Thermodynamics and Kinetics
|
|
|
- Joleen Stone
- 5 years ago
- Views:
Transcription
1 Thermodynamics and Kinetics Lecture 12 Free Energy Applications NC State University
2 Isolated system requires DS > 0 DS sys > 0 Isolated system: Entropy increases for any spontaneous process
3 System and surroundings both play in role in the entropy DS sys + DS surr > 0 DS surr > 0 DS sys < 0 or DS sys > 0 If the system is not isolated, then the entropy can have either sign for the System as long as the surroundings can compensate. Closed system in contact with surroundings: Entropy increases for any spontaneous process
4 We derive one function that includes both sys and surr DS sys + DH surr /T > 0 DH surr /T > 0 DS sys < 0 or DS sys > 0 However, if we now turn to a case where the entropy change in the surroundings compensates for a change in the system, we can include the heat flow from surr to sys.
5 We derive one function that includes both sys and surr DS sys - DH sys /T > 0 -DH sys /T > 0 DS sys < 0 or DS sys > 0 The heat flow to or from the system must opposite to that of the surroundings. Hence, there must be a minus sign In front of the enthalpy term.
6 We derive one function that includes both sys and surr DH sys - TDS sys < 0 -DH sys /T > 0 DS sys < 0 or DS sys > 0 Here we have expressed the total change of system and Surroundings in terms of quantities that relate to the System. Hence, DH sys -TDS sys constitutes a new state function
7 We derive one function that includes both sys and surr DG sys < 0 -DH sys /T > 0 DS sys < 0 or DS sys > 0 Note the sign change when we multiplied by T. Here we have written the new state function explicitly as the free energy change, DG, which is defined for the system.
8 A Definition of Gibbs Free Energy Reactions will proceed in the direction written if DG <0. G decreases as shown in the figure below until G is constant. At equilibrium, DG = 0. DG = DG o + RT ln Q where Q is the reaction quotient. Q changes as the Reaction proceeds. Finally, at equilibrium Q = K. Therefore, DG o = -RT ln K time
9 Alternative form for free energy expression Since DG = DG o + RT ln Q and DG o = -RT ln K we can also write DG = -RT ln K + RT ln Q which can be rearranged to DG = RT ln (Q/K)
10 Van t Hoff plot for drug binding A practical example of the application of the van t Hoff equation can be found in drug binding. The equilibrium constant for drug binding to an active site can be measured by fluorescence, NMR, etc. at various temperatures. Then one may plot ln K vs. 1/T and fit the result to a line. In most cases the binding will be exothermic to that DH o < 0 and then slope of the line will be positive rather than negative as shown in the previous slide.
11 Van t Hoff equation The fundamental relation that leads to the temperature dependence of the equilibrium constant is: If there are more than two data points we can plot the data as ln(k) vs. 1/T.
12 Van t Hoff plot for drug binding Slope = -DH o /R Note: DH o < 0 In this example, the slope is positive because the enthalpy of binding is negative (i.e. binding is exothermic).

 as a")
13 Example: preventing inflammation by binding to prostaglandin synthase 2 COX-2 crystal structure DH o and DS o can be measured using ln(k) as a function 1/T.
14 Example: preventing depression serotonin transport inhibitors DH o and DS o can be measured using ln(k) as a function 1/T. This article shows specific differences in the enthalpy of binding of drugs based on analysis of so-called van t Hoff plots (i.e. plots of ln(k) vs. 1/T). Kinetic and Thermodynamic Assessment of Binding of Serotonin Transporter Inhibitors. J Pharm Exp Tech (2008) vol. 327, pp
15 Basic conclusion: fluvoxamine binds exclusively based on entropic driving force Entropically driven binding is relatively rare. Usually the entropy of binding is unfavorable since a flexible drug molecule (large W) will be forced to adopt a fixed conformation (small W or W = 1) upon binding to a protein. Kinetic and Thermodynamic Assessment of Binding of Serotonin Transporter Inhibitors. J Pharm Exp Tech (2008) vol. 327, pp
16 Thermodynamics of glycolysis Reaction kj/mol D-glucose + ATP D-glucose-6-phosphate + ADP DG o = D-glucose-6-phosphate D-fructose-6-phosphate DG o = +1.7 D-fructose- 6-diphosphate + ATP D-fructose-1,6-diphosphate + ADP DG o = D-fructose-1,6-diphosphate glyceraldehyde-3-phosphate + dihydroxyacetone phosphate DG o = dihydroxyacetone phosphate glyceraldehyde-3-phosphate DG o = glyceraldehyde-3-phosphate + phosphate + NAD+ 1,3-diphosphoglycerate + NADH + H+ DG o = ,3-diphosphoglycerate + ADP 3-phosphoglycerate + ATP DG o = phosphoglycerate 2-phosphoglycerate DG o = phosphoglycerate 2-phosphoenolpyruvate + H 2 O DG o = phosphoenolpyruvate + ADP pyruvate + ATP DG o = pyruvate + NADH + H+ lactate + NAD+ DG o = pyruvate acetaldehyde + CO 2 DG o = acetaldehyde + NADH + H+ ethanol + NAD+ DG o = -23.7
17 Phosphorylation of glucose D-glucose + ATP D-glucose-6-phosphate + ADP DG o = The reaction can be decomposed into two reactions. D-glucose + phosphate D-glucose-6-phosphate + H 2 O DG o = ATP + H 2 O ADP + phosphate DG o = The sum of the two reactions results in an overall negative free energy change under standard conditions. In this manner the strongly spontaneous hydrolysis of ATP is coupled to the otherwise unspontaneous glucose phosphorylation. This reaction is typical of the role played by ATP in the cell. Note that the values for DG o assume a concentration of 1 M. Clearly, the concentrations in the cell are often quite different from the standard state and this will have profound consequences for the direction of spontaneous change.
18 Role of enzymes as catalysts All of the reactions in the glycolytic pathway are catalyzed by enzymes. For example, the phosphorylation of glucose is Catalyzed by hexokinase. The role of the enzyme is to speed up the reaction, but the enzyme does not change thermodynamics of the process. The role of enzymes is the same as that of any catalyst. Catalysts affect the kinetics of the reaction, but not the thermodynamics. We will consider the role of catalysts in the section on kinetics.
19 Equilibrium can be important even when DG o > 0 Notice that DG o for certain steps is positive. For example, D-glucose-6-phosphate D-fructose-6-phosphate DG o = +1.7 is catalyzed by phosphoglucose isomerase. The equilibrium constant for this process is K = exp{-dg o /RT} = exp{-1700/8.31/310} ~ 0.5 The concentration of D-fructose-6-phosphate at equilibrium will be less than that of D-glucose-6-phosphate.
20 Question Given that K ~ 0.5 for the reaction D-glucose-6-phosphate D-fructose-6-phosphate Calculate the concentration of D-fructose-6-phosphate at equilibrium under standard conditions (i.e. for initial concentrations of 1 M). A. 1 M B. 0.5 M C M D M
21 Question Given that K ~ 0.5 for the reaction D-glucose-6-phosphate D-fructose-6-phosphate Calculate the concentration of D-fructose-6-phosphate at equilibrium under standard conditions (i.e. for initial concentrations of 1 M). A. 1 M B. 0.5 M C M K = 1 + x, K(1 x) = 1 + x, K 1 = (1 + K)x 1 x x = K K = = [D gluc 6 phos] = 1 x = 1.33 [D fruc 6 phos] = 1 + x = 0.67 D M
22 Intracellular conditions are not equilibrium conditions If the subsequent step in a series of reactions is spontaneous this will tend to deplete the product for the previous reaction. Thus, more of the product will form by L Chateliers principle. We can observe this quantitatively by considering the value of Q, the reaction quotient. Since, DG = DG o + RT ln Q the value of DG may not be zero. In other words, the coupled Series of reactions in the cell are not at equilibrium. Rather, they proceed under steady state conditions where the concentrations are not at 1 M, but are poised so that the overall of effect on a series of reactions is to produce a net spontaneous change.
23 Sample Problem in Metabolism The enzyme aldolase catalyzes the conversion of fructose 1,6-diphosphate (FDP) to dihydroxyactone phosphate (DHAP) and glyceraldehyde-3-phosphoate (GAP). Under physiological conditions the concentrations of these species in red blood cells (erythrocytes) are [FDP] = 35 mm, [DHAP] = 130 mm and [GAP] = 15 mm. Will the conversion occur spontaneously under these conditions? Solution: The standard free energy change for the reaction is FDP DHAP + GAP DG o = kj and Q = [DHAP][GAP]/[FDP] = (130 x 10-6 )(15 x 10-6 )/ (35 x 10-6 ) = 5.57 x 10-5 DG = DG o + RT ln Q = J/mol + (8.31 J/mol-K)(310 K)ln(5.57 x 10-5 ) = J/mol or kj/mol The reaction will occur spontaneously under the conditions of the cell.
24 Thermodynamics of DNA hybridization A combination of spectroscopy and calorimetry was used to determine the free energies of melting of short oligonucleotides. Based on these measurements the free energy of a helix can be determined based on10 sets of nearest-neighbor pairs shown on the next slide. In addition to these values we need to know the free energy of the initiation (i.e. the first base pair). The overall free energy is then calculated from: DG o = DG o (initiation) + S DG o (nearest neighbors)
25 A 5 A 3 T T T 5 A 3 T A A 5 T 3 A T C 5 A 3 T G A 5 C 3 G T 5 AA 3 TT 5 AT 3 TA 5 TA 3 AT 5 AC 3 TG 5 CA 3 GT DG o DH o DS o Breslauer et al. PNAS (1986)
26 G 5 A 3 T C A 5 G 3 C T G 5 C 3 G C C 5 G 3 C G C 5 C 3 G G 5 AG 3 TC 5 GA 3 CT 5 CG 3 GC 5 GC 3 CG 5 CA 3 GT DG o DH o DS o Breslauer et al. PNAS (1986)
27 Sample problem Determine the melt temperature for the oligonucleotide 5 -ATAGCA TATCGT-5 5 -ATAGCA-3 3 -TATCGT-5 Solution: DG o = DG o (initiation) + S DG o (nearest neighbors) = DG o GC {initial) + DG o AT TA + TA DGo + DG o AG AT TC + GC DGo CG + CA DGo GT = = kj Initiation can be treated as a single value required to start the hybridization. It is unfavorable, but then all other interactions are favorable.
28 Sample problem Determine the melt temperature for the oligonucleotide 5 -ATAGCA TATCGT-5 5 -ATAGCA-3 3 -TATCGT-5 Solution (cont;d): DG o = kj Notice that the the free energy of initiation is positive. Initiation is unfavorable because of the entropy that must be overcome to bring the chains together. To calculate the melt temperature we need the enthalpy of reaction as well. DH o = DH o AT TA + DHo TA AT + DH o AG TC + GC DHo CG + CA DHo GT = = kj
29 Sample problem Given that DG o = kj/mol and DH o = kj/mol for the hexamer, determine the melt temperature. A. 42 o C B. 48 o C C. 52 o C D. 58 o C
30 Sample problem Given that DG o = kj/mol and DH o = kj/mol for the hexamer, determine the melt temperature. A. 42 o C B. 48 o C C. 52 o C D. 58 o C DS o = (DH o - DG o )/T = ( )/298 x1000 = J/mol-K The melt temperature occurs when DG o = 0. T = DH o /DS o = -164,400/(-495.3) = 331 K = 58 o C
31 DNA hyperchromicity

32 DNA hyperchromicity Observed in reverse by fluorescence
33 Folding and hybridization DNA hybridization is similar to protein folding in that they both have a melt temperature. The melt temperature refers to the temperature at which proteins unfold and DNA becomes single-stranded. We next consider the interactions that stabilize folded proteins.

34 Dipole-Dipole Interactions Dipoles often line up in this manner. Example: a-helix

35 Main Chain Electrostatic Interactions Coulomb s Law: V = q 1 q 2 /er Example of a hydrogen bond -N-H.. O=C- Example of a Salt Bridge Main Chain Lysine Glutamate

36 Hydrogen bonding in water
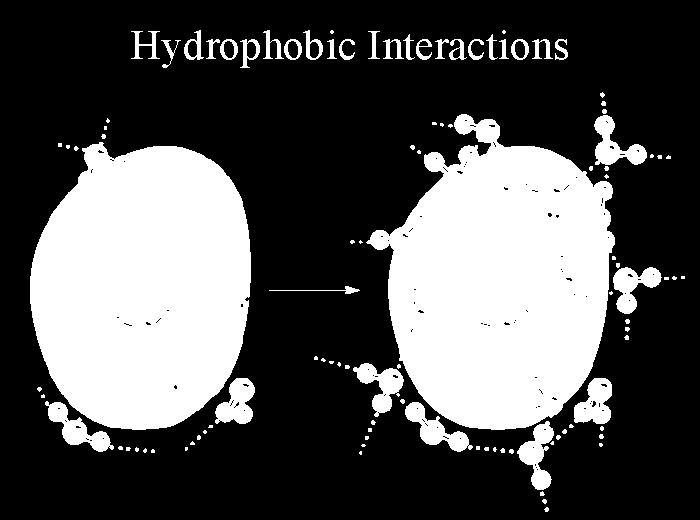
37 Hydrophobic interactions
38 Protein Folding Non-covalent forces in proteins What holds them together? Hydrogen bonds Salt-bridges Dipole-dipole interactions Hydrophobic effect Van der Waals forces What pulls them apart? Conformational Entropy
39 Contributions to DG TDS DH Internal Interactions Conformational Entropy -TDS Hydrophobic Effect Net: DG Folding
40 Protein folding example: Two state model U k f F Unfolded k u Folded K = [F]/[U] K = ff/(1-ff) Fraction folded ff Fraction unfolded 1-ff
41 Thermodynamic model ff = K/(1+K) K = e -DGo /RT ff = 1/(1 + e DGo /RT ) ff = 1/(1 + e DHo /RT e -DSo /R ) The temperature at which the protein is 50% folded can be defined as T m the melt temperature. At T m, DG o = 0 or T m = DH o /DS o.
42 Equilibrium melt curves Mostly folded o o Mostly unfolded T m In this case: T m = 300 K = DH o /DS o
43 Absorbance Delta Absorbance Thermal melt data 90x Temp ( C) Temp ( C) Temp ( C) x Wavenumber (cm -1 ) Wavenumber (cm -1 ) 1720 Infrared absorption spectra in the amide I region of the peptide. This is a probe of degree of folding.
44 The iron in heme is the binding site for oxygen and peroxide Heme is iron protoporphyrin IX. Functional aspects in Mb N O // O N 1. Discrimination against CO binding. N Fe N 2. O 2 is the physiologically relevant ligand, but it can oxidize iron (autooxidation). O O - O O -
45 Ligand binding in myoglobin Ligand binding to myoglobin is described as a chemical equilibrium between a bound state MbCO, a dissociated state Mb:CO and a solvent state Mb + CO. Mb + CO Mb:CO MbCO If we ignore the intermediate state the binding can be described by An overall equilibrium process. Mb + CO with equilibrium constant K. We have: The fraction bound is: f = K = [MbCO] [Mb][CO] MbCO [MbCO] [Mb] + [MbCO] We use CO in Many studies. The same equations Hold for O 2.
46 Ligand binding curve The fraction bound f can be related to the binding constant K: K = [MbCO] [Mb][CO] [MbCO] f = [Mb]+[MbCO] = K[Mb][CO] [Mb] + K[Mb][CO] = K[CO] 1 + K[CO] This type of binding curve is plotted below.

47 Cooperativity: hemoglobin If there are multiple binding sites in a macromolecule there can be two possible cases. First, they can be completely non-interacting and independent. In that case they behave exactly as a collection of monomers. On the other hand, the sites can interact. In that case we can speak of cooperativity or anti-cooperativity of the binding sites. We can use hemoglobin as an example.
48 Multiple equilibria There are four hemes in hemoglobin. Thus, there are four binding sites. The equilbrium constants are: Hb + O 2 Hb(O 2 ) Hb(O 2 ) + O 2 Hb(O 2 ) 2 K 1 = [Hb(O 2)] [Hb][O 2 ] K 2 = [Hb(O 2) 2 ] [Hb(O 2 )][O 2 ] Hb(O 2 ) 2 + O 2 Hb(O 2 ) 3 Hb(O 2 ) 3 + O 2 Hb(O 2 ) 4 K 3 = K 4 = [Hb(O 2 ) 3 ] [Hb(O 2 ) 2 ][O 2 ] [Hb(O 2 ) 4 ] [Hb(O 2 ) 3 ][O 2 ]
49 Combined equilibrium constant There are four hemes in hemoglobin. Thus, there are four binding sites. The equilbrium constants are: [Hb(O 2 )] = K 1 [Hb][O 2 ] [Hb(O 2 ) 2 ] = K 2 [Hb(O 2 )][O 2 ] = K 2 K 1 [Hb][O 2 ] 2 [Hb(O 2 ) 3 ] = K 3 [Hb(O 2 ) 2 ][O 2 ] = K 3 K 2 K 1 [Hb][O 2 ] 3 [Hb(O 2 ) 4 ] = K 4 [Hb(O 2 ) 3 ][O 2 ] = K 4 K 3 K 2 K 1 [Hb][O 2 ] 4 Define K = K 4 K 3 K 2 K 1 which is the equilbriumconstant for the overall reaction: Hb + 4O 2 Hb(O 2 ) 4 K = [Hb(O 2) 4 ] [Hb][O 2 ] 4
50 Cooperative binding isotherm The combined equilibrium treats the binding to hemoglobin as an all-or-nothing process. That is, either there are no O2 ligands or there are four O2 ligands. The is the extreme of cooperative binding. In this case the fraction bound is: f = [Hb(O 2 ) 4 ] [Hb] + [Hb(O 2 ) 4 ] = K [Hb][O 2 ] 4 [Hb] + K [Hb][O 2 ] = K [O 2] K [O 2 ] 4 The isotherm for cooperative binding is shown in the red curve below:
51 The Hill Coefficient In reality the cooperativity is not complete. In other words the binding of ligands does not occur all at once. The binding is sequential such that when then first oxygen (or CO) binds it increases the binding affinity of the second one etc. One way to express the fact that the cooperativity is less than perfect is to use n as a parameter. For hemoglobin n = 1 to 4 depending on the degree of communication between the four subunits. f = K [O 2] n 1 + K [O 2 ] n Experimentally the degree of ligation (O 2 or CO binding) can be followed using UV-vis spectroscopy. The effects of mutations on the local Environment of the bound ligand can be probed using infrared spectroscopy of bound CO.
.")
52 Monod-Wyman-Changeux Theory The original idea of the MWC theory was to describe protein cooperativity in terms of two states R = relaxed state T = tense state In the relaxed state there was little interaction between the protein subunits. A binding event would cause the structure to change and the protein subunits would begin to interact strongly. This would give rise to the tense state (T). In hemoglobin the R state is the oxy form. When oxygen dissociates the heme iron moves out of the heme plane. This occurs because the iron changes from a low spin to a high spin metal. The change in iron position permits the F-helix to move into contact with the CD loop region of the neighboring subunit. T R

53 Structural changes in Hb The change in intersubunit contacts results in a rotation of the a/b subunits so that the overall structure of the tetrameric protein is changed in the R to T transition.
54 The origin of hemoglobin cooperativity The key amino acid residues that change their hydrogen bonding are shown below. The intersubunit contacts are altered by the binding of oxygen to the heme iron because of the change in the position of the heme iron. In-plane iron locks in the R state (pulls subunits apart) Out-of-plane iron iron gives the T state (allows subunits to interact)
, which helps to unload oxygen from the hemoglobin Bohr effect is the increased unloading of O 2 at low ph Figure The black curve is at ph 7.4, The red curve is at ph 7.")
55 The Bohr effect Acid Conditions Help Release O 2 in the Tissues Hemoglobin must bind oxygen tightly in the lungs Some unloading in tissue results from the low po 2 Active tissues make lots of acid (carbonic and lactic), which helps to unload oxygen from the hemoglobin Bohr effect is the increased unloading of O 2 at low ph Figure The black curve is at ph 7.4, The red curve is at ph 7.1 The blue curve is at ph 6.8
 Hemoglobin is a protein")
56 The physiology of Hb and Mb Oxygen Uptake in the Lungs is Increased About 70 times by Hemoglobin in the Red Cells A small amount of oxygen dissolves directly in the serum, but 98.5% of the oxygen is carried by Hb Hb is found within the red blood cells Hb content of blood is ~15 gm/100 ml Each Hemoglobin Can Bind Four O 2 Molecules (100% Saturation) Hemoglobin is a protein molecule with (2 a and 2 b) When hemoglobin is saturated with oxygen it has a bright red color; as it loses oxygen it becomes bluish (cyanosis)
Applications of Free Energy. NC State University
 Chemistry 433 Lecture 15 Applications of Free Energy NC State University Thermodynamics of glycolysis Reaction kj/mol D-glucose + ATP D-glucose-6-phosphate + ADP ΔG o = -16.7 D-glucose-6-phosphate p D-fructose-6-phosphate
Chemistry 433 Lecture 15 Applications of Free Energy NC State University Thermodynamics of glycolysis Reaction kj/mol D-glucose + ATP D-glucose-6-phosphate + ADP ΔG o = -16.7 D-glucose-6-phosphate p D-fructose-6-phosphate
Free Energy. because H is negative doesn't mean that G will be negative and just because S is positive doesn't mean that G will be negative.
 Biochemistry 462a Bioenergetics Reading - Lehninger Principles, Chapter 14, pp. 485-512 Practice problems - Chapter 14: 2-8, 10, 12, 13; Physical Chemistry extra problems, free energy problems Free Energy
Biochemistry 462a Bioenergetics Reading - Lehninger Principles, Chapter 14, pp. 485-512 Practice problems - Chapter 14: 2-8, 10, 12, 13; Physical Chemistry extra problems, free energy problems Free Energy
Chapter 10: Hemoglobin
 Chapter 10: Hemoglobin Voet & Voet: Pages 320-353 Slide 1 Hemoglobin Function Larger aerobic (oxygen utilizing) organism require an O 2 transport system to deliver sufficient O 2 to tissues Dissolved O
Chapter 10: Hemoglobin Voet & Voet: Pages 320-353 Slide 1 Hemoglobin Function Larger aerobic (oxygen utilizing) organism require an O 2 transport system to deliver sufficient O 2 to tissues Dissolved O
= (-22) = +2kJ /mol
 = +2kJ /mol") Lecture 8: Thermodynamics & Protein Stability Assigned reading in Campbell: Chapter 4.4-4.6 Key Terms: DG = -RT lnk eq = DH - TDS Transition Curve, Melting Curve, Tm DH calculation DS calculation van der
Lecture 8: Thermodynamics & Protein Stability Assigned reading in Campbell: Chapter 4.4-4.6 Key Terms: DG = -RT lnk eq = DH - TDS Transition Curve, Melting Curve, Tm DH calculation DS calculation van der
BIOCHEMISTRY. František Vácha. JKU, Linz.
 BIOCHEMISTRY František Vácha http://www.prf.jcu.cz/~vacha/ JKU, Linz Recommended reading: D.L. Nelson, M.M. Cox Lehninger Principles of Biochemistry D.J. Voet, J.G. Voet, C.W. Pratt Principles of Biochemistry
BIOCHEMISTRY František Vácha http://www.prf.jcu.cz/~vacha/ JKU, Linz Recommended reading: D.L. Nelson, M.M. Cox Lehninger Principles of Biochemistry D.J. Voet, J.G. Voet, C.W. Pratt Principles of Biochemistry
Sample Question Solutions for the Chemistry of Life Topic Test
 Sample Question Solutions for the Chemistry of Life Topic Test 1. Enzymes play a crucial role in biology by serving as biological catalysts, increasing the rates of biochemical reactions by decreasing
Sample Question Solutions for the Chemistry of Life Topic Test 1. Enzymes play a crucial role in biology by serving as biological catalysts, increasing the rates of biochemical reactions by decreasing
Biological Chemistry and Metabolic Pathways
 Biological Chemistry and Metabolic Pathways 1. Reaction a. Thermodynamics b. Kinetics 2. Enzyme a. Structure and Function b. Regulation of Activity c. Kinetics d. Inhibition 3. Metabolic Pathways a. REDOX
Biological Chemistry and Metabolic Pathways 1. Reaction a. Thermodynamics b. Kinetics 2. Enzyme a. Structure and Function b. Regulation of Activity c. Kinetics d. Inhibition 3. Metabolic Pathways a. REDOX
Biochemical Pathways
 Biochemical Pathways Living organisms can be divided into two large groups according to the chemical form in which they obtain carbon from the environment. Autotrophs can use carbon dioxide from the atmosphere
Biochemical Pathways Living organisms can be divided into two large groups according to the chemical form in which they obtain carbon from the environment. Autotrophs can use carbon dioxide from the atmosphere
Affinity labels for studying enzyme active sites. Irreversible Enzyme Inhibition. Inhibition of serine protease with DFP
 Irreversible Enzyme Inhibition Irreversible inhibitors form stable covalent bonds with the enzyme (e.g. alkylation or acylation of an active site side chain) There are many naturally-occurring and synthetic
Irreversible Enzyme Inhibition Irreversible inhibitors form stable covalent bonds with the enzyme (e.g. alkylation or acylation of an active site side chain) There are many naturally-occurring and synthetic
Chapter 15 part 2. Biochemistry I Introduction to Metabolism Bioenergetics: Thermodynamics in Biochemistry. ATP 4- + H 2 O ADP 3- + P i + H +
 Biochemistry I Introduction to Metabolism Bioenergetics: Thermodynamics in Biochemistry ATP 4- + 2 ADP 3- + P i 2- + + Chapter 15 part 2 Dr. Ray 1 Energy flow in biological systems: Energy Transformations
Biochemistry I Introduction to Metabolism Bioenergetics: Thermodynamics in Biochemistry ATP 4- + 2 ADP 3- + P i 2- + + Chapter 15 part 2 Dr. Ray 1 Energy flow in biological systems: Energy Transformations
Department of Chemistry and Biochemistry University of Lethbridge. Biochemistry II. Bioenergetics
 Department of Chemistry and Biochemistry University of Lethbridge II. Bioenergetics Slide 1 Bioenergetics Bioenergetics is the quantitative study of energy relationships and energy conversion in biological
Department of Chemistry and Biochemistry University of Lethbridge II. Bioenergetics Slide 1 Bioenergetics Bioenergetics is the quantitative study of energy relationships and energy conversion in biological
Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability
 Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability Dr. Andrew Lee UNC School of Pharmacy (Div. Chemical Biology and Medicinal Chemistry) UNC Med
Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability Dr. Andrew Lee UNC School of Pharmacy (Div. Chemical Biology and Medicinal Chemistry) UNC Med
f) Adding an enzyme does not change the Gibbs free energy. It only increases the rate of the reaction by lowering the activation energy.
 Adding an enzyme does not change the Gibbs free energy. It only increases the rate of the reaction by lowering the activation energy.") Problem Set 2-Answer Key BILD1 SP16 1) How does an enzyme catalyze a chemical reaction? Define the terms and substrate and active site. An enzyme lowers the energy of activation so the reaction proceeds
Problem Set 2-Answer Key BILD1 SP16 1) How does an enzyme catalyze a chemical reaction? Define the terms and substrate and active site. An enzyme lowers the energy of activation so the reaction proceeds
Chem Lecture 3 Hemoglobin & Myoglobin
 Chem 452 - Lecture 3 Hemoglobin & Myoglobin 111003 Hemoglobin (Hb) and Myoglobin (Mb) function as oxygen transport and storage molecules in higher organisms. There functions have been long studied and,
Chem 452 - Lecture 3 Hemoglobin & Myoglobin 111003 Hemoglobin (Hb) and Myoglobin (Mb) function as oxygen transport and storage molecules in higher organisms. There functions have been long studied and,
Lecture 2: Biological Thermodynamics [PDF] Key Concepts
![Lecture 2: Biological Thermodynamics [PDF] Key Concepts](/thumbs/76/73872045.jpg "Lecture 2: Biological Thermodynamics [PDF] Key Concepts") Lecture 2: Biological Thermodynamics [PDF] Reading: Berg, Tymoczko & Stryer: pp. 11-14; pp. 208-210 problems in textbook: chapter 1, pp. 23-24, #4; and thermodynamics practice problems [PDF] Updated on:
Lecture 2: Biological Thermodynamics [PDF] Reading: Berg, Tymoczko & Stryer: pp. 11-14; pp. 208-210 problems in textbook: chapter 1, pp. 23-24, #4; and thermodynamics practice problems [PDF] Updated on:
Principles of Bioenergetics. Lehninger 3 rd ed. Chapter 14
 1 Principles of Bioenergetics Lehninger 3 rd ed. Chapter 14 2 Metabolism A highly coordinated cellular activity aimed at achieving the following goals: Obtain chemical energy. Convert nutrient molecules
1 Principles of Bioenergetics Lehninger 3 rd ed. Chapter 14 2 Metabolism A highly coordinated cellular activity aimed at achieving the following goals: Obtain chemical energy. Convert nutrient molecules
Enzymes I. Dr. Mamoun Ahram Summer semester,
 Enzymes I Dr. Mamoun Ahram Summer semester, 2017-2018 Resources Mark's Basic Medical Biochemistry Other resources NCBI Bookshelf: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=books The Medical Biochemistry
Enzymes I Dr. Mamoun Ahram Summer semester, 2017-2018 Resources Mark's Basic Medical Biochemistry Other resources NCBI Bookshelf: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=books The Medical Biochemistry
Chapter 19 Chemical Thermodynamics Entropy and free energy
 Chapter 19 Chemical Thermodynamics Entropy and free energy Learning goals and key skills: Explain and apply the terms spontaneous process, reversible process, irreversible process, and isothermal process.
Chapter 19 Chemical Thermodynamics Entropy and free energy Learning goals and key skills: Explain and apply the terms spontaneous process, reversible process, irreversible process, and isothermal process.
Lecture 20. Chemical Potential
 Lecture 20 Chemical Potential Reading: Lecture 20, today: Chapter 10, sections A and B Lecture 21, Wednesday: Chapter 10: 10 17 end 3/21/16 1 Pop Question 7 Boltzmann Distribution Two systems with lowest
Lecture 20 Chemical Potential Reading: Lecture 20, today: Chapter 10, sections A and B Lecture 21, Wednesday: Chapter 10: 10 17 end 3/21/16 1 Pop Question 7 Boltzmann Distribution Two systems with lowest
Sample Questions for the Chemistry of Life Topic Test
 Sample Questions for the Chemistry of Life Topic Test 1. Enzymes play a crucial role in biology by serving as biological catalysts, increasing the rates of biochemical reactions by decreasing their activation
Sample Questions for the Chemistry of Life Topic Test 1. Enzymes play a crucial role in biology by serving as biological catalysts, increasing the rates of biochemical reactions by decreasing their activation
a) Write the reaction that occurs (pay attention to and label ends correctly) 5 AGCTG CAGCT > 5 AGCTG 3 3 TCGAC 5
 Write the reaction that occurs (pay attention to and label ends correctly) 5 AGCTG CAGCT > 5 AGCTG 3 3 TCGAC 5") Chem 315 Applications Practice Problem Set 1.As you learned in Chem 315, DNA higher order structure formation is a two step process. The first step, nucleation, is entropically the least favorable. The
Chem 315 Applications Practice Problem Set 1.As you learned in Chem 315, DNA higher order structure formation is a two step process. The first step, nucleation, is entropically the least favorable. The
Chapter 6- An Introduction to Metabolism*
 Chapter 6- An Introduction to Metabolism* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Energy of Life
Chapter 6- An Introduction to Metabolism* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Energy of Life
Lecture #8 9/21/01 Dr. Hirsh
 Lecture #8 9/21/01 Dr. Hirsh Types of Energy Kinetic = energy of motion - force x distance Potential = stored energy In bonds, concentration gradients, electrical potential gradients, torsional tension
Lecture #8 9/21/01 Dr. Hirsh Types of Energy Kinetic = energy of motion - force x distance Potential = stored energy In bonds, concentration gradients, electrical potential gradients, torsional tension
Chapter 16: Spontaneity, Entropy, and Free Energy Spontaneous Processes and Entropy
 Chapter 16: Spontaneity, Entropy, and Free Energy 16.1 Spontaneous Processes and Entropy 1 3 The first law of thermodynamics the law of conservation of energy: Energy can be neither created nor destroyed
Chapter 16: Spontaneity, Entropy, and Free Energy 16.1 Spontaneous Processes and Entropy 1 3 The first law of thermodynamics the law of conservation of energy: Energy can be neither created nor destroyed
The products have more enthalpy and are more ordered than the reactants.
 hapters 7 & 10 Bioenergetics To live, organisms must obtain energy from their environment and use it to do the work of building and organizing cell components such as proteins, enzymes, nucleic acids,
hapters 7 & 10 Bioenergetics To live, organisms must obtain energy from their environment and use it to do the work of building and organizing cell components such as proteins, enzymes, nucleic acids,
Biochemistry 3300 Problems (and Solutions) Metabolism I
 Metabolism I") (1) Provide a reasonable systematic name for an enzyme that catalyzes the following reaction: fructose + ATP > fructose-1 phosphate + ADP (2) The IUBMB has a developed a set of rules for classifying enzymes
(1) Provide a reasonable systematic name for an enzyme that catalyzes the following reaction: fructose + ATP > fructose-1 phosphate + ADP (2) The IUBMB has a developed a set of rules for classifying enzymes
Basic Concepts of Enzyme Action. Enzymes. Rate Enhancement 9/17/2015. Stryer Short Course Chapter 6
 Basic Concepts of Enzyme Action Stryer Short Course Chapter 6 Enzymes Biocatalysts Active site Substrate and product Catalyzed rate Uncatalyzed rate Rate Enhancement Which is a better catalyst, carbonic
Basic Concepts of Enzyme Action Stryer Short Course Chapter 6 Enzymes Biocatalysts Active site Substrate and product Catalyzed rate Uncatalyzed rate Rate Enhancement Which is a better catalyst, carbonic
Energy in Chemical and Biochemical Reactions
 Energy in Chemical and Biochemical Reactions Reaction Progress Diagram for Exothermic Reaction Reactants activated complex Products ENERGY A + B Reactants E a C + D Products Δ rxn Reaction coordinate The
Energy in Chemical and Biochemical Reactions Reaction Progress Diagram for Exothermic Reaction Reactants activated complex Products ENERGY A + B Reactants E a C + D Products Δ rxn Reaction coordinate The
LS1a Fall 2014 Practice Problem Set #1. 1. Consider a hypothetical reaction carried out at constant temperature and pressure:
 LS1a Fall 2014 Practice Problem Set #1 I. Basic Concept Questions 1. Consider a hypothetical reaction carried out at constant temperature and pressure: The Gibbs free energy curve of this reaction is depicted
LS1a Fall 2014 Practice Problem Set #1 I. Basic Concept Questions 1. Consider a hypothetical reaction carried out at constant temperature and pressure: The Gibbs free energy curve of this reaction is depicted
Chapter 17.3 Entropy and Spontaneity Objectives Define entropy and examine its statistical nature Predict the sign of entropy changes for phase
 Chapter 17.3 Entropy and Spontaneity Objectives Define entropy and examine its statistical nature Predict the sign of entropy changes for phase changes Apply the second law of thermodynamics to chemical
Chapter 17.3 Entropy and Spontaneity Objectives Define entropy and examine its statistical nature Predict the sign of entropy changes for phase changes Apply the second law of thermodynamics to chemical
Lecture 3: Thermodynamics
 3 LAWS OF THERMODYNAMICS Lecture 3: Thermodynamics Matter and energy are conserved Margaret A. Daugherty Fall 2004 Entropy always increases Absolute zero is unattainable System and Surroundings 1st Law
3 LAWS OF THERMODYNAMICS Lecture 3: Thermodynamics Matter and energy are conserved Margaret A. Daugherty Fall 2004 Entropy always increases Absolute zero is unattainable System and Surroundings 1st Law
5. Kinetics of Allosteric Enzymes. Sigmoidal Kinetics. Cooperativity Binding Constant
 5. Kinetics of Allosteric Enzymes Sigmoidal Kinetics Cooperativity Binding Constant Kinetics of Allosteric Enzymes Contents Definitions Allosteric enzymes Cooperativity Homoallostery Heteroallostery Biphasic
5. Kinetics of Allosteric Enzymes Sigmoidal Kinetics Cooperativity Binding Constant Kinetics of Allosteric Enzymes Contents Definitions Allosteric enzymes Cooperativity Homoallostery Heteroallostery Biphasic
Other Cells. Hormones. Viruses. Toxins. Cell. Bacteria
 Other Cells Hormones Viruses Toxins Cell Bacteria ΔH < 0 reaction is exothermic, tells us nothing about the spontaneity of the reaction Δ H > 0 reaction is endothermic, tells us nothing about the spontaneity
Other Cells Hormones Viruses Toxins Cell Bacteria ΔH < 0 reaction is exothermic, tells us nothing about the spontaneity of the reaction Δ H > 0 reaction is endothermic, tells us nothing about the spontaneity
Metabolism and Enzymes
 Energy Basics Metabolism and Enzymes Chapter 5 Pgs. 77 86 Chapter 8 Pgs. 142 162 Energy is the capacity to cause change, and is required to do work. Very difficult to define quantity. Two types of energy:
Energy Basics Metabolism and Enzymes Chapter 5 Pgs. 77 86 Chapter 8 Pgs. 142 162 Energy is the capacity to cause change, and is required to do work. Very difficult to define quantity. Two types of energy:
Energy Transformation, Cellular Energy & Enzymes (Outline)
") Energy Transformation, Cellular Energy & Enzymes (Outline) Energy conversions and recycling of matter in the ecosystem. Forms of energy: potential and kinetic energy The two laws of thermodynamic and definitions
Energy Transformation, Cellular Energy & Enzymes (Outline) Energy conversions and recycling of matter in the ecosystem. Forms of energy: potential and kinetic energy The two laws of thermodynamic and definitions
2054, Chap. 8, page 1
 2054, Chap. 8, page 1 I. Metabolism: Energetics, Enzymes, and Regulation (Chapter 8) A. Energetics and work 1. overview a. energy = ability to do work (1) chemical, transport, mechanical (2) ultimate source
2054, Chap. 8, page 1 I. Metabolism: Energetics, Enzymes, and Regulation (Chapter 8) A. Energetics and work 1. overview a. energy = ability to do work (1) chemical, transport, mechanical (2) ultimate source
10/26/2010. An Example of a Polar Reaction: Addition of H 2 O to Ethylene. to Ethylene
 6.5 An Example of a Polar Reaction: Addition of H 2 O to Ethylene Addition of water to ethylene Typical polar process Acid catalyzed addition reaction (Electophilic addition reaction) Polar Reaction All
6.5 An Example of a Polar Reaction: Addition of H 2 O to Ethylene Addition of water to ethylene Typical polar process Acid catalyzed addition reaction (Electophilic addition reaction) Polar Reaction All
BIOLOGICAL SCIENCE. Lecture Presentation by Cindy S. Malone, PhD, California State University Northridge. FIFTH EDITION Freeman Quillin Allison
 BIOLOGICAL SCIENCE FIFTH EDITION Freeman Quillin Allison 8 Lecture Presentation by Cindy S. Malone, PhD, California State University Northridge Roadmap 8 In this chapter you will learn how Enzymes use
BIOLOGICAL SCIENCE FIFTH EDITION Freeman Quillin Allison 8 Lecture Presentation by Cindy S. Malone, PhD, California State University Northridge Roadmap 8 In this chapter you will learn how Enzymes use
Biochemistry 3100 Sample Problems Binding proteins, Kinetics & Catalysis
 (1) Draw an approximate denaturation curve for a typical blood protein (eg myoglobin) as a function of ph. (2) Myoglobin is a simple, single subunit binding protein that has an oxygen storage function
(1) Draw an approximate denaturation curve for a typical blood protein (eg myoglobin) as a function of ph. (2) Myoglobin is a simple, single subunit binding protein that has an oxygen storage function
CHEM Exam 3 - March 31, 2017
 CHEM 3530 - Exam 3 - March 31, 2017 Constants and Conversion Factors NA = 6.02x10 23 mol -1 R = 8.31 J/mol-K = 8.31 kpa-l/mol-k 1 bar = 100 kpa = 750 torr 1 kpa = 7.50 torr 1 J = 1 kpa-l 1 kcal = 4.18
CHEM 3530 - Exam 3 - March 31, 2017 Constants and Conversion Factors NA = 6.02x10 23 mol -1 R = 8.31 J/mol-K = 8.31 kpa-l/mol-k 1 bar = 100 kpa = 750 torr 1 kpa = 7.50 torr 1 J = 1 kpa-l 1 kcal = 4.18
CHEM Exam 3 - March 30, Given in the individual questions on this test.
 CHEM 3530 - Exam 3 - March 30, 2018 Constants and Conversion Factors NA = 6.02x10 23 mol -1 R = 8.31 J/mol-K = 8.31 kpa-l/mol-k 1 bar = 100 kpa = 750 torr 1 kpa = 7.50 torr 1 J = 1 kpa-l 1 kcal = 4.18
CHEM 3530 - Exam 3 - March 30, 2018 Constants and Conversion Factors NA = 6.02x10 23 mol -1 R = 8.31 J/mol-K = 8.31 kpa-l/mol-k 1 bar = 100 kpa = 750 torr 1 kpa = 7.50 torr 1 J = 1 kpa-l 1 kcal = 4.18
What binds to Hb in addition to O 2?
 Reading: Ch5; 158-169, 162-166, 169-174 Problems: Ch5 (text); 3,7,8,10 Ch5 (study guide-facts); 1,2,3,4,5,8 Ch5 (study guide-apply); 2,3 Remember Today at 5:30 in CAS-522 is the second chance for the MB
Reading: Ch5; 158-169, 162-166, 169-174 Problems: Ch5 (text); 3,7,8,10 Ch5 (study guide-facts); 1,2,3,4,5,8 Ch5 (study guide-apply); 2,3 Remember Today at 5:30 in CAS-522 is the second chance for the MB
Thermodynamics. Entropy and its Applications. Lecture 11. NC State University
 Thermodynamics Entropy and its Applications Lecture 11 NC State University System and surroundings Up to this point we have considered the system, but we have not concerned ourselves with the relationship
Thermodynamics Entropy and its Applications Lecture 11 NC State University System and surroundings Up to this point we have considered the system, but we have not concerned ourselves with the relationship
Biomolecules: lecture 10
 Biomolecules: lecture 10 - understanding in detail how protein 3D structures form - realize that protein molecules are not static wire models but instead dynamic, where in principle every atom moves (yet
Biomolecules: lecture 10 - understanding in detail how protein 3D structures form - realize that protein molecules are not static wire models but instead dynamic, where in principle every atom moves (yet
Full file at https://fratstock.eu
 Chapter 03 1. a. DG=DH-TDS Δ G = 80 kj ( 98 K) 0.790 kj = 44.6 kj K b. ΔG = 0 @ T m. Unfolding will be favorable at temperatures above the T m. Δ G =Δ H TΔ S 0 kj kj 80 ( xk) 0.790 K 0 Δ G = = 354.4 K
Chapter 03 1. a. DG=DH-TDS Δ G = 80 kj ( 98 K) 0.790 kj = 44.6 kj K b. ΔG = 0 @ T m. Unfolding will be favorable at temperatures above the T m. Δ G =Δ H TΔ S 0 kj kj 80 ( xk) 0.790 K 0 Δ G = = 354.4 K
The energy of oxidation of 11 g glucose = kj = kg/m 2 s 2
 Chem 350 thermo problems Key 1. How many meters of stairway could a 70kg man climb if all the energy available in metabolizing an 11 g spoonful of sugar to carbon dioxide and water could be converted to
Chem 350 thermo problems Key 1. How many meters of stairway could a 70kg man climb if all the energy available in metabolizing an 11 g spoonful of sugar to carbon dioxide and water could be converted to
Protein Folding & Stability. Lecture 11: Margaret A. Daugherty. Fall How do we go from an unfolded polypeptide chain to a
 Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2004 How do we go from an unfolded polypeptide chain to a compact folded protein? (Folding of thioredoxin, F. Richards) Structure - Function
Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2004 How do we go from an unfolded polypeptide chain to a compact folded protein? (Folding of thioredoxin, F. Richards) Structure - Function
Photosynthetic autotrophs use the energy of sunlight to convert low-g CO 2 and H 2 O into energy-rich complex sugar molecules.
 Chapters 7 & 10 Bioenergetics To live, organisms must obtain energy from their environment and use it to do the work of building and organizing cell components such as proteins, enzymes, nucleic acids,
Chapters 7 & 10 Bioenergetics To live, organisms must obtain energy from their environment and use it to do the work of building and organizing cell components such as proteins, enzymes, nucleic acids,
Thermodynamics. Thermodynamically favored reactions ( spontaneous ) Enthalpy Entropy Free energy
 Enthalpy Entropy Free energy") Thermodynamics Thermodynamically favored reactions ( spontaneous ) Enthalpy Entropy Free energy 1 Thermodynamically Favored Processes Water flows downhill. Sugar dissolves in coffee. Heat flows from hot
Thermodynamics Thermodynamically favored reactions ( spontaneous ) Enthalpy Entropy Free energy 1 Thermodynamically Favored Processes Water flows downhill. Sugar dissolves in coffee. Heat flows from hot
Energy Transformation. Metabolism = total chemical reactions in cells.
 Energy Transformation Metabolism = total chemical reactions in cells. metabole = change Metabolism is concerned with managing the material and energy resources of the cell -Catabolism -Anabolism -Catabolism
Energy Transformation Metabolism = total chemical reactions in cells. metabole = change Metabolism is concerned with managing the material and energy resources of the cell -Catabolism -Anabolism -Catabolism
Lecture Series 9 Cellular Pathways That Harvest Chemical Energy
 Lecture Series 9 Cellular Pathways That Harvest Chemical Energy Reading Assignments Review Chapter 3 Energy, Catalysis, & Biosynthesis Read Chapter 13 How Cells obtain Energy from Food Read Chapter 14
Lecture Series 9 Cellular Pathways That Harvest Chemical Energy Reading Assignments Review Chapter 3 Energy, Catalysis, & Biosynthesis Read Chapter 13 How Cells obtain Energy from Food Read Chapter 14
BMB Lecture 7. Allostery and Cooperativity. A means for exquisite control
 BMB 178 2018 Lecture 7 Allostery and Cooperativity A means for exquisite control Allostery: the basis of enzymatic control From the Greek: allos = other stereos = solid or space Action at a distance Examples
BMB 178 2018 Lecture 7 Allostery and Cooperativity A means for exquisite control Allostery: the basis of enzymatic control From the Greek: allos = other stereos = solid or space Action at a distance Examples
Chapter 9 in Chang Text
 Section 8.0: CHEMICAL EQUILIBRIUM Chapter 9 in Chang Text ..the direction of spontaneous change at constant T and P is towards lower values of Gibbs Energy (G). this also applies to chemical reactions
Section 8.0: CHEMICAL EQUILIBRIUM Chapter 9 in Chang Text ..the direction of spontaneous change at constant T and P is towards lower values of Gibbs Energy (G). this also applies to chemical reactions
BCHS 6229 Protein Structure and Function. Lecture 3 (October 18, 2011) Protein Folding: Forces, Mechanisms & Characterization
 Protein Folding: Forces, Mechanisms & Characterization") BCHS 6229 Protein Structure and Function Lecture 3 (October 18, 2011) Protein Folding: Forces, Mechanisms & Characterization 1 The folding problem One of the greatest unsolved problems of Science The folding
BCHS 6229 Protein Structure and Function Lecture 3 (October 18, 2011) Protein Folding: Forces, Mechanisms & Characterization 1 The folding problem One of the greatest unsolved problems of Science The folding
CHEMISTRY OF THE HUMAN BODY
 CHEMISTRY OF THE HUMAN BODY (Sample Questions Key) WUCT 2018 The three questions below are meant to give a sense of the kinds of questions that might be asked on the exam in April 2018. The actual exam
CHEMISTRY OF THE HUMAN BODY (Sample Questions Key) WUCT 2018 The three questions below are meant to give a sense of the kinds of questions that might be asked on the exam in April 2018. The actual exam
Lecture 11: Protein Folding & Stability
 Structure - Function Protein Folding: What we know Lecture 11: Protein Folding & Stability 1). Amino acid sequence dictates structure. 2). The native structure represents the lowest energy state for a
Structure - Function Protein Folding: What we know Lecture 11: Protein Folding & Stability 1). Amino acid sequence dictates structure. 2). The native structure represents the lowest energy state for a
Protein Folding & Stability. Lecture 11: Margaret A. Daugherty. Fall Protein Folding: What we know. Protein Folding
 Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2003 Structure - Function Protein Folding: What we know 1). Amino acid sequence dictates structure. 2). The native structure represents
Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2003 Structure - Function Protein Folding: What we know 1). Amino acid sequence dictates structure. 2). The native structure represents
Chapter 19. Entropy, Free Energy, and Equilibrium
 Chapter 19 Entropy, Free Energy, and Equilibrium Spontaneous Physical and Chemical Processes A waterfall runs downhill A lump of sugar dissolves in a cup of coffee At 1 atm, water freezes below 0 0 C and
Chapter 19 Entropy, Free Energy, and Equilibrium Spontaneous Physical and Chemical Processes A waterfall runs downhill A lump of sugar dissolves in a cup of coffee At 1 atm, water freezes below 0 0 C and
Thermodynamics. Chem 36 Spring The study of energy changes which accompany physical and chemical processes
 Thermodynamics Chem 36 Spring 2002 Thermodynamics The study of energy changes which accompany physical and chemical processes Why do we care? -will a reaction proceed spontaneously? -if so, to what extent?
Thermodynamics Chem 36 Spring 2002 Thermodynamics The study of energy changes which accompany physical and chemical processes Why do we care? -will a reaction proceed spontaneously? -if so, to what extent?
Enzyme Catalysis & Biotechnology
 L28-1 Enzyme Catalysis & Biotechnology Bovine Pancreatic RNase A Biochemistry, Life, and all that L28-2 A brief word about biochemistry traditionally, chemical engineers used organic and inorganic chemistry
L28-1 Enzyme Catalysis & Biotechnology Bovine Pancreatic RNase A Biochemistry, Life, and all that L28-2 A brief word about biochemistry traditionally, chemical engineers used organic and inorganic chemistry
CHEM 3653 Exam # 1 (03/07/13)
") 1. Using phylogeny all living organisms can be divided into the following domains: A. Bacteria, Eukarya, and Vertebrate B. Archaea and Eukarya C. Bacteria, Eukarya, and Archaea D. Eukarya and Bacteria
1. Using phylogeny all living organisms can be divided into the following domains: A. Bacteria, Eukarya, and Vertebrate B. Archaea and Eukarya C. Bacteria, Eukarya, and Archaea D. Eukarya and Bacteria
Exam 4 April 15, 2005 CHEM 3511 Print Name: KEY Signature
 1) (8 pts) General Properties of Enzymes. Give four properties of enzymaticallycatalyzed reactions. The answers should indicate how enzymatic reactions differ from non-enzymatic reactions. Write four only
1) (8 pts) General Properties of Enzymes. Give four properties of enzymaticallycatalyzed reactions. The answers should indicate how enzymatic reactions differ from non-enzymatic reactions. Write four only
Entropy, Free Energy, and Equilibrium
 Entropy, Free Energy, and Equilibrium Chapter 17 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. 1 Spontaneous Physical and Chemical Processes A waterfall runs
Entropy, Free Energy, and Equilibrium Chapter 17 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. 1 Spontaneous Physical and Chemical Processes A waterfall runs
Basic Concepts of Metabolism. Stages of Catabolism. Key intermediates 10/12/2015. Chapter 15, Stryer Short Course
 Basic Concepts of Metabolism Chapter 15, Stryer Short Course Digestion Formation of key intermediate small molecules Formation of ATP Stages of Catabolism Key intermediates 1 Fundamental Needs for Energy
Basic Concepts of Metabolism Chapter 15, Stryer Short Course Digestion Formation of key intermediate small molecules Formation of ATP Stages of Catabolism Key intermediates 1 Fundamental Needs for Energy
SCORING. The exam consists of 5 questions totaling 100 points as broken down in this table:
 UNIVERSITY OF CALIFORNIA, BERKELEY CHEM C130/MCB C100A MIDTERM EXAMINATION #2 OCTOBER 20, 2016 INSTRUCTORS: John Kuriyan and David Savage THE TIME LIMIT FOR THIS EXAMINATION: 1 HOUR 50 MINUTES SIGNATURE:
UNIVERSITY OF CALIFORNIA, BERKELEY CHEM C130/MCB C100A MIDTERM EXAMINATION #2 OCTOBER 20, 2016 INSTRUCTORS: John Kuriyan and David Savage THE TIME LIMIT FOR THIS EXAMINATION: 1 HOUR 50 MINUTES SIGNATURE:
Bioenergetics and high-energy compounds
 Bioenergetics and high-energy compounds Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Department of Medical Chemistry and Clinical Biochemistry 2nd Faculty of Medicine, Charles University in Prague and Motol
Bioenergetics and high-energy compounds Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Department of Medical Chemistry and Clinical Biochemistry 2nd Faculty of Medicine, Charles University in Prague and Motol
schematic diagram; EGF binding, dimerization, phosphorylation, Grb2 binding, etc.
 Lecture 1: Noncovalent Biomolecular Interactions Bioengineering and Modeling of biological processes -e.g. tissue engineering, cancer, autoimmune disease Example: RTK signaling, e.g. EGFR Growth responses
Lecture 1: Noncovalent Biomolecular Interactions Bioengineering and Modeling of biological processes -e.g. tissue engineering, cancer, autoimmune disease Example: RTK signaling, e.g. EGFR Growth responses
2. In regards to the fluid mosaic model, which of the following is TRUE?
 General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
9/25/2011. Outline. Overview: The Energy of Life. I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V.
 Chapter 8 Introduction to Metabolism Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes Overview: The Energy of Life Figure 8.1 The living cell is a miniature
Chapter 8 Introduction to Metabolism Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes Overview: The Energy of Life Figure 8.1 The living cell is a miniature
Notice that this is an open system!
 Thinking About Energy and Enzymes Case Study: Frank Frank s aldehyde dehydrogenase (ALDH) enzyme has a substitution at position 487. He has the amino acid lysine at this position instead of glutamic acid.
Thinking About Energy and Enzymes Case Study: Frank Frank s aldehyde dehydrogenase (ALDH) enzyme has a substitution at position 487. He has the amino acid lysine at this position instead of glutamic acid.
Macromolecule Stability Curves
 Chem728 page 1 Spring 2012 Macromolecule Stability Curves Macromolecule Transitions - We have discussed in class the factors that determine the spontaneity of processes using conformational transitions
Chem728 page 1 Spring 2012 Macromolecule Stability Curves Macromolecule Transitions - We have discussed in class the factors that determine the spontaneity of processes using conformational transitions
Lecture 27. Transition States and Enzyme Catalysis
 Lecture 27 Transition States and Enzyme Catalysis Reading for Today: Chapter 15 sections B and C Chapter 16 next two lectures 4/8/16 1 Pop Question 9 Binding data for your thesis protein (YTP), binding
Lecture 27 Transition States and Enzyme Catalysis Reading for Today: Chapter 15 sections B and C Chapter 16 next two lectures 4/8/16 1 Pop Question 9 Binding data for your thesis protein (YTP), binding
BMB Lecture 7. Allostery and Cooperativity
 BMB 178 2017 Lecture 7 October 18, 2017 Allostery and Cooperativity A means for exquisite control Allostery: the basis of enzymatic control From the Greek: allos = other stereos = solid or space Action
BMB 178 2017 Lecture 7 October 18, 2017 Allostery and Cooperativity A means for exquisite control Allostery: the basis of enzymatic control From the Greek: allos = other stereos = solid or space Action
Metabolism and enzymes
 Metabolism and enzymes 4-11-16 What is a chemical reaction? A chemical reaction is a process that forms or breaks the chemical bonds that hold atoms together Chemical reactions convert one set of chemical
Metabolism and enzymes 4-11-16 What is a chemical reaction? A chemical reaction is a process that forms or breaks the chemical bonds that hold atoms together Chemical reactions convert one set of chemical
BCH Graduate Survey of Biochemistry
 BCH 5045 Graduate Survey of Biochemistry Instructor: Charles Guy Producer: Ron Thomas Director: Glen Graham Lecture 12 Slide sets available at: http://hort.ifas.ufl.edu/teach/guyweb/bch5045/index.html
BCH 5045 Graduate Survey of Biochemistry Instructor: Charles Guy Producer: Ron Thomas Director: Glen Graham Lecture 12 Slide sets available at: http://hort.ifas.ufl.edu/teach/guyweb/bch5045/index.html
Entropy, Free Energy and the Direction of Chemical Reactions
 Thermodynamics: Entropy, Free Energy and the Direction of Chemical Reactions Dr.ssa Rossana Galassi 320 4381420 rossana.galassi@unicam.it 20-1 Thermodynamics: Entropy, Free Energy, and the Direction of
Thermodynamics: Entropy, Free Energy and the Direction of Chemical Reactions Dr.ssa Rossana Galassi 320 4381420 rossana.galassi@unicam.it 20-1 Thermodynamics: Entropy, Free Energy, and the Direction of
BIOCHEMISTRY - CLUTCH REVIEW 2.
 !! www.clutchprep.com CONCEPT: BINDING AFFINITY Protein-ligand binding is reversible, like a chemical equilibrium [S] substrate concentration [E] enzyme concentration Ligands bind to proteins via the same
!! www.clutchprep.com CONCEPT: BINDING AFFINITY Protein-ligand binding is reversible, like a chemical equilibrium [S] substrate concentration [E] enzyme concentration Ligands bind to proteins via the same
General Biology. The Energy of Life The living cell is a miniature factory where thousands of reactions occur; it converts energy in many ways
 Course No: BNG2003 Credits: 3.00 General Biology 5. An Introduction into Cell Metabolism The Energy of Life The living cell is a miniature factory where thousands of reactions occur; it converts energy
Course No: BNG2003 Credits: 3.00 General Biology 5. An Introduction into Cell Metabolism The Energy of Life The living cell is a miniature factory where thousands of reactions occur; it converts energy
Contents. xiii. Preface v
 Contents Preface Chapter 1 Biological Macromolecules 1.1 General PrincipIes 1.1.1 Macrornolecules 1.2 1.1.2 Configuration and Conformation Molecular lnteractions in Macromolecular Structures 1.2.1 Weak
Contents Preface Chapter 1 Biological Macromolecules 1.1 General PrincipIes 1.1.1 Macrornolecules 1.2 1.1.2 Configuration and Conformation Molecular lnteractions in Macromolecular Structures 1.2.1 Weak
Computational Biology 1
 Computational Biology 1 Protein Function & nzyme inetics Guna Rajagopal, Bioinformatics Institute, guna@bii.a-star.edu.sg References : Molecular Biology of the Cell, 4 th d. Alberts et. al. Pg. 129 190
Computational Biology 1 Protein Function & nzyme inetics Guna Rajagopal, Bioinformatics Institute, guna@bii.a-star.edu.sg References : Molecular Biology of the Cell, 4 th d. Alberts et. al. Pg. 129 190
Energy Transformation and Metabolism (Outline)
") Energy Transformation and Metabolism (Outline) - Definitions & Laws of Thermodynamics - Overview of energy flow ecosystem - Biochemical processes: Anabolic/endergonic & Catabolic/exergonic - Chemical reactions
Energy Transformation and Metabolism (Outline) - Definitions & Laws of Thermodynamics - Overview of energy flow ecosystem - Biochemical processes: Anabolic/endergonic & Catabolic/exergonic - Chemical reactions
Chapter 3: Energy and Work. Energy and Work, con t. BCH 4053 Spring 2003 Chapter 3 Lecture Notes. Slide 1. Slide 2
 BCH 4053 Spring 2003 Chapter 3 Lecture Notes 1 Chapter 3: Thermodynamics of Biological Systems 2 Energy and Work Work = force x distance Energy = ability to do work Mechanical Energy Kinetic Energy = mv
BCH 4053 Spring 2003 Chapter 3 Lecture Notes 1 Chapter 3: Thermodynamics of Biological Systems 2 Energy and Work Work = force x distance Energy = ability to do work Mechanical Energy Kinetic Energy = mv
1 of 8 Class notes lectures 6a, b, c
 1 of 8 Class notes lectures 6a, b, c Last time: 1) entropy calculations 2) Gibb s Free energy Today: Field Trip READ CHAPT 15.11 before coming to field trip. Be prepared to ask questions and take notes.
1 of 8 Class notes lectures 6a, b, c Last time: 1) entropy calculations 2) Gibb s Free energy Today: Field Trip READ CHAPT 15.11 before coming to field trip. Be prepared to ask questions and take notes.
Chapter 8: Energy and Metabolism
 Chapter 8: Energy and Metabolism Why do organisms need energy? How do organisms manage their energy needs? Defining terms and issues: energy and thermodynamics metabolic reactions and energy transfers
Chapter 8: Energy and Metabolism Why do organisms need energy? How do organisms manage their energy needs? Defining terms and issues: energy and thermodynamics metabolic reactions and energy transfers
*The entropy of a system may decrease, but the entropy of the system plus its surroundings must always increase
 AP biology Notes: Metabolism Metabolism = totality of an organism's chemical process concerned with managing cellular resources. Metabolic reactions are organized into pathways that are orderly series
AP biology Notes: Metabolism Metabolism = totality of an organism's chemical process concerned with managing cellular resources. Metabolic reactions are organized into pathways that are orderly series
1. What is an ångstrom unit, and why is it used to describe molecular structures?
 1. What is an ångstrom unit, and why is it used to describe molecular structures? The ångstrom unit is a unit of distance suitable for measuring atomic scale objects. 1 ångstrom (Å) = 1 10-10 m. The diameter
1. What is an ångstrom unit, and why is it used to describe molecular structures? The ångstrom unit is a unit of distance suitable for measuring atomic scale objects. 1 ångstrom (Å) = 1 10-10 m. The diameter
Biochemical bases for energy transformations. Biochemical bases for energy transformations. Nutrition 202 Animal Energetics R. D.
 Biochemical bases for energy transformations Biochemical bases for energy transformations Nutrition 202 Animal Energetics R. D. Sainz Lecture 02 Energy originally from radiant sun energy Captured in chemical
Biochemical bases for energy transformations Biochemical bases for energy transformations Nutrition 202 Animal Energetics R. D. Sainz Lecture 02 Energy originally from radiant sun energy Captured in chemical
An Introduction to Metabolism
 An Introduction to Metabolism I. All of an organism=s chemical reactions taken together is called metabolism. A. Metabolic pathways begin with a specific molecule, which is then altered in a series of
An Introduction to Metabolism I. All of an organism=s chemical reactions taken together is called metabolism. A. Metabolic pathways begin with a specific molecule, which is then altered in a series of
A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility
 proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility") (P&S Ch 5; Fer Ch 2, 9; Palm Ch 10,11; Zub Ch 9) A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility B.
(P&S Ch 5; Fer Ch 2, 9; Palm Ch 10,11; Zub Ch 9) A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility B.
Physical Biochemistry. Kwan Hee Lee, Ph.D. Handong Global University
 Physical Biochemistry Kwan Hee Lee, Ph.D. Handong Global University Week 9 CHAPTER 4 Physical Equilibria Basic Concepts Biological organisms are highly inhomogeneous. Different regions of each cell have
Physical Biochemistry Kwan Hee Lee, Ph.D. Handong Global University Week 9 CHAPTER 4 Physical Equilibria Basic Concepts Biological organisms are highly inhomogeneous. Different regions of each cell have
Giving you the energy you need!
 Giving you the energy you need! Use your dominant hand Open and close the pin (with your thumb and forefinger) as many times as you can for 20 seconds while holding the other fingers straight out! Repeat
Giving you the energy you need! Use your dominant hand Open and close the pin (with your thumb and forefinger) as many times as you can for 20 seconds while holding the other fingers straight out! Repeat
BBS2710 Microbial Physiology. Module 5 - Energy and Metabolism
 BBS2710 Microbial Physiology Module 5 - Energy and Metabolism Topics Energy production - an overview Fermentation Aerobic respiration Alternative approaches to respiration Photosynthesis Summary Introduction
BBS2710 Microbial Physiology Module 5 - Energy and Metabolism Topics Energy production - an overview Fermentation Aerobic respiration Alternative approaches to respiration Photosynthesis Summary Introduction
Energy, Enzymes, and Metabolism. Energy, Enzymes, and Metabolism. A. Energy and Energy Conversions. A. Energy and Energy Conversions
 Energy, Enzymes, and Metabolism Lecture Series 6 Energy, Enzymes, and Metabolism B. ATP: Transferring Energy in Cells D. Molecular Structure Determines Enzyme Fxn Energy is the capacity to do work (cause
Energy, Enzymes, and Metabolism Lecture Series 6 Energy, Enzymes, and Metabolism B. ATP: Transferring Energy in Cells D. Molecular Structure Determines Enzyme Fxn Energy is the capacity to do work (cause
Biology Reading Assignment: Chapter 9 in textbook
 Biology 205 5.10.06 Reading Assignment: Chapter 9 in textbook HTTP://WUNMR.WUSTL.EDU/EDUDEV/LABTUTORIALS/CYTOCHROMES/CYTOCHROMES.HTML What does a cell need to do? propagate itself (and its genetic program)
Biology 205 5.10.06 Reading Assignment: Chapter 9 in textbook HTTP://WUNMR.WUSTL.EDU/EDUDEV/LABTUTORIALS/CYTOCHROMES/CYTOCHROMES.HTML What does a cell need to do? propagate itself (and its genetic program)
An Introduction to Metabolism
 Chapter 8 An Introduction to Metabolism Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Chapter 8 An Introduction to Metabolism Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
CHEMISTRY OF THE HUMAN BODY
 CHEMISTRY OF THE HUMAN BODY (Sample Questions) WUCT 2018 The three questions below are meant to give a sense of the kinds of questions that might be asked on the exam in April 2018. The actual exam is
CHEMISTRY OF THE HUMAN BODY (Sample Questions) WUCT 2018 The three questions below are meant to give a sense of the kinds of questions that might be asked on the exam in April 2018. The actual exam is
b) What is the gradient at room temperature? Du = J/molK * 298 K * ln (1/1000) = kj/mol
 What is the gradient at room temperature? Du = J/molK * 298 K * ln (1/1000) = kj/mol") Chem350 Practice Problems Membranes 1. a) What is the chemical potential generated by the movement of glucose by passive diffusion established by a 1000 fold concentration gradient at physiological temperature?
Chem350 Practice Problems Membranes 1. a) What is the chemical potential generated by the movement of glucose by passive diffusion established by a 1000 fold concentration gradient at physiological temperature?
Chapter 6. Ground Rules Of Metabolism
 Chapter 6 Ground Rules Of Metabolism Alcohol Dehydrogenase An enzyme Breaks down ethanol and other toxic alcohols Allows humans to drink Metabolism Is the totality of an organism s chemical reactions Arises
Chapter 6 Ground Rules Of Metabolism Alcohol Dehydrogenase An enzyme Breaks down ethanol and other toxic alcohols Allows humans to drink Metabolism Is the totality of an organism s chemical reactions Arises
4/19/2016. Chapter 17 Free Energy and Thermodynamics. First Law of Thermodynamics. First Law of Thermodynamics. The Energy Tax.
 Chemistry: A Molecular Approach, 2nd Ed. Nivaldo Tro First Law of Thermodynamics Chapter 17 Free Energy and Thermodynamics You can t win! First Law of Thermodynamics: Energy cannot be created or destroyed
Chemistry: A Molecular Approach, 2nd Ed. Nivaldo Tro First Law of Thermodynamics Chapter 17 Free Energy and Thermodynamics You can t win! First Law of Thermodynamics: Energy cannot be created or destroyed
Chapter Eighteen. Thermodynamics
 Chapter Eighteen Thermodynamics 1 Thermodynamics Study of energy changes during observed processes Purpose: To predict spontaneity of a process Spontaneity: Will process go without assistance? Depends
Chapter Eighteen Thermodynamics 1 Thermodynamics Study of energy changes during observed processes Purpose: To predict spontaneity of a process Spontaneity: Will process go without assistance? Depends