INVESTIGATING THE ROLE OF WNTLESS (WLS) IN WINGLESS (WG) GRADIENT FORMATION. Babak Basiri
|
|
|
- Erick Jackson
- 6 years ago
- Views:
Transcription
1 INVESTIGATING THE ROLE OF WNTLESS (WLS) IN WINGLESS (WG) GRADIENT FORMATION by Babak Basiri A thesis submitted to the Faculty of the University of Delaware in partial fulfillment of the requirements for the degree of Master of Science in Biological Sciences Spring 2012 Copyright 2012 Babak Basiri All Rights Reserved
2 INVESTIGATING THE ROLE OF WNTLESS (WLS) IN WINGLESS (WG) GRADIENT FORMATION by Babak Basiri Approved: Erica M. Selva, Ph.D. Professor in charge of thesis on behalf of the Advisory Committee Approved: Randall L. Duncan, Ph.D. Chair of the Department of Biological Sciences Approved: George H. Watson, Ph.D. Dean of the College of Arts and Sciences Approved: Charles G. Riordan, Ph.D. Vice Provost for Graduate and Professional Education
3 ACKNOWLEDGMENTS First and foremost, I would like to thank Dr. Erica Selva for being a wonderful advisor. She was always there when I needed help, and her help was not only limited to educational support, as she was also willing to offer her unconditional assistance with personal issues. I am very happy that she accepted me into her lab; I could never ask for more. My committee members, Drs. Melinda Duncan and Donna Woulfe were always a source of insight into my project. Without their help and vision, my project would have never reached its current depth. Dr. David Raden walked me through all steps of designing and performing a scientific experiment with an extraordinary patience and diligence. Rich Wittmeyer taught me how to work with fruit flies. Net Ratchasanmuang is definitely a rare gem in our lab. He takes care of everything and volunteers to help others at any time. With him being in the lab, I was never worried about stuff going wrong. I would also like to thank other Selva lab members with whom I had the opportunity to work: Jayasree Padmanabhan, Sencer Tektas, Alicia Liu, Scott Tibbetts, Erica Boetefuer, Allison McCague, Nicholas Lombardi, Rachel Randell, Patrick Shock and Karen Monono. I would like to express my gratefulness to everybody at the University of Delaware Bioimaging Center, specifically Drs. Kirk Czymmek and Jeff Caplan who helped me to perform my microscopy experiments and iii
4 analyze the data. I should also thank University of Delaware and National Science Foundation (NSF) for their professional and financial support that made it possible for me to pursue this project. Many individuals have been responsible for every success I have achieved so far and I can never thank them enough: my family, my lovely friends back home and here in Newark, my teachers and professors, my understanding roommates, my students who have always been an inspiration source for me and I am extremely thankful for having all these wonderful people in my life and wish all the best for them in every step of their lives. iv
5 TABLE OF CONTENTS LIST OF TABLES... viii LIST OF FIGURES... ix ABSTRACT... xi Chapter 1 INTRODUCTION Wnt protein family Strucrure Signal Transduction pathways Wnts act as morphogens during animal development Drosophila Wg is among the most studied Wnt proteins Drosophila wing imaginal disc is a valuable.. model system for analyzing Wg signaling and. gradient formation Activation of short- and long-range targets of. Wg in the wing disc Wg secretion and gradient formation in Drosophila wing. disc Wntless (Wls) is necessary for Wg secretion. from producing cells Two separate pools of Wg exist at the apical. and basolateral membranes of wing disc cells Scope of the thesis MATERIALS AND METHODS Fly stocks Targeted gene expression Wing disc dissection and immunostaining Adult wing phenotypes v
6 2.5 Image processing and statistical analyses RESULTS Effect of Wls overexpression on Wg short- and long-. range signaling Wls overexpression causes loss of wing margin. in the adult wing Alterations of Sens and Dll expression in wing. discs Wg relocalization from apical to basolateral.. membrane The amount and range of expression of.... extracellular Wg increased in wls-.. overexpressing tissue Alteration of Wg intracellular trafficking in response to. wls overexpression Chemical inhibition of vacuolar acidification Disrupted acidification caused Wg. retention in secreting cells wls overexpression was able to.. reduce the negative effects of. acidification inhibition Similar effects were observed following RNAi. inhibition of V-ATPase Vha68-2 RNAi expression at the D/V. boundary caused loss of wing margin. in adults Wg is absent from the basal.. membrane of secreting cells Extracellular Wg is distributed over a. narrower range as a result of V-.. ATPase inhibition in the wing disc Dll expression is noticeably reduced.. after V-ATPase inhibition vi
7 Sens expression around the D/V.. boundary is adversely affected by V-.. ATPse inhibition in the Wg-producing. cells Wls over-expression in the wing disc. did not rescue V-ATPase inhibition.. phenotype DISCUSSION wls overexpression drives Wg molecules toward long-. range gradient Inter-related functions of Wls and vacuolar acidification. in Wg secretion provides insight into Wg intracellular. trafficking Future directions REFERENCES vii
8 LIST OF TABLES Table 2.1: Drosophila stocks used to perform experiments viii
9 LIST OF FIGURES Figure 1.1: Schematic overview of the structure of mouse Wnt3A... 6 Figure 1.2: Overview of Wnt/β-catenin signaling... 8 Figure 1.3: Morphogen target gene activation Figure 1.4: Drosophila imaginal discs Figure 1.5: Wing imaginal disc of third instar Drosophila larva Figure 1.6: Wg short- and long-range target expression in wing imaginal discs Figure 1.7: Proposed models for long-range spread of Wnts Figure 1.8: Model depicting short- and long-range Wg signaling in the wing imaginal disc Figure 2.1: The UAS/GAL4 system in Drosophila Figure 3.1: Development of the Drosophila wing Figure 3.2: wls overexpression in the posterior compartment of the wing causes wing nicks Figure 3.3: Loss of Sens expression in wls-overexpressing compartment Figure 3.4: Dll expression in wls-overexpressing wing discs Figure 3.5: Wg apical to basal redistribution after wls overexpression Figure 3.6: Expansion of extracellular Wg staining in wlsoverexpressing compartment Figure 3.7: Wg retention in bafilomycin A1 treated wing discs ix
10 Figure 3.8: Wls overexpression is able to rescue the effect of V- ATPase inhibition on Wg secretion Figure 3.9: C96 expression in the wing disc Figure 3.10: Adult wings are highly disrupted when acidification is interrupted in the wing disc Figure 3.11: Basal Wg shows a significant decrease after V-ATPase inhibition Figure 3.12: Extracellular Wg reduction caused by V-ATPase inhibition Figure 3.13: Dll expression is negatively affected by V-ATPase inhibition in the Wg-producing cells Figure 3.14: Sens expression is absent around the D/V boundary in the RNAi expressing wing discs Figure 4.1: Proposed model for Wg secretion from producing cells x
11 ABSTRACT Wingless (Wg), the predominant Drosophila Wnt, is a secreted signaling ligand. Like other Wnts in vertebrates, it is required to execute numerous phases of the Drosophila developmental program. Formation of a morphogen gradient is one strategy used by Wg to drive pattern formation during embryonic development. Wntless (Wls) is a multipass transmembrane protein required for Wingless (Wg) secretion from producing cells and a conserved component of the Wnt signaling pathway. The goal of this research was to determine the role of Wls in Wg signaling and morphogen gradient formation. Observed phenotypes caused by overexpression of Wls in the wing imaginal disc of Drosophila support the hypothesis that wls overexpression will change the shape of the Wg morphogen gradient in the wing disc. In the presence of excessive Wls, more Wg is targeted toward the long-range concentration gradient at the expense of short-range signaling. As a result, long-range targets of Wg signaling would be expressed over a longer distance from the secretion source. Expression of short-range targets of Wg, on the other hand, will be attenuated which is best demonstrated by the lack of wing margin in wls-overexpressing flies. I also showed that endosomal acidification is necessary for proper Wg secretion, and wls-overexpressing cells are more resistant to the effects of acidification inhibition. This observation suggests that there is a functional xi
12 relationship between Wls and endosomal acidification in Wg intracellular trafficking and secretion. At the end of this writing, I present a model to explain this interrelation. xii
13 Chapter 1 INTRODUCTION Pattern formation is one of the major constituents of development. Through the process of pattern formation, a spatial and temporal pattern of cellular activities is organized within the embryo so that a well-ordered structure develops. A group of cells need to acquire positional information for an appropriate pattern to emerge. This positional value can be specified by various mechanisms. The simplest is based on a concentration gradient. If the concentration of a molecule decreases from one end of a group of cells to the other end, then the concentration of that molecule in any cell effectively specifies the position of the cell with respect to the boundary. Wingless (Wg) is an extremely important protein for several developmental processes in Drosophila. In wing disc tissue, Wg works with its downstream signaling components to provide positional information necessary to pattern this tissue. Wg is known to be secreted from a narrow stripe of cells at the dorsal/ventral boundary of the developing wing disc where it establishes a concentration gradient within the developing wing disc. Drosophila wing disc is composed of polarized epithelial cells and recent data suggests that a high concentration of Wg molecules is maintained on the apical membrane of secreting cells while the concentration gradient is mainly formed on the basolateral surface of the cells throughout the wing disc. Discovery of Wntless (Wls), a novel multipass transmembrane protein required for Wg release from 1
14 the producing cells, has raised new questions regarding the mechanism of Wg secretion. How do newly synthesized Wg molecules reach both apical and basal membranes within Wg-producing cells? What cellular machinery is involved in distributing Wg molecules between these possible routes? What is the role of Wls in this process? These questions need to be answered to completely understand the molecular mechanism required for Wg secretion and gradient formation. Therefore, this work is focused to elucidate the function of Wls in Wg secretion, gradient formation and subsequent signaling events. 1.1 Wnt protein family The first member of the wnt gene family was discovered in early 1980s (Nusse and Varmus 1982) as a vertebrate gene near several integration sites of Mouse Mammary Tumor Virus (MMTV), hence called Int1 (integration 1). It was an oncogene that upon insertional activation by MMTV caused cell transformation resulting in mammary carcinomas. A few years later wingless (wg), a gene known to be involved in several aspects of Drosophila development, was found to be a homolog of Int1. The name wnt was derived from the combination of these two words, wg and Int1 (van Amerongen and Nusse 2009). Although the mouse gene is called wnt1 at the present time, and the name Int1 is no longer used, the Drosophila homolog is still referred to as wg. Today, it is clear that Wnts are important regulators of growth and development in all Metazoa. In fact, all currently studied model organisms in the animal kingdom from hydra and the round worm C.elegans, to higher order mammals posses multiple wnt genes. For example, there are a 2
15 total of 5 wnt genes in C.elegans, 7 in Drosophila and 19 in mammals. Even such simple organisms as hydra have 11 different wnt homologs (Llimargas and Lawrence 2001; DasGupta, Boutros et al. 2005; MacDonald, Tamai et al. 2009; van Amerongen and Nusse 2009). Considering the fact that all animal species have retained large numbers of wnts throughout evolution, it is not surprising that Wnt proteins play crucial non-redundant functions in development and adult homeostasis. A variety of processes are controlled by Wnt proteins, ranging from cell fate decision and proliferation to organized cell movements and embryonic segmentation. In the adult, Wnt signaling has an important role in tissue homeostasis through the regulation of stem cell population (Wong 2004). As a result, aberrations in Wnt signaling can cause different pathological disorders such as cancer, degenerative diseases and birth defects which can make an attractive target for new therapeutics (Gerlitz and Basler 2002; DasGupta, Boutros et al. 2005; DasGupta, Kaykas et al. 2005; Goldstein, Takeshita et al. 2006; Lorenowicz and Korswagen 2009; MacDonald, Tamai et al. 2009; van Amerongen and Nusse 2009; Port and Basler 2010; Harterink and Korswagen 2011). Not surprisingly, these molecules have been the subject of a tremendous amount of research since their discovery and we have learned a lot about them within the past few decades. While we have learned a great deal about the mechanisms required to transduce the Wnt signal, there is still considerably more research that need to be done to obtain a complete picture of Wnt signaling and their impact on development and disease. 3
16 1.1.1 Structure Wnt proteins have a molecular weight of about 40 KDa with a relatively large number of ( 23) conserved Cysteine residues, most probably required for their structural stability by means of disulfide bonds. They are approximately amino acids in length with an N-terminal signal peptide for endoplasmic reticulum (ER) targeting and secretion. Efforts to unravel the three-dimensional structure of Wnts have yet been unsuccessful, and their purification has turned out to be challenging. The first successful purification of an active Wnt did not occur until 2003 when active mouse Wnt3A was finally isolated (Willert, Brown et al. 2003). Since then, studies have shown that mature Wnt proteins, with the exception of Drosophila WntD (Ching, Hang et al. 2008), are lipid modified at two sites: a palmitate group is attached to a conserved Cysteine (C77 in mouse Wnt3A, C93 in Wg) and a palmitoleic acid is added to a conserved Serine residue located farther away from the N- terminal (Ser209 in mouse Wnt3A and S239 in Wg respectively). I will discuss the role of these lipid moieties in Wnt secretion later in this chapter. Another modification of Wnt proteins is N-linked glycosylation. The number and location of Wnt glycosylation sites vary among Wnt proteins. Wg like Wnt3A is modified by two N-linked additions (Figure 1.1). Glycosylation has been shown to be dispensable for Wg secretion and signaling (Herr and Basler 2012). However, the importance of this modification in other Wnt proteins remains to be elucidated since data gathered from different experiments seem to be contradictory (Mason, Kitajewski et al. 1992; Komekado, Yamamoto et al. 2007; Lorenowicz and Korswagen 2009; Port and Basler 2010; Harterink and Korswagen 2011). 4
17 The ER-resident enzyme Porcupine (Porc) is responsible for lipid modification of Wnts. It is a membrane-bound O-acyltransferase that interacts with Wnts in a region that includes the C77 residue and is also required for the acylation of S209 position (Tanaka, Okabayashi et al. 2000; Tanaka, Kitagawa et al. 2002; Takada, Satomi et al. 2006). There is also some data suggesting that Porc is required for appropriate N-Glycosylation of Wnts (Lorenowicz and Korswagen 2009). It is possible that Porc is directly involved in glycosylation, or it is possible that upon lipid modification Wnts are better targeted to the ER membrane and more accessible for glycosylation Signal transduction pathways Wnts are able to activate a variety of downstream signaling events. The intracellular response triggered by Wnt signaling was originally identified as a β-catenin dependant pathway, also known as the canonical Wnt signaling pathway. During canonical Wnt signaling β-catenin is stabilized, increasing the intracellular β-catenin pool so it can enter the nucleus and activate target genes (Kennell, Gerin et al. 2008). β-catenin, Armadillo in Drosophila, was initially identified as a protein component of cell-cell adherens junctions (AJs) (Tepass, Tanentzapf et al. 2001). However, genetic screens confirmed that it is also a component of Wnt signaling and is capable of activating gene expression by acting as a cofactor for the lymphoid enhancer factor (LEF)/ T- Cell specific transcription factor (TCF) family of high mobility group (HMG)-box transcription factors. Together with the fact that β-catenin has been found to be frequently mutated in human cancers, the Wnt/β-catenin 5
18 Figure Schematic overview of the structure of mouse Wnt3A Wnt proteins do not contain conserved domains, but they harbor between 23 and 25 conserved cysteines indicated by black lines. A cysteine at position 77 is palmitoylated by Porcupine, an enzyme binding the region indicated in orange. A serine at position 209 (green) is modified with a palmitoleic acid group. Wnt3A also contains two predicted N- glycosylation sites. The signal sequence is indicated in blue. (Lorenowicz and Korswagen 2009) canonical signaling is among the most-studied signaling pathways. As shown in figure 1.2 in the absence of Wnt signaling, cytoplasmic β-catenin is constantly degraded by the action of a complex composed of Axin, APC (product of tumor suppressor gene adenomatous polyposis coli which is known to be mutated in most colon cancers), casein kinase1 (CK1) and glycogen synthase kinase3 (GSK3). CK1 and GSK3 phosphorylate the amino 6
19 terminal region of β-catenin resulting in its recognition by β-trcp, a ubiquitin ligase subunit, and subsequent degradation by proteosome complex. Upon Wnt binding to its receptor, the seven-pass transmembrane protein Frizzled (Fz) and its coreceptor, low-density lipoprotein receptor related protein 5 or 6 (LRP5/6; which is called Arrow in Drosophila) and the formation of Wnt-Fz- LRP5/6 complex, Dishevelled (Dvl) is recruited to the site and activated. The cytoplasmic tail of LRP is also phosphorylated. It has been suggested that the product of division abnormally delayed (dally) gene in Drosophila also acts as a coreceptor for Wnt (Lin and Perrimon 1999; Baeg and Perrimon 2000). Dally and Dally-like (Dlp) are the only glypicans in Drosophila (Baeg, Lin et al. 2001). Glypicans are a class of heparan sulfate proteoglycans (HSPGs) that are attached to a glycosyl phosphatidyl inositol (GPI) anchor in the membrane. Nevertheless, formation of this complex between Wnt and its receptors and subsequent LRP phosphorylation reveals an Axin-binding site. The β-catenin destruction complex, in turn, is destabilized as a result of axin depletion. The non-ubiquitinated β-catenin can now enter the nucleus, displace Groucho from TCF and activate Wnt target gene expression in a complex with TCF/LEF transcription factors. (DasGupta, Boutros et al. 2005; Cadigan and Peifer 2009; MacDonald, Tamai et al. 2009; van Amerongen and Nusse 2009; Gong, Bourhis et al. 2010). 7
 (A) In the absence of Wnt, cytoplasmic β-catenin forms a complex with Axin, APC, GSK3, and CK1, and is phosphorylated by CK1 (blue) and subsequently by GSK3 (yellow).")
20 Figure Overview of Wnt/β-Catenin Signaling (MacDonald, Tamai et al. 2009) (A) In the absence of Wnt, cytoplasmic β-catenin forms a complex with Axin, APC, GSK3, and CK1, and is phosphorylated by CK1 (blue) and subsequently by GSK3 (yellow). Phosphorylated β-catenin is recognized by the E3 ubiquitin ligase, β-trcp, which targets β-catenin for proteosomal degradation. Wnt target genes are repressed by TCF-TLE/Groucho and histone deacetylases (HDAC). (B) In the presence of Wnt ligand, a receptor complex forms between Fz and LRP5/6. Dvl recruitment by Fz leads to LRP5/6 phosphorylation and Axin recruitment. This disrupts Axin-mediated phosphorylation/degradation of β-catenin, allowing β-catenin to accumulate in the nucleus where it interacts with TCF to activate Wnt-responsive genes. Wnts are also capable of inducing β-catenin independent (noncanonical) responses inside receiving cells. In this case, Wnts would activate 8
21 small G-proteins and G-protein kinases, including Rac, Rho, JNK and Rho kinase after binding to their receptors leading to changes in cell shape and polarity. Alternatively, they can cause an increase in the intracellular calcium concentration. This leads to the activation of calcium/calmodulin-dependant protein kinase (CaMK) and protein kinase C (PKC) followed by cytoskeletal rearrangement and cell migration events. Although it has been shown that some Wnts are more potent in activating one pathway versus the others, there is a degree of crosstalk between different signaling responses and they are interrelated (DasGupta, Boutros et al. 2005; Kikuchi, Yamamoto et al. 2009; vandenberg and Sassoon 2009). Endocytosis has also been shown to play important roles in Wnt signal transduction. As its simplest function, it can readily down-regulate signaling by removing activated receptor-ligand complexes from the cell surface. However, new evidence shows that endocytosis plays a more sophisticated role in most signaling pathways. This includes an extremely important function in Wnt signaling. Different components of the endocytotic pathway such as dynamin, Rab5 and clathrin are required for Wnt signaling (Strigini and Cohen 2000; Marois, Mahmoud et al. 2006) and although all details are not clear yet, it is well established that after Wnt binding to its receptor and formation of Wnt-Fz-LRP5/6 complex, the whole complex is internalized from the membrane and transported to early endosomal compartments in the presence of Rab5 (Piddini, Marshall et al. 2005). Entrance of the Wnt signaling complex into these vesicles has been reported to be necessary for the subsequent events including inhibition of GSK3 and 9
22 dissociation of β-catenin from Axin (Le Roy and Wrana 2005; Kikuchi, Yamamoto et al. 2009; Andersson 2011). 1.2 Wnts act as morphogens during animal development Wnts are a major class of morphogens with profound importance in establishing tissue patterning, which drives development in the body plan of all multicellular animals (Strigini and Cohen 2000; Baeg, Selva et al. 2004; Tabata and Takei 2004; Hausmann, Banziger et al. 2007; Harterink and Korswagen 2011). Morphogen is a particular type of signaling molecule, which is secreted from a localized source and acts on neighboring cells directly to induce distinct responses in a concentration-dependant manner (Figure 1.3). In order to be considered a morphogen, a molecule should have three distinct properties: first, it should establish a concentration gradient from a fixed spatial source. Second, it should induce several (at least three) discrete cellular states in receiving cells in response to different thresholds of the concentration gradient. These cellular states are usually identified by the expression of different target genes. And third, morphogens should determine the cellular responses directly (Martinez Arias 2003; Tabata and Takei 2004; Wolpert and Tickle 2010). It can be argued that when the Wnt signal is received by the neighbor cells, they have already started to express a specific set of genes and the function of Wnts are solely maintaining the expression of those genes rather than directly inducing their expression. However, neighboring cells clearly adopt distinct states in regard to different thresholds of Wnt concentration which is the main identifier of a morphogen (Martinez Arias 2003). Development of the Drosophila wing has long been used as a 10
23 model to study the role of morphogen gradient formation in tissue patterning (Gilbert 2010). Wg morphogen gradient formation in the wing disc is considered the prototypical for the study of Wnt morphogen gradients. 1.3 Drosophila Wg is among the most studied Wnt proteins It was previously mentioned that Wingless (Wg) is a founding member of the Wnt protein family. The first identified allele of the wg gene was a regulatory mutation that caused absence of wings in mature flies, which explains the reason for its nomenclature (Martinez Arias 2003). Null alleles of wg, however, have more widespread effects during development and are lethal. Wg is involved in the segmentation and patterning of the embryonic epidermis and in the development of eye and head structures, nervous system and Malpighian tubules. It is also used repeatedly during later stages of development in larvae and adult flies (DasGupta, Boutros et al. 2005). The Drosophila genome codes for seven different Wnt proteins, with wg being equivalent to wnt1 in vertebrates (Herr and Basler 2012). Surprisingly, whereas the depletion of wg results in defective β-catenin signaling, depletion of other Drosophila wnt genes results in very targeted loss of function phenotypes with no widespread effects. In fact, except wnt2 mutants that exhibit a gonadal phenotype and wntd mutants who show defects in immune system function, no phenotype has been observed with the deletion of other wnt genes in Drosophila (van Amerongen and Nusse 2009). This lack of redundancy can be seen in many other components of the Wnt pathway in Drosophila and explains why most regulators of Wnt signaling were first identified in flies. In fact, study of Wg signaling in Drosophila eye 11
 elicits differential gene expression (represented by the blue and pink bars) by activating different genes at different")
24 and wing has been one of the most efficient tools to screen for modifiers of signal transduction cascades (DasGupta, Boutros et al. 2005). Figure 1.3 Morphogen target gene activation A morphogen (green) elicits differential gene expression (represented by the blue and pink bars) by activating different genes at different concentrations (Gonzalez-Gaitan 2003). 12
25 1.3.1 Drosophila wing imiginal disc is a valuable model system for analyzing Wg signaling and gradient formation Imiginal discs are two-sided epithelial sacs allocated from the embryonic epidermis and developing at the larval stage. They are found in pairs within the body of larva and develop to adult appendages such as wings, legs and antennae (Figure 1.4). Imiginal discs are composed of a columnar epithelium (which gives rise to eye, leg, wing ) and an overlaying squamous epithelium, or the peripodial membrane. The apical membranes of the epithelial cells of imiginal disc face each other at the center of the sac with basolateral membranes facing outwards. The wing imaginal disc (Figure 1.5) is the largest among discs with the same structural features. It is subdivided into non-intermingling anterior and posterior compartments. The cells in the posterior compartment express engrailed (en); under control of engrailed, these cells are also expressing hedgehog (hh). The Hh protein is secreted into the anterior compartment, where it acts as a morphogen and activates the expression of several target genes. After being subdivided into A and P compartments, the wing disc is further subdivided to Dorsal (D) and Ventral (V) compartments. As a result of Apterous function on the dorsal cells, they express the gene fringe which subsequently activates the Notch receptor pathway at the DV border. Activated Notch induces Wg synthesis at the dorsoventral (DV) border (Tabata and Takei 2004). Wg proteins secreted from the DV boundary move in both dorsal and ventral directions and form a concentration gradient within both compartments. This gradient is maintained during wing development, and provides the positional information required for activating the correct set of genes by surrounding cells. 13
 A) Imaginal discs are paired structures within the body of the larva that give rise to adult")
26 Figure 1.4 Drosophila imaginal discs (Held 2002) A) Imaginal discs are paired structures within the body of the larva that give rise to adult appendages. B) An imiginal wing disc is shown. The precursors of different parts of an adult wing are marked in this diagram. 14
 of third instar Drosophila larva The imaginal disc is a two-sided sac comprising a columnar cell layer that contains presumptive wing blade (wb) and thorax (t) regions,")
27 Figure Wing imaginal disc (red) of third instar Drosophila larva The imaginal disc is a two-sided sac comprising a columnar cell layer that contains presumptive wing blade (wb) and thorax (t) regions, and an overlying squamous peripodial membrane (pm); All these cell layers are polarized epithelia with their apical sides facing to the center of the sac and basolateral sides facing outwards. The imaginal disc is subdivided into anterior (A) and posterior (P) compartments along the anteroposterior axis. hedgehog (hh) is expressed in the posterior compartment; hh mrna is visualized by in situ hybridization here (Tabata and Takei 2004) Activation of short- and long-range targets of Wg in the wing disc Wg secreted from the cells at the DV boundary of the wing imaginal disc forms a concentration gradient in both compartments and acts as a morphogen. How do other cells respond to this concentration gradient? Wg has several short-range and long-range downstream targets activated at 15
 and achaete (ac) have a high threshold of activation and can only be activated up to 4 cells away from the source.")
28 high or low concentrations of Wg, respectively. Short-range targets of Wg, such as senseless (sens) and achaete (ac) have a high threshold of activation and can only be activated up to 4 cells away from the source. Long range target genes, such as distalless (dll) or vestigial (vg), on the other hand, require lower levels of Wg for their expression and can be expressed as far as 30 cell diameters away from the Wg-producing cells (Bartscherer, Pelte et al. 2006). These properties make wing disc an ideal model for the study of Wg gradient formation and target gene activation. There is still another question regarding Wg downstream target activation which needs to be addressed here: Why different cells have different thresholds? Why do cells next to the DV boundary require a very high concentration of Wg for their activation, while more distant cells can respond to a lower Wg concentration? 16
29 Figure 1.6 Wg short- and long range target expression in wing imaginal disc (Fang, Li et al. 2006) A) Wg (blue) is expressed in a narrow stripe of cells at the DV boundry B) Sens (red) is a short-range target of Wg which is expressed in a group of cells only a few cell diameters away from Wg secreting source C) As a long-range target of Wg, Dll (green) can be expressed in the cells far away from the signal-producing source. This question has been already answered in the context of Drosophila wing disc. It has been shown that cells close to the DV boundary possess fewer Frizzled receptors and less glypicans. Moving away from the DV boundary, cells have higher cell surface levels of Frizzled proteins and glypicans. As a result, those cells located next to the DV border have a less efficient response to Wg and require higher concentrations of Wg to trigger an intracellular response. However, since distant cells have more receptors, they can transduce Wg signaling to the cell even in the presence of fewer Wg molecules (Baeg, Selva et al. 2004). 1.4 Wg secretion and gradient formation in Drosophila wing disc It was discussed in the previous sections that Wg forms a concentration gradient and signal-receiving cells activate the expression of a discrete set of genes in response to different concentrations of the signal. The mechanism of signal transduction from membrane to the cell nucleus was also explained in detail. Researchers have spent a considerable amount of time and effort during the past few decades elucidating the mechanism of Wnt 17
30 signaling; events happening inside receiving cells after Wg binding to its receptors at the cell membrane. Wg secretion, on the other hand, has been very poorly studied due to the assumption that it occurs via a simple secretory pathway. However, several lines of evidence suggest that Wg secretion and gradient formation occurs by a series of complex and specific reactions, many of them remained unknown. The structure of Wg, itself, points to the requirement for a specialized transport mechanism. As previously mentioned, Wnts are highly hydrophobic lipid-modified proteins. How can a hydrophobic molecule diffuse over long distances in the hydrophilic environment of the extracellular space? Discovery of several proteins specifically required for Wnt trafficking in recent years further supports the presence of specialized mechanism for Wnt secretion. Additionally, it has been shown that Wg molecules have different compositions and it is hypothesized to have different functions on the apical and basal membranes of the wing disc epithelial cells. This section focuses on what is currently known about the mechanisms of Wg processing in secreting cells Wntless (Wls) is necessary for Wg secretion from producing cells Genetic screens have been a useful tool for discovery of regulators of Wg signaling. Screens performed in Drosophila wing and eye discs have identified a number of new components of the Wg signaling pathway. Several regulators of Wg signaling have been discovered recently are required for Wg secretion from sending cells, and not signal transduction within the receiving 18
31 cell. The latest examples of such proteins are the members of p24 family. These proteins are involved in both retrograde and anterograde transport between ER and Golgi and it is reported that they are required for Wg secretion from Drosophila cultured S2 cells just like Wg-secreting cells of the wing disc (Palmer, Vincent et al. 2011). It has been proposed that these proteins are required for the packaging of Wg proteins to COPII-coated vesicles and sending them out of ER, but additional experiments need to be performed to verify this hypothesis. Wls is another protein found in genetic screens that is only required in sending cells for Wg secretion. Wls (Evi, Srt) was discovered in 2006 by three independent groups (Bänziger, Soldini et al. 2006; Bartscherer, Pelte et al. 2006; Goodman, Thombre et al. 2006). It is evolutionary conserved from worm to human and has been shown to regulate Wnt signaling in several organisms, including planarians, C.elegans, Drosophila, Xenopus, mice and humans (Adell, Salo et al. 2009; Fu, Jiang et al. 2009; Port and Basler 2010). More recent reports indicate that Wls could also be an interesting protein form the clinical point of view. It has been suggested that the morphine-mediated interaction between Wls and the mu-opioid receptor is responsible for decreased Wnt secretion and the subsequent atrophy of dendritic arbors in the CNS (Jin, Kittanakom et al. 2010). Another report has even confirmed that it is overexpressed in astrocytic gliomas (Augustin, Goidts et al. 2011) making the first link between Wls and cancers. Co-IP experiments showed that Wls can bind Wnt (Bänziger, Soldini et al. 2006; Coombs, Yu et al. 2010). It is suggested that the Wnt 19
32 binding domain of Wls resides within the first intralumenal loop of Wls and it folds into a lipocalin like structure (Coombs, Yu et al. 2010). Lipocalins are a large family of small proteins with a low level of sequence identity, but remarkably homogeneous three dimensional structures. The lipocalin folds consist of an eight-stranded antiparallel β-barrel capable of binding to different hydrophobic molecules, including palmitate (Arnoux, Morosinotto et al. 2009). It is also shown that S209 lipid modification is indispensable for Wnt binding to Wls. More recent experiments have shown that the equivalent of S209 (S239 of Wg) is essential for the recognition of all Wnts by Wls (Herr and Basler 2012) supporting the idea that Wls lipocalin domain binds to the palmitoleic acid residue of Wnts. To further support this hypothesis, it has been shown that Drosophila WntD, an unusual member of the Wnt family which does not carry any lipid modifications, doesn t require Wls for secretion (Ching, Hang et al. 2008; Cadigan and Peifer 2009). Based on these observations, a model can be proposed in which Wg is transferred to Wls immediately after palmitoleate addition by Porcupine at the ER and remained bound to it throughout the secretory pathway. But how Wnt is finally released from Wls? It has been reported that endosomal acidification is necessary for Wnt secretion and inhibitors of acidification interfere with Wls-Wnt dissociation. We also know that a plant lipocalin family member dimerizes upon vesicular acidification (Arnoux, Morosinotto et al. 2009). This may lead to the hypothesis that vesicular acidification induces the release of Wnt from Wls by triggering dimerization of the Wnt-binding domains of Wls. 20
33 Wls utilizes a quite sophisticated intracellular trafficking pathway. After it reached the plasma membrane and released Wnt, Wls is transported back to the ER by retromer mediated Golgi targeting and Golgi to ER retrograde transport in order to take part in additional rounds of Wg secretion (Ratchasanmuang and Selva, unpublished). Mutations of the retromer complex that block the retrograde transport of Wls from endosomes to Golgi can dramatically decrease Wg secretion from sending cells. Interestingly, this phenotype can be successfully rescued by wls overexpression (Belenkaya, Wu et al. 2008; Port, Kuster et al. 2008). A gpi deacylase gene in mouse, which encodes Oto, has also been reported to be involved in Wnt secretion. Oto adds a gpi anchor to Wnts, which causes them to be retained in the ER, presumably for controlled release (Zoltewicz, Ashique et al. 2009). The action of oto homologs in other organisms, including Drosophila is not clear yet Two separate pools of Wg exist at the apical and basolateral membranes of wing disc cells How lipid-modified Wg molecules can travel in the hydrophilic environment of the extracellular space? Alternative mechanisms have been suggested to explain this observation and experimental evidence suggests that several of these mechanisms are active at least in some tissues (Figure 1.7). Wg molecules might be distributed by being directly transferred from cell to cell. Heparan sulfate proteoglycans (HSPGs) are located in the extracellular matrix and are known to bind Wg proteins (Lin and Perrimon 2002). These molecules can form a Wg binding matrix on the surface of the tissue and help 21
34 Wg molecules to spread by a lateral diffusion mechanism. In favor of this hypothesis, it has been reported that Wg molecules cannot cross clones of cells defective in HSPG synthesis (Takei, Ozawa et al. 2004). Another interesting observation in Drosophila imaginal discs is that signal receiving cells have been shown to extend long, actin-based protrusions, known as cytonemes towards morphogen-secreting cells (Hsiung, Ramirez-Weber et al. 2005). These cellular extensions might be responsible for morphogen uptake directly from producing cells. A third possible mechanism for the spread of hydrophobic Wg proteins is the formation of Wg protein micelles that would shield their hydrophobic moieties in the middle of the structure. Formation of such structures has been reported for another lipid-modified morphogen, Hedgehog (Hh) (Chen, Li et al. 2004). It is not clear yet if these micelles play a role in the spread of Wnt proteins. The recent discovery of a protein that binds Wnts with high affinity (Nusse, Fuerer et al. 2008) brings about another possible mechanism: Wnt lipid modifications can be concealed by interactions with lipid binding proteins to make a soluble, mobile morphogen (Port and Basler 2010). Wnts can also be transported between cells on exosomal vesicles produced by secreting cells. This phenomenon has been shown to occur at Drosophila neuromuscular junctions. Here, Wg is secreted along with Wls from the presynaptic terminal on exocytic vesicles (Korkut, Ataman et al. 2009; Korkut and Budnik 2009). In conclusion, several pathways might exist for long-range spread of Wg that may work in parallel in individual cells. It also seems likely that tissue type and developmental status will influence which pathways are used and to what extent (Port and Basler 2010; Therond 2012). 22
35 In the wing disc, yet, another mechanism seems to be mainly responsible for the spread of Wg over long-distances; here, the majority of Wg molecules accumulate inside the apical membrane of Wg-producing cells in the wing disc to form a highly concentrated localized morphogen source required for shortrange signaling. Wg molecules at the apical side are largely immobile and seem to be confined in the plasma membrane. Another pool of Wg molecules can be observed on the basolateral side of the wing disc. Unlike apical Wg, the basolateral Wg pool is composed of mobile Wg molecules forming a concentration gradient throughout the wing disc (Figure 1.8). These Wg molecules are present in a more accessible form that can be stained readily without prior permeabilization using extracellular staining protocols (Strigini and Cohen 2000; Hausmann, Banziger et al. 2007). These observations support the idea that Wg molecules on the apical and basolateral sides of the wing disc are present in different structures and may function differently in signal transduction. They may even utilize different secretion pathways. 23
 with the help of HSPGs, or picked up by protrusions sent from receiving cells towards secreting")
36 Figure 1.7 Proposed models for long-range spread of Wnts Wnts can diffuse laterally on the cell surface (A) with the help of HSPGs, or picked up by protrusions sent from receiving cells towards secreting cells (B). They can also form protein micelles (C), or protein-protein complexes (D) in order to conceal their hydrophobic parts. Other possible mechanisms include integration into lipoprotein particles (E) and incorporation into exosomes (F). (Port and Basler 2010) 24

37 Figure Model depicting short and long-range Wg signaling in the wing imaginal disc. A very high concentration of hydrophobic immobile Wg molecules is maintained on the apical surface of the wing disc. These Wg molecules are responsible for the activation of short-range targets. On the basal side, however, there is a separate pool of Wg molecules. These molecules have incorporated their hydrophobic lipid residues into lipoprotein particles and as a result can readily diffuse within the hydrophilic environment of extra-cellular matrix and spread over long distances. This mobile pool of Wg molecules forms the concentration gradient at the basal surface of the wing disc responsible for the activation of long-range signaling targets (Cadigan and Peifer 2009) 25
