Structure and Function of Neisseria gonorrhoeae MtrF Illuminates a Class of Antimetabolite Efflux Pumps
|
|
|
- Christal Dixon
- 5 years ago
- Views:
Transcription
1 Cell Reports Supplemental Information Structure and Function of Neisseria gonorrhoeae MtrF Illuminates a Class of Antimetabolite Efflux Pumps Chih-Chia Su, Jani Reddy Bolla, Nitin Kumar, Abhijith Radhakrishnan, Feng Long, Jared A. Delmar, Tsung-Han Chou, Kanagalaghatta R. Rajashankar, William M. Shafer, and Edward W. Yu
2 Supplemental Figures Fig. S1. Sequence and topology of N.gonorrhoeae MtrF. Related to Figure 1. Alignment of the amino acid sequences of the AbgT family of transporters was done using CLUSTAL W. *, identical residues; :, >60% homologous residues. Secondary structural elements are indicated: TM, transmembrane helix; α, helix. The sequence and topology of N. gonorrhoeae MtrF are shown at the top. Conserved residues involved in lining the channel of the inner core of the protein are highlight with green bars. Fig. S2. Stereo view of the electron density maps of MtrF at a resolution of 3.95 Å. Related to Figure 1. (a) The electron density maps are contoured at 1.2 σ. The Cα traces of the MtrF dimer in the asymmetric unit are included. Anomalous signals of the four Ta 6 Br and six W 6 (µ-o) 6 (µ-cl) 6 Cl 2-6 cluster sites (both contoured at 4 σ) found in the asymmetric unit are colored red and white, respectively. (b) Anomalous maps of the 30 selenium sites (contoured at 4 σ). Two protomers forming a dimer of MtrF are found in the asymmetric unit. Each protomer contributes 15 selenium sites corresponding to the 15 methionines (red). The Cα traces of the two MtrF monomers are colored green and cyan. (c) Representative section of the electron density at the interface of TM2 and TM6 of MtrF. The electron density (colored slate) is contoured at the 1.2 σ level and superimposed with the final refined model (green, carbon; red, oxygen; blue, nitrogen). Fig. S3. Representative gel filtration experiment. Related to Figure 1. The experiment demonstrated that MtrF exists as a dimer in solution. The y axis values were defined as: K av = (V e V 0 )/(V T V 0 ), where V T, V e, and V 0 are the total column volume, elution 1
3 volume, and void volume of the column, respectively. Standards used were the trimeric E. coli CusC channel (M r 12,400) and monomeric N. gonorrhoeae NorM efflux pump (M r 29,000). The void volume was measured using blue dextran (M r 2,000,000). Fig. S4. Surface representation of a cross section of the MtrF protomer. Related to Figure 2. The channel formed within the outer core of the MtrF protomer is colored purple. Fig. S5. Expression level of the MtrF pumps. Related to Figure 3. An immunoblot against MtrF of crude extracts from 50 µg dry cells of strain BL21(DE3) abgt paba expressing the MtrF wild-type and mutant (D193A, S417A, W420A, P438A, R446A, D449A, and P457A) pumps are shown. Fig. S6. Copy numbers of MtrF in the cell. Related to Figure 3. An immunoblot against MtrF of crude extracts from 1.1 x 10 9 cells of BL21(DE3) abgt paba/pet15bωmtrf (lane 1). 6 ng (lane 2), 12 ng (lane 3), 30 ng (lane 4), 60 ng (lane 5), 120 ng (lane 6) and 300 ng (lane 7) of the purified MtrF protein were used as standards. The program ImageJ (Schneider et al., 2012) suggests that the crude cell extracts contain 21 ng MtrF protein, which should correspond to ~200 copies per cell. Fig. S7. Representative isothermal titration calorimetry for the binding of sulfanilamide to MtrF. Related to Figure 5. (a) Each peak corresponds to the injection of 10 µl of 40 µm monomeric MtrF in buffer containing 20 mm Tris-HCl ph 7.5 and 0.03% DDM into 2
4 the reaction containing 1.0 mm sulfanilamide in the same buffer. (b) Cumulative heat of reaction is displayed as a function of the injection number. The solid line is the leastsquare fit to the experimental data, giving a K D of 1.14 ± 0.01 µm. 3
5 Supplemental References Schneider, C.A., Rasband, W.S., and Eliceiri, K.W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9,
6 TM1a TM1b Figure S1 TM2a NgMtrF MSQTDARRSG RFLRTVEWLGNMLPHPVTLFIIFIVLLLIASAVGAYFGLSVPDPRPVGAK---GRADDGL--IHVVSLLDADGLIKILTHTVKNFTGFA 94 EcAbgT -----MSMSSIPSSSQS GKLYGWVERIGNKVPHPFLLFIYLIIVLMVTTAILSAFGVSAKNPTDG TP----VVVKNLLSVEGLHWFLPNVIKNFSGFA 89 AbAbgT MTRTH GWLARLEQLGNRLPHPTLLFVWFCLLLLPLTAVLGALDVTATHPLT-DET ITAHSLLDADGLRYLFTTLVGNFTGFA 81 NmMtrF MSQTDTQRDG RFLRTVEWLGNMLPHPVTLFIIFIVLLLIASAVGAYFGLSVPDPRPVGAK---GRADDGL--IYIVSLLNADGFIKILTHTVKNFTGFA 94 CcAbgT MSDAAPPVSSPPPRQK GLLGVVERLGNLLPEPVMIFVWLILGLMVLSAIGQALGWSASITYAGDEAPQFGELENGVLTYAASSLFSEANLARLFTEMPKTLTSFA 105 SpAbgT MRFLNIVERLGNLLPHPITLFALFCVAVILISGIAGYFELTVADPRPVGSH---GRSADGL--IHVVSLMNAEGLRMIVSNLVTNFTGFT 85 PmMtrF MTTTQQQKKGS KFLHTVEWLGNMLPHPVTLFMIFIVLLLITSALGEYFGLAVADPRPEGVK---GRAADGM--IYVVSLLNAEGLSRILTNLVKNFTNFA 95 VcAbgT MTRREQMSSSASINQNAPKKPLITRFLDGVEYLGNLLPHPITLFAIFCVVLLVASGIAGYFELSVVDPRPEGAK---GRAADGM--IHVVSLLNADGLELIVTNLVKNFVGFA 108 BhAbgT -MKPAPHVELKPNQRG VFVRFLDIIEKYGNKLPDPIMLFVIMAVLILICSAIFSALGTSAVHPGTG EE----IEVVNLLNGEGFILILTELVNNFTSFP 94 PdAbgT MRALNVVERAGNKLPDPVTIFLLLCIIVVILSAVISNLGVEEIHPSTK EV----VKVVNLLEKEQIQSYLGSIVTNFQSFA 77 SaAbgT MTSKHQQKGS IVNRFLNSVEKIGNKLPDPSVLFFLMCVGLAIMTWVISLFNVSVKHPGTH QT----IYIKNIISHDGFTMIMNDTIKNFSEFP 89 TdAbgT MDKKKNKEVS GFLKGVERIGNKLPHPAMIFLILSIIVIIVSALAEAYGTPVTYFDAK--K---GKEVT----IGAVSLLNLDGLRYILNTATKNFTGFA 90 CdAbgT -----MSTTTPPHKTAP SGFLGKIEQLGNRLPDPFWIFAFLAIIVAISSWIGSAIGMTAVNPQDG ST----VEVTNLLTKEGATKMVSEAVNNFVAFP 89 :* ** :*.