Divergent regulatory OsMADS2 functions control size, shape and differentiation of
|
|
|
- Philomena Griffith
- 5 years ago
- Views:
Transcription
1 Genetics: Published Articles Ahead of Print, published on April 3, 2007 as /genetics Divergent regulatory OsMADS2 functions control size, shape and differentiation of the highly derived rice floret second whorl organ Shri Ram Yadav *, #, Kalika Prasad *, #, 1 and Usha Vijayraghavan * * Department of Microbiology and Cell Biology Indian Institute of Science Bangalore India # These authors contributed equally to the paper. 1 Current address: Utrecht University, Padualaan CH Utrecht, Netherlands 1
2 Running title: OsMADS2 regulates rice lodicule development Keywords: Rice PI/GLO, OsMADS2, lodicule differentiation, functional diversification, transcription regulators Corresponding author: Usha Vijayraghavan Tel: Fax: ABSTRACT Functional diversification of duplicated genes can contribute to the emergence of new organ morphologies. Model eudicot plants like Arabidopsis thaliana and Antirrhinum majus have a single PI/GLO gene that together with AP3/DEF regulate petal and stamen formation. Lodicules of grass flowers are morphologically distinct reduced organs occupying the position of petals in other flowers. They serve a distinct function in partial and transient flower opening to allow stamen emergence and cross-pollination. Grasses have duplicated PI/GLO-like genes and in rice (Oryza sativa) one these genes OsMADS2, controls lodicule formation without affecting stamen development. In this study, we investigate the mechanistic roles played by OsMADS2. We ascribe a function for OsMADS2 in controlling cell division and differentiation along the proximal-distal axis. OsMADS2 is required to trigger parenchymatous and lodicule-specific vascular development while maintaining a small organ size. Our data implicate the 2
3 developmentally late spatially restricted accumulation of OsMADS2 transcripts in the differentiating lodicule to control growth of these regions. The global architecture of transcripts regulated by OsMADS2 gives insights on the regulation of cell division and vascular differentiation that can together form this highly modified grass organ with important functions in floret opening and stamen emergence independent of the paralogous gene OsMADS4. INTRODUCTION B-FUNCTION floral organ patterning activity requires a pair of related genes in model laboratory plant species. These are APETALA3 (AP3) and PISTILLATA (PI) in Arabidopsis thaliana and DEFICIENS (DEF) and GLOBOSA (GLO) in Antirrhinum majus. In these species uniform and continued expression of these genes is required for development of petals as dorsiventrally flattened sterile organs (BOWMAN et al. 1989; ZACHGO et al. 1995). Petunia has two AP3/DEF and two PI/GLO-like genes. Mutational analysis of one of the AP3/DEF-like gene: PhDEF/GREEN PETAL (GP) showed its essential role for petal development alone despite its broader expression in both petal and stamen primordia (ANGENENT et al. 1992; VAN DER KROL et al. 1993). The other gene, a paleo-ap3- like gene, PhTM6 has functionally diverged from GP and is required only for determining the stamen identity (RIJPKEMA et al. 2006). Similarly among the duplicated tomato AP3-like genes TAP3 has diverged from TM6 to acquire petal specific functions (DE MARTINO et al. 2006). Functional diversification 3
4 of duplicated orchid (Dendrobium crumenetum) AP3/DEF genes has also been shown recently by their effects upon overexpression in Arabidopsis and by studying their ability to complement ap3 mutants (XU et al., 2006). In contrast the duplicated petunia PI genes, PhGLO1 and PhGLO, have similar expression profiles and redundant activities (VANDENBUSSCHE et al. 2004) particularly with respect to petal development. Studies on B-function gene expression domains in divergent monocots have been informative in understanding floral organ homologies among angiosperms. Florets in grasses, a family of monocot plants, are reduced structures with highly derived organs. Immediately peripheral to the stamens are specialized floret organs called lodicules external to which are two modified bract-like organs- the lemma and palea, that completely enclose all internal organs (SCHMIDT and AMBROSE 1998). Lodicules of rice florets are asymmetrically positioned on the second whorl primordium. The absence of a whorl of lodicules creates the appearance that the palea and lodicules arise from a single whorl. The lodicules are small fleshy cup-shaped non-green organs with a broad base and narrow apex. Rice florets, as in other grasses, are mostly closed and open in a regulated manner for just one to two hours to allow anther emergence. This process requires rapid swelling and shrinking of the lodicules. Grass species have a single AP3/DEF ortholog but have duplicated PI/GLO-like genes (MUNSTER et al. 2001). The rice SPW1and the maize SILKY1genes are the AP3/DEF orthologs are expressed in lodicules and stamens and regulate the identity of these organs (AMBROSE et al. 2000; NAGASAWA et al. 2003). Preliminary RNA expression studies of one of these rice PI/GLO-paralogs, OsMADS2 localized its expression to lodicules and stamens (NANDI et al. 2000; KYOZUKA et al. 2000) but knockdown of its expression impaired only 4
5 lodicule development implicating the second PI/GLO paralog, OsMADS4 to suffice for stamen specification (PRASAD and VIJAYRAGHAVAN 2003). In the present study we report the dynamic expression pattern of OsMADS2 during lodicule development and its unique role in regulating lodicule differentiation and growth. We provide evidence that OsMADS2 functions independently of other rice PI/GLO paralog OsMADS4 to control second whorl cell division and differentiation by regulating genes encoding predicted transcription factors, signaling molecules and cell cycle regulators. MATERIALS AND METHODS In situ hybridisations: RNA in situ hybridization for OsMADS2 and OsMADS4 transcripts was done using gene-specific RNA probes. Probes for detection of OsMADS2 were prepared as in NANDI et al. (2000). XhoI linearized pbluescript KS (+)-OsMADS4 with the 289bp C-terminal and 3 UTR cdna sequences was transcribed with T7 RNA polymerase to make anti-sense RNAs. The same plasmid linearized with EcoRI was the template for sense-strand probe synthesis with T3 RNA polymerase. To generate gene specific probes for OsMADS2 regulated downstream genes, PCR amplified gene-specific regions for each of these genes were cloned in pbluescript KS (+). The resulting clones for AK064240/TCP, AK106784/YABBY1b and AK072687/LEUNIG when linearized with HindIII and transcribed with T7 RNA polymerase generated anti-sense RNA probes. The same plasmids linearized with BamHI were transcribed with T3 RNA polymerase to produce sense probes. The AK070518/Cyclin B clone in pbluescript linearized with BamHI was transcribed with T3 RNA polymerase to make anti-sense RNAs and the same clone digested with XhoI was transcribed with T7 RNA polymerase to produce the sense RNA 5
6 probe. Hybridizations were done as in PRASAD et al. (2005) and signal was visualized using anti-dig- AP conjugated antibodies (Roche) or anti-dig DIG rhodamine conjugated antibody (Roche, gift from Prof. K. Vijayraghavan, NCBS, Bangalore). Microscopy: For histology, florets fixed in 3% glutaraldehyde were dehydrated in a graded ethanol series and embedded in SPURR resin. 0.5µm sections (Ultramicrotome Leica, GmBH) were stained with 0.05% toleudine blue, air dried and mounted. All sections were imaged using a Zeiss Axioscope2 Mot Plus microscope (Carl Zeiss, GmBH) and images processed in Adobe photoshop version 7.0. Microarray and real-time PCR: Total RNA from rice panicles (0.5-4 cm), wild type or dsrnaiosmads2, was extracted with Tri reagent (Sigma) and purified with RNeasy cleanup kit (Qiagen). Two independent pools of mutant RNAs were compared to wild type RNA pool. For the microarray analysis, Agilent Technologies custom rice (22k) microarrays were hybridized with Cy3 and Cy5 labelled cdnas according to manufacturer instructions. The Data were analyzed using GeneSpring GX. An average ratio of mutant to wild type <0.250 for a given gene was the criteria for its differentially expression. For real-time quantitative RT-PCR analysis first strand cdna was synthesized from 1µg of total RNA and Superscript II (Invitrogen) enzyme and used in triplicate qpcr reactions using ABI Prism 7000 system. The difference in Ct value between mutant and wild type for the normalized transcript levels were used to calculate fold down regulation of the deregulated gene. The down-regulation of OsMADS2 in dsrnaiosmads2 panicles and the organ specific expression levels of selected potential OsMADS2 regulated genes were analyzed by semi-quantitative RT-PCR. The primer sequences are in supplementary Table S2. 6
7 RESULTS Dynamic temporal and spatial expression of OsMADS2 RNA during floret development: The reduced grass floret has well developed stamens but lacks obvious large petals. The predicted OsMADS2 protein bears domain organization and relatedness to other PI/GLO-like proteins. Further, like other B-function genes the stable expression of OsMADS2 transcripts is in whorls 2 and 3 primordia and in these developing organs (NANDI et al. 2000; KYOZUKA et al. 2000). We have re-examined domains in the young floret meristem where the early activation of OsMADS2 expression occurs and have carefully interrogated its expression during second and third whorl organ differentiation. This formed the basis for us to decipher the molecular mechanisms underlying the phenotypes created upon its knockdown and its global effects on gene expression. The transcription activation of OsMADS2 is to a great extent localized to the presumptive lodicule and stamen primordia on a newly established floral meristem. In these very young spikelet meristems where only the lemma primordia have been initiated we note low level OsMADS2 expression in the central region of the floral meristem that normally forms the carpel anlagen (Figure 1A). Slightly later in development upon emergence of the palea primordia this weak expression in the floret meristem center is reduced (Figure 1B). The emerging lemma and palea transiently express low levels of OsMADS2 (Figure 1B) which is eliminated slightly later in development (Figure 1C). This temporally early slightly broad domain of OsMADS2 activation was undetected in previous studies perhaps due to its low levels and transient nature (NANDI et al. 2000; KYOZUKA et al. 2000). In fact the 7
8 current analyses show the very early expression profile of OsMADS2 is similar to that of Arabidopsis PI (GOTO and MEYEROWITZ 1994). The temporally later down-regulation of OsMADS2 transcripts from the fourth whorl anlagen is coupled with its high level expression in the second and third whorl anlagen (Figure 1B). At subsequent stages where lemma are well developed hood-shaped organs but when lodicule and stamen primordia are yet to initiate, OsMADS2 expression is completely excluded from developing lemma/palea and carpel primordia (Figure 1C). In rice florets, stamen primordia initiate earlier than the lodicule primordia. OsMADS2 is expressed strongly in these early stage stamen primordia (Figure 1D). The lodicule primordia emerge only when stamen primordia begin to differentiate tetralocular anthers. At this stage high-level OsMADS2 expression is observed in the newly arising second whorl organs; with similar expression levels in developing lodicules and stamens (Figure 1E). Thus, the timing and early localization of OsMADS2 RNAs is consistent with the documented expression patterns for eudicot B-function genes supporting the analogy between petals and lodicules (AMBROSE et al. 2000, MUNSTER et al. 2001; NAGASAWA et al. 2003). However, subsequently during second whorl organogenesis expression levels of OsMADS2 deviate from those reported for PI and several of its orthologs. While expression of OsMADS2 remains high in lodicules its expression is reduced substantially in differentiating stamens (Figure 1, F-H). OsMADS2 transcripts are differentially distributed in the lodicule during its differentiation. They occur at higher levels at the distal lodicule end (Figure 1, F-H). To further examine OsMADS2 transcript distribution along the proximal-distal and the abaxial-adaxial axes we analysed its distribution in whole mount and transverse sections of differentiated lodicules. As seen in longitudinal sections, OsMADS2 expression is greatly reduced in proximal regions of the 8
9 lodicule (Figure 1, I and L) and expression is at progressively higher levels in distal regions (Figure 1I). Peripheral regions that are most distant from the palea also express high levels of OsMADS2 (Figure 1, H, I and M-O). The high level expression in distal portions of the lodicule does not vary in abaxial-adaxial axis (Figure 1, M-O and P-R). The asymmetric distribution of OsMADS2 transcripts on the proximal-distal axis suggests a likely role in the early growth arrest that must occur distally and peripherally in the emerging lodicule perhaps to regulate its size and shape. Effect of OsMADS2 knockdown on second whorl-specific cell differentiation: The second whorl organs of grass florets are small rudimentary fleshy structures due to poor dorsiventral (abaxial-adaxial) flattening coupled with reduced growth along the proximaldistal axis (Figure 2A). The knockdown of OsMADS2 results in continued growth of the distal region forming an extended bract-like structure (Figure 2B; PRASAD and VIJAYRAGHAVAN 2003). The basal or proximal portion of the transformed lodicule is also significantly larger with abnormal, severe dorsiventral flattening (compare Figure 2C vs. H). Because OsMADS2 is expressed at high levels in the presumptive lodicule anlagen even before outgrowth of these second whorl cells from the floral primordia (Figure 1B), we examined its role in triggering lodicule specific cell division and differentiation by serial ultrathin section anatomical analysis of the entire transformed lodicule in OsMADS2 knockdown transgenic spikelets (PRASAD and VIJAYRAGHAVAN 2003). Anatomical features of the wild type glume, lemma and palea are distinctive (Figure 2C and D). The rice palea has a characteristic marginal tissue which is absent in the lemma (Figure 2C). This marginal tissue of palea (mtp) differs from rest of the palea by its unique smooth upper 9
10 epidermis without epicuticular thickenings (Figure 2A) and the underlying are two to three layers of fibrous schlerenchymatous cells (Figure 2C and E). The mtp schlerenchymatous cells differ from those in the glume with regard to their cell wall thickness (compare Figure 2D vs. E). Unlike the palea the wild type lodicule contains a dense population of parenchymatous cells interspersed with vascular tracheae elements (Figure 2E and F). Another striking difference between glume, lemma/palea and lodicules is that while vascular tissues are organized as bundles in the glume (Figure 2D) and lemma/palea but in the lodicule they are dispersed throughout the organ (Figure 2F). This pattern of vascular differentiation is perhaps necessary for the uniform rapid swelling of the lodicule. We studied transverse sections through the proximal (basal) region of the lodicule that is closely juxtaposed with the marginal tissue of palea and find no identifiable border cells demarcating these two organs (Figure 2C and E). Loss of OsMADS2 causes unique lodicule cell differentiation defects notable is the transformation of cells at the proximal regions to cells typical of the immediately adjacent organ- the marginal tissue of palea (Figure 2G- J). On the other hand regions of the transformed lodicule positioned distant from marginal tissue of palea show mixture of schlerenchyma and parenchyma cells (region b in Figure 2B and K). Yet further, the extended distal portions of the transformed lodicule contain schlerenchymatous cells typical of the glume and not the mtp of the palea (Figure 2L and M). We note that in addition to these cellular transformations the lodicule basal parts are flattened comprising very few cell layers and only two to three vascular bundles. These vascular bundles are similar to that of the glume, lemma or palea (Figure 2I-M). Additionally, the continued growth of the lodicule margins is consistently seen in multiple transgenic lines due to which the normal cup-shaped fleshy lodicule structure is always lost ( 10
11 compare Figure 2C vs. H). Thus, the normal differentiation program of wild type lodicule is entirely disrupted in these transformed lodicules with the cells committing to different fates along the proximal-distal axis and we obtain cleistogamous rice lines where spikelets remain unopened. The extent of down-regulation of the endogenous OsMADS2 transcripts in dsrnaiosmads2 panicles was determined by semi-quantitative RT-PCR (Figure 2N). Transcripts levels are extremely low and are barely detected after 45 cycles of PCR amplification indicating near null phenotypes are generated in these transgenic lines. Together, we reason that the likely role for the early expression of OsMADS2 in lodicule primordia is to delineate the boundary between palea and lodicule at its proximal end and to trigger second whorl-specific cell differentiation throughout the lodicule (Figure 2O and P). The temporally later expression of OsMADS2 in specific regions of the lodicule perhaps serves to initiate growth arrest distally and towards the lodicule periphery (Figure 2O and P). Timing and localization of transcripts for the second rice PI/GLO paralog- OsMADS4: OsMADS4 together with the sole rice AP3 homolog SUPERWOMAN (SPW1) specifies lodicules and stamens (KANG et al. 1998; NAGASAWA et al. 2003; LEE et al. 2003b) as indicated from the phenotypic similarities of spw1 flowers and florets with OsMADS4 down regulation. To interrogate OsMADS4 expression pattern we have monitored the spatial and temporal distribution of its transcripts by RNA in situ hybridization. We find OsMADS4 transcription activation occurs very early, and uniformly, during spikelet meristem initiation prior to the emergence of even the first floret organ primordia i.e., the lemma primordia (Figure 3A). The lemma and palea primordia express OsMADS4 from their outgrowth with continuing expression in the 11
12 anlagen for the second, third and fourth whorl organs (Figure 3B). Since these transcripts are excluded from glumes OsMADS4 expression is specific to floret organs (Figure 3B). Thus, the timing and domain of OsMADS4 transcription activation deviates from that of OsMADS2 from very early stages of floral meristem specification. During floret organogenesis high-levels of OsMADS4 expression occur in stamen and carpel whorls (Figure 3C and D) with reduced expression in the differentiating lodicules (Figure 3E and F) and are completely excluded from lemma and palea (Figure 3D-F). Importantly from this stage onwards despite reduced OsMADS4 expression the distribution in the lodicule is uniform (Figure 3E and F) and no expression is detectable in the differentiated lemma or palea (Figure 3D-F). Unlike OsMADS2, strong expression of OsMADS4 is maintained differentiating in fourth whorl primordia (Figure 3D and E). These data show that all the three rice class B-genes share a common conserved expression domain (whorls 2 and 3) as is typical of many B-function genes. But they differ in the onset of their expression in the floret meristem, and in their transcripts levels and distribution within whorls two and three. These rice paralogous genes also differ in their expression patterns in other floret whorls. Because knockdown of OsMADS2 completely perturbs lodicule differentiation we examined if there are any changes in the expression profile of OsMADS4 in the transformed lodicules of Ubi-dsRNAiOsMADS2 transgenic florets (PRASAD and VIJAYRAGHAVAN 2003). RNA in situ hybridization shows OsMADS4 expression is retained in second whorl of these knockdown florets in addition to its normal expression in whorl 3 and whorl 4 (Figure 3G-I). In fact, OsMADS4 transcripts are slightly upregulated in overgrown distal regions of transformed lodicules (Figure 3H). These 12
13 data suggest that whorl-specific expression of OsMADS4 is retained in transgenic spikelet and the abundant OsMADS4 transcripts cannot rescue the second whorl-specific defects caused by the loss of OsMADS2. Potential targets for OsMADS2 during lodicule differentiation: To gain mechanistic insights on OsMADS2 functions we have employed global expression profiling to identify downstream genes regulated by OsMADS2. RNAs from two independent biological pools of dsrnaiosm2 transgenic inflorescences, at various stages of organ specification and differentiation, were compared to two wild type RNA pools. Subsequently these transgenic RNA pools were used in two competitive hybridizations with a wild type RNA pool on rice microarrays (MATERIALS and METHODS). A third hybridization was performed with reciprocally dye-labeled wild type RNA and one of the mutant RNAs. We first identified transcripts down regulated in both the mutant RNA pools and then further screened for transcripts with >2 fold down regulation even after dye-swap labeling of the wild type and mutant RNAs. These transcripts were manually inspected for their predicted or known expression profiles in available databases (KOME i.e., and our unpublished data) to arrive at a list of 385 affected genes whose expression was independently documented in developing wild type florets. Based on the occurrence of protein domains we ascribed functional categories to these differentially expressed genes. We find a preponderance of transcription factors and signaling molecules (10% and 14%) amongst the transcripts affected by the loss of OsMADS2 (Figure 4A). In addition a large set of genes (9%) functioning in diverse aspects of cell division control; including cyclins, 13
14 histones and replication factors are deregulated (Figure 4A; supplemental Table S1). These observations are interesting in light of the global expression profiling data from Arabidopsis pi and ap3 mutants where the affected downstream genes do not include such a large proportion of cell division genes (ZIK and IRISH 2003; WELLMER et al. 2004). On the other hand about 8% of target genes of Antirrhinum DEF are factors affecting cell cycle and DNA processing (BEY et al. 2004). From the subset of potential OsMADS2 regulated genes we chose 10 genes for validation of their down-regulation in absence of OsMADS2. These genes (Table 1) include factors with predicted functions in signaling and cell proliferation (GDSL lipase, F-box factor, Cyclin B) and transcriptional regulators (YABBY domain factors, TCP family of regulators, LEUNIG). Their expression levels, in wild type panicle RNAs and in transgenic dsrnaiosmads2 panicles, were determined by quantitative real time PCR (qrt-pcr). The fold down regulation seen in qrt-pcr was compared with the data from microarray analysis (Figure 4B). Expression of all but one of these genes was dependent on OsMADS2 as seen by both assays. The expression levels of these genes in various wild type floret organs was examined by semi-quantitative RT-PCR specifically in the palea, lodicule, stamens and carpels. From comparison of the normalized expression level we find that all of these genes are expressed in wild type lodicules (Figure 4C) and are expressed at comparable levels in carpels. The expression levels in stamens and palea are somewhat lower. The rice drooping leaf-superman (dl-sup) mutant forms ectopic stamens at the expense of carpel tissues, while lodicules are unaffected (NAGASAWA et al. 2003). We observe the expression domain and localization of OsMADS2 transcripts to specific 14
15 regions of the lodicule in dl-sup flowers remains unaffected (supplemental Figure 1A-C) corroborating the dynamic regulation of OsMADS2 in lodicules of mutant plants of other genotypes. Similarly expression of SPW1 is normal in dl-sup lodicules (NAGASAWA et al. 2003) suggesting that OsMADS2 and SPW1 may suffice to trigger lodicule differentiation. Since lodicule morphology and OsMADS2 expression domains within the lodicule is normal in dl-sup flowers we used this rice mutant, as a control, to verify the expression levels of genes regulated by OsMADS2. We find that the transcription factors AK106784/Yabby1b, AK071176/ZF-SET, AK064240/TCP and the signaling or cell division factors such as AK070518/Cyclin B, AK099104/F-box factor and AK100480/GDSL lipase are expressed at normal or near normal levels in differentiated florets of dl-sup panicles (supplemental Figure 1D). Together these data show these genes to be regulated by OsMADS2. As a first approximation in identifying genes that might be direct targets of OsMADS2 we inspected genomic sequences upto 1kb upstream of all 385 deregulated genes for the presence of cis-acting CArG elements that are binding sites for MADS domain proteins (supplemental Table S1). We find that 47% these genes have CArG elements in this segment of their genomic loci and notably of these about 37% genes have multiple such elements (supplemental Table S1). Spatial transcript distribution for genes regulated by OsMADS2 in developing florets: Four of the genes regulated by OsMADS2 - encoding three transcription regulators and one a cell division regulator were further examined by RNA in situ hybridization to determine their expression domains in developing floral organs. Transcripts for all of these genes are detected in the lodicule right from its emergence, 15
16 expression continues during its development and persists to varying extents in the fully developed lodicules (Figure 6). This profile is in general similar to that of OsMADS2 (Figure 1E-H). However, not unexpectedly, the expression of these genes in other floral organs differs from that of OsMADS2. For example their expression in the fourth whorl is maintained during carpel development as is expression in the mature stamens (compare Figure 6 and Figure 1). These genes also differ from each other in their relative expression levels in stamens, carpels and mature lodicules. The AK064240/TCP and AK070518/cyclin gene transcripts in the lemma, palea and emerging stamens of young florets are at equal levels. The expression in the carpel primordia is somewhat lower (Figure 5A and 5D). While the expression in the lemma and palea is shut down later during their differentiation, high level expression is seen in differentiating stamens particularly in anthers (Figure 5B, C, E and F). Expression levels in the lodicules are maintained uniformly throughout its development (Figure 5B, C, E and F). The transcripts for AK106784/Yabby1b and AK072687/LEUNIG appear to be temporally regulated in differentiating floral organs. They occur at higher abundance early during lodicule development and are at lower levels later during lodicule development (Figure 5H, I, K and L). The expression of both these genes in stamens is maintained throughout stamen differentiation and in later stages is particularly high in the anthers (Figure 5G-L). The recently reported expression levels of YABBY1b (TORIBA et al. 2007), as determined by quantitative RT-PCR, are consistent with our RNA in situ hybridization data (Figure 5G-I) of developing flowers. 16
17 DISCUSSION Developmental innovations by gene duplication: Duplication and diversification of evolutionarily conserved transcription factors can bring about morphological variation between closely related organisms. Mechanistically such functional differences between duplicated genes could arise from changes in the transcription factors themselves, or from their altered expression, or from changes in how they regulate their target genes (reviewed in IRISH and LITT 2005). We demonstrate that of the two rice PI-like genes OsMADS2 controls development of the rice lodicule- a floret organ critical for regulated opening of the rice floret and cross-pollination. Its unique regulatory role is evident from the specific effects, obtained upon OsMADS2 knockdown, on the fate of lodicule cells along proximal vs. distal axis. These distinct functions are not seen on down regulation of its paralog OsMADS4 (KANG et al. 1998). The functional diversification of OsMADS4 with respect to second whorl organ development has been shown using a heterologous plant i.e., tobacco as a model (KANG and AN 2005). Unlike the effect of Antirrhinum GLO over-expression the ectopic over-expression of OsMADS4 in tobacco did not cause any alteration to first whorl organs (KANG and AN 2005). Together with our evidence of the inability of OsMADS4 expression in second whorl organs of dsrnaiosmads2 florets to rescue second whorl lodicule formation we can conclude that OsMADS4 alone cannot confer the unique second whorl organ fate in rice florets. Such differential effects for paralogous factors may arise from alterations in the conserved MADS domain or from changes in the functionally important PI- motif (supplemental Figure 2; LAMB and IRISH 2003) or even from changes in the profile within the conserved B-function expression domains. Interestingly, the early 17
18 transcriptional activation of OsMADS2 in whorls 2 and 3 primordia is a pattern similar to that of many eudicot PI genes but the later sub-domains of OsMADS2 expression differ from that known for Arabidopsis PI and Antirrhinum GLO (GOTO and MEYEROWITZ 1994; TROBNER et al. 1992). In whorl 2 organs high level OsMADS2 transcripts are predominantly localized towards their distal end. We ascribe a function for this expression in regulating growth as this region overgrows upon OsMADS2 knockdown. Recent analysis of several Arabidopsis gene-trap lines reveals that these petals too have underlying differences in gene expression which is reflected in reporter profiles along the proximal/distal axis (NAKAYAMA et al. 2005). However, these molecular signatures are distinct from expression pattern of Arabidopsis B-class genes or their regulators. The restricted late expression of OsMADS2 to specific regions of the lodicule bears similarity to the expression of some lower eudicot PI-like genes of the Ranunculid family where expression is limited to petal edges (KRAMER and IRISH 1999). Such localized expression may serve specific functions in petal development in these species. Expression pattern of OsMADS2 is distinct not only from higher eudicot B class genes but it also diverges from its paralog OsMADS4 in local regions within the second and third whorl. High level OsMADS2 expression is restricted to specific regions of lodicules and expression is generally down-regulated in stamens. In contrast, OsMADS4 expression is down-regulated uniformly in all lodicules cells and its strong uniform expression is maintained in stamens supporting its critical role in defining the stamen identity (KANG et al. 1998; LEE et al. 2003b). Additionally OsMADS4 acquires expression in the fourth whorl a pattern not seen for OsMADS2. These observations also explain the need for repression of SPW1 in fourth whorl 18
19 (NAGASAWA et al. 2003). We deduce that the continued fourth whorl expression of OsMADS4 throughout floret development necessitates the repression of its interacting partner SPW1 to prevent the formation of ectopic fourth whorl stamens (LEE et al. 2003b). Changes in SPW1 affect OsMADS4 transcripts reciprocally without any changes in OsMADS2 transcript levels (XIAO et al. 2003). Therefore rice B-function activity provided by OsMADS4/SPW1 heterodimer may auto regulate its expression as has been reported for the Arabidopsis PI/AP3 and Antirrhinum GLO/DEF heterodimers (TROBNER et al. 1992; KRIZEK and MEYEROWITZ 1996). All of these data suggest divergence in the regulation of the two rice PI/GLO paralogs. Triggering differentiation of the lodicule- a highly derived grass-specific second whorl organ: One of the most intriguing effects of OsMADS2 knockdown is the perturbation of lodicule cell differentiation in a context dependent fashion. In these knockdown florets lodicule cells that are in close proximity to the marginal tissue of palea (mtp) acquire cellular features of the mtp. On the other hand the proximal tissues of the transformed lodicule that are at a distance from mtp acquire a mixed fate of parenchyma and schlerenchyma cells. Further still the distal portions of the transformed lodicule overgrow but acquire exclusively schlerenchymatous cell fate (Figure 2O). This suggests direct cell-cell communication in regions of the floret where the distinctive cells of the mtp and lodicules are juxtaposed. We speculate that the primary role of OsMADS2 in this proximal region is to delineate the boundary between mtp and lodicule, perhaps an early event during commitment of floret meristem cells to the lodicule fate. Our data also show OsMADS2 expression to promote second whorl specific cell differentiation throughout the lodicule as the transformed organs have dramatic alterations in vascular 19
20 differentiation. Our report represents a novel example of a class B gene exerting contextdependent local effects on growth and differentiation. Our studies provide possible explanations for the conversion of lodicules to mtp and not a fully differentiated lemma or palea in the spw1 mutant (NAGASAWA et al. 2003). Loss- of B-function mutants in Arabidopsis and Antirrhinum have petals transformed to the adjacent peripheral organ i.e., sepals. In the rice floret the mtp is placed adjacent to whorl 2 organs; therefore we may interpret the transformation of lodicules to mtp seen in rice B-function mutants as being similar to the phenotypic transformations of Arabidopsis mutants. These homeotic organ transformations support the notion that lodicules are petal homologs as suggested from studies in maize (AMBROSE et al. 2000; WHIPPLE et al. 2004). Targets genes regulated by OsMADS2 encode putative cell proliferation, differentiation and signaling factors: OsMADS2 controls the shape and fleshy characteristics of the second whorl floret organ by regulating cell proliferation and by maintaining a unique differentiation pattern. Several genes deregulated upon knockdown of OsMADS2 are those predicted to regulate cellular growth and differentiation and ultimately organ shape. The transcription activation of these genes must be independent of OsMADS4/SPW1 heterodimer as the normal whorl-specific expression of OsMADS4 does not rescue the phenotype of OsMADS2 knockdown. OsMADS2 regulated genes with potential growth regulatory functions as well as those needed for the development of highly vascularized parenchymatous cells are particularly interesting. Both these features distinguish the lodicule from other sterile whorl organs. Of the 49 rice cyclins identified in rice (LA et al. 2006) we find six cyclins are deregulated in absence of OsMADS2. 20
21 Three of these are B-type cyclins, two are A-type cyclin and another is a D-type cyclin (Table 1; supplemental Table S1). We have validated the down regulation of AK (OsCycB2-2) upon loss of OsMADS2. Intriguingly there are three CArG elements, potential binding site for MADS domain proteins, positioned within 1kb sequences upstream to the predicted OsCycB2-2 transcription start site. Further its spatial expression domain overlaps with OsMADS2. These observations make it one of the candidate genes that may be directly regulated by OsMADS2- investigations on this line are ongoing. Importantly independent studies have shown this cyclin is nuclear localized and its over expression causes increased root growth most likely by promoting cell division (LEE et al. 2003a). We speculate that as a potential target of OsMADS2 perhaps OsCycB2-2 may promote proliferation of parenchymatous cells in the lodicule. The importance of regulated cell proliferation during lodicule development is further reinforced by our discovery of two members of the TCP family of transcription regulators deregulated in absence of OsMADS2 a characteristic not seen among targets of AP3 or PI (WELLMER et al. 2004; ZIK and IRISH, 2003). Among 25 predicted rice TCP factors ( we find AK an uncharacterized Class I factor similar to PCF1 and another PCF7 (AK058570) -a Class II factor are deregulated on OsMADS2 knockdown. The AK064240/TCP is expressed from lodicule emergence with expression continuing during its differentiation. Further sequences upsteam to the predicted transcription start site have one CArG element. The Class II TCP family includes the well studied the Antirrhinum CIN and CYC proteins, the maize TB1 and the rice OsTB1 proteins (CRAWFORD et al. 2004; LUO et al. 1999; DOEBLEY et al. 1997; TAKEDA et al. 2003). Functional studies on CIN and CYC show them to regulate 21
22 growth of lateral organs (NATH et al. 2003; CRAWFORD et al. 2004; COSTA et al. 2005). Similar functions are seen for the Lotus japonicus TCP-box gene LjCYC2 that regulates asymmetric growth of petals and maintains its shape (FENG et al. 2006). The first mechanistic link between TCP factors and cell proliferation came from in vitro studies showing rice PCF1 and PCF2 to bind cis elements in the PCNA gene a key protein of DNA replication complex (KOSUGI and OHASHI 1997, 2002). Another link identified in a recent study shows Arabidopsis TCP20 can bind elements in the promoter of CycB1 in addition to promoters of few of the ribosomal protein genes (LI et al. 2005). Notably, an opposite effect of repression of cell proliferation by a Class I TCP domain factor is exemplified by the interaction between Antirrhinum TIC and CUPULIFORMIS - a NAC-domain protein (WEIR et al. 2004). This study provides an attractive hypothesis that localized expression of TCP and NAC domain transcription factors can repress of cell division to establish boundaries between lateral organs (WEIR et al. 2004) a scenario which may apply in lodicule development as four NAC domain containing factors are deregulated in absence of OsMADS2 (data not shown). Thus multiple alternative mechanisms operate to link this class of transcription regulators with cell proliferation. Together these lines of evidence provide hypotheses to test the impact of OsMADS2 in regulating cell division. Our global transcript profiling shows how OsMADS2 can trigger lodicule-specific differentiation. The Arabidopsis transcription co-regulators SUESS and LEUNIG control petal blade cell number and petal internal vasculature (FRANKS et al. 2006). Similar functions for the Antirrhinum homologues of LUG and SEU i.e., STY and AmSEU1, AmSEU2 and AmSEU3 in organ initiation, laminar growth and venation have been found 22
23 (NAVARRO et al. 2004). STY has also been shown to physically interact with these Antirrhinum SEU-like proteins as well as the Antirrhinum FIL orthologue, GRAMINIFOLIA (GRAM). While SEU and LUG are transcription regulators DNA binding activity is likely provided by their partner- FIL1/YAB1 (NAVARRO et al. 2004). In another study, SEU has been shown to mediate interaction of LUG with MADS box proteins AP1 and SEP3 and thereby acquire DNA binding specificity (SRIDHAR et al. 2006). LUG-SEU has also been shown to acquire DNA binding specificity from MADS box dimers AP1-SVP and AP1-AGL24 (GREGIS et al. 2006) It is striking that the closest rice homologues for these factors (AK OsLEU, AK OsYAB1a, AK OsYAB1b and AK a rice MADS gene) are deregulated in the absence of OsMADS2 hinting at a role for OsMADS2 in regulating vascular development through the actions of a homologous complex. Both, the OsLEU and the OsYAB1b are expressed during lodicule development. While none of the above mentioned rice factors are by themselves functionally characterized, it is noteworthy that a related YABBY domain factor DL-SUP is required for leaf mid-vein development (YAMAGUCHI et al. 2004) and carpel development. DL-SUP is not expressed in lodicules but the YABBY genes identified here are expressed in both lodicules and carpels. Functional studies of these rice genes including understanding their direct vs. indirect regulation by OsMADS2 are required to elucidate their specific roles in lodicule development. Our preliminary comparative survey of genes deregulated on knockdown of OsMADS2, potential targets of Arabidopsis AP3 and PI, or potential targets of Antirrhinum DEF (ZIK and IRISH 2003; BEY et al. 2004; WELLMER et al. 2004) reveal some commonalities and some important differences in the global profile of 23
24 affected genes. Many of commonly affected genes in these diverse species are expressed late in petal differentiation, such as genes for lipases, hydrolases, fatty acid elongation and lipid transfer proteins. Another similarity in the global profile of genes regulated by B function genes is the occurrence of predicted receptor-like kinases and signaling factors. It is noteworthy that a sizable fraction of the genes affected on loss of OsMADS2 are those with functions in cell division, a global profile not seen in targets of PI (WELLMER et al. 2004; ZIK and IRISH 2003). These studies suggest that while some target genes of higher eudicot and grass class B genes many encode molecules with similar functions many downstream genes of OsMADS2 are distinct from those thus far identified for PI or DEF. This likely reveals the differences in the cascade of genes regulated by B-function activity in these diverse species that have distinct petal morphologies and where petals serve different functions. ACKNOWLEDGEMENTS UVR acknowledges grant support from Department of Biotechnology, Government of India for rice functional genomics and expression profiling. Infrastructure support from ICMR, Government of India to Department of MCB is thanked. We are grateful to Prof. Y. Nagato for providing dl-sup seeds. A Senior Research Fellowship from CSIR, Government of India, provided support to SRY. LITERATURE CITED AMBROSE, B. A., D. R. LERNER, P. CICERI, C. M. PADILLA, M. F. YANOFSKY, R. J. SCHMIDT, 2000 Molecular and genetic analyses of the Silky1 gene reveal conservation in floral organ specification between eudicots and monocots. Mol. Cell 5:
25 ANGENENT, C. C., M. BUSSCHER, J. FRANKEN, J. N. MOL, A. J. VAN TUNEN, 1992 Differential expression of two MADS box genes in wild-type and mutant petunia flowers. Plant Cell 4: BEY, M., K., K. STUBER, Z. FELLENBERG, H. SCHWARZ-SOMMER, H. SOMMER, H. SAEDLER, S. ZACHGO, 2004 Characterization of Antirrhinum petal development and identification of target genes of the class B MADS box gene DEFICIENS. Plant Cell 16: BOWMAN, J. L., D. R. SMYTH, E. M. MEYEROWITZ, 1989 Genes directing flower development in Arabidopsis. Plant Cell 1: COSTA, M. M., S. FOX, A. I. HANNA, C. BAXTER, E. COEN, 2005 Evolution of regulatory interactions controlling floral asymmetry. Development 132: CRAWFORD, B. C., U. NATH, R. CARPENETER, E. S. COEN, 2004 CINCINNATA controls both cell differentiation and growth in petal lobes and leaves of Antirrhinum. Plant Physiol. 135: DE MARTINO G., I. PAN, E. EMMANUEL, A. LEVY, V. F. IRISH, 2006 Functional Analyses of Two Tomato APETALA3 Genes Demonstrate Diversification in Their Roles in Regulating Floral Development. Plant Cell 18:
26 DOEBLEY, J., A. STEC, L. HUBBARD, 1997 The evolution of apical dominance in maize. Nature 386: FENG, X., Z. ZHAO, Z. TIAN, S. XU, Y. LUO, Z. CAI, Y. WANG, J. YANG, Z. WANG, L. WENG, J. CHEN, L. ZHENG, X. GUO, J. LUO, S. SATO, S. TABATA, W. MA, X. CAO, X. HU, C. SUN, D. LUO, 2006 Control of petal shape and floral zygomorphy in Lotus japonicus. Proc Natl Acad Sci U S A. 103: FRANKS, R. G., Z. LIU, R. L. FISCHER, 2006 SEUSS and LEUNIG regulate cell proliferation, vascular development and organ polarity in Arabidopsis petals. Planta DOI: /s GOTO, K., E. M. MEYEROWITZ, 1994 Function and regulation of the Arabidopsis floral homeotic gene PISTILLATA. Genes Dev. 8: GREGIS, V., A. SESSA, L. COLOMBO, M. M. KATER, 2006 AGL24, SHORT VEGETATIVE PHASE and APETALA1 redundantly control AGAMOUS during early stages of flower development in Arabidopsis. Plant Cell 18: IRISH, V. F., A. LITT, 2005 Flower development and evolution: gene duplication, diversification and redeployment. Curr. Opin. Genetics and Dev. 15:
27 KANG, H. G., G. AN, 2005 Morphological alterations by ectopic expression of the rice OsMADS4 gene in tobacco plants. Plant Cell Rep. 24: KANG, H. G., J. S. JEON, S. LEE, G. AN, 1998 Identification of class B and class C floral organ identity genes from rice plants. Plant. Mol. Biol. 38: KOSUGI, S., Y. OHASHI, 1997 PCF1 and PCF2 specifically bind to cis elements in the rice proliferating cell nuclear antigen gene. Plant Cell 9: KOSUGI, S., Y. OHASHI, 2002 DNA binding and dimerization specificity and potential targets for the TCP protein family. Plant J. 30: KRAMER, E. M., V. F. IRISH, 1999 Evolution of genetic mechanisms controlling petal development. Nature 399: KRIZEK, B. A., E. M. MEYEROWITZ, 1996 The Arabidopsis homeotic genes APETALA3 and PISTILLATA are sufficient to provide the B class organ identity function. Development 122: KYOZUKA, J., T. KOBAYASHI, M. MORITA, K. SHIMAMOTO, 2000 Spatially and temporally regulated expression of rice MADS box genes with similarity to Arabidopsis class A, B and C genes. Plant Cell Physiol. 41:
28 LA, H., J. LI, Z. JI, Y. CHENG, X. LI, S. JIANG, P. N. VENTAKESH, S. RAMACHANDRAN 2006 Genome-wide analysis of cyclin family in rice (Oryza sativa L.). Mol Genet Genomics 275, LAMB, R. S., V. F. IRISH, 2003 Functional divergence within the APETALA3/PISTILLATA floral homeotic gene lineages. Proc. Natl. Acad. Sci. USA 100: LEE, J., A. DAS, M. YAMAGUCHI, J. HASHIMOTO, N. TSUTSUMI, H. UCHIMIYA, M. UMEDA, 2003a Cell cycle function of a rice B2-type cyclin interacting with a B-type cyclin-dependent kinase. Plant J. 34: LEE, S., J. S. JEON, K. AN, Y. H. MOON, S. LEE, Y. Y. CHUNG, G. AN, 2003b Alteration of floral organ identity in rice through ectopic expression of OsMADS16. Planta. 217: LI, C., T. POTUSCHAK, A. COLON-CARMONA, R. A., GUTIERREZ, P. DOERNER, 2005 Arabidopsis TCP20 links regulation of growth and cell division control pathways. Proc Natl Acad Sci U S A. 102: LUO, D., R. CARPENTER, L. COPSEY, C. VINCENT, J. CLARK, E. COEN, 1999 Control of organ asymmetry in flowers of Antirrhinum. Cell 99:
29 MUNSTER, T., L. U. WINGEN, W. FAIGL, S. WERTH, H. SAEDLER, G. THEISSEN, 2001 Characterization of three GLOBOSA-like MADS-box genes from maize: evidence for ancient paralogy in one class of floral homeotic B-function genes of grasses. Gene 262: NAGASAWA, N., M. MIYOSHI, Y. SANO, H. SATOH, H. HIRANO, H. SAKAI, Y. NAGATO, 2003 SUPERWOMAN1 and DROOPING LEAF genes control floral organ identity in rice. Development 130: NAKAYAMA, N., J. M. ARROYO, J. SIMOROWSKI, B. MAY, R. MARTIENSSEN, V. F. IRISH, 2005 Gene trap lines define domains of gene regulation in Arabidopsis petals and stamens. Plant Cell 17: NANDI, A. K., K. KUSHALAPPA, K. PRASAD, U. VIJAYRAGHAVAN, 2000 A conserved function for Arabidopsis SUPERMAN in regulating floral-whorl cell proliferation in rice, a monocotyledonous plant. Curr. Biol. 10: NATH, U., B. C. CRAWFORD, R. CARPENTER, E. COEN, 2003 Genetic control of surface curvature. Science 299: NAVARRO, C., EFREMOVA, N., H. F. GOLZ, R. RUBIERA, M. KUCKENBERG, R. CASTILLO, O. TIETZ, H. SAEDLER, Z. SCHWARZ-SOMMER, 2004 Molecular and genetic interactions between STYLOSA and GRAMINIFOLIA in the control of Antirrhinum 29
30 vegetative and reproductive development. Development 131: PRASAD, K., S. PARAMESWARAN, U. VIJAYRAGHAVAN, 2005 OsMADS1, a rice MADS-box factor, controls differentiation of specific cell types in the lemma and palea and is an early-acting regulator of inner floral organs. Plant J. 43: PRASAD, K., U. VIJAYRAGHAVAN, 2003 Double-stranded RNA interference of a rice PI/GLO paralog, OsMADS2, uncovers its second-whorl-specific function in floral organ patterning.genetics 165: RIJPKEMA, A. S., S. ROYAERT, J. ZETHOF, G. VAN DER WEERDEN, T. GERATS, M. VANDENBUSSCHE, 2006 Analysis of the Petunia TM6 MADS Box Gene Reveals Functional Divergence within the DEF/AP3 Lineage. Plant Cell 18: SCHMIDT, R., B. A. AMBROSE, 1998 The blooming of grass flower development. Curr. Opin. in Plant Biol. 1: SRIDHAR, V. V., A. SURENDRARAO, Z. LIU, 2006 APETALA1 and SEPALLATA3 interact with SEUSS to mediate transcription repression during flower development. Development 133: TAKEDA, T., Y. SUWA, M. SUZUKI, H. KITANO, M. UEGUCHI-TANAKA, M. ASHIKARI, M. MATSUOKA, C. UEGUCHI, 2003 The OsTB1 gene negatively regulates 30
31 lateral branching in rice. Plant J. 33, TORIBA, T, K. HARADA, A. TAKAMURA, H. NAKAMURA, H. ICHIKAWA, T. SUZAKI, H. Y. HIRANO, 2007 Molecular characterization of YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Mol. Genet. Genomics DOI: /s TROBNER, W., L. RAMIREZ, P. MOTTE, I. HUE, P. HUIJSER, W. E. LONNIG, H. SAEDLER, H. SOMMER, Z. SCHWARZ-SOMMER, 1992 GLOBOSA: a homeotic gene which interacts with DEFICIENS in the control of Antirrhinum floral organogenesis. EMBO J. 11, VANDENBUSSCHE, M., ZETHOF, J., S. ROYAERT, K. WETERINGS, T. GERATS, 2004 The duplicated B-class heterodimer model: whorl-specific effects and complex genetic interactions in Petunia hybrida flower development. Plant Cell 16: VEN DER KROL, A. R., A. BRUNELLE, S. TSUCHIMOTO, N. H. CHUA, 1993 Functional analysis of Petunia floral homeotic MADS box gene pmads1. Genes Dev. 7: WEIR, I., J. LU, H. COOK, B. CAUSIER, Z. SCHWARZ-SOMMER, B. DAVIES, 2004 CUPULIFORMIS establishes lateral organ boundaries in Antirrhinum. Development 131,
32 WELLMER, F., J. L. RIECHMANN, M. ALVES-FERREIRA, E. M. MEYEROWITZ, 2004 Genome-wide analysis of spatial gene expression in Arabidopsis flowers. Plant Cell 16: WHIPPLE, C. J., P. CICERO, C. M. PADILLA, B. A. AMBROSE, S. L. BANDONG, R. J. SCHMIDT, 2004 Conservation of B-class floral homeotic gene function between maize and Arabidopsis. Development 131: XIAO, H., Y. WANG, D. LIU, W. WANG, X. LI, X. ZHAO, J. XU, W. ZHAI, L. ZHU, 2003 Functional analysis of the rice AP3 homologue OsMADS16 by RNA interference. Plant Mol Biol. 52: XU, Y., L. L. TEO, P. P. KUMAR, H. YU, 2006 Floral organ identity genes in orchid Dendrobium crumenatum. Plant J. 46: YAMAGUCHI, T., N. NAGASAWA, S. KAWASAKI, M. MATSUOKA, Y. NAGATO, H. Y. HIRANO, 2004 The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 16: ZACHGO, S., A. SILVA EDE, P. MOTTE, W. TROBNER, H. SAEDLER, Z. SCHWARZ- SOMMER, Functional analysis of the Antirrhinum floral homeotic DEFICIENS gene in vivo and in vitro by using a temperature-sensitive mutant. Development 121,
33 ZIK, M., V. F. IRISH, 2003 Global identification of target genes regulated by APETALA3 and PISTILLATA floral homeotic gene action. Plant Cell 15:
34 TABLE 1. Ten selected genes down-regulated in OsMADS2 knockdown floret Accession Number Annotation/Description Closest Arabidopsis hits AK Axial regulator YABBY1 At2g45190 AK Axial regulator YABBY1 At2g45190 AK GDSL-motif lipase At1g09390 AK cyclin, similar to B-like cyclin At1g76310 AK Cellulose, identical to ATCEL2 At1g02800 AK Transcriptional corepressor At4g32551 LEUNIG AK SET-domain transcription At2g17900 regulator AK F-box family protein At1g53320 AK ZF-HD homeobox family protein At4g24660 AK TCP family transcription factor At1g
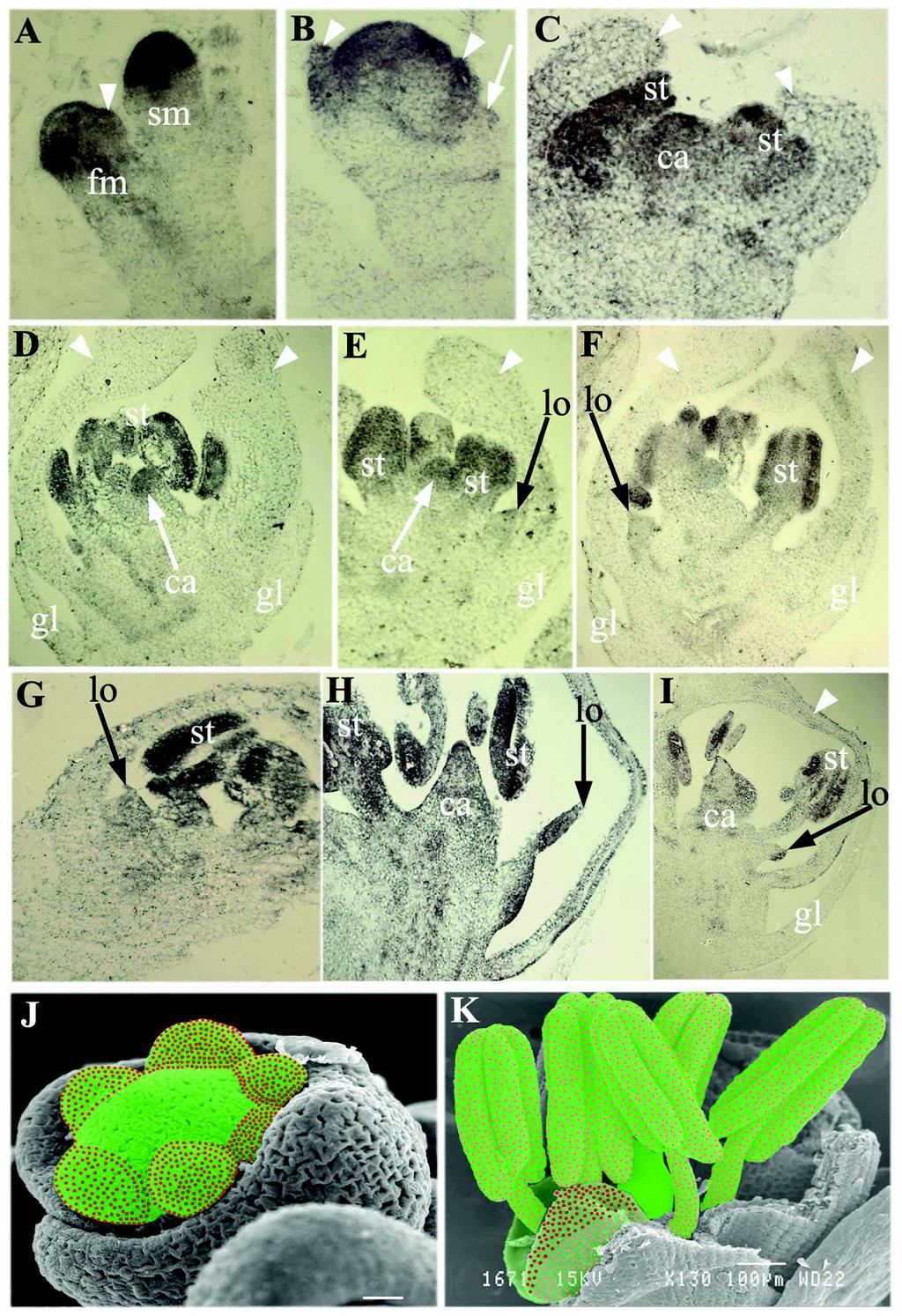
35 FIGURE LEGENDS FIGURE 1. Expression pattern of OsMADS2. (A-H) Longitudinal sections through developing rice spikelets probed with digoxigenin-utp antisense RNAs detected with APconjugated anti-dig antibodies. (A) An incipient floret meristem, black arrowheads mark the lodicule and stamen anlagen and black arrow points to the central floret cells all of which express OsMADS2. (B) Young florets with strong expression in lodicule and stamen anlagen and very low expression in lemma/palea primordia. (C) Developing young florets with emerging lemma/palea. No expression was seen in developing glumes, lemma/palea and the floral center. (D and E) Equal expression in developing stamen (st) and lodicule (lo) primordia. (F and H) Florets during organogenesis show differential OsMADS2 expression. Black arrowheads point to distal and peripheral regions of a differentiated lodicule (lo). Inset in (F) is a magnified lodicule. (I-O) RNA in situ hybridization of whole mount (I-J) and transverse sections (L-O) of mature lodicules detected with anti-dig-rhodamine conjugated antibodies. The section planes are indicated in (I). Probe in (I, L-O) is antisense and in (J) is sense OsMADS2 RNA. (K) Lodicule bright field image. White arrowheads in (M) and (N) mark the lodicule peripheral regions le; lemma, mtp; marginal tissue of palea. (P-R) Longitudinal Optical sections shown are arranged abaxially or adaxially with respect the center of the lodicule. The right (abaxial) and the left sections (adaxial) are 24µm apart. In both these sections accumulation of OsMADS2 transcripts is in distal regions and in peripheral regions that grow away from palea. le; lemma, mtp; marginal tissue of palea. 35
36 FIGURE 2. Ultrathin transverse sections show cell proliferation and differentiation defects in OsMADS2 knockdown florets. (A) Partially dissected wild type. B, OsMADS2 knockdown spikelet. The transverse section planes are marked by lines (j1-j2, a-b, c-d, e-f). (C) Overview of wild type lemma (le), palea (pa), marginal tissue of palea (mtp) and lodicule (lo) cell types. The lodicule peripheral regions are marked by black unfilled boxes. (D) Wild type glume histology. (E) Magnified view of the juxtaposed wild type mtp and lodicule at the floret base. (F) Parachymatous cells and vascular elements (white arrowheads) of a wild type lodicule. (G and H) Overview of OsMADS2 knockdown floret histology. The black unfilled box marks the overgrown peripheral regions of the transformed lodicule (lo). (I and J) Magnified view the base of a transformed lodicule in an OsMADS2 knockdown spikelet, compare (E). (K- M) Sections of the transformed lodicule through plane a-b in (K), c-d in (L) and e-f in (M). Scale bar is 1.0 cm in (A and B), 200 µm in (C, E, and G-J), 50 µm in (D, F and K M). (N) RT-PCR showing down-regulation of endogenous OsMADS2 transcripts in dsrnaiosmads2 panicles. Actin transcript levels are used for normalization. PCR amplification was done for 26 and 45 cycles. (O and P) Representation of the differential distribution of OsMADS2 transcripts and local effects upon its knockdown. (O) Lodicule distal and peripheral regions with high transcripts levels have black dots, regions with low expression have grey dots. (P) Early OsMADS2 expression in developing lodicule (not shown here) may delineate the boundary between the schlrenchymatous cells of the mtp (black dots) and the lodicule cells. Black arrows mark the regions that over grow upon OsMADS2 knockdown. 36
37 FIGURE 3. RNA in situ expression analysis of OsMADS4 during spikelet development. (A) Strong expression of OsMADS4 in spikelet meristem (sm) and floral meristem (fm). (B) Expression of OsMADS4 in young floral primordia, lemma/palea primordia are marked by white arrowheads. White arrow denotes emerging glume. (C) OsMADS4 is expressed in stamen (st) and carpel (ca) primordia and is completely excluded from developing lemma and palea (arrowheads). (D F) OsMADS4 expression patterns in near mature spikelets. High-level of OsMADS4 expression continues in stamens (st) and carpel (ca) but is drastically reduced in lodicules (lo). Note the reduced but uniform expression of OsMADS4 throughout the lodicule (lo). (G I) OsMADS4 expression in differentiating OsMADS2 knockdown spikelets. Spikelet in (G) represents slightly earlier developmental stage than (H) where lodicule is completely transformed. (I) is similar to (H) showing only a small portion of proximal region of transformed lodicule. Whorl-specific expression of OsMADS4 is retained in OsMADS2 knockdown spikelets. (J and K) Schematic representation of OsMADS2 and OsMADS4 expression patterns in developing floral organ primordia (J) and in differentiated floral organs (K). OsMADS2 expression domain is denoted by red dots while green colored region represents OsMADS4 expression domain. Pink dots and light green color represent reduced level of expression. FIGURE 4. Global expression analysis of OsMADS2 regulated genes. (A) Transcripts down regulated in OsMADS2 knockdown florets categorized based on their predicted functions. (B) Comparative and verification of expression data retrieved from microarray experiments with quantitative real time PCR analysis of expression levels. The Bar graphs compare fold down regulation in wild type vs. dsrnaiosmads2 florets for ten transcripts. 37
38 Two biological replicates for the RNAs and at least triplicate qpcrs were performed to compute mean and standard deviation for expression levels determined by qrt-pcr C, Normalized expression levels for a few selected potential OsMADS2 target genes in the wild type floret organs- palea, lodicule, stamen and carpel. FIGURE 5. Spatial expression profile of OsMADS2 regulated genes in wild-type florets. (A-C) Localization of AK064240/TCP transcripts. (D-F) Expression profile of AK070518/Cyclin B. (G-I) Spatial distribution AK106784/Yabby1b transcripts. (J-L) Expression pattern of AK072687/LEUNIG. Spikelets in (A), (D), (G) and (J) have initiated stamen differentiation but lodicules are yet to be formed. In (B), (E), (H) and (K) spikelets are at the early stage of lodicules development. Panels (C), (F), (I) and (L) show florets at late stages of lodicule development. Arrowheads point to carpel primordia and arrows to lodicules. gl; outer glume, pa; palea, lo; lodicule st; stamen and ca; carpel Scale bar in B, C, E, H, I and L 50µm and 100µm in A, D, F, G, J and K. 38

39

40
41

42

43
Divergent Regulatory OsMADS2 Functions Control Size, Shape and Differentiation of the Highly Derived Rice Floret Second-Whorl Organ
 Copyright Ó 2007 by the Genetics Society of America DOI: 10.1534/genetics.107.071746 Divergent Regulatory OsMADS2 Functions Control Size, Shape and Differentiation of the Highly Derived Rice Floret Second-Whorl
Copyright Ó 2007 by the Genetics Society of America DOI: 10.1534/genetics.107.071746 Divergent Regulatory OsMADS2 Functions Control Size, Shape and Differentiation of the Highly Derived Rice Floret Second-Whorl
The Plant Cell, November. 2017, American Society of Plant Biologists. All rights reserved
 The Genetics of Floral Development Teaching Guide Overview The development of flowers in angiosperm plants provided a critical evolutionary advantage, allowing more options for pollen dispersal and seed
The Genetics of Floral Development Teaching Guide Overview The development of flowers in angiosperm plants provided a critical evolutionary advantage, allowing more options for pollen dispersal and seed
75 Development of floral organ identity: stories from the MADS house Günter Theißen Recent studies on AGAMOUS-LIKE2-, DEFICIENS- and GLOBOSA-like MADS
 75 Development of floral organ identity: stories from the MADS house Günter Theißen Recent studies on AGAMOUS-LIKE2-, DEFICIENS- and GLOBOSA-like MADS-box genes in diverse seed plant species have provided
75 Development of floral organ identity: stories from the MADS house Günter Theißen Recent studies on AGAMOUS-LIKE2-, DEFICIENS- and GLOBOSA-like MADS-box genes in diverse seed plant species have provided
Analysis of the Petunia TM6 MADS Box Gene Reveals Functional Divergence within the DEF/AP3 Lineage W
 This article is published in The Plant Cell Online, The Plant Cell Preview Section, which publishes manuscripts accepted for publication after they have been edited and the authors have corrected proofs,
This article is published in The Plant Cell Online, The Plant Cell Preview Section, which publishes manuscripts accepted for publication after they have been edited and the authors have corrected proofs,
Seminars in Cell & Developmental Biology
 Seminars in Cell & Developmental Biology 21 (2010) 73 79 Contents lists available at ScienceDirect Seminars in Cell & Developmental Biology journal homepage: www.elsevier.com/locate/semcdb Review Floral
Seminars in Cell & Developmental Biology 21 (2010) 73 79 Contents lists available at ScienceDirect Seminars in Cell & Developmental Biology journal homepage: www.elsevier.com/locate/semcdb Review Floral
Three TOB1-related YABBY genes are required to maintain proper function of the spikelet and branch meristems in rice
 Research Three TOB1-related YABBY genes are required to maintain proper function of the spikelet and branch meristems in rice Wakana Tanaka, Taiyo Toriba and Hiro-Yuki Hirano Department of Biological Sciences,
Research Three TOB1-related YABBY genes are required to maintain proper function of the spikelet and branch meristems in rice Wakana Tanaka, Taiyo Toriba and Hiro-Yuki Hirano Department of Biological Sciences,
Outline. Leaf Development. Leaf Structure - Morphology. Leaf Structure - Morphology
 Outline 1. Leaf Structure: Morphology & Anatomy 2. Leaf Development A. Anatomy B. Sector analysis C. Leaf Development Leaf Structure - Morphology Leaf Structure - Morphology 1 Leaf Structure - Morphology
Outline 1. Leaf Structure: Morphology & Anatomy 2. Leaf Development A. Anatomy B. Sector analysis C. Leaf Development Leaf Structure - Morphology Leaf Structure - Morphology 1 Leaf Structure - Morphology
Genetics and Evolution of Inflorescence and Flower Development in Grasses
 Plant Cell Physiol. 46(1): 69 78 (2005) doi:10.1093/pcp/pci504, available online at www.pcp.oupjournals.org JSPP 2005 Mini Review Genetics and Evolution of Inflorescence and Flower Development in Grasses
Plant Cell Physiol. 46(1): 69 78 (2005) doi:10.1093/pcp/pci504, available online at www.pcp.oupjournals.org JSPP 2005 Mini Review Genetics and Evolution of Inflorescence and Flower Development in Grasses
Biological Roles of Cytokinins
 Direct Control of Shoot Meristem Activity by a Cytokinin-Activating Enzyme By Kurakawa et. Al. Published in Nature Presented by Boyana Grigorova Biological Roles of Cytokinins Cytokinins are positive regulators
Direct Control of Shoot Meristem Activity by a Cytokinin-Activating Enzyme By Kurakawa et. Al. Published in Nature Presented by Boyana Grigorova Biological Roles of Cytokinins Cytokinins are positive regulators
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
Supplemental Data. Yang et al. (2012). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure 1. Mature flowers of P. heterotricha. (A) An inflorescence of P. heterotricha showing the front view of a zygomorphic flower characterized by two small dorsal petals and only two fertile
Supplemental Figure 1. Mature flowers of P. heterotricha. (A) An inflorescence of P. heterotricha showing the front view of a zygomorphic flower characterized by two small dorsal petals and only two fertile
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family
, HAESA, ERECTA-family") Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family GENES & DEVELOPMENT (2000) 14: 108 117 INTRODUCTION Flower Diagram INTRODUCTION Abscission In plant, the process by which a plant
Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family GENES & DEVELOPMENT (2000) 14: 108 117 INTRODUCTION Flower Diagram INTRODUCTION Abscission In plant, the process by which a plant
A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development
 A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development Xuemei Chen Waksman Institute, Rutgers University, Piscataway, NJ 08854, USA. E-mail: xuemei@waksman.rutgers.edu Plant
A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development Xuemei Chen Waksman Institute, Rutgers University, Piscataway, NJ 08854, USA. E-mail: xuemei@waksman.rutgers.edu Plant
Functional lnteraction between the Homeotic Genes fbpl and pmads7 during Petunia Floral Organogenesis
 The Plant Cell, Vol. 7, 507-516, May 1995 O 1995 American Society of Plant Physiologists Functional lnteraction between the Homeotic Genes fbpl and pmads7 during Petunia Floral Organogenesis Gerco C. Angenent,'
The Plant Cell, Vol. 7, 507-516, May 1995 O 1995 American Society of Plant Physiologists Functional lnteraction between the Homeotic Genes fbpl and pmads7 during Petunia Floral Organogenesis Gerco C. Angenent,'
Ectopic expression of rice OsMADS1 reveals a role in specifying the lemma and palea, grass floral organs analogous to sepals
 Dev Genes Evol (2001) 211:281 290 DOI 10.1007/s004270100153 ORIGINAL ARTICLE Kalika Prasad P. Sriram C. Santhosh Kumar Kumuda Kushalappa Usha Vijayraghavan Ectopic expression of rice OsMADS1 reveals a
Dev Genes Evol (2001) 211:281 290 DOI 10.1007/s004270100153 ORIGINAL ARTICLE Kalika Prasad P. Sriram C. Santhosh Kumar Kumuda Kushalappa Usha Vijayraghavan Ectopic expression of rice OsMADS1 reveals a
The unfolding drama of flower development:
 REVIEW The unfolding drama of flower development: recent results from genetic and molecular analyses Hong Ma Cold Spring Harbor Laboratory, Cold Spring Harbor, New York 11724-2212 USA There has been an
REVIEW The unfolding drama of flower development: recent results from genetic and molecular analyses Hong Ma Cold Spring Harbor Laboratory, Cold Spring Harbor, New York 11724-2212 USA There has been an
Spatially and Temporally Regulated Expression of Rice MADS Box Genes with Similarity to Arabidopsis Class A, B and C Genes
 Plant Cell Physiol. 41(6): 710-718 (2000) JSPP 2000 Spatially and Temporally Regulated Expression of Rice MADS Box Genes with Similarity to Arabidopsis Class A, B and C Genes Junko Kyozuka 1, Takeshi Kobayashi,
Plant Cell Physiol. 41(6): 710-718 (2000) JSPP 2000 Spatially and Temporally Regulated Expression of Rice MADS Box Genes with Similarity to Arabidopsis Class A, B and C Genes Junko Kyozuka 1, Takeshi Kobayashi,
Molecular Genetics of. Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS
 Molecular Genetics of Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS Contents Preface A Word on Genetic Nomenclature page xiii xvii 1 Approaches to the Study of Plant Development 1 Pattern
Molecular Genetics of Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS Contents Preface A Word on Genetic Nomenclature page xiii xvii 1 Approaches to the Study of Plant Development 1 Pattern
Lilium longiflorum and Molecular Floral Development: the ABCDE Model
 Lilium longiflorum and Molecular Floral Development: the ABCDE Model V.A. Benedito Laboratório de Melhoramento de Plantas CENA/USP Av. Centenario, 303 13400-961 Piracicaba-SP Brazil G.C. Angenent, J.M.
Lilium longiflorum and Molecular Floral Development: the ABCDE Model V.A. Benedito Laboratório de Melhoramento de Plantas CENA/USP Av. Centenario, 303 13400-961 Piracicaba-SP Brazil G.C. Angenent, J.M.
Divergence of Function and Regulation of Class B Floral Organ ldentity Genes
 The Plant Cell, Vol. 9, 559-570, April 1997 O 1997 American Society of Plant Physiologists Divergence of Function and Regulation of Class B Floral Organ ldentity Genes Alon Samach,a Susanne E. Kohalmi,b.l
The Plant Cell, Vol. 9, 559-570, April 1997 O 1997 American Society of Plant Physiologists Divergence of Function and Regulation of Class B Floral Organ ldentity Genes Alon Samach,a Susanne E. Kohalmi,b.l
Supplemental Data. Wang et al. (2014). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Genetic Analysis of Streaked and Abnormal Floret Mutant st-fon
 Rice Science, 2013, 20(4): 267 275 Copyright 2013, China National Rice Research Institute Published by Elsevier BV. All rights reserved DOI: 10.1016/S1672-6308(13)60132-8 Genetic Analysis of Streaked and
Rice Science, 2013, 20(4): 267 275 Copyright 2013, China National Rice Research Institute Published by Elsevier BV. All rights reserved DOI: 10.1016/S1672-6308(13)60132-8 Genetic Analysis of Streaked and
The MADS-Box Floral Homeotic Gene Lineages Predate the Origin of Seed Plants: Phylogenetic and Molecular Clock Estimates
 J Mol Evol (1997) 45:392 396 Springer-Verlag New York Inc. 1997 The MADS-Box Floral Homeotic Gene Lineages Predate the Origin of Seed Plants: Phylogenetic and Molecular Clock Estimates Michael D. Purugganan
J Mol Evol (1997) 45:392 396 Springer-Verlag New York Inc. 1997 The MADS-Box Floral Homeotic Gene Lineages Predate the Origin of Seed Plants: Phylogenetic and Molecular Clock Estimates Michael D. Purugganan
Redefining C and D in the Petunia ABC W
 The Plant Cell, Vol. 24: 2305 2317, June 2012, www.plantcell.org ã 2012 American Society of Plant Biologists. All rights reserved. RESEARCH ARTICLES Redefining C and D in the Petunia ABC W Klaas Heijmans,
The Plant Cell, Vol. 24: 2305 2317, June 2012, www.plantcell.org ã 2012 American Society of Plant Biologists. All rights reserved. RESEARCH ARTICLES Redefining C and D in the Petunia ABC W Klaas Heijmans,
Genome-Wide Analysis of Gene Expression during Early Arabidopsis Flower Development
 Genome-Wide Analysis of Gene Expression during Early Arabidopsis Flower Development Frank Wellmer, Márcio Alves-Ferreira a, Annick Dubois b, José Luis Riechmann, Elliot M. Meyerowitz * Division of Biology,
Genome-Wide Analysis of Gene Expression during Early Arabidopsis Flower Development Frank Wellmer, Márcio Alves-Ferreira a, Annick Dubois b, José Luis Riechmann, Elliot M. Meyerowitz * Division of Biology,
Genome-Wide Analysis of Spatial Gene Expression in Arabidopsis Flowers W
 The Plant Cell, Vol. 16, 1314 1326, May 2004, www.plantcell.org ª 2004 American Society of Plant Biologists Genome-Wide Analysis of Spatial Gene Expression in Arabidopsis Flowers W Frank Wellmer, José
The Plant Cell, Vol. 16, 1314 1326, May 2004, www.plantcell.org ª 2004 American Society of Plant Biologists Genome-Wide Analysis of Spatial Gene Expression in Arabidopsis Flowers W Frank Wellmer, José
The MIK region rather than the C-terminal domain of
 The MIK region rather than the C-terminal domain of Blackwell Publishing Ltd AP3-like class B floral homeotic proteins determines functional specificity in the development and evolution of petals Kunmei
The MIK region rather than the C-terminal domain of Blackwell Publishing Ltd AP3-like class B floral homeotic proteins determines functional specificity in the development and evolution of petals Kunmei
The MADS Box Gene FBP2 Is Required for SEPALLATA Function in Petunia
 The Plant Cell, Vol. 15, 914 925, April 2003, www.plantcell.org 2003 American Society of Plant Biologists The MADS Box Gene FBP2 Is Required for SEPALLATA Function in Petunia Silvia Ferrario, a Richard
The Plant Cell, Vol. 15, 914 925, April 2003, www.plantcell.org 2003 American Society of Plant Biologists The MADS Box Gene FBP2 Is Required for SEPALLATA Function in Petunia Silvia Ferrario, a Richard
MADS-box genes and floral development: the dark side
 Journal of Experimental Botany, Vol. 63, No. 15, pp. 5397 5404, 2012 doi:10.1093/jxb/ers233 Advance Access publication 21 August, 2012 Flowering Newsletter Review MADS-box genes and floral development:
Journal of Experimental Botany, Vol. 63, No. 15, pp. 5397 5404, 2012 doi:10.1093/jxb/ers233 Advance Access publication 21 August, 2012 Flowering Newsletter Review MADS-box genes and floral development:
Functional Conservation between CRABS CLAW Orthologues from Widely Diverged Angiosperms
 Annals of Botany 100: 651 657, 2007 doi:10.1093/aob/mcm136, available online at www.aob.oxfordjournals.org Functional Conservation between CRABS CLAW Orthologues from Widely Diverged Angiosperms CHLOE
Annals of Botany 100: 651 657, 2007 doi:10.1093/aob/mcm136, available online at www.aob.oxfordjournals.org Functional Conservation between CRABS CLAW Orthologues from Widely Diverged Angiosperms CHLOE
The ABC model of floral patterning, developed from studies of
 Conservation of B class gene expression in the second whorl of a basal grass and outgroups links the origin of lodicules and petals Clinton J. Whipple*, Michael J. Zanis*, Elizabeth A. Kellogg, and Robert
Conservation of B class gene expression in the second whorl of a basal grass and outgroups links the origin of lodicules and petals Clinton J. Whipple*, Michael J. Zanis*, Elizabeth A. Kellogg, and Robert
Curriculum vitae Xigang Liu
 Curriculum vitae Xigang Liu 1, EDUCATION: 09/1993-07/1997 B.S. Major: Biology. College of Life Sciences, Hebei Normal University Academic Degree Paper: RAPD analysis of Taigu genic male-sterile wheat and
Curriculum vitae Xigang Liu 1, EDUCATION: 09/1993-07/1997 B.S. Major: Biology. College of Life Sciences, Hebei Normal University Academic Degree Paper: RAPD analysis of Taigu genic male-sterile wheat and
Plant Structure, Growth, and Development
 Plant Structure, Growth, and Development Plant hierarchy: Cells Tissue: group of similar cells with similar function: Dermal, Ground, Vascular Organs: multiple kinds of tissue, very diverse function Organ
Plant Structure, Growth, and Development Plant hierarchy: Cells Tissue: group of similar cells with similar function: Dermal, Ground, Vascular Organs: multiple kinds of tissue, very diverse function Organ
New Phytologist. II. Phylogeny and subfunctionalization within the AGAMOUS subfamily
 Review MADS reloaded: evolution of the AGAMOUS subfamily genes Author for correspondence: Martin M. Kater Tel: +39 02 50315050 Email: martin.kater@unimi.it Ludovico Dreni and Martin M. Kater Department
Review MADS reloaded: evolution of the AGAMOUS subfamily genes Author for correspondence: Martin M. Kater Tel: +39 02 50315050 Email: martin.kater@unimi.it Ludovico Dreni and Martin M. Kater Department
Members of the YABBY gene family specify abaxial cell fate in Arabidopsis
 Development 126, 4117-4128 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV0240 4117 Members of the YABBY gene family specify abaxial cell fate in Arabidopsis Kellee R. Siegfried
Development 126, 4117-4128 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV0240 4117 Members of the YABBY gene family specify abaxial cell fate in Arabidopsis Kellee R. Siegfried
Expression Analysis of Flower MADS-box Genes in Saffron Crocus (Crocus sativus L.) Supports a Modified ABCDE Model
 Supports a Modified ABCDE Model") Functional Plant Science and Biotechnology 2010 Global Science Books Expression Analysis of Flower MADS-box Genes in Saffron Crocus (Crocus sativus L.) Supports a Modified ABCDE Model Athanasios S. Tsaftaris
Functional Plant Science and Biotechnology 2010 Global Science Books Expression Analysis of Flower MADS-box Genes in Saffron Crocus (Crocus sativus L.) Supports a Modified ABCDE Model Athanasios S. Tsaftaris
To B or Not to B a Flower: The Role of DEFICIENS and GLOBOSA Orthologs in the Evolution of the Angiosperms
 Journal of Heredity 2005:96(3):1 16 doi:10.1093/jhered/esi033 ª 2005 The American Genetic Association To B or Not to B a Flower: The Role of DEFICIENS and GLOBOSA Orthologs in the Evolution of the Angiosperms
Journal of Heredity 2005:96(3):1 16 doi:10.1093/jhered/esi033 ª 2005 The American Genetic Association To B or Not to B a Flower: The Role of DEFICIENS and GLOBOSA Orthologs in the Evolution of the Angiosperms
Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter
 Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter 9/10/2008 1 Learning Objectives Explain similarities and differences between fungal, mammalian and plant cell cycles Explain
Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter 9/10/2008 1 Learning Objectives Explain similarities and differences between fungal, mammalian and plant cell cycles Explain
Evolution and Development of Flower Diversity. Kelsey Galimba Di Stilio Lab Department of Biology University of Washington
 Evolution and Development of Flower Diversity Kelsey Galimba Di Stilio Lab Department of Biology University of Washington Lecture Outline Evolution of Angiosperms! The first flower! Morphological Diversity
Evolution and Development of Flower Diversity Kelsey Galimba Di Stilio Lab Department of Biology University of Washington Lecture Outline Evolution of Angiosperms! The first flower! Morphological Diversity
Morphogenesis of the flower of Arabidopsis, genes networks and mathematical modelling.
 1 / 10 Morphogenesis of the flower of Arabidopsis, genes networks and mathematical modelling. Samuel Collaudin. Master BioSciences, Département de Biologie,., University of Heidelberg 2012-10-01 Organ
1 / 10 Morphogenesis of the flower of Arabidopsis, genes networks and mathematical modelling. Samuel Collaudin. Master BioSciences, Département de Biologie,., University of Heidelberg 2012-10-01 Organ
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Genetic Framework for Flattened Leaf Blade Formation in Unifacial Leaves of Juncus prismatocarpus C W OA
 This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
The mode of development in animals and plants is different
 The mode of development in animals and plants is different Outcome of animal embryogenesis is a mini edition of the adult Outcome of plant embryogenesis is a simple structure with -root apical meristem
The mode of development in animals and plants is different Outcome of animal embryogenesis is a mini edition of the adult Outcome of plant embryogenesis is a simple structure with -root apical meristem
673 Comparative Genomics of Angiosperm MADS Box Genes Yale University, New Haven, CT. 674 The evolution of plant architecture in Brassicaceae
 673 Comparative Genomics of Angiosperm MADS Box Genes Vivian F. Irish Yale University, New Haven, CT. MADS box genes encode key transcriptional regulators that have been implicated in the control of various
673 Comparative Genomics of Angiosperm MADS Box Genes Vivian F. Irish Yale University, New Haven, CT. MADS box genes encode key transcriptional regulators that have been implicated in the control of various
PANICLE PHYTOMER2 (PAP2 ), encoding a SEPALLATA subfamily MADS-box protein, positively controls spikelet meristem identity in rice
, encoding a SEPALLATA subfamily MADS-box protein, positively controls spikelet meristem identity in rice") PANICLE PHYTOMER2 (PAP2 ), encoding a SEPALLATA subfamily MADS-box protein, positively controls spikelet meristem identity in rice Kaoru Kobayashi1, Masahiko Maekawa2, Akio Miyao3, Hirohiko Hirochika3
PANICLE PHYTOMER2 (PAP2 ), encoding a SEPALLATA subfamily MADS-box protein, positively controls spikelet meristem identity in rice Kaoru Kobayashi1, Masahiko Maekawa2, Akio Miyao3, Hirohiko Hirochika3
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
State Key Laboratory of Crop Stress Biology in Arid Areas, Northwest A&F University, Yangling, Shaanxi, China
 Molecular cloning and expression of the male sterility-related CtYABBY1 gene in flowering Chinese cabbage (Brassica campestris L. ssp chinensis var. parachinensis) X.L. Zhang 1,2,3 and L.G. Zhang 1,2,3
Molecular cloning and expression of the male sterility-related CtYABBY1 gene in flowering Chinese cabbage (Brassica campestris L. ssp chinensis var. parachinensis) X.L. Zhang 1,2,3 and L.G. Zhang 1,2,3
Pea Compound Leaf Architecture Is Regulated by Interactions among the Genes UNIFOLIATA, COCHLEATA, AFILA, and TENDRIL-LESS
 The Plant Cell, Vol. 12, 1279 1294, August 2000, www.plantcell.org 2000 American Society of Plant Physiologists Pea Compound Leaf Architecture Is Regulated by Interactions among the Genes UNIFOLIATA, COCHLEATA,
The Plant Cell, Vol. 12, 1279 1294, August 2000, www.plantcell.org 2000 American Society of Plant Physiologists Pea Compound Leaf Architecture Is Regulated by Interactions among the Genes UNIFOLIATA, COCHLEATA,
Genetic control of branching pattern and floral identity during Petunia inflorescence development
 Development 125, 733-742 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV0153 733 Genetic control of branching pattern and floral identity during Petunia inflorescence development
Development 125, 733-742 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV0153 733 Genetic control of branching pattern and floral identity during Petunia inflorescence development
JAGGED Controls Arabidopsis Petal Growth and Shape by Interacting with a Divergent Polarity Field
 by Interacting with a Divergent Polarity Field Susanna Sauret-Güeto 1, Katharina Schiessl 1, Andrew Bangham 2, Robert Sablowski 1, Enrico Coen 1 * 1 Department of Cell and Developmental Biology, John Innes
by Interacting with a Divergent Polarity Field Susanna Sauret-Güeto 1, Katharina Schiessl 1, Andrew Bangham 2, Robert Sablowski 1, Enrico Coen 1 * 1 Department of Cell and Developmental Biology, John Innes
Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed
 Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed using the Geneious software. Accession numbers of the
Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed using the Geneious software. Accession numbers of the
Cloning and expression analysis of an E-class MADS-box gene from Populus deltoides
 African Journal of Biotechnology Vol. 8 (19), pp. 4789-4796, 5 October, 2009 Available online at http://www.academicjournals.org/ajb ISSN 1684 5315 2009 Academic Journals Full Length Research Paper Cloning
African Journal of Biotechnology Vol. 8 (19), pp. 4789-4796, 5 October, 2009 Available online at http://www.academicjournals.org/ajb ISSN 1684 5315 2009 Academic Journals Full Length Research Paper Cloning
UNIVERSITY OF YORK BIOLOGY. Developmental Biology
 Examination Candidate Number: UNIVERSITY OF YORK BSc Stage 2 Degree Examinations 2017-18 Department: BIOLOGY Title of Exam: Developmental Biology Desk Number: Time allowed: 1 hour and 30 minutes Total
Examination Candidate Number: UNIVERSITY OF YORK BSc Stage 2 Degree Examinations 2017-18 Department: BIOLOGY Title of Exam: Developmental Biology Desk Number: Time allowed: 1 hour and 30 minutes Total
RNAi Suppression of AGAMOUS-like Genes Causes Field Sterility in Populus
 RNAi Suppression of AGAMOUS-like Genes Causes Field Sterility in Populus Haiwei Lu and Steven H. Strauss Oregon State University Forest Tree Workshop PAG XXVI, San Diego, CA, 2018 The containment issue
RNAi Suppression of AGAMOUS-like Genes Causes Field Sterility in Populus Haiwei Lu and Steven H. Strauss Oregon State University Forest Tree Workshop PAG XXVI, San Diego, CA, 2018 The containment issue
Flower Development and Photoperiodic Control of Flowering in Rice
 Rice Science, 2013, 20(2): Copyright 2013, China National Rice Research Institute Published by Elsevier BV. All rights reserved DOI: 10.1016/S1672-6308(13)60112-2 Flower Development and Photoperiodic Control
Rice Science, 2013, 20(2): Copyright 2013, China National Rice Research Institute Published by Elsevier BV. All rights reserved DOI: 10.1016/S1672-6308(13)60112-2 Flower Development and Photoperiodic Control
23-. Shoot and root development depend on ratio of IAA/CK
 Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Genome-wide Identification of Lineage Specific Genes in Arabidopsis, Oryza and Populus
 Genome-wide Identification of Lineage Specific Genes in Arabidopsis, Oryza and Populus Xiaohan Yang Sara Jawdy Timothy Tschaplinski Gerald Tuskan Environmental Sciences Division Oak Ridge National Laboratory
Genome-wide Identification of Lineage Specific Genes in Arabidopsis, Oryza and Populus Xiaohan Yang Sara Jawdy Timothy Tschaplinski Gerald Tuskan Environmental Sciences Division Oak Ridge National Laboratory
MOLECULAR MECHANISMS OF FLOWER DEVELOPMENT: AN ARMCHAIR GUIDE
 MOLCULAR MCHANISMS OF FLOWR DVLOPMNT: AN ARMCHAIR GUID Beth A. Krizek* and Jennifer C. Fletcher Abstract An afternoon stroll through an nglish garden reveals the breathtaking beauty and enormous diversity
MOLCULAR MCHANISMS OF FLOWR DVLOPMNT: AN ARMCHAIR GUID Beth A. Krizek* and Jennifer C. Fletcher Abstract An afternoon stroll through an nglish garden reveals the breathtaking beauty and enormous diversity
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Cell Cycle Regulation by Chlamydomonas Cyclin-Dependent Protein Kinases
 Plant Cell Advance Publication. Published on February 5, 2018, doi:10.1105/tpc.18.00103 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 IN BRIEF Cell Cycle Regulation by Chlamydomonas
Plant Cell Advance Publication. Published on February 5, 2018, doi:10.1105/tpc.18.00103 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 IN BRIEF Cell Cycle Regulation by Chlamydomonas
Gwyneth C. Ingram,a Justin Goodrich,a Mark D. Wilkinson,b Rüdiger Simon,a George W. Haughn,b and Enrico S. Coena,
 The Plant Cell, Vol. 7, 1501-1510, September 1995 O 1995 American Society of Plant Physiologists Parallels between UNUSUAL FLORAL ORGANS and FMBRATA, Genes Controlling Flower Development in Arabidopsis
The Plant Cell, Vol. 7, 1501-1510, September 1995 O 1995 American Society of Plant Physiologists Parallels between UNUSUAL FLORAL ORGANS and FMBRATA, Genes Controlling Flower Development in Arabidopsis
Computational identification and analysis of MADS box genes in Camellia sinensis
 www.bioinformation.net Hypothesis Volume 11(3) Computational identification and analysis of MADS box genes in Camellia sinensis Madhurjya Gogoi*, Sangeeta Borchetia & Tanoy Bandyopadhyay Department of
www.bioinformation.net Hypothesis Volume 11(3) Computational identification and analysis of MADS box genes in Camellia sinensis Madhurjya Gogoi*, Sangeeta Borchetia & Tanoy Bandyopadhyay Department of
THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING. AnitaHajdu. Thesis of the Ph.D.
 THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING AnitaHajdu Thesis of the Ph.D. dissertation Supervisor: Dr. LászlóKozma-Bognár - senior research associate Doctoral
THE ROLE OF THE PHYTOCHROME B PHOTORECEPTOR IN THE REGULATION OF PHOTOPERIODIC FLOWERING AnitaHajdu Thesis of the Ph.D. dissertation Supervisor: Dr. LászlóKozma-Bognár - senior research associate Doctoral
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Tepal formation and expression pattern of B-class paleoap3-like MADS-box genes in crocus (Crocus sativus L.)
") Plant Science 170 (2006) 238 246 www.elsevier.com/locate/plantsci Tepal formation and expression pattern of B-class paleoap3-like MADS-box genes in crocus (Crocus sativus L.) Athanasios S. Tsaftaris a,b,
Plant Science 170 (2006) 238 246 www.elsevier.com/locate/plantsci Tepal formation and expression pattern of B-class paleoap3-like MADS-box genes in crocus (Crocus sativus L.) Athanasios S. Tsaftaris a,b,
Pathways for inflorescence and floral induction in Antirrhinum
 Development 122, 1535-1544 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV3397 1535 Pathways for inflorescence and floral induction in Antirrhinum Desmond Bradley, Coral Vincent,
Development 122, 1535-1544 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV3397 1535 Pathways for inflorescence and floral induction in Antirrhinum Desmond Bradley, Coral Vincent,
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
Lecture 18 June 2 nd, Gene Expression Regulation Mutations
 Lecture 18 June 2 nd, 2016 Gene Expression Regulation Mutations From Gene to Protein Central Dogma Replication DNA RNA PROTEIN Transcription Translation RNA Viruses: genome is RNA Reverse Transcriptase
Lecture 18 June 2 nd, 2016 Gene Expression Regulation Mutations From Gene to Protein Central Dogma Replication DNA RNA PROTEIN Transcription Translation RNA Viruses: genome is RNA Reverse Transcriptase
Prokaryotic Regulation
 Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Supplementary Figure 1. Phenotype of the HI strain.
 Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Gene Trap Lines Define Domains of Gene Regulation in Arabidopsis Petals and Stamens W
 The Plant Cell, Vol. 17, 2486 2506, September 2005, www.plantcell.org ª 2005 American Society of Plant Biologists Gene Trap Lines Define Domains of Gene Regulation in Arabidopsis Petals and Stamens W Naomi
The Plant Cell, Vol. 17, 2486 2506, September 2005, www.plantcell.org ª 2005 American Society of Plant Biologists Gene Trap Lines Define Domains of Gene Regulation in Arabidopsis Petals and Stamens W Naomi
Regulation of Phosphate Homeostasis by microrna in Plants
 Regulation of Phosphate Homeostasis by microrna in Plants Tzyy-Jen Chiou 1 *, Kyaw Aung 1,2, Shu-I Lin 1,3, Chia-Chune Wu 1, Su-Fen Chiang 1, and Chun-Lin Su 1 Abstract Upon phosphate (Pi) starvation,
Regulation of Phosphate Homeostasis by microrna in Plants Tzyy-Jen Chiou 1 *, Kyaw Aung 1,2, Shu-I Lin 1,3, Chia-Chune Wu 1, Su-Fen Chiang 1, and Chun-Lin Su 1 Abstract Upon phosphate (Pi) starvation,
Molecular and Genetic Mechanisms of Floral Control
 The Plant Cell, Vol. 16, S1 S17, Supplement 2004, www.plantcell.org ª 2004 American Society of Plant Biologists Molecular and Genetic Mechanisms of Floral Control Thomas Jack 1 Department of Biological
The Plant Cell, Vol. 16, S1 S17, Supplement 2004, www.plantcell.org ª 2004 American Society of Plant Biologists Molecular and Genetic Mechanisms of Floral Control Thomas Jack 1 Department of Biological
SUPPLEMENTARY INFORMATION
 med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms.
 Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
B-Function Expression in the Flower Center Underlies the Homeotic Phenotype of Lacandonia schismatica (Triuridaceae) C W OA
 C W OA") The Plant Cell, Vol. 22: 3543 3559, November 2010, www.plantcell.org ã 2010 American Society of Plant Biologists B-Function Expression in the Flower Center Underlies the Homeotic Phenotype of Lacandonia
The Plant Cell, Vol. 22: 3543 3559, November 2010, www.plantcell.org ã 2010 American Society of Plant Biologists B-Function Expression in the Flower Center Underlies the Homeotic Phenotype of Lacandonia
Nature Genetics: doi: /ng Supplementary Figure 1. The phenotypes of PI , BR121, and Harosoy under short-day conditions.
 Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
The Role of Developmental Genetics in Understanding Homology and Morphological Evolution in Plants
 The Role of Developmental Genetics in Understanding Homology and Morphological Evolution in Plants The Harvard community has made this article openly available. Please share how this access benefits you.
The Role of Developmental Genetics in Understanding Homology and Morphological Evolution in Plants The Harvard community has made this article openly available. Please share how this access benefits you.
Supplemental Data. Perrella et al. (2013). Plant Cell /tpc
. Plant Cell /tpc") Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
"PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200B Spring 2011
 "PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200B Spring 2011 Evolution and development ("evo-devo") The last frontier in our understanding of biological forms is an understanding
"PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200B Spring 2011 Evolution and development ("evo-devo") The last frontier in our understanding of biological forms is an understanding
Anatomy of Plants Student Notes
 Directions: Fill in the blanks. Anatomy of Plants Student Notes Plant Cell Biology Segment 1. Plants Plants are organisms are incapable of movement produce food through 2. Animals Animals are multicellular
Directions: Fill in the blanks. Anatomy of Plants Student Notes Plant Cell Biology Segment 1. Plants Plants are organisms are incapable of movement produce food through 2. Animals Animals are multicellular
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
Last time: Obtaining information from a cloned gene
 Last time: Obtaining information from a cloned gene Objectives: 1. What is the biochemical role of the gene? 2. Where and when is the gene expressed (transcribed)? 3. Where and when is the protein made?
Last time: Obtaining information from a cloned gene Objectives: 1. What is the biochemical role of the gene? 2. Where and when is the gene expressed (transcribed)? 3. Where and when is the protein made?
Plant Growth and Development Part I. Levels of Organization
 Plant Growth and Development Part I Levels of Organization Whole Plant Organs Tissues Cells Organelles Macromolecules Levels of Organization Whole Plant Organs Tissues Cells Organelles Macromolecules 1
Plant Growth and Development Part I Levels of Organization Whole Plant Organs Tissues Cells Organelles Macromolecules Levels of Organization Whole Plant Organs Tissues Cells Organelles Macromolecules 1
bearded-ear Encodes a MADS Box Transcription Factor Critical for Maize Floral Development W OA
 The Plant Cell, Vol. 21: 2578 2590, September 2009, www.plantcell.org ã 2009 American Society of Plant Biologists bearded-ear Encodes a MADS Box Transcription Factor Critical for Maize Floral Development
The Plant Cell, Vol. 21: 2578 2590, September 2009, www.plantcell.org ã 2009 American Society of Plant Biologists bearded-ear Encodes a MADS Box Transcription Factor Critical for Maize Floral Development
Silvia Ferrario, Jacqueline Busscher, John Franken, Tom Gerats, 1 Michiel Vandenbussche, 2 Gerco C. Angenent, 3 and Richard G.H.
 The Plant Cell, Vol. 16, 1490 1505, June 2004, www.plantcell.org ª 2004 American Society of Plant Biologists Ectopic Expression of the Petunia MADS Box Gene UNSHAVEN Accelerates Flowering and Confers Leaf-Like
The Plant Cell, Vol. 16, 1490 1505, June 2004, www.plantcell.org ª 2004 American Society of Plant Biologists Ectopic Expression of the Petunia MADS Box Gene UNSHAVEN Accelerates Flowering and Confers Leaf-Like
Epigenetics and Flowering Any potentially stable and heritable change in gene expression that occurs without a change in DNA sequence
 Epigenetics and Flowering Any potentially stable and heritable change in gene expression that occurs without a change in DNA sequence www.plantcell.org/cgi/doi/10.1105/tpc.110.tt0110 Epigenetics Usually
Epigenetics and Flowering Any potentially stable and heritable change in gene expression that occurs without a change in DNA sequence www.plantcell.org/cgi/doi/10.1105/tpc.110.tt0110 Epigenetics Usually
Other funding Sources Agency Name: MSU Agricultural Experiment Station /Project GREEEN Amount requested or awarded: 30,000
 FINAL PROJECT REPORT Project Title: Functional genomics of flowering in apple PI: Herb Aldwinckle Co-PI(2): Steve VanNocker Organization: Cornell University Organization: Michigan State University Telephone/email:
FINAL PROJECT REPORT Project Title: Functional genomics of flowering in apple PI: Herb Aldwinckle Co-PI(2): Steve VanNocker Organization: Cornell University Organization: Michigan State University Telephone/email:
Flower symmetry and shape in Antirrhinum
 Int. J. Dev. Biol. 49: 527-537 (2005) doi: 10.1387/ijdb.041967ja Flower symmetry and shape in ntirrhinum JORGE LMEID*,1,2 and LISETE GLEGO 2 1 Instituto Superior de gronomia (IS), Lisboa, Portugal and
Int. J. Dev. Biol. 49: 527-537 (2005) doi: 10.1387/ijdb.041967ja Flower symmetry and shape in ntirrhinum JORGE LMEID*,1,2 and LISETE GLEGO 2 1 Instituto Superior de gronomia (IS), Lisboa, Portugal and
SUMMARY. Keywords: floral development, regulation of gene expression, phytoplasma, petunia, plant responses, floral quartet model.
 The Plant Journal (2011) 67, 971 979 doi: 10.1111/j.1365-313X.2011.04650.x Unique morphological changes in plant pathogenic phytoplasma-infected petunia flowers are related to transcriptional regulation
The Plant Journal (2011) 67, 971 979 doi: 10.1111/j.1365-313X.2011.04650.x Unique morphological changes in plant pathogenic phytoplasma-infected petunia flowers are related to transcriptional regulation
Flower and fruit development in Arabidopsis thaliana
 Int. J. Dev. Biol. 49: 633-643 (2005) doi: 10.1387/ijdb.052020pr Flower and fruit development in Arabidopsis thaliana PEDRO ROBLES 1 and SORAYA PELAZ*,2 1 División de Genética and Instituto de Bioingeniería,
Int. J. Dev. Biol. 49: 633-643 (2005) doi: 10.1387/ijdb.052020pr Flower and fruit development in Arabidopsis thaliana PEDRO ROBLES 1 and SORAYA PELAZ*,2 1 División de Genética and Instituto de Bioingeniería,
Building Beauty: The Genetic Control of Floral Patterning
 Developmental Cell, Vol. 2, 135 142, February, 2002, Copyright 2002 by Cell Press Building Beauty: The Genetic Control of Floral Patterning Review Jan U. Lohmann 1 and Detlef Weigel 1,2,3 1 Plant Biology
Developmental Cell, Vol. 2, 135 142, February, 2002, Copyright 2002 by Cell Press Building Beauty: The Genetic Control of Floral Patterning Review Jan U. Lohmann 1 and Detlef Weigel 1,2,3 1 Plant Biology
Floral homeotic genes were recruited from homologous MADS-box genes preexisting in the common ancestor of ferns and seed plants
 Proc. Natl. Acad. Sci. USA Vol. 94, pp. 2415 2420, March 1997 Evolution Floral homeotic genes were recruited from homologous MADS-box genes preexisting in the common ancestor of ferns and seed plants THOMAS
Proc. Natl. Acad. Sci. USA Vol. 94, pp. 2415 2420, March 1997 Evolution Floral homeotic genes were recruited from homologous MADS-box genes preexisting in the common ancestor of ferns and seed plants THOMAS
The architecture of transcription elongation A crystal structure explains how transcription factors enhance elongation and pausing
 The architecture of transcription elongation A crystal structure explains how transcription factors enhance elongation and pausing By Thomas Fouqueau and Finn Werner The molecular machines that carry out
The architecture of transcription elongation A crystal structure explains how transcription factors enhance elongation and pausing By Thomas Fouqueau and Finn Werner The molecular machines that carry out
Levels of Organization
 Plant Growth and Development Part I Levels of Organization Whole Plant Organs Tissues Cells Organelles Macromolecules Levels of Organization Whole Plant Organs Tissues Cells Organelles Macromolecules Plant
Plant Growth and Development Part I Levels of Organization Whole Plant Organs Tissues Cells Organelles Macromolecules Levels of Organization Whole Plant Organs Tissues Cells Organelles Macromolecules Plant
BIGPETALp,a bhlh transcription factor is involved in the control of Arabidopsis petal size
 The EMBO Journal () 5, 391 39 & European Molecular Biology Organization All Rights Reserved 1-189/ www.embojournal.org BIGPETALp,a bhlh transcription factor is involved in the control of Arabidopsis petal
The EMBO Journal () 5, 391 39 & European Molecular Biology Organization All Rights Reserved 1-189/ www.embojournal.org BIGPETALp,a bhlh transcription factor is involved in the control of Arabidopsis petal
Understanding the Genetic Basis of Floral Diversity
 Understanding the Genetic Basis of Floral Diversity The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Kramer, Elena M.
Understanding the Genetic Basis of Floral Diversity The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Kramer, Elena M.
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps