Title: Pollen killer gene S35 function requires interaction with an activator that maps close to S24, another pollen killer gene in rice
|
|
|
- Colleen Wilkinson
- 5 years ago
- Views:
Transcription
1 G3: Genes Genomes Genetics Early Online, published on March 26, 2016 as doi: /g Title: Pollen killer gene S35 function requires interaction with an activator that maps close to S24, another pollen killer gene in rice Takahiko Kubo, *,,1, Atsushi Yoshimura, and Nori Kurata *, * Plant genetics Laboratory, National Institute of Genetics, Mishima, Shizuoka , Japan Department of Genetics, the Graduate University for Advanced Science (SOKENDAI), Mishima, Shizuoka , Japan Plant breeding Laboratory, Division of Genetics and Plant Breeding, Department of Applied Genetics and Pest Management, Faculty of Agriculture, Kyushu University, Fukuoka , Japan 1 The Author(s) Published by the Genetics Society of America.
2 Running title: S35-activator is linked to S24 Key words: pollen killer, Oryza sativa, epistasis, reproductive isolation 1 Corresponding author: Takahiko Kubo Plant genetics Laboratory, National Institute of Genetics, 1111 Yata, Mishima, Shizuoka , Japan TEL: FAX: takubo@nig.ac.jp 2
3 ABSTRACT Pollen killer genes disable non-carrier pollens and are responsible for male sterility and segregation distortion in hybrid populations of distantly related plant species. The genetic networks and the molecular mechanisms underlying the pollen killer system remain largely unknown. Two pollen killer genes, S24 and S35, have been found in an inter-subspecific cross of Oryza sativa ssp. indica and japonica. The effect of S24 is counteracted by an unlinked locus EFS. Additionally, S35 has been proposed to interact with S24 to induce pollen sterility. These genetic interactions are suggestive of a single S24-centric genetic pathway (EFS S24 S35) for the pollen killer system. To examine this hypothetical genetic pathway, the S35 and the S24 regions were further characterized and genetically dissected in this study. Our results indicated that S35 causes pollen sterility independently of both the EFS and S24 genes but is dependent on a novel gene close to the S24 locus, named incentive for killing pollen (INK). We confirmed the phenotypic effect of the INK gene separately from the S24 gene and identified the INK locus within an interval of less than 0.6 Mb on rice chromosome 5. This study characterized the genetic effect of the two independent genetic pathways of INK S35 and EFS S24 in indica japonica hybrid progeny. Our results provide clear evidence that hybrid male sterility in rice is caused by several pollen killer networks with multiple factors positively and negatively regulating pollen killer genes. 20 3
4 Introduction Hybrid progeny derived from genetically divergent species frequently suffer reproductive dysfunction. This phenomenon, referred to as hybrid sterility, is assumed to play an important role in the development of speciation by triggering postzygotic reproductive isolation. Segregation distorters, also called transmission ratio distortion factors, are evolutionarily selfish genetic elements that distort Mendelian segregation in their favor at the expense of others. Segregation distorters have been reported in a wide variety of reproductive organisms, including animals, plants, and fungi (Burt and Trivers 2009) and are one of the primary origins of hybrid sterility by causing one-half of the gametes to be dysfunctional. Two well-characterized genetic elements are Segregation distorter (Sd) in Drosophila (Larracuente and Presgraves 2012) and t-complex in mice (Lyon 2003). The role of segregation distorters in speciation of reproductive organisms has been the subject of considerable discussion (Burt and Trivers 2009). In plants, diverse types of segregation distorters have been identified in many plant species. Some examples include: pollen killer in Nicotiana (Cameron and Moav 1957), gamete eliminator in tomato (Rick 1966), gametocidal factor in wheat (Endo 2015), chromosomal knobs in maize (Buckler et al. 1999; Kanizay et al. 2013) and female meiotic drive in Mimulus (Fishman et al. 2008, Finseth et al. 2015). Gamete eliminator (Ge) (both pollen and egg are abortive) was found in a tomato intraspecific cross by plant geneticist Charles M. Rick who interpreted Ge as a triallelic system at a single locus (Rick 1966). The gamete eliminator (Ge p ) causes abortion of gametes carrying 4
5 Ge c in the Ge p /Ge c heterozygote. The third neutral allele Ge n is compatible with both Ge p and Ge c in the heterozygous hybrid. Since the neutral allele Ge n is widely distributed, Rick proposed that both Ge c and Ge p could arise by mutation from Ge n without adverse effect on gamete fertility (Rick 1966). Based on this idea, the killer and the abortive allele were proposed to be derived from a common ancestral allele that is compatible with both of them. The egg killer locus in rice, S5, is a type of triallelic system (Ikehashi and Araki 1986). The killer allele S5-i from indica causes female gamete abortion in the heterozygous state with the japonica allele but not with the neutral alleles from other varieties or the wild progenitor. Closer study of the S5 system unveiled the molecular mechanism. The S5 locus is composed of three tightly linked genes, and particular combinations of the alleles lead to endoplasmic reticulum stress, resulting in female gamete abortion (Yang et al. 2012). As a case study of male sterility, two adjacent genes that encode a SUMO E3 ligase-like protein and an F-box protein were identified in an indica/japonica cross (Long et al. 2008). These pioneering studies have provided important insights into the genetic architecture and the evolutionary history of plant gamete killers; namely, the existence of tightly linked multiple genes and cumulative mutations in these genes lead to the gamete killing phenotype in heterozygotes. In this scenario, not only the ancestral haplotype but also some of the derived haplotypes can work as compatible (neutral) haplotypes. In this context, several questions about segregation distorters remain to be explored. (1) What is a common or unique aspect of 5
6 the molecular mechanism? Previous studies have identified highly repetitive and heterochromatic sequences as common causal molecules, e.g. the D locus in Mimulus (Fishman and Saunders 2008), knobs in maize (Buckler et al. 1999), and HSR in mice (Weichenhan et al. 2001). (2) What are the normal functions of these genes within the parental species? (3) Why have a greater number of segregation distorters evolved in plant genomes compared with animal or fungal genomes? (4) Has domestication facilitated the development of segregation distorters? There are many segregation distorters found in crop species, especially in rice, and a previous study proposed an association between reproductive isolation and domestication (Dempewolf et al. 2012). Further studies extended to many more examples are needed to answer these questions. Asian cultivated rice (Oryza sativa L.) is an autogamous diploid species (2n = 24) and has two major subspecies, indica and japonica, whose genomes have been sequenced (Yu et al. 2002, International Rice Genome Sequencing Project 2005). These subspecies are thought to have originated independently from different subpopulations of the closest wild relative O. rufipogon (Morishima 2001; Zhu and Ge 2005; Londo et al. 2006; Kovach et al. 2007). The inter-subspecific cross often exhibits hybrid sterility due to abnormal growth of pollen or/and the embryo sac. Many cases of hybrid sterility in rice are caused by so-called allelic interaction at a single genetic locus rather than epistatic interactions between unlinked loci (Koide et al. 2008, Ouyang et al. 2009). A hybrid male sterility gene S24, also called f5-du (Wang et al. 2006) or S-b (Li et al. 2006), is a typical gene showing an allelic interaction effect that is located on rice 6
7 chromosome 5. S24 causes pollen semi-sterility when the indica allele is introgressed into the japonica background. The indica allele of (S24-i) acts as a pollen killer and produces maldeveloped male gametes bearing the japonica allele exclusively in the heterozygotes (Kubo et al. 2008; Zhao et al. 2011). Consequently, the S24-i allele is preferentially transmitted to the offspring (90 99%). Zhao et al. (2011) identified an ankyrin protein gene as the primary candidate for S24 by a fine-mapping approach. S24 is under the control of an unlinked dominant suppressor, Epistatic Factor for S24 (EFS), located on chromosome 2 (Kubo et al. 2011). Another hybrid male sterility gene S35, located on chromosome 1, acts as a pollen killer in a similar fashion to S24. Interestingly, S35 has been proposed to interact genetically with S24 to cause pollen sterility, implying a killer killer interaction (Kubo et al. 2008). Together with the animal examples, it is plausible that linked and unlinked epistatic factors play key roles in the genetic mechanism of the gamete killer system. In contrast to the S24 locus that has been somewhat characterized, many characteristics of the S35 locus remain unknown. For example, the causal gene and molecular mechanism of S35 have not been elucidated, although the basic genetic characteristics and the approximate position of the S35 locus on chromosome 1 have been identified (Kubo et al. 2008). Also, it remains unclear whether the S24-suppressor gene EFS can restore pollen sterility due to S35, and whether such a killer killer (S24 S35) interaction really exists as previously proposed. The aim of this study was to characterize the S35 gene and its related genetic network more completely. Using 7
8 reciprocal near-isogenic lines for S24 and S35, our results indicated that the mechanism for hybrid male sterility in rice involves multiple genes and more complex gene interactions than expected. Here, we report a novel gene that interacts with S35 and evidence for the genetic independency of the S24 and S35 genes Materials and Methods Plant materials All experimental populations including mapping populations and near-isogenic lines (NILs) were derived from reciprocal chromosome segment substitution lines (CSSLs) and sister lines that were obtained from a cross between the japonica rice variety Asominori and the indica variety IR24 (Kubo et al. 2002). Both of the parents produced fertile pollen (> 90%) in our growth conditions. An overall view of the breeding procedure used to develop the experimental populations is illustrated in Fig. S1. The NILs with the Asominori genetic background were developed from crossed/backcrossed and selfed progeny of AIS 86, a CSSL carrying IR24 segments of chromosomes 1 and 5 (Kubo et al. 2002), if not otherwise specified. To evaluate pollen fertility of all 27 genotype classes generated by different combinations of the S24, S35 and EFS alleles, a triple heterozygous plant for S24, S35 and EFS in the Asominori genetic background (NIL-S24+S35+EFS) was developed by crossing CSSL AIS 86 and AB2-6 (a derivative line of the AIS library) (Fig. S1A). The NIL S24ILH/I carrying a very small IR24 segment around the S24 locus in an otherwise uniform Asominori genetic background 8
9 was developed by two additional backcrosses of AIS 86 with Asominori (BC 5 F 2 ) and marker-assisted selection (MAS) (Fig. S1B). NILs with the IR24 genetic background were developed from crosses of three different CSSLs, IAS 27/IAS 10//IAS 6 (Fig. S1C). Fine mapping of S35 and an S35-activator INK was performed using self-pollinated progeny of AIS 86/Aso and NIL-S24+S35+EFS/Aso. Genotype frequencies were determined using seedling leaves of self-pollinated progeny of a single plant heterozygous for S35 or S24 with different genetic backgrounds. All plant materials for phenotype evaluation were grown under paddy field conditions in in Mishima, Japan DNA analysis Crude DNA extracts from individual leaves were prepared for genotyping using 0.25 M NaOH followed by neutralization with 0.1 M Tris-HCl. These DNA extracts (1.0 µl) were used in PCR reactions (10-µl final volume) performed using GoTaq polymerase (Promega, USA) with the following cycling profile: 94 C for 2 min; followed by 30 cycles of 94 C for 20s, C for 20 s, and 72 C for 30 s. The PCR products were resolved on 2.0% agarose gels and visualized by ethidium bromide staining. PCR-based markers, insertion and deletion (InDel) and simple sequence repeat (SSR) markers were newly designed for mapping of the S35 and INK loci based on DNA sequence polymorphisms between Nipponbare and (MSU6, BGI, 9
10 Also SSR markers reported by McCouch et al. (2002) were used. The primer sequences for DNA markers are listed in Table S1. The whole-genome genotyping data of the reciprocal CSSLs were obtained from Kubo et al. (2002). For genotyping non-target chromosome segments retained in the genetic background, PCR markers were used that were evenly distributed over the 12 rice chromosomes (Mizuta et al. 2010; Kubo et al. 2011). Genes and marker loci are shown on the physical chromosome map based on the Nipponbare sequence (MSU7) Phenotyping and histological experiments Pre-flowering panicles from each individual in the population were collected and fixed in 50% ethanol solutions to examine pollen fertility. Pollen grains from 3 to 6 anthers at one day before anthesis were stained with 1.0% iodine-potassium iodide (I 2 -KI), and the number of stained/unstained pollen grains were counted using a microscope. More than 400 pollen grains were scored for each individual. In this study, we scored pollen phenotypes with <40%, 40 70%, 70 90% and >90% pollen fertility as sterile, semi-sterile, partial sterile and fertile, respectively. To observe the morphology of mature pollen grains, the ethanol-fixed pollen grains were stained with hematoxylin solution by the method of Kindiger and Beckett (1985). Male gametogenesis was analysed using young panicles at different developmental stages collected from Asominori and the pollen sterile plants. The panicles were fixed and stored in FAA solution (45% ethanol, 5% formalin, 5% acetic acid). After fixation, the samples were 10
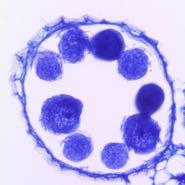
11 embedded in paraffin (Paraplast Plus; McMormick Scientific, St. Louis, MO) and cut at a thickness of 8 µm with a Microm HM355 microtome (Microm, Walldorf, Germany). The sections were stained with 0.05% Toluidine blue O. To evaluate seed fertility, three panicles with fully ripened grains were collected from each plant, and the numbers of filled and unfilled spikelets were counted. The seed setting rate was estimated by the formula: the number of filled grains / total number of filled and unfilled grains Results Characteristics and chromosomal localization of S35 The pollen killer gene S35 has been roughly mapped to the short arm of rice chromosome 1 (Kubo et al. 2008). This previous genetic study indicated that the heterozygous S35 (S35-i/S35-j) caused pollen sterility when the indica segment harboring S24, another pollen killer, was concurrently introgressed into the japonica background (NIL-S35H in Fig. 1A) (Kubo et al. 2008). A single introgression of S35-i in the japonica genome does not cause pollen sterility (NIL-S35al in Fig. 1A). These observations suggested that S35-i interacts with the dominant indica allele of S24 (S24-i) (Kubo et al. 2008). Reciprocal test crossing of NIL-S35H and Asominori showed that S35-i was inherited at a higher frequency (89%) through the male gamete in the S35 heterozygotes (the expected frequency was 50%) (Table 1), indicating the selective abortion of the male gamete bearing S35-j and the incomplete penetrance of the S35 gene. Histological observations showed that the mature anthers of the sterile 11
12 NIL-S35H retained uninucleate and bicellular microspores, suggesting that developmental defects begin from the late uninucleate microspore stage after successful meiosis (Fig. 2A, B). Pollen grains arrested at later developmental stages were also found (Fig. 2C). This finding suggested that evaluating some of the large arrested pollen grains as fertile pollen might have resulted in an overestimation of pollen fertility in the S35 heterozygotes. No apparent defects were found in the tapetum or anther walls of sterile S35-heterozygotes during male gametogenesis (Fig. 2D L). We next performed high-resolution mapping to determine the precise position of the S35 locus. Using a large segregating population (N = 2,800), the location of the S35 locus was narrowed down within a 122-kb region between marker loci 1c300 1c312 (Fig. 1B). The candidate genomic region for S35 encoded 27 putative protein-coding genes and 4 transposable elements (Table S2). Rice transcriptome data (Fujita et al. 2010) indicated that out of the 27 candidate genes, three genes (LOC_Os01g06460/Cys-rich domain protein, LOC_Os01g06580/fasciculin-like arabinogalactan-protein, and LOC_Os01g06590/zinc finger C3HC4 type domain protein) had relatively higher expression or their expression significantly changed around the uninucleate stage (Fig. S2), suggesting that these three genes are the most probable candidates for S35 function Pollen sterility due to S35 is independent of EFS The EFS locus was previously identified as a suppressor of S24; the dominant indica allele Efs-i restores pollen sterility due to S24 (NIL-S24H+Efs in Fig. 1A) (Kubo et al. 12
13 ). A direct or indirect interaction between EFS and S35 was presumed based on the previously identified genetic interactions (S24 S35 and EFS S24) (Kubo et al. 2008, 2011). To examine whether the S35 and EFS genes interact with each other, we phenotyped all 27 allele combinations at the three loci S24, EFS, and S35. The self-pollinated progeny of triple heterozygous plants for S24, EFS and S35 were used to evaluate the 27 genotypes. Consistent with the previous results (Kubo et al. 2008, 2011), S35 caused pollen sterility only in the presence of the S24-i allele (#6 and 18 in Fig. 3), and Efs-i restored the S24-dependent pollen sterility (#13 15 in Fig. 3). However, contrary to our expectations, the heterozygous S35 caused partial pollen sterility regardless of the EFS genotype (#4 6 in Fig. 3). This result indicated that Efs-i is unable to suppress S35 and, therefore, the S24-specific suppressor. Thus, S35 required the IR24 segment harboring S24 to induce pollen sterility but was independent of EFS. It is worth emphasizing that S24-induced sterility was not affected by the S35 genotype (comparison between #9 and 15 in Fig. 3), as previously shown (Kubo et al. 2008) Genetic dissection of the S24 region and an activator of S35 For further characterization of S24 as the pollen killer by itself and as the partner of S35, we genetically dissected the S24 region using a segregating population (n = 485). We obtained a NIL of S24 (called S24ILH/I) carrying a very small segment (181-kb) near the S24 region with an otherwise uniform Asominori genetic background (Fig. 4A, B). The heterozygous segment of S24ILH was able to induce semi-sterility (43.1%) in the 13
14 pollen, indicating that the small segment contained the S24 locus (Fig. 4B). We next reexamined the S24 S35 interaction by analyzing the F 2 and F 3 individuals derived from a cross of S24ILI NIL-S35al. From the self-pollinated progeny of an F 1 plant, a plant homozygous for S24-i and heterozygous for S35 was selected, and the pollen phenotype of its selfed progeny carrying the same parental genotype (5R19) was investigated (Fig. S1B). If interaction between S24 and S35 occurred, the 5R19 would have sterile pollen due to the heterozygous S35-i/S35-j genotype. In contrast, the results indicated that 5R19 plants had good pollen fertility (95.0% fertility) (Fig. 4C D), and the segregation pattern of S35 fit a 1 : 2 : 1 ratio in the selfed progeny (Fig. 5A). These results indicated that S35 does not interact with S24 but with another locus linked to S24. In a similar way, we investigated the effects of other nearby segments on S35-dependent pollen sterility. A distal segment adjacent to the S24 locus had no effect on pollen fertility (5R20 in Figs. 4C D and Fig. 5A). On the other hand, another centromere-side segment resulted in partial sterility for pollen due to S35 (5R21, 77.3% in Fig. 4C D) and selective abortion of the S35-j gamete in the selfed progeny (χ 2 1:2:1 = 65.20, P < 0.001, Fig. 5A). Thus, we were able to confirm that not S24 but the S24-linked gene is essential for S35 activation. This finding was consistent with the aforementioned result that EFS did not suppress S35 and S24 induced pollen semi-sterility regardless of the S35 genotype. The newly identified S35-activator locus was designated as incentive for killing pollen (INK). We next genetically dissected INK using 485 individuals. The INK locus was localized within a 592-kb region between marker loci ms2 and RM13 (Fig. 14
15 D). These results conclusively showed that the dominant Ink-i gene located close to S24 was required to induce S35-dependent pollen sterility Epistasis involving multiple genes controls pollen killers As described above, we genetically dissected the S35 and S24 genes in the japonica genetic background. Through this analysis, we obtained clear evidence for two independent genetic pathways, EFS S24 and INK S35. The question then arises as to whether these genes comprise the full set of genes involved in the pollen killer system. To address this question, we examined another set of NILs with an indica genetic background that have a nearly opposite genomic constitution. We developed two NILs, called NIL-S24H[IR] and NIL-S35H[IR] that carried S24 or S35 heterozygous alleles with the indica genetic background, respectively. Unlike the japonica background populations, pollen semi-sterility due to heterozygous S24 did not occur in the IR24 genetic background (NIL-S24H[IR] in Fig. 5C). Consistent with this normal pollen phenotype, a normal Mendelian segregation of S24 was observed in the IR24 genetic background population (χ 2 1:2:1 = 2.22, P = 0.33) (Fig. 5B). Also, the pollen fertility of the S35-i/S35-j heterozygotes was not significantly reduced in the IR24 genetic background (NIL-S35H[IR] in Fig. 5C). Despite the fertile pollen phenotype, a reduced transmission of the S35-j allele was observed in the selfed progeny of NIL-S35H[IR] (Fig. 5A). The frequency of the S35-j homozygote was slightly recovered in the IR24 genetic background (27/225 = 12.0%, in NIL-S35H[IR]) compared with that in the 15
16 japonica background ( % in #4 and #6). Interestingly, the S35 heterozygote with recessive homozygous ink-j alleles (S35 held inactive) had a higher level of S35-j transmission (The S35-j homozygote emerged at 19.3% (31/161) frequency in the selfed progeny of #18[IR], Fig. 5A). Therefore, we considered that the effect of S35 was slightly diminished, but S35 still remained as a causal agent in the defective fertilization of S35-j pollen in the IR24 genetic background, even though no remarkable visible defects were found in their pollen grains when stained with the I 2 -KI solution. Genotype frequency data of a recombinant inbred (RI) population reported by Huang et al. (2009) supported these results. In the RI population derived from the cross between Nipponbare (japonica) (indica), the genotype frequency distribution peaked at 3.1 Mb on chromosome 1 where S35 is located (Huang et al. 2009). The japonica homozygotes emerged at a reduced frequency (II : JJ = 115 : 33 [22%], the expected Mendelian ratio is 1 : 1 [50%], in Fig. S3A) that was slightly higher than our data based on the japonica background populations (The expected value was estimated as II : JJ = 132 : 16 [11%] when the transmission frequency of the indica S35 allele [k i = 0.89, Table 1] was applied to the RI population). Equal segregation of S24 in the RI population was also in good agreement with our data obtained from NIL-S24[IR]. In total, these results indicated that S35 was active in the indica background population as well as in the F 1 hybrids, and also suggested that some minor modifier(s) existed in the IR24 genome. Furthermore, our results indicated that the action of S24 was repressed by several suppressors encoded by the IR24 genome, including EFS (Fig. 6A). 16
17 Discussion Genetic mechanism of the pollen killer system Although we hypothesized that S35 caused pollen sterility via interaction with the S24 locus (Kubo et al. 2008), the present study revealed that not S24 but a tightly linked locus INK activates S35 to cause pollen sterility. The indica Ink-i allele dominantly activated S35, and conversely S35 had no harmful effect on reproductive or vegetative development in japonica homozygous for ink-j/ink-j. The Ink-i allele did not have a considerable effect on the activity of S24 because S24 caused pollen sterility without Ink-i. The independence of S24 and S35 was consistent with the result that the S24-suppressor Efs-i did not restore pollen sterility caused by S35. Consequently, we concluded that INK S35 and EFS S24 are independent pollen killer systems, rather than a single pathway of EFS S24 S35 (Fig. 6B). In addition to these genes, the present study indicated the presence of additional modifier/suppressor(s) in the genetic background. A few minor modifiers for S35 and a major suppressor for S24 were suggested as the causes of phenotypic changes in the indica background. These findings suggest that the plant gamete killer system is regulated by multiple genetic networks involving linked and unlinked multiple genes with major and minor effects Candidate genes and putative functions of S35 and INK In this study, the S35 locus was delimited within a Mb region of 17
18 chromosome 1 that contained many putative protein-coding genes. Within this chromosomal region ( Mb), a hybrid male sterility locus S-d was found by using other indica/japonica cross combinations (Li et al. 2008). Since S35 and S-d were located within the same narrow interval, these are hypothesized to be identical genes. Allelic differentiation of S24 and S35 seemed to contribute significantly to pollen sterility in the indica-japonica cross because both loci were found in different cross combinations of the indica and japonica varieties (Wang et al. 2006; Li et al. 2006, 2008). It is of great interest to understand the function of these genes in the parental species as well as the harmful effects in the heterozygous hybrids. The S35 locus caused developmental defects after the late uninucleate microspore stage (Fig. 2A), suggesting that the causal gene is expressed and functions around this stage. The uninucleate stage is followed by two mitotic cell divisions in which many different genes are expressed and dynamic changes in expression patterns occur, especially during the bicellular tricellular transition (Fujita et al. 2010). Based on the expression profiles during male gametogenesis in rice, we found three candidate genes for S35 whose expression increased or changed significantly near the uninucleate stage (i.e. Cys-rich domain protein/loc_os01g06460, fasciclin-like arabinogalactan-protein/loc_os01g06580, and zinc finger C3HC4 type domain protein/loc_os01g06590 in Fig. S2). Protein sequence polymorphisms were found in the Cys-rich domain protein and the fasciclin-like arabinogalactan-protein based on the indica variety (data not shown). Therefore, these two genes are strong candidates for S35/S-d. It is noteworthy 18
19 that a rice mutant study has revealed that a fasciclin glycoprotein (Microspore and Tapetum Regulator 1, MTR1) is essential for male gametogenesis in rice (Tan et al. 2012). As is the case with the S-d mapping study (N = 2,160), no recombination events were found within the 122-kb candidate interval of the S35 locus, suggesting a suppression of recombination within this region (a total of about 5,000 individuals were screened for recombination events in the S35/S-d region in these two studies). A large chromosome insertion (~36 kb) and many SNPs were found in the S35/S-d candidate region of the indica variety (data not shown). The INK locus was localized within a 592-kb region where 72 putative protein-encoding genes and 20 transposable elements were found based on the Nipponbare genome sequence. No identical or similar genomic fragments were found between the S35 and INK candidate regions. The INK S35 interaction fits well with an epistasis-based allelic interaction model that hypothesizes that the pollen killer system functions by interaction between sporophytic and gametophytic genes, as proposed previously (Kubo et al. 2011). In the case of INK S35, INK is the sporophytic factor (Fig. 6C). Therefore, the INK gene is predicted to be expressed in sporopytic tissues such as tapetal and anther wall layers. Since tapetal cells supply the major components of the pollen wall and nutrition requisite for pollen development, interaction between tapetal cells and microspore cells is essential for normal pollen development (Tan et al. 2012). Thus, the causal molecule encoded by INK may be involved in such a cooperative interaction between sporophytic and gametophytic gene products. This 19
20 situation resembles the SaF-SaM male sterility system (Long et al. 2008). In that system, two genes, SaF and SaM, interact with each other to cause male sterility in an indica/japonica rice hybrid. The indica SaF + allele acts sporophytically, whereas the japonica SaM allele acts as the gametophytic determinant for the selective pollen killing. A physical protein interaction of SaF + with SaM but not with SaM + was demonstrated (Long et al. 2008). Further cloning analyses will provide valuable information on the molecular interaction of INK S35 and the molecular pathway underlying hybrid male sterility Properties of the S24 and S35 gene blocks In this study, we examined the effect of the S24 INK gene block on chromosome 5 focusing on pollen sterility. Interestingly, a number of studies have detected a major QTL for seed set percentage on the S24 INK region in several indica/japonica populations (Li et al. 1997; Wang et al. 1998; Chin et al. 2011). More recently, Zhao et al. (2007) identified a hybrid female sterility locus S31 at Mb within the S24 INK region by using the same Asominori-IR24 CSSLs. We expect that S31 will facilitate a transmission advantage of the indica S24 INK haplotype through the female gametes. Intriguingly, however, no remarkable phenotype for the S31 gene has been found in our growth conditions (Fukuoka and Mishima in Japan), although S31 caused partial female sterility (about 65%) when the same CSSLs were grown in Nanjing, China (Zhao et al. 2007). This inconsistent gene action may be 20
21 explained by differences in the environments where the experimental populations were grown. If so, this observation means that S31 is strongly influenced by environment and can enhance the transmission of the S24 S31 INK gene complex under certain conditions. In terms of pollen sterility due to S24 and S35, some variations were found within and among the NIL populations (Figs. 1, 3 and 4). Phenotypic variation can result from gene-gene and gene-environment interactions. Linked or unlinked modifier gene(s) located in retained segments in the genetic background is a possible genetic element for phenotypic variation. A straightforward example of this can be found in the difference in pollen fertility among NILs for S24 (Fig. 4B). The pollen fertility of S24ILH with the smallest segment harboring the S24 locus was lower than that of other recombinant lines (5R76 and 5R08), implying the presence of linked minor gene(s) for male-fertility restoration. Also, the S35 locus showed some phenotypic differences (65 85%) between the NIL populations (Fig. 1B and Fig. 3). Environmental effects for pollen sterility due to S35 were seen under different growth conditions (54.3 ± 7.3% in Fukuoka in Kubo et al. (2008); 84.2 ± 4.1% in Mishima in this study). This environmental susceptibility may be a common feature of genes conferring male sterility in plants (Sakata et al. 2010; Ding et al. 2012; Wang et al. 2013; Kubo et al. 2016) The impact of pollen killer genes on the genetic characteristics of hybrid offspring The actions of the S35 and S24 killers were exclusively associated with their 21
22 heterozygotes. Homozygotes for either alleles did not show any notable phenotype in reproductive development or vegetative growth, excluding phenotypic changes by other linked genes. Both pollen sterile S24-heterozygotes and S35-heterozygotes exhibited nearly full seed set (93.3% ± 3.2, n = 10, in S24ILH; 88.4% ± 0.7, n = 3, in NIL-S35H), suggesting that these pollen killers have a transmission advantage for increasing their frequencies with no or a small effect on individual fecundity at least in the japonica genetic background. But as it is, such a strong transmission advantage cannot directly apply to early segregating generations of the indica-japonica hybrid progeny. In view of the results obtained in the RI and the indica background populations, S35-i may weaken its function of eliminating S35-j (japonica allele) but may still keep the killing action to some degree. In contrast, S24 contributed little to the reduction of pollen fertility in the F 1 generation. The action of S24 is substantially suppressed by Efs-i and other suppressor(s). Using indica-japonica hybrid populations, Hiko-Ichi Oka demonstrated that recombination of two independent genes tended to be restricted as if they were linked (referred to as pseudo-linkage or quasi-linkage) (Oka 1988). The same tendency was also observed in O. sativa x Oryza glaberrima (Sano et al. 1980). An example is the inheritance of Phenol reaction (Ph) and Chromogen for anthocyanin (C) located on separate chromosomes. The F 2 population of indica (Ph c) japonica (ph C) had excessive numbers of the parental genotypes (Ph c and ph C) and reduced numbers of recombined genotypes (Ph C and ph c) (Oka 1988). In this study, Oka pointed out that 22
23 a gene set for duplicate gametic lethals is one possible cause for the pseudo-linkage of the phenotypic markers. Theoretically, the INK S35 interaction could also partially contribute to pseudo-linkage. The pollen killer S35-i (indica allele) increases its own transmission with the existence of the indica Ink-i allele through the male gamete, resulting in a high frequency of the indica indica pair for the S35 and INK alleles (S35-i Ink-i) and a reduction of the recombinant pair of the japonica indica alleles (S35-j Ink-i). By this means through male gametes, the parental set of different chromosomes (chromosomal segments) will gradually increase their frequency in the population. Some of this tendency was observed in the two RI populations of Nipponbare (Huang et al. 2009), and Asominori IR24 (Tsunematsu et al. 1996) (Fig. S3B). Thus, several types of hybrid sterility genes may contribute to pseudo-linkage of many chromosomal regions because more than 40 hybrid sterility genes are dispersed throughout the rice genome Evolutionary implications of pollen killers In the traditional triallelic model, both the gamete killer and the sensitive alleles can arise from the original neutral allele without a reduction in fitness and consequently become fixed in diverging populations. Recent studies of the Sa and S5 loci have afforded tangible evidence of the molecular and the evolutionary mechanisms (Long et al. 2008; Yang et al. 2012). These loci contained multiple linked genes. Presumably, sequential mutations within these genes occur in either or both lineages, and these 23
24 divergent haplotypes negatively interact, resulting in gamete dysfunction in the heterozygotes (Ouyang and Zhang 2013). This sequential divergence theory could be applicable to the pollen killer systems by S35 and S24, despite the differences in physical positions of the interacting partner genes. Pollen killing can occur only if the pollen killers are paired with particular alleles at other interacting loci in the hybrid offspring. Such a polygenic system allows the accumulation of mutations at related loci without a significant fitness cost during evolution. In fact, an increasing number of studies have suggested that a gradual accumulation of multiple epistatic genes with no or a small individual effect may play important roles in the development of reproductive isolation in initial speciation (Cabot et al. 1994; Orr 1995; Kao et al. 2010; Kubo et al. 2016). In addition, the existence of third neutral alleles for S24 and S35 have been reported in studies using numerous cultivars and the wild progenitor Oryza rufipogon (Wang et al. 2006; Shi et al. 2009; Liu et al. 2011). This observation implies that these pollen killer loci may contain tightly-linked multiple genes like S5 and Sa, or may be pure trialleles of a single gene. Our study demonstrated the impact of the gene complex S24 S31 INK on hybrid sterility between indica and japonica. Plant species have evolved mainly through repeated duplication of an ancestral genome, chromosome and/or genes. Redundancy of gene sets in plant genomes is important as a prerequisite for diversification of genes adapting to environmental changes. Furthermore, most plants have the ability to reproduce asexually and via autogamy. Therefore, co-adapted gene complexes can be maintained over many generations without breakdown by 24
25 reproductive meiosis and meiotic recombination. These genomic and reproductive characteristics of plants should facilitate the formation of polymorphic gene complexes for reproductive isolation (Dempewolf et al. 2012). Plant genetic studies have traditionally focused on gamete killer loci. Even if interacting partner genes exist in a genetic background, these partners would have largely gone unnoticed. Though much still remains to be elucidated, it is clear that many genetic factors and a genetic network play a pivotal role in the plant gamete killer system. Tracking down the genes one by one can, therefore, shed further light on the molecular and evolutionary mechanisms of gamete killer systems. Knowledge of the chromosomal locations and the nature of gamete killer loci will aid the design of efficient marker-assisted schemes for crop breeding Acknowledgements We thank T. Makino, Y. Gonohe, and H. Kondo (National Institute of Genetics, Mishima, Japan) for technical assistance. This study was partly supported by JSPS KAKENHI Grant Number (T.K.) Literature Cited Buckler E. S., Phelps-Durr T. L., Buckler C. S., Dawe R. K., Doebley J. F., et al., 1999 Meiotic drive of chromosomal knobs reshaped the maize genome. Genetics 153: Burt A., and Trivers R., 2009 Genes in conflict: the biology of selfish genetic elements. Harvard University Press. 25
26 Cabot E. L., Davis A. W., Johnson N. A., and Wu C. I., 1994 Genetics of reproductive isolation in the Drosophila simulans clade: complex epistasis underlying hybrid male sterility. Genetics 137: Cameron D. R., and Moav R. M., 1957 Inheritance in Nicotiana tabacum XXVII. Pollen killer, an alien genetic locus inducing abortion of microspores not carrying it. Genetics 42: Chin J. H., Chu S.-H., Jiang W., Cho Y.-I., Basyirin R., et al., 2011 Identification of QTLs for hybrid fertility in inter-subspecific crosses of rice (Oryza sativa L.). Genes Genomics 33: Dempewolf H., Hodgins K. a., Rummell S. E., Ellstrand N. C., and Rieseberg L. H., 2012 Reproductive isolation during domestication. Plant Cell 24: Ding J., Lu Q., Ouyang Y., Mao H., Zhang P., et al., 2012 A long noncoding RNA regulates photoperiod-sensitive male sterility, an essential component of hybrid rice. Proc. Natl. Acad. Sci. 109: Endo T. R., 2015 Gametocidal Genes. In: Molnár-Láng M, Ceoloni C, Doležel J (Eds.), Alien Introgression in Wheat: Cytogenetics, Molecular Biology, and Genomics, Springer, pp Finseth F. R., Dong Y., Saunders A., and Fishman L., 2015 Duplication and adaptive evolution of a key centromeric protein in Mimulus, a genus with female meiotic drive. Mol. Biol. Evol. 32: Fishman L., and Saunders A., 2008 Centromere-associated female meiotic drive entails male fitness costs in monkeyflowers. Science (80-. ). 322: Fujita M., Horiuchi Y., Ueda Y., Mizuta Y., Kubo T., et al., 2010 Rice expression atlas in reproductive development. Plant Cell Physiol. 51: Huang X., Feng Q., Qian Q., Zhao Q., Wang L., et al., 2009 High-throughput genotyping by whole-genome resequencing. Genome Res 19: Ikehashi H., and Araki H., 1986 Genetics of F 1 sterility in remote crosses of rice. In: Khush GS (Ed.), Rice Genetics, International Rice Research Institute, Los Baños, Philippines, pp