38 What are the structural features of Wg molecules at the basolateral surface of the wing disc that allows them to freely diffuse within the extracellular matrix? Several lines of evidence suggest that Wg molecules are incorporated into lipoprotein particles at the basal side (Panakova, Sprong et al. 2005; Boutros 2008). Lipoprotein particles consist of a central core of neutral lipids and cholesterol and an outer phospholipid monolayer and are scaffolded by large proteins called apolipoproteins. In insects lipoprotein particles are synthesized by the fat body (a tissue functionally related to the liver) using only one apolipoprotein, Lipophorin (Therond 2012). It was reported that Wg derived from wing discs cofractionated and colocalized with lipoprotein particles; Wg could be copurified with apolipoproteins and furthermore Wg signaling was altered in flies with reduced Lipophorin levels (Panakova, Sprong et al. 2005). There is also evidence that glypicans recruit lipophorin to morphogen secreting cells (Eugster, Panakova et al. 2007) which can explain the role of HSPGs in Wg gradient formation. Wg and Hh are present in the Drosophila haemolymph, and in the columnar epithelial cells of the Drosophila wing imaginal disc two pools of Hh have been identified at the apical and basolateral membranes with differential activities at establishing short and long-range gradients (Therond 2012). Hedgehog (Hh) is another lipid-modified morphogen with similar secretion properties to Wg. Based on these data, it can be proposed that newly synthesized Wg molecules are first transported to the apical membrane, where most of them remain in a nonmobile form. Consistent with this model, it has been reported that most wg transcripts are localized near the apical membrane (Manseau 2001; 26
39 Simmonds, dossantos et al. 2001). A portion of Wg molecules are subsequently rerouted from the apical surface. These molecules are loaded onto lipoprotein particles and secreted again from the basolateral membrane. Some observations further support this model. First of all, apical to basal transcytosis has been shown to occur for dally-like protein (Dlp) in the wing discs. Dlp transcytosis also promotes Wg spreading along the basolateral compartment (Gallet, Staccini-Lavenant et al. 2008). As mentioned before, in signal receiving cells Wg is endocytosed in complex with Frizzled, Arrow and most probably Dally. Experimental evidence suggests that Frizzled2 provides the endocytic signals, while Arrow stimulates the targeting of Frizzled-Wg complex to lysosomal compartments (Piddini, Marshall et al. 2005). In sending cells, on the other hand, Wg seems to be endocytosed from the apical membrane in complex with Wls and Dlp (Port and Basler 2010). As a result Wg might rely on glypicans for apical to basal transcytosis and loading into lipoprotein particles. The discovery of an endocytosis motif in an intracellular loop of Wls protein (Gasnereau, Herr et al. 2011) makes this idea even more plausible. Interestingly, Arrow is not able to target Dally like-wingless complexes for degradation (Piddini, Marshall et al. 2005). Further support for this model comes from experiments with Reggie proteins (Katanaev, Solis et al. 2008). Reggie 1/2, or Flotillin 2/1 respectively, are microdomain forming proteins acting as a scaffold for recruitment of gpi-linked proteins, cholesterol and other components of lipid rafts (Langhorst, Reuter et al. 2008; Stuermer 2012). Overexpression of Reggie1 could cause more Wg secretion from the basolateral surface while 27
40 reducing apical Wg. This experiment clearly shows that Wg molecules rely on lipid raft-dependant apical to basal transcytosis to form their long-range gradient at the basolateral surface of the wing disc. 1.5 Scope of the thesis Our current picture of Wg secretion and gradient formation is not complete. Several questions need to be addressed in order to have a satisfactory knowledge of Wg secretion. I will particularly focus on a subset of those questions involving the role of Wls in Wg secretion and gradient formation. Does Wls have differential roles in establishing short and longrange signaling? What is the role of Wls in Wg transcytosis? Is Wls required for loading Wg into lipoprotein particles? What is the effect of acidification on Wls activity specifically its activity regarding Wg gradient formation? Can Wls alter the shape of Wg gradient? These are the questions that I have worked to address in my research. 28
41 Chapter 2 MATERIALS AND METHODS 2.1 Fly Stocks A variety of fly stocks were used during my experiments. Some of them were maintained in our laboratory under standard conditions. The remaining stocks were obtained from the Bloomington Drosophila Stock center at Indiana University which is currently the largest and most diverse of the D. melanogaster collections (Matthews, Kaufman et al. 2005). A complete list of stocks used to perform experiments and their sources is provided in table Targeted gene expression The GAL4/UAS system was used for targeted gene expression in Drosophila wig discs. GAL4 is a yeast protein of 881 amino acids and a regulator of genes induced by galactose. It directly binds to four related 17 bp sites to exert its effect. These sites define an Upstream Activating Sequence (UAS) element, analogous to an enhancer element defined in multicellular eukaryotes. UAS is essential for the transcriptional activation of GAL4 regulated genes. In the GAL4/UAS system, expression of the gene of interest, the responder, is controlled by the presence of an optimized UAS element. 29
42 Table 2.1 Drosophila stocks used to perform experiments Genotype w/w;uaswlscha/uaswlscha;δwls FRT/TM6C Source Function Lab Expressing C- terminal HA tagged version of wls under UAS control w/w;uaswls(cg6210)/tm3 Lab Expressing wls under UAS control w/w;engal4/cyo;hhgal4/tm6c Lab Double driver line; expressing GAL4 in en- and hhexpressing compartment of the wing disc i.e. posterior compartment w/w;hhgal4/tm6b w/w;wggal4/cyo;dr/tm6c Bloom. Expressing GAL4 in hh-expressing compartment Bloom. Expressing GAL4 in Wg-producing cells w/w;wggal4/cyo;uasgfp/tm6c Lab Expressing GAL4 in Wg-producing cells; also expressing GFP under UAS control w/w;vgq-lacz/cyo;uaswls/tm6c Lab Expressing wls under UAS control and B-gal in Vgproducing cells w/w;vgq-lacz/cyo;dr/tm6c Lab Expressing B-gal in Vg-producing cells w/w;c96gal4/tm6c Bloom. Expressing GAL4 in a narrow stripe of cells around the D/V boundary w/w;uasgfp/cyo;c96gal4/tm6 Lab Expressing GAL4 in a narrow stripe 30
43 w[*];p{w[+mw.hs]=gawb}dpr[pgaw]/ CyO w[*]; P{w[+mW.hs]=GawB}c409 y[1] sc[*] v[1]; P{y[+t7.7] v[+t1.8]=trip.hms01056}attp2 y[1] sc[*] v[1]; P{y[+t7.7] v[+t1.8]=trip.hms01442}attp2 y[1]v[1];p{y[+t7.7]v[+t1.8]=trip. JF01821} attp2 y[1]v[1];p{y[+t7.7]v[+t1.8]=trip. JF02029} attp2 y[1]v[1];p{y[+t7.7]v[+t1.8]=trip. JF02059} attp2 y[1]v[1];p{y[+t7.7]v[+t1.8]=trip. HM04032}attP2/TM3,Sb y[1]sc[*]v[1];p{y[+t7.7]v[+t1.8]=trip. HM00821}attP2 of cells around the D/V boundary; also expressing GFP under UAS control Bloom. Expressing GAL4 in the chemosensory bristles of the anterior wing margin Bloom. Expressing GAL4 in eye discs, follicle cells, and anterior wing Bloom. Expressing dsrna for RNAi of Vha68-2 under UAS control Bloom. Expressing dsrna for RNAi of VhaC39-1 under UAS control Bloom. Expressing dsrna for RNAi of Vha16-5 under UAS control Bloom. Expressing dsrna for RNAi of VhaM9.7-1 under UAS control Bloom. Expressing dsrna for RNAi of Vha100-1 under UAS control Bloom. Expressing dsrna for RNAi of Vha100-5 under UAS control Bloom. Expressing dsrna for RNAi of Vha44 under UAS control 31
44 Because transcription of the responder requires the presence of GAL4, the absence of GAL4 in the responder lines maintains them in a transcriptionally silent state (Figure 2.1). To activate their transcription, responder lines are mated to flies expressing GAL4 in a particular pattern, termed driver. The resulting progeny then express the responder in a transcriptional pattern that reflects the GAL4 pattern of the respective driver (Duffy 2002). 32
45 Figure 2.1 The UAS/GAL4 system in Drosophila When females carrying a UAS reporter (UAS-GFP) are mated to males carrying a GAL4 driver progeny containing both elements of the system are produced. The presence of GAL4 in an alternating segmental pattern in the depicted embryos then drives expression of the UAS responder gene in a corresponding pattern (Duffy 2002). 2.3 Wing disc dissection and immunostaining The third instar larvae were harvested and primary dissection was performed in PBS (solution of 137mM NaCl, 2.7mM KCl, 10mM Na₂HPO₄ and 2mM KH₂PO₄ at ph 7.4). Larvae were then fixed in PBS with 4% formaldehyde for 20min at room temperature and washed four times 5-10min each with PBT (PBS+0.1% Triton X100). Blocking was achieved by incubating them for 30min in PBTN (PBT+5% normal horse serum), followed by primary antibody treatment overnight at 4 C. The primary antibodies and their dilutions were as following: Rabbit anti Arrow 1:1000-1:1500, mouse anti Pdi 1:100, rabbit anti HA 1:100, rabbit anti Lamp1 1:500, rabbit anti Rab5 1:50, Chicken anti Bgal 1:500, chicken anti Av1 1:500, mouse anti Dll 1:200, guinea pig anti Sens 1:500, Mouse anti Wg 1:20, Rat anti Ci 1:10 The following day larvae were washed four times 5-10 min each in PBT and incubated with secondary antibody in PBTN for 1 hour. Secondary antibodies were used at the following dilutions: Anti-mouse, -goat and -chicken antibodies were used at a 1:500-1:1000 dilution. Anti-rat antibodies were used at a 1:100 dilution. Depending on the desired colors, secondary 33
46 antibodies were either Alexa Fluor 568, 647 or 488 from Invitrogen. After secondary staining, larvae were washed 4 times with PBT and wing discs were obtained by secondary dissection. The wing discs were then mounted on slides using 70% glycerol solution. Slides were observed and analyzed under 40X oil objective lens of Zeiss LSM 780 confocal microscope. For extracellular staining, primary larval dissection was performed in cold Schneider cell media on ice and larvae were exposed to mouse anti- Wg antibody diluted 1:5 in cold Schneider cell media on ice for 1 hour before proceeding to formaldehyde fixation. They were then fixed, secondary stained and dissected as described. Bafilomycin treatment was performed using the following protocol: After completing primary larval dissection in Schneider cell media (at room temperature), the larvae were exposed to 500 nm solution of Bafilomycin A1 (sigma) in Schneider cell media for 4-5 hours. They were then formaldehyde fixed, stained and dissected as described. 2.4 Adult wing phenotypes Wings were separated from adult flies on CO₂ pads and placed on a slide. Euparal mounting medium was added to the slide followed by a coverslip. Slides were heated overnight at 55 C and imaged at 50X magnification using Zeiss Axiophot microscope. Digital images were processed using Adobe Photoshop CS4. 34
47 2.5 Image processing and statistical analyses The processing of confocal wing disc images to determine pixel intensity and threshold boundaries were performed using Zeiss Zen 2010 software. For comparing the range of extracellular Wg staining between anterior and posterior compartments, an intensity profile was computed along the dorsoventral axes of both compartments and the length of the region showing intensities over 50 was interpreted as the range of extracellular Wg expansion. In the case of Dll, wild type and wls-overexpressing wing discs were placed in the same microcetrifuge tube after primary dissection and stained and imaged under the same conditions. Intensity profiles were calculated along the dorsoventral axis of posterior compartment for wild type and wls-overexpressing wing discs and the same threshold value (50) was used to distinguish dll-expressing cells and measure the length over which Dll is expressed. Wild type flies were carrying a vgq-lacz reporter construct and β-gal staining was used to distinguish them from flies overexpressing Wls in the posterior compartments of their wing discs. Ci staining was used in all cases to distinguish anterior compartment of the wing disc. Obtained data from two groups were compared using two-tailed t-test. Statistical analyses were done using Microsoft Excel and Graphpad programs. All graphs were drawn in Microsoft Excel. 35
48 Chapter 3 RESULTS 3.1 Effect of Wls overexpresion on Wg short- and long-range signaling In order to determine the role of Wls in Wg secretion and signaling, the wls gene was overexpressed using the GAL4/UAS system of gene expression specifically in the posterior compartment of the wing disc with the en-gal4; hh-gal4 double driver line. Using this GAL4 driver line, I monitored wing development and gene expression patterns of Wg targets in the posterior wls-overexpressing compartment and compared these markers to the neighboring anterior compartment, which has endogenous Wls levels. Results obtained from these experiments confirmed a differential role for Wls in shortand long-range Wg signaling Wls overexpression causes loss of wing margin in the adult wing During the third-instar larval stage of Drosophila development, the wing disc is divided into distinct anterior, posterior, dorsal and ventral compartments. The Wg producing cells located at the dorsovetral (D/V) boundary. As wing disc develops into adult wing the cells at the D/V boundary will make the margin of the adult wing (Figure 3.1). And those cells right next 36
 and posterior (P) compartments along the A/P axis, and is further subdivided into dorsal (D) and ventral (V) compartments along the D/V axis (dashed")
49 Figure 3.1 Development of Drosophila wing The adult Drosophila wing develops from the larval imaginal disc a single-layered sac of polarized epithelial cells present in the larva (left). The wing disc is subdivided into anterior (A) and posterior (P) compartments along the A/P axis, and is further subdivided into dorsal (D) and ventral (V) compartments along the D/V axis (dashed line). Cells in the P compartment express hedgehog (Hh, blue). Only part of the imaginal disc, indicated by a bracket, develops into the adult wing (right). The D/V border of the imaginal disc develops into the margin of the adult wing. To visualize this movement, imagine picking up the D/V border in the disc and pulling it towards you, out of the page (Tabata 2001). to the D/V boundary are programmed by Wg short-range signaling to make the sensory bristles of the adult wing. As a result, any perturbation of shortrange Wg signaling can cause a lack of adult wing margin and bristles. This phenotype has been used successfully to screen for the genes that specifically influence Wg short-range signaling. 37

50 Overexpression of wls resulted in a wing nick phenotype and lack of bristles in the posterior half of the adult wing (Figure 3.2). Loss of margin bristles is indicative of defects in Wg short-range signaling. This phenotype was only observed in males, which could be due to hormonal differences between genders. Figure 3.2 wls overexpression in the posterior compartment of the wing causes wing nicks The image on the left shows a wild type wing. The right panel shows the adult wing when wls is overexpressed. Lack of wing margin and bristles can be seen in the same compartment where Wls is being overexpressed. 50 wings from male flies were investigated with 40% of them showing this phenotype. The same phenotype was very rare in females. Scale bar shows 100 microns. 38
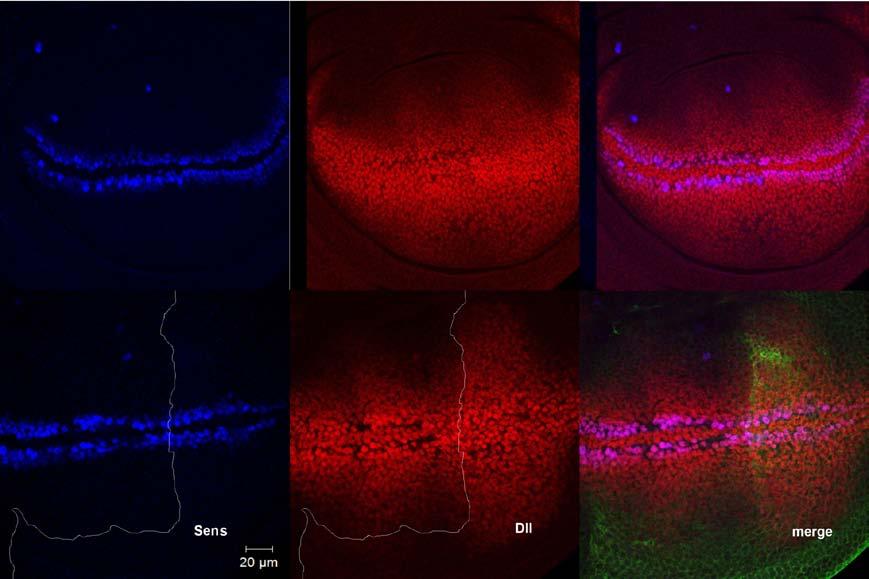
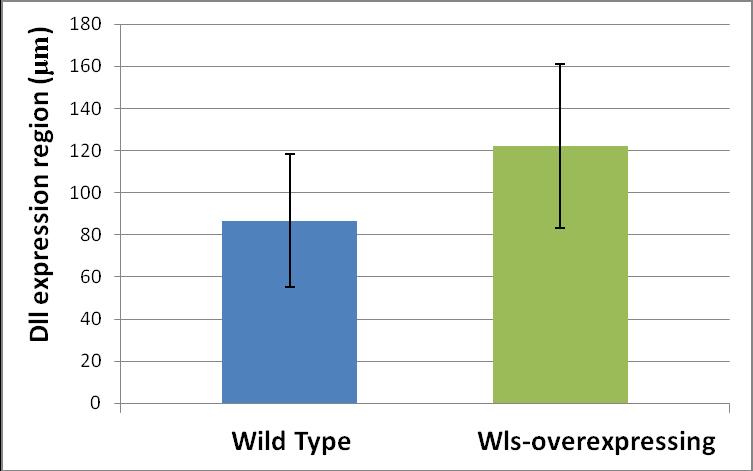
51 3.1.2 Alterations of Sens and Dll expression in wing discs In order to look at Wg short- and long-range signaling more directly, I examined the expression of a short-range and a long-range Wg target and compared them between wls-overexpressing and unaltered compartments of the wing disc. Senseless (Sens) is a short-range target of Wg signaling, while Distalless (Dll) is a long-range target. Loss of Sens expression in the posterior compartment was seen in a few wing discs (Figure 3.3). Dll has a very dynamic expression pattern; both inside a single disc and from one disc to another. As a result of this uneven expression pattern, it was difficult to judge if an expansion of Dll expression had occurred in the posterior compartment only by looking at the micrographs (Figure 3.4 A). My efforts to use another long-range target of Wg signaling, Vestigial (Vg), were also unsuccessful as it filled the whole wing pouch and did not allow evaluation of any differences in the expression range between anterior and posterior compartments. To resolve this problem, I took advantage of statistical approaches. Wls-overexpressing and wild type wing discs were immunostained for Dll and the range of Dll staining was compared between these two groups. Results indicate that there is a significant difference in the range of Dll expression between wild type and wls-overexpressing wing discs (Figure 3.4 B) 39

52 Figure 3.3 Loss of Sens expression in the wls-overexpressing compartment In this confocal image Wg is red, Sens is blue and Wls is stained green. Top: the green part of the wing disc on the far right panel depicts the posterior compartment where wls is being overexpressed. Wg-producing cells at the D/V boundary can be seen on the left, and center, Sens is absent in the posterior compartment in contrast to the wild type discs (bottom) where Sens is expressed in both compartments. This phenotype was seen only in a few wing discs during my observations. 40
53 A B 41
54 Figure 3.4 Dll expression in wls-overexpressing wing discs A: Top panels show a wild type wing disc and bottom panels show a wing disc overexpressing wls in the posterior compartment. Dll is stained in red, Sens in blue and Wls in green to define the borders of wls-overexpressing compartment. It can be observed that Dll has a heterogeneous expression pattern about the D/V boundary even in the wild type wing discs (middle, top) which may account for the apparent expansion of Dll in the posterior compartment viewed in the lower panel. Note that in this wing disc sens expression is intact in the posterior compartment. B: There is a statistically significant difference (P=0.023) between the range of Dll expansion in wild type and wlsoverexpressing wing discs. To perform these measurements, 12 wing discs were obtained from each group and stained together. After acquiring their images under the same condition, the length of Dll expression around the dorsoventral boundary was measured in the posterior compartment for both groups. Numbers are in μm Wg relocalization from apical to the basolateral membrane As previously discussed, there are two different pools of Wg present at the apical and basolateral membranes of the wing disc epithelium with the apical pool being responsible for short-range target activation. Therefore, it is predictable that if a factor alters Wg short-range signaling, it would alter the apical localization of Wg proteins. To evaluate alterations in 42
55 Wg localization, I looked at Wg accumulation at different surfaces of the wing disc by taking individual optical sections across the entire wing disc. Data showed that in the posterior compartment of the wing discs where Wls is overexpressed, less Wg was present at the apical surface. On the basal side, however, the opposite pattern was observed with more Wg punctate structures present in the posterior compartment. These Wg punctate vesicles had also traveled farther from the secreting source in the posterior compartment (Figure 3.5). These observations suggest that upon wls overexpression, Wg molecules are relocated from the apical to the basolateral membrane and as a result more Wg-containing vesicles can be seen on the basal membranes of the wing disc epithelial cells accompanied by a simultaneous reduction of the apical Wg concentration. Since apical Wg is responsible for the activation of short-range signaling targets, these target genes should display reduced expression. The observed loss of wing margin in wls overexpressing cells is consistent with reduced short range signaling (Figure 3.2). At the same time more Wg molecules are present at the basal side and these molecules expand over a wider range. This would predict activation of long-range Wg targets at farther distances from the D/V boundary. Expansion of the expression region of Dll was also observed in wls overexpressing cells (Figure 3.4). 43
 is a protein specifically expressed in the anterior compartment.")
56 Figure 3.5 Wg apical to basal redistribution after wls overexpression Note that in these images another approach has been used to identify the Wls-overexpressing compartment. Wls is overexpressed in the posterior compartment using en-hh- GAL4 driver line. Ci (green, left panel) is a protein specifically expressed in the anterior compartment. Hence, the compartment of wls overexpression is opposite to the Ci expressing compartment. Ci staining will be used for the same purpose in all subsequent images of this chapter. In wls-overexpressing compartment, Wg is absent from the apical membrane at the D/V boundary (center). On the basal side, however, more Wg punctates are observed and at a larger distance from the D/V boundary (right). Expansion of Wg punctates can also be distinguished in figure The amount and range of expansion of extracellular Wg increased in wls-overexpressing tissue When wing discs are exposed to Wg antibody before fixation and permeabilization, the antibody only has access to those Wg molecules that reside on the extracellular cell membranes. It has been shown that by using this method the apically-localized Wg molecules would not be detected, while the mobile Wg-containing structures at the basal membrane are readily stained (Strigini and Cohen 2000). This observation is consistent with our 44
57 proposal that there is a lipoprotein-incorporated Wg pool at the basal membrane, capable of diffusion within the extracellular matrix. Extracellular Wg staining in the posterior compartment showed a stronger staining over a longer distance from the D/V boundary (Figure 3.6 A). I also measured the distance over which staining could be observed in the anterior and posterior compartments of the wing disc and compared them together. Again, the data showed that extracellular Wg has distributed over a longer distance in the posterior compartment in comparison to the anterior (Figure 3.6 B). In summary, when wls is overexpressed in a group of Wg-secreting cells, those cells accumulate less Wg on the apical membrane in comparison to adjacent wild type cells. However, they secrete more diffusible Wg from the basolateral surface making Wg molecules detectable over a longer distance at the basolateral face of the wing disc. Because apical Wg is responsible for activation of short-range signaling targets, and long-range Wg gradient is established on the basolateral side, the net effect of wls overexpression could be interpreted as shifting Wg molecules from short-range to the long-range Wg targets. These results suggest that Wls may play a role in the fate of short- versus long-range Wg targets by altering the subcellular localization of Wg and changing the Wg morphogen gradient and hence provides an interesting example of a molecule that can change the shape of a morphogen gradient. 45


58 A B Figure 3.6 Expansion of extracellular Wg staining in wlsoverexpressing compartment 46
59 A: In this micrograph extracellular Wg is stained red and the Ci-expressing compartment (anterior) is marked in blue. Wls has been over-expressed in posterior compartment. Comparing anterior and posterior compartments, it is clear that in the posterior half of the wing disc the amount of extracellular Wg has increased in comparison to the anterior. Also note that staining is observable over a wider range in the posterior portion. B: The difference between the range of extracellular Wg expansion in anterior and posterior compartments of wlsoverexpressing wing discs is very significant (P=0.004). 12 wing discs were obtained from larva overexpressing Wls in the posterior compartment of their wing discs and the region of extracellular Wg staining was measured and compared between anterior (normal) and posterior (wls-overexpressing) compartments. 3.2 Alteration of Wg intracellular trafficking in response to wls overexpression In the previous section, I discussed that when wls is overexpressed, Wg molecules are re-routed from apical to basolateral membranes. Vacuolar acidification is known to be indispensable for Wnt signaling (George, Leahy et al. 2007; Cruciat, Ohkawara et al. 2010) and recent reports have demonstrated that acidification involved in some aspects of Wg-Wls interaction (Coombs, Yu et al. 2010). With this information, I decided to further inspect the role of acidification in Wg secretion specifically 47
60 in wls-overexpressing tissue. This approach provided valuable insights as I discuss in the following sections Chemical inhibition of vacuolar acidification Bafilomycins are a family of toxic antibiotics that act as specific inhibitors of vacuolar-type H+ ATPases (V-ATPases). They do not have any effect on other ATPases and by inhibiting V-ATPases, they are able to block intracellular acidification (Yoshimori, Yamamoto et al. 1991). The most used bafilomycin is bafilomycin A1. It has been used successfully in numerous studies as a specific inhibitor of acidification and it is commercially available. As a result, I used this reagent to perform my experiments Disrupted acidification caused Wg retention in secreting cells To investigate the role of acidification in Wg secretion, I incubated wild type wing discs in a Bafilomycin A1 solution and looked for any changes in Wg secretion. After a relatively short exposure to Bafilomycin, a phenotype was observed. Wing discs incubated with bafilomycin showed a strong retention of Wg at the apical membrane, while little Wg was observed on the basal side (Figure 3.7). This results show that acidification is required for Wg apical to basal transcytosis and that in the absence of acidification little Wg is available to be secreted from Wg-producing cells at the D/V boundary. 48
, it is clear that bafilomycin treatment has caused Wg retention.")
61 Figure 3.7 Wg retention in bafilomycin A1 treated wing discs Upper panels show an untreated wild type wing disc. Wing discs in the lower panels were incubated in Bafilomycin A1 solution. Wg staining is shown in red. Comparing apical sides (images on the left), it is clear that bafilomycin treatment has caused Wg retention. Stronger Wg staining is observed in treated wing discs with no punctates in receiving cells. Also note that there is little Wg present on the basal side of the treated wing discs. 49
62 wls overexpression was able to reduce the negative effects of acidification inhibition Wing discs observed in the previous experiment were wild type discs. To study the relationship between acidification and Wls activity, I incubated wls-overexpressing discs in bafilomycin. Similar to experiments explained at the beginning of this chapter, I used wing discs that were overexpressing wls at their posterior compartments. These wing discs were exposed to bafilomycin for a short period, and the Wg distribution pattern was subsequently compared between anterior and posterior compartments. Surprisingly, it was observed that in these wing discs, the posterior wlsoverexpressing compartment is less sensitive to the effects of bafilomycin compared to the anterior compartment (Figure 3.8). 50

63 Figure 3.8 Wls overexpression is able to rescue the effect of V-ATPase inhibition on Wg secretion In this micrograph Wg is stained green and blue is used to mark Ci in order to determine the borders of the anterior compartment (Wls is over-expressed in the posterior). Upper panels are optic sections from apical surface of the wing disc and lower panels show the basolateral surface. There is clearly more Wg present in the posterior compartment at the basal side (bottom, left) and also some Wg punctates can be seen inside the receiving cells at the apical side of posterior compartment, while those punctates are absent in the anterior compartment (top, left). 51
64 3.2.2 Similar effects were observed following RNAi inhibition of V-ATPase Experiments with Bafilomycin provided an insight into the role of acidification and Wls function. However, wing discs can only be exposed to bafilomycin following dissection of the larva, which is why this technique doesn t allow us to follow wing discs into later stages of development and observe the long-term effects of V-ATPase inhibition. To tackle this problem, I used RNAi responder lines against multiple components of the V-ATPase complex. Using selected driver lines, I was able to express RNAi molecules in a controlled pattern inside developing wing discs. Out of several RNAi responder lines I examined during my experiments, only two showed a wing phenotype. One of these two responder lines expressed RNAi against Vha68-2 component of the V-ATPase and since Vha 68-2 has been previously reported as an important regulator of Notch signaling (Vaccari, Duchi et al. 2010), I decided to focus on this responder line Vha68-2 RNAi expression at the D/V boundary caused loss of wing margin in adults My goal was to investigate the role of acidification in Wg-secreting cells and as the first line of experiment, I decided to express RNAi against V- ATPase components in Wg-secreting cells using wg-gal4 driver lines. However, expression of Vha 68-2 RNAi in Wg-producing cells turned out to be lethal. Lethality occurred at a very early stage of development and I was unable to acquire wing discs from this cross. C96-GAL4 is another driver line that expresses GAL4 in Wg-secreting cells at the D/V boundary, as well as a few cell diameters around it (Figure 3.9). The difference between C96-GAL4 52
65 and wg-gal4 is that C96-GAL4 only expresses GAL4 in the wing disc, while wg-gal4 is expressing GAL4 in all Wg-producing cells inside the body. Using the C96-GAL4 driver line, I was able to collect some adult progeny which showed very severe wing phenotypes (Figure 3.10). The phenotype expressed by these flies was not only a simple lack of wing margin, as large portions of the wing were not formed. It is more appropriate to consider this phenotype as an indicative of major loss of Wg signaling, possibly as a result of Wg retention inside producing cells and blockage of Wg secretion. 53

66 Figure 3.9 C96 expression in the wing disc In this confocal image red staining on the left shows Wgsecreting cells and green shows C96 expressing cells. It could be seen that C96 is expressed in Wg-secreting cells of the wing disc as well as a few cell diameters around it. To obtain this image the UAS-GFP responder lines were mated to C96-GAL4 driver lines. Figure 3.10 Adult wings are highly disrupted when acidification is interrupted in the wing disc Image on the left shows a wild type wing, while shown on the right side are flies that expressed RNAi against a component of V-ATPase (Vha68-2) in C96-expressing cells (refer to the previous figure). Wing margin is almost completely lost, but other defects, such as reduced wing size are observed. This phenotype was 100% penetrant. Scale bar shows 100 microns. 54
67 Wg is absent from the basal membrane of secreting cells Looking at the wing discs of larva expressing RNAi for V-ATPase under control of C96-GAL4 a very clear phenotype is observable: Wg is absent on the basal membrane of these discs (Figure 3.11). Of course, a very strong Wg retention on the apical membrane, like what observed in Figure 3.7, could not be seen in these wing discs. However, the overall effect is similar to chemical inhibition of V-ATPase with bafilomycins. There is more Wg on the apical surface in comparison to the wild type wing discs, while the basal Wg has been significantly reduced. 55
.")
68 Figure 3.11 Basal Wg shows a significant decrease after V-ATPase inhibition Top panels compare apical Wg between wild type wing discs and wing discs expressing RNAi for V-ATPase. It can be seen that as a result of V-ATPase inhibition, a mild retention of Wg occurs on the apical surface of the wing disc. This retention is not very significant though (Compare with Figure 3.7). On the basal surface, however, a very significant difference between wild type and RNAi-expressing discs can be distinguished (bottom panels). Note the overall similarity between this image and Figure
.")
69 Extracellular Wg is distributed over a narrower range as a result of V-ATPase inhibition in the wing disc The extracellular Wg staining was expanded over a shorter distance and showed less intensity in the wing discs expressing RNAi against V-ATPase subunits compared to the wild-type wing discs (Figure 3.12). This observation is consistent with the idea that intracellular acidification is required for the secretion of mobile Wg molecules from the basolateral membranes of Wg-producing cells. Figure 3.12 Extracellular Wg reduction caused by V-ATPase inhibition In this image, the left panel shows the extracellular Wg staining for a wild-type wing disc and the right panel for a wing disc expressing RNAi against V-ATPase subunits under control of C96-GAL4. V-ATPase inhibition has affected both intensity and expansion of the extracellular Wg. 57

70 Dll expression is noticeably reduced after V-ATPase inhibition Considering the previously mentioned observations of basal and extracellular Wg reduction, a decrease in Dll expression is highly predictable in the RNAi-expressing wing discs. As shown in figure 3.13, this prediction is proved to be correct. Figure 3.13 Dll expression is negatively affected by V-ATPase inhibition in the Wg-producing cells Here, the left panel is showing a wild-type wing disc and the right panel, a wing disc expressing RNAi against V-ATPase under control of C96-GAL4. Both wing discs are stained for Dll under similar conditions. The reduction of Dll expression in the RNAi-expressing wing disc can be easily noticed. 58
.")
71 Sens expression around the D/V boundary is adversely affected by V-ATPase inhibition in the Wgproducing cells After V-ATPase inhibition in the Wg-producing cells using C96- GAL4 driver lines, sens expression was lost in the cells surrounding the D/V boundary (Figure 3.14). Considering the fact that Wg is still present on the apical membranes of secreting cells, and that apical Wg is supposed to be responsible for short-range signaling, this observation was rather unexpected. Figure 3.14 Sens expression is absent around the D/V boundary in the RNAi-expressing wing discs The right panel shows Sens staining for a wing disc expressing RNAi against V-ATPase under control of C96- GAL4. 59
72 As you can see, Sens expression around the D/V boundary is totally lost (compare to the wild-type wing disc on the left). Also notice that other sens-expressing cells throughout the wing disc are stained, showing that lack of signal around the D/V boundary is not due to poor staining Wls over-expression in the wing disc did not rescue V-ATPase inhibition phenotype Due to the less severe effects of bafilomycins on the wlsoverexpressing compared to the wild-type wing discs, I predicted that wls overexpression in the RNAi-expressing wing discs would attenuate the effects of V-ATPase inhibition. In order to overexpress wls and Vha-RNAi simultaneously, I mated double responder lines (UAS-dsRNA;UAS-wls) to C96-GAL4 driver lines. This way, the progeny would express both RNAi and Wls in the cells surrounding the D/V boundary. Contrary to my prediction, over-expression of both RNAi and Wls in the Wg-producing cells turned out to be lethal. However, I believe this lethality in not due to a synergistic interaction between Wls and V-ATPases; it is rather a general response to protein accumulation. C96-GAL4 is a very strong driver and when used to drive the expression of two different proteins, those proteins are expressed at high quantities and their accumulation inside the cells causes lethality. The same effect was observed when I crossed RNAi responder lines to a C96-GAL4 line with GFP in the background (C96- GAL4;UAS-GFP). No adult flies could be obtained from this cross (data not 60
73 shown). Since the progeny of UAS-dsRNA x C96-GAL4 cross were viable, the additional expression of GFP was not expected to affect the viability. However, GFP expression caused lethality, probably due to protein accumulation in Wg-producing cells. 61
74 Chapter 4 DISCUSSION The data gathered throughout the results section reveal some new insights into the function of Wls in Wg secretion and gradient formation. They also provide a framework for future investigations regarding intracellular trafficking pathways of Wg proteins that influence downstream signaling. Here, I summarize the presented data and hypothesize about possible future directions of this project. 4.1 wls overexpression drives Wg molecules toward the long-range gradient Results of several of my experiments are consistent with the idea that upon wls overexpression, more Wg would be driven to the long-range concentration gradient. This movement to the long-range pool occurs at the expense of short-range signaling and a significant decrease could be observed in the amount of Wg molecules dedicated to short-range signaling in wls-overexpressing compared to the wild type tissue. This shift in Wg can best be visualized by looking at Wg localization and extracellular staining. Examination of Wg staining at different optical sections along the apical-basal axis of the wing disc reveals that in the wls-overexpressing compartment less Wg could be found on the apical side in comparison to the neighboring wild type compartment. As previously discussed, the apical Wg pool is responsible 62
75 for the activation of short-range signaling targets. So decreasing the amount of Wg on the apical membranes directly leads to the alteration of short-range Wg signaling. In the same optical series, however, as we continued to move towards the basal surface, the opposite pattern was observed: the amount of Wg was clearly increased on the basal membrane as a result of wls overexpression. Furthermore, Wg punctate vesicles were observable over a wider region indicating that Wg molecules had been taken up and induced signaling in fairly distant cells. This expansion of Wg concentration gradient was easier to visualize by examining the extracellular Wg distribution. Observations suggest that only basally-located, mobile pool of Wg proteins diffuse freely in the extracellular matrix without being confined inside membranes. The apical Wg pool, on the other hand, seems to be intracellular and is not accessible through the extracellular staining protocols (Strigini and Cohen 2000). It is also reported that Wg-containing lipoprotein particles are present in the extracellular space in addition to endocytic compartment in the secreting cells (Panakova, Sprong et al. 2005). As a result, extracellular Wg staining is a powerful tool for solely investigating long-range gradient, as Wg molecules responsible for short-range signaling are completely excluded from detection by this technique. Extracellular Wg staining results confirmed that in wls-overexpressing compartment of the wing disc, the range of Wg gradient has significantly expanded in comparison to the unaltered compartment. Further support for the expansion of a basally-maintained longrange Wg concentration gradient at the expense of apically-accumulated Wg 63
76 molecules that induce short-range signaling, comes from direct investigation of signaling events. Upon wls overexpression, Dll a long-range target of Wg signaling was detected over a wider field of cells than it is normally expressed. This observation indicates an increase in long-range Wg signaling. At the same time, the wing nick phenotype suggested that the short-range Wg signaling had been attenuated at least to some extent. All these data supports the notion that Wls is able to change the shape of Wg concentration gradient so that in the presence of excessive Wls, a reduction in the amount of Wg near the source, together with a flatter slope of the concentration curve results into the activation of long-range targets of Wg signaling over an extended region. 4.2 Inter-related functions of Wls and vacuolar acidification in Wg secretion provides insight into Wg intracellular trafficking Experiments with bafilomycin and RNAi constructs against V- ATPase subunits demonstrated that acidification is necessary for Wg secretion. This requirement for acidification can be surpassed by wls overexpression. As already mentioned, Wls structure is expected to contain a lipocalin fold (Coombs, Yu et al. 2010) and at least one lipocalin family member has been reported to oligomerize in acidic phs (Arnoux, Morosinotto et al. 2009). Experiments conducted in our lab have revealed that Wls forms oligomers (Sencer Tektas, unpublished data) and it seems reasonable to suggest that acidification can enhance Wls oligomerization, as well. Other studies have shown that apical/basal sorting of proteins could be determined by their oligomerization status. In MDCK cells, for example, it has been 64
77 reported that cholesterol addition to the cells can drive the oligomerization and consequent apical sorting of GFP-PrP constructs, which under control conditions do not oligomerise and are basolaterally sorted (Paladino, Lebreton et al. 2008). Based on these data, I propose the following model for Wg apical to basal transcytosis in Drosophila wing disc: Wls proteins escort Wg molecules out of the ER and direct them first to the apical membrane. At this point Wls molecules are monomeric and bound to lipid-modified Wg proteins (Figure 4.1). Wg-Wls complex is then internalized from the apical membrane and passes through the endosomal compartments. Inside these compartments, Wls experiences an acidic environment, which leads to its oligomerization and basal sorting. As a result, when vacuolar acidification is altered, Wls is unable to oligomerize and would be retained at the apical surface. However, when an excessive amount of Wls is present inside the cell, it would be easier for them to make oligomers; which is why wls-overexpressing cells are more resistant to V-ATPase inhibitors. It also explains why more Wg is directed to the basal membrane in wlsoverexpressing tissue. Again, excess Wls drives oligomer formation and Wls oligomers would act as basal sorting signals guiding more Wg to the basolateral membrane for secretion and long-range gradient formation. 65
.")
78 Figure 4.1 Proposed model for Wg secretion from producing cells Wls proteins escort Wg molecules to the apical membrane while still monomerized. For simplicity, Wg molecules are shown extracellular here. However, data suggests that they are present in an undefined intracellular compartment (for more discussion refer to the text). After endocytosis from the apical membrane, Wls proteins experience an acidic environment and oligomerize. This oligomerisation directs them to the basolateral membrane. Adopted from(cadigan and Peifer 2009) 66
Pattern formation: Wingless on the move Robert Howes and Sarah Bray
 R222 Dispatch Pattern formation: Wingless on the move Robert Howes and Sarah Bray Wingless is a key morphogen in Drosophila. Although it is evident that Wingless acts at a distance from its site of synthesis,
R222 Dispatch Pattern formation: Wingless on the move Robert Howes and Sarah Bray Wingless is a key morphogen in Drosophila. Although it is evident that Wingless acts at a distance from its site of synthesis,
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
AT THE EDGE OF DEVELOPMENTAL BIOLOGY: ADVANCES AND MYSTERIES ABOUT THE WNT GENES
 AT THE EDGE OF DEVELOPMENTAL BIOLOGY: ADVANCES AND MYSTERIES ABOUT THE WNT GENES Bénédicte Sanson, University of Cambridge, Department of Genetics, Downing Site, Cambridge CB2 3EH, UK, bs251@mole.bio.cam.ac.uk.
AT THE EDGE OF DEVELOPMENTAL BIOLOGY: ADVANCES AND MYSTERIES ABOUT THE WNT GENES Bénédicte Sanson, University of Cambridge, Department of Genetics, Downing Site, Cambridge CB2 3EH, UK, bs251@mole.bio.cam.ac.uk.
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Reception The target cell s detection of a signal coming from outside the cell May Occur by: Direct connect Through signal molecules
 Why Do Cells Communicate? Regulation Cells need to control cellular processes In multicellular organism, cells signaling pathways coordinate the activities within individual cells that support the function
Why Do Cells Communicate? Regulation Cells need to control cellular processes In multicellular organism, cells signaling pathways coordinate the activities within individual cells that support the function
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
!"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%%
*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%%") !"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%% !"#$%&'(")*++*%,*'-&'./%/,*#01#%-2)#3&)/% 4'(")*++*% % %5"0)%-2)#3&) %%% %67'2#72'*%%%%%%%%%%%%%%%%%%%%%%%4'(")0/./% % 8$+&'&,+"/7 % %,$&7&/9)7$*/0/%%%%%%%%%%
!"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%% !"#$%&'(")*++*%,*'-&'./%/,*#01#%-2)#3&)/% 4'(")*++*% % %5"0)%-2)#3&) %%% %67'2#72'*%%%%%%%%%%%%%%%%%%%%%%%4'(")0/./% % 8$+&'&,+"/7 % %,$&7&/9)7$*/0/%%%%%%%%%%
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Cell Biology Review. The key components of cells that concern us are as follows: 1. Nucleus
 Cell Biology Review Development involves the collective behavior and activities of cells, working together in a coordinated manner to construct an organism. As such, the regulation of development is intimately
Cell Biology Review Development involves the collective behavior and activities of cells, working together in a coordinated manner to construct an organism. As such, the regulation of development is intimately
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Lesson Overview. Gene Regulation and Expression. Lesson Overview Gene Regulation and Expression
 13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
Regulation of Wingless Secretion, Distribution and Signaling
 Regulation of Wingless Secretion, Distribution and Signaling A dissertation submitted to the Division of Research and Advanced Studies of the University of Cincinnati in partial fulfillment of the requirements
Regulation of Wingless Secretion, Distribution and Signaling A dissertation submitted to the Division of Research and Advanced Studies of the University of Cincinnati in partial fulfillment of the requirements
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Explain how cell size and shape affect the overall rate of nutrient intake and the rate of waste elimination. [LO 2.7, SP 6.2]
![Explain how cell size and shape affect the overall rate of nutrient intake and the rate of waste elimination. [LO 2.7, SP 6.2]](/thumbs/77/76271213.jpg "Explain how cell size and shape affect the overall rate of nutrient intake and the rate of waste elimination. [LO 2.7, SP 6.2]") Cells Learning Objectives Use calculated surface area-to-volume ratios to predict which cell(s) might eliminate wastes or procure nutrients faster by diffusion. [LO 2.6, SP 2.2] Explain how cell size and
Cells Learning Objectives Use calculated surface area-to-volume ratios to predict which cell(s) might eliminate wastes or procure nutrients faster by diffusion. [LO 2.6, SP 2.2] Explain how cell size and
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
Morphogens in biological development: Drosophila example
 LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
Wingless, Hedgehog and Heparan Sulfate Proteoglycans
 2509 Wingless, Hedgehog and Heparan Sulfate Proteoglycans Several recent papers published in Development challenge some of the previous views published by our group and others that: (1) the Drosophila
2509 Wingless, Hedgehog and Heparan Sulfate Proteoglycans Several recent papers published in Development challenge some of the previous views published by our group and others that: (1) the Drosophila
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Honors Biology Reading Guide Chapter 11
 Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
The neuron as a secretory cell
 The neuron as a secretory cell EXOCYTOSIS ENDOCYTOSIS The secretory pathway. Transport and sorting of proteins in the secretory pathway occur as they pass through the Golgi complex before reaching the
The neuron as a secretory cell EXOCYTOSIS ENDOCYTOSIS The secretory pathway. Transport and sorting of proteins in the secretory pathway occur as they pass through the Golgi complex before reaching the
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Tuesday, December 27, 16
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
S1 Gene ontology (GO) analysis of the network alignment results
 analysis of the network alignment results") 1 Supplementary Material for Effective comparative analysis of protein-protein interaction networks by measuring the steady-state network flow using a Markov model Hyundoo Jeong 1, Xiaoning Qian 1 and
1 Supplementary Material for Effective comparative analysis of protein-protein interaction networks by measuring the steady-state network flow using a Markov model Hyundoo Jeong 1, Xiaoning Qian 1 and
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins
 13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
CELB40060 Membrane Trafficking in Animal Cells. Prof. Jeremy C. Simpson. Lecture 2 COPII and export from the ER
 CELB40060 Membrane Trafficking in Animal Cells Prof. Jeremy C. Simpson Lecture 2 COPII and export from the ER Today s lecture... The COPII coat - localisation and subunits Formation of the COPII coat at
CELB40060 Membrane Trafficking in Animal Cells Prof. Jeremy C. Simpson Lecture 2 COPII and export from the ER Today s lecture... The COPII coat - localisation and subunits Formation of the COPII coat at
DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS. Scientific Background on the Nobel Prize in Medicine 2013
 DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS Scientific Background on the Nobel Prize in Medicine 2013 Daniela Scalet 6/12/2013 The Nobel Prize in Medicine
DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS Scientific Background on the Nobel Prize in Medicine 2013 Daniela Scalet 6/12/2013 The Nobel Prize in Medicine
Cellular Neuroanatomy I The Prototypical Neuron: Soma. Reading: BCP Chapter 2
 Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
BE 159: Signal Transduction and Mechanics in Morphogenesis
 BE 159: Signal Transduction and Mechanics in Morphogenesis Justin Bois Caltech Winter, 2018 2018 Justin Bois. This work is licensed under a Creative Commons Attribution License CC-BY 4.0. 5 Delta-Notch
BE 159: Signal Transduction and Mechanics in Morphogenesis Justin Bois Caltech Winter, 2018 2018 Justin Bois. This work is licensed under a Creative Commons Attribution License CC-BY 4.0. 5 Delta-Notch
16 CONTROL OF GENE EXPRESSION
 16 CONTROL OF GENE EXPRESSION Chapter Outline 16.1 REGULATION OF GENE EXPRESSION IN PROKARYOTES The operon is the unit of transcription in prokaryotes The lac operon for lactose metabolism is transcribed
16 CONTROL OF GENE EXPRESSION Chapter Outline 16.1 REGULATION OF GENE EXPRESSION IN PROKARYOTES The operon is the unit of transcription in prokaryotes The lac operon for lactose metabolism is transcribed
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis
 18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Robustness of Tissue Patterns*
 MCBU Project II - 2014 Robustness of Tissue Patterns* June, 2014 Frederic Y.M. Wan Mathematics University of California, Irvine Supported by: NIH Grants R01-GM67247 P50-GM66051 Biological Patterning The
MCBU Project II - 2014 Robustness of Tissue Patterns* June, 2014 Frederic Y.M. Wan Mathematics University of California, Irvine Supported by: NIH Grants R01-GM67247 P50-GM66051 Biological Patterning The
Axis determination in flies. Sem 9.3.B.5 Animal Science
 Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
CHAPTER 3. Cell Structure and Genetic Control. Chapter 3 Outline
 CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
Introduction. Wnt signaling in development
 Introduction Wnt signaling in development 7 INTRODUCTION Abstract The complex cell fate determinations and morphogenetic movements that generate the metazoan body plan are controlled by only a handful
Introduction Wnt signaling in development 7 INTRODUCTION Abstract The complex cell fate determinations and morphogenetic movements that generate the metazoan body plan are controlled by only a handful
Reading Assignments. A. Genes and the Synthesis of Polypeptides. Lecture Series 7 From DNA to Protein: Genotype to Phenotype
 Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
2. In regards to the fluid mosaic model, which of the following is TRUE?
 General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Bio 127 Section I Introduction to Developmental Biology. Cell Cell Communication in Development. Developmental Activities Coordinated in this Way
 Bio 127 Section I Introduction to Developmental Biology Cell Cell Communication in Development Gilbert 9e Chapter 3 It has to be EXTREMELY well coordinated for the single celled fertilized ovum to develop
Bio 127 Section I Introduction to Developmental Biology Cell Cell Communication in Development Gilbert 9e Chapter 3 It has to be EXTREMELY well coordinated for the single celled fertilized ovum to develop
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Conclusions. The experimental studies presented in this thesis provide the first molecular insights
 C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
Biological Process Term Enrichment
 Biological Process Term Enrichment cellular protein localization cellular macromolecule localization intracellular protein transport intracellular transport generation of precursor metabolites and energy
Biological Process Term Enrichment cellular protein localization cellular macromolecule localization intracellular protein transport intracellular transport generation of precursor metabolites and energy
Protein Sorting, Intracellular Trafficking, and Vesicular Transport
 Protein Sorting, Intracellular Trafficking, and Vesicular Transport Noemi Polgar, Ph.D. Department of Anatomy, Biochemistry and Physiology Email: polgar@hawaii.edu Phone: 692-1422 Outline Part 1- Trafficking
Protein Sorting, Intracellular Trafficking, and Vesicular Transport Noemi Polgar, Ph.D. Department of Anatomy, Biochemistry and Physiology Email: polgar@hawaii.edu Phone: 692-1422 Outline Part 1- Trafficking
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION
 MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
Biol403 - Receptor Serine/Threonine Kinases
 Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
RANK. Alternative names. Discovery. Structure. William J. Boyle* SUMMARY BACKGROUND
 RANK William J. Boyle* Department of Cell Biology, Amgen, Inc., One Amgen Center Drive, Thousand Oaks, CA 91320-1799, USA * corresponding author tel: 805-447-4304, fax: 805-447-1982, e-mail: bboyle@amgen.com
RANK William J. Boyle* Department of Cell Biology, Amgen, Inc., One Amgen Center Drive, Thousand Oaks, CA 91320-1799, USA * corresponding author tel: 805-447-4304, fax: 805-447-1982, e-mail: bboyle@amgen.com
Developmental roles of heparan sulfate proteoglycans in Drosophila
 Glycoconjugate Journal 19, 363 368, 2003 C 2003 Kluwer Academic Publishers. Manufactured in The Netherlands. Developmental roles of heparan sulfate proteoglycans in Drosophila Xinhua Lin 1 and Norbert
Glycoconjugate Journal 19, 363 368, 2003 C 2003 Kluwer Academic Publishers. Manufactured in The Netherlands. Developmental roles of heparan sulfate proteoglycans in Drosophila Xinhua Lin 1 and Norbert
Course Outcomes Guide (COG)
") Course Outcomes Guide (COG) Directions: Please complete this form to document your progress toward improving student learning. For each item, indicate your progress and your anticipated next steps. Thank
Course Outcomes Guide (COG) Directions: Please complete this form to document your progress toward improving student learning. For each item, indicate your progress and your anticipated next steps. Thank
Cells to Tissues. Peter Takizawa Department of Cell Biology
 Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 2: Phagocytosis Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research Salmonella Gram negative bacteria
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 2: Phagocytosis Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research Salmonella Gram negative bacteria
Advanced Higher Biology. Unit 1- Cells and Proteins 2c) Membrane Proteins
 Membrane Proteins") Advanced Higher Biology Unit 1- Cells and Proteins 2c) Membrane Proteins Membrane Structure Phospholipid bilayer Transmembrane protein Integral protein Movement of Molecules Across Membranes Phospholipid
Advanced Higher Biology Unit 1- Cells and Proteins 2c) Membrane Proteins Membrane Structure Phospholipid bilayer Transmembrane protein Integral protein Movement of Molecules Across Membranes Phospholipid
3/8/ Complex adaptations. 2. often a novel trait
 Chapter 10 Adaptation: from genes to traits p. 302 10.1 Cascades of Genes (p. 304) 1. Complex adaptations A. Coexpressed traits selected for a common function, 2. often a novel trait A. not inherited from
Chapter 10 Adaptation: from genes to traits p. 302 10.1 Cascades of Genes (p. 304) 1. Complex adaptations A. Coexpressed traits selected for a common function, 2. often a novel trait A. not inherited from
Chapter 1. DNA is made from the building blocks adenine, guanine, cytosine, and. Answer: d
 Chapter 1 1. Matching Questions DNA is made from the building blocks adenine, guanine, cytosine, and. Answer: d 2. Matching Questions : Unbranched polymer that, when folded into its three-dimensional shape,
Chapter 1 1. Matching Questions DNA is made from the building blocks adenine, guanine, cytosine, and. Answer: d 2. Matching Questions : Unbranched polymer that, when folded into its three-dimensional shape,
Zool 3200: Cell Biology Exam 5 4/27/15
 Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016
 Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Chapter 18 Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
The Imperatives of Context and Contour for Morphogen Dispersion
 2252 Biophysical Journal Volume 103 December 2012 2252 2256 The Imperatives of Context and Contour for Morphogen Dispersion Thomas B. Kornberg* Cardiovascular Research Institute, University of California,
2252 Biophysical Journal Volume 103 December 2012 2252 2256 The Imperatives of Context and Contour for Morphogen Dispersion Thomas B. Kornberg* Cardiovascular Research Institute, University of California,
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Cell biology: Death drags down the neighbourhood
 Cell biology: Death drags down the neighbourhood The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published Publisher Vasquez,
Cell biology: Death drags down the neighbourhood The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published Publisher Vasquez,
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Cell Death & Trophic Factors II. Steven McLoon Department of Neuroscience University of Minnesota
 Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
2011 The Simple Homeschool Simple Days Unit Studies Cells
 1 We have a full line of high school biology units and courses at CurrClick and as online courses! Subscribe to our interactive unit study classroom and make science fun and exciting! 2 A cell is a small
1 We have a full line of high school biology units and courses at CurrClick and as online courses! Subscribe to our interactive unit study classroom and make science fun and exciting! 2 A cell is a small
Gene Control Mechanisms at Transcription and Translation Levels
 Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
ADAM FAMILY. ephrin A INTERAZIONE. Eph ADESIONE? PROTEOLISI ENDOCITOSI B A RISULTATO REPULSIONE. reverse. forward
 ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
5- Semaphorin-Plexin-Neuropilin
 5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
5- Semaphorin-Plexin-Neuropilin 1 SEMAPHORINS-PLEXINS-NEUROPILINS ligands receptors co-receptors semaphorins and their receptors are known signals for: -axon guidance -cell migration -morphogenesis -immune
Wntless in Wnt secretion: molecular, cellular and genetic aspects
 Front. Biol. DOI 10.1007/s11515-012-1200-8 REVIEW Wntless in Wnt secretion: molecular, cellular and genetic aspects Soumyashree DAS, Shiyan YU, Ryotaro SAKAMORI, Ewa Stypulkowski, Nan GAO ( ) Department
Front. Biol. DOI 10.1007/s11515-012-1200-8 REVIEW Wntless in Wnt secretion: molecular, cellular and genetic aspects Soumyashree DAS, Shiyan YU, Ryotaro SAKAMORI, Ewa Stypulkowski, Nan GAO ( ) Department
TITLE: The Role(s) of Heparan Sulfate Proteoglycan(s) in the wnt- 1 Signaling Pathway
 of Heparan Sulfate Proteoglycan(s) in the wnt- 1 Signaling Pathway") ^ *H AD Award Number: DAMD17-97-1-7220 TITLE: The Role(s) of Heparan Sulfate Proteoglycan(s) in the wnt- 1 Signaling Pathway PRINCIPAL INVESTIGATOR: Xinhua Lin, Ph.D. CONTRACTING ORGANIZATION: Harvard
^ *H AD Award Number: DAMD17-97-1-7220 TITLE: The Role(s) of Heparan Sulfate Proteoglycan(s) in the wnt- 1 Signaling Pathway PRINCIPAL INVESTIGATOR: Xinhua Lin, Ph.D. CONTRACTING ORGANIZATION: Harvard
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018
 Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Cell (Learning Objectives)
") Cell (Learning Objectives) 1. Understand & describe the basic components necessary for a functional cell. 2. Review the order of appearance of cells on earth and explain the endosymbiotic theory. 3. Compare
Cell (Learning Objectives) 1. Understand & describe the basic components necessary for a functional cell. 2. Review the order of appearance of cells on earth and explain the endosymbiotic theory. 3. Compare
PRACTICE EXAM. 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos.
 PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
BILD7: Problem Set. 2. What did Chargaff discover and why was this important?
 BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290
 PLoS Biol 4(9): e290") Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
MOLECULAR CELL BIOLOGY
 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
Exam 2 ID#: November 9, 2006
 Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
5/4/05 Biol 473 lecture
 5/4/05 Biol 473 lecture animals shown: anomalocaris and hallucigenia 1 The Cambrian Explosion - 550 MYA THE BIG BANG OF ANIMAL EVOLUTION Cambrian explosion was characterized by the sudden and roughly simultaneous
5/4/05 Biol 473 lecture animals shown: anomalocaris and hallucigenia 1 The Cambrian Explosion - 550 MYA THE BIG BANG OF ANIMAL EVOLUTION Cambrian explosion was characterized by the sudden and roughly simultaneous
Genes, Development, and Evolution
 14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
Old FINAL EXAM BIO409/509 NAME. Please number your answers and write them on the attached, lined paper.
 Old FINAL EXAM BIO409/509 NAME Please number your answers and write them on the attached, lined paper. Gene expression can be regulated at several steps. Describe one example for each of the following:
Old FINAL EXAM BIO409/509 NAME Please number your answers and write them on the attached, lined paper. Gene expression can be regulated at several steps. Describe one example for each of the following:
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
Nervous Systems: Neuron Structure and Function
 Nervous Systems: Neuron Structure and Function Integration An animal needs to function like a coherent organism, not like a loose collection of cells. Integration = refers to processes such as summation
Nervous Systems: Neuron Structure and Function Integration An animal needs to function like a coherent organism, not like a loose collection of cells. Integration = refers to processes such as summation
Components of a functional cell. Boundary-membrane Cytoplasm: Cytosol (soluble components) & particulates DNA-information Ribosomes-protein synthesis
 & particulates DNA-information Ribosomes-protein synthesis") Cell (Outline) - Components of a functional cell - Major Events in the History of Earth: abiotic and biotic phases; anaerobic and aerobic atmosphere - Prokaryotic cells impact on the biosphere - Origin
Cell (Outline) - Components of a functional cell - Major Events in the History of Earth: abiotic and biotic phases; anaerobic and aerobic atmosphere - Prokaryotic cells impact on the biosphere - Origin
Transport between cytosol and nucleus
 of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
Plant Stimuli pp Topic 3: Plant Behaviour Ch. 39. Plant Behavioural Responses. Plant Hormones. Plant Hormones pp
 Topic 3: Plant Behaviour Ch. 39 Plants exist in environments that are constantly changing. Like animals, plants must be able to detect and react to stimuli in the environment. Unlike animals, plants can
Topic 3: Plant Behaviour Ch. 39 Plants exist in environments that are constantly changing. Like animals, plants must be able to detect and react to stimuli in the environment. Unlike animals, plants can
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Overview of Cells. Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory
 Overview of Cells Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory Prokaryotic Cells Archaea Bacteria Come in many different shapes and sizes.5 µm 2 µm, up to 60 µm long Have large
Overview of Cells Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory Prokaryotic Cells Archaea Bacteria Come in many different shapes and sizes.5 µm 2 µm, up to 60 µm long Have large
The Wingless morphogen gradient is established by the cooperative action of Frizzled and Heparan Sulfate Proteoglycan receptors
 Developmental Biology 276 (2004) 89 100 www.elsevier.com/locate/ydbio The Wingless morphogen gradient is established by the cooperative action of Frizzled and Heparan Sulfate Proteoglycan receptors Gyeong-Hun
Developmental Biology 276 (2004) 89 100 www.elsevier.com/locate/ydbio The Wingless morphogen gradient is established by the cooperative action of Frizzled and Heparan Sulfate Proteoglycan receptors Gyeong-Hun