presumptiv e germ layers during Gastrulatio n and neurulation Somites
|
|
|
- Douglas Parsons
- 6 years ago
- Views:
Transcription
 Principles of Development. 2th ed. London: Oxford university press. Gilbert SF. (2003) Development Biology. 7th ed. Sunderland: Sinaure Associates Inc.")


1 Vertebrate embryos are similar at the phylotypic stage Patterning the Vertebrate Body Plan II: Mesoderm & Early Nervous System Wolpert L, Beddington R, Jessell T, Lawrence P, Meyerowitz E, Smith J. (2001) Principles of Development. 2th ed. London: Oxford university press. Gilbert SF. (2003) Development Biology. 7th ed. Sunderland: Sinaure Associates Inc. Phylotypic stage, the embryo has undergone gastrulation, and the main axial structure (somites, notochord and neural tube), are well develop and already show signs of regional organization both A/P and D/V axis.. Patterning turns mesoderm into repeated structures of skeleton and trunk muscles. Dorsal mesoderm is internalized during gastrulation to eventually become the notochord and the somites. Notochord is transient and becomes part of spinal column. Fig. 4.1 Rearrange ment of the presumptiv e germ layers during Gastrulatio n and neurulation After gastrulation mesoderm internalized rigid rod notochord neural tube spinal, brain side neural crest skeletal, head.. Somites With neurulation, ectoderm overlaying the notochord forms the neural tube to give rise to brain and spinal cord. Somites, mesoderm in blocks flanking notochord, become 1) the vertebrate and ribs 2) muscles of the trunk and limbs and 3) contribute to the dermis. Somite are formed along the antero-posterior axis In the chick, mesoderm forms anterior to the regressing node of the primitive streak (Hensen s node). Pre-somitic mesoderm is the region between the last formed somite and the regressing node. This region will become 4 or 5 somites which h form simultaneously l as pairs on either side of the notochord. No signal specifying A/P position or timing is involved. Direction was A P, (Fig. 4.2) The position of somites were related with early stage of gastrulation Transplant and reverse the pre-somitic mesoderm Reversing the rostral-caudal order of pre-somitic mesoderm does not influence of temporal order of somiteogenesis. Still develop somite 1


 original position (depend on fate). (Fig 4.")

2 The segmental plate mesoderm is determined by its position along the anterior-posterior axis before somitogenesis Somites Form Progressively Side of the anterior primitive streak cell epiblast move, in gastrulation form somitogenic stem cell around Hensen s node leave node and form pre-somitic mesoderm (Fig.4.3) pre-somitic mesoderm The position of somites along A/P axis determines fate. But the related position no changethe mechanisms of somites formation??? May be Notch-Delta signaling pathway or FGF-8 Boundary development somites are formed along the antero-posterior axis Neural tube and somites seen by SEM (scanning electron microscopy) Somite form was A/P dependently and begin during gastrulation. Anterior somites form cervical vertebrate. Posterior ones form ribbed thoracic vertebrate. Somite form was according to its (pre-somitic mesoderm) original position (depend on fate). (Fig 4.5) Develop in a temporal and spatial order. Rearranging pre-somitic mesoderm will not change timing. The pattern is laid down earlier by an A/P axis signal. When surface ectoderm is peeled away Neural tube somitomere Boundary development The Positional Identity of Pre- Somitic Mesoderm Is Determined Paraxial mesoderm rounding somitomere compacted and bound separate from presomitic paraxial mesoderm somites Paraxial mesoderm Will development to thoracic somites 2
3 Somite development is specific gene expression dependent Gastrulation and neurulation in the chick embryo Gastrulation Notochrod neurulation somatic The major lineages of the mesoderm Molecular Mechanisms of Somite Formation The mechanisms are complex. The current view is the clock and wavefront model. splanchnic This model invokes: Cyclical patterns of gene expression A morphogen gradient Scler Myo tome Circulatory body cavity system Cartilage skeletal dermis The important components of somitogenesis are, (1) Periodicity, (2) fissure formation, (3)Epithelialization, (4) specification, and (5) differentiation 3
 expression in somitic")

4 Cyclical Waves of Gene Expression Fig. 4.4 Notch-Delta signaling pathway Somite formation correlates with the wavelike expression of the hairy1 gene in the chick time Cyclical waves of c-hairy (a transcription factor) expression in somitic mesoderm. Oscillations in Notch and Wnt gene expression are at the core of the clock. Pre- Somitic Mesoderm node Expressi on of hairy, Notch, Delta all confined to posterior of each somite Mutation of Notch-Delta, no somites formation The formation of somites was timing and position specifiy Notch signaling involved in somite fissioning A Moving Morphogen Gradient In the Clock and Wavefront Model the node Notch somite boundary formation The node is a source of Fgf 8 and Wnt, morphogens for somite formation. From Aulehla and Hermann (2004) Genes & Development 18:
 Science 301: 328.")
5 Integration of Clock and Wavefront Possible scheme for the regulation of the clock through which an Fgf8 gradient regulates a Wnt oscillating clock, which in turn controls a Notch clock High Axin inhibit Wnt3a Notch did not activate Hairyl x Low Axin did not inhibit Wnt3a Notch activate Hairly expression Notch-related gene From Poriquie (2003) Science 301: 328. Blue FGF8 LOW did not expressed hairly FGF8 high + notch interaction boundary formation white region formation Somitogenesis Regulating Notch activity at the somite boundaries Hairy expression pattern reflects notch activity Multiple regulatory interactions to establish and control Notch in somite Mesoderm becomes notochord and somites The fate of somite cells depends upon adjacent tissue signals In chick-quail trans-species grafts (with distinctive nuclei), somite fate maps have been constructed. (Fig. 4.6) quail chick Gastrulation and neurulation in the chick embryo 5
 and the dermatome (an epithelial sheet that forms dermis).")
 and migrate to surround the notochord and form the vertebrate.")



6 The major lineages of the mesoderm Mesoderm becomes notochord and somites The dorsal and lateral region of somites form the dermomyotome (which expresses Pax3). This is made up of the myotome (forms muscle cells) and the dermatome (an epithelial sheet that forms dermis). Cells from the medial region of the somite (which express MyoD) form axial & back muscles. Lateral region of somites form abdominal and limb muscles. Ventral medial region of the somite contain sclerotome cells (future cartilage which express Pax1) and migrate to surround the notochord and form the vertebrate. Circulatory body cavity system Fig47 Fig.4.7 Scler Myo tome Cartilage skeletal dermis The fate of different somite Dorsal region dermomytome myotome muscle cell epithelial cell dermis Medial region axial and back muscle Ventral region sclerotome migrate g to notochord develop p into vertebrate and ribs cartilage The somites need notochord and neural tube signal regulated No neural tube and notochrod, somites apoptosis Notochord induces sclerotome cells (develops into cartilage) and suppresses the formation dermomyotome. Fig. 4.8 Notochrod/neural tube are induce of cartilage. Later plate mesoderm regulated the lateral part of the dermomyotome. Fig. 4.9 Mesoderm becomes notochord and somites The patterning of somites differentiation From notochord transplantation experiments, an additional notochord induces unsegmented pre-somitic mesoderm to produce greatly increased amount of cartilage. Neural tube (ventral side: the floor plate) induces cartilage. sclerotome Lateral plate mesoderm and the ectoderm induce the dermomyotome. Signals that may pattern the somites are secreted signaling proteins that t may include: 1) Sonic hedgehog which may specify the ventral somites. 2) BMP-4 which may specify the lateral somites. 3) Wnt family proteins which may specify the dorsal somites. 6







7 Many signal modulated the somites differentation The major lineages of the mesoderm From dorsal neural tube, etcoderm,lateral From lateral plate mesoderm From Notochord, floor plate of the neural tube Sonic Hedgehog BMP4 Wnt Fig.4.9 A model for patterning of somite differentiation. Notochord and the floor plate of the neural tube sonic hedgehog protein (diffusible signal) somites sclerotome Dorsal neural tube and ectoderm signal somite dermomytome Lateral plate mesoderm Scler Myo tome Circulatory body cavity system Cartilage skeletal dermis signal signal Somites different cell formation migration specification Back, muscle Back muscle signal Cartilage and rib dermis muscle Cartilage Tend and ligament Diagram of a transverse section through the trunk of a chick embryo 7

 mutants lack limb muscles.")
 genes (invertebrate or vertebrate) Patterning along the A/P axis in all vertebrate involves the expression of a set of genes (HOX) that specify positional identity along the axis.")

