Identification of Sfi1 protein interactions through yeast two-hybrid analysis
|
|
|
- Hugo Andrews
- 5 years ago
- Views:
Transcription
1 University of Colorado, Boulder CU Scholar Undergraduate Honors Theses Honors Program Spring 2013 Identification of Sfi1 protein interactions through yeast two-hybrid analysis Tara Peters University of Colorado Boulder Follow this and additional works at: Recommended Citation Peters, Tara, "Identification of Sfi1 protein interactions through yeast two-hybrid analysis" (2013). Undergraduate Honors Theses This Thesis is brought to you for free and open access by Honors Program at CU Scholar. It has been accepted for inclusion in Undergraduate Honors Theses by an authorized administrator of CU Scholar. For more information, please contact
2 Identification of Sfi1 protein interactions through yeast two-hybrid analysis Tara Peters University of Colorado, Molecular, Cellular, Developmental Biology Department Undergraduate Honors Thesis April 10, 2013 Thesis Advisor: Dr. Mark Winey (MCDB) Committee Members: Dr. Jennifer Martin (MCDB) Dr. Dylan Taatjes (CHEM) Peters 1
3 Abstract The most notable eukaryotic microtubule organizing center, the centrosome, forms the mitotic spindle required for mitosis and meiosis 3. Problems with centrosomal duplication and function can lead to chromosomal missegregation, which has been linked to numerous human diseases and cancer 3. Though the structure has been closely studied in different organisms, the mechanism by which it duplicates is still not well understood 5. In the budding yeast Saccharomyces cerevisiae the spindle pole body is analogous to the human centrosome, and has numerous highly conserved components, making it an excellent model to study to better understand centrosome duplication and function 5. This study aimed to identify protein interactions of Sfi1, one essential and highly conserved SPB component, in order to characterize its role in SPB duplication. Through a yeast two-hybrid screen, the C terminus of Sfi1 was found to interact with RNA II polymerase mediator complex protein Cse2. This interaction provides more evidence for Cse2 being involved at the spindle, and further analysis of the interaction will elucidate the role of Sfi1 in SPB duplication. Peters 2
4 Introduction Microtubule organizing centers (MTOCs) are structures in eukaryotes from which the minus ends of microtubules are anchored and nucleate 1. These structures function to organize cilia and flagella and also to create the mitotic spindle for segregation of the chromosomes during cell division. There are various forms of MTOCs that range from basal bodies, which form cilia and flagella used for cellular motility and signaling, to the more notable eukaryotic centrosomes, which form the mitotic spindle 1. Cells can have numerous MTOCs floating in the cytoplasm or embedded in the nuclear or plasma membrane. They may also have just one MTOC, as is the case in the budding yeast Saccharomyces cerevisiae 1. Though the structure of MTOCs is varied among organisms, many of the components and regulators are highly conserved 5. The main animal MTOC, the centrosome, is involved specifically in chromosome segregation and is duplicated once per cell cycle in a highly regulated process to ensure formation of a bipolar spindle and progression through the cell cycle 2. Despite the critical role centrosomes play in many eukaryotic cells, centrosomes are not essential for cell division in some cell types that are able to go through the cell cycle in centrosome-independent pathways as well 3. Studying the function of centrosomes has been an ongoing area of research due to the severe impacts that have been observed in cells from alterations to the centrosome. Eukaryotes that go through embryogenesis are particularly reliant on proper function of their centrosomes. One example of this comes from mimicking fertilization in Xenopus leavis and allowing the embryo to attempt to divide without being provided the centrosome from the sperm, the egg fails to cleave after cell division and ceases to develop 3. However, lacking centrosomes during embryogenesis is not the only concern. In embryos implanted with extra centrosomes, multipolar rather than bipolar spindles are observed and chromosomes are segregated between them, leading to genomic instability and cellular defects 3. The idea from these and many other studies is that the centrosome allows formation of the mitotic spindle and ensures appropriate separation of genetic material into the daughter cells, and defects with this process can lead to severe phenotypes. Faulty duplication and function of the centrosome has been implicated in many human diseases, including cancer. Genetic instability, which can be caused by missegregation of the Peters 3
5 chromosomes by the centrosomes, can lead to aneuploidy 3 It can manifest as extra copies of chromosomes, chromosomal translocations, deletions, or breaks in the DNA 3. Genetic instability is also one of the hallmarks of cancer cells that contribute to their erratic behavior, and can occur prior to or after cellular transformation 4. Cancer cells have frequently been observed to have extra centrosomes. Extra centrosomes might cause cells to go through a brief multipolar spindle phase before completing division into two daughter cells, which disrupts the normally even pull of chromosomes into the daughter cells and leads to chromosomal missegregation 4. Understanding the mechanisms by which these structures duplicate and function is critical to better understanding cancer and other diseases caused by centrosomal defects and how to treat them. The only MTOC in S. cerevisiae is called the spindle pole body (SPB), and it nucleates cytoplasmic microtubules that orient the mitotic spindle during cell division and spindle microtubules that separate the chromosomes 17. Animal centrosomes also nucleate both cytoplasmic and spindle microtubules 3. The cytoplasmic microtubules from centrosomes orient the spindle during mitosis, and in animal cells there are other sources of cytoplasmic microtubules for purposes unrelated to the spindle as well 3. Variability of MTOCs can be seen when comparing the structure of the SPB to that of the human centrosome. The human centrosome is made of a pair of centrioles, which are complex microtubule structures, surrounded by a matrix of proteins. It does not associate with the nuclear membrane until mitosis, then after nuclear envelope breakdown the centrosomes may form the mitotic spindle 1. Rather than being made of the barrel shaped centrioles, the SPB is a tri-laminar structure embedded in the nuclear envelope throughout the entire cell cycle. The structure of the SPB can be seen in Fig. 1. It is composed of an outer plaque, on the cytoplasmic side of the nuclear envelope, a central plaque that lies in conjunction with the nuclear envelope, an inner plaque that faces the nucleoplasm, and a specialized region of the nuclear envelope called the half-bridge sits on one side of the central plaque 5. The SPB is an excellent model to study centrosome duplication and function because all of the components of this simpler centrosome have been identified, and it shares many of the same components and regulators as the human centrosome 11. Sixteen of the eighteen core SPB components are essential genes 11. The fact that so many of the SPB genes are essential makes it a difficult but interesting centrosome to study, as many unique phenotypes can be observed. Peters 4
6 A model for SPB duplication (Fig. 2) has been developed based on observational electron microscopy data of wild type SPBs and SPBs with mutated components. Duplication begins during early G1 phase of the cell cycle with the elongation of the half-bridge 6. The halfbridge elongates from about 90nm to twice it s original size, 180nm 6. After the half-bridge has elongated, a satellite of four core SPB components, Spc42, Spc29, Cnm67, and Nud1, is then deposited on the cytoplasmic side of the half-bridge, distal to the mother SPB 5. Next the satellite expands to a layered structure that resembles the central plaque of the mother SPB 5. As it expands, the duplication plaque is inserted into the nuclear envelope 5. Finally the nuclear components and gamma tubulin complexes, which are anchored to both the inner and outer plaque and serve as the site for microtubules to bind and nucleate, are assembled to create an identical daughter SPB 5. The duplicated SPBs sit side by side at the end of G1/beginning of S phase, but they must separate and migrate to opposite ends of the nucleus to form the mitotic spindle. Sfi1 is an essential component of the SPB that localizes to the half-bridge, and is highly conserved among organsims 6. It binds the yeast centrin, Cdc31, another SPB component that localizes to the half-bridge, along DNA sequence repeats in the central, alpha-helical domain of the protein 6. Sfi1 has 21 of these repeats and binds one molecule of Cdc31 per repeat 6. Kilmartin and colleagues (2006) found unique localization of the termini of Sfi1 by immunoelectron microscopy, with the amino (N) terminus next to the mother SPB and the carboxy ( C) terminus at the end of the half bridge before duplication and in the center of the full-length bridge. Kilmartin (2006) constructed an Sfi1 molecule with 15 of the centrin binding repeats and found it to be a 60 nm filament. This suggests that one 90nm filament of Sfi1 may span the halfbridge with it s C terminus at the distal end, and then half-bridge elongation is due to the addition of another molecule of Sfi1 with both C termini together and the N terminus free for the satellite to be deposited at the site of new SPB assembly 6. It is thought that the free N terminus of Sfi1 may play a role in recruiting the satellite components to the distal end of the half-bridge 2. There is no published data specifically on the N terminus of Sfi1 and limited data on the other domains of the protein. Mutants in the repeat domain of Sfi1 arrest prior to duplication of the SPBs, indicating the protein is required for duplication 5. Additionally, four alleles of Sfi1 with different point mutations in the C terminus caused arrest of the cell cycle in mitosis 7. Electron microscopy found a novel phenotype in which the cells had fully duplicated side-by- Peters 5
7 side SPBs 7. Another paper found through a synthetic lethal screen that SFI1 and BIK1 genetically interact 17. Bik1 is a homolog of the mammalian CLIP-170 and is a microtubuleassociated protein that stabilizes microtubules during mitosis 17. A mutant in the C terminus of Sfi1 in a Bik1 null background also showed the arrest in mitosis with duplicated side by side SPBs 17. These findings with mutations in the repeats and C terminus of Sfi1 indicate that it may have an important role in initiating duplication of the SPBs and in splitting of the bridge after duplication of the SPBs as well. Based on the data discussed above, it would be interesting to determine whether the termini of Sfi1 play distinct roles in SPB duplication and whether this would give a better understanding of the function of Sfi1. The yeast two-hybrid method is a powerful tool for identifying protein-protein interactions and has been used to identify interactions of many other SPB components already. The two-hybrid system uses the modularity of transcription factors to determine protein interactions 8 (Fig. 3). One protein is fused to the DNA binding domain of a transcription factor and the other is fused to the activation domain of the same transcription factor. If the proteins interact, the transcription factor is brought together, therefore activating transcription of a reporter gene that it regulates 9. In addition to Sfi1 interactions identified with fellow half-bridge component Cdc31, many genetic interactions involving Sfi1 have been found in synthetic lethal screens 6. Fulllength Sfi1 was found to interact with another SPB component, Spc110, in a complex in an affinity capture experiment 6. SFI1 exists within a genetic network of interactions with the genes encoding SPB components BBP1, NDC1, MPS2 and KAR1 as determined by a screen for suppression of the synthetic lethality of a sfi1-65, mad1 Δ double mutant, with the Sfi1 mutation in the C terminus 7. Kar1 is also part of the specialized region of the nuclear envelope called the half-bridge, while Bbp1, Ndc1, and Mps2 are involved in inserting the newly duplicated SPB into the nuclear envelope 7. The results from that screen suggest that mutations in the C terminus of Sfi1 do not result from defective interaction with Cdc31 because Cdc31 overexpression could not rescue the synthetic lethality, but possibly from disrupted interactions of proteins involved in the process of insertion at the nuclear envelope 7. Genetic interactions of SFI1 have been identified with motor associated protein CIK1, and kinetochore protein BUB3 12. SFI1 showed genetic interaction with genes encoding motor associated proteins Cin8 and Kar3, and Bub1, which forms a complex required for the spindle checkpoint 7,17. The use of genetic screens has Peters 6
8 also identified some protein interactions for Sfi1 that would likely not be anticipated otherwise and exemplify the complexity of protein interactions in the cell 12. Though all of the data on interactions does give more evidence for the importance of Sfi1 to the SPB, more research is still needed to fully elucidate Sfi1s critical function. Studying distinct domains of a protein separately is a useful way to understand how a protein functions, and that idea drives this study of Sfi1. The model for SPB duplication and localization data of Sfi1 described above suggest that the N terminus of Sfi1 may interact with core SPB components, specifically, the satellite proteins Spc29, Spc42, Cnm67 and Nud1, while the C terminus may interact with itself or other half-bridge proteins, such as Kar1 or Mps3. Based on that model, this study tested the N terminus for interaction with SPB satellite components Spc29 and Spc42, and the C terminus was tested for interaction with itself and screened against a library containing approximately all of the 6000 open reading frames (ORFs) in the S. cerevisiae genome 18. The C terminus of Sfi1 was also tested for interaction with itself including the two Sfi1 repeats that directly precede the C terminus because those repeats bind Cdc31 and may play a critical role in the function of the protein. The C terminus was not found to interact directly with itself or with itself when two Sfi1 repeats were included before the C terminal domain begins, indicating some other proteins may be involved in duplication and addition of additional molecules of Sfi1 at the half-bridge. The C terminus of Sfi1 was found to interact with RNA polymerase II mediator complex component Cse2 through a screen of the S.cerevisiae genome ORFs. A genetic interaction between SFI1 and CSE2 was confirmed previously, but no data on the C terminus of Sfi1 being sufficient for this interaction has been shown until now 12. Results Confirming expression of the BDSfi1C construct Expression of the bait construct BDSfi1C (plasmids constructed are listed in Table 1) was confirmed through western blot analysis as described in the methods section. Western blot analysis showed the empty pobd2 construct and BDSfi1 constructs were expressed at similar levels (Fig. 4). Directed Two-hybrid Peters 7
9 Prior to actual tests for interaction, the bait strains transformed with BDSfi1N, BDSfi1C and BDSfi1C2rep were tested for autoactivation. Only diploid strains with bait and prey constructs that interact should be able to grow on media lacking histidine because the interaction brings together the BD and AD pieces of GAL4 and activates transcription of the HIS3 reporter 16. The autoactivation test tests the bait strains on media lacking tryptophan and histidine, and with increasing concentrations of the drug 3-amino 1,2,4-triazole (3-AT), which inhibits an enzyme required for hisitidine biosynthesis, in order to eliminate any autoactivaiton of the reporter to allow isolation of strong two-hybrid interactions. All BD constructs grew normally on media lacking tryptophan (Fig. 5). The negative control, the empty BD plasmid, also grew normally on media lacking tryptophan, and showed no growth on media lacking histidine. The BDSfi1N construct showed high levels of autoactivation that required 50mM concentrations of 3-AT to eliminate what? (Fig. 5A). Sequence analysis of the N terminus of Sfi1 showed numerous regions with a net negative charge per 30 amino acids as well as many hydrophobic amino acids, which have both been shown to be important for minimal activating domains and cause autoactivation of the reporter gene 13. From amino acids 147 to 185 there are nine negatively charged residues and only two positively charged residues, and there are also 17 hydrophobic residues in that region. From amino acids 148 to 178 there is a net negative charge of 6, and from amino acid 154 to 185 there is a net negative charge of 5. To determine if the highly acidic and hydrophobic amino acids at the end of the N terminus of Sfi1 were causing the high levels of autoactivation two truncations of the terminus were created at amino acids 154 and 175. The BDSfi1N175 truncation did not decrease autoactivation (Fig. 5A), and growth was still observed on media containing 50mM 3-AT. The BDSfi1N154 truncation did decrease autoactivation (Fig. 5A) from 50mM to 5mM 3-AT. The BDSfi1C and BDSfi1C2rep constructs showed no autoactivation of the reporter so the lowest concentration, 1.5mM, of 3-AT was used (Fig. 5B). Directed two-hybrid tests with BDSfi1N as bait were performed on media lacking leucine, tryptophan and histidine, and with 50mM 3-AT, as determined by the autoactivation tests, and on media without leucine and tryptophan as a control. The following three negative controls were used: BDSfi1N: AD, BD: ADSpc29 and BD: ADSpc42, and BD: AD. All of the negative controls grew normally on diploid selective media (media without leucine and tryptophan; Fig. 6A). Of these negative controls, only BDSfi1N: AD showed a small amount of Peters 8
10 growth on media lacking histidine with 50mM 3-AT. The BDSfi1N: ADSpc29 test did show a small amount of growth on the media lacking histidine with 50mM 3-AT, but it was consistently equal to or less than that seen from the BDSfi1N: AD negative control, indicating no interaction. There was no growth seen on media lacking histidine for the BDSfi1N: ADSpc42 diploids, indicating there is no interaction between these proteins (Fig. 6A). The expression of the BD and AD constructs was not confirmed via western blot analysis for any of the constructs used in this test. The C terminus of Sfi1 was tested for interaction with itself, and the same negative controls were performed (BDSfi1C: AD, BD: ADSfi1C, and BD:AD). All combinations grew normally on diploid selective media (Fig. 6B). No growth was seen on media lacking histidine with 1.5mM 3-AT, as determined by the autoactivation test (Fig. 6B), indicating the C terminus of Sfi1 does not interact with itself. Expression of the BDSfi1C construct was confirmed with western blot analysis, however the ADSfi1C expression was not confirmed. The BDSfi1C2rep construct was also tested for interaction with the ADSfi1C2rep construct to determine if the Sfi1 repeats may be required for interaction. No growth was seen on media lacking histidine with 1.5mM 3-AT, and normal growth was seen on diploid selective media (Fig. 6C). The same negative controls were performed and showed normal growth on diploid selective media but no growth on media lacking histidine also. Expression of these constructs was not confirmed by western blot. Library Screen In this study a yeast two-hybrid screen was performed with the BDSfi1C construct as the bait to find protein interactions specifically with this terminus. The screen was performed against an ORF pool containing approximately all of the ORFs of the S. cerevisiae genome fused in frame to the AD of GAL4. The large scale liquid mating was grown on SC media lacking leucine, tryptophan and histidine, with 1.5mM 3-AT and allowed to grow at 30 C. In this screen 4.84 million clones were screened and there was a mating efficiency of 5.37%. The first two potential positive interaction colonies (called positive 17 and positive 45) were selected from the screen on day two, and the other two potential positives (positive 24 and positive 32) found in this study were selected on day 7 (Fig. 7A). Sequence analysis determined the ORF for positive 17 to be a fragment of GAL4, but not the BD or AD regions. The sequence analysis of positive 45 also matched a fragment of GAL4. Therefore, these were not true Peters 9
11 positive interactions but false positives. The ORF fused to the GAL4 AD in positives 24 and 32 was Cse2. The isolated plasmids from positives 24 and 32 were retransformed in to pj694 mating type A and a directed two-hybrid with BDSfi1C was performed to confirm interaction. The same negative controls were performed in previous two hybrid tests (BDSfi1C:AD, BD:ADCse2, and BD:AD) and showed normal growth on the diploid selective media, and no growth on media lacking leucine, tryptophan, and histidine with 1.5mM 3-AT. The BDSfi1C and ADpositive24 and 32 diploids were able to grow on media lacking leucine, tryptophan and histidine with 1.5mM 3-AT, while no growth was observed from any negative controls, indicating an interaction (Fig. 7B). Discussion This study used the yeast-two hybrid method to identify an interaction of Cse2 with the C terminus of Sfi1. The N terminus of Sfi1 was also tested for interaction with SPB satellite components Spc29 and Spc42, and no interactions were found. The C terminus of Sfi1 was not found to interact with itself through directed two-hybrid, nor did the C terminus including the two Sfi1 repeats preceding the C terminal domain interact with itself. These preliminary results need to be further verified through the methods discussed below. It has been known for some time that transcription activating domains do not frequently share much sequence homology, but share regions of many acidic and hydrophobic amino acids 13. A net negative charge in a region of approximately 30 amino acids has been shown to be a minimal transcription activating domain 13. The yeast two-hybrid system commands the modularity of the domains of a transcription factor in order to find proteins that interact enough to bring the DNA binding and transcription activating domains together and activate a reporter gene, without the domains physically touching. A problem with this system can be that a minimal activating domain within the sequence of a protein being studied through yeast twohybrid is able to activate transcription without truly interacting with the prey protein. This was observed in this study with the N terminus of Sfi1, which contains many stretches of net negative charges in its sequence. Truncating the protein and removing a total of seven negatively charged residues decreased the autoactivation of the HIS3 reporter from requiring 50mM concentrations of 3-AT, to only 5mM concentrations (Fig. 5A). These new truncations could be tested in future Peters 10
12 two-hybrid experiments, however it is not known if they would result in the same interactions in vivo. The directed two-hybrid of the N terminus of Sfi1 with SPB satellite components Spc29 and Spc42 showed no interaction (Fig. 6 ). Growth was observed on media lacking histidine with 50mM 3-AT for BDSfi1N:AD, which is consistent with the high level of autoactivation of the reporter that the BDSfi1N construct showed in the autoactivation tests. The small amount of growth seen on media lacking histidine with 50mM 3-AT for the test of BDSfi1N with ADSpc29 was most likely a result of autoactivation because the growth observed was less than that seen from the negative control. Ultimately a repeat of the experiment with confirmation of expression of the constructs could give more concrete evidence that those proteins do not interact. The C terminus of Sfi1 was not found to interact with itself in this two-hybrid analysis (Fig. 6B). When two Sfi1 repeats were included in the C terminal constructs, the HIS3 reporter gene was still not activated (Fig. 6C). This indicates that some other factors may be necessary for the C termini to interact with each other on the half-bridge. Potentially other half-bridge components Kar1 and Cdc31 are involved. There is also the possibility that the proteins do not directly interact at all, but are merely in close proximity. The results of this two-hybrid test need to be confirmed by verifying expression of the constructs. Once expression of the other constructs has been confirmed, the results of this yeast two-hybrid study provide evidence that the C terminus of Sfi1 or the C terminus of Sfi1 with two Sfi1 repeats do not interact with themselves, and other interactions or requirements for their interaction on the half-bridge of the SPB can be explored. A pull-down assay or variations of directed two-hybrid assays with other half-bridge components could be tested to find C terminal interactions. It would be interesting to test some of the C terminal Sfi1 mutant alleles previously identified that give the duplicated sideby-side SPB phenotype in two-hybrid. The phenotype of these mutant alleles may enhance some interactions and allow for detection of an interaction in this system, or give another interesting result. The yeast two-hybrid screen of Sfi1 came up with four positive hits, and of those two were false positives and two could be true positives but more analysis is needed to confirm that they are not false positives(fig. 7A). Positive 17 was found by sequence analysis to be a fragment of GAL4, and positive 45 matched a region of chromosome 16 with no annotated gene. The other two positives found were actually the ORF Cse2. The fact that interaction was found Peters 11
13 with the same protein twice in the screen is good support that this is not a false positive. One survey of false positives from the two-hybrid system found the most common ones to be heat shock proteins, then ribosomal proteins, or mitochondrial proteins, and Cse2 fits none of these categories 22. Cse2 is an essential part of the RNA polymerase II mediator complex, which is required for transcription of nearly all genes with class II gene promoters in yeast 23. Though it is an unexpected interaction for Sfi1, there are some aspects of Cse2 that lead to the conclusion that this may be a true interaction. First, in a synthetic lethal screen for protein interactions in S. cerevisiae, CSE2 and SFI1 were found to have genetic interaction 12. This interaction was found with the sfi1-3 allele, which contains a mutation in the central domain that binds Cdc Secondly, conditional mutants of Cse2 show abnormal chromosome segregation, a phenotype also observed as a result of mutations of SPB components Cin8 and Ndc1 24,27,28. Cse2 null cells show delayed progression through mitosis and chromosomal abnormalities as well 24. Chromosomal nondisjunction was also observed when mutants in Cse2 were combined with mutants in proteins at the centromere 24. This indicates that Cse2 has a role in chromosome segregation, and means it would possible for Cse2 to be at the spindle interacting with SPB components at some point during the cell cycle. Lastly, CSE2 has also been found to interact genetically with both BIM1 and CIK1 through synthetic genetic arrays 12,25. Bim1 is a microtubule binding protein that helps capture cortical microtubules at the cortex of the bud and prevent progression through mitosis in the event of abnormal spindle orientation 26. Cik1 targets Kar3, a microtubule motor that localizes to the SPB, to the microtubules and aids in proper orientation of the mitotic spindle, and again these interactions provide support for Cse2 being at the spindle 11. The fact that these genetic interactions have been found and that mutants in Cse2 show spindle related defects indicates that the two-hybrid interaction identified in this study should be studied further. This, taken together with the defects in chromosomal segregation of Cse2 mutants, led to the hypothesis that Cse2 could either be involved in interactions between chromosomes and microtubules, such as at the kinetochore, or that Cse2 could be important for expression of proteins involved in chromosomal segregation 24. Sfi1 does not localize to kinetochores or microtubules, but its role in SPB duplication and separation at the half-bridge has been shown 6,7. The genetic interaction of CIK1 with both CSE2 and SFI1 provides an interesting link at the Peters 12
14 SPB. It has been suggested that Cik1 may play a role with Kar3 in separation of the SPBs, and is conceivable, because of the genetic interactions observed, that Cse2 could be actively involved in SPB separation as well 17. It is also possible that Cse2 may exist both at the SPB during separation and at the kinetochore interacting with chromosomes or microtubules during segregation. This would give some explanation of why Sfi1 would interact with Cse, however, the mechanisms by which the SPB functions are still not fully clear. Despite the support for Cse2 truly interacting with Sfi1, there is still the possibility that it could be a false positive and this needs to be ruled out through further testing. Analysis of Cse2 shows it has a potential leucine zipper domain, which is a common structural motif in proteins that is usually found in DNA binding domains of transcription factors. It is possible that the bzip domain could bring the AD Cse2 fusion to the DNA and close enough to the BDSfi1C construct to allow activation of transcription inappropriately, therefore making the interaction a false positive. The interaction of the C terminus of Sfi1 and Cse2 needs confirmation on media lacking leucine, tryptophan and adenine, with adenine as the second reporter that GAL4 can activate. To further confirm these interactions the bait and prey can be swapped, making BDCse2 and ADSfi1C, and again tested for direct two-hybrid interactions. It would also be interesting to test BDSfi1N or full length Sfi1 for direct two-hybrid interaction with Cse2. To rule out random two-hybrid interaction with any SPB components, numerous different components could be tested directly with Cse2. Finally, the in vivo test of protein interaction, co-immunoprecipitation, could be performed. After more evidence is gathered that supports this interaction, then further analysis can be done to understand why Cse2 would interact with a SPB half-bridge component. It would be interesting to study the phenotype of a double mutant of the C terminus of Sfi1 and Cse2 as that may give insight into the interaction. Further analysis can also be done to understand what may control these interactions, such as phosphorylation. Sfi1 is a highly conserved protein involved in centrosome duplication and function, which are imperative cellular processes that need to be better understood. This study identified one novel interaction between the C terminus of Sfi1 and mediator complex protein Cse2. This interaction provides another interesting link to Cse2 and the SPB, and further analysis of it can potentially advance the knowledge of Sfi1s role in SPB duplication. Peters 13
15 Materials and Methods Yeast strains and plasmids The plasmids used in this study were pobd2 (BD), which contains genes encoding ampicillin resistance, tryptophan and the DNA binding domain of GAL4 (amino acids of GAL4), and poad (AD), which contains genes encoding ampicillin resistance, leucine, and the activating domain of GAL4 (amino acids of GAL4) 18. These are centromeric plasmids 18. The bait and prey genes for this study were cloned in translational frame to the C terminus of the GAL4 region of the appropriate plasmid by generating EcoRI restriction sites at the 5 ends and Sal1 sites at the 3 ends using Polymerase Chain Reaction (PCR) with primers containing the restriction sites 15. The N terminus of Sfi1 included amino acids of the full length protein, the C terminus included amino acids , and the C terminus plus two Sfi1 repeats included amino acids , and these were all cloned into the BD plasmid. Full length Spc29, Spc42, the C terminus of Sfi1, and the C terminus of Sfi1 plus two repeats were cloned into the AD plasmid. Constructs were confirmed using sequencing. Table 1 shows the constructs made for this study. All cloned genes were from the w303 sequence. The Saccharomyces cerevisiae strain used for yeast two-hybrid experiments was pj694, trp1-901 leu2-3,112 ura3-52 his3-200 gal4 Δ gal80 Δ LYS2::GAL1-HIS3 GAL2-ADE2 met2::gal7-lacz 18. The binding domain plasmids were transformed into mating type alpha and grown in synthetic complete media (SC) lacking tryptophan (TRP in figures), and the activating domain plasmids were transformed into mating type A and grown in SC media lacking leucine (LEU in figures) in order to select for the plasmids as described previously 19. Strains containing each plasmid designated in Table 1 will be referred to by their plasmid names. The yeast transformation protocol used was the lithium acetate high efficiency transformation method 19. The mating type A strain was also transformed with an AD S. cerevisiae genome ORF library used for two-hybrid screening 19. Confirming expression of the BDSfi1C construct Western blot analysis was used to determine expression of the BDSfi1C construct. Protein samples were collected as previously described by using the rapid protein preparation method 20. Samples were resolved on a 12% gel by SDS-polyacrylamide gel electrophoresis and Peters 14
16 transferred to a PVDF membrane for analysis. The antibody to detect expression of the GAL4 DNA binding domain fusion proteins was a mouse monoclonal IgG 2a from Santa Cruz Biotechnology (RK5C1) at a concentration of 1µg/mL 5% skim milk emulsion in TBS plus.25% Tween, and the membrane was incubated in the primary antibody overnight at 4 C. The secondary antibody used was a goat anti-mouse IgG antibody (Pierce), and Thermo Scientific Supersignal West Femto Chemilluminescent substrate was used for detection. Directed two-hybrid Directed two-hybrid tests were performed as previously described, employing the HIS3 reporter as a primary test for interaction 9,16. The yeast strain used for this study has three possible reporter genes fused to the GAL promoter, HIS3, ADE2, and lacz. For this study only the HIS3 reporter was used. HIS3, which codes for the enzyme imidazoleglycerol-phosphate dehydratase, involved in step 6 of histidine biosynthesis, was the primary reporter used 16. The drug 3-amino1,2,4-triazole (3-AT, purchased from Sigma-Aldrich, catalog number ) competitively inhibits that enzyme, therefore limiting histidine biosynthesis and growth 16. Using predetermined concentrations of 3-AT in the growth media can select for strong yeast two-hybrid interactions that are able to overcome the inhibitory effects of the drug. To determine the concentration of 3-AT to use for each bait construct, an autoactivation test was performed 16. The bait strain was tested for growth at 30 C for up to 7 days on media lacking tryptophan and histidine with increasing concentrations of 3-AT. The concentration that best eliminated autoactivation of the HIS3 reporter and growth on the media was used in all two-hybrid assays with that bait. The media used for the two-hybrid tests lacked leucine and tryptophan to select for both the bait and prey plasmids, and histidine (HIS in figures) to test for interaction and 3-AT. The media was also supplemented with additional adenine, 80mg/mL, for optimal growth in tests employing the HIS3 reporter gene. Plating for single colonies and replica plating techniques were used. Directed two-hybrid assays went for up to 7 days at 30 C, with observation for interaction beginning at day 2. The directed two-hybrid tests were as follows: BDSfi1N for interaction with ADSpc29 and ADSpc42, BDSfi1C for interaction with ADSfi1C, and BDSfi1C2rep for interaction with ADSfi1C2rep. All directed two hybrid tests, including negative controls, were performed by mating an alpha mating type of the pj694 strain that was Peters 15
17 transformed with a BD plasmid, to an A mating type strain that was transformed with an AD plasmid. BD:AD indicates the resultant diploids. All directed two-hybrid tests were done in duplicate, so that multiple transformants of each plasmid strain were tested. Primarily, one representative example of each test is shown in this paper. Library Screening A screen of the BDSfi1C construct was performed against an AD ORF pool containing approximately all of the ~6000 ORFs in the genome created by PCR from the Fields lab 18. The screen was performed as outlined in the BD Biosciences Clontech Matchmaker Pretransformed Libraries User Manual 14, but with the above mentioned plasmids and media. Large scale liquid mating was used, and positive single colonies that grew at 30 C on media with 3-AT lacking leucine, tryptophan and histidine were selected up to 14 days after plating, beginning on day 2. The positive diploids were streaked onto a master plate lacking leucine, tryptophan and histidine with 1.5mM 3-AT and streaked for single colonies to confirm expression of the HIS3 reporter. The plasmids from each potential positive were isolated from yeast and restriction digests were performed to determine if the plasmids were BD or AD 21. All unique AD plasmids were sent for sequencing to determine what ORF gave the positive interaction. AD plasmids giving interaction were isolated as described previously and retransformed into pj694 to confirm interaction through the directed two-hybrid approach 21. Acknowledgements I would like to thank Dr. Mark Winey for his invaluable mentorship for the last two years, Jennifer Avena for her patience in teaching me far more than protocols, as well as the rest of the Winey lab for their support, insight and assistance. I would like to thank the Fields lab for the ORF library and reagents, the Odorizzi lab for the equipment and plasmids, the Espinosa lab for the equipment, and the Yi lab for the strains. I would like to thank Lynn Wolfe and the Bioscience Undergraduate Skills and Training Program and May Penuela and the Undergraduate Research Opportunities Program for funding this project. Peters 16


18 Figures Figure 1. The Spindle Pole Body. The SPB is composed of a central plaque embedded in the nuclear envelope. There is an outer plaque on the cytoplasmic face, and an inner plaque on the nuclear side that nucleate cytoplasmic and nuclear microtubules, respectively, from gamma tubulin complexes anchored to the plaques. A dense structure called the half-bridge emanates off one side of the central plaque and elongates during SPB duplication. The SPB component Sfi1 localizes to the half-bridge structure with its N terminus next to the SPB and the C terminus at the distal end of the cytoplasmic face of the half-bridge. Figure 2. SPB Duplication. Duplication of the SPB begins in G1 of the cell cycle with the mother SPB. The half-bridge of the SPB elongates to twice its original size and then a satellite of the 4 core SPB components (SPC29, SPC42, Cnm67, and Nud1) is deposited on the distal cytoplasmic tip of the bridge. The satellite then expands to a duplication plaque. Finally the duplication plaque is inserted into the nuclear envelope and assembly of the nuclear SPB components and gamma tubulin complexes occurs. Peters 17

19 A B Figure 3. The yeast two-hybrid system. Part A) The plasmids pobd2 (BD) and poad (AD) are used for yeast two-hybrid 18. The BD plasmid contains the DNA binding domain of GAL4 under the expression of the ADH1 promoter and the selectable marker TRP1, and the AD plasmid contains the transcription activating domain of GAL4 under the expression of the ADH1 promoter and the selectable marker LEU2. The bait and prey genes are fused in frame to the BD and AD domains of these plasmids via general cloning procedures. The plasmids are transformed into separate mating types of a yeast strain containing the GAL1 promoter fused to a reporter gene, HIS3. Part B.) If the proteins interact in a diploid strain containing both plasmids as shown, then the binding domain and activating domain of the GAL4 transcription factor activate transcription of the reporter gene and the cells are able to grow on selective media lacking the selectable markers and the reporter gene. Peters 18
, Spc29, entire gene ADSpc29 Selectable marker Spc42, entire gene ADSpc42 LEU2 Sfi1 C terminus, 801-946 ADSfi1C")
20 Plasmid Insert, Amino Acids used Name of plasmid pobd2 (BD), Sfi1 N terminus, BDSfi1N Selectable marker Sfi1 N terminus, BDSfi1N154 TRP1 Sfi1 N terminus, BDSfi1N175 Sfi1 C terminus, BDSfi1C Sfi1C terminus plus two BDSfi1C2rep repeat domains, poad (AD), Spc29, entire gene ADSpc29 Selectable marker Spc42, entire gene ADSpc42 LEU2 Sfi1 C terminus, ADSfi1C Sfi1 C terminus plus two repeat domains, ADSfi1C2rep Table 1. Constructs generated for two-hybrid experiments. Genes were inserted into the appropriate plasmids by generating restriction sites at the 5 and 3 ends of the gene through polymerase chain reaction. All constructs were verified by DNA sequencing. Figure 4. Confirmed expression of BDSfi1C in pj694. Rapid protein preparations of strains transformed with pobd2 and BDSfi1C (-1 and 2 are separate transformants) were loaded onto a 10% SDS-PAGE gel. Proteins were detected using anti-gal4 DBD (Santa Cruz Biotechnology) and anti-mouse HRP conjugated secondary antibody (Pierce). Blots were reblotted with a mouse anti-glucose-6-phosphate dehydrogenase primary loading control. Peters 19


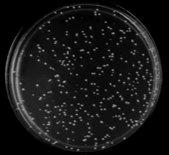






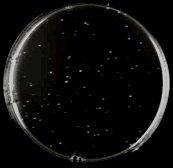

21 A BDSfi1N BDSfi1N154 BDSfi1N175 pobd2 SC-TRP Control SC-TRP- HIS 0mM 3- AT SC-TRP- HIS +5mM 3-AT SC-TRP- HIS +50mM 3-AT SC-TRP- HIS +100mM 3-AT Peters 20


22 B SC-TRP Control BDSfi1C BDSfi1C2rep pobd2 SC-TRP-HIS +0mM 3-AT SC-TRP-HIS +1.5mM 3- AT SC-TRP-HIS +5mM 3-AT Figure 5. Autoactivation of bait constructs. The bait constructs were tested for autoactivation of the HIS3 reporter gene by testing them for growth on media lacking TRP and HIS with increasing concentrations of the drug 3-AT, and comparing this to growth on media lacking TRP, which selects for the BD plasmid. The name of the construct being tested is listed at the top of each column, and media and 3-AT concentration is listed at the start of each row. Part A) shows the autoactivation of the full length N terminus of Sfi1, which required 50mM 3-AT to control autoactivation, and the two truncations, N154 and N175, which required 5mM and 50mM concentrations of 3-AT respectively. The empty pobd2 plasmid was also tested and showed no autoactivation. Part B) shows the autoactivation test of the C terminus of Sfi1 and the C terminus plus two repeat domains. Both of these showed no significant level of autoactivation so the lowest concentration of 3-AT, 1.5mM, was used. Peters 21










23 A SC- LEU- TRP Diploid selective SC- LEU- TRP- HIS +50mM 3-AT BD:AD B SC-LEU- TRP Diploid Selective BDSfi1N: AD BD:ADSpc29 BD:ADSpc42 BDSfi1N: ADSpc29 BD: AD BDSfi1C: AD BD: ADSfi1C BDSfi1C: ADSfi1C BDSfi1N: ADSpc42 SC-LEU- TRP-HIS +1.5mM 3-AT C BD: AD BDSfi1C2rep: AD BD: ADSfi1C2rep BDSfi1C2rep: ADSfi1C2rep SC-LEU- TRP Diploid Selective SC-LEU- TRP-HIS +1.5mM 3-AT Peters 22
24 Figure 6. Directed two-hybrid tests of the N and C termini of Sfi1. Part A) the BDSfi1N and ADSpc29 and ADSpc42 constructs were transformed into separate mating types of pj694 and diploids were selected for (diploids indicated by colon) and tested for interaction through yeast two-hybrid. Negative controls were BDSfi1N:AD, BD:ADSpc29 and BD:ADSpc42, and BD:AD. All strains grew normally on diploid selective media. None of the negative controls or BDSfi1N: ADSpc42 grew on diploid selective media, but BDSfi1N:AD did show some growth. BDSfi1N: ADSpc29 showed a small amount of growth on media selecting for diploids and lacking HIS, but at levels equal to or less than the negative control BDSfi1N:AD. Part B) In part B the BDSfi1C and ADSfi1C constructs were transformed into separate mating types of pj694 and tested for two-hybrid interactions. Negative controls were BDSfi1C: AD, BD:ADSfi1C, and BD:AD. Normal growth was seen for all combinations on media selecting for diploids. No growth was seen for any negative controls or tests for interaction on media selecting for diploids and lacking HIS. Part C) The BDSfi1C2rep and ADSfi1C2 rep constructs were transformed into separate mating types of pj694 and tested for twohybrid interactions. The negative controls were BDSfi1C2rep:AD, BD:ADSfi1C2rep, and BD:AD. Normal growth was seen on media selecting for diploids, and no growth was seen for the negative controls or test for two-hybrid interaction on plates selecting for diploids and lacking HIS. Peters 23











25 B BD: AD BDSfi1C: AD BD: ADpositive24 BD: ADpositive32 BDSfi1C: ADpositive24 BDSfi1C: ADpositive32 SC- LEU- TRP Diploid Selective SC- LEU- TRP- HIS +1.5mM 3-AT Figure 7. Confirmation of true positive from a yeast two-hybrid screen of BDSfi1C. Part A) All four positives, 17, 24, 32, and 45, that came up in a yeast two-hybrid screen for interaction with BDSfi1C were streaked out for single colonies then a single colony for each positive was patched on to a master positive plate lacking LEU, TRP and HIS with 1.5mM 3AT to confirm expression of the HIS3 reporter. Part B) True positives 24 and 32 were isolated and retransformed in to pj694 mating type A, and tested for direct two-hybrid interaction with BDSfi1C. Negative controls performed were empty pobd2 to empty poad, BDSfi1C to empty poad, empty pobd2 to ADpositive24 and empty pobd2 to ADpositive32. All combinations, including tests for interaction, showed normal growth on diploid selective media. None of the negative controls grew on media selecting for diploids and lacking HIS. Both positive tests were able to grow on media lacking HIS also, with less colonies observed than on the diploid selective plates but more than the negative controls. Peters 24
26 References 1. Lüders, Jens, and Tim Stearns (2007). "Microtubule-organizing centres: a reevaluation." Nature Reviews Molecular Cell Biology 8.2: Jones, Michele H. and Winey, Mark (2006). Centrosome duplication: Is asymmetry the clue?. Current Biology: 16(18): R Nigg, E.rich A., and Jordan W. Raff (2009). "Centrioles, centrosomes, and cilia in health and disease." Cell 139.4: Ganem, N.eil J., Susana A. Godinho, and David Pellman(2009). "A mechanism linking extra centrosomes to chromosomal instability." Nature : Jaspersen, S.L., and Winey, M. (2004). The budding yeast spindle pole body: Structure, duplication, and function. Annu. Rev. Cell Dev. Biol. 20, Kilmartin JV. (2003). Sfi1p has conserved centrin-binding sites and an essential function in budding yeast spindle pole body duplication. J. Cell Biol. 162: V.E. Anderson, J. Prudden, S. Prochnik, T.H. Giddings Jr., K.G. Hardwick (2007). Novel sfi1 alleles uncover additional functions for Sfi1p in bi-polar spindle assembly and function. Molecular Biology of the Cell., 18, pp Elliott, Sarah, et al (1999). "Spc29p is a component of the Spc110p subcomplex and is essential for spindle pole body duplication." Proceedings of the National Academy of Sciences 96.11: Fields, S. & Song, O. (1989). A novel genetic system to detect protein-protein interactions. Nature (London) 340, Li, S., Sandercock, A. M., Conduit, P., Robinson, C. V., Williams, R. L. and Kilmartin, J. V. (2006). Structural role of Sfi1 p-centrin filaments in budding yeast spindle pole body duplication. J. Cell Biol. 173, Winey, M. and Bloom, K (2012). Mitotic Spindle Form and Function. Sprague, G., ed., Yeast Book, Genetics Society of America, Bethesda. 12. Costanzo, Michael, et al (2010). "The genetic landscape of a cell." Science Signalling : Ruden, D. M. (1992) Activating regions of yeast transcription factors must have both acidic and hydrophobic amino acids. Chromosoma 101: Clontech (2007). Matchmaker Pretransformed Libraries User Manual. Peters 25
27 15. Finney, M. (2001). Molecular Cloning of PCR Products. Curr. Protoc. Mol. Biol. 56: Gietz, R. Daniel, et al (1997). "Identification of proteins that interact with a protein of interest: applications of the yeast two-hybrid system." Molecular and cellular biochemistry 172.1: Strawn, Lisa A., and Heather L. True (2006). "Deletion of RNQ1 gene reveals novel functional relationship between divergently transcribed Bik1p/CLIP-170 and Sfi1p in spindle pole body separation." Current genetics 50.6: Fields, Stan. Protocols. Laboratory of Stan Fields. University of Washington. Web. 19. Schiestl, Robert H., and R. Daniel Gietz (1996). "Transforming Yeast with DNA. Methods in Molecular and Cellular Biology 16.5: Horvath, Anton, and Howard Riezman (1994). "Rapid protein extraction from Saccharomyces cerevisiae." Yeast 10.10: Robzyk, Kenneth, and Yona Kassir (1992). "A simple and highly efficient procedure for rescuing autonomous plasmids from yeast." Nucleic acids research 20.14: Hengen, Paul H (1997). "Methods and reagents: false positives from the yeast two-hybrid system." Trends in biochemical sciences 22.1: Tomomori-Sato C, et al. (2004) A mammalian mediator subunit that shares properties with Saccharomyces cerevisiae mediator subunit Cse2. J Biol Chem 279(7): Xiao, Z. et al. (1993). "CSE1 and CSE2, two new genes required for accurate mitotic chromosome segregation in Saccharomyces cerevisiae." Molecular and cellular biology 13.8: Collins SR, et al. (2007) Functional dissection of protein complexes involved in yeast chromosome biology using a genetic interaction map. Nature 446(7137): Lee L, et al. (2000) Positioning of the mitotic spindle by a cortical-microtubule capture mechanism. Science 287(5461): Dorer RK, et al. (2005) A small-molecule inhibitor of Mps1 blocks the spindlecheckpoint response to a lack of tension on mitotic chromosomes. Curr Biol 15(11): Thomas JH and Botstein D (1986) A gene required for the separation of chromosomes on the spindle apparatus in yeast. Cell 44(1): Peters 26
2. Yeast two-hybrid system
 2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Three different fusions led to three basic ideas: 1) If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes
 If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes") Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
Reading Assignments. A. Systems of Cell Division. Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces cerevisiae.
 Genetics: Published Articles Ahead of Print, published on July 30, 2012 as 10.1534/genetics.112.143818 A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces
Genetics: Published Articles Ahead of Print, published on July 30, 2012 as 10.1534/genetics.112.143818 A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces
Segregation and the Principle of independent assortment
 . Segregation and the Principle of independent assortment The Principle of Independent Assortment describes how different genes independently separate from one another when reproductive cells develop.
. Segregation and the Principle of independent assortment The Principle of Independent Assortment describes how different genes independently separate from one another when reproductive cells develop.
Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis.
 Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis. The role of kinases and cyclin in the regulation of the cell cycle.
Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis. The role of kinases and cyclin in the regulation of the cell cycle.
The Cell Cycle. Chapter 12
 The Cell Cycle Chapter 12 Why are cells small? As cells get bigger they don t work as well WHY? Difficulties Larger Cells Have: More demands on its DNA Less efficient in moving nutrients/waste across its
The Cell Cycle Chapter 12 Why are cells small? As cells get bigger they don t work as well WHY? Difficulties Larger Cells Have: More demands on its DNA Less efficient in moving nutrients/waste across its
Name Chapter 10: Chromosomes, Mitosis, and Meiosis Mrs. Laux Take home test #7 DUE: MONDAY, NOVEMBER 16, 2009 MULTIPLE CHOICE QUESTIONS
 MULTIPLE CHOICE QUESTIONS 1. A bacterial chromosome consists of: A. a linear DNA molecule many times larger than the cell. B. a circular DNA molecule many times larger than the cell. C. a circular DNA
MULTIPLE CHOICE QUESTIONS 1. A bacterial chromosome consists of: A. a linear DNA molecule many times larger than the cell. B. a circular DNA molecule many times larger than the cell. C. a circular DNA
Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Division Read Chapter 19 pages 651-663 663 only (Benefits of Sex & Meiosis sections these are in Chapter
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Division Read Chapter 19 pages 651-663 663 only (Benefits of Sex & Meiosis sections these are in Chapter
Honors Biology Test Chapter 8 Mitosis and Meiosis
 Honors Biology Test Chapter 8 Mitosis and Meiosis 1. In mitosis, if a parent cell has 16 chromosomes, each daughter cell will have how many chromosomes? a. 64 b. 32 c. 16 d. 8 e. 4 2. Chromatids that are
Honors Biology Test Chapter 8 Mitosis and Meiosis 1. In mitosis, if a parent cell has 16 chromosomes, each daughter cell will have how many chromosomes? a. 64 b. 32 c. 16 d. 8 e. 4 2. Chromatids that are
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Meiosis. Bởi: OpenStaxCollege
 Meiosis Bởi: OpenStaxCollege Sexual reproduction requires fertilization, a union of two cells from two individual organisms. If those two cells each contain one set of chromosomes, then the resulting cell
Meiosis Bởi: OpenStaxCollege Sexual reproduction requires fertilization, a union of two cells from two individual organisms. If those two cells each contain one set of chromosomes, then the resulting cell
ACCELERATE ITS BIOCHEMICAL PROCESSES WHICH WERE SLOWED DOWN BY MITOSIS. THE LENGTH OF THE G1 PHASE CREATES THE DIFFERENCE BETWEEN FAST DIVIDING
 CHAPTER 1: OVERVIEW OF THE CELL CYCLE THE THREE STAGES OF INTERPHASE: INTERPHASE BEFORE A CELL CAN ENTER CELL DIVISION, IT NEEDS TO PREPARE ITSELF BY REPLICATING ITS GENETIC INFORMATION AND ALL OF THE
CHAPTER 1: OVERVIEW OF THE CELL CYCLE THE THREE STAGES OF INTERPHASE: INTERPHASE BEFORE A CELL CAN ENTER CELL DIVISION, IT NEEDS TO PREPARE ITSELF BY REPLICATING ITS GENETIC INFORMATION AND ALL OF THE
2. Which of the following are NOT prokaryotes? A) eubacteria B) archaea C) viruses D) ancient bacteria
 eubacteria B) archaea C) viruses D) ancient bacteria") 1. Which of the following statements is FALSE? A) Errors in chromosome separation are rarely a problem for an organism. B) Errors in chromosome separation can result in a miscarriage. C) Errors in chromosome
1. Which of the following statements is FALSE? A) Errors in chromosome separation are rarely a problem for an organism. B) Errors in chromosome separation can result in a miscarriage. C) Errors in chromosome
Biology: Life on Earth
 Biology: Life on Earth Eighth Edition Lecture for Chapter 11 The Continuity of Life: Cellular Reproduction Cellular Reproduction Intracellular activity between one cell division to the next is the cell
Biology: Life on Earth Eighth Edition Lecture for Chapter 11 The Continuity of Life: Cellular Reproduction Cellular Reproduction Intracellular activity between one cell division to the next is the cell
Chapter 2 Cells and Cell Division
 Chapter 2 Cells and Cell Division MULTIPLE CHOICE 1. The process of meiosis results in: A. the production of four identical cells B. no change in chromosome number from parental cells C. a doubling of
Chapter 2 Cells and Cell Division MULTIPLE CHOICE 1. The process of meiosis results in: A. the production of four identical cells B. no change in chromosome number from parental cells C. a doubling of
E. Incorrect! At telophase II, cells are nearly completed with meiosis, with no cross-over.
 OAT Biology - Problem Drill 06: Mitosis and Meiosis Question No. 1 of 10 1. During meiosis, cross-over between homologous chromosomes occurs at the end of. Question #01 (A) Anaphase II (B) Metaphase I
OAT Biology - Problem Drill 06: Mitosis and Meiosis Question No. 1 of 10 1. During meiosis, cross-over between homologous chromosomes occurs at the end of. Question #01 (A) Anaphase II (B) Metaphase I
Introduction to Molecular and Cell Biology
 Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the molecular basis of disease? What
Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the molecular basis of disease? What
A Few Terms: When and where do you want your cells to divide?
 Today: - Lab 4 Debrief - Mitosis - Lunch -Meiosis Other: Blood Drive Today! TIME: 11:00am 1:00pm + 2:00pm 5:00pm PLACE: Baxter Events Center Thinking About Mitosis When and where do you want your cells
Today: - Lab 4 Debrief - Mitosis - Lunch -Meiosis Other: Blood Drive Today! TIME: 11:00am 1:00pm + 2:00pm 5:00pm PLACE: Baxter Events Center Thinking About Mitosis When and where do you want your cells
2012 Univ Aguilera Lecture. Introduction to Molecular and Cell Biology
 2012 Univ. 1301 Aguilera Lecture Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the
2012 Univ. 1301 Aguilera Lecture Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the
This is DUE: Come prepared to share your findings with your group.
 Biology 160 NAME: Reading Guide 11: Population Dynamics, Humans, Part I This is DUE: Come prepared to share your findings with your group. *As before, please turn in only the Critical Thinking questions
Biology 160 NAME: Reading Guide 11: Population Dynamics, Humans, Part I This is DUE: Come prepared to share your findings with your group. *As before, please turn in only the Critical Thinking questions
Meiosis * OpenStax. This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0.
 OpenStax-CNX module: m45466 1 Meiosis * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section, you will be able to: Abstract
OpenStax-CNX module: m45466 1 Meiosis * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section, you will be able to: Abstract
Unit 6 Test: The Cell Cycle
 Name Date Class Mrs. Knight Biology EHS Unit 6 Test: The Cell Cycle 1. What are the four main stages of the cell cycle (correct order)? A. G 1, S, G 0, M C. G 2, S, G 1, M B. G 1, S, G 2, M D. M, G 2,
Name Date Class Mrs. Knight Biology EHS Unit 6 Test: The Cell Cycle 1. What are the four main stages of the cell cycle (correct order)? A. G 1, S, G 0, M C. G 2, S, G 1, M B. G 1, S, G 2, M D. M, G 2,
Mitosis & Meiosis Practice Questions
 Name: Date: 1. The diagram shown represents a cell that will undergo mitosis. Which diagrams below best illustrate the nuclei of the daughter cells that result from a normal mitotic cell division of the
Name: Date: 1. The diagram shown represents a cell that will undergo mitosis. Which diagrams below best illustrate the nuclei of the daughter cells that result from a normal mitotic cell division of the
Analysis and modelling of protein interaction networks
 Analysis and modelling of protein interaction networks - A study of the two-hybrid experiment Karin Stibius Jensen Analysis and modelling of protein interaction networks - A study of the two-hybrid experiment
Analysis and modelling of protein interaction networks - A study of the two-hybrid experiment Karin Stibius Jensen Analysis and modelling of protein interaction networks - A study of the two-hybrid experiment
Cell Division. Mitosis 11/8/2016
 Cell division consists of two phases, nuclear division followed by cytokinesis. Nuclear division divides the genetic material in the nucleus, while cytokinesis divides the cytoplasm. There are two kinds
Cell division consists of two phases, nuclear division followed by cytokinesis. Nuclear division divides the genetic material in the nucleus, while cytokinesis divides the cytoplasm. There are two kinds
1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine.
 Protein Synthesis & Mutations RNA 1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine. RNA Contains: 1. Adenine 2.
Protein Synthesis & Mutations RNA 1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine. RNA Contains: 1. Adenine 2.
Cell Division. Mitosis
 Cell division consists of two phases, nuclear division followed by cytokinesis. Nuclear division divides the genetic material in the nucleus, while cytokinesis divides the cytoplasm. There are two kinds
Cell division consists of two phases, nuclear division followed by cytokinesis. Nuclear division divides the genetic material in the nucleus, while cytokinesis divides the cytoplasm. There are two kinds
Essential Knowledge: In eukaryotes, heritable information is passed to the next generation via processes that include the cell cycle and mitosis OR
 Essential Knowledge: In eukaryotes, heritable information is passed to the next generation via processes that include the cell cycle and mitosis OR meiosis plus fertilization Objective: You will be able
Essential Knowledge: In eukaryotes, heritable information is passed to the next generation via processes that include the cell cycle and mitosis OR meiosis plus fertilization Objective: You will be able
Chromosome Chr Duplica Duplic t a ion Pixley
 Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
The Microscopic Observation of Mitosis in Plant and Animal Cells
 The Microscopic Observation of Mitosis in Plant and Animal Cells Prelab Assignment Before coming to lab, read carefully the introduction and the procedures for each part of the experiment, and then answer
The Microscopic Observation of Mitosis in Plant and Animal Cells Prelab Assignment Before coming to lab, read carefully the introduction and the procedures for each part of the experiment, and then answer
Genetics Essentials Concepts and Connections 3rd Edition by Benjamin A Pierce Test Bank
 Genetics Essentials Concepts and Connections 3rd Edition by Benjamin A Pierce Test Bank Which of the following statements is FALSE? A) Errors in chromosome separation are rarely a problem for an organism.
Genetics Essentials Concepts and Connections 3rd Edition by Benjamin A Pierce Test Bank Which of the following statements is FALSE? A) Errors in chromosome separation are rarely a problem for an organism.
Spc29p is a component of the Spc110p subcomplex and is essential for spindle pole body duplication
 Proc. Natl. Acad. Sci. USA Vol. 96, pp. 6205 6210, May 1999 Cell Biology Spc29p is a component of the Spc110p subcomplex and is essential for spindle pole body duplication SARAH ELLIOTT*, MICHAEL KNOP*,
Proc. Natl. Acad. Sci. USA Vol. 96, pp. 6205 6210, May 1999 Cell Biology Spc29p is a component of the Spc110p subcomplex and is essential for spindle pole body duplication SARAH ELLIOTT*, MICHAEL KNOP*,
THE CELL CYCLE & MITOSIS. Asexual Reproduction: Production of genetically identical offspring from a single parent.
 THE CELL CYCLE & MITOSIS Asexual Reproduction: Production of genetically identical offspring from a single parent. Sexual Reproduction: The fusion of two separate parent cells that produce offspring with
THE CELL CYCLE & MITOSIS Asexual Reproduction: Production of genetically identical offspring from a single parent. Sexual Reproduction: The fusion of two separate parent cells that produce offspring with
Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells
 Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells Understand how a simple biochemical oscillator can drive the
Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells Understand how a simple biochemical oscillator can drive the
CELL CYCLE UNIT GUIDE- Due January 19, 2016
 CELL CYCLE UNIT GUIDE- Due January 19, 2016 Monday Tuesday Wednesday Thursday Friday January 4- No School 5-Cell Cycle/Mitosis 6-Cell Cycle/ Mitosis 7-Mitosis 8-Meiosis Reading Check Quiz #1 sections 5.1-5.5
CELL CYCLE UNIT GUIDE- Due January 19, 2016 Monday Tuesday Wednesday Thursday Friday January 4- No School 5-Cell Cycle/Mitosis 6-Cell Cycle/ Mitosis 7-Mitosis 8-Meiosis Reading Check Quiz #1 sections 5.1-5.5
Ladies and Gentlemen.. The King of Rock and Roll
 Ladies and Gentlemen.. The King of Rock and Roll Learning Objectives: The student is able to construct an explanation, using visual representations or narratives, as to how DNA in chromosomes is transmitted
Ladies and Gentlemen.. The King of Rock and Roll Learning Objectives: The student is able to construct an explanation, using visual representations or narratives, as to how DNA in chromosomes is transmitted
Meiosis, Sexual Reproduction, & Genetic Variability
 Meiosis, Sexual Reproduction, & Genetic Variability Teachers Guide NARRATION FOR MEIOSIS, SEXUAL REPRODUCTION, AND GENETIC VARIABILITY Since the members of no species, even California redwoods or giant
Meiosis, Sexual Reproduction, & Genetic Variability Teachers Guide NARRATION FOR MEIOSIS, SEXUAL REPRODUCTION, AND GENETIC VARIABILITY Since the members of no species, even California redwoods or giant
AP Biology - Cell cycle / division
 AP Biology - Cell cycle / division Quiz Directions 1. During which stage does DNA replication occur? A. Prophase B. Metaphase C. Anaphase D. none of these 2. At what phase in the cell cycle does DNA replication
AP Biology - Cell cycle / division Quiz Directions 1. During which stage does DNA replication occur? A. Prophase B. Metaphase C. Anaphase D. none of these 2. At what phase in the cell cycle does DNA replication
Topic 8 Mitosis & Meiosis Ch.12 & 13. The Eukaryotic Genome. The Eukaryotic Genome. The Eukaryotic Genome
 Topic 8 Mitosis & Meiosis Ch.12 & 13 The Eukaryotic Genome pp. 244-245,268-269 Genome All of the genes in a cell. Eukaryotic cells contain their DNA in long linear pieces. In prokaryotic cells, there is
Topic 8 Mitosis & Meiosis Ch.12 & 13 The Eukaryotic Genome pp. 244-245,268-269 Genome All of the genes in a cell. Eukaryotic cells contain their DNA in long linear pieces. In prokaryotic cells, there is
Human Biology Chapter 13.4: Meiosis and Genetic Variation
 OpenStax-CNX module: m58013 1 Human Biology Chapter 13.4: Meiosis and Genetic Variation Willy Cushwa Based on Meiosis by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons
OpenStax-CNX module: m58013 1 Human Biology Chapter 13.4: Meiosis and Genetic Variation Willy Cushwa Based on Meiosis by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons
Chapter 2: Chromosomes and cellular reproduction
 Chapter 2: Chromosomes and cellular reproduction I. Contrast between prokaryotes and eukaryotes. See Figure 2.1 Nucleus absent Small diameter 1 to 10 µm Genome usually 1 circular molecule Small genome;
Chapter 2: Chromosomes and cellular reproduction I. Contrast between prokaryotes and eukaryotes. See Figure 2.1 Nucleus absent Small diameter 1 to 10 µm Genome usually 1 circular molecule Small genome;
Mitosis vs Meiosis. Mitosis and Meiosis -- Internet Tutorial
 Mitosis and Meiosis -- Internet Tutorial In this internet lesson, you will review the steps of mitosis and meiosis and view video simulations of cell division. Mitosis: An Interactive Animation (http://www.cellsalive.com/mitosis.htm)
Mitosis and Meiosis -- Internet Tutorial In this internet lesson, you will review the steps of mitosis and meiosis and view video simulations of cell division. Mitosis: An Interactive Animation (http://www.cellsalive.com/mitosis.htm)
A diploid somatic cell from a rat has a total of 42 chromosomes (2n = 42). As in humans, sex chromosomes determine sex: XX in females and XY in males.
. As in humans, sex chromosomes determine sex: XX in females and XY in males.") Multiple Choice Use the following information for questions 1-3. A diploid somatic cell from a rat has a total of 42 chromosomes (2n = 42). As in humans, sex chromosomes determine sex: XX in females and
Multiple Choice Use the following information for questions 1-3. A diploid somatic cell from a rat has a total of 42 chromosomes (2n = 42). As in humans, sex chromosomes determine sex: XX in females and
AP Biology Fall Semester Set 1
 1. During which stage does DNA replication occur? A. Prophase B. Metaphase C. Anaphase D. none of these 2. At what phase in the cell cycle does DNA replication occur? A. G1 B. S C. G2 D. M 3. Which of
1. During which stage does DNA replication occur? A. Prophase B. Metaphase C. Anaphase D. none of these 2. At what phase in the cell cycle does DNA replication occur? A. G1 B. S C. G2 D. M 3. Which of
Cell Structure and Function. The Development of Cell Theory
 Cell Structure and Function The Development of Cell Theory Hooke's original microscope, that he used at Oxford University, in 1665. Robert Hooke first examined a thin piece of cork (see below), and coined
Cell Structure and Function The Development of Cell Theory Hooke's original microscope, that he used at Oxford University, in 1665. Robert Hooke first examined a thin piece of cork (see below), and coined
10 CELL DIVISION AND MITOSIS
 10 CELL DIVISION AND MITOSIS Chapter Outline Why It Matters 10.1 THE CYCLE OF CELL GROWTH AND DIVISION: OVERVIEW The products of mitosis are genetic duplicates of the dividing cell Chromosomes are the
10 CELL DIVISION AND MITOSIS Chapter Outline Why It Matters 10.1 THE CYCLE OF CELL GROWTH AND DIVISION: OVERVIEW The products of mitosis are genetic duplicates of the dividing cell Chromosomes are the
Now starts the fun stuff Cell structure and function
 Now starts the fun stuff Cell structure and function Cell Theory The three statements of the cell theory are: All organisms are composed of one or more cells and the processes of life occur in these cells.
Now starts the fun stuff Cell structure and function Cell Theory The three statements of the cell theory are: All organisms are composed of one or more cells and the processes of life occur in these cells.
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
2:1 Chromosomes DNA Genes Chromatin Chromosomes CHROMATIN: nuclear material in non-dividing cell, composed of DNA/protein in thin uncoiled strands
 Human Heredity Chapter 2 Chromosomes, Mitosis, and Meiosis 2:1 Chromosomes DNA Genes Chromatin Chromosomes CHROMATIN: nuclear material in non-dividing cell, composed of DNA/protein in thin uncoiled strands
Human Heredity Chapter 2 Chromosomes, Mitosis, and Meiosis 2:1 Chromosomes DNA Genes Chromatin Chromosomes CHROMATIN: nuclear material in non-dividing cell, composed of DNA/protein in thin uncoiled strands
Chapter 13 Meiosis and Sexual Reproduction
 Biology 110 Sec. 11 J. Greg Doheny Chapter 13 Meiosis and Sexual Reproduction Quiz Questions: 1. What word do you use to describe a chromosome or gene allele that we inherit from our Mother? From our Father?
Biology 110 Sec. 11 J. Greg Doheny Chapter 13 Meiosis and Sexual Reproduction Quiz Questions: 1. What word do you use to describe a chromosome or gene allele that we inherit from our Mother? From our Father?
Mitosis and Meiosis for AP Biology
 Mitosis and Meiosis for AP Biology by Mark Anestis Practice problems for these concepts can be found at : Cell Division Review Questions for AP Biology Mitosis During mitosis, the fourth stage of the cell
Mitosis and Meiosis for AP Biology by Mark Anestis Practice problems for these concepts can be found at : Cell Division Review Questions for AP Biology Mitosis During mitosis, the fourth stage of the cell
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
Meiosis and Sexual Reproduction. Chapter 9
 Meiosis and Sexual Reproduction Chapter 9 9.1 Genes and Alleles Genes Sequences of DNA that encode heritable traits Alleles Slightly different forms of the same gene Each specifies a different version
Meiosis and Sexual Reproduction Chapter 9 9.1 Genes and Alleles Genes Sequences of DNA that encode heritable traits Alleles Slightly different forms of the same gene Each specifies a different version
BIOLOGY 111. CHAPTER 5: Chromosomes and Inheritance
 BIOLOGY 111 CHAPTER 5: Chromosomes and Inheritance Chromosomes and Inheritance Learning Outcomes 5.1 Differentiate between sexual and asexual reproduction in terms of the genetic variation of the offspring.
BIOLOGY 111 CHAPTER 5: Chromosomes and Inheritance Chromosomes and Inheritance Learning Outcomes 5.1 Differentiate between sexual and asexual reproduction in terms of the genetic variation of the offspring.
Describe the process of cell division in prokaryotic cells. The Cell Cycle
 The Cell Cycle Objective # 1 In this topic we will examine the cell cycle, the series of changes that a cell goes through from one division to the next. We will pay particular attention to how the genetic
The Cell Cycle Objective # 1 In this topic we will examine the cell cycle, the series of changes that a cell goes through from one division to the next. We will pay particular attention to how the genetic
Chapter 8 Lectures by Gregory Ahearn University of North Florida
 Chapter 8 The Continuity of Life: How Cells Reproduce Lectures by Gregory Ahearn University of North Florida Copyright 2009 Pearson Education, Inc. 8.1 Why Do Cells Divide? Cells reproduce by cell division.
Chapter 8 The Continuity of Life: How Cells Reproduce Lectures by Gregory Ahearn University of North Florida Copyright 2009 Pearson Education, Inc. 8.1 Why Do Cells Divide? Cells reproduce by cell division.
Nuclear Functional Organization
 Lecture #4 The Cell as a Machine Nuclear Functional Organization Background readings from Chapters 4 of Alberts et al. Molecular Biology of the Cell (4 th Edition) Description of Functions by Biosystems
Lecture #4 The Cell as a Machine Nuclear Functional Organization Background readings from Chapters 4 of Alberts et al. Molecular Biology of the Cell (4 th Edition) Description of Functions by Biosystems
11.1 The Process of Meiosis
 OpenStax-CNX module: m52675 1 11.1 The Process of Meiosis Shannon McDermott Based on The Process of Meiosis by OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons
OpenStax-CNX module: m52675 1 11.1 The Process of Meiosis Shannon McDermott Based on The Process of Meiosis by OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons
Chapter 8. Introduction. Introduction. The Cellular Basis of Reproduction and Inheritance. Cancer cells. In a healthy body, cell division allows for
 Chapter 8 The Cellular Basis of Reproduction and Inheritance PowerPoint Lectures for Campbell Biology: Concepts & Connections, Seventh Edition Reece, Taylor, Simon, and Dickey Lecture by Edward J. Zalisko
Chapter 8 The Cellular Basis of Reproduction and Inheritance PowerPoint Lectures for Campbell Biology: Concepts & Connections, Seventh Edition Reece, Taylor, Simon, and Dickey Lecture by Edward J. Zalisko
Name: Date: Hour: Unit Four: Cell Cycle, Mitosis and Meiosis. Monomer Polymer Example Drawing Function in a cell DNA
 Unit Four: Cell Cycle, Mitosis and Meiosis I. Concept Review A. Why is carbon often called the building block of life? B. List the four major macromolecules. C. Complete the chart below. Monomer Polymer
Unit Four: Cell Cycle, Mitosis and Meiosis I. Concept Review A. Why is carbon often called the building block of life? B. List the four major macromolecules. C. Complete the chart below. Monomer Polymer
Greenfield Sluder and Dannel McCollum *
 CAROL TRENT Change Password Change User Info CiteTrack Alerts Access Rights Subscription Help Sign Out MOLECULAR BIOLOGY: The Mad Ways of Meiosis Greenfield Sluder and Dannel McCollum * Many organisms,
CAROL TRENT Change Password Change User Info CiteTrack Alerts Access Rights Subscription Help Sign Out MOLECULAR BIOLOGY: The Mad Ways of Meiosis Greenfield Sluder and Dannel McCollum * Many organisms,
Fertilization of sperm and egg produces offspring
 In sexual reproduction Fertilization of sperm and egg produces offspring In asexual reproduction Offspring are produced by a single parent, without the participation of sperm and egg CONNECTIONS BETWEEN
In sexual reproduction Fertilization of sperm and egg produces offspring In asexual reproduction Offspring are produced by a single parent, without the participation of sperm and egg CONNECTIONS BETWEEN
BIOLOGY - CLUTCH CH.13 - MEIOSIS.
 !! www.clutchprep.com CONCEPT: SEXUAL REPRODUCTION Meiosis is a special type of cell division that occurs as part of the sexual life cycle of eukaryotes Sexual reproduction parents donate a unique mixture
!! www.clutchprep.com CONCEPT: SEXUAL REPRODUCTION Meiosis is a special type of cell division that occurs as part of the sexual life cycle of eukaryotes Sexual reproduction parents donate a unique mixture
Meiosis and Sexual Life Cycles
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 13 Meiosis and Sexual Life Cycles
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 13 Meiosis and Sexual Life Cycles
1- Below is a list of cell cycle phases matched with specific processes. Choose the correct pairing:
 Name: NetID: Exam 4 - Version 2 November 13, 2018 Dr. A. Pimentel Instructions: 1- Select the BEST answer for each question 2- Use pencil to mark your responses in the answer sheet. 3- You can mark your
Name: NetID: Exam 4 - Version 2 November 13, 2018 Dr. A. Pimentel Instructions: 1- Select the BEST answer for each question 2- Use pencil to mark your responses in the answer sheet. 3- You can mark your
Meiosis and Sexual Life Cycles
 CAMPBELL BIOLOGY IN FOCUS URRY CAIN WASSERMAN MINORSKY REECE 10 Meiosis and Sexual Life Cycles Lecture Presentations by Kathleen Fitzpatrick and Nicole Tunbridge, Simon Fraser University SECOND EDITION
CAMPBELL BIOLOGY IN FOCUS URRY CAIN WASSERMAN MINORSKY REECE 10 Meiosis and Sexual Life Cycles Lecture Presentations by Kathleen Fitzpatrick and Nicole Tunbridge, Simon Fraser University SECOND EDITION
Chapter 11: The Continuity of Life: Cellular Reproduction. What is Cellular Reproduction?
 Chapter 11: The Continuity of Life: Cellular Reproduction What is Cellular Reproduction? Answer: The division of a parent cell into two daughter cells Requirements of Each Daughter Cell: 1) Necessary genomic
Chapter 11: The Continuity of Life: Cellular Reproduction What is Cellular Reproduction? Answer: The division of a parent cell into two daughter cells Requirements of Each Daughter Cell: 1) Necessary genomic
Transport between cytosol and nucleus
 of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
Prof. Fahd M. Nasr. Lebanese university Faculty of sciences I Department of Natural Sciences.
 Prof. Fahd M. Nasr Lebanese university Faculty of sciences I Department of Natural Sciences fnasr@ul.edu.lb B3206 Microbial Genetics Eukaryotic M. G. The yeast Saccharomyces cerevisiae as a genetic model
Prof. Fahd M. Nasr Lebanese university Faculty of sciences I Department of Natural Sciences fnasr@ul.edu.lb B3206 Microbial Genetics Eukaryotic M. G. The yeast Saccharomyces cerevisiae as a genetic model
Mitosis and Meiosis Cell growth and division
 LIMITS TO CELL GROWTH Mitosis and Meiosis Cell growth and division The larger the cell, the more trouble the cell has moving nutrients and waste across the cell membrane. LIMITS TO CELL GROWTH 1. DNA/information
LIMITS TO CELL GROWTH Mitosis and Meiosis Cell growth and division The larger the cell, the more trouble the cell has moving nutrients and waste across the cell membrane. LIMITS TO CELL GROWTH 1. DNA/information
Meiosis and Sexual Reproduction
 Meiosis and Sexual Reproduction Asexual Reproduction Single parent produces offspring All offspring are genetically identical to one another and to parent Produces identical somatic (body) cells Sexual
Meiosis and Sexual Reproduction Asexual Reproduction Single parent produces offspring All offspring are genetically identical to one another and to parent Produces identical somatic (body) cells Sexual
The geneticist s questions
 The geneticist s questions a) What is consequence of reduced gene function? 1) gene knockout (deletion, RNAi) b) What is the consequence of increased gene function? 2) gene overexpression c) What does
The geneticist s questions a) What is consequence of reduced gene function? 1) gene knockout (deletion, RNAi) b) What is the consequence of increased gene function? 2) gene overexpression c) What does
Dr. Mahmood S. Choudhery, PhD, Postdoc (USA) Assistant Professor Tissue Engineering & Regenerative Medicine King Edward Medical University
 Assistant Professor Tissue Engineering & Regenerative Medicine King Edward Medical University") CELL DIVISION Dr. Mahmood S. Choudhery, PhD, Postdoc (USA) Assistant Professor Tissue Engineering & Regenerative Medicine King Edward Medical University Cell Division The key roles of cell division Unicellular
CELL DIVISION Dr. Mahmood S. Choudhery, PhD, Postdoc (USA) Assistant Professor Tissue Engineering & Regenerative Medicine King Edward Medical University Cell Division The key roles of cell division Unicellular
BIOH111. o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system
 BIOH111 o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 Textbook
BIOH111 o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 Textbook
Chapters 12&13 Notes: DNA, RNA & Protein Synthesis
 Chapters 12&13 Notes: DNA, RNA & Protein Synthesis Name Period Words to Know: nucleotides, DNA, complementary base pairing, replication, genes, proteins, mrna, rrna, trna, transcription, translation, codon,
Chapters 12&13 Notes: DNA, RNA & Protein Synthesis Name Period Words to Know: nucleotides, DNA, complementary base pairing, replication, genes, proteins, mrna, rrna, trna, transcription, translation, codon,
CHAPTER 12 - THE CELL CYCLE (pgs )
") CHAPTER 12 - THE CELL CYCLE (pgs. 228-245) CHAPTER SEVEN TARGETS I. Describe the importance of mitosis in single-celled and multi-cellular organisms. II. Explain the organization of DNA molecules and their
CHAPTER 12 - THE CELL CYCLE (pgs. 228-245) CHAPTER SEVEN TARGETS I. Describe the importance of mitosis in single-celled and multi-cellular organisms. II. Explain the organization of DNA molecules and their
5. As compared to the human sperm cell, the human egg cell contains more... a) cytoplasm c) centrosomes b) mitochondria d) chromosomes
 cytoplasm c) centrosomes b) mitochondria d) chromosomes") BIOLOGY - 3201. Quiz: Cell Reproduction. NAME : Multiple Choice. (1% each) 1. Which of the following is NOT true of mitotic cell division? a) It involves nuclear division. b) It involves division of the
BIOLOGY - 3201. Quiz: Cell Reproduction. NAME : Multiple Choice. (1% each) 1. Which of the following is NOT true of mitotic cell division? a) It involves nuclear division. b) It involves division of the
5.1 Cell Division and the Cell Cycle
 5.1 Cell Division and the Cell Cycle Lesson Objectives Contrast cell division in prokaryotes and eukaryotes. Identify the phases of the eukaryotic cell cycle. Explain how the cell cycle is controlled.
5.1 Cell Division and the Cell Cycle Lesson Objectives Contrast cell division in prokaryotes and eukaryotes. Identify the phases of the eukaryotic cell cycle. Explain how the cell cycle is controlled.
Chromosome duplication and distribution during cell division
 CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
3.a.2- Cell Cycle and Meiosis
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. 3.a.2- Cell Cycle and Meiosis EU 3.A: Heritable information provides for continuity of life.
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. 3.a.2- Cell Cycle and Meiosis EU 3.A: Heritable information provides for continuity of life.
The Cellular Basis of Inheritance
 CHAPTER 9 The Cellular Basis of Inheritance Summary of Key Concepts Concept 9.1 All cells come from cells. (pp. 180 181) Cell reproduction is an important process. Three functions of cell reproduction
CHAPTER 9 The Cellular Basis of Inheritance Summary of Key Concepts Concept 9.1 All cells come from cells. (pp. 180 181) Cell reproduction is an important process. Three functions of cell reproduction
Ch. 13 Meiosis & Sexual Life Cycles
 Introduction Ch. 13 Meiosis & Sexual Life Cycles 2004-05 Living organisms are distinguished by their ability to reproduce their own kind. -Offspring resemble their parents more than they do less closely
Introduction Ch. 13 Meiosis & Sexual Life Cycles 2004-05 Living organisms are distinguished by their ability to reproduce their own kind. -Offspring resemble their parents more than they do less closely
What happens to the replicated chromosomes? depends on the goal of the division
 Segregating the replicated chromosomes What happens to the replicated chromosomes? depends on the goal of the division - to make more vegetative cells: mitosis daughter cells chromosome set should be identical
Segregating the replicated chromosomes What happens to the replicated chromosomes? depends on the goal of the division - to make more vegetative cells: mitosis daughter cells chromosome set should be identical
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Biology. Chapter 12. Meiosis and Sexual Reproduction. Concepts and Applications 9e Starr Evers Starr. Cengage Learning 2015
 Biology Concepts and Applications 9e Starr Evers Starr Chapter 12 Meiosis and Sexual Reproduction 12.1 Why Sex? In asexual reproduction, a single individual gives rise to offspring that are identical to
Biology Concepts and Applications 9e Starr Evers Starr Chapter 12 Meiosis and Sexual Reproduction 12.1 Why Sex? In asexual reproduction, a single individual gives rise to offspring that are identical to
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Learning Objectives Chapter 8
 Learning Objectives Chapter 8 Brief overview of prokaryotic cell replication The three main phases of eukaryotic cell division: Interphase, M phase, C phase Interphase is broken down into three sub-phases
Learning Objectives Chapter 8 Brief overview of prokaryotic cell replication The three main phases of eukaryotic cell division: Interphase, M phase, C phase Interphase is broken down into three sub-phases
Biology I Fall Semester Exam Review 2014
 Biology I Fall Semester Exam Review 2014 Biomolecules and Enzymes (Chapter 2) 8 questions Macromolecules, Biomolecules, Organic Compunds Elements *From the Periodic Table of Elements Subunits Monomers,
Biology I Fall Semester Exam Review 2014 Biomolecules and Enzymes (Chapter 2) 8 questions Macromolecules, Biomolecules, Organic Compunds Elements *From the Periodic Table of Elements Subunits Monomers,
Chapter 11: The Continuity of Life: Cellular Reproduction
 Chapter 11: The Continuity of Life: Cellular Reproduction Chapter 11: Cellular Reproduction What is Cellular Reproduction? Answer: The division of a parent cell into two daughter cells Requirements of
Chapter 11: The Continuity of Life: Cellular Reproduction Chapter 11: Cellular Reproduction What is Cellular Reproduction? Answer: The division of a parent cell into two daughter cells Requirements of
Mitosis, development, regeneration and cell differentiation
 Mitosis, development, regeneration and cell differentiation Mitosis is a type of cell division by binary fission (splitting in two) which occurs in certain eukaryotic cells. Mitosis generates new body
Mitosis, development, regeneration and cell differentiation Mitosis is a type of cell division by binary fission (splitting in two) which occurs in certain eukaryotic cells. Mitosis generates new body
Biology 321 Genetics Winter 2009
 Biology 321 Genetics Winter 2009 Dr. Sandra Schulze Office BI409 Office Hours: WF 4:00PM 5:00PM Grading Exam I: 25% Exam II: 25% Exam III: 25% Problem sets 25%* No specific grades for attendance or participation,
Biology 321 Genetics Winter 2009 Dr. Sandra Schulze Office BI409 Office Hours: WF 4:00PM 5:00PM Grading Exam I: 25% Exam II: 25% Exam III: 25% Problem sets 25%* No specific grades for attendance or participation,
Number of questions TEK (Learning Target) Biomolecules & Enzymes
 Biomolecules & Enzymes") Unit Biomolecules & Enzymes Number of questions TEK (Learning Target) on Exam 8 questions 9A I can compare and contrast the structure and function of biomolecules. 9C I know the role of enzymes and how
Unit Biomolecules & Enzymes Number of questions TEK (Learning Target) on Exam 8 questions 9A I can compare and contrast the structure and function of biomolecules. 9C I know the role of enzymes and how
3.2.2 All cells arise from other cells
 alevelbiology.co.uk SPECIFICATION Within multicellular organisms, not all cells retain the ability to divide. Eukaryotic cells that do retain the ability to divide show a cell cycle. DNA replication occurs
alevelbiology.co.uk SPECIFICATION Within multicellular organisms, not all cells retain the ability to divide. Eukaryotic cells that do retain the ability to divide show a cell cycle. DNA replication occurs
Cellular Division. copyright cmassengale
 Cellular Division 1 Cell Division All cells are derived from pre- existing cells New cells are produced for growth and to replace damaged or old cells Differs in prokaryotes (bacteria) and eukaryotes (protists,
Cellular Division 1 Cell Division All cells are derived from pre- existing cells New cells are produced for growth and to replace damaged or old cells Differs in prokaryotes (bacteria) and eukaryotes (protists,
Honors Biology-CW/HW Cell Biology 2018
 Class: Date: Honors Biology-CW/HW Cell Biology 2018 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Hooke s discovery of cells was made observing a. living
Class: Date: Honors Biology-CW/HW Cell Biology 2018 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Hooke s discovery of cells was made observing a. living