A study on the pattern of prospective somites in the presomitic mesoderm of mouse embryos
|
|
|
- Aileen Tucker
- 5 years ago
- Views:
Transcription

1 /. Embryol. exp. Morph. 9, 9-8 (98) 9 Printed in Great Britain The Company of Biologists Limited 98 A study on the pattern of prospective somites in the presomitic mesoderm of mouse embryos p. P. L. TAM Department of Anatomy, Faculty of Medicine, The Chinese University of Hong Kong, Shatin, NT, Hong Kong SUMMARY Consistently six to seven soniites were formed in explants of presomitic mesoderm of 8--day to --day mouse embryos and this number correlated well with the number of somitomeres previously identified in the same tissue by stereo SEM (Tam, Meier & Jacobson, 98). During this period of development, the size of the presomitic mesoderm varied up to two-fold but the number of prospective somites remained unchanged. This pattern in the presomitic mesoderm was stable with respect to the number and the position of somites that were formed, the craniocaudal sequence and the rate of segmentation. The prospective somite also went through an orderly sequence of expansion and cellular rearrangement prior to segmentation. During the differentiation of the presomitic mesoderm, somites were formed at the expense of somitomeres, suggesting that the somitomere was a morphological manifestation of the prospective somite. INTRODUCTION The earliest expression of a metameric pattern in the embryonic axis is seen in the formation of neuromeres in the neural tube (Neal, 98; Adelman, 9) and the appearance of somites in the paraxial mesoderm (Meier, 98). Somites are formed in a craniocaudal sequence and the last somite in the paraxial mesoderm is contiguous with a longitudinal strip of unsegmented tissue called segmental plate or presomitic mesoderm. Somites are continuously added to the body axis by segmentation of the cranial end of this tissue (Bellairs, 979). The presomitic mesoderm has been regarded as a site of progressive specification of the somitic pattern and one in which a certain degree of regulation of somite size and number is occurring (Flint, Ede, Wilby & Proctor, 978). However, experimental studies in amphibian embryos and chick embryos show that the presomitic mesoderm is distinctively patterned in terms of the number of somites that can be formed and the craniocaudal sequence of segmentation (Deuchar & Burgess, 97; Cooke, 977; Menkes & Sandor, 977; Stern & Bellairs, 98a). This pattern of somitogenic capacity is unperturbed by the removal of epithelial and axial structures that are normally related to the presomitic mesoderm (Packard & Jacobson, 97, 979; Menkes & Sandor, 977; Stern & Bellairs, 98a). Although the segmental plate of avian and snapping turtle embryos varies from two- to three-fold in size Key words: mouse embryo, presomitic mesoderm, somitomere, somite, metameric pattern.
, chick (Packard, 978) and quail (Packard, 98).")
2 7 P. P. L. TAM during development, explants of this tissue always form a relatively constant number of somites (Packard, 978, 98a,b). A pattern of prospective somites is postulated to be present in the segmental plate of snapping turtle (Packard, 98a), chick (Packard, 978) and quail (Packard, 98). Morphological studies using scanning electron microscopy and stereoimaging have recently revealed the presence of a recognizable metameric pattern of cellular arrangement in the segmental plate of the chick (Meier, 979), quail (Packard & Meier, 98) and snapping turtle (Packard & Meier, 98) and in the presomitic mesoderm of the mouse (Tam et al. 98). The pattern presents itself as a tandem series of somitomeres which could be identified by the arrangement of mesenchymal cells in circular swirls about a centre and the boundaries between somitomeres are marked by changes in cellular alignment. In the snapping turtle and avian embryos, there is a good correlation between the number of somitomeres and the number of prospective somites contained in the segmental plate (Packard & Meier, 98, 98). The presomitic mesoderm is therefore fully organized into a metameric pattern consisting of somitomeres which are undergoing further morphogenesis to become somites. Although the presence of somitomeres in the presomitic mesoderm has been documented in the mouse (Tam et al. 98), the existence of prospective somites as suggested by Tam (98) has not been demonstrated. The present study was carried out to investigate the in vitro differentiation of the presomitic mesoderm and to elucidate the developmental relationship between somitomeres and somites in the mouse embryo. MATERIALS AND METHODS Mouse embryos at 8- to - daysp.c. were obtained from ICR strain of mice. Pregnant mice were killed by cervical dislocation and the conceptuses were removed from the uterus to PB medium. The embryos were dissected out from the decidua and the Reichert's membrane and ectoplacental cone were removed. For the study on the somitogenic capacity of the presomitic mesoderm, 8--day and 9--day embryos were subjected to the following operation. Two small incisions were made through the visceral yolk sac and the amnion. Into these openings, a pair of electrolytically polished alloy needles were passed and two transverse cuts were made to isolate a portion of the body containing presomitic mesoderm. The first cut was made at the caudal border of the last-formed somites in the paraxial mesoderm. The second cut was made at a plane at about - ^m from the caudal end of the body axis (Figs, ) so that the primitive streak and tail bud tissues (= caudal tissue) were excised. The operated embryos were then placed in bottles containing a culture medium of equal parts of immediately centrifuged rat serum and Hams F medium (GIBCO). Embryos with incised membranes or intact membranes were also cultured as controls. Four to five embryos were cultured in ml medium in a ml serum bottle (Wheaton) rotated at r.p.m. For 8--day embryos, the culture was gassed with % CO, % O and 9 % N for the first 8 h and then with % CO in air for the next h. For 9--day embryos, a gas phase of % CO, % O and % N was used throughout. At the end of the culture, the embryos were dissected free of membranes and the number of somites present in the body cranial to the first cut, in the segment containing the presomitic mesoderm and in the caudal tissues were counted. The somite numbers of the control embryos and also those of embryos at equivalent ages in vivo were also scored. Selected embryos from the operated and control groups were analysed for protein content (Lowry, Rosebrough, Farr & Randall, 9). Others were fixed in Sanfelice fluid and embedded in paraffin wax (Merck, melting point - C). Serial sections of ym thickness were stained with haematoxylin and eosin.
 embryos showing the position of cuts made through the extraembryonic membranes in order to isolate a segment of the body containing the presomitic mesoderm. Bar, \xm. Figs -.")
3 Differentiation of presomitic mesoderm 7 **v \. -- Figs,. Scanning electron micrographs of 8--day (Fig. ) and 9--day (Fig. ) embryos showing the position of cuts made through the extraembryonic membranes in order to isolate a segment of the body containing the presomitic mesoderm. Bar, \xm. Figs -. Scanning electron micrographs of the caudal region of 8--day (Fig. ), 9-- day (Fig. ) and --day (Fig. ) embryos. Cut a was made usually at the fissure between the last somite and the presomitic mesoderm and sometimes at the intersomitic fissures so that one to two somites were left in the explant. Cut b was made at about - jum from the tip of embryonic axis. The small fragment thus removed was the caudal tissue (ct). For 8--day and 9--day explants, a third cut (c) was made to bisect the explant to yield two symmetrical halves. Bar, jum.

4 7 P. P. L. TAM In another experiment, 8--day to --day embryos were dissected free from the foetal membranes and a portion of the body containing the presomitic mesoderm was isolated by cutting caudal to the last somite and cranial to the caudal tissue (cuts a and b of Figs -). The resultant explants were shown in Figs -8. In another study, which also included --day embryos, cut b was made at various distances away from cut a so that different amounts of presomitic mesoderm were included in the explants. Camera-lucida drawings of original explants containing the entire presomitic mesoderm and caudal tissue, and that of cut pieces were made with a Nikon drawing tube mounted on a Nikon SMZ- dissecting microscope. The various types of explants were cultured under conditions as for the whole embryo. Cultures were examined at -7 h, - h and - h and the number of somites formed was scored. Photographs of the explants before and after culture were taken with a Nikon inverted microscope using phase optics. Some cultured explants were fixed with Sanfelice fluid and processed for wax histology as described previously. Others were fixed with half-strength Karnovsky fixative and % osmium tetroxide, and dehydrated in an ascending alcohol series. Some of the specimens were embedded in epoxy resin, sectioned at -/im and stained with toluidine blue for light-microscopic examination. Some were critical-point dried for scanning electron microscopy using Freon as the exchange fluid. The specimen was coated with gold and examined with a JEOL JSM-CF microscope operating at kv. The longitudinal length of explants was measured on the camera-lucida drawings using a GRAF/BAR sonic digitizer (Science Accessory Corporation) linked to an IBM PC-XT computer that was driven by a Stereometry program (Yucomp Co. Ltd) written in BASIC. The numerical relationship between somitomeres contained in the presomitic mesoderm and somites formed after culture was also studied. The explants of presomitic mesoderm of 8--day and 9--day embryo were prepared by cutting first at positions a and b (Figs, ). Cut a was usually made so that one to three pre-existing somites were included in the explant and this aided later orientation of the craniocaudal axis. A longitudinal cut in the median plane was then made (cut c of Figs, ) to yield two symmetrical portions of the explants. Only pairs of similar length and containing same number of pre-existing somites were used in this study. One portion of the explant was immediately fixed for scanning electron microscopy as described above. The other portion was individually cultured in - ml of medium (equal parts of rat serum and Hams F medium) in a -well Falcon Multiwell culture plate under % CO in air for -7 h. The number of somites formed in the explant was scored and the explant wasfixedand processed for scanning electron microscopy. Stereopair pictures of the presomitic mesoderm were taken at a tilt angle of for stereoscopic examination. Statistical analyses of the data were done using a NWA Statpak program (Multifunction Statistics Library version., Northwest Analytical) run on an IBM PC-XT computer. RESULTS Transecting the body axis did not affect somite formation in the presomitic mesoderm of the operated embryos. About - somites were initially present in the 8--day embryo used for the operation and during the h of culture -8 somites were formed in the presomitic mesoderm portion (Table, Fig. 9). The caudal piece containing primarily the primitive streak grew poorly and in only seven cases an additional - somites were formed. The total number of somites formed in the operated embryo was less than that of the control embryo. The number of somites and the amount of protein contained in the control embryo were comparable to those of 9--day embryos in vivo (Table ). The 9--day embryos used in this experiment have formed somites at the time of operation, and the presomitic mesoderm formed another - somites after h of culture (Table, Fig. ). The caudal piece formed a rudimentary tail but no somites were

5 Differentiation of presomitic mesoderm 7 formed in the scanty paraxial mesoderm. Both the control and operated embryos formed fewer somites and had less protein than --day embryos in vivo. The presomitic mesoderm obtained from embryos at 8- to - days differed in its length. The presomitic mesoderm of 8--day embryo was about 7 jxm and the :\ a*'-'- u Figs -8. The explants from 8--day (Fig. ), 9--day (Fig. 7) and --day (Fig. 8) embryos produced by making cuts a and b. Arrow points cranially. Bar, fxm. Figs 9,. The isolated segments of 8--day (Fig. 9) and 9--day (Fig. ) embryos produced by operations shown in Figs, after h in vitro. The presomitic mesoderm was segmented into somites (s). In Fig. 9, the presomitic mesoderm segment was partly attached to the main trunk (tr) of the embryo. Bar, (im.
6 7 P. P. L. TAM Table. Somite formation in intact and operated embryos in vitro 8--day embryo ( h in vitro) Control Operated 9--day embryo in vivo 9--day embryo ( h in vitro) Control Operated --day embryo in vivo Body Number of somites Presomitic mesoderm Whole body Protein content -9-() -7-(9) --() -8-() 8-- ()t (8)' - + -() --9() --7()* -8- ()t - ±- () - + -(9) 7- ± - ()* 79- ± 7- ()t - - () () Significantly different from in vivo values at * P < -, t P < - and $ P < - by Student's t-test. An additional - ± - somites were formed in the tail segment of seven embryos. size had nearly doubled at 9- days to 97 /im. The size was then reduced to about 8/im at - days (Table ). Despite this variation in the amount of tissues, the presomitic mesoderm formed about ---9 somites after h in vitro (Table ). Only about - % of the 8--day and 9--day explants developed successfully to form recognizable somites (Figs, ). The remaining explants either degenerated or became so stunted that somites could not be definitively identified. Nearly all the --day explants formed distinctive somites (Fig. ). Histological examination of the successfully developed explants showed that proper epithelial somites were formed and some of the somites even began to show sclerotome differentiation (Figs -). The somites were more closely packed than those formed in vivo, and were much taller dorsoventrally but shorter craniocaudally. A comparison of the development of explants without the caudal tissue (PM-PS/TB) and those with intact caudal tissue (PM+PS/TB) was made (Table ). The position of cut b was shifted more cranially than that in previous experiments by about /im to ensure a complete removal of the caudal tissue and the culture was examined at 7-8 h and again at - h. The PM-PS/TB Table. The number of somites formed from explants of presomitic mesoderm of 8--day to --day embryos Age (d) Mean somite no. Size (jum) No. of somites ± -8 (8) ± - () - - ± -7 () 7 ± 8 (8) 97 ± () 8 ± () -9 ± - () -9-() --()

7 Differentiation of presomitic mesoderm 7 explants formed about - to -7 somites after 7-8 h, and further culturing to - h only resulted in a slight increase to about - to -8 somites (Table ). During the same period of time, the PM+PS/TB explants formed significantly more somites (Fig. 7) than those explants without primitive streak or tail bud (Table ), suggesting that the caudal tissue was an important source of somitic cells. The presomitic mesoderm could only segment into a limited number of somites and further somitogenesis ceased when all existing tissues were exhausted. The 8--day and 9--day PM+PS/TB explants formed about one somite in every hour but the --day explants took about -8 h to generate one somite. This rate of somite segmentation was in agreement with a previous estimate based upon in vivo observation (Tarn, 98). Table shows the results of somite formation in explants of presomitic mesoderm that contained different amount of paraxial tissues. Up to seven somites were formed in the largest explant of 8--day and 9--day embryos, and six somites were formed from the entire presomitic mesoderm of --day and --day embryos. Fewer somites were formed in explants containing a smaller cranial portion of the presomitic mesoderm. A statistical analysis of the length of the explant and the number of somites formed after culture revealed a direct correlation between the amount of available presomitic mesoderm and the number of prospective somites (Table ). Based upon the regressional equation, the size of the explant that would generate a specific whole number of somites was calculated. Using these data, the domain occupied by each prospective somite in the presomitic mesoderm could then be mapped (Fig. 8). Six to seven prospective somites were accommodated in the presomitic mesoderm of 8--day to --day embryos. The size of prospective somites varied in accordance with their position in the presomitic mesoderm and to the overall size of this tissue. The caudal prospective somite occupied a smaller domain than the more cranial ones. The variation in the size of the most cranial prospective somite at different embryonic ages followed closely the change in size of newly formed somites observed in mouse embryo in vivo (Tarn, 98). The size of the caudal tissue also changed during development. The primitive streak regressed in both absolute and relative size between 8- and 9- days. The tail bud showed a similar trend of reduction in size between - and - days. When the presomitic mesoderm was examined with SEM and stereoimaging, characteristic somitomeric units were identified (Fig. 9) and about five to six somitomeres were found in explants of 8--day and 9--day embryos (Table ). When the corresponding half of the explant was cultured and examined at various times between and 7 h, somites were seen to form in a proper craniocaudal sequence but at a slower rate than bilaterally intact explants. The newly formed somites were seen to replace the more cranially located somitomeres (Fig. ). The 8--day explants formed one somite every -9 h and the 9--day explants formed one every - h. When the explants were harvested and the number of newly added somites and remaining somitomeres was scored, it was found that the total of these two numbers matched the number of somitomeres initially present


8 - * 7 P. P. L. TAM.. -!,. : as -- - / > \ V ^SH^fjs^i

9 Differentiation ofpresomitic mesoderm 77 (Table ). A comparison of the morphology of the somitomere and the somite derived from it showed that somite formation involved a compaction of the mesenchymal cells and a dorsoventral re-alignment of the cells in the explant (Figs, ). DISCUSSION The present study has provided evidence in support of the existence of a pattern of prospective somites in the presomitic mesoderm of mouse embryos. The foremost of these is the demonstration that six to seven somites were consistently formed from the presomitic mesoderm which varied at least two-fold in size during embryonic development at 8--- days. In the snapping turtle, chick and quail embryos it has been shown that despite a variation in the size of the segmental plate, a relatively constant number of somites would usually be formed in explants of the entire segmental plate. In terms of numbers of somites generated from the presomitic mesoderm, the mouse resembles the snapping turtle where an explant of the entire segmental plate gave five to seven somites (Packard, 98a), but differs from the avian embryos whose segmental plate formed ten to eleven somites (Packard & Jacobson, 97; Packard, 98b). It has also been shown in the present study that reducing the amount of tissue contained in the explant consequently led to fewer somites being formed. There was no evidence of any regulation to restore the full complement of prospective somites, suggesting that the somitogenic pattern in the presomitic mesoderm is a stable one. Both the craniocaudal sequence and the intrinsic rate of segmentation were retained by the cells in the explants and are unaffected by various surgical manipulations. In both the amphibian and avian embryos, experimental manipulation of the presomitic mesoderm such as reversal of the craniocaudal relation, surgical deletion of tissues and disruption of the normal cellular arrangement has little effect on the predetermined direction of somite segmentation and the number of somites formed (Deuchar & Burgess, 97; Cooke, 977; Menkes & Sandor, 977). Somite segmentation was also unperturbed by the transection of the body axis or the segmental plate (Smith, 9; Packard, 978) or by the removal of the axial Figs -. The PM-PS/TB explants of 8--day (Fig. ), 9--day (Fig. ) and -- day (Fig. ) embryos showing the formation of somites in the presomitic mesoderm. Intersomitic boundary is marked by arrow heads. In Fig., the intersomitic fissure marked by > was out of focus. In Fig., the paraxial mesoderm (*) lateral to the bulbous end of the neural tube was a homogeneous tissue mass and was not organized into somites. Arrows point cranially. Bar, jum. Figs -. Histological sections of cultured PM-PS/TB explants of 8--day (Fig. ), 9--day (Fig. ) and --day (Fig. ) embryos. The somites formed in the explants were morphologically comparable to those formed in vivo. Sclerotome dispersion has occurred and individual somites could be discerned by the dermamyotome and the fissures between sclerotomes. Bar, ^m. Fig. 7. A longitudinal section of the PM+PS/TB explant of 9--day embryo showing the presence of eight complete somites, a ninth one which was segmenting and a short unsegmented presomitic mesoderm abutting the tail bud. Bar, //m.
10 Table. The formation of somites in explants ofpresomitic mesoderm containing the caudal tissue (PM+PS/ TB) and without the caudal tissue (PM-PS/TB) No. of somites formed by PM+PS/TB PM-PS/TB 8--day explants 7-8h -h 8- ±- () - ±- () -8 ±- () - ±- () 7-8 h 8- ± - () -7 ±- () 9--day explants -h - ± - () -8 ±- () 7-8 h -7 ± - () - ±- () --day explants -h - ±- () - ±- () Tests for difference in number of somites by Student's t-test: (a) Between types of explants At 7-8h At - h 8--day 9--day P< - P< - P< - P< - --day no P< - (b) At the two time points PM+PS/TB PM-PS/TB P< - no P< - no P< - F<- y r H
11 Table. The somite-forming capacity of different portions ofpresomitic mesoderm of 8--day to --day mouse embryos 8--day 9--day --day --day No. of somites in the embryo -7 ±- () - ± - (7) - ± -9 () - ± - (7) 798 ± 8 () ± (7) 77 ± (9) 7 ± 7 (9) Length (jum) of presomitic mesoderm + caudal tissues Length (jum) of portions of presomitic mesoderm that formed: somites somites somites somites somites 7 somites 8 () ± () ± () ± 9 () ±7 (8) ± () () 8 ± () 9 ± 9 (9) 7 ± 8 () 8 ± 8 (7) 9 ± () () ± 9 () ± 9 () 97 ± () 9 ± 9 (9) ± () ± () 7 ± 7 () 7 ± (7) 7 ± (8) Statistical analysis by single variable logarithmic regression: [Size of explant] = a+b-log[number of somites] Regression coefficient a Regression coefficient b Coefficient of determination Covariance Correlation coefficient Significance at P value -8 ± -9 - ± < - -7 ± -9 - ± < ± -8 - ± <- -7 ± - - ± < - as O VO

12 8 P. P. L. TAM 8--day VI I VII I PS 9--day III IV VII PS/TB --day IV VI TB --day III IV I VI I TB Length 8 Fig. 8. A map of prospective somites in the presomitic mesoderm of 8--day to -- day mouse embryos. The prospective somites were labelled I to VII from the cranial end of the presomitic mesoderm. The position of the boundary between prospective somites was derived from the regressional equations in Table, but that between I and II was estimated by an equal partition of the domain occupied by the two units. In the caudal tissue, PS refers to the primitive streak and TB refers to the tail bud. structures normally associated with the segmental plate (Packard & Jacobson, 97, 979; Stern & Bellairs, 98a). However, the size and shape of the somites were altered under in vitro conditions because of changes in the physical force exerted by the surrounding epithelia and neural tube (Menkes & Sandor, 977; Packard & Jacobson, 979; Stern & Bellairs, 98a; the present study). The other compelling evidence for a prepattern of somites is the demonstration of a developmental relationship between somitomeres initially present in the presomitic mesoderm and the somites formed in the explants. The presence of distinctive clusters of mesenchymal cells which show a unique concentric orientation about a centre was first described in the paraxial mesoderm of the chick embryo (Meier, 979). Subsequently similar cellular patterns are also found in the apparently unsegmented cranial mesoderm of newt (Jacobson & Meier, 98), snapping turtle (Meier & Packard, 98), chick, quail (Meier, 98, 98) and mouse (Meier & Tam, 98) and in the presomitic mesoderm and segmental plate of snapping turtle (Packard & Meier, 98), birds (Packard & Meier, 98) and mouse (Tam etal 98). It has been shown in the present study on mouse embryos and in other studies on avian (Packard & Meier, 98) and snapping turtle embryos (Packard & Meier, 98) that the number of somitomeres identified in the presomitic mesoderm or segmental plate correlates very well with the number of prospective somites contained in the same tissues. The formation of a somite in the explants results in the corresponding disappearance of one somitomere. The somitomeric organization in the presomitic mesoderm is therefore a morphological manifestation of the metameric pattern found in the embryonic axis.

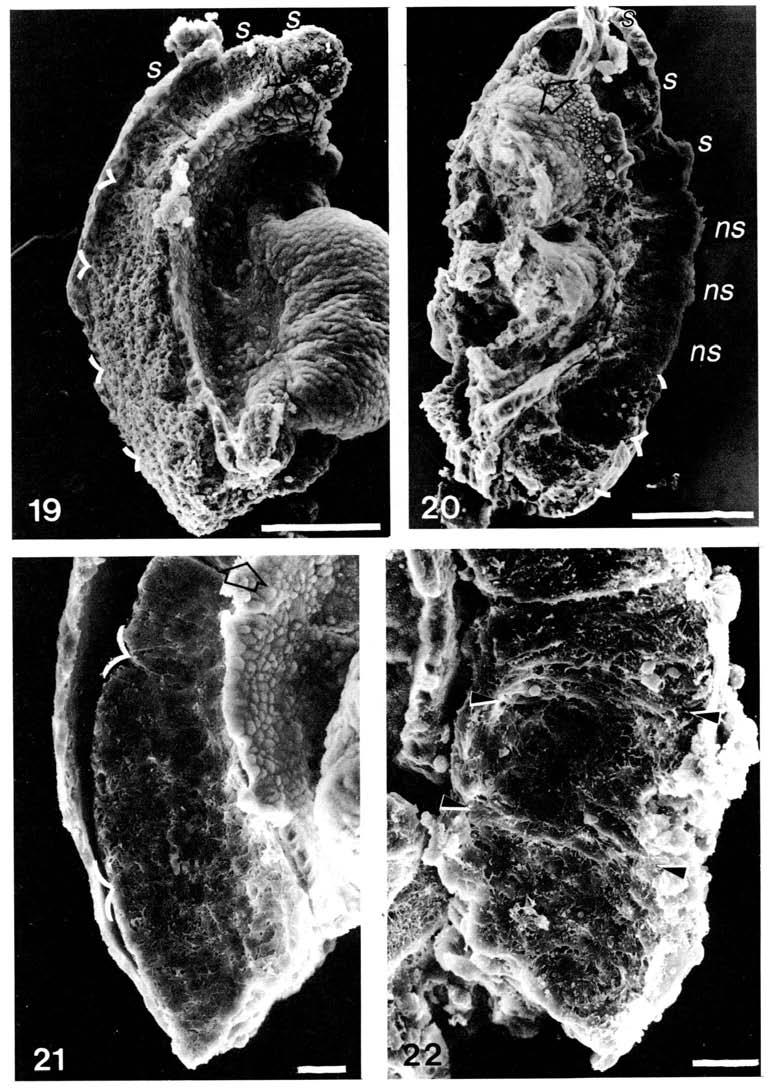
13 Differentiation ofpresomitic mesoderm 8 It becomes apparent from the map shown in Fig. 8 that in order to accommodate all six to seven prospective somites in presomitic mesoderm of different sizes, each and every prospective somite undergoes a proportional change in its dimensions. The more caudal members occupy smaller domains than the cranial counterparts. Since the series of prospective somites represents progressively more mature entities culminating in somite segmentation, an expansion of the prospective somites after their inception might be an essential step in the differentiative process of becoming an epithelial somite. This expansion in size may be coincidental to the burst of cellular proliferation observed in the cranial portion of the segmental plate (Stern & Bellairs, 98ft). Such a transition from a closely packed to an expanded state of prospective somites has recently been described in the tail bud of the Xenopus embryo (Elsdale & Davidson, 98). The so-called packing zone in the tail bud of Xenopus may be equivalent to the primitive streak of the chick (Meier & Jacobson, 98; Stern & Bellairs, 98a) and the caudal tissue of the mouse (Tarn, 98). The zone of extension and zone of prepattern are perhaps analogous to the caudal and cranial portions, respectively, of the presomitic mesoderm in the mouse embryo. The present study also showed that somite formation stopped abruptly in explants without the caudal tissue when the entire presomitic mesoderm has been segmented. A similar truncation of the somitic pattern occurred after the removal of the tail bud in amphibians (Cooke, 97; Elsdale & Davidson, 98), and the primitive streak in birds (Packard & Jacobson, 97; Packard, 98ft; Packard & Meier, 98), snapping turtle (Packard, 98a) and mouse (Smith, 9). The caudal tissue which has extensive histogenetic potential (Tarn, 98) has been suggested as an important source of cells for somitogenesis (Tarn, 98). It has recently been postulated that in the chick embryo the process of somite formation depends upon the constant interaction of a small population of somitic progenitor cells with new cells generated from the primitive streak and the tail bud. The immediate product of such an interaction is the formation of the somitomere or the prospective somite (Bellairs & Veini, 98). It has yet to be shown that there are Fig. 9. Scanning electron micrograph of the bisected explant. The neural tube was dissected away to expose the presomitic mesoderm. Three somites (s) andfivesomitomeres were identified. The intersomitomeric boundary was marked by curved lines. Arrow points cranially. Bar, jum. Fig.. Scanning electron micrograph of the corresponding half of the explant after 8 h in vitro. Three new somites (ns) were formed and two somitomeres were seen at the caudal part of the explants. The somitomeres were marked by curved lines and the three somites originally present were labelled (s). Arrow points cranially. Bar, jum. Fig.. Scanning electron micrograph showing the cellular pattern seen in three somitomeres in the presomitic mesoderm of an explant fixed before culture. The intersomitomeric boundary was marked by curved lines. Cells in the cranial-most somitomere were more closely packed. Arrow points cranially. Bar, jum. Fig.. Scanning electron micrograph showing the medial aspect of somites derived from the three somitomeres showed in Fig.. The somites were compressed craniocaudally because of the reduced elongation of the axial structures. Positions of intersomitic fissures were indicated by arrow heads. Arrow points cranially. Bar, jum.

14 8 P. P. L. TAM 9
15 Differentiation of presomitic mesoderm 8 Table. A comparison of the number of metameric segments in the presomitic mesoderm before and after culture Original explant Explants after culture Somites Somitomeres* New somitesf Somitomeres Total segments* 8--day 8 ± - () 7 9--day l±-() o. 7 * Significant correlation between these two values (r = -8, P< -, d.f. = by linear correlation analysis and Student's t-test). t Number of new somites = (total number of somites present in the explant at the end of culture)-(number of somites originally present before culture). indeed two cell populations within a somitomere or later in a somite, but the primitive streak and the tail bud of the chick embryo have been shown to contribute cells to the somites (Meier & Jacobson, 98; Schoenwolf, 977). The precise role of the caudal tissue in the specification of somitomeres or prospective somites in the mouse embryo remains to be elucidated. REFERENCES ADELMAN, H. B. (9). The development of neural folds and cranial ganglia of the rat. /. comp. Neurol. 9, 9-7.
.")
16 8 P. P. L. TAM BELLAIRS, R. (979). The mechanism of somite segmentation in the chick embryo. /. Embryol. exp. Morph., 7-. BELLAIRS, R. & VEINI, M. (98). Experimental analysis of control mechanisms in somite segmentation in avian embryos. II. Reduction of materials in the gastrula stage of the chick. /. Embryol. exp. Morph. 79, 8-. COOKE, J. (97). Control of somite number during development of a vertebrate, Xenopus laevis. Nature, Lond., COOKE, J. (977). The control of somite number during amphibian development: model and experiment. In Vertebrate Limb and Somite Morphogenesis (ed. D. A. Ede, J. R. Hinchliffe & M. Balls), pp. -8. Cambridge University Press. DEUCHAR, E. M. & BURGESS, A. M. C. (97). Somite segmentation in amphibian embryos: is there a transmitted mechanism?/. Embryol. exp. Morph. 7, 9-8. ELSDALE, T. & DAVIDSON, D. (98). Somitogenesis in amphibian. IV. The dynamics of tail development. /. Embryol. exp. Morph. 7, 7-7. FLINT, O. P., EDE, D. A., WILBY, O. K. & PROCTOR, J. (978). Control of somite number in normal and amputated mutant mouse embryos: an experimental and a theoretical analysis. /. Embryol. exp. Morph., 89-. JACOBSON, A. G. & MEIER, S. (98). Morphogenesis of the head of a newt: mesodermal segments, neuromeres and distribution of neural crest. Devi Biol., 8-9. LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L. & RANDALL, R. J. (9). Protein measurement with the Folin reagent. /. biol. Chem. 9, -7. MEIER, S. (979). Development of the chick embryo mesoblast. Formation of the embryonic axis and establishment of the metameric pattern. Devi Biol. 7, -. MEIER, S. (98). Development of the chick mesoblast: morphogenesis of the prechordal plate and cranial segments. Devi Biol. 8, 9-. MEIER, S. (98). The development of segmentation in the cranial region of vertebrate embryos. Scanning Electron Microscopy in, 9-8. MEIER, S. (98). Somite formation and its relationship to metameric pattern in the mesoderm. CellDiff.,-. MEIER, S. & JACOBSON, A. G. (98). Experimental studies of the origin and expression of metameric pattern in the chick embryo. /. exp. Zool. 9, 7-. MEIER, S. & PACKARD, D. S. JR (98). Morphogenesis of the cranial segments and distribution of the neural crest in the embryos of the snapping turtle, Chelydra serpentina. Devi Biol., 9-. MEIER, S. & TAM, P. P. L. (98). Metameric pattern development in the embryonic axis of the mouse. I. Differentiation of the cranial segments. Differentiation, 9-8. MENKES, B. & SANDOR, S. (977). Somitogenesis, regulation potencies, sequence determination and primordial interactions. In Vertebrate Limb and Somite Morphogenesis (ed. D. A Ede, J. R. Hinchliffe & M. Balls), pp. -9. Cambridge University Press. NEAL, H. V. (98). Neuromeres and metameres. /. Morph., 9-. PACKARD, D. S. JR (978). Chick somite determination: the role of factors in young somites and the segmental plate. /. exp. Zool., 9-. PACKARD, D. S. JR (98a). Somite formation in cultured embryos of the snapping turtle, Chelydra serpentina. J. Embryol. exp. Morph. 9, -. PACKARD, D. S. JR (98). Somitogenesis in cultured embryos of the Japanese quail, Coturnix coturnix japonica. Amer. J. Anat. 8, 8-9. PACKARD, D. S. JR & JACOBSON, A. G. (97). The influence of axial structures on chick somite formation. Devi Biol., -8. PACKARD, D. S. JR & JACOBSON, A. G. (979). Analysis of the physical forces that influence the shape of the chick somites. /. exp. Zool. 7, 8-9. PACKARD, D. S. JR & MEIER, S. (98). An experimental study of the somitomeric organization of the avian segmental plate. Devi Biol. 97,9-. PACKARD, D. S. JR & MEIER, S. (98). Morphological and experimental studies of the somitomeric organization of the segmental plate in snapping turtle embryos. /. Embryol. exp. Morph. 8, -8. SCHOENWOLF, G. C. (977). Tail (end) bud contributions to the posterior region of the chick embryo. J. exp. Zool., 7-.
. The roles of node regression and elongation of the area pellucida in the formation of somites in avian embryos. J. Embryol. exp. Morph. 8, 7-9. STERN, C. D. & BELLAIRS, R. (98b).")
17 Differentiation ofpresomitic mesoderm 8 SMITH, L. J. (9). The effects of transection and extirpation on axis formation and elongation in the young mouse embryo. /. Embryol. exp. Morph., STERN, C. D. & BELLAIRS, R. (98a). The roles of node regression and elongation of the area pellucida in the formation of somites in avian embryos. J. Embryol. exp. Morph. 8, 7-9. STERN, C. D. & BELLAIRS, R. (98b). Mitotic activity during somite segmentation in the early chick embryo. Anat. Embryol. 9, 97-. TAM, P. P. L. (98). The control of somitogenesis in mouse embryos. /. Embryol. exp. Morph., Supplement, -8. TAM, P. P. L. (98). The histogenetic capacity of tissues at the caudal end of the embryonic axis in the mouse. J. Embryol. exp. Morph. 8, -. TAM, P. P. L., MEIER, S. & JACOBSON, A. G. (98). Differentiation of the metameric pattern in the embryonic axis of the mouse. II. Somitomeric organization of the presomitic mesoderm. Differentiation,9-. {Accepted September 98)
18
Differentiation 0 Springer-Verlag Differentiation of the Metameric Pattern in the Embryonic Axis of the Mouse
 Differentiation (1982) 21 : 109-122 Differentiation 0 Springer-Verlag 1982 Differentiation of the Metameric Pattern in the Embryonic Axis of the Mouse II. Somitomeric Organization of the Presodtic Mesoderm
Differentiation (1982) 21 : 109-122 Differentiation 0 Springer-Verlag 1982 Differentiation of the Metameric Pattern in the Embryonic Axis of the Mouse II. Somitomeric Organization of the Presodtic Mesoderm
The allocation of cells in the presomitic mesoderm during somite segmentation in the mouse embryo
 Development 103. 379-390 (1988) Printed in Great Britain The Company of Biologists Limited 1988 379 The allocation of cells in the presomitic mesoderm during somite segmentation in the mouse embryo PATRICK
Development 103. 379-390 (1988) Printed in Great Britain The Company of Biologists Limited 1988 379 The allocation of cells in the presomitic mesoderm during somite segmentation in the mouse embryo PATRICK
Somitomeres: mesodermal segments of vertebrate embryos
 Development 104 Supplement, 209-220 (1988) Printed in Great Britain @ The Company of Biologists Limited 1988 209 Somitomeres: mesodermal segments of vertebrate embryos ANTONE G. JACOBSON Department of
Development 104 Supplement, 209-220 (1988) Printed in Great Britain @ The Company of Biologists Limited 1988 209 Somitomeres: mesodermal segments of vertebrate embryos ANTONE G. JACOBSON Department of
Behavioural properties of chick somitic mesoderm and lateral plate when explanted in vitro
 /. Embryol. exp. Morph. Vol. 56, pp. 41-58,1980 41 Printed in Great Britain Company of Biologists Limited 1980 Behavioural properties of chick somitic mesoderm and lateral plate when explanted in vitro
/. Embryol. exp. Morph. Vol. 56, pp. 41-58,1980 41 Printed in Great Britain Company of Biologists Limited 1980 Behavioural properties of chick somitic mesoderm and lateral plate when explanted in vitro
Sonic hedgehog (Shh) signalling in the rabbit embryo
 signalling in the rabbit embryo") Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
Inhibition of cranial neural crest cell development by vitamin A in the cultured chick embryo
 /. Embryol. exp. Morph. Vol. 39, pp. 267-27J, 1977 267 Printed in Great Britain Inhibition of cranial neural crest cell development by vitamin A in the cultured chick embryo JOHN R. HASSELL, 1 JUDITH H.
/. Embryol. exp. Morph. Vol. 39, pp. 267-27J, 1977 267 Printed in Great Britain Inhibition of cranial neural crest cell development by vitamin A in the cultured chick embryo JOHN R. HASSELL, 1 JUDITH H.
Fish swimbladder: an excellent meso dermal inductor in primary embryonic induction
 /. Embryo/, exp. Morph. Vol 36,, pp. 315-30, 1976 315 Printed in Great Britain Fish swimbladder: an excellent meso dermal inductor in primary embryonic induction IZUMI KAWAKAMI 1 From the Department of
/. Embryo/, exp. Morph. Vol 36,, pp. 315-30, 1976 315 Printed in Great Britain Fish swimbladder: an excellent meso dermal inductor in primary embryonic induction IZUMI KAWAKAMI 1 From the Department of
An increase in cell cell adhesion in the chick segmental plate results in a meristic pattern
 J. Embryol. exp. Morph. 79, 1-10 (1984) Printed in Great Britain (E) The Company of Biologists Limited 1984 An increase in cell cell adhesion in the chick segmental plate results in a meristic pattern
J. Embryol. exp. Morph. 79, 1-10 (1984) Printed in Great Britain (E) The Company of Biologists Limited 1984 An increase in cell cell adhesion in the chick segmental plate results in a meristic pattern
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Somitogenesis in amphibia
 /. Embryol. exp. Morph. 76, 157-176 (1983) 257 Printed in Great Britain The Company of Biologists Limited 1983 Somitogenesis in amphibia IV. The dynamics of tail development By TOM ELSDALE 1 AND DUNCAN
/. Embryol. exp. Morph. 76, 157-176 (1983) 257 Printed in Great Britain The Company of Biologists Limited 1983 Somitogenesis in amphibia IV. The dynamics of tail development By TOM ELSDALE 1 AND DUNCAN
The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis
 Development 99, 19-126 (1987) Printed in Great Britain The Company of Biologists Limited 1987 19 The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis P. P.
Development 99, 19-126 (1987) Printed in Great Britain The Company of Biologists Limited 1987 19 The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis P. P.
Mesoderm Development
 Quiz rules: Spread out across available tables No phones, text books, or (lecture) notes on your desks No consultation with your colleagues No websites open other than the Quiz page No screen snap shots
Quiz rules: Spread out across available tables No phones, text books, or (lecture) notes on your desks No consultation with your colleagues No websites open other than the Quiz page No screen snap shots
The effect of removing posterior apical ectodermal ridge of the chick wing and leg on pattern formation
 /. Embryol. exp. Morph. Vol. 65 {Supplement), pp. 309-325, 1981 309 Printed in Great Britain Company of Biologists Limited 1981 The effect of removing posterior apical ectodermal ridge of the chick wing
/. Embryol. exp. Morph. Vol. 65 {Supplement), pp. 309-325, 1981 309 Printed in Great Britain Company of Biologists Limited 1981 The effect of removing posterior apical ectodermal ridge of the chick wing
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm December 7, 2006 Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of BMPs more lateral mesoderm
Biology 4361 Paraxial and Intermediate Mesoderm December 7, 2006 Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of BMPs more lateral mesoderm
The formation of the gonadal ridge in Xenopus laevis
 /. Embryol. exp. Morph. Vol. 35, 1, pp. 139-148, 1976 \ 39 Printed in Great Britain The formation of the gonadal ridge in Xenopus laevis II. A scanning electron microscope study By C. C. WYL1E, 1 M. BANCROFT
/. Embryol. exp. Morph. Vol. 35, 1, pp. 139-148, 1976 \ 39 Printed in Great Britain The formation of the gonadal ridge in Xenopus laevis II. A scanning electron microscope study By C. C. WYL1E, 1 M. BANCROFT
Myoblasts and notochord influence the orientation of somitic myoblasts from Xenopus laevis
 /. Embryol. exp. Morph. 93, 121-131 (1986) 121 Printed in Great Britain The Company of Biologists Limited 1986 Myoblasts and notochord influence the orientation of somitic myoblasts from Xenopus laevis
/. Embryol. exp. Morph. 93, 121-131 (1986) 121 Printed in Great Britain The Company of Biologists Limited 1986 Myoblasts and notochord influence the orientation of somitic myoblasts from Xenopus laevis
Life Sciences For NET & SLET Exams Of UGC-CSIR. Section B and C. Volume-08. Contents A. BASIC CONCEPT OF DEVELOPMENT 1
 Section B and C Volume-08 Contents 5. DEVELOPMENTAL BIOLOGY A. BASIC CONCEPT OF DEVELOPMENT 1 B. GAMETOGENESIS, FERTILIZATION AND EARLY DEVELOPMENT 23 C. MORPHOGENESIS AND ORGANOGENESIS IN ANIMALS 91 0
Section B and C Volume-08 Contents 5. DEVELOPMENTAL BIOLOGY A. BASIC CONCEPT OF DEVELOPMENT 1 B. GAMETOGENESIS, FERTILIZATION AND EARLY DEVELOPMENT 23 C. MORPHOGENESIS AND ORGANOGENESIS IN ANIMALS 91 0
Positional signalling along the anteroposterior axis of the chick wing. The effect of multiple polarizing region grafts
 /. Embryol exp. Morph. Vol. 6, pp. 5-59, 98 J5 Printed in Great Britain Company of Biologists Limited 98 Positional signalling along the anteroposterior axis of the chick wing. The effect of multiple polarizing
/. Embryol exp. Morph. Vol. 6, pp. 5-59, 98 J5 Printed in Great Britain Company of Biologists Limited 98 Positional signalling along the anteroposterior axis of the chick wing. The effect of multiple polarizing
Somites without a clock
 Dias, AS; de Almeida, I; Belmonte, JM; Glazier, JA; Stern, CD; (2014) Somites Without a Clock. Science 10.1126/science.1247575. ARTICLE Somites without a clock Ana S. Dias 1,, Irene de Almeida 1,, Julio
Dias, AS; de Almeida, I; Belmonte, JM; Glazier, JA; Stern, CD; (2014) Somites Without a Clock. Science 10.1126/science.1247575. ARTICLE Somites without a clock Ana S. Dias 1,, Irene de Almeida 1,, Julio
Mesodermal expansion after arrest of the edge in the area vasculosa of the chick
 /. Embryol. exp. Morph. Vol. 41, pp. 175-188, 1977 175 Printed in Great Britain Company of Biologists Limited 1977 Mesodermal expansion after arrest of the edge in the area vasculosa of the chick By J.
/. Embryol. exp. Morph. Vol. 41, pp. 175-188, 1977 175 Printed in Great Britain Company of Biologists Limited 1977 Mesodermal expansion after arrest of the edge in the area vasculosa of the chick By J.
The growth rate of sensory nerve fibres in the mammalian embryo
 Development 00, 307-3 (987) Printed in Great Britain The Company of Biologists Limited 987 307 The growth rate of sensory nerve fibres in the mammalian embryo ALUN M. DAVIES Department of Anatomy, Si George's
Development 00, 307-3 (987) Printed in Great Britain The Company of Biologists Limited 987 307 The growth rate of sensory nerve fibres in the mammalian embryo ALUN M. DAVIES Department of Anatomy, Si George's
Is there a ventral neural ridge in chick embryos? Implications for the origin of adenohypophyseal and other APUD cells
 /. Embryol. exp. Morph. Vol. 57, pp. 71-78, 1980 7 \ Printed in Great Britain Company of Biologists Limited 1980 Is there a ventral neural ridge in chick embryos? Implications for the origin of adenohypophyseal
/. Embryol. exp. Morph. Vol. 57, pp. 71-78, 1980 7 \ Printed in Great Britain Company of Biologists Limited 1980 Is there a ventral neural ridge in chick embryos? Implications for the origin of adenohypophyseal
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Unit 4 Evaluation Question 1:
 Name: Unit 4 Evaluation Question 1: /7 points A naturally occurring dominant mutant in mice is the Doublefoot (Dbf) mutant. Below is an image of the bones from a wildtype (wt) and Doublefoot mutant mouse.
Name: Unit 4 Evaluation Question 1: /7 points A naturally occurring dominant mutant in mice is the Doublefoot (Dbf) mutant. Below is an image of the bones from a wildtype (wt) and Doublefoot mutant mouse.
Onset of the segmentation clock in the chick embryo: evidence for oscillations in the somite precursors in the primitive streak
 Development 129, 1107-1117 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV2742 1107 Onset of the segmentation clock in the chick embryo: evidence for oscillations in the somite
Development 129, 1107-1117 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV2742 1107 Onset of the segmentation clock in the chick embryo: evidence for oscillations in the somite
Determination of epithelial half-somites in skeletal morphogenesis
 Development 116, 441-445 (1992) Printed in Great Britain The Company of Biologists Limited 1992 441 Determination of epithelial half-somites in skeletal morphogenesis RONALD S. GOLDSTEIN * and CHAYA KALCHEIM
Development 116, 441-445 (1992) Printed in Great Britain The Company of Biologists Limited 1992 441 Determination of epithelial half-somites in skeletal morphogenesis RONALD S. GOLDSTEIN * and CHAYA KALCHEIM
Cell rearrangement and segmentation in Xenopus: direct observation of cultured explants
 Development 105, 155-166 (1989) Printed in Great Britain The Company of Biologists Limited 1989 155 Cell rearrangement and segmentation in Xenopus: direct observation of cultured explants PAUL A. WILSON
Development 105, 155-166 (1989) Printed in Great Britain The Company of Biologists Limited 1989 155 Cell rearrangement and segmentation in Xenopus: direct observation of cultured explants PAUL A. WILSON
2/23/09. Regional differentiation of mesoderm. Morphological changes at early postgastrulation. Segments organize the body plan during embryogenesis
 Regional differentiation of mesoderm Axial Paraxial Intermediate Somatic Splanchnic Chick embryo Morphological changes at early postgastrulation stages Segments organize the body plan during embryogenesis
Regional differentiation of mesoderm Axial Paraxial Intermediate Somatic Splanchnic Chick embryo Morphological changes at early postgastrulation stages Segments organize the body plan during embryogenesis
A cell lineage analysis of segmentation in the chick embryo
 Development 104 Supplement, 231-244 (1988) 231 Printed in Great Britain The Company of Biologists Limited 1988 A cell lineage analysis of segmentation in the chick embryo CLAUDIO D. STERN 1, SCOTT E. FRASER
Development 104 Supplement, 231-244 (1988) 231 Printed in Great Britain The Company of Biologists Limited 1988 A cell lineage analysis of segmentation in the chick embryo CLAUDIO D. STERN 1, SCOTT E. FRASER
Somites in Developing Embryos
 Somites in Developing Embryos NATO ASI Series Advanced Science Institutes Series Aseries presenting the results o( activities sponsored by the NA TO Science Committee, which aims at the dissemination o(
Somites in Developing Embryos NATO ASI Series Advanced Science Institutes Series Aseries presenting the results o( activities sponsored by the NA TO Science Committee, which aims at the dissemination o(
A novel signal induces a segmentation fissure by acting in a ventral-to-dorsal direction in the presomitic mesoderm
 Developmental Biology 282 (2005) 183 191 www.elsevier.com/locate/ydbio A novel signal induces a segmentation fissure by acting in a ventral-to-dorsal direction in the presomitic mesoderm Yuki Sato a, Yoshiko
Developmental Biology 282 (2005) 183 191 www.elsevier.com/locate/ydbio A novel signal induces a segmentation fissure by acting in a ventral-to-dorsal direction in the presomitic mesoderm Yuki Sato a, Yoshiko
Glycosaminoglycan localization and role in maintenance of tissue spaces in the early chick embryo 1
 /. Embryo!'. exp. Morph. Vol. 42, pp. 195-207, 1977 195 Printed in Great Britain Company of Biologists Limited 1977 Glycosaminoglycan localization and role in maintenance of tissue spaces in the early
/. Embryo!'. exp. Morph. Vol. 42, pp. 195-207, 1977 195 Printed in Great Britain Company of Biologists Limited 1977 Glycosaminoglycan localization and role in maintenance of tissue spaces in the early
Role of Organizer Chages in Late Frog Embryos
 Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
N-Cadherin/Catenin-Mediated Morphoregulation of Somite Formation
 DEVELOPMENTAL BIOLOGY 202, 85 102 (1998) ARTICLE NO. DB989025 N-Cadherin/Catenin-Mediated Morphoregulation of Somite Formation Kersti K. Linask,*,1 Cheryl Ludwig,* Ming-Da Han,* Xiao Liu,* Glenn L. Radice,
DEVELOPMENTAL BIOLOGY 202, 85 102 (1998) ARTICLE NO. DB989025 N-Cadherin/Catenin-Mediated Morphoregulation of Somite Formation Kersti K. Linask,*,1 Cheryl Ludwig,* Ming-Da Han,* Xiao Liu,* Glenn L. Radice,
Intercellular contacts at the epithelial-mesenchymal interface of the developing rat submandibular gland in vitro
 /. Embryol. exp. Morph. Vol. 39, pp. 71-77, 1977 71 Printed in Great Britain Intercellular contacts at the epithelial-mesenchymal interface of the developing rat submandibular gland in vitro By LESLIE
/. Embryol. exp. Morph. Vol. 39, pp. 71-77, 1977 71 Printed in Great Britain Intercellular contacts at the epithelial-mesenchymal interface of the developing rat submandibular gland in vitro By LESLIE
Maps of strength of positional signalling activity in the developing chick wing bud
 /. Embryol. exp. Morph. 87, 163-174 (1985) 163 Printed in Great Britain The Company of Biologists Limited 1985 Maps of strength of positional signalling activity in the developing chick wing bud LAWRENCE
/. Embryol. exp. Morph. 87, 163-174 (1985) 163 Printed in Great Britain The Company of Biologists Limited 1985 Maps of strength of positional signalling activity in the developing chick wing bud LAWRENCE
Homeobox genes and the vertebrate head
 Development 103 Supplement, 17-24 (1988) Printed in Great Britain The Company of Biologists Limited 1988 17 Homeobox genes and the vertebrate head PETER W. H. HOLLAND Department of Zoology, University
Development 103 Supplement, 17-24 (1988) Printed in Great Britain The Company of Biologists Limited 1988 17 Homeobox genes and the vertebrate head PETER W. H. HOLLAND Department of Zoology, University
Fate and function of the ventral ectodermal ridge during mouse tail development
 Development 127, 2113-2123 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV3148 2113 Fate and function of the ventral ectodermal ridge during mouse tail development Devorah C.
Development 127, 2113-2123 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV3148 2113 Fate and function of the ventral ectodermal ridge during mouse tail development Devorah C.
Translocation of neural crest cells within a hydrated collagen lattice
 /. Embryol. exp. Morph. Vol. 55, pp. 17-31, 1980 \J Printed in Great Britain Company of Biologists Limited 1980 Translocation of neural crest cells within a hydrated collagen lattice By EDWARD M. DAVIS
/. Embryol. exp. Morph. Vol. 55, pp. 17-31, 1980 \J Printed in Great Britain Company of Biologists Limited 1980 Translocation of neural crest cells within a hydrated collagen lattice By EDWARD M. DAVIS
The dorsal neural tube organizes the dermamyotome and induces axial myocytes in the avian embryo
 Development 122, 231-241 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV3253 231 The dorsal neural tube organizes the dermamyotome and induces axial myocytes in the avian embryo
Development 122, 231-241 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV3253 231 The dorsal neural tube organizes the dermamyotome and induces axial myocytes in the avian embryo
presumptiv e germ layers during Gastrulatio n and neurulation Somites
 Vertebrate embryos are similar at the phylotypic stage Patterning the Vertebrate Body Plan II: Mesoderm & Early Nervous System Wolpert L, Beddington R, Jessell T, Lawrence P, Meyerowitz E, Smith J. (2001)
Vertebrate embryos are similar at the phylotypic stage Patterning the Vertebrate Body Plan II: Mesoderm & Early Nervous System Wolpert L, Beddington R, Jessell T, Lawrence P, Meyerowitz E, Smith J. (2001)
Development of neural tube basal lamina during neurulation and neural crest cell emigration in the trunk of the mouse embryo
 /. Embryol. exp. Morph. 98, 219-236 (1986) 219 Printed in Great Britain (E) The Company of Biologists Limited 1986 Development of neural tube basal lamina during neurulation and neural crest cell emigration
/. Embryol. exp. Morph. 98, 219-236 (1986) 219 Printed in Great Britain (E) The Company of Biologists Limited 1986 Development of neural tube basal lamina during neurulation and neural crest cell emigration
Vital dye analysis of cranial neural crest cell migration in the mouse embryo
 Development 116, 297-307 (1992) Printed in Great Britain The Company of Biologists Limited 1992 297 Vital dye analysis of cranial neural crest cell migration in the mouse embryo GEORGE N. SERBEDZIJA 1,*,
Development 116, 297-307 (1992) Printed in Great Britain The Company of Biologists Limited 1992 297 Vital dye analysis of cranial neural crest cell migration in the mouse embryo GEORGE N. SERBEDZIJA 1,*,
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
Exam 3 (Final Exam) December 20, 2007
 December 20, 2007") Biology 4361 Exam 3 (Final Exam) December 20, 2007 Name: ID: Multiple choice (1 point each. Indicate the best answer.) 1. During Drosophila gastrulation, mesoderm moves in through the a. primitives streak.
Biology 4361 Exam 3 (Final Exam) December 20, 2007 Name: ID: Multiple choice (1 point each. Indicate the best answer.) 1. During Drosophila gastrulation, mesoderm moves in through the a. primitives streak.
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
SIGNIFICANCE OF EMBRYOLOGY
 This lecture will discuss the following topics : Definition of Embryology Significance of Embryology Old and New Frontiers Introduction to Molecular Regulation and Signaling Descriptive terms in Embryology
This lecture will discuss the following topics : Definition of Embryology Significance of Embryology Old and New Frontiers Introduction to Molecular Regulation and Signaling Descriptive terms in Embryology
In ovo time-lapse analysis after dorsal neural tube ablation shows rerouting of chick hindbrain neural crest
 In ovo time-lapse analysis after dorsal neural tube ablation shows rerouting of chick hindbrain neural crest Paul Kulesa, Marianne Bronner-Fraser and Scott Fraser (2000) Presented by Diandra Lucia Background
In ovo time-lapse analysis after dorsal neural tube ablation shows rerouting of chick hindbrain neural crest Paul Kulesa, Marianne Bronner-Fraser and Scott Fraser (2000) Presented by Diandra Lucia Background
Developmental Zoology. Ectodermal derivatives (ZOO ) Developmental Stages. Developmental Stages
 Developmental Stages. Developmental Stages") Developmental Zoology (ZOO 228.1.0) Ectodermal derivatives 1 Developmental Stages Ø Early Development Fertilization Cleavage Gastrulation Neurulation Ø Later Development Organogenesis Larval molts Metamorphosis
Developmental Zoology (ZOO 228.1.0) Ectodermal derivatives 1 Developmental Stages Ø Early Development Fertilization Cleavage Gastrulation Neurulation Ø Later Development Organogenesis Larval molts Metamorphosis
Muscle-forming potential of the non-somitic cells of the early avian limb bud
 /. Embryol exp. Morph. Vol. 54, pp. 209-217, 1979 209 Printed in Great Britain Company of Biologists Limited 1979 Muscle-forming potential of the non-somitic cells of the early avian limb bud By JOHN C.
/. Embryol exp. Morph. Vol. 54, pp. 209-217, 1979 209 Printed in Great Britain Company of Biologists Limited 1979 Muscle-forming potential of the non-somitic cells of the early avian limb bud By JOHN C.
The pattern of cell division during growth of the blastema of regenerating newt forelimbs
 /. Embryo], exp. Morph. Vol. 37, pp. 33-48, 1977 33 Printed in Great Britain The pattern of cell division during growth of the blastema of regenerating newt forelimbs By A. R. SMITH 1 AND A. M. CRAWLEY
/. Embryo], exp. Morph. Vol. 37, pp. 33-48, 1977 33 Printed in Great Britain The pattern of cell division during growth of the blastema of regenerating newt forelimbs By A. R. SMITH 1 AND A. M. CRAWLEY
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm July 28, 2008 Paraxial and Intermediate Mesoderm Overview Development of major mesodermal lineages Somites: formation specification and differentiation Mesodermal
Biology 4361 Paraxial and Intermediate Mesoderm July 28, 2008 Paraxial and Intermediate Mesoderm Overview Development of major mesodermal lineages Somites: formation specification and differentiation Mesodermal
The neural tube/notochord complex is necessary for vertebral but not limb and body wall striated muscle differentiation
 Development 115, 657-672 (1992) Printed in Great Britain The Company of Biologists Limited 1992 657 The neural tube/notochord complex is necessary for vertebral but not limb and body wall striated muscle
Development 115, 657-672 (1992) Printed in Great Britain The Company of Biologists Limited 1992 657 The neural tube/notochord complex is necessary for vertebral but not limb and body wall striated muscle
1/30/2009. Copyright The McGraw Hill Companies, Inc. Permission required for reproduction or display.
 CHAPTER 9 Architectural Pattern of an Animal New Designs for Living Zoologists recognize 34 major phyla of living multicellular animals Survivors of around 100 phyla that appeared 600 million years ago
CHAPTER 9 Architectural Pattern of an Animal New Designs for Living Zoologists recognize 34 major phyla of living multicellular animals Survivors of around 100 phyla that appeared 600 million years ago
Chapter 9. Benefits of Being Large. Levels of Organization in Organismal Complexity. Hierarchical Organization of Animal Complexity. Fig. 9.
 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Chapter 9 Architectural Pattern of an Animal Levels of Organization in Organismal Complexity Zoologists recognize
Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Chapter 9 Architectural Pattern of an Animal Levels of Organization in Organismal Complexity Zoologists recognize
Developmental Biology Lecture Outlines
 Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Mesoderm Divided into three main types - Paraxial (somite) - Intermediate - Lateral (somatic and splanchnic)
 - Intermediate - Lateral (somatic and splanchnic)") Mesoderm Divided into three main types - Paraxial (somite) - Intermediate - Lateral (somatic and splanchnic) Fates of Mesoderm Paraxial - Dermis of skin - Axial Skeleton - Axial and limb muscles/tendons
Mesoderm Divided into three main types - Paraxial (somite) - Intermediate - Lateral (somatic and splanchnic) Fates of Mesoderm Paraxial - Dermis of skin - Axial Skeleton - Axial and limb muscles/tendons
The role of FGF2 in craniofacial skeletogenesis
 The role of FGF2 in craniofacial skeletogenesis P. Ferretti, S. Sarkar, R. Moore, A. Petiot, C. J. Chan and A. Copp Summary E vidence that the major craniosynostosis syndromes are caused by mutations in
The role of FGF2 in craniofacial skeletogenesis P. Ferretti, S. Sarkar, R. Moore, A. Petiot, C. J. Chan and A. Copp Summary E vidence that the major craniosynostosis syndromes are caused by mutations in
Embryogenesis of an insect nervous system II: A second class of neuron precursor cells and the origin of the intersegmental connectives
 J. Embryol. exp. Morph. Vol. 61,pp. 317-330, 1981 3J7 Printed in Great Britain @ Company of Biologists Limited 1981 Embryogenesis of an insect nervous system II: A second class of neuron precursor cells
J. Embryol. exp. Morph. Vol. 61,pp. 317-330, 1981 3J7 Printed in Great Britain @ Company of Biologists Limited 1981 Embryogenesis of an insect nervous system II: A second class of neuron precursor cells
A clock-work somite. Kim J. Dale and Olivier Pourquié* Review articles. 72 BioEssays 22.1 BioEssays 22:72 83, 2000 John Wiley & Sons, Inc.
 A clock-work somite Kim J. Dale and Olivier Pourquié* Summary Somites are transient structures which represent the most overt segmental feature of the vertebrate embryo. The strict temporal regulation
A clock-work somite Kim J. Dale and Olivier Pourquié* Summary Somites are transient structures which represent the most overt segmental feature of the vertebrate embryo. The strict temporal regulation
The Incorporation of Labelled Amino-acids into Amphibian Embryos
 The Incorporation of Labelled Amino-acids into Amphibian Embryos by c. H. WADDINGTON and j. L. SIRLIN 1 Institute of Animal Genetics, Edinburgh INTRODUCTION T H E production of different cell types during
The Incorporation of Labelled Amino-acids into Amphibian Embryos by c. H. WADDINGTON and j. L. SIRLIN 1 Institute of Animal Genetics, Edinburgh INTRODUCTION T H E production of different cell types during
Paul A. Trainor and Patrick P. L. Tam* SUMMARY
 Development 121, 2569-2582 (1995) Printed in Great Britain The Company of Biologists Limited 1995 2569 Cranial paraxial mesoderm and neural crest cells of the mouse embryo: co-distribution in the craniofacial
Development 121, 2569-2582 (1995) Printed in Great Britain The Company of Biologists Limited 1995 2569 Cranial paraxial mesoderm and neural crest cells of the mouse embryo: co-distribution in the craniofacial
Gut Tube Development. For it s rare that a man thinks of anything so seriously as his dinner!
 For it s rare that a man thinks of anything so seriously as his dinner! Ben Johnson Gut Tube Development This chapter will follow the development of the embryonic gut caudal to the developing pharynx,
For it s rare that a man thinks of anything so seriously as his dinner! Ben Johnson Gut Tube Development This chapter will follow the development of the embryonic gut caudal to the developing pharynx,
Cranial paraxial mesoderm: regionalisation of cell fate and impact on craniofacial development in mouse embryos
 Development 120, 2397-2408 (1994) Printed in Great Britain The Company of Biologists Limited 1994 2397 Cranial paraxial mesoderm: regionalisation of cell fate and impact on craniofacial development in
Development 120, 2397-2408 (1994) Printed in Great Britain The Company of Biologists Limited 1994 2397 Cranial paraxial mesoderm: regionalisation of cell fate and impact on craniofacial development in
Interactions between somite cells: the formation and maintenance of segment boundaries in the chick embryo
 Development 99, 261-272 (1987) Printed in Great Britain The Company of Biologists Limited 1987 261 Interactions between somite cells: the formation and maintenance of segment boundaries in the chick embryo
Development 99, 261-272 (1987) Printed in Great Britain The Company of Biologists Limited 1987 261 Interactions between somite cells: the formation and maintenance of segment boundaries in the chick embryo
Aberrant Mitochondria with Longitudinal Cristae Observed in the Normal Rat Hepatic Parenchymal Cell. Takuma Saito and Kazuo Ozawa
 Okajimas Fol. anat. jap., 44 : 357-363, 1968 Aberrant Mitochondria with Longitudinal Cristae Observed in the Normal Rat Hepatic Parenchymal Cell By Takuma Saito and Kazuo Ozawa Department of Anatomy, Kansai
Okajimas Fol. anat. jap., 44 : 357-363, 1968 Aberrant Mitochondria with Longitudinal Cristae Observed in the Normal Rat Hepatic Parenchymal Cell By Takuma Saito and Kazuo Ozawa Department of Anatomy, Kansai
Regulation and potency in the forelimb rudiment of the axolotl embryo
 /. Embryol. exp. Morph. Vol. 57, pp. 23-27, 98 23 Printed in Great Britain Company of Biologists Limited 98 Regulation and potency in the forelimb rudiment of the axolotl embryo By J. M.W. SLACK From the
/. Embryol. exp. Morph. Vol. 57, pp. 23-27, 98 23 Printed in Great Britain Company of Biologists Limited 98 Regulation and potency in the forelimb rudiment of the axolotl embryo By J. M.W. SLACK From the
The patterning and functioning of protrusive activity during convergence and extension of the Xenopus organiser
 Development 1992 Supplement, 81-91 (1992) Printed in Great Britain C The Company of Biologist! Limited 1992 81 The patterning and functioning of protrusive activity during convergence and extension of
Development 1992 Supplement, 81-91 (1992) Printed in Great Britain C The Company of Biologist! Limited 1992 81 The patterning and functioning of protrusive activity during convergence and extension of
Development and degeneration of retina in rds mutant mice: observations in chimaeras of heterozygous mutant and normal genotype
 J. Embryol. exp. Morph. 98, 111-121 (1986) HI Printed in Great Britain The Company of Biologists Limited 1986 Development and degeneration of retina in rds mutant mice: observations in chimaeras of heterozygous
J. Embryol. exp. Morph. 98, 111-121 (1986) HI Printed in Great Britain The Company of Biologists Limited 1986 Development and degeneration of retina in rds mutant mice: observations in chimaeras of heterozygous
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11804 a Tailbud after cutting PSM after cutting b 3500 3000 2500 mean intensity 2000 1500 1000 ROI1 (TB) ROI2 (PSM) 500 0 0 1 2 3 4 5 6 7 8 9 time (h) Supplementary Fig.1 Lfng gene activity
doi:10.1038/nature11804 a Tailbud after cutting PSM after cutting b 3500 3000 2500 mean intensity 2000 1500 1000 ROI1 (TB) ROI2 (PSM) 500 0 0 1 2 3 4 5 6 7 8 9 time (h) Supplementary Fig.1 Lfng gene activity
A study of the growth cones of developing embryonic sensory neurites
 J. Embryol. exp. Morph. 75, 31-47 (1983) 3 \ Printed in Great Britain (G) The Company of Biologists Limited 1983 A study of the growth cones of developing embryonic sensory neurites By ALAN ROBERTS 1 AND
J. Embryol. exp. Morph. 75, 31-47 (1983) 3 \ Printed in Great Britain (G) The Company of Biologists Limited 1983 A study of the growth cones of developing embryonic sensory neurites By ALAN ROBERTS 1 AND
The role of extracellular matrix in the formation of the sclerotome
 /. Embryol. exp. Morph. Vol. 54, pp. 75-98, 1979 75 Printed in Great Britain Company of Biologists Limited 1979 The role of extracellular matrix in the formation of the sclerotome By MICHAEL SOLURSH 1,
/. Embryol. exp. Morph. Vol. 54, pp. 75-98, 1979 75 Printed in Great Britain Company of Biologists Limited 1979 The role of extracellular matrix in the formation of the sclerotome By MICHAEL SOLURSH 1,
Conclusions. The experimental studies presented in this thesis provide the first molecular insights
 C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
Determination of Spatiotemporal Interactions Regulating Head/Trunk Axial Pattern in the Chick Embryo
 Eukaryon, Vol. 2, January 2006, Lake Forest ollege Senior Thesis Determination of Spatiotemporal Interactions Regulating Head/Trunk Axial Pattern in the hick Embryo David Mann * Department of Biology,
Eukaryon, Vol. 2, January 2006, Lake Forest ollege Senior Thesis Determination of Spatiotemporal Interactions Regulating Head/Trunk Axial Pattern in the hick Embryo David Mann * Department of Biology,
A distinct development programme for the cranial paraxial mesoderm
 A distinct development programme for the cranial paraxial mesoderm Article (Published Version) Hacker, A and Guthrie, S (1998) A distinct development programme for the cranial paraxial mesoderm. Development
A distinct development programme for the cranial paraxial mesoderm Article (Published Version) Hacker, A and Guthrie, S (1998) A distinct development programme for the cranial paraxial mesoderm. Development
Tissue interactions affecting the migration and differentiation of neural crest cells in the chick embryo
 Development 113, 207-216 (1991) Printed in Great Britain The Company of Biologists Limited 1991 207 Tissue interactions affecting the migration and differentiation of neural crest cells in the chick embryo
Development 113, 207-216 (1991) Printed in Great Britain The Company of Biologists Limited 1991 207 Tissue interactions affecting the migration and differentiation of neural crest cells in the chick embryo
The Microscopic Observation of Mitosis in Plant and Animal Cells
 The Microscopic Observation of Mitosis in Plant and Animal Cells Prelab Assignment Before coming to lab, read carefully the introduction and the procedures for each part of the experiment, and then answer
The Microscopic Observation of Mitosis in Plant and Animal Cells Prelab Assignment Before coming to lab, read carefully the introduction and the procedures for each part of the experiment, and then answer
THE BEHAVIOUR OF CHLOROPLASTS DURING CELL DIVISION OF ISOETES LACUSTRIS L.
 New Phytol (1974) 73, 139-142. THE BEHAVIOUR OF CHLOROPLASTS DURING CELL DIVISION OF ISOETES LACUSTRIS L. BY JEAN M. WHATLEY Botany School, University of Oxford (Received 2 July 1973) SUMMARY Cells in
New Phytol (1974) 73, 139-142. THE BEHAVIOUR OF CHLOROPLASTS DURING CELL DIVISION OF ISOETES LACUSTRIS L. BY JEAN M. WHATLEY Botany School, University of Oxford (Received 2 July 1973) SUMMARY Cells in
Studies on the Mode of Outgrowth of the Amphibian Pronephric Duct
 Studies on the Mode of Outgrowth of the Amphibian Pronephric Duct by JANE OVERTON 1 From Hull Zoological Laboratory, University of Chicago, Chicago, Illinois WITH ONE PLATE INTRODUCTION ALTHOUGH the manner
Studies on the Mode of Outgrowth of the Amphibian Pronephric Duct by JANE OVERTON 1 From Hull Zoological Laboratory, University of Chicago, Chicago, Illinois WITH ONE PLATE INTRODUCTION ALTHOUGH the manner
An obsession with the chick
 Int. J. Dev. Biol. 62: 15-18 (2018) https://doi.org/10.1387/ijdb.180028rb www.intjdevbiol.com An obsession with the chick RUTH BELLAIRS* Department of Cell and Developmental Biology, University College
Int. J. Dev. Biol. 62: 15-18 (2018) https://doi.org/10.1387/ijdb.180028rb www.intjdevbiol.com An obsession with the chick RUTH BELLAIRS* Department of Cell and Developmental Biology, University College
10/15/09. Tetrapod Limb Development & Pattern Formation. Developing limb region is an example of a morphogenetic field
 Tetrapod Limb Development & Pattern Formation Figure 16.5(1) Limb Bud Formation derived from lateral plate (somatic) & paraxial (myotome) Fig. 16.2 Prospective Forelimb Field of Salamander Ambystoma maculatum
Tetrapod Limb Development & Pattern Formation Figure 16.5(1) Limb Bud Formation derived from lateral plate (somatic) & paraxial (myotome) Fig. 16.2 Prospective Forelimb Field of Salamander Ambystoma maculatum
* School of Biological Sciences, Carslaw Building, University of Sydney, Sydney, N.S.W By VERONICA H. K. Low*
 Aust. J. biol. Sci., 1971, 24, 187-95 * School of Biological Sciences, Carslaw Building, University of Sydney, Sydney, N.S.W. 2006.. NTRODUCTON A detailed survey of the morphological and anatomical effects
Aust. J. biol. Sci., 1971, 24, 187-95 * School of Biological Sciences, Carslaw Building, University of Sydney, Sydney, N.S.W. 2006.. NTRODUCTON A detailed survey of the morphological and anatomical effects
Coordination of Cell Differentiation and Migration in Mathematical Models of Caudal Embryonic Axis Extension
 Coordination of Cell Differentiation and Migration in Mathematical Models of Caudal Embryonic Axis Extension Nigel C. Harrison 1, Ruth Diez del Corral 2 *, Bakhtier Vasiev 1 * 1 Department of Mathematical
Coordination of Cell Differentiation and Migration in Mathematical Models of Caudal Embryonic Axis Extension Nigel C. Harrison 1, Ruth Diez del Corral 2 *, Bakhtier Vasiev 1 * 1 Department of Mathematical
Initial steps of myogenesis in somites are independent of influence from axial structures
 Development 120, 3073-3082 (1994) Printed in Great Britain The Company of Biologists Limited 1994 3073 Initial steps of myogenesis in somites are independent of influence from axial structures Eva Bober
Development 120, 3073-3082 (1994) Printed in Great Britain The Company of Biologists Limited 1994 3073 Initial steps of myogenesis in somites are independent of influence from axial structures Eva Bober
What Is an Animal? Animals come in many shapes, forms, and sizes. About 98 percent of all animals are invertebrates. The Kingdom Animalia
 What Is an Animal? What characteristics do all animals have? Animals come in many shapes, forms, and sizes. Scientists estimate that there are between 1 and 2 million species of animals! Some, like whales
What Is an Animal? What characteristics do all animals have? Animals come in many shapes, forms, and sizes. Scientists estimate that there are between 1 and 2 million species of animals! Some, like whales
(Photo Atlas: Figures 9.147, 9.148, 9.150, 9.1, 9.2, )
") BIOL 221 Concepts of Botany Fall 2007 Topic 07: Primary Plant Body: The Root System (Photo Atlas: Figures 9.147, 9.148, 9.150, 9.1, 9.2, 9.5 9.23) A. Introduction The root has the primary functions of
BIOL 221 Concepts of Botany Fall 2007 Topic 07: Primary Plant Body: The Root System (Photo Atlas: Figures 9.147, 9.148, 9.150, 9.1, 9.2, 9.5 9.23) A. Introduction The root has the primary functions of
The early development of the primary sensory neurones in an amphibian embryo: a scanning electron microscope study
 J. Embryol. exp. Morph. 75, 49-66 (1983) 49 Printed in Great Britain The Company of Biologists Limited 1983 The early development of the primary sensory neurones in an amphibian embryo: a scanning electron
J. Embryol. exp. Morph. 75, 49-66 (1983) 49 Printed in Great Britain The Company of Biologists Limited 1983 The early development of the primary sensory neurones in an amphibian embryo: a scanning electron
Morphogenesis and specification of the muscle lineage during Xenopus laevis embryo development. Armbien F. Sabillo
 Morphogenesis and specification of the muscle lineage during Xenopus laevis embryo development by Armbien F. Sabillo A dissertation submitted in partial satisfaction of the requirements for the degree
Morphogenesis and specification of the muscle lineage during Xenopus laevis embryo development by Armbien F. Sabillo A dissertation submitted in partial satisfaction of the requirements for the degree
The development of the retinotectal projection in Xenopus with one compound eye
 /. Embryol. exp. Morph. Vol 33, 3, pp. 775-787, 1975 775 Printed in Great Britain The development of the retinotectal projection in Xenopus with one compound eye By JOAN D. FELDMAN 2 AND R. M. GAZE 1 From
/. Embryol. exp. Morph. Vol 33, 3, pp. 775-787, 1975 775 Printed in Great Britain The development of the retinotectal projection in Xenopus with one compound eye By JOAN D. FELDMAN 2 AND R. M. GAZE 1 From
On the a clock' mechanism determining the time of tissue-specific enzyme development during ascidian embryogenesis
 /. Embryo!, exp. Morph. Vol. 54, pp. 131-139, 1979 Printed in Great Britain Company of Biologists Limited 1979 On the a clock' mechanism determining the time of tissue-specific enzyme development during
/. Embryo!, exp. Morph. Vol. 54, pp. 131-139, 1979 Printed in Great Britain Company of Biologists Limited 1979 On the a clock' mechanism determining the time of tissue-specific enzyme development during
Patterns of cell motility in the organizer and dorsal mesoderm of Xenopus laevis
 Development 116, 915-930 (1992) Printed in Great Britain The Company of Biologists Limited 1992 915 Patterns of cell motility in the organizer and dorsal mesoderm of Xenopus laevis JOHN SHIH* and RAY KELLER
Development 116, 915-930 (1992) Printed in Great Britain The Company of Biologists Limited 1992 915 Patterns of cell motility in the organizer and dorsal mesoderm of Xenopus laevis JOHN SHIH* and RAY KELLER
Hoxd10 induction and regionalization in the developing lumbosacral spinal
 Development 128, 2255-2268 (2001) Printed in Great Britain The Company of Biologists Limited 2001 DEV9761 2255 Hoxd10 induction and regionalization in the developing lumbosacral spinal cord Cynthia Lance-Jones
Development 128, 2255-2268 (2001) Printed in Great Britain The Company of Biologists Limited 2001 DEV9761 2255 Hoxd10 induction and regionalization in the developing lumbosacral spinal cord Cynthia Lance-Jones
BIOLOGY 340 Exam Study Guide All Exams Comparative Embryology Dr. Stuart S. Sumida California State University San Bernardino; Department of Biology
 BIOLOGY 340 Exam Study Guide All Exams Comparative Embryology Dr. Stuart S. Sumida California State University San Bernardino; Department of Biology Midterm and final exams may include materials studied
BIOLOGY 340 Exam Study Guide All Exams Comparative Embryology Dr. Stuart S. Sumida California State University San Bernardino; Department of Biology Midterm and final exams may include materials studied
BIOLOGY - CLUTCH CH.32 - OVERVIEW OF ANIMALS.
 !! www.clutchprep.com Animals are multicellular, heterotrophic eukaryotes that feed by ingesting their food Most animals are diploid, and produce gametes produced directly by meiosis Animals lack cell
!! www.clutchprep.com Animals are multicellular, heterotrophic eukaryotes that feed by ingesting their food Most animals are diploid, and produce gametes produced directly by meiosis Animals lack cell
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11589 Supplementary Figure 1 Ciona intestinalis and Petromyzon marinus neural crest expression domain comparison. Cartoon shows dorsal views of Ciona mid gastrula (left) and Petromyzon
doi:10.1038/nature11589 Supplementary Figure 1 Ciona intestinalis and Petromyzon marinus neural crest expression domain comparison. Cartoon shows dorsal views of Ciona mid gastrula (left) and Petromyzon
SODIUM-INDUCED CHANGES IN THE NUCLEI OF MONOLAYER HeLa CULTURES
 J. Cell Sci. II, 669-673 (1972) 669 Printed in Great Britain SODIUM-INDUCED CHANGES IN THE NUCLEI OF MONOLAYER HeLa CULTURES K. JOBST AND N. KELLERMAYER Department of Clinical Chemistry, Medical University
J. Cell Sci. II, 669-673 (1972) 669 Printed in Great Britain SODIUM-INDUCED CHANGES IN THE NUCLEI OF MONOLAYER HeLa CULTURES K. JOBST AND N. KELLERMAYER Department of Clinical Chemistry, Medical University
Analysis of the effects of encystment and excystment on incomplete doublets of Oxytricha fallax
 /. Embryo/, exp. Morph. Vol. 59, pp. 19-26, 1980 19 Printed in Great Britain Company of Biologists Limited 1980 Analysis of the effects of encystment and excystment on incomplete doublets of Oxytricha
/. Embryo/, exp. Morph. Vol. 59, pp. 19-26, 1980 19 Printed in Great Britain Company of Biologists Limited 1980 Analysis of the effects of encystment and excystment on incomplete doublets of Oxytricha
Langman's Medical Embryology
 Langman's Medical Embryology Developmental Biology Differentiation Morphogenesis) Epigenetic landscape (Waddington) ips Langman's Medical Embryology Morphogen gradient FGF8 in mouse limb bud Gilbert "Developmental
Langman's Medical Embryology Developmental Biology Differentiation Morphogenesis) Epigenetic landscape (Waddington) ips Langman's Medical Embryology Morphogen gradient FGF8 in mouse limb bud Gilbert "Developmental
Question Set # 4 Answer Key 7.22 Nov. 2002
 Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways
Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways