Natural variation in the flag leaf morphology of rice due to a mutation of the NARROW LEAF 1 gene in Oryza sativa L.
|
|
|
- Blaze Bradley
- 5 years ago
- Views:
Transcription
1 Genetics: Early Online, published on August 1, 01 as.1/genetics Natural variation in the flag leaf morphology of rice due to a mutation of the NARROW LEAF 1 gene in Oryza sativa L. Fumio Taguchi-Shiobara *, Tatsuya Ota, Kaworu Ebana *,1, Taiichiro Ookawa, Masanori Yamasaki, Takanari Tanabata *,, Utako Yamanouchi*, Jianzhong Wu *, Nozomi Ono**, Yasunori Nonoue *,, Kazufumi Nagata *, Shuichi Fukuoka *, Hideyuki Hirabayashi, Toshio Yamamoto *, Masahiro Yano *, * Agrogenomics Research Center, National Institute of Agrobiological Sciences, Tsukuba, Ibaraki 0-0, Japan Department of Evolutionary Studies of Biosystems, SOKENDAI (The Graduate University for Advanced Studies), Hayama, Kanagawa 0-01, Japan Graduate School of Agriculture, Tokyo University of Agriculture and Technology, Fuchu, Tokyo 1-0, Japan Food Resources Education and Research Center, Graduate School of Agricultural Science, Kobe University, Kasai, Hyogo -, Japan ** Institute of the Society for Techno-innovation of Agriculture, Forestry and Fisheries, -1 Ippaizuka, Kamiyokoba, Tsukuba, Ibaraki 0-0, Japan Rice Research Division, NARO Institute of Crop Science, Tsukuba, Ibaraki 0-1, Japan 1 Present address: Genetic Resources Center, National Institute of Agrobiological Sciences, Tsukuba, Ibaraki 0-0, Japan. Present address: Center for Sustainable Resource Science, RIKEN, Tsukuba, Ibaraki 0-00, Japan. Present address: Iwate Biotechnology Research Center, Kitakami, Iwate 0-000, Japan. Present address: NARO Institute of Crop Science, Tsukuba, Ibaraki 0-1, Japan. 0 1 Number of references: 1 Copyright 01.
2 Running title: NAL1 causes flag leaf variations Keywords: rice, flag leaf width, natural variation, Oryza sativa L., NARROW LEAF 1 Corresponding author: Fumio Taguchi-Shiobara Rice Applied Genomics Research Unit (*) Agrogenomics Research Center National Institute of Agrobiological Sciences -1- Kannondai, Tsukuba, Ibaraki 0-0, Japan Tel/Fax: fstagu@affrc.go.jp (*) Present address: Soybean Applied Genomics Research Unit 1
3 ABSTRACT We investigated the natural variations in the flag leaf morphology of rice. We conducted a principal component analysis based on nine flag leaf morphology traits using accessions from the National Institute of Agrobiological Sciences Core Collection. The first component explained % of total variance and the variable with highest loading was the width of the flag leaf (WFL). A genome-wide association analysis of diverse Japanese accessions revealed that marker RM on chromosome was highly associated with WFL. In analyses of progenies derived from a cross between Takanari and Akenohoshi, the most significant quantitative trait locus (QTL) for WFL was in a. kb-region containing the NARROW LEAF 1 (NAL1) gene, located 0. Mb downstream of RM. Analyses of chromosomal segment substitution lines indicated that a mutation (GA single nucleotide mutation, causing an RH amino acid substitution in NAL1) was present at the QTL. This explained 1% and 0% of total variability in WFL and the distance between small vascular bundles, respectively. The mutation apparently occurred during rice domestication and spread into japonica, tropical japonica, and indica subgroups. Notably, one accession, Phulba, had a NAL1 allele encoding only the N-terminal, or one-fourth, of the wild-type peptide. Given that the Phulba allele and the histidine-type allele showed essentially the same phenotype, the histidine-type allele was regarded as malfunctional. The phenotypes of transgenic plants varied depending on the ratio of histidine-type alleles to arginine-type alleles, raising the possibility that H -type products function differently from and compete with R -type products. INTRODUCTION The leaf of grasses typically consists of a relatively narrow blade and sheath enclosing the stem, and venation is parallel in the blade and the sheath (Esau 1). Because large leaves intercept more light, the leaf area of the blade strongly affects final yield in cereal crops (Watson 1). To produce plants that intercept light efficiently, leaf angle has been a target in breeding programs because erect leaves can capture more sunlight (Sinclair and Sheehy 1). It was demonstrated that a brassinosteroid-deficient mutant with erect leaves showed increased grain yield under dense planting conditions
4 (Sakamoto et al. 00). It is also essential to understand the mechanism of development and the natural variations in morphology of the flag leaf, since photosynthesis in the top three leaf blades of the plant, especially flag leaf, makes the largest contribution to the grain yield of rice (Yoshida 1; Tanaka 1). The developmental processes of the flag leaf are the same as those of other leaves. In rice, the longitudinal strands in the leaf comprise the midrib, large vascular bundles, and small vascular bundles (Hoshikawa 1). According to Inosaka (1) and Itoh (00), the midrib and large vascular bundles initiate at the base of the leaf primordium and develop acropetally in the leaf and basipetally in the culm (stage P in leaf development). It takes about one plastochron to initiate a small vascular bundle after the initiation of the midrib. When the leaf sheath and blade start to differentiate (the beginning of stage P), small vascular bundles become visible between the large vascular bundles at the base of leaf primordia. Small vascular bundles form acropetally in the leaf blade and basipetally in the stem. Later (stage P), a small vascular bundle develops between the midrib and a large vascular bundle near the leaf tip, and extends basipetally through the leaf blade. Then, more small vascular bundles form between large vascular bundles sequentially from the midrib towards the leaf margin. After the rapid elongation of the leaf blade (stage P) and the leaf sheath (stage P), the leaf becomes mature and growth is complete (stage P). Natural variations in flag leaf size have been reported for 1 rice accessions from Japan and accessions from other countries (Matsuo 1). Flag leaves were wider in accessions from Java, western China, and Latin America, and narrower in those from north China, central China, and Russia. Flag leaves were longer in accessions from Java and India, but shorter in those from Taiwan, central China, and south China. In genome-wide association studies (GWAS) of 1 diverse accessions, significant loci accounted for about % of variance in the width of the flag leaf (WFL), and three loci on chromosomes 1,, and contributed.%,.0%, and.1% of phenotypic variance, respectively (Zhao et al. 0). The NARROW LEAF 1 (NAL1) gene was located at approximately 1. Mb on chromosome (hereafter, all genomic positions are based on Os-Nipponbare-Reference-IRGSP-1.0), close to one of the single nucleotide
5 polymorphisms contributing to the variation in WFL reported by Zhao et al. (0). NAL1 was originally isolated as a gene affecting vascular patterns in a study on a classic dwarf mutant with a narrow leaf (Qi et al. 00) and was later shown to affect WFL, total spikelet number per panicle, photosynthetic rate, and chlorophyll content (Chen et al. 01; Fujita et al. 01; Takai et al. 01; Zhang et al. 01). NAL1 encodes a plant-specific protein, and the nal1 mutant with an in-frame deletion of amino acids in exon showed reduced basipetal polar auxin transport and fewer longitudinal veins, compared with wild type (Qi et al. 00). A recent analysis of mutant rice with a NAL1 null allele implied that NAL1 is also involved in control of the cell cycle and cell division from the initial stage of leaf development onwards (Jiang et al. 01). The NAL1 gene exhibits natural variations that are associated with plant morphology in rice. The NAL1 allele in Koshihikari, a temperate japonica cultivar, has three amino acid differences (RH, AV, and VI) compared with NAL1 in the indica cultivar Takanari. A study on near-isogenic lines revealed that the Koshihikari NAL1 allele decreased the thickness of the flag leaf and increased the ratio of leaf area to dry mass, i.e., specific leaf area (Takai et al. 01). The NAL1 allele in Daringan, a tropical japonica landrace, has the same predicted amino acid sequence as that of NAL1 in Koshihikari. When the Daringan allele was introduced into an indica cultivar with the same NAL1 protein as Takanari (in an IR background), the progeny showed increased flag leaf width, more vascular bundles, greater root biomass, more spikelets, and increased grain yield per square meter (Fujita et al. 01), suggesting that NAL1 affects various yield-related traits. In this study, we extensively examined world-wide collections of rice accessions using a variety of analyses, including principal component analysis (PCA) of morphological characters, GWAS, quantitative trait loci (QTL) analysis, and positional cloning, to explore the cause of the natural variations in the flag leaf morphology of rice. We found that an amino acid-altering mutation in NAL1, which occurred during rice domestication, significantly contributes to the natural variations in rice flag leaf morphology. MATERIALS AND METHODS
6 Plant materials The rice accessions used in this study are summarized in Table S1 and details are provided in File S1. A total of accessions from the NIAS Core Collection represented natural variations in Oryza sativa L.; accessions were from the Global Core Collection (WRC) (Kojima et al. 00) and accessions were from the Core Collection of Japanese Landraces (JRC) (Ebana et al. 00). Seeds of all of these accessions were planted in 01 to collect data for the PCA (Tables S1 and S). A total of accessions (Tables S1 and S) were selected to represent the breeding history of rice in Japan in the GWAS. These accessions were selected because their population structure was simpler than that of the accessions in the NIAS Core Collection (Yamasaki and Ideta 01). Seeds of these accessions were planted in 00 and 0. Seeds of world-wide collections of rice accessions including those described above, hybrid populations derived from a Takanari Akenohoshi cross, and most of the chromosomal segment substitution lines (CSSLs) were sown in April. The seedlings were transplanted in May in in an experimental field at the National Institute of Agrobiological Sciences (NIAS), Tsukuba, Japan. These plants were used to evaluate flag leaf morphology traits such as the length and width of the flag leaf, number of vascular bundles, distance between vascular bundles, and thickness of the flag leaf. Seeds of the CSSLs that were used to evaluate the Phulba allele of NAL1 were sown in May 01, and seedlings were transplanted in June. Seeds of recombinant fixed lines containing either Akenohoshi or Takanari fragments were sown on April 0 and May 0, and seedlings were transplanted on May 0 and June 1 in 0 and 0, respectively, in an experimental field at the Tokyo University of Agriculture and Technology (TUAT), Fuchu, Japan. These plants were used to evaluate yield and yield-related traits such as flag leaf size, dry weight of leaf and culm, and number of spikelets per plant. Evaluation of flag leaf morphology Plants with a heading date between July 0 and September 1 were used to evaluate flag
7 leaf morphology. Flag leaves were sampled in the field and their width and/or length were measured. In 01, the widest part of the flag leaf was measured as WFL in the field using a ruler. To perform the measurement tasks efficiently, we developed an application program to record measured data using an ipod touch (Apple Inc. Cupertino, CA, USA). Additional details about evaluating flag leaf morphology are provided in File S1. Principal component analysis A flag leaf was sampled from the main or equivalent culm of each plant, and nine traits were scored to identify the primary factors affecting flag leaf morphology by PCA (Table 1). Scores from five plants were averaged to represent each accession. In addition to length, width, and thickness of the flag leaf, the number of small or large vascular bundles and the distance between vascular bundles were recorded. The thickness of the flag leaf was represented by the thickness at three points around the second vascular bundle beside the midrib; i.e., the point of the large vascular bundle, the point of the small vascular bundle, and the point of the motor cell. All nine traits were used for the correlation analysis and PCA, which was conducted using JMP version.0 (SAS Institute, Cary, NC, USA). Genome-wide association analysis We conducted the GWAS of flag leaf morphology using rice accessions (Table S) and 1, DNA markers (Nagasaki et al. 0; Yamasaki and Ideta 01) tagged by Tagger (de Bakker et al. 00). The GWAS model included the effects of multiple QTLs and population structure and was based on the Bayesian method (Iwata et al. 00; Yamasaki and Ideta 01). A marker was regarded as significant if its mean gamma value was higher than the specific threshold of gamma = 0.1. Quantitative trait loci analysis Simple sequence repeat (SSR) markers spread throughout the genome were used for the QTL analysis. The SSR markers were selected from those described in McCouch et al. (00) and the International Rice Genome Sequencing Project (00). Polymorphisms
8 were detected as described by Ebana et al. (0). Genomic DNA was extracted from leaves of each plant by the cetyl trimethylammonium bromide (CTAB) method. A total of and 1 markers were used to analyze F (Akenohoshi/Takanari) plants and backcrossed inbred lines (BILs) (Jarjan/Koshihikari//Koshihikari), respectively. Linkage maps were constructed with MAPMAKER/EXP.0 software (Lander et al. 1) using the Kosambi map function. The length and width of the flag leaf were the averages of the three largest flag leaves per plant in the F population (Akenohoshi/Takanari) or averages of the largest flag leaf of five plants per line in the BIL population (Jarjan/Koshihikari//Koshihikari). The QTL analyses were performed by composite interval mapping as implemented by QTL Cartographer. software ( Genome-wide threshold values (α = 0.0) were used to detect QTLs based on the results of 00 permutations (Churchill and Doerge 1). One-way analysis of variance The accessions from the NIAS Core Collection were divided into two groups according to the th nucleotide ( rd amino acid) of their NAL1 gene; accessions had a guanine (arginine) residue and 1 accessions had an adenine (histidine) residue. To test the equality of the two groups, one-way analysis of variance for length and width of the flag leaf, number of vascular bundles, distance between small vascular bundles, and thickness of the flag leaf was performed using Microsoft Excel 00. The variance between two groups was divided by the total variance to obtain the coefficient of determination (R ). Production and evaluation of transgenic plants Details of the production of transgenic plants are provided in File S1. The copy number of transgenes in T 0 and T 1 plants was determined based on the amount of NAL1 genes relative to ubiquitin (RUBQ, AF) genes, as determined by quantitative real-time PCR. The primers used are shown in Table S. The T 1 plants were classified into groups depending on the copy number of transgenes, and each group was compared to the vector control group using the F-test and t-test in
9 Microsoft Excel 00. To identify the T 1 plant groups that differed significantly from the vector control group, we used Tukey s Honest Significant Difference method in the multcomp package of R v...0 software (R foundation, Vienna, Austria). Molecular phylogenetic analysis The transcribed region of NAL1 was amplified from the accessions from the Global Core Collection (WRC) (Kojima et al. 00) by PCR with the primers shown in Table S. The PCR products were sequenced, and phylogenetic analyses were conducted with MEGA (Tamura et al. 01). The complete deletion option was selected to use only sites shared among aligned sequences, and the T + G model was selected as the best-fit model with the lowest Bayesian information criterion score. The maximum likelihood method was used to infer the phylogenetic tree and ancestral nucleotide sequences. Homology searches in databases of other species Homology searches were conducted using TBLASTN with the predicted NAL1 protein sequence of an accession from the NIAS Core Collection, Bei Khe (WRC 0), as the query. The searches were conducted against the genome databases of plants shown in Table SB and the sequence with the lowest E-value was selected for each species. Real-time PCR analysis Details on determining the transcript levels of NAL1 at various developmental stages and the transgene copy number in transgenic plants are provided in File S1. The primers used for real-time PCR are shown in Table S. TaqMan real-time PCR was performed as described previously (Kojima et al. 00). All TaqMan probes (Operon) were -labeled with Black Hole Quencher-1a dye. The RUBQ probe was -labeled with VIC, and HPT and NAL1 probes were -labeled with the reporter dye FAM. RESULTS First component of principal component analysis explained % of total variance and width of flag leaf (WFL) had highest loading
10 To identify the morphological characters of the flag leaf that primarily contribute to the natural variations in rice, we evaluated the following nine traits: length of the flag leaf (LFL), width of the flag leaf (WFL), number of large vascular bundles (NLVB), number of small vascular bundles (NSVB), number of small vascular bundles between large vascular bundles (NSVB bet LVB), distance between small vascular bundles (DSVB), and thickness of flag leaf at the point of the large vascular bundle (TFL_LVB), at the point of the small vascular bundle (TFL_SVB), and at the point of the motor cell (TFL_MC) (Table S right). There were positive correlations (1) among the four traits WFL, NLVB, NSVB, and NSVB bet LVB, () between DSVB and WFL, TFL_SVB, or TFL_MC (Table S), and () among the three thickness of flag leaf traits. The highest estimated correlation coefficient was 0.. The above nine traits were used in the PCA. This analysis covered accessions from the NIAS Core Collection that represented the natural variations in O. sativa (Table 1). In the PCA, the first three components accounted for more than 0% of the total variance. PC1 and PC accounted for.% and.% of total variance, respectively, and the following variables showed substantial loadings: WFL, NLVB, NSVB, and NSVB bet LVB for PC1; TFL_LVB, TFL_SVB, and TFL_MC for PC; LFL and DSVB for PC. WFL and its correlated traits were, therefore, the most significant characters to distinguish the natural variations in the flag leaf morphology of rice. Scatter plots of PC1/PC or PC/PC exhibited overlapping but distinct distributions of the indica, temperate japonica, and tropical japonica subgroups (Figure S1, see also Table S) and reflected the morphological features of the three cultivars. Flag leaves tended to be thin and short with a moderate width in indica accessions; narrow, thick, and long in temperate japonica accessions; and wide, thick, and short in tropical japonica accessions. QTLs on chromosomes and contributed to natural variations in WFL The GWAS for WFL (Figure 1A) identified a marker RM at 0. Mb on chromosome with a gamma value greater than 0.1 over years. Another marker, Rik1, at. Mb on chromosome, also had a gamma value greater than 0.1 in
 as the donor, one with a temperate japonica accession (LAC) as the donor, and one with Oryza rufipogon (IRGC-ACC1) as the donor, in the Koshihikari background.")

11 , while no marker with gamma values greater than 0.1 were detected for LFL. We conducted whole-genome analyses of four CSSLs; two with indica accessions of O. sativa (Kasalath and IR) as the donor, one with a temperate japonica accession (LAC) as the donor, and one with Oryza rufipogon (IRGC-ACC1) as the donor, in the Koshihikari background. These analyses, except for one using the LAC as the donor, identified a QTL for WFL on the long arm of chromosome but no QTLs on chromosome (Figure 1B). The variation in WFL was further studied by a QTL analysis followed by a mapping study using recombinant fixed lines. The analyses of the F population derived from a Takanari Akenohoshi cross revealed two QTLs, one of which was located in the region near. Mb on chromosome (Table S, Figure SA). Three QTLs, including one located in the region near 1. Mb on chromosome, were identified in the analysis of BILs derived from a BC 1 F 1 (Jarjan/Koshihikari//Koshihikari) plant (Table S). In both analyses, QTLs for WFL were not linked to markers for heading date (HD), whereas QTLs for LFL and for HD on chromosome were linked to the same marker. QTL for WFL on chromosome was located within a.-kb region We analyzed the effect of the QTL on chromosome on WFL using a pair of lines with
12 the same fixed Takanari/Akenohoshi background except at two regions, one at 1..0 Mb on chromosome and the other at.. Mb on chromosome, which has no QTLs (Figure SA and SB). Compared with the line with the Takanari-derived allele, the line with the Akenohoshi-derived allele at the 1..0 Mb region on chromosome showed greater WFL, spikelet number, hulled grain weight per plant, and panicle weight per growing area (Figure SB). This result indicated that the 1..0 Mb region on chromosome likely contained a QTL for WFL and/or yield. We further examined an additional recombinant fixed lines with the same background as that of F line_ or F line_---1- (Figure SB). These analyses further restricted the QTL to a. kb-region within the 1..0 Mb region on chromosome. This region contained only three ORFs; Os0t000 (NARROW LEAF 1), Os0t000, and Os0t00 (Figure SC). Among the three genes, NAL1 had three nucleotide differences that would alter the encoded amino acid (one in exon and the other two in exon ). Another difference between Takanari- and Akenohoshi-derived alleles was the length of a poly-glutamine region, that is, a difference in the number of CTG repeats, in exon of Os0t00 (Table A, Figure SC). Given that NAL1 was the best candidate for the QTL for WFL among the three ORFs, we examined its promoter region, since a previous report suggested that some NAL1 alleles were expressed at different levels (Zhang et al. 01). A pair of recombinant fixed lines, F line_ and - (Figure SD), which had either the Takanari or Akenohoshi fragment in the upstream promoter region but shared the Akenohoshi coding region, exhibited the same WFL and yield. The promoter region accounted for none or little of the variations in WFL and/or yield (Figure SD), suggesting that mutations in the coding region, not the promoter region, caused the variation in WFL in this case. RH amino acid substitution in NAL1 caused variation in WFL and its phenotypic effect depended on ratio of copy number of R -type to H -type Eleven CSSLs with different donor fragments at the.-kb region in the Koshihikari 1

13 background (Figure ) were used to identify the causative mutation of the QTL for WFL. Of the CSSLs shown in Table B, only two had the same WFL as Koshihikari (Table B, right). The donors of these CSSLs were Hayamasari or Tupa-. Among the four differences mentioned above, only the RH amino acid difference in NAL1 coincided with the observed differences in WFL (Table B, left). To gain insight into the RH amino acid mutation, we compared eight traits between Koshihikari and CSSLs whose donor was one of four O. sativa accessions (Hayamasari, Tupa -, Khao Nam Jen, Bei Khe) or one of two O. rufipogon accessions (IRGC-ACC1, IRGC-ACC) (Figure S, Figure ). The seven traits besides WFL were LFL, NLVB, NSVB, TFL_SVB, DSVB, number of cells between motor and epidermal cells (NCME), and number of cells between large vascular bundle and epidermal cells (NCLVBE). We also compared these traits between a pair of recombinant fixed lines that had a Takanari fragment (F _---1-) or an Akenohoshi fragment (F _---1-1) at the 1..0 Mb region (Figure SB). 1



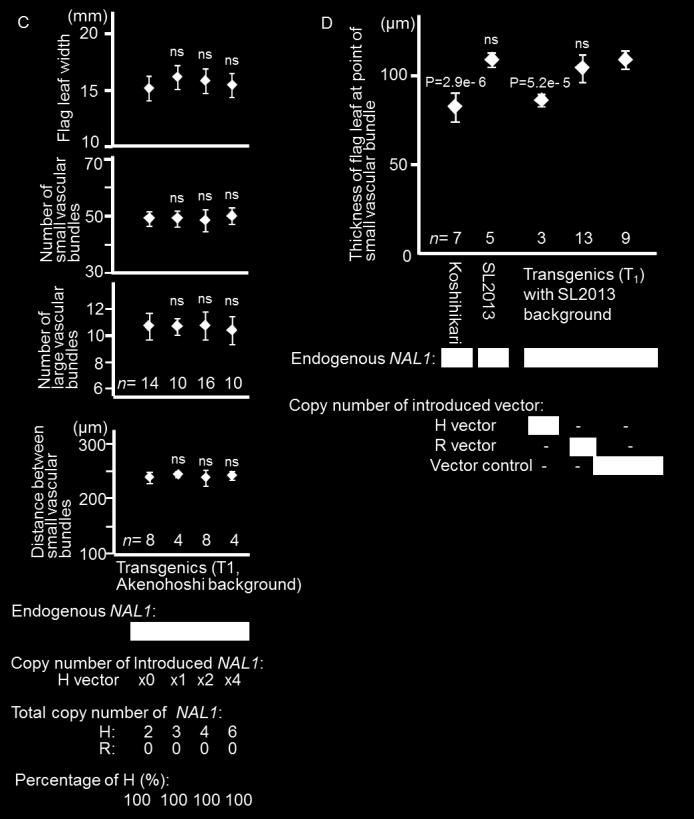
14 1
15 These analyses confirmed that the rd amino acid substitution in NAL1 affected NSVL, NCME, and NCLVBE, as well as WFL (Figure S). To further explore the effect of the RH amino acid mutation on WFL, we transformed two lines (SL01 with R-type NAL1, and Akenohoshi with H-type NAL1) with either Takanari NAL1 or the Takanari NAL1_RH vector (see File S1). We observed the phenotypes of the T 0 and T 1 generations (Figure ), and found that the WFL increased when the H construct was introduced into the R-type background, whereas the WFL decreased when the R construct was introduced into the H-type background. There was no apparent effect when the R construct was introduced into the R-type background, or when the H construct was introduced into the H-type background. In the R-type background (Figure B), WFL was correlated with NSVB and DSVB but not with NLVB. Interestingly, the WFL, NSVB, and DSVB of transgenic plants increased as the copy number of H-type alleles increased from zero to two, but their values did not exceed those of Koshihikari (H-type) even if the copy number of H-type alleles was more than two. In the H-type background (Figure C), the copy number of the H-type allele had little or no effect on traits related to WFL. Therefore, the proportion rather than the actual copy number of introduced vectors (H-type alleles) was important for WFL. Interestingly, the wider flag leaves with increased NSVB and DSVB were thinner at the point of the small vascular bundles (Figure D). RH amino acid mutation occurred during O. sativa domestication, and accounted for 1% of total variation in WFL among rice accessions from the global collection We sequenced the transcribed region of the NAL1 gene from accessions from the NIAS Global Core Collection, the four parents of the CSSLs, and the two parents of the hybrid population used in the QTL analysis (Table SA). The obtained sequences were classified into seven types according to their predicted amino acid sequences (Figure ). Notably, accessions including Nipponbare or Koshihikari had a retrotransposon insertion in exon 1 with duplicated regions at the end of exon 1 and the end of exon (type IV ), although the nucleotide sequences in the coding region were intact and 1

.")
16 essentially the same as that in Akenohoshi (type IV). The type IV allele was also present in LAC and Jarjan, which explained why no QTL on 1 chromosome 1 was detected 1 in the analysis 1 of a CSSL 1 with LAC 1 as donor and 1 from BILs derived from BC 1 F 1 (Jarjan/Koshihikari//Koshihikari). One accession, Phulba, had an allele in which the deletion of a large part of the inserted retrotransposon resulted in a stop codon near the end of exon 1 (type V). Using the nucleotide sequences of only the shared region, and excluding the retrotransposon region, the maximum likelihood tree was estimated by MEGA by inferring ancestral amino acid sequences under the best-fit evolutionary model of T + gamma (Figure ). As for the rd amino acid, accessions with R or H were distributed in all three subgroups; indica, temperate japonica, and tropical japonica. However, most of the accessions with R were in the indica subgroup. Exon of NAL1 was sequenced for accessions from wild rice species, and all had G at the th nucleotide and encoded the amino acid R at the rd residue (Table SA). Interestingly, the survey of NAL1 homologs in the genome databases of plants of various taxa (Table SB) revealed that species in the Streptophyta, including land plants 1


17 (Embryophyta) had NAL1 homologs with R at the corresponding amino acid position, while species in the Rhodophyta or Chlorophyta had no detectable NAL1 homolog. This finding suggested that the original amino acid residue at this position in the rice NAL1 1
.")

18 was R, which later mutated into H. We conducted a one-way analysis of variance of accessions from the NIAS Core Collection to examine the functional significance of the GA nucleotide substitution (RH amino acid substitution). The results showed that the substitution accounted for 1%, 0%, and % of the total variation in WFL, DSVB, and NSVB, respectively (Table ). H -type NAL1 gene is likely to be a malfunctional allele One accession in the NIAS Core Collection, Phulba, had an allele encoding a truncated NAL1 protein of only 1 amino acids, in contrast to the full NAL1 with amino acids (type V in Figure ). The phenotypes were compared between Koshihikari with the H-type allele (type IV in Figure ) and a CSSL containing a Phulba-derived fragment of NAL1 in the Koshihikari background (Figure ). There were no significant differences in any of the traits (Figure ), indicating that the phenotypes of the Phulba allele and H-type allele were essentially the same, and both were likely to be 1
19 malfunctional. NAL1 expression peaked at stage P when small vascular bundles form Immature flag leaves were sampled to survey the transcript levels of NAL1 throughout flag leaf development. Leaves at stage P and at the beginning of stage P were too small to excise intact, and so the whole shoot apex was used instead. In the period from stage P to P, NAL1 transcript levels peaked at stage P. At this stage, there was no difference in NAL1 transcript levels among IR (R-type; type I in Figure ), Koshihikari (H-type; type IV ), and Phulba (truncated protein; type V) (Figure ). DISCUSSION H in NAL1 was likely selected during rice domestication Seven different NAL1 types have been detected among the rice accessions examined so far. The progenitor of cultivated rice had R in its NAL1 protein, given that all accessions from wild rice species including O. rufipogon, from which O. sativa originated, had R (Table SA). The presence of the R-type allele of NAL1 in land plants also suggests that the R-type allele is the ancestral form, and the H-type allele is derived from it (Figure ). The RH amino acid mutation, the insertion of a retrotransposon, and the subsequent deletion of the retrotransposon distinguished type IV from type III, type IV from type IV, and type V from type IV, respectively (Figure ). Our analyses raise the possibility that the three major subgroups of cultivated rice differentiated after the insertion of the retrotransposon into the H-type allele, as both R (type I, II, III, VI and VII) and H (type IV and IV ) were present among the indica, temperate japonica, and tropical japonica subgroups (Figure, Table SA). According to Huang et al. (01), ancient japonica was first domesticated from a specific population of O. rufipogon in southern China. Subsequently, indica originated from crosses between ancient japonica and local O. rufipogon as the initial cultivar of temperate japonica spread into South East and South Asia. Considering this pattern of distribution, we propose an evolutionary scenario that the RH amino acid substitution and the following retrotransposon insertion occurred at the differentiation of 1
20 ancient japonica, and that the temperate japonica, indica, and tropical japonica accessions with H-type alleles originated from crosses between O. rufipogon and ancient japonica harboring the mutation. Although further population genetics studies are required to confirm this scenario, the current data suggest that the H-type allele has been selected for during rice domestication. The evidence for this hypothesis can be summarized as follows: the angiosperms examined so far have retained R-type NAL1 homologs for a long time during evolution, implying a selective advantage of R-type alleles over others in the natural environment. However, both H-type and R-type alleles have persisted among the three rice subgroups during the history of rice cultivation. The H-type allele directly or indirectly leads to increased WFL, number of spikelets per panicle, and often yields, although its effects vary among accessions (see below). Therefore, the H-type allele seems to have provided some desirable characters, e.g., increased grain yields, with little, if any, adverse effects under artificially controlled environments. We propose that NAL1 has been one of the important genes in the domestication of cultivated rice. Amino acid change in NAL1 is important for increased WFL Previous reports have suggested that there are differences among rice accessions in terms of the quantity and quality of NAL1 gene expression. It was reported that the H-type allele in HB but not the R-type allele in D0 was subject to extensive alternative splicing, while both were expressed at similar levels (Chen et al. 01). In Koshihikari (type VI ), only 0% of NAL1 transcripts contained no retrotransposon and a NAL1 protein the same size as that encoded by the R-type allele was synthesized (Takai et al. 01). Furthermore, it was shown that NAL1 expression was significantly higher in Nipponbare (type IV ) than in - (type I), although the expression level was shown to vary among developmental stages (Zhang et al. 01). Overexpression of the Nipponbare allele in Nipponbare increased FWL and LWL through increasing the distance between vascular bundles (Zhang et al. 01), indicating that the NAL1 transcript level is an important factor in leaf morphology. Nonetheless, most of the current evidence indicates that the different effects of NAL1 types on flag leaf phenotypes are mainly due to the amino acid at the rd 0
21 position, rather than changes in transcript levels. First, there was no phenotypic difference between a pair of recombinant fixed lines with the same H -type NAL1 driven by either the Takanari or Akenohoshi promoter (Figure SD). Second, there was no difference in gene expression among the IR allele (type I), the Koshihikari allele (type IV ), and the Phulba allele (type V) at stage P (Figure, Figure B). Third, transformation by two vectors driven by the same Takanari promoter with different amino acids at the rd residue resulted in marked differences in phenotype (Figure ). Products of the malfunctional H -type allele still regulate WFL and compete with R -type products Although the H-type allele is thought to be malfunctional, it must retain some function because the phenotype of plants homozygous for H-type alleles differed from that of plants homozygous for the null allele. The leaves of plants with the null allele were 0% narrower and shorter than those of the H-type plants (Nipponbare, type IV ) (Jiang et al. 01). The H-type allele was neither recessive nor dominant, as transgenic plants with both the R-type allele and the H-type allele exhibited intermediate phenotypes (Figure B). The effect on phenotype did not depend on the dosage of R-type or H-type genes, but on the ratio of the copy numbers of R-type to H-type (Figure ). Therefore, the H -type NAL1 products likely compete with R -type products to regulate WFL. Notably, the deletion of two amino acids in exon 1 resulted in a phenotype similar to that of the null mutant, with a 0% reduction in the width and length of the leaf (Jiang et al. 01). Compared to a deletion in exon 1, a -amino-acid deletion in exon had a milder effect; it reduced the blade width by % and the blade length by 1% (Qi et al. 00). The Phulba allele, which produces a truncated protein consisting only of exon 1 (type V), resulted in a phenotype similar to that of the H-type allele (type IV ), which appeared to have no obvious defects (Figure ). Therefore, the protein domain encoded by exon 1 seems to be essential for the regulation of WFL and LFL, as well as the competition of R -type and H -type products. NAL1 regulates flag leaf width and exhibits various pleiotropic effects NAL1 derived from Nipponbare (type IV ) was shown to promote cell division in the 1
22 anticlinal direction and suppress it in the periclinal direction with no or little effect on cell size in the leaf (Jiang et al. 01). This is consistent with our observations, where WFL, NSVB, DSVB were larger and TFL_SVB was smaller in the line transformed with the H-type allele (Figure ). Takai et al. (01) reported that the R-type allele derived from Takanari (type I) increased leaf thickness and the H-type allele derived from Koshihikari (type IV ) decreased leaf thickness. These results can be explained if the R-type allele is regarded as an allele with a weaker effect than the H-type allele on cell division in the anticlinal and periclinal directions. This would also explain the increase in WFL when the Daringan allele (H-type allele) was introduced into IR (R-type allele) (Fujita et al. 01) and when the Nipponbare allele (H-type allele) was introduced into - (R-type allele) (Zhang et al. 01). However, it could not explain the results of Chen et al. (01), where the R-type allele increased WFL in mapping of QTLs for WFL using hybrid populations originated from a cross between D0 (R-type allele) and HB (H-type allele). This discrepancy might be because of the presence of a large number of alternatively spliced forms of the H-type allele and a low level of the NAL1 protein (Chen et al. 01). However, further analyses are required to identify the exact reason for this discrepancy. Gene expression of NAL1 was observed as early as the formation of leaf primordia (Jiang et al. 01). NAL1 was highly expressed in the vascular bundle, especially in the phloem (Qi et al. 00) and its expression peaked at the P stage when the small vascular bundle formed. Therefore, its main effect appears to be on the formation of vascular bundles, especially small vascular bundles, which subsequently affect leaf width and morphology. Nonetheless, the RH amino acid mutation of NAL1 exhibits a pleiotropic effect. Besides the increase in WFL and the number of vascular bundles, Nal1 increased the total number of spikelets per panicle, root dry weight, and the rate of filled grains (Fujita et al. 01). Improvements to the panicle, such as increased panicle length, more spikelets per panicle, and more secondary branches per panicle, were also observed after introducing the H-type allele (Zhang et al. 01). Since NAL1 is expressed in the culm, coleoptile, crown root, lateral root, and panicles (Qi et al. 01; Fujita et al. 01; Jiang et al. 01), its regulation of the cell cycle and cell division in these tissues likely explain the pleiotropic effects of the Nal1 mutation.
23 With respect to the grain yields, the effects of R-type and H-type alleles seem to vary. Higher photosynthetic rate per area was attained when the Koshihikari allele (H-type) was replaced by the Takanari allele (R-type) due to the increased number of mesophyll cells between vascular bundles and the larger total mesophyll area between the vascular bundles (Takai et al. 01). Meanwhile, when IR (R-type allele) was replaced by the Daringan allele (H-type allele), grain yield per area increased despite the decreased number of panicles per plant and lower 00-grain weight (Fujita et al. 01). When the - allele (R-type) was replaced by the Nipponbare allele (H-type), the yield and chlorophyll content in the leaf increased, and panicle morphology was affected (Zhang et al. 01). Grain yield is also affected by other factors including the primary structure and expression level of NAL1, the background genome, and the growth conditions/environmental factors. Therefore, the outcomes of different types of NAL1 alleles in various rice accessions are not easily predicted. NAL1 may be involved in auxin transport and response It has been speculated that NAL1 is involved in polar auxin transport, since a nal1 mutant lacking amino acids from exon produced fewer vascular bundles and contained lower levels of OsPIN1 protein, compared with wild type (Qi et al. 00). The fact that the expression of the AUXIN RESPONSE FACTOR gene family (ARF1, ARF, and ARF) and PIN1 was reduced in the NAL1 null mutant (Jiang et al. 01) supports the involvement of NAL1 in the auxin response. In this regard, it is notable that a NAL1 homolog was found in Klebsormidium flaccidum, a terrestrial alga with a primitive body plan in the Phylum Streptophyta (Table SB). This species produces multicellular and non-branching filaments without differentiated or specialized cells, but its genome contains most of the genes required for auxin biosynthesis, auxin receptors, auxin sensing, and auxin transport. Also, auxin (indole--acetic acid) was detected in tissues of this terrestrial alga by mass spectrometry (Hori et al. 01). Future research, including studies on the Phylum Streptophyta, will shed light on the function of NAL1 and its involvement in the auxin response. 1
24 Acknowledgments We thank Ms. Akemi Tagiri for thin sectioning and Ms. Haruko Onodera for production of transgenic plants. The wild rice accessions used in this study were obtained from the National Institute of Genetics, supported by the National Bioresource Project, MEXT, Japan. This work was supported by the NIAS technical support system and by grants from the Ministry of Agriculture, Forestry and Fisheries of Japan (Genomics for Agricultural Innovation NVR-0001 and QTL-0, and Genomics-based Technology for Agricultural Improvement IVG-00). Literature Cited Abe, T., Y. Nonoue, N. Ono, M. Omoteno, M. Kuramata et al., 01 Detection of QTLs to reduce cadmium content in rice grains using LAC/Koshihikari chromosome segment substitution lines. Breed Sci. : 1. Abe, T., F. Taguchi-Shiobara, Y. Kojima, T. Ebitani, M. Kuramata et al., 0 Detection of a QTL for accumulating Cd in rice that enables efficient Cd phytoextraction from soil. Breed. Sci. 1: 1. Chen, M., J. Luo, G. Shao, X. Wei, S. Tang, et al., 01 Fine mapping of a major QTL for flag leaf width in rice, qflw, which might be caused by alternative splicing of NAL1. Plant Cell Rep. 1: -. Churchill, G. A., and R. W. Doerge, 1 Empirical threshold values for quantitative trait mapping. Genetics 1: 1. de Bakker P. I. W., R. Yelensky, I. Pe er, S. B. Gabriel, M. J. Daly et al., 00 Efficiency and power in genetic association studies. Nat. Genet. : -1. Ebana, K., Y. Kojima, S. Fukuoka, T. Nagamine, and M. Kawase, 00 Development of mini core collection of Japanese rice landrace. Breed. Sci. : 1 1. Ebana, K., T. Shibaya, J. Wu, K. Matsubara, H. Kanamori, et al., 0 Uncovering of major genetic factors generating naturally occurring variation in heading date among Asian rice cultivars. Theor. Appl. Genet. 1():-0. Ebitani, T., Y. Takeuchi, Y. Nonoue, T. Yamamoto, K. Takeuchi et al., 00, Construction and Evaluation of Chromosome Segment Substitution Lines Carrying
25 Overlapping Chromosome Segments of indica Rice Cultivar 'Kasalath' in a Genetic Background of japonica Elite Cultivar 'Koshihikari'. Breed. Sci. :-. Esau, K., 1, Anatomy of seed plants, nd edition. Wiley & Sons, New York. Fujita, D., K. R. Trijatmiko, A. G. Tagle, M. V. Sapasap, Y. Koide et al., 01 NAL1 allele from a rice landrace greatly increases yield in modern indica cultivars. Proc. Natl. Acad. Sci. USA 0: 01 0 Fuse, T., T. Sasaki, and M. Yano, 001 Ti-plasmid vectors useful for functional analysis of rice genes. Plant Biotechnol. 1: 1-. Hirabayashi, H., H. Sato, Y. Nonoue, Y. Kuno-Takemoto, Y. Takeuchi et al., 0 Development of introgression lines derived from Oryza rufipogon and O. glumaepatula in the genetic background of japonica cultivated rice (O. sativa L.) and evaluation of resistance to rice blast. Breed. Sci. 0: 0 1. Hori, K., F. Maruyama, T. Fujisawa, T. Togashi, N. Yamamoto et al., 01 Klebsormidium flaccidum genome reveals primary factors for plant terrestrial adaptation. Nat. Commun. : doi:./ncomms Hori K, Nonoue Y, Ono N, Shibaya T, Ebana et al., 01 Genetic architecture of variation in heading date among Asian rice accessions. BMC Plant Biology 1:./s x Hoshikawa, K., 1 The Growing Rice Plant: An Anatomical Monograph. Nobunkyo, Tokyo. Huang, X., N. Kurata, X. Wei, Z.-X. Wang, A. Wang et al., 01 A map of rice genome variation reveals the origin of cultivated rice. Nature 0: -01. Inosaka, M., 1 Studies on the development of vascular system in rice plant and the growth of each organ viewed from the vascular connection between them. Miyazaki Daigaku Nogakubu, Kenkyu Hokoku : 1-. International Rice Genome Sequencing Project, 00 The map-based sequence of the rice genome. Nature : -00. Itoh, J.-I., K.-I. Nonomura, K. Ikeda, S. Yamaki, Y. Inukai et al., 00 Rice plant development: from zygote to spikelet. Plant Cell Physiol. :. Iwata, H., Y. Uga, Y. Yoshioka, K. Ebana, and T. Hayashi, 00 Bayesian association mapping of multiple quantitative trait loci and its application to the analyses
26 of genetic variation among Oryza sativa L. germplasms. Theor. Appl. Genet. : 1-1. Jiang D., J. Fang, L. Lou, J. Zhao, S. Yuan et al., 01 Characterization of a null allelic mutant of the rice NAL1 gene reveals its role in regulating cell division. PLoS ONE (): e01. Doi:.11/journal.pone.01. Kojima, Y., K. Ebana, S. Fukuoka, T. Nagamine, and M. Kawase, 00 Development of an RFLP-based rice diversity research set of germplasm. Breed. Sci. : 1 0. Kojima, S., Y. Takahashi, Y. Kobayashi, L. Monna, T. Sasaki et al., 00 Hda, a rice ortholog of the Arabidopsis ft gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. : -0. Lander, E.S., P. Green, J. Abrahamson, A. Barlow, M.J. Daly et al., 1 MAPMAKER: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1: Langdale, J. A., B. Lane, M. Freeling, and T. Nelson, 1 Cell lineage analysis of maize bundle sheath and mesophyll cells. Dev. Biol. 1:1 1. Matsuo, T., 1 Genecological studies on cultivated rice. Bulletin of Nat. Inst. Agr. Sci. D : 1-1. McCouch, S.R., L. Teytelman, Y. Xu, K.B. Lobos, K. Clare, et al., 00 Development and mapping of 0 new SSR markers for rice (Oryza sativa L.). DNA Res. : 1 0. Nagasaki, H., K. Ebana, T. Shibaya, J. Yonemaru, and M. Yano, 0 Core-single nucleotide polymorphisms-a tool for genetic analysis of the Japanese rice population. Breed. Sci. 0: -. Nonoue, Y., K. Matsubara, N. Ono, R. Mizobuchi, T. Shibaya et al., 0 Development of reciprocal chromosome segment substitution lines between a japonica cultivar Koshihikari and an indica cultivar IR in rice. Breed. Sci. Suppl. 1 ():. Qi, J., Q. Qian, Q. Bu, S. Li, Q. Chen et al., 00 Mutation of the rice Narrow leaf1 gene, which encodes a novel protein, affects vein patterning and polar auxin transport. Plant Physiol. 1:1-1. Rasband, W. S., 1-01 ImageJ, U. S. National Institutes of Health, Bethesda,
27 Maryland, USA, Sakamoto, T., Y. Morinaka, T. Ohnishi, H. Sunohara, S. Fujioka et al., 00 Erect leaves caused by brassinosteroid deficiency increase biomass production and grain yield in rice. Nature Biotechnology : -. Sinclair, T. R., and J. E. Sheehy, 1 Erect leaves and photosynthesis in rice. Science :1. Taguchi-Shiobara, F., Y. Kojima, T. Ebitani, M. Yano, K. Ebana, 0 Variation in domesticated rice inflorescence architecture revealed by principal component analysis and quantitative trait locus analysis. Breed. Sci. 1: 0. Taguchi-Shiobara, F., H. Ozaki, H. Sato, H. Maeda, Y. Kojima et al., 01 Mapping and validation of QTLs for rice sheath blight resistance Breed. Sci. :01-0. Takai, T., S. Adachi, F. Taguchi-Shiobara, Y. Sanoh-Arai, N. Iwasawa et al., 01 A natural variant of NAL1, selected in high-yield rice breeding programs, pleiotropically increases photosynthesis rate. Sci. Rep. : 1. Tamura K., G. Stecher, D. Peterson, A. Filipski, S. Kumar, 01 MEGA: Molecular evolutionary genetics analysis version.0. Mol. Biol. Evol. 0: -. Tanaka, A., 1 Studies on the characteristics of the physiological function of leaf at definite position on stem of rice plant. (Part ): comparison of photosynthetic activity of leaves at various position on main stem and translocation of photosynthetic products within plant. Jpn. J. Soil Sci. Plant Nutr. : -. Toki, S., 1 Rapid and efficient Agrobacterium-mediated transformation in rice. Plant Mol. Biol. Rept. 1: 1-1. Watson, D. J., 1 The Physiological Basis of Variation in Yield. Advan. Agron. : 1-1. Yamasaki, M., and O. Ideta, 01 Population structure in Japanese rice population. Breed. Sci. : -. Yonemaru, J., K. Ebana, M. Yano, 01 HapRice, a SNP haplotype database and a web tool for rice. Plant Cell Physiol. (1): e(1 1) doi:./pcp/pct1. Yoshida, S., 1 Physiological Aspects of Grain Yield. Annu. Rev. Plant Physiol. : -. Zhang G.-H., S.-H. Li, L. Wang, W.-J. Ye, D.-L. Zeng, et al., 01 LSCHL from
28 japonica cultivar, which is allelic to NAL1, increase yield of indica super rice -. Mol. Plant : -1. Zhao, K., C.-W. Tung, G. Eizenga, M. Wright, M. L. Ali et al., 0 Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat. Commun. : doi:./ncomms1.
29 Table 1. Eigenvectors for nine flag leaf morphology traits in principal component analysis of rice accessions from NIAS Core Collection Principal component 1st nd rd Eigenvalue Contribution ratio.. 1. Cumulative contribution Trait Abbreviation Length of flag leaf LFL Width of flag leaf WFL Number of large vascular bundles NLVB Number of small vascular bundles NSVB Number of small vascular bundles between large vascular bundles NSVB bet LVB Distance between small vascular bundles DSVB Thickness of flag leaf at point of large vascular bundle a TFL_LVB at point of small vascular bundle b TFL_SVB at point of motor cell b TFL_MC a Second large vascular bundle from midrib. b Near second large vascular bundle from midrib.
30 Table. Four amino acid-altering differences in.-kb region harboring QTL for flag leaf width. A. Amino acid-altering nucleotide differences present in exon regions in.-kb region. NARROW LEAF 1 (NAL1) Os0t00 rd exon th exon nd exon Nucleotide site Number of CTG repeats starting from th nucleotide Amino acid residue Number of glutamine residues starting from th amino acid Parents used in F analysis Takanari G (R: arginine) C(A: alanine) G(V: valine) Akenohoshi A (H: histidine) T(V: valine) A (I: isoleucine) 0
Genetic and physiological approach to elucidation of Cd absorption mechanism by rice plants
 Genetic and physiological approach to elucidation of Cd absorption mechanism by rice plants Satoru Ishikawa National Institute for Agro-Environmental Sciences, 3-1-3, Kannondai, Tsukuba, Ibaraki, 305-8604,
Genetic and physiological approach to elucidation of Cd absorption mechanism by rice plants Satoru Ishikawa National Institute for Agro-Environmental Sciences, 3-1-3, Kannondai, Tsukuba, Ibaraki, 305-8604,
Segregation distortion in F 2 and doubled haploid populations of temperate japonica rice
 c Indian Academy of Sciences RESEARCH NOTE Segregation distortion in F 2 and doubled haploid populations of temperate japonica rice MASUMI YAMAGISHI 1,2,6, YOSHINOBU TAKEUCHI 3,7, ISAO TANAKA 4, IZUMI
c Indian Academy of Sciences RESEARCH NOTE Segregation distortion in F 2 and doubled haploid populations of temperate japonica rice MASUMI YAMAGISHI 1,2,6, YOSHINOBU TAKEUCHI 3,7, ISAO TANAKA 4, IZUMI
Genetic diversity and population structure in rice. S. Kresovich 1,2 and T. Tai 3,5. Plant Breeding Dept, Cornell University, Ithaca, NY
 Genetic diversity and population structure in rice S. McCouch 1, A. Garris 1,2, J. Edwards 1, H. Lu 1,3 M Redus 4, J. Coburn 1, N. Rutger 4, S. Kresovich 1,2 and T. Tai 3,5 1 Plant Breeding Dept, Cornell
Genetic diversity and population structure in rice S. McCouch 1, A. Garris 1,2, J. Edwards 1, H. Lu 1,3 M Redus 4, J. Coburn 1, N. Rutger 4, S. Kresovich 1,2 and T. Tai 3,5 1 Plant Breeding Dept, Cornell
Nature Genetics: doi: /ng Supplementary Figure 1. The phenotypes of PI , BR121, and Harosoy under short-day conditions.
 Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Eiji Yamamoto 1,2, Hiroyoshi Iwata 3, Takanari Tanabata 4, Ritsuko Mizobuchi 1, Jun-ichi Yonemaru 1,ToshioYamamoto 1* and Masahiro Yano 5,6
 Yamamoto et al. BMC Genetics 2014, 15:50 METHODOLOGY ARTICLE Open Access Effect of advanced intercrossing on genome structure and on the power to detect linked quantitative trait loci in a multi-parent
Yamamoto et al. BMC Genetics 2014, 15:50 METHODOLOGY ARTICLE Open Access Effect of advanced intercrossing on genome structure and on the power to detect linked quantitative trait loci in a multi-parent
Classical Selection, Balancing Selection, and Neutral Mutations
 Classical Selection, Balancing Selection, and Neutral Mutations Classical Selection Perspective of the Fate of Mutations All mutations are EITHER beneficial or deleterious o Beneficial mutations are selected
Classical Selection, Balancing Selection, and Neutral Mutations Classical Selection Perspective of the Fate of Mutations All mutations are EITHER beneficial or deleterious o Beneficial mutations are selected
GENOMICS-ASSISTED ALLELE MINING AND ITS INTEGRATION TO BREEDING IN RICE
 GENOMICS-ASSISTED ALLELE MINING AND ITS INTEGRATION TO BREEDING IN RICE Masahiro Yano, Yusaku Uga, Shuichi Fukuoka, Kaoru Ebana, Jun-ichi Yonemaru, Toshio Yamamoto AgroGenomics Research Center, National
GENOMICS-ASSISTED ALLELE MINING AND ITS INTEGRATION TO BREEDING IN RICE Masahiro Yano, Yusaku Uga, Shuichi Fukuoka, Kaoru Ebana, Jun-ichi Yonemaru, Toshio Yamamoto AgroGenomics Research Center, National
RFLP facilitated analysis of tiller and leaf angles in rice (Oryza sativa L.)
") Euphytica 109: 79 84, 1999. 1999 Kluwer Academic Publishers. Printed in the Netherlands. 79 RFLP facilitated analysis of tiller and leaf angles in rice (Oryza sativa L.) Zhikang Li 1,2,3, Andrew H. Paterson
Euphytica 109: 79 84, 1999. 1999 Kluwer Academic Publishers. Printed in the Netherlands. 79 RFLP facilitated analysis of tiller and leaf angles in rice (Oryza sativa L.) Zhikang Li 1,2,3, Andrew H. Paterson
Title. Authors. Characterization of a major QTL for manganese accumulation in rice grain
 Title Characterization of a major QTL for manganese accumulation in rice grain Authors Chaolei Liu, Guang Chen, Yuanyuan Li, Youlin Peng, Anpeng Zhang, Kai Hong, Hongzhen Jiang, Banpu Ruan, Bin Zhang,
Title Characterization of a major QTL for manganese accumulation in rice grain Authors Chaolei Liu, Guang Chen, Yuanyuan Li, Youlin Peng, Anpeng Zhang, Kai Hong, Hongzhen Jiang, Banpu Ruan, Bin Zhang,
Supplementary Methods
 Supplementary Methods Microarray analysis Grains of 7 DAP of the wild-type and gif1 were harvested for RNA preparation. Microarray analysis was performed with the Affymetrix (Santa Clara, CA) GeneChip
Supplementary Methods Microarray analysis Grains of 7 DAP of the wild-type and gif1 were harvested for RNA preparation. Microarray analysis was performed with the Affymetrix (Santa Clara, CA) GeneChip
Principles of QTL Mapping. M.Imtiaz
 Principles of QTL Mapping M.Imtiaz Introduction Definitions of terminology Reasons for QTL mapping Principles of QTL mapping Requirements For QTL Mapping Demonstration with experimental data Merit of QTL
Principles of QTL Mapping M.Imtiaz Introduction Definitions of terminology Reasons for QTL mapping Principles of QTL mapping Requirements For QTL Mapping Demonstration with experimental data Merit of QTL
Improvement of Quantitative Evaluation Method for Plant Type of Rice
 Improvement of Quantitative Evaluation Method for Plant Type of Rice Katsuaki Suzuki 1, Zeyu Zheng 2 and Yutaka Hirata 1 1 Laboratory of Plant Genetics and Biotechnology, Tokyo University of Agriculture
Improvement of Quantitative Evaluation Method for Plant Type of Rice Katsuaki Suzuki 1, Zeyu Zheng 2 and Yutaka Hirata 1 1 Laboratory of Plant Genetics and Biotechnology, Tokyo University of Agriculture
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Genetic Analysis of Heading Date of Japonica Rice Cultivars in Southwest China
 Rice Science, 2011, 18(4): 287 296 Copyright 2011, China National Rice Research Institute Published by Elsevier BV. All rights reserved Genetic Analysis of Heading Date of Japonica Rice Cultivars in Southwest
Rice Science, 2011, 18(4): 287 296 Copyright 2011, China National Rice Research Institute Published by Elsevier BV. All rights reserved Genetic Analysis of Heading Date of Japonica Rice Cultivars in Southwest
Chromosome variations in protoplast-derived calli and in plants regenerated from the calli of
 Jpn. J. Genet. (1989) 64, pp. 355-361 Chromosome variations in protoplast-derived calli and in plants regenerated from the calli of cultivated rice (Oryza sativa L.) Soryu NISHIBAYASHI*, Yasuyuki HAYASHI,
Jpn. J. Genet. (1989) 64, pp. 355-361 Chromosome variations in protoplast-derived calli and in plants regenerated from the calli of cultivated rice (Oryza sativa L.) Soryu NISHIBAYASHI*, Yasuyuki HAYASHI,
Genetic dissection of chlorophyll content at different growth stages in common wheat
 c Indian Academy of Sciences RESEARCH ARTICLE Genetic dissection of chlorophyll content at different growth stages in common wheat KUNPU ZHANG 1,2, ZHIJUN FANG 3, YAN LIANG 1 and JICHUN TIAN 1 1 State
c Indian Academy of Sciences RESEARCH ARTICLE Genetic dissection of chlorophyll content at different growth stages in common wheat KUNPU ZHANG 1,2, ZHIJUN FANG 3, YAN LIANG 1 and JICHUN TIAN 1 1 State
Genetic Analysis and Mapping of Genes for Culm Length and Internode Diameter in Progeny from an Interspecific Cross in Rice
 Plant Breed. Biotech. 2018 (June) 6(2):140~146 https://doi.org/10.9787/pbb.2018.6.2.140 RESEARCH ARTICLE Online ISSN: 2287-9366 Print ISSN: 2287-9358 Genetic Analysis and Mapping of Genes for Culm Length
Plant Breed. Biotech. 2018 (June) 6(2):140~146 https://doi.org/10.9787/pbb.2018.6.2.140 RESEARCH ARTICLE Online ISSN: 2287-9366 Print ISSN: 2287-9358 Genetic Analysis and Mapping of Genes for Culm Length
Analysis of QTLs for panicle exsertion and its relationship with yield and yield-related traits in rice (Oryza sativa L.)
") Analysis of QTLs for panicle exsertion and its relationship with yield and yield-related traits in rice (Oryza sativa L.) C.F. Zhao*, T. Chen*, Q.Y. Zhao, L.H. Zhou, L. Zhao, Y.D. Zhang, Z. Zhu, S. Yao
Analysis of QTLs for panicle exsertion and its relationship with yield and yield-related traits in rice (Oryza sativa L.) C.F. Zhao*, T. Chen*, Q.Y. Zhao, L.H. Zhou, L. Zhao, Y.D. Zhang, Z. Zhu, S. Yao
Identification of quantitative trait loci associated with resistance to brown planthopper in the indica rice cultivar Col.
 Hereditas 144: 48 52 (2007) Identification of quantitative trait loci associated with resistance to brown planthopper in the indica rice cultivar Col.5 Thailand LIHONG SUN 1, YUQIANG LIU 1, LING JIANG
Hereditas 144: 48 52 (2007) Identification of quantitative trait loci associated with resistance to brown planthopper in the indica rice cultivar Col.5 Thailand LIHONG SUN 1, YUQIANG LIU 1, LING JIANG
Outline. Leaf Development. Leaf Structure - Morphology. Leaf Structure - Morphology
 Outline 1. Leaf Structure: Morphology & Anatomy 2. Leaf Development A. Anatomy B. Sector analysis C. Leaf Development Leaf Structure - Morphology Leaf Structure - Morphology 1 Leaf Structure - Morphology
Outline 1. Leaf Structure: Morphology & Anatomy 2. Leaf Development A. Anatomy B. Sector analysis C. Leaf Development Leaf Structure - Morphology Leaf Structure - Morphology 1 Leaf Structure - Morphology
Relationship Between Coleoptile Length and Drought Resistance and Their QTL Mapping in Rice
 Rice Science, 2007, 14(1): 13-20 Copyright 2007, China National Rice Research Institute. Published by Elsevier BV. All rights reserved Relationship Between Coleoptile Length and Drought Resistance and
Rice Science, 2007, 14(1): 13-20 Copyright 2007, China National Rice Research Institute. Published by Elsevier BV. All rights reserved Relationship Between Coleoptile Length and Drought Resistance and
The Relationship between SPAD Values and Leaf Blade Chlorophyll Content throughout the Rice Development Cycle
 JARQ 50 (4), 329-334 (2016) http://www.jircas.affrc.go.jp The Relationship between SPAD Values and Leaf Blade Chlorophyll Content throughout the Rice Development Cycle Yasuyuki WAKIYAMA* National Agriculture
JARQ 50 (4), 329-334 (2016) http://www.jircas.affrc.go.jp The Relationship between SPAD Values and Leaf Blade Chlorophyll Content throughout the Rice Development Cycle Yasuyuki WAKIYAMA* National Agriculture
Title ghd7, on yield components in rice. Citation Molecular breeding (2014), 33(4): 8.
, 33(4): 8.") Title The effects of the photoperiod-inse ghd7, on yield components in rice. Xu, Quan; Saito, Hiroki; Hirose, Ik Author(s) Yoshitake, Yoshihiro; Yokoo, Takayu Teraishi, Masayoshi; Tanisaka, Taka Citation
Title The effects of the photoperiod-inse ghd7, on yield components in rice. Xu, Quan; Saito, Hiroki; Hirose, Ik Author(s) Yoshitake, Yoshihiro; Yokoo, Takayu Teraishi, Masayoshi; Tanisaka, Taka Citation
Evolution of phenotypic traits
 Quantitative genetics Evolution of phenotypic traits Very few phenotypic traits are controlled by one locus, as in our previous discussion of genetics and evolution Quantitative genetics considers characters
Quantitative genetics Evolution of phenotypic traits Very few phenotypic traits are controlled by one locus, as in our previous discussion of genetics and evolution Quantitative genetics considers characters
Exam 1 PBG430/
 1 Exam 1 PBG430/530 2014 1. You read that the genome size of maize is 2,300 Mb and that in this species 2n = 20. This means that there are 2,300 Mb of DNA in a cell that is a. n (e.g. gamete) b. 2n (e.g.
1 Exam 1 PBG430/530 2014 1. You read that the genome size of maize is 2,300 Mb and that in this species 2n = 20. This means that there are 2,300 Mb of DNA in a cell that is a. n (e.g. gamete) b. 2n (e.g.
Computational Biology: Basics & Interesting Problems
 Computational Biology: Basics & Interesting Problems Summary Sources of information Biological concepts: structure & terminology Sequencing Gene finding Protein structure prediction Sources of information
Computational Biology: Basics & Interesting Problems Summary Sources of information Biological concepts: structure & terminology Sequencing Gene finding Protein structure prediction Sources of information
Supplemental Data. Perea-Resa et al. Plant Cell. (2012) /tpc
 /tpc") Supplemental Data. Perea-Resa et al. Plant Cell. (22)..5/tpc.2.3697 Sm Sm2 Supplemental Figure. Sequence alignment of Arabidopsis LSM proteins. Alignment of the eleven Arabidopsis LSM proteins. Sm and
Supplemental Data. Perea-Resa et al. Plant Cell. (22)..5/tpc.2.3697 Sm Sm2 Supplemental Figure. Sequence alignment of Arabidopsis LSM proteins. Alignment of the eleven Arabidopsis LSM proteins. Sm and
QTL analysis for hybrid sterility and plant height in interspecific populations derived from a wild rice relative, Oryza longistaminata
 Breeding Science 59: 441 445 (2009) Note QTL analysis for hybrid sterility and plant height in interspecific populations derived from a wild rice relative, Oryza longistaminata Zhiwei Chen 1,2), Fengyi
Breeding Science 59: 441 445 (2009) Note QTL analysis for hybrid sterility and plant height in interspecific populations derived from a wild rice relative, Oryza longistaminata Zhiwei Chen 1,2), Fengyi
Duplication of an upstream silencer of FZP increases grain yield in rice
 SUPPLEMENTARY INFORMATION Articles https://doi.org/10.1038/s41477-017-0042-4 In the format provided by the authors and unedited. Duplication of an upstream silencer of FZP increases grain yield in rice
SUPPLEMENTARY INFORMATION Articles https://doi.org/10.1038/s41477-017-0042-4 In the format provided by the authors and unedited. Duplication of an upstream silencer of FZP increases grain yield in rice
Genomes and Their Evolution
 Chapter 21 Genomes and Their Evolution PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
Chapter 21 Genomes and Their Evolution PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
Advances in M olecular Genetics of T iller ing Characters in R ice
 2006, 18 (1) : 80 84 Acta Agriculturae J iangxi 1, 2 (1., 330200; 2., 225009) : ( ) : ; ; ; ; : S511. 032 : A : 1001-8581 (2006) 01-0080 - 05 Advances in M olecular Genetics of T iller ing Characters in
2006, 18 (1) : 80 84 Acta Agriculturae J iangxi 1, 2 (1., 330200; 2., 225009) : ( ) : ; ; ; ; : S511. 032 : A : 1001-8581 (2006) 01-0080 - 05 Advances in M olecular Genetics of T iller ing Characters in
National Agricultural Research Center for Tohoku Region, 3 Shimofurumichi, Yotsuya, Omagari, Akita , Japan 2)
") Breeding Science 52 : 275-283 (2002) Quantitative Trait Loci for Nonstructural Carbohydrate Accumulation in Leaf Sheaths and Culms of Rice (Oryza sativa L.) and their Effects on Grain Filling Kenji Nagata*
Breeding Science 52 : 275-283 (2002) Quantitative Trait Loci for Nonstructural Carbohydrate Accumulation in Leaf Sheaths and Culms of Rice (Oryza sativa L.) and their Effects on Grain Filling Kenji Nagata*
MAPPING QUANTITATIVE TRAIT LOCI (QTLS) FOR SALT TOLERANCE IN RICE (ORYZA SATIVA) USING RFLPS
 FOR SALT TOLERANCE IN RICE (ORYZA SATIVA) USING RFLPS") Pak. J. Bot., 36(4): 825-834, 4. MAPPING QUANTITATIVE TRAIT LOCI (QTLS) FOR SALT TOLERANCE IN RICE (ORYZA SATIVA) USING RFLPS M. SHAHID MASOOD, YANAGIHARA SEIJI *, ZABTA K. SHINWARI AND RASHID ANWAR Plant
Pak. J. Bot., 36(4): 825-834, 4. MAPPING QUANTITATIVE TRAIT LOCI (QTLS) FOR SALT TOLERANCE IN RICE (ORYZA SATIVA) USING RFLPS M. SHAHID MASOOD, YANAGIHARA SEIJI *, ZABTA K. SHINWARI AND RASHID ANWAR Plant
Quantitative trait loci mapping of the stigma exertion rate and spikelet number per panicle in rice (Oryza sativa L.)
") Quantitative trait loci mapping of the stigma exertion rate and spikelet number per panicle in rice (Oryza sativa L.) M.H. Rahman, P. Yu, Y.X. Zhang, L.P. Sun, W.X. Wu, X.H. Shen, X.D. Zhan, D.B. Chen,
Quantitative trait loci mapping of the stigma exertion rate and spikelet number per panicle in rice (Oryza sativa L.) M.H. Rahman, P. Yu, Y.X. Zhang, L.P. Sun, W.X. Wu, X.H. Shen, X.D. Zhan, D.B. Chen,
DNA Structure and Function
 DNA Structure and Function Nucleotide Structure 1. 5-C sugar RNA ribose DNA deoxyribose 2. Nitrogenous Base N attaches to 1 C of sugar Double or single ring Four Bases Adenine, Guanine, Thymine, Cytosine
DNA Structure and Function Nucleotide Structure 1. 5-C sugar RNA ribose DNA deoxyribose 2. Nitrogenous Base N attaches to 1 C of sugar Double or single ring Four Bases Adenine, Guanine, Thymine, Cytosine
Genetic analysis of maize kernel thickness by quantitative trait locus identification
 Genetic analysis of maize kernel thickness by quantitative trait locus identification S.S. Wen 1, G.Q. Wen 2, C.M. Liao 3 and X.H. Liu 2 1 Key Laboratory of Southwest Rice Biology and Genetic Breeding,
Genetic analysis of maize kernel thickness by quantitative trait locus identification S.S. Wen 1, G.Q. Wen 2, C.M. Liao 3 and X.H. Liu 2 1 Key Laboratory of Southwest Rice Biology and Genetic Breeding,
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL
 GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
. Supplementary Information
 . Supplementary Information Supplementary Figure S1. Mature embryo sac observations. Supplementary Figure S2. STT observations. Supplementary Figure S3. Comparison of the PTB1 cdna with that of the mutant.
. Supplementary Information Supplementary Figure S1. Mature embryo sac observations. Supplementary Figure S2. STT observations. Supplementary Figure S3. Comparison of the PTB1 cdna with that of the mutant.
Quantitative Genetics & Evolutionary Genetics
 Quantitative Genetics & Evolutionary Genetics (CHAPTER 24 & 26- Brooker Text) May 14, 2007 BIO 184 Dr. Tom Peavy Quantitative genetics (the study of traits that can be described numerically) is important
Quantitative Genetics & Evolutionary Genetics (CHAPTER 24 & 26- Brooker Text) May 14, 2007 BIO 184 Dr. Tom Peavy Quantitative genetics (the study of traits that can be described numerically) is important
Linear Regression (1/1/17)
") STA613/CBB540: Statistical methods in computational biology Linear Regression (1/1/17) Lecturer: Barbara Engelhardt Scribe: Ethan Hada 1. Linear regression 1.1. Linear regression basics. Linear regression
STA613/CBB540: Statistical methods in computational biology Linear Regression (1/1/17) Lecturer: Barbara Engelhardt Scribe: Ethan Hada 1. Linear regression 1.1. Linear regression basics. Linear regression
The Distribution of Japonica
 Journal of Integrative Plant Biology 29 The Distribution of Japonica Rice Cultivars in the Lower Region of the Yangtze River Valley is Determined by Its Photoperiod-sensitivity and Heading Date Genotypes
Journal of Integrative Plant Biology 29 The Distribution of Japonica Rice Cultivars in the Lower Region of the Yangtze River Valley is Determined by Its Photoperiod-sensitivity and Heading Date Genotypes
Small RNA in rice genome
 Vol. 45 No. 5 SCIENCE IN CHINA (Series C) October 2002 Small RNA in rice genome WANG Kai ( 1, ZHU Xiaopeng ( 2, ZHONG Lan ( 1,3 & CHEN Runsheng ( 1,2 1. Beijing Genomics Institute/Center of Genomics and
Vol. 45 No. 5 SCIENCE IN CHINA (Series C) October 2002 Small RNA in rice genome WANG Kai ( 1, ZHU Xiaopeng ( 2, ZHONG Lan ( 1,3 & CHEN Runsheng ( 1,2 1. Beijing Genomics Institute/Center of Genomics and
Useful Propagation Terms. Propagation The application of specific biological principles and concepts in the multiplication of plants.
 Useful Propagation Terms Propagation The application of specific biological principles and concepts in the multiplication of plants. Adventitious Typically describes new organs such as roots that develop
Useful Propagation Terms Propagation The application of specific biological principles and concepts in the multiplication of plants. Adventitious Typically describes new organs such as roots that develop
USDA-DOE Plant Feedstock Genomics for Bioenergy
 USDA-DOE Plant Feedstock Genomics for Bioenergy BERAC Thursday, June 7, 2012 Cathy Ronning, DOE-BER Ed Kaleikau, USDA-NIFA Plant Feedstock Genomics for Bioenergy Joint competitive grants program initiated
USDA-DOE Plant Feedstock Genomics for Bioenergy BERAC Thursday, June 7, 2012 Cathy Ronning, DOE-BER Ed Kaleikau, USDA-NIFA Plant Feedstock Genomics for Bioenergy Joint competitive grants program initiated
Table S1 List of primers used for genotyping and qrt-pcr.
 Table S1 List of primers used for genotyping and qrt-pcr. genotyping! allele! ligomer*! 5'-sequence-3'! rice! d10-2! F! TTGGCTTTGCCTCGTTTC!!! R! AGCCTCCACTTGTACTGTG! Arabidopsis! max2-3, max2-4! F! ACTCTCTCCGACCTCCCTGACG!!!
Table S1 List of primers used for genotyping and qrt-pcr. genotyping! allele! ligomer*! 5'-sequence-3'! rice! d10-2! F! TTGGCTTTGCCTCGTTTC!!! R! AGCCTCCACTTGTACTGTG! Arabidopsis! max2-3, max2-4! F! ACTCTCTCCGACCTCCCTGACG!!!
New insights into the history of rice domestication
 Review TRENDS in Genetics Vol.23 No.11 New insights into the history of rice domestication Michael J. Kovach, Megan T. Sweeney and Susan R. McCouch Cornell University, Department of Plant Breeding and
Review TRENDS in Genetics Vol.23 No.11 New insights into the history of rice domestication Michael J. Kovach, Megan T. Sweeney and Susan R. McCouch Cornell University, Department of Plant Breeding and
Model plants and their Role in genetic manipulation. Mitesh Shrestha
 Model plants and their Role in genetic manipulation Mitesh Shrestha Definition of Model Organism Specific species or organism Extensively studied in research laboratories Advance our understanding of Cellular
Model plants and their Role in genetic manipulation Mitesh Shrestha Definition of Model Organism Specific species or organism Extensively studied in research laboratories Advance our understanding of Cellular
Supplementary Figure 1. Phenotype of the HI strain.
 Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Ti plasmid derived plant vector systems: binary and co - integrative vectors transformation process; regeneration of the transformed lines
 Ti plasmid derived plant vector systems: binary and co - integrative vectors transformation process; regeneration of the transformed lines Mitesh Shrestha Constraints of Wild type Ti/Ri-plasmid Very large
Ti plasmid derived plant vector systems: binary and co - integrative vectors transformation process; regeneration of the transformed lines Mitesh Shrestha Constraints of Wild type Ti/Ri-plasmid Very large
The Plant Cell, November. 2017, American Society of Plant Biologists. All rights reserved
 The Genetics of Floral Development Teaching Guide Overview The development of flowers in angiosperm plants provided a critical evolutionary advantage, allowing more options for pollen dispersal and seed
The Genetics of Floral Development Teaching Guide Overview The development of flowers in angiosperm plants provided a critical evolutionary advantage, allowing more options for pollen dispersal and seed
Regulation of Phosphate Homeostasis by microrna in Plants
 Regulation of Phosphate Homeostasis by microrna in Plants Tzyy-Jen Chiou 1 *, Kyaw Aung 1,2, Shu-I Lin 1,3, Chia-Chune Wu 1, Su-Fen Chiang 1, and Chun-Lin Su 1 Abstract Upon phosphate (Pi) starvation,
Regulation of Phosphate Homeostasis by microrna in Plants Tzyy-Jen Chiou 1 *, Kyaw Aung 1,2, Shu-I Lin 1,3, Chia-Chune Wu 1, Su-Fen Chiang 1, and Chun-Lin Su 1 Abstract Upon phosphate (Pi) starvation,
Sporic life cycles involve 2 types of multicellular bodies:
 Chapter 3- Human Manipulation of Plants Sporic life cycles involve 2 types of multicellular bodies: -a diploid, spore-producing sporophyte -a haploid, gamete-producing gametophyte Sexual Reproduction in
Chapter 3- Human Manipulation of Plants Sporic life cycles involve 2 types of multicellular bodies: -a diploid, spore-producing sporophyte -a haploid, gamete-producing gametophyte Sexual Reproduction in
Plant Structure and Organization - 1
 Plant Structure and Organization - 1 In our first unit of Biology 203 we will focus on the structure and function of the higher plants, in particular the angiosperms, or flowering plants. We will look
Plant Structure and Organization - 1 In our first unit of Biology 203 we will focus on the structure and function of the higher plants, in particular the angiosperms, or flowering plants. We will look
Science 9 - Unit A Review Sheet
 Science 9 - Unit A Review Sheet Learning Outcomes Can you? describe the relative abundance of species on Earth and in different environments describe examples of variation among species and within species
Science 9 - Unit A Review Sheet Learning Outcomes Can you? describe the relative abundance of species on Earth and in different environments describe examples of variation among species and within species
MATHEMATICAL MODELS - Vol. III - Mathematical Modeling and the Human Genome - Hilary S. Booth MATHEMATICAL MODELING AND THE HUMAN GENOME
 MATHEMATICAL MODELING AND THE HUMAN GENOME Hilary S. Booth Australian National University, Australia Keywords: Human genome, DNA, bioinformatics, sequence analysis, evolution. Contents 1. Introduction:
MATHEMATICAL MODELING AND THE HUMAN GENOME Hilary S. Booth Australian National University, Australia Keywords: Human genome, DNA, bioinformatics, sequence analysis, evolution. Contents 1. Introduction:
Gene Action and Combining Ability in Rice (Oryza sativa L.) Involving Indica and Tropical Japonica Genotypes
 Involving Indica and Tropical Japonica Genotypes") International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 6 Number 7 (2017) pp. 8-16 Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2017.607.002
International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 6 Number 7 (2017) pp. 8-16 Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2017.607.002
GENETICS - CLUTCH CH.1 INTRODUCTION TO GENETICS.
 !! www.clutchprep.com CONCEPT: HISTORY OF GENETICS The earliest use of genetics was through of plants and animals (8000-1000 B.C.) Selective breeding (artificial selection) is the process of breeding organisms
!! www.clutchprep.com CONCEPT: HISTORY OF GENETICS The earliest use of genetics was through of plants and animals (8000-1000 B.C.) Selective breeding (artificial selection) is the process of breeding organisms
Drosophila melanogaster and D. simulans, two fruit fly species that are nearly
 Comparative Genomics: Human versus chimpanzee 1. Introduction The chimpanzee is the closest living relative to humans. The two species are nearly identical in DNA sequence (>98% identity), yet vastly different
Comparative Genomics: Human versus chimpanzee 1. Introduction The chimpanzee is the closest living relative to humans. The two species are nearly identical in DNA sequence (>98% identity), yet vastly different
Jie Chen 1), De-Run Huang 1), Lei Wang 1), Guang-Jie Liu 1,2) and Jie-Yun Zhuang* 1) Introduction
, De-Run Huang 1), Lei Wang 1), Guang-Jie Liu 1,2) and Jie-Yun Zhuang* 1) Introduction") Breeding Science 60: 153 159 (2010) Identification of quantitative trait loci for resistance to whitebacked planthopper, Sogatella furcifera, from an interspecific cross Oryza sativa O. rufipogon Jie Chen
Breeding Science 60: 153 159 (2010) Identification of quantitative trait loci for resistance to whitebacked planthopper, Sogatella furcifera, from an interspecific cross Oryza sativa O. rufipogon Jie Chen
2. Der Dissertation zugrunde liegende Publikationen und Manuskripte. 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera)
") 2. Der Dissertation zugrunde liegende Publikationen und Manuskripte 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera) M. Hasselmann 1, M. K. Fondrk², R. E. Page Jr.² und
2. Der Dissertation zugrunde liegende Publikationen und Manuskripte 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera) M. Hasselmann 1, M. K. Fondrk², R. E. Page Jr.² und
Quantitative trait locus analysis for ear height in maize based on a recombinant inbred line population
 Quantitative trait locus analysis for ear height in maize based on a recombinant inbred line population Z.Q. Li 4,5, H.M. Zhang 1,4, X.P. Wu 1,4, Y. Sun 3,4 and X.H. Liu 2 1 Maize Research Institute, Shanxi
Quantitative trait locus analysis for ear height in maize based on a recombinant inbred line population Z.Q. Li 4,5, H.M. Zhang 1,4, X.P. Wu 1,4, Y. Sun 3,4 and X.H. Liu 2 1 Maize Research Institute, Shanxi
Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated
 Relative conversion rates of different plant hormones to their glucosylated") Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated form by recombinant UGT74E2. The naturally occurring auxin
Figure 1. Identification of UGT74E2 as an IBA glycosyltransferase. (A) Relative conversion rates of different plant hormones to their glucosylated form by recombinant UGT74E2. The naturally occurring auxin
What are we learning from genome-wide association studies (GWAS) in rice?
 in rice?") What are we learning from genome-wide association studies (GWAS) in rice? Susan McCouch, Chih Wei Tung, Mark Wright, Adam Famoso, Randy Clark, Anthony Greenberg, Janelle Jung, Hyujung Kim, Josh Cobb, Moni
What are we learning from genome-wide association studies (GWAS) in rice? Susan McCouch, Chih Wei Tung, Mark Wright, Adam Famoso, Randy Clark, Anthony Greenberg, Janelle Jung, Hyujung Kim, Josh Cobb, Moni
Similar traits, different genes? Examining convergent evolution in related weedy rice populations
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Publications from USDA-ARS / UNL Faculty U.S. Department of Agriculture: Agricultural Research Service, Lincoln, Nebraska
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Publications from USDA-ARS / UNL Faculty U.S. Department of Agriculture: Agricultural Research Service, Lincoln, Nebraska
Origin and Dissemination of Cultivated Rice in the Eastern Asia
 Origin and Dissemination of Cultivated Rice in the Eastern Asia Yo-chiro Faculty of Agriculture, Shizuoka University, Japan ntroduction Rice (Oryza sativa L.) and associated behaviors for growing rice
Origin and Dissemination of Cultivated Rice in the Eastern Asia Yo-chiro Faculty of Agriculture, Shizuoka University, Japan ntroduction Rice (Oryza sativa L.) and associated behaviors for growing rice
PRINCIPLES OF MENDELIAN GENETICS APPLICABLE IN FORESTRY. by Erich Steiner 1/
 PRINCIPLES OF MENDELIAN GENETICS APPLICABLE IN FORESTRY by Erich Steiner 1/ It is well known that the variation exhibited by living things has two components, one hereditary, the other environmental. One
PRINCIPLES OF MENDELIAN GENETICS APPLICABLE IN FORESTRY by Erich Steiner 1/ It is well known that the variation exhibited by living things has two components, one hereditary, the other environmental. One
Plant Development. Chapter 31 Part 1
 Plant Development Chapter 31 Part 1 Impacts, Issues Foolish Seedlings, Gorgeous Grapes Gibberellin and other plant hormones control the growth and development of plants environmental cues influence hormone
Plant Development Chapter 31 Part 1 Impacts, Issues Foolish Seedlings, Gorgeous Grapes Gibberellin and other plant hormones control the growth and development of plants environmental cues influence hormone
Outline. Genome Evolution. Genome. Genome Architecture. Constraints on Genome Evolution. New Evolutionary Synthesis 11/8/16
 Genome Evolution Outline 1. What: Patterns of Genome Evolution Carol Eunmi Lee Evolution 410 University of Wisconsin 2. Why? Evolution of Genome Complexity and the interaction between Natural Selection
Genome Evolution Outline 1. What: Patterns of Genome Evolution Carol Eunmi Lee Evolution 410 University of Wisconsin 2. Why? Evolution of Genome Complexity and the interaction between Natural Selection
Regulatory Change in YABBY-like Transcription Factor Led to Evolution of Extreme Fruit Size during Tomato Domestication
 SUPPORTING ONLINE MATERIALS Regulatory Change in YABBY-like Transcription Factor Led to Evolution of Extreme Fruit Size during Tomato Domestication Bin Cong, Luz Barrero, & Steven Tanksley 1 SUPPORTING
SUPPORTING ONLINE MATERIALS Regulatory Change in YABBY-like Transcription Factor Led to Evolution of Extreme Fruit Size during Tomato Domestication Bin Cong, Luz Barrero, & Steven Tanksley 1 SUPPORTING
学位規則第 9 条第 2 項により要約公開 ; 許諾条件により本文は に公開
 Title An active transposon mping facilita flowering time mutant genes in rice Author(s) Xu, Quan Citation Kyoto University ( 京都大学 ) Issue Date 2014-03-24 URL http://dx.doi.org/10.14989/doctor.k Right 学位規則第
Title An active transposon mping facilita flowering time mutant genes in rice Author(s) Xu, Quan Citation Kyoto University ( 京都大学 ) Issue Date 2014-03-24 URL http://dx.doi.org/10.14989/doctor.k Right 学位規則第
Komala, N. T*, Gurumurthy, R and Surendra, P
 ORIGINAL RESEARCH ARTICLE OPEN ACCESS Morphological Characterization of Advanced Lines of Rice (Oryza sativa L.) Derived from Swarna x Ranbir Basmati at Seedling Stage Komala, N. T*, Gurumurthy, R and
ORIGINAL RESEARCH ARTICLE OPEN ACCESS Morphological Characterization of Advanced Lines of Rice (Oryza sativa L.) Derived from Swarna x Ranbir Basmati at Seedling Stage Komala, N. T*, Gurumurthy, R and
Phylogeny and systematics. Why are these disciplines important in evolutionary biology and how are they related to each other?
 Phylogeny and systematics Why are these disciplines important in evolutionary biology and how are they related to each other? Phylogeny and systematics Phylogeny: the evolutionary history of a species
Phylogeny and systematics Why are these disciplines important in evolutionary biology and how are they related to each other? Phylogeny and systematics Phylogeny: the evolutionary history of a species
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
Heterosis Expression of Hybrid Rice in Natural- and Short-Day Length Conditions
 Rice Science, 2015, 22(2): 81 88 Copyright 2015, China National Rice Research Institute Hosting by Elsevier B.V. All rights reserved DOI: 10.1016/S1672-6308(14)60286-9 Heterosis Expression of Hybrid Rice
Rice Science, 2015, 22(2): 81 88 Copyright 2015, China National Rice Research Institute Hosting by Elsevier B.V. All rights reserved DOI: 10.1016/S1672-6308(14)60286-9 Heterosis Expression of Hybrid Rice
TE content correlates positively with genome size
 TE content correlates positively with genome size Mb 3000 Genomic DNA 2500 2000 1500 1000 TE DNA Protein-coding DNA 500 0 Feschotte & Pritham 2006 Transposable elements. Variation in gene numbers cannot
TE content correlates positively with genome size Mb 3000 Genomic DNA 2500 2000 1500 1000 TE DNA Protein-coding DNA 500 0 Feschotte & Pritham 2006 Transposable elements. Variation in gene numbers cannot
J. MITCHELL MCGRATH, LESLIE G. HICKOK, and ERAN PICHERSKY
 P1. Syst. Evol. 189:203-210 (1994) --Plant Systematics and Evolution Springer-Verlag 1994 Printed in Austria Assessment of gene copy number in the homosporous ferns Ceratopteris thalictroides and C. richardii
P1. Syst. Evol. 189:203-210 (1994) --Plant Systematics and Evolution Springer-Verlag 1994 Printed in Austria Assessment of gene copy number in the homosporous ferns Ceratopteris thalictroides and C. richardii
Wheat Genetics and Molecular Genetics: Past and Future. Graham Moore
 Wheat Genetics and Molecular Genetics: Past and Future Graham Moore 1960s onwards Wheat traits genetically dissected Chromosome pairing and exchange (Ph1) Height (Rht) Vernalisation (Vrn1) Photoperiodism
Wheat Genetics and Molecular Genetics: Past and Future Graham Moore 1960s onwards Wheat traits genetically dissected Chromosome pairing and exchange (Ph1) Height (Rht) Vernalisation (Vrn1) Photoperiodism
Through the genetic bottleneck: O. rufipogon as a source of trait-enhancing alleles for O. sativa
 Euphytica (2007) 154:317 339 DOI 10.1007/s10681-006-9210-8 Through the genetic bottleneck: O. rufipogon as a source of trait-enhancing alleles for O. sativa Susan R. McCouch Æ Megan Sweeney Æ Jiming Li
Euphytica (2007) 154:317 339 DOI 10.1007/s10681-006-9210-8 Through the genetic bottleneck: O. rufipogon as a source of trait-enhancing alleles for O. sativa Susan R. McCouch Æ Megan Sweeney Æ Jiming Li
Effect of temperature on endogenous hormone levels and opposite phyllotaxy in maize leaf primordial
 Effect of temperature on endogenous hormone levels and opposite phyllotaxy in maize leaf primordial H. Ye, G.M. Han, Q. Ma, Y.Q. Tan, H.Y. Jiang, S.W. Zhu and B.J. Cheng School of Life Sciences, Anhui
Effect of temperature on endogenous hormone levels and opposite phyllotaxy in maize leaf primordial H. Ye, G.M. Han, Q. Ma, Y.Q. Tan, H.Y. Jiang, S.W. Zhu and B.J. Cheng School of Life Sciences, Anhui
BREEDING, GENETICS, AND PHYSIOLOGY. Phenotypic Analysis of the 2006 MY2 Mapping Population in Arkansas
 BREEDING, GENETICS, AND PHYSIOLOGY Phenotypic Analysis of the 2006 MY2 Mapping Population in Arkansas E.J. Boza, K.A.K. Moldenhauer, R.D. Cartwright, S. Linscombe, J.H. Oard, and M.M. Blocker ABSTRACT
BREEDING, GENETICS, AND PHYSIOLOGY Phenotypic Analysis of the 2006 MY2 Mapping Population in Arkansas E.J. Boza, K.A.K. Moldenhauer, R.D. Cartwright, S. Linscombe, J.H. Oard, and M.M. Blocker ABSTRACT
Curriculum vitae Xigang Liu
 Curriculum vitae Xigang Liu 1, EDUCATION: 09/1993-07/1997 B.S. Major: Biology. College of Life Sciences, Hebei Normal University Academic Degree Paper: RAPD analysis of Taigu genic male-sterile wheat and
Curriculum vitae Xigang Liu 1, EDUCATION: 09/1993-07/1997 B.S. Major: Biology. College of Life Sciences, Hebei Normal University Academic Degree Paper: RAPD analysis of Taigu genic male-sterile wheat and
Curriculum Links. AQA GCE Biology. AS level
 Curriculum Links AQA GCE Biology Unit 2 BIOL2 The variety of living organisms 3.2.1 Living organisms vary and this variation is influenced by genetic and environmental factors Causes of variation 3.2.2
Curriculum Links AQA GCE Biology Unit 2 BIOL2 The variety of living organisms 3.2.1 Living organisms vary and this variation is influenced by genetic and environmental factors Causes of variation 3.2.2
AP Biology Summer 2017
 Directions: Questions 1 and 2 are long free response questions that require about 22 minutes to answer and are worth 10 points each. Questions 3-6 are short free- response questions that require about
Directions: Questions 1 and 2 are long free response questions that require about 22 minutes to answer and are worth 10 points each. Questions 3-6 are short free- response questions that require about
Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed
 Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed using the Geneious software. Accession numbers of the
Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed using the Geneious software. Accession numbers of the
Characteristics and Genetic Segregation of a Rolled Leaf Mutant in Rice
 Kor. J. Breed. Sci. 43(4) : 181~185 (2011) Characteristics and Genetic Segregation of a Rolled Leaf Mutant in Rice Songyee Lee, Minseon Choi, Joohyun Lee, and Hee-Jong Koh* Department of Plant Science,
Kor. J. Breed. Sci. 43(4) : 181~185 (2011) Characteristics and Genetic Segregation of a Rolled Leaf Mutant in Rice Songyee Lee, Minseon Choi, Joohyun Lee, and Hee-Jong Koh* Department of Plant Science,
Ohio Tutorials are designed specifically for the Ohio Learning Standards to prepare students for the Ohio State Tests and end-ofcourse
 Tutorial Outline Ohio Tutorials are designed specifically for the Ohio Learning Standards to prepare students for the Ohio State Tests and end-ofcourse exams. Biology Tutorials offer targeted instruction,
Tutorial Outline Ohio Tutorials are designed specifically for the Ohio Learning Standards to prepare students for the Ohio State Tests and end-ofcourse exams. Biology Tutorials offer targeted instruction,
Q1) Explain how background selection and genetic hitchhiking could explain the positive correlation between genetic diversity and recombination rate.
 Explain how background selection and genetic hitchhiking could explain the positive correlation between genetic diversity and recombination rate.") OEB 242 Exam Practice Problems Answer Key Q1) Explain how background selection and genetic hitchhiking could explain the positive correlation between genetic diversity and recombination rate. First, recall
OEB 242 Exam Practice Problems Answer Key Q1) Explain how background selection and genetic hitchhiking could explain the positive correlation between genetic diversity and recombination rate. First, recall
Lecture WS Evolutionary Genetics Part I 1
 Quantitative genetics Quantitative genetics is the study of the inheritance of quantitative/continuous phenotypic traits, like human height and body size, grain colour in winter wheat or beak depth in
Quantitative genetics Quantitative genetics is the study of the inheritance of quantitative/continuous phenotypic traits, like human height and body size, grain colour in winter wheat or beak depth in
Combining Ability and Heterosis in Rice (Oryza sativa L.) Cultivars
 Cultivars") J. Agr. Sci. Tech. (2010) Vol. 12: 223-231 Combining Ability and Heterosis in Rice (Oryza sativa L.) Cultivars M. Rahimi 1, B. Rabiei 1*, H. Samizadeh 1, and A. Kafi Ghasemi 1 ABSTRACT Quantitative valuations
J. Agr. Sci. Tech. (2010) Vol. 12: 223-231 Combining Ability and Heterosis in Rice (Oryza sativa L.) Cultivars M. Rahimi 1, B. Rabiei 1*, H. Samizadeh 1, and A. Kafi Ghasemi 1 ABSTRACT Quantitative valuations
Genetic dissection of flag leaf morphology in wheat (Triticum aestivum L.) under diverse water regimes
 under diverse water regimes") Yang et al. BMC Genetics (2016) 17:94 DOI 10.1186/s12863-016-0399-9 RESEARCH ARTICLE Open Access Genetic dissection of flag leaf morphology in wheat (Triticum aestivum L.) under diverse water regimes Delong
Yang et al. BMC Genetics (2016) 17:94 DOI 10.1186/s12863-016-0399-9 RESEARCH ARTICLE Open Access Genetic dissection of flag leaf morphology in wheat (Triticum aestivum L.) under diverse water regimes Delong
Supplemental Data. Wang et al. (2014). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Lodging-Resistance Breeding of Platycodon grandiflorus Using Distant Hybridization
 American Journal of Plant Sciences, 2015, 6, 2844-2849 Published Online November 2015 in SciRes. http://www.scirp.org/journal/ajps http://dx.doi.org/10.4236/ajps.2015.618281 Lodging-Resistance Breeding
American Journal of Plant Sciences, 2015, 6, 2844-2849 Published Online November 2015 in SciRes. http://www.scirp.org/journal/ajps http://dx.doi.org/10.4236/ajps.2015.618281 Lodging-Resistance Breeding
Full file at CHAPTER 2 Genetics
 CHAPTER 2 Genetics MULTIPLE CHOICE 1. Chromosomes are a. small linear bodies. b. contained in cells. c. replicated during cell division. 2. A cross between true-breeding plants bearing yellow seeds produces
CHAPTER 2 Genetics MULTIPLE CHOICE 1. Chromosomes are a. small linear bodies. b. contained in cells. c. replicated during cell division. 2. A cross between true-breeding plants bearing yellow seeds produces
How to connect to CGIAR wheat (CIMMYT and ICARDA) CRP??- Public wheat breeding for developing world
 CRP??- Public wheat breeding for developing world") Wheat breeding only exploits 10% of the diversity available The public sector can t breed elite varieties-how to connect to private sector breeders?? How to connect to CGIAR wheat (CIMMYT and ICARDA) CRP??-
Wheat breeding only exploits 10% of the diversity available The public sector can t breed elite varieties-how to connect to private sector breeders?? How to connect to CGIAR wheat (CIMMYT and ICARDA) CRP??-
Genetic Divergence Studies for the Quantitative Traits of Paddy under Coastal Saline Ecosystem
 J. Indian Soc. Coastal Agric. Res. 34(): 50-54 (016) Genetic Divergence Studies for the Quantitative Traits of Paddy under Coastal Saline Ecosystem T. ANURADHA* Agricultural Research Station, Machilipatnam
J. Indian Soc. Coastal Agric. Res. 34(): 50-54 (016) Genetic Divergence Studies for the Quantitative Traits of Paddy under Coastal Saline Ecosystem T. ANURADHA* Agricultural Research Station, Machilipatnam
23-. Shoot and root development depend on ratio of IAA/CK
 Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Science Unit Learning Summary
 Learning Summary Inheritance, variation and evolution Content Sexual and asexual reproduction. Meiosis leads to non-identical cells being formed while mitosis leads to identical cells being formed. In
Learning Summary Inheritance, variation and evolution Content Sexual and asexual reproduction. Meiosis leads to non-identical cells being formed while mitosis leads to identical cells being formed. In
Improving radiation use efficiency in tropical rice
 Improving radiation use efficiency in tropical rice Erik Murchie Agricultural & Environmental Sciences This talk 1. Radiation use efficiency (RUE) in tropical rice 2. Photosynthesis and RUE in the field.
Improving radiation use efficiency in tropical rice Erik Murchie Agricultural & Environmental Sciences This talk 1. Radiation use efficiency (RUE) in tropical rice 2. Photosynthesis and RUE in the field.
Other funding Sources Agency Name: MSU Agricultural Experiment Station /Project GREEEN Amount requested or awarded: 30,000
 FINAL PROJECT REPORT Project Title: Functional genomics of flowering in apple PI: Herb Aldwinckle Co-PI(2): Steve VanNocker Organization: Cornell University Organization: Michigan State University Telephone/email:
FINAL PROJECT REPORT Project Title: Functional genomics of flowering in apple PI: Herb Aldwinckle Co-PI(2): Steve VanNocker Organization: Cornell University Organization: Michigan State University Telephone/email:
You are encouraged to answer/comment on other people s questions. Domestication conversion of plants or animals to domestic uses
 The final exam: Tuesday, May 8 at 4:05-6:05pm in Ruttan Hall B35. 75 multiple choice questions for 150 points 50 questions from Lecture 20 27 25 questions directly from the first two exams. Key for exam
The final exam: Tuesday, May 8 at 4:05-6:05pm in Ruttan Hall B35. 75 multiple choice questions for 150 points 50 questions from Lecture 20 27 25 questions directly from the first two exams. Key for exam