Supplementary text providing additional details of the computer model. I. Diffusive (i.e. not carrier-mediated) membrane permeabilities
|
|
|
- Gregory Hardy
- 5 years ago
- Views:
Transcription
1 Supplementary text providing additional details of the computer model I. Diffusive (i.e. not carrier-mediated) membrane permeabilities Protonated IAA. There is a wide range of published values for the permeability of plant lipid membranes to protonated indole-3-acetic acid (IAAH). Many of these must be discounted however, since they do not control for the likely presence of anion carriers 1,2. The only two studies we know of that control for carriers are both by Delbarre and coworkers 3,4. These report permeabilities of 0.14 cm/hr and 0.18 cm/hr respectively for suspension-cultured tobacco protoplasts. We use the model value P IAAH = 0.2 cm/hr. It should be noted that our model value is an order of magnitude lower than the oftcited result of Gutknecht and Walter 5. However, their permeability is found by fitting a straight line to data that comes almost entirely from experiments on artificial bilayers of egg lecithin. We favor measurements made on plant cell membranes, since Bean et al. 6 have shown that the permeability of bilayers to IAAH depends strongly on the composition of the bilayer (their results range between 0.3 cm/hr and 13 cm/hr). Anionic IAA. Following Refs. [ 5 ] and [ 6 ], we assume the IAA - anion has a negligible permeability. II. Carrier-mediated membrane permeabilities Measured permeability values. We begin our discussion of auxin efflux by noting that the PIN proteins are now commonly described as efflux "facilitators" rather than
2 efflux carriers 7. However, even if PIN's are not carriers, they are components of the efflux machinery, and their patterns of expression and localization are believed to be reliable indicators of auxin efflux 7,8. The permeabilities due to auxin influx carriers (the AUX/LAX gene family 9 ) and to auxin efflux facilitators (the PIN gene family 8,10 ) are not well-known. Perhaps the most useful source of quantitative data is Delbarre et al. 4, who report the diffusive and carrier mediated auxin influx and efflux in suspension-cultured tobacco protoplasts. Based on their data, we estimate an influx carrier permeability of 0.02 cm/hr and an efflux carrier permeability of 0.01 cm/hr. A second useful paper, and the only published value for influx carrier permeability in a plant tissue, is Szponarski et al. 11. They report an influx carrier permeability of cm/hr, measured in plasma membrane vesicles derived from mature Arabidopsis leaves (note the surprising agreement with the tobacco protoplast result). Szponarski's result reflects an average over all cells of the leaf, so the permeability in cells specialized for auxin transport is likely to be larger. Considering these facts, permeability values of order 0.1 cm/hr are expected to be reasonable in the root expansion zone. Model permeability values. Table S2 shows the model parameters used for carriermediated transport in the outer root. In the epidermis, we take the influx permeability to be P AUX1 = 0.2 cm/hr and the efflux permeability to be <P PIN2 > = 0.1 cm/hr. The angle brackets indicate an average over the area of the cell membrane. Note that an average is quoted since the efflux carrier permeabilties vary over the cell membrane due to the polar localization of the carriers. The model ratio of influx to efflux permeabilties (2.0) is chosen using an independent fit to the measured speed of basipetal auxin transport in root
3 tips, about 1 cm/hr 12,13. Blilou et al. 7 describe weak PIN1 expression in the epidermis, co-localized with PIN2. We thus take <P PIN1 > = 0.02 cm/hr. The remaining influx carrier entries in Table S2 are based on Blilou et al. 7. They report strong PIN2 and weak PIN1 expression in the cortex, localized to the apical (i.e. facing the root apex) end of the cells. They also report weak PIN1 expression in the endodermis, localized apically. It should however be noted that there are varying reports on the polarity of PIN2 in the cortex 14,15. We therefore ran model versions with different choices for cortical PIN2 polarity. Surprisingly, model results for the auxin distribution depend only weakly on the localization of PIN2 in the cortex (Tables S3 and S4). The reason is that cortical cells do not express an influx carrier (see Discussion in main text), and so do not accumulate much auxin as compared with the epidermis. The cortical contribution to the net auxin flux is thus relatively weak. A cell with no carrier localized to the plasma membrane is assigned a carrier permeability of zero. For example, the pin2 mutantis modeled using the values in Table S2, with the exception that all PIN2 permeabilities are set equal to zero. Fluorescence immunolabelling shows no AUX1 signal in either the cortex or the endodermis of the Arabidopsis outer root 7,16. We therefore assign zero influx carrier permeability to cells of both the cortex and endodermis. III. Diffusion coefficients Intracellular diffusion coefficient. We do not know of any direct measurements of the cytoplasmic or vacuolar diffusion of IAA (indole-3-acetic acid). Since the vacuole is
4 mostly water, we expect the vacuolar diffusion coefficient of IAA to be comparable to the known aqueous value, D aq = cm 2 /hr 17. In the cytoplasm, comparable small molecules have diffusion coefficients in the range of 10% to 40% of their aqueous value 18. The model used here does not treat the vacuole as a distinct cellular compartment. The proper choice for the diffusion coefficient of IAA inside model cells thus depends on whether IAA can freely enter the central vacuole, and on the fraction of the cell volume occupied by the vacuole. We assume IAA can enter the vacuole (although the carrier family is unknown) 19-21, and so take D = D aq inside model cells. For completeness, however, we have verified that an auxin-impermeable vacuole does not significantly change the model results (data not shown). Apoplastic diffusion coefficient. The permeability of the apoplast to small molecules is not homogeneous, having significant restrictions at Casparian bands, and also possibly within the root meristem However, these restrictions are not present in the zone of the Arabidopsis root tip studied here. Following Refs. [ 25 ] and [ 26 ], we take the apoplastic diffusion coefficient of IAA to be 0.1 D aq. IV. Membrane voltage and ph The ph and membrane voltage of plant cells have been thoroughly studied, for examples see Refs. [ ]. We use the following representative values: cytoplasm ph c = 7.2, apoplast ph w = 5.3, and cell membrane voltage V = -120 mv (negative inside).
5 V. Mathematical and numerical methods Our model extends the techniques of Goldsmith et al. 30 and Kramer 31 to the case of a three dimensional array of rectangular cells (see Fig. 2.A). The main difference with the model cells of Ref. [ 31 ] is that we do not include a separate vacuole. To simplify the model, we generally assume the tonoplast is sufficiently permeable to auxin that the cell interior can be treated as a single compartment As a check, we do conduct some runs with auxin-impermeable vacuoles and verify that they do not cause significant changes (data not shown). The tissue is divided into a rectangular array of boxes (called "control volumes" in Ref. [ 32 ]). The interior of each cell is a set of (usually 5) contiguous boxes, with the remaining boxes making up the apoplastic space. A nonuniform box size allows us to resolve the 0.5 m thick walls, despite cell lengths up to 400 m. The auxin flux between adjacent boxes within each cell, and between boxes in the apoplastic space, is governed by the discrete approximation to Fick's law for diffusion. J 1 2 = D c 2 c 1 L (1) where J 1->2 is the net flux from box 1 to box 2, c j is the concentration of auxin in box j (including both protonated and anion forms), D is the diffusion coefficient of auxin in the compartment, and L is the center-to-center distance between the boxes. The net flux across the cell membrane is governed by the uniform-field approximation 33. The net flux across any one segment of the cell membrane (i.e. between one cell box and one apoplast box) is J 1 2 = J diff + J PIN + J AUX1, where J diff, J PIN, and J AUX1
6 are due to the diffusive membrane permeability of the protonated auxin molecule, PIN family carriers, and AUX1 carriers respectively. c J diff = P 2 IAAH 1+10 c 1 ph 2 pk 1+10 ph 1 pk (2) f ( φ)c J PIN = P 2 PIN 1+10 f (φ)c 1 (3) ph 2 + pk 1+10 ph 1 + pk f (φ)c J AUX1 = P 2 AUX f ( φ)c 1 ph 2 + pk 1+10 ph 1 + pk (4) f (x) = x e x 1 (5) where ph and pk have their usual meanings, the P j are membrane permeabilities, and φ = ± FV/RT where V is the membrane potential, F is the Faraday constant, R is the gas constant, T is the temperature, and the + sign is used if box 1 is interior to box 2 (by definition, the cytoplasm is interior to the cell wall). It is instructive to substitute the parameter values into Eqns. (2) through (5) and sum to find an expression for the net auxin influx across a membrane segment J w c = ( 0.24 P IAAH P AUX P PIN )c w (6) ( P IAAH P AUX P PIN )c c where the subscript w denotes the wall, the subscript c denotes the cytoplasm, and a negative flux would be out of the cell. Note that the influx carrier permeability enters the top line of Eqn. (6) with a prefactor nearly 15 times larger than the diffusive permeability. Thus, although P IAAH and P AUX1 are of the same order of magnitude (see Secs. I and II above), the carrier dominates diffusive influx in all model cells where it is expressed. There is a common misconception that, because protonated auxin is
7 membrane permeable, influx carriers play only a supplemental role in auxin distribution. A quantitative treatment shows that this is not the case. The flux of auxin between the simulation boxes is described by a coupled set of thousands of differential equations. These differential equations are discretized using an explicit scheme as described in Chapter 4 of Ref. [ 32 ]. The equations are solved using the program AuxSim3D, written by the author and run on a PowerBook G4. VI. Assessing the speed of transport. The speed of auxin transport in the elongation zone is assessed in the following way. Figure S7a shows the auxin concentration in the central elongation zone, 500 m proximal to the source of auxin (see model figure, main text), as a function of time. The initial condition is a root empty of auxin. Starting at time zero, a constant, asymmetric auxin source is applied to the distal apoplast of the model root epidermis. After about 5 min, the auxin concentrations in the CEZ have reached a steady state. There are several ways one might assign a speed to this auxin pulse. Our preferred method is to divide the distance traveled (0.05 cm in this case) by the time it takes the auxin concentration to reach half its steady-state value. For data in Fig. S7a, these times are hr along the bottom flank of the root and hr on the top flank. These give speeds of 1.60 cm/hr and 1.46 cm/hr respectively.
8 References 1. Gimmler, H., Heilmann, B., Demming, B. & Hartung, W. The permeability coefficients of the plasmalemma and the chloroplast envelope of spinach mesophyll cells for phytohormones. Z. Naturforsch. 36 c, (1981). 2. Baier, M., Gimmler, H. & Hartung, W. The permeability of the guard cell plasma membrane and tonoplast. Journal of Experimental Botany 41, (1990). 3. Delbarre, A., Muller, P., Imhoff, V., Morgat, J.-L. & Barbier-Brygoo, H. Uptake, accumulation and metabolism of auxins in tobacco leaf protoplasts. Planta 195, (1994). 4. Delbarre, A., Muller, P. & Guern, J. Comparison of mechanisms controlling uptake and accumulation of 2,4-dichlorophenoxy acetic acid, naphthalene-1- acetic acid, and indole-3-acetic acid in suspension-cultured tobacco cells. Planta 198, (1996). 5. Gutknecht, J. & Walter, A. Transport of auxin (indoleacetic acid) through lipid bilayer membranes. Journal of Membrane Biology 56, (1980). 6. Bean, R., Shepherd, W. & Chan, H. Permeability of lipid bilayer membranes to organic solutes. Journal of General Physiology 52, (1968). 7. Blilou, I. et al. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 433, (2005). 8. Benkova, E. et al. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115, (2003). 9. Parry, G. et al. Quick on the uptake: characterization of a family of plant auxin influx carriers. Journal of Plant Growth Regulation 20, (2001). 10. Galweiler, L. et al. Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 282, (1998). 11. Szponarski, W. et al. Reconstitution of an electrogenic auxin transport activity mediated by Arabidopsis thaliana plasma membrane proteins. FEBS Letters 446, (1999). 12. Tsurumi, S. & Ohwaki, Y. Transport of 14 C-labeled indoleacetic acid in Vicia root segments. Plant and Cell Physiology 19, (1978). 13. Monshausen, G. & Sievers, A. Basipetal propagation of gravity-induced surface ph changes along primary roots of Lepidium sativum L. Planta 215, (2002). 14. Muller, A. et al. AtPIN2 defines a locus of Arabidopsis for root gravitropism control. The EMBO Journal 17, (1998). 15. Peer, W. A. et al. Variation in expression and protein localization of the PIN family of auxin efflux facilitator proteins in flavonoid mutants with altered auxin transport in Arabidopsis thaliana. Plant Cell 16, (2004). 16. Swarup, R. et al. Localization of the auxin permease AUX1 suggests two functionally distinct hormone pathways operate in the Arabidopsis root apex. Genes & Development 15, (2001).
9 17. Mitchison, G. J. The dynamics of auxin transport. Proceedings of the Royal Society of London B. Biological Sciences 209, (1980). 18. Paine, P., Moore, L. & Horowitz, S. Nuclear envelope permeability. Nature 254, (1975). 19. Ouitrakul, R. & Hertel, R. Effect of gravity and centrifugal acceleration on auxin transport in corn coleoptiles. Planta 88, (1969). 20. Cande, W., Goldsmith, M. H. M. & Ray, P. M. Polar auxin transport and auxininduced elongation in the absence of cytoplasmic streaming. Planta 111, (1973). 21. Goldsmith, M. H. M. & Ray, P. M. Intracellular localization of the active process in polar transport of auxin. Planta 111, (1973). 22. Wierzbicka, M. Lead accumulation and its translocation barriers in roots of Allium cepa L. - autoradiographic and ultrastructural studies. Plant, Cell and Environment 10, (1987). 23. Enstone, D. & Peterson, C. The apoplastic permeability of root apices. Canadian Journal of Botany 70, (1992). 24. Cholewa, E. & Peterson, C. Detecting exodermal Casparian bands in vivo and fluid-phase endocytosis in onion (Allium cepa L.) roots. Canadian Journal of Botany 79, (2001). 25. Richter, E. & Ehwald, R. Apoplastic mobility of sucrose in storage parenchyma of sugar beet. Physiologia Plantarum 58, (1983). 26. Aloni, R., Enstone, D. & Peterson, C. Indirect evidence for bulk water flow in root cortical cell walls of three dicotyledonous species. Planta 207, 1-7 (1998). 27. Scott, A. C. & Allen, N. S. Changes in the cytosolic ph within Arabidopsis root columella cells play a key role in the early signalling pathway for root gravitropism. Plant Physiology 121, (1999). 28. Sze, H., Li, X. & Palmgren, M. Energization of plant cell membranes by H+ pumping ATPases: regulation and biosynthesis. The Plant Cell 11, (1999). 29. Fasano, J. et al. Changes in root cap ph are required for the gravity response of the Arabidopsis root. Plant Cell 13, (2001). 30. Goldsmith, M. H. M., Goldsmith, T. H. & Martin, M. H. Mathematical analysis of the chemosmotic polar diffusion of auxin through plant tissues. Proc. Nat. Acad. Sci. 78, (1981). 31. Kramer, E. M. PIN and AUX/LAX proteins: their role in auxin accumulation. Trends in Plant Science 9, (2004). 32. Patankar, S. V. Numerical heat transfer and fluid flow (Taylor & Francis, Levittown, PA, 1980). 33. Dainty, J. Ion transport and electrical potentials in plant cells. Annual Review of Plant Physiology 13, (1962).
10 Table S1: t-test: Two-Sample Assuming Equal Variances Variable 1 Variable 2 t Critical twotail P(T<=t) two-tail J0951>>axr3 J J0951>>axr3 Col x C J0951>>axr3 UAS axr J0951>>axr3 Col Col x C24 Col Col x C24 UAS axr C24 Col x C E-09 C24 Col E-10 C24 UAS axr E-09 C24 J Two sample student s t-test assuming equal variance was performed on 5-7 day old roots in three separate experiments. The null hypothesis here is that the means are equal ( P> 0.05), and the alternative hypothesis is that they are not (P< 0.05). The results presented in this table are representative of the three separate experiments. Table S2 Carrier-mediated permeability values. Tissue layer <P PIN1 > <P PIN2 > (cm/hr) PIN polarity P AUX1 (cm/hr) (cm/hr) epidermis To base 0.2 cortex To apex 0 endodermis To apex 0 Carrier-mediated permeabilities and PIN polarity used in the model of wildtype elongation zone tissues. Angle brackets indicate a mean over the area of the cell membrane. "To apex" indicates that PIN permeability is highest on the portion of the cell membrane facing the root apex.
11 Table S3 Variation due to changes in the cortical localization of PIN2. PIN2 polarity in Lateral auxin Fraction of all Fraction of cortex gradient in CEZ auxin in the auxin pulse that (c bottom /c top ) apoplast reaches the CEZ To apex To base To epidermis To endodermis Changes to the wildtype entries of Table 1 resulting from various choices for the polarity of PIN2 in the model cortex. Top row is the version presented in the main text. Table S4 Cytoplasmic auxin concentrations. Cortical Mean auxin concentration in PIN2 polarity epidermis cortex endodermis To apex To base To epidermis To endodermis Model results for the partitioning of auxin between the three cell layers of the outer root, given different choices for the polarity of PIN2 in the cortex. Auxin concentrations have arbitrary units.


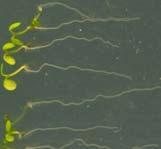


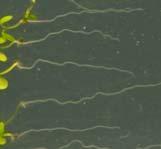

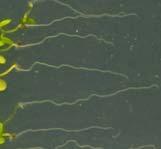
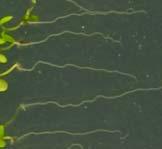



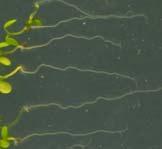

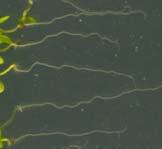



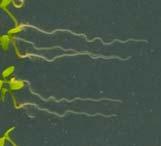







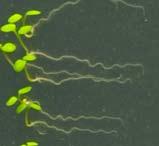








12 Swarup et al Fig S1 Col J0951>Aux Fig. S1 Seedlings were grown vertically for four days on MS plates and the plates were then turned at an angle of 90. Photographs were taken every 30 minute up to 8 hours and then at 24 hours after the change in the direction of the gravity vector. The figure illustrates results from wildtype (Col.) and J0951>AUX1 lines and are representative of data collected for all other lines which is summarised in Fig. 1i.
13 Swarup et al Fig S2 a Auxin concentration (arb. units) t (min) b Auxin concentration (arb. units) t (min) Fig. S2 Time-dependence of the epidermal auxin concentration. Cytoplasmic auxin concentration in two epidermal cells located 500 microns proximal to the lateral root cap. The concentrations on lower and upper sides of the roots are depicted in blue and red, respectively. (a) To assess the speed of the auxin pulse, the initial condition was a root with no auxin. (b) To visualize the gravitropic signal, the initial condition was a vertical root (concentrations were equilibrated to a uniform distal auxin source before time zero). An auxin gradient is supplied to the distal epidermal apoplast beginning at time zero.

 vs agravitropic")

14 Swarup et al Fig S3 a b c d e f Fig. S3 Root gravitropism requires AUX1 to be expressed in more than 5 elongating epidermal cells. Comparison of HA-AUX1 localisation in gravitropic (top) vs agravitropic (bottom) aux >>Aux1 seedlings. Immunolocalisation was performed with anti HA primary antibody and Alexa Fluor 555 coupled secondary antibody (b & e). Background staining was performed with Sytox Green (a & c). Scale bar 40 m.
15 Swarup et al Fig S4 120 % root growth compared to control Col a1-22 axr3 C24 Col x C24 UAS axr3 M0013 M0013>>axr3 J0951 J0951>>axr3 Fig. S4 Disrupting auxin perception in epidermal cells results in increased auxin resistance Seedlings were grown vertically on MS plates for five days and were then transferred on fresh MS, MS + 2,4-D (10-7 M) or MS + IAA plates (2.5 x 10-7 M) for further two days. The delta root growth in presence or absence of auxin was measured and expressed as percent root growth compared to MS control. Error bars represent standard deviation.


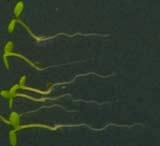



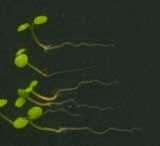


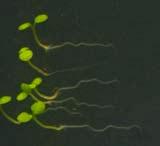
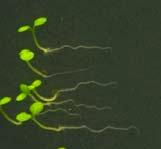








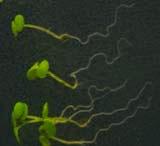







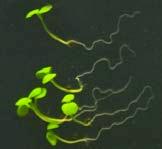






16 Swarup et al Fig S5 J J0951>>axr Fig. S5 Seedlings were grown vertically for four days on MS plates and the plates were then turned at an angle of 90. Photographs were taken every 30 minute up to 8 hours and then at 24 hours after the change in the direction of the gravity vector. The figure illustrates results from wildtype (Col.) and J0951>axr3-1 lines and are representative of data collected for all other lines which is summarised in Fig. 6m.
Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal
 ARTICLES Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal Ranjan Swarup 1, Eric M. Kramer 2, Paula Perry 1, Kirsten Knox 3, H. M. Ottoline
ARTICLES Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal Ranjan Swarup 1, Eric M. Kramer 2, Paula Perry 1, Kirsten Knox 3, H. M. Ottoline
DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Other cellular processes (17%)
 Signal transduction (2%) Development (2%) Other cellular processes (17%)") Fig. 35-24 Other metabolism (18%) DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Unknown (24%) Energy pathways (3%) Cell division and organization (3%) Transport (4%) Transcription
Fig. 35-24 Other metabolism (18%) DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Unknown (24%) Energy pathways (3%) Cell division and organization (3%) Transport (4%) Transcription
Chapter 36: Transport in Vascular Plants - Pathways for Survival
 Chapter 36: Transport in Vascular Plants - Pathways for Survival For vascular plants, the evolutionary journey onto land involved differentiation into roots and shoots Vascular tissue transports nutrients
Chapter 36: Transport in Vascular Plants - Pathways for Survival For vascular plants, the evolutionary journey onto land involved differentiation into roots and shoots Vascular tissue transports nutrients
Actions of auxin. Hormones: communicating with chemicals History: Discovery of a growth substance (hormone- auxin)
") Hormones: communicating with chemicals History- discovery of plant hormone. Auxin Concepts of hormones Auxin levels are regulated by synthesis/degradation, transport, compartmentation, conjugation. Polar
Hormones: communicating with chemicals History- discovery of plant hormone. Auxin Concepts of hormones Auxin levels are regulated by synthesis/degradation, transport, compartmentation, conjugation. Polar
NOTES: CH 36 - Transport in Plants
 NOTES: CH 36 - Transport in Plants Recall that transport across the cell membrane of plant cells occurs by: -diffusion -facilitated diffusion -osmosis (diffusion of water) -active transport (done by transport
NOTES: CH 36 - Transport in Plants Recall that transport across the cell membrane of plant cells occurs by: -diffusion -facilitated diffusion -osmosis (diffusion of water) -active transport (done by transport
Please sit next to a partner. you are an A or a B
 Please sit next to a partner you are an A or a B Plants Transport in Vascular Plants Transport Overview Vascular tissue transports nutrients throughout a plant Such transport may occur over long distances
Please sit next to a partner you are an A or a B Plants Transport in Vascular Plants Transport Overview Vascular tissue transports nutrients throughout a plant Such transport may occur over long distances
Movement of water and solutes in plants Chapter 4 and 30
 Movement of water and solutes in plants Chapter 4 and 30 Molecular Movement Diffusion Molecules or ions moving in the opposite direction = movement against a diffusion gradient. Rates of diffusion are
Movement of water and solutes in plants Chapter 4 and 30 Molecular Movement Diffusion Molecules or ions moving in the opposite direction = movement against a diffusion gradient. Rates of diffusion are
Chapter 36. Transport in Vascular Plants
 Chapter 36 Transport in Vascular Plants Overview: Pathways for Survival For vascular plants The evolutionary journey onto land involved the differentiation of the plant body into roots and shoots Vascular
Chapter 36 Transport in Vascular Plants Overview: Pathways for Survival For vascular plants The evolutionary journey onto land involved the differentiation of the plant body into roots and shoots Vascular
expression of AUX1, LAX1, LAX2 and LAX3 in etiolated hypocotyls.
 Table of Contents Supplementary Material and Methods Supplementary Figure S1: Phototropic response of various aux1lax mutants and expression of AUX1, LAX1, LAX2 and LAX3 in etiolated hypocotyls. Supplementary
Table of Contents Supplementary Material and Methods Supplementary Figure S1: Phototropic response of various aux1lax mutants and expression of AUX1, LAX1, LAX2 and LAX3 in etiolated hypocotyls. Supplementary
Transport in Plants Notes AP Biology Mrs. Laux 3 levels of transport occur in plants: 1. Uptake of water and solutes by individual cells -for
 3 levels of transport occur in plants: 1. Uptake of water and solutes by individual cells -for photosynthesis and respiration -ex: absorption of H 2 O /minerals by root hairs 2. Short distance cell-to-cell
3 levels of transport occur in plants: 1. Uptake of water and solutes by individual cells -for photosynthesis and respiration -ex: absorption of H 2 O /minerals by root hairs 2. Short distance cell-to-cell
Resource Acquisition and Transport in Vascular Plants
 Chapter 36 Resource Acquisition and Transport in Vascular Plants PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Chapter 36 Resource Acquisition and Transport in Vascular Plants PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Compartments and Transport. Three Major Pathways of Transport. Absorp+on of Water and Minerals by Root Cells. Bulk flow
 Plasmodesmata Channels connec+ng neighboring cells Cell membrane and cytosol are con+nuous from cell to cell Symplast Cytoplasmic con+nuum Apoplast Compartments and Transport Through plasmodesmata con+nuum
Plasmodesmata Channels connec+ng neighboring cells Cell membrane and cytosol are con+nuous from cell to cell Symplast Cytoplasmic con+nuum Apoplast Compartments and Transport Through plasmodesmata con+nuum
23-. Shoot and root development depend on ratio of IAA/CK
 Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Chapter 36~ Transport in Plants
 Chapter 36~ Transport in Plants Structural Features Used for Resource Acquistion Roots and stems to do transport of resources Diffusion, active transport, and bulk flow Work in vascular plants to transport
Chapter 36~ Transport in Plants Structural Features Used for Resource Acquistion Roots and stems to do transport of resources Diffusion, active transport, and bulk flow Work in vascular plants to transport
Plant Structure and Function
 Plant Structure and Function A Meridian Biology AP Study Guide by John Ho and Tim Qi Plant Terms Growth: Growth Types Type Location Description Primary Primary Vertical growth (up-down), dominant direction
Plant Structure and Function A Meridian Biology AP Study Guide by John Ho and Tim Qi Plant Terms Growth: Growth Types Type Location Description Primary Primary Vertical growth (up-down), dominant direction
Organs and leaf structure
 Organs and leaf structure Different types of tissues are arranged together to form organs. Structure: 2 parts (Petiole and Leaf Blade) Thin flat blade, large surface area Leaves contain all 3 types of
Organs and leaf structure Different types of tissues are arranged together to form organs. Structure: 2 parts (Petiole and Leaf Blade) Thin flat blade, large surface area Leaves contain all 3 types of
2014 Pearson Education, Inc. 1
 1 CO 2 O 2 Light Sugar O 2 and minerals CO 2 2 Buds 42 29 21 34 13 26 5 18 10 31 23 8 15 28 16 2 24 Shoot apical meristem 7 3 20 1 mm 32 11 19 12 6 4 1 25 17 14 9 40 27 22 3 Cell wall Apoplastic route
1 CO 2 O 2 Light Sugar O 2 and minerals CO 2 2 Buds 42 29 21 34 13 26 5 18 10 31 23 8 15 28 16 2 24 Shoot apical meristem 7 3 20 1 mm 32 11 19 12 6 4 1 25 17 14 9 40 27 22 3 Cell wall Apoplastic route
CHAPTER TRANSPORT
 CHAPTER 2 2.4 TRANSPORT Uptake of CO2 FOCUS: Uptake and transport of water and mineral salts Transport of organic substances Physical forces drive the transport of materials in plants over a range of distances
CHAPTER 2 2.4 TRANSPORT Uptake of CO2 FOCUS: Uptake and transport of water and mineral salts Transport of organic substances Physical forces drive the transport of materials in plants over a range of distances
Resource Acquisition and Transport in Vascular Plants
 Chapter 36 Resource Acquisition and Transport in Vascular Plants PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Chapter 36 Resource Acquisition and Transport in Vascular Plants PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Cytokinin. Fig Cytokinin needed for growth of shoot apical meristem. F Cytokinin stimulates chloroplast development in the dark
 Cytokinin Abundant in young, dividing cells Shoot apical meristem Root apical meristem Synthesized in root tip, developing embryos, young leaves, fruits Transported passively via xylem into shoots from
Cytokinin Abundant in young, dividing cells Shoot apical meristem Root apical meristem Synthesized in root tip, developing embryos, young leaves, fruits Transported passively via xylem into shoots from
Ch. 36 Transport in Vascular Plants
 Ch. 36 Transport in Vascular Plants Feb 4 1:32 PM 1 Essential Question: How does a tall tree get the water from its roots to the top of the tree? Feb 4 1:38 PM 2 Shoot architecture and Light Capture: Phyllotaxy
Ch. 36 Transport in Vascular Plants Feb 4 1:32 PM 1 Essential Question: How does a tall tree get the water from its roots to the top of the tree? Feb 4 1:38 PM 2 Shoot architecture and Light Capture: Phyllotaxy
Mathematical Biology. Multiscale modelling of auxin transport in the plant-root elongation zone. L. R. Band J. R. King
 J. Math. Biol. (2012) 65:743 785 DOI 10.1007/s00285-011-0472-y Mathematical Biology Multiscale modelling of auxin transport in the plant-root elongation zone L. R. Band J. R. King Received: 17 September
J. Math. Biol. (2012) 65:743 785 DOI 10.1007/s00285-011-0472-y Mathematical Biology Multiscale modelling of auxin transport in the plant-root elongation zone L. R. Band J. R. King Received: 17 September
Transport in Vascular Plants
 Chapter 36 Transport in Vascular Plants PowerPoint Lectures for Biology, Seventh Edition Neil Campbell and Jane Reece Lectures by Chris Romero Vascular tissue Transports nutrients throughout a plant; such
Chapter 36 Transport in Vascular Plants PowerPoint Lectures for Biology, Seventh Edition Neil Campbell and Jane Reece Lectures by Chris Romero Vascular tissue Transports nutrients throughout a plant; such
Transport of substances in plants
 Transport of substances in plants We have already looked at why many organisms need transport systems with special reference to surface area and volume. The larger the volume : surface area ratio, the
Transport of substances in plants We have already looked at why many organisms need transport systems with special reference to surface area and volume. The larger the volume : surface area ratio, the
Translocation 11/30/2010. Translocation is the transport of products of photosynthesis, mainly sugars, from mature leaves to areas of growth and
 Translocation Translocation is the transport of products of photosynthesis, mainly sugars, from mature leaves to areas of growth and storage. Phloem is the tissue through which translocation occurs. Sieve
Translocation Translocation is the transport of products of photosynthesis, mainly sugars, from mature leaves to areas of growth and storage. Phloem is the tissue through which translocation occurs. Sieve
CHAPTER 32 TRANSPORT IN PLANTS OUTLINE OBJECTIVES
 CHAPTER 32 TRANSPORT IN PLANTS OUTLINE I. The traffic of water and solutes occurs on cellular, organ, and whole-plant levels: an overview of transport in plants A. Transport at the Cellular Level B. Short
CHAPTER 32 TRANSPORT IN PLANTS OUTLINE I. The traffic of water and solutes occurs on cellular, organ, and whole-plant levels: an overview of transport in plants A. Transport at the Cellular Level B. Short
IB Bio: Plant Biology. Topic 9
 IB Bio: Plant Biology Topic 9 9.1: Transport in xylem How and why does water move up a plant? How do plants conserve water? 9.2: Transport in phloem How and why and where does food move in a plant? 9.3:
IB Bio: Plant Biology Topic 9 9.1: Transport in xylem How and why does water move up a plant? How do plants conserve water? 9.2: Transport in phloem How and why and where does food move in a plant? 9.3:
CONTROL OF PLANT GROWTH AND DEVELOPMENT BI-2232 RIZKITA R E
 CONTROL OF PLANT GROWTH AND DEVELOPMENT BI-2232 RIZKITA R E The development of a plant the series of progressive changes that take place throughout its life is regulated in complex ways. Factors take part
CONTROL OF PLANT GROWTH AND DEVELOPMENT BI-2232 RIZKITA R E The development of a plant the series of progressive changes that take place throughout its life is regulated in complex ways. Factors take part
CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-11 TRANSPORT IN PLANTS
 CHAPTER-11 TRANSPORT IN PLANTS") CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-11 TRANSPORT IN PLANTS Plant transport various substance like gases, minerals, water, hormones, photosynthetes and organic solutes to short distance
CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-11 TRANSPORT IN PLANTS Plant transport various substance like gases, minerals, water, hormones, photosynthetes and organic solutes to short distance
Bio Factsheet. Transport in Plants. Number 342
 Number 342 Transport in Plants This Factsheet: Explains why plants need a transport system Describes what plants transport Describes the tissues which carry out transport Outlines the position of the xylem
Number 342 Transport in Plants This Factsheet: Explains why plants need a transport system Describes what plants transport Describes the tissues which carry out transport Outlines the position of the xylem
OCR (A) Biology A-level
 Biology A-level") OCR (A) Biology A-level Topic 3.3: Transport in plants Notes Plants require a transport system to ensure that all the cells of a plant receive a sufficient amount of nutrients. This is achieved through
OCR (A) Biology A-level Topic 3.3: Transport in plants Notes Plants require a transport system to ensure that all the cells of a plant receive a sufficient amount of nutrients. This is achieved through
 Question 1: What are the factors affecting the rate of diffusion? Diffusion is the passive movement of substances from a region of higher concentration to a region of lower concentration. Diffusion of
Question 1: What are the factors affecting the rate of diffusion? Diffusion is the passive movement of substances from a region of higher concentration to a region of lower concentration. Diffusion of
Plants are sessile. 10d-17/giraffe-grazing.jpg
 Plants are sessile www.mccullagh.org/db9/ 10d-17/giraffe-grazing.jpg Plants have distinct requirements because of their sessile nature Organism-level requirements Must adjust to environment at given location
Plants are sessile www.mccullagh.org/db9/ 10d-17/giraffe-grazing.jpg Plants have distinct requirements because of their sessile nature Organism-level requirements Must adjust to environment at given location
C MPETENC EN I C ES LECT EC UR U E R
 LECTURE 7: SUGAR TRANSPORT COMPETENCIES Students, after mastering the materials of Plant Physiology course, should be able to: 1. To explain the pathway of sugar transport in plants 2. To explain the mechanism
LECTURE 7: SUGAR TRANSPORT COMPETENCIES Students, after mastering the materials of Plant Physiology course, should be able to: 1. To explain the pathway of sugar transport in plants 2. To explain the mechanism
Transport in Plants. Transport in plants. Transport across Membranes. Water potential 10/9/2016
 Transport in Plants Transport in plants How is a plant able to move water and nutrients from roots to the rest of the plant body? Especially tall trees? Sequoia can be over 300 feet tall! Transport across
Transport in Plants Transport in plants How is a plant able to move water and nutrients from roots to the rest of the plant body? Especially tall trees? Sequoia can be over 300 feet tall! Transport across
2014 Pearson Education, Inc. 1. Light. Sugar O 2 H 2 O. and minerals CO Pearson Education, Inc.
 1 CO 2 O 2 Light ugar O 2 and minerals CO 2 2 Buds 34 42 29 26 31 18 21 13 5 10 23 8 15 28 16 24 hoot apical meristem 2 7 3 20 32 11 19 12 6 4 1 25 17 14 9 40 27 22 1 mm 3 Cell wall Apoplastic route Cytosol
1 CO 2 O 2 Light ugar O 2 and minerals CO 2 2 Buds 34 42 29 26 31 18 21 13 5 10 23 8 15 28 16 24 hoot apical meristem 2 7 3 20 32 11 19 12 6 4 1 25 17 14 9 40 27 22 1 mm 3 Cell wall Apoplastic route Cytosol
ENDODERMIS & POLARITY
 https://en.wikipedia.org/wiki/casparian_strip ENDODERMIS & POLARITY Niloufar Pirayesh 13.01.2016 PCDU SEMINAR 2 What is Endodermis? It helps with Regulate the movement of water ions and hormones. (in and
https://en.wikipedia.org/wiki/casparian_strip ENDODERMIS & POLARITY Niloufar Pirayesh 13.01.2016 PCDU SEMINAR 2 What is Endodermis? It helps with Regulate the movement of water ions and hormones. (in and
Roots and Soil Chapter 5
 Roots and Soil Chapter 5 Plant Organs Plant organs are groups of several types of tissues that together perform a particular function. Vegetative organs roots, stems, leaves make and use food, absorb water
Roots and Soil Chapter 5 Plant Organs Plant organs are groups of several types of tissues that together perform a particular function. Vegetative organs roots, stems, leaves make and use food, absorb water
BIO1PS 2012 Plant Science Lecture 4 Hormones Pt. I
 BIO1PS 2012 Plant Science Lecture 4 Hormones Pt. I Dr. Michael Emmerling Department of Botany Room 410 m.emmerling@latrobe.edu.au Hormones and Ghost gum Eucalyptus papuana Coordination ~3 Lectures Leaves
BIO1PS 2012 Plant Science Lecture 4 Hormones Pt. I Dr. Michael Emmerling Department of Botany Room 410 m.emmerling@latrobe.edu.au Hormones and Ghost gum Eucalyptus papuana Coordination ~3 Lectures Leaves
Transport in Plants (Ch. 23.5)
") Transport in Plants (Ch. 23.5) Transport in plants H 2 O & minerals transport in xylem Transpiration Adhesion, cohesion & Evaporation Sugars transport in phloem bulk flow Gas exchange photosynthesis CO
Transport in Plants (Ch. 23.5) Transport in plants H 2 O & minerals transport in xylem Transpiration Adhesion, cohesion & Evaporation Sugars transport in phloem bulk flow Gas exchange photosynthesis CO
Biology Slide 1 of 36
 Biology 1 of 36 2 of 36 Types of Roots Types of Roots What are the two main types of roots? 3 of 36 Types of Roots The two main types of roots are: taproots, which are found mainly in dicots, and fibrous
Biology 1 of 36 2 of 36 Types of Roots Types of Roots What are the two main types of roots? 3 of 36 Types of Roots The two main types of roots are: taproots, which are found mainly in dicots, and fibrous
Essential idea: Plants adapt their growth to environmental conditions.
 9.3 Growth in plants AHL https://c1.staticflickr.com/3/2347/2573372542_a959ecfd4f_b.jpg Essential idea: Plants adapt their growth to environmental conditions. Boxwood, Pivet and Yew are plants commonly
9.3 Growth in plants AHL https://c1.staticflickr.com/3/2347/2573372542_a959ecfd4f_b.jpg Essential idea: Plants adapt their growth to environmental conditions. Boxwood, Pivet and Yew are plants commonly
Introduction to Plant Transport
 Introduction to Plant Transport The algal ancestors of plants were completely immersed in water and dissolved minerals. The adaptation to land involved the differentiation of the plant body into roots,
Introduction to Plant Transport The algal ancestors of plants were completely immersed in water and dissolved minerals. The adaptation to land involved the differentiation of the plant body into roots,
13.2 The Vascular Plant Body (textbook p )
") 13.2 The Vascular Plant Body (textbook p544 550) Learning Goal: Label and explain the anatomy of the Vascular Plant and it's Tissue Types Plants are classified into two main groups: and. Vascular plants
13.2 The Vascular Plant Body (textbook p544 550) Learning Goal: Label and explain the anatomy of the Vascular Plant and it's Tissue Types Plants are classified into two main groups: and. Vascular plants
GFP GAL bp 3964 bp
 Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
The Science of Plants in Agriculture Pl.Sci 102. Getting to Know Plants
 The Science of Plants in Agriculture Pl.Sci 102 Getting to Know Plants Growth and Development of Plants Growth and Development of Plants Why it s important to have knowledge about plant development. What
The Science of Plants in Agriculture Pl.Sci 102 Getting to Know Plants Growth and Development of Plants Growth and Development of Plants Why it s important to have knowledge about plant development. What
E. THE FUNCTIONING OF HORMONES IN PLANT GROWTH AND DEVELOPMENT
 E. THE FUNCTIONING OF HORMONES IN PLANT GROWTH AND DEVELOPMENT E1. The Transport of Auxins David A. Morris 1, Jiří Friml 2, and Eva Zažímalová 3 1 School of Biological Sciences, University of Southampton,
E. THE FUNCTIONING OF HORMONES IN PLANT GROWTH AND DEVELOPMENT E1. The Transport of Auxins David A. Morris 1, Jiří Friml 2, and Eva Zažímalová 3 1 School of Biological Sciences, University of Southampton,
Cells and Tissues PART B
 3 Cells and Tissues PART B PowerPoint Lecture Slide Presentation by Jerry L. Cook, Sam Houston University ESSENTIALS OF HUMAN ANATOMY & PHYSIOLOGY EIGHTH EDITION ELAINE N. MARIEB Cellular Physiology: Membrane
3 Cells and Tissues PART B PowerPoint Lecture Slide Presentation by Jerry L. Cook, Sam Houston University ESSENTIALS OF HUMAN ANATOMY & PHYSIOLOGY EIGHTH EDITION ELAINE N. MARIEB Cellular Physiology: Membrane
Biology 2022 General Botany Exam 1 October 8, 2013
 Biology 2022 General Botany Exam 1 October 8, 2013 Please read the entire question before answering, put your answers on the Scantron form - not on this page 1. Plants. a. are heterotrophic b. have a peptidogycan
Biology 2022 General Botany Exam 1 October 8, 2013 Please read the entire question before answering, put your answers on the Scantron form - not on this page 1. Plants. a. are heterotrophic b. have a peptidogycan
BIOELECTRIC PHENOMENA
 Chapter 11 BIOELECTRIC PHENOMENA 11.3 NEURONS 11.3.1 Membrane Potentials Resting Potential by separation of charge due to the selective permeability of the membrane to ions From C v= Q, where v=60mv and
Chapter 11 BIOELECTRIC PHENOMENA 11.3 NEURONS 11.3.1 Membrane Potentials Resting Potential by separation of charge due to the selective permeability of the membrane to ions From C v= Q, where v=60mv and
AP Biology Chapter 36
 Chapter 36 Chapter 36 Transport in Plants 2006-2007 Transport in plants - Overview H2O & minerals transport in xylem transpiration evaporation, adhesion & cohesion negative pressure Sugars transport in
Chapter 36 Chapter 36 Transport in Plants 2006-2007 Transport in plants - Overview H2O & minerals transport in xylem transpiration evaporation, adhesion & cohesion negative pressure Sugars transport in
LECTURE 4: PHOTOTROPISM
 http://smtom.lecture.ub.ac.id/ Password: https://syukur16tom.wordpress.com/ LECTURE 4: PHOTOTROPISM LECTURE FLOW 1. 2. 3. 4. 5. INTRODUCTION DEFINITION INITIAL STUDY PHOTROPISM MECHANISM PHOTORECEPTORS
http://smtom.lecture.ub.ac.id/ Password: https://syukur16tom.wordpress.com/ LECTURE 4: PHOTOTROPISM LECTURE FLOW 1. 2. 3. 4. 5. INTRODUCTION DEFINITION INITIAL STUDY PHOTROPISM MECHANISM PHOTORECEPTORS
Introduction to Plant Transport
 Introduction to Plant Transport The algal ancestors of plants were completely immersed in water and dissolved minerals. The adaptation to land involved the differentiation of the plant body into roots,
Introduction to Plant Transport The algal ancestors of plants were completely immersed in water and dissolved minerals. The adaptation to land involved the differentiation of the plant body into roots,
23 2 Roots Slide 2 of 36
 2 of 36 Types of Roots Types of Roots What are the two main types of roots? 3 of 36 Types of Roots The two main types of roots are: taproots, which are found mainly in dicots, and fibrous roots, which
2 of 36 Types of Roots Types of Roots What are the two main types of roots? 3 of 36 Types of Roots The two main types of roots are: taproots, which are found mainly in dicots, and fibrous roots, which
35 Transport in Plants
 Transport in Plants 35 Transport in Plants 35.1 How Do Plants Take Up Water and Solutes? 35.2 How Are Water and Minerals Transported in the Xylem? 35.3 How Do Stomata Control the Loss of Water and the
Transport in Plants 35 Transport in Plants 35.1 How Do Plants Take Up Water and Solutes? 35.2 How Are Water and Minerals Transported in the Xylem? 35.3 How Do Stomata Control the Loss of Water and the
Transport in Plants AP Biology
 Transport in Plants 2006-2007 Water & mineral absorption Water absorption from soil osmosis aquaporins Mineral absorption active transport proton pumps active transport of H + aquaporin root hair proton
Transport in Plants 2006-2007 Water & mineral absorption Water absorption from soil osmosis aquaporins Mineral absorption active transport proton pumps active transport of H + aquaporin root hair proton
Class XI Chapter 6 Anatomy of Flowering Plants Biology
 Class XI Chapter 6 Anatomy of Flowering Plants Biology Question 1: State the location and function of different types of meristem. Meristems are specialised regions of plant growth. The meristems mark
Class XI Chapter 6 Anatomy of Flowering Plants Biology Question 1: State the location and function of different types of meristem. Meristems are specialised regions of plant growth. The meristems mark
Figure 18.1 Blue-light stimulated phototropism Blue light Inhibits seedling hypocotyl elongation
 Blue Light and Photomorphogenesis Q: Figure 18.3 Blue light responses - phototropsim of growing Corn Coleoptile 1. How do we know plants respond to blue light? 2. What are the functions of multiple BL
Blue Light and Photomorphogenesis Q: Figure 18.3 Blue light responses - phototropsim of growing Corn Coleoptile 1. How do we know plants respond to blue light? 2. What are the functions of multiple BL
Root System Architecture from Coupling Cell Shape to Auxin Transport
 Root System Architecture from Coupling Cell Shape to Auxin Transport PLoS BIOLOGY Marta Laskowski 1,2[, Verônica A. Grieneisen 3[, Hugo Hofhuis 2[, Colette A. ten Hove 2, Paulien Hogeweg 3, Athanasius
Root System Architecture from Coupling Cell Shape to Auxin Transport PLoS BIOLOGY Marta Laskowski 1,2[, Verônica A. Grieneisen 3[, Hugo Hofhuis 2[, Colette A. ten Hove 2, Paulien Hogeweg 3, Athanasius
The three principal organs of seed plants are roots, stems, and leaves.
 23 1 Specialized Tissues in Plants Seed Plant Structure The three principal organs of seed plants are roots, stems, and leaves. 1 of 34 23 1 Specialized Tissues in Plants Seed Plant Structure Roots: absorb
23 1 Specialized Tissues in Plants Seed Plant Structure The three principal organs of seed plants are roots, stems, and leaves. 1 of 34 23 1 Specialized Tissues in Plants Seed Plant Structure Roots: absorb
ABSORPTION OF WATER MODE OF WATER ABSORPTION ACTIVE AND PASSIVE ABSORPTION AND FACTORS AFFECTING ABSORPTION.
 ABSORPTION OF WATER MODE OF WATER ABSORPTION ACTIVE AND PASSIVE ABSORPTION AND FACTORS AFFECTING ABSORPTION. PRELUDE OF WATER POTENTIAL Most organisms are comprised of at least 70% or more water. Some
ABSORPTION OF WATER MODE OF WATER ABSORPTION ACTIVE AND PASSIVE ABSORPTION AND FACTORS AFFECTING ABSORPTION. PRELUDE OF WATER POTENTIAL Most organisms are comprised of at least 70% or more water. Some
Homework for Monday: Correct potometer questions Complete transport in plants worksheet
 Transport in plants Homework for Monday: Correct potometer questions Complete transport in plants worksheet Transpiration the loss of water from a plant through evaporation Did you know? A 15m maple tree
Transport in plants Homework for Monday: Correct potometer questions Complete transport in plants worksheet Transpiration the loss of water from a plant through evaporation Did you know? A 15m maple tree
Ginkgo leaf. Ginkgo is dioecious, separate sexes: male and female plants are separate. Monoecious plants have both male and female parts.
 Ginkgo leaf Figure 22-30 Ginkgo tree. Ginkgo is dioecious, separate sexes: male and female plants are separate. Monoecious plants have both male and female parts. The vein pattern is dichotomous: Divided
Ginkgo leaf Figure 22-30 Ginkgo tree. Ginkgo is dioecious, separate sexes: male and female plants are separate. Monoecious plants have both male and female parts. The vein pattern is dichotomous: Divided
Bald cypress Taxodium distichum in a swamp in North Carolina
 Bald cypress Taxodium distichum in a swamp in North Carolina Bald cypress is another deciduous gymnosperm. It is native to the SE United States. It can tolerate a wide range of soil conditions. It is not
Bald cypress Taxodium distichum in a swamp in North Carolina Bald cypress is another deciduous gymnosperm. It is native to the SE United States. It can tolerate a wide range of soil conditions. It is not
UNIT 6 - STRUCTURES OF FLOWERING PLANTS & THEIR FUNCTIONS
 6.1 Plant Tissues A tissue is a group of cells with common function, structures or both. In plants we can find 2 types of tissues: Meristem Permanent tissues Meristem is found in regions with continuous
6.1 Plant Tissues A tissue is a group of cells with common function, structures or both. In plants we can find 2 types of tissues: Meristem Permanent tissues Meristem is found in regions with continuous
 Question 1: State the location and function of different types of meristem. Meristems are specialised regions of plant growth. The meristems mark the regions where active cell division and rapid division
Question 1: State the location and function of different types of meristem. Meristems are specialised regions of plant growth. The meristems mark the regions where active cell division and rapid division
Biology Exam #1 Study Guide. True/False Indicate whether the statement is true or false. F 1. All living things are composed of many cells.
 Biology Exam #1 Study Guide True/False Indicate whether the statement is true or false. F 1. All living things are composed of many cells. T 2. Membranes are selectively permeable if they allow only certain
Biology Exam #1 Study Guide True/False Indicate whether the statement is true or false. F 1. All living things are composed of many cells. T 2. Membranes are selectively permeable if they allow only certain
Parenchyma Cell. Magnification 2375X
 Parenchyma Cell The large size of parenchyma cells is due in part to their relatively large vacuole (V) and in part also to the large number of chloroplasts (Cp) they contain. From a crimson clover, Trifolium
Parenchyma Cell The large size of parenchyma cells is due in part to their relatively large vacuole (V) and in part also to the large number of chloroplasts (Cp) they contain. From a crimson clover, Trifolium
AP Biology. Transport in plants. Chapter 36. Transport in Plants. Transport in plants. Transport in plants. Transport in plants. Transport in plants
 Chapter 36. Transport in Plants evaporation, adhesion & cohesion negative pressure evaporation, adhesion & cohesion negative pressure transport in phloem bulk flow Calvin cycle in leaves loads sucrose
Chapter 36. Transport in Plants evaporation, adhesion & cohesion negative pressure evaporation, adhesion & cohesion negative pressure transport in phloem bulk flow Calvin cycle in leaves loads sucrose
Topic 2: Plant Structure & Growth Ch. 35 Angiosperms are the most complex plants. They are composed of cells, tissues, organs and organ systems.
 Topic 2: Plant Structure & Growth Ch. 35 Angiosperms are the most complex plants. They are composed of cells, tissues, organs and organ systems. Fig. 35.8 Plant Cells pp.798-802 Types of plant cells Include:
Topic 2: Plant Structure & Growth Ch. 35 Angiosperms are the most complex plants. They are composed of cells, tissues, organs and organ systems. Fig. 35.8 Plant Cells pp.798-802 Types of plant cells Include:
The plant body has a hierarchy of organs, tissues, and cells. Plants, like multicellular animals:
 Chapter 28 The plant body has a hierarchy of organs, tissues, and cells Plants, like multicellular animals: o Have organs composed of different tissues, which are in turn composed of cells 3 basic organs:
Chapter 28 The plant body has a hierarchy of organs, tissues, and cells Plants, like multicellular animals: o Have organs composed of different tissues, which are in turn composed of cells 3 basic organs:
ARE YOU familiar with the sayings Get to
 Root Anatomy ARE YOU familiar with the sayings Get to the root of the problem or the root of all evil? Both these sayings suggest that the root is an essential part of something. With plants, the essential
Root Anatomy ARE YOU familiar with the sayings Get to the root of the problem or the root of all evil? Both these sayings suggest that the root is an essential part of something. With plants, the essential
Gravity plays a major role in plant morphogenesis by determining
 Gravity-regulated differential auxin transport from columella to lateral root cap cells Iris Ottenschläger*, Patricia Wolff*, Chris Wolverton, Rishikesh P. Bhalerao,Göran Sandberg, Hideo Ishikawa, Mike
Gravity-regulated differential auxin transport from columella to lateral root cap cells Iris Ottenschläger*, Patricia Wolff*, Chris Wolverton, Rishikesh P. Bhalerao,Göran Sandberg, Hideo Ishikawa, Mike
Biology 1030 Winter 2009
 Meeting Tissue Needs II Chapter 36 (738-755) Chapter 37 (756-770) Cellular Currency Plants harvest solar energy Photosynthesis Produces sugars Proteins, nucleic acids, lipids? H 2 O CO 2 Plants cells still
Meeting Tissue Needs II Chapter 36 (738-755) Chapter 37 (756-770) Cellular Currency Plants harvest solar energy Photosynthesis Produces sugars Proteins, nucleic acids, lipids? H 2 O CO 2 Plants cells still
Contains ribosomes attached to the endoplasmic reticulum. Genetic material consists of linear chromosomes. Diameter of the cell is 1 m
 1. (a) Complete each box in the table, which compares a prokaryotic and a eukaryotic cell, with a tick if the statement is correct or a cross if it is incorrect. Prokaryotic cell Eukaryotic cell Contains
1. (a) Complete each box in the table, which compares a prokaryotic and a eukaryotic cell, with a tick if the statement is correct or a cross if it is incorrect. Prokaryotic cell Eukaryotic cell Contains
Membrane transport 1. Summary
 Membrane transport 1. Summary A. Simple diffusion 1) Diffusion by electrochemical gradient no energy required 2) No channel or carrier (or transporter protein) is needed B. Passive transport (= Facilitated
Membrane transport 1. Summary A. Simple diffusion 1) Diffusion by electrochemical gradient no energy required 2) No channel or carrier (or transporter protein) is needed B. Passive transport (= Facilitated
2/25/2013. o Plants take up water and minerals from below ground o Plants take up CO2 and light from above ground THREE BASIC PLANT ORGANS ROOTS
 o Plants take up water and minerals from below ground o Plants take up CO2 and light from above ground THREE BASIC PLANT ORGANS o Roots o Stems o Leaves ROOTS o Anchor plant o Absorb water and minerals
o Plants take up water and minerals from below ground o Plants take up CO2 and light from above ground THREE BASIC PLANT ORGANS o Roots o Stems o Leaves ROOTS o Anchor plant o Absorb water and minerals
CONTROL OF GROWTH BY HORMONES
 CONTROL OF GROWTH BY HORMONES Growth and organogenesis are controlled......by genes (independent of environment): e.g., number of primary vascular bundles, general shape of a leaf or flower...by genes
CONTROL OF GROWTH BY HORMONES Growth and organogenesis are controlled......by genes (independent of environment): e.g., number of primary vascular bundles, general shape of a leaf or flower...by genes
Lecture 3 13/11/2018
 Lecture 3 13/11/2018 1 Plasma membrane ALL cells have a cell membrane made of proteins and lipids. protein channel Cell Membrane Layer 1 Layer 2 lipid bilayer protein pump Lipid bilayer allows water, carbon
Lecture 3 13/11/2018 1 Plasma membrane ALL cells have a cell membrane made of proteins and lipids. protein channel Cell Membrane Layer 1 Layer 2 lipid bilayer protein pump Lipid bilayer allows water, carbon
Plants. Plant Form and Function. Tissue Systems 6/4/2012. Chapter 17. Herbaceous (nonwoody) Woody. Flowering plants can be divided into two groups:
 Woody. Flowering plants can be divided into two groups:") Monocots Dicots 6/4/2012 Plants Plant Form and Function Chapter 17 Herbaceous (nonwoody) In temperate climates, aerial parts die back Woody In temperate climates, aerial parts persist The Plant Body Functions
Monocots Dicots 6/4/2012 Plants Plant Form and Function Chapter 17 Herbaceous (nonwoody) In temperate climates, aerial parts die back Woody In temperate climates, aerial parts persist The Plant Body Functions
NOTES: CH 35 - Plant Structure & Growth
 NOTES: CH 35 - Plant Structure & Growth In their evolutionary journey, plants adapted to the problems of a terrestrial existence as they moved from water to land ANGIOSPERMS (flowering plants) -most diverse
NOTES: CH 35 - Plant Structure & Growth In their evolutionary journey, plants adapted to the problems of a terrestrial existence as they moved from water to land ANGIOSPERMS (flowering plants) -most diverse
Bring Your Text to Lab!!!
 Bring Your Text to Lab!!! Vascular Plant Anatomy: Flowering Plants Objectives: 1. To observe what the basic structure of vascular plants is, and how and where this form originates. 2. To begin to understand
Bring Your Text to Lab!!! Vascular Plant Anatomy: Flowering Plants Objectives: 1. To observe what the basic structure of vascular plants is, and how and where this form originates. 2. To begin to understand
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL
 GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
Types of Plants. Unit 6 Review 5/2/2011. Plants. A. pine B. moss C. corn plant D. bean plant E. liverwort
 Unit 6 Review Plants Initial questions are worth 1 point each. Each question will be followed by an explanation All questions will be asked a second time at the very end, each of those questions will be
Unit 6 Review Plants Initial questions are worth 1 point each. Each question will be followed by an explanation All questions will be asked a second time at the very end, each of those questions will be
1. Transpiration may be defined as the loss of water vapour by diffusion from a plant to its environment.
 1. Transpiration may be defined as the loss of water vapour by diffusion from a plant to its environment. The diagram below shows apparatus that can be used to estimate transpiration rates of a leafy shoot.
1. Transpiration may be defined as the loss of water vapour by diffusion from a plant to its environment. The diagram below shows apparatus that can be used to estimate transpiration rates of a leafy shoot.
Models of long-distance transport: how is carrier-dependent auxin transport regulated in the stem?
 Research Models of long-distance transport: how is carrier-dependent auxin transport regulated in the stem? Michael Renton 1,2, Jim Hanan 3, Brett J. Ferguson 4 and Christine A. Beveridge 5 1 School of
Research Models of long-distance transport: how is carrier-dependent auxin transport regulated in the stem? Michael Renton 1,2, Jim Hanan 3, Brett J. Ferguson 4 and Christine A. Beveridge 5 1 School of
Plant Tissues and Organs. Topic 13 Plant Science Subtopics , ,
 Plant Tissues and Organs Topic 13 Plant Science Subtopics 13.1.2, 13.1.3, 13.1.4 Objectives: List and describe the major plant organs their structure and function List and describe the major types of plant
Plant Tissues and Organs Topic 13 Plant Science Subtopics 13.1.2, 13.1.3, 13.1.4 Objectives: List and describe the major plant organs their structure and function List and describe the major types of plant
Plant Structure and Growth
 Plant Structure and Growth A. Flowering Plant Parts: The flowering plants or are the most diverse group of plants. They are divided into 2 classes and. Examples of monocots: Examples of dicots: The morphology
Plant Structure and Growth A. Flowering Plant Parts: The flowering plants or are the most diverse group of plants. They are divided into 2 classes and. Examples of monocots: Examples of dicots: The morphology
LECTURE 4: SHORTDISTANCE TRANSPORT OF NUTRIENTS
 LECTURE 4: SHORTDISTANCE TRANSPORT OF NUTRIENTS http://vitae-scientia.tumblr.com/ COMPETENCY After completing this Lecture and mastering the lecture materials, the competency expected to develop includes
LECTURE 4: SHORTDISTANCE TRANSPORT OF NUTRIENTS http://vitae-scientia.tumblr.com/ COMPETENCY After completing this Lecture and mastering the lecture materials, the competency expected to develop includes
Cell Review. 1. The diagram below represents levels of organization in living things.
 Cell Review 1. The diagram below represents levels of organization in living things. Which term would best represent X? 1) human 2) tissue 3) stomach 4) chloroplast 2. Which statement is not a part of
Cell Review 1. The diagram below represents levels of organization in living things. Which term would best represent X? 1) human 2) tissue 3) stomach 4) chloroplast 2. Which statement is not a part of
SESSION 6: SUPPORT AND TRANSPORT SYSTEMS IN PLANTS PART 1
 SESSION 6: SUPPORT AND TRANSPORT SYSTEMS IN PLANTS PART 1 KEY CONCEPTS In this session we will focus on summarising what you need to know about: - Anatomy of dicotyledonous plants Root and stem: distribution
SESSION 6: SUPPORT AND TRANSPORT SYSTEMS IN PLANTS PART 1 KEY CONCEPTS In this session we will focus on summarising what you need to know about: - Anatomy of dicotyledonous plants Root and stem: distribution
CELL BIOLOGY - CLUTCH CH. 9 - TRANSPORT ACROSS MEMBRANES.
 !! www.clutchprep.com K + K + K + K + CELL BIOLOGY - CLUTCH CONCEPT: PRINCIPLES OF TRANSMEMBRANE TRANSPORT Membranes and Gradients Cells must be able to communicate across their membrane barriers to materials
!! www.clutchprep.com K + K + K + K + CELL BIOLOGY - CLUTCH CONCEPT: PRINCIPLES OF TRANSMEMBRANE TRANSPORT Membranes and Gradients Cells must be able to communicate across their membrane barriers to materials
Chapter #35~ Plant Structure and Growth
 Chapter #35~ Plant Structure and Growth What part of a plant is represented by each of these: Carrot Celery Red Pepper Tomato Lettuce Garbanzo Bean Angiosperm structure Three basic organs: Roots (root
Chapter #35~ Plant Structure and Growth What part of a plant is represented by each of these: Carrot Celery Red Pepper Tomato Lettuce Garbanzo Bean Angiosperm structure Three basic organs: Roots (root
Chapter 23 Notes Roots Stems Leaves
 Chapter 23 Notes Roots Stems Leaves I. Specialized tissue in plants - effective way to ensure the plant s survival A. Seed plant structure 1. Roots - a. Absorbs water and dissolves nutrients b. anchors
Chapter 23 Notes Roots Stems Leaves I. Specialized tissue in plants - effective way to ensure the plant s survival A. Seed plant structure 1. Roots - a. Absorbs water and dissolves nutrients b. anchors
Honors Biology-CW/HW Cell Biology 2018
 Class: Date: Honors Biology-CW/HW Cell Biology 2018 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Hooke s discovery of cells was made observing a. living
Class: Date: Honors Biology-CW/HW Cell Biology 2018 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Hooke s discovery of cells was made observing a. living
Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated
 Relative conversion rates of different plant hormones to their glucosylated") Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated form by recombinant UGT74E2. The naturally occurring auxin
Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated form by recombinant UGT74E2. The naturally occurring auxin
(17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON
 CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON") (17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON Jim Burton 1 and Marianne Pedersen Abstract. Cyclanilide [1-(2,4-dichlorophenylaminocarbonyl)-cyclopropane carboxylic
(17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON Jim Burton 1 and Marianne Pedersen Abstract. Cyclanilide [1-(2,4-dichlorophenylaminocarbonyl)-cyclopropane carboxylic
Chapter C3: Multicellular Organisms Plants
 Chapter C3: Multicellular Organisms Plants Multicellular Organisms Multicellular organisms have specialized cells of many different types that allow them to grow to a larger size than single-celled organisms.
Chapter C3: Multicellular Organisms Plants Multicellular Organisms Multicellular organisms have specialized cells of many different types that allow them to grow to a larger size than single-celled organisms.
8/25/ Opening Questions: Are all living things made of cells? What are at least five things you know about cells?
 Chapter 3 The Cell: Module Hyperlinks 3.1 Cells are the fundamental units of life 3.2 Plant vs. animal cells 3.3 Membranes: structure 3.4 Membranes: function 3.5 The nucleus 3.6 Organelles in protein production
Chapter 3 The Cell: Module Hyperlinks 3.1 Cells are the fundamental units of life 3.2 Plant vs. animal cells 3.3 Membranes: structure 3.4 Membranes: function 3.5 The nucleus 3.6 Organelles in protein production
Title: Nutrient Movement Towards and Into Plant Roots Speaker: Bill Pan. online.wsu.edu
 Title: Nutrient Movement Towards and Into Plant Roots Speaker: Bill Pan online.wsu.edu Unit 1, Lesson 4 Nutrient Movement Towards and Into Plant Roots http://soils.usda.gov/education/resources/k_12/lessons/profile/
Title: Nutrient Movement Towards and Into Plant Roots Speaker: Bill Pan online.wsu.edu Unit 1, Lesson 4 Nutrient Movement Towards and Into Plant Roots http://soils.usda.gov/education/resources/k_12/lessons/profile/