The sevenless signalling cassette mediates Drosophila EGF receptor function during epidermal development
|
|
|
- Ezra Patrick
- 6 years ago
- Views:
Transcription
1 Development 120, (1994) Printed in Great Britain The Company of Biologists Limited The sevenless signalling cassette mediates Drosophila EGF receptor function during epidermal development Fernando J. Diaz-Benjumea 2 and Ernst Hafen 1 1 Zoologisches Institut, Universität Zürich, Winterthurerstrasse 190, CH-8057 Zürich, Switzerland 2 EMBL, Meyerhoferstrasse 1, Postfach , Heidelberg, Deutschland SUMMARY In Drosophila, Drk, an SH2 adaptor protein, Sos, a putative activator of Ras1, Ras1, raf and rolled/map kinase have been shown to be required for signalling from the sevenless and the torso receptor tyrosine kinase. From these studies, it was unclear whether these components act in a single linear pathway as suggested by the genetic analysis or whether different components serve to integrate different signals. We have analyzed the effects of removing each of these components during the development of the adult epidermal structures by generating clones of homozygous mutant cells in a heterozygous background. Mutations in each of these signalling components produce a very similar set of phenotypes. These phenotypes resemble those caused by loss-of-function mutations in the Drosophila EGF receptor homolog (DER). It appears that these components form a signalling cassette, which mediates all aspects of DER signalling but that is not required for other signalling processes during epidermal development. Key words: Drosophila, sevenless, torso, EGF, DER, receptor, signal transduction, Drk, Sos, Ras1, raf, MAP kinase, wing development, vein formation INTRODUCTION Cellular interactions play an important role during growth and morphogenesis of multicellular organisms. Genetic and biochemical studies in different organisms have shown that a family of cell surface receptors with intrinsic tyrosine kinase activity mediates a wide variety of different responses to extracellular signals. The c-kit receptor tyrosine kinase that is encoded by the White locus of the mouse is required for the migration of germ cells and the differentiation of melanocytes (Chabot et al., 1988; Geissler et al., 1988). In Caenorhabditis elegans, the differentiation of the vulval precursor cells depends on the activation of the let-23 receptor tyrosine kinase (Aroian et al., 1990). In Drosophila, the torso receptor tyrosine kinase is required for the specification of terminal structures in the embryo (Klingler et al., 1988; Casanova and Struhl, 1989; Sprenger et al., 1989) and the sevenless receptor tyrosine kinase controls the specification of the R7 photoreceptor cell fate in the developing eye (Banerjee et al., 1987; Hafen et al., 1987; Basler and Hafen, 1988; Bowtell et al., 1988). In contrast to the single mutant phenotypes associated with torso and sevenless, mutations in the gene encoding the Drosophila homolog of the EGF receptor (DER), produce pleiotropic phenotypes: viable DER alleles prevent dorsoventral patterning of the follicle cells during oogenesis (Schüpbach, 1987). Strong loss-of-function alleles cause embryonic lethality (Price et al., 1989; Schejter and Shilo, 1989; Raz and Shilo, 1993). During postembryonic development, DER function is required for the differentiation of wing veins, the control of cell size and the development of photoreceptor cells (Diaz-Benjumea and Garcia-Bellido, 1990a; Clifford and Schüpbach, 1992; Xu and Rubin, 1993). The gain-of-function allele Ellipse (Elp) causes the formation of a reduced number of ommatidial units in the eye and additional veins in the wing (Baker and Rubin, 1989, 1992). In spite of the plethora of developmental decisions controlled by receptor tyrosine kinases, genetic as well as biochemical characterization of components involved in signal transmission suggests that the different receptors utilize a common set of signal transduction components. The response of rat PC12 cells to treatment by phorbol esters or NGF is mediated by Ras and results in the activation of the c-raf serine/threonine kinase and the mitogen-activated kinase (MAPK or ERK1) (Howe et al., 1992; Thomas et al., 1992; Wood et al., 1992). In C. elegans, the Sem-5 SH2 adaptor protein, let-60 Ras and lin-45 raf are required for signalling from the let-23 receptor during vulval development (Beitel et al., 1990; Han and Sternberg, 1990; Clark et al., 1992; Han et al., 1993). In Drosophila, genetic screens for dominant modifiers of either hypomorphic or gain-of-function sevenless mutations have indicated that Drk, an SH3-SH2-SH3 adaptor protein, Sos, a putative guanine-nucleotide releasing factor, and Ras1 are essential components of the sevenless signalling cascade (Rogge et al., 1991; Simon et al., 1991, 1993; Olivier et al., 1993). Mutations in Drk, Sos and Ras1 also act as dominant modifiers of gain-of-function mutations of torso and DER (Simon et al., 1991; Doyle and Bishop, 1993). Mutations in raf block signalling from torso and sevenless (Ambrosio et al., 1989; Dickson et al., 1992). Recently, it has been shown that MAP kinase, which is encoded by the rolled gene, plays
2 570 F. J. Diaz-Benjumea and E. Hafen an essential role in the torso and the sevenless signalling pathway downstream of raf (Brunner et al., 1994; Biggs et al., unpublished). Constitutively activated versions Ras1 and raf have been constructed and shown to be sufficient to induce differentiation of terminal structures or R7 photoreceptor cells when expressed in the embryo or in the developing eye, respectively (Dickson et al., 1992; Fortini et al., 1992; Lu et al., 1993). Similarly, a gain-of-function mutation in rolled (rl Sem ) results in the activation of the torso, sevenless and DER pathways (Brunner et al., 1994). This suggested that the activation of these components is not only necessary but also sufficient to transduce the signal mediated by sevenless, torso and DER. Since most of the signal transduction components in the sevenless and the torso pathway have been identified as dominant modifiers in individual genetically sensitized pathways, it is not known in how many different signalling processes these components are involved. Animals homozygous for loss-of-function mutations in any of these genes die during embryonic or larval development. We decided to analyze the phenotype of loss-of-function mutations in Drk, Sos, Ras1, raf and rolled in clones of homozygous mutant cells in the adult epidermal structures. The phenotypes observed for loss-of-function mutations in Drk, Sos, Ras1, raf, and rolled/map kinase should reflect the sum of the phenotypes observed for the different receptors that signal via these components. If we observed phenotypes not accounted for by mutations in the known receptor tyrosine kinases, it is likely that there are additional receptors that utilize these signalling components. Furthermore, it is also possible that the phenotypes of the individual signal transduction components differ from each other. This may suggest that some of the components receive input from multiple signalling pathways whereas others do not. The results presented here indicate that the phenotypes of cell clones mutant for either Drk, Sos, Ras1, raf or rolled are identical in all aspects to the phenotypes described for cell clones mutant for DER. This suggests that also during development of the adult epidermis Drk, Sos, Ras, raf and rolled/map kinase act in a linear pathway mediating the different aspects of differentiation controlled by DER. The absence of additional phenotypes for any one of these components indicates that they are not essential for other signalling processes that occur during epidermal development. The gain-of-function allele rl Sem results in a single amino acid substitution in the catalytic domain of MAP kinase and activates the sevenless, torso and DER pathway (Brunner et al., 1994). Two strategies were followed to mark the clones genetically (Fig. 2). In the case of raf and rl, the clones were marked in cis with the mutations f 36a and pwn, respectively. In the case of Drk, Sos and Ras1, the flies were f 36a and the clones were marked by the loss of a P(f + ) insertion in the homologous chromosome. As shown in the Fig. 2, the cell markers used in the clonal analysis of Drk and raf are located between the mutant and the centromere. This permits an unambiguous identification of the mutant cell clones. In the cases of Sos, Ras1 and rl only a subset of the marked clones correspond to events of mitotic recombination proximal to the mutation analyzed. The number, frequency and size of clones in the wing and the tergites are summarized in Table 1. The number of clones analyzed in legs were 9 for Drk, 16 (7 with phenotype) for Sos, 31 (13 with phenotype) for Ras1 and 12 for raf. Minute alleles were used to obtain large clones, which facilitated the analysis of the phenotypes. Mutant clones that are Minute + have an improved viability compared to Minute cells. The developmental parameters of the Minute alleles used in this study can be found in Ferrus, Mitotic recombination was induced by X-rays from a Philips H.T. generator operating at 100 kv, 15 ma, 2 mm filter, at a dose of 1000 rad (=10 2 J. kg 1 ). The age of irradiation was dated in hours after egg laying (A.E.L.). Different parts of the fly were dissected in a mixture of ethanol/glycerol (7/1), fat was removed with KOH (10%) and after dehydration, wings, nota, legs and abdomens were mounted separately in a mixture of lactic acid/ethanol (6/5) and sealed with nail polish. The clones were examined under the compound microscope. Clones were drawn using a digitizing tablet and plotted as shown in Fig. 3. RESULTS We performed a clonal analysis in the adult epidermis of all known lethal mutations that affect the sevenless signal transduction cascade. The phenotypes of mutant clones in the wing were analyzed in greater detail since a similar analysis carried out for DER indicated that DER function is required for a number of different decision during wing development (Diaz- Benjumea and Garcia-Bellido, 1990a). The Drosophila wing is made of a dorsal and a ventral layer of epidermal cells. Each cell differentiates a small hair or trichome. The only visible MATERIALS AND METHODS The clonal analysis was carried out using the strongest available lossof-function mutations at each locus. Drk R1 is a single amino acid substitution in the SH2 domain, which abolishes the ability of Drk to bind the activated receptor (Olivier et al., 1993). Sos X122 behaves genetically as a null mutation and Sos dm7 is a partial loss-of-function mutation (Rogge et al., 1991). Ras1 e2f results in a single amino acid substitution (Simon et al., 1991). It is likely that Drk R1 and Ras1 e2f are not complete loss-of-function mutations. The raf EA75 allele is a strong loss-of-function mutation but has not been characterized molecularly (Melnick et al., 1993). The hypomorphic raf C110 allele was also analyzed in clones (data not shown). It exhibits a very weak variable phenotype similar to that seen for Sos dm7. Three different rolled (rl) alleles were used: Df(2R)rl 10a is a deficiency for rl; rl la8 and rl EMS698 are strong loss-of-function mutations (Hilliker, 1976). Fig. 1. Pattern of veins in the Drosophila wing. Camera-lucida drawing of a wild-type wing showing the dorsal and the ventral character of the different veins. Filled-in veins are dorsal and empty veins are ventral. The longitudinal veins are labelled with roman numerals. Abbreviations: Co, costal vein; cv-a, anterior cross vein; cv-p, posterior cross-veins.
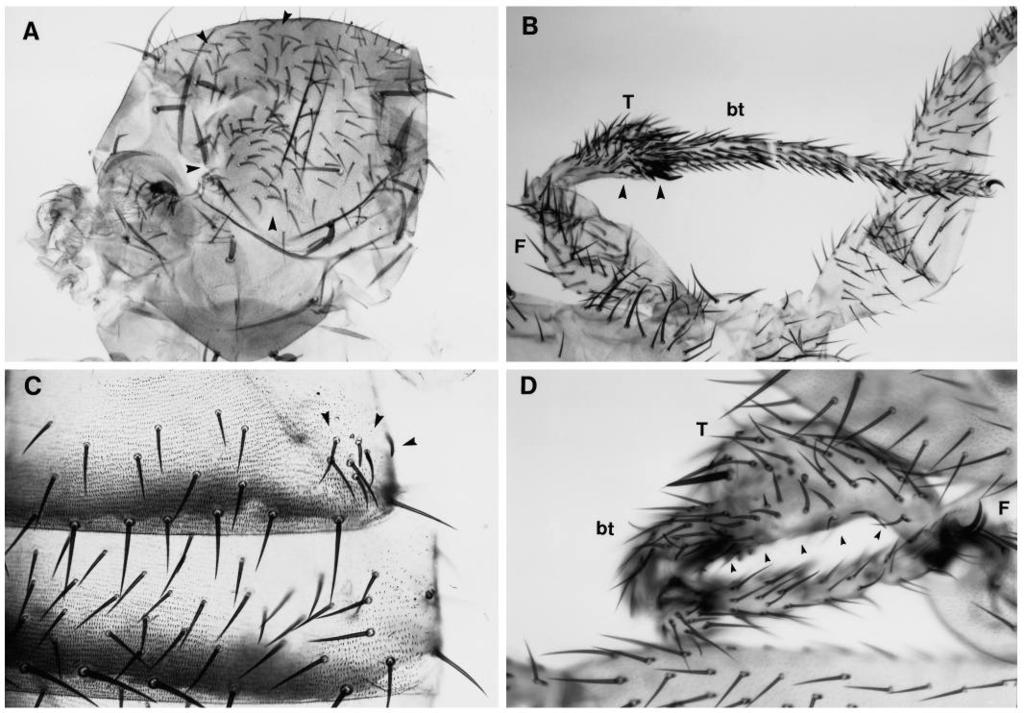
3 sevenless and epidermal development in Drosophila 571 structures on the wing blade are veins and sensory organs. The veins are longitudinal ridges running along the proximodistal axis of the wing on either surface (Fig. 1). The clones of cells homozygous for either Drk, Sos, Ras1, raf or rl exhibit a set of phenotypes that is very similar to the phenotypes observed in the clonal analysis of DER. The phenotypes can be grouped into six classes (see Fig. 7): the six classes are listed in the order of the phenotypes observed with an allelic series of DER mutations (Diaz-Benjumea and Garcia- Bellido, 1990a). The lack-of-vein phenotype (class 1) is observed with weak viable alleles of DER whereas a phenotype in the abdominal tergites (class 6) is only observed with strong loss-of-function mutations. Since the same set of six phenotypes was observed in clones mutant for any of the signalling components tested, we will describe each of these phenotypic classes separately. Class 1: absence of veins Clones of cells mutant for DER or any of the signalling components prevent the formation of veins anywhere on the wing surface. A clone of homozygous Drk cells is shown in Fig. 4A. The dorsal and the ventral component of the vein L4 is removed by the clone. This phenotype is strictly cell autonomous; mutant cells are unable to differentiate veins. Clones in either surface do not affect the differentiation of veins on the other surface suggesting that the proposed inductive mechanism for the differentiation of ventral veins is not affected (Garcia-Bellido 1977). Weak viable alleles of DER and rolled also result in the disruption of the vein L4 suggesting that already a small reduction in the function of these components prevents the differentiation of veins (Fig 6A). The formation of veins is also the only process affected in clones homozygous for the weak Sos dm7 allele. Conversely, gain-offunction mutations in DER and rl result in the formation of extra veins (Fig. 6C). Class 2: non-autonomous differentiation of ectopic veins Narrow clones adjacent to one vein can produce small pieces of extra veins surrounding the clone independently of whether the wild-type vein is removed by the clone. A clone of cells mutant for Sos X122 marked with f is shown in Fig. 4B. No vein is formed within the clone but adjacent to the mutant cells, wild-type (f + ) cells differentiate veins (arrowheads). A similar non-autonomous behaviour for the DER mutations has been observed in the developing eye. Clones of DER mutant cells in the eye imaginal disc prevent the neuronal differentiation of these cells but cause the formation of ommatidia with additional photoreceptor cells adjacent to the clone (Xu and Rubin, 1993). Fig. 2. Chromosomes used in the different clonal analysis experiments. Meiotic (top line) and cytological (bottom line) location of the mutations used in the different clonal analyses. Genotypes involving a P(forked + ) insertion carry on the X chromosome the f 36a allele. This allele permits the analysis of clones in bristles as well as in trichomes. (~) estimated location. Class 3: reduced cell size and class 4: extra bristles Cells homozygous for mutations in DER or any of the signalling components tested produce a higher cell density suggesting that the cells are smaller. As a result of the change in cell density, the wing is folded towards the surface bearing the clone and the dorsal and ventral components of the veins are uncoupled. That is most evident in vein L4, where the vein ridge is dorsal in the distal part of the wing and ventral in the proximal part. An example of a ventral Sos X122 clone producing a fold (arrowheads) in the overlaying dorsal side is shown in Fig. 4C. The difference in cell density is most evident in the Ras e2f clone shown in Fig. 4D. Flies homozygous or hemizygous for the viable rl 1 allele also possess bent down wings (Morgan et al., 1925). The reduced function of rolled/map kinase in rl 1 mutants appears to affect cell size stronger on the ventral than on the dorsal side.
