Internally generated preactivation of single neurons in human medial frontal cortex predicts volition
|
|
|
- Sheena Underwood
- 5 years ago
- Views:
Transcription
1 Internally generated preactivation of single neurons in human medial frontal cortex predicts volition Itzhak Fried, Roy Mukamel, Gabriel Kreiman List of supplementary material Supplementary Tables (2) Supplementary Experimental Procedures Supplementary Figures (8) Supplementary Movie (1) 1
2 Table S1 (expanding on Table 1) PART A Temporal Lobe ACCr ACCd pre-sma SMA proper Total # of recorded cells Increase in rate ( I ) Decrease in rate ( D ) I + D Table S1, PART A: Anatomical distribution of units within the medial frontal lobe showing increase in firing rate ( I ) and decrease in firing rate ( D ) with respect to baseline. I + D indicates the sum of units showing I or D responses. SMA proper supplementary motor area proper; pre-sma pre-supplementary motor area, ACCd dorsal anterior cingulate cortex, ACCr rostral anterior cingulate cortex. See Figure S6 for MR images depicting electrode locations. 2
3 Table S1 (expanding on Table 1) PART B SUB ACCr ACCd SMA proper pre-sma TOTAL All MUA+SU A I D All MUA+SU A I D All MUA+SU A I D All MUA+SUA I D All MUA SUA I D Tot Table S1, PART B: Anatomical distribution of units within the medial frontal lobe in each subject. For each subject and location (see Part A for location abbreviations), we indicate the number of MUA units and SUA units and the number of units that showed an increase ( I ) or decrease ( D ) in firing rate as W was approached. 3
4 Table S2 (expanding on Table 1) Location A H EC ST PHG ACCr 8x10-5 2x10-5 4x10-6 8x10-5 2x10-5 ACCd 7x10-4 3x10-4 6x10-5 3x10-3 7x10-4 psma 9x10-4 3x10-4 5x10-5 4x10-3 9x10-4 SMA 5x10-9 9x x x10-8 5x10-9 Location ACCr ACCd psma SMA ACCr ACCd psma SMA 2x10-2 6x10-3 1x10-3 Comparison of the proportion of units that showed a statistically significant response before W among different areas using a binomial test. A significant response was defined by a ranksum test, p<0.01, 400 ms before W against baseline (Experimental Procedures). Let N 1, N 2 indicate the total number of units recorded from in area 1 and 2 respectively and let n 1, n 2 indicate the number of units that showed a significant response before W. f 1 =n 1 /N 1 and f 2 =n 2 /N 2 define the proportion of responsive units in each area. Under the null hypothesis, we assume that the two areas are indistinguishable and we compute the probability of obtaining n 1 or more responsive units (out of the total of N 1 units) assuming the proportion of responsive units in area 2 as p = N k =n 1 N 1 k (1 f 2 ) k f k 2. This table reports the p values for each pair of areas. Note that SMA has a significantly higher proportion of neurons responding before W compared to all other regions. The corresponding values of n 1, n 2, N 1, N 2 are reported in Table 1. 4
5 Supplementary Experimental Procedures EMG recordings We recorded electromyographic (EMG) signals in three healthy volunteer. We used EMG electrodes (Conmed Corporation, Billerica, MA) placed over the flexor digitorum sperficialis muscles. The data were recorded with the MicrguidePro system from Alpha Omega (Alpharetta, GA) with a 3 khz sampling rate and 500x amplification factor. The EMG data are shown in Figure S3C-D. Data analysis Classification of individual units. Baseline firing rate was defined as the spike count in the window from ms to ms relative to W (see also Figure S3E for other definitions of the baseline interval). We compared the firing rate during baseline with the firing rate in two different time windows using a non-parametric ranksum test and a threshold criterion of p < 0.01 (similar results were observed using a paired two-tailed t-test). The temporal windows were as follows: 1) 400 ms to 0 ms relative to W; 2) 0 ms to 400 ms following W. An analysis using a sliding window is presented in Figure 4. In Figure S1 we compared the changes in firing rate against those expected under three different null hypotheses (5000 iterations): (1) creating for each unit a surrogate spike train with the same firing rate as the actual unit but with the spike times governed by a homogeneous Poisson process (see also Figure 2C); (2) similar to (1) but here the surrogate spike train conserved not only the firing rate but also the interspike interval distribution of the actual spike trains and (3) randomly shifting W. We classified the response of all 128 units responding significantly before W (Table 1) as either showing increase in firing rate with respect to baseline ( I, n = 55) or decrease in firing rate with respect to baseline ( D, n = 73). To plot Figure 4A-C and S1G, the responses were normalized by subtracting the baseline activity and dividing by the maximum firing rate for I cells (or dividing by the absolute value of the minimum firing rate for D cells). After normalization, the responses were averaged. Statistical classifier. Figures 5-7 in the main text as well as Figures S4 use a Support Vector Machine (SVM) (Hung et al., 2005) classifier to quantify whether the neuronal ensemble showed changes in their firing patterns before W. The classifier yields a measure of performance at the single-trial level, as opposed to the typical Bereitschaftspotential averaged over a large number of repetitions (Colebatch, 2007; Erdler et al., 2000; Haggard and Eimer, 1999; Libet et al., 1983; Ohara et al., 2006; Yazawa et al., 2000). In Figures 5B, 5 and S4, we asked whether the classifier could discriminate the neuronal responses from baseline activity at a time t prior to W. Let represent the spike train for a given unit u on a given trial r ( τ i are the spike times). We binned the spikes in windows of size t r (the default value of t r =400 ms was 5
6 used throughout the text; the results were robust to changes in this parameter). Let indicate the spike count in a window of size t r centered at time t. For a population of n neurons, we assumed independent firing and we constructed a population vector by concatenating the responses of all units:. The number of units, n, depends on the specific analysis and is indicted in each figure legend. The baseline response was defined as c -2300, i.e., the spike count from to ms with respect to W. At any given time t, the input to the classifier consisted of the example vectors, r p t, containing the spike counts from the neuronal population for the training trials r associated with a label +1 and the baseline example vectors, r p -2300, associated with the label -1 (Figure S5A shows a simple example in the case of n=2 units). A binary classifier was trained to discriminate between the +1 and -1 examples, that is, to quantify whether the neuronal ensemble activity could be distinguished from baseline at a particular time and on a trial-by-trial basis. We used a linear kernel; in this case, the classifier boundary can be expressed as where the vector p denotes the ensemble response and the weights w are learnt during training. Other statistical classifiers and SVM kernels yielded similar results. The dimension of the input (the number of units n) is indicated for each figure in the text. We used a cross-validation procedure whereby we randomly chose 70% of the trials to be used for training and the remaining 30% of the trials were used to evaluate the classifier performance. This procedure was repeated in each one of 100 iterations. Importantly, the performance of the classifier was evaluated with independent data that was not seen by the classifier during training (i.e., there was no overlap between the training and test data). The performance of the classifier at time t indicates the percentage of test trials correctly discriminated from baseline at a time t prior to W. In those cases where we used a subset of the total number of units, we also randomly chose the units from the available population in each iteration. This was done, for example, in order to ensure that comparisons across areas were fair and not dictated by the higher number of recordings in one area. Error bars in the classifier performance plots denote one standard error and are based on this cross-validation procedure. Right versus left hemisphere. The decoding results discussed throughout the main text pool together units from both the right and left hemisphere. We also considered the performance of the classifier using separate neuronal subpopulations from either the right or left hemispheres in the task where subjects used only the right index finger (Figure 6D). We did not observe any significant difference in decoding performance between these two subpopulations. Single units versus multi-units. We divided the output spiking activity into multi-units (MUA, which cannot reliably be separated any further by the algorithm) and single-units (SUA) using the automatic criteria described in (Tankus et al., 2009). The plots in the main text and supplementary figures do not discriminate between SUA and MUA. We show the results obtained upon separating the SUA and MUA in Figure 6E. Essentially, both types of signals yielded similar results. 6
7 Prediction of W time. Figure 7 in the main text describes the performance of the classifier in predicting the time of volition onset (W). We bin the spike trains in windows of size t r (the default value of t r =400 ms was used throughout the text). Let indicate the spike count for unit u during trial r in a window of size t r centered at time t (Figure 7B). For a population of n neurons, we assumed independent firing and we constructed a population vector by concatenating the responses of all n units:. The time t was shifted in steps of 100 ms and therefore the spike count windows overlapped in time. For an ensemble of simultaneously recorded units, the spike trains were aligned to trial onset at t=0. When considering multiple recording sessions or multiple subjects, responses were aligned to W. Starting at t=-3500 ms, we constructed the vectors r p W-3500, r p W-3400,, r p W Each one of these vectors was associated with an indicator variable L(t) that labeled each window of size t r by +1 or -1 depending on the distance to W: L(t)=-1 if t<w-t b and L(t)=+1 if t W-t b. An SVM classifier was trained to learn the map between p and L (Hung et al., 2005). The performance of the classifier was tested using cross-validation by using new trials not seen by the classifier during training. The predicted urge/decision time,, was defined as the first time point when the classifier prediction on independent test data indicated L(t)=+1 in 3 out of 4 consecutive windows. Figure 7D in the main text shows the distribution of. Accuracy of W. Reporting W accurately is not trivial. Therefore, it is expected that there could be a variation between the reported W and the internal onset of the decision/urge to move. Unfortunately, it is not easy to estimate this variability (Joordens et al., 2002). We first reanalyzed all the data after assuming that W was reported in a random fashion. For this purpose, W was chosen randomly from a uniform distribution starting one clock revolution after trial onset and ending at P. This drastically reduced the number of significant units (Figure S1) and the classifier performance was close to chance levels. In order to further quantify the impact of changes in W time on the spiking responses and our analyses, we simulated inaccuracies in W by adding a fixed temporal bias (Figure S4D1) or random jitter (Figure S4D2) to W. This jitter was taken from a Gaussian distribution with zero mean and a standard deviation σ, with the constraint that W always had to occur before P. This analysis suggests that inaccuracies in W of several hundred ms would have led to a large decrease in the number of neurons responding before W. In particular, inaccuracies of several hundred ms would bring the results very close to chance levels. Yet, it is possible that there could be inaccuracy in W on the order of up to approximately 200 ms without impacting the overall number of significant neurons. Integrate-and-fire model. We speculate in the main text that the urge/decision may arise when a threshold is crossed after a cumulative increase in activity in the medial frontal lobe neuronal ensemble (Crick and Koch, 2003). Here we present a quantitative model of how this mechanism could work by using an integrate and fire model unit receiving input from the medial frontal lobe units recorded from in this study. The basic circuit for a leaky integrate-and-fire unit is shown in Figure S5G. The input current I(t) is integrated through an RC circuit: where C is the capacitance, R is the resistance and V is the voltage. When the voltage reaches a threshold V thres, a spike is generated, the voltage is reset to zero and a refractory period t ref is imposed. Each spike generated an EPSC and I(t) was modeled here as the 7
8 sum of all the input EPSCs. The parameters for the simulation shown in Figure S5H-I are given in the legend. Although the input to the model is given by the I units, it would be easy to also incorporate the D units by adding an additional sign change through an inhibitory interneuron. 8
9 Supplementary Movie 1 (related to Fig. 1): Activity of one neuron in left pre-sma during three trials of the task. Top panel depicts the analogue clock the patients observed. Bottom blue rasters and auditory beeps represent the occurrence of spikes. The black trace represents the spike train depicted by the blue rasters, smoothed with a Gaussian kernel (σ = 80ms). Red bar represents the time of button press and green bar represents the reported time of urge (W). List of Supplementary Figures Figure S1: Number of responsive units under the null hypotheses (related to Table 1). For each unit, we generated surrogate data by considering three possible null model hypotheses: (i) Poisson (green): We created a homogeneous Poisson spike train containing the same number of spikes as the real data. (ii) ISI (blue): We created a surrogate spike train that maintained the interspike interval distribution of the real data. (iii) Random W (red): We kept the original spike trains but the W time was generated randomly. We analyzed the surrogate data using the same methods and criteria applied to the real data (Experimental Procedures) and computed the number of responsive units (x-axis in A-F). A responsive unit was defined as a unit that showed a statistically significant change in firing rate when comparing the 400 ms pre-w and the to ms baseline period based on a ranksum test (p<0.01, Experimental Procedures). The procedure was repeated 5000 times. The plots show the distribution of the number of responsive units in the medial frontal lobe (A) and temporal lobe (B) for all units (A1,B1), I units (A2,B2) and D units (A3,B3). Subplots C-F show the corresponding distributions for the 4 different areas within the frontal lobe (ACCr = rostral anterior cingulate cortex; ACCd = dorsal anterior cingulate cortex; pre-sma = presupplementary motor area; SMA = supplementary motor area). The vertical dashed line shows the mean of the distribution and the dotted lines show 3 standard deviations from the mean. The arrow indicates the number of responsive units in the actual data. G1-G3. Control for Figure 4A based on surrogate data for the Poisson null model (G1), ISI null model (G2) and Random W null model (G3). The procedure to generate surrogate spike trains is described in A-F. Here we use the surrogate data to generate the equivalent to Figure 4A in the main text. The plot shows the normalized firing rate for the I cells (red) and the D cells (blue). Responsive cells were defined based on comparing the pre-w response period with the baseline period (shaded rectangles). For comparison, the dotted lines reproduce the normalized firing rate curves from Figure 4A. Note that the slight increase (decrease) in the red (blue) curve before W is restricted to the time window used in the statistical selection criteria (unlike Figure 4A where the deviations from baseline are more pronounced and start well before the selection window). FIGURE S2: Gradual versus abrupt transitions in single trials (related to Figs. 1-3) A. Schematic example of a hypothetical unit that shows gradual changes in activity in individual trials leading to an gradual increase in firing rate in the average PSTH (right, average of 100 trials). 9
10 B. Schematic example of a hypothetical unit that shows abrupt changes in activity with variable transition times in individual trials leading to a gradual increase in firing rate in the average PSTH (right, average of 100 trials). C. To quantify the degree to which transitions in individual trials should be described as abrupt versus gradual, we considered the spike trains from individual trials in the 2500 ms preceding W after smoothing with a 200 ms width Gaussian. We fit a logistic function, f f (t) = W 1+ exp[ (t t 0 ) / α] + f, (where f B W and f B are the firing rates before W and during the baseline respectively, and t 0, α are free parameters to be fitted. Here we show the shape of the logistic function for different values of α showing abrupt changes (low values of α) or gradual changes (large values of α). D-E. Examples of individual trials (different units) showing the smoothed spike train (blue) and the logistic function fit (red). Note that the function was fit only in the 2500 ms preceding W. In each example, we show the value of α and the Pearson correlation coefficient between the fitted function and the smoothed spike train. The three examples in D show α values larger than the mean (more gradual transitions) whereas the three examples in E show α values smaller than the mean (more abrupt transitions). F. Distribution of fitted α values for n=55 units that showed increases in firing rate as W was approached (Table S1A). The dashed line and the dotted lines show the mean and SD of the distribution respectively. Note the logarithmic scale on the x-axis. G. Distribution of fitted t 0 values for the same n=55 units. H. Distribution of Pearson correlation coefficient (r) between the fitted logistic function and the smoothed spike train. We only considered those trials that yielded r>0.5 for the distributions in F-H. Figure S3: EMG, baseline and neuronal response dynamics (related to Fig. 1-3) A. Fraction of trials where P occurred within x milliseconds after the first revolution of the clock (where x is the value indicated in the x-axis). When x is very short, these trials could be considered to be cued trials where the end of the first revolution of the clock is the cue. As the plot shows, there were very few trials with P<1000 ms after the first revolution of the clock. Error bars indicate the range across all subjects. B. For the 6 responsive units where we had at least 10 repetitions with P < 1500 ms after the first revolution of the clock, we show the PSTHs aligned to W (t=0) for those trials with P<1500 ms (blue), those trials with P>5000 ms (red) and all trials (black). The numbers at the bottom of each subplot indicate the number of trials for each condition. We also indicate the location of each unit at the bottom of each subplot (see Table 1 for abbreviations). C. Example average EMG recordings from one session and one subject. We recorded EMG signals in three subjects (see Experimental Procedures for methods and EMG electrode locations). We note that we did not perform neurophysiological recordings in these three subjects. Here we show the EMG signals aligned to the key press events (t=0) and averaged across all trials (n=158). The gray lines denote one SEM. 10
11 D. Distribution of EMG to Key-Press latencies. In each trial, we computed the first time point when the EMG signal deviated from baseline by more than 5 standard deviations. This time point was defined at the EMG onset. Here we show the distribution of EMG onset to Key Press latencies across all three subjects and all recording sessions. Bin size = 5 ms. The vertical dashed line denotes the mean. E. Fraction of responsive units as a function of the time to W for different definitions of the baseline period (shaded rectangle): E1: to ms; E2: to ms; E3: to ms; E4: to ms (this is the baseline used throughout the text). A unit was considered to be responsive if the firing rate in the 400 ms centered on the time point reported on the x-axis was significantly different from the firing rate in the baseline period (two sided t test, p<0.05). The fraction of responsive units at each time point was computed by dividing the number of responsive units by the maximum number of responsive units across all baselines and time points (filled circle in E3). We indicate the average number of trials for each possible baseline definition. In E4, we point to the baseline and time point used in the numbers reported in the text and in Table 1. F. Comparison among trials when P-W<300 ms (blue), trials when P-W>600 ms (red) and all trials (black). Each subplot shows the PSTH for a separate unit aligned to W (left) or P (right). The format for the PSTHs is the same as in Figure 3 in the main text. The vertical lines indicate the mean W times (left) or mean P times (right); the color code for the means matches the one for the corresponding curves. The number of trials in each condition is indicated at the bottom of each subplot. G. Average normalized PSTH aligned to W (left) or P (right) for all the responsive I units (n=6) that had at least 5 trials with P-W>600 ms. Figure S4: Neuronal response onset correlates with W and P. Estimating the effect of inaccuracies in W (related to Fig.1-4). A. Example trial illustrating the definition of response onset in a single trial. Defining response onsets in single trials is not easy due to the intrinsic variability in neuronal firing. We used the following heuristic to define the response onsets. At each time point, t (step size = 40 ms), we defined fr(t) as the spike count in the time interval from t to t+400 ms and (that is, λ(t) takes the value +1 if the spike count increased in the last 40 ms and -1 otherwise). We considered a window w of 15 consecutive bins and we defined a score. The response onset was defined as the first time point where S 8. This time point is indicated by the blue vertical dashed line in this figure. The conclusions from this figure were not changed when we used different parameters to define the response onset. B. Three example units showing W versus response onset. Each circle corresponds to a separate trial. The black diagonal represents the identity line. The blue dashed line indicates the linear fit. For each responsive unit, we computed a correlation coefficient between W and the response onset and between P and response onset. The red dashed line indicates the time of first clock revolution. C. Distribution of correlation coefficients between W and response onset (C1) and between P and response onset (C2). Bin size = 0.1. The arrow indicates the mean of the distribution. There is no significant difference between these correlation coefficients for W and P. 11
12 D. We estimated the effect of inaccuracies in the W judgment. We simulated two types of inaccuracies: temporal shifts (D1) and random temporal jitter (D2). The effects were evaluated by computing the number of units that passed the same statistical criteria to be considered responsive and normalizing the results by the values reported in the text (which correspond to no temporal shift or temporal jitter). D1. The time of W was shifted by the amount indicated in the x-axis. Negative values indicate shifting to earlier time points and positive values indicate shifting to later time points. In each trial the maximum allowed positive shift was bounded by P. Because P was typically close to W (P- W=193±261 ms, mean±s.d. Figure 1) there are few points after shift=0 ms. The arrow shows the maximum number of significant units, which was obtained for a temporal shift of -50 ms. The red circles correspond to I cells (cells that increase their firing rate as W approaches), the blue circles correspond to D cells (cells that decrease their firing rate as W approaches) and the black circles include both I and D cells. D2. In each trial, the time of W was randomly jittered by adding or subtracting a time taken from a zero-mean Gaussian distribution and standard deviation σ (x-axis). The jitter was constrained to be such that W<P and that W was larger than the first revolution of the clock. After randomly jittering W in each trial, we repeated the same analysis to assess whether each unit shows significant pre-w response with respect to baseline. Here we show the fraction of units that show significance with respect to the σ=0 (no jitter) condition. The red circles correspond to I cells, the blue circles correspond to D cells and the black circles include both I and D cells. Inaccuracies in reporting W on the order of ~ 0 to 200 ms yield only a small decrease in the percentage of responsive units suggesting that subjects reported W with an accuracy of a few hundred ms. Results shown correspond to the mean±sd after 1000 iterations (to avoid clutter, error bars are shown only for the black circles). The horizontal dashed line corresponds to chance levels obtained by repeating the same analysis using surrogate spike trains (corresponding to the green curves in Figure S1). Figure S5: Decoding self-initiated movement in single trials and integrate-and-fire model (related to Fig. 5-6) A. Simplified version of the classifier task for n=2 units to illustrate the algorithm to decode volition in single trials. Responses of two units located in the pre-sma during the 400 ms before W (circles) or during the baseline interval from to ms (crosses). Many of the points overlap due to obtaining the same spike count in different repetitions; therefore, a single point in the figure may correspond to multiple repetitions. Also, the number of spikes for the circles and crosses could overlap; to avoid this and only for the purposes of the figure (not for the classifier) we added here a value of 0.2 to the circles and we subtracted 0.2 from the crosses. A fraction of the data on the left (70% of the repetitions) is used to train a linear classifier to learn to separate the circles and crosses (i.e., to separate the spike counts in the interval before W and baseline interval) by finding an appropriate hyperplane (a line in the 2D case with a linear classifier). On the right we show the same data and the classifier boundary (dashed line). The output of the classifier is indicated by the color of the symbols, red corresponds to circle predictions and blue corresponds to cross predictions. The overall performance of the classifier is given by the fraction of repetitions in the remaining 30% of test data that are correctly classified (red circles and blue crosses). In most of the figures in the main text, the procedure was applied to ensembles of many more than 2 units (see main text and Experimental Procedures for details). 12
13 B. Dependence of the classification performance on the size of the spike count window (referred to as t r in Experimental Procedures). The value highlighted in gray corresponds to the parameter used throughout the text. Here the classification performance is shown at two time points: 500 ms (green) and 1000 ms (red) before W. Error bars denote SEM. C. Classification performance for each individual subject and each location. The thin lines represent individual subjects and the thick line shows the average across all subjects in each location. In each subplot, we only included those subjects with at least 16 units in the corresponding location. D. Classification performance for each subject that had at least 16 units in the frontal lobe. Here we show the classification performance in the [-400,0] ms window. The error bars denote one standard deviation over randomizations of the units and repetitions (see text). The horizontal dashed line and its error bar on the right indicate the mean and range of the chance classification performance obtained by a permutation test (see text). Note that except for subject number 4, the classification performance was well above chance using data from individual subjects. Table S1B also describes the number of units recorded from in each subject and location as well as the number of units that showed significant responses for each individual subject and location. There is a significant amount of subject-to-subject variability. At least partly, these differences can be attributed to different number of electrodes and different recording locations across subjects. E. Decoding accuracy as a function of time to W using classifiers trained with different numbers of units. Spikes were counted in windows of 400 ms centered on the time point indicated on the x-axis. The time on the x-axis is measured with respect to W (dashed vertical line at t=0). Figure 6A in the main text reports the counterpart to this figure where we showed the performance of the classifier at two specific times as a function of the number of units. Here we show the time course of the decoding performance. Overall, it is remarkable that a minuscule fraction of the total number of neurons in the frontal lobe areas is sufficient to achieve strong classifier performance, and that even within this small group, decoding performance improves with the number of neurons used (Figure 6A). It is not unreasonable to expect that a decoder using more neurons could perform at even higher levels and perhaps even make more sophisticated behavioral predictions. However, we should note that the use of a SVM classifier here does not imply that the brain decodes the onset of volition using similar algorithms or classifiers (see also Figure S5). F. Classification performance separated by location (average across subjects). For each location (ACCr=green, ACCd=black, pre-sma=red, SMA=blue), we consider here only those subjects where we had at least 16 units. The numbers next to each curve indicate the number of subjects. The error bars denote one standard deviation across subjects. The horizontal dashed line indicates chance performance and the vertical dashed line denotes W. G. Schematic diagram of a basic model to illustrate how a leaky integration followed by a threshold mechanism could lead to the volitional onset. The input to the integrate-and-fire model here corresponds to the spiking activity of the units recorded from the human frontal lobe (shown schematically on the left). Each spike generates an EPSC and the input current to the model unit is the sum of all the input EPSCs. The leaky integrator fires a spike whenever the voltage crosses a threshold, and then resets the voltage back to zero. A refractory period is imposed 13
14 after each spike. Both excitatory and inhibitory units are used here as input (a minus sign is added to the synaptic weight for the inhibitory units). H. Distribution of the spike times for the I&F model with respect to the W time (n=40 trials). Bin size = 100 ms. The arrow indicates the mean of the distribution. I. Integrate-and-fire model (I&F) performance. Here we show the performance of the integrate and fire model illustrated in part A (parameters: C=32, R=256, α=0.01, v thres =1.6) during 2 trials (out of the 40 trials used to run the simulation). The first column shows the input to the model, consisting of the spikes of the frontal lobe units, including both excitatory and inhibitory ones. The green dashed line indicates W. The second column shows the activity of each unit during the 10 ms preceding a spike in the I&F model. Each row corresponds to a medial frontal cortex unit, each column corresponds to a 1 ms bin and the white marks denote a spike. The third column indicates the EPSCs, the input to the I&F, immediately preceding a spike. The fourth column shows the evolution of the intracellular voltage in the I&F unit as a function of time. The vertical green dashed line at t=0 indicates the W time. The horizontal dashed line shows the threshold for the I&F model, a spike (vertical black dashed line) is emitted when the voltage reaches this threshold. The number to the right in each trial indicates the time of the I&F spike relative to W. Figure S6. Anatomical location of electrodes in the frontal lobe displayed on individual MR images for each subject (related to Table 1). Color code: blue = SMA proper, red = pre-sma, yellow = ACCd, green = ACCr. Figure S7: Hand choice experiment (related to Figures 2-4). Three subjects performed a variant of the main experiment where they were allowed to choose not only the time of action but also which hand to use. A-C. Here we show examples of single unit responses during this variant of the task. The format is similar to the one in Figure 3. The plots show the raster plots and firing rates as a function of the time to W (t=0). The red traces (blue traces) show the average response when the subject executed the movement with the right hand (left hand). Above each subplot, an * indicates whether a two-tailed t-test between the blue and red curves was significant at the p<0.05 level in each time bin. We indicate the location of the electrode in each subplot. A1-A3 show examples of units that showed gradual changes in firing rate that were largely independent of the hand choice. B1-B3 show examples of units that showed enhanced activity when the subject opted to use his right hand. C1-C3 show examples of units that showed enhanced activity when the subject opted to use his left hand. The sharp drop in firing rate after W in C3 constitutes an edge effect artifact. The rightmost column (A4,B4,C4) shows the normalized firing rate for each of the three types of responses illustrated in A-C (the format is similar to the one in Figure 4A). The number of units averaged was 6 (A4), 6 (B4) and 4 (C4). D. The pre-w activity of the neuronal ensemble can extrapolate across hands. Here we show the performance of the classifier in one subject (36 units) that was allowed to freely choose which hand to use to perform the movement (in addition to choosing the time of action). The format is similar to the one in Figure 5B. In the blue curve, we trained the classifier using the ensemble responses from those trials where the 14
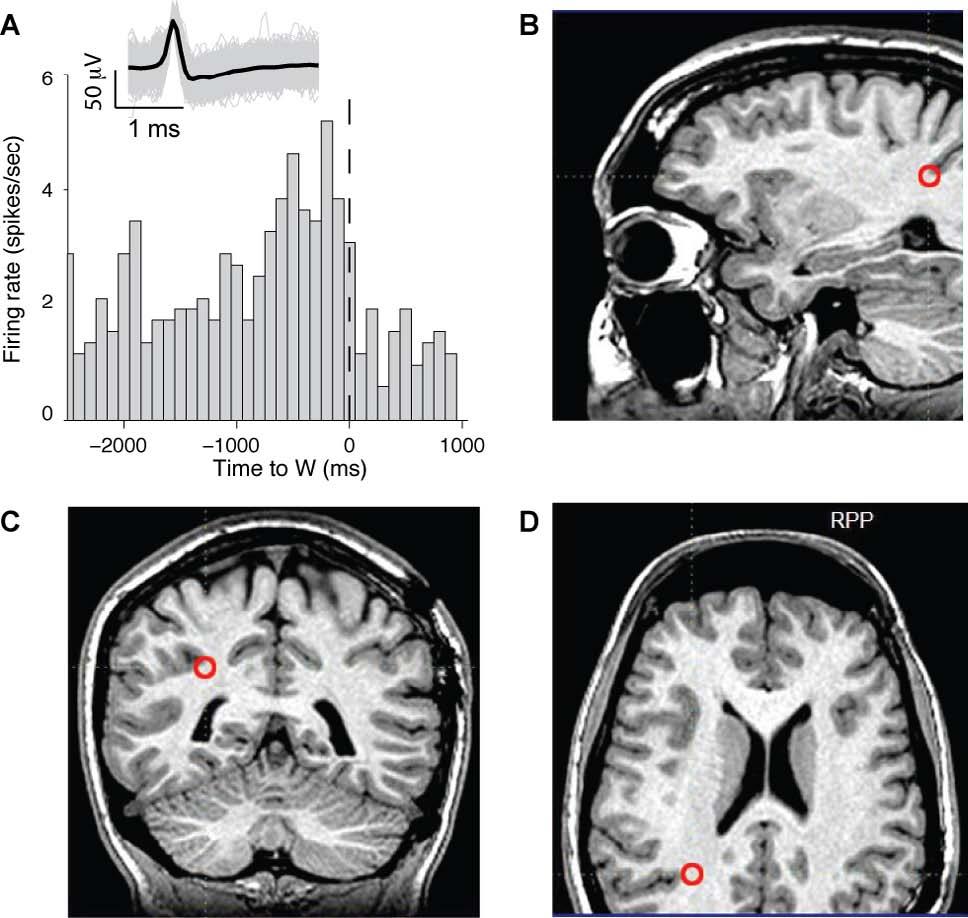
15 subject opted to use his right hand and tested the classification performance using the responses from those trials when the subject opted to use his left hand. In the red curve, we trained the classifier using the left hand trials and tested its performance on the right hand trials. E. The pre-w activity of the neuronal ensemble also allows us to predict which hand the subject will opt to use. Here the classifier was trained to predict the hand choice (blue curve). The black points indicate the performance obtained upon randomly shuffling the right hand / left hand labels. Figure S8: Recordings from parietal cortex. We recorded the activity of units in the right posterior parietal cortex in one additional subject. The unit illustrated in A showed a gradual increase in firing rate as W was approached. There were 3 units that showed this effect in the right posterior parietal cortex (out of a total of 13 recorded units). The MR images in B-D depict the electrode s location (red circle). 15
16 Figure S1
17 FIGURE S2
18 Figure S3
19 Figure S4
20 Figure S5
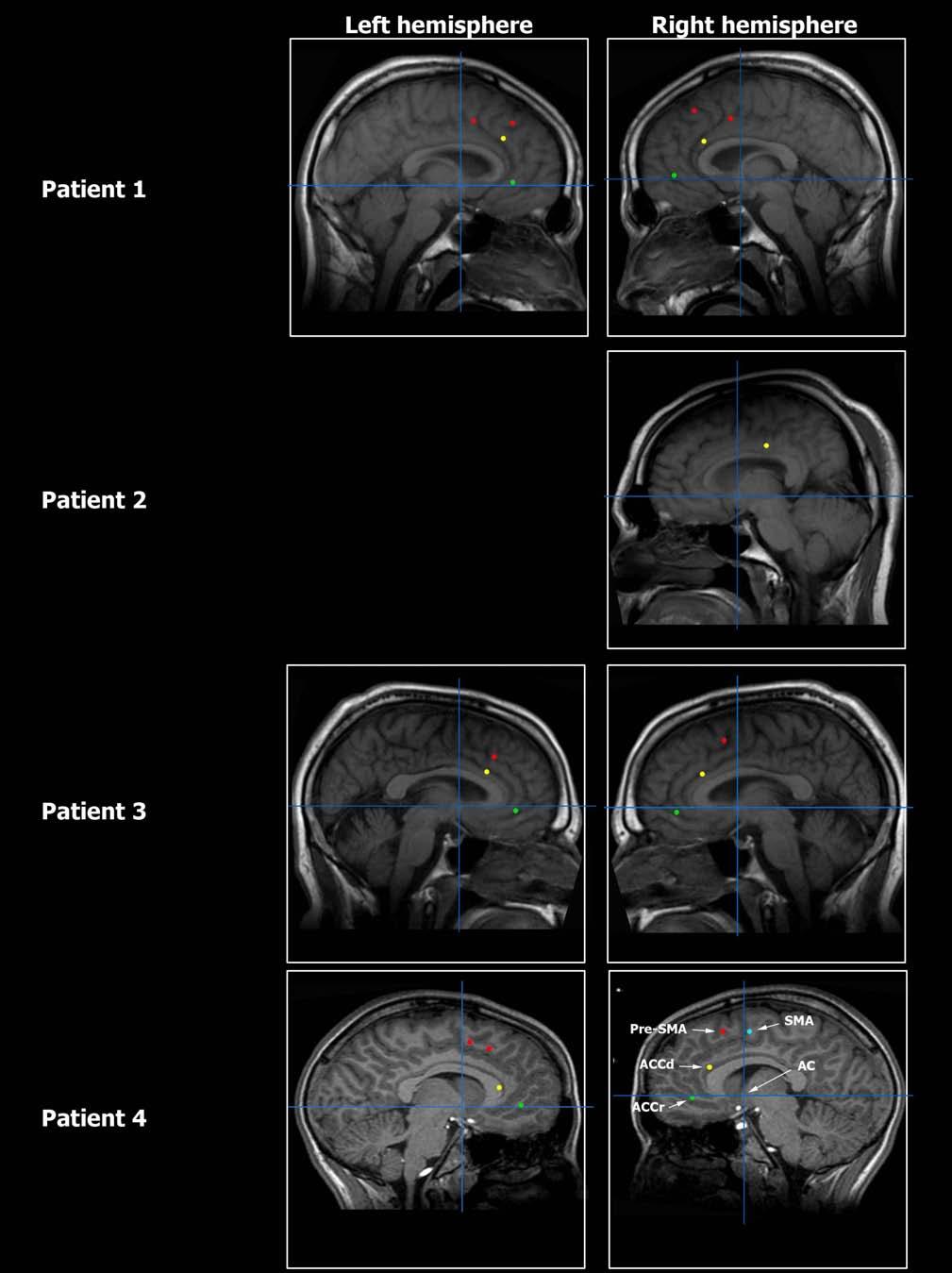
21 Figure S6
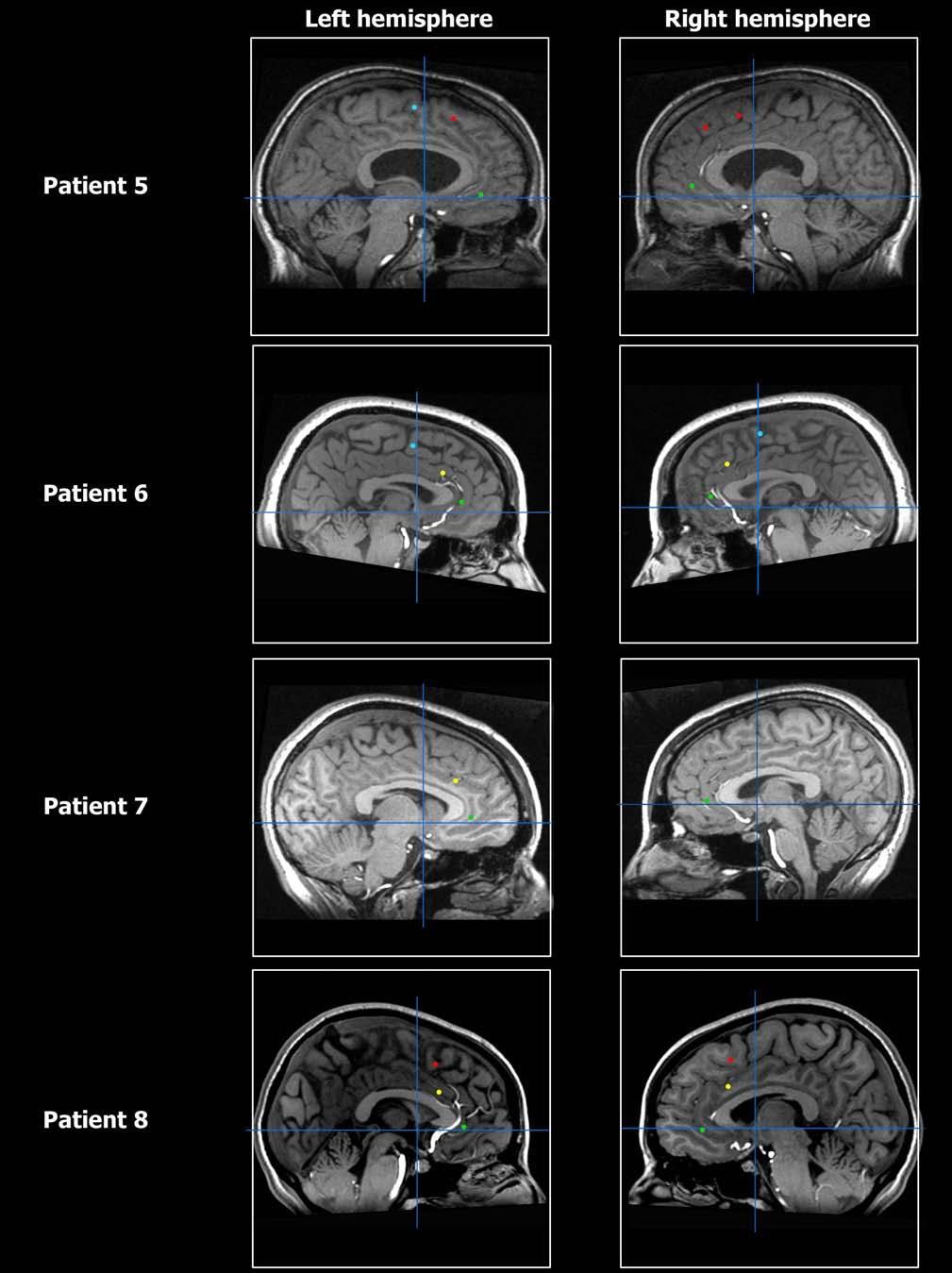
22 Figure S6
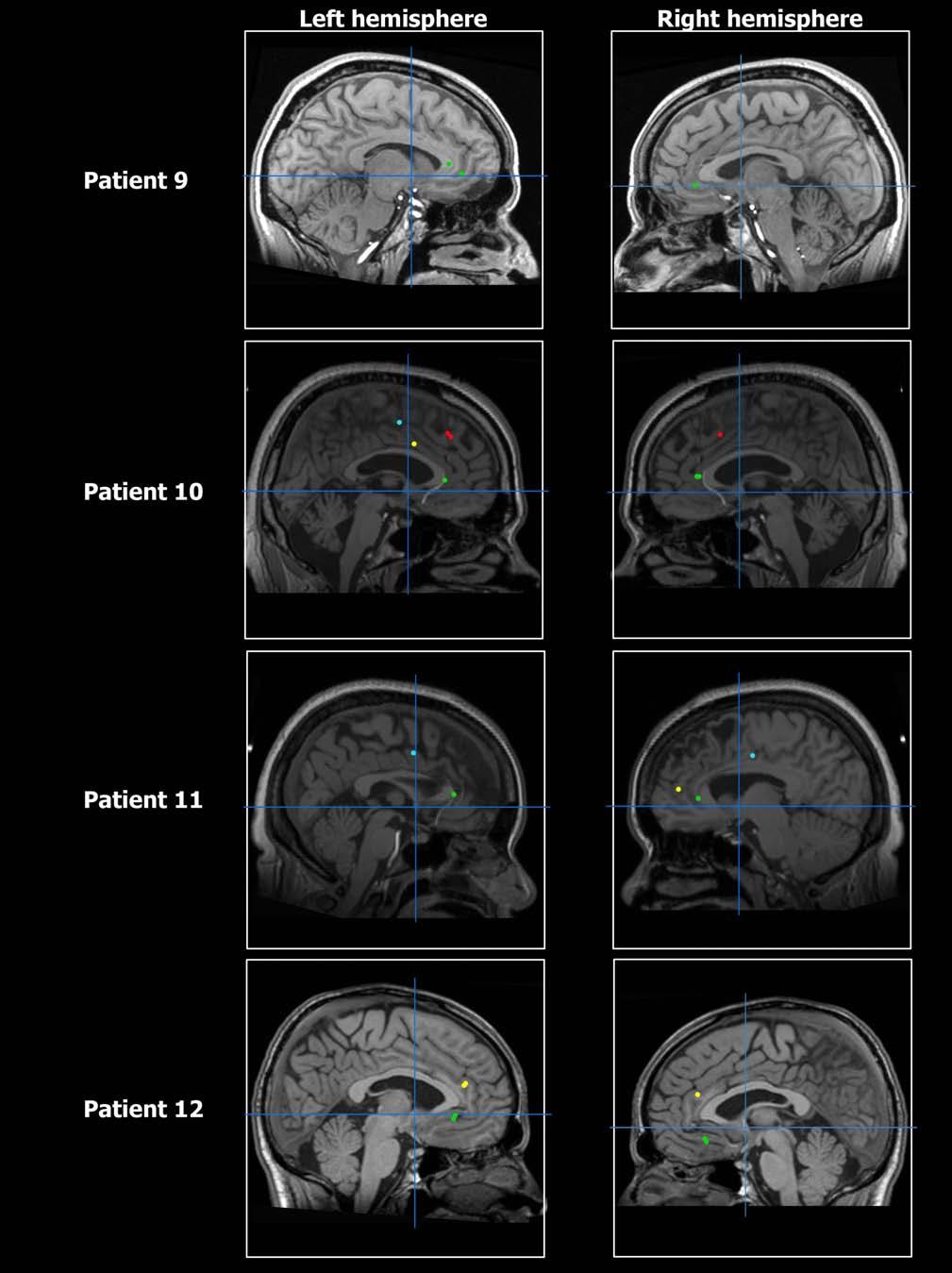
23 Figure S6
24 Figure S7
25 Figure S8
Consider the following spike trains from two different neurons N1 and N2:
 About synchrony and oscillations So far, our discussions have assumed that we are either observing a single neuron at a, or that neurons fire independent of each other. This assumption may be correct in
About synchrony and oscillations So far, our discussions have assumed that we are either observing a single neuron at a, or that neurons fire independent of each other. This assumption may be correct in
Single-Trial Neural Correlates. of Arm Movement Preparation. Neuron, Volume 71. Supplemental Information
 Neuron, Volume 71 Supplemental Information Single-Trial Neural Correlates of Arm Movement Preparation Afsheen Afshar, Gopal Santhanam, Byron M. Yu, Stephen I. Ryu, Maneesh Sahani, and K rishna V. Shenoy
Neuron, Volume 71 Supplemental Information Single-Trial Neural Correlates of Arm Movement Preparation Afsheen Afshar, Gopal Santhanam, Byron M. Yu, Stephen I. Ryu, Maneesh Sahani, and K rishna V. Shenoy
The Spike Response Model: A Framework to Predict Neuronal Spike Trains
 The Spike Response Model: A Framework to Predict Neuronal Spike Trains Renaud Jolivet, Timothy J. Lewis 2, and Wulfram Gerstner Laboratory of Computational Neuroscience, Swiss Federal Institute of Technology
The Spike Response Model: A Framework to Predict Neuronal Spike Trains Renaud Jolivet, Timothy J. Lewis 2, and Wulfram Gerstner Laboratory of Computational Neuroscience, Swiss Federal Institute of Technology
Methods for Estimating the Computational Power and Generalization Capability of Neural Microcircuits
 Methods for Estimating the Computational Power and Generalization Capability of Neural Microcircuits Wolfgang Maass, Robert Legenstein, Nils Bertschinger Institute for Theoretical Computer Science Technische
Methods for Estimating the Computational Power and Generalization Capability of Neural Microcircuits Wolfgang Maass, Robert Legenstein, Nils Bertschinger Institute for Theoretical Computer Science Technische
Disambiguating Different Covariation Types
 NOTE Communicated by George Gerstein Disambiguating Different Covariation Types Carlos D. Brody Computation and Neural Systems Program, California Institute of Technology, Pasadena, CA 925, U.S.A. Covariations
NOTE Communicated by George Gerstein Disambiguating Different Covariation Types Carlos D. Brody Computation and Neural Systems Program, California Institute of Technology, Pasadena, CA 925, U.S.A. Covariations
Lecture on Null Hypothesis Testing & Temporal Correlation
 Lecture on Null Hypothesis Testing & Temporal Correlation CS 590.21 Analysis and Modeling of Brain Networks Department of Computer Science University of Crete Acknowledgement Resources used in the slides
Lecture on Null Hypothesis Testing & Temporal Correlation CS 590.21 Analysis and Modeling of Brain Networks Department of Computer Science University of Crete Acknowledgement Resources used in the slides
The homogeneous Poisson process
 The homogeneous Poisson process during very short time interval Δt there is a fixed probability of an event (spike) occurring independent of what happened previously if r is the rate of the Poisson process,
The homogeneous Poisson process during very short time interval Δt there is a fixed probability of an event (spike) occurring independent of what happened previously if r is the rate of the Poisson process,
+ + ( + ) = Linear recurrent networks. Simpler, much more amenable to analytic treatment E.g. by choosing
 = Linear recurrent networks. Simpler, much more amenable to analytic treatment E.g. by choosing") Linear recurrent networks Simpler, much more amenable to analytic treatment E.g. by choosing + ( + ) = Firing rates can be negative Approximates dynamics around fixed point Approximation often reasonable
Linear recurrent networks Simpler, much more amenable to analytic treatment E.g. by choosing + ( + ) = Firing rates can be negative Approximates dynamics around fixed point Approximation often reasonable
Inventory of Supplemental Information
 Neuron, Volume 71 Supplemental Information Hippocampal Time Cells Bridge the Gap in Memory for Discontiguous Events Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, and Howard Eichenbaum Inventory
Neuron, Volume 71 Supplemental Information Hippocampal Time Cells Bridge the Gap in Memory for Discontiguous Events Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, and Howard Eichenbaum Inventory
Tracking whole-brain connectivity dynamics in the resting-state
 Tracking whole-brain connectivity dynamics in the resting-state Supplementary Table. Peak Coordinates of ICNs ICN regions BA t max Peak (mm) (continued) BA t max Peak (mm) X Y Z X Y Z Subcortical networks
Tracking whole-brain connectivity dynamics in the resting-state Supplementary Table. Peak Coordinates of ICNs ICN regions BA t max Peak (mm) (continued) BA t max Peak (mm) X Y Z X Y Z Subcortical networks
Nature Neuroscience: doi: /nn.2283
 Supplemental Material for NN-A2678-T Phase-to-rate transformations encode touch in cortical neurons of a scanning sensorimotor system by John Curtis and David Kleinfeld Figure S. Overall distribution of
Supplemental Material for NN-A2678-T Phase-to-rate transformations encode touch in cortical neurons of a scanning sensorimotor system by John Curtis and David Kleinfeld Figure S. Overall distribution of
Supplementary Figure 1. Structural MRIs.
 Supplementary Figure 1 Structural MRIs. Coronal and transverse sections of pre-electrode insertion T1 weighted MRIs, illustrating radiologically normal amygdala in the 10 patients for which ierps are presented.
Supplementary Figure 1 Structural MRIs. Coronal and transverse sections of pre-electrode insertion T1 weighted MRIs, illustrating radiologically normal amygdala in the 10 patients for which ierps are presented.
Exercises. Chapter 1. of τ approx that produces the most accurate estimate for this firing pattern.
 1 Exercises Chapter 1 1. Generate spike sequences with a constant firing rate r 0 using a Poisson spike generator. Then, add a refractory period to the model by allowing the firing rate r(t) to depend
1 Exercises Chapter 1 1. Generate spike sequences with a constant firing rate r 0 using a Poisson spike generator. Then, add a refractory period to the model by allowing the firing rate r(t) to depend
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5869/1543/dc1 Supporting Online Material for Synaptic Theory of Working Memory Gianluigi Mongillo, Omri Barak, Misha Tsodyks* *To whom correspondence should be addressed.
www.sciencemag.org/cgi/content/full/319/5869/1543/dc1 Supporting Online Material for Synaptic Theory of Working Memory Gianluigi Mongillo, Omri Barak, Misha Tsodyks* *To whom correspondence should be addressed.
Computing with Inter-spike Interval Codes in Networks of Integrate and Fire Neurons
 Computing with Inter-spike Interval Codes in Networks of Integrate and Fire Neurons Dileep George a,b Friedrich T. Sommer b a Dept. of Electrical Engineering, Stanford University 350 Serra Mall, Stanford,
Computing with Inter-spike Interval Codes in Networks of Integrate and Fire Neurons Dileep George a,b Friedrich T. Sommer b a Dept. of Electrical Engineering, Stanford University 350 Serra Mall, Stanford,
Chapter 1 Statistical Inference
 Chapter 1 Statistical Inference causal inference To infer causality, you need a randomized experiment (or a huge observational study and lots of outside information). inference to populations Generalizations
Chapter 1 Statistical Inference causal inference To infer causality, you need a randomized experiment (or a huge observational study and lots of outside information). inference to populations Generalizations
The Bayesian Brain. Robert Jacobs Department of Brain & Cognitive Sciences University of Rochester. May 11, 2017
 The Bayesian Brain Robert Jacobs Department of Brain & Cognitive Sciences University of Rochester May 11, 2017 Bayesian Brain How do neurons represent the states of the world? How do neurons represent
The Bayesian Brain Robert Jacobs Department of Brain & Cognitive Sciences University of Rochester May 11, 2017 Bayesian Brain How do neurons represent the states of the world? How do neurons represent
AT2 Neuromodeling: Problem set #3 SPIKE TRAINS
 AT2 Neuromodeling: Problem set #3 SPIKE TRAINS Younesse Kaddar PROBLEM 1: Poisson spike trains Link of the ipython notebook for the code Brain neuron emit spikes seemingly randomly: we will aim to model
AT2 Neuromodeling: Problem set #3 SPIKE TRAINS Younesse Kaddar PROBLEM 1: Poisson spike trains Link of the ipython notebook for the code Brain neuron emit spikes seemingly randomly: we will aim to model
Nature Neuroscience: doi: /nn Supplementary Figure 1. Localization of responses
 Supplementary Figure 1 Localization of responses a. For each subject, we classified neural activity using an electrode s response to a localizer task (see Experimental Procedures). Auditory (green), indicates
Supplementary Figure 1 Localization of responses a. For each subject, we classified neural activity using an electrode s response to a localizer task (see Experimental Procedures). Auditory (green), indicates
Recipes for the Linear Analysis of EEG and applications
 Recipes for the Linear Analysis of EEG and applications Paul Sajda Department of Biomedical Engineering Columbia University Can we read the brain non-invasively and in real-time? decoder 1001110 if YES
Recipes for the Linear Analysis of EEG and applications Paul Sajda Department of Biomedical Engineering Columbia University Can we read the brain non-invasively and in real-time? decoder 1001110 if YES
An Introductory Course in Computational Neuroscience
 An Introductory Course in Computational Neuroscience Contents Series Foreword Acknowledgments Preface 1 Preliminary Material 1.1. Introduction 1.1.1 The Cell, the Circuit, and the Brain 1.1.2 Physics of
An Introductory Course in Computational Neuroscience Contents Series Foreword Acknowledgments Preface 1 Preliminary Material 1.1. Introduction 1.1.1 The Cell, the Circuit, and the Brain 1.1.2 Physics of
Temporal whitening by power-law adaptation in neocortical neurons
 Temporal whitening by power-law adaptation in neocortical neurons Christian Pozzorini, Richard Naud, Skander Mensi and Wulfram Gerstner School of Computer and Communication Sciences and School of Life
Temporal whitening by power-law adaptation in neocortical neurons Christian Pozzorini, Richard Naud, Skander Mensi and Wulfram Gerstner School of Computer and Communication Sciences and School of Life
Figure 1: Equivalent electric (RC) circuit of a neurons membrane
 circuit of a neurons membrane") Exercise: Leaky integrate and fire odel of neural spike generation This exercise investigates a siplified odel of how neurons spike in response to current inputs, one of the ost fundaental properties of
Exercise: Leaky integrate and fire odel of neural spike generation This exercise investigates a siplified odel of how neurons spike in response to current inputs, one of the ost fundaental properties of
Bayesian Modeling and Classification of Neural Signals
 Bayesian Modeling and Classification of Neural Signals Michael S. Lewicki Computation and Neural Systems Program California Institute of Technology 216-76 Pasadena, CA 91125 lewickiocns.caltech.edu Abstract
Bayesian Modeling and Classification of Neural Signals Michael S. Lewicki Computation and Neural Systems Program California Institute of Technology 216-76 Pasadena, CA 91125 lewickiocns.caltech.edu Abstract
Supplemental Information. Noise and Correlations. in Parallel Perceptual Decision Making. Thomas U. Otto and Pascal Mamassian
 Current Biology, Volume 22 Supplemental Information Noise and Correlations in Parallel Perceptual Decision Making Thomas U. Otto and Pascal Mamassian Supplemental Inventory Figure S1 (related to Figure
Current Biology, Volume 22 Supplemental Information Noise and Correlations in Parallel Perceptual Decision Making Thomas U. Otto and Pascal Mamassian Supplemental Inventory Figure S1 (related to Figure
Machine Learning Linear Classification. Prof. Matteo Matteucci
 Machine Learning Linear Classification Prof. Matteo Matteucci Recall from the first lecture 2 X R p Regression Y R Continuous Output X R p Y {Ω 0, Ω 1,, Ω K } Classification Discrete Output X R p Y (X)
Machine Learning Linear Classification Prof. Matteo Matteucci Recall from the first lecture 2 X R p Regression Y R Continuous Output X R p Y {Ω 0, Ω 1,, Ω K } Classification Discrete Output X R p Y (X)
Course Review. Kin 304W Week 14: April 9, 2013
 Course Review Kin 304W Week 14: April 9, 2013 1 Today s Outline Format of Kin 304W Final Exam Course Review Hand back marked Project Part II 2 Kin 304W Final Exam Saturday, Thursday, April 18, 3:30-6:30
Course Review Kin 304W Week 14: April 9, 2013 1 Today s Outline Format of Kin 304W Final Exam Course Review Hand back marked Project Part II 2 Kin 304W Final Exam Saturday, Thursday, April 18, 3:30-6:30
Liquid Computing in a Simplified Model of Cortical Layer IV: Learning to Balance a Ball
 Liquid Computing in a Simplified Model of Cortical Layer IV: Learning to Balance a Ball Dimitri Probst 1,3, Wolfgang Maass 2, Henry Markram 1, and Marc-Oliver Gewaltig 1 1 Blue Brain Project, École Polytechnique
Liquid Computing in a Simplified Model of Cortical Layer IV: Learning to Balance a Ball Dimitri Probst 1,3, Wolfgang Maass 2, Henry Markram 1, and Marc-Oliver Gewaltig 1 1 Blue Brain Project, École Polytechnique
THE TRANSFER AND PROPAGATION OF CORRELATED NEURONAL ACTIVITY
 THE TRANSFER AND PROPAGATION OF CORRELATED NEURONAL ACTIVITY A Dissertation Presented to the Faculty of the Department of Mathematics University of Houston In Partial Fulfillment of the Requirements for
THE TRANSFER AND PROPAGATION OF CORRELATED NEURONAL ACTIVITY A Dissertation Presented to the Faculty of the Department of Mathematics University of Houston In Partial Fulfillment of the Requirements for
Emergence of resonances in neural systems: the interplay between adaptive threshold and short-term synaptic plasticity
 Emergence of resonances in neural systems: the interplay between adaptive threshold and short-term synaptic plasticity Jorge F. Mejias 1,2 and Joaquín J. Torres 2 1 Department of Physics and Center for
Emergence of resonances in neural systems: the interplay between adaptive threshold and short-term synaptic plasticity Jorge F. Mejias 1,2 and Joaquín J. Torres 2 1 Department of Physics and Center for
Supplementary Figure 1. Characterization of the single-photon quantum light source based on spontaneous parametric down-conversion (SPDC).
.") .2 Classical light source.8 g (2) ().6.4.2 EMCCD SPAD 2 3.2.4.6.8..2.4.6.8.2 Mean number of photon pairs per pump pulse 4 5 6 7 8 9 2 3 4 Supplementary Figure. Characterization of the single-photon quantum
.2 Classical light source.8 g (2) ().6.4.2 EMCCD SPAD 2 3.2.4.6.8..2.4.6.8.2 Mean number of photon pairs per pump pulse 4 5 6 7 8 9 2 3 4 Supplementary Figure. Characterization of the single-photon quantum
New Machine Learning Methods for Neuroimaging
 New Machine Learning Methods for Neuroimaging Gatsby Computational Neuroscience Unit University College London, UK Dept of Computer Science University of Helsinki, Finland Outline Resting-state networks
New Machine Learning Methods for Neuroimaging Gatsby Computational Neuroscience Unit University College London, UK Dept of Computer Science University of Helsinki, Finland Outline Resting-state networks
A Multivariate Time-Frequency Based Phase Synchrony Measure for Quantifying Functional Connectivity in the Brain
 A Multivariate Time-Frequency Based Phase Synchrony Measure for Quantifying Functional Connectivity in the Brain Dr. Ali Yener Mutlu Department of Electrical and Electronics Engineering, Izmir Katip Celebi
A Multivariate Time-Frequency Based Phase Synchrony Measure for Quantifying Functional Connectivity in the Brain Dr. Ali Yener Mutlu Department of Electrical and Electronics Engineering, Izmir Katip Celebi
#A offered. #B offered. Firing rate. Value of X
 15 12 #A offered 9 6 3 2 1 0 0 1 2 3 4 5 6 8 10 15 20 25 30 35 40 45 #B offered Figure S1. The x-axis and y-axis represent, respectively, the quantities of juices B and A offered in any given trial, and
15 12 #A offered 9 6 3 2 1 0 0 1 2 3 4 5 6 8 10 15 20 25 30 35 40 45 #B offered Figure S1. The x-axis and y-axis represent, respectively, the quantities of juices B and A offered in any given trial, and
Introduction. Chapter 1
 Chapter 1 Introduction In this book we will be concerned with supervised learning, which is the problem of learning input-output mappings from empirical data (the training dataset). Depending on the characteristics
Chapter 1 Introduction In this book we will be concerned with supervised learning, which is the problem of learning input-output mappings from empirical data (the training dataset). Depending on the characteristics
Evaluation. Andrea Passerini Machine Learning. Evaluation
 Andrea Passerini passerini@disi.unitn.it Machine Learning Basic concepts requires to define performance measures to be optimized Performance of learning algorithms cannot be evaluated on entire domain
Andrea Passerini passerini@disi.unitn.it Machine Learning Basic concepts requires to define performance measures to be optimized Performance of learning algorithms cannot be evaluated on entire domain
Continuous detection of weak sensory signals in afferent spike trains: the role of anti-correlated interspike intervals in detection performance
 J Comp Physiol A (2003) 189: 741 759 DOI 10.1007/s00359-003-0449-4 ORIGINAL PAPER J. B. M. Goense Æ R. Ratnam Continuous detection of weak sensory signals in afferent spike trains: the role of anti-correlated
J Comp Physiol A (2003) 189: 741 759 DOI 10.1007/s00359-003-0449-4 ORIGINAL PAPER J. B. M. Goense Æ R. Ratnam Continuous detection of weak sensory signals in afferent spike trains: the role of anti-correlated
Temporal context calibrates interval timing
 Temporal context calibrates interval timing, Mehrdad Jazayeri & Michael N. Shadlen Helen Hay Whitney Foundation HHMI, NPRC, Department of Physiology and Biophysics, University of Washington, Seattle, Washington
Temporal context calibrates interval timing, Mehrdad Jazayeri & Michael N. Shadlen Helen Hay Whitney Foundation HHMI, NPRC, Department of Physiology and Biophysics, University of Washington, Seattle, Washington
Evaluation requires to define performance measures to be optimized
 Evaluation Basic concepts Evaluation requires to define performance measures to be optimized Performance of learning algorithms cannot be evaluated on entire domain (generalization error) approximation
Evaluation Basic concepts Evaluation requires to define performance measures to be optimized Performance of learning algorithms cannot be evaluated on entire domain (generalization error) approximation
Glossary. The ISI glossary of statistical terms provides definitions in a number of different languages:
 Glossary The ISI glossary of statistical terms provides definitions in a number of different languages: http://isi.cbs.nl/glossary/index.htm Adjusted r 2 Adjusted R squared measures the proportion of the
Glossary The ISI glossary of statistical terms provides definitions in a number of different languages: http://isi.cbs.nl/glossary/index.htm Adjusted r 2 Adjusted R squared measures the proportion of the
Evolution of the Average Synaptic Update Rule
 Supporting Text Evolution of the Average Synaptic Update Rule In this appendix we evaluate the derivative of Eq. 9 in the main text, i.e., we need to calculate log P (yk Y k, X k ) γ log P (yk Y k ). ()
Supporting Text Evolution of the Average Synaptic Update Rule In this appendix we evaluate the derivative of Eq. 9 in the main text, i.e., we need to calculate log P (yk Y k, X k ) γ log P (yk Y k ). ()
Neural Encoding: Firing Rates and Spike Statistics
 Neural Encoding: Firing Rates and Spike Statistics Dayan and Abbott (21) Chapter 1 Instructor: Yoonsuck Choe; CPSC 644 Cortical Networks Background: Dirac δ Function Dirac δ function has the following
Neural Encoding: Firing Rates and Spike Statistics Dayan and Abbott (21) Chapter 1 Instructor: Yoonsuck Choe; CPSC 644 Cortical Networks Background: Dirac δ Function Dirac δ function has the following
3 Neural Decoding. 3.1 Encoding and Decoding. (r 1, r 2,..., r N ) for N neurons is a list of spike-count firing rates, although,
 for N neurons is a list of spike-count firing rates, although,") 3 Neural Decoding 3.1 Encoding and Decoding In chapters 1 and 2, we considered the problem of predicting neural responses to known stimuli. The nervous system faces the reverse problem, determining what
3 Neural Decoding 3.1 Encoding and Decoding In chapters 1 and 2, we considered the problem of predicting neural responses to known stimuli. The nervous system faces the reverse problem, determining what
A Model for Real-Time Computation in Generic Neural Microcircuits
 A Model for Real-Time Computation in Generic Neural Microcircuits Wolfgang Maass, Thomas Natschläger Institute for Theoretical Computer Science Technische Universitaet Graz A-81 Graz, Austria maass, tnatschl
A Model for Real-Time Computation in Generic Neural Microcircuits Wolfgang Maass, Thomas Natschläger Institute for Theoretical Computer Science Technische Universitaet Graz A-81 Graz, Austria maass, tnatschl
Mathematical Tools for Neuroscience (NEU 314) Princeton University, Spring 2016 Jonathan Pillow. Homework 8: Logistic Regression & Information Theory
 Princeton University, Spring 2016 Jonathan Pillow. Homework 8: Logistic Regression & Information Theory") Mathematical Tools for Neuroscience (NEU 34) Princeton University, Spring 206 Jonathan Pillow Homework 8: Logistic Regression & Information Theory Due: Tuesday, April 26, 9:59am Optimization Toolbox One
Mathematical Tools for Neuroscience (NEU 34) Princeton University, Spring 206 Jonathan Pillow Homework 8: Logistic Regression & Information Theory Due: Tuesday, April 26, 9:59am Optimization Toolbox One
Semi-Supervised Classification for Intracortical Brain-Computer Interfaces
 Semi-Supervised Classification for Intracortical Brain-Computer Interfaces Department of Machine Learning Carnegie Mellon University Pittsburgh, PA 15213, USA wbishop@cs.cmu.edu Abstract Intracortical
Semi-Supervised Classification for Intracortical Brain-Computer Interfaces Department of Machine Learning Carnegie Mellon University Pittsburgh, PA 15213, USA wbishop@cs.cmu.edu Abstract Intracortical
Non-Bayesian Classifiers Part II: Linear Discriminants and Support Vector Machines
 Non-Bayesian Classifiers Part II: Linear Discriminants and Support Vector Machines Selim Aksoy Department of Computer Engineering Bilkent University saksoy@cs.bilkent.edu.tr CS 551, Fall 2018 CS 551, Fall
Non-Bayesian Classifiers Part II: Linear Discriminants and Support Vector Machines Selim Aksoy Department of Computer Engineering Bilkent University saksoy@cs.bilkent.edu.tr CS 551, Fall 2018 CS 551, Fall
Reducing Multiclass to Binary: A Unifying Approach for Margin Classifiers
 Reducing Multiclass to Binary: A Unifying Approach for Margin Classifiers Erin Allwein, Robert Schapire and Yoram Singer Journal of Machine Learning Research, 1:113-141, 000 CSE 54: Seminar on Learning
Reducing Multiclass to Binary: A Unifying Approach for Margin Classifiers Erin Allwein, Robert Schapire and Yoram Singer Journal of Machine Learning Research, 1:113-141, 000 CSE 54: Seminar on Learning
CORRELATION TRANSFER FROM BASAL GANGLIA TO THALAMUS IN PARKINSON S DISEASE. by Pamela Reitsma. B.S., University of Maine, 2007
 CORRELATION TRANSFER FROM BASAL GANGLIA TO THALAMUS IN PARKINSON S DISEASE by Pamela Reitsma B.S., University of Maine, 27 Submitted to the Graduate Faculty of the Department of Mathematics in partial
CORRELATION TRANSFER FROM BASAL GANGLIA TO THALAMUS IN PARKINSON S DISEASE by Pamela Reitsma B.S., University of Maine, 27 Submitted to the Graduate Faculty of the Department of Mathematics in partial
Logistic Regression: Regression with a Binary Dependent Variable
 Logistic Regression: Regression with a Binary Dependent Variable LEARNING OBJECTIVES Upon completing this chapter, you should be able to do the following: State the circumstances under which logistic regression
Logistic Regression: Regression with a Binary Dependent Variable LEARNING OBJECTIVES Upon completing this chapter, you should be able to do the following: State the circumstances under which logistic regression
3.3 Population Decoding
 3.3 Population Decoding 97 We have thus far considered discriminating between two quite distinct stimulus values, plus and minus. Often we are interested in discriminating between two stimulus values s
3.3 Population Decoding 97 We have thus far considered discriminating between two quite distinct stimulus values, plus and minus. Often we are interested in discriminating between two stimulus values s
Collective Dynamics in Human and Monkey Sensorimotor Cortex: Predicting Single Neuron Spikes
 Collective Dynamics in Human and Monkey Sensorimotor Cortex: Predicting Single Neuron Spikes Supplementary Information Wilson Truccolo 1,2,5, Leigh R. Hochberg 2-6 and John P. Donoghue 4,1,2 1 Department
Collective Dynamics in Human and Monkey Sensorimotor Cortex: Predicting Single Neuron Spikes Supplementary Information Wilson Truccolo 1,2,5, Leigh R. Hochberg 2-6 and John P. Donoghue 4,1,2 1 Department
Linking non-binned spike train kernels to several existing spike train metrics
 Linking non-binned spike train kernels to several existing spike train metrics Benjamin Schrauwen Jan Van Campenhout ELIS, Ghent University, Belgium Benjamin.Schrauwen@UGent.be Abstract. This work presents
Linking non-binned spike train kernels to several existing spike train metrics Benjamin Schrauwen Jan Van Campenhout ELIS, Ghent University, Belgium Benjamin.Schrauwen@UGent.be Abstract. This work presents
Decision-making and Weber s law: a neurophysiological model
 European Journal of Neuroscience, Vol. 24, pp. 901 916, 2006 doi:10.1111/j.14-9568.2006.04940.x Decision-making and Weber s law: a neurophysiological model Gustavo Deco 1 and Edmund T. Rolls 2 1 Institucio
European Journal of Neuroscience, Vol. 24, pp. 901 916, 2006 doi:10.1111/j.14-9568.2006.04940.x Decision-making and Weber s law: a neurophysiological model Gustavo Deco 1 and Edmund T. Rolls 2 1 Institucio
Class 4: Classification. Quaid Morris February 11 th, 2011 ML4Bio
 Class 4: Classification Quaid Morris February 11 th, 211 ML4Bio Overview Basic concepts in classification: overfitting, cross-validation, evaluation. Linear Discriminant Analysis and Quadratic Discriminant
Class 4: Classification Quaid Morris February 11 th, 211 ML4Bio Overview Basic concepts in classification: overfitting, cross-validation, evaluation. Linear Discriminant Analysis and Quadratic Discriminant
Nature Methods: doi: /nmeth Supplementary Figure 1. In vitro screening of recombinant R-CaMP2 variants.
 Supplementary Figure 1 In vitro screening of recombinant R-CaMP2 variants. Baseline fluorescence compared to R-CaMP1.07 at nominally zero calcium plotted versus dynamic range ( F/F) for 150 recombinant
Supplementary Figure 1 In vitro screening of recombinant R-CaMP2 variants. Baseline fluorescence compared to R-CaMP1.07 at nominally zero calcium plotted versus dynamic range ( F/F) for 150 recombinant
Part of the slides are adapted from Ziko Kolter
 Part of the slides are adapted from Ziko Kolter OUTLINE 1 Supervised learning: classification........................................................ 2 2 Non-linear regression/classification, overfitting,
Part of the slides are adapted from Ziko Kolter OUTLINE 1 Supervised learning: classification........................................................ 2 2 Non-linear regression/classification, overfitting,
Dynamical Constraints on Computing with Spike Timing in the Cortex
 Appears in Advances in Neural Information Processing Systems, 15 (NIPS 00) Dynamical Constraints on Computing with Spike Timing in the Cortex Arunava Banerjee and Alexandre Pouget Department of Brain and
Appears in Advances in Neural Information Processing Systems, 15 (NIPS 00) Dynamical Constraints on Computing with Spike Timing in the Cortex Arunava Banerjee and Alexandre Pouget Department of Brain and
Supplementary Information. Brain networks involved in tactile speed classification of moving dot patterns: the. effects of speed and dot periodicity
 Supplementary Information Brain networks involved in tactile speed classification of moving dot patterns: the effects of speed and dot periodicity Jiajia Yang, Ryo Kitada *, Takanori Kochiyama, Yinghua
Supplementary Information Brain networks involved in tactile speed classification of moving dot patterns: the effects of speed and dot periodicity Jiajia Yang, Ryo Kitada *, Takanori Kochiyama, Yinghua
Figure 1-figure supplement 1
 Figure 1-figure supplement 1 a 1 Stroop Task b 1 Reading Task 8 8 Percent Correct 6 4 2 Percent Correct 6 4 2 c 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 Subject Number d 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 Subject
Figure 1-figure supplement 1 a 1 Stroop Task b 1 Reading Task 8 8 Percent Correct 6 4 2 Percent Correct 6 4 2 c 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 Subject Number d 1 2 3 4 5 6 7 8 9 1 11 12 13 14 15 Subject
Phase Response. 1 of of 11. Synaptic input advances (excitatory) or delays (inhibitory) spiking
 or delays (inhibitory) spiking") Printed from the Mathematica Help Browser 1 1 of 11 Phase Response Inward current-pulses decrease a cortical neuron's period (Cat, Layer V). [Fetz93] Synaptic input advances (excitatory) or delays (inhibitory)
Printed from the Mathematica Help Browser 1 1 of 11 Phase Response Inward current-pulses decrease a cortical neuron's period (Cat, Layer V). [Fetz93] Synaptic input advances (excitatory) or delays (inhibitory)
Machine learning comes from Bayesian decision theory in statistics. There we want to minimize the expected value of the loss function.
 Bayesian learning: Machine learning comes from Bayesian decision theory in statistics. There we want to minimize the expected value of the loss function. Let y be the true label and y be the predicted
Bayesian learning: Machine learning comes from Bayesian decision theory in statistics. There we want to minimize the expected value of the loss function. Let y be the true label and y be the predicted
Marr's Theory of the Hippocampus: Part I
 Marr's Theory of the Hippocampus: Part I Computational Models of Neural Systems Lecture 3.3 David S. Touretzky October, 2015 David Marr: 1945-1980 10/05/15 Computational Models of Neural Systems 2 Marr
Marr's Theory of the Hippocampus: Part I Computational Models of Neural Systems Lecture 3.3 David S. Touretzky October, 2015 David Marr: 1945-1980 10/05/15 Computational Models of Neural Systems 2 Marr
Distribution Fitting (Censored Data)
") Distribution Fitting (Censored Data) Summary... 1 Data Input... 2 Analysis Summary... 3 Analysis Options... 4 Goodness-of-Fit Tests... 6 Frequency Histogram... 8 Comparison of Alternative Distributions...
Distribution Fitting (Censored Data) Summary... 1 Data Input... 2 Analysis Summary... 3 Analysis Options... 4 Goodness-of-Fit Tests... 6 Frequency Histogram... 8 Comparison of Alternative Distributions...
The definitions and notation are those introduced in the lectures slides. R Ex D [h
 Mehryar Mohri Foundations of Machine Learning Courant Institute of Mathematical Sciences Homework assignment 2 October 04, 2016 Due: October 18, 2016 A. Rademacher complexity The definitions and notation
Mehryar Mohri Foundations of Machine Learning Courant Institute of Mathematical Sciences Homework assignment 2 October 04, 2016 Due: October 18, 2016 A. Rademacher complexity The definitions and notation
Supervised learning in single-stage feedforward networks
 Supervised learning in single-stage feedforward networks Bruno A Olshausen September, 204 Abstract This handout describes supervised learning in single-stage feedforward networks composed of McCulloch-Pitts
Supervised learning in single-stage feedforward networks Bruno A Olshausen September, 204 Abstract This handout describes supervised learning in single-stage feedforward networks composed of McCulloch-Pitts
What can spike train distances tell us about the neural code?
 What can spike train distances tell us about the neural code? Daniel Chicharro a,b,1,, Thomas Kreuz b,c, Ralph G. Andrzejak a a Dept. of Information and Communication Technologies, Universitat Pompeu Fabra,
What can spike train distances tell us about the neural code? Daniel Chicharro a,b,1,, Thomas Kreuz b,c, Ralph G. Andrzejak a a Dept. of Information and Communication Technologies, Universitat Pompeu Fabra,
!) + log(t) # n i. The last two terms on the right hand side (RHS) are clearly independent of θ and can be
 + log(t) # n i. The last two terms on the right hand side (RHS) are clearly independent of θ and can be") Supplementary Materials General case: computing log likelihood We first describe the general case of computing the log likelihood of a sensory parameter θ that is encoded by the activity of neurons. Each
Supplementary Materials General case: computing log likelihood We first describe the general case of computing the log likelihood of a sensory parameter θ that is encoded by the activity of neurons. Each
A Learning Theory for Reward-Modulated Spike-Timing-Dependent Plasticity with Application to Biofeedback
 A Learning Theory for Reward-Modulated Spike-Timing-Dependent Plasticity with Application to Biofeedback Robert Legenstein, Dejan Pecevski, Wolfgang Maass Institute for Theoretical Computer Science Graz
A Learning Theory for Reward-Modulated Spike-Timing-Dependent Plasticity with Application to Biofeedback Robert Legenstein, Dejan Pecevski, Wolfgang Maass Institute for Theoretical Computer Science Graz
Last updated: Oct 22, 2012 LINEAR CLASSIFIERS. J. Elder CSE 4404/5327 Introduction to Machine Learning and Pattern Recognition
 Last updated: Oct 22, 2012 LINEAR CLASSIFIERS Problems 2 Please do Problem 8.3 in the textbook. We will discuss this in class. Classification: Problem Statement 3 In regression, we are modeling the relationship
Last updated: Oct 22, 2012 LINEAR CLASSIFIERS Problems 2 Please do Problem 8.3 in the textbook. We will discuss this in class. Classification: Problem Statement 3 In regression, we are modeling the relationship
arxiv: v4 [stat.me] 27 Nov 2017
![arxiv: v4 [stat.me] 27 Nov 2017](/thumbs/94/119447143.jpg "arxiv: v4 [stat.me] 27 Nov 2017") CLASSIFICATION OF LOCAL FIELD POTENTIALS USING GAUSSIAN SEQUENCE MODEL Taposh Banerjee John Choi Bijan Pesaran Demba Ba and Vahid Tarokh School of Engineering and Applied Sciences, Harvard University Center
CLASSIFICATION OF LOCAL FIELD POTENTIALS USING GAUSSIAN SEQUENCE MODEL Taposh Banerjee John Choi Bijan Pesaran Demba Ba and Vahid Tarokh School of Engineering and Applied Sciences, Harvard University Center
85. Geo Processing Mineral Liberation Data
 Research Center, Pori / Pertti Lamberg 14024-ORC-J 1 (23) 85. Geo Processing Mineral Liberation Data 85.1. Introduction The Mineral Liberation Analyzer, MLA, is an automated mineral analysis system that
Research Center, Pori / Pertti Lamberg 14024-ORC-J 1 (23) 85. Geo Processing Mineral Liberation Data 85.1. Introduction The Mineral Liberation Analyzer, MLA, is an automated mineral analysis system that
Probing Real Sensory Worlds of Receivers with Unsupervised Clustering
 with Unsupervised Clustering Michael Pfeiffer 1,2 *, Manfred Hartbauer 3, Alexander B. Lang 3, Wolfgang Maass 1, Heinrich Römer 3 1 Institute for Theoretical Computer Science, TU Graz, Graz, Austria, 2
with Unsupervised Clustering Michael Pfeiffer 1,2 *, Manfred Hartbauer 3, Alexander B. Lang 3, Wolfgang Maass 1, Heinrich Römer 3 1 Institute for Theoretical Computer Science, TU Graz, Graz, Austria, 2
HMM and IOHMM Modeling of EEG Rhythms for Asynchronous BCI Systems
 HMM and IOHMM Modeling of EEG Rhythms for Asynchronous BCI Systems Silvia Chiappa and Samy Bengio {chiappa,bengio}@idiap.ch IDIAP, P.O. Box 592, CH-1920 Martigny, Switzerland Abstract. We compare the use
HMM and IOHMM Modeling of EEG Rhythms for Asynchronous BCI Systems Silvia Chiappa and Samy Bengio {chiappa,bengio}@idiap.ch IDIAP, P.O. Box 592, CH-1920 Martigny, Switzerland Abstract. We compare the use
At the Edge of Chaos: Real-time Computations and Self-Organized Criticality in Recurrent Neural Networks
 At the Edge of Chaos: Real-time Computations and Self-Organized Criticality in Recurrent Neural Networks Thomas Natschläger Software Competence Center Hagenberg A-4232 Hagenberg, Austria Thomas.Natschlaeger@scch.at
At the Edge of Chaos: Real-time Computations and Self-Organized Criticality in Recurrent Neural Networks Thomas Natschläger Software Competence Center Hagenberg A-4232 Hagenberg, Austria Thomas.Natschlaeger@scch.at
Reservoir Computing and Echo State Networks
 An Introduction to: Reservoir Computing and Echo State Networks Claudio Gallicchio gallicch@di.unipi.it Outline Focus: Supervised learning in domain of sequences Recurrent Neural networks for supervised
An Introduction to: Reservoir Computing and Echo State Networks Claudio Gallicchio gallicch@di.unipi.it Outline Focus: Supervised learning in domain of sequences Recurrent Neural networks for supervised
The Perceptron algorithm
 The Perceptron algorithm Tirgul 3 November 2016 Agnostic PAC Learnability A hypothesis class H is agnostic PAC learnable if there exists a function m H : 0,1 2 N and a learning algorithm with the following
The Perceptron algorithm Tirgul 3 November 2016 Agnostic PAC Learnability A hypothesis class H is agnostic PAC learnable if there exists a function m H : 0,1 2 N and a learning algorithm with the following
18.9 SUPPORT VECTOR MACHINES
 744 Chapter 8. Learning from Examples is the fact that each regression problem will be easier to solve, because it involves only the examples with nonzero weight the examples whose kernels overlap the
744 Chapter 8. Learning from Examples is the fact that each regression problem will be easier to solve, because it involves only the examples with nonzero weight the examples whose kernels overlap the
COMS 4771 Introduction to Machine Learning. Nakul Verma
 COMS 4771 Introduction to Machine Learning Nakul Verma Announcements HW1 due next lecture Project details are available decide on the group and topic by Thursday Last time Generative vs. Discriminative
COMS 4771 Introduction to Machine Learning Nakul Verma Announcements HW1 due next lecture Project details are available decide on the group and topic by Thursday Last time Generative vs. Discriminative
Probabilistic modeling. The slides are closely adapted from Subhransu Maji s slides
 Probabilistic modeling The slides are closely adapted from Subhransu Maji s slides Overview So far the models and algorithms you have learned about are relatively disconnected Probabilistic modeling framework
Probabilistic modeling The slides are closely adapted from Subhransu Maji s slides Overview So far the models and algorithms you have learned about are relatively disconnected Probabilistic modeling framework
Data Mining. Linear & nonlinear classifiers. Hamid Beigy. Sharif University of Technology. Fall 1396
 Data Mining Linear & nonlinear classifiers Hamid Beigy Sharif University of Technology Fall 1396 Hamid Beigy (Sharif University of Technology) Data Mining Fall 1396 1 / 31 Table of contents 1 Introduction
Data Mining Linear & nonlinear classifiers Hamid Beigy Sharif University of Technology Fall 1396 Hamid Beigy (Sharif University of Technology) Data Mining Fall 1396 1 / 31 Table of contents 1 Introduction
Supplementary Figure 1
 Supplementary Figure 1 Human cerebral vein characterization a, Superficial venous structures were identified and reconstructed using post-contrast brain MRI images (n=50). Scale bar, 3 cm. b-c, Vein diameters
Supplementary Figure 1 Human cerebral vein characterization a, Superficial venous structures were identified and reconstructed using post-contrast brain MRI images (n=50). Scale bar, 3 cm. b-c, Vein diameters
Structure of Materials Prof. Anandh Subramaniam Department of Material Science and Engineering Indian Institute of Technology, Kanpur
 Structure of Materials Prof. Anandh Subramaniam Department of Material Science and Engineering Indian Institute of Technology, Kanpur Lecture - 5 Geometry of Crystals: Symmetry, Lattices The next question
Structure of Materials Prof. Anandh Subramaniam Department of Material Science and Engineering Indian Institute of Technology, Kanpur Lecture - 5 Geometry of Crystals: Symmetry, Lattices The next question
85. Geo Processing Mineral Liberation Data
 Research Center, Pori / Pertti Lamberg 15023-ORC-J 1 (23) 85. Geo Processing Mineral Liberation Data 85.1. Introduction The Mineral Liberation Analyzer, MLA, is an automated mineral analysis system that
Research Center, Pori / Pertti Lamberg 15023-ORC-J 1 (23) 85. Geo Processing Mineral Liberation Data 85.1. Introduction The Mineral Liberation Analyzer, MLA, is an automated mineral analysis system that
CSE/NB 528 Final Lecture: All Good Things Must. CSE/NB 528: Final Lecture
 CSE/NB 528 Final Lecture: All Good Things Must 1 Course Summary Where have we been? Course Highlights Where do we go from here? Challenges and Open Problems Further Reading 2 What is the neural code? What
CSE/NB 528 Final Lecture: All Good Things Must 1 Course Summary Where have we been? Course Highlights Where do we go from here? Challenges and Open Problems Further Reading 2 What is the neural code? What
Neural Networks and Ensemble Methods for Classification
 Neural Networks and Ensemble Methods for Classification NEURAL NETWORKS 2 Neural Networks A neural network is a set of connected input/output units (neurons) where each connection has a weight associated
Neural Networks and Ensemble Methods for Classification NEURAL NETWORKS 2 Neural Networks A neural network is a set of connected input/output units (neurons) where each connection has a weight associated
80% of all excitatory synapses - at the dendritic spines.
 Dendritic Modelling Dendrites (from Greek dendron, tree ) are the branched projections of a neuron that act to conduct the electrical stimulation received from other cells to and from the cell body, or
Dendritic Modelling Dendrites (from Greek dendron, tree ) are the branched projections of a neuron that act to conduct the electrical stimulation received from other cells to and from the cell body, or
Every animal is represented by a blue circle. Correlation was measured by Spearman s rank correlation coefficient (ρ).
.") Supplementary Figure 1 Correlations between tone and context freezing by animal in each of the four groups in experiment 1. Every animal is represented by a blue circle. Correlation was measured by Spearman
Supplementary Figure 1 Correlations between tone and context freezing by animal in each of the four groups in experiment 1. Every animal is represented by a blue circle. Correlation was measured by Spearman
ECE662: Pattern Recognition and Decision Making Processes: HW TWO
 ECE662: Pattern Recognition and Decision Making Processes: HW TWO Purdue University Department of Electrical and Computer Engineering West Lafayette, INDIANA, USA Abstract. In this report experiments are
ECE662: Pattern Recognition and Decision Making Processes: HW TWO Purdue University Department of Electrical and Computer Engineering West Lafayette, INDIANA, USA Abstract. In this report experiments are
Ângelo Cardoso 27 May, Symbolic and Sub-Symbolic Learning Course Instituto Superior Técnico
 BIOLOGICALLY INSPIRED COMPUTER MODELS FOR VISUAL RECOGNITION Ângelo Cardoso 27 May, 2010 Symbolic and Sub-Symbolic Learning Course Instituto Superior Técnico Index Human Vision Retinal Ganglion Cells Simple
BIOLOGICALLY INSPIRED COMPUTER MODELS FOR VISUAL RECOGNITION Ângelo Cardoso 27 May, 2010 Symbolic and Sub-Symbolic Learning Course Instituto Superior Técnico Index Human Vision Retinal Ganglion Cells Simple
arxiv: v1 [q-bio.nc] 1 Jun 2014
![arxiv: v1 [q-bio.nc] 1 Jun 2014](/thumbs/92/108475602.jpg "arxiv: v1 [q-bio.nc] 1 Jun 2014") 1 arxiv:1406.0139v1 [q-bio.nc] 1 Jun 2014 Distribution of Orientation Selectivity in Recurrent Networks of Spiking Neurons with Different Random Topologies Sadra Sadeh 1, Stefan Rotter 1, 1 Bernstein Center
1 arxiv:1406.0139v1 [q-bio.nc] 1 Jun 2014 Distribution of Orientation Selectivity in Recurrent Networks of Spiking Neurons with Different Random Topologies Sadra Sadeh 1, Stefan Rotter 1, 1 Bernstein Center
Discriminative Direction for Kernel Classifiers
 Discriminative Direction for Kernel Classifiers Polina Golland Artificial Intelligence Lab Massachusetts Institute of Technology Cambridge, MA 02139 polina@ai.mit.edu Abstract In many scientific and engineering
Discriminative Direction for Kernel Classifiers Polina Golland Artificial Intelligence Lab Massachusetts Institute of Technology Cambridge, MA 02139 polina@ai.mit.edu Abstract In many scientific and engineering
Testing Problems with Sub-Learning Sample Complexity
 Testing Problems with Sub-Learning Sample Complexity Michael Kearns AT&T Labs Research 180 Park Avenue Florham Park, NJ, 07932 mkearns@researchattcom Dana Ron Laboratory for Computer Science, MIT 545 Technology
Testing Problems with Sub-Learning Sample Complexity Michael Kearns AT&T Labs Research 180 Park Avenue Florham Park, NJ, 07932 mkearns@researchattcom Dana Ron Laboratory for Computer Science, MIT 545 Technology
Glossary for the Triola Statistics Series
 Glossary for the Triola Statistics Series Absolute deviation The measure of variation equal to the sum of the deviations of each value from the mean, divided by the number of values Acceptance sampling
Glossary for the Triola Statistics Series Absolute deviation The measure of variation equal to the sum of the deviations of each value from the mean, divided by the number of values Acceptance sampling
Performance Evaluation and Comparison
 Outline Hong Chang Institute of Computing Technology, Chinese Academy of Sciences Machine Learning Methods (Fall 2012) Outline Outline I 1 Introduction 2 Cross Validation and Resampling 3 Interval Estimation
Outline Hong Chang Institute of Computing Technology, Chinese Academy of Sciences Machine Learning Methods (Fall 2012) Outline Outline I 1 Introduction 2 Cross Validation and Resampling 3 Interval Estimation
Supplementary material: Orbitofrontal and striatal circuits dynamically encode the shift. between goal-directed and habitual actions
 Supplementary material: Orbitofrontal and striatal circuits dynamically encode the shift between goal-directed and habitual actions Christina M. Gremel 1 & Rui M. Costa 1,2 1 Laboratory for Integrative
Supplementary material: Orbitofrontal and striatal circuits dynamically encode the shift between goal-directed and habitual actions Christina M. Gremel 1 & Rui M. Costa 1,2 1 Laboratory for Integrative
Probability Distributions.
 Probability Distributions http://www.pelagicos.net/classes_biometry_fa18.htm Probability Measuring Discrete Outcomes Plotting probabilities for discrete outcomes: 0.6 0.5 0.4 0.3 0.2 0.1 NOTE: Area within
Probability Distributions http://www.pelagicos.net/classes_biometry_fa18.htm Probability Measuring Discrete Outcomes Plotting probabilities for discrete outcomes: 0.6 0.5 0.4 0.3 0.2 0.1 NOTE: Area within
Neural Decoding. Chapter Encoding and Decoding
 Chapter 3 Neural Decoding 3.1 Encoding and Decoding In chapters 1 and 2, we considered the problem of predicting neural responses to known stimuli. The nervous system faces the reverse problem, determining
Chapter 3 Neural Decoding 3.1 Encoding and Decoding In chapters 1 and 2, we considered the problem of predicting neural responses to known stimuli. The nervous system faces the reverse problem, determining
References for online kernel methods
 References for online kernel methods W. Liu, J. Principe, S. Haykin Kernel Adaptive Filtering: A Comprehensive Introduction. Wiley, 2010. W. Liu, P. Pokharel, J. Principe. The kernel least mean square
References for online kernel methods W. Liu, J. Principe, S. Haykin Kernel Adaptive Filtering: A Comprehensive Introduction. Wiley, 2010. W. Liu, P. Pokharel, J. Principe. The kernel least mean square
Algorithm-Independent Learning Issues
 Algorithm-Independent Learning Issues Selim Aksoy Department of Computer Engineering Bilkent University saksoy@cs.bilkent.edu.tr CS 551, Spring 2007 c 2007, Selim Aksoy Introduction We have seen many learning
Algorithm-Independent Learning Issues Selim Aksoy Department of Computer Engineering Bilkent University saksoy@cs.bilkent.edu.tr CS 551, Spring 2007 c 2007, Selim Aksoy Introduction We have seen many learning