Deregulation and Mislocalization of the Cytokinesis Regulator ECT2 Activate. the Rho Signaling Pathways Leading to Malignant Transformation
|
|
|
- Shanon Mosley
- 5 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on November 25, 2003 as Manuscript M Deregulation and Mislocalization of the Cytokinesis Regulator ECT2 Activate the Rho Signaling Pathways Leading to Malignant Transformation Shin ichi Saito 1, Xiu-Fen Liu, Keiju Kamijo, Razi Razziudin, Takashi Tatsumoto 2, Isamu Okamoto, Xiaoyan Chen, Chong-Chou Lee, Matthew V. Lorenzi 3, Naoya Ohara 4, and Toru Miki* From the Molecular Tumor Biology Section, Basic Research Laboratory, National Cancer Institute, 37 Convent Dr. MSC 4255, Bethesda, MD Present address: Loaboratory of Cell Biology, National Cancer Institute, Convent Dr. MSC 4255, Bethesda, MD Present address: Fukuoka Teishin Hospital, Yakuin, Chuo-ku, Fukuoka , Japan. 3 Present address: Oncology Drug Discovery, Pharmaceutical Research Institute Bristol-Myers Squibb, P.O. Box 4000, Princeton, NJ Present address: Department of Bacteriology, Faculty of Dentistry, Nagasaki University, Nagasaki, Japan. *To whom correspondence should be addressed. Tel.: (301) ; Fax.: (301) ; toru@helix.nih.gov. Running Title: Mechanism of ECT2 activation Key words: exchange factor, focus formation, protooncogene, Rho GTPase, transformation 1
2 ABSTRACT The human ECT2 protooncogene encodes a guanine nucleotide exchange factor for the Rho GTPases and regulates cytokinesis. Although the oncogenic form of ECT2 contains an N-terminal truncation, it is not clear how the structural abnormality of ECT2 causes malignant transformation. Here we show that both the removal of the negative regulatory domain and alteration of subcellular localization are required to induce the oncogenic activity of ECT2. The transforming activity of oncogenic ECT2 was strongly inhibited by dominant-negative Rho GTPases, suggesting the involvement of Rho GTPases in ECT2 transformation. While deletion of the N- terminal cell cycle regulator-related domain (N) of ECT2 did not activate its transforming activity, removal of the small central domain (S), which contains two nuclear localization signals (NLSs), significantly induced the activity. The ECT2 N domain interacted with the catalytic domain, and significantly inhibited the focus formation by oncogenic ECT2. Interestingly, the introduction of the NLS mutations in the S domain of N-terminally truncated ECT2 dramatically induced the transforming activity of this otherwise non-oncogenic derivative. Among the known Rho GTPases expressed in NIH 3T3 cells, RhoA was predominantly activated by oncogenic ECT2 in vivo. Therefore, the mislocalization of structurally altered ECT2 might cause the untimely activation of cytoplasmic Rho GTPases leading to the malignant transformation. 2
3 INTRODUCTION The ECT2 oncogene has been isolated in a search for mitogenic signal transducers in epithelial cells, where a murine keratinocyte expression cdna library was introduced into fibroblasts to induce foci of morphologically transformed cells (1). The ECT2 transfectants exhibit anchorage-independent cell growth and efficient tumor formation in nude mice. The transforming ECT2 cdna encodes the C- terminal half of the full-length protein containing Dbl-homology (DH) and pleckstrin homology (PH) domains, which are now found in a number of molecules involved in regulation of the Rho family GTPases. The N-terminal half of ECT2 contains domains related to cell cycle control and repair proteins, including Clb6 and Rad4/Cut5 (2,3). CLB6 encodes a B-type cyclin of the budding yeast, which promotes the transition from G1 into S phase (4). Fission yeast cut5, which is identical to the repair gene rad4, is required for both the onset of S phase and the restraint of M phase before the completion of S phase (5). The Cut5-related domain of ECT2 consists of two repeats (6,7), designated BRCT (BRCA1 C-terminal) repeats, which are widespread in a number of cell-cycle checkpoint control and DNA repair proteins (7). These cell-cycle regulator-related domains of ECT2 play essential roles on the regulation of cytokinesis (2,3). ECT2 catalyzes guanine nucleotide exchange in vitro on three representative Rho GTPases; RhoA, Rac1 and Cdc42 (2). The Rho family of small GTPases function as molecular switches of diverse biological functions including cytoplasmic actin reorganization, cell motility and cell scattering (8). Activation of the Rho proteins is promoted by guanine nucleotide exchange factors (GEFs), which catalyze the replacement of bound GDP by GTP. The GTP-bound form of Rho proteins can 3
4 specifically interact with their effectors or targets and transmit signals to downstream molecules. Rho proteins are inactivated through the hydrolysis of bound GTP to GDP by the intrinsic GTPase activity assisted by GTPase activating proteins (GAPs). RhoA, Rac1 and Cdc42 induce the formation of actin stress fibers, lamellipodia and filopodia, respectively (9). Among the known guanine nucleotide exchange factors for Rho GTPases, ECT2 shows several unique characteristics. ECT2 expression is induced in S phase and reaches to the highest level in G2 and M phases in regenerating mouse liver (10). ECT2 protein is specifically phosphorylated in G2 and M phases (2). ECT2 exhibits nuclear localization in interphase, disperses throughout the cytoplasm in prometaphase, and is condensed in the midbody during cytokinesis. Expression of a dominant negative ECT2 or microinjection of anti-ect2 antibody strongly inhibits cytokinesis, indicating that ECT2 is a critical regulator of cytokinesis (2). Further, the Drosophila pbl gene, whose mutation results in the inhibition of cytokinesis in mitotic cycle 14 during embryogenesis, was found to encode the fly homologue of human ECT2 (11). Although the transforming activity of several DBL family oncogenes is stimulated by N-terminal alterations (12-15), the activation mechanisms are still obscure. Since Rho GTPases play a critical role in cell transformation (16-18), ECT2 may display its transforming activity through the activation of Rho proteins. However, ECT2 is predominantly expressed in the nucleus where no expression of Rho GTPases is reported. In the present study, we examined the activation mechanism of the transforming activity of ECT2. We identified the small central domain containing two tandem nuclear localization signals as a negative regulator of the transforming activity. We show that elimination of these signals and a 4
5 negative regulatory domain from ECT2 resulted in the activation of Rho GTPases in the cytoplasm, leading to malignant transformation of the cells. 5
6 EXPERIMENTAL PROCEDURES DNA constructs. Full-length and N-terminally truncated ECT2 cdnas were amplified by PCR using ECT2 clone 1M (2,3) as template, and subcloned between the BamHI and EcoRI sites of the mammalian expression vector pcev29 or its derivative pcev29f3, which contains three tandem copies of FLAG sequence (19). ECT2 N-terminal derivatives, ECT2-N1 (amino acids 1-421), ECT2-N2 (amino acids 1-378), ECT2-N3 (amino acids 1-360) and ECT2-N4 (amino acids 1-333) have been described (3). ECT2- S mutant lacking the S domain (amino acids ) was created from two PCR products using the same template and primers I, II, III and IV with following sequences: I, 5'-CTC GGA TTC ATG GCT GAA AAT AGT GTA TTA-3'; II, 5'-CAG ACT CGC GGA GTA TTT GCC TTT TCA TA-3'; III, 5'-TCA CTC CGC GGT GGC AAG TTG CAA AAG AG-3'; and IV, 5' ACT GAA TTC GGT AAC GCT TCA TAT CAA ATG 3'. The PCR product synthesized using primer pairs I and II were digested with BamHI and BstUI. The PCR product generated by primers III and IV were digested with BstUI and EcoRI. These products were ligated together with the pcev29 or pcev29f3 vector, which had been digested with BamHI and EcoRI, to create ECT2- S. Two ECT2 mutants, S1 and S2, containing RRR to AAA and R to A mutations in the NLS sequence of the S domain, respectively, were generated by the similar procedure, but following oligonucleotides were used for PCR to introduce mutations: V, 5'-CAG ACT GCG GCC GCT TTG CGA TTG CTG TTA GGG GT-3'; VI, 5'-TCA CTC GCG GCC GCT TTA AAA GAA ACA CTT GCT CAG-3'; VII, 5'-TTT GGC GCG CCC GGG GTG GAA ATG GTG ACA C-3'; and VIII, 5 -TTT GGC GCG CCC ATC AGC TGA GCA TTC CCT T-3'. NotI and AscI were used to create S1 and S2, respectively, instead of BstUI. S3 was created by the 6
7 similar procedure but S1 was used as a PCR template instead of ECT2-F. ECT2-F, S1, S2, S3, S and N5 were also cloned into pegfp-c1 (BD Biosciences Clontech, CA) to express ECT2 as green fluorescent protein (GFP)-fusion proteins. All constructs generated by the use of PCR were sequenced to ensure that no PCR mutation was generated except the desired mutations. An ECT2-DN5 derivative containing PVQR to AAAA mutations (amino acids ) in the DH domain was generated by amplifying two PCR fragments. Primers for the first fragment were; a forward primer with BamHI restriction site, 5'- CCC GGA TTC GCC ACC ATG GTT CCT TCA AAG CAG TCA GCA-3', and a reverse primer with SfiI site; 5'-CAG ACT GGC CGC TGC GGC CCG GAT AAG AAG TTC AAC AAG-3'. Primers for the second fragment were: a forward primer with SfiI site, 5'-TCA CTC GGC CGC AGC GGC CTT ACC CAG TGT TGC ATT ACT-3', and a reverse primer with EcoRI site, 5'-ACT GAA TTC GGT AAC GCT TCA TAT CAA ATG-3'. PCR products were then digested with the indicated restriction enzymes and simultaneously ligated with the pcev29f3 vector digested with BamHI and EcoRI. A new SfiI site was generated as a result of the introduction of PVQR to AAAA mutations. To introduce the SV40 NLS (nuc) into ECT2- N5, ECT2- N5 fragment was subcloned into BamHI site of pecfp-nuc vector (BD Biosciences Clontech), and then a DNA fragment containing the triple repeats of SV40 large T antigen NLS sequence (GAT CCA AAA AAG AAG AGA AAG GTA GAT CCA AAA AAG AAG AGA AAG GTA GAT CCA AAA AAG AAG AGA AAG GTA) and ECT2- N5 sequence was excised and then cloned in pegfp-c1 and pcev29f3 vector. Expression of fusion proteins of expected sizes were confirmed by Western blotting. 7
8 ECT2 NLS sequence (amino acids ) was attached to 3 end of EGFP sequence (BD Biosciences Clontech) by PCR, and subcloned between BamHI and EcoRI sites of the mammalian EGFP expression vector pcaggfp (20) to create an EGFP-EGFP-ECT2 NLS fusion protein. The expression of 60 kda ECT2 NLS-tagged tandem GFP protein was confirmed by Western blotting in U-2 OS cells. Focus forming assays. NIH 3T3 cells were transfected with various amounts (0.01 to 1.0 mg) of the eukaryotic expression vector pcev29 or pcev29f3 (19) containing ECT2 cdnas or vector alone by the calcium phosphate transfection method. Focus formation was observed in unselected plates approximately 14 days after transfection and quantified after Giemsa staining. FLAG-tagged ECT2 variants (in pcev29f3) showed slightly lower transforming activity than non-tagged versions (in pcev29). Transforming activity was expressed as the number of foci per pmol DNA (ffu/pmol). Comparative efficiency of transfection was confirmed by G- 418 resistant colony formation in duplicated plates. Expression levels of the ECT2 variants were examined using anti-flag M2 antibody (Sigma, MO) and anti-gfp (BD Biosciences Clonetech) 48 h after transfection with 10:1 mixture of FLAG-tagged ECT2 expression vector and pegfp-c1. Transient expression reporter gene assays. The construction of SRF-, and AP- 1-luciferase reporter plasmids in pgl2luc containing a minimal c-fos promoter (-56 to +109) has been described previously (21). The SRF binding sequence is derived from the serum response element of the c-fos gene and corresponds to SRE.mutL (22). Firefly luciferase reporter and TK-Renilla luciferase control plasmids were cotransfected with each expression vector into COS cells. Total amount of DNA was adjusted by the addition of vector DNA. Thirty six to forty eight hours posttransfection, cells were lysed and luciferase activities were measured using Dual- 8
9 Luciferase Reporter Assay System (Promega, WI) according to the accompanied protocol. Firefly luciferase activity in the lysates was normalized to Renilla luciferase activity and expressed as a relative luciferase activity. No significant increase in luciferase activity was observed following transfection of each expression vector DNA with a reporter plasmid containing only a luciferase cassette and c-fos minimal promoter. In vitro invasion assays. Invasion assays were performed using Biocoat Matrigel invasion chambers (Becton Dickinson, MA) essentially as described in the manufacturer s protocol. Matrigel invasion chambers (24-well) were rehydrated with DMEM containing 0.1% bovine serum albumin (BSA) for two hours at room temperature. NIH 3T3 cells transfected with ECT2 or control vectors were seeded at 5 x 10 5 / 0.4 ml medium containing 0.1% BSA into the inner well of invasion chambers. The outer chambers were filled with 0.4 ml medium containing 10% calf serum. Cells were allowed to invade Matrigel matrices for hours at 37 C in a CO 2 -incubator. To count cells that migrated onto the membrane at lower surface, the cells on the upper side of the membrane were scraped off with cotton swipe, then the inserts with membrane were stained with Diff-Quick (Dade Diagnostics, PR). Cells on the lower side of membrane were photographed and counted. Activation assays for Rho GTPases. COS cell were transfected by GFP or GFP-ECT2-DN5 expression vectors together with pef4/myc-his (Invitrogen, CA) carrying inserts for AU5-tagged Rho GTPase. Lysates were prepared 24 hr after transfection, and the GTP bound forms of Rho GTPases were determined by pull down assays using GST-Rhotekin (for RhoA, RhoB and RhoC) or GST-PAK PBD according to the manufacturer s protocol (Cytoskeleton, CO). 9
10 Subcellular localization and cell morphology. NIH 3T3 cells were transfected with the GFP- or FLAG-tagged ECT2 expression vectors using the LipofectAMINE PLUS reagent (Invitrogen, CA). GFP -expressing cells were identified by green fluorescence. Actin and DNA were stained with rhodamine-conjugated phalloidin (Sigma, MO) and 4,6 -diamidino-2-phenylindole (DAPI; Sigma, MO), respectively, as reported previously (2,19). FLAG-tagged ECT2 derivatives (0.5 mg each) were transfected into NIH 3T3 cells using the LipofectAMINE PLUS reagent (Invitrogen, CA). Cells were fixed with a freshly prepared mixture of methanol:acetone (1:1) for 2 minutes at room temperature 40 hr after transfection. Expressed proteins were visualized using anti-flag M2 monoclonal antibody-cy3 conjugate (Sigma, MO) in the presence of 1 mg/ml of DAPI. U-2 OS cells were also transiently transfected with the expression vectors for GFP-tagged ECT2 mutants using FuGENE 6 reagent (Roche, IN) to confirm their subcellular localization. In this case, Hoechst dye was added to culture medium at a final concentration of 10 mm and cells were directly observed under the fluorescence microscope. Images were acquired using a Zeiss Axiovert microscope equipped with a Photometrics digital camera and processed with IPLab software (Signal Analytics, VI). Time-lapse video microscopy. The RFP-ECT2-DN5 expression vector was constructed by inserting ECT2-DN5 into prsred2-c1 (BO Biosciences Clontech) at the BglII and EcoRI sites. pegfp-actin was obtained from Clontech. NIH 3T3 was transfected with the equal mixture of both the plasmid using FuGENE 6 transfection reagent (Roche). Cells were cultured on a 35-mm plates and cultured in an environmental chamber on a stage of Zeiss Axiovert S-100 microscope equipped 10
11 with motorized X-Y-Z stages. Images were taken at 3 min intervals by Photometric digital camera controlled by OpenLab software (Improvision, MA). 11
12 RESULTS Deletion of the N-terminal half of human ECT2 induces cell transformation and invasiveness. The mouse ect2 cdna, ect2-t, which carries an N-terminal truncation, exhibits a high transforming activity, while the full-length clone does not significantly induce transformation (23). To test if N-terminal truncation can also activate the transforming activity of human ECT2, we generated a FLAG epitope-tagged fulllength human ECT2 and its derivative ECT2-DN5, which has a similar N-terminal deletion to mouse ect2-t. Like ect2-t, ECT2-DN5 exhibited a high transforming activity in NIH 3T3 cells, whereas the full-length ECT2, ECT2-F, did not show any detectable activity (Figure 1A). Both mouse and human ECT2 similarly induced tiny foci of transforming cells with stellate morphology, which is distinct from ras- or sisinduced foci (Figure 1A, B). We previously reported that ect2-t stimulates anchorage-independent growth of NIH 3T3 cells and tumorigenicity in nude mice (23). To examine additional oncogenic activities of ECT2, we established NIH 3T3 clones expressing FLAG-tagged derivatives of ECT2-F, ECT2-DN5 and ECT2-N2. ECT2-N2 carries the region from the N-terminus to the S domain (see Figure 6A). Western blot analysis with anti-flag antibody showed that these stable transfectants expressed FLAGtagged ECT2-F, ECT2-N2 and ECT2-DN5 at comparable levels (Figure 1C). Upon plating, ECT2-DN5 expressing cells formed secondary foci with stellate morphology, whereas the morphology of ECT2-F and ECT2-N2 clones was indistinguishable from the vector alone transfectants (Figure 1B). When cultured in the presence of 10% 12
13 serum, all of the stable ECT2 clones exhibited similar growth properties (data not shown). However, in medium containing 1% serum, vector alone, ECT2-N2 and ECT2-F transfectants did not grow well and the number of viable cells gradually decreased (Figure 2A). In contrast, ECT2-DN5 transfectants continued to grow for at least 48 h under these conditions, suggesting that these transfectants acquired low serum dependency. To test if ECT2 can induce cell invasiveness, NIH 3T3 cells expressing ECT2- F, ECT2-DN5, or vector alone were placed on the surface of an artificial basement membrane, Matrigel, and the number of the cells, which had migrated through the membrane, was counted. Interestingly, ECT2-DN5 transfectants exhibited a strong invasion activity, whereas ECT2-F or the vector alone transfectants did not show significant activity (Figure 2B). Additionally, mouse ect2-t, which corresponds to human ECT2-DN5, also exhibited a high activity of cell invasiveness. These results indicate that oncogenic ECT2 is an efficient activator of cell invasiveness. Oncogenic ECT2 activates Rho signaling pathways. Since Rho GTPases are known to regulate the JNK and p38 MAP kinase pathways (24,25), we examined whether these pathways are activated in ECT2 transfectants. We first analyzed endogenous JNK activity in these ECT2 transfectants using an antibody specific to the activated form of c-jun, which is phosphorylated at Serine 63. In vector alone transfectants, a very low level of JNK activity was observed (Figure 3A, top). The activity of JNK in these cells was increased by the stimulation with sorbitol, an activator of the JNK pathway. The activity of JNK was also elevated in cells expressing ECT2-DN5. In contrast, ECT2-F or ECT2-N2 expression 13
14 did not significantly affect the JNK activity. The JNK activity was induced by sorbitol to a similar level in all the transfectants (data not shown), indicating that all of the transfectants maintained the ability to induce JNK activity. In contrast, we did not observe significant activation of p38 or Erk MAP kinases by ECT2 and its derivatives (Figure 3A middle and bottom). These results indicate that ECT2-DN5 preferentially activates the JNK signaling pathway. Rho proteins can stimulate the transcriptional activity regulated by serum response factor (SRF) (22). To examine whether ECT2 can stimulate SRF-regulated transcription, a serum response element (SRE)-luciferase plasmid was used as a reporter. Upon coexpression of the reporter plasmid with either the full-length or truncated ECT2 expression vector, luciferase activity was estimated. As shown in Figure 3B upper panel, expression of ECT2-DN5 potently induced the transcriptional activity of the SRE reporter plasmid. In contrast, expression of ECT2-F exhibited the activity slightly higher than the vector alone control. The SRE-regulated transcriptional activity induced by ECT2-DN5 was efficiently inhibited by either of dominant negative RhoA, Rac1 or Cdc42. Moreover, either of constitutively active RhoA, Rac1 or Cdc42 efficiently enhanced SRE-mediated transcription in this system. These results suggest that ECT2-DN5 can stimulate SRE-mediated transcription through the activation of Rho GTPases. We previously showed that Ost, a guanine nucleotide exchange factor for RhoA and Cdc42, activates the transcriptional activity regulated by Activator Protein-1 (AP-1) (19). To examine whether ECT2 can also stimulate AP-1-regulated transcriptional activity, an AP-1-binding site-luciferase plasmid was utilized as a reporter. As shown in Figure 3B lower panel, expression of ECT2-DN5 moderately 14
15 elevated AP-1-regulated transcriptional activity, whereas ECT2-F or ECT2-N2 failed to stimulate the activity. Coexpression of dominant negative RhoA, Rac1 or Cdc42 reduced the ECT2-DN5-mediated stimulation of AP-1-regulated transcription, albeit at lower levels as compared with their effects on SRE-regulated transcription. We also found that constitutively active RhoA, Rac1 or Cdc42 efficiently stimulated AP- 1-mediated transcription. Among these GTPases, Rac1 displayed the highest level of stimulation of AP-1-regulated transcriptional activity. All of these results indicate that ECT2 can regulate the transcriptional events mediated by SRE and AP-1 through the activation of Rho GTPases. Oncogenic ECT2 induces cell rounding in NIH 3T3 fibroblasts. Rho family proteins are involved in the organization of actin-based cytoskeletal structures. In fibroblasts, RhoA activates actin stress fiber formation, whereas Rac1 and Cdc42 induce lamellipodia and filopodia formation, respectively (8). To test which actin-based structures ECT2 can induce, we transiently expressed GFP-tagged ECT2-F, ECT2-DN5, or GFP vector alone in NIH 3T3 cells. A population (~20%) of cells expressing GFP-ECT2-DN5 showed a flat phenotype with moderately enhanced actin stress fiber formation (Figure 4A, GFP-ECT2-DN5 left panesl), suggesting that Rho was preferentially activated by ECT2-DN5 in these cells. However, the majority of NIH 3T3 cells expressing GFP-ECT2-DN5 exhibited a compacted structure with saturated F-actin staining (Figure 4A, GFP-ECT2-DN5 right panels). In contrast, the morphology of the surrounding cells, which were not expressing the GFP-fusion protein, was similar to that of vector alone transfectants. These findings are 15
16 consistent with the previous results that oncogenic ECT2 induces foci containing both fusiforms and rounded cells (23). To examine how these compacted cells were generated by ECT2-DN5 expression, NIH 3T3 cells were transfected with Red Fluorescent Protein (RFP)- fused ECT2-DN5 and GFP-fused actin expression vectors. Cells expressing RFP- ECT2-DN5 were identified with red fluorescence, and their morphologies were examined by time-lapse video microscopy. In most of cells expressing RFP-ECT2- DN5, actin stress fibers were disrupted h after transfection, and the cells rounded up like the M phase cells (Figure 4B, see the cell shown by an arrow), but most of them did not divide in a next few hours. The cell that once rounded nearly completely appeared to flatten to some extent (panels h) and then rounded again (panels h after transfection). RFP-ECT2-DN5 was detected in the entire cell of these transfectants (data not shown). In Figure 4B, another cell expressing ECT2-DN5 at similar level as determined by the red fluorescence also exhibited a similar morphology with additional cortical activities (lower right corner). In contrast, cells expressing ECT2-DN5 weakly did not round up, although they expressed GFP-actin at a similar level (see the rightmost cell in Figure 4B). These results suggest that oncogenic ECT2 stimulates cellular transformation by dramatically changing their actin cytoskeletal morphology in NIH 3T3 cells. ECT2 transformation is dependent on the activation of Rho GTPases. ECT2 can activate RhoA, Rac1 and Cdc42 through guanine nucleotide exchange in vitro (2). Rho GTPases have been shown to play a critical role in cellular transformation (16-18). To test whether ECT2 transformation involves Rho 16
17 activation, we cotransfected NIH 3T3 cells with the ECT2-DN5 expression vector (0.1 mg) and each of the expression vectors encoding the dominant negative (DN) forms of Rho proteins (0.5 mg). Since the addition of a large amount of DNA usually inhibits focus formation presumably due to the competition for the available transcription and translation machinery in the cells, we compared the effects of wild type (WT) and DN constructs whose difference is in a single amino acid residue. As shown in Figure 5A, either of dominant-negative RhoA, Rac1 or Cdc42 more strongly inhibited ECT2-mediated focus formation in NIH 3T3 cells than the WT counterparts. Inhibition of ECT2 transformation by the empty vector was at the similar level by the WT Rho GTPase expression vectors, and a lower amount of WT Rho GTPases did not exhibit the inhibitory effect on ECT2 transformation (data not shown). These results suggest that ECT2-DN5 induces malignant transformation through the activation of Rho GTPases. The S domain of ECT2 plays a critical role in the regulation of transformation. To determine which domains in the N-terminal half of ECT2 regulate the transforming activity, we generated a set of overlapping N-terminal truncation mutants, and expressed these constructs in NIH 3T3 cells as FLAG-tagged proteins. Unexpectedly, the N-terminal deletions extending to the N-terminal most region (N), CLB6-homology domain, or each of the two BRCT domains did not activate the transforming activity of ECT2 (Figure 6A). However, two ECT2 derivatives containing the N-terminal deletions extended to the small central region, designated the S domain, exhibited a markedly high transforming activity (Figure 6A; see DN9 and DN5). To determine if the S domain itself regulates the transforming activity, a 17
18 derivative of ECT2-F lacking the S domain was generated. This mutant (DS) exhibited a high level of transforming activity in NIH 3T3 cells, suggesting that the S domain plays a critical role in the regulation of the transforming activity of ECT2. When the N-terminal deletions extended to the neighboring DH domain, the mutant ECT2 did not show detectable transforming activity (Figure 6A, DN6 and DN8), suggesting that the DH domain is required for the transforming activity of ECT2. To confirm this, point mutations (PVQR AAAA, amino acids ) were introduced into the most conserved region of the DH domain in ECT2-DN5. This mutant plasmid, ECT2-DN5 DH -, did not exhibit any detectable transforming activity (Figure 6A). Therefore, the DH domain, and in turn the exchange activity for the Rho GTPases, appeared to be essential for the transforming activity of ECT2. We also generated a series of C-terminal deletion mutants of ECT2-DN5. A deletion extended to the PH domain (DC1) eliminated the transforming activity from ECT2- DN5, suggesting that the PH domain is also essential for the transforming activity of ECT2. A small C-terminal deletion (DC3) did not affect the transforming activity of ECT2-DN5. A mutant protein lacking most of the C-terminal domain (DC2) was weakly transforming, but the morphology of the foci was less aggressive than the foci induced by ECT2-DN5. The expression level of the exogenous ECT2 proteins in transiently transfected NIH 3T3 cells was examined by immunoblotting using anti- FLAG monoclonal antibody (Figure 6B). All of the ECT2 derivatives were detected as proteins of expected sizes. The expression level of all the ECT2 derivatives was comparable with an exception of ECT2-DN5DC2, which showed markedly high expression. The reason of this high expression is not known, but this expression level might contribute to the transforming activity of ECT2-DN5DC2. 18
19 Transforming ECT2 derivatives partially localize in the cytoplasm. We have found that ECT2 is localized in the nucleus in interphase cells (2). Several putative nuclear localization signals (NLSs) were found in the predicted ECT2 sequence including RKRRR (amino acids ) and PRKRP ( ) located in the S domain (Figure 6A). Like endogenous ECT2, exogenously expressed, FLAG- ECT2-F also localized in the nucleus, whereas FLAG-ECT2-DN5, which lacks both the NLSs, was detected in both the nucleus and cytoplasm (Figure 6C). GFP-tagged ECT2 derivatives also showed similar localization patterns to their FLAG-tagged counterparts (data not shown). ECT2-DN5 contains a putative NLS at the C-terminal domain. GFP-ECT2-C, which consisted of the C-terminal domain alone (amino acids ), was localized predominantly in the nucleus (data not shown), suggesting that the NLS in the C-terminal domain is functional. Like ECT2-DN5, ECT2-DN9, which lacks the N-terminal NLS, had a high transforming activity, whereas ECT2-DN4, which contains both the NLSs in the S domain, did not exhibit detectable transforming activity (Figure 6A). ECT2-DN9 also exhibited both the nuclear and cytoplasmic localization (data not shown). Therefore, the transforming activity of ECT2 derivatives coincided with partial nuclear localization. Loss of nuclear localization signals affects the transforming activity of ECT2. To test whether the NLS sequences located in the S domain are responsible for the activation of the transforming activity of ECT2, we mutated the first (N-terminal) NLS (RKRRRLK) to RKAAALK in ECT2-F. As NLSs are usually rich in the positively charged amino acids Lys and Arg, Arg to Ala changes in NLSs should 19
20 reduce the nuclear localization of the protein. This mutant, ECT2-S1, exhibited a significant transforming activity, while ECT2-F did not have any detectable activity (Figure 6A). ECT2-S2, an ECT2-F derivative containing a mutation in the second NLS also exhibited a weak transforming activity. We also generated ECT2-S3, which contains mutations in both the NLSs in the S domain. This mutant also showed a significant transforming activity in NIH 3T3 cells. Subcellular localization analysis revealed that ECT2-S3 localized in the cytoplasm as well as in the nucleus (Figure 6C). ECT2-S1 and S2 also exhibited a similar localization to ECT2-S3 (data not shown). The expression levels of the wild type and mutant proteins were comparable (Figure 6B). These results suggest that the two NLSs in the S domain can function in vivo, and that the impairment of NLSs can activate the transforming activity of ECT2. To further verify that ECT2 NLSs can function as nuclear localization signals, these NLSs were introduced into tandem GFP. While GFP exhibited both the cytoplasmic and nuclear localization due to its relatively small size, tandem GFP showed reduced nuclear localization (Figure 7A). In contrast, the derivative of tandem GFP containing ECT2 NLSs displayed predominant nuclear localization, indicating that ECT2 NLSs are functional nuclear localization signals. To test whether cytoplasmic localization of ECT2-DN5 can be reduced by the addition of another nuclear localization signal, we introduced SV40 nuclear localization signal (nuc) into ECT2-DN5. As shown in Figure 7B, ECT2-DN5 nuc displayed predominant nuclear localization, whereas ECT2-DN5 exhibited both the cytoplasmic and nuclear localization. Additionally, GFP-tagged ECT2-DN5 displayed a relatively weak but significant transforming activity in NIH 3T3 cells, 20
21 whereas GFP- ECT2-DN5 nuc did not show detectable transforming activity (data not shown). All of these results strongly suggest that cytoplasmic localization of ECT2 is required for its transforming activity. The N-terminal domain of ECT2 interacts with the catalytic domain. While the N-terminal truncations that did not extend to the S domain did not activate the transforming activity of ECT2, the introduction of NLS mutations to ECT2-F stimulated its transforming activity. However, the level of activation by NLS mutations was very low compared with the deletion of the N-terminal half (DN5). The deletion of the entire S domain (DS) markedly stimulated the transforming activity, but the transforming activity of ECT2-DS was still lower than ECT2-DN5, suggesting that the sequence of the S domain other than the NLSs also negatively regulates the transforming activity. We reasoned that although the deletions of N- terminal domains themselves cannot induce the transforming activity of ECT2, these domains may inhibit the catalytic activity of ECT2 by the interaction with the C- terminal half. To test this possibility, GFP-tagged ECT2-N4 was coexpressed with FLAG-tagged ECT2-F, -N4, -DN5, or -DH in COS cells. When GFP-ECT2-N4 was immunoprecipitated with anti-gfp antibody, FLAG-tagged ECT2-F, -DN5 and DH were detected in the immunoprecipitates (Figure 8A). Particularly, FLAG-ECT2-DH exhibited a strong association with GFP-ECT2-N4. In contrast, FLAG-ECT2-N4 was not coimmunoprecipitated with GFP-ECT2-N4. These results suggest that the N- terminal domain of ECT2 can associate with the catalytic domain. We further examined whether ECT2 N-terminal derivatives possess the capability to inhibit the transforming activity of ECT2-DN5. As we previously 21
22 found that ECT2-N4 strongly inhibits cytokinesis (2), we used ECT2-N1, which contains the entire S domain, and ECT2-N3, which contains only the N-terminal NLS in the S domain, for this analysis. As shown in Figure 8B, either of ECT2-N1 or N3 significantly inhibited the transforming activity of ECT2-DN5. In contrast, either ECT2-N1 or N3 did not significantly affect the transforming activity of the H-ras oncogene. Transforming activity of N-terminally truncated ECT2 is strongly stimulated by the loss of nuclear localization signals. As ECT2-N can associate with the catalytic domain, such an intramolecular association may inhibit the catalytic activity of ECT2. This raised the possibility that the loss of NLSs from ECT2-DN4, which lacks the N-terminal domains but retains the S domain, activates the transforming activity. To explore this possibility, the NLS mutations were introduced into the S domain of ECT2-DN4, and their transforming activity was determined (Figure 9). Whereas ECT2-DN4 did not exhibit any detectable activity, ECT2-DN4S1, which contains mutations in the first NLS showed a strong focus formation in NIH3T3 cells. The introduction of S3 mutations (S1+S2) to ECT2-DN4 also exhibited a strong transforming activity. The transforming activity of ECT2-DN4S1 and ECT2-DN4S3 was comparable to that of ECT2-DS. As ECT2-DN5, which lacks the entire S domain, showed a higher activity, the NLS mutations might not strong enough to fully activate the transforming activity. These results suggest that loss of NLSs strongly activates the transforming activity when the N-terminal domain is not present. 22
23 RhoA is strongly activated by cytoplasmic ECT2. The above results suggest that the cytoplasmic localization of ECT2 derivatives lacking the N-terminal cell cycle regulater-related domains activate cytoplasmic Rho GTPases leading to malignant transformation. To test which Rho GTPases are activated in the cytoplasm by such the ECT2 derivatives in vivo, we cotransfected COS cells with expression vectors for AU5-tagged Rho GTPases and GFP-tagged ECT2-DN5 or GFP alone, and GTP-bound Rho GTPases were pulled down with GST-tagged Rho-binding domain of Rhotekin (GST-RBD) or p21-binding domain of PAK (GST-PBD). However, the initial results indicated that the expression level of exogenous Rho GTPases was affected by the ECT2 expression vector and thus the amount of Rho GTPases pulled down with GST-RBD or GST-PBD did not reflect the exchange activity of ECT2 (data not shown). Therefore, we subcloned the AU5-Rho inserts into the EF-1a promoter-based expression vector, and transfected them together with the ECT2-DN5 expression vector into COS cells. As shown in Figure 10A, the expression level of exogenous Rho GTPases were not affected by ECT2-DN5 expression. RhoA was efficiently activated by ECT2-DN5. RhoB and RhoC were also activated by ECT2-DN5 albeit less efficiently. In these experiments we used AU5-tagged Rho GTPases to avoid possible cross reactivity of specific antibodies. However, the detection of activation of endogenous RhoA, RhoB and RhoC by ECT2-DN5 using specific antibodies revealed similar results (data not shown). Rac2 was also dramatically activated by ECT2-DN5, and Rac1 was activated moderately. In contrast, activation of Cdc42 and TC10 was below the detectable level under these conditions (data not shown). 23
24 To test whether ECT2-N affects the exchange activity of ECT2 in vivo, we cotransfected COS cells with ECT2-DN5 and ECT2-N. GTP-bound RhoA was pulled down by GST-RBD and then detected with anti-rhoa antibody (Figure 10B). ECT2- N4, which lacks NLSs, relatively weakly but significantly reduced the accumulation of GTP-RhoA by ECT2-DN5. In contrast, ECT2-N2, which contains NLSs, did not affect the GTP-RhoA accumulation. We also tested a higher amount of ECT2-N DNAs (Figure 10B; ++), but they nonspecifically inhibited GTP-RhoA accumulation. 24
25 DISCUSSION In the present study, we characterized the malignant transformation induced by the human ECT2 protooncogene. Like many other Dbl family proteins, ECT2 also contains the DH and PH domains. However, ECT2 is unique among these proteins in that it contains cell cycle control domains in its N-terminal half and nuclear localization signals in the central and C-terminal domains. The transforming activity of ECT2 can be activated by the deletion of the N-terminal half. In addition to the previously reported oncogenic properties, we also found that ECT2-DN5 transfectants exhibited elevated cell invasiveness and reduced serum dependence. We found that oncogenic ECT2 activates several Rho GTPases-regulated signaling pathways. It has been shown that Rac1 and Cdc42 can activate JNK and p38 MAP kinase cascades (24,25). ECT2 efficiently activated JNK, but not p38 and Erk pathways at a detectable level. Although the differences of experimental conditions and sensitivity of the detection may explain why ECT2 did not activate p38 efficiently, ECT2 may associate with cellular components, which are specifically involved in JNK signaling. It is also possible that stimulation of the cycling of GDPand GTP-bound forms of Rho GTPases by exchange factors may activate the JNK pathways more efficiently than p38 pathway, whereas mutationally activated Rac1 and Cdc42 can efficiently stimulate both the pathways. As Rac2 is specifically expressed in cells of hematopoietic lineages (26), ECT2 may activate JNK through Rac1. However, we cannot rule out the possibility that ECT2 activates JNK through Cdc42 or other Rho family of GTPases, as these GTPases may be activated by ECT2 under certain conditions. 25
26 We have demonstrated that activated ECT2 can induce SRE- or AP-1- regulated transcriptional activity. While the activation of SRE-regulated transcription by ECT2 was efficiently inhibited by DN-Rho GTPases, AP-1-regulated activity was marginally inhibited by these mutant GTPases. This indicates that activation of AP-1-regulated transcription by ECT2 is partly attributed to the activation of Rho GTPases. Therefore, ECT2 may also activate other pathways to stimulate AP-1-mediated transcription. It has been reported that microinjection of porcine aortic endothelial cells with an oncogenic form of mouse ect2 induced lamellipodia formation (27). We also observed a similar phenotype in COS cells expressing ECT2-DN5 (data not shown). However, a population of NIH 3T3 fibroblasts expressing ECT2-DN5 exhibited enhanced stress fiber formation. These results may suggest that different Rho GTPases are activated by ECT2-DN5 in different cell types. We also found that the majority of the NIH 3T3 cells expressing ECT2-DN5 were completely rounded up and actin stress fibers appeared to have been disrupted. These results were consistent with the previous observations that ECT2-induced foci contained both rounded cells and fusiforms (23). Time-lapse microscopy analysis revealed that the morphological change upon ECT2-DN5 expression is a dynamic event, which oscillates between rounded and partially flatten cell shapes. It is possible that activation of Rho GTPases was controlled in a temporal manner: Upon the expression of oncogenic ECT2, Rho might be activated to induce stress fibers, but at a later stage these stress fibers might have been disrupted by unknown mechanisms which can generate rounded cells. As untransformed cells also round up immediately before cell division in their normal cell cycle, it would be of interest to 26
27 investigate whether or not the cytokinesis regulator ECT2 has an additional role in the control of cell rounding. We utilized focus formation assays to determine the region that regulates the oncogenic activity of ECT2. We found that mutations in the conserved amino acids in the DH domain efficiently abolished the transforming activity of ECT2. These results indicate that the activation of Rho GTPases is critical for cell transformation by ECT2. This conclusion is further supported by the finding that the transforming activity of ECT2 was efficiently inhibited by DN Rho GTPases. The DN RhoA, Rac1 and Cdc42 inhibited ECT2 transformation at a similar level and no significant difference was observed among the GTPases. As DN GTPases were thought to tightly bind their exchange factors to inhibit downstream signaling (28), these results are consistent with our previous findings that ECT2 activates RhoA, Rac1 and Cdc42 in vitro. We found that deletion of cell cycle regulator-related domains at the N- terminal half of ECT2 alone did not activate the transforming activity. Unexpectedly, deletion of the S domain was a critical factor for the transforming activity. The S domain contains two tandem nuclear localization signals and the ECT2 derivatives lacking the S domain partially localized in the cytoplasm. Because Rho proteins are known to localize in the cytoplasm and membrane fractions, a spill over of ECT2 into the cytoplasm might result in untimely Rho activation and eventually cause malignant transformation. The result that the ECT2 derivatives containing mutations at the NLSs in the S domain partially localized to the cytoplasm and exhibited an elevated level of the transforming activity further suggests the cytoplasmic localization of ECT2 as a major cause of the transforming activity. However, the activation level of the transforming activity of ECT2 by the 27
28 NLS mutants was very weak when compared with that of ECT2-DN5. Therefore, cytoplasmic localization itself might not be sufficient to fully activate the transforming activity of ECT2. The introduction of the NLS mutants into the N- terminally truncated ECT2, DN4, dramatically induced its transforming activity (Figure 9). These results suggest that although the deletion of the N-terminal cell cycle regulator-related domains itself cannot induce the transforming activity, partial cytoplasmic localization of such the ECT2 derivative efficiently induces cell transformation. Based on these results, we propose a following model on ECT2 activation (Figure 11). ECT2 is localized in the nucleus in interphase cells (2). In this stage, ECT2 may be in an inactive state, where the N-terminal domain interacts with the catalytic domain to inhibit its exchange activity. The deletion of the N-terminal cellcycle domains activates the catalytic activity of ECT2. However, this molecule still cannot activate Rho GTPases in vivo, because such ECT2 derivatives still localize in the nucleus. The removal of the S domain eliminates two NLSs, which causes partial cytoplasmic localization of such the molecules. Cytoplasmic localization of N-terminally truncated ECT2 activates Rho GTPases, resulting in malignant transformation (Figure 11 lower panel). As the introduction of the NLS mutations to the N-terminally truncated ECT2 strongly activated its transforming activity (Figure 9), the loss of the NLSs appeared to be the major cause of the activation of the transforming activity of ECT2. Since the introduction of DS deletion was sufficient to induce the transforming activity of ECT2, the S domain might also be involved in the negative regulation of the catalytic activity. However, as the transforming activity of ECT2-DS was still lower than ECT2-DN5 (Figure 9), the removal of the S 28
29 domain might only partially remove the negative regulation of the catalytic domain, and N-terminal truncation is required for full activation of the transforming activity. Examination of Rho GTPases activation in vivo using the GST-RBD and GST- PBD fusion proteins revealed that RhoA is strongly activated by ECT2-DN5. RhoB and Rac1 were also moderately activated. We also observed a weak activation of RhoC, but Cdc42 activation was under detection. Because the different GST fusion proteins were used to estimate the GTP-bound forms of these GTPases, and affinities of these proteins to these GTPases are also different, it would be difficult to compare the activation levels of the GTPases in accuracy. It is possible that Cdc42 activation in vivo is less efficient than in vitro by some reasons. For example, cytoplasmic activation of Rho may require a scaffold protein, and ECT2 and RhoA, but not Cdc42, may be recruited to such the scaffold. Alternatively, the exchange activity of ECT2 on Cdc42 may be stimulated by a modification such as phosphorylation. Such an example of substrate specificity conversion has been shown in a recent report that phosporylation by Aurora B converts MgcRacGAP to a RhoGAP during cytokinesis (29). As Rac2 expression is restricted is in the cells of hematopoietic lineages (26), and RhoC is not reportedly expressed in NIH 3T3 cells (30), RhoA, RhoB and Rac1 might be involved in transformation by ECT2-DN5. Among them, the highest activation level of RhoA by ECT2-DN5 may indicate the major role of this GTPase in ECT2-mediated cell transformation. While the Rho family of GTPases are involved in various biological functions in cells and tissues, the localization of their exchange factors is strictly restricted to certain cell types and/or specific subcellular structures. In the case of ECT2, its localization to the cleavage furrow and midbody appears to determine its role in the 29
30 regulation of cytokinesis. In interphase cells, ECT2 is localized to the nucleus. This subcellular localization might insulate ECT2 from its substrates and thus prevent malignant transformation. Additionally, ECT2 may have a nuclear function such as DNA damage and replication checkpoint control as suggested by the function of Cut5 (6,31). ECT2 is released to the cytoplasm in prometaphase after nuclear membrane breakdown (2), and subsequent activation of Rho GTPases is thought to trigger cytokinesis. An impediment of such the sophisticated regulatory mechanisms might result in mislocalization of ECT2 and untimely Rho activation leading to malignant transformation of the cells. Footnotes. The abbreviations used are: AP-1, activator protein-1; BRCT, BRCA1 C- terminal; CLB6, cyclin B6; Cut5, cells untimely torn 5; DAPI, 4', 6-diamidino-2'- phenylindole; DH, Dbl homology; ECT2, epithelial cell transforming gene 2 (human); ect2, mouse ECT2; Erk, extracellular signal regulated kinase; GAP, GTPase activating protein; GDP, guanosine diphosphate; GEF, guanine nucleotide exchange factor; GFP, green fluorescent protein; GST, glutathione S-transferase; JNK, c-jun N- terminal kinase; MAPK, mitogen-activated protein kinase; NLS, nuclear localizing signal; PH, pleckstrin homology; RFP, red fluorescent protein; SRE, serum response element; SRF, serum response factor. Acknowledgements. We thank Drs. J. Pierce, L Samelson and D. Lowy for support; V. Kapoor for technical assistance; and Drs. J.S. Gutkind and S. Narumiya for materials. This work was partly supported by a Breast Cancer Think Tank Award from NIH. TT and IO were supported by Japan Society of Promotion of Sciences fellowships for Biomedical and Behavioral Researchers in NIH. 30
31 REFERENCES 1. Miki, T., Fleming, T. P., Bottaro, D. P., Rubin, J. S., Ron, D., and Aaronson, S. A. (1991) Science 251, Tatsumoto, T., Xie, X., Blumenthal, R., Okamoto, I., and Miki, T. (1999) J. Cell Biol. 147, Saito, S., Tatsumoto, T., Lorenzi, M. V., Chedid, M., Kapoor, V., Sakata, H., Rubin, J., and Miki, T. (2003) J Cell Biochem 90, Schwob, E., and Nasmyth, K. (1993) Genes Dev 7, Saka, Y., and Yanagida, M. (1993) Cell 74, Saka, Y., Fantes, P., Sutani, T., McInerny, C., Creanor, J., and Yanagida, M. (1994) Embo J 13, Bork, P., Hofmann, K., Bucher, P., Neuwald, A. F., Altschul, S. F., and Koonin, E. V. (1997) Faseb J 11, Hall, A. (1998) Science 279, Olson, M. F., Ashworth, A., and Hall, A. (1995) Science 269, Sakata, H., Rubin, J. S., Taylor, W. G., and Miki, T. (2000) Hepatology 32, Prokopenko, S. N., Brumby, A., O'Keefe, L., Prior, L., He, Y., Saint, R., and Bellen, H. J. (1999) Genes Dev 13, Horii, Y., Beeler, J. F., Sakaguchi, K., Tachibana, M., and Miki, T. (1994) Embo J 13, Chan, A. M., McGovern, E. S., Catalano, G., Fleming, T. P., and Miki, T. (1994) Oncogene 9,
32 14. Chan, A. M., Takai, S., Yamada, K., and Miki, T. (1996) Oncogene 12, Cerione, R. A., and Zheng, Y. (1996) Curr Opin Cell Biol 8, Prendergast, G. C., Khosravi-Far, R., Solski, P. A., Kurzawa, H., Lebowitz, P. F., and Der, C. J. (1995) Oncogene 10, Qiu, R. G., Chen, J., McCormick, F., and Symons, M. (1995) Proc Natl Acad Sci U S A 92, Qiu, R. G., Chen, J., Kirn, D., McCormick, F., and Symons, M. (1995) Nature 374, Lorenzi, M. V., Castagnino, P., Chen, Q., Hori, Y., and Miki, T. (1999) Oncogene 18, Isaacs, J. S., Saito, S., and Neckers, L. M. (2001) J Biol Chem 276, Tolkacheva, T., Feuer, B., Lorenzi, M. V., Saez, R., and Chan, A. M. (1997) Oncogene 15, Hill, C. S., Wynne, J., and Treisman, R. (1995) Cell 81, Miki, T., Smith, C., Long, J., Eva, A., and Fleming, T. (1993) Nature 362, Coso, O. A., Chiariello, M., Yu, J. C., Teramoto, H., Crespo, P., Xu, N., Miki, T., and Gutkind, J. S. (1995) Cell 81, Minden, A., Lin, A., Claret, F. X., Abo, A., and Karin, M. (1995) Cell 81, Didsbury, J., Weber, R. F., Bokoch, G. M., Evans, T., and Snyderman, R. (1989) J Biol Chem 264,
33 27. Westwick, J. K., Lee, R. J., Lambert, Q. T., Symons, M., Pestell, R. G., Der, C. J., and Whitehead, I. P. (1998) J Biol Chem 273, Feig, L. A. (1999) Nat Cell Biol 1, E Minoshima, Y., Kawashima, T., Hirose, K., Tonozuka, Y., Kawajiri, A., Bao, Y. C., Deng, X., Tatsuka, M., Narumiya, S., May, W. S., Nosaka, T., Semba, K., Inoue, T., Satoh, T., Inagaki, M., and Kitamura, T. (2003) Dev Cell 4, Cheng, L., Rossman, K. L., Mahon, G. M., Worthylake, D. K., Korus, M., Sondek, J., and Whitehead, I. P. (2002) Mol Cell Biol 22, Saka, Y., Esashi, F., Matsusaka, T., Mochida, S., and Yanagida, M. (1997) Genes Dev 11,
34 FIGURE LEGENDS Figure 1. Induction of foci of transforming cells by ECT2 derivatives. A. Morphology of foci induced by ECT2 cdna. One mg each of vector alone (pcev29) or the expression vector for human full-length ECT2 (ECT2-F), human ECT2 lacking the N-terminal half (ECT2-DN5), mouse ect2 lacking the N-terminal half (ect2-t), H-ras V12 or c-sis was used to transfect NIH3T3 cells. Cells were stained by Giemsa and then photographed. B. Cell morphology of stable ECT2-F, N, DN5, and vector alone transfectants. C. Expression of FLAG-tagged ECT2-F, N and DN5 in stable transfectants. FLAG- ECT2 fusion proteins were detected by immunoblotting using anti-flag M2 monoclonal antibody. Figure 2. Characterization of NIH 3T3 cells transfected with ECT2 variants. A. Growth of ECT2 transfectants in low serum conditions. Cells were cultured in DMEM medium containing 1% serum, and viable cells were scored at the indicated time points. B. Induction of cell invasiveness by ECT2. Stable NIH 3T3 transfectants expressing the indicated cdna were used for Matrigel assays to estimate cell invasiveness in vitro. The cells invaded the matrigels were stained and photographed (left half). Number of the cells passed through the matrigel was summarized (right half). ect2- T is a mouse cdna with an N-terminal deletion similar to human ECT2-DN5. 34
35 Figure 3. Stimulation of MAP kinase activity and AP-1 or SRF-regulated transcriptional activation by ECT2. A. Effects of ECT2 derivative expression on MAP kinase activity in NIH 3T3 cells. MAP kinase activity in the stable transfectants expressing indicated plasmids (see Figure 1C) was measured using phosphospecific antibodies, and endogenous proteins were detected by specific antibodies (Cell Signaling Technology, Beverly, MA) according to the accompanied protocols. JNK activation was detected as phosphorylated c-jun at Ser-63, and activated Erk and p38 proteins were detected as their phosphorylated forms. B. Effects of ECT2 derivative expression on AP-1- or SRF-regulated transcription. COS cells were cotransfected with the indicated ECT2 expression vector and SRE- LUC (upper) or AP-1-LUC (lower) reporter plasmid. Activation of AP-1- or SRFregulated transcriptional activity was estimated by the luciferase activity. prk-tk- Renilla lucifirase vector was used to cotransfect the cells together with the reporter plasmid for an internal control to normalize the transfection/expression efficiency. Figure 4. Induction of cytoskeletal actin remodeling by oncogenic ECT2. A. Actin organization of ECT2 transfectants. NIH 3T3 were transfected with the GFP-ECT2-DN5 expression vector or GFP vector. Cells expressing GFP-ECT2-DN5 or GFP (indicated by arrows) were identified by green fluorescence and stained with rhodamine-conjugated phalloidin for F-actin. B. Time-lapse video recording of the morphological changes of NIH 3T3 cells upon oncogenic ECT2 transfection. NIH 3T3 fibroblasts were transiently transfected with GFP-actin and RFP-ECT2-DN5, and cells with green fluorescence were 35
36 photographed using time-lapse video microscopy. An arrow indicates one of the RFP-ECT2-DN5-expressing cells, which showed morphological changes. The number in each panel indicates time after transfection (hr). Bar, 10 mm. Figure 5. Inhibition of ECT2-DN5-induced transformation by dominant-negative Rho GTPases NIH 3T3 cells were cotransfected with ECT2-DN5 and indicated Rho expression vectors. Foci of morphologically transformed cells were scored 14 days after transfection, and expressed as percentages of the number of foci induced by ECT2- DN5 alone. Similar transfection efficiency of NIH 3T3 cells with the vectors used was confirmed by the formation of G418-resistant colonies. Shown is one of the results reproduced three times with an identical pattern. Figure 6. Mapping of the domains that regulate transforming activity of ECT2. A. Deletion mapping of domains that affect transforming activity. Regions carried by the ECT2 derivatives are shown by horizontal bars under a schematic representation of the human ECT2 protein. BRCT-1 and BRCT-2 indicate BRCA1 C- terminal repeats (7). The numbers at the ends of each clone represent the amino acid numbers relative to ECT2-F. DS contains an internal deletion of the S domain. DN5 DH - contains PVQR to AAAA substitutions at amino acids , whose location is also indicated by x. Transforming activity was shown as follows: -, < 1 x 10 0 ffu/pmol; +. 1 x 10 0 ~ 1 x 10 1 ffu/pmol; ++, 1 x 10 3 ~ 1 x 10 4 ffu/pmol; and +++, > 1 x 10 4 ffu/pmol. 36
37 B. Identification of exogenously expressed FLAG-ECT2 fusion proteins by immunoblotting. NIH 3T3 cells were transiently transfected with indicated FLAG-tagged ECT2 derivatives together with the GFP expression vector pegfp- C1. Forty-eight hours after transfection, cells were lysed and the proteins expressed from the vectors were analyzed by immunoblotting with anti-flag (M2), anti-gfp and anti-a tubulin antibodies. The GFP expression level was measured to monitor the transfection efficiency of each ECT2 variant and to detect the possible effect of ECT2 derivatives on protein expression. a-tubulin expression was measured as a loading control. Bands with expected sizes are indicated by dots at the right side. C. Subcellular localization of the FLAG-ECT2 fusion proteins. The fusion proteins (red) and nuclei (blue) were detected by anti-flag antibody and DAPI, respectively. Merged images are shown at the bottom. Figure 7. Functional comparison of ECT2 NLS and SV40 NLS. A. Subcellular localization of ECT2 NLS-tagged tandem GFP protein in U-2 OS cells. U-2 OS cells were transiently transfected with the indicated expression vectors using FuGENE 6 reagent, 24 hr after transfection. Hoechst dye was added to culture medium at a final concentration of 10 mm and cells were directly observed under the fluorescence microscope and photographed. B. Subcellular localization of SV40 NLS (nuc)-tagged ECT2- N5 in U-2 OS cells. U-2 OS and NIH3T3 (data not shown) cells were transiently transfected with the indicated expression vectors using FuGENE 6 and LipofectAMINE PLUS reagent, respectively. Hoechst dye was added to culture medium at a final 37
38 concentration of 10 mm 24 hr after transfection, and cells were directly observed under the fluorescence microscope and photographed. Figure 8. Interaction of the N and C-terminal domains of ECT2. A. Association of ECT2-N with ECT2- N5 or ECT2-DH. Lysates prepared from COS cells expressing indicated proteins were subjected to immunoprecipitation by anti-gfp or anti-flag antibodies, and blotted by anti-flag antibody. * indicates the location of immunoglobulin heavy chain. V, FLAG vector; F, FLAG-ECT2-F; N4, FLAG-ECT2-N4; N, FLAG-ECT2- N5; DH, FLAG-ECT2-DH (amino acids ). B. Inhibition of ECT2- N5 transforming activity in NIH3T3 cells by ECT2-N. NIH 3T3 cells were transfected with ECT2- N5 together with the indicated expression vectors in triplicate, and foci of transforming cells were scored 14 days after transfection. The number of foci induced by ECT2- N5 plus vector alone was normalized to 100 %. Similar experiments were performed using H-rasV12 as control. Shown are representative results reproduced twice with an identical pattern. Figure 9. Effects of mutations in the nuclear localization signals on the transforming activity of ECT2-DN4. NIH 3T3 cells were transfected with the indicated plasmids, and foci of morphologically transformed cells were scored 14 days after transfection. Figure 10. Identification of Rho GTPases activated by oncogenic ECT2. 38
39 A. COS cells were transfected with the indicated AU5-tagged Rho expression vectors together with the GFP vector (-) or GFP-ECT2-DN5 expression vector (+). The GTPbound forms of Rho GTPases were pulled down by GST-RBD (for RhoA, RhoB and RhoC) or GST-PBD (for Rac1 and Rac2) and detected by anti-au5 antibody. The amount of GST-RBD and GST-PBD in the reaction mixtures was also determined by protein staining. B. Effects of ECT2-N derivatives on ECT2-DN5-induced GTP-RhoA accumulation. COS cells were transfected with ECT2-DN5 (0.5 mg) and indicated ECT2 derivatives (+, 1.5 mg; ++, 3.5 mg). The GTP-bound forms of Rho GTPases were analyzed as in panel A. Similar results were reproduced three times. Figure 11. A model of ECT2 activation and malignant transformation. The catalytic domain (DH) of ECT2 is inhibited by the binding of the N-terminal domain. ECT2 cannot activate Rho GTPases as it is sequestered in the nucleus in interphase cells (inactive, nuclear). An ECT2 derivative lacking the N-terminal domain, but retaining the S domain, may be active, but it still cannot activate the Rho GTPases, as it localizes in the nucleus (active, nuclear). An ECT2 derivative having the DS deletion lacks two NLSs, and therefore becomes partially cytoplasmic (active, partially cytoplasmic). This allows the ECT2 derivative to activate Rho GTPases in the cytoplasm and thus causes morphological transformation of the cells (lower). 39











40 Figure 1. Saito et al. A B pcev29 ECT2- N5 H-ras ECT2-F mouse ect2-t sis C vector ECT2-F ECT2-N2 ECT2-DN5 vector ECT2-F 97.4 ECT2-F ECT2-N2 ECT2-DN Blot: Anti-FLAG ECT2-DN5 ECT2-N2
41 Figure 2. Saito et al. A 200 ECT2-DN5 B Viable cells (%) vector ECT2-DN5 vector ECT2-N2 ECT2-F (h) Number of invading cells/filter vector ECT2-F ECT2-DN5 ect2-t
42 Figure 3. Saito et al. A vector vector vector sorbitol anisomycin PDGF-BB ECT2-F ECT2-F ECT2-F ECT2-N2 ECT2-DN5 ECT2-N2 ECT2-N2 ECT2-DN5 ECT2-DN5 c-jun- P (Fold increase) JNK2 JNK1 p38 P p38- Erk1- P Erk2- P Erk1 Erk2 B Relative luciferase activity Relative luciferase activity vector vector ECT2-F ECT2-F ECT2-N2 ECT2-N2 +vector +RhoA N19 +vector +Rac1 N17 ECT2-DN5 +Rac1 N17 +RhoA N19 ECT2-DN5 +Cdc42 N17 RhoA L63 +Cdc42 N Rac1 L61 vector Cdc42 L61 RhoA L63 Rac1 L61 Cdc42 L61







43 Figure 4. Saito et al. A GFP vector GFP-ECT2-DN5 GFP F-actin B
44 Figure 5. Saito et al. 60 Relative number of foci RhoA WT RhoA N19 Rac1 WT Rac1 N17 Cdc42 WT Cdc42 N17


 ( N5 C1) - ( N5 C2) ( N5 C3) +++ ( N6) ( N8) - ( S) ++ (S1) + (S2) ++ (S3)")
45 Figure 6. Saito et al. A B C 1 N kda XR CC1 67 Cyclin Rad4/Cut5 B6 BRCT-1 BRCT-2 vector F 144 N1 N N3.. N10. S NLS NLS N4 N9 N5 N6.... N8. N5 C1 N5 C2 N5 C3 N5 DH -... S S1 S2 S Transforming Activity (F) ( N1) ( N2) - ( N3) ( N10) ( N4) - ( N9) ++ ( N5) +++ ( N5 DH - ) ( N5 C1) - ( N5 C2) ( N5 C3) +++ ( N6) ( N8) - ( S) ++ (S1) + (S2) ++ (S3) x x xx 454 x DH ECT2-F ECT2- N5 ECT2- S ECT2-S3 PH C NLS No plasmid - FLAG GFP a-tubulin FLAG DAPI merge













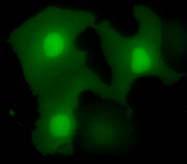

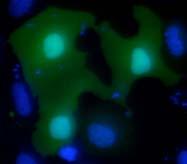


46 Figure 7. Saito et al. A GFP Hoechst merge GFP Tandem GFP B Tandem GFP- ECT2 NLS vector ECT2-F GFP Hoechst merge ECT2- N5 ECT2- N5 nuc


47 Figure 8. Saito et al. A FLAG- ECT2 + GFP-ECT2-N4 V FN4DNDH + GFP-ECT2-N4 FLAG- V FN4DNDH ECT2 B Ttransforming activity (%) IP: anti-gfp IB: anti-flag * IP: anti-flag IB: anti-flag * 0 vector ECT2-N1 ECT2-N3 vector ECT2-N1 ECT2-N3 ECT2- N5 H-rasV12
48 Figure 9. Saito et al. 600 Number of foci / plate vector ECT2-DN4 ECT2-DN4S1 ECT2-DN4S3 ECT2-DS ECT2-DN5

 Pull-down (anti-au5)")
")
")
49 Figure 10. Saito et al. A ECT2-DN5 RhoA RhoB RhoC Rac1 Rac Lysate (anti-au5) Pull-down (anti-au5) GFP-ECT2- N5 GFP GST-RBD or GST-PBD B Lysate (anti-rhoa) vector + ++ ECT2- N4 ECT2- N Pull-down (anti-rhoa) Myc-ECT2-N2 Myc-ECT2-N4 GST-RBD
Deregulation and Mislocalization of the Cytokinesis Regulator ECT2 Activate the Rho Signaling Pathways Leading to Malignant Transformation*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 8, Issue of February 20, pp. 7169 7179, 2004 Printed in U.S.A. Deregulation and Mislocalization of the Cytokinesis Regulator ECT2 Activate the Rho Signaling
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 8, Issue of February 20, pp. 7169 7179, 2004 Printed in U.S.A. Deregulation and Mislocalization of the Cytokinesis Regulator ECT2 Activate the Rho Signaling
SUPPORTING INFORMATION FOR. SEquence-Enabled Reassembly of β-lactamase (SEER-LAC): a Sensitive Method for the Detection of Double-Stranded DNA
: a Sensitive Method for the Detection of Double-Stranded DNA") SUPPORTING INFORMATION FOR SEquence-Enabled Reassembly of β-lactamase (SEER-LAC): a Sensitive Method for the Detection of Double-Stranded DNA Aik T. Ooi, Cliff I. Stains, Indraneel Ghosh *, David J. Segal
SUPPORTING INFORMATION FOR SEquence-Enabled Reassembly of β-lactamase (SEER-LAC): a Sensitive Method for the Detection of Double-Stranded DNA Aik T. Ooi, Cliff I. Stains, Indraneel Ghosh *, David J. Segal
Practical Bioinformatics
 5/2/2017 Dictionaries d i c t i o n a r y = { A : T, T : A, G : C, C : G } d i c t i o n a r y [ G ] d i c t i o n a r y [ N ] = N d i c t i o n a r y. h a s k e y ( C ) Dictionaries g e n e t i c C o
5/2/2017 Dictionaries d i c t i o n a r y = { A : T, T : A, G : C, C : G } d i c t i o n a r y [ G ] d i c t i o n a r y [ N ] = N d i c t i o n a r y. h a s k e y ( C ) Dictionaries g e n e t i c C o
SUPPLEMENTARY DATA - 1 -
 - 1 - SUPPLEMENTARY DATA Construction of B. subtilis rnpb complementation plasmids For complementation, the B. subtilis rnpb wild-type gene (rnpbwt) under control of its native rnpb promoter and terminator
- 1 - SUPPLEMENTARY DATA Construction of B. subtilis rnpb complementation plasmids For complementation, the B. subtilis rnpb wild-type gene (rnpbwt) under control of its native rnpb promoter and terminator
SEQUENCE ALIGNMENT BACKGROUND: BIOINFORMATICS. Prokaryotes and Eukaryotes. DNA and RNA
 SEQUENCE ALIGNMENT BACKGROUND: BIOINFORMATICS 1 Prokaryotes and Eukaryotes 2 DNA and RNA 3 4 Double helix structure Codons Codons are triplets of bases from the RNA sequence. Each triplet defines an amino-acid.
SEQUENCE ALIGNMENT BACKGROUND: BIOINFORMATICS 1 Prokaryotes and Eukaryotes 2 DNA and RNA 3 4 Double helix structure Codons Codons are triplets of bases from the RNA sequence. Each triplet defines an amino-acid.
Supplemental Figure 1.
 A wt spoiiiaδ spoiiiahδ bofaδ B C D E spoiiiaδ, bofaδ Supplemental Figure 1. GFP-SpoIVFA is more mislocalized in the absence of both BofA and SpoIIIAH. Sporulation was induced by resuspension in wild-type
A wt spoiiiaδ spoiiiahδ bofaδ B C D E spoiiiaδ, bofaδ Supplemental Figure 1. GFP-SpoIVFA is more mislocalized in the absence of both BofA and SpoIIIAH. Sporulation was induced by resuspension in wild-type
Advanced topics in bioinformatics
 Feinberg Graduate School of the Weizmann Institute of Science Advanced topics in bioinformatics Shmuel Pietrokovski & Eitan Rubin Spring 2003 Course WWW site: http://bioinformatics.weizmann.ac.il/courses/atib
Feinberg Graduate School of the Weizmann Institute of Science Advanced topics in bioinformatics Shmuel Pietrokovski & Eitan Rubin Spring 2003 Course WWW site: http://bioinformatics.weizmann.ac.il/courses/atib
Number-controlled spatial arrangement of gold nanoparticles with
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2016 Number-controlled spatial arrangement of gold nanoparticles with DNA dendrimers Ping Chen,*
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2016 Number-controlled spatial arrangement of gold nanoparticles with DNA dendrimers Ping Chen,*
Supplemental data. Pommerrenig et al. (2011). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure 1. Prediction of phloem-specific MTK1 expression in Arabidopsis shoots and roots. The images and the corresponding numbers showing absolute (A) or relative expression levels (B) of
Supplemental Figure 1. Prediction of phloem-specific MTK1 expression in Arabidopsis shoots and roots. The images and the corresponding numbers showing absolute (A) or relative expression levels (B) of
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Zn 2+ -binding sites in USP18. (a) The two molecules of USP18 present in the asymmetric unit are shown. Chain A is shown in blue, chain B in green. Bound Zn 2+ ions are shown as
Supplementary Figure 1 Zn 2+ -binding sites in USP18. (a) The two molecules of USP18 present in the asymmetric unit are shown. Chain A is shown in blue, chain B in green. Bound Zn 2+ ions are shown as
Oligomerization of DH Domain Is Essential for Dbl-Induced Transformation
 MOLECULAR AND CELLULAR BIOLOGY, Jan. 2001, p. 425 437 Vol. 21, No. 2 0270-7306/01/$04.00 0 DOI: 10.1128/MCB.21.2.425 437.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Oligomerization
MOLECULAR AND CELLULAR BIOLOGY, Jan. 2001, p. 425 437 Vol. 21, No. 2 0270-7306/01/$04.00 0 DOI: 10.1128/MCB.21.2.425 437.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Oligomerization
Supporting Information
 Supporting Information T. Pellegrino 1,2,3,#, R. A. Sperling 1,#, A. P. Alivisatos 2, W. J. Parak 1,2,* 1 Center for Nanoscience, Ludwig Maximilians Universität München, München, Germany 2 Department of
Supporting Information T. Pellegrino 1,2,3,#, R. A. Sperling 1,#, A. P. Alivisatos 2, W. J. Parak 1,2,* 1 Center for Nanoscience, Ludwig Maximilians Universität München, München, Germany 2 Department of
Supplemental Table 1. Primers used for cloning and PCR amplification in this study
 Supplemental Table 1. Primers used for cloning and PCR amplification in this study Target Gene Primer sequence NATA1 (At2g393) forward GGG GAC AAG TTT GTA CAA AAA AGC AGG CTT CAT GGC GCC TCC AAC CGC AGC
Supplemental Table 1. Primers used for cloning and PCR amplification in this study Target Gene Primer sequence NATA1 (At2g393) forward GGG GAC AAG TTT GTA CAA AAA AGC AGG CTT CAT GGC GCC TCC AAC CGC AGC
High throughput near infrared screening discovers DNA-templated silver clusters with peak fluorescence beyond 950 nm
 Electronic Supplementary Material (ESI) for Nanoscale. This journal is The Royal Society of Chemistry 2018 High throughput near infrared screening discovers DNA-templated silver clusters with peak fluorescence
Electronic Supplementary Material (ESI) for Nanoscale. This journal is The Royal Society of Chemistry 2018 High throughput near infrared screening discovers DNA-templated silver clusters with peak fluorescence
Crick s early Hypothesis Revisited
 Crick s early Hypothesis Revisited Or The Existence of a Universal Coding Frame Ryan Rossi, Jean-Louis Lassez and Axel Bernal UPenn Center for Bioinformatics BIOINFORMATICS The application of computer
Crick s early Hypothesis Revisited Or The Existence of a Universal Coding Frame Ryan Rossi, Jean-Louis Lassez and Axel Bernal UPenn Center for Bioinformatics BIOINFORMATICS The application of computer
Activation of a receptor. Assembly of the complex
 Activation of a receptor ligand inactive, monomeric active, dimeric When activated by growth factor binding, the growth factor receptor tyrosine kinase phosphorylates the neighboring receptor. Assembly
Activation of a receptor ligand inactive, monomeric active, dimeric When activated by growth factor binding, the growth factor receptor tyrosine kinase phosphorylates the neighboring receptor. Assembly
Clay Carter. Department of Biology. QuickTime and a TIFF (Uncompressed) decompressor are needed to see this picture.
 decompressor are needed to see this picture.") QuickTime and a TIFF (Uncompressed) decompressor are needed to see this picture. Clay Carter Department of Biology QuickTime and a TIFF (LZW) decompressor are needed to see this picture. Ornamental tobacco
QuickTime and a TIFF (Uncompressed) decompressor are needed to see this picture. Clay Carter Department of Biology QuickTime and a TIFF (LZW) decompressor are needed to see this picture. Ornamental tobacco
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
SSR ( ) Vol. 48 No ( Microsatellite marker) ( Simple sequence repeat,ssr),
 Vol. 48 No ( Microsatellite marker) ( Simple sequence repeat,ssr),") 48 3 () Vol. 48 No. 3 2009 5 Journal of Xiamen University (Nat ural Science) May 2009 SSR,,,, 3 (, 361005) : SSR. 21 516,410. 60 %96. 7 %. (),(Between2groups linkage method),.,, 11 (),. 12,. (, ), : 0.
48 3 () Vol. 48 No. 3 2009 5 Journal of Xiamen University (Nat ural Science) May 2009 SSR,,,, 3 (, 361005) : SSR. 21 516,410. 60 %96. 7 %. (),(Between2groups linkage method),.,, 11 (),. 12,. (, ), : 0.
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Supplementary Information for
 Supplementary Information for Evolutionary conservation of codon optimality reveals hidden signatures of co-translational folding Sebastian Pechmann & Judith Frydman Department of Biology and BioX, Stanford
Supplementary Information for Evolutionary conservation of codon optimality reveals hidden signatures of co-translational folding Sebastian Pechmann & Judith Frydman Department of Biology and BioX, Stanford
Characterization of Pathogenic Genes through Condensed Matrix Method, Case Study through Bacterial Zeta Toxin
 International Journal of Genetic Engineering and Biotechnology. ISSN 0974-3073 Volume 2, Number 1 (2011), pp. 109-114 International Research Publication House http://www.irphouse.com Characterization of
International Journal of Genetic Engineering and Biotechnology. ISSN 0974-3073 Volume 2, Number 1 (2011), pp. 109-114 International Research Publication House http://www.irphouse.com Characterization of
Supplementary Information
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Directed self-assembly of genomic sequences into monomeric and polymeric branched DNA structures
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Directed self-assembly of genomic sequences into monomeric and polymeric branched DNA structures
Building a Multifunctional Aptamer-Based DNA Nanoassembly for Targeted Cancer Therapy
 Supporting Information Building a Multifunctional Aptamer-Based DNA Nanoassembly for Targeted Cancer Therapy Cuichen Wu,, Da Han,, Tao Chen,, Lu Peng, Guizhi Zhu,, Mingxu You,, Liping Qiu,, Kwame Sefah,
Supporting Information Building a Multifunctional Aptamer-Based DNA Nanoassembly for Targeted Cancer Therapy Cuichen Wu,, Da Han,, Tao Chen,, Lu Peng, Guizhi Zhu,, Mingxu You,, Liping Qiu,, Kwame Sefah,
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
NSCI Basic Properties of Life and The Biochemistry of Life on Earth
 NSCI 314 LIFE IN THE COSMOS 4 Basic Properties of Life and The Biochemistry of Life on Earth Dr. Karen Kolehmainen Department of Physics CSUSB http://physics.csusb.edu/~karen/ WHAT IS LIFE? HARD TO DEFINE,
NSCI 314 LIFE IN THE COSMOS 4 Basic Properties of Life and The Biochemistry of Life on Earth Dr. Karen Kolehmainen Department of Physics CSUSB http://physics.csusb.edu/~karen/ WHAT IS LIFE? HARD TO DEFINE,
Regulatory Sequence Analysis. Sequence models (Bernoulli and Markov models)
") Regulatory Sequence Analysis Sequence models (Bernoulli and Markov models) 1 Why do we need random models? Any pattern discovery relies on an underlying model to estimate the random expectation. This model
Regulatory Sequence Analysis Sequence models (Bernoulli and Markov models) 1 Why do we need random models? Any pattern discovery relies on an underlying model to estimate the random expectation. This model
ydci GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC
 Table S1. DNA primers used in this study. Name ydci P1ydcIkd3 Sequence GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC Kd3ydcIp2 lacz fusion YdcIendP1 YdcItrgP2 GAC AGC
Table S1. DNA primers used in this study. Name ydci P1ydcIkd3 Sequence GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC Kd3ydcIp2 lacz fusion YdcIendP1 YdcItrgP2 GAC AGC
Supporting Information for. Initial Biochemical and Functional Evaluation of Murine Calprotectin Reveals Ca(II)-
-") Supporting Information for Initial Biochemical and Functional Evaluation of Murine Calprotectin Reveals Ca(II)- Dependence and Its Ability to Chelate Multiple Nutrient Transition Metal Ions Rose C. Hadley,
Supporting Information for Initial Biochemical and Functional Evaluation of Murine Calprotectin Reveals Ca(II)- Dependence and Its Ability to Chelate Multiple Nutrient Transition Metal Ions Rose C. Hadley,
Richik N. Ghosh, Linnette Grove, and Oleg Lapets ASSAY and Drug Development Technologies 2004, 2:
 1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
TM1 TM2 TM3 TM4 TM5 TM6 TM bp
 a 467 bp 1 482 2 93 3 321 4 7 281 6 21 7 66 8 176 19 12 13 212 113 16 8 b ATG TCA GGA CAT GTA ATG GAG GAA TGT GTA GTT CAC GGT ACG TTA GCG GCA GTA TTG CGT TTA ATG GGC GTA GTG M S G H V M E E C V V H G T
a 467 bp 1 482 2 93 3 321 4 7 281 6 21 7 66 8 176 19 12 13 212 113 16 8 b ATG TCA GGA CAT GTA ATG GAG GAA TGT GTA GTT CAC GGT ACG TTA GCG GCA GTA TTG CGT TTA ATG GGC GTA GTG M S G H V M E E C V V H G T
Table S1. Primers and PCR conditions used in this paper Primers Sequence (5 3 ) Thermal conditions Reference Rhizobacteria 27F 1492R
 Thermal conditions Reference Rhizobacteria 27F 1492R") Table S1. Primers and PCR conditions used in this paper Primers Sequence (5 3 ) Thermal conditions Reference Rhizobacteria 27F 1492R AAC MGG ATT AGA TAC CCK G GGY TAC CTT GTT ACG ACT T Detection of Candidatus
Table S1. Primers and PCR conditions used in this paper Primers Sequence (5 3 ) Thermal conditions Reference Rhizobacteria 27F 1492R AAC MGG ATT AGA TAC CCK G GGY TAC CTT GTT ACG ACT T Detection of Candidatus
Electronic supplementary material
 Applied Microbiology and Biotechnology Electronic supplementary material A family of AA9 lytic polysaccharide monooxygenases in Aspergillus nidulans is differentially regulated by multiple substrates and
Applied Microbiology and Biotechnology Electronic supplementary material A family of AA9 lytic polysaccharide monooxygenases in Aspergillus nidulans is differentially regulated by multiple substrates and
CELL CYCLE AND DIFFERENTIATION
 CELL CYCLE AND DIFFERENTIATION Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id WHAT IS CELL CYCLE? 09/12/14 d.purnomosari@ugm.ac.id
CELL CYCLE AND DIFFERENTIATION Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id WHAT IS CELL CYCLE? 09/12/14 d.purnomosari@ugm.ac.id
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
SUPPLEMENTARY INFORMATION
 DOI:.8/NCHEM. Conditionally Fluorescent Molecular Probes for Detecting Single Base Changes in Double-stranded DNA Sherry Xi Chen, David Yu Zhang, Georg Seelig. Analytic framework and probe design.. Design
DOI:.8/NCHEM. Conditionally Fluorescent Molecular Probes for Detecting Single Base Changes in Double-stranded DNA Sherry Xi Chen, David Yu Zhang, Georg Seelig. Analytic framework and probe design.. Design
5- Semaphorin-Plexin-Neuropilin
 5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
Analysis and Simulation of Biological Systems
 Analysis and Simulation of Biological Systems Dr. Carlo Cosentino School of Computer and Biomedical Engineering Department of Experimental and Clinical Medicine Università degli Studi Magna Graecia Catanzaro,
Analysis and Simulation of Biological Systems Dr. Carlo Cosentino School of Computer and Biomedical Engineering Department of Experimental and Clinical Medicine Università degli Studi Magna Graecia Catanzaro,
Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter
 Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter 9/10/2008 1 Learning Objectives Explain why a cell cycle was selected for during evolution
Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter 9/10/2008 1 Learning Objectives Explain why a cell cycle was selected for during evolution
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A)
 M-phase nuclei are visualized by a-ph3 labeling in wild-type (A)") Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A) and mutant (B) 4 dpf retinae. The central retina of the
Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A) and mutant (B) 4 dpf retinae. The central retina of the
Mechanisms of Cell Proliferation
 Mechanisms of Cell Proliferation Cell Cycle G 2 S G 1 Multi-cellular organisms depend on cell division/proliferation; Each organism has a developmental plan that determines its behavior and properties;
Mechanisms of Cell Proliferation Cell Cycle G 2 S G 1 Multi-cellular organisms depend on cell division/proliferation; Each organism has a developmental plan that determines its behavior and properties;
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
3.a.2- Cell Cycle and Meiosis
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. 3.a.2- Cell Cycle and Meiosis EU 3.A: Heritable information provides for continuity of life.
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. 3.a.2- Cell Cycle and Meiosis EU 3.A: Heritable information provides for continuity of life.
Ras e la Cascata delle Piccole GTPasi
 Roma 26-27 Giugno 2015 Multidisciplinarietà e Biologia Molecolare applicate alla pratica Clinica i Oncologica: un'opportunità ed un vantaggio per tuttitti Ras e la Cascata delle Piccole GTPasi Alvaro Leone
Roma 26-27 Giugno 2015 Multidisciplinarietà e Biologia Molecolare applicate alla pratica Clinica i Oncologica: un'opportunità ed un vantaggio per tuttitti Ras e la Cascata delle Piccole GTPasi Alvaro Leone
4) Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.
 Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.") Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Protein Threading. Combinatorial optimization approach. Stefan Balev.
 Protein Threading Combinatorial optimization approach Stefan Balev Stefan.Balev@univ-lehavre.fr Laboratoire d informatique du Havre Université du Havre Stefan Balev Cours DEA 30/01/2004 p.1/42 Outline
Protein Threading Combinatorial optimization approach Stefan Balev Stefan.Balev@univ-lehavre.fr Laboratoire d informatique du Havre Université du Havre Stefan Balev Cours DEA 30/01/2004 p.1/42 Outline
The role of the FliD C-terminal domain in pentamer formation and
 The role of the FliD C-terminal domain in pentamer formation and interaction with FliT Hee Jung Kim 1,2,*, Woongjae Yoo 3,*, Kyeong Sik Jin 4, Sangryeol Ryu 3,5 & Hyung Ho Lee 1, 1 Department of Chemistry,
The role of the FliD C-terminal domain in pentamer formation and interaction with FliT Hee Jung Kim 1,2,*, Woongjae Yoo 3,*, Kyeong Sik Jin 4, Sangryeol Ryu 3,5 & Hyung Ho Lee 1, 1 Department of Chemistry,
evoglow - express N kit distributed by Cat.#: FP product information broad host range vectors - gram negative bacteria
 evoglow - express N kit broad host range vectors - gram negative bacteria product information distributed by Cat.#: FP-21020 Content: Product Overview... 3 evoglow express N -kit... 3 The evoglow -Fluorescent
evoglow - express N kit broad host range vectors - gram negative bacteria product information distributed by Cat.#: FP-21020 Content: Product Overview... 3 evoglow express N -kit... 3 The evoglow -Fluorescent
WD Repeat Domain of Dictyostelium Myosin Heavy Chain Kinase C Functions in both Substrate Targeting and Cellular Localization,
 WD Repeat Domain of Dictyostelium Myosin Heavy Chain Kinase C Functions in both Substrate Targeting and Cellular Localization, By: Atiya Franklin, Linzi Hyatt, Alyssa Chowdhury, and Paul A. Steimle* Franklin,
WD Repeat Domain of Dictyostelium Myosin Heavy Chain Kinase C Functions in both Substrate Targeting and Cellular Localization, By: Atiya Franklin, Linzi Hyatt, Alyssa Chowdhury, and Paul A. Steimle* Franklin,
RhoGAP assay kit (Cat. # BK105)
") RhoGAP assay kit (Cat. # BK105) The Ras superfamily of small GTPases (such as Ras, Rho, Rab, Arf and Ran proteins) serve as binary switches cycling between a GDP-bound OFF state and a GTP-bound ON state,
RhoGAP assay kit (Cat. # BK105) The Ras superfamily of small GTPases (such as Ras, Rho, Rab, Arf and Ran proteins) serve as binary switches cycling between a GDP-bound OFF state and a GTP-bound ON state,
Mechanisms of Cell Proliferation
 Mechanisms of Cell Proliferation Cell Cycle G 2 S G 1 Multi-cellular organisms depend on cell division/proliferation; Each organism has a developmental plan that determines its behavior and properties;
Mechanisms of Cell Proliferation Cell Cycle G 2 S G 1 Multi-cellular organisms depend on cell division/proliferation; Each organism has a developmental plan that determines its behavior and properties;
Modelling and Analysis in Bioinformatics. Lecture 1: Genomic k-mer Statistics
 582746 Modelling and Analysis in Bioinformatics Lecture 1: Genomic k-mer Statistics Juha Kärkkäinen 06.09.2016 Outline Course introduction Genomic k-mers 1-Mers 2-Mers 3-Mers k-mers for Larger k Outline
582746 Modelling and Analysis in Bioinformatics Lecture 1: Genomic k-mer Statistics Juha Kärkkäinen 06.09.2016 Outline Course introduction Genomic k-mers 1-Mers 2-Mers 3-Mers k-mers for Larger k Outline
Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells
 Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells Understand how a simple biochemical oscillator can drive the
Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells Understand how a simple biochemical oscillator can drive the
Reading Assignments. A. Systems of Cell Division. Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question.
 Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) The receptors for a group of signaling molecules known as growth factors are
Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) The receptors for a group of signaling molecules known as growth factors are
Direct detection of antibodies in blood plasma using bioluminescent
 Supplementary information Direct detection of antibodies in blood plasma using bioluminescent sensor proteins and a smartphone. Remco Arts, Ilona den Hartog, Stefan Zijlema, Vito Thijssen, Stan van der
Supplementary information Direct detection of antibodies in blood plasma using bioluminescent sensor proteins and a smartphone. Remco Arts, Ilona den Hartog, Stefan Zijlema, Vito Thijssen, Stan van der
evoglow - express N kit Cat. No.: product information broad host range vectors - gram negative bacteria
 evoglow - express N kit broad host range vectors - gram negative bacteria product information Cat. No.: 2.1.020 evocatal GmbH 2 Content: Product Overview... 4 evoglow express N kit... 4 The evoglow Fluorescent
evoglow - express N kit broad host range vectors - gram negative bacteria product information Cat. No.: 2.1.020 evocatal GmbH 2 Content: Product Overview... 4 evoglow express N kit... 4 The evoglow Fluorescent
Describe the process of cell division in prokaryotic cells. The Cell Cycle
 The Cell Cycle Objective # 1 In this topic we will examine the cell cycle, the series of changes that a cell goes through from one division to the next. We will pay particular attention to how the genetic
The Cell Cycle Objective # 1 In this topic we will examine the cell cycle, the series of changes that a cell goes through from one division to the next. We will pay particular attention to how the genetic
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
BIOLOGY. Chapter 10 CELL REPRODUCTION PowerPoint Image Slideshow
 BIOLOGY Chapter 10 CELL REPRODUCTION PowerPoint Image Slideshow FIGURE 10.1 A sea urchin begins life as a single cell that (a) divides to form two cells, visible by scanning electron microscopy. After
BIOLOGY Chapter 10 CELL REPRODUCTION PowerPoint Image Slideshow FIGURE 10.1 A sea urchin begins life as a single cell that (a) divides to form two cells, visible by scanning electron microscopy. After
3. Evolution makes sense of homologies. 3. Evolution makes sense of homologies. 3. Evolution makes sense of homologies
 Richard Owen (1848) introduced the term Homology to refer to structural similarities among organisms. To Owen, these similarities indicated that organisms were created following a common plan or archetype.
Richard Owen (1848) introduced the term Homology to refer to structural similarities among organisms. To Owen, these similarities indicated that organisms were created following a common plan or archetype.
Three different fusions led to three basic ideas: 1) If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes
 If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes") Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
6.047 / Computational Biology: Genomes, Networks, Evolution Fall 2008
 MIT OpenCourseWare http://ocw.mit.edu 6.047 / 6.878 Computational Biology: Genomes, Networks, Evolution Fall 2008 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
MIT OpenCourseWare http://ocw.mit.edu 6.047 / 6.878 Computational Biology: Genomes, Networks, Evolution Fall 2008 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
Cell Division (Outline)
") Cell Division (Outline) 1. Overview of purpose and roles. Comparison of prokaryotic and eukaryotic chromosomes and relation between organelles and cell division. 2. Eukaryotic cell reproduction: asexual
Cell Division (Outline) 1. Overview of purpose and roles. Comparison of prokaryotic and eukaryotic chromosomes and relation between organelles and cell division. 2. Eukaryotic cell reproduction: asexual
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION DOI:.38/NCHEM.246 Optimizing the specificity of nucleic acid hyridization David Yu Zhang, Sherry Xi Chen, and Peng Yin. Analytic framework and proe design 3.. Concentration-adjusted
SUPPLEMENTARY INFORMATION DOI:.38/NCHEM.246 Optimizing the specificity of nucleic acid hyridization David Yu Zhang, Sherry Xi Chen, and Peng Yin. Analytic framework and proe design 3.. Concentration-adjusted
Research Paper 629. Address: Institut de Génétique Moléculaire, C.N.R.S. UMR5535, 1919 route de Mende, F Montpellier cedex 5, France.
 Research Paper 629 The small GTPases Cdc42Hs, Rac1 and RhoG delineate Rafindependent pathways that cooperate to transform NIH3T3 cells Pierre Roux, Cécile Gauthier-Rouvière, Sandrine Doucet-Brutin and
Research Paper 629 The small GTPases Cdc42Hs, Rac1 and RhoG delineate Rafindependent pathways that cooperate to transform NIH3T3 cells Pierre Roux, Cécile Gauthier-Rouvière, Sandrine Doucet-Brutin and
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
RANK. Alternative names. Discovery. Structure. William J. Boyle* SUMMARY BACKGROUND
 RANK William J. Boyle* Department of Cell Biology, Amgen, Inc., One Amgen Center Drive, Thousand Oaks, CA 91320-1799, USA * corresponding author tel: 805-447-4304, fax: 805-447-1982, e-mail: bboyle@amgen.com
RANK William J. Boyle* Department of Cell Biology, Amgen, Inc., One Amgen Center Drive, Thousand Oaks, CA 91320-1799, USA * corresponding author tel: 805-447-4304, fax: 805-447-1982, e-mail: bboyle@amgen.com
Role of tyrosine kinases and the tyrosine phosphatase SptP in the interaction of Salmonella with host cells
 Cellular Microbiology (2001) 3(12), 795±810 Role of tyrosine kinases and the tyrosine phosphatase SptP in the interaction of Salmonella with host cells Sumati Murli, Robert O. Watson and Jorge E. GalaÂn*
Cellular Microbiology (2001) 3(12), 795±810 Role of tyrosine kinases and the tyrosine phosphatase SptP in the interaction of Salmonella with host cells Sumati Murli, Robert O. Watson and Jorge E. GalaÂn*
Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Division Read Chapter 19 pages 651-663 663 only (Benefits of Sex & Meiosis sections these are in Chapter
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Division Read Chapter 19 pages 651-663 663 only (Benefits of Sex & Meiosis sections these are in Chapter
The role of Rho GTPases in the regulation of the rearrangement of actin cytoskeleton and cell movement
 EXPERIMENTAL and MOLECULAR MEDICINE, Vol. 36, No. 4, 358-366, August 2004 The role of Rho GTPases in the regulation of the rearrangement of actin cytoskeleton and cell movement Rokeya Begum 1,3, M.S.A.
EXPERIMENTAL and MOLECULAR MEDICINE, Vol. 36, No. 4, 358-366, August 2004 The role of Rho GTPases in the regulation of the rearrangement of actin cytoskeleton and cell movement Rokeya Begum 1,3, M.S.A.
A. Incorrect! The Cell Cycle contains 4 distinct phases: (1) G 1, (2) S Phase, (3) G 2 and (4) M Phase.
 G 1, (2) S Phase, (3) G 2 and (4) M Phase.") Molecular Cell Biology - Problem Drill 21: Cell Cycle and Cell Death Question No. 1 of 10 1. Which of the following statements about the cell cycle is correct? Question #1 (A) The Cell Cycle contains 3
Molecular Cell Biology - Problem Drill 21: Cell Cycle and Cell Death Question No. 1 of 10 1. Which of the following statements about the cell cycle is correct? Question #1 (A) The Cell Cycle contains 3
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016
 Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
targets. clustering show that different complex pathway
 Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Neurite initiation. Neurite formation begins with a bud that sprouts from the cell body. One or several neurites can sprout at a time.
 Neurite initiation. Neuronal maturation initiation f-actin polarization and maturation tubulin stage 1: "spherical" neuron stage 2: neurons extend several neurites stage 3: one neurite accelerates its
Neurite initiation. Neuronal maturation initiation f-actin polarization and maturation tubulin stage 1: "spherical" neuron stage 2: neurons extend several neurites stage 3: one neurite accelerates its
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Analysis of correlated mutations in Ras G-domain
 www.bioinformation.net Volume 13(6) Hypothesis Analysis of correlated mutations in Ras G-domain Ekta Pathak * Bioinformatics Department, MMV, Banaras Hindu University. Ekta Pathak - E-mail: ektavpathak@gmail.com;
www.bioinformation.net Volume 13(6) Hypothesis Analysis of correlated mutations in Ras G-domain Ekta Pathak * Bioinformatics Department, MMV, Banaras Hindu University. Ekta Pathak - E-mail: ektavpathak@gmail.com;
Transport between cytosol and nucleus
 of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis.
 Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis. The role of kinases and cyclin in the regulation of the cell cycle.
Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis. The role of kinases and cyclin in the regulation of the cell cycle.
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins
 13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
5.1 Cell Division and the Cell Cycle
 5.1 Cell Division and the Cell Cycle Lesson Objectives Contrast cell division in prokaryotes and eukaryotes. Identify the phases of the eukaryotic cell cycle. Explain how the cell cycle is controlled.
5.1 Cell Division and the Cell Cycle Lesson Objectives Contrast cell division in prokaryotes and eukaryotes. Identify the phases of the eukaryotic cell cycle. Explain how the cell cycle is controlled.
CCHS 2015_2016 Biology Fall Semester Exam Review
 Biomolecule General Knowledge Macromolecule Monomer (building block) Function Energy Storage Structure 1. What type of biomolecule is hair, skin, and nails? 2. What is the polymer of a nucleotide? 3. Which
Biomolecule General Knowledge Macromolecule Monomer (building block) Function Energy Storage Structure 1. What type of biomolecule is hair, skin, and nails? 2. What is the polymer of a nucleotide? 3. Which
From DNA to protein, i.e. the central dogma
 From DNA to protein, i.e. the central dogma DNA RNA Protein Biochemistry, chapters1 5 and Chapters 29 31. Chapters 2 5 and 29 31 will be covered more in detail in other lectures. ph, chapter 1, will be
From DNA to protein, i.e. the central dogma DNA RNA Protein Biochemistry, chapters1 5 and Chapters 29 31. Chapters 2 5 and 29 31 will be covered more in detail in other lectures. ph, chapter 1, will be
1- Below is a list of cell cycle phases matched with specific processes. Choose the correct pairing:
 Name: NetID: Exam 4 - Version 2 November 13, 2018 Dr. A. Pimentel Instructions: 1- Select the BEST answer for each question 2- Use pencil to mark your responses in the answer sheet. 3- You can mark your
Name: NetID: Exam 4 - Version 2 November 13, 2018 Dr. A. Pimentel Instructions: 1- Select the BEST answer for each question 2- Use pencil to mark your responses in the answer sheet. 3- You can mark your
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
CHAPTER 12 - THE CELL CYCLE (pgs )
") CHAPTER 12 - THE CELL CYCLE (pgs. 228-245) CHAPTER SEVEN TARGETS I. Describe the importance of mitosis in single-celled and multi-cellular organisms. II. Explain the organization of DNA molecules and their
CHAPTER 12 - THE CELL CYCLE (pgs. 228-245) CHAPTER SEVEN TARGETS I. Describe the importance of mitosis in single-celled and multi-cellular organisms. II. Explain the organization of DNA molecules and their
Evolvable Neural Networks for Time Series Prediction with Adaptive Learning Interval
 Evolvable Neural Networs for Time Series Prediction with Adaptive Learning Interval Dong-Woo Lee *, Seong G. Kong *, and Kwee-Bo Sim ** *Department of Electrical and Computer Engineering, The University
Evolvable Neural Networs for Time Series Prediction with Adaptive Learning Interval Dong-Woo Lee *, Seong G. Kong *, and Kwee-Bo Sim ** *Department of Electrical and Computer Engineering, The University
Answer Key. Cell Growth and Division
 Cell Growth and Division Answer Key SECTION 1. THE CELL CYCLE Cell Cycle: (1) Gap1 (G 1): cells grow, carry out normal functions, and copy their organelles. (2) Synthesis (S): cells replicate DNA. (3)
Cell Growth and Division Answer Key SECTION 1. THE CELL CYCLE Cell Cycle: (1) Gap1 (G 1): cells grow, carry out normal functions, and copy their organelles. (2) Synthesis (S): cells replicate DNA. (3)
Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition
 Supplementary Information to Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition Nadine Czudnochowski 1,2, *, Christian A. Bösken 1, * & Matthias Geyer 1 1 Max-Planck-Institut
Supplementary Information to Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition Nadine Czudnochowski 1,2, *, Christian A. Bösken 1, * & Matthias Geyer 1 1 Max-Planck-Institut
Supplemental Figure 1. Phenotype of ProRGA:RGAd17 plants under long day
 Supplemental Figure 1. Phenotype of ProRGA:RGAd17 plants under long day conditions. Photo was taken when the wild type plant started to bolt. Scale bar represents 1 cm. Supplemental Figure 2. Flowering
Supplemental Figure 1. Phenotype of ProRGA:RGAd17 plants under long day conditions. Photo was taken when the wild type plant started to bolt. Scale bar represents 1 cm. Supplemental Figure 2. Flowering
Biology: Life on Earth
 Biology: Life on Earth Eighth Edition Lecture for Chapter 11 The Continuity of Life: Cellular Reproduction Cellular Reproduction Intracellular activity between one cell division to the next is the cell
Biology: Life on Earth Eighth Edition Lecture for Chapter 11 The Continuity of Life: Cellular Reproduction Cellular Reproduction Intracellular activity between one cell division to the next is the cell
CCHS 2016_2017 Biology Fall Semester Exam Review
 CCHS 2016_2017 Biology Fall Semester Exam Review Biomolecule General Knowledge Macromolecule Monomer (building block) Function Structure 1. What type of biomolecule is hair, skin, and nails? Energy Storage
CCHS 2016_2017 Biology Fall Semester Exam Review Biomolecule General Knowledge Macromolecule Monomer (building block) Function Structure 1. What type of biomolecule is hair, skin, and nails? Energy Storage
THE CELL CYCLE & MITOSIS. Asexual Reproduction: Production of genetically identical offspring from a single parent.
 THE CELL CYCLE & MITOSIS Asexual Reproduction: Production of genetically identical offspring from a single parent. Sexual Reproduction: The fusion of two separate parent cells that produce offspring with
THE CELL CYCLE & MITOSIS Asexual Reproduction: Production of genetically identical offspring from a single parent. Sexual Reproduction: The fusion of two separate parent cells that produce offspring with
Supplemental Information. The Mitochondrial Fission Receptor MiD51. Requires ADP as a Cofactor
 Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Why do we have to cut our hair, nails, and lawn all the time?
 Chapter 5 Cell Reproduction Mitosis Think about this Why do we have to cut our hair, nails, and lawn all the time? EQ: Why is cell division necessary for the growth & development of living organisms? Section
Chapter 5 Cell Reproduction Mitosis Think about this Why do we have to cut our hair, nails, and lawn all the time? EQ: Why is cell division necessary for the growth & development of living organisms? Section
Cellular Signalling 26 (2014) Contents lists available at ScienceDirect. Cellular Signalling
 Contents lists available at ScienceDirect. Cellular Signalling") Cellular Signalling 26 (2014) 2998 3006 Contents lists available at ScienceDirect Cellular Signalling journal homepage: www.elsevier.com/locate/cellsig Review Controlling the switches: Rho GTPase regulation
Cellular Signalling 26 (2014) 2998 3006 Contents lists available at ScienceDirect Cellular Signalling journal homepage: www.elsevier.com/locate/cellsig Review Controlling the switches: Rho GTPase regulation
7.013 Problem Set
 7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.