Genetics: Early Online, published on December 29, 2015 as /genetics
|
|
|
- Camilla Johns
- 5 years ago
- Views:
Transcription
1 Genetics: Early Online, published on December, 01 as.1/genetics The Mediator kinase module restrains epidermal growth factor receptor signaling and represses vulval cell fate specification in Caenorhabditis elegans Jennifer M. Grants *,, Lisa T. L. Ying, Akinori Yoda,1, Charlotte C. You, Hideyuki Okano,, Hitoshi Sawa,**, and Stefan Taubert *, 1 1 * Department of Medical Genetics, University of British Columbia, Vancouver, BC, Canada; Centre for Molecular Medicine and Therapeutics, Child & Family Research Institute, University of British Columbia, Vancouver, BC, Canada; Division of Neuroanatomy, Osaka University Graduate School of Medicine, Osaka, Japan; Department of Physiology, Keio University School of Medicine, Tokyo, Japan; and ** Multicellular Organization Laboratory, National Institute of Genetics, Mishima, Japan Current address: Department of Medical Oncology, Dana-Farber Cancer Institute, Boston, MA, USA 1 1 Copyright 01.
2 Keywords: Mediator complex, CDK, MED, MED1, EGFR, Notch Running title: CDK- inhibits vulval organogenesis Corresponding author: Stefan Taubert -0 W th Avenue, Vancouver, BC, Canada, VZ H Tel: 0--0, taubert@cmmt.ubc.ca
3 Abstract Cell signaling pathways that control proliferation and determine cell fates are tightly regulated to prevent developmental anomalies and cancer. Transcription factors and coregulators are important effectors of signaling pathway output, as they regulate downstream gene programs. In Caenorhabditis elegans, several subunits of the Mediator transcriptional coregulator complex promote or inhibit vulva development, but pertinent mechanisms are poorly defined. Here, we show that Mediator s dissociable Cyclin Dependent Kinase (CDK) Module (CKM), consisting of cdk-, cic-1/cyclin C, mdt-1/dpy-, and mdt-1/let-1, is required to inhibit ectopic vulval cell fates downstream of the epidermal growth factor receptor (EGFR)-Ras-extracellular signal-regulated kinase (ERK) pathway. cdk- inhibits ectopic vulva formation by acting downstream of mpk-1/erk, cell autonomously in vulval cells, and in a kinase-dependent manner. We also provide evidence that the CKM acts as a corepressor for the Ets-family transcription factor LIN-1, as cdk- promotes transcriptional repression by LIN-1. In addition, we find that CKM mutation alters Mediator subunit requirements in vulva development: the mdt-/sur- subunit, which is required for vulva development in wild-type worms, is dispensable for ectopic vulva formation in CKM mutants, which instead display hallmarks of unrestrained Mediator tail module activity. We propose a model whereby the CKM controls EGFR-Ras-ERK transcriptional output by corepressing LIN-1 and by fine-tuning Mediator specificity, thus balancing transcriptional repression vs. activation in a critical developmental signaling pathway. Collectively, these data offer an explanation for CKM repression of EGFR signaling output and ectopic vulva formation and provide the first evidence of Mediator CKM-tail module subunit crosstalk in animals.
4 Introduction Precise regulation of transcription is required to execute developmental programs such as proliferation and cell fate determination. The Mediator complex ( Mediator ) is a conserved eukaryotic transcriptional coregulator of RNA polymerase II (Pol II) transcription (Malik and Roeder 0; Poss et al. 01). Mediator consists of ~0 subunits that assemble into four modules. Core Mediator consists of three of the four modules: the head and middle modules, which contact Pol II, and the tail module, which serves as a docking site for transcription factors. The fourth module, the dissociable CDK kinase module (CKM), interacts with transcription factors, core Mediator, chromatin, and the Pol II machinery to either repress or activate transcription (Malik and Roeder 0; Nemet et al. 01). Whereas many head or middle Mediator subunits are broadly required for Pol II transcription, tail and CKM subunits regulate specific transcriptional programs in animal development or physiology (Malik and Roeder 0; Nemet et al. 01). The CKM consists of enzymatic subunits Cyclin Dependent Kinase (CDK), Cyclin C, and structural subunits MED1, and MED1 that tether the CKM to core Mediator (Tsai et al. 01). CKM subunits regulate many transcriptional programs important for development and/or tumorigenesis, often by directly binding to and influencing the activity of key transcription factors (e.g. β-catenin, Notch, etc.) (Fryer et al. 00; Donner et al. 00; Firestein et al. 00; Zhou et al. 01). Furthermore, in Saccharomyces cerevisiae, the CKM regulates the activity of the Mediator tail module subunits MED, MED, and MED1 (van de Peppel et al. 00; Gonzalez et al. 01). However, whether such intra- Mediator signaling effects occur in metazoans and affect e.g. animal development has not yet been tested. Several Mediator subunits including at least one CKM subunit regulate vulva development in Caenorhabditis elegans (Tuck and Greenwald 1; Singh and Han 1; Kwon and Lee 001; Moghal and Sternberg 00a). The study of cell fate specification in the C. elegans vulva has proven a powerful way to identify the components and regulatory interactions of several evolutionarily
5 conserved signaling pathways (Félix and Barkoulas 01; Schmid and Hajnal 01). Thus, this organogenesis event provides an ideal paradigm to study Mediator subunit specificity and cooperation in a metazoan. C. elegans vulval organogenesis is induced by epidermal growth factor receptor (EGFR) signaling (Moghal and Sternberg 00b), a prominent pathway in animal development that is frequently activated in human cancers (Normanno et al. 00; Baselga and Swain 00). The C. elegans vulva develops from six ventral vulva precursor cells (VPCs), named P.p through P.p from anterior to posterior (Fig. 1). The VPCs form an equivalence group, meaning that all six cells are able to adopt the primary (1 ) vulval cell fate (producing eight descendants), the secondary ( ) vulval cell fate (producing seven descendants), or the tertiary ( ) non-vulval fate (producing two descendants that fuse with the surrounding hypodermis). A signaling cell in the somatic gonad, called the anchor cell, emits a LIN-/EGF-like ligand in close proximity to P.p (Hill and Sternberg 1); therefore, LET- /EGFR and the downstream LET-0/Ras, MPK-1/ Extracellular signal-regulated kinase (ERK) cascade is strongly activated in P.p (Aroian et al. 10). MPK-1/ERK activation in P.p modulates the activity of effector transcription factors such as the ELK1/Ets-family transcription factor LIN-1 and the FoxB transcription factor LIN-1, thereby specifying the 1 vulval fate in P.p (Miller et al. 1; Tan et al. 1; Jacobs et al. 1). The neighbouring P.p and P.p cells are thought to receive a weaker LIN-/EGF signal from the anchor cell (Katz et al. 1) as well as a lateral Notch signal emitted from the 1 cell P.p, inducing them to adopt a vulval fate (Chen and Greenwald 00). Located furthest form the anchor cell, P.p, P.p, and P.p do not receive sufficient EGF signal, and adopt the non-vulval cell fate (Sternberg and Horvitz 1). Mutations that enhance or reduce EGFR or Notch signaling induce ectopic vulval cell fates (Muv, multivulva phenotype) or loss of vulval cell fates (Vul, vulvaless phenotype), respectively (Sternberg and Horvitz 1). These phenotypes are thus powerful indicators of EGFR and Notch signaling pathway activity.
6 Transcriptional regulation is important in maintaining appropriate EGFR signaling pathway output (Fig. 1). For example, transcription factors such as LIN-1/Ets and LIN-1/Forkhead are required to repress 1 cell fate specification in VPCs other than P.p (Miller et al. 1; Beitel et al. 1). In addition, multiple chromatin-modifying complexes, encoded by the synthetic Multivulva (synmuv) genes, redundantly repress ectopic lin-/egf transcription in the hypodermis and other tissues to inhibit 1 cell fate specification in VPCs other than P.p (Myers and Greenwald 00; Cui et al. 00; Saffer et al. 0). Furthermore, the Mediator subunits mdt-/sur-, mdt-/lin-, and mdt- promote vulva development, whereas the CKM subunit mdt-1/dpy- inhibits vulva development in an anchor cell independent manner (reviewed in (Grants et al. 01); for standardized Mediator subunit nomenclature, please see (Bourbon et al. 00)). The mechanism by which mdt-/sur- promotes vulva development has been partially elucidated, as it is a critical coactivator of a target gene downstream of the EGFR signaling pathway, the lag- Notch ligand gene (Zhang and Greenwald 0). The lin-1/ets effector transcription factor is similarly required to repress the lag- gene (Zhang and Greenwald 0), raising the question of whether and how Mediator and LIN-1 interact to control common target genes. The other three Mediator subunits implicated in vulva development (mdt-, mdt- 1/dpy-, and mdt-/lin-) interact genetically with components of the EGFR signaling pathway, but their mode of action within this pathway remain poorly understood. Here, we used the vulva organogenesis paradigm to study the requirements of all four CKM subunits in this process and to interrogate functional interactions with other transcriptional regulators, including the synmuv genes, the key transcription factors lin-1/ets and lin-1/forkhead, and the Mediator subunit mdt-/sur-, an essential effector of EGFR signaling output. We show that all four CKM subunits inhibit ectopic vulval cell fates in C. elegans. We demonstrate that the CKM catalytic subunit cdk- acts downstream of let-/egfr and mpk-1/erk in VPCs, in a kinase-dependent manner. Our data implicate cdk- as a corepressor for the LIN-1/Ets repressive transcription factor to inhibit EGFR signaling-induced transcription. Furthermore, our data indicate that vulval induction in
7 CKM mutants is independent of the mdt-/sur- coactivator, and instead require the Mediator tail module subunits mdt-1, mdt- and mdt- for induction of ectopic vulval cell fates.
8 Results cdk--dependent transcripts overlap with targets of a synmuv gene To define the role of the CKM in metazoan development, we compared transcriptional profiles of developmentally synchronized L larval stage cdk-(tm1) null mutants to wild-type N worms using microarrays (see Materials and Methods and Fig. S1A for cdk- mutant information). We found that genes were upregulated and 1 genes were downregulated more than two-fold in cdk- null mutants, representing ~.% of all C. elegans genes (Table S1). To identify cdk--dependent gene programs, we compared our lists of cdk- regulated genes to other gene lists using EASE (Hosack et al. 00; Engelmann et al. 0). The top hit among genes upregulated in cdk- mutants was a set of genes upregulated in lin-/retinoblastoma (RB) synmuv gene mutants (Kirienko and Fay 00), and lin--repressed genes also overlapped significantly with genes downregulated in cdk- mutants (Fig. A). Importantly, the mrna levels of lin- and the efl-1/dpl-1 transcription factor heterodimer that is repressed by LIN- were not altered in cdk- mutants (Fig. S1B), indicating that cdk- does not affect lin- target gene expression by altering the abundance of lin- or its partners. Together, these data suggest that cdk- could act in parallel to lin- as they regulate similar gene sets cdk- represses EGFR signaling-dependent primary vulval cell fate specification We next investigated whether cdk-, like mdt-1/dpy- (Moghal and Sternberg 00a), also represses vulval induction. Indeed, cdk- and cic-1(tm0) null mutants displayed a low penetrance Muv phenotype, as measured both by VPC induction analysis in L animals (Table 1) and by scoring the occurrence of ectopic vulval protrusions in adult worms (Fig. B; see statistical comparisons between L and adult Muv scores, Table S). cdk- and cic-1 appeared to function redundantly in vulva formation, as cdk-; cic-1 double mutants showed no significant increase in Muv penetrance compared to either single mutant (Fig. B). We then tested if the cdk- mutant Muv phenotype was associated with ectopic EGFR signaling-induced 1 vulval cell fates using an egl-1p::cfp reporter
9 (aris; (Yoo et al. 00)). In wild-type worms, we observed strong egl-1p::cfp expression in the 1 cell descendants, P.px, and occasional weak expression in the cell descendants, P.px or P.px (Fig. C), as reported (Yoo et al. 00). cdk- mutants expressed egl-1p::cfp strongly in P.ps and weakly in P.px and P.px, like wild type (Fig. C). However, some cdk- mutants exhibited egl- 1P::CFP expression in P.px or P.px that was equal in intensity to the expression level in the 1 cell descendants P.px, or exhibited ectopic induction of egl-1p::cfp in VPCs that normally adopt the non-vulval fate (Fig. C). The ectopic expression of egl-1::cfp suggested that the 1 cell fate had been derepressed in presumptive or non-vulval cells. This reflects that the low penetrance Muv phenotype of cdk- mutants is caused in part through derepressed ERK signaling output cdk- interacts genetically with the synmuv repressors of lin-/egf transcription The CKM subunit mdt-1/dpy- was previously shown to act downstream of let-/egfr to modulate vulva development (Moghal and Sternberg 00a). However, the developmental roles of all four CKM subunits are not equivalent (Loncle et al. 00), and our gene expression profiling suggested that cdk- might interact genetically with the synmuv genes to alter vulval cell fate decisions. As synmuv genes encode three redundant chromatin-modifying complexes, a Muv phenotype results when genes in any two complexes are simultaneously mutated (Myers and Greenwald 00; Cui et al. 00; Saffer et al. 0). We therefore studied the simultaneous inactivation of cdk- or cic-1 and representative synmuv class A (lin-1a), B (lin-1b), or C (trr-1) genes. Mutation or RNA interference (RNAi) depletion of all representative synmuv genes enhanced the Muv phenotype of cdk- mutants (Fig. B, Table 1). In addition, loss of the Ras GTPase-activating protein gene gap-1, which causes weak LET-0/Ras derepression, also enhanced the cdk- mutant Muv penetrance (Fig. B, Table 1). Our microarray analysis did not reveal significant downregulation of any known synmuv genes in cdk- mutants (Table S), indicating that cdk- does not simply regulate synmuv mrna levels. Taken
10 together, these results suggest that cdk- and cic-1 act redundantly with synmuv genes to repress vulval cell fates. We observed adjacent VPCs expressing a 1 cell fate marker in cdk- mutants (Fig. C), which is uncharacteristic of synmuv gene mutants. This phenotype instead suggests defects in Notch signaling, which inhibits adjacent 1 fates by inducing EGFR signaling inhibitor genes (Sternberg 1; Berset et al. 001; Yoo et al. 00; Chen and Greenwald 00). Therefore, we examined the expression of the Notch-inducible EGFR signaling inhibitor, lip-1/erk phosphatase, using a lip- 1P::GFP reporter (zhis; (Berset et al. 001)). We used the sensitized lin-1a mutant background to increase the frequency of ectopic VPC induction events. lin-1a single mutants expressed lip-1p::gfp strongly in P.px and P.px, but expression was weak or absent in other Pn.px cells (Fig. D), as reported for wild-type worms (Berset et al. 001). In contrast, some cdk-; lin-1a mutants lost strong lip-1p::gfp expression in P.px and P.px, consistent with loss of the fate (Fig. D). Furthermore, some cdk-; lin-1a mutants ectopically expressed lip-1p::gfp strongly in non-vulval P.p, P.p, or P.p, suggesting ectopic fates (Fig. D). Thus, cdk- mutants display hallmarks of both down- and upregulated Notch signaling, suggesting that CDK- action on the Notch pathway may occur indirectly via the EGFR signaling pathway upstream cdk- regulates lin-/egf transcription in the anchor cell We next tested if cdk- acts redundantly with the synmuv genes to repress lin-/egf transcription (Cui et al. 00; Saffer et al. 0). As the synmuv genes act primarily in the hypodermis to repress lin- transcription (Myers and Greenwald 00; Saffer et al. 0), derepression of lin- in synmuv double mutants is detectable by quantitative PCR in whole-animal preparations (Cui et al. 00). We used Taqman quantitative PCR analysis to quantify whole-animal lin- mrna levels in cdk-, lin-1a, and lin-1b single mutants, and in cdk-; lin-1a and cdk-; lin-1b double mutants; the lin-1ab(n0) mutant served as a positive control, as it is known to upregulate lin- expression
11 1 (Fig. A) (Cui et al. 00). Compared to wild-type worms, cdk- single mutants, cdk-; lin-1a double mutants, and cdk-; lin-1b double mutants showed no statistically significant change in lin- mrna levels (Fig. A). Thus, the enhanced Muv penetrance of cdk-; lin-1a and cdk-; lin-1b mutants compared to cdk- single mutants (Fig. B) likely does not arise from hypodermal lin- derepression. Next, we investigated if cdk- is required to regulate lin- transcription in the signal-emitting anchor cell, which would not be detectable in whole-animal quantitative PCR analysis. In line with this hypothesis, we observed expression of a transcriptional cdk-p::gfp reporter (hdex0) in the anchor cell (Fig. B). To assess lin- transcription in the anchor cell alone, we used a lin- anchor cell-specific enhancer element (ACEL) GFP reporter (syis; (Hwang and Sternberg 00)). We detected a small but significant upregulation of lin- ACEL reporter expression in cdk- mutants compared to wild-type worms at the L larval stage (Fig. C), suggesting that cdk- is required to repress lin-/egf transcription in the anchor cell cdk- acts downstream of mpk-1/erk to regulate VPC induction cell autonomously As the effect size of cdk- loss on lin-/egf anchor cell expression was small, and as mdt- 1/dpy- has been found to act downstream of let-/egfr (Moghal and Sternberg 00a), we next investigated cdk- s role in the EGFR signaling pathway downstream of lin-. We conducted genetic epistasis analyses with strong loss-of-function alleles of EGFR, let-(sy), and ERK, mpk-1(oz); both caused highly penetrant Vul phenotypes due to blockade of the EGFR-Ras-ERK pathway (Fig. D, Table 1). cdk- inactivation significantly rescued the Vul phenotype of let- or mpk-1 single mutants (Fig. D, Table 1). These data suggest that cdk- primarily acts downstream or parallel to mpk- 1/ERK to repress vulval cell fate specification by the EGFR signaling pathway. cdk- s position downstream of mpk-1/erk suggested a cell-autonomous role in VPCs (Fig. 1). Nuclear expression of the MDT-1/DPY- protein in VPCs and in the anchor cell had previously been observed, and gonad-independent vulval induction in mdt-1/dpy- mutants suggested an anchor cell-
12 independent role for MDT-1/DPY- (Moghal and Sternberg 00a). However, the tissue-specific requirements for MDT-1/DPY- in VPCs vs. the hypodermis, two important drivers of VPC cell fate (Fay and Yochem 00; Schmid and Hajnal 01), had not been tested. We used the lin-1a sensitized background to analyze tissue-specific requirements for cdk- in VPCs vs. the hypodermis. First, we demonstrated that a transgene expressing cdk- from its own promoter (cdk-(+)) rescued the cdk-; lin-1a mutant Muv phenotype compared to non-transgenic siblings (Fig. E). This transgene appeared to be broadly expressed and functional, as it rescued two additional phenotypes observed in cdk- mutants: decreased body length (Dumpy phenotype, Dpy) and the low brood size of the cdk- mutant (Fig. SA, B). Expression of cdk- from the lin-1 promoter (lin-1p::cdk-), which drives transgene expression in Pn.ps and some neurons (Tan et al. 1; Kishore and Sundaram 00), also significantly rescued the cdk-; lin-1a Muv phenotype (Fig. E). In contrast, expression of cdk- from the hypodermis-specific dpy- minimal promoter (dpy-p::cdk-) (Gilleard et al. 1) did not significantly rescue the Muv penetrance of cdk-; lin-1a mutant worms (Fig. E), although it was able to rescue the Dpy phenotype (Fig. SA). Unexpectedly, the lin-1p::cdk- transgene partially rescued the cdk-; lin-1a Dpy phenotype compared to non-transgenic worms, albeit to a lesser extent than cdk-(+) or dpy-p::cdk- (Fig. SA). In sum, these experiments provide evidence that cdk- is required cell-autonomously in VPCs but not in the hypodermis to suppress ectopic vulval induction cdk- activity is kinase-dependent We next addressed how cdk- functions downstream of mpk-1/erk. First, we studied CDK- s kinase requirement using a kinase-dead CDK-(D1A) transgene (CDK-(KD)). The D1A mutation is homologous to the previously reported D1A mutation in human CDK and the D0A mutation in budding yeast CDK/SRB, both of which result in loss of enzymatic activity (Liao et al. 1; Gold and Rice 1); however, we note that the kinase activity of C. elegans CDK-(D1A) has not been tested directly. CDK-(KD) did not rescue the cdk-; lin-1a mutant Muv phenotype, and 1
13 actually enhanced Muv penetrance (Fig. A), suggesting that kinase activity is required for transgenic rescue of the Muv phenotype of cdk- null mutants cdk- promotes lin-1/ets repressor activity Next, we investigated transcription factors that repress VPC induction, i.e. the ELK1/Ets-family transcription factor LIN-1 and the Forkhead-family transcription factor LIN-1 (Miller et al. 1; Beitel et al. 1). We hypothesized that the CKM may coregulate LIN-1 and/or LIN-1, and thus tested their genetic interactions with cdk-. Loss of cdk- did not enhance the lin-1 mutant Muv phenotype (Table 1); therefore, we did not investigate lin-1 further. In contrast, cdk- mutation strongly enhanced the low Muv penetrance caused by lin-1 RNAi depletion (Fig. B, Table 1). To corroborate the RNAi experiment, we also studied the lin-1(n10) mutant, which displays both reduction-of-function (ectopic vulval induction of P.p, P.p and P.p due to reduced lin-1 mrna levels) and gain-of-function (reduced vulval induction in P.p-P.p due to impaired LIN-1 ERK binding) phenotypes (Jacobs et al. 1). As seen in lin-1 RNAi worms, loss of cdk- significantly enhanced the Muv penetrance of the lin-1(n10) mutant (Fig. C, Table 1). Together, these results suggest that residual LIN-1 requires cdk- for efficient repression of vulval induction. To test if cdk- promotes transcriptional repression by LIN-1, we investigated whether the direct LIN-1 target gene lag- (Zhang and Greenwald 0) was derepressed in cdk- mutants. In wildtype animals, a lag-p(min)::yfp minimal promoter reporter (arex) is induced by EGFR signaling in P.p, whereas in lin-1 null mutants, it is ectopically induced in additional VPCs (Zhang and Greenwald 0). We again used the lin-1a sensitized background to study cdk- requirements for lag-p(min) repression. In all lin-1a single mutant animals examined, lag-p(min)::yfp exhibited a wild-type expression pattern, as expected (n=; Fig. D). In contrast, in cdk-; lin-1a mutants, lag- P(min)::YFP was occasionally ectopically expressed in VPCs other than P.p (in /1 animals; Fig. D). Thus, cdk- is partially required to repress a direct LIN-1 target gene. Taken together with the 1
14 requirement for cdk- for lin-1 repressor function in vulval induction, these data suggest that CDK- may function as a LIN-1 corepressor, perhaps acting redundantly with other corepressors The CKM subunit mdt-1/let-1 represses primary vulval fate specification Having shown that the enzymatic CKM subunit represses EGFR signaling-induced transcription in the C. elegans vulva, we next investigated whether the structural CKM subunits mdt-1/dpy- and mdt-1/let-1 (Tsai et al. 01) have similar molecular functions. C. elegans mdt-1/dpy- represses ectopic vulva formation downstream of let-/egfr (Moghal and Sternberg 00a), but whether and how it affects vulval cell fate specification is not understood. Thus, we investigated the requirements and mechanisms of structural CKM subunits in the EGFR signaling pathway, focusing primarily on mdt-1/let-1. We first investigated the vulval phenotypes of mdt-1/dpy- and mdt-1/let-1 mutants. mdt- 1/let-1 mutants exhibited a temperature sensitive Muv phenotype (Fig. A) of much higher penetrance than cdk- mutants (Fig. A). We observed a similarly high penetrance Muv phenotype in mdt-1/dpy- reduction-of-function mutants (Fig. A). We note that the os mutant used in this study shows higher Muv penetrance than the -% Muv penetrance reported for dpy-(sy) and dpy-(sy) mutants (Moghal and Sternberg 00a). os may cause stronger loss-of-function than these alleles due to truncation closer to the N-terminus and/or the presence of an additional missense mutation (Yoda et al. 00). Overall, these results demonstrate that mdt-1/dpy- and mdt-1/let-1 are more strongly required than cdk- and cic-1 to repress vulval induction. We next tested if mdt-1/let-1 adopts roles similar to cdk- in vulva development. mdt-1/let- 1 mutants induced egl-1p::cfp expression in / P.px cells examined (Fig. B). In addition, mdt-1/let-1 mutants displayed increased proportions of P.px and P.px weakly expressing egl- 1P::CFP compared to wild-type worms (Fig. B). Furthermore, mdt-1/let-1 mutants displayed strong egl-1p::cfp expression in some P.px and P.px cells, or ectopic expression of egl-1p::cfp 1
15 outside of P.px P.px, suggesting derepression of the 1 fate in these cells (Fig. B). Overall, these data indicated that the Muv phenotype of mdt-1/let-1 mutants was due in part to derepressed ERK signaling output. In support of a role similar to cdk- in regulation of vulval cell fate specification, mdt-1/let-1 interacted genetically with the synmuv genes and with lin-1/ets. Specifically, mdt-1/let-1 mutants interacted genetically with the class A synmuv transcriptional repressor lin-1a, and the class B synmuv gene lin-1b showed a similar trend (Fig. A). In addition, loss of mdt-1/let-1 significantly enhanced the Muv penetrance of lin-1(n10) mutant worms (Fig. C, Table 1), in line with a role for MDT-1/LET-1 as a LIN-1 corepressor. Thus, like cdk-, mdt-1/let-1 interacts genetically with transcriptional regulators in the EGFR signaling pathway. Finally, we investigated the genetic position of mdt-1/let-1 in the EGFR-Ras-ERK pathway. Similarly to cdk-, loss of mdt-1/let-1 suppressed the mpk-1 loss-of-function Vul phenotype (Fig. D, Table 1). Furthermore, the mdt-1/let-1 mutant Muv phenotype appeared epistatic to the mpk-1 mutant Vul phenotype, although this trend was not statistically significant (Fig. D, Table 1). Together, these data support a model in which the CKM acts downstream of mpk-1/erk to inhibit vulval cell fate specification The CKM restricts the specificity of core Mediator subunits As several Mediator subunits positively regulate vulva development (reviewed in (Grants et al. 01)), we hypothesized that the CKM might function in part by inhibiting these subunits. In wild-type worms, the mdt-/sur- tail module subunit is critical for vulva development downstream of Ras (Singh and Han 1), and it is required for activation of the EGFR signaling target gene lag- (Zhang and Greenwald 0); therefore, we tested how a mutation in a CKM subunit affected vulval induction in mdt-/sur- mutants. All mdt-/sur- single mutants examined exhibited a Vul phenotype (Fig. E, Table 1). VPC induction analysis of mdt-1/let-1 single mutants demonstrated that, in addition to 1
16 the Muv phenotype, some animals also displayed decreased proliferation of P.p or P.p (mild Vul; Fig. E, Table 1). Unexpectedly, mdt-1/let-1; mdt-/sur- mutants exhibited a significantly stronger Muv penetrance than mdt-1/let-1 single mutants, and loss of mdt-1/let-1 function significantly suppressed the Vul phenotype of mdt-/sur- single mutants (Fig. D, Table 1), suggesting that loss of mdt-1/let-1 derepresses vulval induction independently of mdt-/sur-. As seen in mdt-1/let-1 single mutants, mdt-1/let-1; mdt-/sur- double mutants showed strong egl-1p::cfp expression in some P.px and P.px cells, or ectopic expression of egl-1p::cfp outside of P.px P.px, suggesting derepression of the 1 fate (Fig. B). Overall, these data indicated that the Muv phenotype of mdt- 1/let-1; mdt-/sur- double mutants was due in part to derepressed ERK signaling output. Together, these findings indicate that loss of the CKM allows activation of EGFR signaling-driven cell fate specification independently of mdt-/sur- activity. Our results suggested that the CKM might influence Mediator subunit(s) other than mdt-/sur-. As Saccharomyces cerevisiae CDK inhibits the Mediator tail module triad composed of MED, MED, and MED1 (van de Peppel et al. 00; Gonzalez et al. 01), we hypothesized that their putative C. elegans orthologs MDT-, MDT-, and MDT-1 (Bourbon 00), might be targets for CKM inhibition. mdt-1 and mdt- knockdown had no effect on vulva formation in wild-type animals (i.e. causing neither Muv nor Vul phenotypes and displaying normal VPC induction; Table 1), but significantly reduced the Muv penetrance of mdt-1/let-1 mutants (Fig. A); mdt- RNAi caused a similar trend (Fig. A). This effect was specific to the tail module triad, as knockdown of the mdt-1.1 tail module subunit in fact increased the Muv penetrance of mdt-1/let-1 mutants (Fig. A). Furthermore, requirement for the tail module triad in ectopic vulva formation appeared to be specific to CKM mutants, as mdt-1 RNAi had no effect on ectopic vulva formation in lin-1(n10) mutants (Fig. B, Table 1). Thus, tail module triad activity appears to be derepressed in a CKM mutant, causing aberrant activation of vulval fate specification. 1
17 Next, we investigated whether the CKM modifies the target gene specificity of the triad. We used qpcr to quantify the expression of cdk--repressed genes identified by our microarray analysis (Table S1) in wild-type worms and cdk- mutants treated with empty vector (EV), mdt-1, mdt-, or mdt- RNAi. On EV RNAi, seven of nine genes tested were upregulated in cdk- mutants compared to wild-type worms, as expected (Fig. C). Upregulation of these cdk--repressed genes was strongly attenuated by mdt-1 depletion, whereas mdt- knockdown only affected fat-, and mdt- depletion caused no significant changes (Fig. C). Thus, for the genes investigated, induction in cdk- mutants appears to specifically require mdt-1, but not the other predicted tail module triad subunits. However, we cannot rule out unequal RNAi efficiency accounting for these differing requirements, although RNAi knockdown of all three genes appeared successful, as we observed partial sterility (not shown) consistent with the essential nature of these core Mediator subunits (Fernandez et al. 00; Sönnichsen et al. 00; Taubert et al. 00). Notably, only two cdk--repressed genes, acd- and fat-, displayed mdt-1 and/or mdt--dependent activation in wild-type worms (Fig. C). Thus, as seen in the genetic analysis of vulva induction, loss of cdk- appears to cause unrestrained tail module activity, i.e. mdt-1 activates novel target genes when cdk- is deleted. Finally, we investigated the molecular cause of unrestrained mdt-1 activity in CKM mutants. Loss of cdk- did not alter mrna levels of any triad subunits (Fig. S). Western blot analysis showed elevated MDT-1 protein levels in cdk- mutants compared to wild type (Fig. D). Taken together, these results demonstrate that cdk- is required for post-transcriptional regulation of MDT
18 Discussion EGFR signaling is critical for cell proliferation and cell fate determination in animal development. Several Mediator subunits positively or negatively regulate EGFR signaling-driven developmental processes (reviewed in (Grants et al. 01)), but pertinent mechanisms remain incompletely understood. Here, we used the well-characterized vulva development paradigm in C. elegans to delineate the role of the Mediator CKM module. Our results suggest a model whereby the CKM acts within the vulval precursor cells, in a kinase-dependent manner, to fine-tune EGFR transcriptional output by modulating two transcriptional regulators: the key downstream transcription factor LIN-1/Ets, and core Mediator (Fig. E). This model is based on four key observations: First, we demonstrate that the primary site of action for the CKM is in the VPCs, as cdk- and mdt-1 repress vulva formation downstream of mpk-1/erk, a key component of the EGFR signaling cascade inside VPCs, and cdk- expression in VPCs is sufficient for this repression. Second, cdk- repression of ectopic vulva formation is kinase-dependent. Third, the CKM appears to act as a corepressor of the Etsfamily transcription factor LIN-1, as loss of cdk- or mdt-1 enhances the ectopic vulval induction caused by lin-1 reduction-of-function, and cdk- is required for full repression of a direct LIN-1 target promoter. Fourth, ectopic vulva formation in mdt-1/let-1 is independent of the Mediator subunit mdt- /sur-, which is critical for EGFR signaling-driven transcription and vulval development in wildtype worms (Singh and Han 1; Zhang and Greenwald 0); instead, mdt-1/let-1 modulates the specificity of the tail module triad subunits mdt-1, mdt-, and mdt-, preventing aberrant activation of downstream transcription. By implicating all CKM subunits and by connecting the CKM to lin-1 and to core Mediator, our data substantially expand on the prior finding that loss of CKM subunit mdt- 1 caused ectopic vulva formation by unknown molecular mechanisms (Moghal and Sternberg 00a). Additionally, our genetic and molecular analysis provide first evidence that CKM-tail module crosstalk, akin to that seen in yeast Mediator (van de Peppel et al. 00; Gonzalez et al. 01), occurs 1
19 in metazoan Mediator, an important experimental finding as tail module subunit sequence conservation between species is extremely poor (Bourbon 00) The CKM inhibits vulva development in a kinase dependent manner We performed unbiased gene expression profiling to define gene programs that depend on cdk- in vivo, which revealed that only.% of C. elegans genes are regulated by cdk- (Fig. A, Table S1). This number agrees with studies in yeast, wherein SRB/CDK regulates only % of genes (Holstege et al. 1). Thus, CDK appears to be a gene program-specific transcriptional coregulator across species. Among cdk--dependent genes, we identified a significant overlap with genes regulated by lin- /RB, a synmuv transcriptional repressor (Fig. A) (Kirienko and Fay 00). We note that as synmuv genes act redundantly, lin- single mutants do not exhibit any defects in EGFR signaling or vulval induction (Myers and Greenwald 00; Cui et al. 00; Kirienko and Fay 00). Thus, the overlap between cdk-- and lin--dependent genes suggested that the CKM and lin- cooperate in multiple aspects of C. elegans development. Similarly, in Drosophila, CDK and RB act in parallel in the Wnt signaling pathway (Morris et al. 00). Therefore, we explored whether the CKM and synmuv genes cooperate in the EGFR signaling pathway to regulate C. elegans vulva development. Both cdk- and mdt-1/let-1 were required to repress C. elegans vulva formation, in a partially redundant manner with the synmuv genes (Figs B, A). However, cdk- did not act redundantly with the synmuv genes to repress lin-/egf transcription (Fig. A), suggesting that the CKM and the synmuv genes regulate EGFR signaling at different junctions, as discussed below. Comparing the vulval phenotypes of CKM mutants, we found evidence that cdk- and cic-1 act redundantly to repress vulval induction, as a cdk-; cic-1 double mutant displayed the same Muv penetrance as cdk- or cic-1 single mutants (Fig. B). In addition, we found that mdt-1/dpy- and mdt-1/let-1 were more strongly required to repress vulva development than cdk- or cic-1. In S. 1
20 cerevisiae Mediator, MED1 and MED1 enable CDK and Cyclin C docking to Mediator (Tsai et al. 01). Loss of MDT-1/DPY- or -1 in C. elegans may similarly disrupt CDK- and CIC-1 function, as well as considerably reducing the size of the CKM. Although CDK- s kinase activity is required to inhibit vulva development (Fig. A), this does not rule out the possibility that the CKM also employs kinase-independent steric mechanisms, as observed in other systems (Knuesel et al. 00). Thus, additional kinase-independent mechanisms could account for the stronger requirement for mdt- 1/dpy- and -1 in vulva development The CKM inhibits the primary vulval cell fate Vulva formation in C. elegans requires both EGFR and Notch signaling (Félix and Barkoulas 01), and human CDK represses Notch signaling-driven transcription by promoting turnover of the Notch intracellular domain (Fryer et al. 00). Therefore, we examined whether the vulval phenotypes in CKM mutants occur due to defects in EGFR signaling, Notch signaling, or both. Using an EGFR- Ras-ERK signaling-induced 1 cell fate reporter, we demonstrated that cdk- and mdt-1/let-1 are required to repress ectopic vulva formation in part by repressing the 1 cell fate (Figs C, B). Using a Notch signaling-induced cell fate reporter, we showed that cdk- is required to represses ectopic fates in non-vulval VPCs, as well as to promote the fate in P.p and P.p (Fig. D). However, cdk- action in the Notch pathway might occur indirectly in this context. EGFR signaling in P.p induces expression of Notch ligands, e.g. lag-, which promote the fate in the neighbouring cells, P.p and P.p (Chen and Greenwald 00). We observed evidence of possible cell fate transformations from to 1 in P.p or P.p in cdk- and mdt-1 mutants, as these cells occasionally exhibited strong expression of the 1 cell fate marker egl-1p::cfp (Figs C, B), expression of the EGFR signaling target gene lag- (Fig. D), or loss of the strong lip-1p::gfp expression characteristic of cells (Fig. D). It is possible that VPCs transformed to the 1 fate could then induce fates in neighbouring VPCs, accounting for our observation of ectopic cells. 0
21 CKM subunits have been implicated as regulators of canonical Wnt signaling (Zhang and Emmons 000; Firestein et al. 00; Morris et al. 00) and cell cycle quiescence (Clayton et al. 00), processes which also contribute to vulva development. Activation of Wnt signaling can bypass requirements for let-/egfr in vulva development (Gleason et al. 00). However, the Muv phenotype of mdt-1/dpy- mutants is independent of bar-1/β-catenin (Moghal and Sternberg 00b), suggesting that the CKM does not repress vulva development through the canonical Wnt signaling pathway. Deregulation of cell cycle quiescence can expand the VPC equivalence group, which are competent to form ectopic vulvae if presented with the appropriate signals (e.g. lin-1/notch gain-offunction employed by (Clayton et al. 00)). Although CKM subunits are required for VPC cell cycle quiescence (Clayton et al. 00), this alone is unlikely to account for the ectopic vulvae observed in these animals. First, the ectopic vulval invaginations observed in cdk- and mdt-1/let-1 animals while scoring VPC induction (Table 1) were positioned in the correct location for P.p, P.p and P.p descendants. Second, ectopic expression of 1 and cell fate markers in cdk- and mdt-1/let-1 mutants (Figs C, D, B) suggests that EGFR and/or Notch signaling indeed drives ectopic vulva formation in these mutants The CKM promotes LIN-1/Ets repressor activity We observed derepression of the lin-/egf anchor cell specific enhancer element (ACEL) in cdk- mutants (Fig. C), implicating cdk- as a novel repressor of lin-/egf transcription in the anchor cell. Albeit interesting, genetic epistasis analysis with let-/egfr and mpk-1/erk loss-of-function alleles clearly demonstrated that cdk- is primarily required downstream of mpk-1/erk to repress vulval induction (Fig. D). The let-(sy) mutant protein is ligand insensitive (Aroian and Sternberg 11; Aroian et al. 1); therefore, weak lin-/egf activation in the anchor cell due to loss of cdk- cannot account for the vulval phenotypes observed in cdk-; let-(sy) mutants (Fig. D). Furthermore, epistasis analysis with mpk-1/erk confirmed that cdk- acts downstream of the core 1
22 EGFR-Ras-ERK pathway to regulate vulval induction (Fig. D). In line with a position downstream of mpk-1/erk, we showed that cdk- is required in VPCs to suppress ectopic vulval induction (Fig. E). A previous report demonstrated that repression of vulval induction by the CKM subunit mdt-1/dpy- is gonad-independent, and thus anchor cell-independent, and that an MDT-1/DPY-::GFP transgene is expressed in VPCs (Moghal and Sternberg 00a), supporting a role for the CKM in VPCs. Downstream of mpk-1/erk, we found evidence that the CKM promotes LIN-1/Ets-mediated repression of vulval induction (Figs B, C, C), and that cdk- promotes transcriptional repression of a direct LIN-1 target, the lag-/notch ligand minimal promoter (Fig. D). The lag- minimal promoter contains activator and repressor elements, VPCact and VPCrep, that cooperatively restrict expression to P.p (Zhang and Greenwald 0). On its own, VPCact is sufficient to drive transcription in all VPCs (P.p-P.p) in a LIN-/EGF ligand-independent manner. VPCrep represses this basal VPCact-driven transcription in VPCs other than P.p, thereby restricting expression of the lag- minimal promoter to the 1 -fated VPC. VPCrep contains an Elk1 consensus site, which is bound by LIN-1 in vitro (Miley et al. 00), and requires lin-1/ets for repression of transcription in VPCs other than P.p (Zhang and Greenwald 0). Our results indicate that cdk- is partially required for transcriptional repression of the lag- minimal promoter (Fig. D), suggesting that the CKM promotes LIN-1-mediated repression at VPCrep. An alternative explanation for the ectopic expression of the lag- minimal promoter observed in cdk- mutants is that the CKM might inhibit a factor that activates transcription through VPCact. The transcription factor(s) that act at VPCact remain poorly defined; however, the mdt-/sur- Mediator subunit is required for VPCact-driven transcription in P.p-P.p (Zhang and Greenwald 0). As we demonstrated that vulval induction in mdt-1/let-1 mutants does not require mdt-/sur- (Figs B, E), this implies that the CKM likely does not inhibit MDT-/SUR- activity at VPCact. Overall, our findings suggest that the CKM may act as a corepressor for LIN-1. In murine embryonic stem cells, Mediator recruitment is important for transcriptional activation by Ets factors, e.g. Elk1 (Stevens et al. 00; Balamotis et al. 00). In this context, activation of Elk1
23 by ERK phosphorylation promotes binding to Mediator in a MED/Sur-dependent manner (Stevens et al. 00). Similarly, in a colon cancer cell line, CDK promotes transcriptional elongation of serum response immediate early genes, which are targeted by multiple transcription factors including Elk1 (Donner et al. 0). However, the role of Mediator in transcriptional repression by an Ets factor has not previously been explored. In the absence of ERK phosphorylation, Ets factors, e.g. LIN-1, can promote transcriptional repression of target genes (Jacobs et al. 1; Zhang and Greenwald 0). Although the SinA-HDAC-1 corepressor complex has been implicated in an epigenetic mechanism that attenuates transcriptional activation by ERK-phosphorylated Elk1(Yang et al. 001), to our knowledge, corepressors of Ets factor-mediated transcriptional repression have not previously been identified. This report provides evidence that the Mediator CKM is required for repression by Ets factors, representing an advance in our understanding of Ets factor repressive mechanisms. Our findings are also of potential clinical interest, as the human CKM is implicated in tumorigenesis (Firestein et al. 00; Donner et al. 0; Mäkinen et al. 0; Lim et al. 01). Loss of MED1 causes cellular resistance to chemotherapeutic agents that inhibit activated BRAF, the human ERK kinase kinase (Shalem et al. 01); this suggests that MED1 represses EGFR signaling downstream of BRAF, in line with our findings for the C. elegans CKM. Furthermore, recurrent MED1 mutations are implicated in uterine leiomyomas and breast fibroadenomas (Mäkinen et al. 0; Lim et al. 01; Mittal et al. 01), but the pathogenic mechanisms of these mutations have not been fully elucidated. Investigation of these mutations using the C. elegans vulva development paradigm may provide insight into their mode of action. 1 The CKM restrains the core Mediator tail module triad Epistatic relationships between Mediator subunits have been identified in S. cerevisiae (van de Peppel et al. 00; Gonzalez et al. 01), but intra-mediator regulation has not been demonstrated in metazoans. Previous studies (reviewed in (Grants et al. 01)) and our data show that several core
24 Mediator subunits promote C. elegans vulva development, whereas CKM subunits inhibit this process. This suggested that intra-mediator regulation might coordinate gene expression downstream of the EGFR-Ras-ERK signaling pathway that drives vulva development. Initially, we hypothesized that the CKM may oppose mdt-/sur--mediated activation of EGFR signaling, as mdt-/sur- is required for vulval induction and activation of EGFR signaling-induced transcription, e.g. lag- (Singh and Han 1; Zhang and Greenwald 0). Unexpectedly, loss of mdt-1/let-1 circumvented the requirement for mdt-/sur- in vulval induction (Fig. E). We therefore explored regulatory interactions between the CKM and the metazoan orthologs of S. cerevisiae MED, MED, and MED1, which are subject to inhibitory post-translational regulation by CDK (van de Peppel et al. 00; Gonzalez et al. 01). Sequence conservation is weak between yeast MED and MED and their putative metazoan homologs, MED and MED, respectively (Bourbon 00). Whether MED and MED function as part of the tail module remains unclear, as structural and biochemical studies locate these subunits between the head and tail modules (Sato et al. 00; Tsai et al. 01). Thus, we were intrigued to find that vulva formation in a C. elegans CKM mutant required mdt-1, mdt-, and mdt- (Fig. A). This requirement appeared specific to the tail module triad, as neither mdt-1.1/med1 nor mdt-/sur- was required for vulval induction in CKM mutants (Figs E, A). Furthermore, the triad did not appear to be generally required for ectopic vulval induction in animals with a wild-type CKM, as mdt-1 knockdown had no effect on ectopic vulval induction in lin-1(n10) mutants (Fig. B). Together, these findings suggest that the CKM restrains triad activity, preventing it from aberrantly activating vulval induction. Gene expression analysis in cdk- mutants identified a requirement for mdt-1, but little or no requirement for mdt- or mdt-, in transcriptional activation of cdk--repressed genes (Fig. C). In the S. cerevisiae tail module triad, both MED and MED are required for overexpression of CDK- repressed genes in CDK mutants, but the requirement for MED1 has not been tested directly (van de Peppel et al. 00; Gonzalez et al. 01). These requirements might be explained by the fact that both
25 MED and MED are necessary to anchor the triad to the tail module (Myers et al. 1; van de Peppel et al. 00; Gonzalez et al. 01). Similar requirements may not in exist in metazoan Mediator, as human Mediator displays more extensive structural contacts between the head and tail modules (Tsai et al. 01), which may result in redundancy for some tail module subunits. Investigating the regulatory relationship between CDK- and MDT-1 further, we found that cdk- is required for post-transcriptional negative regulation of MDT-1, as MDT-1 protein but not mrna levels increase in cdk- mutants (Fig. D). This regulatory relationship resembles that seen in yeast where the three triad subunits MED, MED, and MED1 are negatively regulated posttranslationally by CDK-driven phosphorylation of MED, promoting ubiquitin-proteasome dependent turnover of all three triad subunits (Gonzalez et al. 01). It will be interesting to delineate whether the metazoan CKM regulates MDT-1 protein levels directly, e.g. by phosphorylation leading to ubiquitinmediated degradation, or indirectly through action upon other Mediator subunits. In summary, our findings suggest that the Mediator CKM represses EGFR-Ras-ERK signalingdriven cell fate specification in C. elegans by regulating repressor activity of an Ets-family transcription factor and by promoting specificity of Mediator tail module subunits. 1
26 Materials and Methods Microarrays and data analysis Microarray gene expression profiling was performed at the UCSF SABRE Functional Genomics Facility. We used Agilent C. elegans (V) xk Gene Expression Microarrays (#G1F- 001) and single color labeling. Total RNA was extracted from developmentally synchronized mid- L stage worms as assessed by vulval morphology (wild-type N worms and cdk-(tm1) mutants), as described (Taubert et al. 00). RNA quality was assessed on an Agilent 0 Bioanalyzer using a Pico Chip (Agilent). RNA was amplified and labeled with Cy-CTP using the Agilent low RNA input fluorescent linear amplification kit. Labeled crna was assessed using the Nanodrop ND-0, and equal amounts of Cy labeled target were hybridized to the microarrays for 1 hrs, according to the manufacturers protocol. Arrays were scanned using the Agilent microarray scanner and raw signal intensities were extracted with Feature Extraction v.1 software. The dataset was normalized using quantile normalization (Bolstad et al. 00). No background subtraction was performed, and median feature pixel intensity was used as raw signal before normalization. All arrays were of good quality and had similar foreground and background signal distributions for both mrna and control probes. This suggests that quantile normalization is appropriate. To identify differentially expressed genes, a linear model was fit to the comparison to estimate the mean M values and calculate moderated t-statistic, B statistic, false discovery rate, and p-value for each gene. Adjusted p-values (AdjP) were produced as described (Holm 1). All procedures were carried out using functions in the R package limma in Bioconductor (Gentleman et al. 00; Smyth 00). Using this approach, we identified a total of 10 spots with an AdjP < 0.0 and a fold-change (representing 1 downregulated and upregulated genes) (Table S1). Microarray data have been deposited in Gene Expression Omnibus (GSE0). Differentially expressed genes were compared to published gene expression datasets using EASE (Hosack et al. 00). For best comparison to our data, we reanalyzed published lin- data
27 (Kirienko and Fay 00) to define a set of genes deregulated two-fold or more in L larvae, yielding 1 downregulated and upregulated genes. We compared this set to our cdk- targets and calculated the significance of the overlap using Fisher s exact test C. elegans strains, culture, and genetic methods C. elegans strains were cultured as described (Brenner, 1) at 0 C or C, as indicated. We used nematode growth medium (NGM)-lite (0.% NaCl, 0.% tryptone, 0.% KH PO, 0.0% K HPO ) agar plates seeded with Escherichia coli strain OP0 unless otherwise indicated. Strains are listed in Table S. Wild type was Bristol N. cdk-(tm1) and cic-1(tm0) are likely null alleles that abolish cdk- expression (Fig. S1B) and cic-1 function, respectively (see also (Steimel et al. 01)). For allele details, see mdt-1/dpy- mutants were identified as Dpy, GFP-negative progeny of rescued dpy-(os); osex[dpy-(+)] mothers, and homozygous mdt-1/let-1 mutants were identified as Dpy, GFP-negative progeny of balanced let- 1(mn1)/mIn1 mothers VPC induction VPC induction was scored as described (Han et al. 10), in synchronous mid-l animals under DIC optics at 00X magnification. In wild-type animals, P.p, P.p, and P.p are induced to give a VPC induction score of.0. In Vul animals, these VPCs are not fully induced (VPC induction <.0); in Muv animals, P.p, P.p, or P.p are induced (VPC induction >.0). 1 Multivulva and vulvaless phenotype penetrance Muv and Vul morphologies have been described (Horvitz and Sulston 10; Sulston and Horvitz 11). To facilitate scoring a large number of worms to accurately assess low-penetrance phenotypes, Muv phenotype penetrance was scored in synchronous day 1 adult animals in a dissection
28 microscope at 00x magnification (mdt-1/let-1 mutants) or x magnification (all other strains). To corroborate Muv penetrances scored in adult animals, we also conducted VPC induction analysis in L animals (see above). To assess Vul phenotypes, both Vul and Muv penetrances were extrapolated from VPC induction scores: animals were scored as Vul if VPC induction <.0 in P.p-P.p, and were scored as Muv if VPC induction occurred in P.p, P.p or P.p; using these criteria, animals were occasionally scored as simultaneous Vul and Muv RNA isolation and quantitative real-time PCR Total RNA was extracted from developmentally synchronized mid-l stage worms as assessed by vulval morphology. RNA isolation and qpcr were performed as described (Goh et al. 01). We used t-tests (two-tailed, equal variance) to calculate statistical significance of gene expression changes between mutants (Gaussian distribution). qpcr primers were designed with Primerweb (bioinfo.ut.ee/primer/) and tested on serial cdna dilutions to analyze PCR efficiency (primer sequences in Table S), except lin- (analyzed by TaqMan assay, Invitrogen #, Assay ID Ce011_m1) Fluorescent reporter analysis Synchronous worms were imaged using DIC optics and fluorescence microscopy on a Zeiss Axioplan microscope. Analysis of fluorescence intensity was conducted using ImageJ software, normalizing for cell size and background fluorescence. 1 Generation of transgenic rescue strains cdk- rescue transgenes (steex, -) or cdk-p::gfp were generated by gonad microinjection of a mixture of 0 ng/µl rescue plasmid (cdk-(+), SPD; lin-1p::cdk-, SPD; dpy-p::cdk-, SPD; cdk-(kd), SPD; cdk-p::gfp, SPD0), ng/µl pcfj0[myo-
29 p::mcherry], and ng/µl ppd. empty vector into N worms, then selecting transgenic mcherry-positive progeny. These were then crossed to cdk- and/or cdk-; lin-1a mutants (Table S). Cloning primer sequences are provided (Table S). 1 Feeding RNAi knockdown Feeding RNAi was performed as described (Goh et al. 01), with the following modifications: synchronous mid-l hermaphrodites were allowed to lay eggs at 0 C overnight on RNAi plates (Ahringer library -well format; mdt-1: plate, well C0; lin-1:, G0; Vidal library -well format: mdt-: GHR-0@H0; mdt-: GHR-00@D0; all clones were sequenced to confirm identity; negative control was empty vector L0), after which embryos were isolated by bleach treatment and transferred to fresh RNAi plates. F1 progeny were grown on RNAi plates (0 C or C) until they reached the desired developmental stage Western blot Immunoblot using standard lysis, SDS-PAGE and Western blot techniques was performed, with α-mdt-1 (Taubert et al. 00) and α-gapdh (Calbiochem, CB01) antibodies, as described (Goh et al. 01). 1 1
30 Acknowledgments We thank all Taubert lab members for critical discussions, Shirley Chen for help with experiments, Drs. R. Barbeau, C. Eisley, A. Barczak, and D. Erle from the Sandler Asthma Basic Research (SABRE) Center Functional Genomics Core Facility (UCSF) for help with microarray gene expression profiling, Dr. J. Ewbank for EASE analysis of gene expression arrays, Dr. J. Escobar for advice on scoring VPC induction, Dr. I. Greenwald for GS0 arex[lag-p(min)::yfp], Dr. S. Mitani for cdk-(tm1) and cic-1(tm0) mutants, and Drs. D. Moerman and J. Ward for comments on the manuscript. Some strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P0 OD00). 1 Competing Interests The authors declare no competing financial interests Author Contributions Conceived and designed experiments: JMG HS ST. Performed experiments: JMG LTLY AY CCY DQM HS ST. Analyzed data: JMG AY HO DQM HS ST. Contributed reagents/materials/analysis tools: JMG AY HO HS. Wrote the paper: JMG HS ST Funding This work was supported by grants from CIHR (MOP-1), NSERC (RGPIN -1), CFI (all to ST), and Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (to HS). JMG was supported by Vanier CGS, NSERC CGS- M, CFRI, and UBC scholarships, and ST by a Canada Research Chair. 0
31 References Aroian R. V., Koga M., Mendel J. E., Ohshima Y., Sternberg P. W., 10 The let- gene necessary for Caenorhabditis elegans vulval induction encodes a tyrosine kinase of the EGF receptor subfamily. Nature :. Aroian R. V., Sternberg P. W., 11 Multiple functions of let-, a Caenorhabditis elegans receptor tyrosine kinase gene required for vulval induction. Genetics 1: 1. Aroian R. V., Lesa G. M., Sternberg P. W., 1 Mutations in the Caenorhabditis elegans let- EGFR-like gene define elements important for cell-type specificity and function. EMBO J. 1: Balamotis M. A., Pennella M. A., Stevens J. L., Wasylyk B., Belmont A. S., Berk A. J., 00 Complexity in transcription control at the activation domain-mediator interface. Sci. Signal. : ra Baselga J., Swain S. M., 00 Novel anticancer targets: revisiting ERBB and discovering ERBB. Nat. Rev. Cancer : Beitel G. J., Tuck S., Greenwald I., Horvitz H. R., 1 The Caenorhabditis elegans gene lin-1 encodes an ETS-domain protein and defines a branch of the vulval induction pathway. Genes Dev. : Berset T., Hoier E. F., Battu G., Canevascini S., Hajnal A., 001 Notch inhibition of RAS signaling through MAP kinase phosphatase LIP-1 during C. elegans vulval development. Science 1:. 1
32 Bolstad B. M., Irizarry R. A., Astrand M., Speed T. P., 00 A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinforma. Oxf. Engl. 1: Bourbon H.-M., Aguilera A., Ansari A. Z., Asturias F. J., Berk A. J., Bjorklund S., Blackwell T. K., Borggrefe T., Carey M., Carlson M., Conaway J. W., Conaway R. C., Emmons S. W., Fondell J. D., Freedman L. P., Fukasawa T., Gustafsson C. M., Han M., He X., Herman P. K., Hinnebusch A. G., Holmberg S., Holstege F. C., Jaehning J. A., Kim Y.-J., Kuras L., Leutz A., Lis J. T., Meisterernest M., Naar A. M., Nasmyth K., Parvin J. D., Ptashne M., Reinberg D., Ronne H., Sadowski I., Sakurai H., Sipiczki M., Sternberg P. W., Stillman D. J., Strich R., Struhl K., Svejstrup J. Q., Tuck S., Winston F., Roeder R. G., Kornberg R. D., 00 A unified nomenclature for protein subunits of mediator complexes linking transcriptional regulators to RNA polymerase II. Mol. Cell 1:. 1 1 Bourbon H.-M., 00 Comparative genomics supports a deep evolutionary origin for the large, four- module transcriptional mediator complex. Nucleic Acids Res. : Chen N., Greenwald I., 00 The lateral signal for LIN-1/Notch in C. elegans vulval development comprises redundant secreted and transmembrane DSL proteins. Dev. Cell : Clayton J. E., Heuvel S. J. L. van den, Saito R. M., 00 Transcriptional control of cell-cycle quiescence during C. elegans development. Dev. Biol. 1: Cui M., Chen J., Myers T. R., Hwang B. J., Sternberg P. W., Greenwald I., Han M., 00 SynMuv genes redundantly inhibit lin-/egf expression to prevent inappropriate vulval induction in C. elegans. Dev. Cell :.
33 Donner A. J., Szostek S., Hoover J. M., Espinosa J. M., 00 CDK Is a Stimulus-Specific Positive Coregulator of p Target Genes. Mol. Cell : 1. Donner A. J., Ebmeier C. C., Taatjes D. J., Espinosa J. M., 0 CDK is a positive regulator of transcriptional elongation within the serum response network. Nat. Struct. Mol. Biol. 1: Engelmann I., Griffon A., Tichit L., Montañana-Sanchis F., Wang G., Reinke V., Waterston R. H., Hillier L. W., Ewbank J. J., 0 A comprehensive analysis of gene expression changes provoked by bacterial and fungal infection in C. elegans. PloS One : e10. Fay D. S., Yochem J., 00 The SynMuv genes of Caenorhabditis elegans in vulval development and beyond. Dev. Biol. 0: 1. 1 Félix M.-A., Barkoulas M., 01 Robustness and flexibility in nematode vulva development. Trends Genet. TIG : Fernandez A. G., Gunsalus K. C., Huang J., Chuang L.-S., Ying N., Liang H.-L., Tang C., Schetter A. J., Zegar C., Rual J.-F., Hill D. E., Reinke V., Vidal M., Piano F., 00 New genes with roles in the C. elegans embryo revealed using RNAi of ovary-enriched ORFeome clones. Genome Res. 1: Firestein R., Bass A. J., Kim S. Y., Dunn I. F., Silver S. J., Guney I., Freed E., Ligon A. H., Vena N., Ogino S., Chheda M. G., Tamayo P., Finn S., Shrestha Y., Boehm J. S., Jain S., Bojarski E., Mermel C., Barretina J., Chan J. A., Baselga J., Tabernero J., Root D. E., Fuchs C. S., Loda M., Shivdasani R. A., Meyerson M., Hahn W. C., 00 CDK is a colorectal cancer oncogene that regulates [bgr]-catenin activity. Nature : 1.
34 Fryer C. J., White J. B., Jones K. A., 00 Mastermind recruits CycC:CDK to phosphorylate the Notch ICD and coordinate activation with turnover. Mol. Cell 1: 0 0. Gentleman R. C., Carey V. J., Bates D. M., Bolstad B., Dettling M., Dudoit S., Ellis B., Gautier L., Ge Y., Gentry J., Hornik K., Hothorn T., Huber W., Iacus S., Irizarry R., Leisch F., Li C., Maechler M., Rossini A. J., Sawitzki G., Smith C., Smyth G., Tierney L., Yang J. Y. H., Zhang J., 00 Bioconductor: open software development for computational biology and bioinformatics. Genome Biol. : R0. Gilleard J. S., Barry J. D., Johnstone I. L., 1 cis regulatory requirements for hypodermal cell- specific expression of the Caenorhabditis elegans cuticle collagen gene dpy-. Mol. Cell. Biol. 1: Gleason J. E., Korswagen H. C., Eisenmann D. M., 00 Activation of Wnt signaling bypasses the requirement for RTK/Ras signaling during C. elegans vulval induction. Genes Dev. 1: Goh G. Y. S., Martelli K. L., Parhar K. S., Kwong A. W. L., Wong M. A., Mah A., Hou N. S., Taubert S., 01 The conserved Mediator subunit MDT-1 is required for oxidative stress responses in Caenorhabditis elegans. Aging Cell 1: Gold M. O., Rice A. P., 1 Targeting of CDK to a promoter-proximal RNA element demonstrates catalysis-dependent activation of gene expression. Nucleic Acids Res. : Gonzalez D., Hamidi N., Del Sol R., Benschop J. J., Nancy T., Li C., Francis L., Tzouros M., Krijgsveld J., Holstege F. C. P., Conlan R. S., 01 Suppression of Mediator is regulated by Cdk-dependent Grr1 turnover of the Med coactivator. Proc. Natl. Acad. Sci. U. S. A. 1: 00 0.
35 Grants J. M., Goh G. Y. S., Taubert S., 01 The Mediator complex of Caenorhabditis elegans: insights into the developmental and physiological roles of a conserved transcriptional coregulator. Nucleic Acids Res. :. Han M., Aroian R. V., Sternberg P. W., 10 The let-0 locus controls the switch between vulval and nonvulval cell fates in Caenorhabditis elegans. Genetics 1: 1. Hill R. J., Sternberg P. W., 1 The gene lin- encodes an inductive signal for vulval development in C. elegans. Nature : 0. Holm S., 1 A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. : 0. Holstege F. C., Jennings E. G., Wyrick J. J., Lee T. I., Hengartner C. J., Green M. R., Golub T. R., Lander E. S., Young R. A., 1 Dissecting the regulatory circuitry of a eukaryotic genome. Cell : Horvitz H. R., Sulston J. E., 10 Isolation and genetic characterization of cell-lineage mutants of the nematode Caenorhabditis elegans. Genetics :. 1 1 Hosack D. A., Dennis G. Jr, Sherman B. T., Lane H. C., Lempicki R. A., 00 Identifying biological themes within lists of genes with EASE. Genome Biol. : R Hwang B. J., Sternberg P. W., 00 A cell-specific enhancer that specifies lin- expression in the C. elegans anchor cell for vulval development. Dev. Camb. Engl. : Jacobs D., Beitel G. J., Clark S. G., Horvitz H. R., Kornfeld K., 1 Gain-of-Function Mutations in the Caenorhabditis elegans lin-1 ETS Gene Identify a C-Terminal Regulatory Domain Phosphorylated by ERK MAP Kinase. Genetics 1: 10 1.
36 Katz W. S., Hill R. J., Clandinin T. R., Sternberg P. W., 1 Different levels of the C. elegans growth factor LIN- promote distinct vulval precursor fates. Cell : 0. Kirienko N. V., Fay D. S., 00 Transcriptome profiling of the C. elegans Rb ortholog reveals diverse developmental roles. Dev. Biol. 0:. Kishore R. S., Sundaram M. V., 00 ced- Rac and mig- function redundantly and act with unc- trio to control the orientation of vulval cell divisions and migrations in Caenorhabditis elegans. Dev. Biol. 1:. Knuesel M. T., Meyer K. D., Bernecky C., Taatjes D. J., 00 The human CDK subcomplex is a molecular switch that controls Mediator coactivator function. Genes Dev. : 1. Kwon J. Y., Lee J., 001 Biological significance of a universally conserved transcription mediator in metazoan developmental signaling pathways. Dev. Camb. Engl. 1: Liao S. M., Zhang J., Jeffery D. A., Koleske A. J., Thompson C. M., Chao D. M., Viljoen M., Vuuren H. J. van, Young R. A., 1 A kinase-cyclin pair in the RNA polymerase II holoenzyme. Nature : Lim W. K., Ong C. K., Tan J., Thike A. A., Ng C. C. Y., Rajasegaran V., Myint S. S., Nagarajan S., Nasir N. D. M., McPherson J. R., Cutcutache I., Poore G., Tay S. T., Ooi W. S., Tan V. K. M., Hartman M., Ong K. W., Tan B. K. T., Rozen S. G., Tan P. H., Tan P., Teh B. T., 01 Exome sequencing identifies highly recurrent MED1 somatic mutations in breast fibroadenoma. Nat. Genet. 0 1 Loncle N., Boube M., Joulia L., Boschiero C., Werner M., Cribbs D. L., Bourbon H.-M., 00 Distinct roles for Mediator Cdk module subunits in Drosophila development. EMBO J. :.
37 Mäkinen N., Mehine M., Tolvanen J., Kaasinen E., Li Y., Lehtonen H. J., Gentile M., Yan J., Enge M., Taipale M., Aavikko M., Katainen R., Virolainen E., Böhling T., Koski T. A., Launonen V., Sjöberg J., Taipale J., Vahteristo P., Aaltonen L. A., 0 MED1, the mediator complex subunit 1 gene, is mutated at high frequency in uterine leiomyomas. Science :. Malik S., Roeder R. G., 0 The metazoan Mediator co-activator complex as an integrative hub for transcriptional regulation. Nat. Rev. Genet. : 1. Miley G. R., Fantz D., Glossip D., Lu X., Saito R. M., Palmer R. E., Inoue T., Heuvel S. van den, Sternberg P. W., Kornfeld K., 00 Identification of Residues of the Caenorhabditis elegans LIN-1 ETS Domain That Are Necessary for DNA Binding and Regulation of Vulval Cell Fates. Genetics 1: Miller L. M., Gallegos M. E., Morisseau B. A., Kim S. K., 1 lin-1, a Caenorhabditis elegans HNF- /fork head transcription factor homolog, specifies three alternative cell fates in vulval development. Genes Dev. :. 1 1 Mittal P., Shin Y.-H., Yatsenko S. A., Castro C. A., Surti U., Rajkovic A., 01 Med1 gain-of- function mutation causes leiomyomas and genomic instability. J. Clin. Invest. 1 1 Moghal N., Sternberg P. W., 00a A component of the transcriptional mediator complex inhibits RAS-dependent vulval fate specification in C. elegans. Dev. Camb. Engl. :. 1 1 Moghal N., Sternberg P. W., 00b The epidermal growth factor system in Caenorhabditis elegans. Exp. Cell Res. : Morris E. J., Ji J.-Y., Yang F., Di Stefano L., Herr A., Moon N.-S., Kwon E.-J., Haigis K. M., Naar A. M., Dyson N. J., 00 EF1 represses [bgr]-catenin transcription and is antagonized by both prb and CDK. Nature :.
38 Myers L. C., Gustafsson C. M., Hayashibara K. C., Brown P. O., Kornberg R. D., 1 Mediator protein mutations that selectively abolish activated transcription. Proc. Natl. Acad. Sci. U. S. A. :. Myers T. R., Greenwald I., 00 lin- Rb acts in the major hypodermis to oppose ras-mediated vulval induction in C. elegans. Dev. Cell : 1. Nemet J., Jelicic B., Rubelj I., Sopta M., 01 The two faces of Cdk, a positive/negative regulator of transcription. Biochimie :. Normanno N., De Luca A., Bianco C., Strizzi L., Mancino M., Maiello M. R., Carotenuto A., De Feo G., Caponigro F., Salomon D. S., 00 Epidermal growth factor receptor (EGFR) signaling in cancer. Gene : Peppel J. van de, Kettelarij N., Bakel H. van, Kockelkorn T. T. J. P., Leenen D. van, Holstege F. C. P., 00 Mediator Expression Profiling Epistasis Reveals a Signal Transduction Pathway with Antagonistic Submodules and Highly Specific Downstream Targets. Mol. Cell 1:. 1 1 Poss Z. C., Ebmeier C. C., Taatjes D. J., 01 The Mediator complex and transcription regulation. Crit. Rev. Biochem. Mol. Biol. : Saffer A. M., Kim D. H., Oudenaarden A. van, Horvitz H. R., 0 The Caenorhabditis elegans synthetic multivulva genes prevent ras pathway activation by tightly repressing global ectopic expression of lin- EGF. PLoS Genet. : e Sato S., Tomomori-Sato C., Banks C. A. S., Parmely T. J., Sorokina I., Brower C. S., Conaway R. C., Conaway J. W., 00 A mammalian homolog of Drosophila melanogaster transcriptional coactivator intersex is a subunit of the mammalian Mediator complex. J. Biol. Chem. : 1.
39 Schmid T., Hajnal A., 01 Signal transduction during C. elegans vulval development: a NeverEnding story. Curr. Opin. Genet. Dev. : 1. Shalem O., Sanjana N. E., Hartenian E., Shi X., Scott D. A., Mikkelsen T. S., Heckl D., Ebert B. L., Root D. E., Doench J. G., Zhang F., 01 Genome-scale CRISPR-Cas knockout screening in human cells. Science :. Singh N., Han M., 1 sur-, a novel gene, functions late in the let-0 ras-mediated signaling pathway during Caenorhabditis elegans vulval induction. Genes Dev. : 1. Smyth G. K., 00 Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. : Article Sönnichsen B., Koski L. B., Walsh A., Marschall P., Neumann B., Brehm M., Alleaume A.-M., Artelt J., Bettencourt P., Cassin E., Hewitson M., Holz C., Khan M., Lazik S., Martin C., Nitzsche B., Ruer M., Stamford J., Winzi M., Heinkel R., Röder M., Finell J., Häntsch H., Jones S. J. M., Jones M., Piano F., Gunsalus K. C., Oegema K., Gönczy P., Coulson A., Hyman A. A., Echeverri C. J., 00 Full-genome RNAi profiling of early embryogenesis in Caenorhabditis elegans. Nature : Steimel A., Suh J., Hussainkhel A., Deheshi S., Grants J. M., Zapf R., Moerman D. G., Taubert S., Hutter H., 01 The C. elegans CDK Mediator module regulates axon guidance decisions in the ventral nerve cord and during dorsal axon navigation. Dev. Biol. 1 0 Sternberg P. W., Horvitz H. R., 1 Pattern formation during vulval development in C. elegans. Cell : 1. 1 Sternberg P. W., 1 Lateral inhibition during vulval induction in Caenorhabditis elegans. Nature : 1.
40 Sternberg P. W., Horvitz H. R., 1 The combined action of two intercellular signaling pathways specifies three cell fates during vulval induction in C. elegans. Cell :. Stevens J. L., Cantin G. T., Wang G., Shevchenko A., Shevchenko A., Berk A. J., 00 Transcription control by E1A and MAP kinase pathway via Sur mediator subunit. Science :. Sulston J. E., Horvitz H. R., 11 Abnormal cell lineages in mutants of the nematode Caenorhabditis elegans. Dev. Biol. : 1. Tan P. B., Lackner M. R., Kim S. K., 1 MAP kinase signaling specificity mediated by the LIN-1 Ets/LIN-1 WH transcription factor complex during C. elegans vulval induction. Cell : 0. 1 Taubert S., Van Gilst M. R., Hansen M., Yamamoto K. R., 00 A Mediator subunit, MDT-1, integrates regulation of fatty acid metabolism by NHR--dependent and -independent pathways in C. elegans. Genes Dev. 0:. 1 1 Taubert S., Hansen M., Van Gilst M. R., Cooper S. B., Yamamoto K. R., 00 The Mediator subunit MDT-1 confers metabolic adaptation to ingested material. PLoS Genet. : e Tsai K.-L., Sato S., Tomomori-Sato C., Conaway R. C., Conaway J. W., Asturias F. J., 01 A conserved Mediator-CDK kinase module association regulates Mediator-RNA polymerase II interaction. Nat. Struct. Mol. Biol Tsai K.-L., Tomomori-Sato C., Sato S., Conaway R. C., Conaway J. W., Asturias F. J., 01 Subunit architecture and functional modular rearrangements of the transcriptional mediator complex. Cell 1: 1. 0
41 Tuck S., Greenwald I., 1 lin-, a gene required for vulval induction in Caenorhabditis elegans. Genes Dev. : 1. Yang S. H., Vickers E., Brehm A., Kouzarides T., Sharrocks A. D., 001 Temporal recruitment of the msina-histone deacetylase corepressor complex to the ETS domain transcription factor Elk-1. Mol. Cell. Biol. 1: 0 1. Yoda A., Kouike H., Okano H., Sawa H., 00 Components of the transcriptional Mediator complex are required for asymmetric cell division in C. elegans. Dev. Camb. Engl. 1: 1 1. Yoo A. S., Bais C., Greenwald I., 00 Crosstalk between the EGFR and LIN-1/Notch pathways in C. elegans vulval development. Science 0:. Zhang H., Emmons S. W., 000 A C. elegans mediator protein confers regulatory selectivity on lineage-specific expression of a transcription factor gene. Genes Dev. 1: Zhang X., Greenwald I., 0 Spatial regulation of lag- transcription during vulval precursor cell fate patterning in Caenorhabditis elegans. Genetics 1: Zhou H., Spaeth J. M., Kim N. H., Xu X., Friez M. J., Schwartz C. E., Boyer T. G., 01 MED1 mutations link intellectual disability syndromes with dysregulated GLI-dependent Sonic Hedgehog signaling. Proc. Natl. Acad. Sci. U. S. A. :
42 Tables Table 1. VPC induction scores and Muv/Vul penetrance in L animals (0 C) Genotype VPC induction ± SEM % Muv L a % Vul L b % WT L Vulva N.00 ± CKM cdk-(tm1).0 ± mdt-1(mn1). ± SynMuv lin-1a(n).00 ± cdk-(tm1); lin-1a(n).0 ± lin-1b(n).00 ± cdk-(tm1); lin-1b(n).0 ± N + EV RNAi.00 ± N + trr-1 RNAi.00 ± cdk-(tm1) + EV RNAi.00 ± cdk-(tm1) + trr-1 RNAi.0 ± gap-1(ga1).00 ± cdk-(tm1); gap-1(ga1).0 ± let- let-(sy) 0.00 ± cdk-(tm1); let-(sy). ± 0. 0 mpk-1 mpk-1(oz1).0 ± cdk-(tm1); mpk-1(oz). ± mdt-1(mn1); mpk-1(oz). ± lin-1 lin-1(n10). ± cdk-(tm1); lin-1(n10). ± mdt-1(mn1); lin-1(n10). ± N + lin-1 RNAi. ± cdk-(tm1) + lin-1 RNAi. ± lin-1 lin-1(n01). ± cdk-(tm1); lin-1(n01). ± mdt- mdt- (ku) 1.1 ± mdt-1(mn1); mdt-(ku). ± mdt-1 N + mdt-1 RNAi.00 ± lin-1(n10) + EV RNAi.00 ± lin-1(n10) + mdt-1 RNAi.01 ± a % L animals with ectopic vulval invagination at P.p, P.p or P.p. b % L animals with vulval induction <.0 at P.p-P.p. Note: it is possible for an animal to be scored as both Vul and Muv. n
43 Figure legends Figure 1. Transcriptional regulators in C. elegans vulval induction. The C. elegans vulva is derived from an equivalence group consisting of six vulva precursor cells (VPCs), named P.p through P.p from anterior to posterior. A localized LIN-/EGF signal from the anchor cell (AC) in the somatic gonad activates a LET-/EGFR-LET-0/Ras-MPK-1/ERK signaling cascade strongly in P.p. ERK activation in P.p modulates transcription factor activity in the nucleus (only LIN-1 is shown here for simplicity), leading to induction of the 1 cell fate. In P.p and P.p, a weak LIN-/EGF signal combined with lateral Notch signaling from P.p (not depicted) instead produces the cell fate. In P.p, P.p and P.p, the EGFR signaling cascade is not activated by LIN- /EGF, and cells adopt the non-vulval cell fate. Transcriptional regulators of the EGFR signaling pathway are critical for correct vulval cell fate specification: e.g. the MDT-/SUR- Mediator subunit is a coactivator of EGFR signaling-induced transcription, the SynMuv corepressor complexes are required to inhibit ectopic lin-/egf transcription in the hypodermis (Hyp) surrounding the VPCs, and the MDT-1/DPY- Mediator subunit is required to inhibit vulva development by mechanisms that remain unclear (dashed arrow). The GTPase activating protein GAP-1 that negatively regulates LET-0/Ras activity post-translationally is also shown Figure. cdk- represses vulval cell fates redundantly with synmuv genes. (A) Overlap of cdk- regulated genes with lin-/rb repressed genes. P-values were determined by Fisher s exact test. (B) Adult Muv penetrance in cdk- and mutants with synmuv genes (n ). * p < 0.0, ** p < 0.01, *** p < vs. cdk- mutant or vs. cdk- + empty vector (EV), Fisher s exact test. ND, not determined. See Table S for raw data. (C) Micrographs show wild-type or cdk- mutant worms expressing the aris[egl-1p::cfp] 1 fate marker. Bracket: Pn.px cells expressing reporter. Asterisk: ectopic reporter expression. Scale bar: µm. The graph displays the percentage of animals (n>0) with ectopic egl-1p::cfp expression (defined as expression in P.p/P.p/P.p, or expression
44 in P.p/P.p of equal intensity to P.p) or weak P.p/P.p expression (defined as expression in P.p/P.p that is weaker intensity than P.p). * p < 0.0, Fisher s exact test. (D) Micrographs show lin- 1A and cdk-; lin-1a mutants expressing the Notch-inducible reporter zhis[lip-1p::gfp]. Dashed bracket: P.px. Solid bracket: P.px and P.px. Open circle: loss of expression. Asterisk: ectopic expression. Scale bar: µm. The graph shows the percentage of animals (n 1) with ectopic expression (defined as strong expression in P.p, P.p, or P.p) or loss of expression (defined as weak or no expression in P.p or P.p) of the lip-1p::gfp reporter.* p < 0.0, Fisher s exact test Figure. cdk- acts upstream and downstream of let-/egfr. (A) qpcr analysis of lin- mrna levels in cdk- single mutants and mutants with synmuv genes, relative to wild-type worms. lin-1ab mutant is shown as a positive control (hatched bar). Error bars represent standard error of the mean (SEM, n= independent trials). No statistically significance differences, Wilcoxon signed rank test by method of Pratt. (B) hdex0[cdk-p::gfp] expression in the anchor cell (arrow) during early vulval induction. Top: VPCs divided once (Pn.px). Middle: VPCs divided twice (Pn.pxx). Bottom: Invagination of Pn.pxx epithelium. The fluorescent signal visible near VPCs localizes to neuron cell bodies. Scale bar: µm. (C) Average fluorescence intensity of syis[lin-acel::gfp] in anchor cell of wild-type worms and cdk- mutants. * p < 0.0 vs. N, t- test. A.U., arbitrary units. (D) L Vul penetrance in cdk-, cdk-; let-/egfr, and cdk-; mpk-1/erk mutants (n ). **** p < , Fisher s exact test. See Table 1 for raw data. (E) Tissue specificity of Muv phenotype in cdk-; lin-1a mutant adults expressing wild-type cdk- driven by its own promoter cdk-(+), the Pn.p promoter lin-1p, or the hypodermal promoter dpy-p, compared to non-transgenic siblings (Sib) in each strain (n ). **** p < vs. non-transgenic sibling, Fisher s exact test. See Table S for raw data. Figure. CDK- is a LIN-1/Ets corepressor.
45 (A) Adult Muv penetrance in cdk-; lin-1a mutants expressing kinase dead (KD) cdk-, compared to non-transgenic cdk-; lin-1a (n 1). *** p < vs. non-transgenic sibling, Fisher s exact test. See Table 1 for raw data. (B) Adult Muv penetrance in cdk- mutants on empty vector (EV) or lin-1/ets RNAi (n ) **** p < vs. cdk- + empty vector (EV), p < vs. N + lin-1 RNAi, Fisher s exact test. See Table S for raw data. (C) L Muv penetrance in cdk-; lin-1(n10) mutants (n 1). **** p < vs. cdk-, p < 0.0 vs. lin-1(n10), Fisher s exact test. See Table 1 for raw data. (D) Wild-type expression pattern of arex[lag-p(min)::gfp] in lin-1a mutants, and ectopic expression in cdk-; lin-1a mutants. Bracket: Pn.px. Asterisk: ectopic expression. Scale bar: µm. Number of ectopic expression events over total sample size noted next to genotype Figure. mdt-1/let-1 represses vulval cell fates downstream of mpk-1/erk. (A) Adult Muv penetrance in mdt-1/let-1 mutants and mutants with representative synmuv genes (n ). ** p < 0.01 vs. mdt-1/let-1 single mutant, Fisher s exact test. ND, not determined. See Table S for raw data. (B) Micrographs show mdt-1/let-1 and mdt-1/let-1; mdt-/sur- mutants expressing the aris[egl-1p::cfp] 1 fate marker. Bracket: Pn.px cells expressing reporter. Asterisk: ectopic reporter expression. Scale bar: µm. Graphs show the percentage of animals (n>0) with ectopic egl-1p::cfp expression (defined as expression in P.p/P.p/P.p, or expression in P.p/P.p of equal intensity to P.p) or weak P.p/P.p expression (defined as expression in P.p/P.p that is weaker intensity than P.p). * p < 0.0, Fisher s exact test. (C) Adult Muv penetrance in mdt- 1/let-1; lin-1(n10) mutants (n ). mdt-1/let-1 single mutant Muv penetrance from Fig. A is included for reference. *** p < vs. mdt-1/let-1 single mutant, p < vs. lin-1(n10) single mutant, Fisher s exact test. See Table S for raw data. (D) L Muv and Vul penetrance in mdt- 1/let-1; mpk-1/erk mutants (n 1). p = 0.0 (Muv) and * p < 0.0 (Vul) vs. mpk-1 single mutant, Fisher s exact test. See Table 1 for raw data. (E) L Muv and Vul penetrance in mdt-1/let-1; mdt-
46 /sur- mutants (n ). *** p < vs. mdt-1/let-1 single mutant, p < vs. mdt-/sur- single mutant, Fisher s exact test. See Table S for raw data Figure. The CKM inhibits activation of vulval induction by the mdt-1, mdt-, mdt- triad. (A) Adult Muv penetrance in mdt-1/let-1 mutants on mdt-1, mdt-, or mdt- triad subunit RNAi or mdt-1.1 non-triad subunit RNAi (n 1). * p < 0.0, ** p < 0.01, *** p < vs. mdt-1/let-1 + empty vector (EV), Fisher s exact test. See Table S for raw data. (B) L Muv penetrance in lin- 1(n10) mutants on mdt-1 RNAi (n ). No statistically significant difference, Fisher s exact test. See Table 1 for raw data. (C) qpcr analysis of mrna levels of cdk--repressed genes upon mdt-1, mdt-, or mdt- RNAi. Error bars represent SEM, n = independent trials. * p < 0.0, ** p < 0.01 vs. cdk- + empty vector (EV); p < 0.0, p < 0.01, p < 0.001, p < vs. N + empty vector (EV), Fisher s exact test. (D) α-mdt-1 Western blot in wild-type vs. cdk- mutants, with α- GAPDH loading control. Representative immunoblot from one of three trials. (E) Model of CKM inhibition of EGFR-Ras-MAPK signaling-dependent cell fate specification by repressing lin-/egf in the anchor cell (AC), promoting LIN-1 repressor activity (e.g. at lag-), and inhibiting tail module triad activity (e.g. at cdk--repressed genes) in non-1 VPCs (i.e. VPCs other than P.p). 1
47 Gonad AC LIN-/EGF Figure 1 Hyp LET-/EGFR Cell Fate: P.p P.p P.p P.p P.p P.p 1 LIN-/EGF 1 cell membrane LET-/EGFR Hyp SynMuv lin-/egf LET-0/Ras LIN-/Raf MEK-/MEK MPK-1/ERK Nucleus MPK-1/ERK MDT- LIN-1 GAP-1 cell membrane LET-/EGFR Nucleus MDT-1? LIN-1


")


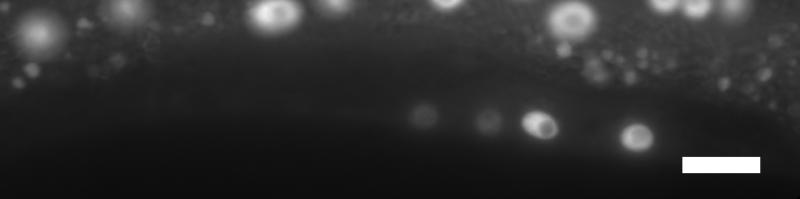

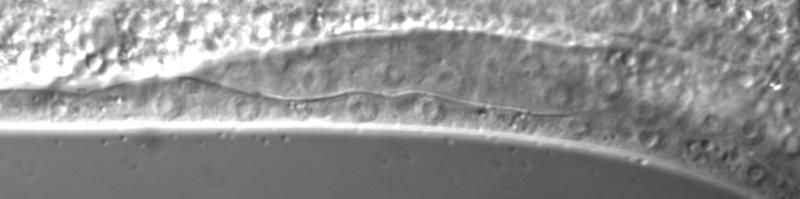
48 A cdk-(0) Up lin-(0) Up B Figure p =.0 - cdk-(0) Down lin-(0) Up C p =.0 - N cdk- C egl-1p::cfp DIC * * D lin-1a cdk-; lin-1a C lip-1p::gfp DIC * * * *
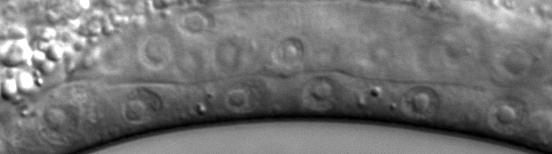




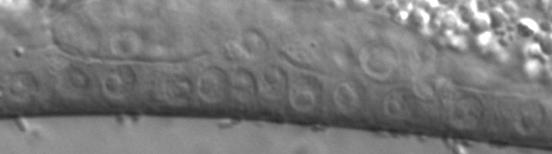
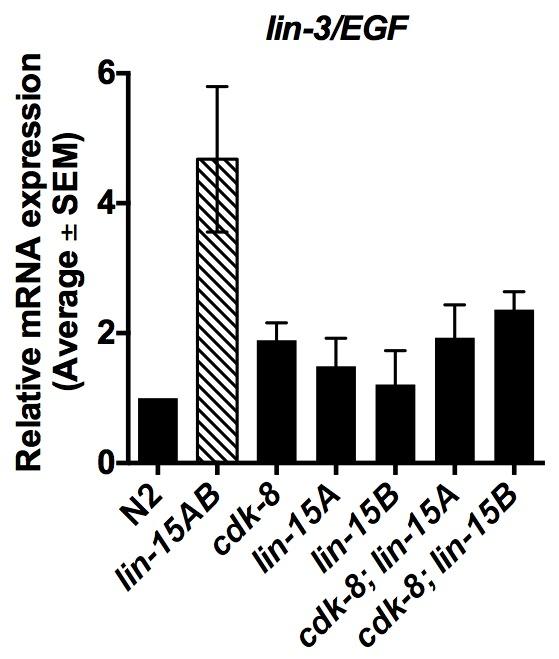
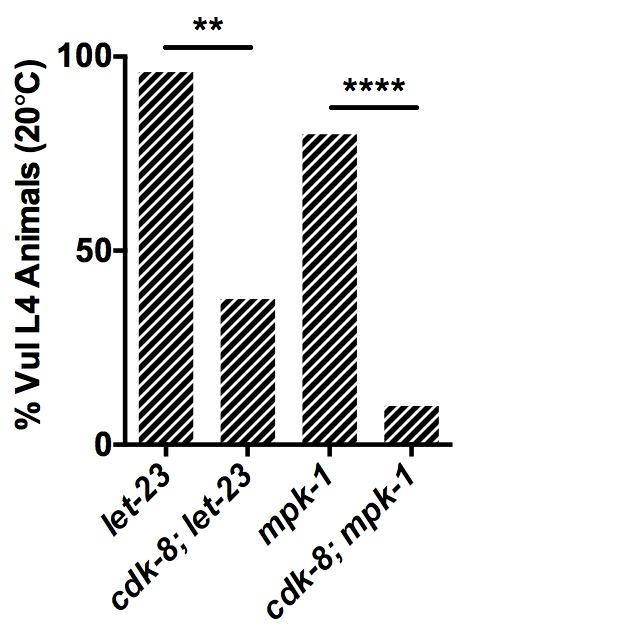
49 A B cdk-p::gfp Figure Pn.pxx Pn.px C D E *

")


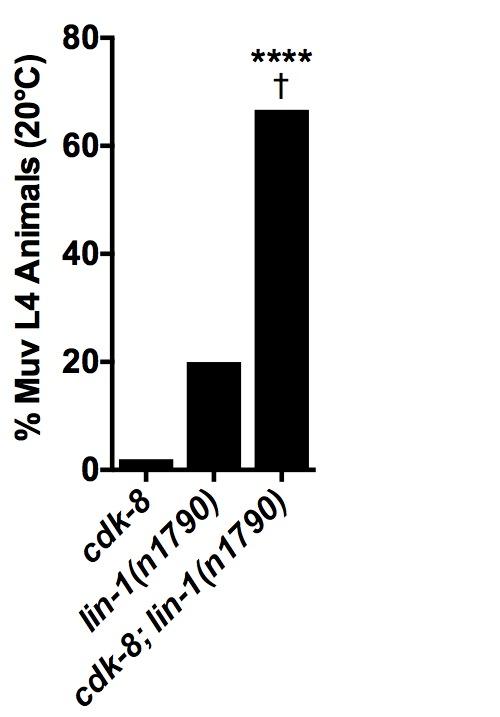
50 A B Figure C D lin-1a (0/) cdk-; lin-1a (/1) C lag-p::yfp DIC * *



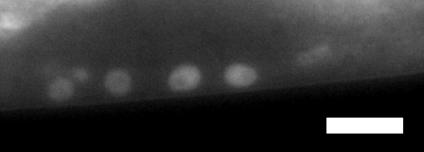
51 A C Figure B mdt-1 mdt-1; mdt- D 0 C egl-1p::cfp DIC * * * * E


52 A B Figure C D MDT-1 WT cdk- E Mediator Kinase Module GAPDH AC lin-/egf ACEL LIN-/EGF Tail Triad Mediator Kinase Module LET-/EGFR LIN-1 non-1 VPC cdk--repressed lag-
Bacterial diet affects vulval organogenesis in Caenorhabditis elegans Mediator kinase module mutants
 Correspondence taubert@cmmt.ubc.ca Disciplines Genetics Keywords Signal Transduction Transcription Gene Expression Models, Genetic Type of Observation Standalone Type of Link Orphan Data Submitted May
Correspondence taubert@cmmt.ubc.ca Disciplines Genetics Keywords Signal Transduction Transcription Gene Expression Models, Genetic Type of Observation Standalone Type of Link Orphan Data Submitted May
Predictive Modeling of Signaling Crosstalk... Model execution and model checking can be used to test a biological hypothesis
 Model execution and model checking can be used to test a biological hypothesis The model is an explanation of a biological mechanism and its agreement with experimental results is used to either validate
Model execution and model checking can be used to test a biological hypothesis The model is an explanation of a biological mechanism and its agreement with experimental results is used to either validate
Bypass and interaction suppressors; pathway analysis
 Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Cell fate specification by Ras-mediated cell signalling in C. elegans. Teresa Tiensuu
 Cell fate specification by Ras-mediated cell signalling in C. elegans Teresa Tiensuu Umeå Centre for Molecular Pathogenesis Umeå University Umeå Sweden 2003 Copyright Teresa Tiensuu 2003 ISBN 91-7305-525-5
Cell fate specification by Ras-mediated cell signalling in C. elegans Teresa Tiensuu Umeå Centre for Molecular Pathogenesis Umeå University Umeå Sweden 2003 Copyright Teresa Tiensuu 2003 ISBN 91-7305-525-5
with%dr.%van%buskirk%%%
 with%dr.%van%buskirk%%% How$to$do$well?$ Before$class:$read$the$corresponding$chapter$ Come$to$class$ready$to$par9cipate$in$Top$Hat$ Don t$miss$an$exam!!!!!!!!!!!!!!!!!!!!!!!!!!$ But$I m$not$good$with$science
with%dr.%van%buskirk%%% How$to$do$well?$ Before$class:$read$the$corresponding$chapter$ Come$to$class$ready$to$par9cipate$in$Top$Hat$ Don t$miss$an$exam!!!!!!!!!!!!!!!!!!!!!!!!!!$ But$I m$not$good$with$science
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
CHAPTER 2. Wnt and EGF pathways act together to induce C. elegans. male hook development
 II-1 CHAPTER 2 Wnt and EGF pathways act together to induce C. elegans male hook development Adeline Seah *, Hui Yu *, Michael A. Herman, Edwin L. Ferguson, H. Robert Horvitz and Paul W. Sternberg *Equal
II-1 CHAPTER 2 Wnt and EGF pathways act together to induce C. elegans male hook development Adeline Seah *, Hui Yu *, Michael A. Herman, Edwin L. Ferguson, H. Robert Horvitz and Paul W. Sternberg *Equal
Welcome to Class 21!
 Welcome to Class 21! Introductory Biochemistry! Lecture 21: Outline and Objectives l Regulation of Gene Expression in Prokaryotes! l transcriptional regulation! l principles! l lac operon! l trp attenuation!
Welcome to Class 21! Introductory Biochemistry! Lecture 21: Outline and Objectives l Regulation of Gene Expression in Prokaryotes! l transcriptional regulation! l principles! l lac operon! l trp attenuation!
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
EGF Signal Propagation during C. elegans Vulval Development Mediated by ROM-1 Rhomboid
 Open access, freely available online EGF Signal Propagation during C. elegans Vulval Development Mediated by ROM-1 Rhomboid PLoS BIOLOGY Amit Dutt [, Stefano Canevascini [, Erika Froehli-Hoier, Alex Hajnal
Open access, freely available online EGF Signal Propagation during C. elegans Vulval Development Mediated by ROM-1 Rhomboid PLoS BIOLOGY Amit Dutt [, Stefano Canevascini [, Erika Froehli-Hoier, Alex Hajnal
MIR-237 is Likely a Developmental Timing Gene that Regulates the L2-to-L3 Transition in C. Elegans
 Marquette University e-publications@marquette Master's Theses (2009 -) Dissertations, Theses, and Professional Projects MIR-237 is Likely a Developmental Timing Gene that Regulates the L2-to-L3 Transition
Marquette University e-publications@marquette Master's Theses (2009 -) Dissertations, Theses, and Professional Projects MIR-237 is Likely a Developmental Timing Gene that Regulates the L2-to-L3 Transition
Developmental Biology
 Developmental Biology 327 (2009) 419 432 Contents lists available at ScienceDirect Developmental Biology jour nal homepage: www. elsevier. com/ developmentalbiology Wnt and EGF pathways act together to
Developmental Biology 327 (2009) 419 432 Contents lists available at ScienceDirect Developmental Biology jour nal homepage: www. elsevier. com/ developmentalbiology Wnt and EGF pathways act together to
CHARACTERIZATION OF let-765/nsh-1 AND ITS ROLE IN RAS SIGNALLING IN Caenorhabditis elegans
 CHARACTERIZATION OF let-765/nsh-1 AND ITS ROLE IN RAS SIGNALLING IN Caenorhabditis elegans by Carrie Leanne Simms MSc, Simon Fraser University 2004 BSc, Simon Fraser University 1999 THESIS SUBMITTED IN
CHARACTERIZATION OF let-765/nsh-1 AND ITS ROLE IN RAS SIGNALLING IN Caenorhabditis elegans by Carrie Leanne Simms MSc, Simon Fraser University 2004 BSc, Simon Fraser University 1999 THESIS SUBMITTED IN
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
OSM-11 Facilitates LIN-12 Notch Signaling during Caenorhabditis elegans Vulval Development
 OSM-11 Facilitates LIN-12 Notch Signaling during Caenorhabditis elegans Vulval Development Hidetoshi Komatsu 1,2[ a, Michael Y. Chao 3[, Jonah Larkins-Ford 1 b, Mark E. Corkins 1, Gerard A. Somers 1, Tim
OSM-11 Facilitates LIN-12 Notch Signaling during Caenorhabditis elegans Vulval Development Hidetoshi Komatsu 1,2[ a, Michael Y. Chao 3[, Jonah Larkins-Ford 1 b, Mark E. Corkins 1, Gerard A. Somers 1, Tim
16 CONTROL OF GENE EXPRESSION
 16 CONTROL OF GENE EXPRESSION Chapter Outline 16.1 REGULATION OF GENE EXPRESSION IN PROKARYOTES The operon is the unit of transcription in prokaryotes The lac operon for lactose metabolism is transcribed
16 CONTROL OF GENE EXPRESSION Chapter Outline 16.1 REGULATION OF GENE EXPRESSION IN PROKARYOTES The operon is the unit of transcription in prokaryotes The lac operon for lactose metabolism is transcribed
Cytokines regulate interactions between cells of the hemapoietic system
 Cytokines regulate interactions between cells of the hemapoietic system Some well-known cytokines: Erythropoietin (Epo) G-CSF Thrombopoietin IL-2 INF thrombopoietin Abbas et al. Cellular & Molecular Immunology
Cytokines regulate interactions between cells of the hemapoietic system Some well-known cytokines: Erythropoietin (Epo) G-CSF Thrombopoietin IL-2 INF thrombopoietin Abbas et al. Cellular & Molecular Immunology
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
SUPPLEMENTARY MATERIAL
 1 SUPPLEMENTARY MATERIAL TABLE OF CONTENTS: Overview of VPC fate specification Biological sources of information for the Core Model Model structure: Objects GUI objects Internal objects Allele representation
1 SUPPLEMENTARY MATERIAL TABLE OF CONTENTS: Overview of VPC fate specification Biological sources of information for the Core Model Model structure: Objects GUI objects Internal objects Allele representation
CHAPTER 3. EGF and Wnt signaling during patterning of the C. elegans
 III-1 CHAPTER 3 EGF and Wnt signaling during patterning of the C. elegans Bγ/δ Equivalence Group Adeline Seah and Paul W. Sternberg Abstract III-2 During development, different signaling pathways interact
III-1 CHAPTER 3 EGF and Wnt signaling during patterning of the C. elegans Bγ/δ Equivalence Group Adeline Seah and Paul W. Sternberg Abstract III-2 During development, different signaling pathways interact
Wnt and EGF Pathways Act Together to Induce C. elegans Male Hook Development
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Faculty Publications in the Biological Sciences Papers in the Biological Sciences 3-2009 Wnt and EGF Pathways Act Together
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Faculty Publications in the Biological Sciences Papers in the Biological Sciences 3-2009 Wnt and EGF Pathways Act Together
GENETIC CONTROL OF CELL INTERACTIONS IN NEMATODE DEVELOPMENT
 Annu. Rev. Genet. 1991. 25. 411-36 Copyright 199l by Annual Reviews Inc. All rights reserved GENETIC CONTROL OF CELL INTERACTIONS IN NEMATODE DEVELOPMENT E. J. Lambie and Judith Kimble Laboratory of Molecular
Annu. Rev. Genet. 1991. 25. 411-36 Copyright 199l by Annual Reviews Inc. All rights reserved GENETIC CONTROL OF CELL INTERACTIONS IN NEMATODE DEVELOPMENT E. J. Lambie and Judith Kimble Laboratory of Molecular
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its
 The Wnt/ -catenin signalling pathway and its") Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Wan-Ju Liu 1,2, John S Reece-Hoyes 3, Albertha JM Walhout 3 and David M Eisenmann 1*
 Liu et al. BMC Developmental Biology 2014, 14:17 RESEARCH ARTICLE Open Access Multiple transcription factors directly regulate Hox gene lin-39 expression in ventral hypodermal cells of the C. elegans embryo
Liu et al. BMC Developmental Biology 2014, 14:17 RESEARCH ARTICLE Open Access Multiple transcription factors directly regulate Hox gene lin-39 expression in ventral hypodermal cells of the C. elegans embryo
Hypodermis to Oppose Ras-Mediated Vulval Induction in C. elegans
 Developmental Cell, Vol. 8, 117 123, January, 2005, Copyright 2005 by Elsevier Inc. DOI 10.1016/j.devcel.2004.11.015 lin-35 Rb Acts in the Major Hypodermis to Oppose Ras-Mediated Vulval Induction in C.
Developmental Cell, Vol. 8, 117 123, January, 2005, Copyright 2005 by Elsevier Inc. DOI 10.1016/j.devcel.2004.11.015 lin-35 Rb Acts in the Major Hypodermis to Oppose Ras-Mediated Vulval Induction in C.
Adam J. Schindler, L. Ryan Baugh, David R. Sherwood* Abstract. Introduction
 Identification of Late Larval Stage Developmental Checkpoints in Caenorhabditis elegans Regulated by Insulin/IGF and Steroid Hormone Signaling Pathways Adam J. Schindler, L. Ryan Baugh, David R. Sherwood*
Identification of Late Larval Stage Developmental Checkpoints in Caenorhabditis elegans Regulated by Insulin/IGF and Steroid Hormone Signaling Pathways Adam J. Schindler, L. Ryan Baugh, David R. Sherwood*
Conservation and diversification of Wnt signaling function during the evolution of nematode vulva development
 Conservation and diversification of Wnt signaling function during the evolution of nematode vulva development Min Zheng 1, Daniel Messerschmidt 1, Benno Jungblut 1,2 & Ralf J Sommer 1 Cell-fate specification
Conservation and diversification of Wnt signaling function during the evolution of nematode vulva development Min Zheng 1, Daniel Messerschmidt 1, Benno Jungblut 1,2 & Ralf J Sommer 1 Cell-fate specification
Upstream Elements Regulating mir-241 and mir-48 Abstract Introduction
 Upstream Elements Regulating mir-241 and mir-48 Hanna Vollbrecht, Tamar Resnick, and Ann Rougvie University of Minnesota: Twin Cities Undergraduate Research Scholarship 2012-2013 Abstract Caenorhabditis
Upstream Elements Regulating mir-241 and mir-48 Hanna Vollbrecht, Tamar Resnick, and Ann Rougvie University of Minnesota: Twin Cities Undergraduate Research Scholarship 2012-2013 Abstract Caenorhabditis
The two steps of vulval induction in Oscheius tipulae CEW1 recruit common regulators including a MEK kinase
 Developmental Biology 265 (2004) 113 126 www.elsevier.com/locate/ydbio The two steps of vulval induction in Oscheius tipulae CEW1 recruit common regulators including a MEK kinase Marie-Laure Dichtel-Danjoy
Developmental Biology 265 (2004) 113 126 www.elsevier.com/locate/ydbio The two steps of vulval induction in Oscheius tipulae CEW1 recruit common regulators including a MEK kinase Marie-Laure Dichtel-Danjoy
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
S1 Gene ontology (GO) analysis of the network alignment results
 analysis of the network alignment results") 1 Supplementary Material for Effective comparative analysis of protein-protein interaction networks by measuring the steady-state network flow using a Markov model Hyundoo Jeong 1, Xiaoning Qian 1 and
1 Supplementary Material for Effective comparative analysis of protein-protein interaction networks by measuring the steady-state network flow using a Markov model Hyundoo Jeong 1, Xiaoning Qian 1 and
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Initiation of translation in eukaryotic cells:connecting the head and tail
 Initiation of translation in eukaryotic cells:connecting the head and tail GCCRCCAUGG 1: Multiple initiation factors with distinct biochemical roles (linking, tethering, recruiting, and scanning) 2: 5
Initiation of translation in eukaryotic cells:connecting the head and tail GCCRCCAUGG 1: Multiple initiation factors with distinct biochemical roles (linking, tethering, recruiting, and scanning) 2: 5
Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
A complementation test would be done by crossing the haploid strains and scoring the phenotype in the diploids.
 Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
University of Massachusetts Medical School Wan-Ju Liu University of Maryland
 University of Massachusetts Medical School escholarship@umms Program in Systems Biology Publications and Presentations Program in Systems Biology 5-13-2014 Multiple transcription factors directly regulate
University of Massachusetts Medical School escholarship@umms Program in Systems Biology Publications and Presentations Program in Systems Biology 5-13-2014 Multiple transcription factors directly regulate
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Tuesday, December 27, 16
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter
 Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter 9/10/2008 1 Learning Objectives Explain why a cell cycle was selected for during evolution
Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter 9/10/2008 1 Learning Objectives Explain why a cell cycle was selected for during evolution
Mutations in cye-1, a Caenorhabditis elegans cyclin E homolog, reveal coordination between cell-cycle control and vulval development
 Development 127, 4049-4060 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV8754 4049 Mutations in cye-1, a Caenorhabditis elegans cyclin E homolog, reveal coordination between
Development 127, 4049-4060 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV8754 4049 Mutations in cye-1, a Caenorhabditis elegans cyclin E homolog, reveal coordination between
Synthesis of Biological Models from Mutation Experiments
 Synthesis of Biological Models from Mutation Experiments Ali Sinan Köksal, Saurabh Srivastava, Rastislav Bodík, UC Berkeley Evan Pu, MIT Jasmin Fisher, Microsoft Research Cambridge Nir Piterman, University
Synthesis of Biological Models from Mutation Experiments Ali Sinan Köksal, Saurabh Srivastava, Rastislav Bodík, UC Berkeley Evan Pu, MIT Jasmin Fisher, Microsoft Research Cambridge Nir Piterman, University
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
elegans vulva precursor cells
 Development 126, 1947-1956 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV5288 1947 Cell cycle-dependent sequencing of cell fate decisions in Caenorhabditis elegans vulva precursor
Development 126, 1947-1956 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV5288 1947 Cell cycle-dependent sequencing of cell fate decisions in Caenorhabditis elegans vulva precursor
Lecture 18 June 2 nd, Gene Expression Regulation Mutations
 Lecture 18 June 2 nd, 2016 Gene Expression Regulation Mutations From Gene to Protein Central Dogma Replication DNA RNA PROTEIN Transcription Translation RNA Viruses: genome is RNA Reverse Transcriptase
Lecture 18 June 2 nd, 2016 Gene Expression Regulation Mutations From Gene to Protein Central Dogma Replication DNA RNA PROTEIN Transcription Translation RNA Viruses: genome is RNA Reverse Transcriptase
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
ADAM FAMILY. ephrin A INTERAZIONE. Eph ADESIONE? PROTEOLISI ENDOCITOSI B A RISULTATO REPULSIONE. reverse. forward
 ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter
 Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter 9/10/2008 1 Learning Objectives Explain similarities and differences between fungal, mammalian and plant cell cycles Explain
Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter 9/10/2008 1 Learning Objectives Explain similarities and differences between fungal, mammalian and plant cell cycles Explain
Chapter 18 Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
SUPPLEMENTARY INFORMATION
 med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
Javier A. Wagmaister a,1, Ginger R. Miley b,2, Corey A. Morris c,3, Julie E. Gleason a, Leilani M. Miller c, Kerry Kornfeld b, David M.
 Developmental Biology 297 (2006) 550 565 www.elsevier.com/locate/ydbio Genomes & Developmental Control Identification of cis-regulatory elements from the C. elegans Hox gene lin-39 required for embryonic
Developmental Biology 297 (2006) 550 565 www.elsevier.com/locate/ydbio Genomes & Developmental Control Identification of cis-regulatory elements from the C. elegans Hox gene lin-39 required for embryonic
Conversion of the LIN-1 ETS protein of C. elegans from a sumoylated transcriptional repressor to a phosphorylated transcriptional activator.
 Genetics: Early Online, published on January 7, 2015 as 10.1534/genetics.114.172668 Conversion of the LIN-1 ETS protein of C. elegans from a sumoylated transcriptional repressor to a phosphorylated transcriptional
Genetics: Early Online, published on January 7, 2015 as 10.1534/genetics.114.172668 Conversion of the LIN-1 ETS protein of C. elegans from a sumoylated transcriptional repressor to a phosphorylated transcriptional
Chapter 2 Organogenesis of the C. elegans Vulva and Control of Cell Fusion
 Chapter 2 Organogenesis of the C. elegans Vulva and Control of Cell Fusion Nathan Weinstein and Benjamin Podbilewicz Abstract The vulva of Caenorhabditis elegans is widely used as a paradigm for the study
Chapter 2 Organogenesis of the C. elegans Vulva and Control of Cell Fusion Nathan Weinstein and Benjamin Podbilewicz Abstract The vulva of Caenorhabditis elegans is widely used as a paradigm for the study
Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday
 Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday 1. What is the Central Dogma? 2. How does prokaryotic DNA compare to eukaryotic DNA? 3. How is DNA
Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday 1. What is the Central Dogma? 2. How does prokaryotic DNA compare to eukaryotic DNA? 3. How is DNA
Caenorhabditis elegans Development
 MOLECULAR REPRODUCTION AND DEVELOPMENT 42523528 (1995) LET-23-Mediated Signal Transduction During Caenorhabditis elegans Development PAUL W. STERNBERG, GIOVANNI LESA, JUNHO LEE, WENDY S. KATZ, CHARLES
MOLECULAR REPRODUCTION AND DEVELOPMENT 42523528 (1995) LET-23-Mediated Signal Transduction During Caenorhabditis elegans Development PAUL W. STERNBERG, GIOVANNI LESA, JUNHO LEE, WENDY S. KATZ, CHARLES
Caenorhabditis elegans
 Caenorhabditis elegans Why C. elegans? Sea urchins have told us much about embryogenesis. They are suited well for study in the lab; however, they do not tell us much about the genetics involved in embryogenesis.
Caenorhabditis elegans Why C. elegans? Sea urchins have told us much about embryogenesis. They are suited well for study in the lab; however, they do not tell us much about the genetics involved in embryogenesis.
Bi Lecture 8 Genetic Pathways and Genetic Screens
 Bi190-2013 Lecture 8 Genetic Pathways and Genetic Screens WT A 2X:2A her-1 tra-1 1X:2A her-1 tra-1 Female body Male body Female body Male body her-1(lf) B 2X:2A her-1(lf) tra-1 1X:2A her-1(lf) tra-1 Female
Bi190-2013 Lecture 8 Genetic Pathways and Genetic Screens WT A 2X:2A her-1 tra-1 1X:2A her-1 tra-1 Female body Male body Female body Male body her-1(lf) B 2X:2A her-1(lf) tra-1 1X:2A her-1(lf) tra-1 Female
The Eukaryotic Genome and Its Expression. The Eukaryotic Genome and Its Expression. A. The Eukaryotic Genome. Lecture Series 11
 The Eukaryotic Genome and Its Expression Lecture Series 11 The Eukaryotic Genome and Its Expression A. The Eukaryotic Genome B. Repetitive Sequences (rem: teleomeres) C. The Structures of Protein-Coding
The Eukaryotic Genome and Its Expression Lecture Series 11 The Eukaryotic Genome and Its Expression A. The Eukaryotic Genome B. Repetitive Sequences (rem: teleomeres) C. The Structures of Protein-Coding
Nature Biotechnology: doi: /nbt Supplementary Figure 1. Overexpression of YFP::GPR-1 in the germline.
 Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Written Exam 15 December Course name: Introduction to Systems Biology Course no
 Technical University of Denmark Written Exam 15 December 2008 Course name: Introduction to Systems Biology Course no. 27041 Aids allowed: Open book exam Provide your answers and calculations on separate
Technical University of Denmark Written Exam 15 December 2008 Course name: Introduction to Systems Biology Course no. 27041 Aids allowed: Open book exam Provide your answers and calculations on separate
V- 1. Chapter 5. Summary
 V- 1 Chapter 5 Summary V- 2 This body of work combines molecular and genetic techniques to analyze IP 3 signaling downstream of the Caenorhabditis elegans LET-23 epidermal growth factor receptor homolog.
V- 1 Chapter 5 Summary V- 2 This body of work combines molecular and genetic techniques to analyze IP 3 signaling downstream of the Caenorhabditis elegans LET-23 epidermal growth factor receptor homolog.
lecture 10. Genetic Interactions synthetic lethality high copy suppression suppression and enhancement mechanisms of suppression
 lecture 10. Genetic Interactions synthetic lethality high copy suppression suppression and enhancement mechanisms of suppression actin act1-1 act1-4 actin non- complementer anc4 anc1 anc3 anc2 actin-binding
lecture 10. Genetic Interactions synthetic lethality high copy suppression suppression and enhancement mechanisms of suppression actin act1-1 act1-4 actin non- complementer anc4 anc1 anc3 anc2 actin-binding
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biol403 - Receptor Serine/Threonine Kinases
 Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Prokaryotic Regulation
 Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Afundamental issue in developmental biology is how
 Copyright Ó 2006 by the Genetics Society of America DOI: 10.1534/genetics.106.056465 Identification and Classification of Genes That Act Antagonistically to let-60 Ras Signaling in Caenorhabditis elegans
Copyright Ó 2006 by the Genetics Society of America DOI: 10.1534/genetics.106.056465 Identification and Classification of Genes That Act Antagonistically to let-60 Ras Signaling in Caenorhabditis elegans
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
The β-catenin homolog BAR-1 and LET-60 Ras coordinately regulate the Hox gene lin-39 during Caenorhabditis elegans vulval development
 Development 125, 3667-3680 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5223 3667 The β-catenin homolog BAR-1 and LET-60 Ras coordinately regulate the Hox gene lin-39 during
Development 125, 3667-3680 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5223 3667 The β-catenin homolog BAR-1 and LET-60 Ras coordinately regulate the Hox gene lin-39 during
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Cell Death & Trophic Factors II. Steven McLoon Department of Neuroscience University of Minnesota
 Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Homology and the hierarchy of biological systems
 Homology and the hierarchy of biological systems Ralf J. Sommer Summary Homology is the similarity between organisms due to common ancestry. Introduced by Richard Owen in 1843 in a paper entitled Lectures
Homology and the hierarchy of biological systems Ralf J. Sommer Summary Homology is the similarity between organisms due to common ancestry. Introduced by Richard Owen in 1843 in a paper entitled Lectures
Gene regulation I Biochemistry 302. Bob Kelm February 25, 2005
 Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
7.013 Problem Set
 7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
SUPPLEMENTARY INFORMATION
 Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Stochastic simulations
 Stochastic simulations Application to molecular networks Literature overview Noise in genetic networks Origins How to measure and distinguish between the two types of noise (intrinsic vs extrinsic)? What
Stochastic simulations Application to molecular networks Literature overview Noise in genetic networks Origins How to measure and distinguish between the two types of noise (intrinsic vs extrinsic)? What
CS-E5880 Modeling biological networks Gene regulatory networks
 CS-E5880 Modeling biological networks Gene regulatory networks Jukka Intosalmi (based on slides by Harri Lähdesmäki) Department of Computer Science Aalto University January 12, 2018 Outline Modeling gene
CS-E5880 Modeling biological networks Gene regulatory networks Jukka Intosalmi (based on slides by Harri Lähdesmäki) Department of Computer Science Aalto University January 12, 2018 Outline Modeling gene
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
23-. Shoot and root development depend on ratio of IAA/CK
 Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells
 Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells Understand how a simple biochemical oscillator can drive the
Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells Understand how a simple biochemical oscillator can drive the
COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION
 COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION I. Aksan 1, M. Sen 2, M. K. Araz 3, and M. L. Kurnaz 3 1 School of Biological Sciences, University of Manchester,
COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION I. Aksan 1, M. Sen 2, M. K. Araz 3, and M. L. Kurnaz 3 1 School of Biological Sciences, University of Manchester,
Eukaryotic vs. Prokaryotic genes
 BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 18: Eukaryotic genes http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Eukaryotic vs. Prokaryotic genes Like in prokaryotes,
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 18: Eukaryotic genes http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Eukaryotic vs. Prokaryotic genes Like in prokaryotes,
State Machine Modeling of MAPK Signaling Pathways
 State Machine Modeling of MAPK Signaling Pathways Youcef Derbal Ryerson University yderbal@ryerson.ca Abstract Mitogen activated protein kinase (MAPK) signaling pathways are frequently deregulated in human
State Machine Modeling of MAPK Signaling Pathways Youcef Derbal Ryerson University yderbal@ryerson.ca Abstract Mitogen activated protein kinase (MAPK) signaling pathways are frequently deregulated in human
Examination paper for Bi3016 Molecular Cell Biology
 Department of Biology Examination paper for Bi3016 Molecular Cell Biology Academic contact during examination: Per Winge Phone: 99369359 Examination date: 20 th December 2017 Examination time (from-to):
Department of Biology Examination paper for Bi3016 Molecular Cell Biology Academic contact during examination: Per Winge Phone: 99369359 Examination date: 20 th December 2017 Examination time (from-to):
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Three types of RNA polymerase in eukaryotic nuclei
 Three types of RNA polymerase in eukaryotic nuclei Type Location RNA synthesized Effect of α-amanitin I Nucleolus Pre-rRNA for 18,.8 and 8S rrnas Insensitive II Nucleoplasm Pre-mRNA, some snrnas Sensitive
Three types of RNA polymerase in eukaryotic nuclei Type Location RNA synthesized Effect of α-amanitin I Nucleolus Pre-rRNA for 18,.8 and 8S rrnas Insensitive II Nucleoplasm Pre-mRNA, some snrnas Sensitive
THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING. AnitaHajdu. Thesis of the Ph.D.
 THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING AnitaHajdu Thesis of the Ph.D. dissertation Supervisor: Dr. LászlóKozma-Bognár - senior research associate Doctoral
THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING AnitaHajdu Thesis of the Ph.D. dissertation Supervisor: Dr. LászlóKozma-Bognár - senior research associate Doctoral
CELL CYCLE AND DIFFERENTIATION
 CELL CYCLE AND DIFFERENTIATION Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id WHAT IS CELL CYCLE? 09/12/14 d.purnomosari@ugm.ac.id
CELL CYCLE AND DIFFERENTIATION Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id WHAT IS CELL CYCLE? 09/12/14 d.purnomosari@ugm.ac.id
REVIEW SESSION. Wednesday, September 15 5:30 PM SHANTZ 242 E
 REVIEW SESSION Wednesday, September 15 5:30 PM SHANTZ 242 E Gene Regulation Gene Regulation Gene expression can be turned on, turned off, turned up or turned down! For example, as test time approaches,
REVIEW SESSION Wednesday, September 15 5:30 PM SHANTZ 242 E Gene Regulation Gene Regulation Gene expression can be turned on, turned off, turned up or turned down! For example, as test time approaches,
bi lecture 17 integration of genetic interaction data
 bi190 2013 lecture 17 integration of genetic interaction data Genes interact to specify phenotype 20,000 genes 200,000,000 potential binary interactions (2 locus physiological epistasis) How will we efficiently
bi190 2013 lecture 17 integration of genetic interaction data Genes interact to specify phenotype 20,000 genes 200,000,000 potential binary interactions (2 locus physiological epistasis) How will we efficiently
9/4/2015 INDUCTION CHAPTER 1. Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology. Fig 1.
 INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
Developmental Biology Lecture Outlines
 Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
BIS &003 Answers to Assigned Problems May 23, Week /18.6 How would you distinguish between an enhancer and a promoter?
 Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Regulation of gene expression. Premedical - Biology
 Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Honors Biology Reading Guide Chapter 11
 Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Question Set # 4 Answer Key 7.22 Nov. 2002
 Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways
Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways