Biochemistry, The University of Western Australia, Crawley WA 6009, Australia.
|
|
|
- Catherine Russell
- 5 years ago
- Views:
Transcription
1 Manuscript (MUST INCLUDE TITLE PAGE AND ABSTRACT) Click here to download Manuscript (MUST INCLUDE TITLE PAGE AND ABSTRACT): Lopez et al revised2.docx 1 Estrogen-mediated regulation of mitochondrial gene expression 2 3 Maria I.G. Lopez Sanchez 1, Anne-Marie J. Shearwood 1, Tiong-Sun Chia 1, Stefan M.K. Davies 1, Oliver Rackham 1, 2 1, 2# and Aleksandra Filipovska Harry Perkins Institute of Medical Research and Centre for Medical Research, Level 7 QQ Block, QEII Medical Centre, 6 Verdun Street, Nedlands WA 6009 and 2 School of Chemistry and Biochemistry, The University of Western Australia, Crawley WA 6009, Australia # Correspondence to: Aleksandra Filipovska. Address: The School of Chemistry and Biochemistry and Harry Perkins Institute for Medical Research, Level 7 QQ Block, QEII Medical Centre, 6 Verdun Street, Nedlands WA 6009, Australia. aleksandra.filipovska@uwa.edu.au Abbreviated title: Estrogen affects mitochondrial gene expression 12 Keywords: Estrogen, estrogen receptor alpha, gene expression, RNA, mitochondria. 13 Abbreviations: E2, 17β-estradiol; ERα, estrogen receptor alpha. 14 Word count (excluding abstract, figure captions and references): This project was supported by fellowships and project grants (to A.F and O.R.) from the National Health and Medical Research Council (APP , APP , APP , APP , APP ) and the Australian Research Council (FT , FT , DP ) DISCLOSURE STATEMENT: The authors have nothing to disclose
2 Abstract Estrogens, in particular 17β-estradiol, are well known regulators of essential cellular functions; however, discrepancies remain over the mechanisms by which they act on mitochondria. Here, we propose a novel mechanism for direct regulation of mitochondrial gene expression by estrogen under metabolic stress. We show that in serum-depleted medium estrogen stimulates a rapid relocation of estrogen receptor alpha (ERα) to mitochondria, where it elicits a cellular response resulting in an increase in mitochondrial RNA abundance. Mitochondrial RNA levels are regulated through the association of ERα with hydroxysteroid (17-β) dehydrogenase 10 (HSD17B10), a multifunctional protein involved in steroid metabolism that is also a core subunit of the mitochondrial RNase P complex responsible for the cleavage of mitochondrial polycistronic transcripts. Processing of mitochondrial transcripts affects mitochondrial gene expression by controlling the levels of mature RNAs available for translation. This work provides the first mechanism linking RNA processing and estrogen activation in mitochondrial gene expression and underscores the coordinated response between the nucleus and mitochondria in response to stress
3 Introduction Estrogens, in particular 17β-estradiol (E2), are responsible for the regulation of key cellular processes such as proliferation and apoptosis, inducing changes in nuclear gene expression or by activating cell signaling pathways (1,2). Estrogen receptors (ERs) can function as receptors and transcription factors upon E2 exposure in the nucleus and as signaling molecules in the plasma membrane (3). In the nucleus, ERs regulate the expression of E2-responsive genes through binding to estrogen-responsive elements and inducing changes in transcription (4). In the plasma membrane ERs participate in rapid signal transduction mechanisms and activate G proteins, leading to calcium and cyclic adenosine monophosphate generation and the activation of proximal and distal kinases, which results in the phosphorylation of proteins involved in the modulation of cell migration, survival and proliferation (4,5) More recently, research has focused on the effects of estrogen on mitochondrial function, as mitochondria are involved in crucial cellular processes in health and in disease. It has been reported that E2 treatment increases the abundance of mitochondrially encoded RNAs that results in elevated respiration (6,7), indicating that E2 plays a role in the modulation of mitochondrial gene expression. It has been suggested that E2 effects on mitochondrial function are indirectly mediated by transcription factors acting on nuclear genes that regulate mitochondrial gene expression (7). In particular, it has been suggested that E2 stimulates the nuclear respiratory factor 1 (NRF-1), promoting the transcription of mitochondrial transcription factor A (TFAM), which subsequently increases mitochondrial DNA (mtdna) transcription (7,8). The localization of ERs to mitochondria in cells has initiated interest in their specific roles within these organelles. ERs have been localized to mitochondria of several cell lines including the breast cancer-derived MCF-7, and hepatoma-derived HepG2 cells (9-11), where they have been suggested to act as transcription factors affecting mitochondrial gene expression directly (11-13). Other studies have suggested that the interaction between ERs and putative estrogen-responsive elements in mitochondrial DNA is responsible for the induction of mitochondrial transcription by E2 (14,15). There are different modes suggested by which 3
4 73 74 estrogens exert their effects on the mitochondrial genome; however a general agreement over the mechanism by which ERs may affect mitochondrial gene expression is lacking Most of the genetic information necessary for mitochondrial function is encoded by the nuclear genome whereas mitochondrial DNA encodes 11 mrnas, 2 rrnas and 22 trnas that are transcribed as long polycistronic precursor transcripts from both DNA strands (16). Despite their common polycistronic origin, there is wide variation in the abundance of individual mitochondrial trnas, mrnas, and rrnas in different tissues and cell types (17). This highlights the importance of post-transcriptional processing mechanisms in the regulation of mitochondrial gene expression. RNA processing is a crucial stage in the regulation of mitochondrial RNA metabolism and consequently mitochondrial function (18-20). Two nuclear DNA-encoded protein complexes account for the majority of the cleavage events involved in mitochondrial RNA processing, the mitochondrial RNase P complex that cleaves the 5' end of trnas and (18) the mitochondrial RNase Z, encoded by the ELAC2 gene, which cleaves the 3' ends of trnas (19,20) The mitochondrial RNase P complex is composed of three proteins: trna methyltransferase 10 C (TRMT10C), (17-β) dehydrogenase 10 (HSD17B10) and proteinaceous RNase P (PRORP) that are all necessary for efficient cleavage of the 5' trna ends. TRMT10C and HSD17B10 have additional functions suggesting that recruitment into the RNase P complex is an additional role for these proteins (20-22). Recently it was shown that TRMT10C, also known as mitochondrial RNase P protein 1 (MRPP1), is a methyltransferase responsible for a common methylation modification of trnas and it may be responsible for RNA-recognition for RNase P cleavage (20,21). HSD17B10 is a member of the short-chain dehydrogenase/reductase family known under a range of names including MRPP2, ABAD, ERAB, HADH2, MHBD or SCHAD (Table 1). It has been described as a multifunctional protein that has a NAD + -binding domain, although this is not required for RNase P activity (21). In addition, reports suggest that HSD17B10 participates in the degradation of the amino acid isoleucine via its 2-methyl-3-hydroxybutyryl-CoA dehydrogenase activity (23,24). PRORP, also known as mitochondrial RNase P protein 3 (MRPP3), belongs to the mammalian pentatricopeptide repeat (PPR) domain protein family (25). PRORP is composed of a PPR domain and a putative metallonuclease 4
5 domain that is hypothesized to harbor the catalytic site of the RNase P complex. In mammalian mitochondria it was shown that ELAC2 is the mitochondrial RNase Z responsible for processing the 3' end of trnas and that it associates with PTCD1, another PPR protein that also acts as a negative regulator of leucine trnas (20,26,27) Interestingly, HSD17B10 catalyzes the conversion of 17β-estradiol to the less potent estrogen metabolite estrone in mitochondria (28). It has previously been reported that the estrogen receptor alpha (ERα) interacts with HSD17B10 in a bacterial two-hybrid screen (29), however, no further investigation into the relevance of this interaction has been carried out. Therefore we investigated the biological significance of the interaction between HSD17B10 and ERα in mitochondria of estrogenresponsive breast adenocarcinoma MCF-7 cells and how this affects mitochondrial gene expression. Here we show that upon exposure to E2, ERα rapidly localizes to mitochondria, where it interacts with HSD17B10 to modulate the expression of mitochondrial RNA transcripts. We show that changes in the expression of proteins involved in the processing of mitochondrial transcripts can have profound effects on mitochondrial gene expression, by affecting the levels of mature species, the final processing of the different RNAs, and overall protein synthesis in the presence of estrogen
6 124 Materials and Methods 125 Cell culture and estrogen treatment Estrogen-responsive MCF-7, and ER-negative MDA-468 and MDA-231 human breast adenocarcinoma cells were grown at 37 C under humidified 95% air/5% CO 2 in Dulbecco s phenol red-free modified Eagle medium (DMEM, Invitrogen) supplemented with 25 mm glucose, 1mM pyruvate, 2 mm glutamine, uridine (0.05 g.l -1 ), penicillin (0.1 U.l -1 ), streptomycin sulphate (0.1 g.l -1 ) and 10% fetal bovine serum (FBS). To evaluate the effects of 17β-estradiol (E2), cells were cultured in serum-depleted medium containing 5% charcoal-stripped FBS for 6 days prior to treatment with 100 nm estradiol in ethanol (Sigma-Aldrich). 133 Mammalian expression plasmids All expression vectors were based on pcdna3 (Invitrogen). Full length human TRMT10C (NCBI accession number NP_ ), HSD17B10 (NCBI accession number NP_ ), PRORP (NCBI accession number NP_ ), PTCD1 (NCBI accession number NP_ ), ELAC2 (NCBI accession number NP_ ) or ERα (NCBI accession number NP_ ) were expressed with their native termination codon or fused to a tandem affinity purification tag (TAP tag, ABO76910) or EGFP (BD Biosciences) at the C-terminus. All plasmids were tested for expression by transfection and immunoblotting. 141 sirna and plasmid transfections MCF-7 cells were plated at 60% confluence in 6-well plates or 10 cm dishes and transfected with annealed sirnas in OptiMEM medium (Invitrogen) according to manufacturer's instructions. 125 nm (for 6-well plates) or 145 nm (for 10 cm dishes) of TRMT10C, HSD17B10, PRORP, PTCD1, ELAC2 or non-targeting (NT) control sirna (Thermo Scientific) was transfected using Lipofectamine 2000 (Invitrogen). Cells were incubated 72 h following transfection and then re-transfected for an additional 72 h to enhance knockdown efficiency, followed by a further incubation in the absence or presence of 100 nm E2 for 18 h and then harvested. 6
7 MCF-7 cells were plated at 60% confluence in 10 cm dishes in serum-depleted medium and transfected with mammalian expression plasmids in OptiMEM medium (Invitrogen) containing puromycin (1µg.mlˉ¹) to maintain plasmid expression. 158 ng/cm 2 of TRMT10C-TAP, HSD17B10- TAP, PRORP-TAP, PTCD1-TAP, ELAC2-TAP, ERα-TAP or control EGFP-TAP plasmid DNA was transfected using Fugene HD (Roche). Cells were incubated for 48 h following transfection to allow integration and expression of the plasmid, in serum-depleted medium containing puromycin (1µg.mlˉ¹) to maintain plasmid expression. This was followed by a further incubation in the absence or presence of 100 nm E2 for 18 h before cells were harvested. 157 Mitochondrial isolation Mitochondria were prepared from 2 x 10 7 MCF-7 cells grown in basal, non serum-depleted or serumdepleted medium or transfected with sirna and treated with E2 (as described above). Cells in PBS were collected by centrifugation at 150 g for 5 min at 4 C and the pellet was suspended in 4 ml of icecold 10 mm NaCl, 1.5 mm MgCl 2 and 10 mm Tris-HCl, ph 7.5. Cells were allowed to swell for 5 min on ice and briefly homogenized with a 7 ml glass homogenizer. The nuclei from this suspension were sedimented at 1300 g for 3 min at 4 C, and this centrifugation step was repeated for the supernatant once more. The mitochondria from this suspension were sedimented (10,000 g for 10 min at 4 C) and suspended in 120 mm KCl, 10 mm HEPES, 1mM EGTA. Protein concentration was determined by the bicinchoninic acid assay as described previously (30). 167 Mitochondrial subfractionation Mitochondria were prepared from MCF-7 cells grown in serum-depleted medium and treated with E2 for 18 h as described above. Protein concentration was normalized to 100 mg.mlˉ¹ in STE containing 0.1% (w/v) fat-free BSA (STE/BSA) and determined by the bicinchoninic acid assay. The mitochondrial outer membrane was solubilized by adding 0.2 mg digitonin (12.5 mg.mlˉ¹ in STE) /mg protein to the mitochondrial preparation followed by incubation for 15 min at 4 C. Three volumes STE/BSA were added to each sample, mixed and gently homogenized using micro pestles (Ambion). Mitoplasts were sedimented by centrifugation (10,000 g for 10 min at 4 C) and the pellet was 7
8 suspended in 20 µl STE/BSA, while the supernatant was retained for further fractionation. The mitochondrial outer membrane fraction was collected as a pellet following centrifugation (144,000 g for 20 min at 4 C) and suspended in 70 µl STE containing 1% lauryl maltoside representing the membrane, insoluble fraction. The supernatant from this centrifugation step contained the soluble fraction. 180 Immunoblotting Protein expression was detected by immunoblotting using Sigma-Aldrich antibodies according to manufacturer s recommendations: TRMT10C (Cat. No. HPA036671), HSD17B10 (Cat. No. HPA001432), PRORP (Cat. No. HPA020459), PTCD1 (Cat. No. HPA020106), ELAC2 (Cat. No. HPA019535), ERα (Cat. No. HPA000450). Specific mitochondrial proteins were detected using MitoSciences antibodies: COX1 (Cat. No. MS404), COX2 (Cat. No. MS405) and complex II subunit 70 KDa, SDHA (Cat. No. MS204). Porin (Abcam, Cat. No. ab15895) was used as a control to confirm normalized protein loading. The primary antibodies were detected using the Odyssey Infrared IRDye 680RD goat anti-rabbit or IRDye 800CW goat anti-mouse secondary antibodies (Li-Cor Biosciences) and visualized using the Odyssey imaging system (Li-Cor Biosciences). 190 RNA harvesting and northern blotting RNA (5 µg) was extracted from sirna-transfected MCF-7 cells, MDA-468 or MDA-231, followed by E2 treatment as described above, using the mirneasy RNA extraction kit (Qiagen) with a DNase I on-column digestion step included to remove DNA, according to manufacturer s instructions. Northern blotting was carried out as described in (31). The signal was detected using IRDye labelled streptavidin (Li-Cor Biosciences) diluted 1:10000 in 3x SSC, 5% SDS, 25 mm Na 2 HPO 4, ph 7.5 and visualized using the Odyssey (Li-Cor Biosciences) imaging system. 197 Quantitative RT-PCR The transcript abundance of mrnas of interest was measured on RNA isolated from MCF-7 cells following sirna treatment. cdna was prepared using ThermoScript reverse transcriptase 8
9 (Invitrogen) and random hexamers and used as a template in the subsequent PCR that was performed using a Corbett Rotorgene 3000 using Platinum UDG SYBR Green mastermix (Invitrogen) and normalized to 18S rrna. 203 Tandem affinity purification Tandem affinity purification was carried out from plasmid-transfected MCF-7 cell lysates previously treated with E2 as described previously (27,31). 206 Microscopy MCF-7 cells were plated onto 13 mm diameter glass coverslips and allowed to attach overnight. Before transfection, the cells were placed in serum-depleted medium as described above. Cells were transfected with ERα-EGFP or HSD17B10-EGFP plasmid for 48 h, followed by incubation with 50 nm MitoTracker Orange in PBS for 15 min. In experiments examining the effects of E2, the cells were cultured in serum-depleted medium for 6 days prior to E2 treatment. After washing with PBS, the cells were fixed with 4% paraformaldehyde in PBS for 30 min and washed again with PBS. Cells were mounted in DABCO/PVA medium and images were acquired using a Nikon Ellipse Ti fluorescent inverted microscope using a Nikon 60x or 100x objective. 215 Mitochondrial protein synthesis MCF-7 cells were grown in six-well plates, transfected, incubated in the presence or absence of E2 and de novo protein synthesis was analyzed as described in (32). 218 Mitochondrial respiration Mitochondrial respiration using 0.5 mm N,N,N,N -Tetramethyl- p-phenylenediamine dihydrochloride (TMPD) and 2 mm ascorbate was measured in permeabilized cells as described in (20). 222 Native PAGE 9
10 Mitochondria were prepared from MCF-7 cells grown in serum-depleted medium and incubated in the presence or absence of 100 nm E2 for 18 h. Mitochondrial proteins were solubilized in sample buffer containing 1% dodecyl maltoside for 15 min followed by centrifugation ( g, 30 min at 4 C) and the supernatant resolved by one-dimensional native PAGE in the NativePAGE Novex Bis-Tris Gel System (Life Technologies). Electrophoresis was carried out in an XCell SureLock Mini-cell (Life Technologies) and followed by transfer to PVDF membrane (Immobilon FL, Millipore) using a Mini- Protean Trans blot system (Bio-Rad). Transfer was followed by incubation in Odyssey Blocking buffer (Li-Cor Biosciences) for 1 h, followed by overnight incubation with primary antibodies at 4 C. The primary antibodies were detected using the Odyssey Infrared IRDye 680RD goat anti-rabbit or IRDye 800CW goat anti-mouse secondary antibodies and visualized using the Odyssey imaging system (Li-Cor Biosciences). 234 Immunoblot densitometry and statistical analysis Band intensity from electronic (tiff) images of immunoblots was calculated by densitometry using Image J freeware (rsb.info.nih.gov/ij). Data are presented as the mean and standard deviation. The statistical significance was examined using a two-tailed Student s t-test
11 246 Results 247 HSD17B10 and ERα expression in MCF-7 cells Previous studies have indicated that the E2 effects on mitochondria are mediated by ERα (33), since ERα has been found associated with mitochondria in cells (9-11). To determine if there is an E2- dependent effect in the abundance of ERα the endogenous distribution of ERα was assessed by immunoblotting experiments carried out on mitochondria isolated from MCF-7 cells grown in basal, non serum-depleted medium, compared with mitochondria from cells grown in serum-depleted medium, followed by E2 treatment for 18h (Fig. 1A). We found ERα associated with mitochondria of MCF-7 cells independent of E2 treatment, however ERα is significantly enriched in mitochondria from cells grown in serum-depleted medium. This suggests that the serum-depletion may enhance the localization of ERα to mitochondria Mitochondria contain the HSD17B10 enzyme that is involved in E2 degradation and this protein has been suggested to interact with ERα in a bacterial two-hybrid system (29). The levels of this protein are not affected by growth in basal or serum-depleted medium (Fig. 1A). However, we wanted to gain further insight into the distribution of HSD17B10 and ERα in mitochondria and in response to E2. We carried out cell and mitochondrial subfractionation experiments to analyze the distribution of ERα and HSD17B10 in mitochondria from MCF-7 cells grown in basal (Fig. 1B) and serum-depleted medium (Fig. 1C), followed by incubation in the presence or absence of 100 nm E2 for 18h. Enriched fractions were obtained containing whole cell lysates, intact mitochondria, mitoplasts (mitochondria devoid of outer membrane), mitochondrial insoluble fraction (membranes) and mitochondrial soluble fraction. Immunoblotting for ERα and HSD17B10 indicates that both proteins are enriched in mitoplasts under normal and metabolic stress conditions, however, ERα has a significantly stronger association with mitochondria following serum starvation. 269 HSD17B10 interacts with ERα in MCF-7 cells Next we investigated possible association of ERα with HSD17B10 within mitochondria of MCF-7 cells. The C-terminus of HSD17B10 was fused to a tandem affinity purification (TAP) tag to isolate it 11
12 and determine if it associates with ERα by immunoblotting (Fig. 2). We found that endogenous ERα associated with HSD17B10-TAP, in the presence or absence of E2 (Fig. 2A), although the presence of E2 this association was weaker as suggested previously (29). These interactions were further assessed by reverse TAP, in which the C-terminus of ERα was fused to a TAP tag to determine if it interacts with HSD17B10 (Fig. 2B). Endogenous HSD17B10 was detected in association with ERα-TAP, and this association was weaker in the presence of E2. These results indicate that ERα associates with HSD17B10 and that this association can be affected by E2, validating a previous report of this interaction identified in a bacterial two-hybrid screen, hereby establishing the relevance of this interaction in mammalian cells. EGFP was used as a control to show specific antibody detection and that neither HSD17B10 (Fig. 2C) nor ERα (Fig. 2D) associate with EGFP, indicating the specificity of the interactions between ERα and HSD17B HSD17B10 and ERα co-localize in mitochondria of MCF-7 cells The localization of HSD17B10 in mitochondria has been found in rat cardiac myocytes (29), brain vascular smooth muscle cells (34) and monkey kidney fibroblast-like COS-7 cells (35), whereas ERα has been reported to be present in the nucleus and mitochondria of MCF-7 cells (10). Confocal fluorescence microscopy analyses were carried out to determine the specific localization of HSD17B10 and ERα in MCF-7 cells grown in serum-depleted medium, where we found ERα enriched in our immunoblotting experiments (Fig. 1). We transfected plasmids expressing either ERα or HSD17B10 fused to EGFP at the C-terminus and incubated the cells with 100 nm E2 for 5 min, 3 h or 18 h to investigate possible changes in their intracellular localization relative to control, untreated MCF-7 cells. We used MitoTracker Orange to specifically stain mitochondria within the cells. In the absence of E2, ERα is found predominantly in the nuclei of MCF-7 cells, although we also observed it associated with mitochondria (Fig. 3A). Next we observed the distribution of ERα following the addition of 100 nm E2 after 5 minutes and found that ERα is rapidly enriched within mitochondria (Fig. 3A, 5 min E2 treatment). ERα is found localized in the nucleus after prolonged, 3 and 18 h treatments with E2 (Fig. 3A, 3 h and 18 h E2 treatment). This suggests that E2 has a rapid effect on mitochondrial function via the recruitment of ERα to mitochondria. Next, we investigated the 12
13 localization of HSD17B10 in MCF-7 cells to determine if its distribution follows a similar pattern to ERα, since these two proteins associate with each other (Fig. 2). HSD17B10 was found in the cytoplasm and in specific puncta within mitochondria of MCF-7 cells, however its distribution is not changed as a result of E2 treatment (Fig. 3B), confirming our immunoblot experiment (Fig. 1A). Together with our immunoblotting and TAP experiments, these results show that ERα is rapidly recruited to mitochondria where it associates with HSD17B10 and this association is affected by E In addition to its role in estrogen degradation, HSD17B10 is an essential subunit of the mitochondrial RNase P complex that cleaves the 5' ends of trnas. Since ERα translocates to mitochondria in response to E2, where it interacts with HSD17B10, we investigated whether E2 treatment led to a differential recruitment of HSD17B10 that could affect the formation or stability of the mitochondrial RNase P complex. To maintain mitochondrial proteins in their native complexes we resolved mitochondrial lysates from MCF-7 cells by native electrophoresis, followed by immunoblotting to detect the three protein subunits of the RNase P complex, TRMT10C, HSD17B10 or PRORP (Supplementary Fig. 1). We found that E2 treatment did not affect the stability of the multimeric conformations of the components of the RNase P complex. 314 Estrogen-dependent regulation of mitochondrial transcripts by RNA processing proteins Since HSD17B10 is part of the RNase P complex and RNA processing enzymes regulate mitochondrial gene expression (20,36) we sought to identify the role of these proteins upon E2 treatment. To examine the role of the RNA processing proteins TRMT10C, HSD17B10, PRORP, PTCD1 or ELAC2 on RNA metabolism in MCF-7 cells, we assessed the effects of the sirnamediated knockdown of these proteins in the presence of E2. We carried out qrt-pcr to measure the efficiency of knockdown and found that the sirnas significantly decreased the mrna levels of each RNA processing protein (Fig 4A). In addition, treatment of MCF-7 cells with sirnas against each of the mitochondrial RNA processing enzymes leads to a significant decrease in their protein abundance compared to cells treated with control sirnas in the presence and absence of E2 (Fig. 4B and C). Interestingly, knockdown of HSD17B10 also leads to a decrease in TRMT10C, suggesting 13
14 that the stability of TRMT10C depends on the presence of HSD17B10. These two proteins have been shown to form a stable sub-complex of the mitochondrial RNase P enzyme previously (18,21). In addition, knockdown of HSD17B10 also leads to a subtle decrease of PRORP compared to control indicating the association of these proteins as part of the RNase P complex. Furthermore we observed that knockdown of ELAC2 leads to an increase in PTCD1 abundance in MCF-7 cells (Fig 4B). We have shown that PTCD1 interacts with ELAC2 and affects the 3' end processing of mitochondrial trnas as a negative regulator (20,27) and this increase may represent a compensatory response Next we investigated the effects of knocking down TRMT10C, HSD17B10, PRORP, PTCD1 or ELAC2, relative to controls, on the steady state levels of mitochondrial RNA transcripts by northern blotting in the presence or absence of E2 (Fig. 5A and B). We observed an increase in many mitochondrial RNAs in response to E2 treatment in the control (NT) sirna-treated cells (Supplementary Fig. 2). In contrast, mitochondrially encoded cytochrome c oxidase subunit 1 (MT- CO1) mrna abundance was decreased upon E2 treatment, and the levels of mitochondrially encoded cytochrome b (MT-CYB) and the 12S rrna were not significantly affected (Supplementary Fig. 2). Knockdown of TRMT10C or ELAC2 led to a dramatic reduction of most RNA transcripts (Fig. 5B and Supplementary Fig. 2) and E2 treatment only subtly increased their abundance as in the control samples, providing further evidence that these two enzymes are required for RNA processing in mitochondria. The significant reduction of mature mitochondrial transcripts upon TRMT10C or ELAC2 knockdown is consistent with that observed in HeLa cells (20). In contrast, trna Trp steady state levels in MCF-7 cells were not reduced by the knockdown of any of the mitochondrial RNA processing enzymes (Fig. 5B and Supplementary Fig. 2) Knockdown of HSD17B10 did not result in significant changes in the levels of mitochondrial RNAs (Fig. 5D), however upon addition of E2 we observed a significant increase in the abundance of a particular subset of mitochondrial RNAs (Fig. 5B and Supplementary Fig. 2). In the absence of E2 treatment, PRORP silencing resulted in the up-regulation of some mitochondrial trnas and mrnas, including trna Glu, trna His and trna SerAGY, compared to NT sirna control (Fig. 5D). Knockdown of PTCD1 resulted in an overall increase in the steady state levels of RNA transcripts compared to NT 14
15 sirna control upon estrogen treatment (Fig. 5E). These findings indicate that the levels of many mitochondrial RNA transcripts are affected by both the hormonal status of the cellular environment and the levels of the RNA processing proteins Mutations in the gene encoding HSD17B10 cause 2-methyl-3-hydroxybutyryl-CoA dehydrogenase deficiency, an X-linked neurodegenerative inborn error of isoleucine metabolism which mimics the symptoms of mitochondrial disease (23,37). This indicates that HSD17B10 has an additional role to mitochondrial RNA processing, in the regulation of isoleucine metabolism. Therefore, we investigated the steady state levels of mitochondrial trna Ile in MCF-7 cells compared to cells treated with TRMT10C, HSD17B10, PRORP or NT sirnas in the absence or presence of E2 (Fig. 5B). TRMT10C knockdown led to dramatically decreased trna Ile levels, similar to the changes observed in other RNAs upon knockdown of this protein (Fig. 5B). Interestingly, we observed E2-dependent changes in trna Ile abundance upon knockdown of HSD17B10, that were not observed for any other trna investigated in this study. In the absence of E2 treatment, we observed increased levels of trna Ile upon HSD17B10 knockdown compared to NT sirna control and PRORP knockdown, whereas E2 treatment led to a significant decrease in the steady state levels of trna Ile compared to control and PRORP knockdown (Fig 5F). This pattern was not observed for other mitochondrial RNA transcripts investigated, which indicates that HSD17B10 may play a specific role in the regulation of the trna Ile transcript in an estrogen-dependent manner Next we investigated the effects of E2 on the processing of mitochondrial RNAs by qrt-pcr (Fig. 6). Knockdown of the RNA processing proteins causes an accumulation of pre-processed mitochondrial transcripts (Fig. 6A and B) relative to controls, however E2 can decrease the abundance of specific pre-processed transcripts by affecting the RNase P complex (Fig. 6A), but not the 3' trna processing proteins ELAC2 and PTCD1 (Fig. 6B), suggesting that E2 stimulation can partially rescue processing defects, possibly by recruiting HSD17B10 into the RNAse P complex. Taken together these results show that ERα is rapidly recruited to mitochondria upon E2 activation where it associates with HSD17B10 and stimulates mitochondrial RNA processing. 15
16 Since we observed E2-stimulated changes in mitochondrial transcripts when PTCD1 was knocked down we investigated if this protein associates with ERα by TAP pull down (Fig. 7). We found that endogenous ERα associated with PTCD1-TAP (Fig. 7A), however endogenous PTCD1 could not be detected in association with ERα-TAP (Fig. 7B), suggesting that the TAP-tag may obscure the site through which it associates with PTCD1. Next we examined if PTCD1 and HSD17B10 associated with each other but we could not detect an association between them by TAP pull down, independent of E2 treatment (Supplementary Fig. 3A). This finding indicates that RNase P and ELAC2/PTCD1 complexes are distinct within mitochondria. Additional TAP pull down experiments confirmed the association between TRMT10C and HSD17B10, HSD17B10 and PRORP (Supplementary Fig. 3B) and between PTCD1 and ELAC2 (Supplementary Fig. 3C), independent of E2 treatment. These results confirm the co-existence of TRMT10C, HSD17B10 and PRORP in the RNase P complex in cells and confirm the association between PTCD1 and ELAC2, both originally observed in HeLa cells (18,20). 391 Impaired processing of mitochondrial transcripts leads to decreased protein expression Changes in mitochondrial trna levels as a result of defects in RNA processing can affect the translation of mitochondrially-encoded mrnas (20,38). To investigate the effects of TRMT10C, HSD17B10, PRORP, PTCD1 or ELAC2 knockdown on mitochondrial protein synthesis in the presence and absence of E2, we carried out a de novo incorporation assay using 35 S methionine and cysteine pulse labeling (Fig. 8A and 8B). Coomassie staining of pulse labeling gels showing normalized protein loading is shown in Supplementary Fig. 4. As observed for mitochondrial RNA levels (Supplementary Figure 2), we also found a significant increase in mitochondrial protein synthesis in response to E2 treatment in the control cells (Fig. 7C and D). In contrast, we observed decreased mitochondrial protein synthesis when TRMT10C and ELAC2 were knocked down (Fig. 8C and D), consistent with the most significant decrease in mitochondrial RNAs observed by northern blotting (Fig. 5A). Knockdown of HSD17B10 or PRORP did not have a significant effect on mitochondrial protein synthesis (Fig 8C). We did not observe changes in mitochondrial protein synthesis upon knockdown of PTCD1, independent of E2 treatment (Fig. 8D). PTCD1 knockdown 16
17 increases the abundance of mature trnas that are further stimulated upon E2 treatment (Fig. 5E), however this increase does not affect the rate of protein synthesis, suggesting that these trnas are not normally present at rate-limiting amounts for mitochondrial protein synthesis Next we investigated the steady state levels of mitochondrial proteins by immunoblotting from MCF- 7 cells grown in serum-depleted medium and transfected with TRMT10C, HSD17B10, PRORP, PTCD1 or ELAC2 sirnas and control NT sirnas in the presence and absence of E2. We observed a decrease in the levels of mtdna-encoded COX1 and COX2 cytochrome c oxidase polypeptides upon TRMT10C and ELAC2 knockdown relative to controls, independent of E2 treatment (Fig. 8E and F), consistent with our northern blots (Fig. 5A) and translation measurements (Fig. 8A and 8B). A decrease was also observed in the expression of nuclear-encoded complex II subunit SDHA upon TRMT10C knockdown, independent of E2 and likely as a response to feedback from the decrease in mitochondrially-encoded polypeptides of the electron transport chain. Knockdown of HSD17B10, PRORP or PTCD1 did not markedly affect the steady state levels of mitochondrial proteins (Supplementary Figure 5). Together, the pulse labeling and immunoblotting experiments show a significant decrease of mitochondrial protein expression upon TRMT10C or ELAC2 knockdown. These decreases reflect their crucial role in the processing of mitochondrial transcripts as core subunits of the RNase P and RNase Z complexes and their requirement for mitochondrial gene expression. The dramatic effects of TRMT10C knockdown may be due to the dual role of this protein as a methyltransferase responsible for a trna modification and as a component of the RNase P complex (18,20,21). The E2-dependent nature of the changes observed upon HSD17B10 or PRORP knockdown may reflect additional roles of these proteins in mitochondrial function, including HSD17B10 s enzymatic activity in estrogen metabolism and as a subunit of the RNase P processing complex We investigated the effects of E2 on mitochondrial oxygen consumption in MCF7 cells treated with NT sirnas and upon knockdown of HSD17B10. Mitochondrial respiration is increased upon E2 treatment, which correlates well with increased mitochondrial gene expression (Fig. 8G). Knockdown 17
18 of HSD17B10 led to decreased oxygen consumption, indicating the requirement of this protein for mitochondrial function, however the addition of E2 in cells where HSD17B10 was knocked down improved mitochondrial respiration (Fig. 8G). We carried out qrt-pcr to show that the increase in mitochondrial gene expression in the control MCF7 cells was not due to increased biogenesis driven by nuclear encoded master regulators such as TFAM, PGC-1 and PPAR as their expression is not increased upon E2 treatment (Fig. 8H). Instead, we observe upregulated master regulators in E2- treated cells only when HSD17B10 is knocked down. Taken together these data suggest a possible mechanism by which HSD17B10 can be recruited into the RNase P complex upon E2 stimulation under stress to increase mitochondrial gene expression that consequently improves energy production. However if HSD17B10 is decreased or lost mitochondrial gene expression can be upregulated by increasing the levels of transcriptional activators of mitochondrial biogenesis in the nucleus. 443 Discussion Traditionally, estrogens regulate mitochondrial function by modulating the transcription of nuclear genes involved in mitochondrial gene expression or by acting on receptor-dependent signaling cascades that might affect mitochondrial function. This work explored a new avenue for the regulation of mitochondrial gene expression upon estrogen activation. The results presented here reveal a role for direct action of E2 on mitochondrial RNA metabolism via HSD17B10 in association with ERα. A direct action would allow the hormone to induce rapid changes in mitochondrial gene expression, suggesting that E2 might directly influence energy production by the organelle, by regulating the availability of subunits of the respiratory chain. This would be of particular relevance under metabolic stress conditions, where changes in the energy demand might require a quick, adaptive response by the cell The function of HSD17B10 as a hydroxysteroid dehydrogenase involved in the regulation of estrogen levels is well established (39,40). The association of HSD17B10 with ERα and their co-localization in mitochondria of MCF-7 cells in the presence of estrogen suggests that this hormone affects mitochondrial gene expression via RNA processing. The estrogen response was rapid and stimulated 18
19 the distribution of ERα to mitochondria, suggesting that the cellular stress response in MCF-7 cells is mediated by ERα through its association with HSD17B10. The interaction of HSD17B10 with ERα was initially found in a bacterial two-hybrid screen (29). In that study, a putative mechanism for the interaction between ERα and HSD17B10 was proposed, in which, under low E2 conditions, ERα s binding to HSD17B10 inhibits its catalytic activity in estradiol degradation. In contrast, in the presence of high concentrations of estradiol, HSD17B10 is proposed to dissociate from the HSD17B10- ERα complex and metabolize estradiol to estrone. Although HSD17B10 s binding to ERα could indeed lead to its inactivation in its role in estradiol degradation, this model does not take into account HSD17B10 s fundamental role in mitochondrial RNA processing, as a core subunit of the RNase P processing enzyme. It is reasonable to hypothesize that the recruitment of HSD17B10 by the RNase P complex or its association with ERα upon E2 activation, may underlie the changes observed in this work on mitochondrial RNA transcript levels. In an E2-responsive cellular context, such as that of MCF-7 cells, HSD17B10 s role in estradiol degradation or as a component of the RNase P might result in changes in RNase P activity that in turn lead to variations in mitochondrial transcript abundance We have investigated the roles of TRMT10C, HSD17B10, PRORP, PTCD1 and ELAC2 in the regulation of mitochondrial gene expression in MCF-7 cells and showed that mitochondrial RNA processing can be affected by E2 treatment in cells expressing ERα. Some of the changes observed upon knockdown of these proteins on steady state RNA levels indicate that there are discrete, cell type-specific variations in their action on mitochondrial RNA processing. For instance, in contrast with previous studies in HeLa cells, trna Trp was not affected by the knockdown of TRMT10C or ELAC2 in MCF-7 cells (20) and it remains to be determined how the levels of trna Trp are controlled in MCF-7 cells. Knockdown of TRMT10C decreased the levels of most mitochondrial RNAs dramatically, consistent with previous reports that suggest this may be due to its dual role as a trnamethyltransferase and as part of the mitochondrial RNase P complex in RNA processing (18,20,21). It has been shown that all three components of the mammalian mitochondrial RNase P complex are required for its activity and here we observe that knockdown of each component leads to a decrease of 19
20 mitochondrial RNAs, albeit at different levels. The metallonuclease domain of PRORP provides the catalytic component of the RNase P complex; however, the effects of its knockdown on mitochondrial RNA abundance are not particularly dramatic. With the exception of trna Ile, E2 treatment of cells where HSD17B10 was knocked down did not result in significant changes in mitochondrial RNAs. Control cells showed an increase in many mitochondrial RNAs in response to E2 treatment; interestingly, in the absence of E2 treatment, MCF-7 cells where PRORP was knocked down increased mitochondrial RNA abundance. The estrogen-dependent changes observed upon PRORP knockdown may reflect the dual role of HSD17B10, as a subunit of the RNase P enzyme and as a binding partner of ERα. In the presence of estrogen, HSD17B10 may be preferentially recruited to the steroid metabolism pathway rather than its role as a core subunit of the RNase P complex. Hence, in the absence of E2, mitochondrial transcript levels may be recovered as the protein is available to join the RNase P complex and carry out the processing activity that results in increased mature transcript levels. The changes observed at the RNA level were consistent with changes in mitochondrial protein translation, with decreases in mitochondrial protein synthesis upon TRMT10C or ELAC2 knockdown The increase in PTCD1 abundance in response to ELAC2 knockdown suggests that the two proteins have opposing roles in trna processing, whereby decreased abundance of PTCD1 and consequent increase in ELAC2 should result in increased abundance of mature mitochondrial transcripts. Alternatively, it could be that the association of the two proteins is required for 3' trna processing and the increase in ELAC2 abundance is a compensatory response to decreased PTCD1 levels. The association of ERα with both HSD17B10 and PTCD1 provides the means to affect both 5' and 3' mitochondrial RNA processing in response to E2 and enables coordinated control of mitochondrial gene expression. The effects on mitochondrial gene expression by E2 and ERα under stress growth conditions are specific, since no significant changes in mitochondrial gene expression were observed in cells lacking ERα that were grown in basal and serum depleted medium (Supplementary Fig. 6) Since its identification as a component of the mitochondrial RNase P complex, the role of HSD17B10 in RNA processing has remained obscure. The association between HSD17B10 and TRMT10C into a subcomplex is required for TRMT10C to carry out an important nucleotide modification of 20
21 mitochondrial trnas (18,20,21). Initially, it was suggested that the NAD+ binding site present in HSD17B10 was necessary for the processing activity of RNase P. However, recent studies showed that both HSD17B10 s dehydrogenase activity and NAD+ binding site are not necessary for RNase P activity (21). HSD17B10 s role in the conversion of 17β-estradiol to estrone within mitochondria suggests that mitochondria may be able to directly regulate the intracellular levels of the hormone. This is of particular importance given estrogen s role in cellular proliferation and tumor progression in cancer (41,42). Furthermore, a number of mutations in the gene encoding HSD17B10 have been associated with disease (43). For instance, patients carrying mutations in HSD17B10 present with neurological abnormalities and neuromotor disorders associated with defects in the metabolism of isoleucine. Accordingly, our work shows a specific role for HSD17B10 in trna Ile transcript abundance. The role of HSD17B10 on isoleucine availability may explain the effect of HSD17B10 knockdown on trna Ile abundance. HSD17B10 s role in isoleucine degradation represents yet another function of this versatile, multifunctional protein that requires further investigation Mitochondria play a range of key roles in the cell, including the integration of extracellular signals, such as hormones, and the execution of intracellular responses. There is increasing evidence suggesting a crosstalk between mitochondria and the nucleus, which is directly related to cellular function and reveals the existence of signaling pathways that may provide new therapeutic targets in disease. Estrogen mediated carcinogenesis caused by spontaneous mutations that lead to enhanced cell proliferation has been associated with increased levels of estrogen and long-term stimulation of estrogen signaling pathways (44). Although the exact mechanisms underlying these signaling pathways are not fully understood, the results presented here indicate that increased estrogen levels affect mitochondrial gene expression by modulating the activities of mitochondrial RNA processing proteins via ERα, further supported by the evidence that the master regulators of mitochondrial biogenesis (TFAM, PGC-1 and PPAR ) are not upregulated with ERα treatment. Instead these factors are upregulated when the levels of HSD17B10 are compromised, suggesting that there is a secondary mechanism that can compensate for decreased mitochondrial function. These findings illustrate the existence of feedback mechanisms between the nucleus and mitochondria directly modulating 21
22 mitochondrial gene expression. Here, HSD17B10 is described as a novel target for the regulation of mitochondrial gene expression by E2 and a clear example of a moonlighting enzyme, a multifunctional protein performing multiple, unrelated functions and involved in different aspects of mitochondrial metabolism. 543 Acknowledgements The authors thank Moira Hibbs for technical assistance and Jacky Bentel for providing MCF-7 cells and critical review of the manuscript
23 References 1. Hall JM, Couse JF, Korach KS. The multifaceted mechanisms of estradiol and estrogen receptor signaling. J Biochem. 2001;276(40): Vasconsuelo A, Pronsato L, Ronda AC, Boland R, Milanesi L. Role of 17beta-estradiol and testosterone in apoptosis. Steroids. 2011;76(12): Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science. 2005;307(5715): Losel R, Wehling M. Nongenomic actions of steroid hormones. Nat Rev Mol Cell Biol 2003;4(1): Levin ER. Plasma membrane estrogen receptors. Trends Endocrinol Metab. 2009;20(10): Stirone C, Duckles SP, Krause DN, Procaccio V. Estrogen increases mitochondrial efficiency and reduces oxidative stress in cerebral blood vessels. Mol Pharmacol. 2005;68(4): Klinge CM. Estrogenic control of mitochondrial function and biogenesis. J Cell Biochem. 2008;105(6): Mattingly KA, Ivanova MM, Riggs KA, Wickramasinghe NS, Barch MJ, Klinge CM. Estradiol stimulates transcription of nuclear respiratory factor-1 and increases mitochondrial biogenesis. Mol Endocrinol. 2008;22(3): Yager JD, Chen JQ. Mitochondrial estrogen receptors--new insights into specific functions. Trends Endocrinol Metab. 2007;18(3): Chen JQ. Mitochondrial localization of ER and ER in human MCF7 cells. Am J Physiol. 2004;286(6):E1011 E Pedram A, Razandi M, Wallace DC, Levin ER. Functional estrogen receptors in the mitochondria of breast cancer cells. Mol Biol Cell. 2006;17(5): Psarra AM, Sekeris CE. Nuclear receptors and other nuclear transcription factors in mitochondria: regulatory molecules in a new environment. Biochim Biophys Acta. 2008;1783(1): Chen JQ, Yager JD. Estrogen's effects on mitochondrial gene expression: mechanisms and potential contributions to estrogen carcinogenesis. Ann N Y Acad Sci. 2004;1028: Sekeris CE. The mitochondrial genome: a possible primary site of action of steroid hormones. In Vivo. 1990;4(5): Chen JQ, Eshete M, Alworth WL, Yager JD. Binding of MCF-7 cell mitochondrial proteins and recombinant human estrogen receptors alpha and beta to human mitochondrial DNA estrogen response elements. J Cell Biochem. 2004;93(2): Ojala D, Montoya J, Attardi G. trna punctuation model of RNA processing in human mitochondria. Nature. 1981;290(5806):
24 Mercer TR, Neph S, Dinger ME, Crawford J, Smith MA, Shearwood A-MJ, Haugen E, Bracken CP, Rackham O, Stamatoyannopoulos JA, Filipovska A, Mattick JS. The Human Mitochondrial Transcriptome. Cell. 2011;146(4): Holzmann J, Frank P, Löffler E, Bennett KL, Gerner C, Rossmanith W. RNase P without RNA: Identification and Functional Reconstitution of the Human Mitochondrial trna Processing Enzyme. Cell. 2008;135(3): Brzezniak L, Stepien PP, Bijata M, Szczesny RJ. Human ELAC2 gene encodes the trnasez responsible for mitochondrial trna 3 processing which acts on precursors already cleaved by RNAseP. RNA Biol. 2011;8(4): Lopez Sanchez MI, Mercer TR, Davies SM, Shearwood A-MJ, Nygård KK, Richman TR, Mattick JS, Rackham O, Filipovska A. RNA processing in human mitochondria. Cell Cycle. 2011;10(17): Vilardo E, Nachbagauer C, Buzet A, Taschner A, Holzmann J, Rossmanith W. A subcomplex of human mitochondrial RNase P is a bifunctional methyltransferase--extensive moonlighting in mitochondrial trna biogenesis. Nucleic Acids Res. 2012;40(22): Rauschenberger K, Schöler K, Sass JO, Sauer S, Djuric Z, Rumig C, Wolf NI, Okun JG, Kölker S, Schwarz H, Fischer C, Grziwa B, Runz H, Nümann A, Shafqat N, Kavanagh KL, Hämmerling G, Wanders RJA, Shield JPH, Wendel U, Stern D, Nawroth P, Hoffmann GF, Bartram CR, Arnold B, Bierhaus A, Oppermann U, Steinbeisser H, Zschocke J. A non-enzymatic function of 17β-hydroxysteroid dehydrogenase type 10 is required for mitochondrial integrity and cell survival. EMBO Mol Med. 2010;2(2): Ofman R, Ruiter JP, Feenstra M, Duran M, Poll-The BT, Zschocke J, Ensenauer R, Lehnert W, Sass JO, Sperl W, Wanders RJ. 2-Methyl-3-hydroxybutyryl-CoA dehydrogenase deficiency is caused by mutations in the HADH2 gene. Am J Hum Genet. 2003;72(5): Korman SH. Inborn errors of isoleucine degradation: a review. Mol Genet Metab. 2006;89(4): Rackham O, Filipovska A. The role of mammalian PPR domain proteins in the regulation of mitochondrial gene expression. Biochim Biophys Acta. 2011;1819(9-10): Liu G, Mercer TR, Shearwood A-MJ, Siira SJ, Hibbs ME, Mattick JS, Rackham O, Filipovska A. Mapping of mitochondrial RNA-protein interactions by digital RNase footprinting. Cell Reports. 2013;5(3): Rackham O, Davies SMK, Shearwood A-MJ, Hamilton KL, Whelan J, Filipovska A. Pentatricopeptide repeat domain protein 1 lowers the levels of mitochondrial leucine trnas in cells. Nucleic Acids Res. 2009;37(17): He XY, Merz G, Yang YZ, Mehta P, Schulz H, Yang SY. Characterization and localization of human type10 17beta-hydroxysteroid dehydrogenase. Eur J Biochem. 2001;268(18): Jazbutyte V, Kehl F, Neyses L, Pelzer T. Estrogen receptor alpha interacts with 17betahydroxysteroid dehydrogenase type 10 in mitochondria. Biochem Biophys Res Commun. 2009;384(4):
