Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal
|
|
|
- Debra Carr
- 5 years ago
- Views:
Transcription
1 ARTICLES Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal Ranjan Swarup 1, Eric M. Kramer 2, Paula Perry 1, Kirsten Knox 3, H. M. Ottoline Leyser 3, Jim Haseloff 4, Gerrit T. S. Beemster 5, Rishikesh Bhalerao 6 and Malcolm J. Bennett 1,7 Re-orientation of Arabidopsis seedlings induces a rapid, asymmetric release of the growth regulator auxin from gravity-sensing columella cells at the root apex. The resulting lateral auxin gradient is hypothesized to drive differential cell expansion in elongationzone tissues. We mapped those root tissues that function to transport or respond to auxin during a gravitropic response. Targeted expression of the auxin influx facilitator AUX1 demonstrated that root gravitropism requires auxin to be transported via the lateral root cap to all elongating epidermal cells. A three-dimensional model of the root elongation zone predicted that AUX1 causes the majority of auxin to accumulate in the epidermis. Selectively disrupting the auxin responsiveness of expanding epidermal cells by expressing a mutant form of the AUX/IAA17 protein, axr3-1, abolished root gravitropism. We conclude that gravitropic curvature in Arabidopsis roots is primarily driven by the differential expansion of epidermal cells in response to an influx-carrier-dependent auxin gradient. Gravity represents an important environmental signal for plants that profoundly influences their growth and development 1 5. Roots sense changes in their orientation using specialized gravity-sensing cells that are located within the columella root cap 6 9 (Fig. 1a). They correct changes in their orientation relative to gravity by means of a differential growth response that is termed root gravitropism; cell expansion on the lower side of the elongation zone is reduced relative to the upper side, which causes the root to bend downwards 10. The root columella and elongation-zone tissues, that respectively sense 6 9 and respond 10 to the gravity stimulus, are spatially distinct (Fig. 1a), necessitating the transmission of a gravitropic signal(s). The plant growth regulator auxin represents a strong candidate for providing such a gravitropic signal, as a large number of auxin transport and response mutants exhibit root gravitropic defects Auxin transport components that are functionally required for root gravitropism include the auxin efflux facilitator PIN3; the gravity-induced retargeting of PIN3 to the lower face of columella cells helps create the initial lateral auxin gradient 16. Similarly, the expression pattern of the auxin influx and efflux facilitators AUX1 (ref. 11) and PIN2 (AGR1/EIR1/WAV6) seem to channel auxin from the root cap to the elongation zone. Nevertheless, it remains unclear whether auxin represents the gravitropic signal that acts directly on the elongation-zone cells or whether this involves other signalling intermediates Indeed, signalling molecules, such as cytokinin and nitric oxide, have recently been reported to be asymmetrically redistributed 25 or functionally required 26 during a root gravitropic response. Auxin-responsive reporters have been used successfully to visualize dynamic asymmetric changes in gravity-sensing tissues 13,27 29, which is consistent with the creation of a lateral auxin gradient. However, determining whether auxin acts as the gravitropic signal has, so far, been hampered by the insensitivity of auxin-responsive reporters to visualize dynamic expression changes in gravity-responsive elongation-zone tissues. We have adopted an alternative approach to address this important issue by directly manipulating the ability of selected root tissues to transport and/or respond to the auxin signal using a targeted expression approach 30, followed by monitoring the phenotypic effects on root gravitropism. Tissues were selected for manipulation based, in part, on computer simulations of auxin flux in a three-dimensional model of the root. In this study, we provide definitive evidence that auxin acts as the intercellular gravitropic signal by functionally mapping the root tissues that transport and respond to auxin during a gravitropic response. Our results highlight the effectiveness of adopting a predictive biology-based approach that integrates modelling and experimental studies. RESULTS Root gravitropism requires AUX1 expression in the lateral root cap and epidermis We initially addressed the question of whether root gravitropism requires auxin to be transported from the root cap to the elongation zone using the auxin influx facilitator AUX1 (ref. 31). Mutational studies had 1 School of Biosciences, University of Nottingham, Nottingham, LE12 5RD, UK. 2 Physics Department, Simon s Rock College, Great Barrington, MA 01230, USA. 3 Department of Biology, University of York, York, YO10 5YW, UK. 4 Department of Plant Sciences, University of Cambridge, Cambridge, CB2 3EA, UK. 5 Plant Systems Biology, Flemish Institute of Biotechnology (VIB)/University of Ghent, Ghent, B-9052, Belgium. 6 Umeå Plant Science Centre, SLU, Umeå, SE-90183, Sweden. 7 Correspondence should be addressed to M.J.B. ( malcolm.bennett@nottingham.ac.uk) Received 19 August 2005; accepted 26 August 2005; published online 23 October 2005; DOI: /ncb1316 NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER

 Localization of HA-AUX1 (ref.")
 Root gravitropic phenotypes of (g) aux1, AUX1>>HA-AUX1; (h) aux1, previously revealed that Arabidopsis root gravitropism required AUX1 (ref. 11).")
, we attempted to determine which of these root tissues requires the auxin influx facilitator to mediate gravitropic signalling.")
.")
, and then transformed into a null aux1 background (see Methods).")
 correctly expressed HA-AUX1 in stele, columella, LRC and epidermal tissues (Fig.")
 in columella and lower lateral root cap (LRC) cells")

2 ARTICLES a b c d e f g h i j k LRC Columella Epidermis Cortex Endodermis Pericycle Vasculature AUX1>>AUX1 M0028>>AUX1 J1071>>AUX1 J0951>>AUX1 M0013>>AUX1 l Percentage of seedlings responding to gravity stimulus Col aux1m0028>>aux1 aux1j1701>>aux1 aux1j0951>>aux1 aux1m0013>>aux1 aux1uas: AUX1 aux1m0028 aux1j1701 aux1j0951 aux1m0013 aux1-22 Time after gravity stimulus (h) Figure 1 Mapping Arabidopsis root tissues that transport auxin during a gravitropic response. (a) Schematic diagram of tissues in the Arabidopsis root apex with colour-coded key. (b f) Localization of HA-AUX1 (ref. 31) expression (red) in root tissues, counter stained with cytox green (green) of (b) aux1, AUX1>>HA-AUX1; (c) aux1, M0028>>HA-AUX1; (d) aux1, J1701>>HA- AUX1; (e) aux1, J0951>>HA-AUX1; and (f) aux1, M0013>>HA-AUX1. (g k) Root gravitropic phenotypes of (g) aux1, AUX1>>HA-AUX1; (h) aux1, previously revealed that Arabidopsis root gravitropism required AUX1 (ref. 11). Given the broad pattern of AUX1 expression in stele, columella, lateral root cap (LRC) and epidermal tissues 31 (Fig. 1b), we attempted to determine which of these root tissues requires the auxin influx facilitator to mediate gravitropic signalling. This was performed by targeting the expression of AUX1 to specific tissue domains in aux1 roots and examining the rescue of the mutant s agravitropic root phenotype (Fig. 1 and see Supplementary Information, Fig. S1). The targeted expression of a functional HA-epitope-tagged AUX1 sequence (HA-AUX1) 31 was achieved using a GAL4-based transactivation expression approach 30. The HA-AUX1 coding sequence was fused to the GAL4 recognition motif (termed the upstream activating sequence [UAS]), and then transformed into a null aux1 background (see Methods). The aux1, UAS:HA-AUX1 line was initially crossed with an AUX1 pro :GAL4 line (also in an aux1 mutant background) to demonstrate that transactivation did not interfere with AUX1-dependent gravitropic bending. Localization and gravitropic assays revealed that F1 progeny (termed aux1, AUX1>>AUX1) correctly expressed HA-AUX1 in stele, columella, LRC and epidermal tissues (Fig. 1b) and fully restored the aux1 root gravitropic defect (Fig. 1g). M0028>>HA-AUX1; (i) aux1, J1701>>HA-AUX1; (j) aux1, J0951>>HA- AUX1; and (k) aux1, M0013>>HA-AUX1. Note: Due to the requirement for limited proteolysis in the whole-mount immunolocalization procedure that was used 31, the signal for AUX1 (red) in columella and lower lateral root cap (LRC) cells seems to be less than expected compared with an AUX1 YFP fusion protein 48. (l) Kinetics of root bending for selected wild-type Col., aux1 and aux1 GAL4>>AUX1 lines. The aux1, UAS:HA-AUX1 transgenic line was crossed with a selection of tissue-specific GAL4 driver lines (see ac.uk/haseloff/genecontrol/gal4frame.html; these driver lines had also been introgressed (a backcrossing method) into an aux1 mutant background) in an attempt to rescue the mutant phenotype (see Fig. 1). Expressing HA-AUX1 in columella, LRC and epidermal tissues (Fig. 1c; M0028>>AUX1) successfully rescued aux1 root gravitropism (Fig. 1h). By contrast, expressing HA-AUX1 in stele and columella tissues (J1701>>AUX1; Fig. 1d) did not rescue aux1 gravitropism (Fig. 1i). Our M0028>>AUX1 results highlight the functional importance of basipetal (that is, root apex to base) auxin transport during a root gravitropic response. We next defined the domains of root tissues that are required to express AUX1 to rescue the aux1 phenotype. Expressing AUX1 in LRC and expanding epidermal tissues (Fig. 1e; J0951>>AUX1) rescued aux1 root gravitropism (Fig. 1j). By contrast, expressing AUX1 in just the LRC (M0013>>AUX1; Fig. 1f) was not able to rescue aux1 root gravitropism (Fig. 1k). Our targeted expression results are consistent with auxin acting as the intercellular signal that requires AUX1 to facilitate its transport via LRC and expanding epidermal cells during a gravitropic response NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER 2005
3 ARTICLES a b Pulse profile Cell number 100 µm Auxin concentration (arbitrary units) c Distance from LRC (cm) Figure 2 Modelling carrier-mediated transport of the lateral auxin gradient in the elongation zone. (a) Sketch showing a cylindrical section of the root elongation zone. During a gravistimulus, the lateral root cap (LRC; at left, not shown) deposits auxin into the lower side of the epidermis (blue arrow). Active transport then carries auxin basipetally. Shades of blue approximate the auxin concentration in the epidermis. Note that, from left to right, diffusion tends to smooth out the lateral gradient and also to reduce the overall auxin content. Inset: sketch of the model geometry, showing the three cell layers (top to bottom: epidermis, cortex and endodermis) and carrier localization. Green: cytoplasm; yellow: apoplast; grey: plasma membrane without carriers; pink: Our targeted expression results indicate that the root gravitropic response is dependent on AUX1 LRC and epidermal (but not columella) expression domains. Nevertheless, it remains possible that AUX1 columella expression is required during the early root bending response, given that the lateral auxin gradient is generated in this tissue 13, Root curvature was measured every 30 min following a 90 gravity stimulus in wild-type and aux1 J0951>>AUX1 roots expressing AUX1 in LRC and epidermal tissues (Fig. 1l and see Supplementary Information, Fig. S1). Our measurements revealed that the kinetics of the root gravitropic bending response in aux1, J0951>>AUX1 roots was as fast as in the wild type (Fig. 1l). Hence, AUX1 columella expression was not required for root gravitropism. Instead, our targeted expression studies demonstrated that root gravitropism required AUX1 expression in just LRC and epidermal tissues, presumably to facilitate the delivery of the gravity-induced lateral auxin gradient to elongation-zone tissues. Root gravitropism is dependent on AUX1 epidermal expression throughout the elongation zone As auxin moves through the elongation zone, diffusion around the circumference will diminish the lateral gradient, and radial diffusion will PIN2 in epidermis and cortex 14 plus weak PIN1 expression in epidermis, cortex and endodermis 32 ; orange: AUX1 in epidermis 31. (b) Cytoplasmic auxin concentration in all 20 epidermal cells around the circumference of the outer root. Dashed line is the profile of the auxin source applied at the LRC. Solid lines are profiles in the elongation zone, after 500 µm of basipetal transport, for the wild type (red) and aux1 mutant (blue). Each profile is normalized to a mean of 0.5. (c) Cytoplasmic auxin concentration along the length of the model wild type root (blue) and aux1 root (red). The upper red and blue lines denote auxin concentrations on the bottom side of the root, whereas the lower red and blue lines denote auxin concentrations along the top side. recycle auxin from the epidermis back to the stele 32. The auxin influx and efflux facilitators AUX1 (ref. 11) and PIN2 (refs 12 15) may function to minimize the effect of radial diffusion as well as facilitating basipetal auxin transport (that is, root apex to base; Fig. 2a). We probed the functional importance of AUX1 and PIN on the gravity-induced auxin pulse using a three-dimensional model of the Arabidopsis root elongation zone (see Methods; and see Supplementary Information text for further details). The model simulated the outer three elongation-zone tissues, encompassing ~1,200 cells and incorporating known auxin-carrier expression and localization patterns (Fig. 2a): PIN2 in the epidermis and cortex 14 ; weak PIN1 expression in the epidermis, cortex and endodermis 32 ; and AUX1 in the epidermis 11. For the tissues under study, other members of the Arabidopsis AUX1 and PIN gene families are not included in our gravitropic model based on their measured expression patterns and gravitropic phenotypes of knockout mutants (R.S. and M.J.B., unpublished observations) 32. We simulated the gravitropic signal by supplying an auxin asymmetry (a higher concentration at the bottom) to the apical end of our virtual root model (Fig. 2a, b), then monitoring the movement of the lateral auxin gradient through the elongation-zone tissues (Fig. 2c and see NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER


.")
4 ARTICLES Table 1a Characteristics of auxin transport in the epidermis of WT, aux1, pin1 and pin1pin2 mutants Lateral auxin gradient in CEZ (c bottom /c top ) Fraction of all auxin in the apoplast WT aux pin pin1pin2 NA < 10 8 Fraction of auxin pulse that reaches the CEZ The central elongation zone (CEZ) is here defined to be the portion of the root 500 µm behind the lateral root cap. Table 1b Cytoplasmic auxin concentration ratios showing the partitioning of auxin between the three cell layers of the outer root Epidermis Cortex Endodermis WT aux pin pin1pin2 NA NA NA Values normalized to the concentration in the endodermis. Supplementary Information, Fig. S2). Our modelling studies indicate that the wild-type root epidermis is competent to transport the auxin pulse with only moderate diffusion (Fig. 2b, c). Wild-type root epidermal cells can retain more than 40% of the initial auxin asymmetry 500 µm into the elongation zone (Table 1a), and can maintain the lateral auxin gradient at least 2 mm from the root apex, which is consistent with auxin transport measurements 33. Our simulations reveal that the pin2 mutant root is still able to transport the auxin pulse in elongation-zone tissues (Table 1a) due to weak epidermal PIN1 expression 32, which is consistent with auxin transport measurements 27. Instead, we predict that the pin2 agravitropic phenotype results from a block in the auxin efflux activity of LRC cells (where PIN1 is not expressed) 34, which is consistent with elevated auxin-responsive reporter expression in the pin2 LRC cells (Fig. 3c). Model roots lacking AUX1 are not competent to transport the auxin pulse (Table 1a), which is in agreement with the reduced root basipetal auxin transport 27, altered pattern of auxin-responsive reporter expression (Fig. 3b) and agravitropic root phenotype in the mutant 11. In the absence of influx-carrier activity, the majority of auxin remains in the apoplast (Table 1a), where it is relatively free to diffuse back to the pericycle 32,35. Apoplastic diffusion significantly diminishes the scale of the auxin pulse that is delivered to the aux1 central elongation zone (Table 1a), resulting in a shallow auxin gradient that is insufficient to drive a differential growth response in the aux1 root (Fig. 2b, c). Simulations using the virtual root model predict that auxin levels drop rapidly (by at least two orders of magnitude) in the basal half of the aux1 elongation zone (Fig. 2c). Such a reduction in auxin levels is likely to impact the rate of growth as expanding cells progress through the aux1 elongation zone, prompting us to perform kinematic analysis (see Methods) 36 to provide detailed measurements of root growth at different positions within the aux1 elongation zone (Fig. 4). These studies revealed that the velocity profile in the aux1 distal elongation zone was equivalent to that in the wild type, but was curtailed from the central elongation zone onwards, corresponding with the region where the LRC is absent (Fig. 4a). Plotting the derivative of velocity values in Fig. 4a to provide relative expansion rates reveals that aux1 cells do not expand to the same extent as does the wild type (Fig. 4b). However, a rate of growth a b c WT aux1 eir1 Figure 3 Auxin-responsive IAA2:uidA reporter expression. IAA2:uidA reporter expression in (a) wild-type (WT), (b) aux1 and (c) eir1/pin2 backgrounds. Note that the IAA2:uidA reporter was undetectable in lateral root cap cells of aux1, but was stained more intensely in eir1/pin2 compared with the wild type. Scale bar, 20 µm. almost the same as that in the wild type could be restored in the aux1 elongation zone by expressing AUX1 in LRC/expanding epidermal cells using the GAL4 driver line J0951 (Fig. 4a, b). Our kinematic results highlight the importance of AUX1 epidermal expression to auxin-regulated root growth, as expanding cells migrate through the elongation zone. Detailed growth measurements in Arabidopsis roots following a gravitropic stimulus have revealed that the initial curvature originated in the distal elongation zone 10. The Arabidopsis primary root elongation zone contains ~16 rapidly expanding cells in each longitudinal file of cells 36. We investigated how many of these expanding cells were required to be exposed to the lateral auxin gradient to result in a gravitropic bending response. This was determined by attempting to rescue aux1 root gravitropism by expressing AUX1 in a variable number of expanding epidermal cells using the J1092 GAL4 driver line. Localization studies revealed that ~90% of J1092>>AUX1 F1 seedling roots expressed AUX1 in just columella and LRC tissues (Fig. 5a), but AUX1 was also expressed in up to five elongating epidermal cells in ~10% of roots (Fig. 5b). Nevertheless, gravitropic assays failed to observe any evidence of a root bending response in ~50 F1 seedlings (Fig. 5c, d). Gravitropic assays were subsequently performed on a larger number of F2 aux1 J1092>>AUX1 seedlings (n = 700), which allowed identification of individuals that exhibited a wild-type gravitropic response (Fig. 5e, f). Localization studies revealed that gravitropic aux1 J1092>>AUX1 seedlings strongly expressed AUX1 in the LRC plus expanding epidermal cells throughout the elongation zone (Fig. 5g and see Supplementary Information, Fig. S3a). By contrast, agravitropic aux1 J1092>>AUX1 seedlings strongly expressed AUX1 in the columella, LRC and less than five expanding epidermal cells (Fig. 5h and see Supplementary Information, Fig. S3b). We therefore concluded from our studies on F1 and F2 aux1 J1092>>AUX1 seedlings that the root gravitropic response requires AUX1 expression in every expanding epidermal cell, thereby ensuring that the lateral auxin gradient persists as it migrates through the elongation zone and drives asymmetric root growth. The epidermal auxin response is essential for differential root growth Simulations using our root elongation zone model suggest that times more of the lateral auxin gradient accumulates in the 1060 NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER 2005
 Figure 4 Kinematic analysis of AUX1-dependent root growth.")
.")
.")
.")

 caused every root cell to express the auxin-responsive reporter IAA2:GUS (Fig.")
, several of which have been demonstrated to regulate differential")

5 ARTICLES Col aux1 UAS:AUX1 aux1 J0951>>AUX1 a b c 0 d Velocity (µm h 1 ) e 0 g h f * * 24 Strain rate (h 1 ) Distance from QC (µm) Figure 4 Kinematic analysis of AUX1-dependent root growth. Profiles of velocity (a) and relative rates of cell expansion (b) along the root elongation zone of the wild type (circle), aux1 (diamond) and aux1 J0951>>AUX1 (triangle) obtained from kinematic analysis 36 (see Methods). Note that the growth rates in aux1 are similar in the apical part of the root, but decrease below that of the controls from the end of the lateral root cap (denoted by the arrow). QC denotes quiescent centre at the root apex. epidermis compared with the underlying cortical and endodermal tissues (Table 1b). The large difference in wild-type auxin distribution that was predicted to occur in inner versus outer root tissues primarily results from AUX1 epidermal expression, as the simulated ratio of cytoplasmic auxin concentrations in the root epidermis:cortex:endodermis of the aux1 mutant was 1.55:0.78:1.00, respectively (Table 1b). Partitioning the lateral auxin gradient in wild-type roots in such a manner is predicted to result in root gravitropic bending being driven primarily by differential cell elongation in the epidermis. We tested the predicted functional importance of the epidermis for root gravitropism by selectively disrupting the auxin response in this tissue using the axr3-1 protein 17,37. The axr3-1 protein is able to disrupt auxin responses in a wide range of root tissues, including in the epidermis (Fig. 6c). For example, although external addition of the auxin indole-3-acetic acid (IAA) caused every root cell to express the auxin-responsive reporter IAA2:GUS (Fig. 6b), axr3-1 significantly disrupted reporter expression in every root tissue (Fig. 6d). As an AUX/IAA protein 17, axr3-1 negatively regulates the activity of transcription factors termed auxin response factors 38 (ARFs), several of which have been demonstrated to regulate differential growth processes 39,40. The ability of axr3-1 to disrupt ARF function was illustrated when an UAS:axr3-1 line was expressed under the control of the GAL4 driver line J1701, resulting in a root-less phenotype (R.S. and * * Figure 5 Root gravitropism requires AUX1 expression in all expanding epidermal cells. (a b) Confocal images of F1 aux1 J1092>>AUX1 seedlings expressing AUX1 (red) in (a) columella and lateral root cap and (b) up to five expanding epidermal cells in the elongation zone. Note that root tissues were counter-stained with cytox green (green). (c d) All F1 aux1 J1092>>AUX1 seedlings that were characterized were agravitropic (c) even 24 h after a gravity stimulus (d). (e f) F2 aux1 J1092>>AUX1 seedling roots were agravitropic (e) even 24 h after a gravity stimulus (f), with the exception of those seedlings denoted with an asterisk. (g h) Localization studies revealed that (g) gravitropic F2 aux1 J1092>>AUX1 seedling roots expressed AUX1 (red) in epidermal cells throughout the elongation zone, whereas (h) agravitropic F2 aux1 J1092>>AUX1 seedling roots expressed AUX1 in only a few epidermal cells. M.J.B., unpublished observations), which mimicks the ARF5 mutant, monopteros 41. The UAS:axr3-1 line was next crossed with the epidermaland/or LRC-expressed GAL4 driver lines J0951 and M0013 (Fig. 6e, f) in an attempt to disrupt auxin responses in these tissues. Consistent with this experimental goal, root bioassays revealed that J0951>>axr3-1 seedlings (but not UAS:axr3-1, J0951, M0013 or M0013>>axr3-1 lines) exhibited a growth-resistance phenotype to the auxins IAA and 2,4D (see Supplementary Information, Fig. S4). Gravitropic assays revealed that disrupting the auxin response in LRC and expanding epidermal cells in J0951>>axr3-1 seedlings completely blocked root gravitropism (Fig. 6j and see Supplementary Information, Fig. S5). By contrast, expressing axr3-1 in just LRC cells in M0013>>axr3-1 seedlings did not affect root gravitropism (Fig. 6l). Kinetic studies revealed that the gravitropic defect in J0951>>axr3-1 seedlings was comparable to the axr3-1 mutant phenotype (Fig. 6m). However, J0951>>axr3-1 exhibited only a small reduction in basal root elongation compared with its equivalent Col C24 control (Fig. 6n; Student s t-test, P = ; see Supplementary Information, Table S1). Note that the genetic background of the GAL4 driver lines (C24) versus the UAS:axr3-1 line (Col) had a far more significant effect on root growth (Fig. 6n; Student s t-test, P = 9.08, ; see Supplementary Information, Table S1). Our results demonstrate that disrupting auxin response in the expanding epidermal cells is sufficient to block differential growth during root gravitropism, without greatly affecting basal-cell elongation. NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER

 Col C24 Col C24 UAS: axr3-1 J0951 M0013")

 Auxin-responsive IAA2:GUS reporter 13,31 expression in (a, b) wild-type")
.")
; and (f) M0013 in")
 Root growth behaviour of wild type (g), J0951 (h), UAS: axr3-1 (i) and")
 Kinetics of root bending for selected wild-type Col.")

 Root growth measurements 7 d after germination for wild type (Col.")
, aux1 and axr3-1 mutants, UAS:axr3-1 and M0013 and J0951 GAL4")

6 ARTICLES a b c d e f WT g WT axr3-1 axr3-1 J0951 M0013 h i j k l Col UAS:axr3-1 J0951 J0951>>axr3-1 M0013 M0013>>axr3-1 m Percentage of seedlings responding to gravity stimulus n Time after gravity stimulus (h) Col C24 Col C24 UAS: axr3-1 J0951 M0013 J0951>>axr3-1 M0013>>axr3-1 axr3-1 Root growth (mm) 25 0 Col C24 Col C24 aux1-22 arx3-1 UAS: axr3-1 M0013 M0013>>axr3-1 J0951 J0951>>axr3-1 Figure 6 Mapping root tissues that respond to the lateral auxin gradient using axr3-1. (a d) Auxin-responsive IAA2:GUS reporter 13,31 expression in (a, b) wild-type (WT) and (c, d) axr3-1 backgrounds in response to endogenous (a, c) and external 50 µm IAA stimuli for 16 h (b, d). (e, f) Tissue-specific UAS GFP expression (green) driven by GAL4 lines (e) J0951 in expanding epidermis plus lateral root cap (LRC); and (f) M0013 in LRC alone. Background root tissues are stained with propidium iodide. (g i) Root growth behaviour of wild type (g), J0951 (h), UAS: axr3-1 (i) and J0951>>axr3-1 (j), M0013 (k) and M0013>>axr3-1 (l). (m) Kinetics of root bending for selected wild-type Col., axr3-1 mutant, UAS:axr3-1 and M0013 and J0951 GAL4 driver lines, plus F1 seedlings expressing axr3-1, in epidermal and/or LRC tissues (J0951>>axr3-1 and M0013>>axr3-1), respectively. (n) Root growth measurements 7 d after germination for wild type (Col., C24 and F1 Col. C24 control), aux1 and axr3-1 mutants, UAS:axr3-1 and M0013 and J0951 GAL4 driver lines, plus F1 seedlings expressing axr3-1, in epidermal and/or LRC tissues (J0951>>axr3-1 and M0013>>axr3-1), respectively. Error bars denote standard error. Statistical significance of differences observed are displayed in Supplementary Information, Table S1. Scale bars represent 20 µm for panels a f and 200 µm for panels g l NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER 2005
7 ARTICLES g Figure 7 A schematic model for auxin-regulated root gravitropism. A gravity stimulus (denoted by g) causes auxin to be asymmetrically redistributed (denoted by arrow width) at the root apex due to the gravity-induced retargeting of PIN efflux facilitators, such as PIN3 (ref. 16), to the lower side of the gravitysensing columella cells (coloured in grey). The resulting lateral auxin gradient is mobilized through the combined action of auxin influx and efflux facilitators AUX1 (ref. 31) and PIN2 (refs 14, 34) via lateral root cap cells (coloured in yellow) to expanding epidermal cells in the elongation zone (coloured in lilac). The lateral auxin gradient inhibits the expansion of epidermal cells on the lower side of the root relative to the upper side, causing a differential growth response that ultimately results in root curvature downwards. DISCUSSION For more than 80 years, auxin has been hypothesized to represent the signal that regulates gravitropic responses in plants 1 5. Recent work has demonstrated gravity-induced changes in auxin distribution at the root apex 13, This current study provides conclusive evidence that auxin functions as the primary gravitropic signal linking columella tissues that perceive a gravitropic stimulus and elongation-zone tissues that mediate the root bending response (Fig. 7). Our targeted expression experiments initially demonstrated that root gravitropism is dependent on the activity of the auxin-influx facilitator AUX1 in LRC and elongating epidermal cells (Fig. 1), which is consistent with the basipetal transmission of an auxin signal from the root tip to gravity-responsive tissues in the elongation zone 42 (Fig. 7). We subsequently demonstrated the functional importance of the auxin response in the elongation zone for root gravitropism through targeted expression of axr3-1 (Fig. 6). Auxin is therefore linked with the gravitropic functions of tissues that mediate the perception, transmission and response to a gravity stimulus, which is consistent with its role as the primary gravitropic signal. The targeted expression approach has also enabled us to probe the gravitropic signalling function(s) of root apical tissues (Fig. 7). Our demonstration that root gravitropism requires expression of the auxin influx facilitator AUX1 by both the LRC and the epidermis (Fig. 1) implies that both tissues perform auxin transport functions. However, LRC cells do not seem to be required for the subsequent gravitropic response phase. Disrupting the auxin response in LRC cells had no effect on root gravitropism (Fig. 6m). Therefore, LRC cells seem to function primarily to facilitate the transmission of the auxin signal from columella to elongation-zone tissues. By contrast, disrupting the auxin response in expanding epidermal (plus LRC) cells was observed to abolish root gravitropism (Fig. 6m). Taken together, these results indicate that expanding epidermal cells are required to perform both auxin transport and response functions, whereas LRC is required for auxin transport following a gravitropic stimulus. AUX1 (ref. 11) and PIN (refs 12 16) auxin influx and efflux facilitators are crucial for epidermal auxin transport function during a root gravitropic response. As only the epidermis expresses an auxin influx facilitator (AUX1) in the elongation zone 31, simulations indicate that this tissue accumulates significantly more auxin than the cortex and endodermis (Table 1b), which is in agreement with experimental observations 32,33,43. Hence, the polar localization of PIN auxin efflux facilitators in the epidermis establishes the basipetal direction of flux through the outer root. Flux may be regarded as the product of two terms a speed (set by the carrier permeabilities and the cell size) multiplied by a concentration. The relative depletion of auxin from the cortex and endodermis makes these tissues minor players in the auxin flux. Models that are run with different choices for the direction of efflux from cortical cells show only small changes in the auxin distribution (see Supplementary Information, Tables S2, S3). Thus, whereas PIN efflux facilitators establish the direction of transport, the AUX1 influx facilitator determines which cell types participate in the transport stream. There is a common misconception that, because protonated IAA is membrane permeable, influx carriers play only a supplementary role in auxin distribution. However, we estimate that cells expressing AUX1 can have a carrier-mediated IAA influx 15 times greater than the diffusive contribution (see Supplementary Information). In the absence of AUX1, the rate of IAA membrane diffusion is too slow for root gravitropism to occur. This biophysical constraint in the aux1 mutant can be overcome through the addition of the membrane-permeable auxin 1-NAA (naphthalene-1-acetic acid), which rescues the gravitropic defect 44,45. The fact that the majority of the lateral auxin gradient accumulates in the epidermis indicates that this is the root tissue that is vital for a gravitropic response. Indeed, results from our axr3-1 targeted expression studies clearly demonstrate that the auxin response in expanding epidermal cells is essential for the differential root growth response, gravitropism (Fig. 6). By contrast, the basal rate of root growth is only slightly reduced NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER
8 ARTICLES when the auxin response is disrupted in this tissue (Fig. 6n). These contrasting growth effects could be explained if the epidermis primarily regulates differential root growth, whereas basal root growth was driven by the expansion of the inner tissues. The idea that the basal rate of organ elongation is established by the inner tissues, rather than the epidermis, has received support from experiments with sunflower hypocotyls 46. In such a model, the epidermis would function to restrain basal root growth in response to a lateral auxin gradient. As the outermost root tissue, the epidermis is ideally positioned to regulate bending 47. Generating sufficient force to modify the pattern of growth of the underlying tissues is likely to necessitate the involvement of multiple epidermal cells in the elongation zone. Indeed, our J1092>>AUX1 results (Fig. 5) highlight the importance of an integrated differential growth response that involves multiple expanding epidermal cells that are resident in both distal and central elongation zones. METHODS DNA constructs and transgenic materials. The GAL4 recognition sequence UAS was sub-cloned from pic UAS tnos 30 (a gift from Dolf Weijers) into pbc (Stratagene, La Jolla, CA) to create pbc UAS. The UAS:HA-AUX1 construct contained UAS:HA-Aux1 and UAS:uidA cassettes, which were originally created as two separate constructs in pbc UAS. Full-length GUS gene (uida) containing the 5 untranslated region (UTR), and the Nos terminator and HA-tagged AUX1 genomic sequence 31 containing the 5 and 3 UTRs, were fused downstream of the UAS. The resulting UAS:uidA and UAS:HA-AUX1 cassettes were then cloned into a BIN19-based kanamycin-resistant plant transformation vector to create the final construct UAS:HA-AUX1. The axr3-1 sequence 17 was subcloned downstream of the UAS to create UAS:axr3-1. The AUX1pro:GAL4 construct was created by cloning the GAL4-VP16 (ref. 30) sequence (a gift from Dolf Weijers) into a BIN19-based kanamycin-resistant plant transformation vector, followed by insertion of a 2-kb AUX1 promoter sequence upstream of the GAL4 coding sequence. The UAS:axr3-1 was transformed into the Col background, whereas UAS:HA-AUX1 and AUX1pro:Gal4 constructs were transformed into an aux1 mutant background 48. In parallel, existing tissue-specific-expressing GAL4 driver lines (see Fig. 1) were introgressed into an aux1 mutant background. GAL4- dependent transactivation of UAS:HA-AUX1 was verified by GUS staining (as the UAS:HA-AUX1 construct also encoded an UAS:uidA gene), as well as by immunolocalization of HA-AUX1. Immunolocalization. Tissue-specific AUX1 expression was assayed by immunolocalization of HA-AUX1 (ref. 31) using anti-ha primary antibody (Roche, Lewes, UK) and Alexa-Fluor-555-coupled secondary antibody (Molecular Probes, Carlsbad, CA), then visualised using confocal microscopy. Background staining was performed with Sytox Green (Molecular Probes). Root growth measurements. To assess basal root growth, seedlings were grown vertically for 5 d and the root length measured. Two-tail t-tests were performed using Microsoft Excel software. For 2,4-dichlorophenoxyacetic acid (2,4-D) and IAA root bioassays, seedlings were grown vertically on MS plates for 5 d and then transferred to fresh MS, MS + 2,4-D (10 7 M) or MS + IAA plates ( M) for a further 2 d. The delta root growth in the presence or absence of auxin was measured and expressed as percentage root growth compared with the MS control. Kinematic analyses were performed 3 d after germination of seedlings on vertically placed seedlings. To this end, a series of overlapping time-lapse images were obtained that covered the entire growth zone using a vertically oriented microscope (Axiolab, Zeiss) with a charge-coupled device camera connected to a PC fitted with a Scion LC3 framegrabber board and ScionImage software. For each treatment, at least five replicate roots were imaged nine times at 10-s intervals. The obtained image stacks were used for calculation of velocity and strain rate profiles using RootflowRT software 49. Gravitropic assays. Seedlings were grown vertically for 4 d on MS plates and the plates were then turned at an angle of 90. Photographs were taken every 30 min up to 8 h after the gravity stimulus and then finally at 24 h (see Supplementary Information, Fig. S1, S4). The kinetics of the root gravitropic response was expressed as a percentage of seedlings responding to the gravity at a given time point. Seedlings were scored as positively responding to the gravity stimulus once they had reoriented by 45. Computer simulations. Our computer model of auxin flux in root tissue used the program AuxSim, written by E.M.K. 35 The new version modelled cells in three-dimensions, rather than in two-dimensions. The root tip was modelled as a rectangular array of cells that was 20 long 20 wide 3 deep, with periodic boundary conditions in y to mimic a cylindrical geometry. The three cell layers in z were the epidermis, cortex and endodermis. In transverse view, epidermal and cortical cells were µm 2 and endodermal cells were 14 7 µm 2. Cell lengths increased exponentially from the apex back (doubling every third row), with the first row being 10 µm long (see Fig. 2a). Cell walls were 0.5 µm thick. An additional description of the model, including model membrane permeabilities, can be found in the Supplementary Information text and Supplemental Table S4. Note: Supplementary Information is available on the Nature Cell Biology website. ACKNOWLEDGEMENTS We would like to thank the Nottingham Arabidopsis Stock Centre (NASC) for providing selected GAL4 enhancer trap lines used in this study and D. Weijers for the GAL4-related constructs. We also thank M. Broadley, J. Friml, D. Grierson, C. Hodgman, M. Holdsworth, L. Laplaze, J. Roberts, P.J. White, Z. Wilson and anonymous referees for helpful comments about the manuscript. The work was supported by the Biotechnology and Biological Sciences Research Council (R.S., P.P., K.K., H.M.O.L. and M.J.B.); European Space Agency (R.S. and M.J.B.); EU Training site grant HTMC-CT awarded to P.P.; Gatsby Charitable Foundation (J.H. and M.J.B.); Formas and V.R. (R.B.). COMPETING FINANCIAL INTERESTS The authors declare that they have no competing financial interests. Published online at Reprints and permissions information is available online at reprintsandpermissions/ 1. Muday, G. K. Auxin and tropisms. J. Plant Growth Regul. 20, (2001). 2. Moore, I. Gravitropism: Lateral thinking in auxin transport. Curr. Biol. 12, (2002). 3. Boonsirichai, K, Guan, C., Chen, R. & Masson, P. H. Root gravitropism: An experimental tool to investigate basic cellular and molecular processes underlying mechanosensing and signal transmission in plants. Annu. Rev. Plant Biol. 53, (2002). 4. Blancaflor, E. B. & Masson, P. H. Plant gravitropism. Unravelling the ups and downs of a complex process. Plant Physiol. 133, (2003). 5. Morita, M. T. & Tasaka, M. Gravity sensing and signalling. Curr. Opin. Plant Biol. 7, (2004). 6. Sack, F. D. Plant gravity sensing. Intl. Rev. Cytol. 127, (1991). 7. Blancfluor, E. B., Fasano, J. M. & Gilroy, S. Mapping the functional roles of cap cells in the response of Arabidopsis primary roots to gravity. Plant Physiol. 116, (1998). 8. Weise, S. E., Kuznetsov, O. A., Hasenstein, K. H. & Kiss, J. Z. Curvature in Arabidopsis inflorescence stems is limited to the region of amyloplast displacement. Plant Cell Physiol. 41, (2000). 9. Tanaka, A., Kobayashi, Y., Hase, Y. & Watanabe, H. Positional effect of cell inactivation on root gravitropism using heavy-ion microbeams. J. Exp. Bot. 53, (2002). 10. Mullen, J. L., Ishikawa, H. & Evans, M. L. Analysis of changes in relative elemental growth rate patterns in the elongation zone of Arabidopsis roots upon gravistimulation. Planta 206, (1991). 11. Bennett, M. J. et al. Arabidopsis AUX1 gene: A permease-like regulator of root gravitropism. Science 273, (1996). 12. Chen, R. J. et al. The Arabidopsis thaliana AGRAVITROPIC 1 gene encodes a component of the polar-auxin-transport efflux carrier. Proc. Natl Acad. Sci. USA 95, (1998). 13. Luschnig, C., Gaxiola, R., Grisafi, P. & Fink, G. EIR1, a root specific protein involved in auxin transport, is required for gravitropism in Arabidopsis thaliana. Genes Dev. 12, (1998). 14. Muller, A. et al. AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J. 17, (1998). 15. Utsuno, K., Shikanai, T., Yamada, Y. & Hashimoto, T. AGR, an agravitropic locus of Arabidopsis thaliana, encodes a novel membrane-protein family member. Plant Cell Physiol. 39, (1998). 16. Friml, J., Wisniewska, J., Benkova, E., Mendgen, K. & Palme, K. Lateral relocation of the auxin efflux regulator AtPIN3 mediates tropism in Arabidopsis. Nature 415, (2002). 17. Rouse, D., Mackay, P., Stirnberg, P., Estelle, M. & Leyser, H. M. O. Changes in auxin response from mutations in an AUX/IAA gene. Science 279, (1998). 18. Tian, Q. & Reed, J. W. Control of auxin regulated root development by the Arabidopsis thaliana SHY2/IAA3 gene. Development 126, (1999) NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER 2005
9 ARTICLES 19. Nagpal, P. et al. AXR2 encodes a member of the Aux/IAA protein family. Plant Physiol. 123, (2000). 20. Leyser, H. M. O. et al. Arabidopsis auxin-resistance gene AXR1 encodes a protein related to ubiquitin activating enzyme E1. Nature 364, (1993). 21. Fasano, J. M., Massa, G. D. & Gilroy, S. Ionic signalling in plant responses to gravity and touch. J. Plant Growth Regul. 21, (2002). 22. Plieth, C. & Trewavas, A. J. Reorientation of seedlings in the Earth s gravitational field induces cytosolic calcium transients. Plant Physiol. 129, (2002). 23. Monshausen, G. B. & Sievers, A. Basipetal propagation of gravity-induced surface ph changes along primary roots of Lepidium sativum L. Planta 215, (2002). 24. Wolverton, C., Mullen, J. L., Ishikawa, H. & Evans, M. L. Root gravitropism in response to a signal originating outside of the cap. Planta 215, (2002). 25. Aloni R., Langhans M., Aloni E. & Ullrich C. I. Role of cytokinin in the regulation of root gravitropism. Planta 220, (2004). 26. Hu, X. Y., Neill, S. J., Tang, Z. C. & Cai, W. M. Nitric oxide mediates gravitropic bending in soybean roots. Plant Physiol. 137, (2005). 27. Rashotte, A. M., DeLong, A. & Muday, G. K. Genetic and chemical reductions in protein phosphatase activity alter auxin transport, gravity response and lateral root growth. Plant Cell 13, (2001). 28. Boonsirichai, K., Sedbrook, J. C., Chen, R., Gilroy, S. & Masson, P. H. ALTERED RESPONSE TO GRAVITY is a peripheral membrane protein that modulates gravityinduced cytoplasmic alkalinization and lateral auxin transport in plant statocytes. Plant Cell 15, (2003). 29. Ottenslager, I. et al. Gravity-regulated differential auxin transport from columella to lateral root cap cells. Proc. Natl Acad. Sci. USA 100, (2003). 30. Weijers, D., van Hamburg, J.-P., van Rijn, E., Hooykaas, P. J. J. & Offringa, R. Diphtheria toxin-mediated cell ablation reveals interregional communication during Arabidopsis seed development. Plant Physiol. 133, (2003). 31. Swarup, R. et al. Localization of the auxin permease AUX1 suggests two functionally distinct hormone transport pathways operate in the Arabidopsis root apex. Genes Dev. 15, (2001). 32. Blilou, I. et al. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 433, (2005). 33. Tsurumi, S. & Ohwaki, Y. Transport of 14 C-labeled indoleacetic acid in Vicia root segments. Plant Cell Physiol. 19, (1978). 34. Friml, J., Benkova, E., Mayer, U., Palme, K. & Muster, G. Automated whole mount localisation techniques for plant seedlings. Plant J. 34, (2003). 35. Kramer, E. M. PIN and AUX/LAX proteins: their role in auxin accumulation. Trends Plant Sci. 9, (2004). 36. Beemster, G. & Baskins, T. Stunted Plant1 mediates effects of cytokinin, not auxin, on cell division and expansion in the root of Arabidopsis. Plant Physiol. 124, (2001). 37. Knox, K., Grierson, C. S. & Leyser, H. M. O. AXR3 and SHY2 interact to regulate root hair development. Development 130, (2003). 38. Tiwari, S. B., Hagen, G. & Guilfoyle, T. The role of auxin response factor domains in auxin-responsive transcription. Plant Cell 15, (2003). 39. Li, H., Johnson, P., Stepanova, A., Alonso, J. M. & Ecker, J. R. Convergence of signalling pathways in the control of differential cell growth in Arabidopsis. Dev. Cell 7, (2004). 40. Harper, R. M. et al. The NPH4 locus encodes the auxin response factor ARF7, a conditional regulator of differential growth in aerial Arabidopsis tissues. Plant Cell 12, (2000). 41. Hardtke, C. S. & Berleth, T. The Arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. EMBO J. 17, (1998). 42. Konings, H. On the mechanism of tranverse distribution of auxin in geotropically exposed pea roots. Acta Bot. Neerl. 16, (1967). 43. Ohwaki, Y. & Tsurumi, S. Auxin transport and growth in intact roots of Vicia faba. Plant Cell Physiol. 17, (1976). 44. Yamamoto, M. & Yamamoto, K. Differential effects of 1-naphthalenic acid, indole-3- acetic acid and 2,4-dichlorophenoxyacetic acid on the gravitropic response of roots in an auxin resistant mutant of Arabidopsis, aux1. Plant Cell Physiol. 39, (1998). 45. Marchant, A. et al. AUX1 regulates root gravitropism in Arabidopsis by facilitating auxin uptake. EMBO J. 18, (1999). 46. Peters, W. S. & Tomos, A. D. The mechanic state of inner tissue in the growing zone of sunflower hypocotyls and the regulation of its growth rate following excision. Plant Physiol. 123, (2000). 47. Kutschera, U. Tissue stresses in growing plant organs. Phys. Plant. 77, (1989). 48. Swarup, R. et al. Structure-function analysis of the presumptive Arabidopsis auxin permease AUX1. Plant Cell 16, (2004). 49. van der Weele, C. M. et al. A new algorithm for computational image analysis of deformable motion at high spatial and temporal resolution applied to root growth. Roughly uniform elongation in the meristem and also, after an abrupt acceleration, in the elongation zone. Plant Physiol. 132, (2003). NATURE CELL BIOLOGY VOLUME 7 NUMBER 11 NOVEMBER
10 Supplementary text providing additional details of the computer model I. Diffusive (i.e. not carrier-mediated) membrane permeabilities Protonated IAA. There is a wide range of published values for the permeability of plant lipid membranes to protonated indole-3-acetic acid (IAAH). Many of these must be discounted however, since they do not control for the likely presence of anion carriers 1,2. The only two studies we know of that control for carriers are both by Delbarre and coworkers 3,4. These report permeabilities of 0.14 cm/hr and 0.18 cm/hr respectively for suspension-cultured tobacco protoplasts. We use the model value P IAAH = 0.2 cm/hr. It should be noted that our model value is an order of magnitude lower than the oftcited result of Gutknecht and Walter 5. However, their permeability is found by fitting a straight line to data that comes almost entirely from experiments on artificial bilayers of egg lecithin. We favor measurements made on plant cell membranes, since Bean et al. 6 have shown that the permeability of bilayers to IAAH depends strongly on the composition of the bilayer (their results range between 0.3 cm/hr and 13 cm/hr). Anionic IAA. Following Refs. [ 5 ] and [ 6 ], we assume the IAA - anion has a negligible permeability. II. Carrier-mediated membrane permeabilities Measured permeability values. We begin our discussion of auxin efflux by noting that the PIN proteins are now commonly described as efflux "facilitators" rather than
11 efflux carriers 7. However, even if PIN's are not carriers, they are components of the efflux machinery, and their patterns of expression and localization are believed to be reliable indicators of auxin efflux 7,8. The permeabilities due to auxin influx carriers (the AUX/LAX gene family 9 ) and to auxin efflux facilitators (the PIN gene family 8,10 ) are not well-known. Perhaps the most useful source of quantitative data is Delbarre et al. 4, who report the diffusive and carrier mediated auxin influx and efflux in suspension-cultured tobacco protoplasts. Based on their data, we estimate an influx carrier permeability of 0.02 cm/hr and an efflux carrier permeability of 0.01 cm/hr. A second useful paper, and the only published value for influx carrier permeability in a plant tissue, is Szponarski et al. 11. They report an influx carrier permeability of cm/hr, measured in plasma membrane vesicles derived from mature Arabidopsis leaves (note the surprising agreement with the tobacco protoplast result). Szponarski's result reflects an average over all cells of the leaf, so the permeability in cells specialized for auxin transport is likely to be larger. Considering these facts, permeability values of order 0.1 cm/hr are expected to be reasonable in the root expansion zone. Model permeability values. Table S2 shows the model parameters used for carriermediated transport in the outer root. In the epidermis, we take the influx permeability to be P AUX1 = 0.2 cm/hr and the efflux permeability to be <P PIN2 > = 0.1 cm/hr. The angle brackets indicate an average over the area of the cell membrane. Note that an average is quoted since the efflux carrier permeabilties vary over the cell membrane due to the polar localization of the carriers. The model ratio of influx to efflux permeabilties (2.0) is chosen using an independent fit to the measured speed of basipetal auxin transport in root
12 tips, about 1 cm/hr 12,13. Blilou et al. 7 describe weak PIN1 expression in the epidermis, co-localized with PIN2. We thus take <P PIN1 > = 0.02 cm/hr. The remaining influx carrier entries in Table S2 are based on Blilou et al. 7. They report strong PIN2 and weak PIN1 expression in the cortex, localized to the apical (i.e. facing the root apex) end of the cells. They also report weak PIN1 expression in the endodermis, localized apically. It should however be noted that there are varying reports on the polarity of PIN2 in the cortex 14,15. We therefore ran model versions with different choices for cortical PIN2 polarity. Surprisingly, model results for the auxin distribution depend only weakly on the localization of PIN2 in the cortex (Tables S3 and S4). The reason is that cortical cells do not express an influx carrier (see Discussion in main text), and so do not accumulate much auxin as compared with the epidermis. The cortical contribution to the net auxin flux is thus relatively weak. A cell with no carrier localized to the plasma membrane is assigned a carrier permeability of zero. For example, the pin2 mutantis modeled using the values in Table S2, with the exception that all PIN2 permeabilities are set equal to zero. Fluorescence immunolabelling shows no AUX1 signal in either the cortex or the endodermis of the Arabidopsis outer root 7,16. We therefore assign zero influx carrier permeability to cells of both the cortex and endodermis. III. Diffusion coefficients Intracellular diffusion coefficient. We do not know of any direct measurements of the cytoplasmic or vacuolar diffusion of IAA (indole-3-acetic acid). Since the vacuole is
13 mostly water, we expect the vacuolar diffusion coefficient of IAA to be comparable to the known aqueous value, D aq = cm 2 /hr 17. In the cytoplasm, comparable small molecules have diffusion coefficients in the range of 10% to 40% of their aqueous value 18. The model used here does not treat the vacuole as a distinct cellular compartment. The proper choice for the diffusion coefficient of IAA inside model cells thus depends on whether IAA can freely enter the central vacuole, and on the fraction of the cell volume occupied by the vacuole. We assume IAA can enter the vacuole (although the carrier family is unknown) 19-21, and so take D = D aq inside model cells. For completeness, however, we have verified that an auxin-impermeable vacuole does not significantly change the model results (data not shown). Apoplastic diffusion coefficient. The permeability of the apoplast to small molecules is not homogeneous, having significant restrictions at Casparian bands, and also possibly within the root meristem However, these restrictions are not present in the zone of the Arabidopsis root tip studied here. Following Refs. [ 25 ] and [ 26 ], we take the apoplastic diffusion coefficient of IAA to be 0.1 D aq. IV. Membrane voltage and ph The ph and membrane voltage of plant cells have been thoroughly studied, for examples see Refs. [ ]. We use the following representative values: cytoplasm ph c = 7.2, apoplast ph w = 5.3, and cell membrane voltage V = -120 mv (negative inside).
14 V. Mathematical and numerical methods Our model extends the techniques of Goldsmith et al. 30 and Kramer 31 to the case of a three dimensional array of rectangular cells (see Fig. 2.A). The main difference with the model cells of Ref. [ 31 ] is that we do not include a separate vacuole. To simplify the model, we generally assume the tonoplast is sufficiently permeable to auxin that the cell interior can be treated as a single compartment As a check, we do conduct some runs with auxin-impermeable vacuoles and verify that they do not cause significant changes (data not shown). The tissue is divided into a rectangular array of boxes (called "control volumes" in Ref. [ 32 ]). The interior of each cell is a set of (usually 5) contiguous boxes, with the remaining boxes making up the apoplastic space. A nonuniform box size allows us to resolve the 0.5 m thick walls, despite cell lengths up to 400 m. The auxin flux between adjacent boxes within each cell, and between boxes in the apoplastic space, is governed by the discrete approximation to Fick's law for diffusion. J 1 2 = D c 2 c 1 L (1) where J 1->2 is the net flux from box 1 to box 2, c j is the concentration of auxin in box j (including both protonated and anion forms), D is the diffusion coefficient of auxin in the compartment, and L is the center-to-center distance between the boxes. The net flux across the cell membrane is governed by the uniform-field approximation 33. The net flux across any one segment of the cell membrane (i.e. between one cell box and one apoplast box) is J 1 2 = J diff + J PIN + J AUX1, where J diff, J PIN, and J AUX1
15 are due to the diffusive membrane permeability of the protonated auxin molecule, PIN family carriers, and AUX1 carriers respectively. c J diff = P 2 IAAH 1+10 c 1 ph 2 pk 1+10 ph 1 pk (2) f ( φ)c J PIN = P 2 PIN 1+10 f (φ)c 1 (3) ph 2 + pk 1+10 ph 1 + pk f (φ)c J AUX1 = P 2 AUX f ( φ)c 1 ph 2 + pk 1+10 ph 1 + pk (4) f (x) = x e x 1 (5) where ph and pk have their usual meanings, the P j are membrane permeabilities, and φ = ± FV/RT where V is the membrane potential, F is the Faraday constant, R is the gas constant, T is the temperature, and the + sign is used if box 1 is interior to box 2 (by definition, the cytoplasm is interior to the cell wall). It is instructive to substitute the parameter values into Eqns. (2) through (5) and sum to find an expression for the net auxin influx across a membrane segment J w c = ( 0.24 P IAAH P AUX P PIN )c w (6) ( P IAAH P AUX P PIN )c c where the subscript w denotes the wall, the subscript c denotes the cytoplasm, and a negative flux would be out of the cell. Note that the influx carrier permeability enters the top line of Eqn. (6) with a prefactor nearly 15 times larger than the diffusive permeability. Thus, although P IAAH and P AUX1 are of the same order of magnitude (see Secs. I and II above), the carrier dominates diffusive influx in all model cells where it is expressed. There is a common misconception that, because protonated auxin is
16 membrane permeable, influx carriers play only a supplemental role in auxin distribution. A quantitative treatment shows that this is not the case. The flux of auxin between the simulation boxes is described by a coupled set of thousands of differential equations. These differential equations are discretized using an explicit scheme as described in Chapter 4 of Ref. [ 32 ]. The equations are solved using the program AuxSim3D, written by the author and run on a PowerBook G4. VI. Assessing the speed of transport. The speed of auxin transport in the elongation zone is assessed in the following way. Figure S7a shows the auxin concentration in the central elongation zone, 500 m proximal to the source of auxin (see model figure, main text), as a function of time. The initial condition is a root empty of auxin. Starting at time zero, a constant, asymmetric auxin source is applied to the distal apoplast of the model root epidermis. After about 5 min, the auxin concentrations in the CEZ have reached a steady state. There are several ways one might assign a speed to this auxin pulse. Our preferred method is to divide the distance traveled (0.05 cm in this case) by the time it takes the auxin concentration to reach half its steady-state value. For data in Fig. S7a, these times are hr along the bottom flank of the root and hr on the top flank. These give speeds of 1.60 cm/hr and 1.46 cm/hr respectively.
17 References 1. Gimmler, H., Heilmann, B., Demming, B. & Hartung, W. The permeability coefficients of the plasmalemma and the chloroplast envelope of spinach mesophyll cells for phytohormones. Z. Naturforsch. 36 c, (1981). 2. Baier, M., Gimmler, H. & Hartung, W. The permeability of the guard cell plasma membrane and tonoplast. Journal of Experimental Botany 41, (1990). 3. Delbarre, A., Muller, P., Imhoff, V., Morgat, J.-L. & Barbier-Brygoo, H. Uptake, accumulation and metabolism of auxins in tobacco leaf protoplasts. Planta 195, (1994). 4. Delbarre, A., Muller, P. & Guern, J. Comparison of mechanisms controlling uptake and accumulation of 2,4-dichlorophenoxy acetic acid, naphthalene-1- acetic acid, and indole-3-acetic acid in suspension-cultured tobacco cells. Planta 198, (1996). 5. Gutknecht, J. & Walter, A. Transport of auxin (indoleacetic acid) through lipid bilayer membranes. Journal of Membrane Biology 56, (1980). 6. Bean, R., Shepherd, W. & Chan, H. Permeability of lipid bilayer membranes to organic solutes. Journal of General Physiology 52, (1968). 7. Blilou, I. et al. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 433, (2005). 8. Benkova, E. et al. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115, (2003). 9. Parry, G. et al. Quick on the uptake: characterization of a family of plant auxin influx carriers. Journal of Plant Growth Regulation 20, (2001). 10. Galweiler, L. et al. Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 282, (1998). 11. Szponarski, W. et al. Reconstitution of an electrogenic auxin transport activity mediated by Arabidopsis thaliana plasma membrane proteins. FEBS Letters 446, (1999). 12. Tsurumi, S. & Ohwaki, Y. Transport of 14 C-labeled indoleacetic acid in Vicia root segments. Plant and Cell Physiology 19, (1978). 13. Monshausen, G. & Sievers, A. Basipetal propagation of gravity-induced surface ph changes along primary roots of Lepidium sativum L. Planta 215, (2002). 14. Muller, A. et al. AtPIN2 defines a locus of Arabidopsis for root gravitropism control. The EMBO Journal 17, (1998). 15. Peer, W. A. et al. Variation in expression and protein localization of the PIN family of auxin efflux facilitator proteins in flavonoid mutants with altered auxin transport in Arabidopsis thaliana. Plant Cell 16, (2004). 16. Swarup, R. et al. Localization of the auxin permease AUX1 suggests two functionally distinct hormone pathways operate in the Arabidopsis root apex. Genes & Development 15, (2001).
18 17. Mitchison, G. J. The dynamics of auxin transport. Proceedings of the Royal Society of London B. Biological Sciences 209, (1980). 18. Paine, P., Moore, L. & Horowitz, S. Nuclear envelope permeability. Nature 254, (1975). 19. Ouitrakul, R. & Hertel, R. Effect of gravity and centrifugal acceleration on auxin transport in corn coleoptiles. Planta 88, (1969). 20. Cande, W., Goldsmith, M. H. M. & Ray, P. M. Polar auxin transport and auxininduced elongation in the absence of cytoplasmic streaming. Planta 111, (1973). 21. Goldsmith, M. H. M. & Ray, P. M. Intracellular localization of the active process in polar transport of auxin. Planta 111, (1973). 22. Wierzbicka, M. Lead accumulation and its translocation barriers in roots of Allium cepa L. - autoradiographic and ultrastructural studies. Plant, Cell and Environment 10, (1987). 23. Enstone, D. & Peterson, C. The apoplastic permeability of root apices. Canadian Journal of Botany 70, (1992). 24. Cholewa, E. & Peterson, C. Detecting exodermal Casparian bands in vivo and fluid-phase endocytosis in onion (Allium cepa L.) roots. Canadian Journal of Botany 79, (2001). 25. Richter, E. & Ehwald, R. Apoplastic mobility of sucrose in storage parenchyma of sugar beet. Physiologia Plantarum 58, (1983). 26. Aloni, R., Enstone, D. & Peterson, C. Indirect evidence for bulk water flow in root cortical cell walls of three dicotyledonous species. Planta 207, 1-7 (1998). 27. Scott, A. C. & Allen, N. S. Changes in the cytosolic ph within Arabidopsis root columella cells play a key role in the early signalling pathway for root gravitropism. Plant Physiology 121, (1999). 28. Sze, H., Li, X. & Palmgren, M. Energization of plant cell membranes by H+ pumping ATPases: regulation and biosynthesis. The Plant Cell 11, (1999). 29. Fasano, J. et al. Changes in root cap ph are required for the gravity response of the Arabidopsis root. Plant Cell 13, (2001). 30. Goldsmith, M. H. M., Goldsmith, T. H. & Martin, M. H. Mathematical analysis of the chemosmotic polar diffusion of auxin through plant tissues. Proc. Nat. Acad. Sci. 78, (1981). 31. Kramer, E. M. PIN and AUX/LAX proteins: their role in auxin accumulation. Trends in Plant Science 9, (2004). 32. Patankar, S. V. Numerical heat transfer and fluid flow (Taylor & Francis, Levittown, PA, 1980). 33. Dainty, J. Ion transport and electrical potentials in plant cells. Annual Review of Plant Physiology 13, (1962).
19 Table S1: t-test: Two-Sample Assuming Equal Variances Variable 1 Variable 2 t Critical twotail P(T<=t) two-tail J0951>>axr3 J J0951>>axr3 Col x C J0951>>axr3 UAS axr J0951>>axr3 Col Col x C24 Col Col x C24 UAS axr C24 Col x C E-09 C24 Col E-10 C24 UAS axr E-09 C24 J Two sample student s t-test assuming equal variance was performed on 5-7 day old roots in three separate experiments. The null hypothesis here is that the means are equal ( P> 0.05), and the alternative hypothesis is that they are not (P< 0.05). The results presented in this table are representative of the three separate experiments. Table S2 Carrier-mediated permeability values. Tissue layer <P PIN1 > <P PIN2 > (cm/hr) PIN polarity P AUX1 (cm/hr) (cm/hr) epidermis To base 0.2 cortex To apex 0 endodermis To apex 0 Carrier-mediated permeabilities and PIN polarity used in the model of wildtype elongation zone tissues. Angle brackets indicate a mean over the area of the cell membrane. "To apex" indicates that PIN permeability is highest on the portion of the cell membrane facing the root apex.
20 Table S3 Variation due to changes in the cortical localization of PIN2. PIN2 polarity in Lateral auxin Fraction of all Fraction of cortex gradient in CEZ auxin in the auxin pulse that (c bottom /c top ) apoplast reaches the CEZ To apex To base To epidermis To endodermis Changes to the wildtype entries of Table 1 resulting from various choices for the polarity of PIN2 in the model cortex. Top row is the version presented in the main text. Table S4 Cytoplasmic auxin concentrations. Cortical Mean auxin concentration in PIN2 polarity epidermis cortex endodermis To apex To base To epidermis To endodermis Model results for the partitioning of auxin between the three cell layers of the outer root, given different choices for the polarity of PIN2 in the cortex. Auxin concentrations have arbitrary units.


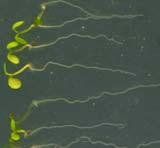
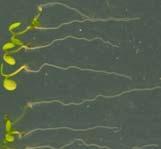









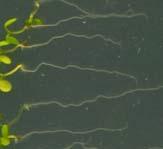
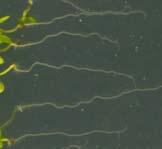



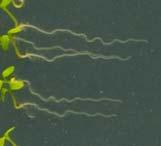
















21 Swarup et al Fig S1 Col J0951>Aux Fig. S1 Seedlings were grown vertically for four days on MS plates and the plates were then turned at an angle of 90. Photographs were taken every 30 minute up to 8 hours and then at 24 hours after the change in the direction of the gravity vector. The figure illustrates results from wildtype (Col.) and J0951>AUX1 lines and are representative of data collected for all other lines which is summarised in Fig. 1i.
22 Swarup et al Fig S2 a Auxin concentration (arb. units) t (min) b Auxin concentration (arb. units) t (min) Fig. S2 Time-dependence of the epidermal auxin concentration. Cytoplasmic auxin concentration in two epidermal cells located 500 microns proximal to the lateral root cap. The concentrations on lower and upper sides of the roots are depicted in blue and red, respectively. (a) To assess the speed of the auxin pulse, the initial condition was a root with no auxin. (b) To visualize the gravitropic signal, the initial condition was a vertical root (concentrations were equilibrated to a uniform distal auxin source before time zero). An auxin gradient is supplied to the distal epidermal apoplast beginning at time zero.

 vs agravitropic")

23 Swarup et al Fig S3 a b c d e f Fig. S3 Root gravitropism requires AUX1 to be expressed in more than 5 elongating epidermal cells. Comparison of HA-AUX1 localisation in gravitropic (top) vs agravitropic (bottom) aux >>Aux1 seedlings. Immunolocalisation was performed with anti HA primary antibody and Alexa Fluor 555 coupled secondary antibody (b & e). Background staining was performed with Sytox Green (a & c). Scale bar 40 m.
24 Swarup et al Fig S4 120 % root growth compared to control Col a1-22 axr3 C24 Col x C24 UAS axr3 M0013 M0013>>axr3 J0951 J0951>>axr3 Fig. S4 Disrupting auxin perception in epidermal cells results in increased auxin resistance Seedlings were grown vertically on MS plates for five days and were then transferred on fresh MS, MS + 2,4-D (10-7 M) or MS + IAA plates (2.5 x 10-7 M) for further two days. The delta root growth in presence or absence of auxin was measured and expressed as percent root growth compared to MS control. Error bars represent standard deviation.


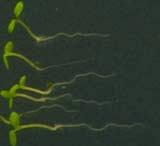
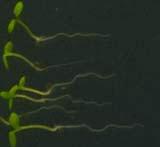



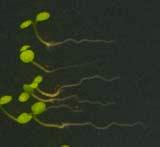


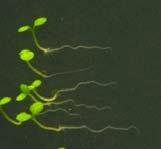
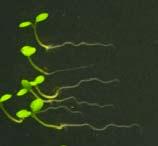
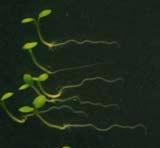


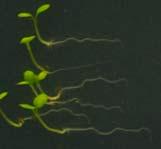















25 Swarup et al Fig S5 J J0951>>axr Fig. S5 Seedlings were grown vertically for four days on MS plates and the plates were then turned at an angle of 90. Photographs were taken every 30 minute up to 8 hours and then at 24 hours after the change in the direction of the gravity vector. The figure illustrates results from wildtype (Col.) and J0951>axr3-1 lines and are representative of data collected for all other lines which is summarised in Fig. 6m.
Supplementary text providing additional details of the computer model. I. Diffusive (i.e. not carrier-mediated) membrane permeabilities
 membrane permeabilities") Supplementary text providing additional details of the computer model I. Diffusive (i.e. not carrier-mediated) membrane permeabilities Protonated IAA. There is a wide range of published values for the
Supplementary text providing additional details of the computer model I. Diffusive (i.e. not carrier-mediated) membrane permeabilities Protonated IAA. There is a wide range of published values for the
23-. Shoot and root development depend on ratio of IAA/CK
 Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Actions of auxin. Hormones: communicating with chemicals History: Discovery of a growth substance (hormone- auxin)
") Hormones: communicating with chemicals History- discovery of plant hormone. Auxin Concepts of hormones Auxin levels are regulated by synthesis/degradation, transport, compartmentation, conjugation. Polar
Hormones: communicating with chemicals History- discovery of plant hormone. Auxin Concepts of hormones Auxin levels are regulated by synthesis/degradation, transport, compartmentation, conjugation. Polar
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
Gravity plays a major role in plant morphogenesis by determining
 Gravity-regulated differential auxin transport from columella to lateral root cap cells Iris Ottenschläger*, Patricia Wolff*, Chris Wolverton, Rishikesh P. Bhalerao,Göran Sandberg, Hideo Ishikawa, Mike
Gravity-regulated differential auxin transport from columella to lateral root cap cells Iris Ottenschläger*, Patricia Wolff*, Chris Wolverton, Rishikesh P. Bhalerao,Göran Sandberg, Hideo Ishikawa, Mike
GFP GAL bp 3964 bp
 Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
Abscisic Acid Represses Growth of the Arabidopsis Embryonic Axis after Germination by Enhancing Auxin Signaling W
 The Plant Cell, Vol. 21: 2253 2268, August 2009, www.plantcell.org ã 2009 American Society of Plant Biologists Abscisic Acid Represses Growth of the Arabidopsis Embryonic Axis after Germination by Enhancing
The Plant Cell, Vol. 21: 2253 2268, August 2009, www.plantcell.org ã 2009 American Society of Plant Biologists Abscisic Acid Represses Growth of the Arabidopsis Embryonic Axis after Germination by Enhancing
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
BIO1PS 2012 Plant Science Lecture 4 Hormones Pt. I
 BIO1PS 2012 Plant Science Lecture 4 Hormones Pt. I Dr. Michael Emmerling Department of Botany Room 410 m.emmerling@latrobe.edu.au Hormones and Ghost gum Eucalyptus papuana Coordination ~3 Lectures Leaves
BIO1PS 2012 Plant Science Lecture 4 Hormones Pt. I Dr. Michael Emmerling Department of Botany Room 410 m.emmerling@latrobe.edu.au Hormones and Ghost gum Eucalyptus papuana Coordination ~3 Lectures Leaves
LECTURE 4: PHOTOTROPISM
 http://smtom.lecture.ub.ac.id/ Password: https://syukur16tom.wordpress.com/ LECTURE 4: PHOTOTROPISM LECTURE FLOW 1. 2. 3. 4. 5. INTRODUCTION DEFINITION INITIAL STUDY PHOTROPISM MECHANISM PHOTORECEPTORS
http://smtom.lecture.ub.ac.id/ Password: https://syukur16tom.wordpress.com/ LECTURE 4: PHOTOTROPISM LECTURE FLOW 1. 2. 3. 4. 5. INTRODUCTION DEFINITION INITIAL STUDY PHOTROPISM MECHANISM PHOTORECEPTORS
Plants are sessile. 10d-17/giraffe-grazing.jpg
 Plants are sessile www.mccullagh.org/db9/ 10d-17/giraffe-grazing.jpg Plants have distinct requirements because of their sessile nature Organism-level requirements Must adjust to environment at given location
Plants are sessile www.mccullagh.org/db9/ 10d-17/giraffe-grazing.jpg Plants have distinct requirements because of their sessile nature Organism-level requirements Must adjust to environment at given location
Apical dominance models can generate basipetal patterns of bud activation
 Apical dominance models can generate basipetal patterns of bud activation Przemyslaw Prusinkiewicz 1, Richard S. Smith 1 and Ottoline Leyser 2 1 Department of Computer Science, University of Calgary 2
Apical dominance models can generate basipetal patterns of bud activation Przemyslaw Prusinkiewicz 1, Richard S. Smith 1 and Ottoline Leyser 2 1 Department of Computer Science, University of Calgary 2
Ethylene Regulates Root Growth through Effects on Auxin Biosynthesis and Transport-Dependent Auxin Distribution W
 The Plant Cell, Vol. 19: 2197 2212, July 2007, www.plantcell.org ª 2007 American Society of Plant Biologists Ethylene Regulates Root Growth through Effects on Auxin Biosynthesis and Transport-Dependent
The Plant Cell, Vol. 19: 2197 2212, July 2007, www.plantcell.org ª 2007 American Society of Plant Biologists Ethylene Regulates Root Growth through Effects on Auxin Biosynthesis and Transport-Dependent
How plants respond to their environment
 Travis Lick Biology How plants respond to their environment Plants, with their roots firmly fixed in the earth, seem immobile and vulnerable compared to animals, but this does not prevent them from reacting
Travis Lick Biology How plants respond to their environment Plants, with their roots firmly fixed in the earth, seem immobile and vulnerable compared to animals, but this does not prevent them from reacting
expression of AUX1, LAX1, LAX2 and LAX3 in etiolated hypocotyls.
 Table of Contents Supplementary Material and Methods Supplementary Figure S1: Phototropic response of various aux1lax mutants and expression of AUX1, LAX1, LAX2 and LAX3 in etiolated hypocotyls. Supplementary
Table of Contents Supplementary Material and Methods Supplementary Figure S1: Phototropic response of various aux1lax mutants and expression of AUX1, LAX1, LAX2 and LAX3 in etiolated hypocotyls. Supplementary
Cytokinin. Fig Cytokinin needed for growth of shoot apical meristem. F Cytokinin stimulates chloroplast development in the dark
 Cytokinin Abundant in young, dividing cells Shoot apical meristem Root apical meristem Synthesized in root tip, developing embryos, young leaves, fruits Transported passively via xylem into shoots from
Cytokinin Abundant in young, dividing cells Shoot apical meristem Root apical meristem Synthesized in root tip, developing embryos, young leaves, fruits Transported passively via xylem into shoots from
Root System Architecture from Coupling Cell Shape to Auxin Transport
 Root System Architecture from Coupling Cell Shape to Auxin Transport PLoS BIOLOGY Marta Laskowski 1,2[, Verônica A. Grieneisen 3[, Hugo Hofhuis 2[, Colette A. ten Hove 2, Paulien Hogeweg 3, Athanasius
Root System Architecture from Coupling Cell Shape to Auxin Transport PLoS BIOLOGY Marta Laskowski 1,2[, Verônica A. Grieneisen 3[, Hugo Hofhuis 2[, Colette A. ten Hove 2, Paulien Hogeweg 3, Athanasius
Gravitropism of Arabidopsis thaliana Roots Requires the Polarization of PIN2 toward the Root Tip in Meristematic Cortical Cells C W
 This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
10/4/2017. Chapter 39
 Chapter 39 1 Reception 1 Reception 2 Transduction CYTOPLASM CYTOPLASM Cell wall Plasma membrane Phytochrome activated by light Cell wall Plasma membrane Phytochrome activated by light cgmp Second messenger
Chapter 39 1 Reception 1 Reception 2 Transduction CYTOPLASM CYTOPLASM Cell wall Plasma membrane Phytochrome activated by light Cell wall Plasma membrane Phytochrome activated by light cgmp Second messenger
CONTROL OF PLANT GROWTH AND DEVELOPMENT BI-2232 RIZKITA R E
 CONTROL OF PLANT GROWTH AND DEVELOPMENT BI-2232 RIZKITA R E The development of a plant the series of progressive changes that take place throughout its life is regulated in complex ways. Factors take part
CONTROL OF PLANT GROWTH AND DEVELOPMENT BI-2232 RIZKITA R E The development of a plant the series of progressive changes that take place throughout its life is regulated in complex ways. Factors take part
ENDODERMIS & POLARITY
 https://en.wikipedia.org/wiki/casparian_strip ENDODERMIS & POLARITY Niloufar Pirayesh 13.01.2016 PCDU SEMINAR 2 What is Endodermis? It helps with Regulate the movement of water ions and hormones. (in and
https://en.wikipedia.org/wiki/casparian_strip ENDODERMIS & POLARITY Niloufar Pirayesh 13.01.2016 PCDU SEMINAR 2 What is Endodermis? It helps with Regulate the movement of water ions and hormones. (in and
Cell Polarity Signaling in Arabidopsis Involves a BFA-Sensitive Auxin Influx Pathway
 Current Biology, Vol. 12, 329 334, February 19, 2002, 2002 Elsevier Science Ltd. All rights reserved. PII S0960-9822(02)00654-1 Cell Polarity Signaling in Arabidopsis Involves a BFA-Sensitive Auxin Influx
Current Biology, Vol. 12, 329 334, February 19, 2002, 2002 Elsevier Science Ltd. All rights reserved. PII S0960-9822(02)00654-1 Cell Polarity Signaling in Arabidopsis Involves a BFA-Sensitive Auxin Influx
Research. Simon Moore 1 *, Xiaoxian Zhang 2 *, Anna Mudge 1, James H. Rowe 1, Jennifer F. Topping 1, Junli Liu 1 and Keith Lindsey 1.
 Research Spatiotemporal modelling of hormonal crosstalk explains the level and patterning of hormones and gene expression in Arabidopsis thaliana wild-type and mutant roots Simon Moore 1 *, Xiaoxian Zhang
Research Spatiotemporal modelling of hormonal crosstalk explains the level and patterning of hormones and gene expression in Arabidopsis thaliana wild-type and mutant roots Simon Moore 1 *, Xiaoxian Zhang
Lecture 4: Radial Patterning and Intercellular Communication.
 Lecture 4: Radial Patterning and Intercellular Communication. Summary: Description of the structure of plasmodesmata, and the demonstration of selective movement of solutes and large molecules between
Lecture 4: Radial Patterning and Intercellular Communication. Summary: Description of the structure of plasmodesmata, and the demonstration of selective movement of solutes and large molecules between
A developmental geneticist s guide to roots Find out about the hidden half of plants
 the Centre for Plant Integrative Biology A developmental geneticist s guide to roots Find out about the hidden half of plants What do roots look like from the inside? How do roots form? Can we improve
the Centre for Plant Integrative Biology A developmental geneticist s guide to roots Find out about the hidden half of plants What do roots look like from the inside? How do roots form? Can we improve
Plant Responses. NOTE: plant responses involve growth and changes in growth. Their movement is much slower than that of animals.
 Plant Responses A stimulus is anything that causes a reaction in an organism. Examples: light, gravity and temperature A response is the activity of an organism as a result of a stimulus. Examples: Growth,
Plant Responses A stimulus is anything that causes a reaction in an organism. Examples: light, gravity and temperature A response is the activity of an organism as a result of a stimulus. Examples: Growth,
DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Other cellular processes (17%)
 Signal transduction (2%) Development (2%) Other cellular processes (17%)") Fig. 35-24 Other metabolism (18%) DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Unknown (24%) Energy pathways (3%) Cell division and organization (3%) Transport (4%) Transcription
Fig. 35-24 Other metabolism (18%) DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Unknown (24%) Energy pathways (3%) Cell division and organization (3%) Transport (4%) Transcription
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL
 GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
Elisabeth J Chapman and Mark Estelle
 Minireview Cytokinin and auxin intersection in root meristems Elisabeth J Chapman and Mark Estelle Address: Division of Biology, 9500 Gilman Drive, La Jolla, CA 92093-0116, USA. Correspondence: Mark Estelle.
Minireview Cytokinin and auxin intersection in root meristems Elisabeth J Chapman and Mark Estelle Address: Division of Biology, 9500 Gilman Drive, La Jolla, CA 92093-0116, USA. Correspondence: Mark Estelle.
Topic 14. The Root System. II. Anatomy of an Actively Growing Root Tip
 Topic 14. The Root System Introduction. This is the first of two lab topics that focus on the three plant organs (root, stem, leaf). In these labs we want you to recognize how tissues are organized in
Topic 14. The Root System Introduction. This is the first of two lab topics that focus on the three plant organs (root, stem, leaf). In these labs we want you to recognize how tissues are organized in
Supplemental Data. Wang et al. (2014). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Useful Propagation Terms. Propagation The application of specific biological principles and concepts in the multiplication of plants.
 Useful Propagation Terms Propagation The application of specific biological principles and concepts in the multiplication of plants. Adventitious Typically describes new organs such as roots that develop
Useful Propagation Terms Propagation The application of specific biological principles and concepts in the multiplication of plants. Adventitious Typically describes new organs such as roots that develop
Last time: Obtaining information from a cloned gene
 Last time: Obtaining information from a cloned gene Objectives: 1. What is the biochemical role of the gene? 2. Where and when is the gene expressed (transcribed)? 3. Where and when is the protein made?
Last time: Obtaining information from a cloned gene Objectives: 1. What is the biochemical role of the gene? 2. Where and when is the gene expressed (transcribed)? 3. Where and when is the protein made?
Synthesis of IAA is not always site of action, so must be moved to other locations: R-COO- R-COO-
 AUXIN TRANSPORT Synthesis of IAA is not always site of action, so must be moved to other locations: IAA lipid soluble in undissociated form, weakly acidic. R-COOH ph5 R-COOH R-COO- R-COO- keep ph gradient.
AUXIN TRANSPORT Synthesis of IAA is not always site of action, so must be moved to other locations: IAA lipid soluble in undissociated form, weakly acidic. R-COOH ph5 R-COOH R-COO- R-COO- keep ph gradient.
SUPPLEMENTARY INFORMATION
 PRC2 represses dedifferentiation of mature somatic cells in Arabidopsis Momoko Ikeuchi 1 *, Akira Iwase 1 *, Bart Rymen 1, Hirofumi Harashima 1, Michitaro Shibata 1, Mariko Ohnuma 1, Christian Breuer 1,
PRC2 represses dedifferentiation of mature somatic cells in Arabidopsis Momoko Ikeuchi 1 *, Akira Iwase 1 *, Bart Rymen 1, Hirofumi Harashima 1, Michitaro Shibata 1, Mariko Ohnuma 1, Christian Breuer 1,
Potassium carrier TRH1 is required for auxin transport in Arabidopsis roots
 The Plant Journal (2004) 40, 523 535 doi: 10.1111/j.1365-313X.2004.02230.x Potassium carrier TRH1 is required for auxin transport in Arabidopsis roots Francisco Vicente-Agullo 1, Stamatis Rigas 2, Guilhem
The Plant Journal (2004) 40, 523 535 doi: 10.1111/j.1365-313X.2004.02230.x Potassium carrier TRH1 is required for auxin transport in Arabidopsis roots Francisco Vicente-Agullo 1, Stamatis Rigas 2, Guilhem
Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated
 Relative conversion rates of different plant hormones to their glucosylated") Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated form by recombinant UGT74E2. The naturally occurring auxin
Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated form by recombinant UGT74E2. The naturally occurring auxin
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
Supplemental Data. Perrella et al. (2013). Plant Cell /tpc
. Plant Cell /tpc") Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
Supplementary Material For: Modeling halotropism: A key role for root tip architecture and reflux loop remodeling in redistributing auxin
 Supplementary Material For: Modeling halotropism: A key role for root tip architecture and reflux loop remodeling in redistributing auxin Thea van den Berg, Ruud Korver, Christa Testerink, Kirsten ten
Supplementary Material For: Modeling halotropism: A key role for root tip architecture and reflux loop remodeling in redistributing auxin Thea van den Berg, Ruud Korver, Christa Testerink, Kirsten ten
POTASSIUM IN PLANT GROWTH AND YIELD. by Ismail Cakmak Sabanci University Istanbul, Turkey
 POTASSIUM IN PLANT GROWTH AND YIELD by Ismail Cakmak Sabanci University Istanbul, Turkey Low K High K High K Low K Low K High K Low K High K Control K Deficiency Cakmak et al., 1994, J. Experimental Bot.
POTASSIUM IN PLANT GROWTH AND YIELD by Ismail Cakmak Sabanci University Istanbul, Turkey Low K High K High K Low K Low K High K Low K High K Control K Deficiency Cakmak et al., 1994, J. Experimental Bot.
The mode of development in animals and plants is different
 The mode of development in animals and plants is different Outcome of animal embryogenesis is a mini edition of the adult Outcome of plant embryogenesis is a simple structure with -root apical meristem
The mode of development in animals and plants is different Outcome of animal embryogenesis is a mini edition of the adult Outcome of plant embryogenesis is a simple structure with -root apical meristem
COMPETITIVE CANALIZATION OF AUXIN IN PEA CAN BE INVOLVED IN INITIATION OF AXILLARY BUD OUTGROWTH
 COMPETITIVE CANALIZATION OF AUXIN IN PEA CAN BE INVOLVED IN INITIATION OF AXILLARY BUD OUTGROWTH Medveďová Z. 1, Balla J. 1, 2, Procházka S. 1 1 CEITEC - Central European Institute of Technology, Mendel
COMPETITIVE CANALIZATION OF AUXIN IN PEA CAN BE INVOLVED IN INITIATION OF AXILLARY BUD OUTGROWTH Medveďová Z. 1, Balla J. 1, 2, Procházka S. 1 1 CEITEC - Central European Institute of Technology, Mendel
Mathematical Biology. Multiscale modelling of auxin transport in the plant-root elongation zone. L. R. Band J. R. King
 J. Math. Biol. (2012) 65:743 785 DOI 10.1007/s00285-011-0472-y Mathematical Biology Multiscale modelling of auxin transport in the plant-root elongation zone L. R. Band J. R. King Received: 17 September
J. Math. Biol. (2012) 65:743 785 DOI 10.1007/s00285-011-0472-y Mathematical Biology Multiscale modelling of auxin transport in the plant-root elongation zone L. R. Band J. R. King Received: 17 September
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
Ethylene Upregulates Auxin Biosynthesis in Arabidopsis Seedlings to Enhance Inhibition of Root Cell Elongation W
 The Plant Cell, Vol. 19: 2186 2196, July 2007, www.plantcell.org ª 2007 American Society of Plant Biologists Ethylene Upregulates Auxin Biosynthesis in Arabidopsis Seedlings to Enhance Inhibition of Root
The Plant Cell, Vol. 19: 2186 2196, July 2007, www.plantcell.org ª 2007 American Society of Plant Biologists Ethylene Upregulates Auxin Biosynthesis in Arabidopsis Seedlings to Enhance Inhibition of Root
Plant morphogenesis: long-distance coordination and local patterning Thomas Berleth* and Tsvi Sachs
 57 Plant morphogenesis: long-distance coordination and local patterning Thomas Berleth* and Tsvi Sachs The overall morphology of a plant is largely determined by developmental decisions taken within or
57 Plant morphogenesis: long-distance coordination and local patterning Thomas Berleth* and Tsvi Sachs The overall morphology of a plant is largely determined by developmental decisions taken within or
Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha
 Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha Plasmids 1. Extrachromosomal DNA, usually circular-parasite 2. Usually encode ancillary
Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha Plasmids 1. Extrachromosomal DNA, usually circular-parasite 2. Usually encode ancillary
 DOI: 10.1038/ncb2819 Gαi3 / Actin / Acetylated Tubulin Gαi3 / Actin / Acetylated Tubulin a a Gαi3 a Actin Gαi3 WT Gαi3 WT Gαi3 WT b b Gαi3 b Actin Gαi3 KO Gαi3 KO Gαi3 KO # # Figure S1 Loss of protein
DOI: 10.1038/ncb2819 Gαi3 / Actin / Acetylated Tubulin Gαi3 / Actin / Acetylated Tubulin a a Gαi3 a Actin Gαi3 WT Gαi3 WT Gαi3 WT b b Gαi3 b Actin Gαi3 KO Gαi3 KO Gαi3 KO # # Figure S1 Loss of protein
Chapter 6. General discussion
 Chapter 6 General discussion 67 Chapter 6 General discussion Plants react in various ways on environmental stress conditions. The riverplain species Rumex palustris responds to submergence with an upward
Chapter 6 General discussion 67 Chapter 6 General discussion Plants react in various ways on environmental stress conditions. The riverplain species Rumex palustris responds to submergence with an upward
Formation of polarity convergences underlying shoot outgrowths
 RESEARCH ARTICLE Formation of polarity convergences underlying shoot outgrowths Katie Abley, Susanna Sauret-Güeto, Athanasius FM Marée, Enrico Coen* John Innes Centre, Norwich Research Park, Norwich, United
RESEARCH ARTICLE Formation of polarity convergences underlying shoot outgrowths Katie Abley, Susanna Sauret-Güeto, Athanasius FM Marée, Enrico Coen* John Innes Centre, Norwich Research Park, Norwich, United
Polar auxin transport old questions and new concepts?
 Plant Molecular Biology 49: 273 284, 2002. Perrot-Rechenmann and Hagen (Eds.), Auxin Molecular Biology. 2002 Kluwer Academic Publishers. Printed in the Netherlands. 273 Polar auxin transport old questions
Plant Molecular Biology 49: 273 284, 2002. Perrot-Rechenmann and Hagen (Eds.), Auxin Molecular Biology. 2002 Kluwer Academic Publishers. Printed in the Netherlands. 273 Polar auxin transport old questions
Essential idea: Plants adapt their growth to environmental conditions.
 9.3 Growth in plants AHL https://c1.staticflickr.com/3/2347/2573372542_a959ecfd4f_b.jpg Essential idea: Plants adapt their growth to environmental conditions. Boxwood, Pivet and Yew are plants commonly
9.3 Growth in plants AHL https://c1.staticflickr.com/3/2347/2573372542_a959ecfd4f_b.jpg Essential idea: Plants adapt their growth to environmental conditions. Boxwood, Pivet and Yew are plants commonly
CONTROL SYSTEMS IN PLANTS
 AP BIOLOGY PLANTS FORM & FUNCTION ACTIVITY #5 NAME DATE HOUR CONTROL SYSTEMS IN PLANTS HORMONES MECHANISM FOR HORMONE ACTION Plant Form and Function Activity #5 page 1 CONTROL OF CELL ELONGATION Plant
AP BIOLOGY PLANTS FORM & FUNCTION ACTIVITY #5 NAME DATE HOUR CONTROL SYSTEMS IN PLANTS HORMONES MECHANISM FOR HORMONE ACTION Plant Form and Function Activity #5 page 1 CONTROL OF CELL ELONGATION Plant
Sequential induction of auxin efflux and influx carriers regulates lateral root emergence
 Molecular Systems Biology 9; Article number 699; doi:1.138/msb.213.43 Citation: Molecular Systems Biology 9:699 www.molecularsystemsbiology.com Sequential induction of auxin efflux and influx carriers
Molecular Systems Biology 9; Article number 699; doi:1.138/msb.213.43 Citation: Molecular Systems Biology 9:699 www.molecularsystemsbiology.com Sequential induction of auxin efflux and influx carriers
Chapter 33 Plant Responses
 Chapter 33 Plant Responses R. Cummins 1 Chapter 33 Plant Responses External Factors Light, Day Length, Gravity, Temperature Internal Factors Hormones R. Cummins 2 Tropisms R. Cummins 3 Phototropism and
Chapter 33 Plant Responses R. Cummins 1 Chapter 33 Plant Responses External Factors Light, Day Length, Gravity, Temperature Internal Factors Hormones R. Cummins 2 Tropisms R. Cummins 3 Phototropism and
Functional redundancy of PIN proteins is accompanied by auxindependent cross-regulation of PIN expression
 Research article 4521 Functional redundancy of PIN proteins is accompanied by auxindependent cross-regulation of PIN expression Anne Vieten 1, Steffen Vanneste 2, Justyna Wiśniewska 1,3, Eva Benková 1,
Research article 4521 Functional redundancy of PIN proteins is accompanied by auxindependent cross-regulation of PIN expression Anne Vieten 1, Steffen Vanneste 2, Justyna Wiśniewska 1,3, Eva Benková 1,
Molecular Genetics of. Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS
 Molecular Genetics of Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS Contents Preface A Word on Genetic Nomenclature page xiii xvii 1 Approaches to the Study of Plant Development 1 Pattern
Molecular Genetics of Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS Contents Preface A Word on Genetic Nomenclature page xiii xvii 1 Approaches to the Study of Plant Development 1 Pattern
Biological Roles of Cytokinins
 Direct Control of Shoot Meristem Activity by a Cytokinin-Activating Enzyme By Kurakawa et. Al. Published in Nature Presented by Boyana Grigorova Biological Roles of Cytokinins Cytokinins are positive regulators
Direct Control of Shoot Meristem Activity by a Cytokinin-Activating Enzyme By Kurakawa et. Al. Published in Nature Presented by Boyana Grigorova Biological Roles of Cytokinins Cytokinins are positive regulators
A. Stimulus Response:
 Plant Hormones A. Stimulus Response: A house plant on a windowsill grows light. If you rotate the plant, it reorients its growth until its leaves face the window again. The growth of a shoot towards light
Plant Hormones A. Stimulus Response: A house plant on a windowsill grows light. If you rotate the plant, it reorients its growth until its leaves face the window again. The growth of a shoot towards light
Ron et al SUPPLEMENTAL DATA
 Ron et al SUPPLEMENTAL DATA Hairy root transformation using Agrobacterium rhizogenes as a tool for exploring cell type-specific gene expression and function using tomato as a model Mily Ron, Kaisa Kajala,
Ron et al SUPPLEMENTAL DATA Hairy root transformation using Agrobacterium rhizogenes as a tool for exploring cell type-specific gene expression and function using tomato as a model Mily Ron, Kaisa Kajala,
The auxin influx carrier is essential for correct leaf positioning
 The Plant Journal (2002) 32, 509 517 The auxin influx carrier is essential for correct leaf positioning Pia A. Stieger, Didier Reinhardt and Cris Kuhlemeier Institute of Plant Sciences, University of Berne,
The Plant Journal (2002) 32, 509 517 The auxin influx carrier is essential for correct leaf positioning Pia A. Stieger, Didier Reinhardt and Cris Kuhlemeier Institute of Plant Sciences, University of Berne,
PLANT HORMONES-Introduction
 PLANT HORMONES-Introduction By convention hormone are said to be a substances whose site of synthesis and site of action are different; the two events are separated by space and time. Hormones are known
PLANT HORMONES-Introduction By convention hormone are said to be a substances whose site of synthesis and site of action are different; the two events are separated by space and time. Hormones are known
Nature Genetics: doi: /ng Supplementary Figure 1. The phenotypes of PI , BR121, and Harosoy under short-day conditions.
 Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Brassinosteroids Stimulate Plant Tropisms through Modulation of Polar Auxin Transport in Brassica and Arabidopsis
 The Plant Cell, Vol. 17, 2738 2753, October 2005, www.plantcell.org ª 2005 American Society of Plant Biologists Brassinosteroids Stimulate Plant Tropisms through Modulation of Polar Auxin Transport in
The Plant Cell, Vol. 17, 2738 2753, October 2005, www.plantcell.org ª 2005 American Society of Plant Biologists Brassinosteroids Stimulate Plant Tropisms through Modulation of Polar Auxin Transport in
JAGGED Controls Arabidopsis Petal Growth and Shape by Interacting with a Divergent Polarity Field
 by Interacting with a Divergent Polarity Field Susanna Sauret-Güeto 1, Katharina Schiessl 1, Andrew Bangham 2, Robert Sablowski 1, Enrico Coen 1 * 1 Department of Cell and Developmental Biology, John Innes
by Interacting with a Divergent Polarity Field Susanna Sauret-Güeto 1, Katharina Schiessl 1, Andrew Bangham 2, Robert Sablowski 1, Enrico Coen 1 * 1 Department of Cell and Developmental Biology, John Innes
E. THE FUNCTIONING OF HORMONES IN PLANT GROWTH AND DEVELOPMENT
 E. THE FUNCTIONING OF HORMONES IN PLANT GROWTH AND DEVELOPMENT E1. The Transport of Auxins David A. Morris 1, Jiří Friml 2, and Eva Zažímalová 3 1 School of Biological Sciences, University of Southampton,
E. THE FUNCTIONING OF HORMONES IN PLANT GROWTH AND DEVELOPMENT E1. The Transport of Auxins David A. Morris 1, Jiří Friml 2, and Eva Zažímalová 3 1 School of Biological Sciences, University of Southampton,
Plant Development. Chapter 31 Part 1
 Plant Development Chapter 31 Part 1 Impacts, Issues Foolish Seedlings, Gorgeous Grapes Gibberellin and other plant hormones control the growth and development of plants environmental cues influence hormone
Plant Development Chapter 31 Part 1 Impacts, Issues Foolish Seedlings, Gorgeous Grapes Gibberellin and other plant hormones control the growth and development of plants environmental cues influence hormone
Analysis of regulatory function of circadian clock. on photoreceptor gene expression
 Thesis of Ph.D. dissertation Analysis of regulatory function of circadian clock on photoreceptor gene expression Tóth Réka Supervisor: Dr. Ferenc Nagy Biological Research Center of the Hungarian Academy
Thesis of Ph.D. dissertation Analysis of regulatory function of circadian clock on photoreceptor gene expression Tóth Réka Supervisor: Dr. Ferenc Nagy Biological Research Center of the Hungarian Academy
Root gravitropism and root hair development constitute coupled developmental responses regulated by auxin homeostasis in the Arabidopsis root apex
 Research Root gravitropism and root hair development constitute coupled developmental responses regulated by auxin homeostasis in the Arabidopsis root apex Stamatis Rigas 1 *, Franck Anicet Ditengou 2
Research Root gravitropism and root hair development constitute coupled developmental responses regulated by auxin homeostasis in the Arabidopsis root apex Stamatis Rigas 1 *, Franck Anicet Ditengou 2
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
Cytokinins influence root gravitropism via differential regulation of auxin transporter expression and localization in Arabidopsis
 Research Cytokinins influence root gravitropism via differential regulation of auxin transporter expression and localization in Arabidopsis Marketa Pernisova 1,2, Tomas Prat 1, Peter Grones 3, Danka Harustiakova
Research Cytokinins influence root gravitropism via differential regulation of auxin transporter expression and localization in Arabidopsis Marketa Pernisova 1,2, Tomas Prat 1, Peter Grones 3, Danka Harustiakova
Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family
, HAESA, ERECTA-family") Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family GENES & DEVELOPMENT (2000) 14: 108 117 INTRODUCTION Flower Diagram INTRODUCTION Abscission In plant, the process by which a plant
Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family GENES & DEVELOPMENT (2000) 14: 108 117 INTRODUCTION Flower Diagram INTRODUCTION Abscission In plant, the process by which a plant
Bio 100 Guide 27.
 Bio 100 Guide 27 http://www.offthemarkcartoons.com/cartoons/1994-11-09.gif http://www.cneccc.edu.hk/subjects/bio/album/chapter20/images/plant_growth.jpg http://pgjennielove.files.wordpress.com/2008/06/apical_meristem.png
Bio 100 Guide 27 http://www.offthemarkcartoons.com/cartoons/1994-11-09.gif http://www.cneccc.edu.hk/subjects/bio/album/chapter20/images/plant_growth.jpg http://pgjennielove.files.wordpress.com/2008/06/apical_meristem.png
TOPIC 9.3 GROWTH IN PLANTS
 TOPIC 9.3 GROWTH IN PLANTS 9.3 A Growth INTRO http://cdn2.hubspot.net/hubfs/18130/social-suggested-images/plant_growing.jpeg IB BIO 9.3 3 In general, plants are able to grow indeterminately. This means
TOPIC 9.3 GROWTH IN PLANTS 9.3 A Growth INTRO http://cdn2.hubspot.net/hubfs/18130/social-suggested-images/plant_growing.jpeg IB BIO 9.3 3 In general, plants are able to grow indeterminately. This means
Title: Cyclic programmed cell death stimulates hormone signaling and root development in Arabidopsis
 Title: Cyclic programmed cell death stimulates hormone signaling and root development in Arabidopsis Authors: Wei Xuan 1,2,3, Leah R. Band 4#, Robert P. Kumpf 1,2#, Daniël Van Damme 1,2, Boris Parizot
Title: Cyclic programmed cell death stimulates hormone signaling and root development in Arabidopsis Authors: Wei Xuan 1,2,3, Leah R. Band 4#, Robert P. Kumpf 1,2#, Daniël Van Damme 1,2, Boris Parizot
Plant Stimuli pp Topic 3: Plant Behaviour Ch. 39. Plant Behavioural Responses. Plant Hormones. Plant Hormones pp
 Topic 3: Plant Behaviour Ch. 39 Plants exist in environments that are constantly changing. Like animals, plants must be able to detect and react to stimuli in the environment. Unlike animals, plants can
Topic 3: Plant Behaviour Ch. 39 Plants exist in environments that are constantly changing. Like animals, plants must be able to detect and react to stimuli in the environment. Unlike animals, plants can
Supplementary Figure 1. Phenotype of the HI strain.
 Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
From Genome to Phenotype: Modeling the interaction of physical and chemical signals in plant meristems. Meyerowitz Lab and many collaborators
 From Genome to Phenotype: Modeling the interaction of physical and chemical signals in plant meristems Meyerowitz Lab and many collaborators Needs to understand tissues, morphogenesis and development:
From Genome to Phenotype: Modeling the interaction of physical and chemical signals in plant meristems Meyerowitz Lab and many collaborators Needs to understand tissues, morphogenesis and development:
Roots and Soil Chapter 5
 Roots and Soil Chapter 5 Plant Organs Plant organs are groups of several types of tissues that together perform a particular function. Vegetative organs roots, stems, leaves make and use food, absorb water
Roots and Soil Chapter 5 Plant Organs Plant organs are groups of several types of tissues that together perform a particular function. Vegetative organs roots, stems, leaves make and use food, absorb water
ORGANISMS RESPOND TO CHANGES IN THEIR INTERNAL AND EXTERNAL ENVIRONMENTS
 MODULE 6 ORGANISMS RESPOND TO CHANGES IN THEIR INTERNAL AND EXTERNAL ENVIRONMENTS BIOLOGY NOTES I have designed and compiled these beautiful notes to provide a detailed but concise summary of this module.
MODULE 6 ORGANISMS RESPOND TO CHANGES IN THEIR INTERNAL AND EXTERNAL ENVIRONMENTS BIOLOGY NOTES I have designed and compiled these beautiful notes to provide a detailed but concise summary of this module.
Chapter 39. Plant Response. AP Biology
 Chapter 39. Plant Response 1 Plant Reactions Stimuli & a Stationary Life u animals respond to stimuli by changing behavior move toward positive stimuli move away from negative stimuli u plants respond
Chapter 39. Plant Response 1 Plant Reactions Stimuli & a Stationary Life u animals respond to stimuli by changing behavior move toward positive stimuli move away from negative stimuli u plants respond
Root gravitropism is regulated by a crosstalk between para-aminobenzoic acid, ethylene, and auxin
 Plant Physiology Preview. Published on October 1, 2018, as DOI:10.1104/pp.18.00126 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 Root gravitropism is regulated by a crosstalk
Plant Physiology Preview. Published on October 1, 2018, as DOI:10.1104/pp.18.00126 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 Root gravitropism is regulated by a crosstalk
Travis Lick. Plant movement
 Travis Lick Plant movement Plants, with their roots firmly fixed in the earth, seem immobile and vulnerable compared to animals, but this does not prevent them from reacting to stimuli in their environment.
Travis Lick Plant movement Plants, with their roots firmly fixed in the earth, seem immobile and vulnerable compared to animals, but this does not prevent them from reacting to stimuli in their environment.
Aida, M., Vernoux, X., Furutani, M., Traas, J. and Tasaka, M. (2002). Development 129, On page 3972 of this article, the sentence This
. Development 129, On page 3972 of this article, the sentence This") ERRATUM Roles of PIN-FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo Aida, M., Vernoux, X., Furutani, M., Traas, J. and Tasaka, M. (2002). Development 129, 3965-3974.
ERRATUM Roles of PIN-FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo Aida, M., Vernoux, X., Furutani, M., Traas, J. and Tasaka, M. (2002). Development 129, 3965-3974.
Name Class Date. In the space provided, write the letter of the description that best matches the term or phrase.
 Assessment Chapter Test B Plant Responses In the space provided, write the letter of the description that best matches the term or phrase. 1. thigmonasty 2. auxin 3. ethylene 4. phytochrome 5. abscisic
Assessment Chapter Test B Plant Responses In the space provided, write the letter of the description that best matches the term or phrase. 1. thigmonasty 2. auxin 3. ethylene 4. phytochrome 5. abscisic
Major Plant Hormones 1.Auxins 2.Cytokinins 3.Gibberelins 4.Ethylene 5.Abscisic acid
 Plant Hormones Lecture 9: Control Systems in Plants What is a Plant Hormone? Compound produced by one part of an organism that is translocated to other parts where it triggers a response in target cells
Plant Hormones Lecture 9: Control Systems in Plants What is a Plant Hormone? Compound produced by one part of an organism that is translocated to other parts where it triggers a response in target cells
Plant Growth and Development
 Plant Growth and Development Concept 26.1 Plants Develop in Response to the Environment Factors involved in regulating plant growth and development: 1. Environmental cues (e.g., day length) 2. Receptors
Plant Growth and Development Concept 26.1 Plants Develop in Response to the Environment Factors involved in regulating plant growth and development: 1. Environmental cues (e.g., day length) 2. Receptors
Arabidopsis ASA1 Is Important for Jasmonate-Mediated Regulation of Auxin Biosynthesis and Transport during Lateral Root Formation W OA
 The Plant Cell, Vol. 21: 1495 1511, May 2009, www.plantcell.org ã 2009 American Society of Plant Biologists Arabidopsis ASA1 Is Important for Jasmonate-Mediated Regulation of Auxin Biosynthesis and Transport
The Plant Cell, Vol. 21: 1495 1511, May 2009, www.plantcell.org ã 2009 American Society of Plant Biologists Arabidopsis ASA1 Is Important for Jasmonate-Mediated Regulation of Auxin Biosynthesis and Transport
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-15 PLANT GROWTH AND DEVELOPMENT
 CHAPTER-15 PLANT GROWTH AND DEVELOPMENT") CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-15 PLANT GROWTH AND DEVELOPMENT Root, stem leaves, flower, fruits and seeds arise in orderly manner in plants. The sequence of growth is as follows-
CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-15 PLANT GROWTH AND DEVELOPMENT Root, stem leaves, flower, fruits and seeds arise in orderly manner in plants. The sequence of growth is as follows-
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/9/452/ra106/dc1 Supplementary Materials for Stem-piped light activates phytochrome B to trigger light responses in Arabidopsis thaliana roots Hyo-Jun Lee, Jun-Ho
www.sciencesignaling.org/cgi/content/full/9/452/ra106/dc1 Supplementary Materials for Stem-piped light activates phytochrome B to trigger light responses in Arabidopsis thaliana roots Hyo-Jun Lee, Jun-Ho
Topic 15. The Shoot System
 Topic 15. The Shoot System Introduction. This is the second of two lab topics that focus on the three plant organs (root, stem, leaf). In these labs we want you to recognize how tissues are organized in
Topic 15. The Shoot System Introduction. This is the second of two lab topics that focus on the three plant organs (root, stem, leaf). In these labs we want you to recognize how tissues are organized in
Responses to Light. Responses to Light
 Sensory Systems in Plants Chapter 41 Pigments other than those used in photosynthesis can detect light and mediate the plant s response to it Photomorphogenesis refers to nondirectional, light-triggered
Sensory Systems in Plants Chapter 41 Pigments other than those used in photosynthesis can detect light and mediate the plant s response to it Photomorphogenesis refers to nondirectional, light-triggered
insights into root deveiopment from Arabidopsis root mutants
 Plant, Cell and Environment { \994)A7, 675-680 COMMISSIONED REVIEW insights into root deveiopment from Arabidopsis root mutants p. N. B E N F E Y ' & J. W. SCHIEFELBEIN^ 'Department of Biology, New York
Plant, Cell and Environment { \994)A7, 675-680 COMMISSIONED REVIEW insights into root deveiopment from Arabidopsis root mutants p. N. B E N F E Y ' & J. W. SCHIEFELBEIN^ 'Department of Biology, New York
Auxin is not asymmetrically distributed in initiating Arabidopsis leaves. *Author for correspondence: Marcus G Heisler
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 Auxin is not asymmetrically distributed in initiating Arabidopsis leaves Neha Bhatia 1 and Marcus G. Heisler 1* Affiliations
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 Auxin is not asymmetrically distributed in initiating Arabidopsis leaves Neha Bhatia 1 and Marcus G. Heisler 1* Affiliations
Figure 18.1 Blue-light stimulated phototropism Blue light Inhibits seedling hypocotyl elongation
 Blue Light and Photomorphogenesis Q: Figure 18.3 Blue light responses - phototropsim of growing Corn Coleoptile 1. How do we know plants respond to blue light? 2. What are the functions of multiple BL
Blue Light and Photomorphogenesis Q: Figure 18.3 Blue light responses - phototropsim of growing Corn Coleoptile 1. How do we know plants respond to blue light? 2. What are the functions of multiple BL
Chapter 39. Plant Reactions. Plant Hormones 2/25/2013. Plants Response. What mechanisms causes this response? Signal Transduction Pathway model
 Chapter 39 Plants Response Plant Reactions Stimuli & a Stationary life Animals respond to stimuli by changing behavior Move toward positive stimuli Move away from negative stimuli Plants respond to stimuli
Chapter 39 Plants Response Plant Reactions Stimuli & a Stationary life Animals respond to stimuli by changing behavior Move toward positive stimuli Move away from negative stimuli Plants respond to stimuli
Valley Central School District 944 State Route 17K Montgomery, NY Telephone Number: (845) ext Fax Number: (845)
 ext Fax Number: (845)") Valley Central School District 944 State Route 17K Montgomery, NY 12549 Telephone Number: (845)457-2400 ext. 18121 Fax Number: (845)457-4254 Advance Placement Biology Presented to the Board of Education
Valley Central School District 944 State Route 17K Montgomery, NY 12549 Telephone Number: (845)457-2400 ext. 18121 Fax Number: (845)457-4254 Advance Placement Biology Presented to the Board of Education
(17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON
 CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON") (17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON Jim Burton 1 and Marianne Pedersen Abstract. Cyclanilide [1-(2,4-dichlorophenylaminocarbonyl)-cyclopropane carboxylic
(17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON Jim Burton 1 and Marianne Pedersen Abstract. Cyclanilide [1-(2,4-dichlorophenylaminocarbonyl)-cyclopropane carboxylic