27 International Rice GenomeSequencing Project, 2005 The map-based sequence of the rice genome. Nature 436: Kanizay L. B., Albert P. S., Birchler J. a, and Dawe R. K., 2013 Intragenomic conflict between the two major knob repeats of maize. Genetics 194: Kao K. C., Schwartz K., and Sherlock G., 2010 A genome-wide analysis reveals no nuclear Dobzhansky-Muller pairs of determinants of speciation between S. cerevisiae and S. paradoxus, but suggests more complex incompatibilities. PLoS Genet. 6: Kindiger B., and Beckett J. B., 1985 A hematoxylin staining procedure for maize pollen grain chromosomes. Biotech. Histochem. 60: Koide Y., Onishi K., Kanazawa A., and Sano Y., 2008 Genetics of Speciation in Rice. In: Hirano H, Sano Y, Hirai A, Sasaki T (Eds.), Rice Biology in the Genomics Era. Biotechnology in Agriculture and Forestry, Springer-Verlag, Berlin Heidelberg, pp Kovach M. J., Sweeney M. T., and McCouch S. R., 2007 New insights into the history of rice domestication. Trends Genet. 23: Kubo T., Aida Y., Nakamura K., Tsunematsu H., Doi K., et al., 2002 Reciprocal chromosome segment substitution series derived from Japonica and Indica cross of rice (Oryza sativa L.). Breed. Sci. 52: Kubo T., Yamagata Y., Eguchi M., and Yoshimura A., 2008 A novel epistatic interaction at two loci causing hybrid male sterility in an inter-subspecific cross of rice (Oryza sativa L.). Genes Genet. Syst. 83: Kubo T., Yoshimura A., and Kurata N., 2011 Hybrid male sterility in rice is due to epistatic interactions with a pollen killer locus. Genetics 189: Kubo T., Takashi T., Ashikari M., Yoshimura A., and Kurata N., 2016 Two tightly linked genes at the hsa1 locus cause both F 1 and F 2 hybrid sterility in rice. Mol. Plant 9: Larracuente A. M., and Presgraves D. C., 2012 The selfish segregation distorter gene complex of Drosophila melanogaster. Genetics 192:
28 Li Z., Pinson S. R. M., Paterson A. H., Park W. D., and Stancel J. W., 1997 Genetics of hybrid sterility and hybrid breakdown in an intersubspecific rice (Oryza sativa L.) populations. Genetics 145: Li W., Zeng R., Zhang Z., Ding X., and Zhang G., 2006 Fine mapping of locus S-b for F 1 pollen sterility in rice (Oryza sativa L.). Chinese Sci. Bull. 51: Li W., Zeng R., Zhang Z., Ding X., and Zhang G., 2008 Identification and fine mapping of S-d, a new locus conferring the partial pollen sterility of intersubspecific F 1 hybrids in rice (Oryza sativa L.). Theor. Appl. Genet. 116: Liu B., Li J. Q., Liu X. D., Shahid M. Q., Shi L. G., et al., 2011 Identification of neutral genes at pollen sterility loci Sd and Se of cultivated rice (Oryza sativa) with wild rice (O. rufipogon) origin. Genet. Mol. Res. 10: Londo J. P., Chiang Y.-C., Hung K.-H., Chiang T.-Y., and Schaal B. A., 2006 Phylogeography of Asian wild rice, Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. Proc. Natl. Acad. Sci. U. S. A. 103: Long Y., Zhao L., Niu B., Su J., Wu H., et al., 2008 Hybrid male sterility in rice controlled by interaction between divergent alleles of two adjacent genes. Proc Natl Acad Sci USA 105: Lyon M. F., 2003 Transmission ratio distortion in mice. Annu. Rev. Genet. 37: McCouch S. R., Teytelman L., Xu Y., Lobos K. B., Clare K., et al., 2002 Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res. 9: Mizuta Y., Harushima Y., and Kurata N., 2010 Rice pollen hybrid incompatibility caused by reciprocal gene loss of duplicated genes. Proc Natl Acad Sci USA 107: Morishima H., 2001 Evolution and domestication of rice. In: Khushu GS, Brar DS, Hardy B (Eds.), Rice Genetics IV, International Rice Research Institute, Los Baños, Philippines, pp
29 Oka H., 1988 Origin of cultivated rice. Japan Scientific Societies Press, Tokyo, Japan. Orr H. A., 1995 The population genetics of speciation: The evolution of hybrid incompatibilities. Genetics 139: Ouyang Y., Chen J., Ding J., and Zhang Q., 2009 Advances in the understanding of inter-subspecific hybrid sterility and wide-compatibility in rice. Chinese Sci. Bull. 54: Ouyang Y., and Zhang Q., 2013 Understanding reproductive isolation based on the rice model. Annu. Rev. Plant Biol. 64: Rick C. M., 1966 Abortion of male and female gametes in the tomato determined by allelic interaction. Genetics 53: Sakata T., Oshino T., Miura S., Tomabechi M., Tsunaga Y., et al., 2010 Auxins reverse plant male sterility caused by high temperatures. Proc. Natl. Acad. Sci. U. S. A. 107: Sano Y., Chu Y., and Oka H., 1980 Genetic studies of speciation in cultivated rice. 2. Character variations in backcross derivatives between Oryza sativa and O. graberrima: MV linkage and key. Japanese J. Genet. 55: Shi L., Liu X., LIiu B., Zhao X., Wang L., et al., 2009 Identifying neutral allele Sb at pollen-sterility loci in cultivated rice with Oryza rufipogon origin. Chinese Sci. Bull. 54: Tan H., Liang W., Hu J., and Zhang D., 2012 MTR1 encodes a secretory fasciclin glycoprotein required for male reproductive development in rice. Dev. Cell 22: Tsunematsu H., Yoshimura A., Harushima Y., Nagamura Y., Kurata N., et al., 1996 RFLP framework map using recombinant inbred lines in rice. Japanese J. Breed. 46: Wang J., Liu K. D., Xu C. G., Li X. H., and Zhang Q., 1998 The high level of wide-compatibility of variety Dular has a complex genetic basis. Theor. Appl. Genet. 97:
30 Wang G. W., He Y. Q., Xu C. G., and Zhang Q., 2006 Fine mapping of f5-du, a gene conferring wide-compatibility for pollen fertility in inter-subspecific hybrids of rice (Oryza sativa L.). Theor. Appl. Genet. 112: Wang H., Lu Y., Jiang T., Berg H., Li C., et al., 2013 The Arabidopsis U-box/ARM repeat E3 ligase AtPUB4 influences growth and degeneration of tapetal cells, and its mutation leads to conditional male sterility. Plant J. 74: Weichenhan D., Kunze B., Winking H., Geel M. van, Osoegawa K., et al., 2001 Source and component genes of a Mb gene cluster in the house mouse. Mamm. Genome 12: Yang J., Zhao X., Cheng K., Du H., Ouyang Y., et al., 2012 A killer-protector system regulates both hybrid sterility and segregation distortion in rice. Science (80-. ). 337: Yu J., Hu S., Wang J., Wong G. K., Li S., et al., 2002 A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 296: Zhao Z. G., Jiang L., Zhang W. W., Yu C. Y., Zhu S. S., et al., 2007 Fine mapping of S31, a gene responsible for hybrid embryo-sac abortion in rice (Oryza sativa L.). Planta 226: Zhao Z. G., Zhu S. S., Zhang Y. H., Bian X. F., Wang Y., et al., 2011 Molecular analysis of an additional case of hybrid sterility in rice (Oryza sativa L.). Planta 233: Zhu Q., and Ge S., 2005 Phylogenetic relationships among A-genome species of the genus Oryza revealed by intron sequences of four nuclear genes. New Phytol. 167: Figure caption Figure 1. Genetic basis of hybrid male sterility genes and localization of the S35 locus. (A) Pollen phenotypes of NILs for S35, S24, and EFS genes associated with hybrid 30
31 male sterility. Graphical genotypes (left) and pollen grains (right) of the NILs stained by I 2 -KI are shown. Black and white bars denote IR24 (indica) and Asominori (japonica) chromosomes, respectively. The NIL carrying S35 alone (NIL-S35al) were fertile, but co-introgression of the S24 segment from IR24 dominantly activated S35 leading to pollen sterility in the Asominori genetic background (NIL-S35H). The S24-suppressor Efs gene from IR24 dominantly inactivated S24 (NIL-S24H+Efs). (B) Chromosome position of S35. Upper panel: Physical chromosome positions of the S35 locus and DNA markers (1cXXX). Lower panel: Diagram showing the recombination breakpoints and pollen phenotypes of the lines. All recombinant plants carried IR24 homozygous alleles for S24 to activate S35-i. The pollen fertility is shown as the mean (%) ± SD (n = 3 7) Figure 2. Histological characterization of the sterile S35 heterozygotes. (A C) Mature pollen grains stained with hematoxylin. (A) Pollen grains from Asominori. (B and C) Pollen grains from the sterile S35 heterozygotes (NIL-S35H). White and black arrowheads indicate normal sperm cells and nuclei of abortive pollen grains, respectively. Pollen grains arrested at the bicellular stage (black arrowheads) and vacuolated pollen at the uninucleate stage are shown in panel B. Panel C shows that these two pollen grains from NIL-S35H were well filled and similar in size, but the pollen grain on the right has a single bold nucleus instead of two sperm cells as would be observed in normal pollen grains. (D M) Transverse sections showing anther 31
32 development of Asominori and the S35 heterozygote (NIL-S35H). Five stages of anther development in Asominori and NIL-S35H were compared. Transverse sections were stained with 0.05% Toluidine blue O. (D) to (H) are Asominori, and (I) to (M) are NIL-S35H. (D) and (I) The pollen mother cell stage, (E) and (J) the tetrad stage, (F) and (K) the young microspore stage, (G) and (L) vacuolated pollen stage, (H) and (M) the mature pollen stage. The solid black arrows indicate degenerated pollen grains. PMC, pollen mother cell; T, tapetal layer; Td, tetrad cell; Msp, microspore; Mp, mature pollen. Scale bars = 50 µm for A M Figure 3. Pollen fertility of 27 genotypes determined by three genes S24, S35, and EFS. Pollen fertility (right bar chart) for each genotype class (left panel) is expressed by the mean (%) ± SD (n = 3 10). Effective genes for pollen sterility are marked with red frames on the genotype panel. White and black bars in the bar chart represent the sterile and fertile pollen phenotypes, respectively. Micrographs of the pollen grains for each fertility class are shown in the bottom panel. Plants of each genotype were selected from self-pollinated progeny of NIL-S24+S35+Efs by genotyping using the linked markers ms2 for S24, 1c305 for S35, and 2c2015 for EFS. I: indica (IR24) homozygote, J: japonica (Asominori) homozygote, H: heterozygote, F: fertile, PS: partial-sterile, SS: semi-sterile, S: sterile Figure 4. Identification and localization of the S35-activator. (A) Graphical genotype of 32
33 the near-isogenic line S24ILI. S24ILI was homozygous for S24 and carried a very small IR24 segment of less than 181-kb (ms1 ms3, see also Fig. 4B) within a uniform Asominori background. (B) Pollen phenotype of the most informative recombinants and the NILs S24ILI/H. These lines carried Asominori homozygous alleles for both the EFS and S35 loci, representing the pollen sterility dependent exclusively on S24. Bars show the mean (%) with SD (n = 4 9). (C) Pollen fertility of a set of substitution lines covering the S24 region on chromosome 5. All the lines were heterozygous for S35. The white arrows in the photo indicate degenerated pollen grains. (D) Chromosome position of the S35-activator INK locus on chromosome 5. The diagram shows the recombination breakpoints of the obtained plants/lines. All plants/lines carried heterozygous alleles for S35 to evaluate the effect of the recombinant segment on the S35 phenotype. The pollen phenotype of each line was determined using selfed progeny (BC 4 F 3 ) of the recombinant individuals (BC 4 F 2 ) excluding 5R18. 5R18 was a single BC 5 F 1 plant derived from a backcross of the recombinant BC 4 F 2 plant carrying homozygous S35-i alleles. The 5R02 line had the Efs-i homozygous genotype that prevented the influence of heterozygous S24 on pollen fertility. The mean (%) ± SD (n = 3 10) are shown on the right Figure 5. Effect of the genetic background on pollen sterility. (A, B) Frequencies of the S35 and S24 genotypes in the selfed progeny of the NILs with reciprocal genetic backgrounds. The stacked bar chart represents the frequency of the S35 genotype (A) 33
34 and the S24 genotype (B) in the selfed progeny of the heterozygotes. Population size and chi-square values for the deviation from 1 : 2 : 1 are shown on the top. The numbers of plants in each genotype (black: Asominori homozygote, grey: heterozygote, white: IR24 homozygote) are shown inside the bars. The parental genotype is shown at the bottom. DNA markers 1c350 or 1c305 (for S35) and ms2 (for S24) were used for genotyping (the red line on chromosomes). The plant line ID is identical to that shown in Figs. 3, 4 and 6. Parental plants #4, #6, #15, and 5R21 had sterile pollen. All other plants had fertile pollen. ns: not significant, **: P < 0.01, ***: P < (C) Graphical genotype (left) and pollen fertility (right) of the NILs that carried heterozygous segments harboring the S24 or S35 locus in either the Asominori or the IR24 genetic backgrounds. The NILs carried the appropriate genotype for the interacting partner gene (efs-j/efs-i for the S24 activation or Ink-i/Ink-i for the S35 activation). Bars show the mean (%) with SD (n = 5) Figure 6. Diagram showing the genetic interactions between hybrid male sterility loci. (A) Schematic representation of the chromosome position and genetic interactions of hybrid male sterility genes. Arrows and bars represent positive and negative regulation, respectively. Indica has the killer alleles for the S24 and S35 loci (S24-i and S35-i), and these genes kill the male gamete bearing the japonica alleles in heterozygous hybrid progeny. Activation of these pollen killer genes is dependent on the sporophytic factors EFS for S24 (B) and INK for S35 (C). Additional unknown suppressors/modifiers for 34
35 712 S24 and S35 exist within the indica genome
36 Table 1 Transmission frequency of S35-i-bearing gametes in reciprocal test crosses between NIL-S35H and Asominori Cross combination No. of plants a Total japonica Heterozygote x 2 (1:1) Frequency of the indica allele (k i ) b Asominori NIL-S35H *** 0.89 NIL-S35H Asominori NS 0.61 NS not significant; *** P < a DNA marker 1c350 linked to S35 was used for genotyping. b The expected frequency is




37 Fertile Sterile Figure 1 A Sterile S35 S24? NIL-S35H B S NIL-S24H S EFS S Fertile NIL-S35al NIL-S24H+Efs B Line # 1R10 1R05 1R31 1R08 1R35 1R c275 Asominori IR24 1c Chr. 1 Chr. 5 S35 1c300 1c305 1c310 1c312 Heterozygous 1c c318 1c c340 (Mb) Pollen fertility (%) 97.6± ± ± ± ± ±10.3












38 Figure 2 A B C D E F G H PMC Td Msp Msp Mp T T I J K L M PMC Td Msp Msp T T Mp




39 Figure 3 # EFS S24 S35 I I I H I I J I I I I H H I H J I H I H I H H I J H I I H H H H H J H H I H J H H J J H J I J H H J H J J H I J I H J I J J I I J J H J J J J J I I J H I J J I J F F F PS PS PS F F SS PS PS S F F SS F F F F F F F F F F F F Pollen fertility (%) Sterile Semi-sterile Partial sterile Fertile

 ms1 5c132 5c137 ms2 ms3 5R19 S24 5R20 INK 5R21 Asominori IR24 Heterozygous 2298 (2.")

40 Sterile Fertile Figure 4 A S35 S EFS S24ILI Asominori IR24 B Pollen fertility (%) 100 Line # 50 0 S24ILI S24ILH 5R76 5R ms1 5c132 5c135 5c137 ms ms3 (Mb) S24 76 kb Asominori Heterozygous IR24 C Chr.1 5 S35 RM6320 (0.44 Mb) ms1 5c132 5c137 ms2 ms3 5R19 S24 5R20 INK 5R21 Asominori IR24 Heterozygous 2298 (2.30Mb) Fertile Fertile Partially sterile D Chr. 1 Chr. 5 S24 INK ms1 5c132 5c137 ms2 ms3 5c178 RM13 5c212 5c229 5c256 RM267 Line # 5R02 5R21 5R18* 5R11 5R19 5R (Mb) Pollen fertility (%) 71.9± ± ± ± ±3.3 Asominori IR24 Heterozygous
Pollen Killer Gene S35 Function Requires Interaction with an Activator That Maps Close to S24, Another Pollen Killer Gene in Rice
 INVESTIGATION Pollen Killer Gene S35 Function Requires Interaction with an Activator That Maps Close to S24, Another Pollen Killer Gene in Rice Takahiko Kubo,*,,1 Atsushi Yoshimura, and Nori Kurata*, *Plant
INVESTIGATION Pollen Killer Gene S35 Function Requires Interaction with an Activator That Maps Close to S24, Another Pollen Killer Gene in Rice Takahiko Kubo,*,,1 Atsushi Yoshimura, and Nori Kurata*, *Plant
Segregation distortion in F 2 and doubled haploid populations of temperate japonica rice
 c Indian Academy of Sciences RESEARCH NOTE Segregation distortion in F 2 and doubled haploid populations of temperate japonica rice MASUMI YAMAGISHI 1,2,6, YOSHINOBU TAKEUCHI 3,7, ISAO TANAKA 4, IZUMI
c Indian Academy of Sciences RESEARCH NOTE Segregation distortion in F 2 and doubled haploid populations of temperate japonica rice MASUMI YAMAGISHI 1,2,6, YOSHINOBU TAKEUCHI 3,7, ISAO TANAKA 4, IZUMI
Exam 1 PBG430/
 1 Exam 1 PBG430/530 2014 1. You read that the genome size of maize is 2,300 Mb and that in this species 2n = 20. This means that there are 2,300 Mb of DNA in a cell that is a. n (e.g. gamete) b. 2n (e.g.
1 Exam 1 PBG430/530 2014 1. You read that the genome size of maize is 2,300 Mb and that in this species 2n = 20. This means that there are 2,300 Mb of DNA in a cell that is a. n (e.g. gamete) b. 2n (e.g.
Genetic diversity and population structure in rice. S. Kresovich 1,2 and T. Tai 3,5. Plant Breeding Dept, Cornell University, Ithaca, NY
 Genetic diversity and population structure in rice S. McCouch 1, A. Garris 1,2, J. Edwards 1, H. Lu 1,3 M Redus 4, J. Coburn 1, N. Rutger 4, S. Kresovich 1,2 and T. Tai 3,5 1 Plant Breeding Dept, Cornell
Genetic diversity and population structure in rice S. McCouch 1, A. Garris 1,2, J. Edwards 1, H. Lu 1,3 M Redus 4, J. Coburn 1, N. Rutger 4, S. Kresovich 1,2 and T. Tai 3,5 1 Plant Breeding Dept, Cornell
UNIT 8 BIOLOGY: Meiosis and Heredity Page 148
 UNIT 8 BIOLOGY: Meiosis and Heredity Page 148 CP: CHAPTER 6, Sections 1-6; CHAPTER 7, Sections 1-4; HN: CHAPTER 11, Section 1-5 Standard B-4: The student will demonstrate an understanding of the molecular
UNIT 8 BIOLOGY: Meiosis and Heredity Page 148 CP: CHAPTER 6, Sections 1-6; CHAPTER 7, Sections 1-4; HN: CHAPTER 11, Section 1-5 Standard B-4: The student will demonstrate an understanding of the molecular
Sporic life cycles involve 2 types of multicellular bodies:
 Chapter 3- Human Manipulation of Plants Sporic life cycles involve 2 types of multicellular bodies: -a diploid, spore-producing sporophyte -a haploid, gamete-producing gametophyte Sexual Reproduction in
Chapter 3- Human Manipulation of Plants Sporic life cycles involve 2 types of multicellular bodies: -a diploid, spore-producing sporophyte -a haploid, gamete-producing gametophyte Sexual Reproduction in
2. Der Dissertation zugrunde liegende Publikationen und Manuskripte. 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera)
") 2. Der Dissertation zugrunde liegende Publikationen und Manuskripte 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera) M. Hasselmann 1, M. K. Fondrk², R. E. Page Jr.² und
2. Der Dissertation zugrunde liegende Publikationen und Manuskripte 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera) M. Hasselmann 1, M. K. Fondrk², R. E. Page Jr.² und
CHAPTER 23 THE EVOLUTIONS OF POPULATIONS. Section C: Genetic Variation, the Substrate for Natural Selection
 CHAPTER 23 THE EVOLUTIONS OF POPULATIONS Section C: Genetic Variation, the Substrate for Natural Selection 1. Genetic variation occurs within and between populations 2. Mutation and sexual recombination
CHAPTER 23 THE EVOLUTIONS OF POPULATIONS Section C: Genetic Variation, the Substrate for Natural Selection 1. Genetic variation occurs within and between populations 2. Mutation and sexual recombination
Classical Selection, Balancing Selection, and Neutral Mutations
 Classical Selection, Balancing Selection, and Neutral Mutations Classical Selection Perspective of the Fate of Mutations All mutations are EITHER beneficial or deleterious o Beneficial mutations are selected
Classical Selection, Balancing Selection, and Neutral Mutations Classical Selection Perspective of the Fate of Mutations All mutations are EITHER beneficial or deleterious o Beneficial mutations are selected
QTL analysis for hybrid sterility and plant height in interspecific populations derived from a wild rice relative, Oryza longistaminata
 Breeding Science 59: 441 445 (2009) Note QTL analysis for hybrid sterility and plant height in interspecific populations derived from a wild rice relative, Oryza longistaminata Zhiwei Chen 1,2), Fengyi
Breeding Science 59: 441 445 (2009) Note QTL analysis for hybrid sterility and plant height in interspecific populations derived from a wild rice relative, Oryza longistaminata Zhiwei Chen 1,2), Fengyi
Solutions to Problem Set 4
 Question 1 Solutions to 7.014 Problem Set 4 Because you have not read much scientific literature, you decide to study the genetics of garden peas. You have two pure breeding pea strains. One that is tall
Question 1 Solutions to 7.014 Problem Set 4 Because you have not read much scientific literature, you decide to study the genetics of garden peas. You have two pure breeding pea strains. One that is tall
Chapter 13 Meiosis and Sexual Reproduction
 Biology 110 Sec. 11 J. Greg Doheny Chapter 13 Meiosis and Sexual Reproduction Quiz Questions: 1. What word do you use to describe a chromosome or gene allele that we inherit from our Mother? From our Father?
Biology 110 Sec. 11 J. Greg Doheny Chapter 13 Meiosis and Sexual Reproduction Quiz Questions: 1. What word do you use to describe a chromosome or gene allele that we inherit from our Mother? From our Father?
Life Cycles, Meiosis and Genetic Variability24/02/2015 2:26 PM
 Life Cycles, Meiosis and Genetic Variability iclicker: 1. A chromosome just before mitosis contains two double stranded DNA molecules. 2. This replicated chromosome contains DNA from only one of your parents
Life Cycles, Meiosis and Genetic Variability iclicker: 1. A chromosome just before mitosis contains two double stranded DNA molecules. 2. This replicated chromosome contains DNA from only one of your parents
Name Class Date. KEY CONCEPT Gametes have half the number of chromosomes that body cells have.
 Section 1: Chromosomes and Meiosis KEY CONCEPT Gametes have half the number of chromosomes that body cells have. VOCABULARY somatic cell autosome fertilization gamete sex chromosome diploid homologous
Section 1: Chromosomes and Meiosis KEY CONCEPT Gametes have half the number of chromosomes that body cells have. VOCABULARY somatic cell autosome fertilization gamete sex chromosome diploid homologous
A species of plants. In maize 20 genes for male sterility are listed by
 THE INTERRELATION OF PLASMAGENES AND CHROMO- GENES IN POLLEN PRODUCTION IN MAIZE DONALD F. JONES Connecticut Agricultural Experiment Station, New Haven, Connecticut Received January 24, 195 IV INHERITED
THE INTERRELATION OF PLASMAGENES AND CHROMO- GENES IN POLLEN PRODUCTION IN MAIZE DONALD F. JONES Connecticut Agricultural Experiment Station, New Haven, Connecticut Received January 24, 195 IV INHERITED
Supplementary Figure 1. Phenotype of the HI strain.
 Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Chapter 2: Extensions to Mendel: Complexities in Relating Genotype to Phenotype.
 Chapter 2: Extensions to Mendel: Complexities in Relating Genotype to Phenotype. please read pages 38-47; 49-55;57-63. Slide 1 of Chapter 2 1 Extension sot Mendelian Behavior of Genes Single gene inheritance
Chapter 2: Extensions to Mendel: Complexities in Relating Genotype to Phenotype. please read pages 38-47; 49-55;57-63. Slide 1 of Chapter 2 1 Extension sot Mendelian Behavior of Genes Single gene inheritance
Reinforcement Unit 3 Resource Book. Meiosis and Mendel KEY CONCEPT Gametes have half the number of chromosomes that body cells have.
 6.1 CHROMOSOMES AND MEIOSIS KEY CONCEPT Gametes have half the number of chromosomes that body cells have. Your body is made of two basic cell types. One basic type are somatic cells, also called body cells,
6.1 CHROMOSOMES AND MEIOSIS KEY CONCEPT Gametes have half the number of chromosomes that body cells have. Your body is made of two basic cell types. One basic type are somatic cells, also called body cells,
Meiosis and Mendel. Chapter 6
 Meiosis and Mendel Chapter 6 6.1 CHROMOSOMES AND MEIOSIS Key Concept Gametes have half the number of chromosomes that body cells have. Body Cells vs. Gametes You have body cells and gametes body cells
Meiosis and Mendel Chapter 6 6.1 CHROMOSOMES AND MEIOSIS Key Concept Gametes have half the number of chromosomes that body cells have. Body Cells vs. Gametes You have body cells and gametes body cells
Family resemblance can be striking!
 Family resemblance can be striking! 1 Chapter 14. Mendel & Genetics 2 Gregor Mendel! Modern genetics began in mid-1800s in an abbey garden, where a monk named Gregor Mendel documented inheritance in peas
Family resemblance can be striking! 1 Chapter 14. Mendel & Genetics 2 Gregor Mendel! Modern genetics began in mid-1800s in an abbey garden, where a monk named Gregor Mendel documented inheritance in peas
Mutation, Selection, Gene Flow, Genetic Drift, and Nonrandom Mating Results in Evolution
 Mutation, Selection, Gene Flow, Genetic Drift, and Nonrandom Mating Results in Evolution 15.2 Intro In biology, evolution refers specifically to changes in the genetic makeup of populations over time.
Mutation, Selection, Gene Flow, Genetic Drift, and Nonrandom Mating Results in Evolution 15.2 Intro In biology, evolution refers specifically to changes in the genetic makeup of populations over time.
Ladies and Gentlemen.. The King of Rock and Roll
 Ladies and Gentlemen.. The King of Rock and Roll Learning Objectives: The student is able to construct an explanation, using visual representations or narratives, as to how DNA in chromosomes is transmitted
Ladies and Gentlemen.. The King of Rock and Roll Learning Objectives: The student is able to construct an explanation, using visual representations or narratives, as to how DNA in chromosomes is transmitted
Ch. 13 Meiosis & Sexual Life Cycles
 Introduction Ch. 13 Meiosis & Sexual Life Cycles 2004-05 Living organisms are distinguished by their ability to reproduce their own kind. -Offspring resemble their parents more than they do less closely
Introduction Ch. 13 Meiosis & Sexual Life Cycles 2004-05 Living organisms are distinguished by their ability to reproduce their own kind. -Offspring resemble their parents more than they do less closely
Science Unit Learning Summary
 Learning Summary Inheritance, variation and evolution Content Sexual and asexual reproduction. Meiosis leads to non-identical cells being formed while mitosis leads to identical cells being formed. In
Learning Summary Inheritance, variation and evolution Content Sexual and asexual reproduction. Meiosis leads to non-identical cells being formed while mitosis leads to identical cells being formed. In
Full file at CHAPTER 2 Genetics
 CHAPTER 2 Genetics MULTIPLE CHOICE 1. Chromosomes are a. small linear bodies. b. contained in cells. c. replicated during cell division. 2. A cross between true-breeding plants bearing yellow seeds produces
CHAPTER 2 Genetics MULTIPLE CHOICE 1. Chromosomes are a. small linear bodies. b. contained in cells. c. replicated during cell division. 2. A cross between true-breeding plants bearing yellow seeds produces
Principles of QTL Mapping. M.Imtiaz
 Principles of QTL Mapping M.Imtiaz Introduction Definitions of terminology Reasons for QTL mapping Principles of QTL mapping Requirements For QTL Mapping Demonstration with experimental data Merit of QTL
Principles of QTL Mapping M.Imtiaz Introduction Definitions of terminology Reasons for QTL mapping Principles of QTL mapping Requirements For QTL Mapping Demonstration with experimental data Merit of QTL
IT is an accepted concept that biological species are for reproductive isolation. For example, floral traits that
 Copyright 2001 by the Genetics Society of America A Genome-Wide Survey of Reproductive Barriers in an Intraspecific Hybrid Yoshiaki Harushima,* Masahiro Nakagahra, Masahiro Yano, Takuji Sasaki and Nori
Copyright 2001 by the Genetics Society of America A Genome-Wide Survey of Reproductive Barriers in an Intraspecific Hybrid Yoshiaki Harushima,* Masahiro Nakagahra, Masahiro Yano, Takuji Sasaki and Nori
Meiosis and Sexual Life Cycles
 CAMPBELL BIOLOGY IN FOCUS URRY CAIN WASSERMAN MINORSKY REECE 10 Meiosis and Sexual Life Cycles Lecture Presentations by Kathleen Fitzpatrick and Nicole Tunbridge, Simon Fraser University SECOND EDITION
CAMPBELL BIOLOGY IN FOCUS URRY CAIN WASSERMAN MINORSKY REECE 10 Meiosis and Sexual Life Cycles Lecture Presentations by Kathleen Fitzpatrick and Nicole Tunbridge, Simon Fraser University SECOND EDITION
Genetic proof of chromatin diminution under mitotic agamospermy
 Genetic proof of chromatin diminution under mitotic agamospermy Evgenii V. Levites Institute of Cytology and Genetics, Siberian Branch of the Russian Academy of Sciences, Novosibirsk, Russia Email: levites@bionet.nsc.ru
Genetic proof of chromatin diminution under mitotic agamospermy Evgenii V. Levites Institute of Cytology and Genetics, Siberian Branch of the Russian Academy of Sciences, Novosibirsk, Russia Email: levites@bionet.nsc.ru
PRINCIPLES OF MENDELIAN GENETICS APPLICABLE IN FORESTRY. by Erich Steiner 1/
 PRINCIPLES OF MENDELIAN GENETICS APPLICABLE IN FORESTRY by Erich Steiner 1/ It is well known that the variation exhibited by living things has two components, one hereditary, the other environmental. One
PRINCIPLES OF MENDELIAN GENETICS APPLICABLE IN FORESTRY by Erich Steiner 1/ It is well known that the variation exhibited by living things has two components, one hereditary, the other environmental. One
Processes of Evolution
 15 Processes of Evolution Forces of Evolution Concept 15.4 Selection Can Be Stabilizing, Directional, or Disruptive Natural selection can act on quantitative traits in three ways: Stabilizing selection
15 Processes of Evolution Forces of Evolution Concept 15.4 Selection Can Be Stabilizing, Directional, or Disruptive Natural selection can act on quantitative traits in three ways: Stabilizing selection
Evolution of phenotypic traits
 Quantitative genetics Evolution of phenotypic traits Very few phenotypic traits are controlled by one locus, as in our previous discussion of genetics and evolution Quantitative genetics considers characters
Quantitative genetics Evolution of phenotypic traits Very few phenotypic traits are controlled by one locus, as in our previous discussion of genetics and evolution Quantitative genetics considers characters
Nature Genetics: doi: /ng Supplementary Figure 1. The phenotypes of PI , BR121, and Harosoy under short-day conditions.
 Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Meiosis and Sexual Life Cycles
 Chapter 13 Meiosis and Sexual Life Cycles Lecture Outline Overview Living organisms are distinguished by their ability to reproduce their own kind. Offspring resemble their parents more than they do less
Chapter 13 Meiosis and Sexual Life Cycles Lecture Outline Overview Living organisms are distinguished by their ability to reproduce their own kind. Offspring resemble their parents more than they do less
Essential Questions. Meiosis. Copyright McGraw-Hill Education
 Essential Questions How does the reduction in chromosome number occur during meiosis? What are the stages of meiosis? What is the importance of meiosis in providing genetic variation? Meiosis Vocabulary
Essential Questions How does the reduction in chromosome number occur during meiosis? What are the stages of meiosis? What is the importance of meiosis in providing genetic variation? Meiosis Vocabulary
RFLP facilitated analysis of tiller and leaf angles in rice (Oryza sativa L.)
") Euphytica 109: 79 84, 1999. 1999 Kluwer Academic Publishers. Printed in the Netherlands. 79 RFLP facilitated analysis of tiller and leaf angles in rice (Oryza sativa L.) Zhikang Li 1,2,3, Andrew H. Paterson
Euphytica 109: 79 84, 1999. 1999 Kluwer Academic Publishers. Printed in the Netherlands. 79 RFLP facilitated analysis of tiller and leaf angles in rice (Oryza sativa L.) Zhikang Li 1,2,3, Andrew H. Paterson
Supplementary Methods
 Supplementary Methods Microarray analysis Grains of 7 DAP of the wild-type and gif1 were harvested for RNA preparation. Microarray analysis was performed with the Affymetrix (Santa Clara, CA) GeneChip
Supplementary Methods Microarray analysis Grains of 7 DAP of the wild-type and gif1 were harvested for RNA preparation. Microarray analysis was performed with the Affymetrix (Santa Clara, CA) GeneChip
Meiosis and Sexual Life Cycles
 Chapter 13 Meiosis and Sexual Life Cycles Lecture Outline Overview: Variations on a Theme Living organisms are distinguished by their ability to reproduce their own kind. Offspring resemble their parents
Chapter 13 Meiosis and Sexual Life Cycles Lecture Outline Overview: Variations on a Theme Living organisms are distinguished by their ability to reproduce their own kind. Offspring resemble their parents
Genetics 275 Notes Week 7
 Cytoplasmic Inheritance Genetics 275 Notes Week 7 Criteriafor recognition of cytoplasmic inheritance: 1. Reciprocal crosses give different results -mainly due to the fact that the female parent contributes
Cytoplasmic Inheritance Genetics 275 Notes Week 7 Criteriafor recognition of cytoplasmic inheritance: 1. Reciprocal crosses give different results -mainly due to the fact that the female parent contributes
Meiosis and Life Cycles - 1
 Meiosis and Life Cycles - 1 We have just finished looking at the process of mitosis, a process that produces cells genetically identical to the original cell. Mitosis ensures that each cell of an organism
Meiosis and Life Cycles - 1 We have just finished looking at the process of mitosis, a process that produces cells genetically identical to the original cell. Mitosis ensures that each cell of an organism
Chapter 10 Sexual Reproduction and Genetics
 Sexual Reproduction and Genetics Section 1: Meiosis Section 2: Mendelian Genetics Section 3: Gene Linkage and Polyploidy Click on a lesson name to select. Chromosomes and Chromosome Number! Human body
Sexual Reproduction and Genetics Section 1: Meiosis Section 2: Mendelian Genetics Section 3: Gene Linkage and Polyploidy Click on a lesson name to select. Chromosomes and Chromosome Number! Human body
Name: Period: EOC Review Part F Outline
 Name: Period: EOC Review Part F Outline Mitosis and Meiosis SC.912.L.16.17 Compare and contrast mitosis and meiosis and relate to the processes of sexual and asexual reproduction and their consequences
Name: Period: EOC Review Part F Outline Mitosis and Meiosis SC.912.L.16.17 Compare and contrast mitosis and meiosis and relate to the processes of sexual and asexual reproduction and their consequences
UON, CAS, DBSC, General Biology II (BIOL102) Dr. Mustafa. A. Mansi. The Origin of Species
 Dr. Mustafa. A. Mansi. The Origin of Species") The Origin of Species Galápagos Islands, landforms newly emerged from the sea, despite their geologic youth, are filled with plants and animals known no-where else in the world, Speciation: The origin
The Origin of Species Galápagos Islands, landforms newly emerged from the sea, despite their geologic youth, are filled with plants and animals known no-where else in the world, Speciation: The origin
genome a specific characteristic that varies from one individual to another gene the passing of traits from one generation to the next
 genetics the study of heredity heredity sequence of DNA that codes for a protein and thus determines a trait genome a specific characteristic that varies from one individual to another gene trait the passing
genetics the study of heredity heredity sequence of DNA that codes for a protein and thus determines a trait genome a specific characteristic that varies from one individual to another gene trait the passing
Genetic and physiological approach to elucidation of Cd absorption mechanism by rice plants
 Genetic and physiological approach to elucidation of Cd absorption mechanism by rice plants Satoru Ishikawa National Institute for Agro-Environmental Sciences, 3-1-3, Kannondai, Tsukuba, Ibaraki, 305-8604,
Genetic and physiological approach to elucidation of Cd absorption mechanism by rice plants Satoru Ishikawa National Institute for Agro-Environmental Sciences, 3-1-3, Kannondai, Tsukuba, Ibaraki, 305-8604,
DNA Structure and Function
 DNA Structure and Function Nucleotide Structure 1. 5-C sugar RNA ribose DNA deoxyribose 2. Nitrogenous Base N attaches to 1 C of sugar Double or single ring Four Bases Adenine, Guanine, Thymine, Cytosine
DNA Structure and Function Nucleotide Structure 1. 5-C sugar RNA ribose DNA deoxyribose 2. Nitrogenous Base N attaches to 1 C of sugar Double or single ring Four Bases Adenine, Guanine, Thymine, Cytosine
Biology, 7e (Campbell) Chapter 13: Meiosis and Sexual Life Cycles
 Chapter 13: Meiosis and Sexual Life Cycles") Biology, 7e (Campbell) Chapter 13: Meiosis and Sexual Life Cycles Chapter Questions 1) What is a genome? A) the complete complement of an organism's genes B) a specific sequence of polypeptides within
Biology, 7e (Campbell) Chapter 13: Meiosis and Sexual Life Cycles Chapter Questions 1) What is a genome? A) the complete complement of an organism's genes B) a specific sequence of polypeptides within
THE WORK OF GREGOR MENDEL
 GENETICS NOTES THE WORK OF GREGOR MENDEL Genetics-. - Austrian monk- the father of genetics- carried out his work on. Pea flowers are naturally, which means that sperm cells fertilize the egg cells in
GENETICS NOTES THE WORK OF GREGOR MENDEL Genetics-. - Austrian monk- the father of genetics- carried out his work on. Pea flowers are naturally, which means that sperm cells fertilize the egg cells in
Chapter 6 Meiosis and Mendel
 UNIT 3 GENETICS Chapter 6 Meiosis and Mendel 1 hairy ears (hypertrichosis)- due to holandric gene. (Y chromosome)-only occurs in males. Appears in all sons. 2 Polydactyly- having extra fingers Wendy the
UNIT 3 GENETICS Chapter 6 Meiosis and Mendel 1 hairy ears (hypertrichosis)- due to holandric gene. (Y chromosome)-only occurs in males. Appears in all sons. 2 Polydactyly- having extra fingers Wendy the
Chromosome Chr Duplica Duplic t a ion Pixley
 Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
GENETICS 603 Exam 3, Dec 2, 2011
 GENETICS 603 Exam 3, Dec 2, 2011 Name I. Chromosome IV in Drosophila is a very small chromosome with few genes. Fruitflies that are trisomic for chromosome IV have no apparent phenotypic abnormalities,
GENETICS 603 Exam 3, Dec 2, 2011 Name I. Chromosome IV in Drosophila is a very small chromosome with few genes. Fruitflies that are trisomic for chromosome IV have no apparent phenotypic abnormalities,
Meiosis and Sexual Life Cycles
 13 Meiosis and Sexual Life Cycles Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Variations on a Theme Living
13 Meiosis and Sexual Life Cycles Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Variations on a Theme Living
Lecture WS Evolutionary Genetics Part I 1
 Quantitative genetics Quantitative genetics is the study of the inheritance of quantitative/continuous phenotypic traits, like human height and body size, grain colour in winter wheat or beak depth in
Quantitative genetics Quantitative genetics is the study of the inheritance of quantitative/continuous phenotypic traits, like human height and body size, grain colour in winter wheat or beak depth in
is the scientific study of. Gregor Mendel was an Austrian monk. He is considered the of genetics. Mendel carried out his work with ordinary garden.
 11-1 The 11-1 Work of Gregor Mendel The Work of Gregor Mendel is the scientific study of. Gregor Mendel was an Austrian monk. He is considered the of genetics. Mendel carried out his work with ordinary
11-1 The 11-1 Work of Gregor Mendel The Work of Gregor Mendel is the scientific study of. Gregor Mendel was an Austrian monk. He is considered the of genetics. Mendel carried out his work with ordinary
A Stochastic Model for the Development of. Bateson-Dobzhansky-Muller Incompatibilities that Incorporates. Protein Interaction Networks
 A Stochastic Model for the Development of Bateson-Dobzhansky-Muller Incompatibilities that Incorporates Protein Interaction Networks Garner Cochran,, Andrius Dagilis,, Karen MacPherson,, Kerry Seitz,,
A Stochastic Model for the Development of Bateson-Dobzhansky-Muller Incompatibilities that Incorporates Protein Interaction Networks Garner Cochran,, Andrius Dagilis,, Karen MacPherson,, Kerry Seitz,,
Animal Genetics - MENDELU
 Mendel and his experiments Animal Genetics Gregor Johann Mendel (1822-1884) was born in Heinzendorf, (nowadays in the Czech Republic). During the period in which Mendel developed his theory of heredity,
Mendel and his experiments Animal Genetics Gregor Johann Mendel (1822-1884) was born in Heinzendorf, (nowadays in the Czech Republic). During the period in which Mendel developed his theory of heredity,
Genetically Engineering Yeast to Understand Molecular Modes of Speciation
 Genetically Engineering Yeast to Understand Molecular Modes of Speciation Mark Umbarger Biophysics 242 May 6, 2004 Abstract: An understanding of the molecular mechanisms of speciation (reproductive isolation)
Genetically Engineering Yeast to Understand Molecular Modes of Speciation Mark Umbarger Biophysics 242 May 6, 2004 Abstract: An understanding of the molecular mechanisms of speciation (reproductive isolation)
Darwinian Selection. Chapter 7 Selection I 12/5/14. v evolution vs. natural selection? v evolution. v natural selection
 Chapter 7 Selection I Selection in Haploids Selection in Diploids Mutation-Selection Balance Darwinian Selection v evolution vs. natural selection? v evolution ² descent with modification ² change in allele
Chapter 7 Selection I Selection in Haploids Selection in Diploids Mutation-Selection Balance Darwinian Selection v evolution vs. natural selection? v evolution ² descent with modification ² change in allele
Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section B and ONE question from Section C.
 UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2017-2018 GENETICS BIO-5009A Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section
UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2017-2018 GENETICS BIO-5009A Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section
Mendel and the Gene Idea. Biology Exploring Life Section Modern Biology Section 9-1
 Mendel and the Gene Idea Biology Exploring Life Section 10.0-10.2 Modern Biology Section 9-1 Objectives Summarize the Blending Hypothesis and the problems associated with it. Describe the methods used
Mendel and the Gene Idea Biology Exploring Life Section 10.0-10.2 Modern Biology Section 9-1 Objectives Summarize the Blending Hypothesis and the problems associated with it. Describe the methods used
Model plants and their Role in genetic manipulation. Mitesh Shrestha
 Model plants and their Role in genetic manipulation Mitesh Shrestha Definition of Model Organism Specific species or organism Extensively studied in research laboratories Advance our understanding of Cellular
Model plants and their Role in genetic manipulation Mitesh Shrestha Definition of Model Organism Specific species or organism Extensively studied in research laboratories Advance our understanding of Cellular
Title. Authors. Characterization of a major QTL for manganese accumulation in rice grain
 Title Characterization of a major QTL for manganese accumulation in rice grain Authors Chaolei Liu, Guang Chen, Yuanyuan Li, Youlin Peng, Anpeng Zhang, Kai Hong, Hongzhen Jiang, Banpu Ruan, Bin Zhang,
Title Characterization of a major QTL for manganese accumulation in rice grain Authors Chaolei Liu, Guang Chen, Yuanyuan Li, Youlin Peng, Anpeng Zhang, Kai Hong, Hongzhen Jiang, Banpu Ruan, Bin Zhang,
Lesson Overview Meiosis
 11.4 THINK ABOUT IT As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located. They expected genes to be carried on structures inside the cell, but which structures?
11.4 THINK ABOUT IT As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located. They expected genes to be carried on structures inside the cell, but which structures?
Chromosome duplication and distribution during cell division
 CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
MEIOSIS LAB INTRODUCTION PART I: SIMULATION OF MEIOSIS EVOLUTION. Activity #9
 AP BIOLOGY EVOLUTION Unit 1 Part 7 Chapter 13 Activity #9 NAME DATE PERIOD MEIOSIS LAB INTRODUCTION Meiosis involves two successive nuclear divisions that produce four haploid cells. Meiosis I is the reduction
AP BIOLOGY EVOLUTION Unit 1 Part 7 Chapter 13 Activity #9 NAME DATE PERIOD MEIOSIS LAB INTRODUCTION Meiosis involves two successive nuclear divisions that produce four haploid cells. Meiosis I is the reduction
Chapter 11 INTRODUCTION TO GENETICS
 Chapter 11 INTRODUCTION TO GENETICS 11-1 The Work of Gregor Mendel I. Gregor Mendel A. Studied pea plants 1. Reproduce sexually (have two sex cells = gametes) 2. Uniting of male and female gametes = Fertilization
Chapter 11 INTRODUCTION TO GENETICS 11-1 The Work of Gregor Mendel I. Gregor Mendel A. Studied pea plants 1. Reproduce sexually (have two sex cells = gametes) 2. Uniting of male and female gametes = Fertilization
Review of Plant Cytogenetics
 Review of Plant Cytogenetics Updated 2/13/06 Reading: Richards, A.J. and R.K. Dawe. 1998. Plant centromeres: structure and control. Current Op. Plant Biol. 1: 130-135. R.K. Dawe. 2005. Centromere renewal
Review of Plant Cytogenetics Updated 2/13/06 Reading: Richards, A.J. and R.K. Dawe. 1998. Plant centromeres: structure and control. Current Op. Plant Biol. 1: 130-135. R.K. Dawe. 2005. Centromere renewal
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/331/6019/876/dc1 Supporting Online Material for Synthetic Clonal Reproduction Through Seeds Mohan P. A. Marimuthu, Sylvie Jolivet, Maruthachalam Ravi, Lucie Pereira,
www.sciencemag.org/cgi/content/full/331/6019/876/dc1 Supporting Online Material for Synthetic Clonal Reproduction Through Seeds Mohan P. A. Marimuthu, Sylvie Jolivet, Maruthachalam Ravi, Lucie Pereira,
CSS 350 Midterm #2, 4/2/01
 6. In corn three unlinked dominant genes are necessary for aleurone color. The genotypes B-D-B- are colored. If any of these loci is homozygous recessive the aleurone will be colorless. What is the expected
6. In corn three unlinked dominant genes are necessary for aleurone color. The genotypes B-D-B- are colored. If any of these loci is homozygous recessive the aleurone will be colorless. What is the expected
Duplication of an upstream silencer of FZP increases grain yield in rice
 SUPPLEMENTARY INFORMATION Articles https://doi.org/10.1038/s41477-017-0042-4 In the format provided by the authors and unedited. Duplication of an upstream silencer of FZP increases grain yield in rice
SUPPLEMENTARY INFORMATION Articles https://doi.org/10.1038/s41477-017-0042-4 In the format provided by the authors and unedited. Duplication of an upstream silencer of FZP increases grain yield in rice
AP Biology Unit 6 Practice Test 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8
 AP Biology Unit 6 Practice Test Name: 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8 picograms of DNA per nucleus. How many picograms
AP Biology Unit 6 Practice Test Name: 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8 picograms of DNA per nucleus. How many picograms
Homework Assignment, Evolutionary Systems Biology, Spring Homework Part I: Phylogenetics:
 Homework Assignment, Evolutionary Systems Biology, Spring 2009. Homework Part I: Phylogenetics: Introduction. The objective of this assignment is to understand the basics of phylogenetic relationships
Homework Assignment, Evolutionary Systems Biology, Spring 2009. Homework Part I: Phylogenetics: Introduction. The objective of this assignment is to understand the basics of phylogenetic relationships
Pollen fertility Vs Spikelet fertility in F2 of a CMS based hybrids in rice (Oryza sativa L.) under Aerobic condition
 under Aerobic condition") Research Article Pollen fertility Vs Spikelet fertility in F of a CMS based hybrids in rice (Oryza sativa L.) under Aerobic condition N.Naresh Babu,N.Shivakumar and Shailaja Hittalmani Abstract : Identification
Research Article Pollen fertility Vs Spikelet fertility in F of a CMS based hybrids in rice (Oryza sativa L.) under Aerobic condition N.Naresh Babu,N.Shivakumar and Shailaja Hittalmani Abstract : Identification
BIOLOGY. Meiosis and Sexual Life Cycles CAMPBELL. Reece Urry Cain Wasserman Minorsky Jackson
 CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson 13 Meiosis and Sexual Life Cycles Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick Variations on a Theme Living
CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson 13 Meiosis and Sexual Life Cycles Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick Variations on a Theme Living
Meiosis and Sexual Life Cycles
 Chapter 13 Meiosis and Sexual Life Cycles Lecture Outline Overview: Variations on a Theme Living organisms are distinguished by their ability to reproduce their own kind. Offspring resemble their parents
Chapter 13 Meiosis and Sexual Life Cycles Lecture Outline Overview: Variations on a Theme Living organisms are distinguished by their ability to reproduce their own kind. Offspring resemble their parents
2 Numbers in parentheses refer to literature cited.
 A Genetic Study of Monogerm and Multigerm Characters in Beets V. F. SAVITSKY 1 Introduction Monogerm beets were found in the variety Michigan Hybrid 18 in Oregon in 1948. Two of these monogerm plants,
A Genetic Study of Monogerm and Multigerm Characters in Beets V. F. SAVITSKY 1 Introduction Monogerm beets were found in the variety Michigan Hybrid 18 in Oregon in 1948. Two of these monogerm plants,
CAPE Biology Unit 1 Scheme of Work
 CAPE Biology Unit 1 Scheme of Work 2011-2012 Term 1 DATE SYLLABUS OBJECTIVES TEXT PAGES ASSIGNMENTS COMMENTS Orientation Introduction to CAPE Biology syllabus content and structure of the exam Week 05-09
CAPE Biology Unit 1 Scheme of Work 2011-2012 Term 1 DATE SYLLABUS OBJECTIVES TEXT PAGES ASSIGNMENTS COMMENTS Orientation Introduction to CAPE Biology syllabus content and structure of the exam Week 05-09
1. they are influenced by many genetic loci. 2. they exhibit variation due to both genetic and environmental effects.
 October 23, 2009 Bioe 109 Fall 2009 Lecture 13 Selection on quantitative traits Selection on quantitative traits - From Darwin's time onward, it has been widely recognized that natural populations harbor
October 23, 2009 Bioe 109 Fall 2009 Lecture 13 Selection on quantitative traits Selection on quantitative traits - From Darwin's time onward, it has been widely recognized that natural populations harbor
Speciation. Mechanisms of Speciation. Title goes here. Some Key Tenets of the Modern Synthesis
 Carol Eunmi Lee 11/9/17 Speciation Increasing genetic distance Fitness Mating between different species Mating between relatives Inbreeding Depression Hybrid Vigor Outbreeding Depression 2 Darwin s Origin
Carol Eunmi Lee 11/9/17 Speciation Increasing genetic distance Fitness Mating between different species Mating between relatives Inbreeding Depression Hybrid Vigor Outbreeding Depression 2 Darwin s Origin
Genetics (patterns of inheritance)
") MENDELIAN GENETICS branch of biology that studies how genetic characteristics are inherited MENDELIAN GENETICS Gregory Mendel, an Augustinian monk (1822-1884), was the first who systematically studied
MENDELIAN GENETICS branch of biology that studies how genetic characteristics are inherited MENDELIAN GENETICS Gregory Mendel, an Augustinian monk (1822-1884), was the first who systematically studied
MEIOSIS C H A P T E R 1 3
 MEIOSIS CHAPTER 13 CENTRAL DOGMA OF BIOLOGY DNA RNA Protein OFFSPRING ACQUIRE GENES FROM PARENTS Genes are segments of DNA that program specific traits. Genetic info is transmitted as specific sequences
MEIOSIS CHAPTER 13 CENTRAL DOGMA OF BIOLOGY DNA RNA Protein OFFSPRING ACQUIRE GENES FROM PARENTS Genes are segments of DNA that program specific traits. Genetic info is transmitted as specific sequences
Legend: S spotted Genotypes: P1 SS & ss F1 Ss ss plain F2 (with ratio) 1SS :2 WSs: 1ss. Legend W white White bull 1 Ww red cows ww ww red
 1SS :2 WSs: 1ss. Legend W white White bull 1 Ww red cows ww ww red") On my honor, this is my work GENETICS 310 EXAM 1 June 8, 2018 I. Following are 3 sets of data collected from crosses: 1. Spotted by Plain gave all spotted in the F1 and 9 spotted and 3 plain in the F2.
On my honor, this is my work GENETICS 310 EXAM 1 June 8, 2018 I. Following are 3 sets of data collected from crosses: 1. Spotted by Plain gave all spotted in the F1 and 9 spotted and 3 plain in the F2.
Designer Genes C Test
 Northern Regional: January 19 th, 2019 Designer Genes C Test Name(s): Team Name: School Name: Team Number: Rank: Score: Directions: You will have 50 minutes to complete the test. You may not write on the
Northern Regional: January 19 th, 2019 Designer Genes C Test Name(s): Team Name: School Name: Team Number: Rank: Score: Directions: You will have 50 minutes to complete the test. You may not write on the
Cell Division: the process of copying and dividing entire cells The cell grows, prepares for division, and then divides to form new daughter cells.
 Mitosis & Meiosis SC.912.L.16.17 Compare and contrast mitosis and meiosis and relate to the processes of sexual and asexual reproduction and their consequences for genetic variation. 1. Students will describe
Mitosis & Meiosis SC.912.L.16.17 Compare and contrast mitosis and meiosis and relate to the processes of sexual and asexual reproduction and their consequences for genetic variation. 1. Students will describe
Major questions of evolutionary genetics. Experimental tools of evolutionary genetics. Theoretical population genetics.
 Evolutionary Genetics (for Encyclopedia of Biodiversity) Sergey Gavrilets Departments of Ecology and Evolutionary Biology and Mathematics, University of Tennessee, Knoxville, TN 37996-6 USA Evolutionary
Evolutionary Genetics (for Encyclopedia of Biodiversity) Sergey Gavrilets Departments of Ecology and Evolutionary Biology and Mathematics, University of Tennessee, Knoxville, TN 37996-6 USA Evolutionary
Drosophila melanogaster and D. simulans, two fruit fly species that are nearly
 Comparative Genomics: Human versus chimpanzee 1. Introduction The chimpanzee is the closest living relative to humans. The two species are nearly identical in DNA sequence (>98% identity), yet vastly different
Comparative Genomics: Human versus chimpanzee 1. Introduction The chimpanzee is the closest living relative to humans. The two species are nearly identical in DNA sequence (>98% identity), yet vastly different
EXPRESSION OF THE FIS2 PROMOTER IN ARABIDOPSIS THALIANA
 EXPRESSION OF THE FIS2 PROMOTER IN ARABIDOPSIS THALIANA Item Type text; Electronic Thesis Authors Bergstrand, Lauren Janel Publisher The University of Arizona. Rights Copyright is held by the author. Digital
EXPRESSION OF THE FIS2 PROMOTER IN ARABIDOPSIS THALIANA Item Type text; Electronic Thesis Authors Bergstrand, Lauren Janel Publisher The University of Arizona. Rights Copyright is held by the author. Digital
- interactions between alleles. - multiple phenotypic effects of one gene. - phenotypic variability in single genes. - interactions between genes
 Genotype Phenotype - interactions between alleles - dominance - multiple phenotypic effects of one gene - phenotypic variability in single genes - interactions between genes amount of product threshold
Genotype Phenotype - interactions between alleles - dominance - multiple phenotypic effects of one gene - phenotypic variability in single genes - interactions between genes amount of product threshold
Eiji Yamamoto 1,2, Hiroyoshi Iwata 3, Takanari Tanabata 4, Ritsuko Mizobuchi 1, Jun-ichi Yonemaru 1,ToshioYamamoto 1* and Masahiro Yano 5,6
 Yamamoto et al. BMC Genetics 2014, 15:50 METHODOLOGY ARTICLE Open Access Effect of advanced intercrossing on genome structure and on the power to detect linked quantitative trait loci in a multi-parent
Yamamoto et al. BMC Genetics 2014, 15:50 METHODOLOGY ARTICLE Open Access Effect of advanced intercrossing on genome structure and on the power to detect linked quantitative trait loci in a multi-parent
The Origin of Species
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 24 The Origin of Species Lectures
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 24 The Origin of Species Lectures
Chromosome variations in protoplast-derived calli and in plants regenerated from the calli of
 Jpn. J. Genet. (1989) 64, pp. 355-361 Chromosome variations in protoplast-derived calli and in plants regenerated from the calli of cultivated rice (Oryza sativa L.) Soryu NISHIBAYASHI*, Yasuyuki HAYASHI,
Jpn. J. Genet. (1989) 64, pp. 355-361 Chromosome variations in protoplast-derived calli and in plants regenerated from the calli of cultivated rice (Oryza sativa L.) Soryu NISHIBAYASHI*, Yasuyuki HAYASHI,
Outline for today s lecture (Ch. 13)
") Outline for today s lecture (Ch. 13) Sexual and asexual life cycles Meiosis Origins of Genetic Variation Independent assortment Crossing over ( recombination ) Heredity Transmission of traits between generations
Outline for today s lecture (Ch. 13) Sexual and asexual life cycles Meiosis Origins of Genetic Variation Independent assortment Crossing over ( recombination ) Heredity Transmission of traits between generations
Meiosis and Sexual Life Cycles
 Chapter 13 Meiosis and Sexual Life Cycles PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
Chapter 13 Meiosis and Sexual Life Cycles PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
Ch. 10 Sexual Reproduction and Genetics. p
 Ch. 10 Sexual Reproduction and Genetics p. 270 - 10.1 Meiosis p. 270-276 Essential Question Main Idea! Meiosis produces haploid gametes Where are the instructions for each trait located in a cell?! On
Ch. 10 Sexual Reproduction and Genetics p. 270 - 10.1 Meiosis p. 270-276 Essential Question Main Idea! Meiosis produces haploid gametes Where are the instructions for each trait located in a cell?! On
Q Expected Coverage Achievement Merit Excellence. Punnett square completed with correct gametes and F2.
 NCEA Level 2 Biology (91157) 2018 page 1 of 6 Assessment Schedule 2018 Biology: Demonstrate understanding of genetic variation and change (91157) Evidence Q Expected Coverage Achievement Merit Excellence
NCEA Level 2 Biology (91157) 2018 page 1 of 6 Assessment Schedule 2018 Biology: Demonstrate understanding of genetic variation and change (91157) Evidence Q Expected Coverage Achievement Merit Excellence
Jie Chen 1), De-Run Huang 1), Lei Wang 1), Guang-Jie Liu 1,2) and Jie-Yun Zhuang* 1) Introduction
, De-Run Huang 1), Lei Wang 1), Guang-Jie Liu 1,2) and Jie-Yun Zhuang* 1) Introduction") Breeding Science 60: 153 159 (2010) Identification of quantitative trait loci for resistance to whitebacked planthopper, Sogatella furcifera, from an interspecific cross Oryza sativa O. rufipogon Jie Chen
Breeding Science 60: 153 159 (2010) Identification of quantitative trait loci for resistance to whitebacked planthopper, Sogatella furcifera, from an interspecific cross Oryza sativa O. rufipogon Jie Chen
AP Biology Essential Knowledge Cards BIG IDEA 1
 AP Biology Essential Knowledge Cards BIG IDEA 1 Essential knowledge 1.A.1: Natural selection is a major mechanism of evolution. Essential knowledge 1.A.4: Biological evolution is supported by scientific
AP Biology Essential Knowledge Cards BIG IDEA 1 Essential knowledge 1.A.1: Natural selection is a major mechanism of evolution. Essential knowledge 1.A.4: Biological evolution is supported by scientific
Introduction to Genetics
 Introduction to Genetics The Work of Gregor Mendel B.1.21, B.1.22, B.1.29 Genetic Inheritance Heredity: the transmission of characteristics from parent to offspring The study of heredity in biology is
Introduction to Genetics The Work of Gregor Mendel B.1.21, B.1.22, B.1.29 Genetic Inheritance Heredity: the transmission of characteristics from parent to offspring The study of heredity in biology is
Chapter 13: Meiosis and Sexual Life Cycles Overview: Hereditary Similarity and Variation
 Chapter 13: Meiosis and Sexual Life Cycles Overview: Hereditary Similarity and Variation Living organisms Are distinguished by their ability to reproduce their own kind Biology, 7 th Edition Neil Campbell
Chapter 13: Meiosis and Sexual Life Cycles Overview: Hereditary Similarity and Variation Living organisms Are distinguished by their ability to reproduce their own kind Biology, 7 th Edition Neil Campbell
Chapter 13- Reproduction, Meiosis, and Life Cycles. Many plants and other organisms depend on sexual reproduction.
 Chapter 13- Reproduction, Meiosis, and Life Cycles Many plants and other organisms depend on sexual reproduction. Flowers are the sexual reproductive organ systems of angiosperms. Sexual reproduction gametes
Chapter 13- Reproduction, Meiosis, and Life Cycles Many plants and other organisms depend on sexual reproduction. Flowers are the sexual reproductive organ systems of angiosperms. Sexual reproduction gametes