Functional Analysis of the RabGAP RN-Tre in Drosophila. Lei Shi
|
|
|
- Virginia Reeves
- 6 years ago
- Views:
Transcription
1 Functional Analysis of the RabGAP RN-Tre in Drosophila Lei Shi Department of Biology McGill University Montreal, Quebec, Canada August 5, 2009 A thesis submitted to McGill University in partial fulfillment of the requirements for the degree of Master of Science Lei Shi,
2 Table of contents List of figures 4 List of abbreviations used 5 Abstract 6 Résumé 7 Acknowledgements 9 Chapter 1. Introduction Rab family small GTPases and their regulators The function of Rab RabGAP RN-Tre Drosophila as a genetic model to study the in vivo function of Rab5 and RN-Tre Drosophila wing development Drosophila compound eye development The Drosophila RN-Tre gene 26 Chapter 2. Results Overexpression analysis Loss-of-function analysis Western blot analysis of RN-Tre mutant Analysis of membrane trafficking defects in RN-Tre mutants Pulse-chase experiments under non-permeable condition Pulse-chase experiments under permeable condition RNA interference analysis Investigating the potential genetic interaction between RN-Tre and Notch Investigating the potential genetic interaction between RN-Tre and Rab Materials and Methods Western blot analysis 53 2
3 2.6.2 Pulse chase experiments Genetics 56 Chapter 3. Discussion 58 References 68 3
4 List of figures Chapter 1, Figure 1.1. Functions of Rabs in endocytic pathways 12 Chapter 1, Figure 1.2. The structure of RN-Tre. 16 Chapter 1, Figure 1.3. One previously proposed model 19 Chapter 1, Figure Another proposed model 21 Chapter 1, Figure 1.5. RN-Tre mutants used in my experiments 27 Chapter 2, Figure 2.1. Western blot analysis of RN-Tre GSV mutants 32 Chapter 2, Figure 2.2. Overexpression of RN-Tre in third-instar eyes 35 Chapter 2, Figure 2.3. Schematic drawing of pulse chase. 38 Chapter 2, Figure 2.4. Pulse chase of Notch in the wing disc. 41 Chapter 2, Figure 2.5. Schematic drawing of pulse chase.. 43 Chapter 2, Figure 2.6. Pulse chase of Notch in the wing disc. 45 Chapter 2, Figure 2.7. Pulse chase of Notch in the wing disc 48 Chapter 2, Figure 2.8. Reducing the dosage of RN-Tre. 51 Chapter 2, Figure 2.9. Rab5 mutations enhanced the RN-tre 54 Chapter 3, Figure A proposed model for the role. 65 4
5 List of abbreviations used EGFR EMS FasII GAP GDF GDI GDP GTP MDCK NGS PBS Rab RNAi R-cells WT Epidermal growth factor receptor Ethylmethane Sulphonate Fasciculin II GTPase-activating-protein GDI-displacement factor GDP dissociation inhibitor Guanosine diphosphate Guanosine triphosphate Madin-Darby Canine Kidney Cells Normal goat serum Phosphate buffer saline Ras-like in rat brain RNA interference Photoreceptor cells Wild type 5
6 Abstract Rab-family small monomeric GTP-binding proteins are well known for their role in regulating intracellular vesicular transport required for signal transduction, cell growth, cell differentiation, and development. GTP-hydrolysis activating proteins (i.e. GAPs) function as negative regulators of Rabs for controlling the lifetime of Rabs at active state. RN-tre is an evolutionarily conserved Rab GAP and has been shown to possess GAP activity towards Rab5 in vitro. Previous cell culture studies on mammalian RN-tre, however, show controversial results regarding whether RN-Tre is involved in regulating endocytosis. As genetic analysis of RN-tre has not been performed in any experimental organism, the biological function of RN-tre remains unknown. In my M. Sc project, I performed genetic experiments to determine the function of RN-Tre in Drosophila. The results from overexpression experiments show that increasing the level of RN-Tre decreased the surface level of cell surface proteins and induced the accumulation of membrane proteins in vesicle-like structures. Mutations in the RN-Tre gene, or knocking down the expression of RN-Tre with RNA interference, significantly inhibited Notch endocytosis. Consistently, reducing the dosage of the RN-Tre gene suppressed the wing phenotype caused by Notch haploinsufficiency. These results support that RN-Tre regulates the steady-state level of cell surface receptors by promoting endocytosis. The genetic interaction between RN-Tre and Rab5 suggests a model in which RN-Tre promotes endocytosis by facilitating the recycling of Rab5. My M. Sc. study thus reveals for the first time the in vivo function of RN-Tre, and provides novel and important insights into functional relationship between Rabs and GAPs during development. 6
7 Résumé Les protéines de liaison du GTP de la famille Rab sont bien connues pour le rôle qu'elles jouent dans la régulation du transport vésiculaire intracellulaire requis lors de la transduction de signaux, ainsi que la croissance, la différentiation et le développement cellulaire. Les protéines d'hydrolyse du GTP (les GAPs) agissent comme régulateurs négatifs pour contrôler la durée de vie des Rabs à l'état actif. RN-Tre est un GAP Rab conservé évolutivement et il a été démontré que cette protéine était GAP-active sur Rab5 in vitro. Cependant, des cultures cellulaires précédentes sur le RN-Tre mammifère ont montré des résultats controversés par rapport au rôle du RN-Tre dans la régulation de l'endocytose. Étant donné que l'analyse génétique du RN-Tre n'a pas été effectué chez aucun organisme expérimental, la fonction biologique du RN-Tre demeure inconnue. Au cours de mon projet de maîtrise, j'ai réalisé des expériences génétiques pour déterminer la fonction du RN-Tre chez la drosophile. Les résultats des expériences de surexpression démontrent que l'augmentation des niveaux de RN-Tre réduisait le niveau de surface des protéines de surface et induisait l'accumulation de protéines membranaires dans des structures ressemblant à des vésicules. Des mutations dans le gène du RN-Tre ou un knock-down de l'expression du RN-Tre grâce à l'arn interférent inhiba de façon significative l'endocytose de Notch. De la même manière, en réduisant le dosage du gène RN-Tre suppréssait le phénotype ailes causé par une haplo-insuffisance de Notch. Ces résultats supportent l'hypothèse que le RN-Tre régule l'équilibre dynamique des récepteurs en surface cellulaire en promouvant l'endocytose. L'interaction génétique entre RN-Tre et Rab5 suggère un model selon lequel RN-Tre promouvoit l'endocytose en facilitant le recyclage de Rab5. Mon étude de maîtrise révèle pour la première fois le 7
8 fonctionnement in vivo du RN-Tre et fournit de nouvelles et importantes connaissances sur la relation fonctionnelle entre les Rabs et GAPs lors du développement. 8
9 Acknowledgements I would like to acknowledge and extend my heartfelt gratitude to those who have given their generous support to help me make the completion of this work possible. I wish to thank: My supervisor, Dr. Yong Rao, for giving me the opportunity to work in his lab, and providing me with vital guidance, advice and incredible support all along my study. My graduate research advisory committee member, Dr. Don Van meyel, for his encouragement, guidance and support. Dr. Laura Nilson, for her guidance as a member of my graduate research advisory committee. Dr. Tarek Houalla for initiating work on RN-Tre and Ms Wen-tzu Chang for her technical help I am grateful to everyone that supported me during my years of study at McGill. I would like to thank my parents for giving me the most touching and selfless love that encourage me at both good and difficult times, as well as my friends and lab colleagues for giving me precious friendships and kind help. Finally I would like to thank the One who has given me all this goodness in my life. 9
10 Chapter 1 INTRODUCTION 10
11 1.1 Rab family small GTPases and their regulators Rab proteins are small GTPases (21-25 kda) involved in regulating intracellular vesicular transport in many cellular processes such as endocytosis, exocytosis, signal transduction and cell differentiation (Molendijk et al., 2004; Seabra and Coudrier, 2004; Spang, 2004) (Fig.1.1). The first Rab family protein YPT1p was identified in yeast in 1983 (Gallwitz et al., 1983). Mutations in the YPT1 gene caused the accumulation of abnormal Golgi structures and arrested growth (Segev and Botstein, 1987). Later study revealed that YPT1 is required for vesicular trafficking from endoplasmic reticulum (ER) to the Golgi complex (Baker et al., 1990). Genetic screen for mutations affecting yeast secretion also identified another Rab family protein SEC4p (Novick et al., 1980). In 1987, Salminen and Novick demonstrated that the SEC4p protein is required for post-golgi secretion (Salminen and Novick, 1987). In 1987, Tavitian and colleagues identified vertebrate homologs of YPT1 and SEC4 (Touchot et al., 1987). They used DNA probes encoding a highly conserved sequence Asp-Thr-Ala-Gly-Gln-Glu found in the Ras protein to screen a rat brain cdna library. From this screen, they identified four cdna clones that encode for proteins displaying high homology to yeast YPT1 and SEC4. They named the four proteins as Rab1, 2, 3 and 4 (for ras genes from rat brain). Rab proteins are present in two forms, active GTP-bound form and inactive GDPbound form (Seabra and Coudrier, 2004). In vitro studies show that the activity of Rab proteins can be stimulated by guanine nucleotide exchange factors (GEFs) that promote the exchange of GDP with GTP, and can also be inhibited by GTP-hydrolysis activating 11
12 12
13 Figure 1.1. Functions of Rabs in endocytic pathways (Rodman and Wandingerness, 2000). 13
14 proteins (GAPs), which enhance the GTPase activity of Rabs leading to the inactivation of Rabs. In vivo, Rabs may be activated temporally and spatially by GEFs, while GAPs may control the period of Rabs in active state. Thus, fully understanding the action of Rabs in vivo requires the identification and characterization of GEFs and GAPs that specifically control the activity of each Rab protein. My M.Sc. project is to study the in vivo role of RN-Tre, an evolutionarily conserved RabGAP, in endocytic pathways during development. A large number of Rabs have been found in the genome of human as well as experimental model systems such as mouse and Drosophila (Adams et al., 2000; Armstrong, 2000b; Pereira-Leal and Seabra, 2001). Similarly, a large number of GAPs are also found in both vertebrates and invertebrates (Bernards, 2003). There are ~52 potential GAPs in the human genome and 24 in the fly genome (Bernards, 2003), each may regulate the lifetime of a specific Rab in activation state. One important question is how to examine in vivo specificity of individual GAPs towards each member of Rabs. This is a difficult job because of the existence of large numbers of Rabs and GAPs in the genome. One such example is demonstrated by several recent studies of the functional relationship between Rab5 and its potential GAP RN-tre, which produced very controversial results (Haas et al., 2005; Lanzetti et al., 2000) The function of Rab5 Rab5 was originally identified by Tavitian and colleagues in 1989 (Zahraoui et al., 1989). They used a probe corresponding to the yeast SEC4 gene to screen a human pheochromocytoma cdna library. From this screen, they identified a cdna sequence 14
15 encoding for a 214-amino acid protein. This protein was named Rab5 that displays more homology to other human Rabs than SEC4. A later study by Zerial and coworkers (Chavrier et al., 1990) also independently cloned Rab5 from a human MDCK cdna library. Their immunocytochemical analysis revealed the association of Rab5 with the cytoplasmic side of the plasma membrane and the early endosome, suggesting the potential involvement of Rab5 in the early endocytic pathway. Subsequent studies indicate a key role for Rab5 in the early endocytic pathway (Bucci et al., 1992). Rab5 plays an important role in mediating receptor internalization, subsequent vesicular transport and fusion with the early endosome in C-elegans, Drosophila and mammals (reviewed in (Armstrong, 2000a; Mohrmann and van der Sluijs, 1999; Simpson and Jones, 2005; Somsel Rodman and Wandinger-Ness, 2000)). Accumulated studies demonstrate the involvement of Rab5 in modulating a number of important and evolutionarily conserved signaling pathways such as transforming growth factor- (TGF- ), epidermal growth factor (EGF), Wnt and Notch signaling in cell proliferation, cell differentiation, pattern formation and synaptic transmission at both developmental and adult stages (reviewed in (Armstrong, 2000a; Mohrmann and van der Sluijs, 1999; Simpson and Jones, 2005; Somsel Rodman and Wandinger-Ness, 2000)) RabGAP RN-Tre RN-tre, like most of RabGAPs, contains a Tre2/Bub2/Cdc16 (TBC) domain (or GAP domain) that accelerates the GTPase activity of Rabs with an Arg-Glu dual-finger mechanism (Pan et al., 2006) (Fig. 1.2). RN-Tre was originally identified by Di Fiore and colleagues as a binding partner of Eps8, a substrate of the epidermal growth factor 15
16 N GAP C N GAP C N GAP C 16
17 Figure 1.2. The structure of RN-Tre. There are three alternatively spliced forms of RN- Tre consistsing of 571, 457, 485 amino acid residues. Each has a N-terminal Rab GTPase-activating domain (GAP) and a C-terminal regulatory region. Three isoforms share the amino-acid sequence and differ in their C-terminal sequence (colored rectangles). 17
18 receptor (EGFR) (Matoskova et al., 1996). Subsequent studies by the same group show that RN-tre specifically accelerates GTP hydrolysis by Rab5 in vitro (Lanzetti et al., 2000). Consistently, overexpression of RN-tre in cultured cells were shown to block Rab- 5-dependent EGFR internalization (Lanzetti et al., 2000) (Fig. 1.3). That RN-tre functions as a Rab5 GAP, however, is challenged by a recent study, which argues that another RabGAP (i.e. RabGAP-5), but not RN-tre, negatively regulates the function of Rab5 in endocytic trafficking in cultured cells (Haas et al., 2005) (Fig. 1.4). Bar and coworkers show that RabGAP-5 can bind directly to Rab5. Interfering with the function of RabGAP-5 in cultured cells disrupted the trafficking of EGF and LAMP1, and caused the increase in the size of endosomes. Although their in vitro study also showed that RN-Tre was capable of promoting the GTPase activity of Rab5, they found that overexpression of RN-Tre failed to induce the redistribution of Rab5 effectors to the cytoplasm. Since they observed that RN-Tre also promote the GTPase activity of Rab41, they suggested that RN-tre is involved in trans-golgi vesicle transport, but not required in endocytosis. These controversial studies leave two unresolved issues. First, whether RNtre truly functions as a Rab5 GAP in vivo remains an open question. Second, as genetic analysis of RN-tre has not been performed in any model organism (all previous studies were done in cultured cells), the biological function of RN-tre remains unknown Drosophila as a genetic model to study the in vivo function of Rab5 and RN-Tre My M. Sc. project is to use the powerful Drosophila genetics to determine the in vivo function of RN-tre, for instance, whether RN-tre is required for endocytosis in vivo. There are several advantages to use Drosophila as an in vivo model to study the function 18
19 19
20 Figure 1.3. One previously proposed model for the role of RN-Tre in mammals. Studies in cultured cells suggest that RN-Tre enhances the GTPase activity of Rab5 and thus inhibits its function in endocytosis of EGFR. 20

21 21
22 Figure Another proposed model for the role of RN-Tre in mammals. A recent report suggests that Rab5GAP, but not RN-Tre, enhances the GTPase activity of Rab5, thus inhibiting its function in endocytosis. They suggested that RN-Tre is not involved in endocytosis, but may be involved in trans-golgi vesicular trafficking (Haas et al., Nature Cell Biol., 2005). 22
23 of Rab proteins and their regulators. First, Drosophila is a well-established genetic model system. A large collection of genetic material such as genetic markers, deficiency and balancer lines are available for genetic studies. Second, the Drosophila genome project has been completed. EST cdna clones for most genes in the Drosophila genome are also available for molecular analysis. Third, a large collection of mutant lines such as P- element insertions and transgenic RNAi lines are available for loss-of-function analysis. And fourth, the availability of a large number of tissue-specific GAL4 lines allows the over-expression or misexpression of a target gene in a tissue-specific manner for gain-offunction analysis. In my project, I used Drosophila wing and compound eye development to study the function of RN-tre Drosophila wing development The Drosophila wing develops from the wing imaginal disc. The wing imaginal disc has a sac-like structure consisting of the wing epithelial cell monolayer and the peripodial membrane. The wing disc is compartmentalized along both anterior-posterior and dorsal-ventral axes. At the larval stage, the wing disc begins to proliferate and differentiate in response to the pulse of ecdysone. Pattern formation in the wing development is controlled by several signaling pathways. Pattern formation along the anterior-posterior axis is controlled by Decapentaplegic (DPP), a morphogen produced by a stripe of anterior compartment cells located immediately adjacent to the posterior compartment (Morata and Sanchez-Herrero, 1999). The dorsal-ventral compartmentalization is controlled by the Notch signaling pathway. The Notch ligand 23
24 Serrate (Ser) and glycosyltransferase Fringe (Fng) are expressed in the dorsal compartment (Couso et al., 1995), which ensures the activation of Notch in ventral cells by Ser and the activation of Notch in dorsal cells by another ligand Delta (Fleming et al., 1997; Haines and Irvine, 2003; Panin et al., 1997). Pattern formation along the proximaldistal axis is controlled by Wingless, which is produced by a stripe of cells located at the dorsal-ventral boundary region (Morata and Sanchez-Herrero, 1999). DPP and Wingless also function together to control the uniform proliferation of cells in the entire wing disc. Several studies indicate a key role for Rab5-mediated endocytosis in regulating the action of DPP and Wingless in the wing development. In 2000, González-Gaitán and colleagues showed that the establishment of the long-range DPP gradient involves the Rab5-mediated endocytotic pathway (Entchev et al., 2000). Their results suggest a planar transcytosis mechanism for the establishment of the DPP gradient in which Rab5 and Dynamin mediate the endocytosis of DPP and its receptor complex in cells exposed to the morphogen, and subsequently DPP is recycled onto cell surface for the further spreading of DPP (Entchev et al., 2000). Later studies by other groups also implicate the Rab5- mediated endocytosis in the regulation of the function of the Wingless signaling pathway in the Drosophila wing. For instance, it is shown that knocking down the expression of Rab5 in the developing wing disc reduced the wingless signaling level (Seto and Bellen, 2006). This result indicates that internalization and endosomal trafficking of Wingless is required for the activation of downstream effectors that control the expression of Wingless-target genes. Another study by Eaton and coworkers showed that Rab5- mediated endocytosis restricts the spread of Wingless along the promixal-distal axis, 24
25 which facilitates the formation of appropriate Wingless gradient for controlling the expression of target gene expression (Marois et al., 2006). Rab5-mediated endocytosis has also been shown to be involved in regulating the function of Notch in the Drosophila wing disc. Some studies show that Rab-mediated endocytosis is required for the full activation of Notch in the Drosophila wing disc (Hori et al., 2004). Other studies suggest that Notch endocytosis is also required for the clearance of Notch from cell surface, thus preventing the hyper-activation of downstream signaling (Maitra et al., 2006) Drosophila compound eye development The development of the Drosophila adult compound eye begins at the third instar larval stage, when precursor cells differentiate into photoreceptor cells (R cells) and accessory cells in the eye-imaginal discs. The third-instar eye disc has a sac-like morphology consisting of the single epithelial cell monolayer and the peripodial membrane. The development of eye discs is marked by the posterior-to-anterior movement of the morphogenetic furrow across the eye disc. The movement of the furrow involves cycles of dynamic nuclear movement along the apical-basal axis. Initially, anterior precursor cell nuclei translocate basally, leading to the formation of the furrow. After they exit the furrow, cells differentiate and become R cells and accessory cells (Tomlinson, 1985). The posterior-to-anterior movement of the furrow is regulated by Hedgehog, which is produced by differentiating cells posterior to the furrow (Burke and Basler, 1997). The subsequent development of R cells and accessory also involves the 25
26 action of several other signaling pathways such as EGF, Notch and Sevenless (Basler and Hafen, 1991; Dominguez et al., 1998). A recent study by Lu and Bilder demonstrated the importance of Rab5-mediated endocytic pathway in controlling the epithelial polarity and proliferation of the thirdinstar eye disc (Lu and Bilder, 2005). They showed that loss of Rab5 disrupted the endocytic pathway, leading to the abnormal accumulation of cell surface receptors such as Notch and Crumbs on the cell surface. The increased level of these cell surface proteins in Rab5 null mutants disrupted the apico-basal polarity and thus caused the overproliferation of the eye disc The Drosophila RN-Tre gene The fly genome contains a single RN-tre gene (Yedvobnick and Smoller, 1998) (Fig. 1.5). The RN-Tre gene is located at cytological region 50C23 on the right arm of the second chromosome. The RN-Tre gene spans a genomic region of ~6 kb, containing eight exons and seven introns. The expression of the RN-Tre gene generates three transcripts with the length of 1987 bp, 2971 bp and 2697 bp, which encode for 571 amino-acid (RN-Tre-RA), 457 amino-acid (RN-Tre-RB) and 485 amino-acid proteins (RN-Tre-RC) respectively. Three RN-Tre isoforms share the amino-acid sequence containing the GAP domain and portion of the C-terminal regulatory region, and differ in the C-terminal sequence Fig Tarek Houalla, another graduate student in the Rao lab, worked on RN-tre previously. He found that overexpression of RN-tre caused nuclear migration defect in the eye (Houalla and Rao, unpublished data). To examine if RN-tre is indeed required for 26
27 27
28 Figure 1.5. RN-Tre mutants used in my experiments. The GSV allele carrying a nonsense mutation that is predicted to convert the codon for Trp 287 into a stop codon In the RN- Tre gene induced by EMS.The GSV allele was generated by another graduate student Tarek Houalla in the Rao lab. 28
29 nuclear migration, he performed loss-of-function analysis. He generated the RN-tre lossof-function mutant allele by EMS mutagenesis (Fig. 1.5). In a genetic screen, Tarek isolated the GSV mutation in the RN-tre gene that suppresses an adult rough eye phenotype resulting from the overexpression of the endogenous RN-tre gene. He then sequenced the RN-tre gene in the GSV allele and found that this mutation is a non-sense mutation as it converts a codon for Trp at amino-acid residue 287 into a stop codon, which leads to the production of a truncated RN-tre protein lacking the C-terminal regulatory sequence and part of GAP domain sequence. Tarek examined nuclear migration in RN-tre mutants, but did not observe a phenotype. He then focused on studying the role of Rab proteins in nuclear migration and thus no longer work on the RN-tre project. In my M. Sc project, I performed genetic studies to determine the in vivo function of RN-Tre. My results suggest that RN-tre is involved in endocytosis, arguing against previous report that RN-tre is not required for endocytosis (Haas et al., 2005). My results also suggest that RN-tre promotes endocytosis, which is in contrast to several previous reports that RN-tre negatively regulates endocytosis in cultured mammalian cells (Lanzetti et al., 2004; Lanzetti et al., 2000; Martinu et al., 2002). Genetic studies also provided promising data suggesting the interactions between RN-Tre and Notch, and between RN-Tre and Rab5. 29
30 Chapter 2 RESULTS 30
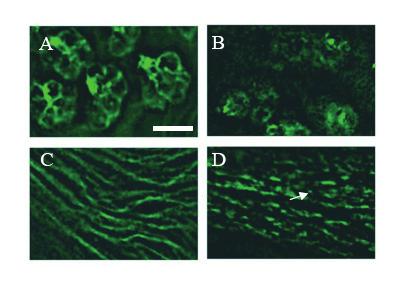
31 2.1 Overexpression analysis Previous studies on mammalian RN-Tre showed that overexpression of RN-Tre in cultured cells blocked the endocytosis of EGFR, suggesting a role for RN-Tre to negatively regulate endocytosis by inhibiting the activity of Rab5 (Lanzetti et al., 2000) (Martinu et al., 2002). To determine if fly RN-Tre also plays a similar role in Drosophila, we performed overexpression experiments to determine if overexpression of fly RN-Tre in the developing tissue blocks the endocytosis of cell surface receptors. It was shown that blocking endocytosis could cause the abnormal accumulation of cell surface receptors on the cell surface (Lu and Bilder, 2005; Wilkin et al., 2004). If fly RN-Tre also negatively regulates endocytosis, one would predict that overexpression of RN-Tre should also increase the surface level of cell surface receptors. To overexpress the fly RN-Tre in the third-instar eye disc, flies carrying the UAS- RN-Tre transgene were crossed with flies carrying the GAL4 gene under control of the eyespecific GMR promoter. Overexpression of RN-Tre was confirmed by immunostaining (Houalla and Rao, unpublished data). To examine the effect of RN-Tre overexpression on the distribution of cell surface receptors, third-instar eye imaginal discs were stained with MAb24B10, which recognizes the R-cell-specific cell adhesion molecule Chaoptin (Van Vactor et al., 1988). In wild type, Chaoptin staining was observed predominantly on the surface of R cells (Fig. 2.1A). Interestingly, in third-instar eye discs overexpressing RN-Tre, the surface staining of Chaoptin was significantly reduced (100%, n>10 eye discs. Fig. 2.1B). The surface Chaoptin staining appeared to be discontinuous. Chaoptin staining was also observed in vesicle-like structures within R-cell bodies and their axons (Fig. 2.1B and 2.1D). However, the nature of Chaoptin-positive vesicle-like structures is unknown. 31
32 32
33 Figure 2.1. Overexpression of RN-Tre in third-instar eyes disrupted the membrane distribution of cell adhesion molecule Chaoptin. (A and C) In wild type, Chaoptin is predominantly localized on the plasma membrane of R-cell bodies (A) and their axons (C) (n=8 discs). (B) In eye discs overexpressing RN-Tre, the surface level of Chaoptin was significantly decreased (n=10). (D) Overexpression of RN-Tre caused puncta-like (arrow) distribution of Chaoptin in R-cell axons. Scale bar: 5 μm. 33
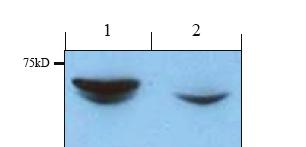
34 The above result thus does not support a role for RN-Tre to negatively regulate endocytosis. However, the appearance of Chaoptin in vesicle-like structures suggests that RN-Tre is required for vesicular transport in Drosophila. 2.2 Loss-of-function analysis To directly address the question if RN-Tre is required for the endocytosis of cell surface receptors in vivo, I performed loss-of-function analysis to determine the potential effect of RN-Tre mutations on the endocytosis of cell surface receptors. Tarek Houalla, another graduate student in the Rao lab, generated a RN-tre loss-offunction mutant allele called GSV by EMS mutagenesis (Fig.1.5). In a genetic screen, Tarek isolated the GSV mutation in the RN-tre gene that suppresses an adult rough eye phenotype resulting from the overexpression of the endogenous RN-Tre gene. He then sequenced the RN-tre gene in the GSV allele and found that this mutation is a non-sense mutation as it converts a codon for Trp at amino-acid residue 287 into a stop codon, which is predicted to generate a truncated RN-tre protein lacking the C-terminal regulatory sequence and part of GAP domain sequence (Fig.1.5) Western blot analysis of RN-Tre mutants To confirm that the GSV mutation in the RN-Tre gene indeed affects the production of the RN-Tre protein, I performed western blot analysis. The peptide corresponding to the amino-acid sequence of the RN-Tre protein was synthesized and used to raise polyclonal antibodies by Genscript Corporation (Piscataway, New Jersey). The affinitypurified anti-rn-tre antibody was then used in western blot analysis. In wild type, this antibody recognized one major band at ~65 kd, and a minor band at ~62kD. In GSV mutants, while the 62 kd band was still detected by the anti-rn-tre antibody and showed 34

35 35
36 Figure 2.2. Western blot analysis of RN-Tre GSV mutants. Lane 1, wild-type lysates. Lane 2, lysates prepared from GSV mutants. Similar results were obtained in two independent experiments. Since my antibody only recognizes the C-terminal region of RN-Tre, it remains possible that a truncated RN-Tre containing the portion of the GAP domain could be produced, and thus the RN-tre GSV allele may not be a null allele. 36
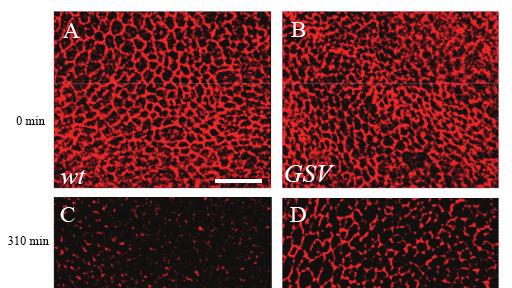
37 similar intensity compared to that in wild-type, the 65 kd band completely disappeared (Fig. 2.2). This result indicates that the 65 kd protein is the RN-Tre gene product. Since the 62 kd band could be found in both wild-type and GSV mutants, it likely represents a nonspecific protein that cross-reacted with the anti-rn-tre antibody. This result thus confirms that the GSV mutation disrupted the production of RN-Tre protein. Future preparation of anti-n-terminal antibody may allow us to detect the truncated protein and determine whether the GSV mutation is a null allele Analysis of membrane trafficking defects in RN-Tre mutants To directly address the question if RN-Tre is involved in regulating endocytosis, I performed pulse-chase experiments to determine the effect of loss of RN-Tre on membrane trafficking of Notch, a cell surface receptor, in the third-instar wing imaginal disc. Due to the availability of antibodies against the extracellular region of Notch, membrane trafficking of Notch has been used to study the in vivo control of endocytosis by a number of groups (e.g. (Lu and Bilder, 2005; Wilkin et al., 2004)). In my experiments, I performed pulse chase under both non-permeable and permeable conditions to examine the clearance and trafficking of membrane Notch in live third-instar wing imaginal discs Pulse-chase experiments under non-permeable condition To examine the effect of RN-Tre mutations on the clearance of Notch from membrane due to endocytosis, I used the mouse anti-notch C458.2H antibody that recognizes the extracellular region of Notch in pulse-chase experiments under non-permeable condition. Dissected third-instar wing discs were cultured in medium and pulse-labeled by anti-notch antibodies, which only labeled Notch on the cell surface. After pulse-labeling, wing discs were chased with medium that did not contain antibodies. After the chase period, wing discs were fixed under non-permeable condition (without Triton-X-100). Notch that remained on the cell surface was visualized by incubating wing discs with anti-mouse secondary antibodies (Fig. 2.3). In wild type, after a 310 min chase, most of cell surface Notch 37
38 38
39 Figure 2.3. Schematic drawing of pulse chase of Notch in the wing disc without Triton. 39
40 disappeared, and Notch that remained on the surface showed discontinuous puncta-like staining pattern, suggesting that most of labeled Notch were internalized (Fig. 2.4C). Compared to that in wild type, however, significant higher level of surface Notch was observed in GSV mutants after a 310 min chase, and most cells still showed continuous membrane Notch staining pattern in the cell periphery (6 out of 9 discs examined; Fig. 2.4D). This result suggests that RN-Tre is required for the clearance of Notch from cell surface and may do so by promoting Notch endocytosis Pulse-chase experiments under permeable condition To further confirm that loss of RN-Tre increased the internalization of Notch, I also performed pulse chase experiments under permeable condition (Fig. 2.5). After pulselabeling live third-instar wing discs with the mouse anti-notch C458.2H antibody, wing disces were chased with medium lacking antibodies and then fixed under permeable condition (with Triton-X-100). Labeled Notch on the surface and internalized into the cytoplasm was then visualized by incubating wing discs with anti-mouse secondary antibodies. In wild type, after a 120 min chase, I found that significant portion of Notch was found in cytoplasmic puncta (Fig. 2.6C). In GSV mutants, however, most of Notch still remained on the surface after a 120 min chase (Fig. 2.6D). Thus, the results from pulse-chase experiments under both permeable and nonpermeable conditions suggest that RN-Tre is required for regulating the turn over of membrane Notch by promoting its endocytosis. 2.3 RNA interference analysis Since expression of transgenic RN-Tre caused a gain-of-function phenotype, I did not perform rescue experiments to confirm that the Notch endocytosis phenotype was due to loss of RN-Tre. Instead, I performed RNA interference (RNAi) experiments to examine the effects of knocking down the expression of RN-Tre on Notch endocytosis. 40
41 41
42 Figure 2.4. Pulse chase of Notch in the wing disc without Triton at 0 min (A and B) and 310 min (C and D). Anti-Notch antibody was used at 1:30 dilution. A and C, wild type. B and D, GSV. At 0 min, Notch staining was associated with membrane in both wild type (A) and GSV mutants (B). At 310 min, Notch staining was significantly reduced in wildtype wing discs (n=10) (C). (D) In GSV mutant discs (n=9), most still showed strong membrane Notch staining. Three independent experiments were performed. The data was quantified by measuring the ratio of retained Notch staining intensity over time. In wild type, 8±3% of Notch staining was retained after 310 min, which was significantly different from that (i.e. 21.7±5.9%) of GSV mutants (p<0.001). Scale bar: 10 μm. 42
43 43
44 Figure 2.5. Schematic drawing of pulse chase of Notch in the wing disc with Triton. 44
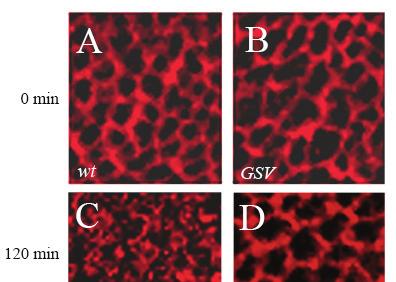
45 45
46 Figure 2.6. Pulse chase of Notch in the wing disc with Triton at 0 min (A and B) and 120 min (C and D). Anti-Notch antibody was used at 1:30 dilution. A and C, wild type. B and D, GSV. At 0 min, Notch staining was largely associated with membrane. At 120 min, cells in wild-type discs (C) showed loss of membrane staining and the accumulation of vesiclelike staining (n=9). In GSV mutant cells (n=7 discs)(d), Notch staining was still largely associated with membrane. Three independent experiments were performed. Scale bar: 5 μm. 46
47 UAS-RN-Tre RNAi line was obtained from VDRC. The Gal4 driver Apterous-Gal4 (AP- GAL4) was used to drive the expression of UAS-RN-Tre-RNAi. Apterous-Gal4 (AP- GAL4) specifically turns on the expression of UAS-RN-Tre-RNAi only in the dorsal region of third-instar wing discs, and thus knocking down the expression of RN-Tre in the dorsal region. Pulse-chase experiments were performed similarly to examine the effect of knocking down RN-Tre expression on Notch endocytosis. In this case, Notch endocytosis in the dorsal region with decreased RN-Tre expression was compared to that in the ventral region as well as that in wild-type. In wild type, after a 310 min chase, the level of labeled membrane Notch was significantly reduced in both dorsal and ventral regions of wing discs (Fig. 2.7C). There was no significant difference in the level of labeled membrane Notch between dorsal and ventral regions. By contrast, most of labeled Notch was found to be still associated with the cell periphery in the dorsal region of wing discs expressing UAS-RN- Tre-RNAi after a 310 min chase (Fig. 2.7D), whereas a lower level of labeled membrane Notch was observed in the ventral region (Fig. 2.7D). In summary, the results from RNAi experiments are consistent with loss-of-function analysis, and support that RN-Tre functions to promote Notch endocytosis. 2.4 Investigating the potential genetic interaction between RN-Tre and Notch The results from above pulse-chase experiments suggest a role for RN-Tre to regulate the steady-state level of membrane Notch by promoting Notch endocytosis. To further test this, I performed genetic experiments to examine if RN-Tre interacts with Notch to regulate the wing development. Previous studies showed that reducing the dosage of Notch by 50% induced a dominant wing phenotype (i.e. notching wing margin) (Sakata et al., 2004). Genetic modulation of this haploinsufficient Notch wing phenotype has been used by several groups to test the potential role of candidate genes in regulating the steadystate level of Notch (e.g. (Maitra et al., 2006; Sakata et al., 2004)). If RN-Tre promtes Notch 47
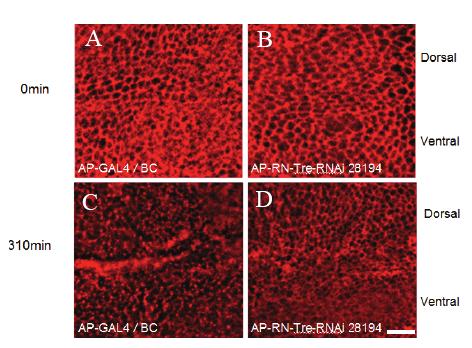
48 48
49 Figure 2.7. Pulse chase of Notch in the wing disc with Triton at 0 min (A and B) and 310 min (C and D). Anti-Notch antibody was used at 1:30 dilution. A and C, AP-GAL4/Bc control. B and D, wing discs expressing UAS-RN-Tre-RNAi transgene under control of AP-GAL4. At 0 min, Notch staining was largely associated with membrane. At 310 min, like wild type, cells in AP-GAL4/Bc Discs (C) showed loss of membrane staining and the accumulation of vesicle-like staining (n=10). In RN-Tre-RNAi-expressed cells in the dorsal region (n=7 discs) (D), Notch staining was still largely associated with membrane. Three independent experiments were performed. The data was quantified by counting the percentage of cells in which the cell periphery still retained continuous Notch staining. In AP-GAL4/Bc, only 7.1±5.7% cells showed continuous Notch membrane staining after 310 min, while 69.8±10% RN-Tre-RNAi cells still showed continuous Notch membrane staining. P< Scale bar: 10 μm. 49
50 endocytosis, one would predict that reducing the dosage of RN-Tre should increase the steady-state level of Notch in flies that are heterozygous for a Notch null mutation, and thus suppress the notching wing margin phenotype. To test this, I performed genetics crosses to reduce the dosage of wild-type RN-Tre by 50% in Notch heterozygous flies. In Notch heterozygous flies carrying two copies of wild-type RN-Tre, all adult wings displayed the notching wing margin phenotype (n>20; Fig. 2.8A). Interestingly, when the dosage of RN-Tre was reduced by 50%, most adult wings in Notch heterozygotes displayed normal wing margin (~60%, n=35; Fig. 2.8B). This result thus supports that RN-Tre functions to reduce the steady-state level of Notch, and is consistent with a role for RN-Tre to promote Notch endocytosis as suggested by previous pulse-chase experiments (see Fig. 2.4 and 2.6). 2.5 Investigating the potential genetic interaction between RN-Tre and Rab5 Mammalian RN-Tre has been shown to possess GAP activity and negatively regulate the activity of Rab5 (Haas et al., 2005; Lanzetti et al., 2000). To determine the functional relationship between RN-Tre and Rab5 in Drosophila, I performed genetic experiments to examine the potential interaction between RN-Tre and Rab5 in the fly compound eye. I used the longgmr-gal driver to overexpress UAS-RN-tre in the fly eye. This eye-specific driver is more specific to photoreceptor cell than the regular GMR-GAL4 driver (Wernet et al., 2003)(see previous section). Overexpression of UAS-RN-tre under control of the longgmr-gal4 driver caused a rough eye phenotype (Fig. 2.9; 100%, n>30 eyes). We reasoned that if this rough eye phenotype is due to mis-regulation of the activity of Rab5 protein by overexpression of RN-tre, reducing the dosage of Rab5 would modify the rough eye phenotype. To test this, I examined the effect of reducing the 50
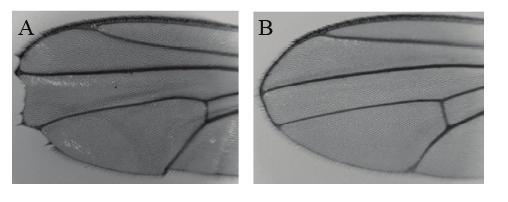
51 51
52 Figure 2.8. Reducing the dosage of RN-Tre suppressed the notching wing margin phenotype caused by Notch haploinsufficiency. (A) In flies heterozygous for a Notch null mutation (i.e. N 54L9 /+), all adult wings showed the notching wing margin phenotype (n>20). Genotype: N 54L9 /+. (B) When the dosage of RN-Tre was reduced by 50% in Notch heterozygotes, the notching wing margin phenotype was suppressed (~60%, n=35). Genotype: N 54L9 /+; RN-Tre GSV /+. 52
53 dosage of Rab5 by 50% on the RN-tre-induced eye phenotype. I found that reducing the dosage of Rab5 significantly enhanced the RN-tre-induced rough eye phenotype (52%, n=76 eyes, Fig. 2.9B), whereas no such enhancement was observed when the dosage of Rab6 was reduced (0%, n=24 eyes, Fig. 2.9C). This result indicates that overexpression of RN-tre negatively regulates the activity of Rab5 in the fly eye. Since Rab41 mutant is not available, I could not test if Rab41 also interacts with RN-tre in the fly eye. 2.6 Materials and Methods Western blot analysis Lysates were prepared from 10 wild-type and 10 RN-tre GSV mutant fly heads. After homogenizing the heads in 100ul 1x loading dye, the samples were boiled for 10min and then centrifuged. The supernatant was loaded on SDS-PAGE, underwent electrophoresis and then transferred onto the nitrocellulose membrane. The membrane was blocked with 5% skim milk in TBS at room temperature for 1hour, and then washed with 3X TTBS. The membrane was then incubated with anti-rn-tre (1:1000) in TTBS overnight. After wash 3 times with TTBS, the membrane was incubated with anti-rabbit secondary antibody (1:3000) at room temperature for 2 hours, washed three times with TTBS, and then followed by color development Pulse Chase experiments Third-instar larval wing discs were dissected in 1% FBS in Schneider s medium on ice. After quick rinse in above medium, the discs were incubated with medium (1% FBS in Schneider s medium with 1ug/ml 20-OH ecdysome) containing anti-notch 53
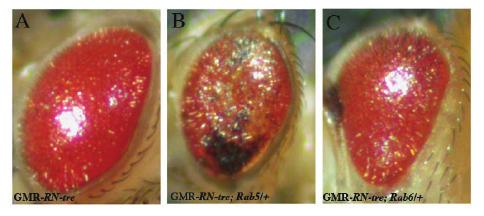
54 54
55 Figure 2.9. Rab5 mutations enhanced the RN-tre overexpression eye phenotype. (A) Ovexpression of RN-tre under control of the longgmr-gal4 driver caused a mild rough eye phenotype. The eye appeared glassy and smooth. Genotype: UAS-RN-tre/+; longgmr-gal4/+. (B) Reducing the dosage of Rab5 by 50% enhanced the RN-tre overexpression eye phenotype. The eye became flat and displayed a necrotic phenotype. Genotype: UAS-RN-tre/Rab5 2 ; longgmr-gal4/+. (C) Reducing the dosage of Rab6 did not enhance the RN-tre overexpression eye phenotype. Genotype: UAS-RN-tre/Rab6 D23D ; longgmr-gal4/+. 55
56 antibody (1:30 dilution) in on ice for 30min. The discs were then washed three times with 1% FBS in Schneider s medium on ice, 10min each time. Samples were chased for different period of time (0min, 120min, and 310min) by incubation with above medium lacking Notch antibody on ice. The discs were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) at room temperature for 30min, blocked with 0.1% Triton in 1% normal goat serum (NGS) in PBS (i.e. under -permeable condition) or 1% NGS in PBS (i.e. under non-permeable condition) at room temperature for 30min. The discs were then incubated with Texas-red-or FITC-conjugated goat anti-mouse secondary antibody (1:200 dilution) in PBT (0.5% Triton-X-100 in PBS) (under permeable condition) or in PBS (under non-permeable condition) at 4 C overnight. The discs were washed four times with PBT and once with PBS, 10min each, and incubated with SlowFade Gold antifade reagent (Invitrogen) at room temperature for 20 min before mounting on glass glides for observation Genetics GMR-GAL4 was provided by the Bloomington Drosophila stock center. Rab5 2, FRT 40 was provided by D.Bilder. UAS-RN-Tre-RNAi was provided by Vienna Drosophila RNAi Centre (VDRC). Rab6 D23D was provided by A. Guichet. AP-GAL4 was obtained from D. van Meyel. The full-length RN-Tre cdna (LD38355) was subcloned into the puast vector. The resulting construct was used to generate UAS-RN-Tre transgenic lines (BestGene Inc., CA). Regional-specific RNAi knock down was performed by crossing flies carrying AP-GAL4 with flies carrying UAS-RN-Tre RNAi. The dosage of RN-Tre was reduced by 50% in flies heterozygous for a null Notch mutation by crossing 56
57 female flies N 54L9 /FM6 with male flies w; RN-Tre GSV. The dosage of Rab5 or Rab6 was reduced by 50% in flies overexpressing RN-tre by crossing female flies yw; Cyo/UAS- RN-tre; long-gmr-gal4/tm3, Tb with male flies w; Cyo/Rab5 2, or yw; Cyo/ Rab6 D23D, respectively. 57
58 CHAPTER 3 DISCUSSION 58
59 The vertebrate RN-tre has been identified as a RabGAP for more than one decade (Matoskova et al., 1996). However, the function of RN-Tre remains highly controversial. While in vitro studies demonstrated that RN-Tre is capable of promoting the GTPase activity of Rab5 (Lanzetti et al., 2000) (Haas et al., 2005), the in vivo function of vertebrate RN-Tre remains unclear. Two groups reported that overexpression of RN-Tre blocked the endocytosis of ligand-induced internalization of EGFR in Hela cells and fibroblast cells, suggesting that RN-Tre functions to negatively regulate endocytosis by promoting the GTPase activity of Rab5 (Lanzetti et al., 2000) (Martinu et al., 2002). However, the study by Barr and coworkers presented evidence to support that RabGAP- 5, but not RN-Tre, negatively regulates Rab5 in ligand-induced EGFR endocytosis in Hela cells (Haas et al., 2005). They suggested that RN-Tre regulates the function of Rab41 or Rab30 in the Golgi complex. Since all above data were obtained from overexpression studies using cell culture system, it is unclear if the results truly reflect the function of RN-Tre in vivo. In my M. Sc project, I performed a combination of loss-of-function and gain-offunction analyses to determine the in vivo function of RN-Tre in the genetic model system Drosophila melanogaster. Several lines of evidence support the unexpected finding that RN-Tre promotes endocytosis of cell surface receptors. First, misexpression of RN-Tre in the Drosophila eye-imaginal disc decreased the cell surface level of Chaoptin and induced the accumulation of Chaoptin in vesicle-like structures. Second, mutations in the RN-Tre gene prevented the internalization of cell surface Notch in the developing Drosophila wing imaginal disc. Third, knocking down the expression of RN- 59
60 Tre in the wing disc also decreased the rate of Notch endocytosis. And fourth, reducing the dosage of RN-Tre suppressed the wing phenotype caused by Notch haploinsufficiency. I also examined the potential genetic interaction between RN-Tre and Rab5, which provides preliminary data that RN-Tre negatively regulates Rab5. Since previous reports showed that overexpression of mammalian RN-Tre blocked the endocytosis of cell surface receptors such as EGFR (Lanzetti et al., 2000) (Martinu et al., 2002), we examined if misexpression or overexpression of the fly RN-Tre also causes a similar phenotype. If the fly RN-Tre negatively regulates endocytosis, one prediction is that overexpression of the fly RN-Tre would block endocytosis and consequently the accumulation of receptor proteins on the cell surface. However, when RN-Tre was overexpressed in the fly third-instar eye-imaginal disc, we did not observe an increase in the surface level of cell adhesion molecule Chaoptin. Instead, we found that the surface level of Chaoptin was decreased (Chapter 2). We also observed the accumulation of Chaoptin in vesicle-like structures in both R-cell bodies and their axons (Chapter 2). While this result supports the involvement of RN-Tre in intracellular vesicular transport, it argues against the model in which RN-Tre negatively regulates endocytosis. There are several possible explanations for this gain-of-function result. First, RN-Tre may promote endocytosis of cell surface receptors leading to their clearance on the surface and accumulation within intracellular endosomes. Second, RN-Tre may negatively regulate the targeting of newly synthesized cell surface proteins towards the plasma membrane, thus overexpression of RN-Tre could block the transportation of cargoes containing cell surface receptors leading to the accumulation of those vesicles within the cytoplasm. And third, overexpression of RN-Tre may affect multiple vesicular 60
61 trafficking pathways, for instance, the recycling of Chaoptin and exocytosis. Elucidation of the nature of Chaoptin-positive vesicle-like structures will allow us to distinguish among those possibilities. To specifically address the question if RN-Tre is required for endocytosis, I performed loss-of-function analysis to examine if loss of RN-Tre affects the endocytosis of Notch in the third-instar wing imaginal disc. My results showed that mutation in the RN-Tre gene decreased the rate of the internalization of cell-surface Notch significantly. Similar results were obtained when the expression of endogenous RN-Tre was knocked down with RNA interference. These results thus argue convincingly that RN-Tre promotes Notch endocytosis in vivo and is required for regulating the steady-state level of Notch on the cell surface. In addition to Notch, RN-Tre may also promote the endocytosis of other cell surface receptors. That overexpression of RN-Tre could cause the accumulation of cell adhesiom molecules Chaoptin in intracellular vesicle-like structures in the third-instar eye imaginal disc is consistent with this view (Chapter 2). That reducing the dosage of RN-Tre significantly suppressed the notching wing phenotype caused by Notch haploinsufficiency, suggests a mechanism in which RN-Tremediated endocytosis of Notch allows the clearance of Notch from the cell surface to restrict the Notch signal. Such potentiation of Notch signaling by decreasing Notch endocytosis was not unprecedented. A previous study by another group showed that mutations in the Merlin gene, a cytoskeletal regulator, also decreased the endocytosis of Notch and thus increased the steady-state level of Notch on the cell surface (Maitra et al., 2006). Similarly, they found that reducing the dosage of Merlin suppressed the notching wing phenotype caused by Notch haploinsufficiency. 61
62 Although it is clear that RN-Tre is required for Notch endocytosis and interacts with Notch genetically (Chapter 2), homozygous RN-Tre mutants did not display any obvious developmental defects in the wing imaginal disc and visible defects in the adult wing (data not shown). This result indicates that while the increased steady-state level of Notch caused by reducing the dosage of RN-Tre is sufficient for rescuing the wing phenotype due to Notch haploinsufficiency, it is not sufficient to hyper-activate the Notch signaling pathway above a threshold level for causing a gain-of-function phenotype in wild-type background. This result is consistent with the previous report that overexpression of Notch did not cause any obvious phenotype in wild-type flies (Rebay et al., 1993). One possible explanation is the existence of other redundant genetic pathways that prevent the hyper-activation of the Notch pathway. In this context, it is notable that two previous studies demonstrated that the HECT domain-containing E3 ubiquitin ligase Nedd4 antagonizes Notch signaling by promoting degradation of Notch (Sakata et al., 2004; Wilkin et al., 2004). This Nedd4-mediated Notch degradation is suggested to be necessary for preventing the ligand-independent activation of Notch in unstimulated cells. Previous studies showed that mammalian RN-Tre could promote the GTPase activity of Rab5 and thus negatively regulate the activity of Rab5 in vitro (Haas et al., 2005; Lanzetti et al., 2000). My genetic data showing that reducing the dosage of Rab5 enhanced the rough eye phenotype caused by overexpression of RN-Tre is consistent with that RN-Tre negatively regulates the activity of Rab5. Since Rab5 is the key regulator of endocytosis, this raises the question how negative regulation of Rab5 activity by RN-Tre contributes to the positive role of RN-Tre in promoting endocytosis. One 62
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins
 13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
CELB40060 Membrane Trafficking in Animal Cells. Prof. Jeremy C. Simpson. Lecture 2 COPII and export from the ER
 CELB40060 Membrane Trafficking in Animal Cells Prof. Jeremy C. Simpson Lecture 2 COPII and export from the ER Today s lecture... The COPII coat - localisation and subunits Formation of the COPII coat at
CELB40060 Membrane Trafficking in Animal Cells Prof. Jeremy C. Simpson Lecture 2 COPII and export from the ER Today s lecture... The COPII coat - localisation and subunits Formation of the COPII coat at
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
10/2/2015. Chapter 4. Determination and Differentiation. Neuroanatomical Diversity
 Chapter 4 Determination and Differentiation Neuroanatomical Diversity 1 Neurochemical diversity: another important aspect of neuronal fate Neurotransmitters and their receptors Excitatory Glutamate Acetylcholine
Chapter 4 Determination and Differentiation Neuroanatomical Diversity 1 Neurochemical diversity: another important aspect of neuronal fate Neurotransmitters and their receptors Excitatory Glutamate Acetylcholine
CHAPTER 3. Cell Structure and Genetic Control. Chapter 3 Outline
 CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
Multiple Choice Review- Eukaryotic Gene Expression
 Multiple Choice Review- Eukaryotic Gene Expression 1. Which of the following is the Central Dogma of cell biology? a. DNA Nucleic Acid Protein Amino Acid b. Prokaryote Bacteria - Eukaryote c. Atom Molecule
Multiple Choice Review- Eukaryotic Gene Expression 1. Which of the following is the Central Dogma of cell biology? a. DNA Nucleic Acid Protein Amino Acid b. Prokaryote Bacteria - Eukaryote c. Atom Molecule
Honors Biology Reading Guide Chapter 11
 Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
7.06 Problem Set
 7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
A complementation test would be done by crossing the haploid strains and scoring the phenotype in the diploids.
 Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
Cell Biology Review. The key components of cells that concern us are as follows: 1. Nucleus
 Cell Biology Review Development involves the collective behavior and activities of cells, working together in a coordinated manner to construct an organism. As such, the regulation of development is intimately
Cell Biology Review Development involves the collective behavior and activities of cells, working together in a coordinated manner to construct an organism. As such, the regulation of development is intimately
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Cellular Neuroanatomy I The Prototypical Neuron: Soma. Reading: BCP Chapter 2
 Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Pattern formation: Wingless on the move Robert Howes and Sarah Bray
 R222 Dispatch Pattern formation: Wingless on the move Robert Howes and Sarah Bray Wingless is a key morphogen in Drosophila. Although it is evident that Wingless acts at a distance from its site of synthesis,
R222 Dispatch Pattern formation: Wingless on the move Robert Howes and Sarah Bray Wingless is a key morphogen in Drosophila. Although it is evident that Wingless acts at a distance from its site of synthesis,
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Supplementary Information
 Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
!"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%%
*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%%") !"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%% !"#$%&'(")*++*%,*'-&'./%/,*#01#%-2)#3&)/% 4'(")*++*% % %5"0)%-2)#3&) %%% %67'2#72'*%%%%%%%%%%%%%%%%%%%%%%%4'(")0/./% % 8$+&'&,+"/7 % %,$&7&/9)7$*/0/%%%%%%%%%%
!"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%% !"#$%&'(")*++*%,*'-&'./%/,*#01#%-2)#3&)/% 4'(")*++*% % %5"0)%-2)#3&) %%% %67'2#72'*%%%%%%%%%%%%%%%%%%%%%%%4'(")0/./% % 8$+&'&,+"/7 % %,$&7&/9)7$*/0/%%%%%%%%%%
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
The neuron as a secretory cell
 The neuron as a secretory cell EXOCYTOSIS ENDOCYTOSIS The secretory pathway. Transport and sorting of proteins in the secretory pathway occur as they pass through the Golgi complex before reaching the
The neuron as a secretory cell EXOCYTOSIS ENDOCYTOSIS The secretory pathway. Transport and sorting of proteins in the secretory pathway occur as they pass through the Golgi complex before reaching the
AP Biology Gene Regulation and Development Review
 AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016
 Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Protein Sorting, Intracellular Trafficking, and Vesicular Transport
 Protein Sorting, Intracellular Trafficking, and Vesicular Transport Noemi Polgar, Ph.D. Department of Anatomy, Biochemistry and Physiology Email: polgar@hawaii.edu Phone: 692-1422 Outline Part 1- Trafficking
Protein Sorting, Intracellular Trafficking, and Vesicular Transport Noemi Polgar, Ph.D. Department of Anatomy, Biochemistry and Physiology Email: polgar@hawaii.edu Phone: 692-1422 Outline Part 1- Trafficking
Nature Neuroscience: doi: /nn.2662
 Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice Supplementary Figure 2.
 Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice. Percentage of marginal zone B cells in the spleen of wild-type mice (+/+), mice homozygous for cpm or pri
Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice. Percentage of marginal zone B cells in the spleen of wild-type mice (+/+), mice homozygous for cpm or pri
Activation of a receptor. Assembly of the complex
 Activation of a receptor ligand inactive, monomeric active, dimeric When activated by growth factor binding, the growth factor receptor tyrosine kinase phosphorylates the neighboring receptor. Assembly
Activation of a receptor ligand inactive, monomeric active, dimeric When activated by growth factor binding, the growth factor receptor tyrosine kinase phosphorylates the neighboring receptor. Assembly
Richik N. Ghosh, Linnette Grove, and Oleg Lapets ASSAY and Drug Development Technologies 2004, 2:
 1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
S1 Gene ontology (GO) analysis of the network alignment results
 analysis of the network alignment results") 1 Supplementary Material for Effective comparative analysis of protein-protein interaction networks by measuring the steady-state network flow using a Markov model Hyundoo Jeong 1, Xiaoning Qian 1 and
1 Supplementary Material for Effective comparative analysis of protein-protein interaction networks by measuring the steady-state network flow using a Markov model Hyundoo Jeong 1, Xiaoning Qian 1 and
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p
 Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p.110-114 Arrangement of information in DNA----- requirements for RNA Common arrangement of protein-coding genes in prokaryotes=
Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p.110-114 Arrangement of information in DNA----- requirements for RNA Common arrangement of protein-coding genes in prokaryotes=
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
Molecular Cell Biology 5068 In Class Exam 1 September 30, Please print your name:
 Molecular Cell Biology 5068 In Class Exam 1 September 30, 2014 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 1 September 30, 2014 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
SUPPLEMENTARY INFORMATION
 med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
7.06 Cell Biology EXAM #3 April 21, 2005
 7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290
 PLoS Biol 4(9): e290") Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Transport between cytosol and nucleus
 of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes
 9 The Nucleus Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes Explain general structures of Nuclear Envelope, Nuclear Lamina, Nuclear Pore Complex Explain movement of proteins
9 The Nucleus Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes Explain general structures of Nuclear Envelope, Nuclear Lamina, Nuclear Pore Complex Explain movement of proteins
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Heather Currinn, Benjamin Guscott, Zita Balklava, Alice Rothnie and Thomas Wassmer*
 Online Resources APP controls the formation of PI(3,5)P 2 vesicles through its binding of the PIKfyve complex. Cellular and Molecular Life Sciences Heather Currinn, Benjamin Guscott, Zita Balklava, Alice
Online Resources APP controls the formation of PI(3,5)P 2 vesicles through its binding of the PIKfyve complex. Cellular and Molecular Life Sciences Heather Currinn, Benjamin Guscott, Zita Balklava, Alice
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
Drosophila wing. Temporal regulation of Apterous activity during development of the. Marco Milán and Stephen M. Cohen* SUMMARY
 Development 127, 3069-3078 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2547 3069 Temporal regulation of Apterous activity during development of the Drosophila wing Marco Milán
Development 127, 3069-3078 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2547 3069 Temporal regulation of Apterous activity during development of the Drosophila wing Marco Milán
BIS &003 Answers to Assigned Problems May 23, Week /18.6 How would you distinguish between an enhancer and a promoter?
 Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
BILD7: Problem Set. 2. What did Chargaff discover and why was this important?
 BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BE 159: Signal Transduction and Mechanics in Morphogenesis
 BE 159: Signal Transduction and Mechanics in Morphogenesis Justin Bois Caltech Winter, 2018 2018 Justin Bois. This work is licensed under a Creative Commons Attribution License CC-BY 4.0. 5 Delta-Notch
BE 159: Signal Transduction and Mechanics in Morphogenesis Justin Bois Caltech Winter, 2018 2018 Justin Bois. This work is licensed under a Creative Commons Attribution License CC-BY 4.0. 5 Delta-Notch
Morphogens in biological development: Drosophila example
 LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Biol403 - Receptor Serine/Threonine Kinases
 Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Supplementary Materials for
 www.sciencemag.org/cgi/content/full/science.1244624/dc1 Supplementary Materials for Cytoneme-Mediated Contact-Dependent Transport of the Drosophila Decapentaplegic Signaling Protein Sougata Roy, Hai Huang,
www.sciencemag.org/cgi/content/full/science.1244624/dc1 Supplementary Materials for Cytoneme-Mediated Contact-Dependent Transport of the Drosophila Decapentaplegic Signaling Protein Sougata Roy, Hai Huang,
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Name: TF: Section Time: LS1a ICE 5. Practice ICE Version B
 Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
ADAM FAMILY. ephrin A INTERAZIONE. Eph ADESIONE? PROTEOLISI ENDOCITOSI B A RISULTATO REPULSIONE. reverse. forward
 ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
7.06 Spring 2004 PS 6 KEY 1 of 14
 7.06 Spring 2004 PS 6 KEY 1 of 14 Problem Set 6. Question 1. You are working in a lab that studies hormones and hormone receptors. You are tasked with the job of characterizing a potentially new hormone
7.06 Spring 2004 PS 6 KEY 1 of 14 Problem Set 6. Question 1. You are working in a lab that studies hormones and hormone receptors. You are tasked with the job of characterizing a potentially new hormone
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Bypass and interaction suppressors; pathway analysis
 Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Importance of Protein sorting. A clue from plastid development
 Importance of Protein sorting Cell organization depend on sorting proteins to their right destination. Cell functions depend on sorting proteins to their right destination. Examples: A. Energy production
Importance of Protein sorting Cell organization depend on sorting proteins to their right destination. Cell functions depend on sorting proteins to their right destination. Examples: A. Energy production
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway
 Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
with%dr.%van%buskirk%%%
 with%dr.%van%buskirk%%% How$to$do$well?$ Before$class:$read$the$corresponding$chapter$ Come$to$class$ready$to$par9cipate$in$Top$Hat$ Don t$miss$an$exam!!!!!!!!!!!!!!!!!!!!!!!!!!$ But$I m$not$good$with$science
with%dr.%van%buskirk%%% How$to$do$well?$ Before$class:$read$the$corresponding$chapter$ Come$to$class$ready$to$par9cipate$in$Top$Hat$ Don t$miss$an$exam!!!!!!!!!!!!!!!!!!!!!!!!!!$ But$I m$not$good$with$science
THE CELL 3/15/15 HUMAN ANATOMY AND PHYSIOLOGY I THE CELLULAR BASIS OF LIFE
 HUMAN ANATOMY AND PHYSIOLOGY I Lecture: M 6-9:30 Randall Visitor Center Lab: W 6-9:30 Swatek Anatomy Center, Centennial Complex Required Text: Marieb 9 th edition Dr. Trevor Lohman DPT (949) 246-5357 tlohman@llu.edu
HUMAN ANATOMY AND PHYSIOLOGY I Lecture: M 6-9:30 Randall Visitor Center Lab: W 6-9:30 Swatek Anatomy Center, Centennial Complex Required Text: Marieb 9 th edition Dr. Trevor Lohman DPT (949) 246-5357 tlohman@llu.edu
5- Semaphorin-Plexin-Neuropilin
 5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
Chapter 18 Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Conclusions. The experimental studies presented in this thesis provide the first molecular insights
 C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain
 Development 127, 367-379 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV5342 367 Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain Wei Du
Development 127, 367-379 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV5342 367 Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain Wei Du
IN the Drosophila ventral nerve cord, there are 20
 Copyright Ó 2007 by the Genetics Society of America DOI: 10.1534/genetics.107.075085 Regulation of Axon Guidance by Slit and Netrin Signaling in the Drosophila Ventral Nerve Cord Krishna Moorthi Bhat,
Copyright Ó 2007 by the Genetics Society of America DOI: 10.1534/genetics.107.075085 Regulation of Axon Guidance by Slit and Netrin Signaling in the Drosophila Ventral Nerve Cord Krishna Moorthi Bhat,
Student Handout Fruit Fly Ethomics & Genomics
 Student Handout Fruit Fly Ethomics & Genomics Summary of Laboratory Exercise In this laboratory unit, students will connect behavioral phenotypes to their underlying genes and molecules in the model genetic
Student Handout Fruit Fly Ethomics & Genomics Summary of Laboratory Exercise In this laboratory unit, students will connect behavioral phenotypes to their underlying genes and molecules in the model genetic
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its
 The Wnt/ -catenin signalling pathway and its") Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
CAPE Biology Unit 1 Scheme of Work
 CAPE Biology Unit 1 Scheme of Work 2011-2012 Term 1 DATE SYLLABUS OBJECTIVES TEXT PAGES ASSIGNMENTS COMMENTS Orientation Introduction to CAPE Biology syllabus content and structure of the exam Week 05-09
CAPE Biology Unit 1 Scheme of Work 2011-2012 Term 1 DATE SYLLABUS OBJECTIVES TEXT PAGES ASSIGNMENTS COMMENTS Orientation Introduction to CAPE Biology syllabus content and structure of the exam Week 05-09
cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody
 Based on Monoclonal Anti-cGMP Antibody") (FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
(FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2647 Figure S1 Other Rab GTPases do not co-localize with the ER. a, Cos-7 cells cotransfected with an ER luminal marker (either KDEL-venus or mch-kdel) and mch-tagged human Rab5 (mch-rab5,
DOI: 10.1038/ncb2647 Figure S1 Other Rab GTPases do not co-localize with the ER. a, Cos-7 cells cotransfected with an ER luminal marker (either KDEL-venus or mch-kdel) and mch-tagged human Rab5 (mch-rab5,
Biological Process Term Enrichment
 Biological Process Term Enrichment cellular protein localization cellular macromolecule localization intracellular protein transport intracellular transport generation of precursor metabolites and energy
Biological Process Term Enrichment cellular protein localization cellular macromolecule localization intracellular protein transport intracellular transport generation of precursor metabolites and energy
Gene Control Mechanisms at Transcription and Translation Levels
 Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
Reading: Chapter 5, pp ; Reference chapter D, pp Problem set F
 Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
The Pax and large Maf families of genes in mammalian eye development
 The Pax and large Maf families of genes in mammalian eye development Vertebrate eye development is dependent on the coordinated action of thousands of genes. A specific group of over one hundred of regulatory
The Pax and large Maf families of genes in mammalian eye development Vertebrate eye development is dependent on the coordinated action of thousands of genes. A specific group of over one hundred of regulatory
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila
 Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A)
 M-phase nuclei are visualized by a-ph3 labeling in wild-type (A)") Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A) and mutant (B) 4 dpf retinae. The central retina of the
Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A) and mutant (B) 4 dpf retinae. The central retina of the
DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS. Scientific Background on the Nobel Prize in Medicine 2013
 DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS Scientific Background on the Nobel Prize in Medicine 2013 Daniela Scalet 6/12/2013 The Nobel Prize in Medicine
DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS Scientific Background on the Nobel Prize in Medicine 2013 Daniela Scalet 6/12/2013 The Nobel Prize in Medicine
RhoGAP assay kit (Cat. # BK105)
") RhoGAP assay kit (Cat. # BK105) The Ras superfamily of small GTPases (such as Ras, Rho, Rab, Arf and Ran proteins) serve as binary switches cycling between a GDP-bound OFF state and a GTP-bound ON state,
RhoGAP assay kit (Cat. # BK105) The Ras superfamily of small GTPases (such as Ras, Rho, Rab, Arf and Ran proteins) serve as binary switches cycling between a GDP-bound OFF state and a GTP-bound ON state,
Drosophila IAP1-mediated ubiquitylation controls activation of the initiator caspase DRONC independent of protein degradation
 University of Massachusetts Medical School escholarship@umms Molecular, Cell and Cancer Biology Publications Molecular, Cell and Cancer Biology 9-1-2011 Drosophila IAP1-mediated ubiquitylation controls
University of Massachusetts Medical School escholarship@umms Molecular, Cell and Cancer Biology Publications Molecular, Cell and Cancer Biology 9-1-2011 Drosophila IAP1-mediated ubiquitylation controls
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
S Phase Coupled E2f1 Destruction Ensures Homeostasis in Proliferating Tissues
 S Phase Coupled E2f1 Destruction Ensures Homeostasis in Proliferating Tissues Jean M. Davidson 1, Robert J. Duronio 1,2,3 * 1 Department of Biology, The University of North Carolina at Chapel Hill, Chapel
S Phase Coupled E2f1 Destruction Ensures Homeostasis in Proliferating Tissues Jean M. Davidson 1, Robert J. Duronio 1,2,3 * 1 Department of Biology, The University of North Carolina at Chapel Hill, Chapel
TBC1D9B functions as a GTPase-activating protein for Rab11a in polarized MDCK cells
 MBoC ARTICLE TBC1D9B functions as a GTPase-activating protein for Rab11a in polarized MDCK cells Luciana I. Gallo a, Yong Liao b, Wily G. Ruiz a, Dennis R. Clayton a, Min Li b,c, Yong-Jian Liu d, Yu Jiang
MBoC ARTICLE TBC1D9B functions as a GTPase-activating protein for Rab11a in polarized MDCK cells Luciana I. Gallo a, Yong Liao b, Wily G. Ruiz a, Dennis R. Clayton a, Min Li b,c, Yong-Jian Liu d, Yu Jiang
Designer Genes C Test
 Northern Regional: January 19 th, 2019 Designer Genes C Test Name(s): Team Name: School Name: Team Number: Rank: Score: Directions: You will have 50 minutes to complete the test. You may not write on the
Northern Regional: January 19 th, 2019 Designer Genes C Test Name(s): Team Name: School Name: Team Number: Rank: Score: Directions: You will have 50 minutes to complete the test. You may not write on the