 develop by another gene; It means mutation causes transformation of one body part into another body part.")
8 Mesoderm becomes notochord and somites Regulation of the Pax homeobox genes (transcription factors) Pax genes are regulated by signals from the notochord and neural tube to control the somitic cell fate. Notochord/neural signal pre-somitic cell regulate Pax gene expression Pax3 is expressed early in all cells that will form somites. Pax3 is modulated by BMP-4 and Wnt to confine it to muscle precursors. Pax3 is further down-regulated in back muscle precursors but remains active in future limb muscle cells. In mice, Splotch (Pax3-minus) mutants lack limb muscles. Defined by the presence of a conserved paired-box that codes for a 128-amino-acid paired domain, a DNA binding domain 9 Pax genes: Pax-1 to -9 Positional somites along the A/P axis specified by homeobox (HOX) genes (invertebrate or vertebrate) Patterning along the A/P axis in all vertebrate involves the expression of a set of genes (HOX) that specify positional identity along the axis. Homeobox genes: (Box 4A) Homeobox about 180 base pair (DNA) 60 amino acids (homeodomain) transcription factor (DNA binding protein) bind to DNA (helix-turnhelix DNA binding motif) turn on gene Homebox gene encode a large family of transcription factors. The transcription factor also called homeodomain proteins. Share a similar 60 amino acid DNA binding homeodomain which is encoded by 180 basepair homeobox sequence. Homeotic transformation: One mutation replace by adjacent another gene (structure) develop by another gene; It means mutation causes transformation of one body part into another body part. Homeodoamin protein activate batteries of gene specify the particular properties of each segment There are four separate clusters of Hox genes (subset of the homeobox genes) in all vertebrates. Hox genes start to be start to be expressed in mesoderm cells at an early stage of gastrulation when they begin to leave the primitive streak. Hox genes: anterior genes are expressed first. The posterior pattern develops later; Clearly defined patterns of Hox gene expression are most easily seen in the mesoderm and neural tube, after somites formation and neurulation. Box 4A Homeobox genes Paralogy groups helix-turn-helix DNA binding motif Orthology groups 8
 gene in the four different hox complexes - genes with maximum sequence identity orthology same gene in a different species Homeobox is a highly-conserved, 180 bp DNA sequence only")
9 Paralogy group -are thehox complexes - natural clusters of Hox genes on a chromosome - order relates to the duplication that occurred within a chromosomal region Orthology group - appropriate (pseudo-orthologs) gene in the four different hox complexes - genes with maximum sequence identity orthology same gene in a different species Homeobox is a highly-conserved, 180 bp DNA sequence only DNA has a homeobox homeodomain is the protein product of the homeobox and proteins encoded by homeobox genes are homeodomain proteins homeotic genes act tin a cell autonomous manner - an essential gene encoding a body part or segment - a gene whose mutant allele causes replacement of a certain body part with a body part normally found elsewhere - best described by homeotic mutants (transformation) - the mutation causes transformation of one body part into another body part only one mutant affect phenotype all homeotic genes are homeobox genes; but not all homeobox genes are homeotic genes The homeotic genes in the HOM (Drosophila) and HOX (vertebrate) complexes are all homeodomain transcription factors Hox gene clusters Hox genes ( Hox gene clusters) are a subset of the homeobox genes encoding of transcription factors. homeodomain Might have arisen by rounds of duplication of ancestral genes (to form a cluster), followed by further duplication of that cluster in mammals. Paralogous group are composed of the most similar members of each cluster, in the same chromosome. Mouse paralogous subgroup=13 Partially overlapping zones of expression which vary in the anterior extent of their expression define distinct regions. Various genes respond to the combination of gene products expressed. Homeotic gene are involved in specifying regional identity along the A/P axis. How Hox gene can specify the identity of axis? (Fig.4.10) Gene activity can provide positional values Anterior posterior Tissue W: only need gene1 Z: need gene 1-4 expression Different combine Hox gene expression The pattern of expression defines four distinct regions, coded for by the expression of different combinations of genes. Different gene expression produced different interaction specification of A/P. 9

 mesoderm (after somite formation) and 2) neural tube (neurulation). Fig. 4.")

10 Hox genes pattern the A/P axis The differences between vertebrae (i.e. anterior -attach to skull; cervical; thoracic have ribs; lumbar, sacral and caudal ) clearly demonstrate that identity of somites differ along the A/P axis. Hox genes are expressed along the A/P axis in mouse. First, anterior Hox genes expressed in early gastrulation as mesoderm begins to leave the primitive streak. The anterior gene are expressed first. As the posterior pattern develop latter. More posterior Hox genes turn on as development continues. Defined patterns of Hox gene expression are seen in: 1) mesoderm (after somite formation) and 2) neural tube (neurulation). Fig For different antibody to stain The arrowheads indicated the anterior boundary of expression of each gene within the neural tube Hox genes pattern the somites Hox genes are expressed as gastrulation proceeds. Hox genes provide a code for regional identity. Hox genes show a sharp anterior border of expression and a much less defined posterior border. Lot of overlap by every region but each has distinct pattern of Hox gene expression. The pattern unfolds (Fig. 4.12) (open) over time with Most anterior (hindbrain) express Hoxa1 different genes and Hoxb1 only. expressed at different Posterior regions express all Hox genes (over lapping). The anterior head, forebrain and midbrain do not express Hox genes but have other homeobox genes (etx & otx.) times. The pattern of expression is quite different in anterior and posterior regions of the body axis The same chromosome, Hox gene different : spatial, temporal expressed A-P axis Hox gene expression is co-linear Hoxa1 has its most anterior expression in the posterior head. Hoxa11 has its most anterior expression in the sacral (lower back) region. Hox gene expression is co-linear (from different chromosome) as order of genes on the chromosome (per cluster) reflects the order of spatial and temporal expression along the A/P axis. Hox gene are involved in controlling regional identity. (Fig.4.13) Combination of Hox gene provide positional identity. Hox gene expression is conserved between mouse and chick. Different Hox gene regulated specific tissue (region) d12 and d13 may switched Technique: Insertional mutagenesis (gene knock-out) A targeting vector is constructed that has the central (functional) region of a gene replaced with a drug resistance gene. This is transfected into ES cells and selected by drug exposure. By homologous recombination, a fraction of the transformants will have one copy of the original gene replaced with the altered (non-functional) form. These cells are injected into the inner cell mass of a blastocyst. Resultant chimeric mice give rise to heterozygous mutants which can be bred to generate mutant homozygotes. Different Hox gene, control the same somites 10
 gene knock-out (non-specific) gene knock-out (tissue-specific) EC cell; Gene transfection insert randomly by homologous recombination")
 to target gene transgenic mice Cre/loxP System crossed with transgenic mice (has recombinase Cre) offspring Cre is a DNA")



11 Gene Physiological function: 1. transgenic mice 2. gene knock out Transgenic: 1. Injection DNA (interesting gene) to the fertilized eggs 2. Alter or add a gene to the genome of embryonic cell (EC cell) gene knock-out (non-specific) gene knock-out (tissue-specific) EC cell; Gene transfection insert randomly by homologous recombination mutant cell (inactive gene; gene knock out). It radonly, insert to blastocyst, usually altered the gene function but not knock gene. Why use EC cell: 1. less umber cell effect all body 2. can produced germ cells EC cell micro-injection Both side insert loxp (34 bp) to target gene transgenic mice Cre/loxP System crossed with transgenic mice (has recombinase Cre) offspring Cre is a DNA recombinase Cre expressed cut loxp all loxp sites are 34 bp knock-out Cre recognizes loxp sites With tissue specific gene offspring tissue specific expression of Cre cut specific loxp tissue specific knock out Cre tissue specific promoter The lox-cre recombination system can knock out genes Altering Hox gene expression alters axial patterning loxp inserted on each side of target gene produced a chimera mice; Chimera mice mating with another chimera mice (has recombinase Cre) produced lop X plus Cre chimera mice loxp + cre specific knock out loxp + normal nothing happen No loxp nothing happen With cell typespecific promoter: produce tissue specific knout out In mice, gene knock-out experiments produce mutants. There is redundancy, where a missing gene can be at least partially compensated for the expression of related genes. Paralogous genes from another Hox complex may compensate for gene loss. Posterior prevalence : mutation affects the anterior extent of gene expression. Homeotic transformations result from Hox gene loss. For replace by another Hox gene expression different and specific tissue Loss leads to cells assuming a more anterior value i.e. Hoxc8 mutant mice have extra ribs. Loss one Hox gene more anterior tissue expressed Abnormal expression of Hox genes in anterior regions lead to tissues becoming more like posterior positioned tissues. 11
 Retinoic acid can alter positional value Loss of Hox gene, the conversion of one body part into")


 RA cross membrane bind receptor complex transcription factor turn on gene Retinoic")


12 Homeotic transformation (Fig.4.14) Retinoic acid can alter positional value Loss of Hox gene, the conversion of one body part into another. Its absence gives them a more anterior positional value, and they develop accordingly Retinoic acid is a derivative of vitamin A. It has very important role in signaling vertebrate development. In early development, retinoic i acid can cause homeotic transformation of the vertebrate. It can diffuse across plasma membranes to bind protein receptors and form an active transcription factor. (steroid molecule) RA cross membrane bind receptor complex transcription factor turn on gene Retinoic acid interferes with the normal expression of Hox genes. Later, it can alter positional development in limb development. Neural induction and the role of organizer Organizer vs. variety animal Neural induction: Form ectoderm During gastrulation ectoderm lying along the dorsal midline form neural plate neural tube brain/spinal cord Nervous system must be linked to the mesoderm patterning Organizer-primary embryonic induction Amphibians: spemann organizer Zebrafish: shield bird/chick: Hensen s node Mouse: equivalent node region Mouse: equivalent node region Amphibians: spemann organizer Zebrafish: shield bird/chick: Hensen s node 12




")


13 The development of mouse embryo Primary neurulation: neural tube formation in the chick embryo MHP: medial neural hinge point Primary neurulation: neural tube formation in the chick embryo Neural crest cells Neural crest cells migrate away from the neural tube to develop into... 1) skull (bone) 2) sensory and autonomic nervous systems 3) pigment cells Somites are formed after gastrulation along the antero-posterior axis. Primary neurulation DLHP: dorsolateral hinge point 13

 pigment cells Somites are formed after")


14 Neural crest cells Neural crest cells migrate away from the neural tube to develop into... 1) skull (bone) 2) sensory and autonomic nervous systems 3) pigment cells Somites are formed after gastrulation along the antero-posterior t axis. N C muscle Heart, kidney Gaunard, gut muscle Blood froming tissue Dorsal-Ventral Specification of the Neural Tube Cascade of Inductions Initiated by the Notochord in the Ventral Neural Tube Shh: Sonic Hedgehog 14

.")
 Mid-gastrula induces trunk and tail (no head). Late gastrula induces only tail.")

, at the anterior of the primitive streak contributes t to notochord and somites and can induce another axis. Fig.4.")
15 The role of spemann organizer: axis development (A/P; D/V) Spemann Organizer is temporally dependent Fig Transplantation of organizer in different gastrulation stage Spemann organizer as primary embryonic induction. Spemann Organizer (dorsal blastopore lip) grafted to the ventral side of the marginal zone results in a twinned embryo. Fig.4.15 The second embryo can have head, trunk and, sometimes, a tail but will be joined to primary embryo along the axis. Blastopore transplants vary with time (timing dependent). In different gastrulation stage, transplantation of organizer Early gastrula induces a full 2nd embryo. (completed body axis and CNS,but only older organizer induce-posterior structure) Mid-gastrula induces trunk and tail (no head). Late gastrula induces only tail. Different stage of embryo, spemann organizer plays different induction of axis. May different gene expression at different stage in organizer. Early gastrulation: A/P Late gastrulation: D/V tail The Spemann Organizer has conserved functions As gastrulation proceeds, the A/P axis become specified and as it progresses the blastopore lip can only induce more posterior structures. Hensen's node (the chick Organizer), at the anterior of the primitive streak contributes t to notochord and somites and can induce another axis. Fig.4.16 Organizer can induce many region (Fig.4.17) is timing dependent Early stage: head Mid stage: no head, other yes Late stage: tail In early gastrulation stage, the inducting region as determined form the results of transplantation t ti of small pieces of tissue form the dorsal marginal zone. Fig 4.16 Hensen s node can induce a new in bird embryos Xenopus and mouse share a number of genes that are expressed in the organizer: 1) Brachyury 2) FGF (fibroblast growth factor) Control of Hox genes is unknown Fig Genes expressed in the spemann organizer. 15


, it also positional specification.")
 Organizer can secret many neural")
 BMP-4, secreted growth factor, inhibits cells from forming neural tissue.")
 inhibits BMP-4 and acts to dorsalize the mesoderm.")
16 Neural plate is induced in the ectoderm (by mesoderm; positional specification) Dorsal lip transplantation experiments demonstrate that a nervous system can be induced from ectoderm.(fig.4.19) Original development The nervous system of Xenopus is induced during gastrulation. Specific position induced to neural system; not dependent on cell fate Different region develop Ventral tissue can be different structure induced neural tissue The formation of the nervous system is dependent on an inductive signal (is induced ), it also positional specification. The specific gene turn on (specific time and position) induced neural plate Neural plate is induced in the ectoderm by secreted proteins (specific region secreted specific protein to ectoderm formed neural plate) Organizer can secret many neural induction protein. Chordin, noggin, cerberus are BMP-4 antagonists. (BMP- 4 mesoderm) BMP-4, secreted growth factor, inhibits cells from forming neural tissue. Inhibition of BMP-4 allows neural tissue formation. noggin (secreted by the organizer) inhibits BMP-4 and acts to dorsalize the mesoderm. noggin also induces neural tissue (neural induction factor). BMP is ventral induction. chordin expressed by the future neural plate cells of the organizer. It can bind BMP-4 and preventing its action. Both noggin and chordin directly bind BMP-4 and inactive it to allow the induction of neural tissue. Knock-out noggin and chordin in zebrafish, no neural plate developing Others proteins: FGF (induce), Wnt (block) 16
")

 from mesoderm (positional specification) Nervous system can be patterned by signals from the mesoderm.")
 when transplanted beneath the ectoderm of a gastrula.")

17 - Cortical cytoplasmic rotation causes translocation of the dishevel protein to the dorsal side of the embryo - Dishevel inhibits GSK-3 (glycogen synthase kinase 3) -catenin is not degraded in blastomeres of the future dorsal side - -catenin enters the nuclei and binds with tcf3 to form a transcription factor - b-catenin/tcf3 complex activates genes such as siamois (drosalize cause factor) - siamois along with factors stimulated by the VegT and Vg1 (nodal) cumulate to activated goosecoid+ gene in the cell of the blastopore lip - goosecoid+ turns on the expression of genes that code for dorsalizing molecules, like noggin and chordin - the organizer is now functional, directly dorsalization The hypothesis of induction of the organizer in the dorsal mesoderm Cortical rotation translocation of Dsh move to dorsal region Dsh + GSK-3 -catenin accumulate catenin enter to nuclear and bind to Tcf-3 protein formation a complex transcription factor activate gene encoding siamois protein Dorsal formation Neural plate is induced by signaling mechanism(s) from mesoderm (positional specification) Nervous system can be patterned by signals from the mesoderm. It is positional specificity in this induction. Fig.4.20 In newt neurula, mesoderm transplantation into younger newt embryos, anterior explants induce head and brain. Posterior explants induce trunk & spinal cord. Neural plate explants induce specific neural structures (depending upon position) when transplanted beneath the ectoderm of a gastrula. Induction of the nervous system by the mesoderm is position specific Different region of mesoderm induced different neural tissue Neural tissue formation, depend on the development of axis. Hox gene expression regulated the formation of somites, but it can not detected in the anterior-most neural tissue. Hox gene expression vs. somites development Different HOX gene different somite induced different neural tissue Spinal cord Brain Hox gene regulated Otx, Emc regulated Form hindbrain Neural crest cell Gastrulation (HOX gene turn) somite neural tissue 17


18 How the mesoderm signal induced neural patting (Model: The two signal model of neural patterning) Signal 1 from the mesoderm induces ectoderm to become anterior neural tissue. Directly turn on the formation of neural tissue. (chordin and noggin are good candidates) Signal 2 turns part of this into posterior neural tissue in a graded manner Regulated the neural patterning (FGF, Wnts & retinoic i acid are candidates). Signal 2 Signal 1 Fig Models of neural patterning by induction They are related with Hox gene expression Head organizer: BMP Inhibitors Cordin &Noggin, Wnt inhibitors mesoderm Cerberus, Dickkopf & frzb1 to "anteriorize " neural tube The two signal model of neural patterning (mixing) Different part and graded signal mesoderm Tail organizer: FGF, WNT, RA &BMP inhibitors Are posteriorizing signaling molecules Not only mesoderm but also Hensen s node can induced gene expression characteristic of neural tissue Mouse and chick grafts of the primitive streak (ie node or Hensen's node) can also induce neural tissue. This model differs from another model which suggests that there may exist a number of region-specific inducer molecules Hensen's s node (chick) can induce neural gene expression in Xenopus ectoderm. This (Fig.4.22) demonstrates the evolutionary conservation of neural induction signals and confirm similarity of Hensen's node and Spemann Organizer. Specific region may has specific induced-signal. Early nodes induce anterior neuron structures. Later nodes induce posterior neuron structures. The nodes can specify different A/P positional values over time. The capacity to produce signals that generate anterior structure is lost. Neural induction is time, position, mesoderm and Hensen s dependent Fig Hensen s node can induced neuron specific marker in Xenopus ectoderm Neurogenic factor-3 Neural cell adhesion molecular Common transcription factor Only Hensen s node can induced neural specific gene factor (NF-3) in ectoderm. It may be some specific induced-signal of specific region, such as Hensen s node. neuronal specific marker induced by specific region 18


 Posterior head and hindbrain development requires")

 Rhombomeres: the region do not cross form one side of a boundary to the other.")
19 Neural plate signals travel within the neural plate (did not cross mesoderm) The signal (planar: route from the mesoderm to the overlying ectoderm, whereas the other is) being generated within the neural plate itself and traveling within the ectodermal sheet Fig.4.23 Induction and patterning of the nervous system involves planar signal originating with in the ectoderm Specific gene (engrailed-2 and_krox-20) are only expressed in the neural tissue Did not across N-CAM, a neural cell-cell adhesion protein, neurogenic factors and other neural specific proteins can be expressed in the ectoderm in the correct A/P order in exogastrula which suggests that the inducing signal can travel a relatively long distance through the tissues and did not across the mesoderm. Hindbrain rhombomeres restrict cell lineage (The hindbrain is segmented into rhombomeres by boundaries of cell-lineage restriction) Posterior head and hindbrain development requires segmentation of the anterior neural tube, it along A/P axis. It did not occur elsewhere along the spinal cord. Other spinal region, the pattern of dorsal root ganglia and ventral motor nerves is imposed by the somites. Segmentation events in the 3 day chick embryo's posterior head include... Fig ) somite formation from mesoderm on either side of notochord, 2) the hindbrain (rhombocephalon) is divided into 8 rhombomeres, and 3) the lateral mesoderm forms the branchial arches. (Note: spinal cord is segmented into dorsal root ganglia and ventral motor nerves by the somites) Rhombomeres: the region do not cross form one side of a boundary to the other. It formed by cell-lineage restriction. Fig Rhombomeres share some adhesive property that prevent mixing with those of adjacent cell. Fig Division of the hindbrain into rhombomeres: a unique identity and develop 19





.")
20 Early human brain development Regional specification in the developing brain Fig.4.24 Fig Lineage restriction in rhombomere Pharyngeal arch Rhombomeres formation different development Why? For different function development Branchial In early stage of neurulation Can spread In late stage of neurulation Nerve cross boundaries Cells in each rhombomere may be under the control of the same genes, and that the rhombomere is a developmental unit. Fig Signaling by ephrins and their receptor It self generated signal Via receptor activation generated signal Ephrin and its receptor separately express, thus preventing cell mixing at the rhombomere. It mean that cells in each rhombomere may be under the control of the same genes, and that the rhombomere is a development unit. Development of posterior head involves interactions Neural crest cells innervate the face and neck to form the segmental cranial nerves. Neural tube forms segmentally arranged the segmental cranial nerves. Neural crest cells also give rise to peripheral nerves and bones including jaw (from the first branchial arch) and the bony parts of the ear (from the second arch). Eight rhombomeres form by constricting the freshly closed neural tube into eight evenly spaced sections. Lineage restriction occurs with cells and their descendants remaining in their parent rhombomere. Cell movement restriction depends on adhesive properties. Within each rhombomere, the cells are under the control of the same genes and act as a developmental unit. 20

by rhombomere 6 cells. Neural crest cells from rhombomeres 3 and 5 die by apoptosis (programmed cell death).")

21 Neural crest cells have positional values from different rhombomeres different gene expression produced different neural crest cell different tissue development Chick neural crest cells can be labeled and their fate mapped. The cranial neural crest cells migrate out from the rhombomeres of the dorsal region of the hindbrain. Branchial arch 1 (formed jawbones, ear bone face bones)is populated by cells from rhombomere 2, Branchial arch 2 (formed neck cargilage, middle ear bone) by rhombomere 4 cells and Branchial arch 3 ( thymus, parathyroid, thyroid gland)by rhombomere 6 cells. Neural crest cells from rhombomeres 3 and 5 die by apoptosis (programmed cell death). Transplantation of rhombomere 2 cells to where rhombomere 4 cells should be results in formation of a second jaw. Lineage restriction Rhombomeres developed independently, produces different neural crest. Different neural crest can develop to different tissue Pharyngeal Hindbrain Hox gene expression Mouse embryonic hindbrain Hox gene expression is well defined and correlates well with the segmental pattern. Paralogous group 1 (Hoxa1, b1 etc.) is expressed anterior to paralogou 2 (Hoxa2, b2 etc.) followed by paralog 3. Fig Control of gene expression is complex (i.e. Hoxb2 is controlled in rhombomere 3 and 5 by one enhancer which is activated by Krox-20 and in r4 by another enhancer). Hoxa2 knock-out mice show head skeletal defects that are consistent with a homeotic transformation of the inner ear (2nd branchial arch) to jaw (1st arch). (By neurula stage, the embryo is divided into regulating organforming regions.) Fig Expression of Hox genes in the branchial region of the head. Fig 4.28 Gene expression in the hindbrain. 21




22 The embryo is patterned by the neural stage into organ-forming regions At gastrulation state, body plane had established. At the neurula stage, the regions of embryo have become determined, however it can regulated. Developmental Biology Patterning the Vertebrate Body Plan II: Mesoderm & Early Nervous System Vertebrate embryos are similar at the phylotypic stage Somites, notochord and neural tube show A/P organization. Patterning turns mesoderm into repeated structures of skeleton and trunk muscles. Dorsal mesoderm is internalized during gastrulation to eventually become the notochord and the somites. Notochord is transient and becomes part of spinal column. With neuralation, ectoderm overlaying the notochord forms the neural tube to give rise to brain and spinal cord. Somites, mesoderm in blocks flanking notochord, become 1) the vertebrate and ribs 2) muscles of the trunk and limbs and 3) contribute to the dermis. 22
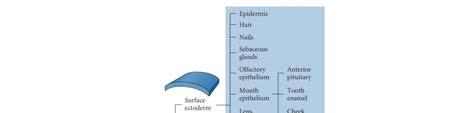



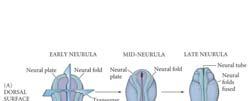
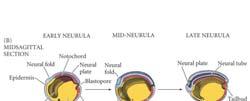
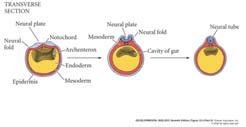
23 Primary Neurulation: Neural Tube Formation in the Chick Embryo Secondary Neurulation in the Caudal Region of a 25-Somite Chick Embryo Three Views of Neurulation in an Amphibian Embryo 23


 skull (bone) 2) sensory and")


24 Neurulation in the Human Embryo Early Human Brain Development Dorsal-Ventral Specification of the Neural Tube Neural crest cells migrate away from the neural tube to develop into... 1) skull (bone) 2) sensory and autonomic nervous systems 3) pigment cells Somites are formed after gastrulation along the antero-posterior axis. Neural crest cells N C muscle Heart, kidney Gaunard, gut muscle Blood froming tissue 24




25 Cascade of Inductions Initiated by the Notochord in the Ventral Neural Tube Notochord and somite development in chick In the chick, mesoderm forms anterior to the regressing node of the primitive streak. Pre-somitic mesoderm is the region between the last formed somite and the regressing node. This region will become 4 or 5 somites which form simultaneously as pairs on either side of the notochord. The position of somites along A/P axis determines fate. Anterior somites form cervical vertebrate. Posterior ones form ribbed thoracic vertebrate. Develop in a temporal and spatial order. Rearranging pre-somitic mesoderm will not change timing. The pattern is laid down earlier by an A/P axis signal. Ephrin and Its Receptor Constitute a Possible Cut Site for Somite Formation The connection between the proteins oscillations and somatic formation is not clear; maybe Delta-Notch signaling pathway The pre-somatic mesoderm has a positional identity before somatic formation 25
, somite fate maps have been constructed.")
 and the dermatome (an epithelial sheet that forms dermis).")




26 Mesoderm becomes notochord and somites The fate of somite cells depends upon adjacent tissue signals In chick-quail trans-species grafts (with distinctive nuclei), somite fate maps have been constructed. The dorsal and lateral region of somites form the dermomyotome (which expresses Pax3). This is made up of the myotome (forms muscle cells) and the dermatome (an epithelial sheet that forms dermis). Cells from the medial region of the somite (which express MyoD) form axial & back muscles. Lateral region of somites form abdominal and limb muscles. Ventral medial region of the somite contain sclerotome cells (future cartilage which express Pax1) and migrate to surround dthe notochord and dform the vertebrate. t Notochord induces sclerotome cells. The Major Lineages of the Amniote Mesoderm Mesoderm becomes notochord and somites From notochord transplantation experiments, an additional notochord induces unsegmented pre-somitic mesoderm to produce greatly increased amount of cartilage. Neural tube (ventral side: the floor plate) induces cartilage. Lateral plate mesoderm and the ectoderm induce the dermamyotome. Signals that may pattern the somites are secreted signaling proteins that may include: 1) Sonic hedgehog which may specify the ventral somites. 2) BMP-4 which may specify the lateral somites. 3) Wnt family proteins which may specify the dorsal somites. 26


 in vertebrates.")
27 Major Postulated Interactions in the Patterning of the Somite Mesoderm and homeobox genes Regulation of the Pax homeobox genes (transcription factors) Pax genes are regulated by signals from the notochord and neural tube to control the somitic cell fate. Pax3 is expressed early in all cells that will form somites. Pax3 is modulated by BMP-4 and Wnt to confine it to muscle precursors. Pax3 is further down-regulated in back muscle precursors but remains active in future limb muscle cells. In mice, Splotch (Pax3-minus) mutants lack limb muscles. Homeobox genes: Encode a large family of transcription factors. Share a similar il 60 amino acid DNA binding homeodomain which is encoded by 180 basepair homeobox sequence. Homeotic transformation is often observed in mutants of genes that have this domain. Identified first in Drosophila (Bithorax and Antennapedia complexes) as a split cluster. There are four separate clusters of Hox genes (subset of the homeobox genes) in vertebrates. Gene activity can provide a positional identities Hox gene clusters Hox genes ( Hox gene clusters) are a subset of the homeobox genes encoding of transcription factors. Might have arisen by rounds of duplication of ancestral genes (to form a cluster), followed by further duplication of that cluster in mammals. Paralogous group are composed of the most similar members of each cluster. Partially overlapping zones of expression which vary in the anterior extent of their expression define distinct regions. Various genes respond to the combination of gene products expressed. Most homeobox genes are not Hox genes (i.e. Pax genes) Hox genes pattern the A/P axis The differences between vertebrae (i.e. anterior -attach to skull; cervical; thoracic have ribs; lumbar, sacral and caudal) clearly demonstrate that identity of somites differ along the A/P axis. Hox genes are expressed along the A/P axis in mouse. First, anterior Hox genes expressed in early gastrulation as mesoderm begins to leave the primitive streak. More posterior Hox genes turn on as development continues. Defined patterns of Hox gene expression are seen in: 1) mesoderm (after somite formation) and 2) neural tube (neuralation). Hox genes show a sharp anterior border of expression and a much less defined posterior border. Lot of overlap by every region but each has distinct pattern of Hox gene expression. Most anterior somites express Hoxa1 and Hoxb1 only. Posterior regions express all Hox genes. The anterior head, forebrain and midbrain do not express Hox genes but have other homeobox genes (etx & otx.) 27
 reflects the order of spatial and temporal expression along the A/P axis.")
 A targeting vector is constructed that has the central (functional) region of a gene replaced with a drug resistance gene.")
 form.")
28 Hox gene expression is co-linear Hoxa1 has its most anterior expression in the posterior head. Hoxa11 has its most anterior expression in the sacral (lower back) region. Hox gene expression is co-linear as order of genes on the chromosome (per cluster) reflects the order of spatial and temporal expression along the A/P axis. Hox gene expression is conserved between mouse and chick. Technique: Insertional mutagenesis (gene knock-out) A targeting vector is constructed that has the central (functional) region of a gene replaced with a drug resistance gene. This is transfected into ES cells and selected by drug exposure. By homologous recombination, a fraction of the transformants will have one copy of the original gene replaced with the altered (non-functional) form. These cells are injected into the inner cell mass of a blastocyst. Resultant chimeric mice give rise to heterozygous mutants which can be bred to generate mutant homozygotes. Altering Hox gene expression alters axial patterning In mice, gene knock-out experiments produce mutants. There is redundancy, where a missing gene can be at least partially compensated for the expression of related genes. Paralogous genes from another Hox complex may compensate for gene loss. Posterior prevalence: mutation affects the anterior extent of gene expression. Homeotic transformations result from Hox gene loss. Loss leads to cells assuming a more anterior value i.e. Hoxc8 mutant mice have extra ribs. Abnormal expression of Hox genes in anterior regions lead to tissues becoming more like posterior positioned tissues. Retinoic acid can alter positional value Retinoic acid is a derivative of vitamin A. It has very important role in signaling vertebrate development It has very important role in signaling vertebrate development. In early development, retinoic acid can cause homeotic transformation of the vertebrate. It can diffuse across plasma membranes to bind protein receptors and form an active transcription factor. Retinoic acid interferes with the normal expression of Hox genes. Later, it can alter positional development in limb development. 28


 Brachyury 2) FGF (fibroblast growth factor) Control of Hox genes is unknown.")


29 The Spemann Organizer is temporally dependent Spemann Organizer (dorsal blastopore lip) grafted to the ventral side of the marginal zone results in a twinned embryo. The second embryo can have head, trunk and, sometimes, a tail but will be joined to primary embryo along the axis. Blastopore transplants vary with time. Early gastrula induces a full 2nd embryo. Mid-gastrula induces trunk and tail (no head). Late gastrula induces only tail. The Spemann Organizer has conserved functions As gastrulation proceeds, the A/P axis become specified and as it progresses the blastopore lip can only induce more posterior structures. Hensen's node (the chick Organizer), at the anterior of the primitive streak contributes to notochord and somites and can induce another axis (more difficult than frogs) Xenopus and mouse share a number of genes that are expressed in the organizer: 1) Brachyury 2) FGF (fibroblast growth factor) Control of Hox genes is unknown. Neural plate is induced by mesoderm and in the ectorderm Neural plate is induced by signaling mechanism(s) Dorsal lip transplantation experiments demonstrate that a nervous system can be induced from ectoderm. BMP-4, secreted growth factor, inhibits cells from forming neural tissue. Inhibition of BMP-4 allows neural tissue formation. noggin (secreted by the organizer) inhibits BMP-4 and acts to dorsalize the mesoderm. noggin also induces neural tissue. chordin expressed by the future neural plate cells of the organizer. Both noggin and chordin directly bind BMP-4 and inactive it to allow the induction of neural tissue. Nervous system can be patterned by signals from the mesoderm In newt neurula, mesoderm transplantation into younger newt embryos, anterior explants induce head and brain. Posterior explants induce trunk & spinal cord. Neural plate explants induce specific neural structures (depending upon position) when transplanted beneath the ectoderm of a gastrula. Otx (orthodenticle in Drosophila), Emc are expressed in anterior to hindbrain 29
 Signal 2 turns part of this into posterior neural tissue in a graded manner (FGF, Wnts & retinoic acid are candidates).")



30 Model: The two signal model of neural patterning Signal 1 from the mesoderm induces ectoderm to become anterior neural tissue. (chordin and noggin are good candidates) Signal 2 turns part of this into posterior neural tissue in a graded manner (FGF, Wnts & retinoic acid are candidates). Mouse and chick grafts of the primitive streak (ie node or Hensen's node) can also induce neural tissue. This model differs from another model which suggests that there may exist a number of region-specific inducer molecules. Hensen's node is the chick organizer Hensen's node (chick) can induce neural gene expression in Xenopus ectoderm. This demonstrates the evolutionary conservation of neural induction signals and confirm similarity of Hensen's node and Spemann Organizer. Early nodes induce anterior structures. Later nodes induce posterior structures. The nodes can specify different A/P positional values over time. The capacity to produce signals that generate anterior structure is lost. Neural plate signals travel within the neural plate Mesoderm does not have to lie in contact with ectoderm to induce it. N-CAM, a neural cell-cell adhesion protein, neurogenic factors and other neural specific proteins can be expressed in the ectoderm in the correct A/P order in exogastrula which suggests that the inducing signal can travel a relatively long distance through the tissues. Hindbrain rhombomeres restrict cell lineage Posterior head and hindbrain development requires segmentation of the anterior neural tube. Segmentation events in the 3 day chick embryo's posterior head include... 1) somite formation from mesoderm on either side of notochord, 2) the hindbrain (rhombocephalon) is divided into 8 rhombomeres, and 3) the lateral mesoderm forms the branchial arches. (Note: spinal cord is segmented into dorsal root ganglia and ventral motor nerves by the somites) Neural tube forms segmentally arranged the segmental cranial nerves. Neural crest cells also give rise to peripheral nerves and bones including jaw (from the first branchial arch) and the bony parts of the ear (from the second arch). Eight rhombomeres form by constricting the freshly closed neural tube into eight evenly spaced sections. Lineage restriction occurs with cells and their descendants remaining in their parent rhombomere. Cell movement restriction depends on adhesive properties. Within each rhombomere, the cells are under the control of the same genes and act as a developmental unit. 30

31 Positional Identity of the rhomomere anf neural crest Chick neural crest cells can be labeled and their fate mapped. The cranial neural crest cells migrate out from the rhombomeres of the dorsal region of the hindbrain. Branchial arch 1 is populated by cells from rhombomere 2, Branchial arch 2 by rhombomere 4 cells and Branchial arch 3 by rhombomere 6 cells. Neural crest cells from rhombomeres 3 and 5 die by apoptosis (programmed cell death). Transplantation of rhombomere 2 cells to where rhombomere 4 cells should be results in formation of a second jaw. Hindbrain Hox gene expression Mouse embryonic hindbrain Hox gene expression is well defined and correlates well with the segmental pattern. Paralogous group 1 (Hoxa1, b1 etc.) is expressed anterior to paralog 2 (Hoxa2, b2 etc.) followed by paralog 3. Control of gene expression is complex (i.e. Hoxb2 is controlled in rhombomere 3 and 5 by one enhancer which is activated by Krox-20 and in r4 by another enhancer). Hoxa2 knock-out mice show head skeletal defects that are consistent with a homeotic transformation of the inner ear (2nd branchial arch) to jaw (1st arch). (By neurula stage, the embryo is divided into regulating organ-forming regions.) 31
Role of Organizer Chages in Late Frog Embryos
 Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm December 7, 2006 Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of BMPs more lateral mesoderm
Biology 4361 Paraxial and Intermediate Mesoderm December 7, 2006 Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of BMPs more lateral mesoderm
2/23/09. Regional differentiation of mesoderm. Morphological changes at early postgastrulation. Segments organize the body plan during embryogenesis
 Regional differentiation of mesoderm Axial Paraxial Intermediate Somatic Splanchnic Chick embryo Morphological changes at early postgastrulation stages Segments organize the body plan during embryogenesis
Regional differentiation of mesoderm Axial Paraxial Intermediate Somatic Splanchnic Chick embryo Morphological changes at early postgastrulation stages Segments organize the body plan during embryogenesis
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Life Sciences For NET & SLET Exams Of UGC-CSIR. Section B and C. Volume-08. Contents A. BASIC CONCEPT OF DEVELOPMENT 1
 Section B and C Volume-08 Contents 5. DEVELOPMENTAL BIOLOGY A. BASIC CONCEPT OF DEVELOPMENT 1 B. GAMETOGENESIS, FERTILIZATION AND EARLY DEVELOPMENT 23 C. MORPHOGENESIS AND ORGANOGENESIS IN ANIMALS 91 0
Section B and C Volume-08 Contents 5. DEVELOPMENTAL BIOLOGY A. BASIC CONCEPT OF DEVELOPMENT 1 B. GAMETOGENESIS, FERTILIZATION AND EARLY DEVELOPMENT 23 C. MORPHOGENESIS AND ORGANOGENESIS IN ANIMALS 91 0
Question Set # 4 Answer Key 7.22 Nov. 2002
 Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways
Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Mesoderm Development
 Quiz rules: Spread out across available tables No phones, text books, or (lecture) notes on your desks No consultation with your colleagues No websites open other than the Quiz page No screen snap shots
Quiz rules: Spread out across available tables No phones, text books, or (lecture) notes on your desks No consultation with your colleagues No websites open other than the Quiz page No screen snap shots
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018
 Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
!!!!!!!! DB3230 Midterm 2 12/13/2013 Name:
 1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
Biology 218, practise Exam 2, 2011
 Figure 3 The long-range effect of Sqt does not depend on the induction of the endogenous cyc or sqt genes. a, Design and predictions for the experiments shown in b-e. b-e, Single-cell injection of 4 pg
Figure 3 The long-range effect of Sqt does not depend on the induction of the endogenous cyc or sqt genes. a, Design and predictions for the experiments shown in b-e. b-e, Single-cell injection of 4 pg
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
9/4/2015 INDUCTION CHAPTER 1. Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology. Fig 1.
 INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm July 28, 2008 Paraxial and Intermediate Mesoderm Overview Development of major mesodermal lineages Somites: formation specification and differentiation Mesodermal
Biology 4361 Paraxial and Intermediate Mesoderm July 28, 2008 Paraxial and Intermediate Mesoderm Overview Development of major mesodermal lineages Somites: formation specification and differentiation Mesodermal
MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning
 MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning Learning Goals for Lecture 29 4.1 Describe the contributions of early developmental events in the embryo to the formation
MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning Learning Goals for Lecture 29 4.1 Describe the contributions of early developmental events in the embryo to the formation
Lecture 3 - Molecular Regulation of Development. Growth factor signaling, Hox genes and the body plan
 Lecture 3 - Molecular Regulation of Development. Growth factor signaling, Hox genes and the body plan Lecture Objectives Outline August 18, 2015, M.D., Ph.D. To understand how cell differentiation and
Lecture 3 - Molecular Regulation of Development. Growth factor signaling, Hox genes and the body plan Lecture Objectives Outline August 18, 2015, M.D., Ph.D. To understand how cell differentiation and
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION
 MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
Name. Biology Developmental Biology Winter Quarter 2013 KEY. Midterm 3
 Name 100 Total Points Open Book Biology 411 - Developmental Biology Winter Quarter 2013 KEY Midterm 3 Read the Following Instructions: * Answer 20 questions (5 points each) out of the available 25 questions
Name 100 Total Points Open Book Biology 411 - Developmental Biology Winter Quarter 2013 KEY Midterm 3 Read the Following Instructions: * Answer 20 questions (5 points each) out of the available 25 questions
Name KEY. Biology Developmental Biology Winter Quarter Midterm 3 KEY
 Name KEY 100 Total Points Open Book Biology 411 - Developmental Biology Winter Quarter 2009 Midterm 3 KEY All of the 25 multi-choice questions are single-answer. Choose the best answer. (4 pts each) Place
Name KEY 100 Total Points Open Book Biology 411 - Developmental Biology Winter Quarter 2009 Midterm 3 KEY All of the 25 multi-choice questions are single-answer. Choose the best answer. (4 pts each) Place
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8
 Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Chapter 10 Development and Differentiation
 Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Questions in developmental biology. Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration
 Questions in developmental biology Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration Representative cell types of a vertebrate zygote => embryo => adult differentiation
Questions in developmental biology Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration Representative cell types of a vertebrate zygote => embryo => adult differentiation
Developmental processes Differential gene expression Introduction to determination The model organisms used to study developmental processes
 Date Title Topic(s) Learning Outcomes: Sept 28 Oct 3 1. What is developmental biology and why should we care? 2. What is so special about stem cells and gametes? Developmental processes Differential gene
Date Title Topic(s) Learning Outcomes: Sept 28 Oct 3 1. What is developmental biology and why should we care? 2. What is so special about stem cells and gametes? Developmental processes Differential gene
Exam 3 (Final Exam) December 20, 2007
 December 20, 2007") Biology 4361 Exam 3 (Final Exam) December 20, 2007 Name: ID: Multiple choice (1 point each. Indicate the best answer.) 1. During Drosophila gastrulation, mesoderm moves in through the a. primitives streak.
Biology 4361 Exam 3 (Final Exam) December 20, 2007 Name: ID: Multiple choice (1 point each. Indicate the best answer.) 1. During Drosophila gastrulation, mesoderm moves in through the a. primitives streak.
Bio Section III Organogenesis. The Neural Crest and Axonal Specification. Student Learning Objectives. Student Learning Objectives
 Bio 127 - Section III Organogenesis The Neural Crest and Axonal Specification Gilbert 9e Chapter 10 Student Learning Objectives 1. You should understand that the neural crest is an evolutionary advancement
Bio 127 - Section III Organogenesis The Neural Crest and Axonal Specification Gilbert 9e Chapter 10 Student Learning Objectives 1. You should understand that the neural crest is an evolutionary advancement
Sonic hedgehog (Shh) signalling in the rabbit embryo
 signalling in the rabbit embryo") Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
Limb Development Involving the development of the appendicular skeleton and muscles
 Limb Development Involving the development of the appendicular skeleton and muscles 1 Objectives Timing and location of limb bud development The tissues from which limb buds are made Determining the position
Limb Development Involving the development of the appendicular skeleton and muscles 1 Objectives Timing and location of limb bud development The tissues from which limb buds are made Determining the position
Maternal Control of GermLayer Formation in Xenopus
 Maternal Control of GermLayer Formation in Xenopus The zygotic genome is activated at the mid-blastula transition mid-blastula fertilized egg Xenopus gastrulae early-gastrula 7 hrs 10 hrs control not VP
Maternal Control of GermLayer Formation in Xenopus The zygotic genome is activated at the mid-blastula transition mid-blastula fertilized egg Xenopus gastrulae early-gastrula 7 hrs 10 hrs control not VP
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
10/15/09. Tetrapod Limb Development & Pattern Formation. Developing limb region is an example of a morphogenetic field
 Tetrapod Limb Development & Pattern Formation Figure 16.5(1) Limb Bud Formation derived from lateral plate (somatic) & paraxial (myotome) Fig. 16.2 Prospective Forelimb Field of Salamander Ambystoma maculatum
Tetrapod Limb Development & Pattern Formation Figure 16.5(1) Limb Bud Formation derived from lateral plate (somatic) & paraxial (myotome) Fig. 16.2 Prospective Forelimb Field of Salamander Ambystoma maculatum
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
BIOLOGY - CLUTCH CH.32 - OVERVIEW OF ANIMALS.
 !! www.clutchprep.com Animals are multicellular, heterotrophic eukaryotes that feed by ingesting their food Most animals are diploid, and produce gametes produced directly by meiosis Animals lack cell
!! www.clutchprep.com Animals are multicellular, heterotrophic eukaryotes that feed by ingesting their food Most animals are diploid, and produce gametes produced directly by meiosis Animals lack cell
4. Neural tube cells are specified by opposing dorsal-ventral gradients of a. Wnts and Nodal. b. FGF and Shh. c. BMPs and Wnts. d. BMPs and Shh.
 Biology 4361 Name: KEY Exam 4 ID#: August 1, 2008 Multiple choice (one point each; indicate the best answer) 1. Neural tube closure is accomplished by movement of the a. medial hinge point cells. b. medial
Biology 4361 Name: KEY Exam 4 ID#: August 1, 2008 Multiple choice (one point each; indicate the best answer) 1. Neural tube closure is accomplished by movement of the a. medial hinge point cells. b. medial
Developmental Biology Lecture Outlines
 Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Bio 127 Section I Introduction to Developmental Biology. Cell Cell Communication in Development. Developmental Activities Coordinated in this Way
 Bio 127 Section I Introduction to Developmental Biology Cell Cell Communication in Development Gilbert 9e Chapter 3 It has to be EXTREMELY well coordinated for the single celled fertilized ovum to develop
Bio 127 Section I Introduction to Developmental Biology Cell Cell Communication in Development Gilbert 9e Chapter 3 It has to be EXTREMELY well coordinated for the single celled fertilized ovum to develop
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Developmental Zoology. Ectodermal derivatives (ZOO ) Developmental Stages. Developmental Stages
 Developmental Stages. Developmental Stages") Developmental Zoology (ZOO 228.1.0) Ectodermal derivatives 1 Developmental Stages Ø Early Development Fertilization Cleavage Gastrulation Neurulation Ø Later Development Organogenesis Larval molts Metamorphosis
Developmental Zoology (ZOO 228.1.0) Ectodermal derivatives 1 Developmental Stages Ø Early Development Fertilization Cleavage Gastrulation Neurulation Ø Later Development Organogenesis Larval molts Metamorphosis
Homeotic Genes and Body Patterns
 Homeotic Genes and Body Patterns Every organism has a unique body pattern. Although specialized body structures, such as arms and legs, may be similar in makeup (both are made of muscle and bone), their
Homeotic Genes and Body Patterns Every organism has a unique body pattern. Although specialized body structures, such as arms and legs, may be similar in makeup (both are made of muscle and bone), their
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis
 18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
10/2/2015. Chapter 4. Determination and Differentiation. Neuroanatomical Diversity
 Chapter 4 Determination and Differentiation Neuroanatomical Diversity 1 Neurochemical diversity: another important aspect of neuronal fate Neurotransmitters and their receptors Excitatory Glutamate Acetylcholine
Chapter 4 Determination and Differentiation Neuroanatomical Diversity 1 Neurochemical diversity: another important aspect of neuronal fate Neurotransmitters and their receptors Excitatory Glutamate Acetylcholine
Genes, Development, and Evolution
 14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
Skeletal Development in Human
 Atlas of Genetics and Cytogenetics in Oncology and Haematology Skeletal Development in Human Skeletal development in human - Long version I. Introduction I.1 Developmental genes in Drosophila I.2 Skeletal
Atlas of Genetics and Cytogenetics in Oncology and Haematology Skeletal Development in Human Skeletal development in human - Long version I. Introduction I.1 Developmental genes in Drosophila I.2 Skeletal
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
3/8/ Complex adaptations. 2. often a novel trait
 Chapter 10 Adaptation: from genes to traits p. 302 10.1 Cascades of Genes (p. 304) 1. Complex adaptations A. Coexpressed traits selected for a common function, 2. often a novel trait A. not inherited from
Chapter 10 Adaptation: from genes to traits p. 302 10.1 Cascades of Genes (p. 304) 1. Complex adaptations A. Coexpressed traits selected for a common function, 2. often a novel trait A. not inherited from
Neural development its all connected
 Neural development its all connected How do you build a complex nervous system? How do you build a complex nervous system? 1. Learn how tissue is instructed to become nervous system. Neural induction 2.
Neural development its all connected How do you build a complex nervous system? How do you build a complex nervous system? 1. Learn how tissue is instructed to become nervous system. Neural induction 2.
UNIVERSITY OF YORK BIOLOGY. Developmental Biology
 Examination Candidate Number: UNIVERSITY OF YORK BSc Stage 2 Degree Examinations 2017-18 Department: BIOLOGY Title of Exam: Developmental Biology Desk Number: Time allowed: 1 hour and 30 minutes Total
Examination Candidate Number: UNIVERSITY OF YORK BSc Stage 2 Degree Examinations 2017-18 Department: BIOLOGY Title of Exam: Developmental Biology Desk Number: Time allowed: 1 hour and 30 minutes Total
PRACTICE EXAM. 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos.
 PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
2. Fertilization activates the egg and bring together the nuclei of sperm and egg
 2. Fertilization activates the egg and bring together the nuclei of sperm and egg Sea urchins (what phylum?) are models for the study of the early development of deuterostomes (like us, right?). Sea urchin
2. Fertilization activates the egg and bring together the nuclei of sperm and egg Sea urchins (what phylum?) are models for the study of the early development of deuterostomes (like us, right?). Sea urchin
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Exam 3 ID#: July 31, 2009
 Biology 4361 Name: KEY Exam 3 ID#: July 31, 2009 Multiple choice (one point each; indicate the best answer) 1. Neural tube closure is accomplished by movement of the a. medial hinge point cells. b. medial
Biology 4361 Name: KEY Exam 3 ID#: July 31, 2009 Multiple choice (one point each; indicate the best answer) 1. Neural tube closure is accomplished by movement of the a. medial hinge point cells. b. medial
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its
 The Wnt/ -catenin signalling pathway and its") Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Bi 117 Final (60 pts) DUE by 11:00 am on March 15, 2012 Box by Beckman Institute B9 or to a TA
 DUE by 11:00 am on March 15, 2012 Box by Beckman Institute B9 or to a TA") Bi 117 Final (60 pts) DUE by 11:00 am on March 15, 2012 Box by Beckman Institute B9 or to a TA Instructor: Marianne Bronner Exam Length: 6 hours plus one 30-minute break at your discretion. It should take
Bi 117 Final (60 pts) DUE by 11:00 am on March 15, 2012 Box by Beckman Institute B9 or to a TA Instructor: Marianne Bronner Exam Length: 6 hours plus one 30-minute break at your discretion. It should take
Determination of Spatiotemporal Interactions Regulating Head/Trunk Axial Pattern in the Chick Embryo
 Eukaryon, Vol. 2, January 2006, Lake Forest ollege Senior Thesis Determination of Spatiotemporal Interactions Regulating Head/Trunk Axial Pattern in the hick Embryo David Mann * Department of Biology,
Eukaryon, Vol. 2, January 2006, Lake Forest ollege Senior Thesis Determination of Spatiotemporal Interactions Regulating Head/Trunk Axial Pattern in the hick Embryo David Mann * Department of Biology,
Homeotic genes in flies. Sem 9.3.B.6 Animal Science
 Homeotic genes in flies Sem 9.3.B.6 Animal Science So far We have seen that identities of each segment is determined by various regulators of segment polarity genes In arthopods, and in flies, each segment
Homeotic genes in flies Sem 9.3.B.6 Animal Science So far We have seen that identities of each segment is determined by various regulators of segment polarity genes In arthopods, and in flies, each segment
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Early Development in Invertebrates
 Developmental Biology Biology 4361 Early Development in Invertebrates October 25, 2006 Early Development Overview Cleavage rapid cell divisions divisions of fertilized egg into many cells Gastrulation
Developmental Biology Biology 4361 Early Development in Invertebrates October 25, 2006 Early Development Overview Cleavage rapid cell divisions divisions of fertilized egg into many cells Gastrulation
Paraxial and Intermediate Mesoderm
 12 Paraxial and Intermediate Mesoderm chapters 10 and 11 followed the formation of various tissues from the vertebrate ectoderm. In this chapter and the next, we will follow the development of the mesodermal
12 Paraxial and Intermediate Mesoderm chapters 10 and 11 followed the formation of various tissues from the vertebrate ectoderm. In this chapter and the next, we will follow the development of the mesodermal
Making Headway: The Roles of Hox Genes and Neural Crest Cells in Craniofacial Development
 Mini-Review TheScientificWorldJOURNAL (2003) 3, 240 264 ISSN 1537-744X; DOI 10.1100/tsw.2003.11 Making Headway: The Roles of Hox Genes and Neural Crest Cells in Craniofacial Development Paul A Trainor
Mini-Review TheScientificWorldJOURNAL (2003) 3, 240 264 ISSN 1537-744X; DOI 10.1100/tsw.2003.11 Making Headway: The Roles of Hox Genes and Neural Crest Cells in Craniofacial Development Paul A Trainor
Mesoderm induction: from caps to chips
 Mesoderm induction: from caps to chips David Kimelman Abstract Vertebrate mesoderm induction is one of the classical problems in developmental biology. Various developmental biology approaches, particularly
Mesoderm induction: from caps to chips David Kimelman Abstract Vertebrate mesoderm induction is one of the classical problems in developmental biology. Various developmental biology approaches, particularly
Unit 4 Evaluation Question 1:
 Name: Unit 4 Evaluation Question 1: /7 points A naturally occurring dominant mutant in mice is the Doublefoot (Dbf) mutant. Below is an image of the bones from a wildtype (wt) and Doublefoot mutant mouse.
Name: Unit 4 Evaluation Question 1: /7 points A naturally occurring dominant mutant in mice is the Doublefoot (Dbf) mutant. Below is an image of the bones from a wildtype (wt) and Doublefoot mutant mouse.
THE PROBLEMS OF DEVELOPMENT. Cell differentiation. Cell determination
 We emphasize these points from Kandel in Bi/CNS 150 Bi/CNS/NB 150: Neuroscience Read Lecture Lecture Friday, October 2, 2015 Development 1: pp 5-10 Introduction Brains evolved All higher animals have brains
We emphasize these points from Kandel in Bi/CNS 150 Bi/CNS/NB 150: Neuroscience Read Lecture Lecture Friday, October 2, 2015 Development 1: pp 5-10 Introduction Brains evolved All higher animals have brains
Mesoderm Divided into three main types - Paraxial (somite) - Intermediate - Lateral (somatic and splanchnic)
 - Intermediate - Lateral (somatic and splanchnic)") Mesoderm Divided into three main types - Paraxial (somite) - Intermediate - Lateral (somatic and splanchnic) Fates of Mesoderm Paraxial - Dermis of skin - Axial Skeleton - Axial and limb muscles/tendons
Mesoderm Divided into three main types - Paraxial (somite) - Intermediate - Lateral (somatic and splanchnic) Fates of Mesoderm Paraxial - Dermis of skin - Axial Skeleton - Axial and limb muscles/tendons
Polarity and Segmentation
 P618621-002.qxd 8/29/05 4:57 PM Page 29 C H P T E R 2 Polarity and Segmentation REGIONL IDENTITY OF THE NERVOUS SYSTEM Like the rest of the body in most metazoans, the nervous system is regionally specialized.
P618621-002.qxd 8/29/05 4:57 PM Page 29 C H P T E R 2 Polarity and Segmentation REGIONL IDENTITY OF THE NERVOUS SYSTEM Like the rest of the body in most metazoans, the nervous system is regionally specialized.
7.013 Problem Set
 7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
Coordination of Cell Differentiation and Migration in Mathematical Models of Caudal Embryonic Axis Extension
 Coordination of Cell Differentiation and Migration in Mathematical Models of Caudal Embryonic Axis Extension Nigel C. Harrison 1, Ruth Diez del Corral 2 *, Bakhtier Vasiev 1 * 1 Department of Mathematical
Coordination of Cell Differentiation and Migration in Mathematical Models of Caudal Embryonic Axis Extension Nigel C. Harrison 1, Ruth Diez del Corral 2 *, Bakhtier Vasiev 1 * 1 Department of Mathematical
Head and Face Development
 Head and Face Development Resources: http://php.med.unsw.edu.au/embryology/ Larsen s Human Embryology The Developing Human: Clinically Oriented Embryology Dr Annemiek Beverdam School of Medical Sciences,
Head and Face Development Resources: http://php.med.unsw.edu.au/embryology/ Larsen s Human Embryology The Developing Human: Clinically Oriented Embryology Dr Annemiek Beverdam School of Medical Sciences,
Lesson 10. References: Chapter 8: Reading for Next Lesson: Chapter 8:
 Lesson 10 Lesson Outline: General Features of the Skeletal System Axial Skeleton (Vertebral Column) Ontogenetic Origins (Morphogenesis) of the Skeletal System Functions of the Axial Skeleton Phylogenetic
Lesson 10 Lesson Outline: General Features of the Skeletal System Axial Skeleton (Vertebral Column) Ontogenetic Origins (Morphogenesis) of the Skeletal System Functions of the Axial Skeleton Phylogenetic
Chapter 32. Objectives. Table of Contents. Characteristics. Characteristics, continued. Section 1 The Nature of Animals
 Introduction to Animals Table of Contents Objectives Identify four important characteristics of animals. List two kinds of tissues found only in animals. Explain how the first animals may have evolved
Introduction to Animals Table of Contents Objectives Identify four important characteristics of animals. List two kinds of tissues found only in animals. Explain how the first animals may have evolved
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11589 Supplementary Figure 1 Ciona intestinalis and Petromyzon marinus neural crest expression domain comparison. Cartoon shows dorsal views of Ciona mid gastrula (left) and Petromyzon
doi:10.1038/nature11589 Supplementary Figure 1 Ciona intestinalis and Petromyzon marinus neural crest expression domain comparison. Cartoon shows dorsal views of Ciona mid gastrula (left) and Petromyzon
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Lecture 2 - Making babies: Organ formation in the Ectoderm, Mesoderm, Endoderm and Neural Crest. Outline August 15, 2016 Eddy De Robertis, M.D., Ph.D.
 Lecture 2 - Making babies: Organ formation in the Ectoderm, Mesoderm, Endoderm and Neural Crest Lecture Objectives Outline August 15, 2016, M.D., Ph.D. - To examine how the main organ systems are formed
Lecture 2 - Making babies: Organ formation in the Ectoderm, Mesoderm, Endoderm and Neural Crest Lecture Objectives Outline August 15, 2016, M.D., Ph.D. - To examine how the main organ systems are formed
AP Biology Gene Regulation and Development Review
 AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
Avian hairy Gene Expression Iden4fies a Molecular Clock Linked to Vertebrate Segmenta4on and Somitogenesis
 Avian hairy Gene Expression Iden4fies a Molecular Clock Linked to Vertebrate Segmenta4on and Somitogenesis Pourquie, et. al., 1997 Swetha Mummini, Victoria Fong What is a somite? Defini4on: Segmental block
Avian hairy Gene Expression Iden4fies a Molecular Clock Linked to Vertebrate Segmenta4on and Somitogenesis Pourquie, et. al., 1997 Swetha Mummini, Victoria Fong What is a somite? Defini4on: Segmental block
Morphogenesis and specification of the muscle lineage during Xenopus laevis embryo development. Armbien F. Sabillo
 Morphogenesis and specification of the muscle lineage during Xenopus laevis embryo development by Armbien F. Sabillo A dissertation submitted in partial satisfaction of the requirements for the degree
Morphogenesis and specification of the muscle lineage during Xenopus laevis embryo development by Armbien F. Sabillo A dissertation submitted in partial satisfaction of the requirements for the degree
Somites without a clock
 Dias, AS; de Almeida, I; Belmonte, JM; Glazier, JA; Stern, CD; (2014) Somites Without a Clock. Science 10.1126/science.1247575. ARTICLE Somites without a clock Ana S. Dias 1,, Irene de Almeida 1,, Julio
Dias, AS; de Almeida, I; Belmonte, JM; Glazier, JA; Stern, CD; (2014) Somites Without a Clock. Science 10.1126/science.1247575. ARTICLE Somites without a clock Ana S. Dias 1,, Irene de Almeida 1,, Julio
THE MAKING OF THE SOMITE: MOLECULAR EVENTS IN VERTEBRATE SEGMENTATION. Yumiko Saga* and Hiroyuki Takeda
 THE MAKING OF THE SOMITE: MOLECULAR EVENTS IN VERTEBRATE SEGMENTATION Yumiko Saga* and Hiroyuki Takeda The reiterated structures of the vertebrate axial skeleton, spinal nervous system and body muscle
THE MAKING OF THE SOMITE: MOLECULAR EVENTS IN VERTEBRATE SEGMENTATION Yumiko Saga* and Hiroyuki Takeda The reiterated structures of the vertebrate axial skeleton, spinal nervous system and body muscle
Exam 2 ID#: November 9, 2006
 Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Molecular Cell Biology of Brain Development
 1 Molecular Cell Biology of Brain Development Takayoshi Inoue National Institute of Neuroscience, 4-1-1, Ogawahigashi, Kodaira, Tokyo 187-8502, Japan 1 Neural Induction The First Step Involved in Forming
1 Molecular Cell Biology of Brain Development Takayoshi Inoue National Institute of Neuroscience, 4-1-1, Ogawahigashi, Kodaira, Tokyo 187-8502, Japan 1 Neural Induction The First Step Involved in Forming
A systems approach to biology
 A systems approach to biology SB200 Lecture 7 7 October 2008 Jeremy Gunawardena jeremy@hms.harvard.edu Recap of Lecture 6 In phage lambda, cooperativity leads to bistability and hysteresis In HIV-1, sequestration
A systems approach to biology SB200 Lecture 7 7 October 2008 Jeremy Gunawardena jeremy@hms.harvard.edu Recap of Lecture 6 In phage lambda, cooperativity leads to bistability and hysteresis In HIV-1, sequestration
Tsukushi Modulates Xnr2, FGF and BMP Signaling: Regulation of Xenopus Germ Layer Formation
 Tsukushi Modulates Xnr2, FGF and BMP Signaling: Regulation of Xenopus Germ Layer Formation Samantha A. Morris 1 *, Alexandra D. Almeida 1, Hideaki Tanaka 2, Kunimasa Ohta 2, Shin-ichi Ohnuma 1 * 1 Department
Tsukushi Modulates Xnr2, FGF and BMP Signaling: Regulation of Xenopus Germ Layer Formation Samantha A. Morris 1 *, Alexandra D. Almeida 1, Hideaki Tanaka 2, Kunimasa Ohta 2, Shin-ichi Ohnuma 1 * 1 Department
THE SPECIFICATION OF DORSAL CELL FATES IN THE VERTEBRATE CENTRAL NERVOUS SYSTEM
 Annu. Rev. Neurosci. 1999. 22:261 94 Copyright c 1999 by Annual Reviews. All rights reserved THE SPECIFICATION OF DORSAL CELL FATES IN THE VERTEBRATE CENTRAL NERVOUS SYSTEM Kevin J. Lee and Thomas M. Jessell
Annu. Rev. Neurosci. 1999. 22:261 94 Copyright c 1999 by Annual Reviews. All rights reserved THE SPECIFICATION OF DORSAL CELL FATES IN THE VERTEBRATE CENTRAL NERVOUS SYSTEM Kevin J. Lee and Thomas M. Jessell
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
In ovo time-lapse analysis after dorsal neural tube ablation shows rerouting of chick hindbrain neural crest
 In ovo time-lapse analysis after dorsal neural tube ablation shows rerouting of chick hindbrain neural crest Paul Kulesa, Marianne Bronner-Fraser and Scott Fraser (2000) Presented by Diandra Lucia Background
In ovo time-lapse analysis after dorsal neural tube ablation shows rerouting of chick hindbrain neural crest Paul Kulesa, Marianne Bronner-Fraser and Scott Fraser (2000) Presented by Diandra Lucia Background
SIGNIFICANCE OF EMBRYOLOGY
 This lecture will discuss the following topics : Definition of Embryology Significance of Embryology Old and New Frontiers Introduction to Molecular Regulation and Signaling Descriptive terms in Embryology
This lecture will discuss the following topics : Definition of Embryology Significance of Embryology Old and New Frontiers Introduction to Molecular Regulation and Signaling Descriptive terms in Embryology
Nature Neuroscience: doi: /nn.2662
 Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
inhibition of embryonic myogenesis
 Dermomyotome development and myotome patterning require Tbx6 inhibition of embryonic myogenesis Stefanie E. Windner 1,2, Nathan C. Bird 1, Sara Elizabeth Patterson 1, Rosemarie Doris 1, and Stephen Henri
Dermomyotome development and myotome patterning require Tbx6 inhibition of embryonic myogenesis Stefanie E. Windner 1,2, Nathan C. Bird 1, Sara Elizabeth Patterson 1, Rosemarie Doris 1, and Stephen Henri
Organization of Vertebrate Body. Organization of Vertebrate Body
 The Animal Body and Principles of Regulation Chapter 43 There are four levels of organization: 1. Cells 2. Tissues 3. Organs 4. Organ systems Bodies of vertebrates are composed of different cell types
The Animal Body and Principles of Regulation Chapter 43 There are four levels of organization: 1. Cells 2. Tissues 3. Organs 4. Organ systems Bodies of vertebrates are composed of different cell types
BONE MORPHOGENETIC PROTEINS AS REGULATORS OF NEURAL TUBE DEVELOPMENT
 Papers on Anthropology XX, 2011, pp. 314 326 BONE MORPHOGENETIC PROTEINS AS REGULATORS OF NEURAL TUBE DEVELOPMENT Aimar Namm 1,2, Andres Arend 1, Marina Aunapuu 1,2 1 Department of Anatomy, University
Papers on Anthropology XX, 2011, pp. 314 326 BONE MORPHOGENETIC PROTEINS AS REGULATORS OF NEURAL TUBE DEVELOPMENT Aimar Namm 1,2, Andres Arend 1, Marina Aunapuu 1,2 1 Department of Anatomy, University
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.138/nature1237 a b retinol retinal RA OH RDH (retinol dehydrogenase) O H Raldh2 O R/R.6.4.2 (retinaldehyde dehydrogenase 2) RA retinal retinol..1.1 1 Concentration (nm)
SUPPLEMENTARY INFORMATION doi:1.138/nature1237 a b retinol retinal RA OH RDH (retinol dehydrogenase) O H Raldh2 O R/R.6.4.2 (retinaldehyde dehydrogenase 2) RA retinal retinol..1.1 1 Concentration (nm)
Biology 224 Human Anatomy and Physiology - II Week 1; Lecture 1; Monday Dr. Stuart S. Sumida. Review of Early Development of Humans.
 Biology 224 Human Anatomy and Physiology - II Week 1; Lecture 1; Monday Dr. Stuart S. Sumida Review of Early Development of Humans Special Senses Review: Historical and Developmental Perspectives Ontogeny
Biology 224 Human Anatomy and Physiology - II Week 1; Lecture 1; Monday Dr. Stuart S. Sumida Review of Early Development of Humans Special Senses Review: Historical and Developmental Perspectives Ontogeny