25 Smith P, Krohn R, Hermanson G, Mallia A, Gartner F, Provenzano M, Fujimoto E, Goeke N, Olson B, Klenk D. Measurement of protein using bicinchoninic acid. Anal Biochem. 1985;150: Davies SMK, Lopez Sanchez MIG, Narsai R, Shearwood A-MJ, Razif MFM, Small ID, Whelan J, Rackham O, Filipovska A. MRPS27 is a pentatricopeptide repeat domain protein required for the translation of mitochondrially encoded proteins. FEBS Letts. 2012;586(20): Davies SMK, Rackham O, Shearwood A-MJ, Hamilton KL, Narsai R, Whelan J, Filipovska A. Pentatricopeptide repeat domain protein 3 associates with the mitochondrial small ribosomal subunit and regulates translation. FEBS Letters. 2009;583(12): Razmara A, Sunday L, Stirone C, Wang XB, Krause DN, Duckles SP, Procaccio V. Mitochondrial effects of estrogen are mediated by estrogen receptor alpha in brain endothelial cells. J pharm exp ther. 2008;325(3): Frackowiak J, Mazur-Kolecka B, Kaczmarski W, Dickson D. Deposition of Alzheimer's vascular amyloid-beta is associated with decreased expression of brain L-3-hydroxyacylcoenzyme A dehydrogenase (ERAB). Brain Res. 2001;907(1-2): He XY, Merz G, Mehta P, Schulz H, Yang SY. Human brain short chain L-3-hydroxyacyl coenzyme A dehydrogenase is a single-domain multifunctional enzyme. Characterization of a novel 17beta-hydroxysteroid dehydrogenase. J Biochem. 1999;274(21): Brzezniak LK, Bijata M, Szczesny RJ, Stepien PP. Involvement of human ELAC2 gene product in 3' end processing of mitochondrial trnas. RNA Biol. 2011;8(4): Perez-Cerda C, Garcia-Villoria J, Ofman R, Sala PR, Merinero B, Ramos J, Garcia-Silva MT, Beseler B, Dalmau J, Wanders RJ, Ugarte M, Ribes A. 2-Methyl-3-hydroxybutyryl- CoA dehydrogenase (MHBD) deficiency: an X-linked inborn error of isoleucine metabolism that may mimic a mitochondrial disease. Pediatr Res. 2005;58(3): Hayashi J, Ohta S, Kagawa Y, Takai D, Miyabayashi S, Tada K, Fukushima H, Inui K, Okada S, Goto Y, al E. Functional and morphological abnormalities of mitochondria in human cells containing mitochondrial DNA with pathogenic point mutations in trna genes. J Biol Chem. 1994;269(29): Yang S, He X, Schulz H. Multiple functions of type 10 17beta-hydroxysteroid dehydrogenase. Trends Endocrinol Metab. 2005;16(4): Moeller G, Adamski J. Multifunctionality of human 17beta-hydroxysteroid dehydrogenases. Mol Cell Endocrinol. 2006;248(1-2): Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson G, Enmark E, Pettersson K, Warner M, Gustafsson JA. Mechanisms of estrogen action. Physiol Rev. 2001;81(4): Yue W, Yager JD, Wang JP, Jupe ER, Santen RJ. Estrogen receptor-dependent and independent mechanisms of breast cancer carcinogenesis. Steroids. 2013;78(2): Zschocke J. HSD10 disease: clinical consequences of mutations in the HSD17B10 gene. J Inherit Metab Dis. 2012;35(1): Feigelson HS, Henderson BE. Estrogens and breast cancer. Carcinogenesis. 25
26 ;17(11):
27 691 Table and Figure legends Table 1. Nomenclature and synonyms of the studied proteins Nuclear encoded proteins used in this work are listed with their UniProtKB identifiers a, gene symbol according to HUGO Nomenclature Committee b, common synonyms c and their function d. Figure 1. HSD17B10 and ERα expression in MCF-7 cells A. ERα was detected by immunoblotting of mitochondrial lysates from MCF-7 cells grown in basal, non serum-depleted medium and cells grown in serum-depleted medium, followed by incubation with 100 nm E2 for 18 h. Porin was used as a control to confirm normalized protein loading. Bars represent mean ± SD of relative mitochondrial ERα levels or HSD17B10 in serum-depleted cells, normalized to porin and compared with cells grown in basal, non-depleted medium in the presence or absence of E2. Statistical significance was examined using a two-tailed Student s t-test, * p < 0.05, n= ERα and HSD17B10 cellular distribution was assessed by immunoblotting from cell lysates and mitochondrial fractions isolated from MCF-7 cells grown in basal medium (B) or serum-depleted medium (C), followed by incubation with 100 nm E2 for 18 h. 707 Figure 2. HSD17B10 interacts with ERα A. Endogenous ERα was isolated from MCF-7 cells grown in serum-depleted medium, expressing HSD17B10-TAP (48 kda) in the presence or absence of 100 nm estradiol (E2) for 18 h, using IgG agarose beads, followed by TEV protease cleavage B. TAP analyses show endogenous HSD17B10 (27 kda) isolated from cells expressing ERα-TAP (88 kda). IgG heavy (50 kda) and light chains (23 kda) can be seen on the immunoblots and are marked with asterisks. 714 C and D. EGFP was used as a control to show the specificity of the TAP procedure. 715 Figure 3. HSD17B10 and ERα co-localize in mitochondria of MCF-7 cells 27
28 A. ERα is predominantly nuclear but is rapidly recruited to mitochondria of MCF-7 cells upon E2 treatment. Cells grown in serum-depleted medium were transfected with a plasmid expressing ERα fused to EGFP at the C-terminus, followed by incubation with 100 nm E2 for 5 min, 3 h or 18 h. Cells were then incubated with MitoTracker Orange (MT Orange) to stain mitochondria in live cells and fixed. ERα-EGFP (green) can be seen in the nucleus and also in mitochondria stained with MT Orange (red) in the overlaid images (yellow). Scale bar is 10 µm B. HSD17B10 is present in the cytoplasm and in puncta within mitochondria of MCF-7 cells. Cells grown in serum-depleted medium were transfected with a plasmid expressing HSD17B10 fused to EGFP at the C-terminus, followed by incubation with 100 nm E2 for 5 min, 3 h or 18 h. Cells were then incubated with MitoTracker Orange (MT Orange) to stain mitochondria in live cells and fixed. HSD17B10-EGFP (green) can be seen in the cytoplasm and also in mitochondria stained with MT Orange (red) in the overlaid images (yellow). Scale bar is 10 µm. 728 Figure 4. Knockdown of mitochondrial RNA processing proteins A. The mrna knockdown levels of the mitochondrial RNA processing proteins in MCF-7 cells were determined by qrt- PCR relative to control non-targeting (NT) sirna-treated cells and normalized to 18S rrna. Bars represent mean ± SD. Student's 2-tailed t test for paired data was carried out and showed that mrnas were significantly decreased relative to controls, p < 0.05 n= B. Immunoblotting of mitochondria isolated from MCF-7 cells transfected with control non-targeting (NT), TRMT10C, HSD17B10, PRORP, PTCD1 or ELAC2 sirnas for 72 h in serum-depleted medium, followed by an additional re-transfection for 72 h to enhance knockdown efficiency, and in the presence or absence of 100 nm estradiol (E2) for 18 h. Antibodies were used to detect the expression levels of TRMT10C, HSD17B10, PRORP, PTCD1 or ELAC2 upon sirna-mediated knockdown. Porin was used as a control to confirm equal protein loading. Blots are representative of 4 independent biological experiments C. Densitometry analysis of immunoblotting experiments show that sirna treatment significantly decreases protein abundance compared to control cells, in the presence and absence of E2. Graph 28
29 shows relative protein levels normalized to porin and compared to non-targeting (NT) control. Bars represent relative mean ± SD. Student's 2-tailed t test for paired data was carried out and showed that protein levels were significantly decreased relative to controls, p < Figure 5. E2-dependent effects of RNA processing proteins on mitochondrial RNA levels A. RNA was isolated from MCF-7 cells transfected with control non-targeting (NT), TRMT10C, HSD17B10, PRORP, PTCD1 or ELAC2 sirna, in the presence or absence of 100 nm estradiol (E2) for 18 h, to determine steady state levels of mitochondrial transcripts by northern blotting. Nuclear 18S rrna abundance was used as an internal control for total RNA loading. Data shown are typical results repeated on three independent RNA preparations. Each blot was probed for the different RNA species by stripping and re-probing B. Northern blot showing the effects of RNA processing protein knockdown on the steady state levels of trna Ile in the presence and absence of 100 nm E Figure 6. E2-dependent effects on mitochondrial RNA processing RNA was isolated from MCF-7 cells transfected with control non-targeting (NT), TRMT10C, HSD17B10, PRORP (A) PTCD1 or ELAC2 (B) sirna, in the presence or absence of 100 nm estradiol (E2) for 18 h, to determine the effects on mitochondrial RNA processing by qrt-pcr relative to control NT sirna-treated cells and normalized to 18S rrna. Data shown are typical results repeated on three independent RNA preparations. Data are means ± S.D. of three separate experiments, p < 0.05 is marked with an asterisk. 761 Figure 7. PTCD1 associates with ERα A. Endogenous ERα (67 kda) was isolated from MCF-7 cells grown in serum-depleted medium, expressing PTCD1-TAP (98 kda) in the presence or absence of 100 nm estradiol (E2) for 18 h, using IgG agarose beads, followed by TEV protease cleavage. 29
30 B. Reverse TAP analyses were carried out to detect endogenous PTCD1 (77 kda) isolated from cells expressing ERα-TAP (88 kda) using a PTCD1 specific monoclonal antibody Figure 8. Impaired processing of mitochondrial transcripts leads to decreased protein expression Mitochondrial protein synthesis was analyzed by the incorporation of 35 S-labeled methionine and cysteine in the 13 proteins synthesized by the mitochondrial translation machinery. MCF-7 cells were grown in serum-depleted medium and transfected with sirnas targeting TRMT10C, HSD17B10, PRORP or NT control (A) and sirnas targeting PTCD1, ELAC2 or NT control (B) in serumdepleted medium, followed by treatment with 100 nm E2 for 18 h. The protein synthesis of cytoplasmic proteins was inhibited with emetine. Coomassie stained gels showing equal loading of the cell lysates are shown in Supplementary Fig. 4A C. Knockdown of TRMT10C, in the presence of estrogen, decreases mitochondrial DNA-encoded protein synthesis. Graph shows relative protein levels normalized to porin. Bars represent relative mean ± SD. Statistical significance was examined using a two-tailed Student s t-test, and indicated by * (* p < 0.05), n= D. Knockdown of ELAC2 decreases mitochondrial DNA-encoded protein synthesis. Graph shows relative protein levels normalized to porin. Bars represent relative mean ± SD. Statistical significance was examined using a two-tailed Student s t-test, and indicated by * (* p < 0.05), n= Immunoblotting analyses of mitochondrial proteins isolated from MCF-7 cells grown in serumdepleted medium and transfected with sirnas targeting TRMT10C, HSD17B10, PRORP or NT control (E) and sirnas targeting PTCD1, ELAC2 or NT control (F), followed by treatment with 100 nm E2 for 18 h. Antibodies were used to detect the expression levels of mtdna-encoded cytochrome c oxidase subunits COX1 and COX2 and nuclear DNA-encoded complex II SDHA subunit. Porin was used as a control to confirm normalized protein loading. 30
31 G. Mitochondrial oxygen consumption was measured in MCF-7 cells grown in serum-depleted medium, in the presence or absence of 100 nm E2 for 18 h. Data are means ± S.D. of three separate experiments, p < 0.05, n= H. The mrna levels of TFAM, PGC-1 and PPAR were measured in MCF-7 cells treated with HSD17B10 or NT control sirnas in the presence or absence of 100 nm E2 by qrt- PCR. The fold change in expression upon addition of E2 compared to absence of E2 is shown. Data were normalized to 18S rrna and bars represent mean ± SD. Student's 2-tailed t test for paired data was carried out and showed that mrnas were significantly changed relative to controls, p < 0.05, n= Supplementary Figure 1. E2 treatment does not affect the stability of the RNase P complex Mitochondria were prepared from MCF-7 cells grown in serum-depleted medium and incubated in the presence or absence of 100 nm E2 for 18 h. Mitochondrial proteins were then solubilized with 1% dodecyl maltoside, sedimented and the supernatant resolved by native PAGE. Immunoblotting was carried to detect TRMT10C, HSD17B10 or PRORP with antibodies specific to these proteins. Probing with a monoclonal TRMT10C antibody revealed the presence of two bands at ~240 kda and ~300 kda. The same two bands were observed after probing an independent blot with PRORP antibody. In contrast, a band of ~150 kda was detected using a monoclonal antibody against HSD17B10. Proteins detected in native PAGE experiments have a size estimation error or ±15%. The theoretical, combined molecular weight of the protein subunits of RNase P is ~140 kda (TRMT10C has a molecular weight of 47 kda, HSD17B10 27 kda and PRORP 67 kda). It has been reported that HSD17B10 exists as a 108 kda homotetramer (He, 2001), which might correspond to the band detected by native PAGE using a monoclonal antibody specific to HSD17B10, taking into account the size estimation error. The higher molecular weight bands observed with TRMT10C or PRORP antibodies might represent multimeric associations between the RNA processing subunits Supplementary Figure 2. E2 effects on steady-state levels of mitochondrial RNAs in control, and gene-specific sirna treated cells. 31
32 A. Effects of knockdown of TRMT10C, HSD17B10 and PRORP on the steady-state levels of mitochondrial RNA transcripts in the presence and absence of E B. Effects of knockdown of ELAC2 and PTCD1 on the steady-state levels of mitochondrial RNA transcripts in the presence and absence of E The data are expressed relative to transcripts from control samples. Bars represent relative mean ± SD. Student's 2-tailed t test for paired data was carried out, p < C. HSD17B10 knockdown results in E2-dependent changes in trna Ile abundance. Data are expressed as the ratio of trna Ile transcript normalized to 18S rrna. Bars represent relative mean ± SD. Statistical significance was examined using a two-tailed Student s t-test, and indicated by * (* p < 0.05). 824 Supplementary Figure 3. Additional protein interactions detected by TAP 825 A. PTCD1 and HSD17B10 do not interact, as assessed by TAP followed by immunoblotting B. Endogenous HSD17B10 was isolated from MCF-7 cells expressing TRMT10C-TAP or PRORP- TAP, independent of E2 treatment C. Endogenous ELAC2 or PTCD1 isolated from MCF-7 cells expressing PTCD1-TAP or ELAC2- TAP respectively, independent of E2 treatment. 830 Supplementary Figure 4. Normalized protein loading of pulse labeling gels 831 A and B. Blue Coomassie staining of pulse labeling gels showing normalized protein loading Supplementary Figure 5. Knockdown of trna processing proteins and their effects on the steady state levels of mitochondrial proteins in MCF7 cells Bars represent mean ± SD of relative protein expression, normalized to porin and compared with NT controls. 32
33 Supplementary Figure 6. The levels of mitochondrial RNA are not affected by E2 in ER negative cells RNA was isolated from MDA-468 and MDA-231 grown in basal or serum depleted medium in the presence or absence of 100 nm estradiol (E2) for 18 h, to determine steady state levels of mitochondrial transcripts by northern blotting. Nuclear 18S rrna abundance was used as an internal control for total RNA loading. Data shown are typical results repeated on three independent RNA preparations. Each blot was probed for the different RNA species by stripping and re-probing and the signal was quantified from three independent experiments
34 Figure Click here to download Figure: Figure 1.eps Figure 1 A 100 nm E B 67 kda 26 kda 31 kda ERα Porin Cell lysate Mitochondria Mitoplast Insoluble fraction Soluble fraction 100 nm E kda Basal medium Depleted medium HSD17B10 Mitochondrial ERα levels relative to control in the presence of E E2 +E2 Cell lysate Mitochondria Mitoplast Insoluble fraction Soluble fraction * Basal ERα * Depleted Mitochondrial HSD17B10 levels relative to control E2 +E2 Basal Depleted C 26 kda HSD17B nm E kda 26 kda ERα HSD17B10





35 Figure Click here to download Figure: Figure 2.eps A B 67 kda 48 kda -100 nm E nm E2 Cell lysate Flow through Blank Eluate Cell lysate Flow through Blank Eluate Cell lysate Flow through Blank Eluate ERα 67 kda ERα * * HSD17B10-TAP 48 kda HSD17B10-TAP -100 nm E nm E2 Cell lysate Flow through Blank Eluate 88 kda ER α-tap 88 kda ER α-tap * * 26 kda HSD17B10 26 kda HSD17B10 C 48 kda -100 nm E nm E2 Cell lysate Blank Flow through Eluate Cell lysate Flow through Blank Eluate * HSD17B10-TAP * 48 kda HSD17B10-TAP 27 kda EGFP 27 kda EGFP D -100 nm E nm E2 Cell lysate Flow through Blank Eluate Cell lysate Flow through Blank Eluate 88 kda ER α-tap * 88 kda * ER α-tap 27 kda EGFP 27 kda EGFP Figure 2








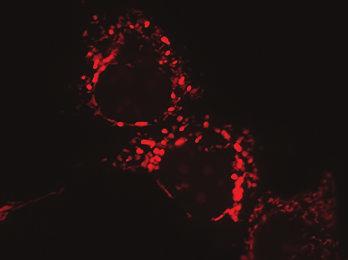





36 A Untreated cells ERα-EGFP MT Orange Overlay Figure Click here to download Figure: Figure 3.eps 100 nm E2 for 18 h 100 nm E2 for 3 h 100 nm E2 for 5 min B HSD17B10-EGFP MT Orange Overlay 100 nm E2 for 18 h 100 nm E2 for 3 h 100 nm E2 for 5 min Untreated cells Figure 3
37 Figure Click here to download Figure: Figure 4.eps A TRMT10C HSD17B10 PRORP PTCD1 ELAC2 B sirna sirna 100 nm E nm E2 47 kda TRMT10C 77 kda 26 kda HSD17B10 92 kda 67 kda PRORP 31 kda 31 kda Porin control TRMT10C HSD17B10 PRORP control PTCD ELAC2 PTCD1 ELAC2 Porin C Normalized protein expression relative to control Fold RNA change relative to control Figure 4
38 Figure Click here to download Figure: Figure 5.eps Figure 5 A sirna 100 nm E2 NT TRMT10C HSD17B PRORP NT PTCD1 ELAC trna Ala trna Glu trna His trna LeuUUR trna SerAGY trna Trp MT-CO1 MT-CYB 12S rrna B sirna 100 nm E2 NT TRMT10C HSD17B10 PRORP NT PTCD1 ELAC S rrna 18S rrna trna Ile
39 A Figure Click here to download Figure: Figure 6.eps Fold RNA change relative to control * * * * * * * ND1-ND2 ND2-tRNA Ala CO1-CO2 sitrmt10c -E2 sitrmt10c +E2 sihsd17b10 -E2 sihsd17b10 +E2 siprorp -E2 siprorp +E2 B Fold RNA change relative to control * ND1-ND2 ND2-tRNA Ala CO1-CO2 sielac2 -E2 sielac2 +E2 siptcd1 -E2 siptcd1 +E2 Figure 6

40 Figure Click here to download Figure: Figure 7.eps Figure 7 A 98 kda 67 kda -100 nm E nm E2 Cell lysate Flow through Blank Eluate PTCD1-TAP ER α Cell lysate Flow through Blank Eluate 98 kda PTCD1-TAP 67 kda ER α B -100 nm E nm E2 Cell lysate Flow through Blank Eluate 88 kda ERα -TAP 88 kda 77 kda PTCD1 77 kda Cell lysate Flow through Blank Eluate ER α -TAP PTCD1


41 A Figure Click here to download Figure: Figure 8.eps sirna 100 nm E2 NT TRMT10C HSD17B10 PRORP B sirna 100 nm E2 NT PTCD1 ELAC ND5 CO1 ND4 Cyt b ND2 ND1 CO3 CO2 ATP6 ND3 ND4L ATP8 ND5 CO1 ND4 Cyt b ND2 ND1 CO3 CO2 ATP6 ND3 ND4L ATP8 C E Normalized protein expression sirna 100 nm E NT * * NT -E2 NT +E2 TRMT10C -E2 TRMT10C +E2 HSD17B10 -E2 HSD17B10 +E2 PRORP -E2 PRORP +E2 TRMT10C HSD17B PRORP sirna 100 nm E2 37 kda COX1 37 kda D F Normalized protein expression NT * NT -E2 NT +E2 PTCD1 -E2 PTCD1 +E2 ELAC2 -E2 ELAC2 +E2 PTCD * * ELAC2 COX1 26 kda COX2 26 kda COX2 70 kda SDHA 70 kda SDHA 31 kda Porin 31 kda Porin G Oxygen consumption (nmol O/min/10 6 cells) nm E NT sirna MRPP2 sirna H * * 14 NT sirna * 12 * HSD17B10 sirna 10 8 * 6 4 * 2 0 Figure 8 Fold increase in the presence of estrogen TFAM PPARγ PGC-1
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
BME 5742 Biosystems Modeling and Control
 BME 5742 Biosystems Modeling and Control Lecture 24 Unregulated Gene Expression Model Dr. Zvi Roth (FAU) 1 The genetic material inside a cell, encoded in its DNA, governs the response of a cell to various
BME 5742 Biosystems Modeling and Control Lecture 24 Unregulated Gene Expression Model Dr. Zvi Roth (FAU) 1 The genetic material inside a cell, encoded in its DNA, governs the response of a cell to various
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Tuesday, December 27, 16
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Three types of RNA polymerase in eukaryotic nuclei
 Three types of RNA polymerase in eukaryotic nuclei Type Location RNA synthesized Effect of α-amanitin I Nucleolus Pre-rRNA for 18,.8 and 8S rrnas Insensitive II Nucleoplasm Pre-mRNA, some snrnas Sensitive
Three types of RNA polymerase in eukaryotic nuclei Type Location RNA synthesized Effect of α-amanitin I Nucleolus Pre-rRNA for 18,.8 and 8S rrnas Insensitive II Nucleoplasm Pre-mRNA, some snrnas Sensitive
CHAPTER 3. Cell Structure and Genetic Control. Chapter 3 Outline
 CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
What Organelle Makes Proteins According To The Instructions Given By Dna
 What Organelle Makes Proteins According To The Instructions Given By Dna This is because it contains the information needed to make proteins. assemble enzymes and other proteins according to the directions
What Organelle Makes Proteins According To The Instructions Given By Dna This is because it contains the information needed to make proteins. assemble enzymes and other proteins according to the directions
Reading Assignments. A. Genes and the Synthesis of Polypeptides. Lecture Series 7 From DNA to Protein: Genotype to Phenotype
 Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1
 and Ksp/Cre;Pkd1") Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
1. In most cases, genes code for and it is that
 Name Chapter 10 Reading Guide From DNA to Protein: Gene Expression Concept 10.1 Genetics Shows That Genes Code for Proteins 1. In most cases, genes code for and it is that determine. 2. Describe what Garrod
Name Chapter 10 Reading Guide From DNA to Protein: Gene Expression Concept 10.1 Genetics Shows That Genes Code for Proteins 1. In most cases, genes code for and it is that determine. 2. Describe what Garrod
Supplementary Figure 1.
 Supplementary Figure 1. Characterisation of IHG-1 overexpressing and knockdown cell lines. (A) Total cellular RNA was prepared from HeLa cells stably overexpressing IHG-1 or mts-ihg-1. IHG-1 mrna was quantified
Supplementary Figure 1. Characterisation of IHG-1 overexpressing and knockdown cell lines. (A) Total cellular RNA was prepared from HeLa cells stably overexpressing IHG-1 or mts-ihg-1. IHG-1 mrna was quantified
7.06 Cell Biology EXAM #3 April 21, 2005
 7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
16 CONTROL OF GENE EXPRESSION
 16 CONTROL OF GENE EXPRESSION Chapter Outline 16.1 REGULATION OF GENE EXPRESSION IN PROKARYOTES The operon is the unit of transcription in prokaryotes The lac operon for lactose metabolism is transcribed
16 CONTROL OF GENE EXPRESSION Chapter Outline 16.1 REGULATION OF GENE EXPRESSION IN PROKARYOTES The operon is the unit of transcription in prokaryotes The lac operon for lactose metabolism is transcribed
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
Chapter 16 Lecture. Concepts Of Genetics. Tenth Edition. Regulation of Gene Expression in Prokaryotes
 Chapter 16 Lecture Concepts Of Genetics Tenth Edition Regulation of Gene Expression in Prokaryotes Chapter Contents 16.1 Prokaryotes Regulate Gene Expression in Response to Environmental Conditions 16.2
Chapter 16 Lecture Concepts Of Genetics Tenth Edition Regulation of Gene Expression in Prokaryotes Chapter Contents 16.1 Prokaryotes Regulate Gene Expression in Response to Environmental Conditions 16.2
9/2/17. Molecular and Cellular Biology. 3. The Cell From Genes to Proteins. key processes
 Molecular and Cellular Biology Animal Cell ((eukaryotic cell) -----> compare with prokaryotic cell) ENDOPLASMIC RETICULUM (ER) Rough ER Smooth ER Flagellum Nuclear envelope Nucleolus NUCLEUS Chromatin
Molecular and Cellular Biology Animal Cell ((eukaryotic cell) -----> compare with prokaryotic cell) ENDOPLASMIC RETICULUM (ER) Rough ER Smooth ER Flagellum Nuclear envelope Nucleolus NUCLEUS Chromatin
MITOCHONDRIAL BIOGENESIS AND REDOX REGULATION
 MITOCHONDRIAL BIOGENESIS AND REDOX Claude A. Piantadosi, MD Professor of Medicine and Pathology Duke University Medical Center Durham, N.C. USA Objectives Provide an overview of the physiological and pathological
MITOCHONDRIAL BIOGENESIS AND REDOX Claude A. Piantadosi, MD Professor of Medicine and Pathology Duke University Medical Center Durham, N.C. USA Objectives Provide an overview of the physiological and pathological
Arginase Assay Kit. Catalog Number KA assays Version: 05. Intended for research use only.
 Arginase Assay Kit Catalog Number KA1609 200 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Arginase Assay Kit Catalog Number KA1609 200 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Chapter 12. Genes: Expression and Regulation
 Chapter 12 Genes: Expression and Regulation 1 DNA Transcription or RNA Synthesis produces three types of RNA trna carries amino acids during protein synthesis rrna component of ribosomes mrna directs protein
Chapter 12 Genes: Expression and Regulation 1 DNA Transcription or RNA Synthesis produces three types of RNA trna carries amino acids during protein synthesis rrna component of ribosomes mrna directs protein
Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Old FINAL EXAM BIO409/509 NAME. Please number your answers and write them on the attached, lined paper.
 Old FINAL EXAM BIO409/509 NAME Please number your answers and write them on the attached, lined paper. Gene expression can be regulated at several steps. Describe one example for each of the following:
Old FINAL EXAM BIO409/509 NAME Please number your answers and write them on the attached, lined paper. Gene expression can be regulated at several steps. Describe one example for each of the following:
GCD3033:Cell Biology. Transcription
 Transcription Transcription: DNA to RNA A) production of complementary strand of DNA B) RNA types C) transcription start/stop signals D) Initiation of eukaryotic gene expression E) transcription factors
Transcription Transcription: DNA to RNA A) production of complementary strand of DNA B) RNA types C) transcription start/stop signals D) Initiation of eukaryotic gene expression E) transcription factors
Full-length GlpG sequence was generated by PCR from E. coli genomic DNA. (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
,") Supplementary Methods Protein expression and purification Full-length GlpG sequence was generated by PCR from E. coli genomic DNA (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
Supplementary Methods Protein expression and purification Full-length GlpG sequence was generated by PCR from E. coli genomic DNA (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
What are mitochondria?
 What are mitochondria? What are mitochondria? An intracellular organelle. There are 100 to 1000s of mitochondria/cell. Most mitochondria come from the mother. Mitochondria have their own DNA Mitochondria
What are mitochondria? What are mitochondria? An intracellular organelle. There are 100 to 1000s of mitochondria/cell. Most mitochondria come from the mother. Mitochondria have their own DNA Mitochondria
Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday
 Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday 1. What is the Central Dogma? 2. How does prokaryotic DNA compare to eukaryotic DNA? 3. How is DNA
Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday 1. What is the Central Dogma? 2. How does prokaryotic DNA compare to eukaryotic DNA? 3. How is DNA
U.S. Patent No. 9,051,563 and other pending patents. Ver
 INSTRUCTION MANUAL Direct-zol 96 RNA Catalog Nos. R2054, R2055, R2056 & R2057 Highlights Quick, 96-well purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and all other
INSTRUCTION MANUAL Direct-zol 96 RNA Catalog Nos. R2054, R2055, R2056 & R2057 Highlights Quick, 96-well purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and all other
Molecular Biology (9)
") Molecular Biology (9) Translation Mamoun Ahram, PhD Second semester, 2017-2018 1 Resources This lecture Cooper, Ch. 8 (297-319) 2 General information Protein synthesis involves interactions between three
Molecular Biology (9) Translation Mamoun Ahram, PhD Second semester, 2017-2018 1 Resources This lecture Cooper, Ch. 8 (297-319) 2 General information Protein synthesis involves interactions between three
Prokaryotic Regulation
 Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Hybrid Gold Superstructures: Synthesis and. Specific Cell Surface Protein Imaging Applications
 Supporting Information Hybrid Gold Nanocube@Silica@Graphene-Quantum-Dot Superstructures: Synthesis and Specific Cell Surface Protein Imaging Applications Liu Deng, Ling Liu, Chengzhou Zhu, Dan Li and Shaojun
Supporting Information Hybrid Gold Nanocube@Silica@Graphene-Quantum-Dot Superstructures: Synthesis and Specific Cell Surface Protein Imaging Applications Liu Deng, Ling Liu, Chengzhou Zhu, Dan Li and Shaojun
Energy and Cellular Metabolism
 1 Chapter 4 About This Chapter Energy and Cellular Metabolism 2 Energy in biological systems Chemical reactions Enzymes Metabolism Figure 4.1 Energy transfer in the environment Table 4.1 Properties of
1 Chapter 4 About This Chapter Energy and Cellular Metabolism 2 Energy in biological systems Chemical reactions Enzymes Metabolism Figure 4.1 Energy transfer in the environment Table 4.1 Properties of
9/11/18. Molecular and Cellular Biology. 3. The Cell From Genes to Proteins. key processes
 Molecular and Cellular Biology Animal Cell ((eukaryotic cell) -----> compare with prokaryotic cell) ENDOPLASMIC RETICULUM (ER) Rough ER Smooth ER Flagellum Nuclear envelope Nucleolus NUCLEUS Chromatin
Molecular and Cellular Biology Animal Cell ((eukaryotic cell) -----> compare with prokaryotic cell) ENDOPLASMIC RETICULUM (ER) Rough ER Smooth ER Flagellum Nuclear envelope Nucleolus NUCLEUS Chromatin
Reception The target cell s detection of a signal coming from outside the cell May Occur by: Direct connect Through signal molecules
 Why Do Cells Communicate? Regulation Cells need to control cellular processes In multicellular organism, cells signaling pathways coordinate the activities within individual cells that support the function
Why Do Cells Communicate? Regulation Cells need to control cellular processes In multicellular organism, cells signaling pathways coordinate the activities within individual cells that support the function
Toxicological Targets. Russell L. Carr Department of Basic Sciences College of Veterinary Medicine
 Toxicological Targets Russell L. Carr Department of Basic Sciences College of Veterinary Medicine Toxicology Definitions = study of poisons Poison = any agent capable of producing a deleterious response
Toxicological Targets Russell L. Carr Department of Basic Sciences College of Veterinary Medicine Toxicology Definitions = study of poisons Poison = any agent capable of producing a deleterious response
Effect of enzyme deficiencies on oxidative phosphorylation: from isolated mitochondria to intact tissues. Theoretical studies.
 Effect of enzyme deficiencies on oxidative phosphorylation: from isolated mitochondria to intact tissues. Theoretical studies. Bernard Korzeniewski Institute of Molecular Biology and Biotechnology, Jagiellonian
Effect of enzyme deficiencies on oxidative phosphorylation: from isolated mitochondria to intact tissues. Theoretical studies. Bernard Korzeniewski Institute of Molecular Biology and Biotechnology, Jagiellonian
Name Period The Control of Gene Expression in Prokaryotes Notes
 Bacterial DNA contains genes that encode for many different proteins (enzymes) so that many processes have the ability to occur -not all processes are carried out at any one time -what allows expression
Bacterial DNA contains genes that encode for many different proteins (enzymes) so that many processes have the ability to occur -not all processes are carried out at any one time -what allows expression
Name: SBI 4U. Gene Expression Quiz. Overall Expectation:
 Gene Expression Quiz Overall Expectation: - Demonstrate an understanding of concepts related to molecular genetics, and how genetic modification is applied in industry and agriculture Specific Expectation(s):
Gene Expression Quiz Overall Expectation: - Demonstrate an understanding of concepts related to molecular genetics, and how genetic modification is applied in industry and agriculture Specific Expectation(s):
camp Direct Immunoassay Kit
 camp Direct Immunoassay Kit Catalog Number KA0886 100 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
camp Direct Immunoassay Kit Catalog Number KA0886 100 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Cellular Neuroanatomy I The Prototypical Neuron: Soma. Reading: BCP Chapter 2
 Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
-14. -Abdulrahman Al-Hanbali. -Shahd Alqudah. -Dr Ma mon Ahram. 1 P a g e
 -14 -Abdulrahman Al-Hanbali -Shahd Alqudah -Dr Ma mon Ahram 1 P a g e In this lecture we will talk about the last stage in the synthesis of proteins from DNA which is translation. Translation is the process
-14 -Abdulrahman Al-Hanbali -Shahd Alqudah -Dr Ma mon Ahram 1 P a g e In this lecture we will talk about the last stage in the synthesis of proteins from DNA which is translation. Translation is the process
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
Quiz answers. Allele. BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA)
 Lecture 17: The Quiz (and back to Eukaryotic DNA)") BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA) http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Quiz answers Kinase: An enzyme
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA) http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Quiz answers Kinase: An enzyme
Topic 4 - #14 The Lactose Operon
 Topic 4 - #14 The Lactose Operon The Lactose Operon The lactose operon is an operon which is responsible for the transport and metabolism of the sugar lactose in E. coli. - Lactose is one of many organic
Topic 4 - #14 The Lactose Operon The Lactose Operon The lactose operon is an operon which is responsible for the transport and metabolism of the sugar lactose in E. coli. - Lactose is one of many organic
Scale in the biological world
 Scale in the biological world 2 A cell seen by TEM 3 4 From living cells to atoms 5 Compartmentalisation in the cell: internal membranes and the cytosol 6 The Origin of mitochondria: The endosymbion hypothesis
Scale in the biological world 2 A cell seen by TEM 3 4 From living cells to atoms 5 Compartmentalisation in the cell: internal membranes and the cytosol 6 The Origin of mitochondria: The endosymbion hypothesis
Regulation of Transcription in Eukaryotes
 Regulation of Transcription in Eukaryotes Leucine zipper and helix-loop-helix proteins contain DNA-binding domains formed by dimerization of two polypeptide chains. Different members of each family can
Regulation of Transcription in Eukaryotes Leucine zipper and helix-loop-helix proteins contain DNA-binding domains formed by dimerization of two polypeptide chains. Different members of each family can
Eukaryotic vs. Prokaryotic genes
 BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 18: Eukaryotic genes http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Eukaryotic vs. Prokaryotic genes Like in prokaryotes,
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 18: Eukaryotic genes http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Eukaryotic vs. Prokaryotic genes Like in prokaryotes,
What is the central dogma of biology?
 Bellringer What is the central dogma of biology? A. RNA DNA Protein B. DNA Protein Gene C. DNA Gene RNA D. DNA RNA Protein Review of DNA processes Replication (7.1) Transcription(7.2) Translation(7.3)
Bellringer What is the central dogma of biology? A. RNA DNA Protein B. DNA Protein Gene C. DNA Gene RNA D. DNA RNA Protein Review of DNA processes Replication (7.1) Transcription(7.2) Translation(7.3)
Arabidopsis PPR40 connects abiotic stress responses to mitochondrial electron transport
 Ph.D. thesis Arabidopsis PPR40 connects abiotic stress responses to mitochondrial electron transport Zsigmond Laura Supervisor: Dr. Szabados László Arabidopsis Molecular Genetic Group Institute of Plant
Ph.D. thesis Arabidopsis PPR40 connects abiotic stress responses to mitochondrial electron transport Zsigmond Laura Supervisor: Dr. Szabados László Arabidopsis Molecular Genetic Group Institute of Plant
Welcome to Class 21!
 Welcome to Class 21! Introductory Biochemistry! Lecture 21: Outline and Objectives l Regulation of Gene Expression in Prokaryotes! l transcriptional regulation! l principles! l lac operon! l trp attenuation!
Welcome to Class 21! Introductory Biochemistry! Lecture 21: Outline and Objectives l Regulation of Gene Expression in Prokaryotes! l transcriptional regulation! l principles! l lac operon! l trp attenuation!
The mir-301a story: Regulation of mitochondria by microrna
 The mir-301a story: Regulation of mitochondria by microrna INSTITUTE OF BIOTECHNOLOGY, ASCR, LABORATORY OF TUMOUR RESISTANCE SANDRA LETTLOVÁ, ZUZANA RYCHTARČÍKOVÁ, VERONIKA TOMKOVÁ, MAGDALENA VONDRUSOVÁ,
The mir-301a story: Regulation of mitochondria by microrna INSTITUTE OF BIOTECHNOLOGY, ASCR, LABORATORY OF TUMOUR RESISTANCE SANDRA LETTLOVÁ, ZUZANA RYCHTARČÍKOVÁ, VERONIKA TOMKOVÁ, MAGDALENA VONDRUSOVÁ,
MOLECULAR CELL BIOLOGY
 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p
 Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p.110-114 Arrangement of information in DNA----- requirements for RNA Common arrangement of protein-coding genes in prokaryotes=
Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p.110-114 Arrangement of information in DNA----- requirements for RNA Common arrangement of protein-coding genes in prokaryotes=
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Multiple Choice Review- Eukaryotic Gene Expression
 Multiple Choice Review- Eukaryotic Gene Expression 1. Which of the following is the Central Dogma of cell biology? a. DNA Nucleic Acid Protein Amino Acid b. Prokaryote Bacteria - Eukaryote c. Atom Molecule
Multiple Choice Review- Eukaryotic Gene Expression 1. Which of the following is the Central Dogma of cell biology? a. DNA Nucleic Acid Protein Amino Acid b. Prokaryote Bacteria - Eukaryote c. Atom Molecule
Lecture 7 Cell Biolog y ٢٢٢ ١
 Lecture 7 ١ Mitochondria ٢ Mitochondria Mitochondria are the energy factories of the cells. The energy currency for the work that animals must do is the energy-rich molecule adenosine triphosphate (ATP).
Lecture 7 ١ Mitochondria ٢ Mitochondria Mitochondria are the energy factories of the cells. The energy currency for the work that animals must do is the energy-rich molecule adenosine triphosphate (ATP).
Introduction to molecular biology. Mitesh Shrestha
 Introduction to molecular biology Mitesh Shrestha Molecular biology: definition Molecular biology is the study of molecular underpinnings of the process of replication, transcription and translation of
Introduction to molecular biology Mitesh Shrestha Molecular biology: definition Molecular biology is the study of molecular underpinnings of the process of replication, transcription and translation of
Honors Biology Reading Guide Chapter 11
 Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section B and ONE question from Section C.
 UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2015-2016 FUNDAMENTALS OF CELL BIOLOGY AND BIOCHEMISTRY BIO-4004B Time allowed: 2 hours Answer ALL questions in Section
UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2015-2016 FUNDAMENTALS OF CELL BIOLOGY AND BIOCHEMISTRY BIO-4004B Time allowed: 2 hours Answer ALL questions in Section
Chapter 15 Active Reading Guide Regulation of Gene Expression
 Name: AP Biology Mr. Croft Chapter 15 Active Reading Guide Regulation of Gene Expression The overview for Chapter 15 introduces the idea that while all cells of an organism have all genes in the genome,
Name: AP Biology Mr. Croft Chapter 15 Active Reading Guide Regulation of Gene Expression The overview for Chapter 15 introduces the idea that while all cells of an organism have all genes in the genome,
Written Exam 15 December Course name: Introduction to Systems Biology Course no
 Technical University of Denmark Written Exam 15 December 2008 Course name: Introduction to Systems Biology Course no. 27041 Aids allowed: Open book exam Provide your answers and calculations on separate
Technical University of Denmark Written Exam 15 December 2008 Course name: Introduction to Systems Biology Course no. 27041 Aids allowed: Open book exam Provide your answers and calculations on separate
Degeneracy. Two types of degeneracy:
 Degeneracy The occurrence of more than one codon for an amino acid (AA). Most differ in only the 3 rd (3 ) base, with the 1 st and 2 nd being most important for distinguishing the AA. Two types of degeneracy:
Degeneracy The occurrence of more than one codon for an amino acid (AA). Most differ in only the 3 rd (3 ) base, with the 1 st and 2 nd being most important for distinguishing the AA. Two types of degeneracy:
RANK. Alternative names. Discovery. Structure. William J. Boyle* SUMMARY BACKGROUND
 RANK William J. Boyle* Department of Cell Biology, Amgen, Inc., One Amgen Center Drive, Thousand Oaks, CA 91320-1799, USA * corresponding author tel: 805-447-4304, fax: 805-447-1982, e-mail: bboyle@amgen.com
RANK William J. Boyle* Department of Cell Biology, Amgen, Inc., One Amgen Center Drive, Thousand Oaks, CA 91320-1799, USA * corresponding author tel: 805-447-4304, fax: 805-447-1982, e-mail: bboyle@amgen.com
7.06 Problem Set
 7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
NAD + /NADH Assay [Colorimetric]
![NAD + /NADH Assay [Colorimetric]](/thumbs/91/105496846.jpg "NAD + /NADH Assay [Colorimetric]") G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name NAD + /NADH Assay [Colorimetric] (Cat. #786 1539, 786 1540) think proteins! think G-Biosciences
G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name NAD + /NADH Assay [Colorimetric] (Cat. #786 1539, 786 1540) think proteins! think G-Biosciences
9 The Process of Translation
 9 The Process of Translation 9.1 Stages of Translation Process We are familiar with the genetic code, we can begin to study the mechanism by which amino acids are assembled into proteins. Because more
9 The Process of Translation 9.1 Stages of Translation Process We are familiar with the genetic code, we can begin to study the mechanism by which amino acids are assembled into proteins. Because more
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
Chapter
 Chapter 17 17.4-17.6 Molecular Components of Translation A cell interprets a genetic message and builds a polypeptide The message is a series of codons on mrna The interpreter is called transfer (trna)
Chapter 17 17.4-17.6 Molecular Components of Translation A cell interprets a genetic message and builds a polypeptide The message is a series of codons on mrna The interpreter is called transfer (trna)
2. Yeast two-hybrid system
 2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
Quantification of Protein Half-Lives in the Budding Yeast Proteome
 Supporting Methods Quantification of Protein Half-Lives in the Budding Yeast Proteome 1 Cell Growth and Cycloheximide Treatment Three parallel cultures (17 ml) of each TAP-tagged strain were grown in separate
Supporting Methods Quantification of Protein Half-Lives in the Budding Yeast Proteome 1 Cell Growth and Cycloheximide Treatment Three parallel cultures (17 ml) of each TAP-tagged strain were grown in separate
Proteomics. Areas of Interest
 Introduction to BioMEMS & Medical Microdevices Proteomics and Protein Microarrays Companion lecture to the textbook: Fundamentals of BioMEMS and Medical Microdevices, by Prof., http://saliterman.umn.edu/
Introduction to BioMEMS & Medical Microdevices Proteomics and Protein Microarrays Companion lecture to the textbook: Fundamentals of BioMEMS and Medical Microdevices, by Prof., http://saliterman.umn.edu/
Supplemental material
 Supplemental material THE JOURNAL OF CELL BIOLOGY Mourier et al., http://www.jcb.org/cgi/content/full/jcb.201411100/dc1 Figure S1. Size and mitochondrial content in Mfn1 and Mfn2 knockout hearts. (A) Body
Supplemental material THE JOURNAL OF CELL BIOLOGY Mourier et al., http://www.jcb.org/cgi/content/full/jcb.201411100/dc1 Figure S1. Size and mitochondrial content in Mfn1 and Mfn2 knockout hearts. (A) Body
Biology 112 Practice Midterm Questions
 Biology 112 Practice Midterm Questions 1. Identify which statement is true or false I. Bacterial cell walls prevent osmotic lysis II. All bacterial cell walls contain an LPS layer III. In a Gram stain,
Biology 112 Practice Midterm Questions 1. Identify which statement is true or false I. Bacterial cell walls prevent osmotic lysis II. All bacterial cell walls contain an LPS layer III. In a Gram stain,
Chapter 17. From Gene to Protein. Biology Kevin Dees
 Chapter 17 From Gene to Protein DNA The information molecule Sequences of bases is a code DNA organized in to chromosomes Chromosomes are organized into genes What do the genes actually say??? Reflecting
Chapter 17 From Gene to Protein DNA The information molecule Sequences of bases is a code DNA organized in to chromosomes Chromosomes are organized into genes What do the genes actually say??? Reflecting
Types of RNA. 1. Messenger RNA(mRNA): 1. Represents only 5% of the total RNA in the cell.
: 1. Represents only 5% of the total RNA in the cell.") RNAs L.Os. Know the different types of RNA & their relative concentration Know the structure of each RNA Understand their functions Know their locations in the cell Understand the differences between prokaryotic
RNAs L.Os. Know the different types of RNA & their relative concentration Know the structure of each RNA Understand their functions Know their locations in the cell Understand the differences between prokaryotic
Lesson Overview. Ribosomes and Protein Synthesis 13.2
 13.2 The Genetic Code The first step in decoding genetic messages is to transcribe a nucleotide base sequence from DNA to mrna. This transcribed information contains a code for making proteins. The Genetic
13.2 The Genetic Code The first step in decoding genetic messages is to transcribe a nucleotide base sequence from DNA to mrna. This transcribed information contains a code for making proteins. The Genetic
4) Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.
 Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.") Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Chapter 17 The Mechanism of Translation I: Initiation
 Chapter 17 The Mechanism of Translation I: Initiation Focus only on experiments discussed in class. Completely skip Figure 17.36 Read pg 521-527 up to the sentence that begins "In 1969, Joan Steitz..."
Chapter 17 The Mechanism of Translation I: Initiation Focus only on experiments discussed in class. Completely skip Figure 17.36 Read pg 521-527 up to the sentence that begins "In 1969, Joan Steitz..."
NucView TM 488 Caspase-3 Assay Kit for Live Cells
 NucView TM 488 Caspase-3 Assay Kit for Live Cells Catalog Number: 30029 (100-500 assays) Contact Information Address: Biotium, Inc. 3423 Investment Blvd. Suite 8 Hayward, CA 94545 USA Telephone: (510)
NucView TM 488 Caspase-3 Assay Kit for Live Cells Catalog Number: 30029 (100-500 assays) Contact Information Address: Biotium, Inc. 3423 Investment Blvd. Suite 8 Hayward, CA 94545 USA Telephone: (510)
Biochemical bases for energy transformations. Biochemical bases for energy transformations. Nutrition 202 Animal Energetics R. D.
 Biochemical bases for energy transformations Biochemical bases for energy transformations Nutrition 202 Animal Energetics R. D. Sainz Lecture 02 Energy originally from radiant sun energy Captured in chemical
Biochemical bases for energy transformations Biochemical bases for energy transformations Nutrition 202 Animal Energetics R. D. Sainz Lecture 02 Energy originally from radiant sun energy Captured in chemical
Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition
 Supplementary Information to Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition Nadine Czudnochowski 1,2, *, Christian A. Bösken 1, * & Matthias Geyer 1 1 Max-Planck-Institut
Supplementary Information to Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition Nadine Czudnochowski 1,2, *, Christian A. Bösken 1, * & Matthias Geyer 1 1 Max-Planck-Institut
U.S. Patent No. 9,051,563 and other pending patents. Ver
 INSTRUCTION MANUAL Direct-zol RNA MiniPrep Catalog Nos. R050, R05, R05, & R053 Highlights Quick, spin column purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and other
INSTRUCTION MANUAL Direct-zol RNA MiniPrep Catalog Nos. R050, R05, R05, & R053 Highlights Quick, spin column purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and other
Gene Control Mechanisms at Transcription and Translation Levels
 Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
Regulation of gene expression. Premedical - Biology
 Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Lecture 13: PROTEIN SYNTHESIS II- TRANSLATION
 http://smtom.lecture.ub.ac.id/ Password: https://syukur16tom.wordpress.com/ Password: Lecture 13: PROTEIN SYNTHESIS II- TRANSLATION http://hyperphysics.phy-astr.gsu.edu/hbase/organic/imgorg/translation2.gif
http://smtom.lecture.ub.ac.id/ Password: https://syukur16tom.wordpress.com/ Password: Lecture 13: PROTEIN SYNTHESIS II- TRANSLATION http://hyperphysics.phy-astr.gsu.edu/hbase/organic/imgorg/translation2.gif
M i t o c h o n d r i a
 M i t o c h o n d r i a Dr. Diala Abu-Hassan School of Medicine dr.abuhassand@gmail.com Mitochondria Function: generation of metabolic energy in eukaryotic cells Generation of ATP from the breakdown of
M i t o c h o n d r i a Dr. Diala Abu-Hassan School of Medicine dr.abuhassand@gmail.com Mitochondria Function: generation of metabolic energy in eukaryotic cells Generation of ATP from the breakdown of
Nitric Oxide Synthase Ultrasensitive Colorimetric Assay
 Package Insert Nitric Oxide Synthase Ultrasensitive Colorimetric Assay 96 Wells For Research Use Only v. 2.0 09.20.17 Eagle Biosciences, Inc. 20A NW Blvd., Suite 112, Nashua, NH 03063 Phone: 866-419-2019
Package Insert Nitric Oxide Synthase Ultrasensitive Colorimetric Assay 96 Wells For Research Use Only v. 2.0 09.20.17 Eagle Biosciences, Inc. 20A NW Blvd., Suite 112, Nashua, NH 03063 Phone: 866-419-2019
Biology I Fall Semester Exam Review 2014
 Biology I Fall Semester Exam Review 2014 Biomolecules and Enzymes (Chapter 2) 8 questions Macromolecules, Biomolecules, Organic Compunds Elements *From the Periodic Table of Elements Subunits Monomers,
Biology I Fall Semester Exam Review 2014 Biomolecules and Enzymes (Chapter 2) 8 questions Macromolecules, Biomolecules, Organic Compunds Elements *From the Periodic Table of Elements Subunits Monomers,
Chapter 6- An Introduction to Metabolism*
 Chapter 6- An Introduction to Metabolism* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Energy of Life
Chapter 6- An Introduction to Metabolism* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Energy of Life
Supplementary Information
 Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Number of questions TEK (Learning Target) Biomolecules & Enzymes
 Biomolecules & Enzymes") Unit Biomolecules & Enzymes Number of questions TEK (Learning Target) on Exam 8 questions 9A I can compare and contrast the structure and function of biomolecules. 9C I know the role of enzymes and how
Unit Biomolecules & Enzymes Number of questions TEK (Learning Target) on Exam 8 questions 9A I can compare and contrast the structure and function of biomolecules. 9C I know the role of enzymes and how
REVIEW SESSION. Wednesday, September 15 5:30 PM SHANTZ 242 E
 REVIEW SESSION Wednesday, September 15 5:30 PM SHANTZ 242 E Gene Regulation Gene Regulation Gene expression can be turned on, turned off, turned up or turned down! For example, as test time approaches,
REVIEW SESSION Wednesday, September 15 5:30 PM SHANTZ 242 E Gene Regulation Gene Regulation Gene expression can be turned on, turned off, turned up or turned down! For example, as test time approaches,
Lipniacki 2004 Ground Truth
 Abstract Lipniacki 2004 Ground Truth The two-feedback-loop regulatory module of nuclear factor kb (NF-kB) signaling pathway is modeled by means of ordinary differential equations. signaling pathway: https://en.wikipedia.org/wiki/signaling_pathway
Abstract Lipniacki 2004 Ground Truth The two-feedback-loop regulatory module of nuclear factor kb (NF-kB) signaling pathway is modeled by means of ordinary differential equations. signaling pathway: https://en.wikipedia.org/wiki/signaling_pathway
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
CHAPTER : Prokaryotic Genetics
 CHAPTER 13.3 13.5: Prokaryotic Genetics 1. Most bacteria are not pathogenic. Identify several important roles they play in the ecosystem and human culture. 2. How do variations arise in bacteria considering
CHAPTER 13.3 13.5: Prokaryotic Genetics 1. Most bacteria are not pathogenic. Identify several important roles they play in the ecosystem and human culture. 2. How do variations arise in bacteria considering
mrna Isolation Kit for Blood/Bone Marrow For isolation mrna from blood or bone marrow lysates Cat. No
 For isolation mrna from blood or bone marrow lysates Cat. No. 1 934 333 Principle Starting material Application Time required Results Key advantages The purification of mrna requires two steps: 1. Cells
For isolation mrna from blood or bone marrow lysates Cat. No. 1 934 333 Principle Starting material Application Time required Results Key advantages The purification of mrna requires two steps: 1. Cells
Figure S1: Extracellular nicotinic acid, but not tryptophan, is sufficient to maintain
 SUPPLEMENTAL INFORMATION Supplemental Figure Legends Figure S1: Extracellular nicotinic acid, but not tryptophan, is sufficient to maintain mitochondrial NAD +. A) Extracellular tryptophan, even at 5 µm,
SUPPLEMENTAL INFORMATION Supplemental Figure Legends Figure S1: Extracellular nicotinic acid, but not tryptophan, is sufficient to maintain mitochondrial NAD +. A) Extracellular tryptophan, even at 5 µm,