* :* : : : : :.::..: *. TM2b TM2c HP1a HP1b TM3a TM3b NgMtrF PLGTVLVSLLGVGIAEKSGLISALMRLLLTKSPRKLTTFMVVFTGILSNTASELGYVVLIPLSAVIFHSLGRHPLAGLAAAFAGVSGGYSANLFLGTIDPLLAGITQQAAQIIHPDYVVG 214 EcAbgT PLGAILALVLGAGLAERVGLLPALMVKMASHVNARYASYMVLFIAFFSHISSDAALVIMPPMGALIFLAVGRHPVAGLLAAIAGVGCGFTANLLIVTTDVLLSGISTEAAAAFNPQMHVS 209 AbAbgT PLGVVLVAMLGLGVAEQSGLLSVSLASLVRRSSGGALVFTVAFAGVLSSLTVDAGYVVLIPLAGLVFQLAGRPPIAGIATAFAAVSGGFSANLLVGPVDATLAGLSTEAAHIIDPDRTVA 201 NmMtrF PLGTVLVSLLGVGIAEKSGLISALMRLLLTKSPRKLTTFMVVFTGILSNTASELGYVVLIPLSAIIFHSLGRHPLAGLAAAFAGVSGGYSANLFLGTIDPLLAGITQQAAQIIHPDYVVG 214 CcAbgT PLGLVLVVILGAAVAERSGLFSALIRASLREAPKRILTPLVVIIGMVSHHASDAAYVVFIPLAGLLYAAVGRHPLAGIAAGFAAVSGGFAGNLTPGQFDVVLFGFTQEAARIIDPTWTMN 225 SpAbgT PLGTVLVALLGVGIAERSGLLSAAMRALVMGASKRLVTVTIVFAGIMSNTAAELGYVVLIPMAAMIFHSLGRHPLAGLAAAFAGVSGGYSANLLLGTVDPLLSGITEAAARMIDPDYSVG 205 PmMtrF PLGTVLVALLGVGIAEKAGLLSAVMRLLVTKSPRKLTTFTIVFAGILSNTASELGYVVLIPLAAIIFHSLGRHPLAGLAAAFAGVSGGYSANLLLGTIDPLLAGITQQAAQIIDPTYTVG 215 VcAbgT PLGTVLVAMLGVAIAEHSGLLSAAMRGLVMGASKRMVTFTVVFAGIISNTASELGYVVLIPLAAMLFHSLGRHPLAGLAAAFAGVSGGYSANLLIGTVDPLLSGITETAARMIDPTYSVG 228 BhAbgT PLGLVLVVMLGVGVAESTGLLSALMKTTILNAPRKLILPTIVLVAMLGNAAADAAMVVLPPIVAMIFIALGRHPLAGLAAAYASVAGGFSANLILSMLDPLVAGFTQTGAQMIDPDYVAN 214 PdAbgT PLGLVLVTMLGAGVAEKSGFMEVLMKKGISKVPQKLVTVAIVFAGMLSHTAADVGFIILPPLAALVFLGIGRHPLVGMFAAFAGVAGGFAANVMLSTTDVLLAGFTIPAAQMMDPSYQGN 197 SaAbgT ALGLVLAVMIGIGVAEKTGYFDKLMISVVNRAPRFLILPTIILIGILGSTAGDAATIILPPLAAMLFIKIGYHPIAGLTMAYASAVGGFAANIVVGMQDALVYSFTEPATRIVSDSIKTN 209 TdAbgT PLGTVLVAMLGVGVAEWTGLINTSLKKLLSGVHPRLLTVVVVFAGIMSNVASDAGYVVVIPLGAIVFANAGRHPMAGLAAAFAGVSGGFSANLMLGTIDPLLTGITVEALHNAGMDIAID 210 CdAbgT PLGVIITVMLGVSVAEHSGFISALVRAMVAKVGPKTLTYVVALAGVTGSIASDAVYVILISLGAMSFRALGRSPIVGAMVAFAASSAGFNASLILNITDVLLSGISTSAAQLVDPEYHVS 209.** ::. ::*.:** * : : : :... : : ::..:.: : * *:.*. *. *:..: * :.::. TM4!1 TM5 NgMtrF PEANWFFMAASTFVIALIGYFVTEKIVEPQLGPYQSDLSQEEKDIRHSNE ITPLEYKGLIWAGVVFIALSALLAWSIV-----PADGILRHPET--GLVAGS 309 EcAbgT VIDNWYFMASSVVVLTIVGGLITDKIIEPRLGQWQG-----NSDEKLQT LTESQRFGLRIAGVVSLLFIAAIALMVI-----PQNGILRDPIN--HTVMPS 298 AbAbgT ATGNYWFIIASTFLVTGLVTLITRTLTEPRLAHANTVADASVDAPQIHS RAMKWTGLT-LAILLAGLALLV----LPNDAPLR--HPDTGSVLGS 289 NmMtrF PEANWFFMVASTFVIALIGYFVTEKIVEPQLGPYQSDLSQEEKDIRHSNE ITPLEYKGLIWAGVVFVALSALLAWSIV-----PADGILRHPET--GLVSGS 309 CcAbgT PLGNWWYILAIVVVFTPIAWFLTDKVVEPRLGPWGGQADDALKAELAKSA VTADEKRGLKFAGLAALAIVALFAALSL----IPGFTPLID-ETKTGPAQLT 322 SpAbgT PEVNWYFMFVSTFVITFLGAFVTEKIVEPKLGKYQDGDADETVLQS-MES VSAIEKRGLKWAGLSVLILAIMLALLVV-----PEGAPLRHPDT--GLVSGS 299 PmMtrF PEANWFFMAASTFVIAFIGYFITEKIVEPQLGPYNSHLSQEELDLQHSNE VSPLERKGLRYAGLVFLILCALLAWTVV-----PENGILRDPKT--GLVTGS 310 VcAbgT PEVNWYFMAASTFVIAILGAFVTEKIVEPKLGKYDVSEASDDLSQDKMGS LTALEKKALAYAGLAVVVVSALLAWTIV-----PADGVLRGED---GLVSGS 322 BhAbgT PAMNYYFLVASCLVLVPVAVWVTTKIVEPRLGTYTG-----EVEEITG VTKEEKKGLRWAGISVVILAVVFLFLTV-----PEQALLRDPETGS--LTVS 302 PdAbgT PAMNLYFAFISAIVLTFAGAFVTEKFIAPRFTKYNDS----TNDKDIQE LSELENKGIKYALLSLLVVVVVIVALCI-----GDNAFMKDPETGSILSSNA 289 SaAbgT VAMNWYFIAASVVVLLPTILLVTTKLIIPRLGKYDDSLMHDDHEETSSH ITDKEAHALKWANISFIVTIILLIITAI-----PEHSFLRNAKTGSL-LDDA 304 TdAbgT PTCNWFFMIVSTFILTVVGTFVTEKIVEKNLGTYNGSYKPDNMPIS DVENKGLKLAGISVLIMAVILIIGLFGLPKLPGLAVLREIDPKTGESSIS 306 CdAbgT TLANYFFVVASFLVLALIITAVTELFVKNRARQLVDHDHIDHSELSFRDDDHPDLGAKTDEELAEEIALHSGEIRALTIAGVAFIGMLAVYFALLF----VPASPFYSE-ES----AMSS 320 * ::.:. :*...: : : :. : TM6 TM7a TM7b TM7c HP2a HP2b NgMtrF PFLKS-IVVFIFLLFALPGIVYGRITRSLRGEREVVNAMAESMSTLGLYLVIIFFAAQFVAFFNWTNIGQYIAVKGAVFLKEVGLAGSVLFIGFILICAFINLMIGSASAQWAVTAPIFV 428 EcAbgT PFIKG-IVPLIILFFFVVSLAYGIATRTIRRQADLPHLMIEPMKEMAGFIVMVFPLAQFVAMFNWSNMGKFIAVGLTDILESSGLSGIPAFVGLALLSSFLCMFIASGSAIWSILAPIFV 417 AbAbgT PFIHG-LVVIVALIAGICGAVYGRVSGQFRNSGAVITAMEVTMASMAGYLVLMFFAAQFVAWFNYSQLGLLLAVKGAAWLGALTVPKVVLLLLFVVLTALINLMIGSASAKWSILAPVFI 408 NmMtrF PFLKS-IVVFIFLLFALPGIVYGRVTRSLRGEQEVVNAMAESMSTLGLYLVIIFFAAQFVAFFNWTNIGQYIAVKGATFLKEVGLGGSVLFIGFILICAFINLMIGSASAQWAVTAPIFV 428 CcAbgT PFYGA-LIAGFMMLFLAGGVAYGVGVGTVKTEGDVVNMMADGVRSVAPYIVFAFFAAHFVAMFNWSRLGPIAAIHGAETLKAMNLPAPLLLVSVLGFSSVLDLFIGSASAKWSALAPVVV 441 SpAbgT PFLKG-IVAFIFICFAIPGFVYGKVVGSINNDRDVINAMSHSMSTMGLYIVLVFFAAQFVAFFNWSNLGAVLAVLGADALQSIGLTGPILFLFFIMLCGFINLMLGSASAQWAITAPIFV 418 PmMtrF PFLKS-IVAFIFILFAIPGTVYGIVTKSIRSEKDVVNAMAEAMSTLGLYLVIIFFASQFVAFFNWTNIGQYIAVKGANFLNEVGLHGGILFMGFILICAFINLMIGSASAQWAVTAPIFV 429 VcAbgT PFLKS-IVAFIFIFFAIPGYVYGRVVGTMKTDRDVINAMAKSMSSMGMYIVLVFFAAQFVAFFSWTKFGQVLAVLGADFLKDIGLTGPMLFFAFILMCGYINLMIGSASAQWAVTAPIFV 441 BhAbgT PFMTG-IVPIMMVFFLVPALVYGFVAKVFTSSKDVADHLAKSMSNMGTYIVIAFVAAQMIAFFNWSQLGPIVAIKGANFLQTIGFTGLPLLLGFIVIAALINLMVASASAKWAILAPVFV 421 PdAbgT PLMKG-IVPIITIIFLTPGLVYGKVSKKIKSDKDLVSMMGSSMSDMGGYIVLAFIASQFINLFNLSNLGTILSITGAKLLAESGIPSYGLIIGFILLSGFINLFVGSASAKWAILAPIFV 408 SaAbgT PLING-VGLIILVVFLVPGLVYGILSKEIKNTKDLGKMFGDAVGSMGTFIVIVFFAAQLLAYLKWSNLGIIAAVKGAKLLEHQ--NGIVLILGIIVLSAMVNMLIGSASAKWGILGPIFV 421 TdAbgT NFMHGGLLPVILLLFLIPGLIYGKKTGKINSSHDLVKGMSQAMSSMGGYLVLSFFAAQFVNYFGKTNLGTIISVNGANFLKSIGFTGLPLIISFVIISAFLNLFMGSASAKWAIMAPIFV 426 CdAbgT PLVKA-VTVPISLMFLGLGVVYGITIKSITSLGDIPAFMAKGLTTLIPMVVLFFMVAQFLAWFQWSNLGIWTAIKGAELLQRWDLPVYVLFAAVVLAVALLNLTITSGSAQWALMAPVIV 439 :. :. :. **. : : : : :*: * :::: : :.:* :: : * :.. : : : *.** *..*:.: TM8a TM8b TM9 NgMtrF PMLMLAGYAPEVIQAAYRIGDSVTNIITPMMSYFGLIMATVMKYKKDAGVGTLISMMLPYSAFFLIAWIALFCIWVFVLGLPVGPGAPTLYPAP EcAbgT PMFMLLGFHPAFAQILFRIADSSVLPLAPVSPFVPLFLGFLQRYKPDAKLGTYYSLVLPYPLIFLVVWLLMLLAW-YLVGLPIGPGIYPRLS AbAbgT PMLMLLGISPEASQAAYRVGDSSTNIITPLMPYFVLVLGFARRYQPETGIGTLIALMLPYSLTLLLGWSVLLGVW-IGFGWPLGP NmMtrF PMLMLAGYAPEVIQAAYRIGDSVTNIITPMMSYFGLIMATVIKYKKDAGVGTLISMMLPYSAFFLIAWIALFCIWVFVLGLPVGPGAPTFYPAP CcAbgT PMFMLLGISPEMTTAAYRMGDSFTNLMTPLMSYFPLVLAMTRRWDPSMGVGSLLALMLPYALAFMVAGVAMTLAW-VAFDWPLGPAAQVHYTPPGGLLK 539 SpAbgT PMLMLVGYAPETIQAAYRIGDSVTNLVTPMMSYFGLILAVACRYQKNLGIGTLIATMLPYTLVFFVGWTAFFFLWVFGFGLPVGPGAATYYTP PmMtrF PMLMLTGYSPEIIQAAYRIGDSVTNIITPMMSYFGLIMATVLKYKKDAGVGTLVSMMLPYSVAFLVAWSAMFFIWVFVLGLPVGPNSPMFYPAQ VcAbgT PMLMLVGYAPEVIQAAYRIGDSVTNIITPMMSYFGLILAVATRYMKNLGIGTLIATMLPYSVVFMVGWSLLFYVWVFVLGLPVGPGAATYYTP BhAbgT PMFMLLEYSPAFTQAAYRVGDSITNPITPMLPYFAIALTFAKKYDPKIGIGTFMSSLLPYSIAFAVIWIILFTVW-YLLGLPLGPGEYIHLHT PdAbgT PMFMLLDFNPALTQIAYRIGDASTNPISPLFPYFPVILAFARRYDKDIGIGTVISNMIPYSVVFTLIEIIILLLF-MGIGIPLGPGGGISYVL SaAbgT PMLILIGFHPAFTQVIYRVGDSITNPITPMMPYLPLLLTYAQKYDKRMKLGALLSSLMPYSIALSIVWTVFVIIW-FLLGIPVGPGGPIFVK TdAbgT PMMVNLGLSPALTQVAYRIGDSSTNLITPLMSYFAMIVVFMKKYDEDSGLGTLISTMLPYSIAFLLSWIGLMIIW-YIFGLPLGPGAFIHI CdAbgT PMMMYVGISPEVTQMLFRIGDSPTNIITPMSPYFALALTFLQRYYKPAGVGTLVSLALPYSIAMLVGWFVFFIVW-YALGVPLGPGTPMHFQQG **:: * :*:.*:. ::*:.:. : : :: :*: : :**. : : : :.. *:**!




7 Figure S2 a G92 T91 F90 A94 P95 F93 L96 T98 V316 L100 V101 V317 b L319 L103 F318 L104 F320 V106 L321 L322 G107 I108 G92 T91 c A94 T98 V316 F318 V101 V317 L319 F320 L321 L322 F90 F93 P95 L96 L100 L103 L104 V106 G107 I108
8 Figure S NorM monomer 0.22 K av MtrF dimer CusC trimer Log (M.W.)
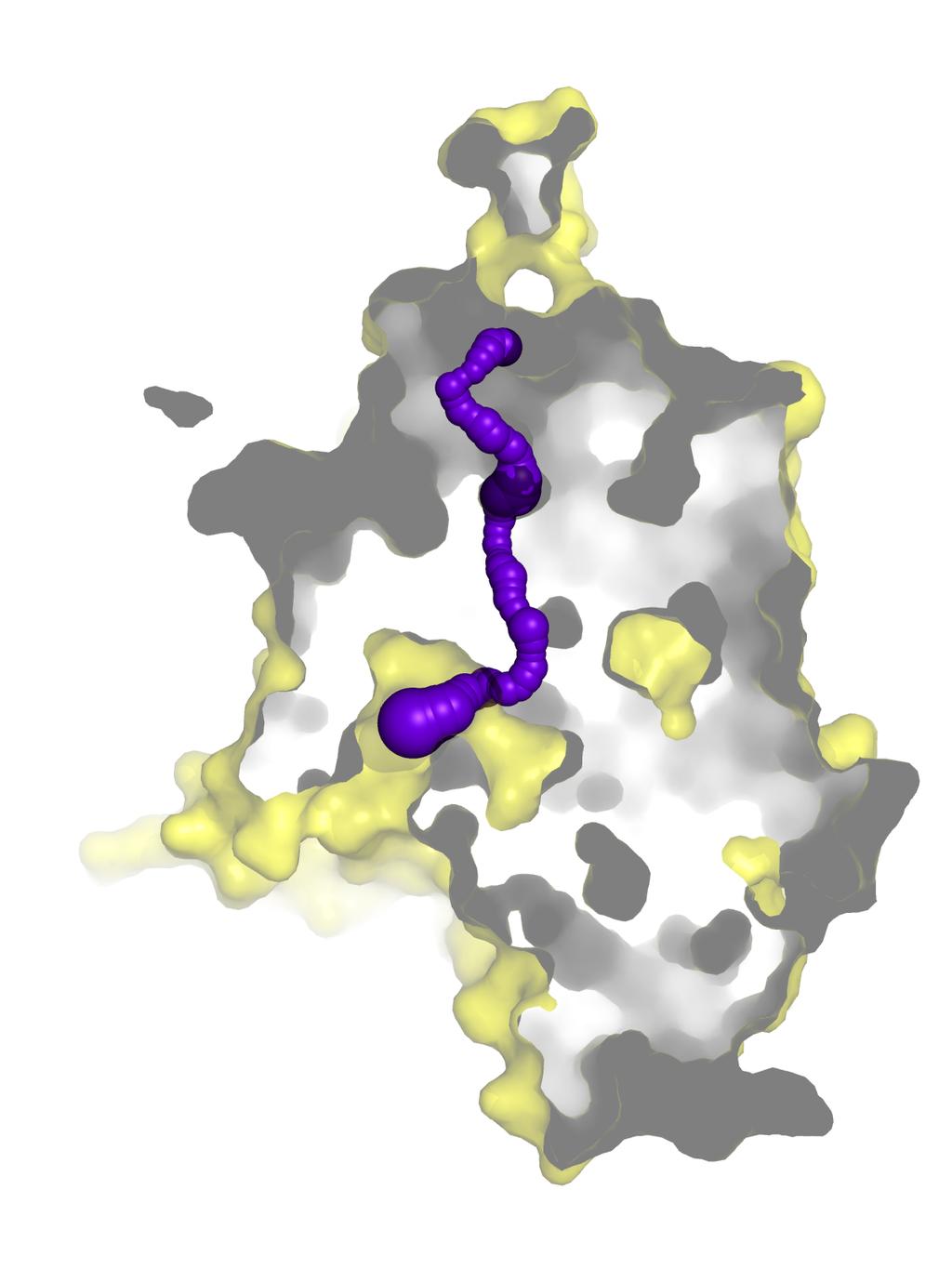
9 Figure S4
10 Figure S5
11 Figure S6
12 Time (min) a Figure S µcal/sec kcal/mole of injectant b Molar Ratio
13 Table S1. Data collection, phasing and structural refinement statistics. Related to Figure 1. Data set Native MtrF Ta 6 Br W 6 (µ-o) 6 (µ- Cl) 6 Cl 6 2- Se (peak) Data Collection Wavelength (Å) Space group P6 5 P6 5 P6 5 P6 5 Resolution (Å) ( ) ( ) ( ) ( ) Cell constants (Å) a b c α, β, γ ( ) 90, 90, , 90, , 90, , 90, 120 Molecules in ASU Redundancy 4.7 (4.0) 5.7 (5.8) 5.6 (6.5) 4.4 (3.8) Total reflections 868,348 28, , ,647 Unique reflections 16, Completeness (%) 98.2 (92.8) 99.9 (99.9) 99.7 (99.0) 99.9 (100) R merge (%) 5.8 (46.8) 5.3 (49.5) 8.6 (48.7) 9.4 (59.0) I / σ (0.88) (2.60) (1.7) (1.95) Phasing Number of sites Phasing power (acentric/centric) 1.71/0.99 Figure of merit 0.59 Refinement MtrF Resolution (Å) R work (%) 29.9 R free (%) 33.6 RMSD bond lengths (Å) RMSD bond angles ( ) 1.52 Ramachandran plot most favoured (%) 89.0 additional allowed (%) 10.0 generously allowed (%) 1.0 disallowed (%) 0.0
14 Table S2. Primers for site-directed mutagenesis. Related to Figure 3. Primer D193A-forward D193A-reverse S417A-forward S417A-reverse W420A-forward W420A-reverse P438A-forward P438A-reverse R446A-forward R446A-reverse D449A-forward D449A-reverse P457A-forward P457A-reverse Sequence 5ʹ -CTGTTCCTGGGCACCATTGCTCCGCTGCTGGCCGGTATC-3ʹ 5ʹ -GATACCGGCCAGCAGCGGAGCAATGGTGCCCAGGAACAG-3ʹ 5ʹ -GGTAGTGCTGCCGCACAATGGGCAGTGACCGCACCGATCT-3ʹ 5ʹ -CCATTGTGCGGCAGCACTACCGATCATCAGGTTAATAAA-3ʹ 5ʹ -CGCACAAGCGGCAGTGACCGCACCGATCTTCGTTCCG-3ʹ 5ʹ -GGTCACTGCCGCTTGTGCGGAAGCACTACCGATCATCAG-3ʹ 5ʹ -GGCTATGCTGCGGAAGTCATTCAGGCCGCATACCGC-3ʹ 5ʹ -GACTTCCGCAGCATAGCCTGCCAGCATCAGCATCGG-3ʹ 5ʹ -GTCATTCAGGCCGCATACGCCATCGGTGATTCAGTTACC-3ʹ 5ʹ -GGTAACTGAATCACCGATGGCGTATGCGGCCTGAATGAC-3ʹ 5ʹ -GCCGCATACCGCATCGGTGCTTCAGTTACCAATATTATC-3ʹ 5ʹ -GATAATATTGGTAACTGAAGCACCGATGCGGTATGCGGC-3ʹ 5ʹ -ATCACGGCGATGATGTCGTATTTTGGTCTGATTATG-3ʹ 5ʹ -CGACATCATCGCCGTGATAATATTGGTAACTGAATC-3ʹ
15 Table S3. MICs of sulfamethazine, sulfadiazine, sulfathiazole and sulfanilamide for different MtrF variants expressed in E. coli BL21(DE3) abgt paba. Related to Figure 5. Gene in BL21(DE3) abgt paba Sulfamethazine (µg/ml) Sulfadiazine (µg/ml) Sulfathiazole (µg/ml) Sulfanilamide (µg/ml) Empty vector mtrf (wild-type) 2000 >250 > mtrf (D193A) mtrf (S417A) mtrf (W420A) mtrf (P438A) mtrf (R446A) 1000 >250 > mtrf (D449A) mtrf (P457A)
16 Table S4. Binding of sulfamethazine, sulfadiazine, sulfathiazole, sulfanilamide and p- aminobenzoic acid by MtrF. Related to Figure 5. K D (µm) H (kcal mol -1 ) S (cal mol deg -1 ) Sulfamethazine 0.33 ± ± Sulfadiazine ± ± Sulfathiazole 1.52 ± ± Sulfanilamide 1.14 ± ± PABA 0.54 ± ±
17 Table S5. Binding of sulfamethazine, sulfadiazine, sulfathiazole, sulfanilamide and p- aminobenzoic acid by the W420A mutant. Related to Figure 5. K D (µm) H (kcal mol -1 ) S (cal mol deg -1 ) Sulfamethazine ± ± Sulfadiazine ± ± Sulfathiazole ± ± Sulfanilamide 6.80 ± ± PABA ± ±
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
SUPPLEMENTARY INFORMATION
 Supplementary materials Figure S1 Fusion protein of Sulfolobus solfataricus SRP54 and a signal peptide. a, Expression vector for the fusion protein. The signal peptide of yeast dipeptidyl aminopeptidase
Supplementary materials Figure S1 Fusion protein of Sulfolobus solfataricus SRP54 and a signal peptide. a, Expression vector for the fusion protein. The signal peptide of yeast dipeptidyl aminopeptidase
REVIEWS. The AbgT family: A novel class of antimetabolite transporters. Jared A. Delmar 1 and Edward W. Yu 1,2 *
 REVIEWS The AbgT family: A novel class of antimetabolite transporters Jared A. Delmar 1 and Edward W. Yu 1,2 * 1 Department of Physics and Astronomy, Iowa State University, Ames, Iowa 50011 2 Department
REVIEWS The AbgT family: A novel class of antimetabolite transporters Jared A. Delmar 1 and Edward W. Yu 1,2 * 1 Department of Physics and Astronomy, Iowa State University, Ames, Iowa 50011 2 Department
Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1.
 Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1. PDZK1 constru cts Amino acids MW [kda] KD [μm] PEPT2-CT- FITC KD [μm] NHE3-CT- FITC KD [μm] PDZK1-CT-
Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1. PDZK1 constru cts Amino acids MW [kda] KD [μm] PEPT2-CT- FITC KD [μm] NHE3-CT- FITC KD [μm] PDZK1-CT-
SUPPLEMENTARY INFORMATION. doi: /nature07461
 Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
Supplementary Information. Structural basis for precursor protein-directed ribosomal peptide macrocyclization
 Supplementary Information Structural basis for precursor protein-directed ribosomal peptide macrocyclization Kunhua Li 1,3, Heather L. Condurso 1,3, Gengnan Li 1, Yousong Ding 2 and Steven D. Bruner 1*
Supplementary Information Structural basis for precursor protein-directed ribosomal peptide macrocyclization Kunhua Li 1,3, Heather L. Condurso 1,3, Gengnan Li 1, Yousong Ding 2 and Steven D. Bruner 1*
SUPPLEMENTARY INFORMATION
 www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
SUPPLEMENTARY INFORMATION
 Supplementary Table 1: Amplitudes of three current levels. Level 0 (pa) Level 1 (pa) Level 2 (pa) TrkA- TrkH WT 200 K 0.01 ± 0.01 9.5 ± 0.01 18.7 ± 0.03 200 Na * 0.001 ± 0.01 3.9 ± 0.01 12.5 ± 0.03 200
Supplementary Table 1: Amplitudes of three current levels. Level 0 (pa) Level 1 (pa) Level 2 (pa) TrkA- TrkH WT 200 K 0.01 ± 0.01 9.5 ± 0.01 18.7 ± 0.03 200 Na * 0.001 ± 0.01 3.9 ± 0.01 12.5 ± 0.03 200
Table 1. Crystallographic data collection, phasing and refinement statistics. Native Hg soaked Mn soaked 1 Mn soaked 2
 Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
SUPPLEMENTARY INFORMATION
 Supplementary Table 1: Data collection, phasing and refinement statistics ChbC/Ta 6 Br 12 Native ChbC Data collection Space group P4 3 2 1 2 P4 3 2 1 2 Cell dimensions a, c (Å) 132.75, 453.57 132.81, 452.95
Supplementary Table 1: Data collection, phasing and refinement statistics ChbC/Ta 6 Br 12 Native ChbC Data collection Space group P4 3 2 1 2 P4 3 2 1 2 Cell dimensions a, c (Å) 132.75, 453.57 132.81, 452.95
Supplementary Figure 1. Biochemical and sequence alignment analyses the
 Supplementary Figure 1. Biochemical and sequence alignment analyses the interaction of OPTN and TBK1. (a) Analytical gel filtration chromatography analysis of the interaction between TBK1 CTD and OPTN(1-119).
Supplementary Figure 1. Biochemical and sequence alignment analyses the interaction of OPTN and TBK1. (a) Analytical gel filtration chromatography analysis of the interaction between TBK1 CTD and OPTN(1-119).
Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R)
") Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R) Shown in cyan and green are two adjacent tetramers from the crystallographic lattice of COP, forming the only unique inter-tetramer
Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R) Shown in cyan and green are two adjacent tetramers from the crystallographic lattice of COP, forming the only unique inter-tetramer
SUPPLEMENTARY INFORMATION
 Table of Contents Page Supplementary Table 1. Diffraction data collection statistics 2 Supplementary Table 2. Crystallographic refinement statistics 3 Supplementary Fig. 1. casic1mfc packing in the R3
Table of Contents Page Supplementary Table 1. Diffraction data collection statistics 2 Supplementary Table 2. Crystallographic refinement statistics 3 Supplementary Fig. 1. casic1mfc packing in the R3
SUPPLEMENTARY INFORMATION
 doi:1.138/nature1737 Supplementary Table 1 variant Description FSEC - 2B12 a FSEC - 6A1 a K d (leucine) c Leucine uptake e K (wild-type like) K (Y18F) K (TS) K (TSY) K288A mutant, lipid facing side chain
doi:1.138/nature1737 Supplementary Table 1 variant Description FSEC - 2B12 a FSEC - 6A1 a K d (leucine) c Leucine uptake e K (wild-type like) K (Y18F) K (TS) K (TSY) K288A mutant, lipid facing side chain
Full-length GlpG sequence was generated by PCR from E. coli genomic DNA. (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
,") Supplementary Methods Protein expression and purification Full-length GlpG sequence was generated by PCR from E. coli genomic DNA (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
Supplementary Methods Protein expression and purification Full-length GlpG sequence was generated by PCR from E. coli genomic DNA (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
SUPPLEMENTARY FIGURES
 SUPPLEMENTARY FIGURES Supplementary Figure 1 Protein sequence alignment of Vibrionaceae with either a 40-residue insertion or a 44-residue insertion. Identical residues are indicated by red background.
SUPPLEMENTARY FIGURES Supplementary Figure 1 Protein sequence alignment of Vibrionaceae with either a 40-residue insertion or a 44-residue insertion. Identical residues are indicated by red background.
THE CRYSTAL STRUCTURE OF THE SGT1-SKP1 COMPLEX: THE LINK BETWEEN
 THE CRYSTAL STRUCTURE OF THE SGT1-SKP1 COMPLEX: THE LINK BETWEEN HSP90 AND BOTH SCF E3 UBIQUITIN LIGASES AND KINETOCHORES Oliver Willhoft, Richard Kerr, Dipali Patel, Wenjuan Zhang, Caezar Al-Jassar, Tina
THE CRYSTAL STRUCTURE OF THE SGT1-SKP1 COMPLEX: THE LINK BETWEEN HSP90 AND BOTH SCF E3 UBIQUITIN LIGASES AND KINETOCHORES Oliver Willhoft, Richard Kerr, Dipali Patel, Wenjuan Zhang, Caezar Al-Jassar, Tina
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Chemical structure of LPS and LPS biogenesis in Gram-negative bacteria. a. Chemical structure of LPS. LPS molecule consists of Lipid A, core oligosaccharide and O-antigen. The polar
Supplementary Figure 1 Chemical structure of LPS and LPS biogenesis in Gram-negative bacteria. a. Chemical structure of LPS. LPS molecule consists of Lipid A, core oligosaccharide and O-antigen. The polar
SUPPLEMENTARY INFORMATION
 Fig. 1 Influences of crystal lattice contacts on Pol η structures. a. The dominant lattice contact between two hpol η molecules (silver and gold) in the type 1 crystals. b. A close-up view of the hydrophobic
Fig. 1 Influences of crystal lattice contacts on Pol η structures. a. The dominant lattice contact between two hpol η molecules (silver and gold) in the type 1 crystals. b. A close-up view of the hydrophobic
Structural basis of PROTAC cooperative recognition for selective protein degradation
 SUPPLEMENTARY INFORMATION Structural basis of PROTAC cooperative recognition for selective protein degradation Morgan S. Gadd 1, Andrea Testa 1, Xavier Lucas 1, Kwok-Ho Chan, Wenzhang Chen, Douglas J.
SUPPLEMENTARY INFORMATION Structural basis of PROTAC cooperative recognition for selective protein degradation Morgan S. Gadd 1, Andrea Testa 1, Xavier Lucas 1, Kwok-Ho Chan, Wenzhang Chen, Douglas J.
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10458 Active Site Remodeling in the Bifunctional Fructose-1,6- bisphosphate aldolase/phosphatase Juan Du, Rafael F. Say, Wei Lü, Georg Fuchs & Oliver Einsle SUPPLEMENTARY FIGURES Figure
doi:10.1038/nature10458 Active Site Remodeling in the Bifunctional Fructose-1,6- bisphosphate aldolase/phosphatase Juan Du, Rafael F. Say, Wei Lü, Georg Fuchs & Oliver Einsle SUPPLEMENTARY FIGURES Figure
SUPPLEMENTARY INFORMATION
 Supplementary Table S1 Kinetic Analyses of the AMSH-LP mutants AMSH-LP K M (μm) k cat x 10-3 (s -1 ) WT 71.8 ± 6.3 860 ± 65.4 T353A 76.8 ± 11.7 46.3 ± 3.7 F355A 58.9 ± 10.4 5.33 ± 0.30 proximal S358A 75.1
Supplementary Table S1 Kinetic Analyses of the AMSH-LP mutants AMSH-LP K M (μm) k cat x 10-3 (s -1 ) WT 71.8 ± 6.3 860 ± 65.4 T353A 76.8 ± 11.7 46.3 ± 3.7 F355A 58.9 ± 10.4 5.33 ± 0.30 proximal S358A 75.1
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12045 Supplementary Table 1 Data collection and refinement statistics. Native Pt-SAD X-ray source SSRF BL17U SPring-8 BL41XU Wavelength (Å) 0.97947 1.07171 Space group P2 1 2 1 2 1 P2
doi:10.1038/nature12045 Supplementary Table 1 Data collection and refinement statistics. Native Pt-SAD X-ray source SSRF BL17U SPring-8 BL41XU Wavelength (Å) 0.97947 1.07171 Space group P2 1 2 1 2 1 P2
Supplemental Information. Molecular Basis of Spectral Diversity. in Near-Infrared Phytochrome-Based. Fluorescent Proteins
 Chemistry & Biology, Volume 22 Supplemental Information Molecular Basis of Spectral Diversity in Near-Infrared Phytochrome-Based Fluorescent Proteins Daria M. Shcherbakova, Mikhail Baloban, Sergei Pletnev,
Chemistry & Biology, Volume 22 Supplemental Information Molecular Basis of Spectral Diversity in Near-Infrared Phytochrome-Based Fluorescent Proteins Daria M. Shcherbakova, Mikhail Baloban, Sergei Pletnev,
According to the manufacture s direction (Pierce), RNA and DNA
, RNA and DNA") Supplementary method Electrophoretic Mobility-shift assay (EMSA) According to the manufacture s direction (Pierce), RNA and DNA oligonuleotides were firstly labeled by biotin. TAVb (1pM) was incubated
Supplementary method Electrophoretic Mobility-shift assay (EMSA) According to the manufacture s direction (Pierce), RNA and DNA oligonuleotides were firstly labeled by biotin. TAVb (1pM) was incubated
Plasmid Relevant features Source. W18N_D20N and TrXE-W18N_D20N-anti
 Table S1. E. coli plasmids Plasmid Relevant features Source pdg680 T. reesei XynII AA 2-190 with C-terminal His 6 tag optimized for E. coli expression in pjexpress401 Wan et al. (in press) psbn44d psbn44h
Table S1. E. coli plasmids Plasmid Relevant features Source pdg680 T. reesei XynII AA 2-190 with C-terminal His 6 tag optimized for E. coli expression in pjexpress401 Wan et al. (in press) psbn44d psbn44h
Supplementary Information
 Supplementary Information The direct role of selenocysteine in [NiFeSe] hydrogenase maturation and catalysis Marta C. Marques a, Cristina Tapia b, Oscar Gutiérrez-Sanz b, Ana Raquel Ramos a, Kimberly L.
Supplementary Information The direct role of selenocysteine in [NiFeSe] hydrogenase maturation and catalysis Marta C. Marques a, Cristina Tapia b, Oscar Gutiérrez-Sanz b, Ana Raquel Ramos a, Kimberly L.
Table S1. Primers used for the constructions of recombinant GAL1 and λ5 mutants. GAL1-E74A ccgagcagcgggcggctgtctttcc ggaaagacagccgcccgctgctcgg
 SUPPLEMENTAL DATA Table S1. Primers used for the constructions of recombinant GAL1 and λ5 mutants Sense primer (5 to 3 ) Anti-sense primer (5 to 3 ) GAL1 mutants GAL1-E74A ccgagcagcgggcggctgtctttcc ggaaagacagccgcccgctgctcgg
SUPPLEMENTAL DATA Table S1. Primers used for the constructions of recombinant GAL1 and λ5 mutants Sense primer (5 to 3 ) Anti-sense primer (5 to 3 ) GAL1 mutants GAL1-E74A ccgagcagcgggcggctgtctttcc ggaaagacagccgcccgctgctcgg
SUPPLEMENTARY INFORMATION
 Data collection Supplementary Table 1 Statistics of data collection, phasing and refinement Native Se-MAD Space group P2 1 2 1 2 1 P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 50.4, 94.2, 115.4 49.8, 94.2,
Data collection Supplementary Table 1 Statistics of data collection, phasing and refinement Native Se-MAD Space group P2 1 2 1 2 1 P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 50.4, 94.2, 115.4 49.8, 94.2,
T H E J O U R N A L O F G E N E R A L P H Y S I O L O G Y. jgp
 S u p p l e m e n ta l m at e r i a l jgp Lee et al., http://www.jgp.org/cgi/content/full/jgp.201411219/dc1 T H E J O U R N A L O F G E N E R A L P H Y S I O L O G Y S u p p l e m e n ta l D I S C U S
S u p p l e m e n ta l m at e r i a l jgp Lee et al., http://www.jgp.org/cgi/content/full/jgp.201411219/dc1 T H E J O U R N A L O F G E N E R A L P H Y S I O L O G Y S u p p l e m e n ta l D I S C U S
SUPPLEMENTARY INFORMATION
 Dph2 SeMet (iron-free) # Dph2 (iron-free) Dph2-[4Fe-4S] Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 58.26, 82.08, 160.42 58.74, 81.87, 160.01 55.70, 80.53,
Dph2 SeMet (iron-free) # Dph2 (iron-free) Dph2-[4Fe-4S] Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 58.26, 82.08, 160.42 58.74, 81.87, 160.01 55.70, 80.53,
Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions. Implicated in Dimerization and Autoinhibition
 Reveals Regions. Implicated in Dimerization and Autoinhibition") JBC Papers in Press. Published on November 1, 2000 as Manuscript M006502200 Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions Implicated in Dimerization and Autoinhibition 1 Copyright
JBC Papers in Press. Published on November 1, 2000 as Manuscript M006502200 Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions Implicated in Dimerization and Autoinhibition 1 Copyright
type GroEL-GroES complex. Crystals were grown in buffer D (100 mm HEPES, ph 7.5,
 Supplementary Material Supplementary Materials and Methods Structure Determination of SR1-GroES-ADP AlF x SR1-GroES-ADP AlF x was purified as described in Materials and Methods for the wild type GroEL-GroES
Supplementary Material Supplementary Materials and Methods Structure Determination of SR1-GroES-ADP AlF x SR1-GroES-ADP AlF x was purified as described in Materials and Methods for the wild type GroEL-GroES
of the Guanine Nucleotide Exchange Factor FARP2
 Structure, Volume 21 Supplemental Information Structural Basis for Autoinhibition of the Guanine Nucleotide Exchange Factor FARP2 Xiaojing He, Yi-Chun Kuo, Tyler J. Rosche, and Xuewu Zhang Inventory of
Structure, Volume 21 Supplemental Information Structural Basis for Autoinhibition of the Guanine Nucleotide Exchange Factor FARP2 Xiaojing He, Yi-Chun Kuo, Tyler J. Rosche, and Xuewu Zhang Inventory of
Supplementary figure 1. Comparison of unbound ogm-csf and ogm-csf as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine
 Supplementary figure 1. Comparison of unbound and as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine GM-CSF (slate) with bound GM-CSF in the GIF:GM-CSF complex (GIF: green,
Supplementary figure 1. Comparison of unbound and as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine GM-CSF (slate) with bound GM-CSF in the GIF:GM-CSF complex (GIF: green,
Supplemental Information
 Supplemental Information Combinatorial Readout of Unmodified H3R2 and Acetylated H3K14 by the Tandem PHD Finger of MOZ Reveals a Regulatory Mechanism for HOXA9 Transcription Yu Qiu 1, Lei Liu 1, Chen Zhao
Supplemental Information Combinatorial Readout of Unmodified H3R2 and Acetylated H3K14 by the Tandem PHD Finger of MOZ Reveals a Regulatory Mechanism for HOXA9 Transcription Yu Qiu 1, Lei Liu 1, Chen Zhao
Nature Structural and Molecular Biology: doi: /nsmb Supplementary Figure 1. Definition and assessment of ciap1 constructs.
 Supplementary Figure 1 Definition and assessment of ciap1 constructs. (a) ciap1 constructs used in this study are shown as primary structure schematics with domains colored as in the main text. Mutations
Supplementary Figure 1 Definition and assessment of ciap1 constructs. (a) ciap1 constructs used in this study are shown as primary structure schematics with domains colored as in the main text. Mutations
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Identification of the ScDcp2 minimal region interacting with both ScDcp1 and the ScEdc3 LSm domain. Pull-down experiment of untagged ScEdc3 LSm with various ScDcp1-Dcp2-His 6 fragments.
Supplementary Figure 1 Identification of the ScDcp2 minimal region interacting with both ScDcp1 and the ScEdc3 LSm domain. Pull-down experiment of untagged ScEdc3 LSm with various ScDcp1-Dcp2-His 6 fragments.
Supporting Protocol This protocol describes the construction and the force-field parameters of the non-standard residue for the Ag + -site using CNS
 Supporting Protocol This protocol describes the construction and the force-field parameters of the non-standard residue for the Ag + -site using CNS CNS input file generatemetal.inp: remarks file generate/generatemetal.inp
Supporting Protocol This protocol describes the construction and the force-field parameters of the non-standard residue for the Ag + -site using CNS CNS input file generatemetal.inp: remarks file generate/generatemetal.inp
Supplementary materials. Crystal structure of the carboxyltransferase domain. of acetyl coenzyme A carboxylase. Department of Biological Sciences
 Supplementary materials Crystal structure of the carboxyltransferase domain of acetyl coenzyme A carboxylase Hailong Zhang, Zhiru Yang, 1 Yang Shen, 1 Liang Tong Department of Biological Sciences Columbia
Supplementary materials Crystal structure of the carboxyltransferase domain of acetyl coenzyme A carboxylase Hailong Zhang, Zhiru Yang, 1 Yang Shen, 1 Liang Tong Department of Biological Sciences Columbia
Supplemental Methods. Protein expression and purification
 Supplemental Methods Protein expression and purification The isolated collagen-binding domain of hlair-1, amino acid 22-122, was cloned into pet3xa using introduced BamHI and NotI sites at the 5 and 3
Supplemental Methods Protein expression and purification The isolated collagen-binding domain of hlair-1, amino acid 22-122, was cloned into pet3xa using introduced BamHI and NotI sites at the 5 and 3
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11539 Supplementary Figure 1 Schematic representation of plant (A) and mammalian (B) P 2B -ATPase domain organization. Actuator (A-), nucleotide binding (N-),
SUPPLEMENTARY INFORMATION doi:10.1038/nature11539 Supplementary Figure 1 Schematic representation of plant (A) and mammalian (B) P 2B -ATPase domain organization. Actuator (A-), nucleotide binding (N-),
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1: The HpUreI crystal used for collection of native diffraction data. The crystal belongs to spacegroup P4 2 2 1 2 and has an approximate maximal dimension of 0.25 mm. Supplementary
Supplementary Figure 1: The HpUreI crystal used for collection of native diffraction data. The crystal belongs to spacegroup P4 2 2 1 2 and has an approximate maximal dimension of 0.25 mm. Supplementary
CH 3 CH 2 OH +H 2 O CHO. 2e + 2H + + O 2 H 2 O +HCOOH
 2 4 H CH 3 2e + 2H + + 2 H 2 2 H CH 2 H 2e + 2H + + 2 H 2 2 H +H 2 CH 2e + 2H + + 2 H 2 2 H +HCH Supplemental Figure S. The three-step 4DM reaction, each step requires two reducing equivalents from ADPH
2 4 H CH 3 2e + 2H + + 2 H 2 2 H CH 2 H 2e + 2H + + 2 H 2 2 H +H 2 CH 2e + 2H + + 2 H 2 2 H +HCH Supplemental Figure S. The three-step 4DM reaction, each step requires two reducing equivalents from ADPH
for Molecular Biology and Neuroscience and Institute of Medical Microbiology, Rikshospitalet-Radiumhospitalet
 SUPPLEMENTARY INFORMATION TO Structural basis for enzymatic excision of N -methyladenine and N 3 -methylcytosine from DNA Ingar Leiros,5, Marivi P. Nabong 2,3,5, Kristin Grøsvik 3, Jeanette Ringvoll 2,
SUPPLEMENTARY INFORMATION TO Structural basis for enzymatic excision of N -methyladenine and N 3 -methylcytosine from DNA Ingar Leiros,5, Marivi P. Nabong 2,3,5, Kristin Grøsvik 3, Jeanette Ringvoll 2,
SUPPLEMENTARY INFORMATION
 Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Cryo-EM data collection, refinement and validation statistics
 1 Table S1 Cryo-EM data collection, refinement and validation statistics Data collection and processing CPSF-160 WDR33 (EMDB-7114) (PDB 6BM0) CPSF-160 WDR33 (EMDB-7113) (PDB 6BLY) CPSF-160 WDR33 CPSF-30
1 Table S1 Cryo-EM data collection, refinement and validation statistics Data collection and processing CPSF-160 WDR33 (EMDB-7114) (PDB 6BM0) CPSF-160 WDR33 (EMDB-7113) (PDB 6BLY) CPSF-160 WDR33 CPSF-30
Structural insights into Aspergillus fumigatus lectin specificity - AFL binding sites are functionally non-equivalent
 Acta Cryst. (2015). D71, doi:10.1107/s1399004714026595 Supporting information Volume 71 (2015) Supporting information for article: Structural insights into Aspergillus fumigatus lectin specificity - AFL
Acta Cryst. (2015). D71, doi:10.1107/s1399004714026595 Supporting information Volume 71 (2015) Supporting information for article: Structural insights into Aspergillus fumigatus lectin specificity - AFL
Stabilizing the CH2 domain of an Antibody by Engineering in an Enhanced Aromatic Sequon
 Stabilizing the CH2 domain of an Antibody by Engineering in an Enhanced Aromatic Sequon Wentao Chen,, Leopold Kong, Stephen Connelly, Julia M. Dendle,, Yu Liu,, Ian A. Wilson,#, Evan T. Powers, *, Jeffery
Stabilizing the CH2 domain of an Antibody by Engineering in an Enhanced Aromatic Sequon Wentao Chen,, Leopold Kong, Stephen Connelly, Julia M. Dendle,, Yu Liu,, Ian A. Wilson,#, Evan T. Powers, *, Jeffery
Acta Cryst. (2017). D73, doi: /s
. D73, doi: /s") Acta Cryst. (2017). D73, doi:10.1107/s2059798317010932 Supporting information Volume 73 (2017) Supporting information for article: Designing better diffracting crystals of biotin carboxyl carrier protein
Acta Cryst. (2017). D73, doi:10.1107/s2059798317010932 Supporting information Volume 73 (2017) Supporting information for article: Designing better diffracting crystals of biotin carboxyl carrier protein
SUPPLEMENTARY INFORMATION
 doi:10.108/nature11899 Supplementar Table 1. Data collection and refinement statistics (+TPMP, native) (-TPMP, native) (+TPMP, recombinant) (MgCl ) (MgSO ) Data collection Space group C P 1 C P 1 1 P 1
doi:10.108/nature11899 Supplementar Table 1. Data collection and refinement statistics (+TPMP, native) (-TPMP, native) (+TPMP, recombinant) (MgCl ) (MgSO ) Data collection Space group C P 1 C P 1 1 P 1
Diphthamide biosynthesis requires a radical iron-sulfur enzyme. Pennsylvania State University, University Park, Pennsylvania 16802, USA
 Diphthamide biosynthesis requires a radical iron-sulfur enzyme Yang Zhang, 1,4 Xuling Zhu, 1,4 Andrew T. Torelli, 1 Michael Lee, 2 Boris Dzikovski, 1 Rachel Koralewski, 1 Eileen Wang, 1 Jack Freed, 1 Carsten
Diphthamide biosynthesis requires a radical iron-sulfur enzyme Yang Zhang, 1,4 Xuling Zhu, 1,4 Andrew T. Torelli, 1 Michael Lee, 2 Boris Dzikovski, 1 Rachel Koralewski, 1 Eileen Wang, 1 Jack Freed, 1 Carsten
Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing
 Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing 0.1% SDS without boiling. The gel was analyzed by a fluorescent
Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing 0.1% SDS without boiling. The gel was analyzed by a fluorescent
Crystal lattice Real Space. Reflections Reciprocal Space. I. Solving Phases II. Model Building for CHEM 645. Purified Protein. Build model.
 I. Solving Phases II. Model Building for CHEM 645 Purified Protein Solve Phase Build model and refine Crystal lattice Real Space Reflections Reciprocal Space ρ (x, y, z) pronounced rho F hkl 2 I F (h,
I. Solving Phases II. Model Building for CHEM 645 Purified Protein Solve Phase Build model and refine Crystal lattice Real Space Reflections Reciprocal Space ρ (x, y, z) pronounced rho F hkl 2 I F (h,
Supplementary Figures
 1 Supplementary Figures Supplementary Figure 1 Type I FGFR1 inhibitors (a) Chemical structures of a pyrazolylaminopyrimidine inhibitor (henceforth referred to as PAPI; PDB-code of the FGFR1-PAPI complex:
1 Supplementary Figures Supplementary Figure 1 Type I FGFR1 inhibitors (a) Chemical structures of a pyrazolylaminopyrimidine inhibitor (henceforth referred to as PAPI; PDB-code of the FGFR1-PAPI complex:
SI Text S1 Solution Scattering Data Collection and Analysis. SI references
 SI Text S1 Solution Scattering Data Collection and Analysis. The X-ray photon energy was set to 8 kev. The PILATUS hybrid pixel array detector (RIGAKU) was positioned at a distance of 606 mm from the sample.
SI Text S1 Solution Scattering Data Collection and Analysis. The X-ray photon energy was set to 8 kev. The PILATUS hybrid pixel array detector (RIGAKU) was positioned at a distance of 606 mm from the sample.
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature17991 Supplementary Discussion Structural comparison with E. coli EmrE The DMT superfamily includes a wide variety of transporters with 4-10 TM segments 1. Since the subfamilies of the
doi:10.1038/nature17991 Supplementary Discussion Structural comparison with E. coli EmrE The DMT superfamily includes a wide variety of transporters with 4-10 TM segments 1. Since the subfamilies of the
Expanded View Figures
 The EMBO Journal Structure of a Dm peptide bound to the OT module Tobias Raisch et al Expanded View Figures A Hs Dm 262 297 685 8 HEAT HEAT MIF4G 9BD 1SHD 761 91 193 169 1152 1317 16 1376 1467 HEAT HEAT
The EMBO Journal Structure of a Dm peptide bound to the OT module Tobias Raisch et al Expanded View Figures A Hs Dm 262 297 685 8 HEAT HEAT MIF4G 9BD 1SHD 761 91 193 169 1152 1317 16 1376 1467 HEAT HEAT
Table S1. Theoretical and apparent molecular weights of the proteins and protein complexes used for ITC analysis
 Table S1. Theoretical and apparent molecular weights of the proteins and protein complexes used for ITC analysis Sample Theoretical molecular weight (without glycosylation) Apparent molecular weight R-2
Table S1. Theoretical and apparent molecular weights of the proteins and protein complexes used for ITC analysis Sample Theoretical molecular weight (without glycosylation) Apparent molecular weight R-2
Supporting Information
 Supporting Information Oxaliplatin binding to human copper chaperone Atox1 and protein dimerization Benny D. Belviso, 1 Angela Galliani, 2 Alessia Lasorsa, 2 Valentina Mirabelli, 1,3 Rocco Caliandro, 1
Supporting Information Oxaliplatin binding to human copper chaperone Atox1 and protein dimerization Benny D. Belviso, 1 Angela Galliani, 2 Alessia Lasorsa, 2 Valentina Mirabelli, 1,3 Rocco Caliandro, 1
Supplementary Information
 1 Supplementary Information Figure S1 The V=0.5 Harker section of an anomalous difference Patterson map calculated using diffraction data from the NNQQNY crystal at 1.3 Å resolution. The position of the
1 Supplementary Information Figure S1 The V=0.5 Harker section of an anomalous difference Patterson map calculated using diffraction data from the NNQQNY crystal at 1.3 Å resolution. The position of the
Supplementary Information. The protease GtgE from Salmonella exclusively targets. inactive Rab GTPases
 Supplementary Information The protease GtgE from Salmonella exclusively targets inactive Rab GTPases Table of Contents Supplementary Figures... 2 Supplementary Figure 1... 2 Supplementary Figure 2... 3
Supplementary Information The protease GtgE from Salmonella exclusively targets inactive Rab GTPases Table of Contents Supplementary Figures... 2 Supplementary Figure 1... 2 Supplementary Figure 2... 3
Bacterial protease uses distinct thermodynamic signatures for substrate recognition
 Bacterial protease uses distinct thermodynamic signatures for substrate recognition Gustavo Arruda Bezerra, Yuko Ohara-Nemoto, Irina Cornaciu, Sofiya Fedosyuk, Guillaume Hoffmann, Adam Round, José A. Márquez,
Bacterial protease uses distinct thermodynamic signatures for substrate recognition Gustavo Arruda Bezerra, Yuko Ohara-Nemoto, Irina Cornaciu, Sofiya Fedosyuk, Guillaume Hoffmann, Adam Round, José A. Márquez,
Supplemental Data SUPPLEMENTAL FIGURES
 Supplemental Data CRYSTAL STRUCTURE OF THE MG.ADP-INHIBITED STATE OF THE YEAST F 1 C 10 ATP SYNTHASE Alain Dautant*, Jean Velours and Marie-France Giraud* From Université Bordeaux 2, CNRS; Institut de
Supplemental Data CRYSTAL STRUCTURE OF THE MG.ADP-INHIBITED STATE OF THE YEAST F 1 C 10 ATP SYNTHASE Alain Dautant*, Jean Velours and Marie-France Giraud* From Université Bordeaux 2, CNRS; Institut de
Structural insights into WcbI, a novel polysaccharide-biosynthesis enzyme
 Volume 1 (2014) Supporting information for article: Structural insights into WcbI, a novel polysaccharide-biosynthesis enzyme Mirella Vivoli, Emily Ayres, Edward Beaumont, Michail N. Isupov and Nicholas
Volume 1 (2014) Supporting information for article: Structural insights into WcbI, a novel polysaccharide-biosynthesis enzyme Mirella Vivoli, Emily Ayres, Edward Beaumont, Michail N. Isupov and Nicholas
ml. ph 7.5 ph 6.5 ph 5.5 ph 4.5. β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS
 a UV28 absorption (mau) 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS β 2 AR-Gs complex dissociated complex excess nucleotides b 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex
a UV28 absorption (mau) 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS β 2 AR-Gs complex dissociated complex excess nucleotides b 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex
Supporting Information
 Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
Supplementary Figure S1. MscS orientation in spheroplasts and liposomes (a) Current-voltage relationship for wild-type MscS expressed in E.
 Current-voltage relationship for wild-type MscS expressed in E.") a b c Supplementary Figure S1. MscS orientation in spheroplasts and liposomes (a) Current-voltage relationship for wild-type MscS expressed in E. coli giant spheroplasts (MJF465) and reconstituted into
a b c Supplementary Figure S1. MscS orientation in spheroplasts and liposomes (a) Current-voltage relationship for wild-type MscS expressed in E. coli giant spheroplasts (MJF465) and reconstituted into
Microcalorimetry for the Life Sciences
 Microcalorimetry for the Life Sciences Why Microcalorimetry? Microcalorimetry is universal detector Heat is generated or absorbed in every chemical process In-solution No molecular weight limitations Label-free
Microcalorimetry for the Life Sciences Why Microcalorimetry? Microcalorimetry is universal detector Heat is generated or absorbed in every chemical process In-solution No molecular weight limitations Label-free
The copper active site in CBM33 polysaccharide oxygenases
 Supporting Information for: The copper active site in CBM33 polysaccharide oxygenases Glyn R. Hemsworth, Edward J. Taylor, Robbert Q. Kim, Rebecca C. Gregory, Sally J. Lewis, Johan P. Turkenburg, Alison
Supporting Information for: The copper active site in CBM33 polysaccharide oxygenases Glyn R. Hemsworth, Edward J. Taylor, Robbert Q. Kim, Rebecca C. Gregory, Sally J. Lewis, Johan P. Turkenburg, Alison
Structure and RNA-binding properties. of the Not1 Not2 Not5 module of the yeast Ccr4 Not complex
 Structure and RNA-binding properties of the Not1 Not2 Not5 module of the yeast Ccr4 Not complex Varun Bhaskar 1, Vladimir Roudko 2,3, Jerome Basquin 1, Kundan Sharma 4, Henning Urlaub 4, Bertrand Seraphin
Structure and RNA-binding properties of the Not1 Not2 Not5 module of the yeast Ccr4 Not complex Varun Bhaskar 1, Vladimir Roudko 2,3, Jerome Basquin 1, Kundan Sharma 4, Henning Urlaub 4, Bertrand Seraphin
RNA Polymerase I Contains a TFIIF-Related DNA-Binding Subcomplex
 Molecular Cell, Volume 39 Supplemental Information RNA Polymerase I Contains a TFIIFRelated DNABinding Subcomplex Sebastian R. Geiger, Kristina Lorenzen, Amelie Schreieck, Patrizia Hanecker, Dirk Kostrewa,
Molecular Cell, Volume 39 Supplemental Information RNA Polymerase I Contains a TFIIFRelated DNABinding Subcomplex Sebastian R. Geiger, Kristina Lorenzen, Amelie Schreieck, Patrizia Hanecker, Dirk Kostrewa,
Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits. AhpC and AhpF from Escherichia coli
 Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits AhpC and AhpF from Escherichia coli Phat Vinh Dip 1,#, Neelagandan Kamariah 2,#, Malathy Sony Subramanian Manimekalai
Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits AhpC and AhpF from Escherichia coli Phat Vinh Dip 1,#, Neelagandan Kamariah 2,#, Malathy Sony Subramanian Manimekalai
Building a Homology Model of the Transmembrane Domain of the Human Glycine α-1 Receptor
 Building a Homology Model of the Transmembrane Domain of the Human Glycine α-1 Receptor Presented by Stephanie Lee Research Mentor: Dr. Rob Coalson Glycine Alpha 1 Receptor (GlyRa1) Member of the superfamily
Building a Homology Model of the Transmembrane Domain of the Human Glycine α-1 Receptor Presented by Stephanie Lee Research Mentor: Dr. Rob Coalson Glycine Alpha 1 Receptor (GlyRa1) Member of the superfamily
Advanced Certificate in Principles in Protein Structure. You will be given a start time with your exam instructions
 BIRKBECK COLLEGE (University of London) Advanced Certificate in Principles in Protein Structure MSc Structural Molecular Biology Date: Thursday, 1st September 2011 Time: 3 hours You will be given a start
BIRKBECK COLLEGE (University of London) Advanced Certificate in Principles in Protein Structure MSc Structural Molecular Biology Date: Thursday, 1st September 2011 Time: 3 hours You will be given a start
Supplementary Figure 1
 A R R RA-selective pocket Cl Adenine pocket and hinge-binding moiety Cl ulfonamide series PLX7 PLX Br BR BR TV PLX RI TQ D RI9 C B PLX7 M ulfonamide concentration Monomer Dimer RA-elective Pocket Unoccupied
A R R RA-selective pocket Cl Adenine pocket and hinge-binding moiety Cl ulfonamide series PLX7 PLX Br BR BR TV PLX RI TQ D RI9 C B PLX7 M ulfonamide concentration Monomer Dimer RA-elective Pocket Unoccupied
The Fic protein Doc uses an inverted substrate to phosphorylate and. inactivate EF-Tu
 The Fic protein Doc uses an inverted substrate to phosphorylate and inactivate EF-Tu Daniel Castro-Roa 1, Abel Garcia-Pino 2,3 *, Steven De Gieter 2,3, Nico A.J. van Nuland 2,3, Remy Loris 2,3, Nikolay
The Fic protein Doc uses an inverted substrate to phosphorylate and inactivate EF-Tu Daniel Castro-Roa 1, Abel Garcia-Pino 2,3 *, Steven De Gieter 2,3, Nico A.J. van Nuland 2,3, Remy Loris 2,3, Nikolay
Supporting Information. UV-induced ligand exchange in MHC class I protein crystals
 Supporting Information for the article entitled UV-induced ligand exchange in MHC class I protein crystals by Patrick H.N. Celie 1, Mireille Toebes 2, Boris Rodenko 3, Huib Ovaa 3, Anastassis Perrakis
Supporting Information for the article entitled UV-induced ligand exchange in MHC class I protein crystals by Patrick H.N. Celie 1, Mireille Toebes 2, Boris Rodenko 3, Huib Ovaa 3, Anastassis Perrakis
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11744 Supplementary Table 1. Crystallographic data collection and refinement statistics. Wild-type Se-Met-BcsA-B SmCl 3 -soaked EMTS-soaked Data collection Space
SUPPLEMENTARY INFORMATION doi:10.1038/nature11744 Supplementary Table 1. Crystallographic data collection and refinement statistics. Wild-type Se-Met-BcsA-B SmCl 3 -soaked EMTS-soaked Data collection Space
Nature Structural and Molecular Biology: doi: /nsmb.2783
 Supplementary Figure 1: Crystallized chimera construct (mhv1cc). (a) Sequence alignment between mhv1cc and other VSDs. These sequences (mhv1cc, Kv1.2 Kv2.1; shaker family voltage gated potassium channel
Supplementary Figure 1: Crystallized chimera construct (mhv1cc). (a) Sequence alignment between mhv1cc and other VSDs. These sequences (mhv1cc, Kv1.2 Kv2.1; shaker family voltage gated potassium channel
FW 1 CDR 1 FW 2 CDR 2
 Supplementary Figure 1 Supplementary Figure 1: Interface of the E9:Fas structure. The two interfaces formed by V H and V L of E9 with Fas are shown in stereo. The Fas receptor is represented as a surface
Supplementary Figure 1 Supplementary Figure 1: Interface of the E9:Fas structure. The two interfaces formed by V H and V L of E9 with Fas are shown in stereo. The Fas receptor is represented as a surface
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10955 Supplementary Figures Supplementary Figure 1. Electron-density maps and crystallographic dimer structures of the motor domain. (a f) Stereo views of the final electron-density maps
doi:10.1038/nature10955 Supplementary Figures Supplementary Figure 1. Electron-density maps and crystallographic dimer structures of the motor domain. (a f) Stereo views of the final electron-density maps
Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate
 and IDH2 (bottom) with isocitrate") SUPPLEMENTARY FIGURE LEGENDS Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate dehydrogenase from Escherichia coli [ICD, pdb 1PB1, Mesecar, A. D., and Koshland,
SUPPLEMENTARY FIGURE LEGENDS Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate dehydrogenase from Escherichia coli [ICD, pdb 1PB1, Mesecar, A. D., and Koshland,
Supplementary Figure 1. Stability constants of metal monohydroxides. The log K values are summarized according to the atomic number of each element
 Supplementary Figure 1. Stability constants of metal monohydroxides. The log K values are summarized according to the atomic number of each element as determined in a previous study 1. The log K value
Supplementary Figure 1. Stability constants of metal monohydroxides. The log K values are summarized according to the atomic number of each element as determined in a previous study 1. The log K value
Structural insights into bacterial flagellar hooks similarities and specificities
 Supplementary information Structural insights into bacterial flagellar hooks similarities and specificities Young-Ho Yoon, Clive S. Barker, Paula V. Bulieris, Hideyuki Matsunami, Fadel A. Samatey* Affiliation:
Supplementary information Structural insights into bacterial flagellar hooks similarities and specificities Young-Ho Yoon, Clive S. Barker, Paula V. Bulieris, Hideyuki Matsunami, Fadel A. Samatey* Affiliation:
Acta Crystallographica Section F
 Supporting information Acta Crystallographica Section F Volume 70 (2014) Supporting information for article: Chemical conversion of cisplatin and carboplatin with histidine in a model protein crystallised
Supporting information Acta Crystallographica Section F Volume 70 (2014) Supporting information for article: Chemical conversion of cisplatin and carboplatin with histidine in a model protein crystallised
Structural basis for catalytically restrictive dynamics of a high-energy enzyme state
 Supplementary Material Structural basis for catalytically restrictive dynamics of a high-energy enzyme state Michael Kovermann, Jörgen Ådén, Christin Grundström, A. Elisabeth Sauer-Eriksson, Uwe H. Sauer
Supplementary Material Structural basis for catalytically restrictive dynamics of a high-energy enzyme state Michael Kovermann, Jörgen Ådén, Christin Grundström, A. Elisabeth Sauer-Eriksson, Uwe H. Sauer
Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a
 To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a") Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a series of tmfret-pairs comprised of single cysteine mutants
Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a series of tmfret-pairs comprised of single cysteine mutants
Supplementary Figure 1 Structure of the Orai channel. (a) The hexameric Drosophila Orai channel structure derived from crystallography 1 comprises
 The hexameric Drosophila Orai channel structure derived from crystallography 1 comprises") Supplementary Figure 1 Structure of the Orai channel. (a) The hexameric Drosophila Orai channel structure derived from crystallography 1 comprises six Orai subunits, each with identical amino acid sequences
Supplementary Figure 1 Structure of the Orai channel. (a) The hexameric Drosophila Orai channel structure derived from crystallography 1 comprises six Orai subunits, each with identical amino acid sequences
Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets
 Supporting information Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets Wan-Na Chen, Christoph Nitsche, Kala Bharath Pilla, Bim Graham, Thomas
Supporting information Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets Wan-Na Chen, Christoph Nitsche, Kala Bharath Pilla, Bim Graham, Thomas
The structure of vanadium nitrogenase reveals an unusual bridging ligand
 SUPPLEMENTARY INFORMATION The structure of vanadium nitrogenase reveals an unusual bridging ligand Daniel Sippel and Oliver Einsle Lehrstuhl Biochemie, Institut für Biochemie, Albert-Ludwigs-Universität
SUPPLEMENTARY INFORMATION The structure of vanadium nitrogenase reveals an unusual bridging ligand Daniel Sippel and Oliver Einsle Lehrstuhl Biochemie, Institut für Biochemie, Albert-Ludwigs-Universität
A conserved P-loop anchor limits the structural dynamics that mediate. nucleotide dissociation in EF-Tu.
 Supplemental Material for A conserved P-loop anchor limits the structural dynamics that mediate nucleotide dissociation in EF-Tu. Evan Mercier 1,2, Dylan Girodat 1, and Hans-Joachim Wieden 1 * 1 Alberta
Supplemental Material for A conserved P-loop anchor limits the structural dynamics that mediate nucleotide dissociation in EF-Tu. Evan Mercier 1,2, Dylan Girodat 1, and Hans-Joachim Wieden 1 * 1 Alberta
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature12242 C. thermophilum 666 RPAVLDNVYIRPALE-GKRVPGKVEIHQNGIRYQSPLSTTQRVDVLFSNIRHLFFQPCQN S. pombe 659 RPAHINDVYVRPAID-GKRLPGFIEIHQNGIRYQSPLRSDSHIDLLFSNMKHLFFQPCEG
SUPPLEMENTARY INFORMATION doi:10.1038/nature12242 C. thermophilum 666 RPAVLDNVYIRPALE-GKRVPGKVEIHQNGIRYQSPLSTTQRVDVLFSNIRHLFFQPCQN S. pombe 659 RPAHINDVYVRPAID-GKRLPGFIEIHQNGIRYQSPLRSDSHIDLLFSNMKHLFFQPCEG
Experimental and Computational Mutagenesis to Investigate the. Positioning of a General Base within an Enzyme Active Site
 Experimental and Computational Mutagenesis to Investigate the Positioning of a General Base within an Enzyme Active Site Jason P. Schwans, Philip Hanoian, Benjamin J. Lengerich, Fanny Sunden, Ana Gonzalez
Experimental and Computational Mutagenesis to Investigate the Positioning of a General Base within an Enzyme Active Site Jason P. Schwans, Philip Hanoian, Benjamin J. Lengerich, Fanny Sunden, Ana Gonzalez
Biophysics 490M Project
 Biophysics 490M Project Dan Han Department of Biochemistry Structure Exploration of aa 3 -type Cytochrome c Oxidase from Rhodobacter sphaeroides I. Introduction: All organisms need energy to live. They
Biophysics 490M Project Dan Han Department of Biochemistry Structure Exploration of aa 3 -type Cytochrome c Oxidase from Rhodobacter sphaeroides I. Introduction: All organisms need energy to live. They
Sunhats for plants. How plants detect dangerous ultraviolet rays
 Sunhats for plants How plants detect dangerous ultraviolet rays Anyone who has ever suffered sunburn will know about the effects of too much ultraviolet (UV) radiation, in particular UV-B (from 280-315
Sunhats for plants How plants detect dangerous ultraviolet rays Anyone who has ever suffered sunburn will know about the effects of too much ultraviolet (UV) radiation, in particular UV-B (from 280-315
Structure and mechanism of an intramembrane liponucleotide synthetase central for phospholipid biosynthesis
 Structure and mechanism of an intramembrane liponucleotide synthetase central for phospholipid biosynthesis Xiuying Liu 1,3, Yan Yin 1,2,3, Jinjun Wu 1 and Zhenfeng Liu 1 1 National Laboratory of Biomacromolecules,
Structure and mechanism of an intramembrane liponucleotide synthetase central for phospholipid biosynthesis Xiuying Liu 1,3, Yan Yin 1,2,3, Jinjun Wu 1 and Zhenfeng Liu 1 1 National Laboratory of Biomacromolecules,
Supplementary Materials for
 www.advances.sciencemag.org/cgi/content/full/1/7/e1500263/dc1 Supplementary Materials for Newton s cradle proton relay with amide imidic acid tautomerization in inverting cellulase visualized by neutron
www.advances.sciencemag.org/cgi/content/full/1/7/e1500263/dc1 Supplementary Materials for Newton s cradle proton relay with amide imidic acid tautomerization in inverting cellulase visualized by neutron