Liu, Yang (2012) The characterization of a novel abscission-related gene in Arabidopsis thaliana. PhD thesis, University of Nottingham.
|
|
|
- Arline Hill
- 5 years ago
- Views:
Transcription

1 Liu, Yang (2012) The characterization of a novel abscission-related gene in Arabidopsis thaliana. PhD thesis, University of Nottingham. Access from the University of Nottingham repository: Copyright and reuse: The Nottingham eprints service makes this work by researchers of the University of Nottingham available open access under the following conditions. This article is made available under the University of Nottingham End User licence and may be reused according to the conditions of the licence. For more details see: For more information, please contact eprints@nottingham.ac.uk
2 Chapter 4 Functional analysis of G2 4.1 Manipulation of G2 Expression The expression analysis described in chapter 3 indicates that G2 is expressed specifically in AZ and in cortical cells overlying the lateral root primordial. The approach that was applied to determine the function of G2 was to manipulate its expression. A reverse genetics strategy was applied to investigate the role of G2 in plant development. Two T-DNA insertion lines associated with G2 were identified and seeds were obtained from Nottingham Arabidopsis Stock Centre (NASC). Homozygous knock out lines were determined by PCR. Unfortunately none of them showed a reduction in expression of G2 in floral tissues. An RNAi strategy was then undertaken and homozygous lines were isolated and the phenotypes of down-regulated lines were studied. To further investigate the potential function of G2, the gene was ectopically expressed under the regulation of a 35SCaMV promoter. The resulting G2 ectopic expression lines were characterized in detail. 113
 were identified and obtained from NASC (European Arabidopsis Stock Centre) (Scholl RL, et al., 2000).")
3 Generation of G2 null lines using a T-DNA strategy A T-DNA insertion strategy was applied to generate null lines of G2. T3 T-DNA insertion lines SALK_ and SALK_ (Figure 4.1) were identified and obtained from NASC (European Arabidopsis Stock Centre) (Scholl RL, et al., 2000). The T-DNA insertions were at 310 bp (SALK_065404) and 360 bp (SALK_055356) upstream of the translation start site. Genomic PCR was used to identify the homozygous KOs of the two T-DNA insertion lines (Figure 4.1). Figure 4.1 A schematic diagram of T-DNA insertion positions of G2. The diagrams are not drawn to scale. For T-DNA insertion line SALK_065404, the genomic PCR result indicates that samples 3 and 6 are the T-DNA insertion homozygotes (Figure 4.2) (Shahid, unpubished data). 114
.")

. Figure 4.3 PCR analysis of putative T-DNA insertion lines (SALK_55356) of G2.")

4 Figure 4.2 PCR analysis of putative T-DNA insertion lines (SALK_565404) of G2. Group Insertion specific shows the PCR products amplified by primers LBb1 and G2_reverse (Table 1.1). Group Gene Specific shows the PCR products amplified by primers G2PR_Forward & G2_reverse. L: ladder. Lines 3 and 6 were identified as homozygous insertion lines. For T-DNA insertion line SALK_055356, the PCR result indicates that sample 1, 2, and 4 are the T-DNA insertion homozygotes (Figure 4.3). Figure 4.3 PCR analysis of putative T-DNA insertion lines (SALK_55356) of G2. Group Insertion specific shows the PCR products amplified by primers G2PR_F & LBb1. Group Gene Specific shows the PCR products amplified by primers G2PR_F & G2_rev. wt: wild type. Lines 1, 2 and 4 are identified as homozygous insertion lines. A Reverse-transcription PCR (RTPCR) was then carried out to confirm that G2 expression was silenced in the KO lines. Total RNA was isolated from flowers in position 5 8. HK-CBP primers (Table 1.1) were used to amplify 115


5 housekeeping transcripts. The results showed that neither of the T-DNA insertion lines exhibited reduced the expression of G2 (Figure 4.4). Figure 4.4 RT-PCR analysis of putative T-DNA insertion lines, SALK_65404 and SALK_ Group HK-CBP shows the RT-PCR product amplified by HK-CBP primers and group G2 Specific shows the RT-PCR product amplified by G2_Forward and G2_Reverse primers. L: ladder Generation of G2 null lines using an RNAi strategy Fragments with 357 base pairs covering the full length cdna of G2, were fused into an entry vector pdonr221 (Invitrogen TM ) by using a Gateway TM Technology (Figure 4.5). Because the gene does not have an intron, 116
 were designed and the amplified fragments from PCR by using Phusion TM DNA Polymerase were purified before fusing them into the vector pdonr221 by the Gateway TM BP-reaction strategy which was")
6 primers with attb1.1 and attb2.1 sites (Table 1.1) were designed and the amplified fragments from PCR by using Phusion TM DNA Polymerase were purified before fusing them into the vector pdonr221 by the Gateway TM BP-reaction strategy which was described in section The resulting construct was called G2Entry (Figure 4.6). The plasmids were then transformed into E.coli strain DH5α using a heat-shock method (Sambrook et al., 1989) as described in section The transformed cells were selected by spreading the cells on LB medium plates containing 50 μg/ml kanamycin and the resistant plasmids were isolated and purified using a NucleoSpin TM Plasmid mini kit which has been described in section The plasmids were confirmed by PCR (Figure 4.7) and then sent for sequencing. Figure 4.5 Diagram of GateWay TM pdonr TM 221 entry vector, which contains a puc origin, a Kanamycin gene, and M13 primers sites for sequencing (Invitrogen TM ). 117
.")
7 Figure 4.6 Schematic diagram of G2Entry plasmid construct. The 357 bp G2 cdna insert was fused into pdonr TM 221 by BP reaction. A kanamycin-resistance gene was used as a selectable marker in bacteria. Figure 4.7 PCR analysis of the colonies that contain the G2Entry plasmid using primers M13_Forward and G2_reverse (Table 1.1). The positions of the primers are shown in figure 4.6. L: ladder. 118
8 The RNAi construct was then made by fusing the fragments into a destination vector pk7gwiwg2 which has been described by Karimi et al (2005). The Gateway TM technology LR reaction was applied following the procedure described in section The final construct was named RNAi:G2. A diagrammatic representation of the construct is shown in Figure 4.8. Prior to the transformation, the constructs both in E.coli DH5α and Agrobacterium tumefaciens C58 were confirmed by PCR using respective primers (Table 1.1). DNA bands with the expected size were successfully amplified (Figure 4.9). Figure 4.8 The G2 cdna fragment in the Entry Clone was transferred to a destination vector, pk7gwiwg2, which contains compatible recombination sites, attr1 and AttR2, in a reaction mediated by Gateway TM LR Clonase TM Enzyme Mix (Invitrogen). The resulting construct was named RNAI:G2. This construct was expected to produce hairpin RNA from the inserted G2 fragments, triggering post-transcriptional gene silencing. The diagrams are not drawn to scale. 119


9 Figure 4.9 PCR analysis of plasmids from the two colonies transformed with Agrobacterium tumefaciens C58. Group1 shows the PCR products amplified by primers G2AttB_forward & PK7_Reverse (Table 1.1). Group2 shows the PCR products amplified by primers 35S_Forward & G2AttB_reverse. The positions of primers are shown in Figure 4.8. The plasmid carrying constructs RNAi:G2 in Agrobacterium tumefaciens C58 were transformed into Arabidopsis thaliana flowers by the Floral dip method (Clough and Bent, 1998) which has been described in section A total of 14 independent primary transformants for RNAi:G2 were obtained from the Kanamycin-resistant screening. In order to confirm the down-regulation of G2 expression, an RT-PCR was carried out by using G2_forward and G2_reverse primers with HK-CBP primers as global control (Table 1.1). Total RNA was isolated from flowers at positions from 5 8 where G2 has the highest amount of expression. Compared to wild type, weak or no RT-PCR product was observed. Two lines without any observed band were selected to generate T2 and T3 to select homozygotes (Figure 4.10). 120
 of both wild type and primary transformants of RNAi:G2. L: ladder. 4.1.3.")
10 Figure 4.10 RT-PCR analysis of RNAi:G2 Plants. Group Gene Specific shows the RT-PCR product amplified by using primer G2_Forward and G2_Reverse. Group HK-CBP shows the RT-PCR product amplified using HK-CBP primers. Total RNA was isolated from mature flowers (P5-8) of both wild type and primary transformants of RNAi:G2. L: ladder Generation of ectopically expressing lines of G2 The next strategy was to generate overexpression lines of G2. The full length cdna (357 bp) of the gene was inserted into a destination vector pgwb8 which was described in section The Gateway TM technology LR reaction was then carried out following the procedure described in section To confirm that transformation was successful, a genomic PCR was carried out using 35S_forward primer and G2_reverse primer (Table 1.1) and a band with expected size was obtained. The final construct was named 35S:G2. A diagrammatic representation of the construct is 121
11 shown in Figure Prior to the transformation, the constructs both in E.coli DH5α and Agrobacterium tumefaciens C58 were confirmed by PCR using specific primers. Bands of the expected size were successfully amplified (Figure 4.12). Figure The G2 fragment in the Entry Clone was transferred to a destination vector, pgwb8, which contains compatible recombination sites, attr1 and AttR2, in a reaction mediated by Gateway TM LR Clonase TM Enzyme Mix (Invitrogen). The resulting construct was named 35S:G2. This construct is expected to make G2 express intensively in the transformed plants. The diagrams are not drawn to scale. 122

12 Figure 4.12 PCR analysis of plasmids from the six colonies of transformed Agrobacterium tumefaciens C58. C: Control group from the plasmid transformed into Agrobacterium tumefaciens C58. L: ladder. PCR products were amplified using primers 35S_Forward & G2AttB_rev. The position of primers is shown in Figure The plasmid constructs in Agrobacterium tumefaciens C58 were transformed into Arabidopsis thaliana flowers by the Floral dip method (Clough and Bent, 1998) which has been described in section The primary transformants of 35S:G2 construct were screened from Kanamycin-resistant screening. A total of 12 independent transformants for 35S:G2 were obtained and transferred into pots containing Levington M3 compost. The T1 transformants were confirmed by genomic PCR using 35S_forward and G2_reverse primers (Table 1.1). In order to confirm the ectopic expression of G2, an RT-PCR analysis was carried out using G2_forward and G2_reverse primers (Table 1.1), with HK-CBP primers as global control. Total RNA was isolated from cauline leaves, where G2 does not naturally express. RT-PCR amplification was clearly observed in all the 12 T1 transformants (Figure 4.13). T2 homozygotes were selected by screening T3 progenies in MS media with Kanamycin and 100% resistant lines were considered as homozygotes. 123

13 Figure 4.13 RT-PCR analysis of 35S:G2 Plants. Group Gene Specific shows the RT-PCR product amplified using primer G2_Forward and G2_Reverse. Group HK-CBP shows the RT-PCR product amplified using HK-CBP primers. Total RNA was isolated from cauline leaves of mature Arabidopsis both wild type and 35S:G2 plants. No expression of G2 was observed in wild type (WT) comparing to significant expression in Transformed 35S:G2 plants. L: ladder. 124
14 4.2 Characterization of Knock Out and overexpression lines of G2 and analysis of phenotypes Phenotypes of RNAi:G2 plants An in-depth comparative analysis of floral organ abscission was carried out between RNAi:G2 homozygous and wild type plants. RNAi:G2 lines did not show any visible difference in either the timing of abscission or in the development of floral organs. An analysis of pod development in RNAi:G2 plants revealed that seeds in mature siliques did not all fully develop (Figure A, B). Figure 4.14 (C) shows the percentage of normally developed seeds in a wild type and RNAi:G2 plants. Fourteen pods at position 12 were collected from three individuals of wild type and RNAi:G2 plants respectively and the percentage of fully developed seeds was calculated. The average final percentage of fully developed seeds in wild type plants was 92% whereas in RNAi:G2 it was 25%. The abnormal seeds were observed to have aborted. 125

15 Figure 4.14 Undeveloped seeds identified in siliques of RNAi:G2 (A) and wild type (B) and the percentage of fully developed seeds (C). Scale bar: 100 µm. A pollen viability test was performed using Alexander staining to determine the viability of the RNAi:G2 pollen. Pollen grains stained in purple indicate viable cytoplasm, whereas undeveloped pollen is stained green. In comparison to wild type, RNAi:G2 pollen was only partially functional (Figure 4.15). The percentage of undeveloped pollen per anther was determined by calculating the undeveloped pollen in twenty anthers. The result showed that the average percentage of undeveloped pollen was 21.78%, comparing to 2.45% in wild type. 126
. Error bars: standard error. 4.2.2 Phenotype of 35S:G2 plants A phenotypic characterization was carried out in 35S:G2 homozygous lines and wild type plants.")
16 Picture 4.15 A: Pollen of RNAi:G2 and wild type stained with Alexander solution. The arrow shows the percentage of undeveloped pollen grains. Scale bar: 50 µm B: The percentage of undeveloped pollen grainsrnai:g2 plant and wild type (WT). Error bars: standard error Phenotype of 35S:G2 plants A phenotypic characterization was carried out in 35S:G2 homozygous lines and wild type plants. The 35S:G2 and wild type plants were grown under the identical conditions as described in section
17 An in-depth comparative analysis of floral organ abscission was carried out between 35S:G2 homozygous and wild type plants. The first flower on the inflorescence with visible white petals was considered as position 1. The wild type Arabidopsis plant normally abscises its floral organs at position 5 7 in natural conditions. Compared to wild type, 35S:G2 lines did not show any significant difference either in the timing of abscission or in the development of floral organs. An analysis of root development in 35S:G2 seedlings in different stages under the microscope revealed a striking phenotype in root hair development compared to wild type plants. In general the 35S:G2 plants developed significantly shorten and more swollen root hairs. The swollen root hairs were observed as the primary roots emerged from the germinating seeds and this phenotype was maintained throughout root development. However the level of the bulging was observed to be reduced as it came near to the elongation zone. The bulging was observed in trichoblasts and not all the trichoblasts were affected. In some 10-day-old seedlings, the development of root hairs near the elongation zone was indistinguishable to wild type (Figure 4.16). 128

. Type I root hairs, which locate furthest from elongation zone, are extremely swollen.")
18 Figure 4.16 Phenotypic analysis of the roots of wild type (A, C and E) and 35S:G2 plant (B, D and F). The 7 day old Arabidopsis thaliana primary roots were analysed under the microscope. Scale bar: A, B, C, and D: 500 µm. E, F: 50 µm Based on the morphological differences, the phenotypes of 35S:G2 root hairs were classified into three types: type I, II, and III (Figure 4.17). Type I root hairs, which locate furthest from elongation zone, are extremely swollen. The degree of swelling varies with some trichoblasts being 129
19 completely swollen while some were relatively unaffected. No tip growth or elongation was detected in type I hairs. Type II hairs were normally located between type I and type III hairs. Tip growth and elongation was detected but the hairs were much shorter than wild type. The diameters of the hairs varied along the length of the hairs and extra bulges were formed during the elongation process. Type III hairs were located near the root elongation zone and the length was longer than type II hairs but shorter than wild type. Type III hairs should be distinguished from type II hairs by that the diameters of the type III hairs are consistent along the length. Branches were detected in some type III hairs. Some type III hairs also displayed a crooked appearance (Figure 4.17). 130

20 Figure 4.17 Phenotype of wild type and 35S:G2 root hairs. 7 day old Arabidopsis roots were analysed under the microscope. WT: wild type. I, II, III: type I, type II and type III. Arrows in 35S:G2 II show the hairs with diameters varied along the length of the hairs. Arrows in 35S:G2 III show the hairs with branches. Scale bar: 500 µm. Thirty 5-day-old root hairs from two 35S:G2 T3 lines were selected and root hair diameter, length and hair density were measured under a microscope. The same number of 5-day-old wild type seedlings was used for the control. A bar chart describing the average diameter of the three types of root hairs is shown below. The average diameters of 35S:G2 type I, II and III root hairs were µm, µm and µm compared to µm in wild type (Figure 4.18). 131
.")
21 Figure 4.18 The diameters of 35S:G2 type I, type II and type III root hairs. The diameters were determined by calculating the average diameter of thirty root hairs. Error bar: stand error. The average length of 35S:G2 type I, II and III root hairs was µm, µm and µm compared to µm in wild type (Figure). The average length of type I hairs was lower than 40 µm, which indicates that there was no tip growth in type I hairs (Figure 4.19). 132

22 Figure 4.19 The length of 35S:G2 type I, type II and type III root hairs. The length was determined by calculating the average length of thirty root hairs. Error bars: standard error. The density of root hairs was measured by counting the quantity of root hairs per 1 mm in the elongation zone of roots. The value was determined by calculating the average number per 1 mm of twenty root hairs. 35S:G2 seedlings were shown to have root hairs per 1 mm, which was 0.75 greater than wild type (18.6 per 1 mm) (Figure 4.20). 133

23 Figure 4.20 The density of 35S:G2 and wild type root hairs in the elongation zone. The density was determined by calculating the average number per 1 mm of twenty root hairs. Error bars: Standard error. 134
24 4.3 Crossing 35S:G2 with 35S:IDA In chapter 3 it was demonstrated that G2 is an abscission-related gene on the basis of expression analysis. A gene expression manipulation strategy was carried out in order to investigate the function of G2. However neither down-regulating nor up-regulating the expression of G2 has revealed a phenotype modified in the floral organ abscission. The expression analysis of G2 discussed in chapter 3 revealed a negative correlation between G2 and INFLORESCENCE DEFICIENT IN ABSCISSION (IDA) which is an important abscission-related gene. It has been shown that IDA controls floral organ abscission and the null mutant without the presence of IDA fails to shed its floral organs (Butenko et al., 2003). Further analysis of plants with ectopically expressed IDA showed that such material had an earlier abscission and exhibited an extended AZ (Stenvik et al., 2006). 35S:IDA plants were also associated with enhanced production of arabinogalactan protein (AGP). This was substantially secreted in the AZ and the amount secreted was much more than that found in wild type. AGP was absent in the AZ of ida (Stenvik et al., 2006). Further expression analysis by micro array and RT-PCR showed that one of the AGPs in Arabidopsis, AGP24 was up-regulated in 35S:IDA and down-regulated in ida (Stenvik et al., 2006). IDA is not only expressed specifically in the AZ but also in the cortex and epidermal cells overlying lateral root primordial and a mutation in IDA also leads to a delay in lateral root emergence (Kumpf et al. 2010). 135
25 G2 and IDA have a similar expression patterning with both of them expressing in AZ and in the cortical cells adjacent to emerging lateral roots and both of them are strongly up-regulated by IAA. Expression analysis showed a negative correlation between G2 and IDA. Given the above observation, a cross between 35S:G2 and 35S:IDA was performed in order to further investigate the interactions between G2 and IDA Screening and isolation of homozygous lines of 35S:G2 35S:IDA To investigate the potential relationship between G2 and IDA, a cross was carried out between 35S:G2 and 35S:IDA. As both 35S:G2 and 35S:IDA are dominant, the genotype can not be characterized by phenotype. In order to select the homozygotes, genomic PCR screening was carried out to determine the genotype of F2 and F3 progenies. DNA isolated from individual plants was analyzed using 35S-promoter-specific and gene-specific primers. In the F2 generation, individuals with both PCR amplifications of 35S:G2 and 35S:IDA were selected (Figure 4.21) and F3 lines generated. PCR was performed on F3 lines with at least 24 individuals per line. Lines with 100% individuals having both PCR amplifications from 35S forward and G2/IDA specific primers (Table 1.1) were selected as homozygotes. Figure 4.22 shows that the F3 line B13 was homozygous for both genes. 136

, Group A shows the amplification from 35S_Forward and IDA_Reverse primers and Group B")
26 Figure 4.21 Genomic PCR to select F2 generation of 35S:G2 x 35S:IDA, Group A shows the amplification from 35S_Forward and IDA_Reverse primers and Group B shows the amplification from 35S_Forward and G2_Reverse primers. Figure 4.22 Genomic PCR to select F3 homozygotes of 35S:G2 x 35S:IDA (line B13), Group A shows the amplification from 35S_Forward and IDA_Reverse primers and Group B shows the amplification from 35S_Forward and G2_Reverse primers Phenotype of 35S:IDA x 35S:G2 lines Overexpression of IDA leads to a series of phenotypes. Firstly, 35S:IDA individuals show earlier abscission from position 4 compared to position 6 137
27 8 of wild type. Secondly, 35S:IDA plants show an extended abscission zone. Thirdly, a large amount of AGP is secreted at the site of abscission and this is maintained until the silique is mature (Stenvik et al., 2006). 35S:G2 does not show any visible phenotype in floral organ abscission. An in-depth analysis of floral organ abscission was carried out on 35S:IDA, 35S:G2 and 35S:G2 35S:IDA plants with wild type as control. In terms of the timing of abscission, 35S:IDA lines showed early abscission at position 3, compared to position 6 in wild type. Both 35S:G2 and 35S:G2 35S:IDA showed abscission at position 6, which is the same as wild type (Figure 4.23). In terms of the development of AZ, 35S:IDA individuals showed significant extended AZs which were easy to be distinguished, whereas 35S:G2 and 35S:G2 35S:IDA showed no visible difference from wild type (Figure 4.24). 138


28 Figure 4.23 Time course analysis of floral organ abscission of wild type (WT), 35S:G2, 35S:IDA and 35S:IDA x 35S:G2 plants. The shedding position was analyzed by gentle shaking of the flowers. P: position. Scale bar: 1 mm. 139

29 Figure 4.24 Floral AZ of wild type (WT) (A), 35S:G2 (B), 35S:IDA (C) and 35S:IDA x 35S:G2 (D). 35S:IDA shows a significantly extended AZ and with white material covering the AZ whereas 35S:G2 and 35S:IDA x 35S:G2 show no difference from wild type. Scale bar: 500 µm. In terms of the secreted AGP, in 35S:IDA a white substance (AGP) was detected covering the extended AZ. No white substance was detected either in 35S:G2 or 35S:G2 35S:IDA, which was the same as the wild type. A synthetic chemical reagent β-d-glucosyl Yariv (β-glcy) was used to determine production of AGP. 35S:IDA showed a higher amount of AGP covering the AZ whereas in wild type only a small amount of AGP was detected at position 4. Interestingly, no AGP or less AGP was detected in both 35S:G2 and 35S:G2 x 35S:IDA plants (Figure 4.25). 140
30 These results reveal that by overexpressing G2 in a 35S:IDA background, in terms of floral organ abscission and AZ development, the phenotypic characteristics are lost and 35S:IDA x 35S:G2 plants look essentially like wild type. The series of effects indicate that overexpression of G2 can rescue 35S:IDA phenotypes. This observation suggests that G2 and IDA could function in the same pathway or that expression of IDA is down-regulated by G2. 141
 wild type (WT), (C, D) 35S:IDA, (E,F) 35S:G2, (G,H) 35S:G2 x 35S:IDA.")
31 Figure Identification of Arabinogalactan using the Yariv Reagent β-glcy. Staining areas are shown in red. (A, B) wild type (WT), (C, D) 35S:IDA, (E,F) 35S:G2, (G,H) 35S:G2 x 35S:IDA. (A, C, D, E) were selected from position 4 (P4) and (B, D, F, H) were selected from position 8 (P8). Scale bar: 500 µm. 142
32 To test the hypothesis that IDA is down-regulated by G2, a quantitative-reverse Transcription-PCR (qpcr) was performed to determine the expression pattern of G2 and IDA respectively in the backgrounds of 35S:G2, 35S:IDA, 35SG2 35S:IDA, and wild type, following the protocol which was described in chapter Total RNA was isolated from both flowers (P5 P8) and cauline leaves. Two biological duplicates and two independent samples from each duplicate were examined. The relative expression levels were normalized to the expression of ACTIN 2 (ACT2). The qrt-pcr results showed that G2 expression was significantly up-regulated in the 35S:G2 plant compared to wild type whereas in the 35S:IDA background G2 expression was observed to be very slightly down-regulated. The G2 expression in the background 35S:G2 x 35S:IDA was unaffected compared to the wild type but the expression in the flowers of 35S:G2 x 35S:IDA plants was slightly lower than in 35S:G2 background, which suggests that the up-regulation of IDA in a 35S:G2 background down-regulated G2. IDA expression was detected in the flowers of the 35S:G2 and wild type backgrounds and the expression was up-regulated in the 35S:IDA background. Compared to the expression in both the flowers and cauline leaves of 35S:IDA, IDA expression in the 35S:G2 x 35S:IDA background was found to be slightly down-regulated, suggesting that over-expression of IDA is down-regulated by ectopic expression of G2 (Figure 4.26). However the observed expression changes were very slight and further experimentation is required to determine if they reflect biologically significant expression changes. 143
33 144
, IDA (efficiency 1.")
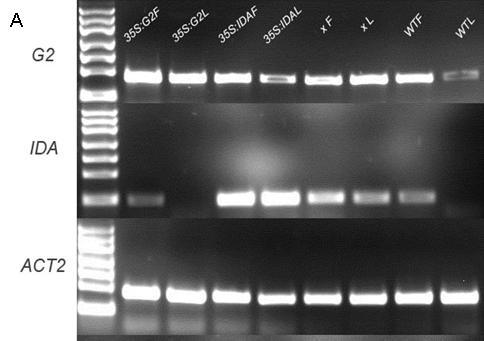
34 Figure 4.26: qrt-pcr of mrna from both flowers (F) (P5 P8) and cauline leaves (L) of 35S:G2, 35S:IDA, 35S:IDA x 35S:G2 (x) and wild type. A: RT-PCR result before qrt-pcr performed. B: qrt-pcr result of G2 expression in different backgrounds. C: qrt-pcr result of IDA expression in different backgrounds. D: Standard curves of G2 (efficiency 1.975), IDA (efficiency 1.686) and ACT2 (efficiency 1.992). Error bar: Stand error. 145
35 S:IDA x 35S:G2 homozygous plants were observed to be normal in morphology and development compared to 35S:IDA. Apart from the morphological and developmental changes in floral organ AZ, 35S:IDA also showed other phenotypic characteristics. Vestigial AZs were found at the bases of pedicels, branches and cauline leaves (Stenvik et al., 2006). In addition, 35S:IDA plants are significantly smaller in structure. In most of the 35S:IDA plants rosette leaves were observed to senesce more rapidly. As described in the previous sections, 35S:G2 plants showed no significant difference compared to wild type except in the phenotype of root hairs. 35S:G2 x 35S:IDA homozygous plants showed no visible difference comparing to wild type, which indicated that the non-abscission-related effects of overexpression of IDA are also rescued by 35S:G2 (Figure 4.27). 146
, B")
36 Figure 4.27 Phenotypes of whole plants of A wild type (WT), B 35S:G2, C 35S:IDA and D 35S:IDA x 35S:G2. 147

37 S:IDA rescues the phenotype of swelling root hairs of 35S:G2 As the result that has been described in chapter 3, overexpression of G2 leads to a phenotype of root hair swelling. In 35S:IDA x 35S:G2 homozygous plants the phenotype of extremely swollen root hair was not observed, while shorter and branched hairs were detected, which indicated that 35S:IDA rescues the root hairs bugling effect of type I hairs but not type II and type III hairs (Figure 4.25). Figure 4.28 Arabidopsis root hairs. A: wild type (WT). B: 35S:G2 x 35S:IDA. C: 35S:IDA. D: 35S:G2. The arrow shows the branched root hair in 35S:G2 x 35S:IDA plant. Scale bar: 500 µm 148
38 35S:IDA plants showed significant smaller seedlings and rosette leaves than wild type while 35S:G2 plants showed no difference. Interestingly, compared to wild type, 35S:G2 and 35S:IDA, the 35S:G2 x 35S:IDA homozygotes showed a different rosette leaf morphology with a decrease in width and increase in length which is consistent even until the plant becomes mature, while the size of the whole seedling was similar compared to wild type (Figure 4.29). 30 leaves from 21-day-old seedlings of 35S:G2 x 35S:IDA, 35S:G2, 35S:IDA and wild type were isolated and measured for the leaf length to width ratio. The results showed that the ratio of leaf length to width were wild type: 2.12, 35S:G2: 2.14, 35S:IDA: 1.93 and 35S:G2 x 35S:IDA: 3.09, which suggested that 35S:G2 x 35S:IDA plants developed a different rosette leaf morphology. 149


39 Figure 4.29 The morphology of rosette leaves (Group A: whole seedlings, Group B: rosette leaves from the same position). WT: wild type. 150
40 4.3.5 Immunolocalization of 35S:G2 root hairs It has been shown that the amount of AGP secretion in AZ is reduced in 35S:G2 comparing to both 35S:IDA and the wild type. AGPs were predicted to act as a mediator between the cell wall and cortical microtubules in Arabidopsis root tissue and the absence of AGP leads to swollen root hairs by disrupting the microtubules (MT) (Andeme-Onzighi et al., 2002), therefore it is possible that the down-regulatation of AGP in 35S:G2 root hairs may lead to the phenotype of swollen root hairs. In order to investigate if the MT structure was changed in root hairs, an immunolocalization strategy was applied to localize MT. The immunolocalization was carried out as shown in chapter using 5-day-seedlings of 35S:G2 and wild type plants. Antibody Antitublin (Abnova TM ) was used as primary antibody and antibody Anti-rabbit IgG (Abnova TM ) was used as the secondary antibody. The results are shown in figure

41 Figure 4.30: Immunolocalization of the microtubules (MTs) in root hair cells of wild type (W.T.) and 35S:G2. MTs are stained in Green Figure 4.30 shows that the MT could be visualized using this technique but that the internal structure of the root hairs was disrupted and appeared to be shrunk. The reason could be that in this analysis, the fixing system for the tissue was not suitable for fixing root hairs. In a future study methods such as freeze substitution could be applied to overcome this issue Generation of a translational fusion by fusing G2 protein with GFP. In order to localize G2 protein at a cellular level, G2 protein was fused with the GFP marker. Fragments with 1841 base pairs covering the full length of G2 cdna and 1504 base paris of promoter area, were fused into an entry vector pdonr221 (Invitrogen TM ) by using a Gateway TM Technology (Figure 4.5). Because the gene does not have an intron, primers with attb1.1 and attb2.1 sites were designed and the amplified fragments from PCR by using 152
. The plasmids were then transformed into E.coli strain DH5α using a heat-shock method (Sambrook et al., 1989")
42 Phusion TM DNA Polymerase were purified before fusing them into the vector pdonr221 by the Gateway TM BP-reaction strategy which was described in section The resulting construct was called PrG2Entry (Figure 4.31). The plasmids were then transformed into E.coli strain DH5α using a heat-shock method (Sambrook et al., 1989) as described in section The transformed cells were selected by spreading the cells on LB medium plates containing 50 μg/ml kanamycin and the resistant plasmids were isolated and purified using a NucleoSpin TM Plasmid mini kit which has been described in section The plasmids were confirmed by PCR (Figure 4.32) and then sent for sequencing. Figure 4.31: Schematic diagram of G2Entry plasmid construct. The 1841 bp G2 genomic DNA insert was fused into pdonr TM 221 by BP reaction. A kanamycin-resistance gene was used as a selectable marker in bacteria. The diagrams are not drawn to scale. 153

43 Figure 4.32: PCR analysis of the colonies that contain the PrG2Entry plasmid using primers PrG2_Forward and M13_reverse. The positions of the primers are shown in figure L: ladder. 154


44 The full length fragment was then inserted into a destination vector pgwb4 which was described in section The Gateway TM technology LR reaction was then carried out following the procedure described in section To confirm that transformation was successful, a genomic PCR was carried out using G2_forward primer and GFP_reverse primer and a band with expected size was obtained. The final construct was named prg2:pgwb4. A diagrammatic representation of the construct is shown in Figure Prior to the transformation, the constructs both in E.coli DH5α and Agrobacterium tumefaciens C58 were confirmed by PCR using specific primers. Bands of the expected size were successfully amplified (Figure 4.34). Figure 4.33: The fragment in the Entry Clone PrG2Entry was transferred to a destination vector, pgwb4, which contains compatible recombination sites, attr1 and AttR2, in a reaction mediated by GatewayTM LR ClonaseTM Enzyme Mix (Invitrogen). The resulting construct was named PrG2:GFP. This construct is expected to fuse the GFP marker at the C terminal of G2 protein therefore to locate G2 protein in the cellular level. The diagrams are not drawn to scale. 155

45 Figure 4.34 PCR analysis of plasmids from the 4 colonies of transformed Agrobacterium tumefaciens C58. L: ladder. PCR products were amplified using primers PrG2_Forward & GFP_reverse. The position of primers is shown in Figure PrG2:GFP were obtained and transferred into pots containing Levington M3 compost and GFP expression examined during floral organ shedding. However no GFP signal was identified. The reason for this could be that the fusion of G2 protein and GFP disrupt the GFP structure so that no GFP signal could be visualised. To solve this issue, in a future study the GFP protein could be placed at the N terminal of the G2 protein. However, apart from antibiotic selection, there was no further confirmation (eg, genomic PCR, sequencing or expression analysis) to show that the construct was successfully transferred into the plants, therefore future studies, are needed to confirm the nature of the problems encountered. 156
46 4.4 Discussion Generation and analysis of lines down-regulated in the expression of G2. An insertional mutagenesis strategy was pursued to obtain KO lines of G2. The insertions were located 310 bp or 360 bp upstream of the translation start site. Although homozygous lines were isolated, RT-PCR results indicated that G2 expression was not affected by the insertion. The reason could be that both of the SALK line T-DNA insertions were located in G2 promoter area and the insertions did not disrupt the transcription initiation. No T-DNA insertion line was available in the exon region of G2 from the database searching ( Therefore an RNAi strategy was designed and implemented in order to knock down G2 expression. Two homozygous lines were confirmed by RT-PCR to be silenced. A phenotypic consequence of silencing G2 was found in developing pods. Seeds from the transgenic line were observed to be aborted in mature siliques of the RNAi:G2 homozygous plants compared to wild type under normal growing conditions. Statistical analysis in the development rate of seeds showed that only 25% of visible seed in siliques developed in siliques of RNAi:G2 lines compared to 92% in wild type. Study of the morphology of the undeveloped seeds indicated that the ovules were failed to develop. Genevestor_V3 showed that G2 was expressed in ovary and the level of 157
47 expression was medium, which suggested that G2 could play a role in ovary development. Interestingly, some pollen grains were found to be undeveloped in RNAi:G2 plants. G2 is expressed in the stamen in accordance with Genevestor_V3 data, suggesting that the gene may be involved in pollen development. G2 may be functional redundant with one or more of the other 9 genes in abscission process (chapter 3), which could also be the reason of that no abscission-related phenotype was detected in G2 null lines. The gene At1g10530, which is predicted by Genevestigator_V3 that it is expressed specifically in AZ with a medium level of expression, could be functional redundant with G2. To test this hypothesis, a double-ko of G2 and At1g10530 could be generated and the abscission properties of the material studied. 158
48 4.4.2 Overexpression of G2 in 35S:IDA background rescues the effects of 35S:IDA in floral organ abscission G2 has been shown to be expressed highly in AZ (Chapter 3). However no significant difference was observed in either the KO line or the overexpression line in terms of floral organ abscission comparing to wild type. Firstly, no abscission-related phenotype was detected in RNAi:G2 lines. The bioinformatics analysis described in chapter 3 revealed nine proteins in Arabidopsis that shared domain I and III with G2 and both of the domains were predicted to be potentially responsible for the function of G2. These genes could form a new family in Arabidopsis and be functionally redundant with G2, which could be the reason why there was no phenotype in mutants with the absence of G2. Another explanation is that G2 could be involved in the protection process against pathogens on cell wall synthesis, so that no physical phenotype could be detected. Secondly, no abscission-related phenotype was observed in 35S:G2 plants. The reporter gene and RT-PCR analysis showed that the expression level of G2 was high and exceeded the threshold in AZ in a wild type background, therefore overexpression of G2 might not make a significant difference. The expression analysis of G2 which has been described in chapter 3 has shown an inverse correlation between IDA and G2 with the wounding induced G2:GUS signal down-regulated by the overexpression of IDA. IDA 159
49 has been reported to play an important role in regulating floral organ abscission and absence of IDA leads to partial degradation of the middle lamella and an inhibition of abscission (Butenko et al. 2003). Overexpression of IDA leads to a series of phenotypes including early abscission, extended AZ development and secreted AGP. These features are easy to distinguish from the wild type (Stenvik et al., 2006), whereas in 35S:G2 plants no change in either the timing of abscission or the development of the AZ was observed. To further investigate the relation between G2 and IDA, a cross between 35S:G2 and 35S:IDA was carried out. Homozygotes of 35S:G2 x 35S:IDA were selected by PCR using 35S_Forward and G2/IDA_reverse primers (Table 1.1). The timing of abscission and the development of AZ were examined in the 35S:G2 x 35S:IDA homozygotes and the result showed that 35S:G2 x 35S:IDA homozygous plants did not exhibit the 35S:IDA phenotype. This observation suggests that overexpression of G2 can rescue the effects of 35S:IDA in terms of floral organ abscission. Compared to the wild type, homozygous plants did not show either an earlier abscission or an extended AZ region. In addition, vestigial AZs were not found at the bases of pedicels, branches in 35S:IDA x 35S:G2 plants and the partial shedding of siliques was also not observed. Finally, the plants retained a wild type size and appeared to be much healthier than 35S:IDA plants. The above observations indicate that overexpression of G2 rescues the effects of 35S:IDA. The RT-PCR result suggests that when ectopically expressed G2 160
50 is introduced into a 35S:IDA background, overexpressed IDA is down-regulated and the amount of expression of IDA was similar as in wild type background. This could be the reason why the 35S:G2 x 35S:IDA homozygote did not show a 35S:IDA phenotype. Interestingly, no, or a lower amount of AGPs was detected in 35S:G2 and 35S:G2 x 35S:IDA background compared to that observed in a 35S:IDA background. AGP24 was reported to be up-regulated in 35S:IDA plant and absent in ida mutant background (Stenvik et al., 2006). The function of AGPs in the abscission process is unknown but there are reports showing that AGPs may be involved in molecular interactions and cellular signalling (Showalter, 2001). In addition, AGPs were predicted to act as a mediator between the cell wall and cortical microtubules in Arabidopsis root tissue (Andeme-Onzighi et al., 2002). Whether AGPs are involved in the similar process during abscission remains unknown, however it would be worthwhile investigating microtubule orientation during abscission which could be achieved by an immunolocalization strategy. Leaves of the rosette leaf of the 35S:G2 x 35S:IDA cross display a reduction in width compared to wild type and this phenotype is prolonged until the plant becomes mature. This effect can not be detected in either 35S:G2 or 35S:IDA plants. In addition, 35S:IDA can partially recover the phenotype of the swollen root hairs in 35S:G2 (this observation will be discussed in the following sections), indicating that 35S:G2 can not fully 161
51 suppress the 35S:IDA phenotype. The RT-PCR analysis shows that G2 and IDA expression in 35S:G2 x 35S:IDA plants are down-regulated compared to 35S:G2 and 35S:IDA pants. This could be one of the reasons that the cross has a unique morphology of its rosette leaves Overexpression of G2 leads to swollen root hairs The overexpression lines of G2 were obtained by fusing G2 cdna with a 35SCaMV promoter. RT-PCR was performed in order to confirm the ectopic expression of 35S:G2. The 35S:G2 root hair was classified into three types based on the morphology. Type I hairs were extremely swollen and failed to transfer to tip growth. Type II hairs were short and vary in diameter along their length. Type III hairs were not only short but also crooked and display branches. Rachel and Dolan (2002) separated root hair development into four phases: (1) the specification of hair-producing cells; (2) the initiation of root hair growth; (3) tip growth and elongation; and (4) cessation of mature root hairs growth. In plants with ectopic expression of G2, the position and amount of root hairs was not affected, which indicates a change in the morphology of root hairs occurs under the impact of 35S:G2 and that the transfer to tip growth and the tip growth/elongation process is disrupted. Cellulose 162
52 microfibrils have been shown to control the direction of expansion (Green, 1962) and the deposition of microfibrils is believed to be under the control of the cytoskeleton (Heath & Seagull, 1982). Microtubule cytoskeleton has been reported to control the tip-growth of root hairs (Heath & Seagull 1982, Sieberer 2005). The mutant root epidermal bulger 1 (reb1-1) (Andeme-Onzighi et al., 2002) shows a similar phenotype to type I hairs in 35S:G2 plants with bulging trichoblasts and this has been shown to be accompanied by disrupted cortical microtubules. This mutant has been shown to lack arabinogalactan-protein (AGP). Although attempts to investigate the microtubules in root hairs using an immunolocalzation strategy were not successful, it is possible that overexpression of G2 leads to a loss of arabinogalactan-protein which could result in the bulging of root hairs. G2 may play a role in interaction with IDA and IDL genes In 35S:IDA plants, secreted AGPs were detected and this process was blocked by overexpressing G2. Our staining results with the chemical reagent β-glcy on 35S:G2 and 35S:G2 x 35S:IDA flowers showed that less AGPs were detected compared to the wild type in the AZ. RT-PCR results showed that overexpression of G2 resulted in a down-regulation of IDA and down-regulation of IDA leads to the absence of AGP in AZ. This AGP has been shown to be AGP24 (Stenvik et al. 2006). AGP24 is most highly expressed in pollen and highly expressed in root hair zone, endodermis and cortex, according to Genevestigator_V3 data ( (Figure 4.35). 163
53 Figure 4.35 Genevestigator V3 result of the expression value of AGP24 in tissues of Arabidopsis. AGPs have been reported to mediate interactions between the cell wall and cortical microtubules and the mutation of root epidermal bulger 1-1 (reb1-1) leads to a reduction of AGP in root, and a disruption of cortical microtubules which results in a bugling of trichoblasts (Andeme-Onzighi et al. 2002). It has been shown in chapter 3 that there is a negative correlation between the expression of G2 and IDA and the RT-PCR result presented in this chapter suggests that IDA is down-regulated by ectopically expressed G2. 164
54 Overexpression of G2 leads to bulging root hairs and a hypothesis to account for this may be a change in microtubule orientation. Given the evidence that IDA promotes secretion of AGPs and G2 down-regulates IDA, it is possible that the bulging of root hairs in 35S:G2 could be consequence of down-regulating IDA, which results in the absence of AGP in roots and a disruption in microtubule organisation. In the 21 st International Conference on Arabidopsis Research, Kumpf et al. (2010) showed that IDA played an important role in lateral root (LR) development despite the Genevestigator_V3 data suggesting a low expression of IDA in Arabidopsis root tissues (data not shown). The absence of IDA leads to a delay in lateral development (Kumpf et al., 2010). IDA has been shown to be auxin-induced and expressed in cortical cells overlying the primordial (Kumpf et al., 2010), and both of the features are shared with G2. However, in terms of root hair development, IDA is not expressed in epidermal cells and based on our observation, the IDA knock out mutants do not show a swelling in the root hairs, therefore it is not clear that the swelling of root hairs effect in 35S:G2 might be caused by a down-regulation of IDA. IDA encodes a novel family of putative ligands and in Arabidopsis there are 5 IDA like genes IDA-LIKE 1 (AtIDL1) to AtIDL5 (Butenko et al., 2003). Further analysis has shown that the effect of mutating IDA could be largely rescued in plants with AtIDL1 fused to the IDA promoter (Stenvik et al., 165
55 2008). AtIDL1 is strongly expressed in Arabidopsis root tissues (Genevestigator data Figure 4.36). Although there has been no report about phenotypic consequences of down-regulating AtIDL1, it might be predicted that the absence of AtIDL1 could lead to a similar effect as overexpression of G2. Figure 4.36 Genevestigator result of the expression value of AtIDL1 in tissues of Arabidopsis. 166
56 IDA has been reported to be functionally dependent on the presence of the receptor like protein kinase (RLKs) HAESA and HAESA-LIKE2 (HSL2) (Stenvik et al., 2008). IDA, HAESA and HSL2 are functional in the common pathway and IDA and IDL proteins are acting through RLKs in regulating other processes during plant development (Stenvik et al., 2008). However whether IDL genes are functional in the same pathway with HAESA and HSL2 is not yet known and there has been no report about the characterization of IDL genes. In addition, the bioinformatic analysis of G2 protein in chapter 3 suggested that G2 could contain a spectrin repeats like domain which acts to bind and interact with other proteins. Therefore it is possible that G2 play a role in binding proteins and involved in the IDA-HAE/HSL2 signalling pathway. To test this hypothesis, an RT-PCR could be performed to test the expression level of AtIDL1 in ectopically expressing G2 lines. Phenotypic analysis could also be carried out on the null lines of AtIDL1 plants to investigate whether there is change in the morphology of root hair development. Methods such as yeast two-hybrid could also be applied to investigate potential interactions between G2 and IDA/IDL proteins. The mutant kojak (kjk) also fails to transfer to tip growth and hairs become so expanded that they burst at the end (Favery et al., 2001). KOJAK was 167
57 suggested to be involved in the synthesis of cell wall polymers (Favery et al., 2001). However it is less likely that overexpression of G2 has an effect on this process for the reasons that type I hairs in 35S:G2 plants do not burst at the end and some of the hairs can successfully transfer to tip growth and elongation. Overexpression of G2 has an impact on the tip growth process of root hairs The phenotypic change in the initiation process of root hair development of 35S:G2 was similar to the mutant reb1-1 (rhd1), however reports of this mutant have also shown that the total length of the root hair was not affected (Schiefelbein and Somerville, 1990; Andeme-Onzighi et al., 2002). In 35S:G2 plants, the disruption of initiation of root hair growth is accompanied by an increase in the diameter of the root hair and a reduction compared to the wild type, suggesting that the process of root hair elongation and tip growth was affected by ectopically expressing G2. In type II hairs of 35S:G2 plants, bulges were observed along the length which causes the hairs to vary in diameter. Type III root hairs show branching and are sometimes crooked. The type II and III hairs are shorter than wild type but exceed the length of 40 µm which suggests that the transfer to tip growth has been successful. Microtubules (MTs) could be a reason for branching and crooked effects. Under normal condition, MTs are longitudinally organized and parallel to the direction of growth of a hair (Baluska et al., 2000; Bibikova et al., 1999). When an Arabidopsis root is 168
58 treated with MT stabilization drugs, hairs have been shown to be wavy and branched (Bibikova et al., 1999). New growth points were detected on the hair treated with MT stabilization drugs, which suggested that MT could be involved in determination of the apical position of calcium influx (Bibikova et al., 1999). Therefore the disruption of the MTs in 35S:G2 roots could be responsible for the effects observed. Future work might include immunolocalization strategy using anti-tublin as the primary antibody to investigate MT organisation in the 35S:G2 root hairs. It has been shown that the genes BST1, CEN1, CEN2, CEN3, RHD3, RHD4, COW1 and SCN1 play a role in controlling root hair shapes during tip growth (Grierson et al., 1997; Ryan et al., 1998; Galway et al., 1997; Parker et al., 2000). Root hairs in plants without the presence of these genes are short and often altered in morphology (Parker et al., 2000). Mutation of RHD3 or RHD4 results in wavy and crooked root hairs (Schiefelbein and Somerville, 1990). Mutation of CEN1, CEN2 or CEN3 results in curled root hairs. bst1 mutants displayed short and straight root hairs which in scn1 mutants are wide and curled. rhd3 mutants not only showed short and curled root hairs, but also a shorter root compared to wild type (Schiefelbein and Somerville, 1990). Plants without TIP1 displayed not only shorter root hair but also disrupted pollen tube growth compared to wild type (Schiefelbein et al., 1993). Some root hairs in 35S:G2 roots were observed to be crooked, but no wavy nor curled hair was detected, suggesting that the curve level was much lower than it was in cen, rhd3. 169
59 It has been shown that mutation of RHD4, COW1, CEN3 and SCN1 result in a similar phenotype to the type II and type III hairs identified in 35S:G2 plants. Further work to study the impact of ectopically expressing G2 could include a series of RT-PCRs to determine the expression of these genes. When overexpression of IDA was introduced into 35S:G2 background, the root hair phenotype was partially recued and no type I hairs were detected, while type II and III hairs were still apparent. An explanation for this observation might be that overexpression of IDA leads to the rescuing of AGPs, which rescues the disruption of cortical microtubules. Branched root hairs were sometimes detected in 35S:IDA roots (Figure 4.37), which suggested that overexpression of IDA may also have an impact on the tip growth and the elongation process of root hairs. 170

60 Figure 4.37 Root hair of 35S:IDA plant. Yellow arrow points to a normally developed hair. White arrow points to a branched hair. Scale bar: 500 µm During root development, a hair cell is initiated and then a bulge is formed before tip growth and elongation taking place. The two phases are distinct and could be controlled by independent pathways. Ectopic expression of G2 is likely to disrupt both phases, but whether 35S:G2 achieve this effect through controlling the expression of IDA and IDLs solely or through other root hair development related genes is unclear. Given the above information, a predicted model has been developed to describe this potential pathway (Figure 4.38) and future work has been designed to test the predicted model. 171
61 Firstly, ectopically expressed G2 down-regulates IDA or IDLs (possibly AtIDL1) in root, which prevents cortical AGPs from being secreted in the root cortical cells. A lack of cortical AGPs affects cortical and epidermal MTs, which disrupts bulge formation (type I hairs) (Figure 4.38A). To test this hypothesis, the expression of IDA and IDLs could be quantified by RT-PCR or QPCR using total RNA from root tissue; additionally, a yeast-two-hybrid strategy could be applied to investigate the interaction between G2 and IDA/IDLs. Secondly, overexpression of G2 leads to crooked, branched hairs, or hairs forming bulges along the length of the hair (type II and type III hairs). One explanation for this could be that ectopic expression of G2 disrupts the root hair MTs, which leads to these effects (Figure 4.38B). To test this, an MT could be immunolocalized in roots using an anti-tubulin antibody in wild type and 35S:G2 plants. Overexpression of G2 may also down-regulates some genes such as RHD4, COW1, CEN3 and SCN1 etc, which are critical for root hair development and this might lead to the formation of type II and type III hairs. An RT-PCR or QPCR could be undertaken on root tissue to investigate this possibility. 172

62 Figure 4.38 Proposed model of how overexpression of G2 leads to the swollen, crooked and branched phenotypes. suggests the gene is up-regulated and suggests the gene is down-regulated. 173
Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family
, HAESA, ERECTA-family") Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family GENES & DEVELOPMENT (2000) 14: 108 117 INTRODUCTION Flower Diagram INTRODUCTION Abscission In plant, the process by which a plant
Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family GENES & DEVELOPMENT (2000) 14: 108 117 INTRODUCTION Flower Diagram INTRODUCTION Abscission In plant, the process by which a plant
Supplementary Figure 1. Phenotype of the HI strain.
 Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
23-. Shoot and root development depend on ratio of IAA/CK
 Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Supplemental Data. Wang et al. (2014). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Supplemental Data. Perrella et al. (2013). Plant Cell /tpc
. Plant Cell /tpc") Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
GFP GAL bp 3964 bp
 Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
EXPRESSION OF THE FIS2 PROMOTER IN ARABIDOPSIS THALIANA
 EXPRESSION OF THE FIS2 PROMOTER IN ARABIDOPSIS THALIANA Item Type text; Electronic Thesis Authors Bergstrand, Lauren Janel Publisher The University of Arizona. Rights Copyright is held by the author. Digital
EXPRESSION OF THE FIS2 PROMOTER IN ARABIDOPSIS THALIANA Item Type text; Electronic Thesis Authors Bergstrand, Lauren Janel Publisher The University of Arizona. Rights Copyright is held by the author. Digital
Arabidopsis Class I KNOTTED-Like Homeobox Proteins Act Downstream in the IDA-HAE/HSL2 Floral Abscission Signaling Pathway C W OA
 This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated
 Relative conversion rates of different plant hormones to their glucosylated") Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated form by recombinant UGT74E2. The naturally occurring auxin
Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated form by recombinant UGT74E2. The naturally occurring auxin
Department of Molecular, Cellular, and Developmental Biology, University of Michigan, Ann Arbor, Michigan
 The Plant Cell, Vol. 20: 1805 1817, July 2008, www.plantcell.org ã 2008 American Society of Plant Biologists The EPIP Peptide of INFLORESCENCE DEFICIENT IN ABSCISSION Is Sufficient to Induce Abscission
The Plant Cell, Vol. 20: 1805 1817, July 2008, www.plantcell.org ã 2008 American Society of Plant Biologists The EPIP Peptide of INFLORESCENCE DEFICIENT IN ABSCISSION Is Sufficient to Induce Abscission
Nature Genetics: doi: /ng Supplementary Figure 1. The phenotypes of PI , BR121, and Harosoy under short-day conditions.
 Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Biological Roles of Cytokinins
 Direct Control of Shoot Meristem Activity by a Cytokinin-Activating Enzyme By Kurakawa et. Al. Published in Nature Presented by Boyana Grigorova Biological Roles of Cytokinins Cytokinins are positive regulators
Direct Control of Shoot Meristem Activity by a Cytokinin-Activating Enzyme By Kurakawa et. Al. Published in Nature Presented by Boyana Grigorova Biological Roles of Cytokinins Cytokinins are positive regulators
SUPPLEMENTARY INFORMATION
 reverse 3175 3175 F L C 318 318 3185 3185 319 319 3195 3195 315 8 1 315 3155 315 317 Supplementary Figure 3. Stability of expression of the GFP sensor constructs return to warm conditions. Semi-quantitative
reverse 3175 3175 F L C 318 318 3185 3185 319 319 3195 3195 315 8 1 315 3155 315 317 Supplementary Figure 3. Stability of expression of the GFP sensor constructs return to warm conditions. Semi-quantitative
Lipid transfer proteins confer resistance to trichothecenes
 Lipid transfer proteins confer resistance to trichothecenes John McLaughlin and Anwar Bin-Umer Tumer Laboratory National Fusarium Head Blight Forum December 6th, 2012 FY09-11: Identify trichothecene resistance
Lipid transfer proteins confer resistance to trichothecenes John McLaughlin and Anwar Bin-Umer Tumer Laboratory National Fusarium Head Blight Forum December 6th, 2012 FY09-11: Identify trichothecene resistance
Characterisation of abiotic stress inducible plant promoters and bacterial genes for osmotolerance using transgenic approach
 Characterisation of abiotic stress inducible plant promoters and bacterial genes for osmotolerance using transgenic approach ABSTRACT SUBMITTED TO JAMIA MILLIA ISLAMIA NEW DELHI IN PARTIAL FULFILMENT OF
Characterisation of abiotic stress inducible plant promoters and bacterial genes for osmotolerance using transgenic approach ABSTRACT SUBMITTED TO JAMIA MILLIA ISLAMIA NEW DELHI IN PARTIAL FULFILMENT OF
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Supplemental Data. Chen and Thelen (2010). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Data. Chen and Thelen (2010). Plant Cell 10.1105/tpc.109.071837 1 C Total 5 kg 20 kg 100 kg Transmission Image 100 kg soluble pdtpi-gfp Plastid (PDH-alpha) Mito (PDH-alpha) GFP Image vector
Supplemental Data. Chen and Thelen (2010). Plant Cell 10.1105/tpc.109.071837 1 C Total 5 kg 20 kg 100 kg Transmission Image 100 kg soluble pdtpi-gfp Plastid (PDH-alpha) Mito (PDH-alpha) GFP Image vector
Major Plant Hormones 1.Auxins 2.Cytokinins 3.Gibberelins 4.Ethylene 5.Abscisic acid
 Plant Hormones Lecture 9: Control Systems in Plants What is a Plant Hormone? Compound produced by one part of an organism that is translocated to other parts where it triggers a response in target cells
Plant Hormones Lecture 9: Control Systems in Plants What is a Plant Hormone? Compound produced by one part of an organism that is translocated to other parts where it triggers a response in target cells
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL
 GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
Supplemental material
 Supplemental material Table 1- Segregation analysis of sgt1a sgt1b double mutant plants Parental genotype No. of lines Normal seeds Phenotype Abnormal seeds Ratio of abnormal seeds (%) WT 3 171 2 1.16
Supplemental material Table 1- Segregation analysis of sgt1a sgt1b double mutant plants Parental genotype No. of lines Normal seeds Phenotype Abnormal seeds Ratio of abnormal seeds (%) WT 3 171 2 1.16
SUPPLEMENTARY INFORMATION
 Supplemental Methods Isolation and mapping of SPCH An EMS-mutagenized population of tmm-1 (Col);E1728 (an enhancer trap GFP marking guard cells) was created. M2 seeds were collected from M1 s planted in
Supplemental Methods Isolation and mapping of SPCH An EMS-mutagenized population of tmm-1 (Col);E1728 (an enhancer trap GFP marking guard cells) was created. M2 seeds were collected from M1 s planted in
Table S1 List of primers used for genotyping and qrt-pcr.
 Table S1 List of primers used for genotyping and qrt-pcr. genotyping! allele! ligomer*! 5'-sequence-3'! rice! d10-2! F! TTGGCTTTGCCTCGTTTC!!! R! AGCCTCCACTTGTACTGTG! Arabidopsis! max2-3, max2-4! F! ACTCTCTCCGACCTCCCTGACG!!!
Table S1 List of primers used for genotyping and qrt-pcr. genotyping! allele! ligomer*! 5'-sequence-3'! rice! d10-2! F! TTGGCTTTGCCTCGTTTC!!! R! AGCCTCCACTTGTACTGTG! Arabidopsis! max2-3, max2-4! F! ACTCTCTCCGACCTCCCTGACG!!!
** * * * Col-0 cau1 CAU1. Actin2 CAS. Actin2. Supplemental Figure 1. CAU1 affects calcium accumulation.
 Ca 2+ ug g -1 DW Ca 2+ ug g -1 DW Ca 2+ ug g -1 DW Supplemental Data. Fu et al. Plant Cell. (213). 1.115/tpc.113.113886 A 5 4 3 * Col- cau1 B 4 3 2 Col- cau1 ** * * ** C 2 1 25 2 15 1 5 Shoots Roots *
Ca 2+ ug g -1 DW Ca 2+ ug g -1 DW Ca 2+ ug g -1 DW Supplemental Data. Fu et al. Plant Cell. (213). 1.115/tpc.113.113886 A 5 4 3 * Col- cau1 B 4 3 2 Col- cau1 ** * * ** C 2 1 25 2 15 1 5 Shoots Roots *
Plant Growth and Development
 Plant Growth and Development Concept 26.1 Plants Develop in Response to the Environment Factors involved in regulating plant growth and development: 1. Environmental cues (e.g., day length) 2. Receptors
Plant Growth and Development Concept 26.1 Plants Develop in Response to the Environment Factors involved in regulating plant growth and development: 1. Environmental cues (e.g., day length) 2. Receptors
Supplemental Data. Perea-Resa et al. Plant Cell. (2012) /tpc
 /tpc") Supplemental Data. Perea-Resa et al. Plant Cell. (22)..5/tpc.2.3697 Sm Sm2 Supplemental Figure. Sequence alignment of Arabidopsis LSM proteins. Alignment of the eleven Arabidopsis LSM proteins. Sm and
Supplemental Data. Perea-Resa et al. Plant Cell. (22)..5/tpc.2.3697 Sm Sm2 Supplemental Figure. Sequence alignment of Arabidopsis LSM proteins. Alignment of the eleven Arabidopsis LSM proteins. Sm and
Last time: Obtaining information from a cloned gene
 Last time: Obtaining information from a cloned gene Objectives: 1. What is the biochemical role of the gene? 2. Where and when is the gene expressed (transcribed)? 3. Where and when is the protein made?
Last time: Obtaining information from a cloned gene Objectives: 1. What is the biochemical role of the gene? 2. Where and when is the gene expressed (transcribed)? 3. Where and when is the protein made?
Development 143: doi: /dev : Supplementary information
 Supplementary Materials and Methods Plant materials The mutants and transgenic plants used in the present study were as follows: E361 (from Alex Webb s laboratory); tmm-1, ptmm::tmm-gfp and flp-1 (from
Supplementary Materials and Methods Plant materials The mutants and transgenic plants used in the present study were as follows: E361 (from Alex Webb s laboratory); tmm-1, ptmm::tmm-gfp and flp-1 (from
Old FINAL EXAM BIO409/509 NAME. Please number your answers and write them on the attached, lined paper.
 Old FINAL EXAM BIO409/509 NAME Please number your answers and write them on the attached, lined paper. Gene expression can be regulated at several steps. Describe one example for each of the following:
Old FINAL EXAM BIO409/509 NAME Please number your answers and write them on the attached, lined paper. Gene expression can be regulated at several steps. Describe one example for each of the following:
DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Other cellular processes (17%)
 Signal transduction (2%) Development (2%) Other cellular processes (17%)") Fig. 35-24 Other metabolism (18%) DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Unknown (24%) Energy pathways (3%) Cell division and organization (3%) Transport (4%) Transcription
Fig. 35-24 Other metabolism (18%) DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Unknown (24%) Energy pathways (3%) Cell division and organization (3%) Transport (4%) Transcription
Cytokinin. Fig Cytokinin needed for growth of shoot apical meristem. F Cytokinin stimulates chloroplast development in the dark
 Cytokinin Abundant in young, dividing cells Shoot apical meristem Root apical meristem Synthesized in root tip, developing embryos, young leaves, fruits Transported passively via xylem into shoots from
Cytokinin Abundant in young, dividing cells Shoot apical meristem Root apical meristem Synthesized in root tip, developing embryos, young leaves, fruits Transported passively via xylem into shoots from
Supplemental Data. Fernández-Calvo et al. Plant Cell. (2011) /tpc
 /tpc") Supplemental Data. Fernández-Calvo et al. Plant Cell. (2011). 10.1105/tpc.110.080788 Supplemental Figure S1. Phylogenetic tree of MYC2-related proteins from Arabidopsis and other plants. Phenogram representation
Supplemental Data. Fernández-Calvo et al. Plant Cell. (2011). 10.1105/tpc.110.080788 Supplemental Figure S1. Phylogenetic tree of MYC2-related proteins from Arabidopsis and other plants. Phenogram representation
Heterosis and inbreeding depression of epigenetic Arabidopsis hybrids
 Heterosis and inbreeding depression of epigenetic Arabidopsis hybrids Plant growth conditions The soil was a 1:1 v/v mixture of loamy soil and organic compost. Initial soil water content was determined
Heterosis and inbreeding depression of epigenetic Arabidopsis hybrids Plant growth conditions The soil was a 1:1 v/v mixture of loamy soil and organic compost. Initial soil water content was determined
The Plant Cell, November. 2017, American Society of Plant Biologists. All rights reserved
 The Genetics of Floral Development Teaching Guide Overview The development of flowers in angiosperm plants provided a critical evolutionary advantage, allowing more options for pollen dispersal and seed
The Genetics of Floral Development Teaching Guide Overview The development of flowers in angiosperm plants provided a critical evolutionary advantage, allowing more options for pollen dispersal and seed
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DEVELOPMENTAL GENETICS OF ARABIDOPSIS THALIANA
 DEVELOPMENTAL GENETICS OF ARABIDOPSIS THALIANA CHASE BALLARD LINDA EAN HECTOR LOPEZ DR. JOANNA WERNER-FRACZEK IN COLLABORATION WITH DR. PATRICIA SPRINGER S LAB AT UCR AND ROBERT KOBLE PURPOSE OF RESEARCH
DEVELOPMENTAL GENETICS OF ARABIDOPSIS THALIANA CHASE BALLARD LINDA EAN HECTOR LOPEZ DR. JOANNA WERNER-FRACZEK IN COLLABORATION WITH DR. PATRICIA SPRINGER S LAB AT UCR AND ROBERT KOBLE PURPOSE OF RESEARCH
SUPPLEMENTARY INFORMATION
 Figure S1. Haploid plant produced by centromere-mediated genome elimination Chromosomes containing altered CENH3 in their centromeres (green dots) are eliminated after fertilization in a cross to wild
Figure S1. Haploid plant produced by centromere-mediated genome elimination Chromosomes containing altered CENH3 in their centromeres (green dots) are eliminated after fertilization in a cross to wild
Arabidopsis thaliana. Lucia Strader. Assistant Professor, Biology
 Arabidopsis thaliana Lucia Strader Assistant Professor, Biology Arabidopsis as a genetic model Easy to grow Small genome Short life cycle Self fertile Produces many progeny Easily transformed HIV E. coli
Arabidopsis thaliana Lucia Strader Assistant Professor, Biology Arabidopsis as a genetic model Easy to grow Small genome Short life cycle Self fertile Produces many progeny Easily transformed HIV E. coli
RNAi Suppression of AGAMOUS-like Genes Causes Field Sterility in Populus
 RNAi Suppression of AGAMOUS-like Genes Causes Field Sterility in Populus Haiwei Lu and Steven H. Strauss Oregon State University Forest Tree Workshop PAG XXVI, San Diego, CA, 2018 The containment issue
RNAi Suppression of AGAMOUS-like Genes Causes Field Sterility in Populus Haiwei Lu and Steven H. Strauss Oregon State University Forest Tree Workshop PAG XXVI, San Diego, CA, 2018 The containment issue
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2. thaliana. regulate senescence and floral organ abscission in Arabidopsis.
 Research article 4563 AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana Christine M. Ellis 1, Punita Nagpal 1, Jeffery C. Young 3,
Research article 4563 AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana Christine M. Ellis 1, Punita Nagpal 1, Jeffery C. Young 3,
Regulation of Phosphate Homeostasis by microrna in Plants
 Regulation of Phosphate Homeostasis by microrna in Plants Tzyy-Jen Chiou 1 *, Kyaw Aung 1,2, Shu-I Lin 1,3, Chia-Chune Wu 1, Su-Fen Chiang 1, and Chun-Lin Su 1 Abstract Upon phosphate (Pi) starvation,
Regulation of Phosphate Homeostasis by microrna in Plants Tzyy-Jen Chiou 1 *, Kyaw Aung 1,2, Shu-I Lin 1,3, Chia-Chune Wu 1, Su-Fen Chiang 1, and Chun-Lin Su 1 Abstract Upon phosphate (Pi) starvation,
CHAPTER : Prokaryotic Genetics
 CHAPTER 13.3 13.5: Prokaryotic Genetics 1. Most bacteria are not pathogenic. Identify several important roles they play in the ecosystem and human culture. 2. How do variations arise in bacteria considering
CHAPTER 13.3 13.5: Prokaryotic Genetics 1. Most bacteria are not pathogenic. Identify several important roles they play in the ecosystem and human culture. 2. How do variations arise in bacteria considering
Ti plasmid derived plant vector systems: binary and co - integrative vectors transformation process; regeneration of the transformed lines
 Ti plasmid derived plant vector systems: binary and co - integrative vectors transformation process; regeneration of the transformed lines Mitesh Shrestha Constraints of Wild type Ti/Ri-plasmid Very large
Ti plasmid derived plant vector systems: binary and co - integrative vectors transformation process; regeneration of the transformed lines Mitesh Shrestha Constraints of Wild type Ti/Ri-plasmid Very large
Bio 100 Guide 27.
 Bio 100 Guide 27 http://www.offthemarkcartoons.com/cartoons/1994-11-09.gif http://www.cneccc.edu.hk/subjects/bio/album/chapter20/images/plant_growth.jpg http://pgjennielove.files.wordpress.com/2008/06/apical_meristem.png
Bio 100 Guide 27 http://www.offthemarkcartoons.com/cartoons/1994-11-09.gif http://www.cneccc.edu.hk/subjects/bio/album/chapter20/images/plant_growth.jpg http://pgjennielove.files.wordpress.com/2008/06/apical_meristem.png
CONTROL SYSTEMS IN PLANTS
 AP BIOLOGY PLANTS FORM & FUNCTION ACTIVITY #5 NAME DATE HOUR CONTROL SYSTEMS IN PLANTS HORMONES MECHANISM FOR HORMONE ACTION Plant Form and Function Activity #5 page 1 CONTROL OF CELL ELONGATION Plant
AP BIOLOGY PLANTS FORM & FUNCTION ACTIVITY #5 NAME DATE HOUR CONTROL SYSTEMS IN PLANTS HORMONES MECHANISM FOR HORMONE ACTION Plant Form and Function Activity #5 page 1 CONTROL OF CELL ELONGATION Plant
Penghui Li, Beibei Chen, Gaoyang Zhang, Longxiang Chen, Qiang Dong, Jiangqi Wen, Kirankumar S. Mysore and Jian Zhao
 New Phytologist Supporting Information Regulation of anthocyanin and proanthocyanidin biosynthesis by Medicago truncatula bhlh transcription factor MtTT8 Penghui Li, Beibei Chen, Gaoyang Zhang, Longxiang
New Phytologist Supporting Information Regulation of anthocyanin and proanthocyanidin biosynthesis by Medicago truncatula bhlh transcription factor MtTT8 Penghui Li, Beibei Chen, Gaoyang Zhang, Longxiang
10/4/2017. Chapter 39
 Chapter 39 1 Reception 1 Reception 2 Transduction CYTOPLASM CYTOPLASM Cell wall Plasma membrane Phytochrome activated by light Cell wall Plasma membrane Phytochrome activated by light cgmp Second messenger
Chapter 39 1 Reception 1 Reception 2 Transduction CYTOPLASM CYTOPLASM Cell wall Plasma membrane Phytochrome activated by light Cell wall Plasma membrane Phytochrome activated by light cgmp Second messenger
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
Reproduction, Seeds and Propagation
 Reproduction, Seeds and Propagation Diploid (2n) somatic cell Two diploid (2n) somatic cells Telophase Anaphase Metaphase Prophase I One pair of homologous chromosomes (homologues) II Homologues condense
Reproduction, Seeds and Propagation Diploid (2n) somatic cell Two diploid (2n) somatic cells Telophase Anaphase Metaphase Prophase I One pair of homologous chromosomes (homologues) II Homologues condense
Useful Propagation Terms. Propagation The application of specific biological principles and concepts in the multiplication of plants.
 Useful Propagation Terms Propagation The application of specific biological principles and concepts in the multiplication of plants. Adventitious Typically describes new organs such as roots that develop
Useful Propagation Terms Propagation The application of specific biological principles and concepts in the multiplication of plants. Adventitious Typically describes new organs such as roots that develop
Genetic Characterization and Functional Analysis of the GID1 Gibberellin Receptors in Arabidopsis W
 The Plant Cell, Vol. 18, 3399 3414, December 2006, www.plantcell.org ª 2006 American Society of Plant Biologists Genetic Characterization and Functional Analysis of the GID1 Gibberellin Receptors in Arabidopsis
The Plant Cell, Vol. 18, 3399 3414, December 2006, www.plantcell.org ª 2006 American Society of Plant Biologists Genetic Characterization and Functional Analysis of the GID1 Gibberellin Receptors in Arabidopsis
Sporic life cycles involve 2 types of multicellular bodies:
 Chapter 3- Human Manipulation of Plants Sporic life cycles involve 2 types of multicellular bodies: -a diploid, spore-producing sporophyte -a haploid, gamete-producing gametophyte Sexual Reproduction in
Chapter 3- Human Manipulation of Plants Sporic life cycles involve 2 types of multicellular bodies: -a diploid, spore-producing sporophyte -a haploid, gamete-producing gametophyte Sexual Reproduction in
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING. AnitaHajdu. Thesis of the Ph.D.
 THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING AnitaHajdu Thesis of the Ph.D. dissertation Supervisor: Dr. LászlóKozma-Bognár - senior research associate Doctoral
THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING AnitaHajdu Thesis of the Ph.D. dissertation Supervisor: Dr. LászlóKozma-Bognár - senior research associate Doctoral
Utilizing Illumina high-throughput sequencing technology to gain insights into small RNA biogenesis and function
 Utilizing Illumina high-throughput sequencing technology to gain insights into small RNA biogenesis and function Brian D. Gregory Department of Biology Penn Genome Frontiers Institute University of Pennsylvania
Utilizing Illumina high-throughput sequencing technology to gain insights into small RNA biogenesis and function Brian D. Gregory Department of Biology Penn Genome Frontiers Institute University of Pennsylvania
By Jonathan I. Watkinson. Virginia Polytechnic Institute and State University. Doctor of Philosophy Horticulture
 Characterization of two genes, trehalose-6-phosphate synthase/phosphatase and nucleotide binding protein, shown to be differentially regulated in roots of Cypripedium parviflorum var. pubescens grown with
Characterization of two genes, trehalose-6-phosphate synthase/phosphatase and nucleotide binding protein, shown to be differentially regulated in roots of Cypripedium parviflorum var. pubescens grown with
. Supplementary Information
 . Supplementary Information Supplementary Figure S1. Mature embryo sac observations. Supplementary Figure S2. STT observations. Supplementary Figure S3. Comparison of the PTB1 cdna with that of the mutant.
. Supplementary Information Supplementary Figure S1. Mature embryo sac observations. Supplementary Figure S2. STT observations. Supplementary Figure S3. Comparison of the PTB1 cdna with that of the mutant.
Transitivity-dependent and transitivity-independent cell-to-cell movement of RNA
 Himber et al. Transitivity-dependent and transitivity-independent cell-to-cell movement of RNA silencing SUPPLEMENTAL MATERIAL Supplemental material 1. Short-range movement of GFP silencing affects a nearly
Himber et al. Transitivity-dependent and transitivity-independent cell-to-cell movement of RNA silencing SUPPLEMENTAL MATERIAL Supplemental material 1. Short-range movement of GFP silencing affects a nearly
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
Arabidopsis COMPASS-Like Complexes Mediate Histone H3 Lysine-4 Trimethylation to Control Floral Transition and Plant Development
 Arabidopsis COMPASS-Like Complexes Mediate Histone H3 Lysine-4 Trimethylation to Control Floral Transition and Plant Development Danhua Jiang 1,2, Nicholas C. Kong 1,2, Xiaofeng Gu 2, Zicong Li 1, Yuehui
Arabidopsis COMPASS-Like Complexes Mediate Histone H3 Lysine-4 Trimethylation to Control Floral Transition and Plant Development Danhua Jiang 1,2, Nicholas C. Kong 1,2, Xiaofeng Gu 2, Zicong Li 1, Yuehui
Supplementary Figure 1 Characterization of wild type (WT) and abci8 mutant in the paddy field.
 and abci8 mutant in the paddy field.") Supplementary Figure 1 Characterization of wild type (WT) and abci8 mutant in the paddy field. A, Phenotypes of 30-day old wild-type (WT) and abci8 mutant plants grown in a paddy field under normal sunny
Supplementary Figure 1 Characterization of wild type (WT) and abci8 mutant in the paddy field. A, Phenotypes of 30-day old wild-type (WT) and abci8 mutant plants grown in a paddy field under normal sunny
Genetic interaction and phenotypic analysis of the Arabidopsis MAP kinase pathway mutations mekk1 and mpk4 suggests signaling pathway complexity
 Genetic interaction and phenotypic analysis of the Arabidopsis MAP kinase pathway mutations mekk1 and mpk4 suggests signaling pathway complexity Shih-Heng Su, Maria Cristina Suarez-Rodriguez, Patrick Krysan
Genetic interaction and phenotypic analysis of the Arabidopsis MAP kinase pathway mutations mekk1 and mpk4 suggests signaling pathway complexity Shih-Heng Su, Maria Cristina Suarez-Rodriguez, Patrick Krysan
Plant transformation
 Plant transformation Objectives: 1. What is plant transformation? 2. What is Agrobacterium? How and why does it transform plant cells? 3. How is Agrobacterium used as a tool in molecular genetics? References:
Plant transformation Objectives: 1. What is plant transformation? 2. What is Agrobacterium? How and why does it transform plant cells? 3. How is Agrobacterium used as a tool in molecular genetics? References:
Edward M. Golenberg Wayne State University Detroit, MI
 Edward M. Golenberg Wayne State University Detroit, MI Targeting Phragmites Success As Invasive Species Phragmites displays multiple life history parameters High seed output and small seed size Rapid growth
Edward M. Golenberg Wayne State University Detroit, MI Targeting Phragmites Success As Invasive Species Phragmites displays multiple life history parameters High seed output and small seed size Rapid growth
A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development
 A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development Xuemei Chen Waksman Institute, Rutgers University, Piscataway, NJ 08854, USA. E-mail: xuemei@waksman.rutgers.edu Plant
A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development Xuemei Chen Waksman Institute, Rutgers University, Piscataway, NJ 08854, USA. E-mail: xuemei@waksman.rutgers.edu Plant
Three different fusions led to three basic ideas: 1) If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes
 If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes") Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Supplemental Data. Hou et al. (2016). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Data. Hou et al. (216). Plant Cell 1.115/tpc.16.295 A Distance to 1 st nt of start codon Distance to 1 st nt of stop codon B Normalized PARE abundance 8 14 nt 17 nt Frame1 Arabidopsis inflorescence
Supplemental Data. Hou et al. (216). Plant Cell 1.115/tpc.16.295 A Distance to 1 st nt of start codon Distance to 1 st nt of stop codon B Normalized PARE abundance 8 14 nt 17 nt Frame1 Arabidopsis inflorescence
Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed
 Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed using the Geneious software. Accession numbers of the
Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed using the Geneious software. Accession numbers of the
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
GCD3033:Cell Biology. Transcription
 Transcription Transcription: DNA to RNA A) production of complementary strand of DNA B) RNA types C) transcription start/stop signals D) Initiation of eukaryotic gene expression E) transcription factors
Transcription Transcription: DNA to RNA A) production of complementary strand of DNA B) RNA types C) transcription start/stop signals D) Initiation of eukaryotic gene expression E) transcription factors
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Plant Structure, Growth, and Development
 Plant Structure, Growth, and Development Plant hierarchy: Cells Tissue: group of similar cells with similar function: Dermal, Ground, Vascular Organs: multiple kinds of tissue, very diverse function Organ
Plant Structure, Growth, and Development Plant hierarchy: Cells Tissue: group of similar cells with similar function: Dermal, Ground, Vascular Organs: multiple kinds of tissue, very diverse function Organ
Supplemental Figure 1: Increased Fe deficiency gene expression in roots of nas4x-2
 Supplemental Figure 1: Increased Fe deficiency gene expression in roots of nas4x-2 IRT1, FRO2 and FIT expression levels in roots of the wild-type, nas4x- 1 and nas4x-2, showing that in both nas mutants
Supplemental Figure 1: Increased Fe deficiency gene expression in roots of nas4x-2 IRT1, FRO2 and FIT expression levels in roots of the wild-type, nas4x- 1 and nas4x-2, showing that in both nas mutants
A Mutation in MRH2 Kinesin Enhances the Root Hair Tip Growth Defect Caused by Constitutively Activated ROP2 Small GTPase in Arabidopsis
 City University of New York (CUNY) CUNY Academic Works Publications and Research Lehman College October 2007 A Mutation in MRH2 Kinesin Enhances the Root Hair Tip Growth Defect Caused by Constitutively
City University of New York (CUNY) CUNY Academic Works Publications and Research Lehman College October 2007 A Mutation in MRH2 Kinesin Enhances the Root Hair Tip Growth Defect Caused by Constitutively
Actions of auxin. Hormones: communicating with chemicals History: Discovery of a growth substance (hormone- auxin)
") Hormones: communicating with chemicals History- discovery of plant hormone. Auxin Concepts of hormones Auxin levels are regulated by synthesis/degradation, transport, compartmentation, conjugation. Polar
Hormones: communicating with chemicals History- discovery of plant hormone. Auxin Concepts of hormones Auxin levels are regulated by synthesis/degradation, transport, compartmentation, conjugation. Polar
Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering
 Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering by Valverde et. Al Published in Science 2004 Presented by Boyana Grigorova CBMG 688R Feb. 12, 2007 Circadian Rhythms: The Clock Within
Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering by Valverde et. Al Published in Science 2004 Presented by Boyana Grigorova CBMG 688R Feb. 12, 2007 Circadian Rhythms: The Clock Within
Introduction to Molecular and Cell Biology
 Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the molecular basis of disease? What
Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the molecular basis of disease? What
Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha
 Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha Plasmids 1. Extrachromosomal DNA, usually circular-parasite 2. Usually encode ancillary
Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha Plasmids 1. Extrachromosomal DNA, usually circular-parasite 2. Usually encode ancillary
Other funding Sources Agency Name: MSU Agricultural Experiment Station /Project GREEEN Amount requested or awarded: 30,000
 FINAL PROJECT REPORT Project Title: Functional genomics of flowering in apple PI: Herb Aldwinckle Co-PI(2): Steve VanNocker Organization: Cornell University Organization: Michigan State University Telephone/email:
FINAL PROJECT REPORT Project Title: Functional genomics of flowering in apple PI: Herb Aldwinckle Co-PI(2): Steve VanNocker Organization: Cornell University Organization: Michigan State University Telephone/email:
A Genome-Wide Analysis of Arabidopsis Rop-Interactive CRIB Motif Containing Proteins That Act as Rop GTPase Targets
 The Plant Cell, Vol. 13, 2841 2856, December 2001, www.plantcell.org 2001 American Society of Plant Biologists A Genome-Wide Analysis of Arabidopsis Rop-Interactive CRIB Motif Containing Proteins That
The Plant Cell, Vol. 13, 2841 2856, December 2001, www.plantcell.org 2001 American Society of Plant Biologists A Genome-Wide Analysis of Arabidopsis Rop-Interactive CRIB Motif Containing Proteins That
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/331/6019/876/dc1 Supporting Online Material for Synthetic Clonal Reproduction Through Seeds Mohan P. A. Marimuthu, Sylvie Jolivet, Maruthachalam Ravi, Lucie Pereira,
www.sciencemag.org/cgi/content/full/331/6019/876/dc1 Supporting Online Material for Synthetic Clonal Reproduction Through Seeds Mohan P. A. Marimuthu, Sylvie Jolivet, Maruthachalam Ravi, Lucie Pereira,
2012 Univ Aguilera Lecture. Introduction to Molecular and Cell Biology
 2012 Univ. 1301 Aguilera Lecture Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the
2012 Univ. 1301 Aguilera Lecture Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the
SMALL AUXIN UP RNA (SAUR) genes are the largest class of primary auxin (growth
 genes are the largest class of primary auxin (growth") Abstract SMALL AUXIN UP RNA (SAUR) genes are the largest class of primary auxin (growth hormone) responsive genes in all land plants, many of which are expressed in actively growing plant tissues. This
Abstract SMALL AUXIN UP RNA (SAUR) genes are the largest class of primary auxin (growth hormone) responsive genes in all land plants, many of which are expressed in actively growing plant tissues. This
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Ethylene is critical to the maintenance of primary root growth and Fe. homeostasis under Fe stress in Arabidopsis
 Ethylene is critical to the maintenance of primary root growth and Fe homeostasis under Fe stress in Arabidopsis Guangjie Li, Weifeng Xu, Herbert J. Kronzucker, Weiming Shi * Supplementary Data Supplementary
Ethylene is critical to the maintenance of primary root growth and Fe homeostasis under Fe stress in Arabidopsis Guangjie Li, Weifeng Xu, Herbert J. Kronzucker, Weiming Shi * Supplementary Data Supplementary
2. Yeast two-hybrid system
 2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
Reading Assignments. A. Genes and the Synthesis of Polypeptides. Lecture Series 7 From DNA to Protein: Genotype to Phenotype
 Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Model plants and their Role in genetic manipulation. Mitesh Shrestha
 Model plants and their Role in genetic manipulation Mitesh Shrestha Definition of Model Organism Specific species or organism Extensively studied in research laboratories Advance our understanding of Cellular
Model plants and their Role in genetic manipulation Mitesh Shrestha Definition of Model Organism Specific species or organism Extensively studied in research laboratories Advance our understanding of Cellular
Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice Supplementary Figure 2.
 Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice. Percentage of marginal zone B cells in the spleen of wild-type mice (+/+), mice homozygous for cpm or pri
Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice. Percentage of marginal zone B cells in the spleen of wild-type mice (+/+), mice homozygous for cpm or pri
Anatomy of Plants Student Notes
 Directions: Fill in the blanks. Anatomy of Plants Student Notes Plant Cell Biology Segment 1. Plants Plants are organisms are incapable of movement produce food through 2. Animals Animals are multicellular
Directions: Fill in the blanks. Anatomy of Plants Student Notes Plant Cell Biology Segment 1. Plants Plants are organisms are incapable of movement produce food through 2. Animals Animals are multicellular
Plant Development. Chapter 31 Part 1
 Plant Development Chapter 31 Part 1 Impacts, Issues Foolish Seedlings, Gorgeous Grapes Gibberellin and other plant hormones control the growth and development of plants environmental cues influence hormone
Plant Development Chapter 31 Part 1 Impacts, Issues Foolish Seedlings, Gorgeous Grapes Gibberellin and other plant hormones control the growth and development of plants environmental cues influence hormone
EST1 Homology Domain. 100 aa. hest1a / SMG6 PIN TPR TPR. Est1-like DBD? hest1b / SMG5. TPR-like TPR. a helical. hest1c / SMG7.
 hest1a / SMG6 EST1 Homology Domain 100 aa 853 695 761 780 1206 hest1 / SMG5 -like? -like 109 145 214 237 497 165 239 1016 114 207 212 381 583 hest1c / SMG7 a helical 1091 Sc 57 185 267 284 699 Figure S1:
hest1a / SMG6 EST1 Homology Domain 100 aa 853 695 761 780 1206 hest1 / SMG5 -like? -like 109 145 214 237 497 165 239 1016 114 207 212 381 583 hest1c / SMG7 a helical 1091 Sc 57 185 267 284 699 Figure S1:
Supporting Information
 Supporting Information Cao et al. 10.1073/pnas.1306220110 Gram - bacteria Hemolymph Cytoplasm PGRP-LC TAK1 signalosome Imd dfadd Dredd Dnr1 Ikk signalosome P Relish Nucleus AMP and effector genes Fig.
Supporting Information Cao et al. 10.1073/pnas.1306220110 Gram - bacteria Hemolymph Cytoplasm PGRP-LC TAK1 signalosome Imd dfadd Dredd Dnr1 Ikk signalosome P Relish Nucleus AMP and effector genes Fig.
Figure 18.1 Blue-light stimulated phototropism Blue light Inhibits seedling hypocotyl elongation
 Blue Light and Photomorphogenesis Q: Figure 18.3 Blue light responses - phototropsim of growing Corn Coleoptile 1. How do we know plants respond to blue light? 2. What are the functions of multiple BL
Blue Light and Photomorphogenesis Q: Figure 18.3 Blue light responses - phototropsim of growing Corn Coleoptile 1. How do we know plants respond to blue light? 2. What are the functions of multiple BL
Host-Pathogen Interaction. PN Sharma Department of Plant Pathology CSK HPKV, Palampur
 Host-Pathogen Interaction PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 PATHOGEN DEFENCE IN PLANTS A BIOLOGICAL AND MOLECULAR VIEW Two types of plant resistance response to potential
Host-Pathogen Interaction PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 PATHOGEN DEFENCE IN PLANTS A BIOLOGICAL AND MOLECULAR VIEW Two types of plant resistance response to potential
Biology 112 Practice Midterm Questions
 Biology 112 Practice Midterm Questions 1. Identify which statement is true or false I. Bacterial cell walls prevent osmotic lysis II. All bacterial cell walls contain an LPS layer III. In a Gram stain,
Biology 112 Practice Midterm Questions 1. Identify which statement is true or false I. Bacterial cell walls prevent osmotic lysis II. All bacterial cell walls contain an LPS layer III. In a Gram stain,
SUPPLEMENTARY INFORMATION
 Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
CONTROL OF GROWTH BY HORMONES
 CONTROL OF GROWTH BY HORMONES Growth and organogenesis are controlled......by genes (independent of environment): e.g., number of primary vascular bundles, general shape of a leaf or flower...by genes
CONTROL OF GROWTH BY HORMONES Growth and organogenesis are controlled......by genes (independent of environment): e.g., number of primary vascular bundles, general shape of a leaf or flower...by genes