4 572 F. J. Diaz-Benjumea and E. Hafen Table 1. Number, frequency and size of the clones obtained in the wings and the abdominal tergites in the different experiments Wing Abdomen Number of Cells per Number of Clones per Expt. Genotype Age (h.ael) Clones (*) Clone ( ) clones (*) Abdomen A f 36a ; Drk / f +44A M(2R)l 2 63+/ (38) (1) 0.26 B f 36a ; Sos X122 / M(2L)Z f +30B 63+/ (61) (420) 24 (0) 0.34 C f 36a ; Sos dm7 / M(2L)Z f +30B 63+/ (4) (2.030) 16 (0) 0,41 D f 36a ; Ras1 e2f / f +87D M(3R)w / (52) (8) 0,15 E Raf EA75 f 36a / M(1)o sp 42+/ (76) (0) 0.83 F f 36a ; rl 10a / f +44A M(2R)l 2 63+/ (0) G rl EMS698 pwn / M(2R)S7 63+/ (15) (330) 130 (2) 0,53 H f 36a ; Sem / f +44A M(2R)l 2 63+/ (0) (0) 0,50 *Number of clones that show a phenotype (in parenthesis). Note that in the wing this is 100% when the cell marker used is more proximal on the chromosome than the mutation analysed in each case (see Fig. 2). and <100% when the cell marker is distal to the mutation. This suggests that the penetrance of the phenotype is 100% and the clones without a phenotype correspond to the mitotic recombination events between the cell marker and the mutation analysed in each case. Average number of cells per clone in clones that show a phenotype (in parenthesis). Note the reduction in the size of the clones mutant for the strong Sos, raf and rolled alleles used in the experiments B, E, and G in comparison with the size of wild-type clones (around 2,000 cells) induced at the same time. A higher cell density is also observed in clones in the notum, the legs and the head. Examples are shown in Fig. 5A for Ras1 and in Fig. 5B for Sos. In addition to the higher cell density of epidermal cells, the number of bristles is also increased in the clone compared to the regions of wild-type tissue surrounding the clone (arrowheads in Fig. 5A and Diaz-Benjumea and Garcia-Bellido, 1990a). Class 5: cell viability Strong loss-of-function alleles are cell lethal in clones in imaginal discs. However, using the Minute + technique, it was possible to recover very small clones. These clones are very narrow, only two or three cells wide and are recovered at low frequency. This observation suggests that the absence of mutant clones under normal circumstances is caused by the competition with faster growing heterozygous cells. In the case of rolled/map kinase for which complete loss-of-function mutations are available, it was not possible to detect any clones suggesting that the complete removal of MAP kinase function is lethal for the cell. Similarly, with strong loss-of-function mutations in DER, it is also only possible to recover very small clones. Due to the strongly reduced viability of rolled clones, it was not possible to cover the entire wing surface (Fig. 2). To further establish the similarity between DER and rolled phenotypes, we used as a complementary approach an analysis of genetic interactions between rolled lack- and gain-of-function alleles and other mutations affecting the wing vein pattern. The expectation was that both mutations, rolled and DER, show interactions with the same wing vein mutants. As shown in Fig. 6, genetic combinations of either lack-of-function or gain-offunction alleles of rolled and DER show a strong enhancement of the wing phenotype. The results of this analysis are summarized in Table 2. Essentially the same mutations that showed an interaction with DER (Diaz-Benjumea and Garcia-Bellido, 1990a), also interact with rolled. The results from the clonal analysis and the genetic interactions indicate that DER and rolled/map kinase function in the same pathway. Class 6: abdominal tergites Strong loss-of-function alleles of DER and rolled produce small narrow clones in the abdominal tergites. In the Ras e2f analysis, we have found a similar phenotype but with a low penetrance. An example of a Ras e2f clone is shown in Fig. 5C. Interestingly, wild-type bristles from the region around the clone shift their position and come to lie inside the clone. Consequently, the mutant clones contain both mutant and wild-type bristles, whereas the surrounding region contains fewer bristles. This last phenotype is not observed with the alleles of Drk, Sos and raf tested. It is possible that in these mutations there is sufficient residual activity left for the normal development of the epidermis in the tergites. DISCUSSION The clonal analysis of the signalling components Drk, Sos, Ras1, raf and rolled/map kinase during the development of the adult epidermis presented here indicates that mutations in each of these components produce the same set of six phenotypes as mutations in DER. Furthermore, for each of the genes for which multiple alleles were tested, the six phenotypes can be ordered in the same phenotypic series. This indicates that these components, initially identified by their role in mediating signalling in the developing eye, also mediate every aspects of DER function during the development of epidermal structures. In addition to the lethal mutations analyzed here, mutations in Gap1 and sina have been shown to be involved in the sevenless signalling pathway (Carthew and Rubin, 1990; Gaul et al., 1992). The existing alleles of Gap1 and sina are viable in homozygous condition and do not display phenotypes characteristic of hypomorphic mutations in DER or rolled. The gene products of Gap1 and sina may therefore act specifically in sevenless pathway. DER signalling controls two different aspects of differentiation in epidermal cells In the wing, cells homozygous for loss-of-function mutations in DER or for any of the downstream signalling components are smaller and are unable to differentiate veins. It is possible that the two phenotypes are linked, if smaller cells are unable to differentiate vein structures. There are, however, a number
me10 produces small cells similar to those produced in DER clones but it does not affect the formation of veins (Ferrus and Garcia-Bellido, 1976).")
5 sevenless and epidermal development in Drosophila 573 of other mutations that cause a reduction in cell size without preventing the formation of veins. For example the mutation l(3)me10 produces small cells similar to those produced in DER clones but it does not affect the formation of veins (Ferrus and Garcia-Bellido, 1976). Likewise there are many mutations that affect the differentiation of veins without affecting cell size. The mutation veinlet, a viable allele of rhomboid, removes the distal portions of veins without altering the cell size (Diaz- Benjumea and García-Bellido, 1990b; Sturtevant et al., 1993). Furthermore veinlet clones on the dorsal side affect in a nonautonomous manner the differentiation of ventral veins (Garcia- Bellido, 1977). This is not the case for the mutations analyzed here. None of the clones on one side of the wing affect the formation of veins on the other side (Fig. 3). It is likely then that cell size and vein differentiation are two independent characteristics acquired by cells in the wing and that DER activity, mediated via the same signalling cascade, controls these two different aspects of differentiation in each cell. 1992). In the mutations analyzed here, the non-autonomous phenotype consists of the differentiation of a normal-sized vein adjacent to the mutant clone. The differentiation of the wing margin is dependent on wingless function (Phillips and Whittle, 1993). Although many of the mutant clones tested crossed the wing margin its differentiation was not affected The sevenless signalling cassette is exclusively required for signalling from DER during epidermal development All phenotypes resulting from the reduction of the function of either Drk, Sos, Ras1, raf or rolled/map kinase resemble those caused by loss-of-function mutations of DER. This suggests that none of these components is involved in other signalling processes that can be revealed by clonal analysis during the development of the adult epidermis. The process of lateral inhibition mediated by the products of N and Dl plays a role not only in the differentiation of bristles but also in the differentiation of wing veins. In contrast to the phenotypes detected in the clonal analysis for mutations in the TK pathway, clones of cells mutant for loss-offunction alleles of Dl and N produce thicker veins. In the case of Dl, the proposed ligand, this phenotype is non-autonomous and a row of cells surrounding the clone also differentiates as vein tissue (Garcia-Bellido and de Celis, Fig. 3. Regions of the wing covered in the different clonal analyses. Shaded areas represent areas that were covered by mutant clones. Veins whose differentiation is prevented in the different clones are not drawn. In each case analyzed, the mutant clone removed the veins in the surface of the wing where the clone was located but did not affect the vein on the opposite surface.
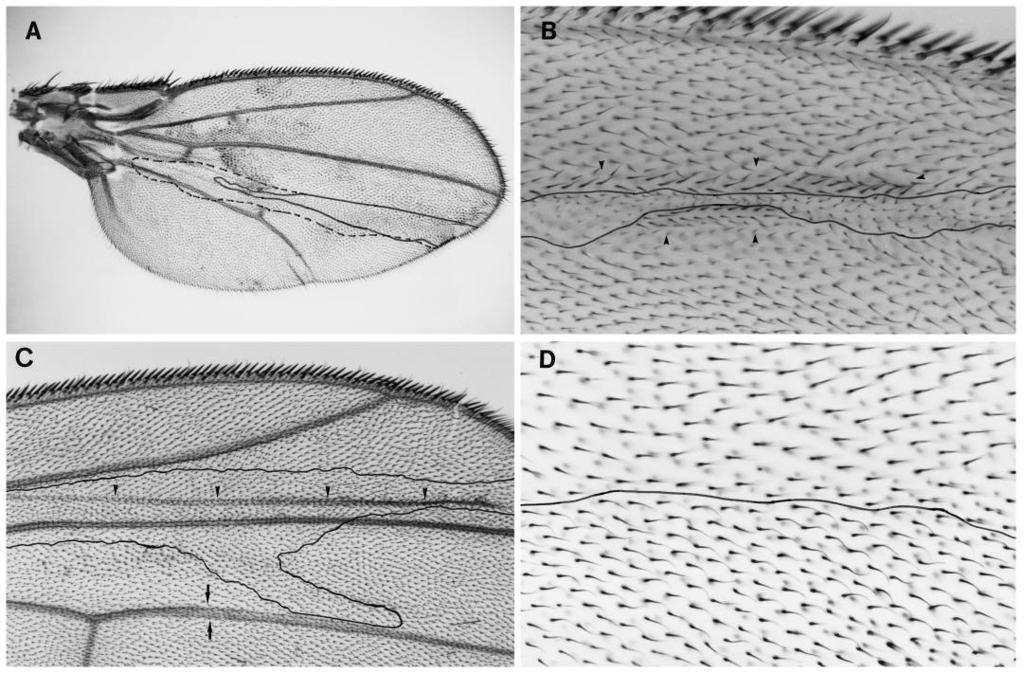
6 574 F. J. Diaz-Benjumea and E. Hafen 4 5
 and ventral surfaces (dashed line) of the posterior compartment.")
