Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate
|
|
|
- Juniper Gilmore
- 5 years ago
- Views:
Transcription
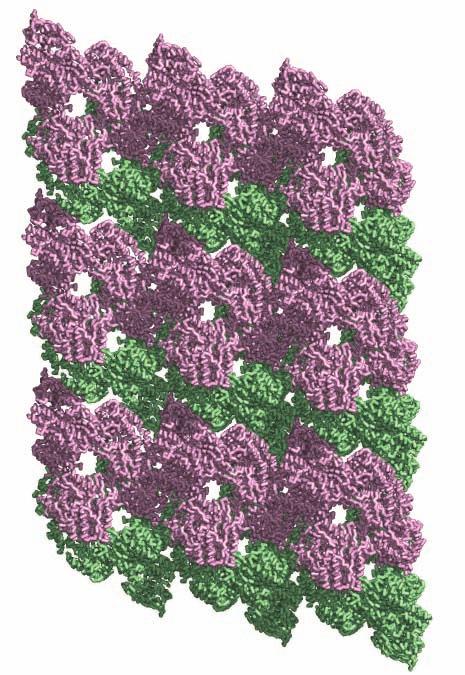
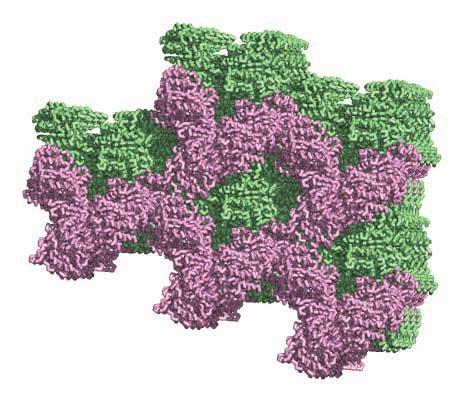
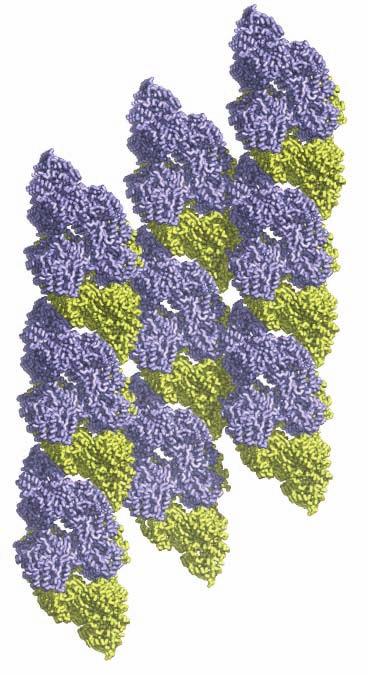
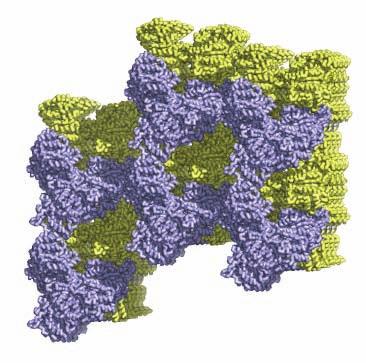
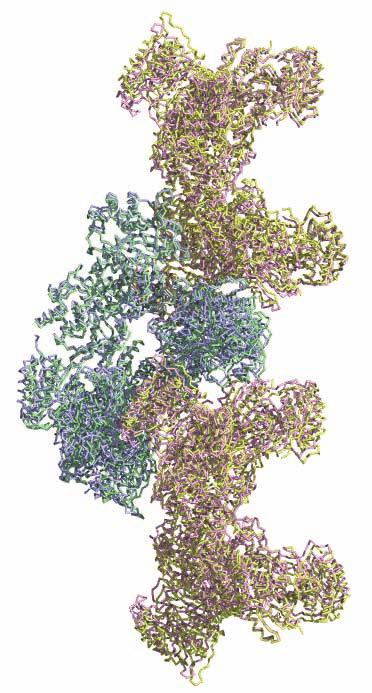
1 SUPPLEMENTARY FIGURE LEGENDS Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate dehydrogenase from Escherichia coli [ICD, pdb 1PB1, Mesecar, A. D., and Koshland, D. E., Jr. (2000) Nature 403, ], homoisocitrate dehydrogenase from Thermus thermophilus (HICD, pdb 1X0L) (37) and isopropyl malate dehydrogenase from Sulfolobus tokodaii (IMDH, pdb 1WPW) (Hirose et al. unpublished). The alignment of these sequences obtained from a BLAST search was modified based on the structural data and prepared using ESPript [Gouet, P., Robert, X., and Courcelle, E. (2003) Nucleic Acids Res. 31, ]. Red and green arrows/helices represent secondary structural elements for IDH1 and IDH2, respectively. Identical residues are boxed and highlighted in blue. Similar residues are boxed and highlighted in yellow. Indicated are the residues that form the catalytic sites (C), regulatory sites (R in red), and polar interactions in the clasp β-barrel (I) of yeast IDH. Residues that participate in AMP binding in the regulatory site are shown as light blue (R) symbols. Supplementary Figure 2: Crystal packing of the IDH heterooctamer in the citrate-bound C2 and unliganded P1 crystal forms. a) Columns of IDH heterooctamers are shown running vertically in the plane of the page. Consecutive heterooctamers in each column are shown in different colors. In the left image, three adjacent columns of citrate-bound IDH in space group C2 are shown with heterooctamers colored pink and green. In the right image, three adjacent columns of unliganded IDH in space group P1 are shown with heterooctamers colored blue and yellow. The columns are in approximately the same orientation such that pink and blue heterooctamers and green and yellow heterooctamers are in equivalent positions in each column. Note that the columns are slightly translated with respect to each other in the two crystal forms causing them to engage in different protein-protein interactions horizontally in the plane of the page. b) Columns of IDH heterooctamers looking end-on. The view is 1
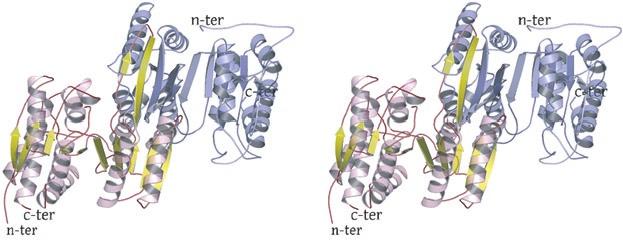
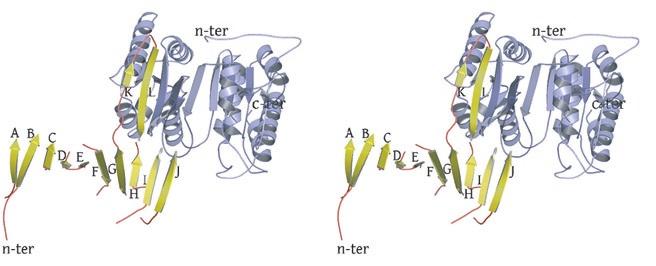
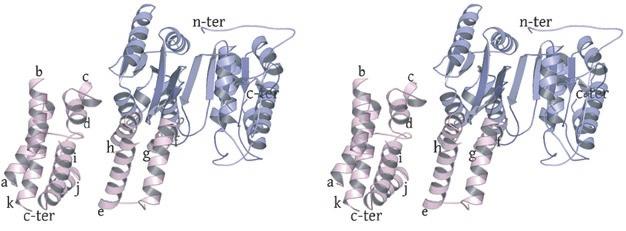
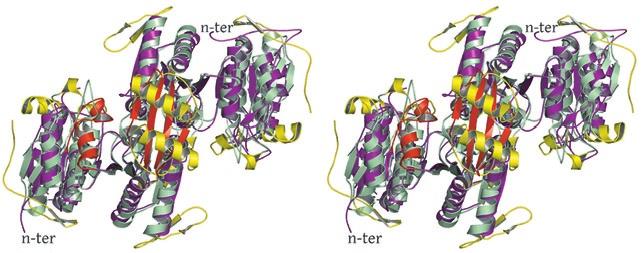
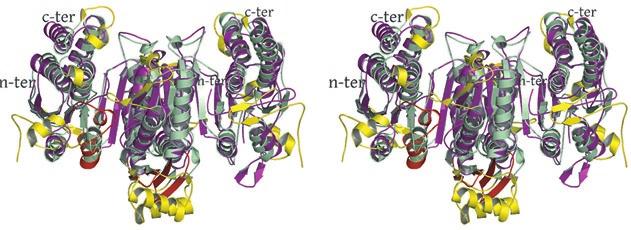
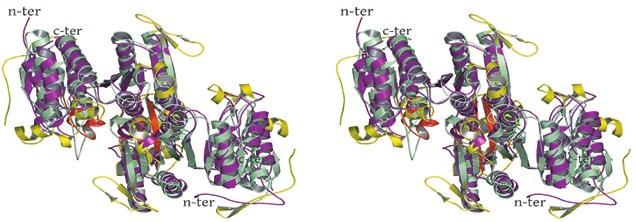
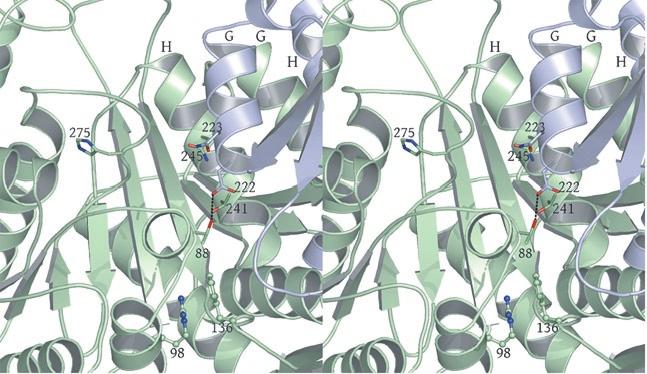
2 rotated 90 o relative to panel a). The ends of five columns are shown for the citrate-bound IDH structure in space group C2 on the left and the ends of six columns are shown for the unliganded IDH structure in space group P1 on the right. The translation of columns relative to each other in the two crystal forms is evident. c) Superposition of three consecutive heterooctamers in a column coming from each crystal form. The color coding is as in panels a) and b). Supplementary Figure 3: Secondary structural elements in yeast mitochondrial NAD + -specific IDH. a) The yeast IDH1/IDH2 heterodimer. The regulatory IDH1 subunit is shown at the left, with α-helices colored light pink and β-strands colored yellow. The catalytic IDH2 subunit is shown at the right and is colored slate blue. The homologous IDH1 and IDH2 subunits of the heterodimer each contain 12 β- strands and 11 α-helices related by an approximate 180 o axis of rotation. b) The β-strands in the IDH1/IDH2 heterodimer. Strands A-H from each subunit form a continuous 10-standed β-sheet. In each subunit, the outermost β-strand is designated "A" and the innermost β-strand is designated "J". β-strands K and L are offset from the 10-stranded β-sheet in each subunit and associate to form a 4-stranded β- sheet centered on the pseudo-two-fold axis of rotation. c) The α-helices in the IDH1/IDH2 heterodimer. Helices a-k are labeled sequentially in the order they appear in the primary sequence of each subunit. Supplementary Figure 4: Structural alignment of the unliganded yeast NAD + -specific IDH heterodimer with the homologous E. coli NADP + -specific IDH homodimer (pdb code 1PB1) (49) in three orthogonal views. a) Superposition of yeast IDH (purple) with E. coli IDH (light green). The view is along the yeast IDH pseudo-molecular two-fold axis of rotation, which corresponds to the true two-fold axis of rotation in the E. coli homodimer. Structural elements differing in the E. coli and yeast IDH structures are highlighted by showing the E. coli elements in yellow. The "clasp" β-strands that form one-half of the yeast heterotetrameric interface are in the very center of the image and are shown in red. Residues 2
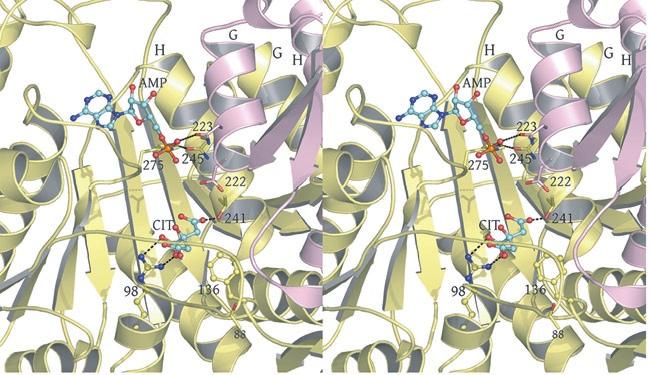
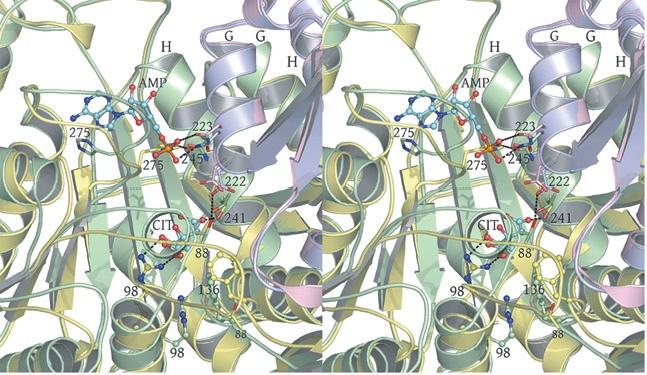
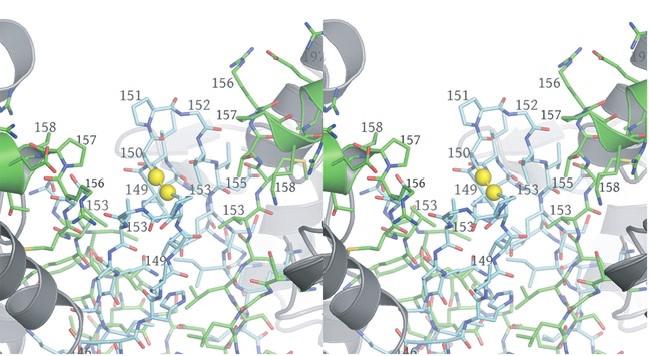
3 in IDH1 that undergo a conformational change upon isocitrate binding site are also shown in red. The corresponding residues in the E. coli structure form a loop that is similar to the conformation of these residues in the liganded yeast IDH structure. b) This view is rotated 90 o relative to panel a) around an axis that runs horizontally in plane of the page. Note that the E. coli IDH structure contains two α- helices (yellow) that have been lost in yeast IDH to be replaced by β-strands K and L (red) that comprise the yeast heterotetrameric interface. c) This view is rotated 90 o relative to panel b) and 180 o relative to panel a) around an axis that runs horizontally in the plane of the page. The four-helix bundle at the heart of the heterodimeric yeast IDH and homodimeric E. coli interfaces is located at the very center of the image. Supplementary Figure 5: Participation of residues of the 4-helix bundle in allosteric communication in the IDH heterodimer. a) The ligand-bound yeast IDH structure. IDH1 is shown in yellow and IDH2 is shown in pink. Upon binding of (iso)citrate and AMP, Thr241 and Asn245 of IDH1 helix h make hydrogen bonds to these ligands. Asn223 from helix g of IDH2 makes a hydrogen bond to the phosphate moiety of AMP. The binding of citrate and AMP in the regulatory site therefore is likely communicated through the four-helix bundle at the heterodimer interface to the active site. Previous kinetic and mutagenesis studies in which apolar residues in the core of the 4-helix bundle were altered slightly resulted in yeast IDH proteins with altered allosteric properties (35). b) The unliganded yeast IDH structure in the same orientation as panel a). IDH1 is shown in pale green and IDH2 is shown in slate blue. c) Superposition of the liganded and unliganded IDH structures shown in the same orientation as in panels a) and b). Supplementary Figure 6: The local environment near IDH2 Cys150 in the unliganded yeast IDH structure. a) IDH2 Cys150 residues form a disulfide bond at the heterotetrameric interface. IDH2 3
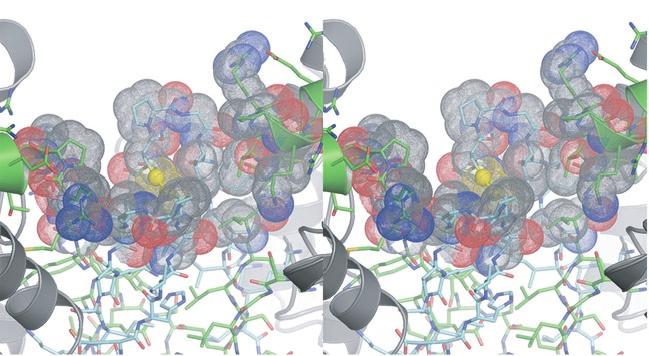
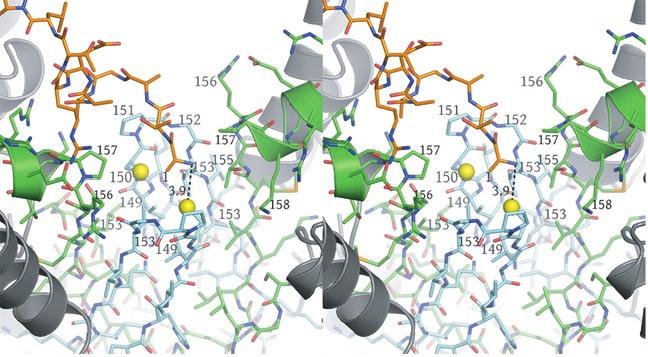
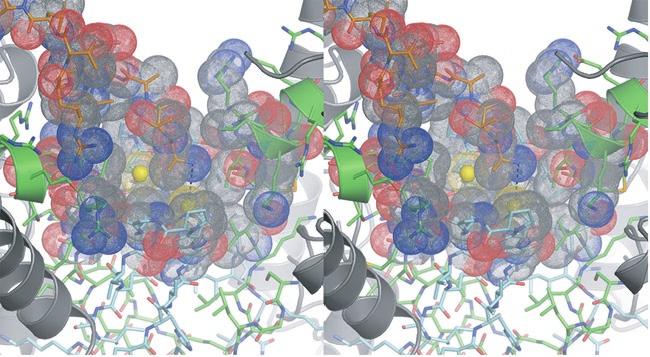
4 residues forming the clasp β-barrel are shown in cyan. IDH1 residues are shown in green. The N- termini of IDH1 helix e are shown as ribbons with side chains represented as sticks. The Sγ atoms of IDH2 Cys150 are "sunken" into the clasp β-barrel and are surrounded by apolar side chains. These include IDH2 Val149, Pro151, and Val153 and IDH1 Val153 and Pro157. Each IDH2 Cys150 Sγ atom is positioned on the long axis of helix e coming from its cognate IDH1 subunit in the heterodimer. The combination of the positive charge coming from basic residues at the N-terminus of the uncapped helix e coupled with the generally apolar environment is predicted to substantially lower the pka values of these Cys150 residues. b) Surface representation of the image shown in panel a). The colors reflect the type of atom. Nitrogen atoms are blue, oxygen atoms are red, sulfur atoms are yellow and carbon atoms are gray. Note that each Sγ atom of Cys150 is in contact with carbon atoms only. Supplementary Figure 7: The local environment near IDH2 Cys150 in the citrate-bound yeast IDH structure. The view and the color scheme is the same as in Supplementary Figure 6 except that the N- terminus of IDH1 chain C from the cognate heterotetramer of the heterooctamer is shown as orange sticks. a) The disulfide bond observed between IDH2 Cys150 residues in the unliganded structure is reduced and the positively charged primary amino group of IDH1 chain C resides 3.9 Å from one of the Cys150 Sγ atoms. The proximity of helix e from the cognate IDH1 subunit and the IDH1 chain C amino terminus are predicted to stabilize the thiolate form of the Cys150 side chain. b) Surface representation of the image shown in panel a). The colors reflect the type of atom. Nitrogen atoms are blue, oxygen atoms are red, sulfur atoms are yellow and carbon atoms are gray. 4
5 Supplementary Figure IDH1 A T A A Q A E.... R T L P K K Y G G R F T V T L I P G D G V G K E I T D S V R T I F E A E. N I P I D W E T I N.... I K Q T D H.. K E.... ICD... M E S K V V V P A Q G K K I T L Q N G K L N V P E N P I I P Y I E G D G I G V D V T P A M L K V V D A A V E K A Y K G E R K I S W M E I Y T G E K S T Q V Y G Q D V W.... HICD A Y R I C L I E G D G I G H E V I P A A R R V L E A T. G L P L E F V E A E.... A G W E.... T F E R R G IMDH G F T V A L I Q G D G I G P E I V S K S K R I L A K I N E L Y S... L P I E Y I E V E.... A G D R.... A L A R Y G IDH2 A T V K Q P S I G R Y T G K P N P S T G K Y T V S F I E G D G I G P E I S K S V K K I F S A A. N V P I E W E S C D.... V S P I F V N G R 9 0 R R R R R I I IDH1.. G V Y E. A V E S L K R N K I G L K G L W H T P A D Q T G H. G S L N V A L R K Q L D I Y A N V A L F K S L K G V K T. R. I P D. I D L I V I R E N T E G E F S G L E H E S V ICD.. L P A E. T L D L I R E Y R V A I K G P L T T P. V G G G I. R S L N V A L R Q E L D L Y I C L R P V R Y Y Q G T P S P V K H P E L T D M V I F R E N S E D I Y A G I E W K A D HICD. T S V P E E T V E K I L S C H A T L F G A A T S P T R K V P G F F G A I R Y L R R R L D L Y A N V R P A K S R P. V P G. S. R P G. V D L V I V R E N T E G L Y V E Q E R R Y. IMDH. E A L P K D S L K I I D K A D I I L K G P V G E S A A D V V V K L R Q I Y D M Y A N I R P A K S I P. G I D. T K Y G N. V D I L I V R E N T E D L Y K G F E H I V S IDH2 L T T I P D P A V Q S I T K N L V A L K G P L A T P. I G K G H. R S L N L T L R K T F G L F A N V R P A K S I E G F K T. T. Y E N. V D L V L I R E N T E G E Y S G I E H I V C C C C C C C C C I I I C C IDH P G V V E S L K V M T R P K T E R I A R F A F D F A K K Y N R K S V T A V H K A N I M K L G D G L F R N I I T E I G Q K E Y..... ICD S A D A E K V I K F L R E E M G V K K I R F P E H C G I G I K P C S E E G T K R L V R A A I E Y A I A N D R D S V T L V H K G N I M K F T E G A F K D W G Y Q L A R E E F G G E L I HICD L D V A I A D A V I S K K A S E R I G R A A L R I A E G R P R K T L H I A H K A N V L P L T Q G L F L D T V K E V A. K D F..... IMDH D G V A V G M K I I T R F A S E R I A K V G L N F A L R R. R K K V T C V H K A N V M R I T D G L F A E A C R S V L. K G K..... IDH P G V V Q S I K L I T R D A S E R V I R Y A F E Y A R A I G R P R V I V V H K S T I Q R L A D G L F V N V A K E L S. K E Y..... I R C R R R R R R R IDH P D I D V S S I I V D N A S M Q A V A K P H Q F.. D V L V T P S M Y G T I L G N I G A A L I. G G P G L V A G A N F G R D Y A V F E P G S R H V G L D ICD D G G P W L K V K N P N T G K E I V I K D V I A D A F L Q Q I L L R P A E Y.. D V I A C M N L N G D Y I S D A L A A Q V. G G I G I A P G A N I G D E C A L F E A. T H G T A P K HICD P L V N V Q D I I V D N C A M Q L V M R P E R F.. D V I V T T N L L G D I L S D L A A G L V. G G L G L A P S G N I G D T T A V F E P.. V H G S A P IMDH V E Y S E M Y V D A A A A N L V R N P Q M F.. D V I V T E N V Y G D I L S D E A S Q I A. G S L G I A P S A N I G D K K A L F E P.. V H G A A F IDH P D L T L E T E L I D N S V L K V V T N P S A Y T D A V S V C P N L Y G D I L S D L N S G L S A G S L G L T P S A N I G H K I S I F E A.. V H G S A P R C C C C R R IDH1 I K G. Q N V A N P T A M I L S S T L M L N H L G L N E Y A T R I S. K A V H E T I A E. G K H T T R D I G..... G. S S S T T D F T N E I I N K L S T M ICD Y A G. Q D K V N P G S I I L S A E M M L R H M G W T E A A D L I V. K G M E G A I N A. K. T V T Y D F E R L M D G A K L L K C S E F G D A I I E N M... HICD D I A G K G I A N P T A A I L S A A M M L D Y L G E K E A A K R V E K. A V D L V L E R. G. P R T P D L G..... G. D A T T E A F T E A V V E A L K S L IMDH D I A G K N I G N P T A F L L S V S M M Y E R M Y E L S N D D R Y I K A S R A L E N. A I Y L V Y K E R K. A L T P D V G..... G. N A T T D D L I N E I Y N K L G.. IDH2 D I A G Q D K A N P T A L L L S S V M M L N H M G L T N H A D Q I Q N A V L S T I A S G. P E N R T G D L A..... G. T A T T S S F T E A V I K R L
6 Supplementary Figure 2 a) b) c)
7 Supplementary Figure 3 A B C
8 Supplementary Figure 4 A B C
9 Supplementary Figure 5 A B C
10 Supplementary Figure 6 A B
11 Supplementary Figure 7 A B
SUPPLEMENTARY FIGURES
 SUPPLEMENTARY FIGURES Supplementary Figure 1 Protein sequence alignment of Vibrionaceae with either a 40-residue insertion or a 44-residue insertion. Identical residues are indicated by red background.
SUPPLEMENTARY FIGURES Supplementary Figure 1 Protein sequence alignment of Vibrionaceae with either a 40-residue insertion or a 44-residue insertion. Identical residues are indicated by red background.
SUPPLEMENTARY INFORMATION
 Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Nitrogenase MoFe protein from Clostridium pasteurianum at 1.08 Å resolution: comparison with the Azotobacter vinelandii MoFe protein
 Acta Cryst. (2015). D71, 274-282, doi:10.1107/s1399004714025243 Supporting information Volume 71 (2015) Supporting information for article: Nitrogenase MoFe protein from Clostridium pasteurianum at 1.08
Acta Cryst. (2015). D71, 274-282, doi:10.1107/s1399004714025243 Supporting information Volume 71 (2015) Supporting information for article: Nitrogenase MoFe protein from Clostridium pasteurianum at 1.08
LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor
 LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor Note: Adequate space is given for each answer. Questions that require a brief explanation should
LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor Note: Adequate space is given for each answer. Questions that require a brief explanation should
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
Model Mélange. Physical Models of Peptides and Proteins
 Model Mélange Physical Models of Peptides and Proteins In the Model Mélange activity, you will visit four different stations each featuring a variety of different physical models of peptides or proteins.
Model Mélange Physical Models of Peptides and Proteins In the Model Mélange activity, you will visit four different stations each featuring a variety of different physical models of peptides or proteins.
SUPPLEMENTARY INFORMATION
 Supplementary materials Figure S1 Fusion protein of Sulfolobus solfataricus SRP54 and a signal peptide. a, Expression vector for the fusion protein. The signal peptide of yeast dipeptidyl aminopeptidase
Supplementary materials Figure S1 Fusion protein of Sulfolobus solfataricus SRP54 and a signal peptide. a, Expression vector for the fusion protein. The signal peptide of yeast dipeptidyl aminopeptidase
Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase Cwc27
 Acta Cryst. (2014). D70, doi:10.1107/s1399004714021695 Supporting information Volume 70 (2014) Supporting information for article: Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase
Acta Cryst. (2014). D70, doi:10.1107/s1399004714021695 Supporting information Volume 70 (2014) Supporting information for article: Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10955 Supplementary Figures Supplementary Figure 1. Electron-density maps and crystallographic dimer structures of the motor domain. (a f) Stereo views of the final electron-density maps
doi:10.1038/nature10955 Supplementary Figures Supplementary Figure 1. Electron-density maps and crystallographic dimer structures of the motor domain. (a f) Stereo views of the final electron-density maps
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Cryo-EM structure and model of the C. thermophilum 90S preribosome. a, Gold standard FSC curve showing the average resolution of the 90S preribosome masked and unmasked (left). FSC
Supplementary Figure 1 Cryo-EM structure and model of the C. thermophilum 90S preribosome. a, Gold standard FSC curve showing the average resolution of the 90S preribosome masked and unmasked (left). FSC
Table 1. Crystallographic data collection, phasing and refinement statistics. Native Hg soaked Mn soaked 1 Mn soaked 2
 Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits. AhpC and AhpF from Escherichia coli
 Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits AhpC and AhpF from Escherichia coli Phat Vinh Dip 1,#, Neelagandan Kamariah 2,#, Malathy Sony Subramanian Manimekalai
Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits AhpC and AhpF from Escherichia coli Phat Vinh Dip 1,#, Neelagandan Kamariah 2,#, Malathy Sony Subramanian Manimekalai
SUPPLEMENTARY INFORMATION
 Dph2 SeMet (iron-free) # Dph2 (iron-free) Dph2-[4Fe-4S] Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 58.26, 82.08, 160.42 58.74, 81.87, 160.01 55.70, 80.53,
Dph2 SeMet (iron-free) # Dph2 (iron-free) Dph2-[4Fe-4S] Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 58.26, 82.08, 160.42 58.74, 81.87, 160.01 55.70, 80.53,
SUPPLEMENTARY INFORMATION
 Fig. 1 Influences of crystal lattice contacts on Pol η structures. a. The dominant lattice contact between two hpol η molecules (silver and gold) in the type 1 crystals. b. A close-up view of the hydrophobic
Fig. 1 Influences of crystal lattice contacts on Pol η structures. a. The dominant lattice contact between two hpol η molecules (silver and gold) in the type 1 crystals. b. A close-up view of the hydrophobic
SUPPLEMENTARY INFORMATION
 Figure S1. Secondary structure of CAP (in the camp 2 -bound state) 10. α-helices are shown as cylinders and β- strands as arrows. Labeling of secondary structure is indicated. CDB, DBD and the hinge are
Figure S1. Secondary structure of CAP (in the camp 2 -bound state) 10. α-helices are shown as cylinders and β- strands as arrows. Labeling of secondary structure is indicated. CDB, DBD and the hinge are
Supplementary figure 1. Comparison of unbound ogm-csf and ogm-csf as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine
 Supplementary figure 1. Comparison of unbound and as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine GM-CSF (slate) with bound GM-CSF in the GIF:GM-CSF complex (GIF: green,
Supplementary figure 1. Comparison of unbound and as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine GM-CSF (slate) with bound GM-CSF in the GIF:GM-CSF complex (GIF: green,
SUPPLEMENTARY INFORMATION
 Supplementary Table 1: Amplitudes of three current levels. Level 0 (pa) Level 1 (pa) Level 2 (pa) TrkA- TrkH WT 200 K 0.01 ± 0.01 9.5 ± 0.01 18.7 ± 0.03 200 Na * 0.001 ± 0.01 3.9 ± 0.01 12.5 ± 0.03 200
Supplementary Table 1: Amplitudes of three current levels. Level 0 (pa) Level 1 (pa) Level 2 (pa) TrkA- TrkH WT 200 K 0.01 ± 0.01 9.5 ± 0.01 18.7 ± 0.03 200 Na * 0.001 ± 0.01 3.9 ± 0.01 12.5 ± 0.03 200
Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1.
 Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1. PDZK1 constru cts Amino acids MW [kda] KD [μm] PEPT2-CT- FITC KD [μm] NHE3-CT- FITC KD [μm] PDZK1-CT-
Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1. PDZK1 constru cts Amino acids MW [kda] KD [μm] PEPT2-CT- FITC KD [μm] NHE3-CT- FITC KD [μm] PDZK1-CT-
Supporting Information
 Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
SUPPLEMENTARY INFORMATION. doi: /nature07461
 Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
Design of a Novel Globular Protein Fold with Atomic-Level Accuracy
 Design of a Novel Globular Protein Fold with Atomic-Level Accuracy Brian Kuhlman, Gautam Dantas, Gregory C. Ireton, Gabriele Varani, Barry L. Stoddard, David Baker Presented by Kate Stafford 4 May 05 Protein
Design of a Novel Globular Protein Fold with Atomic-Level Accuracy Brian Kuhlman, Gautam Dantas, Gregory C. Ireton, Gabriele Varani, Barry L. Stoddard, David Baker Presented by Kate Stafford 4 May 05 Protein
Supplementary Figures
 1 Supplementary Figures Supplementary Figure 1 Type I FGFR1 inhibitors (a) Chemical structures of a pyrazolylaminopyrimidine inhibitor (henceforth referred to as PAPI; PDB-code of the FGFR1-PAPI complex:
1 Supplementary Figures Supplementary Figure 1 Type I FGFR1 inhibitors (a) Chemical structures of a pyrazolylaminopyrimidine inhibitor (henceforth referred to as PAPI; PDB-code of the FGFR1-PAPI complex:
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Identification of the ScDcp2 minimal region interacting with both ScDcp1 and the ScEdc3 LSm domain. Pull-down experiment of untagged ScEdc3 LSm with various ScDcp1-Dcp2-His 6 fragments.
Supplementary Figure 1 Identification of the ScDcp2 minimal region interacting with both ScDcp1 and the ScEdc3 LSm domain. Pull-down experiment of untagged ScEdc3 LSm with various ScDcp1-Dcp2-His 6 fragments.
SUPPLEMENTARY INFORMATION
 www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
1. What is an ångstrom unit, and why is it used to describe molecular structures?
 1. What is an ångstrom unit, and why is it used to describe molecular structures? The ångstrom unit is a unit of distance suitable for measuring atomic scale objects. 1 ångstrom (Å) = 1 10-10 m. The diameter
1. What is an ångstrom unit, and why is it used to describe molecular structures? The ångstrom unit is a unit of distance suitable for measuring atomic scale objects. 1 ångstrom (Å) = 1 10-10 m. The diameter
Supplementary Information
 1 Supplementary Information Figure S1 The V=0.5 Harker section of an anomalous difference Patterson map calculated using diffraction data from the NNQQNY crystal at 1.3 Å resolution. The position of the
1 Supplementary Information Figure S1 The V=0.5 Harker section of an anomalous difference Patterson map calculated using diffraction data from the NNQQNY crystal at 1.3 Å resolution. The position of the
HIV protease inhibitor. Certain level of function can be found without structure. But a structure is a key to understand the detailed mechanism.
 Proteins are linear polypeptide chains (one or more) Building blocks: 20 types of amino acids. Range from a few 10s-1000s They fold into varying three-dimensional shapes structure medicine Certain level
Proteins are linear polypeptide chains (one or more) Building blocks: 20 types of amino acids. Range from a few 10s-1000s They fold into varying three-dimensional shapes structure medicine Certain level
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11085 Supplementary Tables: Supplementary Table 1. Summary of crystallographic and structure refinement data Structure BRIL-NOP receptor Data collection Number of crystals 23 Space group
doi:10.1038/nature11085 Supplementary Tables: Supplementary Table 1. Summary of crystallographic and structure refinement data Structure BRIL-NOP receptor Data collection Number of crystals 23 Space group
Pymol Practial Guide
 Pymol Practial Guide Pymol is a powerful visualizor very convenient to work with protein molecules. Its interface may seem complex at first, but you will see that with a little practice is simple and powerful
Pymol Practial Guide Pymol is a powerful visualizor very convenient to work with protein molecules. Its interface may seem complex at first, but you will see that with a little practice is simple and powerful
Section III - Designing Models for 3D Printing
 Section III - Designing Models for 3D Printing In this section of the Jmol Training Guide, you will become familiar with the commands needed to design a model that will be built on a 3D Printer. As you
Section III - Designing Models for 3D Printing In this section of the Jmol Training Guide, you will become familiar with the commands needed to design a model that will be built on a 3D Printer. As you
Basics of protein structure
 Today: 1. Projects a. Requirements: i. Critical review of one paper ii. At least one computational result b. Noon, Dec. 3 rd written report and oral presentation are due; submit via email to bphys101@fas.harvard.edu
Today: 1. Projects a. Requirements: i. Critical review of one paper ii. At least one computational result b. Noon, Dec. 3 rd written report and oral presentation are due; submit via email to bphys101@fas.harvard.edu
Secondary and sidechain structures
 Lecture 2 Secondary and sidechain structures James Chou BCMP201 Spring 2008 Images from Petsko & Ringe, Protein Structure and Function. Branden & Tooze, Introduction to Protein Structure. Richardson, J.
Lecture 2 Secondary and sidechain structures James Chou BCMP201 Spring 2008 Images from Petsko & Ringe, Protein Structure and Function. Branden & Tooze, Introduction to Protein Structure. Richardson, J.
Diphthamide biosynthesis requires a radical iron-sulfur enzyme. Pennsylvania State University, University Park, Pennsylvania 16802, USA
 Diphthamide biosynthesis requires a radical iron-sulfur enzyme Yang Zhang, 1,4 Xuling Zhu, 1,4 Andrew T. Torelli, 1 Michael Lee, 2 Boris Dzikovski, 1 Rachel Koralewski, 1 Eileen Wang, 1 Jack Freed, 1 Carsten
Diphthamide biosynthesis requires a radical iron-sulfur enzyme Yang Zhang, 1,4 Xuling Zhu, 1,4 Andrew T. Torelli, 1 Michael Lee, 2 Boris Dzikovski, 1 Rachel Koralewski, 1 Eileen Wang, 1 Jack Freed, 1 Carsten
Protein Structure. W. M. Grogan, Ph.D. OBJECTIVES
 Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Introduction to Comparative Protein Modeling. Chapter 4 Part I
 Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Chemical structure of LPS and LPS biogenesis in Gram-negative bacteria. a. Chemical structure of LPS. LPS molecule consists of Lipid A, core oligosaccharide and O-antigen. The polar
Supplementary Figure 1 Chemical structure of LPS and LPS biogenesis in Gram-negative bacteria. a. Chemical structure of LPS. LPS molecule consists of Lipid A, core oligosaccharide and O-antigen. The polar
Supplemental Data SUPPLEMENTAL FIGURES
 Supplemental Data CRYSTAL STRUCTURE OF THE MG.ADP-INHIBITED STATE OF THE YEAST F 1 C 10 ATP SYNTHASE Alain Dautant*, Jean Velours and Marie-France Giraud* From Université Bordeaux 2, CNRS; Institut de
Supplemental Data CRYSTAL STRUCTURE OF THE MG.ADP-INHIBITED STATE OF THE YEAST F 1 C 10 ATP SYNTHASE Alain Dautant*, Jean Velours and Marie-France Giraud* From Université Bordeaux 2, CNRS; Institut de
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis Sergio de Cima, Luis M. Polo, Carmen Díez-Fernández, Ana I. Martínez, Javier
SUPPLEMENTARY INFORMATION Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis Sergio de Cima, Luis M. Polo, Carmen Díez-Fernández, Ana I. Martínez, Javier
Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions. Implicated in Dimerization and Autoinhibition
 Reveals Regions. Implicated in Dimerization and Autoinhibition") JBC Papers in Press. Published on November 1, 2000 as Manuscript M006502200 Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions Implicated in Dimerization and Autoinhibition 1 Copyright
JBC Papers in Press. Published on November 1, 2000 as Manuscript M006502200 Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions Implicated in Dimerization and Autoinhibition 1 Copyright
Thermophilic organism
 Thermophilic organism Thermophiles are organisms that grow and thrive at temperatures (60 80 C) that are often too high for mesophiles most thermophiles are Archaea Some organisms grow at even higher temperatures
Thermophilic organism Thermophiles are organisms that grow and thrive at temperatures (60 80 C) that are often too high for mesophiles most thermophiles are Archaea Some organisms grow at even higher temperatures
Structural insights into Aspergillus fumigatus lectin specificity - AFL binding sites are functionally non-equivalent
 Acta Cryst. (2015). D71, doi:10.1107/s1399004714026595 Supporting information Volume 71 (2015) Supporting information for article: Structural insights into Aspergillus fumigatus lectin specificity - AFL
Acta Cryst. (2015). D71, doi:10.1107/s1399004714026595 Supporting information Volume 71 (2015) Supporting information for article: Structural insights into Aspergillus fumigatus lectin specificity - AFL
Physiochemical Properties of Residues
 Physiochemical Properties of Residues Various Sources C N Cα R Slide 1 Conformational Propensities Conformational Propensity is the frequency in which a residue adopts a given conformation (in a polypeptide)
Physiochemical Properties of Residues Various Sources C N Cα R Slide 1 Conformational Propensities Conformational Propensity is the frequency in which a residue adopts a given conformation (in a polypeptide)
Cks1 CDK1 CDK1 CDK1 CKS1. are ice- lobe. conserved. conserved
 Cks1 d CKS1 Supplementary Figure 1 The -Cks1 crystal lattice. (a) Schematic of the - Cks1 crystal lattice. -Cks1 crystallizes in a lattice that contains c 4 copies of the t - Cks1 dimer in the crystallographic
Cks1 d CKS1 Supplementary Figure 1 The -Cks1 crystal lattice. (a) Schematic of the - Cks1 crystal lattice. -Cks1 crystallizes in a lattice that contains c 4 copies of the t - Cks1 dimer in the crystallographic
Examples of Protein Modeling. Protein Modeling. Primary Structure. Protein Structure Description. Protein Sequence Sources. Importing Sequences to MOE
 Examples of Protein Modeling Protein Modeling Visualization Examination of an experimental structure to gain insight about a research question Dynamics To examine the dynamics of protein structures To
Examples of Protein Modeling Protein Modeling Visualization Examination of an experimental structure to gain insight about a research question Dynamics To examine the dynamics of protein structures To
SUPPLEMENTARY INFORMATION
 Supplementary Table 1: Data collection, phasing and refinement statistics ChbC/Ta 6 Br 12 Native ChbC Data collection Space group P4 3 2 1 2 P4 3 2 1 2 Cell dimensions a, c (Å) 132.75, 453.57 132.81, 452.95
Supplementary Table 1: Data collection, phasing and refinement statistics ChbC/Ta 6 Br 12 Native ChbC Data collection Space group P4 3 2 1 2 P4 3 2 1 2 Cell dimensions a, c (Å) 132.75, 453.57 132.81, 452.95
SUPPLEMENTARY INFORMATION
 SUPPLMTARY IFORMATIO a doi:10.108/nature10402 b 100 nm 100 nm c SAXS Model d ulers assigned to reference- Back-projected free class averages class averages Refinement against single particles Reconstructed
SUPPLMTARY IFORMATIO a doi:10.108/nature10402 b 100 nm 100 nm c SAXS Model d ulers assigned to reference- Back-projected free class averages class averages Refinement against single particles Reconstructed
RNA Polymerase I Contains a TFIIF-Related DNA-Binding Subcomplex
 Molecular Cell, Volume 39 Supplemental Information RNA Polymerase I Contains a TFIIFRelated DNABinding Subcomplex Sebastian R. Geiger, Kristina Lorenzen, Amelie Schreieck, Patrizia Hanecker, Dirk Kostrewa,
Molecular Cell, Volume 39 Supplemental Information RNA Polymerase I Contains a TFIIFRelated DNABinding Subcomplex Sebastian R. Geiger, Kristina Lorenzen, Amelie Schreieck, Patrizia Hanecker, Dirk Kostrewa,
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11744 Supplementary Table 1. Crystallographic data collection and refinement statistics. Wild-type Se-Met-BcsA-B SmCl 3 -soaked EMTS-soaked Data collection Space
SUPPLEMENTARY INFORMATION doi:10.1038/nature11744 Supplementary Table 1. Crystallographic data collection and refinement statistics. Wild-type Se-Met-BcsA-B SmCl 3 -soaked EMTS-soaked Data collection Space
Nature Structural and Molecular Biology: doi: /nsmb.2938
 Supplementary Figure 1 Characterization of designed leucine-rich-repeat proteins. (a) Water-mediate hydrogen-bond network is frequently visible in the convex region of LRR crystal structures. Examples
Supplementary Figure 1 Characterization of designed leucine-rich-repeat proteins. (a) Water-mediate hydrogen-bond network is frequently visible in the convex region of LRR crystal structures. Examples
B O C 4 H 2 O O. NOTE: The reaction proceeds with a carbonium ion stabilized on the C 1 of sugar A.
 hbcse 33 rd International Page 101 hemistry lympiad Preparatory 05/02/01 Problems d. In the hydrolysis of the glycosidic bond, the glycosidic bridge oxygen goes with 4 of the sugar B. n cleavage, 18 from
hbcse 33 rd International Page 101 hemistry lympiad Preparatory 05/02/01 Problems d. In the hydrolysis of the glycosidic bond, the glycosidic bridge oxygen goes with 4 of the sugar B. n cleavage, 18 from
Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R)
") Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R) Shown in cyan and green are two adjacent tetramers from the crystallographic lattice of COP, forming the only unique inter-tetramer
Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R) Shown in cyan and green are two adjacent tetramers from the crystallographic lattice of COP, forming the only unique inter-tetramer
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
Protein Data Bank Contents Guide: Atomic Coordinate Entry Format Description. Version Document Published by the wwpdb
 Protein Data Bank Contents Guide: Atomic Coordinate Entry Format Description Version 3.30 Document Published by the wwpdb This format complies with the PDB Exchange Dictionary (PDBx) http://mmcif.pdb.org/dictionaries/mmcif_pdbx.dic/index/index.html.
Protein Data Bank Contents Guide: Atomic Coordinate Entry Format Description Version 3.30 Document Published by the wwpdb This format complies with the PDB Exchange Dictionary (PDBx) http://mmcif.pdb.org/dictionaries/mmcif_pdbx.dic/index/index.html.
Bacterial protease uses distinct thermodynamic signatures for substrate recognition
 Bacterial protease uses distinct thermodynamic signatures for substrate recognition Gustavo Arruda Bezerra, Yuko Ohara-Nemoto, Irina Cornaciu, Sofiya Fedosyuk, Guillaume Hoffmann, Adam Round, José A. Márquez,
Bacterial protease uses distinct thermodynamic signatures for substrate recognition Gustavo Arruda Bezerra, Yuko Ohara-Nemoto, Irina Cornaciu, Sofiya Fedosyuk, Guillaume Hoffmann, Adam Round, José A. Márquez,
Dr. Yonca Yuzugullu PERG (Protein Engineering Research Group)
") Dr. Yonca Yuzugullu PERG (Protein Engineering Research Group) BSc, 1997 Ankara University, Turkey PhD, 2010 Middle East Technical University, Turkey Lecturer in Department of Biology Kocaeli University,
Dr. Yonca Yuzugullu PERG (Protein Engineering Research Group) BSc, 1997 Ankara University, Turkey PhD, 2010 Middle East Technical University, Turkey Lecturer in Department of Biology Kocaeli University,
SUPPLEMENTARY FIGURES. Figure S1
 SUPPLEMENTARY FIGURES Figure S1 The substrate for DH domain (2R,3R,4R,6R,7S,8S,9R)-3,7,9-trihydroxy-5-oxo-2,4,6,8 tetramethylundecanoate) was docked as two separate fragments shown in magenta and blue
SUPPLEMENTARY FIGURES Figure S1 The substrate for DH domain (2R,3R,4R,6R,7S,8S,9R)-3,7,9-trihydroxy-5-oxo-2,4,6,8 tetramethylundecanoate) was docked as two separate fragments shown in magenta and blue
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
Modeling for 3D structure prediction
 Modeling for 3D structure prediction What is a predicted structure? A structure that is constructed using as the sole source of information data obtained from computer based data-mining. However, mixing
Modeling for 3D structure prediction What is a predicted structure? A structure that is constructed using as the sole source of information data obtained from computer based data-mining. However, mixing
Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA
 Online Supplemental Results Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA Structure solution and overall architecture
Online Supplemental Results Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA Structure solution and overall architecture
Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6
 Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6 Hsuan-Liang Liu* and Chin-Wen Chen Department of Chemical Engineering and Graduate Institute
Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6 Hsuan-Liang Liu* and Chin-Wen Chen Department of Chemical Engineering and Graduate Institute
Bioinformatics. Macromolecular structure
 Bioinformatics Macromolecular structure Contents Determination of protein structure Structure databases Secondary structure elements (SSE) Tertiary structure Structure analysis Structure alignment Domain
Bioinformatics Macromolecular structure Contents Determination of protein structure Structure databases Secondary structure elements (SSE) Tertiary structure Structure analysis Structure alignment Domain
Gene regulation I Biochemistry 302. Bob Kelm February 25, 2005
 Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Peptides And Proteins
 Kevin Burgess, May 3, 2017 1 Peptides And Proteins from chapter(s) in the recommended text A. Introduction B. omenclature And Conventions by amide bonds. on the left, right. 2 -terminal C-terminal triglycine
Kevin Burgess, May 3, 2017 1 Peptides And Proteins from chapter(s) in the recommended text A. Introduction B. omenclature And Conventions by amide bonds. on the left, right. 2 -terminal C-terminal triglycine
Structural insights into WcbI, a novel polysaccharide-biosynthesis enzyme
 Volume 1 (2014) Supporting information for article: Structural insights into WcbI, a novel polysaccharide-biosynthesis enzyme Mirella Vivoli, Emily Ayres, Edward Beaumont, Michail N. Isupov and Nicholas
Volume 1 (2014) Supporting information for article: Structural insights into WcbI, a novel polysaccharide-biosynthesis enzyme Mirella Vivoli, Emily Ayres, Edward Beaumont, Michail N. Isupov and Nicholas
Tracking Protein Allostery in Evolution
 Tracking Protein Allostery in Evolution Glycogen phosphorylase frees sugars to provide energy GP orthologs diverged 600,000,000 years can respond to transcription controls, metabolite concentrations and
Tracking Protein Allostery in Evolution Glycogen phosphorylase frees sugars to provide energy GP orthologs diverged 600,000,000 years can respond to transcription controls, metabolite concentrations and
What binds to Hb in addition to O 2?
 Reading: Ch5; 158-169, 162-166, 169-174 Problems: Ch5 (text); 3,7,8,10 Ch5 (study guide-facts); 1,2,3,4,5,8 Ch5 (study guide-apply); 2,3 Remember Today at 5:30 in CAS-522 is the second chance for the MB
Reading: Ch5; 158-169, 162-166, 169-174 Problems: Ch5 (text); 3,7,8,10 Ch5 (study guide-facts); 1,2,3,4,5,8 Ch5 (study guide-apply); 2,3 Remember Today at 5:30 in CAS-522 is the second chance for the MB
Bonds and Structural Supports
 Bonds and Structural Supports Part of the Jmol Training Guide from the MSOE Center for BioMolecular Modeling Interactive version available at http://cbm.msoe.edu/teachingresources/jmol/jmoltraining/struts.html
Bonds and Structural Supports Part of the Jmol Training Guide from the MSOE Center for BioMolecular Modeling Interactive version available at http://cbm.msoe.edu/teachingresources/jmol/jmoltraining/struts.html
Structures of Human Cytosolic NADP-dependent Isocitrate Dehydrogenase Reveal a Novel Self-regulatory Mechanism of Activity*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 32, Issue of August 6, pp. 33946 33957, 2004 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Structures of
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 32, Issue of August 6, pp. 33946 33957, 2004 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Structures of
Objective: Students will be able identify peptide bonds in proteins and describe the overall reaction between amino acids that create peptide bonds.
 Scott Seiple AP Biology Lesson Plan Lesson: Primary and Secondary Structure of Proteins Purpose:. To understand how amino acids can react to form peptides through peptide bonds.. Students will be able
Scott Seiple AP Biology Lesson Plan Lesson: Primary and Secondary Structure of Proteins Purpose:. To understand how amino acids can react to form peptides through peptide bonds.. Students will be able
Supporting Information
 Supporting Information Oxaliplatin binding to human copper chaperone Atox1 and protein dimerization Benny D. Belviso, 1 Angela Galliani, 2 Alessia Lasorsa, 2 Valentina Mirabelli, 1,3 Rocco Caliandro, 1
Supporting Information Oxaliplatin binding to human copper chaperone Atox1 and protein dimerization Benny D. Belviso, 1 Angela Galliani, 2 Alessia Lasorsa, 2 Valentina Mirabelli, 1,3 Rocco Caliandro, 1
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12045 Supplementary Table 1 Data collection and refinement statistics. Native Pt-SAD X-ray source SSRF BL17U SPring-8 BL41XU Wavelength (Å) 0.97947 1.07171 Space group P2 1 2 1 2 1 P2
doi:10.1038/nature12045 Supplementary Table 1 Data collection and refinement statistics. Native Pt-SAD X-ray source SSRF BL17U SPring-8 BL41XU Wavelength (Å) 0.97947 1.07171 Space group P2 1 2 1 2 1 P2
Protein Structure. Hierarchy of Protein Structure. Tertiary structure. independently stable structural unit. includes disulfide bonds
 Protein Structure Hierarchy of Protein Structure 2 3 Structural element Primary structure Secondary structure Super-secondary structure Domain Tertiary structure Quaternary structure Description amino
Protein Structure Hierarchy of Protein Structure 2 3 Structural element Primary structure Secondary structure Super-secondary structure Domain Tertiary structure Quaternary structure Description amino
The Structure and Functions of Proteins
 Wright State University CORE Scholar Computer Science and Engineering Faculty Publications Computer Science and Engineering 2003 The Structure and Functions of Proteins Dan E. Krane Wright State University
Wright State University CORE Scholar Computer Science and Engineering Faculty Publications Computer Science and Engineering 2003 The Structure and Functions of Proteins Dan E. Krane Wright State University
Three-dimensional structure of a viral genome-delivery portal vertex
 Three-dimensional structure of a viral genome-delivery portal vertex Adam S. Olia 1, Peter E. Prevelige Jr. 2, John E. Johnson 3 and Gino Cingolani 4 1 Department of Biological Sciences, Purdue University,
Three-dimensional structure of a viral genome-delivery portal vertex Adam S. Olia 1, Peter E. Prevelige Jr. 2, John E. Johnson 3 and Gino Cingolani 4 1 Department of Biological Sciences, Purdue University,
NB-DNJ/GCase-pH 7.4 NB-DNJ+/GCase-pH 7.4 NB-DNJ+/GCase-pH 4.5
 SUPPLEMENTARY TABLES Suppl. Table 1. Protonation states at ph 7.4 and 4.5. Protonation states of titratable residues in GCase at ph 7.4 and 4.5. Histidine: HID, H at δ-nitrogen; HIE, H at ε-nitrogen; HIP,
SUPPLEMENTARY TABLES Suppl. Table 1. Protonation states at ph 7.4 and 4.5. Protonation states of titratable residues in GCase at ph 7.4 and 4.5. Histidine: HID, H at δ-nitrogen; HIE, H at ε-nitrogen; HIP,
Supplementary Figure 3 a. Structural comparison between the two determined structures for the IL 23:MA12 complex. The overall RMSD between the two
 Supplementary Figure 1. Biopanningg and clone enrichment of Alphabody binders against human IL 23. Positive clones in i phage ELISA with optical density (OD) 3 times higher than background are shown for
Supplementary Figure 1. Biopanningg and clone enrichment of Alphabody binders against human IL 23. Positive clones in i phage ELISA with optical density (OD) 3 times higher than background are shown for
Lecture 10 (10/4/17) Lecture 10 (10/4/17)
 Lecture 10 (10/4/17)") Lecture 10 (10/4/17) Reading: Ch4; 125, 138-141, 141-142 Problems: Ch4 (text); 7, 9, 11 Ch4 (study guide); 1, 2 NEXT Reading: Ch4; 125, 132-136 (structure determination) Ch4; 12-130 (Collagen) Problems:
Lecture 10 (10/4/17) Reading: Ch4; 125, 138-141, 141-142 Problems: Ch4 (text); 7, 9, 11 Ch4 (study guide); 1, 2 NEXT Reading: Ch4; 125, 132-136 (structure determination) Ch4; 12-130 (Collagen) Problems:
COMP 598 Advanced Computational Biology Methods & Research. Introduction. Jérôme Waldispühl School of Computer Science McGill University
 COMP 598 Advanced Computational Biology Methods & Research Introduction Jérôme Waldispühl School of Computer Science McGill University General informations (1) Office hours: by appointment Office: TR3018
COMP 598 Advanced Computational Biology Methods & Research Introduction Jérôme Waldispühl School of Computer Science McGill University General informations (1) Office hours: by appointment Office: TR3018
Experimental and Computational Mutagenesis to Investigate the. Positioning of a General Base within an Enzyme Active Site
 Experimental and Computational Mutagenesis to Investigate the Positioning of a General Base within an Enzyme Active Site Jason P. Schwans, Philip Hanoian, Benjamin J. Lengerich, Fanny Sunden, Ana Gonzalez
Experimental and Computational Mutagenesis to Investigate the Positioning of a General Base within an Enzyme Active Site Jason P. Schwans, Philip Hanoian, Benjamin J. Lengerich, Fanny Sunden, Ana Gonzalez
CH 3 CH 2 OH +H 2 O CHO. 2e + 2H + + O 2 H 2 O +HCOOH
 2 4 H CH 3 2e + 2H + + 2 H 2 2 H CH 2 H 2e + 2H + + 2 H 2 2 H +H 2 CH 2e + 2H + + 2 H 2 2 H +HCH Supplemental Figure S. The three-step 4DM reaction, each step requires two reducing equivalents from ADPH
2 4 H CH 3 2e + 2H + + 2 H 2 2 H CH 2 H 2e + 2H + + 2 H 2 2 H +H 2 CH 2e + 2H + + 2 H 2 2 H +HCH Supplemental Figure S. The three-step 4DM reaction, each step requires two reducing equivalents from ADPH
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/5/243/ra68/dc1 Supplementary Materials for Superbinder SH2 Domains Act as Antagonists of Cell Signaling Tomonori Kaneko, Haiming Huang, Xuan Cao, Xing Li, Chengjun
www.sciencesignaling.org/cgi/content/full/5/243/ra68/dc1 Supplementary Materials for Superbinder SH2 Domains Act as Antagonists of Cell Signaling Tomonori Kaneko, Haiming Huang, Xuan Cao, Xing Li, Chengjun
Outline. Levels of Protein Structure. Primary (1 ) Structure. Lecture 6:Protein Architecture II: Secondary Structure or From peptides to proteins
 Structure. Lecture 6:Protein Architecture II: Secondary Structure or From peptides to proteins") Lecture 6:Protein Architecture II: Secondary Structure or From peptides to proteins Margaret Daugherty Fall 2003 Outline Four levels of structure are used to describe proteins; Alpha helices and beta sheets
Lecture 6:Protein Architecture II: Secondary Structure or From peptides to proteins Margaret Daugherty Fall 2003 Outline Four levels of structure are used to describe proteins; Alpha helices and beta sheets
Secondary Structure. Bioch/BIMS 503 Lecture 2. Structure and Function of Proteins. Further Reading. Φ, Ψ angles alone determine protein structure
 Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
Packing of Secondary Structures
 7.88 Lecture Notes - 4 7.24/7.88J/5.48J The Protein Folding and Human Disease Professor Gossard Retrieving, Viewing Protein Structures from the Protein Data Base Helix helix packing Packing of Secondary
7.88 Lecture Notes - 4 7.24/7.88J/5.48J The Protein Folding and Human Disease Professor Gossard Retrieving, Viewing Protein Structures from the Protein Data Base Helix helix packing Packing of Secondary
Central Dogma. modifications genome transcriptome proteome
 entral Dogma DA ma protein post-translational modifications genome transcriptome proteome 83 ierarchy of Protein Structure 20 Amino Acids There are 20 n possible sequences for a protein of n residues!
entral Dogma DA ma protein post-translational modifications genome transcriptome proteome 83 ierarchy of Protein Structure 20 Amino Acids There are 20 n possible sequences for a protein of n residues!
Supersecondary Structures (structural motifs)
") Supersecondary Structures (structural motifs) Various Sources Slide 1 Supersecondary Structures (Motifs) Supersecondary Structures (Motifs): : Combinations of secondary structures in specific geometric
Supersecondary Structures (structural motifs) Various Sources Slide 1 Supersecondary Structures (Motifs) Supersecondary Structures (Motifs): : Combinations of secondary structures in specific geometric
Impact of the crystallization condition on importin-β conformation
 Supporting information Volume 72 (2016) Supporting information for article: Impact of the crystallization condition on importin-β conformation Marcel J. Tauchert, Clément Hémonnot, Piotr Neumann, Sarah
Supporting information Volume 72 (2016) Supporting information for article: Impact of the crystallization condition on importin-β conformation Marcel J. Tauchert, Clément Hémonnot, Piotr Neumann, Sarah
Structure and RNA-binding properties. of the Not1 Not2 Not5 module of the yeast Ccr4 Not complex
 Structure and RNA-binding properties of the Not1 Not2 Not5 module of the yeast Ccr4 Not complex Varun Bhaskar 1, Vladimir Roudko 2,3, Jerome Basquin 1, Kundan Sharma 4, Henning Urlaub 4, Bertrand Seraphin
Structure and RNA-binding properties of the Not1 Not2 Not5 module of the yeast Ccr4 Not complex Varun Bhaskar 1, Vladimir Roudko 2,3, Jerome Basquin 1, Kundan Sharma 4, Henning Urlaub 4, Bertrand Seraphin
Transmembrane Domains (TMDs) of ABC transporters
 of ABC transporters") Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Supporting Information. Structural and functional characterization of human and murine C5a anaphylatoxins
 Supporting Information Structural and functional characterization of human and murine C5a anaphylatoxins Janus Asbjørn Schatz-Jakobsen a, Laure Yatime a, Casper Larsen a, Steen Vang Petersen b, Andreas
Supporting Information Structural and functional characterization of human and murine C5a anaphylatoxins Janus Asbjørn Schatz-Jakobsen a, Laure Yatime a, Casper Larsen a, Steen Vang Petersen b, Andreas
of the Guanine Nucleotide Exchange Factor FARP2
 Structure, Volume 21 Supplemental Information Structural Basis for Autoinhibition of the Guanine Nucleotide Exchange Factor FARP2 Xiaojing He, Yi-Chun Kuo, Tyler J. Rosche, and Xuewu Zhang Inventory of
Structure, Volume 21 Supplemental Information Structural Basis for Autoinhibition of the Guanine Nucleotide Exchange Factor FARP2 Xiaojing He, Yi-Chun Kuo, Tyler J. Rosche, and Xuewu Zhang Inventory of
Supplementary information
 Supplementary information The structural basis of modularity in ECF-type ABC transporters Guus B. Erkens 1,2, Ronnie P-A. Berntsson 1,2, Faizah Fulyani 1,2, Maria Majsnerowska 1,2, Andreja Vujičić-Žagar
Supplementary information The structural basis of modularity in ECF-type ABC transporters Guus B. Erkens 1,2, Ronnie P-A. Berntsson 1,2, Faizah Fulyani 1,2, Maria Majsnerowska 1,2, Andreja Vujičić-Žagar
Bahnson Biochemistry Cume, April 8, 2006 The Structural Biology of Signal Transduction
 Name page 1 of 6 Bahnson Biochemistry Cume, April 8, 2006 The Structural Biology of Signal Transduction Part I. The ion Ca 2+ can function as a 2 nd messenger. Pick a specific signal transduction pathway
Name page 1 of 6 Bahnson Biochemistry Cume, April 8, 2006 The Structural Biology of Signal Transduction Part I. The ion Ca 2+ can function as a 2 nd messenger. Pick a specific signal transduction pathway
Part 7 Bonds and Structural Supports
 Part 7 Bonds and Structural Supports http://cbm.msoe.edu/newwebsite/learntomodel Introduction In addition to covalent bonds between atoms in a molecule, Jmol has the ability to render Hydrogen Bonds and
Part 7 Bonds and Structural Supports http://cbm.msoe.edu/newwebsite/learntomodel Introduction In addition to covalent bonds between atoms in a molecule, Jmol has the ability to render Hydrogen Bonds and
Acta Crystallographica Section D
 Supporting information Acta Crystallographica Section D Volume 70 (2014) Supporting information for article: Structural basis of the heterodimerization of the MST and RASSF SARAH domains in the Hippo signalling
Supporting information Acta Crystallographica Section D Volume 70 (2014) Supporting information for article: Structural basis of the heterodimerization of the MST and RASSF SARAH domains in the Hippo signalling
Supporting Information
 Electronic Supplementary Material (ESI) for Physical Chemistry Chemical Physics. This journal is the Owner Societies 2016 Supporting Information Lipid molecules can induce an opening of membrane-facing
Electronic Supplementary Material (ESI) for Physical Chemistry Chemical Physics. This journal is the Owner Societies 2016 Supporting Information Lipid molecules can induce an opening of membrane-facing
Orientational degeneracy in the presence of one alignment tensor.
 Orientational degeneracy in the presence of one alignment tensor. Rotation about the x, y and z axes can be performed in the aligned mode of the program to examine the four degenerate orientations of two
Orientational degeneracy in the presence of one alignment tensor. Rotation about the x, y and z axes can be performed in the aligned mode of the program to examine the four degenerate orientations of two
Biomolecules: lecture 10
 Biomolecules: lecture 10 - understanding in detail how protein 3D structures form - realize that protein molecules are not static wire models but instead dynamic, where in principle every atom moves (yet
Biomolecules: lecture 10 - understanding in detail how protein 3D structures form - realize that protein molecules are not static wire models but instead dynamic, where in principle every atom moves (yet
ml. ph 7.5 ph 6.5 ph 5.5 ph 4.5. β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS
 a UV28 absorption (mau) 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS β 2 AR-Gs complex dissociated complex excess nucleotides b 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex
a UV28 absorption (mau) 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS β 2 AR-Gs complex dissociated complex excess nucleotides b 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex
Heteropolymer. Mostly in regular secondary structure
 Heteropolymer - + + - Mostly in regular secondary structure 1 2 3 4 C >N trace how you go around the helix C >N C2 >N6 C1 >N5 What s the pattern? Ci>Ni+? 5 6 move around not quite 120 "#$%&'!()*(+2!3/'!4#5'!1/,#64!#6!,6!
Heteropolymer - + + - Mostly in regular secondary structure 1 2 3 4 C >N trace how you go around the helix C >N C2 >N6 C1 >N5 What s the pattern? Ci>Ni+? 5 6 move around not quite 120 "#$%&'!()*(+2!3/'!4#5'!1/,#64!#6!,6!