Journal of Cell Science Accepted manuscript
|
|
|
- Deborah Annabelle Flowers
- 5 years ago
- Views:
Transcription
1 2013. Published by The Company of Biologists Ltd. Title: Direct Binding of TUBB3 with DCC Couples Netrin-1 Signaling to Intracellular Microtubule Dynamics in Axon Outgrowth and Guidance Running Title: Modulation of Microtubule Dynamics in Netrin Signaling Chao Qu, Trisha Dwyer, Qiangqiang Shao, Tao Yang, Huai Huang, and Guofa Liu Department of Biological Sciences, University of Toledo, M.S. 601, 2801 W. Bancroft St., Toledo, OH 43606, USA Author for correspondence: Guofa Liu, Department of Biological Sciences, University of Toledo, 2801 West Bancroft St., Toledo, OH 43606, USA. Phone: Total: 8 figures, 27 pages Conflict of Interest: The authors declare no competing financial interests. 1 JCS Advance Online Article. Posted on 2 May 2013
2 Summary The coupling of axon guidance cues, such as netrin-1, to microtubule (MT) dynamics is essential for growth cone navigation in the developing nervous system. However, whether axon guidance signaling regulates MT dynamics directly or indirectly is unclear. Here, we report that TUBB3, the most dynamic β tubulin isoform in neurons, directly interacts with the netrin receptor DCC, and that netrin-1 induces this interaction in primary neurons. TUBB3 colocalizes with DCC in the growth cones of primary neurons and MT dynamics is required for netrin-1-promoted association of TUBB3 with DCC. Netrin-1 not only increases cosedimentation of DCC with polymerized MT, but also promotes MT dynamics in the growth cone. Knocking down TUBB3 inhibits netrin-1-induced MT dynamics, axon outgrowth and attraction in vitro and causes defects in commissural axon projection in the embryo. These results indicate that TUBB3 directly links netrin signaling pathways to MT dynamics and plays an important role in guiding commissural axons in vivo. Introduction In the nervous system, temporal and spatial information processing relies on a variety of functional neural circuitries. The formation of neural circuits during brain development depends on the precise coordination of different guidance cues, their receptors and intracellular signal transduction cascades (Dent et al., 2011; Guan and Rao, 2003; Kolodkin and Tessier-Lavigne, 2011), which eventually converges on orchestrating the cytoskeleton dynamics to maneuver growth cone navigation (Kalil and Dent, 2005; Lowery and Van Vactor, 2009; Vitriol and Zheng, 2012). The coordination of actin filament and microtubule (MT) dynamics in neurons plays a crucial role in axon pathfinding in the developing nervous system (Buck and Zheng, 2002; Dent and Gertler, 2003; Dent et al., 2011; Lowery and Van Vactor, 2009; Vitriol and Zheng, 2012). However, whether MT dynamics are directly regulated by guidance cues and how intracellular signaling initiated by the activation of guidance receptors modulates MT dynamics are still unclear. Netrins, a family of canonical guidance cues, is a typical model for studying molecular mechanisms of axon outgrowth and guidance in the developing nervous system (Hedgecock et al., 1990; Ishii et al., 1992; Kennedy et al., 1994; Kolodziej et al., 1996; Tessier-Lavigne et al., 1988). As bifunctional guidance cues, netrins act as either chemoattractants or chemorepellents 2
3 for different cell types (Alcantara et al., 2000; Colamarino and Tessier-Lavigne, 1995). The mammalian receptors of netrins are Deleted in Colorectal Cancer (DCC) (Fazeli et al., 1997; Keino-Masu et al., 1996), neogenin (Keeling et al., 1997; Keino-Masu et al., 1996), Uncoordinated-5 (UNC5) (Ackerman et al., 1997; Leonardo et al., 1997), and Down Syndrome Cell Adhesion Molecule (DSCAM) (Liu et al., 2009; Ly et al., 2008). The phenotypic defects in DCC-deficient mouse embryos are similar to those in netrin-1 -/- mice embryos with aberrant commissural axon pathfinding and, ultimately, the inability of most commissural axons to reach the floor plate and cross the midline (Fazeli et al., 1997; Serafini et al., 1994). DCC collaborates with DSCAM mediating netrin-1-induced axon outgrowth and chemoattraction (Liu et al., 2009; Ly et al., 2008), whereas interaction of UNC-5 with DCC or DSCAM mediates repulsion (Finger et al., 2002; Hong et al., 1999; Keino-Masu et al., 1996; Kolodziej et al., 1996; Leonardo et al., 1997; Purohit et al., 2012), indicating the coordination of different netrin receptors is crucial for their ability to mediate attraction or repulsion. Recent studies have shown that mutations in TUBB3, a neuronal β-tubulin isotype III, result in commissural axon malformation both in human patients and mouse models with dysgenesis of the corpus callosum, anterior commissure, and internal capsule, suggesting TUBB3 is required for axon guidance and neuronal development (Poirier et al., 2010; Tischfield et al., 2010). Here, we have found that DCC interacts directly with TUBB3, and netrin-1 induces this interaction depending on MT dynamics in primary neurons. TUBB3 is required for netrininduced axon outgrowth and pathfinding in the developing nervous system. These results demonstrate, to our knowledge for the first time, that guidance receptors directly couple MT dynamics in axon guidance. Results TUBB3 interacts with the netrin receptor DCC To examine the potential interaction of TUBB3 with DCC in the developing brain, primary neurons from the cerebral cortex of embryonic day 15 (E15) mice were isolated, cultured and treated with conditioned media from control HEK cells or HEK cells stably expressing human netrin-1. The cell extracts were immunoprecipitated with anti-dcc antibody before probing the blots with the anti-tubb3 antibody. Anti-DCC antibody immunoprecipitated TUBB3 (Fig. 1A). In contrast, as predicted, TUBB3 was not present when primary antibody was 3
4 omitted (see the IgG control lanes in Fig. 1A). Treatment with netrin-1 conditioned media increased the interaction of TUBB3 with DCC (Fig. 1A, quantification in Fig. 1C, upper panel). Netrins play a crucial role in promoting commissural neuron axon outgrowth and pathfinding in the developing spinal cord (Kennedy et al., 1994; Serafini et al., 1994). To determine whether endogenous TUBB3 interacted with DCC in spinal cord neurons, dissociated cells from the dorsal half of E13 mouse spinal cords were cultured and treated with netrin-1 conditioned media. The immunoprecipitation results indicated that DCC interacted with TUBB3 and netrin-1 dramatically increased this interaction (Fig. 1B, quantification in Fig. 1C, lower panel). Netrin-1 increased the interaction of TUBB3 with DCC within 5 min and the induction lasted up to 20 min after netrin-1 stimulation (Fig. 1D). Similar time-courses of netrin-1 stimulation were observed in E15 cortical neurons (data not shown). To confirm that netrin-1 was directly responsible for increasing the interaction of TUBB3 with DCC, we tested purified netrin-1 from two sources: chicken netrin-1 from a commercial source and human netrin-1 purified from a stable HEK cell line established in our lab. Both sources of netrin-1 increased the interaction of TUBB3 with DCC in E15 primary cortical neurons in a dose-dependent manner (Fig. 1E). In contrast, DCC could not interact with TUBB1 and TUBB2, two other β-tubulin subunits, in the absence or presence of purified netrin-1 (Fig. 1F). These results strongly suggest that TUBB3 specifically associates with DCC in primary neurons. To determine whether TUBB3 could interact directly with DCC, cdnas expressing HAtagged human TUBB3 were co-transfected with DCC-Myc into HEK293 cells that do not share the same signaling machinery with primary neurons. Anti-HA antibody immunoprecipitated DCC (Fig. 1G), detected by probing the blots with anti-myc antibody. This result suggests that TUBB3 may directly interact with DCC. To further characterize this direct interaction, the isolated intracellular domain of DCC (DCC-ICD-MBP) was purified and incubated with purified TUBB3. TUBB3 appeared to interact directly with the intracellular domains of DCC (Fig. 1H). DCC has three conserved intracellular domains (P1, P2, P3) that are required for netrin-mediated signal transduction. To identify the DCC domain(s) that interact with TUBB3, TUBB3 was cotransfected with three distinct DCC truncation mutants, P1 (deletion of residues ) or P2 (deletion of residues ) or P3 (deletion of residues ) into HEK293 cells. TUBB3 bound to a truncated DCC lacking the P1 domain, but did not bind to truncated 4
5 DCCs lacking either the P2 or P3 domains (Fig. 1I). These results suggest that the intracellular P2 and P3 domains of DCC may be required for its interaction with TUBB3. Axon growth cones are highly motile sensory structures in developing neurons that respond to extracellular guidance cues. To determine whether TUBB3 is subcellularly colocalized with DCC in the growth cone of primary neurons, tissues from the E11 dorsal spinal cord and E15 cortex were dissociated and cultured. The localization of DCC and TUBB3 overlapped in the growth cones of E11 commissural neurons after 4-day culture (Fig. S1A-C). DCC colocalized with TUBB3 in the peripheral (P) region of growth cones, including lamellipodia and filopodia (Fig. S1A-C). Colocalization of TUBB3 with DCC was also observed in growth cones of E15 cortical neurons (Fig. S1D-F). MT dynamics modulates the interaction of TUBB3 with DCC The modulation of MT dynamics in growth cones plays a crucial role in axon guidance (Dent et al., 2011). To investigate whether MT dynamics is required for the binding of TUBB3 with DCC, primary E15 cortical neurons were treated with paclitaxel (taxol) or nocodazole, drugs that disrupt MT dynamics. As expected based on the results described above, netrin-1 increased the interaction of endogenous TUBB3 with DCC (Fig. 2A-C). The netrin-1-induced interaction was inhibited by both the MT-stabilizing drug taxol and the MT-destabilizing drug nocodazole (Fig. 2A-C), indicating that MT dynamics is required for the netrin-1 dependent interaction of TUBB3 with DCC. To further determine whether monomeric or polymeric TUBB3 binds to DCC, a MT cosedimentation assay was performed on E15 cortical neuron cell lysates. Because MT in cell lysates becomes unstable in vitro and constantly depolymerizes under cold conditions, incubation of the lysate on ice induced MT depolymerization and yielded monomerized TUBB3 subunits in the soluble supernatant and polymerized TUBB3 in the pellet after centrifugation (Fig. 2D and 2F). Most of the endogenous DCC also remained in the supernatants with or without netrin-1 stimulation (Fig. 2D-E). In contrast, in the presence of taxol in cell lysate stabilizing MT against depolymerization by cold treatment, DCC cosedimented with polymerized MT with a large quantity of DCC and TUBB3 in the pellet (Fig. 2D and quantification in Fig. 2E-F). As predicted, in the presence of taxol, netrin-1 further increased DCC and TUBB3 in the pellets (Fig. 2D and quantification in Fig. 2E-2F). These results indicate that netrin-1 induces the interaction of endogenous DCC with polymerized 5
6 TUBB3 in primary neurons, suggesting that the interaction may be dependent on MT dynamics. To determine whether netrin-1 directly modulates MT dynamics, EB3-GFP, a marker of MT dynamics in live cells, was transfected into either E13 dorsal spinal cord neurons (Fig. S2, see movie 1-4 in supplementary material) or E15 cortical neurons (data not shown). Results from live cell imaging revealed that netrin-1 increased the percentage of moving EB3-GFP comets in the growth cone of primary spinal cord neurons (Fig. S2, see movie 1-2 in supplementary material) and cortical neurons (data not shown), and TUBB3 knockdown blocked this induction (Fig. S2C, see movie 3-4 in supplementary material, data not shown for cortical neurons). These data suggest that netrin-1 directly modulates MT dynamics via TUBB3. Tyrosine phosphorylation is required for netrin signaling and the post-translation modification (PTM) of MT subunits is crucial for intracellular MT organization and dynamics. To investigate whether tyrosine phosphorylation of TUBB3 is involved in netrin signaling, primary neurons from E13 spinal cords and E15 cortexes were dissociated and examined for the TUBB3 tyrosine phosphorylation in the presence of or absence of netrin-1 (Fig. 3A-C and Fig. S3A-B). Immunoprecipitation experiments demonstrated that the tyrosine phosphorylation of TUBB3 was increased by netrin-1 in a dose- and time-dependent manner (Fig. 3A-C and Fig. S3A). Netrin-1-induced TUBB3 tyrosine phosphorylation was inhibited by PP2, an inhibitor of the Src family kinases, but not PP3, an inactive control for PP2 (Fig. 3D and Fig. S3B). The netrin-1-dependent induction of the endogenous interaction of TUBB3 with DCC in E15 neurons was also inhibited by PP2 (Fig. 3E-F). These findings indicate that Src family kinases are required for the netrin-1-induced tyrosine phosphorylation of TUBB3 and the interaction of TUBB3 with DCC. To further examine the role of Src family kinases in the netrin-1-induced binding of endogenous DCC to dynamic TUBB3, these key proteins were cosedimented from lysates of dissociated E15 primary cortical neurons after netrin-1 stimulation (Fig. 3G-H). As expected, netrin-1 increased the amount of DCC in the MT-sedimented pellets (Fig. 3G-H) and PP2, not PP3, inhibited the netrin-1-induced cosedimentation of DCC and MTs (Fig. 3G-H). These results suggest that Src family kinases play an important role in regulating the dynamic interaction of TUBB3 with DCC in primary neurons. 6
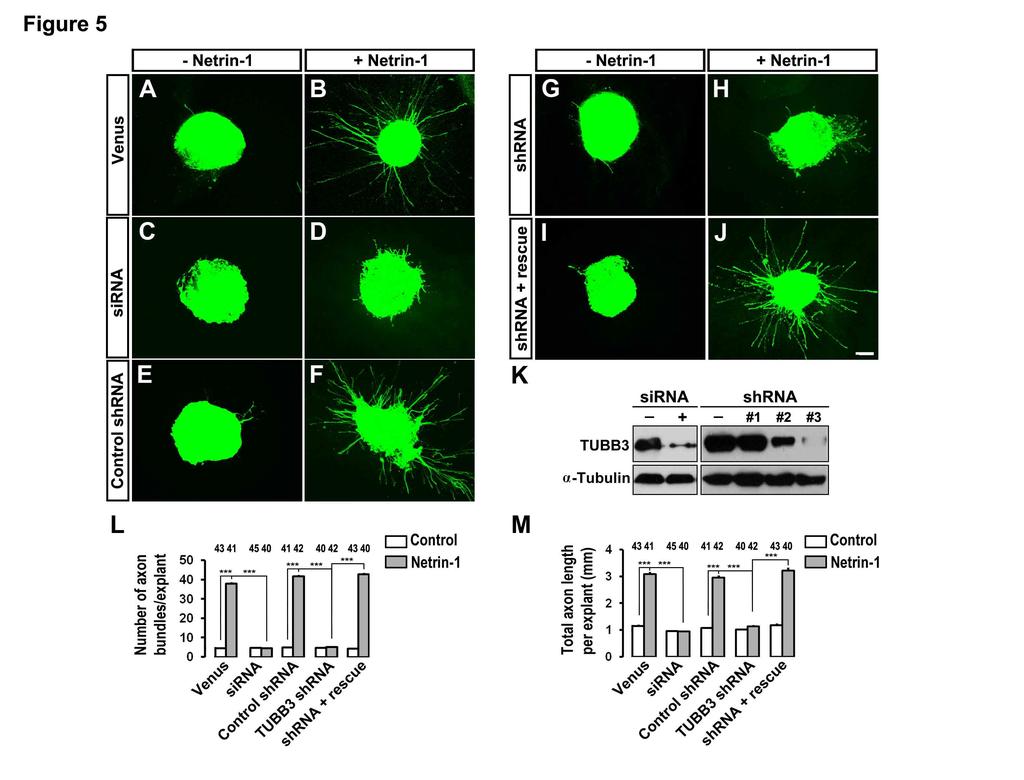
7 TUBB3 is required for netrin-1-induced neurite outgrowth To study the function of TUBB3 in netrin-1 signaling, a TUBB3 sirna pool (Dharmacon) or short hairpin-based RNA interference constructs (TUBB3 shrnas, gift from Dr. David L. Turner, University of Michigan, targeting a sequence common to mouse and chicken TUBB3) (Yu et al., 2002) were transfected into E15 mouse cortical neurons (Fig. 4K) and chicken neural tubes at stage (Fig. 5K). The sirna pool and one shrna construct significantly reduced the level of endogenous TUBB3 in these neurons (Fig. 4K and Fig. 5K) and were used in subsequent experiments. To examine whether TUBB3 is involved in netrin-1-induced neurite outgrowth, primary cortical neurons from E15 mice were dissociated and transfected with a construct expressing Venus yellow fluorescent protein (Venus YFP) only (Fig. 4A-B) or Venus YFP together with the TUBB3 sirna pool (Fig. 4C-D), the control shrna (Fig. 4E-F), the TUBB3 shrna (Fig. 4G- H) or the TUBB3 shrna plus the RNAi-resistant rescue constructs (Fig. 4I-J), respectively, as we have described previously (Li et al., 2008; Liu et al., 2007; Liu et al., 2009). These neurons were stimulated with netrin-1 and cultured for 20 h. In neurons transfected with the Venus YFP only, neurite outgrowth was stimulated by netrin-1 (Fig. 4A-B, quantification in Fig. 4L). As predicted, either the TUBB3 sirna pool (Fig. 4C-D) or TUBB3 shrna (Fig. 4G-H), but not control TUBB3 shrna (Fig. 4E-F), inhibited netrin-1-induced neurite outgrowth (quantification in Fig. 4L). Importantly, the expression of the wild-type human RNAi-resistant TUBB3 rescued netrin-1-promoted neurite outgrowth in neurons treated with TUBB3 sirna (Fig. 4I-J, quantification in Fig. 4L). To examine the role of TUBB3 in netrin-1-induced commissural axon outgrowth, we examined cultured chick dorsal spinal cord explants, as described previously (Liu et al., 2009). The TUBB3 sirna pool or shrna together with Venus YFP plasmids were introduced into chicken neural tubes at stage 12-15, and the YFP-labeled dorsal spinal cord segments were dissected at stage Axon outgrowth was quantified by measuring the numbers of axon bundles and the total axon length per explant. In explants transfected with Venus YFP only (Fig. 5A-B) or with Venus YFP plus control shrna (Fig. 5E-F), netrin-1 significantly induced axon outgrowth (quantification in Fig. 5L-M). TUBB3 sirna (Fig. 5C-D) or shrna (Fig. 5G-H), but not control shrna (Fig. 5E-F), significantly inhibited netrin-1-induced axon outgrowth (quantification in Fig. 5L-M). The expression of wild-type human TUBB3, which is resistant to 7
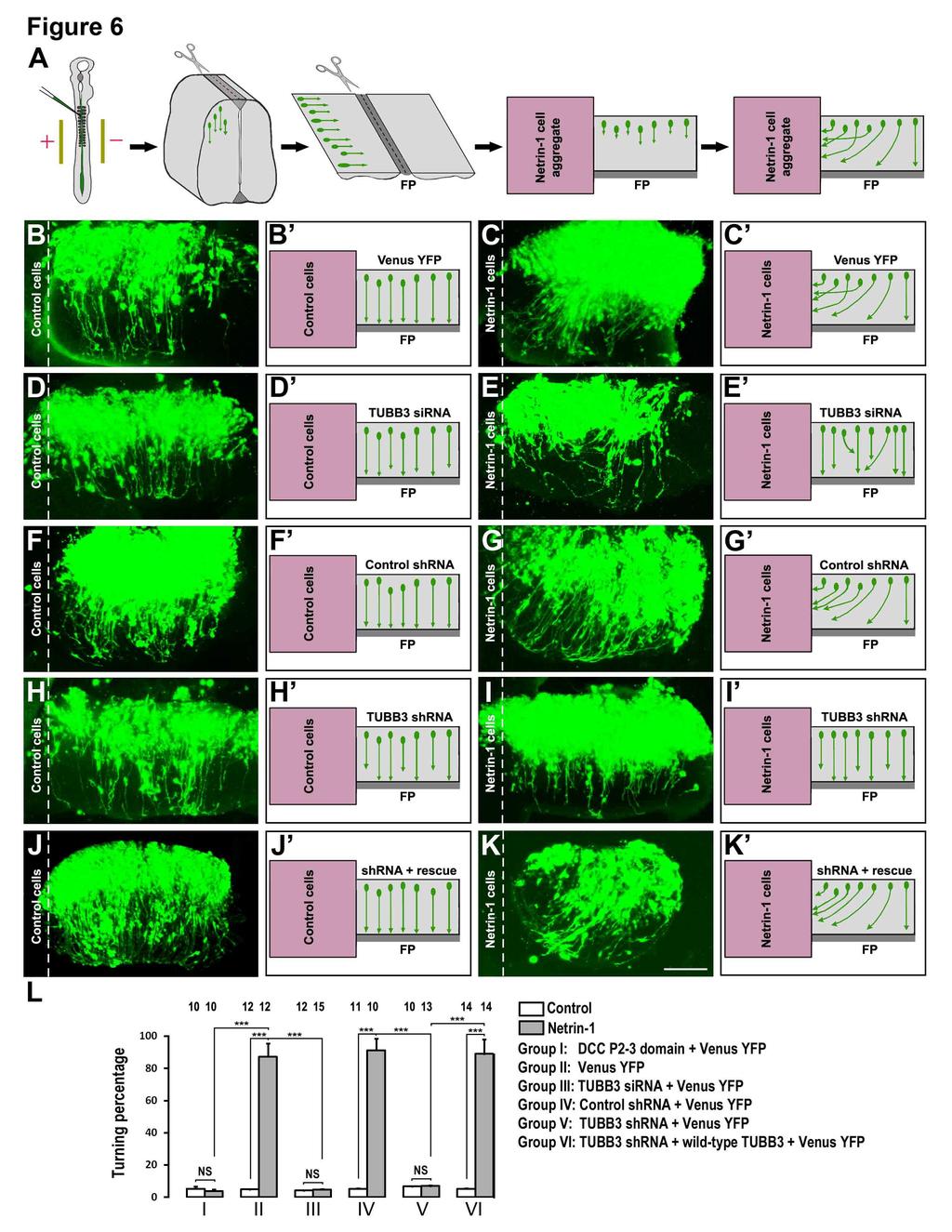
8 TUBB3 shrna, rescued the effect of TUBB3 knockdown on netrin-1-induced axon outgrowth (Fig. 5I-J, quantification in Fig. 5L-M). These results indicate that TUBB3 is required for netrin- 1-induced commissural axon outgrowth in vitro. Basal axon outgrowth of either dissociated E15 mouse cortical neurons or chick spinal cord commissural neurons was not affected by TUBB3 knockdown (Figs 4, 5 and Fig. S4). In addition, TrkB, a receptor of BDNF, could not interact with endogenous TUBB3 in the absence or presence of BDNF (Fig. S4A-B) and knockdown of TUBB3 did not affect BDNF-induced axon outgrowth of primary cortical neurons (Fig. S4C-I). These data suggest that TUBB3 is specifically involved in netrin-1-induced axon outgrowth. Furthermore, overexpression of DCC intracellular P2-3 domain not only inhibited the interaction of TUBB3 with full-length wild-type DCC in HEK293 cells (Fig. S5A-B), but also abolished netrin-1-induced neurite outgrowth of E15 cortical neurons (Fig. S5C-5G), suggesting that DCC- TUBB3 interaction is required for netrin-1-induced axon outgrowth. TUBB3 is required for axon attraction by netrin-1 Netrin-1 plays a crucial role in attracting the commissural axon projection in the developing neural tube (Kennedy et al., 1994; Serafini et al., 1996; Serafini et al., 1994; Tessier- Lavigne et al., 1988). To determine whether TUBB3 is required for commissural axon turning towards netrin-1, we used the open-book assay with commissural axons from chick embryos as illustrated in Fig. 6A (Li et al., 2008; Liu et al., 2004; Liu et al., 2007; Liu et al., 2009). The Venus YFP construct was electroporated either alone or with the TUBB3 sirna pool, TUBB3 control shrna, TUBB3 shrna, and TUBB3 shrna plus TUBB3 RNAi-resistant constructs, respectively, into the chick neural tube at stages The electroporated neural tube (visualized by green fluorescence) was then isolated and laid out as an open book at stage (Fig. 6A). A rectangle of neural tube explant containing the floor plate was prepared and cocultured with an aggregate of control HEK293 cells or HEK293 cells that stably secreted netrin-1 for 40 h (Fig. 6A). When the neural tube was electroporated with Venus YFP alone, 87.3 ± 8.2% of commissural axons turned towards netrin-1 secreting cell aggregates (Fig. 6C-C and quantification in Fig. 6L), whereas only 4.9 ± 0.1% of axons projected towards the control cell aggregates not secreting netrin-1 (Fig. 6B-B and Fig. 6L). TUBB3 sirna significantly inhibited commissural axons turning toward netrin-1: the turning percentage was reduced from 87.3 ± 8.2% 8
9 in the Venus YFP with netrin-1 group to 4.8 ± 0.2% in the TUBB3 sirna with netrin-1 group (Fig. 6C-C, 6E-E and Fig. 6L). To confirm the role of TUBB3 in netrin-1-induced axon turning, TUBB3 shrna or the control shrna were introduced into the neural tube by electroporation. When the control shrna was electroporated into the neural tube together with Venus YFP, 91.3 ± 7.3% of commissural axons turned towards netrin-1 (Fig. 6F-G and Fig. 6L). In contrast, the co-transfection of TUBB3 shrna with Venus YFP significantly inhibited netrin-1-induced attraction with only 7.1 ± 0.3% of axons turning towards netrin-1 (Fig. 6H-I and Fig. 6L). To further determine the specificity of shrna knockdown, TUBB3 shrna was electroporated with plasmids encoding a wild-type human RNAi-resistant TUBB3 into the chick neural tube. The wild-type RNAi-resistant TUBB3 transgene rescued the netrin-1 dependent phenotype with 89.1 ± 9.0% of axons turning towards netrin-1 (Fig. 6J-L). In addition, expression of DCC P2-3 domain also inhibited netrin-1-induced axon turning (Fig. S5H-I, quantification in Fig. 6L). Together, these results support the hypothesis that TUBB3 is specifically required for netrin-1- induced commissural axon attraction. TUBB3 is required for spinal commissural axon projection in vivo The results discussed above have shown that TUBB3 plays an important role in the in vitro functions of netrin-1 in neurite outgrowth and axon attraction. To determine the in vivo role of TUBB3, the effects of TUBB3 sirna and TUBB3 shrna were examined on commissural axon projection in the developing chicken spinal cord (Fig. 7) (Li et al., 2008; Liu et al., 2007; Liu et al., 2009). Venus YFP was introduced by electroporation into the neural tube of stage 12 chick embryos in ovo and the embryos were allowed to develop until stages 23 (Fig. 7A). The YFP-labeled lumbosacral segments of the spinal cord were isolated and laid out as an open book (Fig. 7A). By stage 23, 81.1 ± 3.3% of the commissural axons expressing Venus YFP alone reached the floor plate (Fig. 7B, quantification in Fig. 7G) (Li et al., 2008; Liu et al., 2007; Liu et al., 2009). In contrast, only 15.5 ± 2.6% of the commissural axons transfected with the TUBB3 sirna pool reached the floor plate (Fig. 7C and Fig. 7G). Similarly, TUBB3 shrna also significantly inhibited the projection of commissural axons towards the floor plate, while TUBB3 control shrna had no effect on the projection of commissural axons (Fig. 7D-E and Fig. 7G). The percentage of the YFP-labeled commissural axons per embryo reaching the floor plate was decreased from 79.1 ± 5.3% in the control shrna group to 24.3 ± 1.3% in the TUBB3 9
10 shrna group. The effect of TUBB3 shrna on commissural axon projection was reversed by co-transfecting the wild-type RNAi-resistant human TUBB3 plasmid (Fig. 7F-G) with 82.6 ± 4.7% of the YFP-labeled commissural axons per embryo reaching the floor plate. Although the open-book preparation demonstrated obvious defects in commissural axon projection in vivo, it was difficult to effectively assess effects on axon turning. To examine whether knockdown of TUBB3 disrupted commissural axon pathfinding in addition to inhibiting axon extension in vivo, either Venus YFP alone or Venus YFP with the TUBB3 sirna pool were electroporated into chick spinal cord and transverse sections of the chick spinal cord at stage 23 were prepared (Fig. 8A-C). In addition to the inhibition of axon extension, some commissural axons transfected with TUBB3 sirnas were misguided (Fig. 8C and quantification in Fig. 8G-I) instead of projecting normally towards the floor plate (Fig. 8B and Fig. 8G-I). These phenotypes were further confirmed by expressing Venus YFP and the TUBB3 shrna in commissural axons that also exhibited shortened and misguided axons in vivo (Fig. 8E and Fig. 8G-I). In contrast, the expression of the TUBB3 control shrna had no effect on commissural axons projection (Fig. 8D and Fig. 8G-I). As predicted, the expression of the wild-type RNAiresistant human TUBB3 rescued the effects of TUBB3 RNAi knock-down on commissural axon extension and turning (Fig. 8F-I). These results indicate that TUBB3 is required for both the projection and pathfinding of commissural axons in vivo in the developing spinal cord. Discussion TUBB3 is the most dynamic β tubulin isotype (Katsetos et al., 2003) and mutations in TUBB3 result in neurological disorders associated with abnormal neuronal migration, differentiation and axon guidance (Poirier et al., 2010; Tischfield et al., 2010). Our results here indicate that TUBB3 interacts directly with DCC and is required for netrin-1-induced axon outgrowth and turning in the developing nervous system. These results not only untangle the role of TUBB3 in netrin signaling, but also provide a working model for the direct involvement of MT dynamics in axon guidance. Direct coupling of netrin/dcc signaling to MT dynamics The neuronal cytoskeleton plays an essential role in axon outgrowth and pathfinding. Although most of the research has focused on the role of actin dynamics in growth cone protrusion, recent studies suggest that MTs also play an instructive role in growth cone steering 10
11 (Buck and Zheng, 2002; Dent et al., 2004; Dent et al., 2011; Suter and Forscher, 2000; Tanaka and Sabry, 1995). However, whether MT dynamics are directly regulated by guidance cues is still unclear. In this study, we have found that TUBB3, the neuronal β tubulin isotype III, colocalizes with DCC in the peripheral region, including both lamellipodia and filopodia, of the growth cones of developing commissural and cortical neurons (Fig. S1). TUBB3 interacts directly with DCC and that netrin-1 induces these interactions both in vitro and in vivo (Fig. 1). The netrin-1-induced interaction of TUBB3 with DCC is abolished by the disruption of MT dynamics either with taxol or nocodazole, suggesting MT dynamics are required for these interactions (Fig. 2). DCC cosediments with stabilized MTs and netrin-1 increases the ratio of DCC in the pellet versus the supernatant fraction (Fig. 2). Netrin-1 stimulation also increases the ratio of polymerized TUBB3 in the pellet. Live cell imaging of moving EB3 comets reveals that netrin-1 directly modulates MT dynamics in the growth cones of both primary E13 dorsal spinal cord (Fig. S2, see movie 1-4 in supplemental material) and E15 cortical neurons (data not shown). More importantly, our functional data indicate that TUBB3 is required for netrin-1- induced MT dynamics (Fig. S2, see movie 1-4 in supplemental material) as well as axon outgrowth and guidance both in vitro and in vivo (Fig. 4-8). In addition, DCC intracellular P2-3 domain interacts with TUBB3 (Fig. 1I, Fig. S5A-B) and this domain is required for netrin-1- induced neurite outgrowth and attraction (Fig. S5C-I) (Gitai et al., 2003; Stein and Tessier- Lavigne, 2001; Stein et al., 2001). These results lead to a generalizable model suggesting netrin- 1 signaling directly regulates MT dynamics through coupling its receptor DCC to TUBB3. In this model, in response to netrin-1, dynamic MTs are captured by DCC in the growth cone, stabilizing filopodia against retraction and promoting axon outgrowth and turning. Previous studies on the role of MTs dynamics in the regulation of growth cone turning also support this model. For instance, 1) dynamic MTs became oriented and stabilized preferentially in the direction of the growth cone turn (Tanaka and Kirschner, 1995), 2) the local stabilization of MTs in one side of the growth cone caused the growth cone to turn towards that side (Buck and Zheng, 2002), 3) the local disruption of MT stabilization on one side of a growth cone was sufficient to induce growth cone turning away from that side (Buck and Zheng, 2002), 4) the application of low concentration of taxol enhanced axon outgrowth in vitro and in vivo via MT stabilization, promoting their polymerization at plus ends (Sengottuvel et al., 2011). These studies suggest that 11
12 intrinsically polarized MT dynamics in the growth cone may be directly involved in axon guidance. TUBB3 is an essential downstream component coupling netrin-1 signaling to MT dynamics and axon outgrowth/guidance The spectrum of TUBB3-mutation phenotypes includes malformations of oculomotor nerves, the corpus callosum, anterior commissural, cortical spinal tracts and neuronal migration defects, suggesting that TUBB3 is required for axon guidance in the developing brain (Poirier et al., 2010; Tischfield et al., 2010). However, it is unclear why TUBB3 mutations are only associated with specific axon projection defects, such as those involved in commissural axon guidance, when it is widely expressed in all neurons in the developing nervous system. Although mutations in TUBB3 result in the perturbation of MT dynamics (Poirier et al., 2010; Tischfield et al., 2010), the specific role of TUBB3 in axon guidance in the developing nervous system is unknown. In this study, we propose that TUBB3 plays an essential role in netrin-1 signaling, involved in netrin-1-promoted axon outgrowth and projection in the developing nervous system. We have described a functional role of TUBB3 in netrin signaling both in vitro and in vivo. Netrin-1 increased neurite growth from primary cortical neurons and chick dorsal spinal cord explants (Fig. 4-5), as reported in previous studies (Li et al., 2008; Liu et al., 2004; Liu et al., 2007; Liu et al., 2009). Both a TUBB3 sirna pool and a specific TUBB3 shrna inhibited neurite outgrowth induced by netrin-1 (Fig. 4-5). In addition, TUBB3 knockdown by these same sirna pool or shrna also inhibited netrin-1-induced axon attraction in the chick open-book turning assay (Fig. 6). These results demonstrate that TUBB3 is involved in netrin-induced axon outgrowth and attraction in vitro. TUBB3 is also required for spinal commissural axon projection in vivo, as indicated by in ovo electroporation studies with chick spinal cords (Fig. 7-8). Together, these studies strongly suggest that TUBB3 plays an essential role in netrin-1-mediated axon outgrowth and guidance in the developing nervous system. TUBB3 mutations primarily affect MT function in a dominant fashion through altering heterodimer incorporation, MT stability, motor protein trafficking and kinesin-mt interactions (Tischfield et al., 2010). In future studies, it will be interesting to determine whether these TUBB3 mutations can affect the netrin-1-dependent interaction of TUBB3 with DCC. Although DCC could not interact with TUBB1 and TUBB2, heterozygous missense mutations in TUBA1A and TUBB2B share certain phenotypic similarities in brain malformations, such as dysgenesis of 12
13 the corpus callosum, basal ganglia dysmorphisms and neuronal migration defects, suggesting that these tubulin isotypes may have important overlapping functions (Jaglin et al., 2009; Keays et al., 2007; Poirier et al., 2010; Tischfield et al., 2010). It remains to be determined whether TUBA1A and TUBB2B can associate with other netrin receptors, such as DSCAM and UNC5C, in netrin signaling. Src family kinases are required for coupling netrin signaling to MT dynamics. Src family kinases are involved in netrin/dcc signaling (Li et al., 2004; Liu et al., 2004; Liu et al., 2007; Liu et al., 2009; Meriane et al., 2004). Netrin-1 can stimulate the tyrosine phosphorylation of TUBB3 by Src family kinases and this TUBB3 phosphorylation appears to be required for the subsequent interaction of TUBB3 with DCC and modulation of MT dynamics. For example, the inhibition of Src kinases by PP2, a Src family kinase inhibitor, 1) decreases netrin-1-induced TUBB3 phosphorylation (Fig. 3D and Fig. S3B), 2) decreases the netrin-1- stimulated interaction of DCC with TUBB3 in primary neurons (Fig. 3E-F) and 3) blocks netrin- 1-induced cosedimentation of DCC with polymerized MT (Fig. 3G-H). Together, these results suggest that Src family kinases are essential in regulating MT dynamics in netrin-1 signaling and may function as a key signaling component of DCC/TUBB3 complex. Indeed, Src family kinases play an important role in growth cone steering via regulating MT dynamics (Suter et al., 2004). Therefore, these results suggest a model in which DCC serves as a signaling platform for recruitment of a multiprotein complex, including TUBB3, Src family kinases and other key signaling molecules to modulate MT dynamics in netrin-1-induced axon outgrowth and turning. In addition to Src family kinases, many other signaling molecules have been identified in netrin/dcc signaling (Guan and Rao, 2003; Kolodkin and Tessier-Lavigne, 2011; Sun et al., 2011) including cyclic nucleotides, phospholipase C, phosphoinositol-3-kinase, mitogenactivated protein kinases, TRIO, DOCK180, transient receptor potential channels, calcium, myosin-x, PAK1 and Enabled/vasodilator-stimulated phosphoprotein. Whether these signaling molecules may also be involved in directly regulating MT dynamics through coupling netrin receptors to TUBB3 remains to be investigated. It has long been recognized that the different signal transduction cascades initiated by extracellular guidance cues converge on the cytoskeleton to manipulate growth cone behavior (Buck and Zheng, 2002; Dent and Gertler, 2003; Dent et al., 2011; Lowery and Van Vactor, 2009; Vitriol and Zheng, 2012). The coordination of dynamic MTs and actin filaments in growth 13
14 cones is necessary for proper axon outgrowth and guidance (Buck and Zheng, 2002; Dent and Gertler, 2003; Dent et al., 2011; Lowery and Van Vactor, 2009; Vitriol and Zheng, 2012). Although actin polymerization is not necessary for neurite outgrowth, the disruption of actin dynamics by cytochalasin D blocks growth cone turning induced by local taxol application, suggesting that the actin cytoskeleton plays an essential role in MT stabilization in growth cone turning (Buck and Zheng, 2002; Marsh and Letourneau, 1984). It will be interesting to investigate whether the interaction of DCC with TUBB3 will be regulated by actin dynamics, such as polymerization-depolymerization, treadmilling and retrograde flow in response to netrin signaling. Materials & Methods Materials. The following antibodies were used: rabbit anti-flag, mouse anti-phospho-tyrosine antibody 4G10 (anti-py) and rabbit anti-tubb3 (Abcam, Cambridge, MA, USA); rabbit anti- MBP and rabbit anti-hemagglutinin (HA) (Santa Cruz, CA, USA); mouse anti-dcc (BD Biosciences, San Jose, CA, USA ); mouse anti-tubb3 (Covance, Princeton, New Jersey, USA); mouse anti-gst (Cell Signaling, Danvers, MA, USA ); Alexa Fluor 488 goat anti-mouse IgG and Alexa Fluor 647 goat anti-rabbit IgG (Invitrogen, Grand Island, NY, USA); mouse anti- Myc, and mouse functional blocking anti-dcc (Calbiochem, Rockland, MA, USA). Taxol and nocodazole were obtained from MP Biochemicals (Solon, OH, USA). PP2 and PP3 were from Calbiochem. Purified TUBB3 was purchased from Origene (Rockville, MD, USA) and TUBB3- GST from Abnova (Walnut, CA, USA). Plasmids encoding TUBB3, DCC, DCC- P1 ( ), P2 ( ), and P3 ( ) have been described previously (Li et al., 2004). The sequence-verified DCC intracellular P2-3 domain was subcloned into pt-rex -Dest 30 vector via Gateway technology (Invitrogen). TUBB3 ON-TARGETplus SMARTpool was obtained from Dharmacon (Waltham, MA, USA). The targeted sequences of control shrna and DCC shrna are: AATGCATCTCTGCAAGAGGTA and CATCCGATGTGCGACTGTA, respectively. The target sequence was inserted into pavu6+27 between Sal I site and Xba I site. TUBB3 shrna and control shrna are gifts from Dr. David L. Turner (Yu et al., 2002), EB3- GFP constructs are from Dr. Niels Galjart and TUBB3-V5 are from Dr. Elizabeth C Engle. For TUBB3 RNAi rescue experiments, an RNAi-resistant construct was created by introducing seven silent point mutations in the target sequences. 14
15 Netrin-1 protein was either obtained from R&D (Minneapolis, MN, USA) or purified with anti-myc tag affinity matrix from the conditioned media of HEK cells stably secreting netrin-1. The control was made by sham-purification from the conditioned media from HEK cells that had not be transfected with a cdna expressing the Myc-tagged netrin-1. Recombinant DCC intracellular domain tagged with maltose-binding protein (DCC-ICD-MBP) was produced from BL21 competent E. coli and purified using amylose resin (New England Biolabs, Ipswich, MA, USA). The fusion protein was eluted with maltose and analyzed by SDS-PAGE, followed by Coomassie blue staining. Primary neuron cultures and nucleofection. The dissociated primary neuron culture and nucleofection procedures were performed as described previously (Liu et al., 2004; Liu et al., 2007; Liu et al., 2009). For examining the effect of RNAi knock-down, dissociated primary neurons were cultured on PLL-coated dishes for 2 days after nucleofection and cell lysates then analyzed by Western blotting. For examining neurite outgrowth, cortical neurons were left settling on the coverslips for 2 h and then cultured in DMEM with B27 (Invitrogen) and penicillin/streptomycin at 37ºC with 5% CO 2 for 20 h with purified netrin-1 (250 ng/ml) or the sham-purified control. Cells were then fixed with 4% pre-warmed paraformaldehyde (PFA) for 20 min and stained with the Alexa Fluor 555 phalloidin (Molecular Probes, Grand Island, NY, USA). Nuclei were visualized with Hoechst dye or DAPI (Invitrogen). Images were taken under a fluorescent microscope (Olympus IX81, Pittsburgh, PA, USA). The longest and total neurite length was examined using NIH Image J program. Immunoprecipitation and Western analysis. HEK293 cells were transfected with the PEI method and cultured 48 h after transfection. For netrin-1 stimulation, primary neurons and transfected HEK293 cells were starved for 8 h in serum-free DMEM media and followed by incubation with purified netrin-1 protein (500 ng/ml) or sham purified control up to 30 min. For immunoprecipitation, primary neurons from E15 mouse cortex, E13 dorsal spinal cord and HEK293 cells were lysed as described previously (Li et al., 2008; Liu et al., 2004; Liu et al., 2007; Liu et al., 2009). Cell lysates were incubated with specific antibodies for 2 h before protein A/G-agarose beads (Santa Cruz Biotechnology) were added. For the immunoblotting, protein extracts were separated with 7.5% SDS-polyacrylamide gel electrophoresis and Western blots were visualized with the enhanced chemiluminescence kit (Fisher, Pittsburgh, PA, USA). 15
16 Microtubule cosedimentation assay. E15 mouse cortical neurons were dissociated and cultured as previously described (Li et al., 2008; Liu et al., 2004; Liu et al., 2007; Liu et al., 2009). Primary neurons were stimulated by purified netrin-1 (500 ng/ml) or sham-purified control for 20 min. To detect the effects of Src family kinases on MT dynamics, PP2 (5 nm) or PP3 (5 nm) was added 6 h before netrin-1 stimulation. Primary neurons were lysed in the MLB buffer and cell lysates centrifuged at 100,000 g for 1 h at 4 C. The supernatant was incubated with 40 µm taxol or DMSO in PEMG buffer (100 mm PIPES, 1 mm EGTA, 1 mm MgSO4, 1 mm GTP, ph 6.8) at room temperature for 30 min. MTs were pelleted by centrifugation through a 10% sucrose cushion at 50,000 g for 30 min at 4 C. The pellet and supernatant fractions were collected separately, and the pellet was resuspended with tubulin buffer (50 mm HEPES, 1 mm MgCl2, 1 mm EGTA, 10% glycerol, 150 mm KCl, 40 µm taxol, 1 mm GTP, 5 mm Mg-ATP, 1 mm PMSF, 1 protease inhibitor mixture). Proteins in the supernatant and the pellet were separated by SDS-PAGE and analyzed by Western blotting. Immunocytochemistry. Dissociated neurons from E15 mouse cortexes and E11 dorsal spinal cords were fixed either in pre-warmed 4% PFA in 1 stabilization buffer (127 mm NaCl, 5 mm KCl, 1.1 mm NaH 2 PO 4, 0.4 mm KH 2 PO 4, 2 mm MgCl 2, 5.5 mm Glucose, 1mM EGTA, 10 mm Pipes) or ice-cold methanol and permeabilized with 0.5% Triton X-100 for 15 min. Cells were blocked with 0.25% BSA and 0.1% Triton in PBS at room temperature for 30 min and then incubated with primary antibody solution containing mouse anti-dcc (1:100) and rabbit anti- TUBB3 (1:300) antibodies overnight at 4 C. Neurons were incubated with secondary antibodies (anti-mouse-488 and anti-rabbit-647) and mounted onto glass slides with Fluorogel (Electron Microscopy Sciences). Images of growth cones were taken sequentially using a confocal microscope (Olympus IX71 Fluoview, Pittsburgh, PA, USA) with the same exposure settings. Chick spinal cord explant culture and analysis of axon outgrowth. Fertilized White Leghorn chicken eggs were incubated and chicken embryos staged as described previously (Liu et al., 2004; Liu et al., 2007; Liu et al., 2009). At stage 12-15, plasmids or sirnas plus Venus YFP were injected into the neural tube of chicken embryos and the in ovo electroporation was performed with the following program: 25 V, 5 ms, 5 pulses (BTX ECM830, Holliston, MA, USA) (Liu et al., 2004; Liu et al., 2007; Liu et al., 2009). Embryos at stage were collected and examined under a fluorescent microscope. The dorsal one-third of the spinal cord labeled with YFP fluorescence was isolated, trimmed to 200 µm in size, and transferred into the mixed 16
17 gel (3:2:1 collagen: matrigel: medium) as described previously (Liu et al., 2009). After gel polymerization, explants were cultured in DMEM with B27 (Invitrogen) and penicillin/streptomycin at 37ºC with 5% CO 2 overnight with purified netrin-1 (250 ng/ml) or the sham-purified control. Explants were fixed with 4% PFA in 1 PBS, and the images of axons with YFP fluorescence were obtained under a confocal microscope (Olympus IX70). The numbers of axons and total axon length per explant were measured using the NIH Image J software. Chicken commissural axon turning assay. The in ovo electroporation procedures were essentially done as described above (Liu et al., 2004; Liu et al., 2007; Liu et al., 2009; Qu et al., 2013). Embryos were harvested at stages and samples showing YFP fluorescence were isolated under the fluorescence microscope. The spinal cord was opened at the roof plate (openbook preparation) and explants from the half spinal cord with YFP fluorescence were co-cultured with an aggregate of control or netrin-1 secreting HEK cells as described previously (Liu et al., 2004; Liu et al., 2007; Liu et al., 2009). Axons with the turning angle towards the cell aggregate more than 5 were counted as attraction and the percentage of attractive axons was calculated from the numbers of fluorescent axons turning towards the HEK cell aggregate divided by the total numbers of fluorescent axons within 300 µm of the HEK cell aggregates. Images were acquired under an Olympus IX70 confocal microscope. Chicken commissural axon projection in vivo. Chick spinal cords with YFP fluorescence were collected until stage 23 after electroporation as described previously (Li et al., 2008; Liu et al., 2007; Liu et al., 2009; Qu et al., 2013). The lumbosacral region of the spinal cord was isolated and the open-book preparation of the spinal cord was performed by opening the roof plate. After fixation, the tissues in the open-book preparation were mounted in Gel/Mount (Biomeda, Pittsburgh, MA, USA) for analysis of commissural axon projection in vivo. Images were taken under the confocal microscope. The percentage of axons reaching the floor plate was calculated from the numbers of fluorescent axons crossing the ipsilateral floor plate divided by the total numbers of fluorescent axons within 100 µm from the floor plate (Li et al., 2008; Liu et al., 2007; Liu et al., 2009). To examine commissural axon pathfinding in vivo, the lumbosacral segments of the chick spinal cords expressing YFP at stage 23 were isolated and transverse sections of 200 µm were cut by a vibrotome. The spinal cord slices were mounted in Gel/Mount for confocal fluorescent 17
18 microscopy. The percentage of axons reaching the midline was quantified from the numbers of fluorescent axons arriving at or crossing over the midline divided by the total numbers of fluorescent axons within 200 µm from the floor plate. The axon distance from the midline was calculated from the average distance of fluorescent axons away from the midline. Acknowledgements: We thank Dr. Andrea L. Kalinoski for help with confocal imaging, Dr. David L. Turner for TUBB3 shrna constructs, Dr. Niels Galjart for EB3-GFP constructs, Dr. Elizabeth C Engle for TUBB3-V5 constructs,dr. Bruce Bamber for image analysis, Dr. Richard Komuniecki for comments on the manuscript, and the Whitehall Foundation and NIH for support. 18
19 References Ackerman, S.L., Kozak, L.P., Przyborski, S.A., Rund, L.A., Boyer, B.B., and Knowles, B.B. (1997). The mouse rostral cerebellar malformation gene encodes an UNC-5-like protein. Nature 386, Alcantara, S., Ruiz, M., De Castro, F., Soriano, E., and Sotelo, C. (2000). Netrin 1 acts as an attractive or as a repulsive cue for distinct migrating neurons during the development of the cerebellar system. Development 127, Buck, K.B., and Zheng, J.Q. (2002). Growth cone turning induced by direct local modification of microtubule dynamics. J Neurosci 22, Colamarino, S.A., and Tessier-Lavigne, M. (1995). The axonal chemoattractant netrin-1 is also a chemorepellent for trochlear motor axons. Cell 81, Dent, E.W., Barnes, A.M., Tang, F., and Kalil, K. (2004). Netrin-1 and semaphorin 3A promote or inhibit cortical axon branching, respectively, by reorganization of the cytoskeleton. J Neurosci 24, Dent, E.W., and Gertler, F.B. (2003). Cytoskeletal dynamics and transport in growth cone motility and axon guidance. Neuron 40, Dent, E.W., Gupton, S.L., and Gertler, F.B. (2011). The growth cone cytoskeleton in axon outgrowth and guidance. Cold Spring Harb Perspect Biol 3. Fazeli, A., Dickinson, S.L., Hermiston, M.L., Tighe, R.V., Steen, R.G., Small, C.G., Stoeckli, E.T., Keino-Masu, K., Masu, M., Rayburn, H., et al. (1997). Phenotype of mice lacking functional Deleted in colorectal cancer (Dcc) gene. Nature 386, Finger, J.H., Bronson, R.T., Harris, B., Johnson, K., Przyborski, S.A., and Ackerman, S.L. (2002). The netrin 1 receptors Unc5h3 and Dcc are necessary at multiple choice points for the guidance of corticospinal tract axons. J Neurosci 22, Gitai, Z., Yu, T.W., Lundquist, E.A., Tessier-Lavigne, M., and Bargmann, C.I. (2003). The netrin receptor UNC-40/DCC stimulates axon attraction and outgrowth through enabled and, in parallel, Rac and UNC-115/AbLIM. Neuron 37, Guan, K.L., and Rao, Y. (2003). Signalling mechanisms mediating neuronal responses to guidance cues. Nat Rev Neurosci 4,
20 Hedgecock, E.M., Culotti, J.G., and Hall, D.H. (1990). The unc-5, unc-6, and unc-40 genes guide circumferential migrations of pioneer axons and mesodermal cells on the epidermis in C. elegans. Neuron 4, Hong, K., Hinck, L., Nishiyama, M., Poo, M.M., Tessier-Lavigne, M., and Stein, E. (1999). A ligand-gated association between cytoplasmic domains of UNC5 and DCC family receptors converts netrin-induced growth cone attraction to repulsion. Cell 97, Ishii, N., Wadsworth, W.G., Stern, B.D., Culotti, J.G., and Hedgecock, E.M. (1992). UNC-6, a laminin-related protein, guides cell and pioneer axon migrations in C. elegans. Neuron 9, Jaglin, X.H., Poirier, K., Saillour, Y., Buhler, E., Tian, G., Bahi-Buisson, N., Fallet-Bianco, C., Phan-Dinh-Tuy, F., Kong, X.P., Bomont, P., et al. (2009). Mutations in the beta-tubulin gene TUBB2B result in asymmetrical polymicrogyria. Nat Genet 41, Kalil, K., and Dent, E.W. (2005). Touch and go: guidance cues signal to the growth cone cytoskeleton. Curr Opin Neurobiol 15, Katsetos, C.D., Legido, A., Perentes, E., and Mork, S.J. (2003). Class III beta-tubulin isotype: a key cytoskeletal protein at the crossroads of developmental neurobiology and tumor neuropathology. Journal of child neurology 18, ; discussion 867. Keays, D.A., Tian, G., Poirier, K., Huang, G.-J., Siebold, C., Cleak, J., Oliver, P.L., Fray, M., Harvey, R.J., Molnar, Z., et al. (2007). Mutations in alpha-tubulin cause abnormal neuronal migration in mice and lissencephaly in humans. Cell 128, Keeling, S.L., Gad, J.M., and Cooper, H.M. (1997). Mouse Neogenin, a DCC-like molecule, has four splice variants and is expressed widely in the adult mouse and during embryogenesis. Oncogene 15, Keino-Masu, K., Masu, M., Hinck, L., Leonardo, E.D., Chan, S.S., Culotti, J.G., and Tessier- Lavigne, M. (1996). Deleted in Colorectal Cancer (DCC) encodes a netrin receptor. Cell 87, Kennedy, T.E., Serafini, T., de la Torre, J.R., and Tessier-Lavigne, M. (1994). Netrins are diffusible chemotropic factors for commissural axons in the embryonic spinal cord. Cell 78, Kolodkin, A.L., and Tessier-Lavigne, M. (2011). Mechanisms and molecules of neuronal wiring: a primer. Cold Spring Harb Perspect Biol 3. 20
21 Kolodziej, P.A., Timpe, L.C., Mitchell, K.J., Fried, S.R., Goodman, C.S., Jan, L.Y., and Jan, Y.N. (1996). frazzled encodes a Drosophila member of the DCC immunoglobulin subfamily and is required for CNS and motor axon guidance. Cell 87, Leonardo, E.D., Hinck, L., Masu, M., Keino-Masu, K., Ackerman, S.L., and Tessier-Lavigne, M. (1997). Vertebrate homologues of C. elegans UNC-5 are candidate netrin receptors. Nature 386, Li, W., Lee, J., Vikis, H.G., Lee, S.H., Liu, G., Aurandt, J., Shen, T.L., Fearon, E.R., Guan, J.L., Han, M., et al. (2004). Activation of FAK and Src are receptor-proximal events required for netrin signaling. Nat Neurosci 7, Li, X., Gao, X., Liu, G., Xiong, W., Wu, J., and Rao, Y. (2008). Netrin signal transduction and the guanine nucleotide exchange factor DOCK180 in attractive signaling. Nat Neurosci 11, Liu, G., Beggs, H., Jurgensen, C., Park, H.T., Tang, H., Gorski, J., Jones, K.R., Reichardt, L.F., Wu, J., and Rao, Y. (2004). Netrin requires focal adhesion kinase and Src family kinases for axon outgrowth and attraction. Nat Neurosci 7, Liu, G., Li, W., Gao, X., Li, X., Jurgensen, C., Park, H.T., Shin, N.Y., Yu, J., He, M.L., Hanks, S.K., et al. (2007). p130cas is required for netrin signaling and commissural axon guidance. J Neurosci 27, Liu, G., Li, W., Wang, L., Kar, A., Guan, K.L., Rao, Y., and Wu, J.Y. (2009). DSCAM functions as a netrin receptor in commissural axon pathfinding. Proc Natl Acad Sci U S A 106, Lowery, L.A., and Van Vactor, D. (2009). The trip of the tip: understanding the growth cone machinery. Nat Rev Mol Cell Biol 10, Ly, A., Nikolaev, A., Suresh, G., Zheng, Y., Tessier-Lavigne, M., and Stein, E. (2008). DSCAM is a netrin receptor that collaborates with DCC in mediating turning responses to netrin-1. Cell 133, Marsh, L., and Letourneau, P.C. (1984). Growth of neurites without filopodial or lamellipodial activity in the presence of cytochalasin B. J Cell Biol 99, Meriane, M., Tcherkezian, J., Webber, C.A., Danek, E.I., Triki, I., McFarlane, S., Bloch-Gallego, E., and Lamarche-Vane, N. (2004). Phosphorylation of DCC by Fyn mediates Netrin-1 signaling in growth cone guidance. J Cell Biol 167,
22 Poirier, K., Saillour, Y., Bahi-Buisson, N., Jaglin, X.H., Fallet-Bianco, C., Nabbout, R., Castelnau-Ptakhine, L., Roubertie, A., Attie-Bitach, T., Desguerre, I., et al. (2010). Mutations in the neuronal ss-tubulin subunit TUBB3 result in malformation of cortical development and neuronal migration defects. Hum Mol Genet 19, Purohit, A.A., Li, W., Qu, C., Dwyer, T., Shao, Q., Guan, K.-L., and Liu, G. (2012). Down Syndrome Cell Adhesion Molecule (DSCAM) Associates with Uncoordinated-5C (UNC5C) in Netrin-1-mediated Growth Cone Collapse. The Journal of biological chemistry 287, Qu, C., Li, W., Shao, Q., Dwyer, T., Huang, H., Yang, T., and Liu, G. (2013). c-jun N-terminal Kinase 1 (JNK1) Is Required for Coordination of Netrin Signaling in Axon Guidance. The Journal of biological chemistry 288, Sengottuvel, V., Leibinger, M., Pfreimer, M., Andreadaki, A., and Fischer, D. (2011). Taxol facilitates axon regeneration in the mature CNS. The Journal of neuroscience : the official journal of the Society for Neuroscience 31, Serafini, T., Colamarino, S.A., Leonardo, E.D., Wang, H., Beddington, R., Skarnes, W.C., and Tessier-Lavigne, M. (1996). Netrin-1 is required for commissural axon guidance in the developing vertebrate nervous system. Cell 87, Serafini, T., Kennedy, T.E., Galko, M.J., Mirzayan, C., Jessell, T.M., and Tessier-Lavigne, M. (1994). The netrins define a family of axon outgrowth-promoting proteins homologous to C. elegans UNC-6. Cell 78, Stein, E., and Tessier-Lavigne, M. (2001). Hierarchical organization of guidance receptors: silencing of netrin attraction by slit through a Robo/DCC receptor complex. Science 291, Stein, E., Zou, Y., Poo, M., and Tessier-Lavigne, M. (2001). Binding of DCC by netrin-1 to mediate axon guidance independent of adenosine A2B receptor activation. Science 291, Sun, K.L., Correia, J.P., and Kennedy, T.E. (2011). Netrins: versatile extracellular cues with diverse functions. Development 138, Suter, D.M., and Forscher, P. (2000). Substrate-cytoskeletal coupling as a mechanism for the regulation of growth cone motility and guidance. J Neurobiol 44,
23 Suter, D.M., Schaefer, A.W., and Forscher, P. (2004). Microtubule dynamics are necessary for SRC family kinase-dependent growth cone steering. Curr Biol 14, Tanaka, E., and Kirschner, M.W. (1995). The role of microtubules in growth cone turning at substrate boundaries. J Cell Biol 128, Tanaka, E., and Sabry, J. (1995). Making the connection: cytoskeletal rearrangements during growth cone guidance. Cell 83, Tessier-Lavigne, M., Placzek, M., Lumsden, A.G., Dodd, J., and Jessell, T.M. (1988). Chemotropic guidance of developing axons in the mammalian central nervous system. Nature 336, Tischfield, M.A., Baris, H.N., Wu, C., Rudolph, G., Van Maldergem, L., He, W., Chan, W.M., Andrews, C., Demer, J.L., Robertson, R.L., et al. (2010). Human TUBB3 mutations perturb microtubule dynamics, kinesin interactions, and axon guidance. Cell 140, Vitriol, E.A., and Zheng, J.Q. (2012). Growth cone travel in space and time: the cellular ensemble of cytoskeleton, adhesion, and membrane. Neuron 73, Yu, J.-Y., DeRuiter, S.L., and Turner, D.L. (2002). RNA interference by expression of shortinterfering RNAs and hairpin RNAs in mammalian cells. Proceedings of the National Academy of Sciences of the United States of America 99, Figure legends Figure 1. Interaction of TUBB3 with DCC (A-B) Interaction of endogenous TUBB3 with DCC in E15 cortical (A) and E13 dorsal spinal cord (B) neurons. (C) Quantification of A and B from three independent experiments. * indicates p< 0.05 (two-tailed Student s t-test). (D) Netrin-1 increased the interaction of endogenous TUBB3 with DCC in a time-dependent manner. Lysates of dissociated neurons from E13 mouse spinal cords were immunoprecipitated with either anti-dcc and analyzed with anti-tubb3. (E) Netrin-1 increased the interaction of endogenous TUBB3 with DCC in a dose-dependent manner. E15 primary cortical neurons were treated with purified netrin-1 from 10, 50, to 200 ng/ml. (F) Endogenous DCC interacted with TUBB3, not TUBB1 and TUBB2, in E15 cortical neurons with or without netrin-1 treatment. Cell lysates of dissociated neurons from E15 mouse cortexes were immunoprecipitated with anti-dcc and followed by probing with anti-tubb1, anti- TUBB2 or anti-tubb3. (G) Interaction of TUBB3 with DCC in HEK293 cells. TUBB3-HA was 23
24 co-transfected with DCC-Myc into HEK293 cells. Anti-HA (TUBB3) precipitated DCC-Myc. (H) Direct interaction of TUBB3 with DCC. Purified TUBB3 was incubated with purified intracellular domain of DCC tagged with MBP in vitro. The anti-gst antibody was used to immunoprecipitate proteins and the blot was analyzed with anti-mbp. (I) P2 and P3 domains in DCC are required for the interaction of DCC and TUBB3. TUBB3-HA was co-transfected with different truncated DCCs (DCC- P1, P2 and P3) tagged with Myc in HEK293 cells. Figure 2. The induction of the interaction of TUBB3 with DCC by netrin-1 depends on MT dynamics. (A-C) Taxol and nocodazole (Noc) inhibited netrin-1-induced interactions of endogenous TUBB3 with DCC. E15 cortical neurons were treated with purified netrin-1 in the presence of different concentrations of taxol or nocodazole (1µM taxol and 3 µm nocodazole in A). (B) Quantification of (A) from three independent experiments showing relative binding of DCC and TUBB3. (D-F) E15 cortical neurons were stimulated with netrin-1 and the cosedimentation assay of cell lysates was performed in the absence or presence of taxol. DCC and TUBB3 in the pellet (P) and supernatant (S) fractions were examined by Western blot analysis using anti-dcc and anti-tubb3, respectively. (E and F) Quantification of three independent experiments showing P/S ratio of DCC (E) and TUBB3 (F), respectively. ns, not significant ; ***, p<0.001 (two-tailed Student s t-test). Netrin-1 increased the cosedimentation of DCC and TUBB3 with polymerized MTs in primary neurons. Figure 3. Src family kinase activity is required for the induction of TUBB3 tyrosine phosphorylation and the interaction of TUBB3 with DCC by netrin-1. (A-B) Induction of TUBB3 tyrosine phosphorylation in dissociated cortical neurons by netrin-1. E15 cortical neurons were treated with purified netrin-1 from 5 to 20 min. The anti-py antibody was used to immunoprecipitate proteins and the immunoblot was analyzed with anti-tubb3. (C) Induction of TUBB3 tyrosine phosphorylation in dissociated E13 dorsal spinal cord neurons by netrin-1. Primary neurons were treated with purified netrin-1 (200 ng/ml). (D) Src family kinasespecific inhibitor PP2 inhibited netrin-1-induced TUBB3 tyrosine phosphorylation. E15 cortical cells were stimulated with netrin-1 in the presence of PP2 or PP3. Quantification was shown in the lower panel. * indicates p<0.05 (Student s t-test). (E) PP2, but not PP3, blocked netrin-1-24
25 induced interaction of endogenous TUBB3 with DCC in primary neurons. (F) Quantification showing relative binding of DCC and TUBB3 in (E). (G-H) Netrin-1-stimulated cosedimentation of DCC with MTs was inhibited by PP2, not PP3. E15 cortical neurons were stimulated with netrin-1 in the presence of PP2 or PP3. DCC in the pellet and supernatant fractions was examined by Western blot. Quantification of (G) was shown in (H). *** indicates p<0.001 (Oneway ANOVA and Fisher LSD post-hoc comparisons). Figure 4. Inhibition of netrin-1-induced axon outgrowth of cortical neurons by TUBB3 knockdown. E15 mouse cortical neurons were transfected with Venus YFP only (A-B), Venus YFP plus the TUBB3 sirna pool (C-D), Venus YFP plus control shrna (E-F), Venus YFP plus TUBB3 shrna (G-H), and Venus YFP plus shrna and the wild-type human RNAi-resistant TUBB3 (I- J), respectively. Neurite outgrowth from YFP-positive neurons was assessed in the presence of purified netrin-1 (B, D, F, H and J) and in the sham-purified control (A, C, E, G and I). Scale bar, 10 µm. (K) Both TUBB3 sirna pool and shrna (#3) reduced endogenous TUBB3 protein levels in E15 cortical neurons. ***, p<0.001 (Student s t-test). (L) Quantification of netrin-1- induced neurite outgrowth. Only the neurites of YFP-positive neurons not in contact with other cells were measured and used in the statistical analyses. Data are mean ± s.e.m. from three separate experiments. The numbers on the top of each bar indicate the numbers of neurons tested in the corresponding groups. ***, p<0.001 (One-way ANOVA with Fischer LSD for post-hoc comparisons). Figure 5. Inhibition of netrin-1-induced axon outgrowth of chick dorsal spinal cord explants by TUBB3 knockdown. The chick neural tubes were electroporated in ovo at stage by Venus YFP only (A-B), Venus YFP plus the TUBB3 sirna pool (C-D), Venus YFP plus control shrna (E-F), Venus YFP plus TUBB3 shrna (G-H), and Venus YFP plus TUBB3 shrna and the wild-type human RNAi-resistant TUBB3 (I-J), respectively. Netrin-1 increased commissural axon outgrowth with longer and more axon bundles in the netrin-1 group (B and F) than the control group (A and E). Netrin-1-induced axon outgrowth was inhibited either by the TUBB3 sirna pool (C-D) or shrna (G-H). The expression of the wild-type human RNAi-resistant TUBB3 plasmids 25
26 reversed the effect of TUBB3 shrna (I-J). Scale bar is 100 µm. (K) Both TUBB3 sirna pool and shrna efficiently knocked down endogenous TUBB3 in chick spinal cords. (L-M) Quantification of netrin-1-induced commissural axon outgrowth. Only YFP-positive axon bundles were measured and used in the statistical analyses. The numbers on the top of each bar indicate the numbers of explants tested in the corresponding groups. Data are mean ± s.e.m. from three separate experiments. ***, p<0.001 (One-way ANOVA with Fischer LSD for posthoc comparisons). Figure 6. Requirement of TUBB3 for netrin-1 attraction of spinal commissural axons. (A) Schematic diagram of the in ovo electroporation and the coculture assay with the open-book preparation. Electroporation of Venus YFP into the neural tube of chick embryos allowed visualization of axons. In the left panels (B, B, D, D, F, F, H, H, J, J ), neural tube explants were co-cultured with control HEK293 cells and commissural axons projected straight toward the floor plate (FP). In the right panels (C, C, E, E, G, G, I, I, K, K ), neural tube explants were co-cultured with aggregates of HEK293 cells secreting netrin-1. The neural tube was electroporated with Venus YFP alone (B-B and C-C ); Venus YFP together with the TUBB3 sirna pool (D-D and E-E ); Venus YFP together with the control shrna (F-F and G-G ); Venus YFP with the TUBB3 shrna (H-H and I-I ) and Venus YFP with the TUBB3 shrna plus the wild-type human RNAi-resistant TUBB3 (J-J and K-K ). Expression of either TUBB3 sirnas or shrna inhibited commissural axon turning towards the netrin-1 source. The commissural axon turning defect of RNAi knock-down could be rescued by expressing RNAiresistant TUBB3. The scale bar is 100 µm. (L) Quantification of axon turning. The numbers on the top of each bar indicate the numbers of explants tested in the corresponding groups. Data are mean ± s.e.m. from group I-VI. *** indicates p<0.001 (One-way ANOVA and Fisher LSD posthoc comparisons). NS, not significant. Figure 7. Inhibition of commissural axon projection in vivo by TUBB3 RNAi. (A) Schematic diagram showing the experimental design. Different combinations of plasmids and sirnas were electroporated into the chick neural tube in ovo at stage 12 and the lumbosacral region of the spinal cord was isolated at stages 23. (B) Neurons electroporated with Venus YFP only. (C) Neurons electroporated with Venus YFP plus the TUBB3 sirna pool. (D) Neurons 26
27 with Venus YFP plus control shrna. (E) Neurons with Venus YFP plus TUBB3 shrna. (F) Neurons with Venus YFP plus shrna and wild-type human RNAi-resistant TUBB3. The red arrowheads point to shortened axons. Scale bar, 100 µm. (G) Quantification of the percentage of axons reaching the FP. The numbers on the top of each bar indicate the numbers of embryos tested in the corresponding groups. ***, p<0.001 (One-way ANOVA with Fischer LSD for posthoc comparisons). Figure 8. TUBB3 is essential for spinal cord commissural axon pathfinding in vivo. (A) Schematic diagram showing the transverse section of the chick spinal cord after electroporation. (B-F) The chick neural tube was electroporated with Venus YFP only (B), Venus YFP plus the TUBB3 sirna pool (C), Venus YFP plus control shrna (D), Venus YFP plus TUBB3 shrna (E), or Venus YFP plus shrna and wild-type human RNAi-resistant TUBB3 (F). Expression of TUBB3 sirnas or shrna not only inhibited the commissural axon extension but also caused aberrant pathfinding (C, E, G-I). The wild-type RNAi-resistant TUBB3 rescued the defect of TUBB3 shrna knockdown on commissural axon projection and turning (F and G-I). The red arrows point to misguided axons. Scale bar, 100 µm. (G) Quantification of the percentage of axons reaching the midline of the chick spinal cord. (H) Quantification of the average distance of axons away from the midline. ***, p<0.001 (One-way ANOVA with Fischer LSD for post-hoc comparisons). (I) The percentage of embryos with misguided axons. The numbers of embryos tested were: 17 for the Venus YFP group; 14 for the sirna pool group; 12 for the control shrna group; 15 for the TUBB3 shrna group; 14 for the rescue group. 27

28
29
30

31
32
33

34

35
Axon Guidance. Multiple decision points along a growing axon s trajectory Different types of axon guidance cues:
 Axon Guidance Multiple decision points along a growing axon s trajectory Different types of axon guidance cues: Contact mediated - requires direct contact by growth cone Long range - growth cone responds
Axon Guidance Multiple decision points along a growing axon s trajectory Different types of axon guidance cues: Contact mediated - requires direct contact by growth cone Long range - growth cone responds
Neurite initiation. Neurite formation begins with a bud that sprouts from the cell body. One or several neurites can sprout at a time.
 Neurite initiation. Neuronal maturation initiation f-actin polarization and maturation tubulin stage 1: "spherical" neuron stage 2: neurons extend several neurites stage 3: one neurite accelerates its
Neurite initiation. Neuronal maturation initiation f-actin polarization and maturation tubulin stage 1: "spherical" neuron stage 2: neurons extend several neurites stage 3: one neurite accelerates its
Netrin signal transduction and the guanine nucleotide exchange factor DOCK180 in attractive signaling
 28 Nature Publishing Group http://www.nature.com/natureneuroscience signal transduction and the guanine nucleotide exchange factor in attractive signaling Xiaoling Li 1,5, Xue Gao 1,5, Guofa Liu 1,5, Wencheng
28 Nature Publishing Group http://www.nature.com/natureneuroscience signal transduction and the guanine nucleotide exchange factor in attractive signaling Xiaoling Li 1,5, Xue Gao 1,5, Guofa Liu 1,5, Wencheng
Axon guidance I. Paul Garrity March 15, /9.013
 Axon guidance I Paul Garrity March 15, 2004 7.68/9.013 Neuronal Wiring: Functional Framework of the Nervous System Stretch reflex circuit Early theories of axonogenesis Schwann: many neurons link to form
Axon guidance I Paul Garrity March 15, 2004 7.68/9.013 Neuronal Wiring: Functional Framework of the Nervous System Stretch reflex circuit Early theories of axonogenesis Schwann: many neurons link to form
Reading. Lecture VI. Making Connections 9/17/12. Bio 3411 Lecture VI. Making Connections. Bio 3411 Monday September 17, 2012
 Lecture VI. Making Connections Bio 3411 Monday September 17, 2012!! 1! Reading NEUROSCIENCE: 5 th ed, pp!507?536! 4 th ed, pp 577-609 Bentley, D., & Caudy, M. (1983). Nature, 304(5921), 62-65. Dickson,
Lecture VI. Making Connections Bio 3411 Monday September 17, 2012!! 1! Reading NEUROSCIENCE: 5 th ed, pp!507?536! 4 th ed, pp 577-609 Bentley, D., & Caudy, M. (1983). Nature, 304(5921), 62-65. Dickson,
Neurite formation & neuronal polarization. The cytoskeletal components of neurons have characteristic distributions and associations
 Mechanisms of neuronal migration & Neurite formation & neuronal polarization Paul Letourneau letou001@umn.edu Chapter 16; The Cytoskeleton; Molecular Biology of the Cell, Alberts et al. 1 The cytoskeletal
Mechanisms of neuronal migration & Neurite formation & neuronal polarization Paul Letourneau letou001@umn.edu Chapter 16; The Cytoskeleton; Molecular Biology of the Cell, Alberts et al. 1 The cytoskeletal
Phosphorylation of DCC by Fyn mediates Netrin-1 signaling in growth cone guidance
 Published Online: 22 November, 2004 Supp Info: http://doi.org/10.1083/jcb.200405053 Downloaded from jcb.rupress.org on April 8, 2018 JCB: ARTICLE Phosphorylation of DCC by Fyn mediates Netrin-1 signaling
Published Online: 22 November, 2004 Supp Info: http://doi.org/10.1083/jcb.200405053 Downloaded from jcb.rupress.org on April 8, 2018 JCB: ARTICLE Phosphorylation of DCC by Fyn mediates Netrin-1 signaling
Neurite formation & neuronal polarization
 Neurite formation & neuronal polarization Paul Letourneau letou001@umn.edu Chapter 16; The Cytoskeleton; Molecular Biology of the Cell, Alberts et al. 1 An immature neuron in cell culture first sprouts
Neurite formation & neuronal polarization Paul Letourneau letou001@umn.edu Chapter 16; The Cytoskeleton; Molecular Biology of the Cell, Alberts et al. 1 An immature neuron in cell culture first sprouts
Hierarchical Organization of Guidance Receptors: Silencing of Netrin Attraction by Slit Through a Robo/DCC Receptor Complex
 Hierarchical Organization of Guidance Receptors: Silencing of Netrin Attraction by Slit Through a Robo/DCC Receptor Complex Elke Stein and Marc Tessier-Lavigne* Axonal growth cones that cross the nervous
Hierarchical Organization of Guidance Receptors: Silencing of Netrin Attraction by Slit Through a Robo/DCC Receptor Complex Elke Stein and Marc Tessier-Lavigne* Axonal growth cones that cross the nervous
Reference: Forscher, P., Kaczmarek, L.K., Buchanan, J. and Smith, S.J. (1987) Cyclic AMP induces changes in distribution and transport of organelles
 Cyclic AMP induces changes in distribution and transport of organelles") Reference: Forscher, P., Kaczmarek, L.K., Buchanan, J. and Smith, S.J. (1987) Cyclic AMP induces changes in distribution and transport of organelles within growth cones of Aplysia bag cell neurons. J.
Reference: Forscher, P., Kaczmarek, L.K., Buchanan, J. and Smith, S.J. (1987) Cyclic AMP induces changes in distribution and transport of organelles within growth cones of Aplysia bag cell neurons. J.
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8
 Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
Supplementary Figure 1.
 Supplementary Figure 1. Characterisation of IHG-1 overexpressing and knockdown cell lines. (A) Total cellular RNA was prepared from HeLa cells stably overexpressing IHG-1 or mts-ihg-1. IHG-1 mrna was quantified
Supplementary Figure 1. Characterisation of IHG-1 overexpressing and knockdown cell lines. (A) Total cellular RNA was prepared from HeLa cells stably overexpressing IHG-1 or mts-ihg-1. IHG-1 mrna was quantified
Pioneer midbrain longitudinal axons navigate using a balance of Netrin attraction and Slit repulsion
 Pioneer midbrain longitudinal axons navigate using a balance of Netrin attraction and Slit repulsion Kim et al. Kim et al. Neural Development 2014, 9:17 Kim et al. Neural Development 2014, 9:17 RESEARCH
Pioneer midbrain longitudinal axons navigate using a balance of Netrin attraction and Slit repulsion Kim et al. Kim et al. Neural Development 2014, 9:17 Kim et al. Neural Development 2014, 9:17 RESEARCH
Nature Neuroscience: doi: /nn.2662
 Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Control of Growth Cone Polarity, Microtubule Accumulation, and Protrusion by UNC- 6/Netrin and its Receptors in C. elegans
 Genetics: Early Online, published on July 25, 2018 as 10.1534/genetics.118.301234 1 2 3 4 5 6 7 8 9 10 11 12 Control of Growth Cone Polarity, Microtubule Accumulation, and Protrusion by UNC- 6/Netrin and
Genetics: Early Online, published on July 25, 2018 as 10.1534/genetics.118.301234 1 2 3 4 5 6 7 8 9 10 11 12 Control of Growth Cone Polarity, Microtubule Accumulation, and Protrusion by UNC- 6/Netrin and
4) Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.
 Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.") Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
CELL-CELL COMMUNICATION
 CELL-CELL COMMUNICATION paracrine & juxtacrine signalling autocrine & intracrine signalling methods to study cell-cell communication: attraction & repulsion chemotaxis & chemokinesis substrate preference
CELL-CELL COMMUNICATION paracrine & juxtacrine signalling autocrine & intracrine signalling methods to study cell-cell communication: attraction & repulsion chemotaxis & chemokinesis substrate preference
RACK-1 Acts with Rac GTPase Signaling and UNC-115/ ablim in Caenorhabditis elegans Axon Pathfinding and Cell Migration
 RACK-1 Acts with Rac GTPase Signaling and UNC-115/ ablim in Caenorhabditis elegans Axon Pathfinding and Cell Migration Rafael S. Demarco, Erik A. Lundquist* Programs in Genetics and Molecular, Cellular,
RACK-1 Acts with Rac GTPase Signaling and UNC-115/ ablim in Caenorhabditis elegans Axon Pathfinding and Cell Migration Rafael S. Demarco, Erik A. Lundquist* Programs in Genetics and Molecular, Cellular,
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Draxin Inhibits Axonal Outgrowth through the Netrin Receptor DCC
 14018 The Journal of Neuroscience, September 28, 2011 31(39):14018 14023 Brief Communications Draxin Inhibits Axonal Outgrowth through the Netrin Receptor DCC Giasuddin Ahmed, 1,2,3 Yohei Shinmyo, 1,2
14018 The Journal of Neuroscience, September 28, 2011 31(39):14018 14023 Brief Communications Draxin Inhibits Axonal Outgrowth through the Netrin Receptor DCC Giasuddin Ahmed, 1,2,3 Yohei Shinmyo, 1,2
Neurite Outgrowth Assay Kit (1 µm)
") Neurite Outgrowth Assay Kit (1 µm) 12 Tests Catalog No. NS225 FOR RESEARCH USE ONLY Not for use in diagnostic procedures USA & Canada Phone: +1(800) 437-7500 Fax: +1 (951) 676-9209 Europe +44 (0) 23 8026
Neurite Outgrowth Assay Kit (1 µm) 12 Tests Catalog No. NS225 FOR RESEARCH USE ONLY Not for use in diagnostic procedures USA & Canada Phone: +1(800) 437-7500 Fax: +1 (951) 676-9209 Europe +44 (0) 23 8026
Supplementary Information
 Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Conclusions. The experimental studies presented in this thesis provide the first molecular insights
 C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
ROLE OF RACK1 IN AXONAL OUTGROWTH OF DEVELOPING NEURONS
 ROLE OF RACK1 IN AXONAL OUTGROWTH OF DEVELOPING NEURONS A thesis submitted to the Kent State University Honors College In partial fulfillment of the requirements For University Honors by Joel Serre May
ROLE OF RACK1 IN AXONAL OUTGROWTH OF DEVELOPING NEURONS A thesis submitted to the Kent State University Honors College In partial fulfillment of the requirements For University Honors by Joel Serre May
C. elegans L1 cell adhesion molecule functions in axon guidance
 C. elegans L1 cell adhesion molecule functions in axon guidance Biorad Lihsia Chen Dept. of Genetics, Cell Biology & Development Developmental Biology Center C. elegans embryogenesis Goldstein lab, UNC-Chapel
C. elegans L1 cell adhesion molecule functions in axon guidance Biorad Lihsia Chen Dept. of Genetics, Cell Biology & Development Developmental Biology Center C. elegans embryogenesis Goldstein lab, UNC-Chapel
Report. CED-10/Rac1 Mediates Axon Guidance by Regulating the Asymmetric Distribution of MIG-10/Lamellipodin
 Current Biology 18, 808 813, June 3, 2008 ª2008 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2008.04.050 CED-10/Rac1 Mediates Axon Guidance by Regulating the Asymmetric Distribution of MIG-10/Lamellipodin
Current Biology 18, 808 813, June 3, 2008 ª2008 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2008.04.050 CED-10/Rac1 Mediates Axon Guidance by Regulating the Asymmetric Distribution of MIG-10/Lamellipodin
THE ROLE OF NCK IN DCC-SENSITIVE SPINAL CORD CIRCUITS. Ciaran Lane
 THE ROLE OF NCK IN DCC-SENSITIVE SPINAL CORD CIRCUITS by Ciaran Lane Submitted in partial fulfilment of the requirements for the degree of Master of Science at Dalhousie University Halifax, Nova Scotia
THE ROLE OF NCK IN DCC-SENSITIVE SPINAL CORD CIRCUITS by Ciaran Lane Submitted in partial fulfilment of the requirements for the degree of Master of Science at Dalhousie University Halifax, Nova Scotia
targets. clustering show that different complex pathway
 Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
TNFα 18hr. Control. CHX 18hr. TNFα+ CHX 18hr. TNFα: 18 18hr (KDa) PARP. Cleaved. Cleaved. Cleaved. Caspase3. Pellino3 shrna. Control shrna.
 PARP. Cleaved. Cleaved. Cleaved. Caspase3. Pellino3 shrna. Control shrna.") Survival ( %) a. TNFα 18hr b. Control sirna Pellino3 sirna TNFα: 18 18hr c. Control shrna Pellino3 shrna Caspase3 Actin Control d. Control shrna Pellino3 shrna *** 100 80 60 CHX 18hr 40 TNFα+ CHX 18hr
Survival ( %) a. TNFα 18hr b. Control sirna Pellino3 sirna TNFα: 18 18hr c. Control shrna Pellino3 shrna Caspase3 Actin Control d. Control shrna Pellino3 shrna *** 100 80 60 CHX 18hr 40 TNFα+ CHX 18hr
Wnt5a Induces Simultaneous Cortical Axon Outgrowth and Repulsive Axon Guidance through Distinct Signaling Mechanisms
 The Journal of Neuroscience, May 6, 2009 29(18):5873 5883 5873 Development/Plasticity/Repair Wnt5a Induces Simultaneous Cortical Axon Outgrowth and Repulsive Axon Guidance through Distinct Signaling Mechanisms
The Journal of Neuroscience, May 6, 2009 29(18):5873 5883 5873 Development/Plasticity/Repair Wnt5a Induces Simultaneous Cortical Axon Outgrowth and Repulsive Axon Guidance through Distinct Signaling Mechanisms
RabGDI controls axonal midline crossing by regulating Robo1 surface expression
 Philipp et al. Neural Development 2012, 7:36 RESEARCH ARTICLE RabGDI controls axonal midline crossing by regulating Robo1 surface expression Open Access Melanie Philipp 1, Vera Niederkofler 2, Marc Debrunner
Philipp et al. Neural Development 2012, 7:36 RESEARCH ARTICLE RabGDI controls axonal midline crossing by regulating Robo1 surface expression Open Access Melanie Philipp 1, Vera Niederkofler 2, Marc Debrunner
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins
 13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
Myosin X regulates netrin receptors and functions in axonal path-finding
 Myosin X regulates netrin receptors and functions in axonal path-finding Xiao-Juan Zhu 1,4, Cheng-Zhong Wang 2,4, Peng-Gao Dai 1,4, Yi Xie 3, Ning-Ning Song 2, Yu Liu 1, Quan-Sheng Du 1, Lin Mei 1, Yu-Qiang
Myosin X regulates netrin receptors and functions in axonal path-finding Xiao-Juan Zhu 1,4, Cheng-Zhong Wang 2,4, Peng-Gao Dai 1,4, Yi Xie 3, Ning-Ning Song 2, Yu Liu 1, Quan-Sheng Du 1, Lin Mei 1, Yu-Qiang
SDQR migrations in Caenorhabditis elegans are controlled by multiple guidance cues and changing responses to netrin UNC-6
 Development 126, 3881-389 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV8558 3881 SDQR migrations in Caenorhabditis elegans are controlled by multiple guidance cues and changing
Development 126, 3881-389 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV8558 3881 SDQR migrations in Caenorhabditis elegans are controlled by multiple guidance cues and changing
Cell Death & Trophic Factors II. Steven McLoon Department of Neuroscience University of Minnesota
 Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
The N-terminal Leucine-Rich Regions in Slit Are Sufficient To Repel Olfactory Bulb Axons and Subventricular Zone Neurons
 The Journal of Neuroscience, March 1, 2001, 21(5):1548 1556 The N-terminal Leucine-Rich Regions in Slit Are Sufficient To Repel Olfactory Bulb Axons and Subventricular Zone Neurons Jin-hui Chen, 1 Leng
The Journal of Neuroscience, March 1, 2001, 21(5):1548 1556 The N-terminal Leucine-Rich Regions in Slit Are Sufficient To Repel Olfactory Bulb Axons and Subventricular Zone Neurons Jin-hui Chen, 1 Leng
Report. Functional Diversity of Robo Receptor Immunoglobulin Domains Promotes Distinct Axon Guidance Decisions
 Current Biology 20, 567 572, March 23, 2010 ª2010 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2010.02.021 Functional Diversity of Robo Receptor Immunoglobulin Domains Promotes Distinct Axon Guidance
Current Biology 20, 567 572, March 23, 2010 ª2010 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2010.02.021 Functional Diversity of Robo Receptor Immunoglobulin Domains Promotes Distinct Axon Guidance
5- Semaphorin-Plexin-Neuropilin
 5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
Supporting Information
 Supporting Information Wang et al. 10.1073/pnas.0804871105 SI Materials and Methods Cell Culture and Transfection. Human neuroblastoma M17 cells were grown as described before (1). Transfection was performed
Supporting Information Wang et al. 10.1073/pnas.0804871105 SI Materials and Methods Cell Culture and Transfection. Human neuroblastoma M17 cells were grown as described before (1). Transfection was performed
Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition
 Supplementary Information to Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition Nadine Czudnochowski 1,2, *, Christian A. Bösken 1, * & Matthias Geyer 1 1 Max-Planck-Institut
Supplementary Information to Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition Nadine Czudnochowski 1,2, *, Christian A. Bösken 1, * & Matthias Geyer 1 1 Max-Planck-Institut
Cellular/Molecular The Journal of Neuroscience, March 23, (12):
:") 3132 The Journal of Neuroscience, March 23, 2005 25(12):3132 3141 Cellular/Molecular Deleted in Colorectal Cancer Binding Netrin-1 Mediates Cell Substrate Adhesion and Recruits Cdc42, Rac1, Pak1, and N-WASP
3132 The Journal of Neuroscience, March 23, 2005 25(12):3132 3141 Cellular/Molecular Deleted in Colorectal Cancer Binding Netrin-1 Mediates Cell Substrate Adhesion and Recruits Cdc42, Rac1, Pak1, and N-WASP
Migration of Neurons During Embryonic Development. Christine Simmons Saint Louis University April 15 th, 2008
 Migration of Neurons During Embryonic Development Christine Simmons Saint Louis University April 15 th, 2008 Embryonic Development of the CNS Central Nervous System (CNS) arises from Surface ectoderm Skin,
Migration of Neurons During Embryonic Development Christine Simmons Saint Louis University April 15 th, 2008 Embryonic Development of the CNS Central Nervous System (CNS) arises from Surface ectoderm Skin,
Transmembrane Receptor DCC Associates with Protein Synthesis Machinery and Regulates Translation
 Transmembrane Receptor DCC Associates with Protein Synthesis Machinery and Regulates Translation Joseph Tcherkezian, 1,5 Perry A. Brittis, 1,5, * Franziska Thomas, 2,6 Philippe P. Roux, 3,4 and John G.
Transmembrane Receptor DCC Associates with Protein Synthesis Machinery and Regulates Translation Joseph Tcherkezian, 1,5 Perry A. Brittis, 1,5, * Franziska Thomas, 2,6 Philippe P. Roux, 3,4 and John G.
Activation of ADF/Cofilin Mediates Attractive Growth Cone Turning Toward Nerve Growth Factor and Netrin-1
 Activation of ADF/Cofilin Mediates Attractive Growth Cone Turning Toward Nerve Growth Factor and Netrin-1 Bonnie M. Marsick, 1,2 Kevin C. Flynn, 3 * Miguel Santiago-Medina, 4 James R. Bamburg, 3,5 Paul
Activation of ADF/Cofilin Mediates Attractive Growth Cone Turning Toward Nerve Growth Factor and Netrin-1 Bonnie M. Marsick, 1,2 Kevin C. Flynn, 3 * Miguel Santiago-Medina, 4 James R. Bamburg, 3,5 Paul
Inactivation of Ras by p120gap via Focal Adhesion Kinase Dephosphorylation Mediates RGMa-Induced Growth Cone Collapse
 The Journal of Neuroscience, May 20, 2009 29(20):6649 6662 6649 Cellular/Molecular Inactivation of Ras by p120gap via Focal Adhesion Kinase Dephosphorylation Mediates RGMa-Induced Growth Cone Collapse
The Journal of Neuroscience, May 20, 2009 29(20):6649 6662 6649 Cellular/Molecular Inactivation of Ras by p120gap via Focal Adhesion Kinase Dephosphorylation Mediates RGMa-Induced Growth Cone Collapse
Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing
 Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing 0.1% SDS without boiling. The gel was analyzed by a fluorescent
Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing 0.1% SDS without boiling. The gel was analyzed by a fluorescent
Turning of Retinal Growth Cones in a Netrin-1 Gradient Mediated by the Netrin Receptor DCC
 Neuron, Vol. 19, 1211 1224, December, 1997, Copyright 1997 by Cell Press Turning of Retinal Growth Cones in a Netrin-1 Gradient Mediated by the Netrin Receptor DCC José R. de la Torre,* Veit H. Höpker,
Neuron, Vol. 19, 1211 1224, December, 1997, Copyright 1997 by Cell Press Turning of Retinal Growth Cones in a Netrin-1 Gradient Mediated by the Netrin Receptor DCC José R. de la Torre,* Veit H. Höpker,
Microtubule dynamics in axon guidance
 Neurosci Bull June, 2014. http://www.neurosci.cn DOI: 10.1007/s12264-014-1444-6 1 Microtubule dynamics in axon guidance Guofa Liu, Trisha Dwyer Department of Biological Sciences, University of Toledo,
Neurosci Bull June, 2014. http://www.neurosci.cn DOI: 10.1007/s12264-014-1444-6 1 Microtubule dynamics in axon guidance Guofa Liu, Trisha Dwyer Department of Biological Sciences, University of Toledo,
Mapping Netrin Receptor Binding Reveals Domains of Unc5 Regulating Its Tyrosine Phosphorylation
 10826 The Journal of Neuroscience, December 1, 2004 24(48):10826 10834 Cellular/Molecular Mapping Netrin Receptor Binding Reveals Domains of Unc5 Regulating Its Tyrosine Phosphorylation Robert P. Kruger,
10826 The Journal of Neuroscience, December 1, 2004 24(48):10826 10834 Cellular/Molecular Mapping Netrin Receptor Binding Reveals Domains of Unc5 Regulating Its Tyrosine Phosphorylation Robert P. Kruger,
Varicose and cheerio collaborate with pebble to mediate semaphorin-1a reverse signaling in Drosophila
 Varicose and cheerio collaborate with pebble to mediate semaphorin-1a reverse signaling in Drosophila Sangyun Jeong a,1, Da-som Yang a, Young Gi Hong a, Sarah P. Mitchell b,c, Matthew P. rown b,c, and
Varicose and cheerio collaborate with pebble to mediate semaphorin-1a reverse signaling in Drosophila Sangyun Jeong a,1, Da-som Yang a, Young Gi Hong a, Sarah P. Mitchell b,c, Matthew P. rown b,c, and
MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning
 MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning Learning Goals for Lecture 29 4.1 Describe the contributions of early developmental events in the embryo to the formation
MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning Learning Goals for Lecture 29 4.1 Describe the contributions of early developmental events in the embryo to the formation
The Cytoskeleton of the Neuron An Essay in Celebration of Paul Letourneau s Career
 The Cytoskeleton of the Neuron An Essay in Celebration of Paul Letourneau s Career Daphney C. Jean, 1 Mark M. Black, 2 Peter W. Baas 1 1 Department of Neurobiology and Anatomy, Drexel University College
The Cytoskeleton of the Neuron An Essay in Celebration of Paul Letourneau s Career Daphney C. Jean, 1 Mark M. Black, 2 Peter W. Baas 1 1 Department of Neurobiology and Anatomy, Drexel University College
Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A)
 M-phase nuclei are visualized by a-ph3 labeling in wild-type (A)") Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A) and mutant (B) 4 dpf retinae. The central retina of the
Fig. S1. Proliferation and cell cycle exit are affected by the med mutation. (A,B) M-phase nuclei are visualized by a-ph3 labeling in wild-type (A) and mutant (B) 4 dpf retinae. The central retina of the
Supplementary Figure 1. Biochemical and sequence alignment analyses the
 Supplementary Figure 1. Biochemical and sequence alignment analyses the interaction of OPTN and TBK1. (a) Analytical gel filtration chromatography analysis of the interaction between TBK1 CTD and OPTN(1-119).
Supplementary Figure 1. Biochemical and sequence alignment analyses the interaction of OPTN and TBK1. (a) Analytical gel filtration chromatography analysis of the interaction between TBK1 CTD and OPTN(1-119).
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
Monitoring neurite morphology and synapse formation in primary neurons for neurotoxicity assessments and drug screening
 APPLICATION NOTE ArrayScan High Content Platform Monitoring neurite morphology and synapse formation in primary neurons for neurotoxicity assessments and drug screening Suk J. Hong and Richik N. Ghosh
APPLICATION NOTE ArrayScan High Content Platform Monitoring neurite morphology and synapse formation in primary neurons for neurotoxicity assessments and drug screening Suk J. Hong and Richik N. Ghosh
Supplemental Information. The Mitochondrial Fission Receptor MiD51. Requires ADP as a Cofactor
 Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila
 Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Supplementary Figure 1-10 with Figure legends
 332 coordinates mechanotransduction and growth cone bifurcation in sensory neurons Li Yang Chiang 1, Kate Poole, Beatriz E. Oliveira, Neuza Duarte,Yinth Andrea Bernal Sierra, Leena Bruckner Tuderman, Manuel
332 coordinates mechanotransduction and growth cone bifurcation in sensory neurons Li Yang Chiang 1, Kate Poole, Beatriz E. Oliveira, Neuza Duarte,Yinth Andrea Bernal Sierra, Leena Bruckner Tuderman, Manuel
Neural development its all connected
 Neural development its all connected How do you build a complex nervous system? How do you build a complex nervous system? 1. Learn how tissue is instructed to become nervous system. Neural induction 2.
Neural development its all connected How do you build a complex nervous system? How do you build a complex nervous system? 1. Learn how tissue is instructed to become nervous system. Neural induction 2.
7.013 Problem Set
 7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
Crk-associated substrate (Cas) signaling protein functions with integrins to specify axon guidance during development
 signaling protein functions with integrins to specify axon guidance during development") RESEARCH ARTICLE 2337 Development 134, 2337-2347 (2007) doi:10.1242/dev.004242 Crk-associated substrate (Cas) signaling protein functions with integrins to specify axon guidance during development Zhiyu
RESEARCH ARTICLE 2337 Development 134, 2337-2347 (2007) doi:10.1242/dev.004242 Crk-associated substrate (Cas) signaling protein functions with integrins to specify axon guidance during development Zhiyu
Role of Organizer Chages in Late Frog Embryos
 Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2647 Figure S1 Other Rab GTPases do not co-localize with the ER. a, Cos-7 cells cotransfected with an ER luminal marker (either KDEL-venus or mch-kdel) and mch-tagged human Rab5 (mch-rab5,
DOI: 10.1038/ncb2647 Figure S1 Other Rab GTPases do not co-localize with the ER. a, Cos-7 cells cotransfected with an ER luminal marker (either KDEL-venus or mch-kdel) and mch-tagged human Rab5 (mch-rab5,
JOURNAL OF NEUROCHEMISTRY doi: /j x
 JOURNAL OF NEUROCHEMISTRY 2008 107 398 417 doi: 10.1111/j.1471-4159.2008.05609.x, *Department of Neurology and Neurosurgery, Montreal Neurological Institute, McGill University, Montreal, Quebec, Canada
JOURNAL OF NEUROCHEMISTRY 2008 107 398 417 doi: 10.1111/j.1471-4159.2008.05609.x, *Department of Neurology and Neurosurgery, Montreal Neurological Institute, McGill University, Montreal, Quebec, Canada
Netrin- DCC Signaling Regulates Corpus Callosum Formation Through Attraction of Pioneering Axons and by Modulating Slit2- Mediated Repulsion
 Netrin- DCC Signaling Regulates Corpus Callosum Formation Through Attraction of Pioneering Axons and by Modulating Slit2- Mediated Repulsion Thomas Fothergill1,5, Amber- Lee S. Donahoo1, Amelia Douglass1,
Netrin- DCC Signaling Regulates Corpus Callosum Formation Through Attraction of Pioneering Axons and by Modulating Slit2- Mediated Repulsion Thomas Fothergill1,5, Amber- Lee S. Donahoo1, Amelia Douglass1,
JCB. A novel Dbl family RhoGEF promotes Rho-dependent axon attraction to the central nervous system midline in Drosophila and overcomes Robo repulsion
 JCB Report A novel Dbl family RhoGEF promotes Rho-dependent axon attraction to the central nervous system midline in Drosophila and overcomes Robo repulsion Greg J. Bashaw, 1,2 Hailan Hu, 2 Catherine D.
JCB Report A novel Dbl family RhoGEF promotes Rho-dependent axon attraction to the central nervous system midline in Drosophila and overcomes Robo repulsion Greg J. Bashaw, 1,2 Hailan Hu, 2 Catherine D.
Avoidance of posterior tectal membranes by temporal retinal axons
 Development 101. 909-913 (1987) Printed in Great Britain The Company of Biologists Limited 1987 909 Avoidance of posterior tectal membranes by temporal retinal axons JOCHEN WALTER, SIGRID HENKE-FAHLE and
Development 101. 909-913 (1987) Printed in Great Britain The Company of Biologists Limited 1987 909 Avoidance of posterior tectal membranes by temporal retinal axons JOCHEN WALTER, SIGRID HENKE-FAHLE and
NOVA regulates Dcc alternative splicing during neuronal migration and axon guidance in the spinal cord
 RESEARCH ARTICLE NOVA regulates Dcc alternative splicing during neuronal migration and axon guidance in the spinal cord Janelle C Leggere 1, Yuhki Saito 2, Robert B Darnell 2, Marc Tessier-Lavigne 3, Harald
RESEARCH ARTICLE NOVA regulates Dcc alternative splicing during neuronal migration and axon guidance in the spinal cord Janelle C Leggere 1, Yuhki Saito 2, Robert B Darnell 2, Marc Tessier-Lavigne 3, Harald
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018
 Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Deleted in Colorectal Cancer (DCC) Encodes a Netrin Receptor
 Encodes a Netrin Receptor") Cell, Vol. 87, 175 185, October 18, 1996, Copyright 1996 by Cell Press Deleted in Colorectal Cancer (DCC) Encodes a Netrin Receptor Kazuko Keino-Masu,* Masayuki Masu,* developing axons, but the receptors
Cell, Vol. 87, 175 185, October 18, 1996, Copyright 1996 by Cell Press Deleted in Colorectal Cancer (DCC) Encodes a Netrin Receptor Kazuko Keino-Masu,* Masayuki Masu,* developing axons, but the receptors
Selective Targeting of ER Exit Sites Supports Axon Development
 Traffic 2009; 10: 1669 1684 2009 John Wiley & Sons A/S doi:10.1111/j.1600-0854.2009.00974.x Selective Targeting of ER Exit Sites Supports Axon Development Meir Aridor 1 and Kenneth N. Fish 2, 1 Department
Traffic 2009; 10: 1669 1684 2009 John Wiley & Sons A/S doi:10.1111/j.1600-0854.2009.00974.x Selective Targeting of ER Exit Sites Supports Axon Development Meir Aridor 1 and Kenneth N. Fish 2, 1 Department
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
THE PROBLEMS OF DEVELOPMENT. Cell differentiation. Cell determination
 We emphasize these points from Kandel in Bi/CNS 150 Bi/CNS/NB 150: Neuroscience Read Lecture Lecture Friday, October 2, 2015 Development 1: pp 5-10 Introduction Brains evolved All higher animals have brains
We emphasize these points from Kandel in Bi/CNS 150 Bi/CNS/NB 150: Neuroscience Read Lecture Lecture Friday, October 2, 2015 Development 1: pp 5-10 Introduction Brains evolved All higher animals have brains
1. The plasma membrane of eukaryotic cells is supported by a. actin filaments. b. microtubules. c. lamins. d. intermediate filaments.
 ANALYSIS AND MODELING OF CELL MECHANICS Homework #2 (due 1/30/13) This homework involves comprehension of key biomechanical concepts of the cytoskeleton, cell-matrix adhesions, and cellcell adhesions.
ANALYSIS AND MODELING OF CELL MECHANICS Homework #2 (due 1/30/13) This homework involves comprehension of key biomechanical concepts of the cytoskeleton, cell-matrix adhesions, and cellcell adhesions.
Axon Guidance at the Midline: From Mutants to Mechanisms
 Critical Reviews in Biochemistry and Molecular Biology, 39:319 341, 2004 Copyright c Taylor & Francis Inc. ISSN: 1040-9238print / 1549-7798online DOI: 10.1080/10409230490906797 Axon Guidance at the Midline:
Critical Reviews in Biochemistry and Molecular Biology, 39:319 341, 2004 Copyright c Taylor & Francis Inc. ISSN: 1040-9238print / 1549-7798online DOI: 10.1080/10409230490906797 Axon Guidance at the Midline:
Supplementary Figures
 Supplementary Figures Supplementary Fig. S1: Normal development and organization of the embryonic ventral nerve cord in Platynereis. (A) Life cycle of Platynereis dumerilii. (B-F) Axonal scaffolds and
Supplementary Figures Supplementary Fig. S1: Normal development and organization of the embryonic ventral nerve cord in Platynereis. (A) Life cycle of Platynereis dumerilii. (B-F) Axonal scaffolds and
Waithe et al Supplementary Figures
 Waithe et al Supplementary Figures Supplementary Figure 1 Expression and properties of WT and W391A mutant YFP- Ca V 2.2. A Immunoblot using Ca V 2.2 Ab for untransfected cells (UT, lane 1), YFP-Ca V 2.2
Waithe et al Supplementary Figures Supplementary Figure 1 Expression and properties of WT and W391A mutant YFP- Ca V 2.2. A Immunoblot using Ca V 2.2 Ab for untransfected cells (UT, lane 1), YFP-Ca V 2.2
Metadata of the chapter that will be visualized online
 Metadata of the chapter that will be visualized online ChapterTitle Chapter Sub-Title Regulation of Actin Filaments During Neurite Extension and Guidance Chapter CopyRight - Year Springer Science+Business
Metadata of the chapter that will be visualized online ChapterTitle Chapter Sub-Title Regulation of Actin Filaments During Neurite Extension and Guidance Chapter CopyRight - Year Springer Science+Business
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Chapter 16. Cellular Movement: Motility and Contractility. Lectures by Kathleen Fitzpatrick Simon Fraser University Pearson Education, Inc.
 Chapter 16 Cellular Movement: Motility and Contractility Lectures by Kathleen Fitzpatrick Simon Fraser University Two eukaryotic motility systems 1. Interactions between motor proteins and microtubules
Chapter 16 Cellular Movement: Motility and Contractility Lectures by Kathleen Fitzpatrick Simon Fraser University Two eukaryotic motility systems 1. Interactions between motor proteins and microtubules
Motor Axon Subpopulations Respond Differentially to the Chemorepellents Netrin-1 and Semaphorin D
 Neuron, Vol. 18, 193 207, February, 1997, Copyright 1997 by Cell Press Motor Axon Subpopulations Respond Differentially to the Chemorepellents Netrin-1 and Semaphorin D Alfredo Varela-Echavarría,* Anna
Neuron, Vol. 18, 193 207, February, 1997, Copyright 1997 by Cell Press Motor Axon Subpopulations Respond Differentially to the Chemorepellents Netrin-1 and Semaphorin D Alfredo Varela-Echavarría,* Anna
Vertebrate Slit, a Secreted Ligand for the Transmembrane Protein Roundabout, Is a Repellent for Olfactory Bulb Axons
 Cell, Vol. 96, 807 818, March 19, 1999, Copyright 1999 by Cell Press Vertebrate Slit, a Secreted Ligand for the Transmembrane Protein Roundabout, Is a Repellent for Olfactory Bulb Axons Hua-shun Li,*#
Cell, Vol. 96, 807 818, March 19, 1999, Copyright 1999 by Cell Press Vertebrate Slit, a Secreted Ligand for the Transmembrane Protein Roundabout, Is a Repellent for Olfactory Bulb Axons Hua-shun Li,*#
The role of ephrins and structured retinal activity in the development of visual map topography
 The role of ephrins and structured retinal activity in the development of visual map topography David Feldheim, UC Santa Cruz KITP Brain08 March 21, 2008 Topographic map development in the mouse visual
The role of ephrins and structured retinal activity in the development of visual map topography David Feldheim, UC Santa Cruz KITP Brain08 March 21, 2008 Topographic map development in the mouse visual
The majority of cells in the nervous system arise during the embryonic and early post
 Introduction Introduction The majority of cells in the nervous system arise during the embryonic and early post natal period. These cells are derived from population of neural stem cells first shown by
Introduction Introduction The majority of cells in the nervous system arise during the embryonic and early post natal period. These cells are derived from population of neural stem cells first shown by
The cellular events that precede myelination in the
 JCB: ARTICLE ErbB2 directly activates the exchange factor Dock7 to promote Schwann cell migration Junji Yamauchi, 1 Yuki Miyamoto, 1 Jonah R. Chan, 2 and Akito Tanoue 1 1 Department of Pharmacology, National
JCB: ARTICLE ErbB2 directly activates the exchange factor Dock7 to promote Schwann cell migration Junji Yamauchi, 1 Yuki Miyamoto, 1 Jonah R. Chan, 2 and Akito Tanoue 1 1 Department of Pharmacology, National
A Theoretical Model of Axon Guidance by the Robo Code
 NOTE Communicated by Arjen van Ooyen A Theoretical Model of Axon Guidance by the Robo Code Geoffrey J. Goodhill geoff@georgetown.edu Department of Neuroscience, Georgetown University Medical Center, Washington,
NOTE Communicated by Arjen van Ooyen A Theoretical Model of Axon Guidance by the Robo Code Geoffrey J. Goodhill geoff@georgetown.edu Department of Neuroscience, Georgetown University Medical Center, Washington,
WD Repeat Domain of Dictyostelium Myosin Heavy Chain Kinase C Functions in both Substrate Targeting and Cellular Localization,
 WD Repeat Domain of Dictyostelium Myosin Heavy Chain Kinase C Functions in both Substrate Targeting and Cellular Localization, By: Atiya Franklin, Linzi Hyatt, Alyssa Chowdhury, and Paul A. Steimle* Franklin,
WD Repeat Domain of Dictyostelium Myosin Heavy Chain Kinase C Functions in both Substrate Targeting and Cellular Localization, By: Atiya Franklin, Linzi Hyatt, Alyssa Chowdhury, and Paul A. Steimle* Franklin,
Introduction Principles of Signaling and Organization p. 3 Signaling in Simple Neuronal Circuits p. 4 Organization of the Retina p.
 Introduction Principles of Signaling and Organization p. 3 Signaling in Simple Neuronal Circuits p. 4 Organization of the Retina p. 5 Signaling in Nerve Cells p. 9 Cellular and Molecular Biology of Neurons
Introduction Principles of Signaling and Organization p. 3 Signaling in Simple Neuronal Circuits p. 4 Organization of the Retina p. 5 Signaling in Nerve Cells p. 9 Cellular and Molecular Biology of Neurons
We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists. International authors and editors
 We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists 3,800 116,000 120M Open access books available International authors and editors Downloads Our
We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists 3,800 116,000 120M Open access books available International authors and editors Downloads Our
The Abelson tyrosine kinase, the Trio GEF and Enabled interact with the Netrin receptor Frazzled in Drosophila
 1983 The Abelson tyrosine kinase, the Trio GEF and Enabled interact with the Netrin receptor Frazzled in Drosophila David J. Forsthoefel 1, Eric C. Liebl 2, Peter A. Kolodziej 3, * and Mark A. Seeger 1,
1983 The Abelson tyrosine kinase, the Trio GEF and Enabled interact with the Netrin receptor Frazzled in Drosophila David J. Forsthoefel 1, Eric C. Liebl 2, Peter A. Kolodziej 3, * and Mark A. Seeger 1,
NOVA regulates Dcc alternative splicing during neuronal migration and axon guidance in the spinal cord.
 University of Colorado, Boulder CU Scholar Molecular, Cellular, and Developmental Biology Faculty Contributions Molecular, Cellular, and Developmental Biology 5-25-2016 NOVA regulates Dcc alternative splicing
University of Colorado, Boulder CU Scholar Molecular, Cellular, and Developmental Biology Faculty Contributions Molecular, Cellular, and Developmental Biology 5-25-2016 NOVA regulates Dcc alternative splicing
Nature Biotechnology: doi: /nbt Supplementary Figure 1. Overexpression of YFP::GPR-1 in the germline.
 Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
The Molecular Biology of Axon Guidance
 Thursday, January 25, 2001 Science -- Tessier-Lavigne and Goodman 274 (5290): 1123 Page: 1 Institution: HARVARD UNIVERSITY Sign In as Individual FAQ The Molecular Biology of Axon Guidance Marc Tessier-Lavigne
Thursday, January 25, 2001 Science -- Tessier-Lavigne and Goodman 274 (5290): 1123 Page: 1 Institution: HARVARD UNIVERSITY Sign In as Individual FAQ The Molecular Biology of Axon Guidance Marc Tessier-Lavigne
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1
 and Ksp/Cre;Pkd1") Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR