Journal of Cell Science Accepted manuscript
|
|
|
- Rose Adams
- 5 years ago
- Views:
Transcription
1 2015. Published by The Company of Biologists Ltd. Bit-1 is an essential regulator of myogenic differentiation Genevieve S. Griffiths a, Jinger Doe b, Mayumi Jijiwa c, Pam Van Ry b, Vivian Cruz b, Michelle de la Vega c, Joe W. Ramos c, Dean J. Burkin b and Michelle L. Matter a,c,1 a John A. Burns School of Medicine University of Hawaii Honolulu, HI USA b Department of Pharmacology University of Nevada Medical School Reno, NV USA c University of Hawaii Cancer Center University of Hawaii Honolulu, HI USA 1 To whom correspondence should be addressed: Michelle L. Matter University of Hawaii Cancer Center Honolulu, HI USA matter@hawaii.edu Voice: Running Title: Bit-1 regulates muscle differentiation JCS Advance Online Article. Posted on 13 March 2015
2 Summary Muscle differentiation requires a complex signaling cascade that leads to multinucleated myofibers. Genes regulating the intrinsic mitochondrial apoptotic pathway also function in controlling cell differentiation. How such signaling pathways are regulated during differentiation is not fully understood. Bit-1 mutations in humans cause infantile-onset multisystem disease with muscle weakness. We demonstrate here that Bit-1 controls skeletal myogenesis through a caspase-mediated signaling pathway. Bit-1 null mice exhibit a myopathy with hypotrophic myofibers. Bit-1 null myoblasts prematurely express muscle-specific proteins. Similarly, knockdown of Bit-1 expression in C2C12 myoblasts promotes early differentiation whereas overexpression delays differentiation. In wild type mice Bit-1 levels increase during differentiation. Bit-1 null myoblasts exhibited increased caspase 9 and caspase 3 without increased apoptosis*. Bit-1 reexpression partially rescued differentiation. In Bit-1 null muscle, Bcl-2 is reduced suggesting Bcl-2-mediated inhibition of caspase 9 and caspase 3 is decreased. Bcl-2 re-expression rescued Bit-1-mediated early differentiation in Bit-1 null myoblasts and C2C12s with knockdown of Bit-1 expression. These results support an unanticipated yet essential role for Bit-1 in controlling myogenesis through regulation of Bcl-2.
3 Introduction Bit-1 (also known as Peptidyl-tRNA hydrolase 2; Ptrh2) is part of an integrin-specific signaling pathway responsible for the survival effects of cell-extracellular matrix interactions (Griffiths et al., 2011; Jan et al., 2004). It mediates attachment-dependent cell survival through an interaction with FAK and subsequent activation of the PI3K- AKT-NFkB pathway. Upon integrin-mediated attachment Bit-1 expression induces bcl-2 transcription (Griffiths et al., 2011) and blocks the intrinsic mitochondrial apoptotic pathway. When integrins are not ligated Bit-1 complexes with Groucho/TLE to promote apoptosis in a variety of cell types including myoblasts (Bouchentouf et al., 2007; Jan et al., 2004). Bit-1 is phosphorylated by PKD1 and Bit-1 expression impairs ERK phosphorylation (Biliran et al., 2008; Kairouz-Wahbe et al., 2008). Thus, Bit-1 is a phosphoprotein that can modulate NFkB and ERK signaling and thereby regulate integrin-mediated apoptotic signals. Initial characterization of Bit-1 null mice revealed that they are smaller than wildtype littermates, develop a runting syndrome and die within the first 2 weeks of life. Indeed, normal development is impaired in the Bit-1 nulls as kidney subcapsular glomeruli and epaxial muscle fiber diameters are smaller than controls (Kairouz-Wahbe et al., 2008). Recently we reported that a homozygous mutation in the Bit-1 gene causes infantileonset multisystem neurologic, endocrine and pancreatic disease (IMNEPD) with corresponding muscle weakness (Hu et al., 2014). The more severely affected patient became wheel chair dependent due to muscle weakness and ataxia by age 15.
4 However, the physiological function of Bit-1 has not been fully investigated in any tissue including skeletal muscle. Skeletal muscle differentiation is a multi-step process that requires myoblast progenitor cells to withdraw from the cell cycle, commit to a myogenic phenotype and differentiate into multinucleated myofibers that comprise mature muscle cells (Anderson, 1998; Grounds, 1991; Lluis et al., 2006a). Signal transduction pathways that regulate apoptosis are also crucial in controlling cell differentiation. For example, caspase 3 activation is necessary for the maturation of skeletal muscle (Fernando and Megeney, 2007; Larsen et al., 2010; Li and Yuan, 2008). Caspase 3 may also regulate differentiation indirectly by targeting and inactivating proteins that are involved in stemcell renewal (Fujita et al., 2008; Janzen et al., 2008). Other apoptotic regulators also play a role in muscle differentiation including Bcl-xL and the PI3K/AKT pathway (Briata et al., 2011; Majmundar et al., 2011; Murray et al., 2008; Wilson and Rotwein, 2007). The mechanism that activates these biochemical pathways in a non-death setting are not yet completely elucidated. Here we investigated the physiological function of Bit-1 using KO mice. We find that Bit- 1 levels increased during differentiation in wild type mice suggesting that Bit-1 plays a key role in myogenesis. Indeed, Bit-1 null mice exhibited a myopathy with hypotrophic myofibers without affecting apoptosis. This myopathy is due to early differentiation that occurs in the absence of Bit-1 expression. Both Bit-1 and its downstream effector Bcl-2 rescued Bit-1 mediated early differentiation. These findings reveal that Bit-1, which
5 regulates integrin-mediated apoptosis, can be engaged in the completely different function of muscle differentiation.
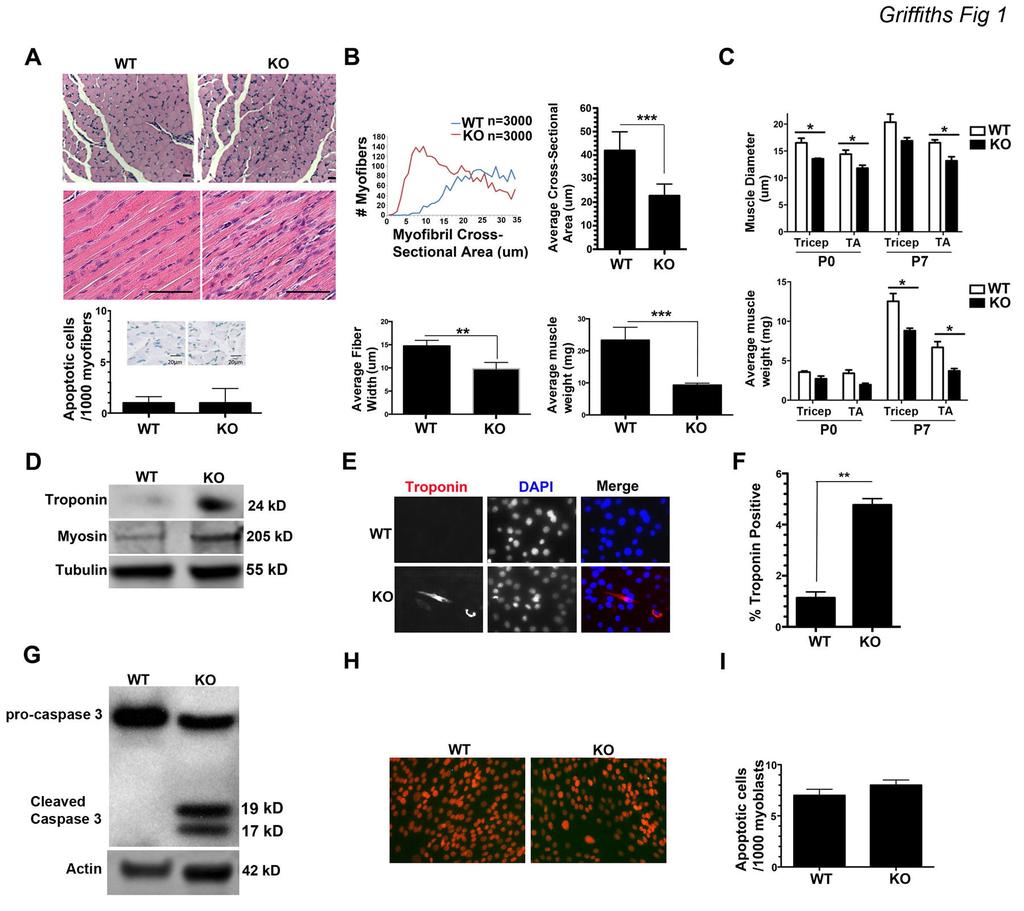
6 Results Loss of Bit-1 Promotes Early Myogenesis We investigated in detail the phenotype of Bit-1 null mice with particular focus on skeletal muscle. Bit-1 knockouts (KO) display reduced muscle mass and smaller gastrocnemius muscle fibers. As such, we reasoned that myogenesis may be altered in Bit-1 KOs. H&E stained gastrocnemius muscle tissue from Bit-1 KO mice at P7 demonstrated smaller myofibers and increased number of nuclei (Fig. 1A). Because Bit- 1 mediates cell survival and apoptosis in tumor cells (Griffiths et al., 2011; Jan et al., 2004), we examined apoptosis in KO and WT tissue sections by TUNEL staining. Apoptosis was similar in KO and WT gastrocnemius muscle tissue at P7 (Fig. 1A lower graph). Average myofiber cross-sectional area and longitudinal fiber width were significantly decreased in Bit-1 KO gastrocnemius muscle tissue compared to agematched WT littermates at P7 (Fig. 1B). Muscle diameter, as measured by Feret s diameter, was significantly decreased in Bit-1 KO tibialis anterior (TA) muscle at P0 and P7 and in tricep muscle at P0 (Fig. 1C top graph). The average tricep and TA muscle weights at P7 were significantly decreased in the KO mice compared to age-matched WT littermates (Fig. 1C lower graph). The smaller myofibers and increased nuclei observed in Bit-1 KO tissue may be due to abnormal differentiation. Therefore, to examine the effect of Bit-1 ablation on myogenesis, primary myoblast cultures were generated from 7 day-old Bit-1 KO and WT mice. Upon isolation of KO myoblasts and prior to induction of differentiation, the expression of the differentiation specific proteins, troponin T and myosin II, were substantially increased in Bit-1 KO myoblasts relative to WT (Fig. 1D). Troponin positive cells were significantly increased in Bit-1 KO myoblasts
7 compared to WT at day 0 pre-differentiation (Fig. 1E,F). These findings suggest that loss of Bit-1 was permissive to inducing myoblast differentiation irrespective of growthstimulating conditions (i.e. high serum). Caspase 3 Activity Is Increased In Bit-1 Null Myoblasts Without Increased Apoptosis We next assessed caspase 3 activity in isolated Bit-1 KO and WT skeletal myoblasts prior to inducing differentiation. Indeed, caspase 3 activity was increased in Bit-1 KO myoblasts compared to WT prior to inducing differentiation (Fig. 1G). Caspase 3 activity is required and sufficient for myoblast differentiation (Fernando et al., 2002). Our data suggest that the early differentiation in Bit-1 KO myoblasts may be due to increased caspase 3 activity. To rule out caspase 3 activity enhanced apoptosis in these cells we assayed for apoptosis via TUNEL staining. A comparable proportion of apoptotic cells were observed for WT and Bit-1 KO myoblasts prior to inducing differentiation (Fig. 1H,I). These observations suggest elevated caspase 3 activity in the Bit-1 KO promotes premature differentiation in the early stages of skeletal myogenesis. The increase in caspase 3 activity does not promote apoptosis. To further delineate the effect of Bit-1 on myogenesis, we isolated myoblasts from Bit-1 KO and WT age-matched littermates and transiently transfected wildtype Bit-1 (WTBit-1; Fig. 2A; top blot). WT, WTBit-1 or Bit-1 KO myoblasts were placed in low serum to induce differentiation over 7 days. Caspase 3 activity was assessed by immunoblot at differentiation day 3. As expected Bit-1 KO myoblasts revealed enhanced active
8 caspase 3 compared to WT controls (Fig. 2A; lower blot). A partial reduction of increased caspase 3 activity was obtained with expression of wildtype Bit-1 (WTBit-1; Fig. 2A; lower blot). Fluorometric analyses of caspase 3 activity revealed a 1.7-fold increase in caspase 3 activity at pre-differentiation day 0 in the isolated Bit-1 KO myoblasts compared to WT controls (Fig. 2B; left graph). A partial rescue of Bit-1 KO myoblasts with wildtype Bit-1 expression reduced caspase 3 activity to midway between KO and WT activity levels (Fig. 2B; left graph). To confirm that Bit-1 KO myoblasts withdraw from the cell cycle prior to inducing differentiation, colormetric analysis of proliferating myoblasts as assessed by BRDu incorporation was determined. Bit-1 KO myoblasts incorporated 1-fold less BRDu at day 0 pre-differentiation compared to WT (Fig. 2B; right graph). In contrast, expression of wildtype Bit-1 induced a partial rescue by increasing BRDu incorporation to midway between Bit-1 KO and WT levels (Fig. 2B; right graph). A comparable proportion of apoptotic cells were observed for both WT and Bit-1 null myoblasts during differentiation (Fig. 2C). Moreover, re-expressing Bit-1 in Bit-1 KO myoblasts returned differentiation levels to control levels (Fig. 2D). Therefore, Bit-1 KO myoblasts are withdrawing from the cell cycle and beginning to differentiate even in high serum. Expression of Bit-1 blocks these cells from cell cycle withdraw. *Myogenin accumulation is delayed in caspase 3 null primary myoblasts (Fernando et al., 2002), whereas Bit-1 KO primary myoblasts induce early expression of myogenin at day 0 pre-differentiation and increased myogenin accumulation during differentiation
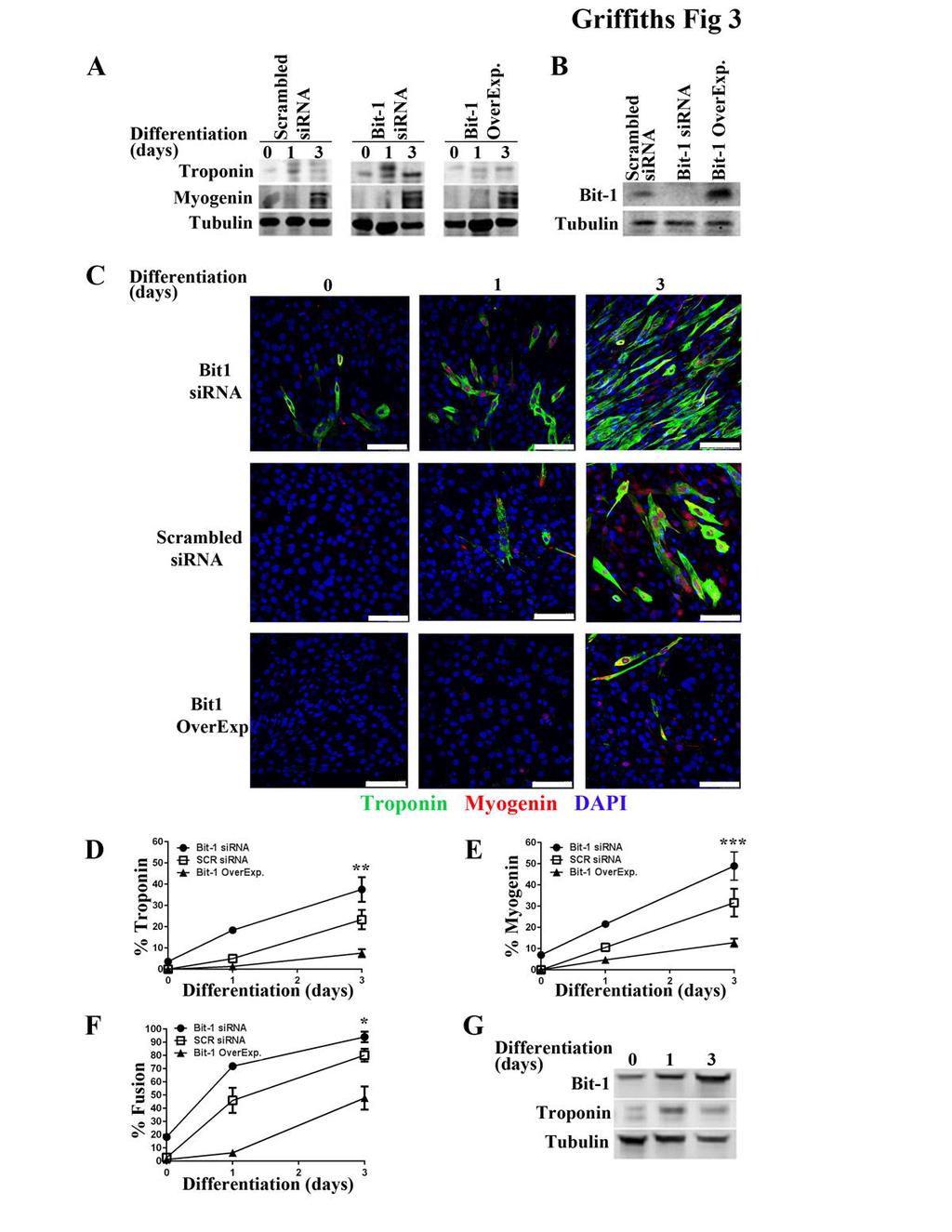
9 (Fig 2D). *Bit-1 KO primary myoblasts express higher Troponin levels and increased %Fusion compared to control wild type myoblasts. In addition, re-expression of Bit-1 returns Troponin levels and %Fusion back to control levels (Fig 2D). Taken together, these findings point to Bit-1 as a negative regulator of differentiation by modulating caspase 3 activity. sirna Knockdown of Bit-1 Promotes Early Differentiation In a C2C12 Differentiation Model To test whether specifically loss of Bit-1 in KO skeletal muscle led to premature differentiation rather than other developmental effects, we explored whether knockdown of Bit-1 in a myoblast cell line had similar results. C2C12 cells were engineered to express reduced Bit-1, normal levels of Bit-1, or enhanced levels of Bit-1 (Fig. 3B) and induced to undergo differentiation in low serum (Fernando et al., 2002). To assess muscle differentiation, we examined expression of the differentiation markers troponin T, myosin II, and myogenin. C2C12 cells transfected with Bit-1 targeting sirna showed premature differentiation as demonstrated by enhanced troponin T expression at predifferentiation day 0 and differentiation day 1 (Fig. 3A) compared to normal differentiation in C2C12 cells transfected with scrambled sirna (Fig. 3A). Myosin II expression was comparable in C2C12 cells transfected with Bit-1 targeting sirna or scrambled sirna (Fig. 3A). In contrast, C2C12 cells overexpressing Bit-1 showed increased levels of troponin T on differentiation day 3 indicating delayed differentiation (Fig. 3A). Similarly, immunostaining and quantitation for the differentiation-specific marker troponin T revealed that loss of Bit-1 promoted troponin expression at pre-
10 differentiation day 0 and enhanced expression at differentiation days 2 and 3 compared to scrambled sirna controls (Fig. 3C-D). Increased myogenin expression was also observed at differentiation day 1 in C2C12 cells transfected with Bit-1 sirna compared to scrambled sirna (Fig. 3C,E). In contrast, immunostaining of cells overexpressing Bit-1 showed delayed troponin expression, which was visible at differentiation day 3 (Fig. 3C-D). Finally, upon induction of differentiation, a significant increase in myotube formation was evident at earlier time-points in C2C12 cells transfected with Bit-1 sirna compared to scrambled sirna (Fig. 3F; compare a fusion index of 90% vs. 70%, knockdown to control, respectively). Upon differentiation of C2C12 myoblasts overexpressing Bit-1, a delay in myotube formation was evident (Fig. 3F). Notably, during normal C2C12 differentiation Bit-1 expression increases as differentiation progresses (Fig. 3G), indicating a differentiation-specific role for Bit-1. These findings confirm that loss of Bit-1 promotes early differentiation while Bit-1 overexpression delays differentiation as measured by differentiation-specific markers troponin, myosin II and myogenin and fusion index. sirna Knockdown of Bit-1 Promotes Increased Caspase 3 Activity In a C2C12 Differentiation Model We next examined how loss of Bit-1 affected caspase 3 activity in C2C12 cells during differentiation. Proliferating C2C12 cells transfected with Bit-1 sirna (Fig. 3B) or scrambled sirna (Fig. 3B) were placed in low serum and allowed to differentiate for three days. At respective time points, cells were washed extensively to ensure lysates were free of non-adherent apoptotic cells. Immunoblotting demonstrated enhanced
11 caspase 3 activity at day 0 pre-differentiation in C2C12 cells transfected with Bit-1 sirna compared to scrambled sirna (Fig. 4A). To confirm the immunoblotting analysis a fluorometric assay of caspase 3 activity was performed. Fluorometric analyses revealed a 4-fold increase in caspase 3 activity at pre-differentiation day 0 and a 2.0- fold increase within 24 h, which declined as differentiation progressed in the C2C12 cells transfected with Bit-1 sirna (Fig. 4B; left graph). Thus, loss of Bit-1 increased caspase 3 activity prior to inducing differentiation. Re-expression of Bit-1 Rescues Caspase 3 Activity To Wild Type Levels Rescue of C2C12 cells, in which Bit-1 levels had been knocked down, with wildtype Bit- 1 re-expression, returned caspase 3 activity back to control levels at pre-differentiation day 0 and during differentiation (Fig. 4B; left graph). *A comparable proportion of apoptotic cells were observed for C2C12 myoblasts transfected with Bit-1 sirna or scrambled sirna (Fig. 4C). Caspase 3 activity is necessary for myoblast differentiation while caspase 8 activity has a minimal effect (Fernando et al., 2002). Fluorometric analysis of caspase 8 activity showed an increase in activity during differentiation in agreement with previous reports (Fernando et al., 2002); Fig. 4D; left graph); however cells transfected with Bit-1 sirna or scrambled sirna had a similar trend (Fig. 4D; left graph). Treatment with a pharmacological inhibitor selective for caspase 8 (z-letd.fmk) completely inhibited caspase 8 activity in all cells (Fig. 4D; left graph). These findings suggest that upon Bit-1 knockdown, increased caspase 3 activity but not caspase 8 promotes myogenic differentiation.
12 Upon differentiation, myoblasts withdraw from the cell cycle and differentiate into multinucleate myofibers. Colorimetric analysis of proliferating cells as assessed by BRDu incorporation was determined during differentiation. C2C12s transfected with Bit- 1 sirna demonstrated significant reduced proliferation at differentiation day 1 compared to scrambled sirna control (Fig. 4B; right graph). Rescue of C2C12s, in which Bit-1 expression had been knocked down, with re-expression of wildtype Bit-1 returned BRDu levels back to control levels during differentiation (Fig. 4B; right graph).* Treatment with the caspase 8 inhibitor, z-letd.fmk, did not alter the observed cell cycle withdraw in the Bit-1 knockdown C2C12s or in the corresponding controls (Fig. 4D; right graph). Moreover, rescue of C2C12 cells, in which Bit-1 levels had been knocked down, with wildtype Bit-1 re-expression returned differentiation back to control levels as shown by decreased %Troponin, %Myogenin and %Fusion (Fig. 4E; all 3 graphs). Taken together, these findings demonstrate that prior to inducing differentiation C2C12 cells with reduced Bit-1 are withdrawing from the cell cycle and beginning to differentiate.* Thus, loss of Bit-1 promotes myoblast differentiation via increased caspase 3 activity and not through caspase 8.* Bcl-2 Levels Are Reduced In Bit-1 Null Skeletal Muscle The Bcl-2 family regulates caspase activity by blocking release of cytochrome c and inhibiting caspase 9 and 3 activation. The BH4 domain of Bcl-2 and Bcl-xL binds to the C terminal part of Apaf-1, and inhibits caspase 9 activation and therefore caspase 3 activity (Hu et al., 1998; Huang et al., 1998; Pan et al., 1998). We have reported that Bit-1 functions to upregulate Bcl-2 expression in a Hela tumor cell line (Griffiths et al.,
13 2011). Because we observed an increase in caspase 3 activity in Bit-1 KO skeletal muscle tissue, in isolated Bit-1 KO primary myoblasts and in C2C12 cells transfected with Bit-1 sirna, we next examined Bcl-2 levels in gastrocnemius muscle from Bit-1 KO and aged-matched WT littermates (Fig. 5A). In Bit-1 KO muscle tissue Bcl-2 levels are reduced suggesting Bcl-2-mediated inhibition of caspase 3 activity is decreased. Consistent with this result knockdown of Bit-1 expression in C2C12 cells also reduces Bcl-2 levels (Fig. 5B). Our data suggests loss of Bit-1 decreases Bcl-2 levels and promotes increased caspase 3 activity. This increase in caspase 3 activity would, thereby, initiate the early differentiation observed in isolated Bit-1 KO myoblasts and in C2C12 cells transfected with Bit-1 sirna. Bcl-2 Expression Rescues Bit-1 Null-Mediated Early Myogenesis Rescue of C2C12 cells, in which Bit-1 levels had been knocked down, with Bcl-2 reexpression returned caspase 3 activity to control levels at pre-differentiation day 0 and during differentiation (Fig. 5C). Re-expression of Bcl-2 in these cells also reduced differentiation specific Troponin expression (Fig. 5D left graph) and % Fusion (Fig. 5D right graph) to control levels during differentiation. Similarly, Bit-1 null myoblasts demonstrate decreased Bcl-2 expression levels (Fig. 6A). Re-expression of Bcl-2 in Bit- 1 null myoblasts rescued caspase 3 activity back to control levels (Fig. 6B). Moreover, caspase 9 cleavage was increased in Bit-1 null myoblasts compared to WT controls (Fig. 6C). Caspase 9 activity was higher in Bit-1 null myoblasts and re-expression of Bcl-2 in Bit-1 null myoblasts decreased caspase 9 activity to WT control levels (Fig. 6D). The pattern of caspase 9 activity was similar to that of caspase 3. These findings
14 suggest that Bit-1 regulates skeletal muscle differentiation by modulating Bcl-2 expression and therefore downstream caspase activity.
15 Discussion In this study we provide evidence that Bit-1, a regulator of Bcl-2, is a mediator of muscle differentiation. Upon examination and further characterization of the skeletal muscle of Bit-1 null mice we found these mice developed a muscle myopathy with hypotrophic myofibers. The observed early differentiation in Bit-1 null muscle was due to decreased Bcl-2 expression and subsequent increased caspase 3 activity without increased apoptosis. Re-expression of Bit-1 or of Bcl-2 blocked premature differentiation of Bit-1 null myoblasts. Our data demonstrate for the first time that Bit-1 plays a regulatory role in normal muscle differentiation outside of its function in regulating apoptosis. Muscle differentiation requires myoblast precursor cells to withdraw from the cell cycle and differentiate into multinucleated myofibers (Anderson, 1998; Grounds, 1991; Lluis et al., 2006b). Several of the biochemical pathways that regulate apoptosis also play key roles in terminal cell differentiation. Bcl-2 family members including Bcl-xL are required for skeletal muscle differentiation (Murray et al., 2008). Caspase 3 is another such mediator and is also required for skeletal muscle differentiation (Fernando et al., 2005; Fernando et al., 2002; Li and Yuan, 2008). These findings point to a critical regulatory role of genes that control the intrinsic mitochondrial apoptotic pathway in muscle differentiation. However, how such signaling pathways are regulated is not yet fully understood. We have previously identified Bit-1 as a regulator of integrin-mediated apoptosis (Griffiths et al., 2011; Jan et al., 2004). Integrins are a family of transmembrane
16 receptors that bind to extracellular matrix (ECM) proteins. These receptors promote cell ECM adhesion and mediate adhesion-initiated signal transduction pathways. A subset of integrins is essential for myogenesis including the β3 and α7 integrins (Burkin and Kaufman, 1999; Liu et al., 2011). The β3 integrin acts on myogenic differentiation through activation of p130 Cas and MAPK (Kawauchi et al., 2012). α7 integrin mutations result in congenital myopathies in humans and induce muscular dystrophy in mice (Hayashi et al., 1998; Mayer et al., 1997). Exactly how downstream signals are activated by integrin attachment to promote muscle differentiation is not yet fully known. Integrin adhesion activates the tyrosine kinase focal adhesion kinase (FAK), which in turn activates a number of downstream signaling pathways that promote cell proliferation, survival and muscle differentiation. Integrin-dependent myoblast differentiation requires FAK activation (Luo et al., 2009). We have previously reported that Bit-1 interacts in a complex with FAK in a myoblast cell line (Griffiths et al., 2011). We propose integrins signal through Bit-1 to control myoblast differentiation by linking adhesion and apoptosis pathways. The molecular mechanism of how integrins induce Bit-1 signaling is currently under investigation. Our data show that the myopathy characterized by hypotrophic myofibers observed in Bit-1 nulls is due, in part, to increased caspase 3 signaling. We have previously reported that loss of Bit-1 expression induces increased caspase 3 activity in normal endothelial cells and cancer cells (Griffiths et al., 2011). Our data support previous findings on the essential role of caspase 3 activity for muscle differentiation (Fernando et al., 2002) and extends them by demonstrating that Bit-1 negatively regulates caspase 3 activity during muscle differentiation. We show here that loss of Bit-1 promotes
17 increased caspase 3 activity in myoblasts even in pre-differentiation high serum conditions. While re-expression of Bit-1 in the primary Bit-1 null myoblasts showed minimal effect on returning caspase 3 levels to control levels we did observe a significant rescue of caspase 3 activity upon re-expression of Bit-1 in C2C12 cells that had been treated with Bit-1 sirna. Moreover, in both cell types re-expression of Bit-1 induced a significant rescue of the differentiation phenotype. The reduced effect on caspase 3 activity by re-expressing Bit-1 in the primary Bit-1 null myoblasts may be due to the lower transfection efficiency that occurs with primary cells. Our findings are in agreement with the report that caspase 3 over-expression induces myoblast differentiation in pre-differentiation high serum conditions (Fernando et al., 2002). * To determine if the increase in early differentiation upon loss of Bit-1 was due to other caspases such as caspase 8, we used a pharmacolgical inhibitor for caspase 8. Pharmacological inhibition of caspase 8 did not block the observed early differentiation in C2C12 myoblasts with sirna knockdown of Bit-1. While the pharmacological inhibitor tested is selective and not specific, caspase 8 did not appear to play a role. This finding is in agreement with previous reports that myoblast differentiation requires caspase 3 but not caspase 8 (Fernando et al., 2002). Furthermore, caspase 9, an initiator caspase in the intrinsic mitochondrial apoptotic pathway, functions in a non-apoptotic role by activating caspase 3 in C2C12 myoblast differentiation (Murray et al., 2008). Our data demonstrate that caspase 9 processing and activity increased in Bit-1 null myoblasts similar to caspase 3 activity suggesting that caspase 9 is the intermediate step that activates caspase 3 during differentiation. Our data also confirm that similar to caspase 9 and 3, Bit-1 functions in a non-apoptotic role during myogenesis.
18 We have shown Bit-1 regulates caspase 3 activity is through its effects on Bcl-2. The Bcl-2 family controls caspase activity by inhibiting cytochrome c release and subsequent caspase 3 activation. The BH4 domain of Bcl-2 and Bcl-xL binds to the c- terminal region of Apaf-1, inhibiting caspase 9 activation and subsequent caspase 3 activity (Hu et al., 1998; Huang et al., 1998; Pan et al., 1998). Bcl-xL overexpression blocks myoblast differentiation by inhibiting downstream caspase activation (Murray et al., 2008). It is therefore possible that other Bcl-2 family members like Bcl-xL could mediate the effects of Bit-1 in muscle differentiation. This remains to be determined. We have reported previously that upon integrin-mediated attachment Bit-1 upregulates Bcl- 2 expression (Griffiths et al., 2011). We report here that Bcl-2 levels were significantly reduced in gastrocnemius muscle from Bit-1 null mice compared to age-matched WT littermate controls. Moreover, re-expression of Bcl-2 in Bit-1 null myoblasts returned caspase 3 activity to WT control levels and rescued early differentiation. Bcl-2 reexpression in these cells also decreased caspase 9 activity to WT control levels. These findings suggest that Bcl-2 mediated inhibition of caspase 3 activity is decreased in Bit-1 null muscle through modulation of the initiator caspase 9 and may be, at least in part, the reason that the Bit-1 null mice present with hypotrophic myofibers. Therefore, Bit-1 regulates myoblast differentiation by controlling Bcl-2 expression to restrain caspase 3 in a non-death setting. Overexpression of Bcl-2 ameliorates muscular dystrophy disease progression in an integrin ligand-laminin alpha 2 null mouse model of muscular dystrophy (Dominov et al., 2005) and in a mouse model of oculopharngeal muscular dystrophy (Davies and Rubinsztein, 2011). It will be informative to determine if overexpression of Bit-1 in
19 laminin alpha 2 null mice may attenuate disease progression in these mice. Functional and physiological studies are needed in these mice to demonstrate the therapeutic potential of Bit-1 overexpression. Recently, we used whole exome sequencing to identify mutations in Bit-1 that cause progressive congenital muscle weakness in humans (Hu et al., 2014) suggesting that Bit-1 plays a key role in muscle function. Our data here points to Bit-1 as an important regulator of muscle differentiation. Bit-1 is a potential new therapeutic target for muscle disease such as muscular dystrophy. We postulate that Bit-1 is the missing signal between integrins and the intrinsic mitochondrial apoptotic pathway in muscle differentiation.
20 Materials and Methods Animals. Mouse tissue was obtained from c57bl/6 Bit-1-/- and age-matched WT littermates. All animals received care in compliance with the principles of laboratory animal care and use formulated by the Institutional Animal Care & Use Committee. Genotyping- Genotyping of Bit-1 null mice has been described previously (Kairouz- Wahbe et al., 2008). Briefly, DNA for genotyping was isolated from 8 day old mouse tails or toes, and genotypes were determined via PCR using the primer combination G2F/G1R/30d; G2F (5 TGG GTC TTT GAA TCA ACT AG 3 ), G1R (5 ACA TGC CAC AAG CAA CTC CA 3 ), 30d (5 TTT GAG ACC CTA TCA CTC CAC ACG). PCR was performed under conditions of initial 2 min denaturation at 95 C, followed by 30 cycles consisting of 94 C for 30 sec, 55 C for 30 sec and 72 C for one minute with a final extension of 72 C for 5 min. H&E Staining. Gastrocnemius muscle from Bit-1 KO and WT mice were embedded in OCT cutting compound (Sakuara, Torrance, CA) and sectioned to 8 µm using a LeicaCM 1850 cryostat. Sections were placed onto prewashed Surgipath slides (Surgipath, Richmond, IL). Sections were stained via Hematoxylin and Eosin and used to determine myofiber cross-sectional area and longitudinal fiber width. Myofiber Area Determination.Hematoxylin and Eosin stained slides were used to determine myofiber cross-sectional area and longitudinal myofiber width. A minimum of 1000 fibers per animal from 3 Bit-1 KO and 3 WT mice were counted. Myofiber cross-
21 sectional area was determined with a Zeiss Axioskop 2 Plus fluorescent microscope and images were captured with a Zeiss AxioCam HRc digital camera with Axiovision 4.1 software. Minimal Feret s Diameter. Quantitative determination of muscle fiber cross-section diameter was obtained using minimal Feret s diameter analysis for triceps and TA muscles of Bit-1 KO and age-matched WT litter-mate controls at P0 and P7. Using a Leica CM1850 cryostat 10-µm sections of Tissue-TEK Optimal cutting Temperature compound (Sakura Finetek USA Inc.) embedded tissues from mice were placed on Surgipath microscope slides (Surgipath Medical Industries). Hemotoxylin and eosin (H&E) staining was performed on triceps and TA muscles from Bit-1 KO and WT agematched littermates at P0 and P7. Images were taken using an Olympus Fluoview FV1000 Laser Confocal Microscope and blinded evaluation of sections was performed. Pictures of a tricep or TA section were taken at 100x using a Zeiss Axioskop 2 Plus fluorescent microscope, Zeiss AxioCam HRc digital camera, and Axiovision 4.8 software. Compiled images were used to reconstruct a view of the entire tricep or TA muscle as described previously (Van Ry et al., 2014). This compilation was used for calculating minimum Feret s diameter. There was a minimum of n=3 mice for each treatment group and a minimum of n=3000 fibers Myoblast isolation. Primary myoblasts were isolated from Bit-1 KO and WT mice from gastrocnemius skeletal muscle as previously described (Megeney et al., 1996). Isolated muscle was treated with Penstrep (Invitrogen) for 10 min at 37 C to prevent bacterial contamination during culture. Muscle tissue was rinsed 3 times in PBS and minced in
22 Ham s F-12 media supplemented with 20% FBS and 2% Penstrep, 1% L-Glutamine and 1% non-essential amino acids (Invitrogen). Fusion Index. To determine the fusion index differentiation cultures were analyzed microscopically as previously described (Quach et al., 2009). A 20 x objective and phase contrast (Zeiss Axioskop 2 Plus microscope) were used to analyze random fields of WT control, Bit-1 null primary myoblasts or C2C12 myoblasts. Myotubes with three or more nuclei were counted. Fusion index was then determined as the percentage of nuclei in myotubes compared with the total number of nuclei in the field. Approximately 100 myotubes were counted per dish. Measurements were preformed in triplicate in three independent experiments per genotype. Cell culture. The C2C12 myoblast cell line was maintained in DMEM supplemented with 15% FBS, 1% Penstrep, 1% MEM non-essential amino acids, and 1% L-Glutamine. Cells were incubated at 5% CO 2 ; 37 C. For differentiation assays, media was changed to DMEM supplemented with 1.5% horse serum, 1% Penstrep, 1% MEM non-essential amino acids, and 1% L-Glutamine and incubated 5% CO 2 ; 37 C. Primary myoblasts were isolated from Bit-1 null and age-matched WT littermates as described above and induced to differentiate in low serum media. Caspase 8 Inhibition*. For caspase 8 inhibition primary KO and WT myoblasts or C2C12 cells were treated with 20 µm inhibitor selective for caspase 8 (z-letd.fmk; BD Parmingen) or negative control peptide (Z-FA-FMK; BD Pharmingen) for 3 h prior to
23 differentiation when fresh inhibitor or control peptide was added; fresh inhibitor or control peptide was added every 24 h throughout the experiments. RNA Interference of Bit-1. A Dharmacon si-genome Smartpool (Thermo Fisher Scientific) consisting of 4 Bit-1 specific sirnas and control scrambled sirnas were used to knockdown Bit-1 expression levels. For transient transfection experiments, 2 x 10 5 C2C12 cells were transfected with 25 µm of Bit-1 Smartpool using the LipofectAMINE 2000 transfection reagent (Invitrogen). At various time points cells were subjected to immunostaining or enzymatic assays as described below. Cell lysates were also collected at corresponding time points for immunoblotting. For over-expression and re-expression experiments 3 µg of the WT-Bit-1 or WT Bcl-2 constructs were transfected into KO and WT primary myoblasts or C2C12 cells using LipofectAMINE Immunoblotting. Isolated primary Bit-1 KO myoblasts, aged matched WT littermate control primary myoblasts and C2C12 cells were lysed with lysis buffer containing 50mM Tris-HCL, 3mM EDTA, 0.5% Triton X-100, ph 7.0, 0.5 mm dithiothreitol and protease inhibitors (Complete Protease Inhibitor Tablets, Roche Applied Science). Cell lysates were sonicated and cleared by centrifugation at 28,000 rpm for 10 min at 4 C. Equal amounts of cell lysates were resolved on 4-12% SDS-polyacrylamide gels. After electrophoresis, proteins were transferred to Immobilon-P nylon membrane (Millipore Corp., Bedford, MA) and immunoblotted. Membranes were blocked for 1 h with blocking buffer (3% bovine serum albumin/pbs-t) and incubated with various antibodies at a
24 dilution of 1:1000 unless specified otherwise including rabbit polyclonal anti-bit-1 antibody (developed at Washington Biotechnology Columbia, MD using the Bit-1 amino acid sequence GPADLIDKVTAGHLKL), mouse anti-troponin antibody (Santa Cruz Biotechnology Inc. Santa Cruz, CA), rabbit anti-myogenin antibody (Santa Cruz Biotechnology Inc.), anti-myosin (Abcam), anti-microtubulin (BioLegend), anti-caspase 3 (Cell Signaling) or anti-caspase 9 (Cell Signaling) for 1 h. Membranes were washed and incubated with Li-Cor secondary antibodies (ant-rabbit IR 800 CW and anti-mouse IR 680 CW; Li-Cor) for 1 h. Immunoblots were analyzed via Oddessy software, provided with the Li-Cor system. Caspase fluorometric assays. Primary Bit-1 KO and WT myoblasts and C2C12 cells were analyzed for caspase 3 activity, caspase 8 activity or caspase 9 activity via fluorometric assays (BioVision Inc., San Francisco, CA) as per the manufacturers instructions. Briefly, 1x 10 5 cells were plated/well in a 96 well plate. Cells were exposed to non-differentiation or differentiation media prior to being lysed and incubated with DEVD-AFC substrate for 1 h at 37 C. Fluorescence was assessed using a plate reader with 400/505 nm excitation/emission. *Cells incubated with a caspase 8 inhibitor (BD Pharmingin) at a concentration of 20 µm were used as a negative control for caspase 8 activity. BrdU Proliferation Assay. Proliferation rates of Bit-1 KO primary myoblasts, WT primary myoblasts and C2C12 cells were determined using a BrdU incorporation assay (Cell Signaling) per manufacturers instructions. Bit-1 or WT primary myoblasts or
25 C2C12 cells were exposed to non-differentiation or differentiation media prior analysis using the BrdU kit reagents. Cells were washed 3x, fixed and incubated with an anti- BrdU antibody for 1h at RT. Cells were next washed 3x and incubated with an HRPconjugated secondary antibody for 30 min at RT., followed by addition of TMB substrate. After a 30 min incubation, a STOP solution was added, and Absorbance was read at 450 nm in a plate reader. *Cells incubated in the presence of a caspase 8 selective inhibitor (BD Biosciences) at a concentration of 20 µm were used as a negative control. Immunostaining. Bit-1 and WT primary myoblasts or C2C12 cells were fixed in 4% paraformaldehyde, permeabilized and incubated with an anti-troponin T or antimyogenin antibody (Santa Cruz), followed by an Alexa Fluor 594 or 488 secondary antibody (Molecular Probes) and counterstained with DAPI. Images were obtained using a Zeiss Axiovert 200m fluorescent microscope. APOPtag. Isolated primary myoblasts from Bit-1 KO and aged-matched WT littermates or gastrocnemius muscle tissue fixed in 1% paraformaldehyde in PBS, permeabilized in 70% ethanol and stained with ApopTag reagent (apoptosis in situ detection kit; Millipore, Temecula, CA) and propidium iodide per kit protocol. Images were obtained using a Zeiss Axiovert 200m fluorescent microscope. Statistical Analyses. Myofiber cross sectional area analysis, % myoblast fusion, ELISA and densitometric readings were subjected to statistical analysis. Differences
26 between the mean values and the densitometric readings were analyzed by ANOVA followed by Bonferroni s test for multiple comparisons between pairs or a Student s t test. Values of P<0.05 indicated statistical significance. At least three independent experiments were performed for each data set and combined for statistical analysis.
27 Acknowledgements. We thank S. Young-Robbins and Anna Leychenko for expert technical assistance. This work was supported in part by grants from the National Institutes of Health (RO1-GM to M.L.M; NCRR P20-RR to M.L.M; R01- GM to J.W.R.; R01AR to D.J.B) and a grant from the Ingeborg Foundation (M.L.M). Conflict of Interest. The authors declare they have no competing financial interests.
28 References: ƒ ƒ ƒ ˆ Š ƒ ˆ Ž ƒž Ž ƒ Š ƒ ƒ Š ŽŽ Ž Ž ƒƒš ƒ ƒž ƒ ŽƒŠ ƒ ƒ Žƒ ˆ ƒ ˆ ŽŠ Š ˆ ƒ ƒžžƒš ƒ Š ƒ œƒ Žƒ ˆ ˆ ŽŽ Žƒ ƒ Žƒ ƒ Ž Š ƒ ƒ Š ƒ ƒ Œ ƒž ˆ ƒ Žƒ ƒ ˆˆ ƒžœ ƒž ˆ Š ƒ ˆ ƒ Žƒ ƒ ƒ Š ƒ ˆ ƒ Žƒ ƒ ƒ ƒ ŽŽ ƒ Š ƒ Š ˆ Ž Š ƒ Š œ ƒž ƒ ƒ Š ˆ ˆƒ Ž ƒž ŽŽ ƒ Šƒ ˆˆ ƒ ƒ ƒ ˆ ƒ Š ƒž Šƒ ƒ Ž Ž ƒ ƒ ŽŽ ƒ ƒ œ š ˆ Ž ƒ ƒ Ž ˆ Ž Šƒ ƒž Žƒ Š ƒ Ž Žƒ ƒ œ Ž ƒƒ ŽŽ Ž ˆ š ƒ Ž ƒ Ž ƒ Žƒ ƒž Šƒ ˆ Š ˆ ƒ Ž Žƒ ƒ ƒ ƒž ŽŽ ˆˆ ƒ ƒ ƒ ƒ Š Œ ƒž ˆˆ ƒž Ž ƒ ˆ Š ƒ ˆ ƒ ˆ š ƒž Ž ƒ ŽŽ ƒžƒœ Žƒ ƒ ƒ ƒ ƒ ˆ Ž ƒž Ž ˆˆ ƒ ˆ Š ƒ ƒž ƒ ˆ ˆ Š ƒ ˆ ƒ ƒ ƒ ƒ ƒ ƒ Ž ŽŽ ˆˆ ƒ ƒ Š š Š Œ ƒž ˆˆ ƒž Ž ƒ ˆ Š ƒ ˆ ƒ ˆ š ƒž Ž Œ ƒ ƒ œƒ Œ œ ƒ Žƒ ŽŽŠ ƒ ƒ ƒƒ ƒ ƒ ƒ Š ˆˆ ƒ ˆ ŽŽ ŽŽ ŽŽ ˆˆ Š Ž Š Žœ ƒ ƒž ƒƒ ƒ ƒ ƒ ŽŽ ƒž Š Š ƒ ˆ Š ƒ ƒ ƒ Š ƒ Š ƒž ˆ Ž ƒž Š ƒ ƒ Ž ƒž Ž ƒ ƒ Š Ž ƒ ƒ ƒ Š Š ƒžž ƒ ƒ ƒ ƒ ƒ ƒ ƒ Š Š ƒ ƒ ˆ ƒ ƒž ƒ Š ƒž Šƒ ƒ ƒž ƒ Š ƒ ƒ ƒ ƒš Œ ƒ ƒ ƒ Žƒ ƒ ƒ Š Ž ƒ ƒ ƒ ƒž ƒ ƒ Ž ˆƒ Ž Ž ƒ Š ŽŽ ƒž ƒ Ž ŠƒŽ ƒ ƒš ƒ ƒ Ž ƒ ƒž ˆ Ž ƒžƒ ƒ Žƒ ƒž Ž
29 Šƒ ƒ ƒ œ Ž ƒ Š ƒˆ ƒ Š ƒˆ ƒ ƒ ƒ ƒ ˆ Š ƒ ƒž ƒ ˆ ˆ Š ƒ ˆ ƒ ƒ ƒ ƒ Š ƒž ƒ ˆ Ž Š Ž ƒžˆ Š ˆƒ ƒ ƒ Š Š Œ ƒž ƒƒ ƒ Š Ž Š ƒ ƒ ƒ ŽƒŠ Š ƒž ƒ ƒ Žƒ ƒ Š ŽŽ ƒ œ Ž ƒ Ž Ž Ž Ž ƒ ƒ Ž ƒž ƒ ŽŽ š ƒž Ž ƒ ƒ ŽŽ ŽŽ ƒ œ ƒš Ž ƒ Š ƒ ŽŽ Š ŽŽ ˆ ƒ ƒž Š ƒ ƒ ŽƒŠ ˆˆ ƒ Ž Žƒ ƒ ƒ Ž ƒ ƒ ƒ Š ƒ ƒ ƒ ƒ Œ ƒ ƒ ƒ ƒ ƒ ƒ ƒ ƒ ƒ ŽŽ Žƒ ˆˆ ƒ Š ƒ ƒ ƒžž Ž Š ƒ ƒ ƒ ƒ ƒ ƒ ƒ ƒ ŽŽ ˆˆ ƒ ƒ ƒ ˆ Š ƒ ƒž ƒ ˆ ˆ Š ƒ ˆ ƒ ƒ ƒƒ ƒ ƒ ƒ Š ƒ ƒ ƒ ƒ ŽŽ ŽŽ ˆˆ ƒ ƒ Ž Ž ƒƒ œ ƒ ƒ Žƒ ˆ Ž ƒž Ž š ƒ ŽŽ Ž Ž ƒƒ œ ƒ Žƒ ˆ Ž ƒž Ž š ƒ ŽŽ Ž Šƒ Šƒ ƒ Žƒ ˆŠ Š ƒ ŽŽ ƒ š Ž ˆˆ ƒ ƒ Š ƒœ ƒ Ž ƒ Š ƒ ƒ Žƒ Ž ƒž Ž ˆˆ ƒ Š Š ƒž Ž Žƒ ƒ ŽŽ Žƒ Ž ƒ ƒš ƒ Ž ƒ Š ƒ ŠŽ ƒ ƒ ˆ ƒž Šƒ ƒ ƒ Žˆ ˆ Žƒ Š ƒ ƒ Žƒ ƒ ƒ ˆ ŽŽˆ ƒ Ž Ž ƒž Ž ƒ ƒš Ž ƒ Ž Š ƒ ƒ ƒ ƒ Š ƒ ƒ Ž ˆ ƒ ƒ Ž ˆˆ ƒ ŽŽ ƒ ƒ š ƒ ƒ Žƒ ƒˆˆ ƒ ƒ Ž š Š ƒž ˆ Ž ƒž Š
30 ƒ Š Šƒ ŽŽ ƒ ƒ ƒžƒ Š ƒ ƒž Žƒ Š š ˆ ƒ Ž ƒ ƒ ƒž ˆ ƒž Žƒ ˆ Ž Ž ŽŽ ƒ ƒ ƒ Ž ƒ ƒ Ž ˆ ˆ ƒž Žƒ Š ƒ Ž Žƒ Ž ƒ Ž Ž ˆ Ž ƒž Ž ˆˆ ƒ Š ƒž ˆ Ž ƒž Š
31 Figure Legends. Figure 1. Bit-1 null mice exhibit a myopathy with hypotrophic muscle fibers. A) H&E stained gastrocnemius muscle tissue sections (top: cross-sections; bottom: longitudinal). Lower graph: Apoptag stained gastrocnemius muscle tissue sections for detection of apoptotic myofibers. A comparable proportion of apoptotic myoblasts was observed for both WT and Bit-1 KO. (Bit-1 KO n=3000; WT n=3000. B) top graphs: Quantitation of myofibril number (Y-axis) and myofibril cross-sectional area (μm; X-axis) and average cross-sectional area (top right graph; μm) of Bit-1 KO and age-matched WT littermate controls at P7. Bottom graphs: Quantitation of average longitudinal fiber width (bottom left graph; μm; Bit-1 KO n=3000; WT n=3000) and average muscle weight (bottom right graph; mg; Bit-1 KO n=5; WT n=5) of Bit-1 KO and age-matched WT littermate controls. For A-B: **P<0.001; ***P< C) Quantitation of triceps and tibialis anterior (TA) muscle diameter using minimal Feret s diameter analysis at P0 and P7 of Bit-1 KO and age-matched WT littermate controls (top graph; Bit-1 KO n=3000; WT n=3000). Average muscle weight (bottom graph; mg; Bit-1 KO n=5; WT n=5) of Bit- 1 KO and age-matched WT littermate controls at P0 and P7. *P<0.05. D) Primary myoblast cultures were derived from WT and Bit-1 KO mice (n=3 for each genotype). Western blot analysis for the differentiation-specific proteins Troponin T and Myosin II were assessed at pre-differentiation day 0 (cultured in high serum). Tubulin was used as a loading control. Data shown is representative of 3 independent experiments. E) Bit- 1 null myoblasts express the differentiation-specific marker Troponin T earlier than WT age-matched littermate controls. Immunostaining for Troponin T (red) and nuclear DAPI (blue) was performed at pre-differentiation day 0 (40X). Data shown is representative of
32 3 independent experiments. F) Quantitation of isolated primary Bit-1 KO myoblasts and WT age-matched littermate controls at pre-differentiation day 0. Statistically significant differences are reported in the graph as P values (Student s t-test), **, P <0.01. (n=3 for each genotype). G) Western blot of pro-caspase 3 and cleaved caspase 3 in primary myoblasts isolated from Bit-1 KO and age-matched WT littermates at day 0 predifferentiation. Actin was used as a loading control. Blot is representative of 3 independent experiments. H) Apoptag stained primary myoblasts isolated from Bit-1 KO and WT age-matched controls at day 0 pre-differentiation (40X). I) Quantitation of apoptotic cells. Apoptosis was comparable for KO and WT controls at day 0 predifferentiation. n=3 for each genotype. Figure 2. Expression of Bit-1 in Bit-1 null primary myoblasts inhibits early differentiation and reduces caspase 3 activity. A) Western blot analysis of: Top blot: Bit-1 expression levels in WT, Bit-1 null and expression of wildtype Bit-1 in isolated KO myoblasts at pre-differentiation day 0. Actin was used as a loading control. Bottom blot: Pro-caspase 3 and cleaved caspase 3 (cleaved Casp 3) was assessed at differentiation day 3 (differentiation media) in myoblasts from Bit-1 KO and age-matched WT littermate controls or Bit-1 null myoblasts transfected with wildtype Bit-1. The 19- and 17-kD fragments are indicative of caspase 3 cleavage, which is more abundant in Bit-1 KO myoblasts. For blots data is representative of 3 independent experiments. B) Left graph: A sharp increase in caspase 3 activity, as determined by a fluorometric protease assay that directly measures caspase 3 activity, was found at pre-differentiation day 0 and differentiation days 3, 5, and 7 in Bit-1 KO myoblasts (open square) compared to age-
33 matched WT littermates (circle). Expression of wildtype Bit-1 (triangle) reduced caspase 3 activity to mid-way between KO and control levels. Right graph: Assessment of BRDu incorporation in Bit-1 KO (open square) and WT (circle) myoblasts indicate that there is a sharp decrease in BRDu incorporation at pre-differentiation day 0 and differentiation days 3, 5, and 7 in the Bit-1 KO myoblasts. Expression of Bit-1 (triangle) reduced BRDu incorporation to mid-way between the KO and WT controls. C) Quantitation of myoblast apoptosis at day 3 differentiation (cultured in differentiation media). Apoptosis was comparable for Bit-1 KO and WT controls (n=5 for each myoblast type)*. D) Quantitation of %Troponin T, %Myogenin, and %Fusion Index. Bit-1 KO myoblasts demonstrated earlier and increased troponin T and myogenin expression in addition to increased myoblast fusion at pre-differentiation day 0 and over three days of differentiation. Statistically significant differences are reported in the graphs as P values (Student s t- test), *, P <0.01; Bit-1 KO vs. WT or WTBit-1. (n=5 for each genotype). Figure 3. Loss of Bit-1 expression induces differentiation in growing C2C12 cells. A) C2C12s were transfected with either a scrambled sirna, Bit-1 sirna or wildtype Bit- 1 (Bit-1 OverExp.) and analyzed via Western blot for the differentiation-specific proteins Troponin T and Myosin II at pre-differentiation day 0 (cultured in high serum) and differentiation days 1 and 3 (cultured in low serum differentiation media). An increase in Troponin T expression was found in C2C12s with low Bit-1 expression (Bit-1 sirna) at pre-differentiation day 0. Myosin II expression levels were comparable between scrambled sirna, Bit-1 sirna and Bit-1 Overexpressors. Tubulin was used as a loading control. Data shown is representative of 3 independent experiments. B)
34 Western blot analysis of Bit-1 expression levels in scrambled sirna, Bit-1 sirna, and Bit-1 Overexpressors at pre-differentiation day 0. Tubulin was used as a loading control. Data shown is representative of 3 independent experiments. C) Immunostaining for Troponin T (red), nuclear DAPI (blue), and myogenin (green) was performed at predifferentiation day 0 and differentiation days 1 and 3 (40X). Data shown is representative of 3 independent experiments. Quantitation of %Troponin T (D), %Myogenin (E), and %Fusion Index (F). % Fusion index was assessed as a percentage of cells containing two or more nuclei within a differentiated myotube. Statistically significant differences are reported in the graphs as P values (Student s t-test), *, P <0.01; Bit-1 KO vs. Scr sirna. (n=5 per genotype). G) Bit-1 and Troponin expression levels were assessed during normal C2C12 myogenic differentiation over 3 days in low serum conditions. Tubulin was used as a loading control. Blot is representative of 3 independent experiments. Figure 4. Caspase 3 activity is increased in growing C2C12 cells transfected with Bit-1 targeting sirna. A) C2C12s were transfected with either a scrambled sirna or Bit-1 sirna and 48 hours later analyzed by Western blot for pro-caspase 3 and cleaved caspase 3 at pre-differentiation day 0 (high serum) and differentiation days 1 and 3 (differentiation media). The 19- and 17-kD fragments are indicative of caspase 3 activation, which is more abundant in pre-differentiation day 0 in C2C12s with knockdown levels of Bit-1. Data is representative of 3 independent experiments. B) Left graph: A sharp increase in caspase 3 activity, as determined by a fluorometric protease assay that directly measures caspase 3 activity, was found at pre-differentiation day 0
35 and differentiation days 1 and 3 in C2C12s expressing low levels of Bit-1 (Bit-1 sirna;open square) compared to scrambled sirna (Scr sirna;black circle). Reexpression of wildtype Bit-1 (WTBit-1;black triangle) reduced caspase 3 activity back to control levels*. Right graph: Assessment of BRDu incorporation in C2C12s transfected with Bit-1 sirna and scrambled sirna controls demonstrate that there is a sharp decrease in BRDu incorporation at pre-differentiation day 0 and differentiation days 1 and 3 in C2C12s expressing low levels of Bit-1 (open square). Re-expression of Bit-1 (black triangle) reduced BRDu incorporation back to control levels. C) Quantitation of C2C12 apoptosis at day 3 differentiation (cultured in differentiation media). Apoptosis was comparable for Bit-1 sirna and scrambled sirna controls (n=3 for each). D) Left graph: Caspase 8 activation, as determined by a fluorometric protease assay that directly measures caspase 8 activity, displayed a minimal increase upon induction of differentiation, which was similar in the Bit-1 sirna (open square) and scrambled sirna (black circle) C2C12s. Cells were incubated in low serum with a caspase 8 selective inhibitor (z-letd.fmk) or DMSO-carrier control and assessed over 3 days differentiation. Right graph: Treatment of C2C12s with a caspase 8 selective inhibitor did not change BRDu incorporation in differentiating C2C12s. E) Quantitation of %Troponin T, %Myogenin, and %Fusion Index. C2C12s expressing knockdown levels of Bit-1 demonstrated earlier and increased Troponin T and Myogenin expression in addition to increased myoblast fusion (%Fusion) over three days of differentiation. Reexpression of WTBit-1 returned %Troponin T, %Myogenin, and %Fusion to control levels. Incubation with a caspase 8 selective inhibitor did not significantly alter C2C12 differentiation. Statistically significant differences are reported in the graphs as P values
36 (Student s t-test), *, P <0.01; Bit-1 KO vs. Scr sirna. (n=5 for each variant: Bit-1 sirna; Scrambled sirna; Bit-1 sirna + WTBit-1). Figure 5. Bcl-2 expression rescues Bit-1 null-induced early differentiation. A) Western blot of Bit-1 KO and age-matched WT littermate control gastrocnemius muscle tissue. Bit-1 KO cannot achieve WT levels of Bcl-2 expression. Tubulin was used as a loading control. Blot is representative of 3 independent experiments. B) C2C12s were transfected with either a scrambled sirna or Bit-1 sirna or Bit-1 sirna and Bcl-2 plasmid and 48 hours later analyzed by Western blot for Bit-1 and Bcl-2 expression. Tubulin was used as a loading control. Blot is representative of 3 independent experiments. C) A sharp increase in caspase 3 activity, as determined by a fluorometric protease assay that directly measures caspase 3 activity, was found at predifferentiation day 0 and differentiation days 1 and 3 in C2C12s expressing low levels of Bit-1 (Bit-1 sirna;open square) compared to scrambled sirna (Scr sirna;black square) or scrambled sirna and Bcl-2 (Scr sirna/bcl-2; black triangle). Re-expression of Bcl-2 (Bit-1 sirna/bcl-2;open circle) reduced caspase 3 activity back to control levels. D) Quantitation of %Troponin T, and % Fusion Index. Re-expression of Bcl-2 in C2C12s expressing low levels of Bit-1 reduced troponin T and % Fusion back to control levels over three days of differentiation. Statistically significant differences are reported in the graphs as P values (Student s t-test), *, P <0.01; Bit-1 KO vs. Bcl-2. (n=5 for each variant: Bit-1 sirna; Scrambled sirna; Scrambled sirna + Bcl-2; Bit-1 sirna + Bcl-2).
37 Figure 6. Bcl-2 expression rescues Bit-1 null-mediated early myogenesis. A) Western blot analysis of Bcl-2 expression levels in isolated WT and Bit-1 KO myoblasts at pre-differentiation day 0. Actin was used as a loading control. B) An increase in caspase 3 activity, as determined by a fluorometric protease assay that directly measures caspase 3 activity, was found at pre-differentiation day 0 and differentiation days 1 and 3 in Bit-1 KO myoblasts (open circle) compared to agematched WT littermates (closed diamond). Expression of Bcl-2 (open square) reduced caspase 3 activity to control levels. Statistically significant differences are reported in the graphs as P values (Student s t-test), *, P <0.01; Bit-1 KO vs. Bcl-2. (n=5 for each genotype). C) Pro-caspase 9 and cleaved caspase 9 were increased in isolated Bit-1 KO myoblasts at pre-differentiation day 0. Tubulin was used as a loading control. D) An increase in caspase 9 activity was found at pre-differentiation day 0 and differentiation days 1 and 3 in Bit-1 KO myoblasts (open circle) compared to age-matched WT littermates (closed diamond). Expression of Bcl-2 (open square) reduced caspase 9 activity towards control levels. Statistically significant differences are reported in the graphs as P values (Student s t-test), *, P <0.01; Bit-1 KO vs. Bcl-2. (n=5 for each genotype).
38
39
40
ƒ ƒ ƒ ƒ Š ƒ Š Ž Ž ƒ ƒ ƒ ˆ ƒ ƒž ƒ ƒ ƒž ƒ Š ƒ Š ƒ ƒ ƒ ƒ ŽŽ ƒ ƒ ƒ ƒ ƒ ƒ ƒ ˆ Š Š ƒ ƒ ƒ Žƒ Ž ƒ Š ƒ ƒž Ž ƒ Ž ƒ ƒ Š ƒ ˆ Š Š Š ƒ š Ž ƒ ƒ Š Š ƒ Ž ƒ Ž š ƒž š Š
 Š ƒžˆ Š Ž ƒ ƒ Ž Š ˆ Š ƒ d KEdydd d &hed/ke>^z/wd/kek&d,^zs/dk/dw>ded d d d &hed/ke>>k/dzKEdZK>E^^hZE dd d d Z,/s/E' dd d d WZKhd/^^D/Ed/KEdd d d h^zde'ded dd d e /EdZ&^ de d e ^Zs/sK>hd/KE
Š ƒžˆ Š Ž ƒ ƒ Ž Š ˆ Š ƒ d KEdydd d &hed/ke>^z/wd/kek&d,^zs/dk/dw>ded d d d &hed/ke>>k/dzKEdZK>E^^hZE dd d d Z,/s/E' dd d d WZKhd/^^D/Ed/KEdd d d h^zde'ded dd d e /EdZ&^ de d e ^Zs/sK>hd/KE
The Caspase System: a potential role in muscle proteolysis and meat quality? Tim Parr
 The Caspase System: a potential role in muscle proteolysis and meat quality? Tim Parr Caroline Kemp, Ron Bardsley,, Peter Buttery Division of Nutritional Sciences, School of Biosciences, University of
The Caspase System: a potential role in muscle proteolysis and meat quality? Tim Parr Caroline Kemp, Ron Bardsley,, Peter Buttery Division of Nutritional Sciences, School of Biosciences, University of
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
Cell Death & Trophic Factors II. Steven McLoon Department of Neuroscience University of Minnesota
 Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
F ST. ] = p anc Var(p s s. p anc. (1 p anc ) = F ST + F ST. ] p anc
![F ST. ] = p anc Var(p s s. p anc. (1 p anc ) = F ST + F ST. ] p anc](/thumbs/74/71246024.jpg "F ST. ] = p anc Var(p s s. p anc. (1 p anc ) = F ST + F ST. ] p anc") ƒ ƒ Š ƒ ˆƒ ƒ ƒ ƒ ƒ Šƒ ƒ ƒ ƒ ƒƒ ƒ ƒ ƒ Ž ƒ ƒ ƒ ƒ Š ˆ Š Ž ˆ ƒž Š ƒ Š Ž ƒ ƒ ˆƒ ƒ ƒ ƒ ƒ ˆ Ž ƒ ƒ Š Ž ˆ Ž ƒž Š ƒ ˆ ƒ ƒ ƒ Š Ž ˆ Ž ƒž Š ƒ ˆ ƒ ƒ ƒž Š Ž Š ƒ Š ƒžž Š Š Ž ƒ Šƒ ƒ ƒ ƒ Š Š Šƒ ƒ Ž ƒ ƒ Žƒ Žƒ ˆˆ ƒ ƒ ƒž Ž
ƒ ƒ Š ƒ ˆƒ ƒ ƒ ƒ ƒ Šƒ ƒ ƒ ƒ ƒƒ ƒ ƒ ƒ Ž ƒ ƒ ƒ ƒ Š ˆ Š Ž ˆ ƒž Š ƒ Š Ž ƒ ƒ ˆƒ ƒ ƒ ƒ ƒ ˆ Ž ƒ ƒ Š Ž ˆ Ž ƒž Š ƒ ˆ ƒ ƒ ƒ Š Ž ˆ Ž ƒž Š ƒ ˆ ƒ ƒ ƒž Š Ž Š ƒ Š ƒžž Š Š Ž ƒ Šƒ ƒ ƒ ƒ Š Š Šƒ ƒ Ž ƒ ƒ Žƒ Žƒ ˆˆ ƒ ƒ ƒž Ž
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Supplemental Figures S1 S5
 Beyond reduction of atherosclerosis: PON2 provides apoptosis resistance and stabilizes tumor cells Ines Witte (1), Sebastian Altenhöfer (1), Petra Wilgenbus (1), Julianna Amort (1), Albrecht M. Clement
Beyond reduction of atherosclerosis: PON2 provides apoptosis resistance and stabilizes tumor cells Ines Witte (1), Sebastian Altenhöfer (1), Petra Wilgenbus (1), Julianna Amort (1), Albrecht M. Clement
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1
 and Ksp/Cre;Pkd1") Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
FSC-W FSC-H CD4 CD62-L
 Supplementary Fig. 1 a SSC-A FSC-A FSC-W FSC-H SSC-W SSC-H CD4 CD62-L b SSC-A FSC-A FSC-W FSC-A FSC-A 7-AAD FSC-A CD4 IL-9 CD4 c SSC-A FSC-A FSC-W FSC-H SSC-W SSC-H 7-AAD KI67 Annexin-V 7-AAD d I L -5
Supplementary Fig. 1 a SSC-A FSC-A FSC-W FSC-H SSC-W SSC-H CD4 CD62-L b SSC-A FSC-A FSC-W FSC-A FSC-A 7-AAD FSC-A CD4 IL-9 CD4 c SSC-A FSC-A FSC-W FSC-H SSC-W SSC-H 7-AAD KI67 Annexin-V 7-AAD d I L -5
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11419 Supplementary Figure 1 Schematic representation of innate immune signaling pathways induced by intracellular Salmonella in cultured macrophages. a, During the infection Salmonella
doi:10.1038/nature11419 Supplementary Figure 1 Schematic representation of innate immune signaling pathways induced by intracellular Salmonella in cultured macrophages. a, During the infection Salmonella
Caspase Substrate Assay Kit (Colorimetric)
") Caspase Substrate Assay Kit (Colorimetric) Catalog Number KA3694 7 x 25 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Caspase Substrate Assay Kit (Colorimetric) Catalog Number KA3694 7 x 25 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
NucView TM 488 Caspase-3 Assay Kit for Live Cells
 NucView TM 488 Caspase-3 Assay Kit for Live Cells Catalog Number: 30029 (100-500 assays) Contact Information Address: Biotium, Inc. 3423 Investment Blvd. Suite 8 Hayward, CA 94545 USA Telephone: (510)
NucView TM 488 Caspase-3 Assay Kit for Live Cells Catalog Number: 30029 (100-500 assays) Contact Information Address: Biotium, Inc. 3423 Investment Blvd. Suite 8 Hayward, CA 94545 USA Telephone: (510)
TNFα 18hr. Control. CHX 18hr. TNFα+ CHX 18hr. TNFα: 18 18hr (KDa) PARP. Cleaved. Cleaved. Cleaved. Caspase3. Pellino3 shrna. Control shrna.
 PARP. Cleaved. Cleaved. Cleaved. Caspase3. Pellino3 shrna. Control shrna.") Survival ( %) a. TNFα 18hr b. Control sirna Pellino3 sirna TNFα: 18 18hr c. Control shrna Pellino3 shrna Caspase3 Actin Control d. Control shrna Pellino3 shrna *** 100 80 60 CHX 18hr 40 TNFα+ CHX 18hr
Survival ( %) a. TNFα 18hr b. Control sirna Pellino3 sirna TNFα: 18 18hr c. Control shrna Pellino3 shrna Caspase3 Actin Control d. Control shrna Pellino3 shrna *** 100 80 60 CHX 18hr 40 TNFα+ CHX 18hr
HIF-1 can act as a tumor suppressor gene in murine Acute Myeloid Leukemia.
 HIF-1 can act as a tumor suppressor gene in murine Acute Myeloid Leukemia. Velasco, Talia; Hyrenius Wittsten, Axel; Rehn, Matilda; Bryder, David; Cammenga, Jörg Published in: Blood DOI: 1.1182/blood-14-4-56765
HIF-1 can act as a tumor suppressor gene in murine Acute Myeloid Leukemia. Velasco, Talia; Hyrenius Wittsten, Axel; Rehn, Matilda; Bryder, David; Cammenga, Jörg Published in: Blood DOI: 1.1182/blood-14-4-56765
CELL REPLICATION. Fluorescent light microscopy showing mitosis, especially immunolabelled cytoskeleton and tubulin
 CELL REPLICATION Fluorescent light microscopy showing mitosis, especially immunolabelled cytoskeleton and tubulin Cell REPLICATION PROLIFERATION MUTIPLICATION DIVISION CELL REPLICATION Fluorescent light
CELL REPLICATION Fluorescent light microscopy showing mitosis, especially immunolabelled cytoskeleton and tubulin Cell REPLICATION PROLIFERATION MUTIPLICATION DIVISION CELL REPLICATION Fluorescent light
For the rapid, sensitive and accurate measurement of Caspase 3 Activity in cell and tissue lysates
 ab39383 Caspase 3 Assay Kit (Fluorometric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase 3 Activity in cell and tissue lysates This product is for research use only
ab39383 Caspase 3 Assay Kit (Fluorometric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase 3 Activity in cell and tissue lysates This product is for research use only
RayBio CaspGLOW TM Fluorescein Active Caspase-3 Staining Kit
 RayBio CaspGLOW TM Fluorescein Active Caspase-3 Staining Kit User Manual Version 1.0 May 10 th, 2015 RayBio Caspase-3 Fluorometric Assay Kit Protocol (Cat#: 68FLS-Casp3-S) RayBiotech, Inc. We Provide You
RayBio CaspGLOW TM Fluorescein Active Caspase-3 Staining Kit User Manual Version 1.0 May 10 th, 2015 RayBio Caspase-3 Fluorometric Assay Kit Protocol (Cat#: 68FLS-Casp3-S) RayBiotech, Inc. We Provide You
Death Ligand- Death Receptor. Annexin V. Fas PI MKK7 PIP3. pro-caspase-3. Granzyme B. Caspase Activation. Bid. Bcl-2 Caspase-3 Caspase-12
 International Corporation life. science. discovery. { Apoptosis Apoptosis Kits International Corporation Apoptosis Signal Cascade and Detection Kits Annexin V Kits MEBCYTO Apoptosis Kit Annexin V Apoptosis
International Corporation life. science. discovery. { Apoptosis Apoptosis Kits International Corporation Apoptosis Signal Cascade and Detection Kits Annexin V Kits MEBCYTO Apoptosis Kit Annexin V Apoptosis
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates
 ab65607 Caspase 9 Assay Kit (Fluorometric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates This product is for research use only
ab65607 Caspase 9 Assay Kit (Fluorometric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates This product is for research use only
Supplementary Figure 1.
 Supplementary Figure 1. Characterisation of IHG-1 overexpressing and knockdown cell lines. (A) Total cellular RNA was prepared from HeLa cells stably overexpressing IHG-1 or mts-ihg-1. IHG-1 mrna was quantified
Supplementary Figure 1. Characterisation of IHG-1 overexpressing and knockdown cell lines. (A) Total cellular RNA was prepared from HeLa cells stably overexpressing IHG-1 or mts-ihg-1. IHG-1 mrna was quantified
Apoptosis Detection Using the BD Accuri C6 Flow Cytometer
 Apoptosis Detection Using the BD Accuri C6 Flow Cytometer Stacey Roys, Marketing Applications Specialist, BD Biosciences Cyndy Lane, Senior Product Manager, BD Biosciences 23-14519-00 Outline What is apoptosis?
Apoptosis Detection Using the BD Accuri C6 Flow Cytometer Stacey Roys, Marketing Applications Specialist, BD Biosciences Cyndy Lane, Senior Product Manager, BD Biosciences 23-14519-00 Outline What is apoptosis?
Supplemental Data. Perrella et al. (2013). Plant Cell /tpc
. Plant Cell /tpc") Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody
 Based on Monoclonal Anti-cGMP Antibody") (FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
(FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
ydci GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC
 Table S1. DNA primers used in this study. Name ydci P1ydcIkd3 Sequence GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC Kd3ydcIp2 lacz fusion YdcIendP1 YdcItrgP2 GAC AGC
Table S1. DNA primers used in this study. Name ydci P1ydcIkd3 Sequence GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC Kd3ydcIp2 lacz fusion YdcIendP1 YdcItrgP2 GAC AGC
For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates.
 ab65608 Caspase 9 Assay Kit (Colorimetric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates. This product is for research use only
ab65608 Caspase 9 Assay Kit (Colorimetric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates. This product is for research use only
Caspase 3 (active) Red Staining Kit
 Red Staining Kit") ab65617 Caspase 3 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 3 in living cells. This product is for research use only and is not
ab65617 Caspase 3 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 3 in living cells. This product is for research use only and is not
Zool 3200: Cell Biology Exam 5 4/27/15
 Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Apoptosis: Death Comes for the Cell
 Apoptosis: Death Comes for the Cell Joe W. Ramos joeramos@hawaii.edu From Ingmar Bergman s The Seventh Seal 1 2 Mutations in proteins that regulate cell proliferation, survival and death can contribute
Apoptosis: Death Comes for the Cell Joe W. Ramos joeramos@hawaii.edu From Ingmar Bergman s The Seventh Seal 1 2 Mutations in proteins that regulate cell proliferation, survival and death can contribute
Nature Neuroscience: doi: /nn.2662
 Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Information
 Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Nature Medicine: doi: /nm.3776
 C terminal Hsp90 inhibitors restore glucocorticoid sensitivity and relieve a mouse allograft model of Cushing s disease Mathias Riebold, Christian Kozany, Lee Freiburger, Michael Sattler, Michael Buchfelder,
C terminal Hsp90 inhibitors restore glucocorticoid sensitivity and relieve a mouse allograft model of Cushing s disease Mathias Riebold, Christian Kozany, Lee Freiburger, Michael Sattler, Michael Buchfelder,
DNA DAMAGE-INDUCED APOPTOSIS: INHIBITION BY CALMODULIN ANTAGONIST, FAS RECEPTOR ANTIBODY AND CASPASE INHIBITORS
 DNA DAMAGE-INDUCED APOPTOSIS: INHIBITION BY CALMODULIN ANTAGONIST, FAS RECEPTOR ANTIBODY AND CASPASE INHIBITORS R. Ray, B. J. Benton, M. E. Burke, T. Rockwood, K. R. Bhat, D. R. Anderson, J. P. Petrali,
DNA DAMAGE-INDUCED APOPTOSIS: INHIBITION BY CALMODULIN ANTAGONIST, FAS RECEPTOR ANTIBODY AND CASPASE INHIBITORS R. Ray, B. J. Benton, M. E. Burke, T. Rockwood, K. R. Bhat, D. R. Anderson, J. P. Petrali,
Supplemental Information. The Mitochondrial Fission Receptor MiD51. Requires ADP as a Cofactor
 Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Technical Bulletin. Caspase 9 Fluorescein (FLICA) Assay. Catalog Number CS0300 Storage Temperature 2-8 C
 Assay. Catalog Number CS0300 Storage Temperature 2-8 C") 1 Caspase 9 Fluorescein (FLICA) Assay Catalog Number CS0300 Storage Temperature 2-8 C Technical Bulletin Product Description Caspases are detected by immunoprecipitation, immunoblotting with caspase specific
1 Caspase 9 Fluorescein (FLICA) Assay Catalog Number CS0300 Storage Temperature 2-8 C Technical Bulletin Product Description Caspases are detected by immunoprecipitation, immunoblotting with caspase specific
SUPPLEMENTAL MATERIAL
 SUPPLEMENTAL MATERIAL Figure S1. Mitochondrial morphology in Fis1-null, Mff-null and Fis1/Mff-null MEF cells. (A) Western blotting of lysates from Fis1-null, Mff-null and Fis1/Mff-null cells. Lysates were
SUPPLEMENTAL MATERIAL Figure S1. Mitochondrial morphology in Fis1-null, Mff-null and Fis1/Mff-null MEF cells. (A) Western blotting of lysates from Fis1-null, Mff-null and Fis1/Mff-null cells. Lysates were
Caspase 7 Inhibitor Drug Detection Kit
 ab102495 Caspase 7 Inhibitor Drug Detection Kit Instructions for Use For the rapid, sensitive and accurate detection of Caspase inhibitors in various samples This product is for research use only and is
ab102495 Caspase 7 Inhibitor Drug Detection Kit Instructions for Use For the rapid, sensitive and accurate detection of Caspase inhibitors in various samples This product is for research use only and is
Caspase 8 (active) Red Staining Kit
 Red Staining Kit") ab65618 Caspase 8 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 8 in living cells This product is for research use only and is not
ab65618 Caspase 8 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 8 in living cells This product is for research use only and is not
Caspase Substrate Kit (Colorimetric)
") ab102486 Caspase Substrate Kit (Colorimetric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase activity in cell culture This product is for research use only and is not
ab102486 Caspase Substrate Kit (Colorimetric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase activity in cell culture This product is for research use only and is not
RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit
 ELISA Kit") RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit Catalog #: PEL-EIF2A-S52 User Manual Last revised August 10, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway
RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit Catalog #: PEL-EIF2A-S52 User Manual Last revised August 10, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway
For the rapid, sensitive and accurate detection of active Caspase 9 in living cells
 ab65615 Caspase 9 (active) FITC Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of active Caspase 9 in living cells This product is for research use only and is not intended
ab65615 Caspase 9 (active) FITC Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of active Caspase 9 in living cells This product is for research use only and is not intended
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
For the rapid, sensitive and accurate detection of Caspase 5 inhibition by various compounds
 ab102493 Caspase 5 Inhibitor Drug Detection Kit Instructions for Use For the rapid, sensitive and accurate detection of Caspase 5 inhibition by various compounds This product is for research use only and
ab102493 Caspase 5 Inhibitor Drug Detection Kit Instructions for Use For the rapid, sensitive and accurate detection of Caspase 5 inhibition by various compounds This product is for research use only and
Nexcelom ViaStain Live Caspase 3/7 Detection for 2D/3D Culture
 Nexcelom ViaStain Live Caspase 3/7 Detection for 2D/3D Culture Product Numbers: CSK-V0002-1, CSK-V0003-1 This product is for RESEARCH USE ONLY and is not approved for diagnostic or therapeutic use. Table
Nexcelom ViaStain Live Caspase 3/7 Detection for 2D/3D Culture Product Numbers: CSK-V0002-1, CSK-V0003-1 This product is for RESEARCH USE ONLY and is not approved for diagnostic or therapeutic use. Table
Technical Bulletin. Caspase 13 Fluorescein (FLICA) Assay. Catalog Number CS0320 Storage Temperature 2-8 C
 Assay. Catalog Number CS0320 Storage Temperature 2-8 C") Caspase 13 Fluorescein (FLICA) Assay Catalog Number CS0320 Storage Temperature 2-8 C Technical Bulletin Product Description Caspases are detected by immunoprecipitation, immunoblotting with caspase specific
Caspase 13 Fluorescein (FLICA) Assay Catalog Number CS0320 Storage Temperature 2-8 C Technical Bulletin Product Description Caspases are detected by immunoprecipitation, immunoblotting with caspase specific
Version 1.1. Caspase-9 Fluorometric Assay Kit KC test
 Caspase-9 Fluorometric Assay Kit KC-04-012 100-200-500 test Index Introduction Pag. 1 Materials Pag. 2 Assay Principle Pag. 3 Reagent preparation Pag. 4 Assay protocol Pag. 5 Data analysis Pag. 6 Warranties
Caspase-9 Fluorometric Assay Kit KC-04-012 100-200-500 test Index Introduction Pag. 1 Materials Pag. 2 Assay Principle Pag. 3 Reagent preparation Pag. 4 Assay protocol Pag. 5 Data analysis Pag. 6 Warranties
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Supplementary Figure 1. AnnexinV FITC and Sytox orange staining in wild type, Nlrp3 /, ASC / and casp1/11 / TEC treated with TNF /CHX.
 Supplementary Figure 1. AnnexinV FITC and Sytox orange staining in wild type, Nlrp3 /, ASC / and casp1/11 / TEC treated with TNF /CHX. Phase contrast and widefield fluorescence microscopy (20x magnification).
Supplementary Figure 1. AnnexinV FITC and Sytox orange staining in wild type, Nlrp3 /, ASC / and casp1/11 / TEC treated with TNF /CHX. Phase contrast and widefield fluorescence microscopy (20x magnification).
Examination paper for Bi3016 Molecular Cell Biology
 Department of Biology Examination paper for Bi3016 Molecular Cell Biology Academic contact during examination: Per Winge Phone: 99369359 Examination date: 20 th December 2017 Examination time (from-to):
Department of Biology Examination paper for Bi3016 Molecular Cell Biology Academic contact during examination: Per Winge Phone: 99369359 Examination date: 20 th December 2017 Examination time (from-to):
Apoptosis in Mammalian Cells
 Apoptosis in Mammalian Cells 7.16 2-10-05 Apoptosis is an important factor in many human diseases Cancer malignant cells evade death by suppressing apoptosis (too little apoptosis) Stroke damaged neurons
Apoptosis in Mammalian Cells 7.16 2-10-05 Apoptosis is an important factor in many human diseases Cancer malignant cells evade death by suppressing apoptosis (too little apoptosis) Stroke damaged neurons
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Proteome-wide High Throughput Cell Based Assay for Apoptotic Genes
 John Kenten, Doug Woods, Pankaj Oberoi, Laura Schaefer, Jonathan Reeves, Hans A. Biebuyck and Jacob N. Wohlstadter 9238 Gaither Road, Gaithersburg, MD 2877. Phone: 24.631.2522 Fax: 24.632.2219. Website:
John Kenten, Doug Woods, Pankaj Oberoi, Laura Schaefer, Jonathan Reeves, Hans A. Biebuyck and Jacob N. Wohlstadter 9238 Gaither Road, Gaithersburg, MD 2877. Phone: 24.631.2522 Fax: 24.632.2219. Website:
BIOH111. o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system
 BIOH111 o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 Textbook
BIOH111 o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 Textbook
Programmed Cell Death
 Programmed Cell Death Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id What is apoptosis? a normal component of the development
Programmed Cell Death Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id What is apoptosis? a normal component of the development
protein biology cell imaging Automated imaging and high-content analysis
 protein biology cell imaging Automated imaging and high-content analysis Automated imaging and high-content analysis Thermo Fisher Scientific offers a complete portfolio of imaging platforms, reagents
protein biology cell imaging Automated imaging and high-content analysis Automated imaging and high-content analysis Thermo Fisher Scientific offers a complete portfolio of imaging platforms, reagents
Mitochondrial Dynamics Is a Distinguishing Feature of Skeletal Muscle Fiber Types and Regulates Organellar Compartmentalization
 Cell Metabolism Supplemental Information Mitochondrial Dynamics Is a Distinguishing Feature of Skeletal Muscle Fiber Types and Regulates Organellar Compartmentalization Prashant Mishra, Grigor Varuzhanyan,
Cell Metabolism Supplemental Information Mitochondrial Dynamics Is a Distinguishing Feature of Skeletal Muscle Fiber Types and Regulates Organellar Compartmentalization Prashant Mishra, Grigor Varuzhanyan,
Cells to Tissues. Peter Takizawa Department of Cell Biology
 Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Human amyloid beta peptide 1-42, Aβ1-42 ELISA Kit
 Human amyloid beta peptide 1-42, Aβ1-42 ELISA Kit Catalog No: E0946h 96 Tests Operating instruction www.eiaab.com FOR RESEARCH USE ONLY; NOT FOR THERAPEUTIC OR DIAGNOSTIC APPLICATIONS! PLEASE READ THROUGH
Human amyloid beta peptide 1-42, Aβ1-42 ELISA Kit Catalog No: E0946h 96 Tests Operating instruction www.eiaab.com FOR RESEARCH USE ONLY; NOT FOR THERAPEUTIC OR DIAGNOSTIC APPLICATIONS! PLEASE READ THROUGH
CC2.04_t.txt The Caspase System and Muscle Degradation Tim Parr
 The Caspase System and Muscle Degradation Tim Parr [1]Our last speaker of the day is Dr. Tim Parr. Dr. Parr graduated in 1987 from the University of Southampton with a degree in physiology, biochemistry,
The Caspase System and Muscle Degradation Tim Parr [1]Our last speaker of the day is Dr. Tim Parr. Dr. Parr graduated in 1987 from the University of Southampton with a degree in physiology, biochemistry,
Biology 580 Cellular Physiology Spring 2007 Course Syllabus
 Class # 17416 MW 0800-0850 Biology 580 Cellular Physiology Spring 2007 Course Syllabus Course Text: Course Website: Molecular Biology of the Cell by Alberts et al.; 4th edition, Garland Science, 2002 (ISBN
Class # 17416 MW 0800-0850 Biology 580 Cellular Physiology Spring 2007 Course Syllabus Course Text: Course Website: Molecular Biology of the Cell by Alberts et al.; 4th edition, Garland Science, 2002 (ISBN
Caspase 3 (active) Red Staining Kit
 Red Staining Kit") ab65617 Caspase 3 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 3 in living cells. This product is for research use only and is not
ab65617 Caspase 3 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 3 in living cells. This product is for research use only and is not
Supplemental Data. Chen and Thelen (2010). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Data. Chen and Thelen (2010). Plant Cell 10.1105/tpc.109.071837 1 C Total 5 kg 20 kg 100 kg Transmission Image 100 kg soluble pdtpi-gfp Plastid (PDH-alpha) Mito (PDH-alpha) GFP Image vector
Supplemental Data. Chen and Thelen (2010). Plant Cell 10.1105/tpc.109.071837 1 C Total 5 kg 20 kg 100 kg Transmission Image 100 kg soluble pdtpi-gfp Plastid (PDH-alpha) Mito (PDH-alpha) GFP Image vector
SUPPORTING INFORMATION FOR. SEquence-Enabled Reassembly of β-lactamase (SEER-LAC): a Sensitive Method for the Detection of Double-Stranded DNA
: a Sensitive Method for the Detection of Double-Stranded DNA") SUPPORTING INFORMATION FOR SEquence-Enabled Reassembly of β-lactamase (SEER-LAC): a Sensitive Method for the Detection of Double-Stranded DNA Aik T. Ooi, Cliff I. Stains, Indraneel Ghosh *, David J. Segal
SUPPORTING INFORMATION FOR SEquence-Enabled Reassembly of β-lactamase (SEER-LAC): a Sensitive Method for the Detection of Double-Stranded DNA Aik T. Ooi, Cliff I. Stains, Indraneel Ghosh *, David J. Segal
U.S. Patent No. 9,051,563 and other pending patents. Ver
 INSTRUCTION MANUAL Direct-zol 96 RNA Catalog Nos. R2054, R2055, R2056 & R2057 Highlights Quick, 96-well purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and all other
INSTRUCTION MANUAL Direct-zol 96 RNA Catalog Nos. R2054, R2055, R2056 & R2057 Highlights Quick, 96-well purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and all other
Signal transduction by Cell-Cell and Cell-Matrix contacts
 Signal transduction by Cell-Cell and Cell-Matrix contacts - Integrins as central adhesion receptors - - Focal Adhesion Kinase (FAK) as a central signal molecule - Structure of Cell/Cell and Cell/Matrix
Signal transduction by Cell-Cell and Cell-Matrix contacts - Integrins as central adhesion receptors - - Focal Adhesion Kinase (FAK) as a central signal molecule - Structure of Cell/Cell and Cell/Matrix
Protease Inhibitor Cocktail A (1 tablet / 7 10 ml, Roche Cat# ) Protease inhibitor Cocktail B (0.5ml per 250ml, Calbiochem Cat# )
 Protease inhibitor Cocktail B (0.5ml per 250ml, Calbiochem Cat# )") Protocol for Western Blotting Tissue/Cell Sample Preparation Lysis Buffer 1 (ph8.0) o 50mM Tris-Cl o 150mM NaCl o 1% v/v NP40 o protease inhibitor cocktail A/B Lysis Buffer 2 (RIPA) (ph 8.0) o 50mM Tris-Cl
Protocol for Western Blotting Tissue/Cell Sample Preparation Lysis Buffer 1 (ph8.0) o 50mM Tris-Cl o 150mM NaCl o 1% v/v NP40 o protease inhibitor cocktail A/B Lysis Buffer 2 (RIPA) (ph 8.0) o 50mM Tris-Cl
4) Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.
 Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.") Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
Scholarly Activity Improvement Fund (SAIF)-A 2012/2013 Final Report. Blocking signal pathway inter-connectors to reverse cell death
-A 2012/2013 Final Report. Blocking signal pathway inter-connectors to reverse cell death") Scholarly Activity Improvement Fund (SAIF)-A 2012/2013 Final Report Blocking signal pathway inter-connectors to reverse cell death Chanaka Mendis Professor of Chemistry Department of Chemistry & Engineering
Scholarly Activity Improvement Fund (SAIF)-A 2012/2013 Final Report Blocking signal pathway inter-connectors to reverse cell death Chanaka Mendis Professor of Chemistry Department of Chemistry & Engineering
Published Ahead of Print on May 24, 2018, as doi: /haematol Copyright 2018 Ferrata Storti Foundation.
 Published Ahead of Print on May 24, 2018, as doi:10.3324/haematol.2018.193151. Copyright 2018 Ferrata Storti Foundation. Reducing mortality in newly diagnosed standard-risk acute promyelocytic leukemia
Published Ahead of Print on May 24, 2018, as doi:10.3324/haematol.2018.193151. Copyright 2018 Ferrata Storti Foundation. Reducing mortality in newly diagnosed standard-risk acute promyelocytic leukemia
Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering
 Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering by Valverde et. Al Published in Science 2004 Presented by Boyana Grigorova CBMG 688R Feb. 12, 2007 Circadian Rhythms: The Clock Within
Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering by Valverde et. Al Published in Science 2004 Presented by Boyana Grigorova CBMG 688R Feb. 12, 2007 Circadian Rhythms: The Clock Within
Proteomics. Areas of Interest
 Introduction to BioMEMS & Medical Microdevices Proteomics and Protein Microarrays Companion lecture to the textbook: Fundamentals of BioMEMS and Medical Microdevices, by Prof., http://saliterman.umn.edu/
Introduction to BioMEMS & Medical Microdevices Proteomics and Protein Microarrays Companion lecture to the textbook: Fundamentals of BioMEMS and Medical Microdevices, by Prof., http://saliterman.umn.edu/
Cell Survival Curves (Chap. 3)
") Cell Survival Curves (Chap. 3) Example of pedigress of EMT6 mouse cells Clone The in vitro cell survival curve Cell Survival Assay Dye exclusion assay: membrane integrity MTT assay: respiratory activity
Cell Survival Curves (Chap. 3) Example of pedigress of EMT6 mouse cells Clone The in vitro cell survival curve Cell Survival Assay Dye exclusion assay: membrane integrity MTT assay: respiratory activity
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
FINAL REPORT For Japan-Korea Joint Research Project AREA
 (Form4-2) FINAL REPORT For Japan-Korea Joint Research Project AREA 1. Mathematics & Physics 2. Chemistry & Material Science 3. Biology 4. Informatics & Mechatronics 5. Geo-Science & Space Science 6. Medical
(Form4-2) FINAL REPORT For Japan-Korea Joint Research Project AREA 1. Mathematics & Physics 2. Chemistry & Material Science 3. Biology 4. Informatics & Mechatronics 5. Geo-Science & Space Science 6. Medical
Caspase 3 Inhibitor Drug Detection Kit
 ab102491 Caspase 3 Inhibitor Drug Detection Kit Instructions for Use For the rapid, sensitive and accurate detection of Caspase 3 inhibition by various compounds This product is for research use only and
ab102491 Caspase 3 Inhibitor Drug Detection Kit Instructions for Use For the rapid, sensitive and accurate detection of Caspase 3 inhibition by various compounds This product is for research use only and
Human Toll-like receptor 4, TLR4 ELISA Kit
 Human Toll-like receptor 4, TLR4 ELISA Kit Catalog No: E0753h 96 Tests Operating instruction www.eiaab.com FOR RESEARCH USE ONLY; NOT FOR THERAPEUTIC OR DIAGNOSTIC APPLICATIONS! PLEASE READ THROUGH ENTIRE
Human Toll-like receptor 4, TLR4 ELISA Kit Catalog No: E0753h 96 Tests Operating instruction www.eiaab.com FOR RESEARCH USE ONLY; NOT FOR THERAPEUTIC OR DIAGNOSTIC APPLICATIONS! PLEASE READ THROUGH ENTIRE
 http://cwp.embo.org/w09-13/index.html Roads to Ruin is s o t p o ap healthy cell autophagic cell death ne cr os is Thanks to Seamus Martin! Evolution of apoptosis signalling cascades Inhibitor Adopted
http://cwp.embo.org/w09-13/index.html Roads to Ruin is s o t p o ap healthy cell autophagic cell death ne cr os is Thanks to Seamus Martin! Evolution of apoptosis signalling cascades Inhibitor Adopted
Supporting Information
 Supporting Information Wang et al. 10.1073/pnas.0804871105 SI Materials and Methods Cell Culture and Transfection. Human neuroblastoma M17 cells were grown as described before (1). Transfection was performed
Supporting Information Wang et al. 10.1073/pnas.0804871105 SI Materials and Methods Cell Culture and Transfection. Human neuroblastoma M17 cells were grown as described before (1). Transfection was performed
RayBio Protease Activity Assay Kit
 RayBio Protease Activity Assay Kit User Manual Version 1.0 Mar 25, 2013 RayBio Protease Activity Assay (Cat#: 68AT-Protease-S100) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
RayBio Protease Activity Assay Kit User Manual Version 1.0 Mar 25, 2013 RayBio Protease Activity Assay (Cat#: 68AT-Protease-S100) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
What are mitochondria?
 What are mitochondria? What are mitochondria? An intracellular organelle. There are 100 to 1000s of mitochondria/cell. Most mitochondria come from the mother. Mitochondria have their own DNA Mitochondria
What are mitochondria? What are mitochondria? An intracellular organelle. There are 100 to 1000s of mitochondria/cell. Most mitochondria come from the mother. Mitochondria have their own DNA Mitochondria
Supplemental material
 Supplemental material THE JOURNAL OF CELL BIOLOGY Mourier et al., http://www.jcb.org/cgi/content/full/jcb.201411100/dc1 Figure S1. Size and mitochondrial content in Mfn1 and Mfn2 knockout hearts. (A) Body
Supplemental material THE JOURNAL OF CELL BIOLOGY Mourier et al., http://www.jcb.org/cgi/content/full/jcb.201411100/dc1 Figure S1. Size and mitochondrial content in Mfn1 and Mfn2 knockout hearts. (A) Body
Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.
 Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.441/HST522J INTEGRINS I.V. Yannas, Ph.D. and M. Spector, Ph.D. Regulator
Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.441/HST522J INTEGRINS I.V. Yannas, Ph.D. and M. Spector, Ph.D. Regulator
Bio-Plex Pro RBM Apoptosis Assays
 Acute Phase Apoptosis Cancer Cardiovascular Disease Cytokines Chemokines, Growth Factors Diabetes Gene Expression Genotyping Immunoglobulin Isotyping MicroRNA Expression Bio-Plex Pro RBM Apoptosis Assays
Acute Phase Apoptosis Cancer Cardiovascular Disease Cytokines Chemokines, Growth Factors Diabetes Gene Expression Genotyping Immunoglobulin Isotyping MicroRNA Expression Bio-Plex Pro RBM Apoptosis Assays
Figure S1: Extracellular nicotinic acid, but not tryptophan, is sufficient to maintain
 SUPPLEMENTAL INFORMATION Supplemental Figure Legends Figure S1: Extracellular nicotinic acid, but not tryptophan, is sufficient to maintain mitochondrial NAD +. A) Extracellular tryptophan, even at 5 µm,
SUPPLEMENTAL INFORMATION Supplemental Figure Legends Figure S1: Extracellular nicotinic acid, but not tryptophan, is sufficient to maintain mitochondrial NAD +. A) Extracellular tryptophan, even at 5 µm,
RayBio Rat TNF-alpha ELISA Kit (For Lysates)
") RayBio Rat TNF-alpha ELISA Kit (For Lysates) Catalog #: ELR-TNFa-CL User Manual Last revised April 15, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite
RayBio Rat TNF-alpha ELISA Kit (For Lysates) Catalog #: ELR-TNFa-CL User Manual Last revised April 15, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite
targets. clustering show that different complex pathway
 Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Richik N. Ghosh, Linnette Grove, and Oleg Lapets ASSAY and Drug Development Technologies 2004, 2:
 1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
mrna Isolation Kit for Blood/Bone Marrow For isolation mrna from blood or bone marrow lysates Cat. No
 For isolation mrna from blood or bone marrow lysates Cat. No. 1 934 333 Principle Starting material Application Time required Results Key advantages The purification of mrna requires two steps: 1. Cells
For isolation mrna from blood or bone marrow lysates Cat. No. 1 934 333 Principle Starting material Application Time required Results Key advantages The purification of mrna requires two steps: 1. Cells
Massive loss of neurons in embryos occurs during normal development (!)
") Types of Cell Death Apoptosis (Programmed Cell Death) : Cell-Autonomous Stereotypic Rapid Clean (dead cells eaten) Necrosis : Not Self-Initiated Not Stereotypic Can Be Slow Messy (injury can spread) Apoptosis
Types of Cell Death Apoptosis (Programmed Cell Death) : Cell-Autonomous Stereotypic Rapid Clean (dead cells eaten) Necrosis : Not Self-Initiated Not Stereotypic Can Be Slow Messy (injury can spread) Apoptosis
Phospholipase C Fluorometric/Colorimetric Assay Kit
 Phospholipase C Fluorometric/Colorimetric Assay Kit Catalog No. KM0129 Detection and Quantification of Phospholipase C Concentrations in Biological Samples. Research Purposes Only. Not Intended for Diagnostic
Phospholipase C Fluorometric/Colorimetric Assay Kit Catalog No. KM0129 Detection and Quantification of Phospholipase C Concentrations in Biological Samples. Research Purposes Only. Not Intended for Diagnostic
Human papillomavirus,hpv ELISA Kit
 Human papillomavirus,hpv ELISA Kit Catalog No: E0787h 96 Tests Operating instructions www.eiaab.com FOR RESEARCH USE ONLY; NOT FOR THERAPEUTIC OR DIAGNOSTIC APPLICATIONS! PLEASE READ THROUGH ENTIRE PROCEDURE
Human papillomavirus,hpv ELISA Kit Catalog No: E0787h 96 Tests Operating instructions www.eiaab.com FOR RESEARCH USE ONLY; NOT FOR THERAPEUTIC OR DIAGNOSTIC APPLICATIONS! PLEASE READ THROUGH ENTIRE PROCEDURE
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Mechanisms of Cell Death
 Mechanisms of Cell Death CELL DEATH AND FORMATION OF THE SEMICIRCULAR CANALS Carol M. Troy, MD PhD August 24, 2009 FROM: Fekete et al., Development 124: 2451 (1997) PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT
Mechanisms of Cell Death CELL DEATH AND FORMATION OF THE SEMICIRCULAR CANALS Carol M. Troy, MD PhD August 24, 2009 FROM: Fekete et al., Development 124: 2451 (1997) PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT
** * * * Col-0 cau1 CAU1. Actin2 CAS. Actin2. Supplemental Figure 1. CAU1 affects calcium accumulation.
 Ca 2+ ug g -1 DW Ca 2+ ug g -1 DW Ca 2+ ug g -1 DW Supplemental Data. Fu et al. Plant Cell. (213). 1.115/tpc.113.113886 A 5 4 3 * Col- cau1 B 4 3 2 Col- cau1 ** * * ** C 2 1 25 2 15 1 5 Shoots Roots *
Ca 2+ ug g -1 DW Ca 2+ ug g -1 DW Ca 2+ ug g -1 DW Supplemental Data. Fu et al. Plant Cell. (213). 1.115/tpc.113.113886 A 5 4 3 * Col- cau1 B 4 3 2 Col- cau1 ** * * ** C 2 1 25 2 15 1 5 Shoots Roots *
ab Caspase 3, Caspase 8 and Caspase 9 Multiplex Activity Assay Kit (Fluorometric)
") Version 1 Last updated 6 April 2017 ab219915 Caspase 3, Caspase 8 and Caspase 9 Multiplex Activity Assay Kit (Fluorometric) For monitoring multiple caspase activation in live cells. This product is for
Version 1 Last updated 6 April 2017 ab219915 Caspase 3, Caspase 8 and Caspase 9 Multiplex Activity Assay Kit (Fluorometric) For monitoring multiple caspase activation in live cells. This product is for
Apoptosis: Death comes for the Cell. Apoptosis: Programmed Cell Death. Apoptosis. Necrosis
 poptosis: eath comes for the Cell Mutations in proteins that regulate cell proliferation, survival and death can contribute to oncogenesis Joe W. Ramos jramos@crch.hawaii.edu From Ingmar Bergman s The
poptosis: eath comes for the Cell Mutations in proteins that regulate cell proliferation, survival and death can contribute to oncogenesis Joe W. Ramos jramos@crch.hawaii.edu From Ingmar Bergman s The