7 sevenless and epidermal development in Drosophila 575 Fig. 4. Clonal phenotypes in the wing. (A) f Drk (Minute + ) clone in the wing. The clone extends as a stripe of cells in both the dorsal (solid line) and ventral surfaces (dashed line) of the posterior compartment. Both the dorsal and the ventral component of the vein L4 is removed. (B) Ectopic veins produced by a f Sos X122 clone. The f trichomes occupy a narrow stripe which splits the presumptive vein region and induces the differentiation of two veins (arrowheads), which have the same width as the wild-type veins. Note that both veins are formed by wild-type cells adjacent to the clone. (C) f Sos X122 (Minute + ) clone in the ventral surface of the wing (outline) produces a fold in the dorsal surface (arrowhead). As a result of this clone, the wing is folded ventrally. Note that the dorsal and ventral components of the vein L4 (arrows), which overlap in the wild-type wing, are shifted as a result of the change in cell density in the ventral surface. The viable allele rolled 1 shows a similar phenotype. (D) High magnification of a Ras1 e2f f clone. The clone covers the bottom part of the figure as a transverse stripe. The black line outlines the boundary between the clone and the wild-type tissue. Note the higher density of f trichomes compared with the wild-type trichomes. Fig. 5. Clonal phenotypes in the notum, leg and tergites. (A) Ras1 e2f f clone in the left hemi-notum. Note the higher density of f bristles (arrowheads) as compared with the wild-type bristles in the right half of the notum. (B) Low magnification of a f Sos X122 (Minute + ) clone in the tibia (arrow). Note the reduction in size. (C) Ras1 e2f f clone in an abdominal tergite. Note the higher density of f and wild-type bristles (arrowheads) within the clone and the region devoid of bristles surrounding the clone. (D) High magnification of a f Sos X122 (Minute + ) clone (arrowhead) covering part of the femur (F), the tibia (T) and the basitarsus (bt) of the second leg. Note the deformation and the reduction in size of the segments affected by the clone in comparison with the normal tarsal segments. (Figs 3, 4C). Clones of dpp cells prevent the formation of wing veins and affect the development of the imaginal disc cells in a non-autonomous fashion when the mutant cells occupy a region anterior to the AP compartment boundary (Posakony et al., 1990). It is unlikely that the lack of vein phenotype and the reduced viability observed for mutations analyzed here are caused by blocking the dpp signalling pathway. dpp mutant clones do not produce extra veins outside the clone as it is observed for all mutations analyzed here. Furthermore, the allelic series of dpp mutations and hypomorphic mutations in DER and rolled exhibit a different set of phenotypes (Segal and Gelbart, 1985; Clifford and Schüpbach, 1989). It appears therefore that none of the signalling components that are common to different receptor tyrosine kinases are required in the as yet unknown signalling pathways controlled by wingless and dpp. It has been proposed that the different steps in the signal transduction cascade serve to integrate different intracellular signals. Studies in vertebrate cell culture using dominant negative forms of p21-ras suggest that Ras functions not only downstream of receptor tyrosine kinases but also downstream of src-related kinases (Kremer et al., 1991) as well as protein kinase C (Mulcahy et al., 1985). Furthermore, MAP kinase can be activated by a Ras-dependent and a Ras-independent pathway. In vertebrates, the dual specificity kinase Mek (MAPK or ERK kinase) activates MAP kinase by phosphorylation on both tyrosine and threonine. Mek can be activated by raf or by Mek kinase. Mek kinase, similar to its yeast homolog STE11, mediates signals from heterotrimeric G protein coupled receptors (Crews and Erikson, 1993). Therefore Mek, like Ras, appears to act as an integration point of signals from Fig. 6. Genetic interaction between rolled and alleles of DER. (A) Wings of flies with the following genotypes are shown: rl 1 /rl 10a (A), rl 1 DER top1 /rl 10a DER top4a (B), Elp B1 /+ (C), and Elp B1 /rl Sem (D). Wings of flies hemizygous for rl 1 are slightly reduced in size and exhibit a disruption of vein L4 (A). The phenotype of top 1 hemizygous flies is very similar (not shown). The lack of veins phenotype and size reduction is strongly enhanced in the rl top double mutant (B). Similarly, the formation of additional veins in the gain-of-function mutation Elp B1 (C) and rl Sem (not shown) is strongly enhanced in the Elp B1 /rl Sem double mutant (D). All the pictures were made at the same magnification.
8 576 F. J. Diaz-Benjumea and E. Hafen Table 2. Genetic interactions between rolled alleles and other mutations affecting the wing vein morphogenesis Allele Locus rl 1 /rl 10a Sem / + top 1 /top 4A (2-100) Elp B1 (Elipse) (2-100) * ras-2 (1- ) ast (asteroid) (2-3.8) ve (veinlet) (3-0.2) vn (vein) (3-16.2) ci (cubitus interruptus) (4-0) Vno /+ (Vein off) (3-Rear.) ri (radius incompletus) (3-47.0) + tt (tilt) (3-40.0) vvl M3 /In sep (ventral veinless) (3-Rear.) net (net) (2-0.0) * px (plexus) ( ) * Dl M1 /+ (Delta) (3-66.2) + N 55E11 /+ (Notch) (1-3.0) Ax /+ (Abruptex) (1-3.0) The results of these interactions with DER alleles have been described previously (Diaz-Benjumea and Garcia-Bellido, 1990a)., strong enhancement;, suppression; +, additive; *, studied in heterozygous condition;, Drosophila activated ras-2 expressed under the control of the actin promoter (Bishop III and Corces, 1988). The phenotypes of viable combinations of different rl alleles can be ordered according their strength as follows: Phenotypic series of different viable allelic combinations of rl: rl 10a /rl 1 rl EMS64 /rl 1 >rl la8 /rl 1 >rl EMS698 /rl 1 >rl 1 /rl 1. different signalling pathways. In light of this high degree of integration of different signals observed in vertebrate cell culture systems, it is surprising that in our studies the mutant phenotypes for the different signalling components are so similar. The fact that mutations in Drk, coding for an SH2 adaptor protein that is likely to be involved primarily in signalling from receptor tyrosine kinases and mutations in Ras1 and rolled/map kinase produce similar phenotypes suggests that during epidermal development, Ras1 and MAP kinase are not responding to a larger variety of signals than Drk. The results presented here are consistent with the finding that also during the development of the compound eye clones of Drk and Ras1 produce identical phenotypes (Simon et al., 1991). Therefore during the development of neither the compound eye nor the adult epidermis is there functional evidence that this signalling cascade may serve to integrate different intracellular signals. Different cellular responses to a common signalling cassette The components analyzed here have been shown to mediate signalling from sevenless, torso and DER in the eye (Simon et al., 1991; Doyle and Bishop, 1993; Olivier et al., 1993). Our results indicate that even within the same cell two aspects of differentiation, cell size and vein formation, are controlled by the same receptor signalling system. In rat PC12 cells, stimulation with EGF induces cell proliferation whereas stimulation with NGF induces neural differentiation. These different responses are mediated by a very similar signalling cascade including Ras, raf and MAP kinase (Chao, 1992). It is possible that the strength of the signal or its duration triggers the qualitatively different responses. Consistent with this hypothesis is the finding that the activation profile of MAP kinase upon EGF and NGF stimulation differs significantly. Whereas MAP Fig. 7. Summary of the classes of phenotypes observed. The phenotypes are ordered according to the strength of the allele required to produce it. Lack of veins (1) is seen with very weak hypomorphic alleles whereas the recruitment of wild-type bristles into the clone in tergites is only observed with strong loss-offunction alleles. Filled dots indicate that the phenotype has been observed. Open circles indicate that the phenotype was not seen in every clone analyzed. (+) The phenotype has not been observed with the alleles tested. (?) The results were ambiguous. kinase activity increases within 2 minutes of stimulation with either growth factor, the activity decreases very rapidly upon EGF stimulation but is sustained for minutes after NGF stimulation (Traverse et al., 1992). In the wing, weak alleles of DER and Sos affect only the differentiation of veins but do not reduce cell size. This suggests that these two cellular responses are also dependent on different levels of DER signalling. It is clear, however, that simple quantitative differences in the amount of signalling are not sufficient to explain the variety of phenotypes observed in mutations of the different receptor tyrosine kinases. The gain-of-function mutation in rolled, rl Sem, mimics the gain-of-function phenotypes of torso in the
9 sevenless and epidermal development in Drosophila 577 embryo, sevenless in the eye and DER in the wing (Brunner et al., 1994). This suggests that the different interpretation of MAP kinase activity is likely to occur downstream of MAP kinase. In vertebrates, MAP kinase has been shown to activate transcription factors such as Elk-1, a component of the serum response factor complex (Marais et al., 1993). In the developing eye, the sina gene encodes a nuclear protein required for R7 development that is specifically expressed in the cells competent to become R7 cells (Carthew and Rubin, 1990). Although this factor may not be a direct target of the sevenless signalling cascade, it appears to be essential for the correct interpretation of the signal in these cells. Genetic screens to identify components that act downstream of rolled/map kinase should help to identify mutations with phenotypes corresponding to only a subset of those observed for mutations used in the present study and thereby help to identify genes involved in the tissue- and stage-specific interpretation of a common signal. We would like to thank M. Simon for providing Ras1 and Sos alleles, N. Perrimon for raf EA75, A. Hilliker for the rolled alleles, V. Corces for the P(forked + ) insertions and A. Garcia-Bellido for the stocks of P(f + ) insertions with Minutes. We thank K. Basler, S. Cohen, B. Dickson, M. Dominguez and M. Mlodzik for comments on the manuscript. F. D-B. was supported by an EMBO short-term fellowship. The work was supported by a grant of the Swiss National Science Foundation to E. H. REFERENCES Ambrosio, L., Mahowald, A. P. and Perrimon, N. (1989). Requirement of the Drosophila raf homologue for torso function. Nature 342, Aroian, R. V., Koga, M., Mendel, J. E., Ohshima, Y. and Sternberg, P. W. (1990). The let-23 gene necessary for Caenorhabditis elegans vulval induction encodes a tyrosine kinase of the EGF receptor subfamily. Nature 348, Baker, N. E. and Rubin, G. M. (1989). Effect on eye development of dominant mutations in Drosophila homologue of the EGF receptor. Nature 340, Baker, N. E. and Rubin, G. M. (1992). Ellipse mutations in the Drosophila homologue of the EGF receptor affect pattern formation, cell division, and cell death in eye imaginal discs. Dev. Biol. 150, Banerjee, U., Renfranz, P. J., Pollock, J. A. and Benzer, S. (1987). Molecular characterization and expression of sevenless, a gene involved in neuronal pattern formation in the Drosophila eye. Cell 49, Basler, K. and Hafen, E. (1988). Control of photoreceptor cell fate by the sevenless protein requires a functional tyrosine kinase domain. Cell 54, Beitel, G. J., Clark, S. G. and Horvitz, R. (1990). Caenorhabditis elegans ras gene let-60 acts as a switch in the pathway of vulval induction. Nature 348, Bishop III, J. G. and Corces, V. (1988). Expression of an activated ras gene causes developmental abnormalities in transgenic Drosophila melanogaster. Genes Dev. 2, Bowtell, D., Simon, M. A. and Rubin, G. M. (1988). Nucleotide sequence and structure of the sevenless gene of Drosophila melanogaster. Genes Dev. 2, Brunner, D., Oellers, N., Szabad, J., Biggs III, W. H., Zipursky, S. L. and Hafen, E. (1994). A gain of function mutation in Drosophila MAP kinase activates multiple receptor tyrosine kinase signalling pathways. Cell in press. Carthew, R. W. and Rubin, G. M. (1990). seven in absentia, a gene required for specification of R7 cell fate in the Drosophila eye. Cell 63, Casanova, J. and Struhl, G. (1989). Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern in Drosophila. Genes Dev. 3, Chabot, B., Stephenson, D. A., Chapman, V. M., Besmer, P. and Bernstein, A. (1988). The proto-oncogene c-kit encoding a transmembrane tyrosine kinase receptor maps to the mouse W locus. Nature 335, Chao, M. V. (1992). Growth factor signaling: Where is the specificity? Cell 68, Clark, S. G., Stern, M. J. and Horvitz, H. R. (1992). C. elegans cellsignalling gene sem-5 encodes a protein with SH2 and SH3 domains. Nature 356, Clifford, R. and Schüpbach, T. (1992). The torpedo (DER) receptor tyrosine kinase is required at multiple times during Drosophila embryogenesis. Development 115, Clifford, R. J. and Schüpbach, T. (1989). Coordinately and differentially mutable activities of torpedo, the Drosophila melanogaster homolog of the vertebrate EGF receptor gene. Genetics 123, Crews, C. M. and Erikson, R. L. (1993). Extracellular signals and reversible protein phosphorylation: What to Mek of it all. Cell 74, Diaz-Benjumea, F. J. and Garcia-Bellido, A. (1990a). Behaviour of cells mutant for an EGF receptor homologue of Drosophila in genetic mosaics. Proc. R. Soc. Lond. B 242, Diaz-Benjumea, F. J. and García-Bellido, A. (1990b). Genetic analysis of the wing vein pattern of Drosophila. Roux s Arch. Dev. Biol. 198, Dickson, B., Sprenger, F., Morrison, D. and Hafen, E. (1992). Raf functions downstream of Ras1 in the Sevenless signal transduction pathway. Nature 360, Doyle, H. J. and Bishop, J. M. (1993). Torso, a receptor tyrosine kinase required for embryonic pattern formation, shares substrates with the Sevenless and EGF-R pathways in Drosophila. Genes Dev. 7, Ferrus, A. (1974). Parameters of mitotic recombination of Minute mutants of Drosophila melanogaster. Genetics 79, Ferrus, A. and Garcia-Bellido, A. (1976). Morphogenetic mutants detected in mitotic recombination clones. Nature 260, Fortini, M. E., Simon, M. A. and Rubin, G. M. (1992). Signalling by the sevenless protein tyrosine kinase is mimicked by Ras1 activation. Nature 355, Garcia-Bellido, A. (1977). Inductive mechanisms in the process of wing vein formation in Drosophila. Roux s Arch. Dev. Biol. 182, Garcia-Bellido, A. and de Celis, J. F. (1992). Developmental genetics of the venation pattern of Drosophila. Annu Rev Genet 26, Gaul, U., Mardon, G. and Rubin, G. M. (1992). A putative Ras GTPase activating protein acts as a negative regulator of signaling by the sevenless receptor tyrosine kinase. Cell 68, Geissler, E. N., Ryan, M. A. and Housman, D. E. (1988). The dominant white spotting (W) locus of the mouse encodes the c-kit proto-oncogene. Cell 55, Hafen, E., Basler, K., Edstroem, J. E. and Rubin, G. M. (1987). sevenless, a cell-specific homeotic gene of Drosophila, encodes a putative transmembrane receptor with a tyrosine kinase domain. Science 236, Han, M., Golden, A., Han, Y. and Sternberg, P. W. (1993). C. elegans lin-45 raf gene participates in let-60 ras-stimulated vulval differentiation. Nature 363, Han, M. and Sternberg, P. W. (1990). let-60, a gene that specifies cell fates during C. elegans vulval induction, encodes a ras protein. Cell 63, Hilliker, A. J. (1976). Genetic analysis of the centromeric heterochromatin of chromosome 2 of Drosophila melanogaster: deficiency mapping of EMSinduced lethal complementation groups. Genetics 83, Howe, L. R., Leevers, S. J., Gómez, N., Nakielny, S., Cohen, P. and Marshall, C. J. (1992). Activation of the MAP kinase pathway by the protein kinase raf. Cell 71, Klingler, M., Erdélyi, M., Szabad, J. and Nüsslein-Volhard, C. (1988). Function of torso in determining the terminal anlagen of the Drosophila embryo. Nature 335, Kremer, N. E., D, A. G., Thomas, S. M., DeMarco, M., Brugge, J. S. and Halegoua, S. (1991). Signal transduction by nerve growth factor and fibroblast growth factor in PC12 cells requires a sequence of src and ras actions. J. Cell Biol. 115, Lu, X., Chou, T. B., Williams, N. G., Roberts, T. and Perrimon, N. (1993). Control of cell fate determination by p21ras/ras1, an essential component of torso signaling in Drosophila. Genes Dev. 7, Marais, R., Wynne, J. and Treisman, R. (1993). The SRF accessory protein Elk-1 contains a growth factor-regulated transcriptional activation domain. Cell 73, Melnick, M. B., Perkins, L. A., Lee, M., Ambrosio, L. and Perrimon, N. (1993). Developmental and molecular characterization of mutations in the Drosophila-raf serine/threonine protein kinase. Development 118, Morgan, T. H., Bridges, C. B. and Sturtevant, A. H. (1925). The genetics of Drosophila. Biobliog Genet 2, 233. Mulcahy, L. S., Smith, M. R. and Stacey, D. W. (1985). Requirement for ras
10 578 F. J. Diaz-Benjumea and E. Hafen proto-oncogene function during serum-stimulated growth of NIH3T3 cells. Nature 313, Olivier, J. P., Raabe, T., Henkemeyer, M., Dickson, B., Mbamalu, G., Margolis, B., Schlessinger, J., Hafen, E. and Pawson, T. (1993). A Drosophila SH2-SH3 adaptor protein implicated in coupling the Sevenless tyrosine kinase to an activator of Ras guanine nucleotide exchange, Sos. Cell 73, Phillips, R. G. and Whittle, J. R. S. (1993). wingless expression mediates determination of peripheral nervous system elements in late stages of Drosophila wing disc development. Development 118, Posakony, L. G., Raftery, L. A. and Gelbart, W. M. (1990). Wing formation in Drosophila melanogaster requires decapentaplegic gene function along the anterior-posterior compartment boundary. Mech. Dev. 33, Price, J. V., Clifford, R. J. and Schüpbach, T. (1989). The maternal ventralizing locus torpedo is allelic to faint little ball, an embryonic lethal, and encodes the Drosophila EGF receptor homolog. Cell 56, Raz, E. and Shilo, B.-Z. (1993). Establishment of ventral cell fates in the Drosophila embryonic extoderm requries the EGF receptor homolog (DER). Genes Dev. in press. Rogge, R. D., Karlovich, C. A. and Banerjee, U. (1991). Genetic dissection of a neurodevelopmental pathway: Son of sevenless functions downstream of the sevenless and EGF receptor tyrosine kinases. Cell 64, Schejter, E. D. and Shilo, B. Z. (1989). The Drosophila EGF receptor homolog (DER) gene is allelic to faint little ball, a locus essential for embryonic development. Cell 56, Schüpbach, T. (1987). Germ line and soma cooperate during oogenesis to establish the dorsoventral pattern of egg shell and embryo in Drosophila melanogaster. Cell 49, Segal, D. and Gelbart, W. M. (1985). shortvein, a new component of the decapentaplegic gene complex in Drosophila melanogaster. Genetics 109, Simon, M. A., Bowtell, D., Dodson, G. S., Laverty, T. R. and Rubin, G. M. (1991). Ras1 and a putative guanine nucleotide exchange factor perform crucial steps in signaling by the sevenless protein tyrosine kinase. Cell 67, Simon, M. A., Dodson, G. S. and Rubin, G. M. (1993). An SH3-SH2-SH3 protein is required for p21ras1 activation and binds to sevenless and Sos proteins in vitro. Cell 73, Sprenger, F., Stevens, L. M. and Nüsslein-Volhard, C. (1989). The Drosophila gene torso encodes a putative receptor tyrosine kinase. Nature 338, Sturtevant, M. A., Roark, M. and Bier, E. (1993). The Drosophila rhomboid gene mediates the localized formation of wing veins and interacts genetically with components of the EGF-R signaling pathway. Genes Dev. 7, Thomas, S. M., DeMarco, M., D Arcangelo, G., Halegoua, S. and Brugge, J. S. (1992). Ras is essential for nerve growth factor- and phorbol esterinduced tyrosine phosphorylation of MAP kinases. Cell 68, Traverse, S., Gomez, N., Paterson, H., Marshall, C. and Cohen, P. (1992). Sustained activation of the mitogen-activated protein (MAP) kinase cascade may be required for differentiation of PC12 cells. Comparison of the effects of nerve growth factor and epidermal growth factor. Biochem. J. 288, Wood, K. W., Sarnecki, C., Roberts, T. M. and Blenis, J. (1992). ras mediates nerve growth factor receptor modulation of three signaltransducing protein kinases: MAP kinase, Raf-1, and RSK. Cell 68, Xu, T. and Rubin, G. M. (1993). Analysis of genetic mosaics in developing and adult Drosophila tissues. Development 117, (Accepted 3 December 1993)
Reading: Chapter 5, pp ; Reference chapter D, pp Problem set F
 Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
Replacement of posterior by anterior structures in the Drosophila wing caused by the mutation apterous-blot
 J. Embryo!, exp. Morph. Vol. 53, pp. 291-303, 1979 291 Printed in Great Britain Company of Biologists Limited 1979 Replacement of posterior by anterior structures in the Drosophila wing caused by the mutation
J. Embryo!, exp. Morph. Vol. 53, pp. 291-303, 1979 291 Printed in Great Britain Company of Biologists Limited 1979 Replacement of posterior by anterior structures in the Drosophila wing caused by the mutation
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
ventral veinless, the gene encoding the Cf1a transcription factor, links
 Development 121, 3405-3416 (1995) Printed in Great Britain The Company of Biologists Limited 1995 3405 ventral veinless, the gene encoding the Cf1a transcription factor, links positional information and
Development 121, 3405-3416 (1995) Printed in Great Britain The Company of Biologists Limited 1995 3405 ventral veinless, the gene encoding the Cf1a transcription factor, links positional information and
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Bypass and interaction suppressors; pathway analysis
 Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Dissecting the roles of the Drosophila EGF receptor in eye development and MAP kinase activation
 Development 125, 3875-3885 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5225 3875 Dissecting the roles of the Drosophila EGF receptor in eye development and MAP kinase activation
Development 125, 3875-3885 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5225 3875 Dissecting the roles of the Drosophila EGF receptor in eye development and MAP kinase activation
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
EGFR signalling inhibits Capicua-dependent repression during specification of Drosophila wing veins
 Development 129, 993-1002 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV7917 993 EGFR signalling inhibits Capicua-dependent repression during specification of Drosophila wing
Development 129, 993-1002 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV7917 993 EGFR signalling inhibits Capicua-dependent repression during specification of Drosophila wing
The Drosophila orphan nuclear receptor Seven-up requires the Ras pathway for its function in photoreceptor determination
 Development 121, 225-235 (1995) Printed in Great Britain The Company of Biologists Limited 1995 225 The Drosophila orphan nuclear receptor Seven-up requires the Ras pathway for its function in photoreceptor
Development 121, 225-235 (1995) Printed in Great Britain The Company of Biologists Limited 1995 225 The Drosophila orphan nuclear receptor Seven-up requires the Ras pathway for its function in photoreceptor
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION
 MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
MCB 141 Midterm I Feb. 19, 2009
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Drosophila. The wingless signalling pathway and the patterning of the wing margin in. Juan Pablo Couso*, Sarah A. Bishop and Alfonso Martinez Arias
 Development 120, 621-636 (1994) Printed in Great Britain The Company of Biologists Limited 1994 621 The wingless signalling pathway and the patterning of the wing margin in Drosophila Juan Pablo Couso*,
Development 120, 621-636 (1994) Printed in Great Britain The Company of Biologists Limited 1994 621 The wingless signalling pathway and the patterning of the wing margin in Drosophila Juan Pablo Couso*,
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Analysis of the genetic hierarchy guiding wing vein development in
 Development 121, 785-801 (1995) Printed in Great Britain The Company of Biologists Limited 1995 785 Analysis of the genetic hierarchy guiding wing vein development in Drosophila Mark A. Sturtevant and
Development 121, 785-801 (1995) Printed in Great Britain The Company of Biologists Limited 1995 785 Analysis of the genetic hierarchy guiding wing vein development in Drosophila Mark A. Sturtevant and
Cytokines regulate interactions between cells of the hemapoietic system
 Cytokines regulate interactions between cells of the hemapoietic system Some well-known cytokines: Erythropoietin (Epo) G-CSF Thrombopoietin IL-2 INF thrombopoietin Abbas et al. Cellular & Molecular Immunology
Cytokines regulate interactions between cells of the hemapoietic system Some well-known cytokines: Erythropoietin (Epo) G-CSF Thrombopoietin IL-2 INF thrombopoietin Abbas et al. Cellular & Molecular Immunology
The EGF Signaling Pathway! Introduction! Introduction! Chem Lecture 10 Signal Transduction & Sensory Systems Part 3. EGF promotes cell growth
 Chem 452 - Lecture 10 Signal Transduction & Sensory Systems Part 3 Question of the Day: Who is the son of Sevenless? Introduction! Signal transduction involves the changing of a cell s metabolism or gene
Chem 452 - Lecture 10 Signal Transduction & Sensory Systems Part 3 Question of the Day: Who is the son of Sevenless? Introduction! Signal transduction involves the changing of a cell s metabolism or gene
Genetic interactions and cell behaviour in blistered mutants during proliferation and differentiation of the Drosophila wing
 Development 125, 1823-1832 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV2195 1823 Genetic interactions and cell behaviour in blistered mutants during proliferation and differentiation
Development 125, 1823-1832 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV2195 1823 Genetic interactions and cell behaviour in blistered mutants during proliferation and differentiation
10/2/2015. Chapter 4. Determination and Differentiation. Neuroanatomical Diversity
 Chapter 4 Determination and Differentiation Neuroanatomical Diversity 1 Neurochemical diversity: another important aspect of neuronal fate Neurotransmitters and their receptors Excitatory Glutamate Acetylcholine
Chapter 4 Determination and Differentiation Neuroanatomical Diversity 1 Neurochemical diversity: another important aspect of neuronal fate Neurotransmitters and their receptors Excitatory Glutamate Acetylcholine
COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION
 COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION I. Aksan 1, M. Sen 2, M. K. Araz 3, and M. L. Kurnaz 3 1 School of Biological Sciences, University of Manchester,
COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION I. Aksan 1, M. Sen 2, M. K. Araz 3, and M. L. Kurnaz 3 1 School of Biological Sciences, University of Manchester,
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Developmental Biology Lecture Outlines
 Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Cell Death & Trophic Factors II. Steven McLoon Department of Neuroscience University of Minnesota
 Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
with%dr.%van%buskirk%%%
 with%dr.%van%buskirk%%% How$to$do$well?$ Before$class:$read$the$corresponding$chapter$ Come$to$class$ready$to$par9cipate$in$Top$Hat$ Don t$miss$an$exam!!!!!!!!!!!!!!!!!!!!!!!!!!$ But$I m$not$good$with$science
with%dr.%van%buskirk%%% How$to$do$well?$ Before$class:$read$the$corresponding$chapter$ Come$to$class$ready$to$par9cipate$in$Top$Hat$ Don t$miss$an$exam!!!!!!!!!!!!!!!!!!!!!!!!!!$ But$I m$not$good$with$science
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
EGF receptor signaling induces pointed P1 transcription and inactivates Yan protein in the Drosophila embryonic ventral ectoderm
 Development 122, 3355-3362 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV8344 3355 EGF receptor signaling induces pointed P1 transcription and inactivates Yan protein in the
Development 122, 3355-3362 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV8344 3355 EGF receptor signaling induces pointed P1 transcription and inactivates Yan protein in the
TGF-β/BMP superfamily members, Gbb-60A and Dpp, cooperate to provide pattern information and establish cell identity in the Drosophila wing
 Development 125, 2723-2734 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5189 2723 TGF-β/BMP superfamily members, Gbb-60A and Dpp, cooperate to provide pattern information and
Development 125, 2723-2734 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5189 2723 TGF-β/BMP superfamily members, Gbb-60A and Dpp, cooperate to provide pattern information and
Biol403 - Receptor Serine/Threonine Kinases
 Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Drosophila wing. Temporal regulation of Apterous activity during development of the. Marco Milán and Stephen M. Cohen* SUMMARY
 Development 127, 3069-3078 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2547 3069 Temporal regulation of Apterous activity during development of the Drosophila wing Marco Milán
Development 127, 3069-3078 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2547 3069 Temporal regulation of Apterous activity during development of the Drosophila wing Marco Milán
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
Dissection of the Torso Signal Transduction Pathway in Drosophila
 MOLECULAR REPRODUCTION AND DEVELOPMENT 42:515c522 (1995) Dissection of the Torso Signal Transduction Pathway in Drosophila NORBERT PERRIMON, XIANGYI LU, XIANYU STEVEN HOU, JUI-CHOU HSU, MICHAEL B. MELNICK,
MOLECULAR REPRODUCTION AND DEVELOPMENT 42:515c522 (1995) Dissection of the Torso Signal Transduction Pathway in Drosophila NORBERT PERRIMON, XIANGYI LU, XIANYU STEVEN HOU, JUI-CHOU HSU, MICHAEL B. MELNICK,
BILD7: Problem Set. 2. What did Chargaff discover and why was this important?
 BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
Conclusions. The experimental studies presented in this thesis provide the first molecular insights
 C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
5- Semaphorin-Plexin-Neuropilin
 5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Axis determination in flies. Sem 9.3.B.5 Animal Science
 Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Bi Lecture 8 Genetic Pathways and Genetic Screens
 Bi190-2013 Lecture 8 Genetic Pathways and Genetic Screens WT A 2X:2A her-1 tra-1 1X:2A her-1 tra-1 Female body Male body Female body Male body her-1(lf) B 2X:2A her-1(lf) tra-1 1X:2A her-1(lf) tra-1 Female
Bi190-2013 Lecture 8 Genetic Pathways and Genetic Screens WT A 2X:2A her-1 tra-1 1X:2A her-1 tra-1 Female body Male body Female body Male body her-1(lf) B 2X:2A her-1(lf) tra-1 1X:2A her-1(lf) tra-1 Female
Caenorhabditis elegans Development
 MOLECULAR REPRODUCTION AND DEVELOPMENT 42523528 (1995) LET-23-Mediated Signal Transduction During Caenorhabditis elegans Development PAUL W. STERNBERG, GIOVANNI LESA, JUNHO LEE, WENDY S. KATZ, CHARLES
MOLECULAR REPRODUCTION AND DEVELOPMENT 42523528 (1995) LET-23-Mediated Signal Transduction During Caenorhabditis elegans Development PAUL W. STERNBERG, GIOVANNI LESA, JUNHO LEE, WENDY S. KATZ, CHARLES
Drosophila Fos mediates ERK and JNK signals via distinct phosphorylation sites
 Drosophila Fos mediates ERK and JNK signals via distinct phosphorylation sites Laura Ciapponi, 1,2 David B.Jackson, 1,4 Marek Mlodzik, 1,3 and Dirk Bohmann 1,2,5 1 European Molecular Biology Laboratory,
Drosophila Fos mediates ERK and JNK signals via distinct phosphorylation sites Laura Ciapponi, 1,2 David B.Jackson, 1,4 Marek Mlodzik, 1,3 and Dirk Bohmann 1,2,5 1 European Molecular Biology Laboratory,
Morphogens in biological development: Drosophila example
 LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Homeotic genes in flies. Sem 9.3.B.6 Animal Science
 Homeotic genes in flies Sem 9.3.B.6 Animal Science So far We have seen that identities of each segment is determined by various regulators of segment polarity genes In arthopods, and in flies, each segment
Homeotic genes in flies Sem 9.3.B.6 Animal Science So far We have seen that identities of each segment is determined by various regulators of segment polarity genes In arthopods, and in flies, each segment
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
BIS &003 Answers to Assigned Problems May 23, Week /18.6 How would you distinguish between an enhancer and a promoter?
 Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Richik N. Ghosh, Linnette Grove, and Oleg Lapets ASSAY and Drug Development Technologies 2004, 2:
 1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
b. The maximum binding will decrease.
 Cell Signaling Receptors are a. proteins that change conformation upon interaction with a stimulus b. genes that change expression in response to a stimulus c. phosphorylation cascades that control cellular
Cell Signaling Receptors are a. proteins that change conformation upon interaction with a stimulus b. genes that change expression in response to a stimulus c. phosphorylation cascades that control cellular
Chem Lecture 10 Signal Transduction
 Chem 452 - Lecture 10 Signal Transduction 111202 Here we look at the movement of a signal from the outside of a cell to its inside, where it elicits changes within the cell. These changes are usually mediated
Chem 452 - Lecture 10 Signal Transduction 111202 Here we look at the movement of a signal from the outside of a cell to its inside, where it elicits changes within the cell. These changes are usually mediated
Genes controlling segmental specification in the Drosophila thorax (homeosis/insect segments/determination/clonal analysis)
") Proc. NatL Acad. Sci. USA Vol. 79, pp. 7380-7384, December 1982 Developmental Biology Genes controlling segmental specification in the Drosophila thorax (homeosis/insect segments/determination/clonal analysis)
Proc. NatL Acad. Sci. USA Vol. 79, pp. 7380-7384, December 1982 Developmental Biology Genes controlling segmental specification in the Drosophila thorax (homeosis/insect segments/determination/clonal analysis)
1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms.
 Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
ADAM FAMILY. ephrin A INTERAZIONE. Eph ADESIONE? PROTEOLISI ENDOCITOSI B A RISULTATO REPULSIONE. reverse. forward
 ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos by the polyhomeotic gene
 Development 103, 733-741 (1988) Printed in Great Britain The Company of Biologists Limited 1988 733 Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos
Development 103, 733-741 (1988) Printed in Great Britain The Company of Biologists Limited 1988 733 Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos
Drosophila receptor tyrosine kinase signaling
 Development 129, 71-82 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV5966 71 MASK, a large ankyrin repeat and KH domain-containing protein involved in Drosophila receptor tyrosine
Development 129, 71-82 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV5966 71 MASK, a large ankyrin repeat and KH domain-containing protein involved in Drosophila receptor tyrosine
blistered: a gene required for vein/intervein formation in wings of Drosophila
 Development 120, 2661-2671 (1994) Printed in Great Britain The Company of Biologists Limited 1994 2661 blistered: a gene required for vein/intervein formation in wings of Drosophila Dianne Fristrom 1,
Development 120, 2661-2671 (1994) Printed in Great Britain The Company of Biologists Limited 1994 2661 blistered: a gene required for vein/intervein formation in wings of Drosophila Dianne Fristrom 1,
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290
 PLoS Biol 4(9): e290") Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Cell fate specification by Ras-mediated cell signalling in C. elegans. Teresa Tiensuu
 Cell fate specification by Ras-mediated cell signalling in C. elegans Teresa Tiensuu Umeå Centre for Molecular Pathogenesis Umeå University Umeå Sweden 2003 Copyright Teresa Tiensuu 2003 ISBN 91-7305-525-5
Cell fate specification by Ras-mediated cell signalling in C. elegans Teresa Tiensuu Umeå Centre for Molecular Pathogenesis Umeå University Umeå Sweden 2003 Copyright Teresa Tiensuu 2003 ISBN 91-7305-525-5
Activation of a receptor. Assembly of the complex
 Activation of a receptor ligand inactive, monomeric active, dimeric When activated by growth factor binding, the growth factor receptor tyrosine kinase phosphorylates the neighboring receptor. Assembly
Activation of a receptor ligand inactive, monomeric active, dimeric When activated by growth factor binding, the growth factor receptor tyrosine kinase phosphorylates the neighboring receptor. Assembly
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8
 Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
Wingless transduction by the Frizzled and Frizzled2 proteins of Drosophila
 Development 126, 5441-5452 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV7755 5441 Wingless transduction by the Frizzled and Frizzled2 proteins of Drosophila Chiann-mun Chen
Development 126, 5441-5452 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV7755 5441 Wingless transduction by the Frizzled and Frizzled2 proteins of Drosophila Chiann-mun Chen
Genes, Development, and Evolution
 14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila
 Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila Jordi Casanova and Gary Struhl Howard Hughes Medical Institute, Center for Neurobiology and
Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila Jordi Casanova and Gary Struhl Howard Hughes Medical Institute, Center for Neurobiology and
Chapter 18 Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis
 18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Bio 3411, Fall 2006, Lecture 19-Cell Death.
 Types of Cell Death Questions : Apoptosis (Programmed Cell Death) : Cell-Autonomous Stereotypic Rapid Clean (dead cells eaten) Necrosis : Not Self-Initiated Not Stereotypic Can Be Slow Messy (injury can
Types of Cell Death Questions : Apoptosis (Programmed Cell Death) : Cell-Autonomous Stereotypic Rapid Clean (dead cells eaten) Necrosis : Not Self-Initiated Not Stereotypic Can Be Slow Messy (injury can
Supplemental table S7.
 Supplemental table S7. GO terms significantly enriched in significantly up-regulated genes of the microarray. K: number of genes from the input cluster in the given category. F: number of total genes in
Supplemental table S7. GO terms significantly enriched in significantly up-regulated genes of the microarray. K: number of genes from the input cluster in the given category. F: number of total genes in
A complementation test would be done by crossing the haploid strains and scoring the phenotype in the diploids.
 Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Drosophila Somatic Anterior-Posterior Axis (A-P Axis) Formation
 Formation") Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics
Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
PRACTICE EXAM. 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos.
 PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
Glia in the fly wing are clonally related to epithelial cells and use the nerve as a pathway for migration
 Development 120, 523-534 (1994) Printed in Great Britain The Company of Biologists Limited 1994 523 Glia in the fly wing are clonally related to epithelial cells and use the nerve as a pathway for migration
Development 120, 523-534 (1994) Printed in Great Britain The Company of Biologists Limited 1994 523 Glia in the fly wing are clonally related to epithelial cells and use the nerve as a pathway for migration
Phenotypic Plasticity in Drosophila Pigmentation Caused by Temperature Sensitivity of a Chromatin Regulator Network
 Phenotypic Plasticity in Drosophila Pigmentation Caused by Temperature Sensitivity of a Chromatin Regulator Network Jean-Michel Gibert 1*, Frédérique Peronnet 2, Christian Schlötterer 1,3 1 Institut für
Phenotypic Plasticity in Drosophila Pigmentation Caused by Temperature Sensitivity of a Chromatin Regulator Network Jean-Michel Gibert 1*, Frédérique Peronnet 2, Christian Schlötterer 1,3 1 Institut für
Mosaic analyses reveal the function of Drosophila Ras in embryonic dorsoventral patterning and dorsal follicle cell morphogenesis
 Development 129, 2209-2222 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV3618 2209 Mosaic analyses reveal the function of Drosophila Ras in embryonic dorsoventral patterning
Development 129, 2209-2222 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV3618 2209 Mosaic analyses reveal the function of Drosophila Ras in embryonic dorsoventral patterning
Functions and Mechanisms of Receptor Tyrosine Kinase Torso Signaling: Lessons From Drosophila Embryonic Terminal Development
 DEVELOPMENTAL DYNAMICS 232:656 672, 2005 REVIEWS A PEER REVIEWED FORUM Functions and Mechanisms of Receptor Tyrosine Kinase Torso Signaling: Lessons From Drosophila Embryonic Terminal Development Willis
DEVELOPMENTAL DYNAMICS 232:656 672, 2005 REVIEWS A PEER REVIEWED FORUM Functions and Mechanisms of Receptor Tyrosine Kinase Torso Signaling: Lessons From Drosophila Embryonic Terminal Development Willis
Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain
 Development 127, 367-379 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV5342 367 Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain Wei Du
Development 127, 367-379 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV5342 367 Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain Wei Du
Mechanisms of Human Health and Disease. Developmental Biology
 Mechanisms of Human Health and Developmental Biology Joe Schultz joe.schultz@nationwidechildrens.org D6 1 Dev Bio: Mysteries How do fertilized eggs become adults? How do adults make more adults? Why and
Mechanisms of Human Health and Developmental Biology Joe Schultz joe.schultz@nationwidechildrens.org D6 1 Dev Bio: Mysteries How do fertilized eggs become adults? How do adults make more adults? Why and
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway
 Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
Solutions to Problem Set 4
 Question 1 Solutions to 7.014 Problem Set 4 Because you have not read much scientific literature, you decide to study the genetics of garden peas. You have two pure breeding pea strains. One that is tall
Question 1 Solutions to 7.014 Problem Set 4 Because you have not read much scientific literature, you decide to study the genetics of garden peas. You have two pure breeding pea strains. One that is tall
Reading. Lecture VI. Making Connections 9/17/12. Bio 3411 Lecture VI. Making Connections. Bio 3411 Monday September 17, 2012
 Lecture VI. Making Connections Bio 3411 Monday September 17, 2012!! 1! Reading NEUROSCIENCE: 5 th ed, pp!507?536! 4 th ed, pp 577-609 Bentley, D., & Caudy, M. (1983). Nature, 304(5921), 62-65. Dickson,
Lecture VI. Making Connections Bio 3411 Monday September 17, 2012!! 1! Reading NEUROSCIENCE: 5 th ed, pp!507?536! 4 th ed, pp 577-609 Bentley, D., & Caudy, M. (1983). Nature, 304(5921), 62-65. Dickson,
lecture 10. Genetic Interactions synthetic lethality high copy suppression suppression and enhancement mechanisms of suppression
 lecture 10. Genetic Interactions synthetic lethality high copy suppression suppression and enhancement mechanisms of suppression actin act1-1 act1-4 actin non- complementer anc4 anc1 anc3 anc2 actin-binding
lecture 10. Genetic Interactions synthetic lethality high copy suppression suppression and enhancement mechanisms of suppression actin act1-1 act1-4 actin non- complementer anc4 anc1 anc3 anc2 actin-binding
Caenorhabditis elegans
 Caenorhabditis elegans Why C. elegans? Sea urchins have told us much about embryogenesis. They are suited well for study in the lab; however, they do not tell us much about the genetics involved in embryogenesis.
Caenorhabditis elegans Why C. elegans? Sea urchins have told us much about embryogenesis. They are suited well for study in the lab; however, they do not tell us much about the genetics involved in embryogenesis.
GENETIC CONTROL OF CELL INTERACTIONS IN NEMATODE DEVELOPMENT
 Annu. Rev. Genet. 1991. 25. 411-36 Copyright 199l by Annual Reviews Inc. All rights reserved GENETIC CONTROL OF CELL INTERACTIONS IN NEMATODE DEVELOPMENT E. J. Lambie and Judith Kimble Laboratory of Molecular
Annu. Rev. Genet. 1991. 25. 411-36 Copyright 199l by Annual Reviews Inc. All rights reserved GENETIC CONTROL OF CELL INTERACTIONS IN NEMATODE DEVELOPMENT E. J. Lambie and Judith Kimble Laboratory of Molecular
9/4/2015 INDUCTION CHAPTER 1. Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology. Fig 1.
 INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
5/4/05 Biol 473 lecture
 5/4/05 Biol 473 lecture animals shown: anomalocaris and hallucigenia 1 The Cambrian Explosion - 550 MYA THE BIG BANG OF ANIMAL EVOLUTION Cambrian explosion was characterized by the sudden and roughly simultaneous
5/4/05 Biol 473 lecture animals shown: anomalocaris and hallucigenia 1 The Cambrian Explosion - 550 MYA THE BIG BANG OF ANIMAL EVOLUTION Cambrian explosion was characterized by the sudden and roughly simultaneous
NIH Public Access Author Manuscript Curr Opin Genet Dev. Author manuscript; available in PMC 2009 August 1.
 NIH Public Access Author Manuscript Published in final edited form as: Curr Opin Genet Dev. 2008 August ; 18(4): 342 347. doi:10.1016/j.gde.2008.06.002. Dynamics of maternal morphogen gradients in the
NIH Public Access Author Manuscript Published in final edited form as: Curr Opin Genet Dev. 2008 August ; 18(4): 342 347. doi:10.1016/j.gde.2008.06.002. Dynamics of maternal morphogen gradients in the
The function of engrailed and the specification of Drosophila wing pattern
 Development 121, 3447-3456 (1995) Printed in Great Britain The Company of Biologists Limited 1995 3447 The function of engrailed and the specification of Drosophila wing pattern Isabel Guillén 1, *, Jose
Development 121, 3447-3456 (1995) Printed in Great Britain The Company of Biologists Limited 1995 3447 The function of engrailed and the specification of Drosophila wing pattern Isabel Guillén 1, *, Jose
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018
 Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental