MOLECULAR CELL BIOLOGY
|
|
|
- Charlotte Adele Johns
- 5 years ago
- Views:
Transcription
1 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 11 Transmembrane Transport of Ions and Small Molecules Copyright 2013 by W. H. Freeman and Company
2 Transport mechanism 1. Passive transport 1) simple diffusion (passive diffusion) 2) facilitated diffusion 2. Active transport 1) primary active transport (ATP-powered pump) 2) secondary active transport (cotransport) gradient passive active
3
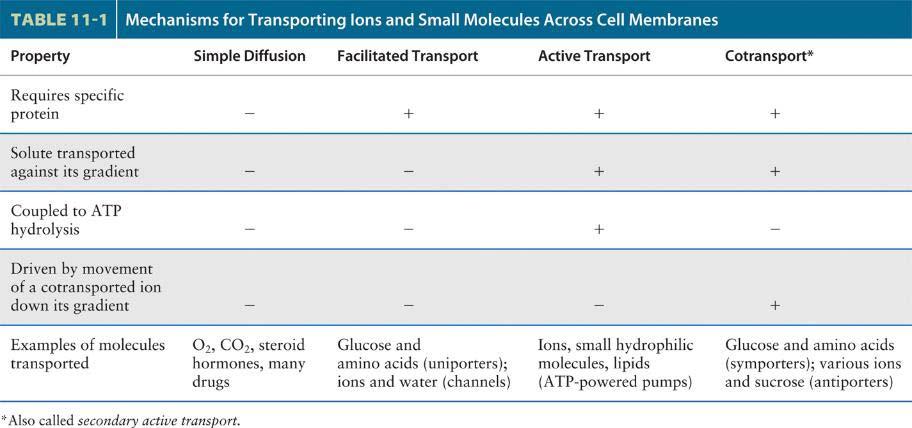
4 Transport mechanism 1. Active and passive transport are biological processes that move oxygen, water and nutrients into cells and remove waste products. 1 2 Active transport requires chemical energy because it is the movement of biochemicals from areas of lower concentration to areas of higher concentration. On the other hand, passive trasport moves biochemicals from areas of high concentration to areas of low concentration; so it does not require energy. 2. Table 11-1 summarizes the four mechanisms (simple diffusion, facilitated transport, active transport, cotransport) by which small molecules and ions are transported across cellular membranes.
5

6 Figure 11.2 Overview of membrane transport proteins.
7 Membrane transport proteins Thus transport of most molecules into and out of cells requires the assistance of specialized membrane proteins (Figure 11-2). 1. Pumps utilize the energy released by ATP hydrolysis to power movement of specific ions or small molecules against their electrochemical gradient. 2. Channels permit movement of specific ions (or water) down their electrochemical gradient. 3. Transporters, which fall into three groups, facilitate movement of specific small molecules or ions. Uniporters transport a single type of molecule down its concentration gradient. Cotransport proteins (symporters, and antiporters) catalyze the movement of one molecule against its concentration gradient (black circles), driven by movement of one or more ions down an electrochemical gradient (red circles). Differences in the mechanisms of transport by these three major classes of proteins account for their varying rates of solute movement.
8

9 Simple diffusion Figure 11.1 Relative permeability of a pure phospholipid bilayer to various molecules and ions.
10 Simple diffusion With its dense hydrophobic core, a phospholipid bilayer is largely impermeable to water-soluble molecules and ions. Only gases, such as O 2 and CO 2, and small, uncharged polar molecules, such as urea and ethanol, can readily move by simple diffusion across an artificial membrane composed of pure phospholipid or of phospholipid and cholesterol. Such molecules also can diffuse across cellular membranes without the aid of transport proteins (Figure 11-1).
11

12 Facilitated diffusion 1. glucose 2. amino acids 3. ions: ion channel 4. water Experimental Figure 11.4 Cellular uptake of glucose mediated by GLUT proteins exhibits simple enzyme kinetics. Figure 11.5 Model of uniport transport by GLUT1.

13 Ion channel: facilitated diffusion
14 Facilitated diffusion 1. The protein-mediated transport of a single type of molecule, such as glucose or other small hydrophilic molecules, down a concentration gradient across a cellular membrane is known as uniport (Experimental Figure 11-4, Figure 11-5). 1 2 The rate of substrate movement by uniporters is far higher than simple diffusion through a pure phospholipid bilayer. A measure of the affinity of a tranporter for its substrate is Km, which is the concentration of substrate at which transport is half maximal. Uniport proteins, such as the glucose transporters (GLUTs), are thought to shuttle between two conformational states, one in which the substrate-binding site faces outward and one in which the binding site faces inward. 2. Channels transport water, specific ions, or hydrophilic small molecules across membranes down their concentration or electric potential gradients. Because this process requires transport proteins but not energy, it is sometimes referred to as passive transport or facilitated diffusion, but it is more properly called facilitated transport. Voltage-gated ion channels open and close in response to membrane potential. Ligand-gated ion channels are a group of transmembrane ion channel proteins which open to allow ions such as Na +, K +, Ca 2+, or Cl to pass through the membrane in response to the binding of a chemical messenger (i.e. a ligand), such as a neurotransmitter.
15

16 Figure 11.8 Structure of the water-channel protein aquaporin.

17 Figure 11.6 Osmotic pressure.
18 Aquaporin Aquaporins are water-channel proteins that specifically increase the permeability of biomembranes for water. Aquaporin 2 in the plasma membrane of certain kidney cells is essential for resorption of water from the forming urine; its absence leads to a form of diabetes. In its functional form, aquaporin is a tetramer of identical 28-kDa subunits (a). Each subunit contains six membrane-spanning α helices that form a central pore through which water moves (b, c). Water tends to move across a semipermeable membrane from a solution of low solute concentration to one of high concentration, a process termed osmosis, or osmotic flow. In other words, since solutions with a high concentration of dissolved solute have a lower concentration of water, water will spontaneously move from a solution of high water concentration to one of lower. In effect, osmosis is equivalent to diffusion of water. Osmotic pressure is defined as the hydrostatic pressure required to stop the net flow of water across a membrane separating solutions of different compositions. In this context, the membrane may be a layer of cells or a plasma membrane that is permeable to water but not to the solutes. The osmotic pressure is directly proportional to the difference in the concentration of the total number of solute molecules on each side of the membrane.
19
20 2 Active transport 1) primary active transport (ATP-powered pump) 2) secondary active transport (cotransport)

21 Figure 11.9 The four classes of ATP-powered transport proteins.
22 The four classes of ATP-powered transport proteins All ATP-powered pumps are transmembrane proteins with one or more binding sites for ATP located on the cytosolic face of the membrane. Although these proteins commonly are called ATPases, they normally do not hydrolyze ATP into ADP and Pi unless ions or other molecules are simultaneously transported. Because of this tight coupling between ATP hydrolysis and transport, the energy stored in the phosphoanhydride bond is not dissipated but rather used to move ions or other molecules uphill against an electrochemical gradient. The general structures of the four classes of ATP-powered pumps are depicted in Figure 11-9, with specific examples in each class listed below. Note that the members of three classes (P, F, and V) transport ions only, whereas members of the ABC superfamily primarily transport small molecules.
23

24 P-class pump
25 P-class pump All P-class ion pumps possess two identical catalytic α subunits that contain an ATP-binding site. Most also have two smaller β subunits that usually have regulatory functions. During the transport process, at least one of the α subunits is phosphorylated (hence the name P class), and the transported ions are thought to move through the phosphorylated subunit. 1. This class includes the Na + /K + ATPase in the plasma membrane, which maintains the low cytosolic Na + and high cytosolic K + concentrations typical of animal cells. 2. Certain Ca 2+ ATPases pump Ca 2+ ions out of the cytosol into the external medium; others pump Ca 2+ from the cytosol into the endoplasmic reticulum or into the specialized ER called the sarcoplasmic reticulum, which is found in muscle cells. 3. Another member of the P class, found in acid-secreting cells of the mammalian stomach, transports protons (H + ions) out of and K + ions into the cell. 4. The H + pump that generates and maintains the membrane electric potential in plant, fungal, and bacterial cells also belongs to this class.
26

27 Na +, K +, Ca 2+ gradient outside inside Na + Cl - Ca 2+ K + ER & SR Ca 2+
28 Na +, K +, Ca 2+ gradient The specific ionic composition of the cytosol usually differs greatly from that of the surrounding extracellular fluid. Also, the cytosolic concentration of K + is much higher than that of Na +. Some Ca 2+ in the cytosol is bound to the negatively charged groups in ATP and other molecules, but it is the concentration of free, unbound Ca 2+ that is critical to its functions in signaling pathways and muscle contraction (Table 11-2).
29

30 Na + /K + ATPase

31 Figure Operational model of the plasma membrane Na + /K + ATPase.

")

32 The Nobel Prize in Chemistry 1997: elucidation of the enzymatic mechanism underlying the synthesis of adenosine triphosphate (ATP) The Nobel Prize in Chemistry 1997: discovery of an ion-transporting enzyme, Na+, K+ -ATPase
33 Na + /K + ATPase 1. An important P-class ion pump present in the plasma membrane of all animal cells is the Na+/K+ ATPase. This ion pump is a tetramer of subunit composition α2β2, and shares structural homology with the Ca2+ pump (Figure 11-13) During the catalytic cycle of the Na+/K+ ATPase it moves three Na+ ions out of and two K+ ions into the cell per ATP molecule hydrolyzed. In its E1 conformation, the Na+/K+ ATPase has three high-affinity Na+-binding sites and two lowaffinity K+-binding sites accessible to the cytosolic surface of the protein. Conversely, the affinity of the cytosolic K+-binding sites is low enough that K+ ions, transported inward through the protein, dissociate from E1 into the cytosol despite the high intracellular K+ concentration. During the E1 E2 transition, the three bound Na+ ions become accessible to the exoplasmic face, and stimultaneously the affinity of the three Na+-binding sites drops. The three Na+ ions now bound to low-affinity Na+ sites dissociate one at a time into the extracellular medium despite the high extracellular Na+ concentration. Transition to the E2 conformation also generates two high-affinity K+ sites accessible to the exoplasmic face. Similarly, during the subsequent E2 E1 transition, the two bound K+ ions are transported inward and then released into the cytosol. 2. Ion pumping by the Na+/K+ ATPase involves phosphorylation, dephosphorylation, and conformational changes similar to those in the muscle Ca2+ ATPase. In this case, hydrolysis of the E2 P intermediate powers the E2 E1 conformational change and concomitant transport of two K+ ions inward (Figure 11-13).
34
35 3 Physilogical roles of Na + /K + gradient 1. Osmolarity & cell volume maintenance 2. Resting membrane potential (K+) 3. Energy for secondary active transport : electrochemical gradient of Na+
36 The Na+/K+-ATPase helps maintain resting potential, effect transport, and regulate cellular volume. 1. Resting potential : In order to maintain the cell membrane potential, cells keep a low concentration of sodium ions and high levels of potassium ions within the cell (intracellular). The sodiumpotassium pump moves 3 sodium ions out and moves 2 potassium ions in, thus, in total, removing one positive charge carrier from the intracellular space. The action of the sodium-potassium pump is not the only mechanism responsible for the generation of the resting membrane potential. 2. Transport : Export of sodium from the cell provides the driving force for several secondary active transporters membrane transport proteins, which import glucose, amino acids, and other nutrients into the cell by use of the sodium gradient. Another important task of the Na+-K+ pump is to provide a Na+ gradient that is used by certain carrier processes. 3. Controlling cell volume : Failure of the Na+-K+ pumps can result in swelling of the cell. A cell's osmolarity is the sum of the concentrations of the various ion species and many proteins and other organic compounds inside the cell. When this is higher than the osmolarity outside of the cell, water flows into the cell through osmosis. This can cause the cell to swell up and lyse. The Na+-K+ pump helps to maintain the right concentrations of ions.
37

 depends on")
38 Resting membrane potential Experimental Figure The electric potential across the plasma membrane of living cells can be measured. Experimental Figure Generation of a transmembrane electric potential (voltage) depends on the selective movement of ions across a semipermeable membrane.
39 Resting membrane potential 1. The plasma membranes of animal cells contain many open K+ channels but few open Na+, Cl-, or Ca2+ channels. As a result, the major ionic movement across the plasma membrane is that of K+ from the inside outward, powered by the K+ concentration gradient, leaving an excess of negative charge on the inside and creating an excess of positive charge on the outside, similar to the experimental system shown in Figure 11-18c. Thus the outward flow of K+ ions through these channels, called resting K+ channels, is the major determinant of the inside-negative membrane potential (Experimental Figure 11-18). 2. The potential across the plasma membrane of large cells can be measured with a microelectrode inserted inside the cell and a reference electrode placed in the extracellular fluid. The two are connected to a potentiometer capable of measuring small potential differences. In virtually all cells the inside (cytosolic face) of the cell membrane is negative relative to the outside; typical membrane potentials range between 30 and 70 mv. The potential across the surface membrane of most animal cells generally does not vary with time. In contrast, neurons and muscle cells the principal types of electrically active cells undergo controlled changes in their membrane potential that we discuss later (Experimental Figure 11-19).
40 Resting K + channels 1. Like all other K + channels, bacterial K + channels are built of four identical subunits symmetrically arranged around a central pore. Each subunit contains two membrane-spanning α helices (S5 and S6) and a short P (pore domain) segment that partly penetrates the membrane bilayer. Four extended loops that are part of the P segments form the actual ion-selectivity filter in the narrow part of the pore near the exoplasmic surface above the vestibule (Figure 11-20). 2. The ability of the ion-selectivity filter in K + channels to select K + over Na + is due mainly to backbone carbonyl oxygens on glycine residues located in a Gly-Tyr-Gly sequence that is found in an analogous position in the P segment in every known K + channel. As a K + ion enters the narrow selectivity filter, it loses its water of hydration but becomes bound to eight backbone carbonyl oxygens, two from the extended loop in each P segment lining the channel. As a result, a relatively low activation energy is required for passage of K + ions through the channel. Because a dehydrated Na + ion is too small to bind to all eight carbonyl oxygens that line the selectivity filter, the activation energy for passage of Na + ions is relatively high. This difference in activation energies favors passage of K + ions over Na + by a factor of thousand. Calcium ions are too large to pass through a K + channel with or without their bound water (Figure 11-21).
41
42 Energy for secondary active transport Figure Transmembrane forces acting on Na + ions.
43 Energy for secondary active transport The electrochemical gradient across a semipermeable membrane determines the direction of ion movement through channel proteins. The two forces constituting the electrochemical gradient, the membrane electric potential and the ion concentration gradient, may act in the same or opposite directions. As with all ions, the movement of Na+ ions across the plasma membrane is governed by the sum of two separate forces the ion concentration gradient and the membrane electric potential. At the internal and external Na+ concentrations typical of mammalian cells, these forces usually act in the same direction, making the inward movement of Na+ ions energetically favorable (Figure 11-25).
44

45 Figure Operational model of the Ca 2+ ATPase in the SR membrane of skeletal muscle cells.
46 Ca 2+ ATPase 1. In skeletal muscle cells, Ca2+ ions are concentrated and stored in the sarcoplasmic reticulum (SR), a specialized type of endoplasmic reticulum (ER). The release via ion channels of stored Ca2+ ions from the SR lumen into the cytosol causes muscle contraction. A P-class Ca2+ ATPase located in the SR membrane of skeletal muscle pumps Ca2+ from the cytosol back into the lumen of the SR, thereby inducing muscle relaxation (Figure 11-10). 1 2 The current model for the mechanism of action of the Ca2+ ATPase in the SR membrane involves multiple conformational states. For simplicity, we group these into E1 states, in which the two binding sites for Ca2+, located in the center of the membrane-spanning domain, face the cytosol, and E2 states, in which these binding sites face the exoplasmic face of the membrane, pointing into the lumen of the SR. Coupling of ATP hydrolysis with ion pumping involves several conformational changes in the protein that must occur in a defined order. When the protein is in the E1 conformation, two Ca2+ ions bind to two high-affinity binding sites accessible from the cytosolic side; even though the cytosolic Ca2+ concentration is low, calcium ions still fill these sites. All P-class ion pumps, regardless of which ion they transport, are phosphorylated on a highly concserved aspartate residue during the transport process. 2. This suggestion is borne out by the similarities of the three-dimensional structures of the membranespanning segments of the Na+/K+ ATPase with that of the Ca2+ pump; the molecular structures of the three cytoplasmic domains are also very similar. Thus, the operational model in Figure is generally applicable to all of the P-class ATP-powered ion pumps.
47

48 H + /K + ATPase in stomach parietal cell Figure Acidification of the stomach lumen by parietal cells in the gastric lining.
49 H + /K + ATPase in stomach parietal cell The overall process by which parietal cells acidify the stomach lumen is illustrated in Figure In a reaction catalyzed by carbonic anhydrase the excess cytosolic OH - combines with CO 2 that diffuses in from the blood, forming HCO 3-. Catalyzed by the basolateral anion antiporter, this bicarbonate ion is exported across the basolateral membrane (and ultimately into the blood) in exchange for a Cl - ion. The Cl - ions then exit through Cl - channels in the apical membrane, entering the stomach lumen. To preserve electroneutrality, each Cl - ion that moves into the stomach lumen across the apical membrane is accompanied by a K + ion that moves outward through a separate K + channel. In this way, the excess K + ions pumped inward by the H + /K + ATPase are returned to the stomach lumen, thus maintaining the normal intracellular K + concentration. The net result is secretion of equal amounts of H + and Cl - ions (i.e., HCl) into the stomach lumen, while the ph of the cytosol remains neutral and the excess OH - ions, as HCO3 -, are transported into the blood.
50


51 V-class H + pump lysosome & endosome 4 Figure Concentration of ions and sucrose by the plant vacuole. Figure Effect of V-class H + pumps on H + concentration gradients and electric potential gradients Figure Dissolution of bone by polarized osteoclast cells requires a V-class proton pump and the ClC-7 chloride channel protein.
52 Figure Effect of V-class H + pumps on H + concentration gradients and electric potential gradients across cellular
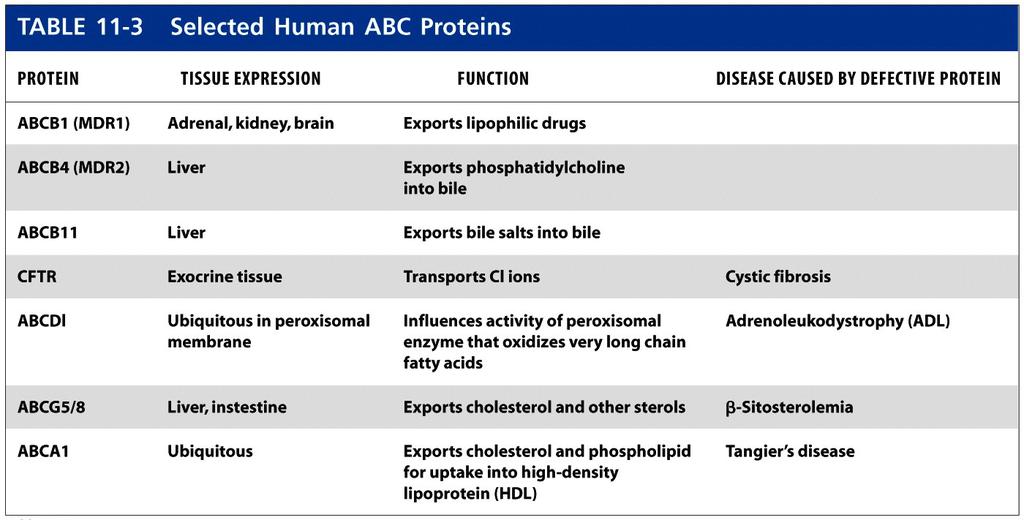
53 V-Class H ATPases Pump Protons Across Lysosomal and Vacuolar Membranes By themselves, ATP-powered proton pumps cannot acidify the lumen of an organelle (or the extracellular space) because these pumps are electrogenic; that is, a net movement of electric charge occurs during transport. Pumping of just a few protons causes a buildup of positively charged H+ ions on the exoplasmic (inside) face of the organelle membrane. For each H+ pumped across, a negative ion (e.g., OH- or Cl-) will be left behind on the cytosolic face, causing a buildup of negatively charged ions there. These oppositely charged ions attract each other on opposite faces of the membrane, generating a charge separation, or electric potential, across the membrane. The lysosome membrane thus functions as a capacitor in an electric circuit, storing opposing charges (anions and cations) on opposite sides of a barrier impermeable to movement of charged particles. As more and more protons are pumped and build up excess positive charge on the exoplasmic face, the energy required to move additional protons against this rising electric potential gradient increases dramatically and prevents pumping of additional protons long beforea significant transmembrane H+ concentration gradient is established (Figure 11-14a). In fact, this is the way that P-class H+ pumps generate a cytosol-negative potential across plant and yeast plasma membranes. In order for an organelle lumen or an extracellular space (e.g., the lumen of the stomach) to become acidic, movement of protons must be accompanied either by (1) movement of an equal number of anions (e.g., Cl-) in the same direction or by (2) movement of equal numbers of a different cation in the opposite direction. The first process occurs in lysosomes and plant vacuoles, whose membranes contain V-class H+ ATPases and anion channels through which accompanying Clions move (Figure 11-14b). The second process occurs in the lining of the stomach, which contains a P-class H+/K+ ATPase that is not electrogenic and pumps one H+ outward and one K+ inward. Operation of this pump is discussed later in the chapter. The ATP-powered proton pumps in lysosomal and vacuolar membranes have been solubilized, purified, and incorporated into liposomes. As shown in Figure 11-9, these V-class proton pumps contain two discrete domains: a cytosolic hydrophilic domain (V1) and a transmembrane domain (V0) with multiple subunits forming each domain. Binding and hydrolysis of ATP by the B subunits in V1 provide the energy for pumping of H+ ions through the proton-conducting channel formed by the c and a subunits in V0. Unlike P-class ion pumps, V-class proton pumps are not phosphorylated and dephosphorylated during proton transport. The structurally similar F-class proton pumps, which we describe in Chapter 12, normally operate in the reverse direction to generate ATP rather than pump protons; their structure and mechanism of action is understood in great detail.
54 Figure Concentration of ions and sucrose by the plant vacuole.

55 Figure Dissolution of bone by polarized osteoclast cells requires a V-class proton pump and the ClC-7 chloride channel protein.
56 V-class H + pump All V-class ATPase transport only H+ ions. These proton pumps, present in the membranes of lysosomes, endosomes, and plant vacuoles, function to acidify the lumen of these organelles. 1. If an intracellular organelle contains only V-class pumps, proton pumping generates an electric potential across the membrane (cytosol-facing side negative and luminal-side positive) but no significant change in the intraluminal ph. If the organelle membrane also contains Cl- channels, anions passively follow the pumped protons, resulting in an accumulation of H+ and Cl- ions in the lumen (low luminal ph) but no electric potential across the membrane (Figure 11-14). 2. The vacuolar membrane contains two types of proton pumps: a V-class H+ ATPase and a pyrophosphate-hydrolyzing proton pump that differs from all other ion transport proteins and probably is unique to plants. These pumps generate a low luminal ph as well as an inside-positive electric potential across the vacuolar membrane owing to the inward pumping of H+ ions. The inside-positive potential powers the movement of Cl- and NO3- from the cytosol through separate channel proteins. Proton antiporters powered by the H+ gradient, accumulate Na+, Ca2+, and sucrose inside the vacuole (Figure 11-29). 3. The osteoclast plasma membrane is divided into two domains separated by the tight seal between a ring of membrane and the bone surface. The membrane domain facing the bone contains V-class proton pumps and ClC-7 Cl- channels. The opposing membrane domain contains anion antiporters that exchange HCO3- and Cl- ions. The combined operation of these three transport proteins and carbonic anhydrase acidifies the enclosed space and allows bone resorption while maintaining the neutral ph of the cytosol (Figure 11-32).
57

58 F-class pump
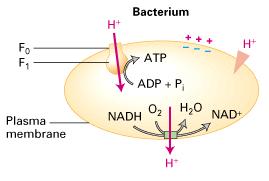
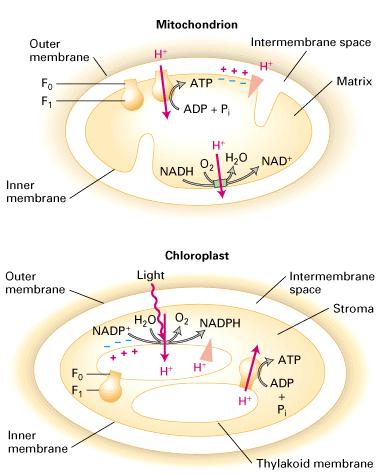
59 F-class pump 1. F-class pumps are found in bacterial plasma membranes and in mitochondria and chloroplasts. In contrast to V pumps, they generally function to power the synthesis of ATP from ADP and Pi by movement of protons from the exoplasmic to the cytosolic face of the membrane down the proton electrochemical gradient. Because of their importance in ATP synthesis in chloroplasts and mitochondria, F-class proton pumps, commonly called ATP synthases, are treated separately in Chapter. 2. In chemiosmosis, a proton-motive force generated by proton pumping across a membrane is used to power ATP synthesis. The mechanism and membrane orientation of the process are similar in bacteria, mitochondria, and chloroplasts. 1 2 During electron transport, protons are always pumped from the cytosolic face to the exoplasmic face, creating a proton concentration gradient (exoplasmic face > cytosolic face) and an electric potential (negative cytosolic face and positive exoplasmic face) across the membrane. During the synthesis of ATP, protons flow in the reverse direction (down their electrochemical gradient) through ATP synthase (F0F1 complex), which protrudes in a knob at the cytosolic face in all cases.
60

61 ABC superfamily
62 ABC superfamily The final class of ATP-powered pumps contains more members and is more diverse than the other classes. Referred to as the ABC (ATP-binding cassette) superfamily, this class includes several hundred different transport proteins found in organisms ranging from bacteria to humans. Each ABC protein is specific for a single substrate or group of related substrates, which may be ions, sugars, amino acids, phospholipids, peptides, polysaccharides, or even proteins. All ABC transport proteins share a structural organization consisting of four core domains: two transmembrane (T) domains, forming the passageway through which transported molecules cross the membrane, and two cytosolic ATP-binding (A) domains. In some ABC proteins, mostly in bacteria, the core domains are present in four separate polypeptides; in others, the core domains are fused into one or two multidomain polypeptides (Table 11-3).
63
64 5 Active transport 1) primary active transport (ATP-powered pump) 2) secondary active transport (cotransport)
65 Secondary active transport: cotransport Energy 1) Na + motive force 2) H + motive force Transported molecules 1) Glucose 2) Amino acids 3) Ca 2+ Figure Transmembrane forces acting on Na + ions.

66 Figure Transcellular transport of glucose from the intestinal lumen into the blood. [Glucose] low high low
67 Cotransport 1. Cotransporters use the energy released by movement of an ion (usually H+ or Na+) down its electrochemical gradient to power the import or export of a small molecule or different ion against its concentration gradient. The cells lining the small intestine and kidney tubules express symport proteins that couple the energetically favorable entry of Na+ to the import of glucose and amino acids against their concentration gradients (Figure 11-25). 2. The Na+/K+ ATPase in the basolateral surface membrane generates Na+ and K+ concentration gradients (step1). The outward movement of K+ ions through nongated K+ channels generates an inside-negative membrane potential. Both the Na+ concentration gradient and the membrane potential are used to drive the uptake of glucose from the intestinal lumen by the two-na+/one-glucose symporter located in the apical surface membrane (step2). Glucose leaves the cell via facilitated diffusion catalyzed by GLUT2, a glucose uniporter located in the basolateral membrane (step3) (Figure 11-30).
68

69 Figure Operational model for the two-na + /one-glucose symporter.
70 Two-Na + /one-glucose symporter Thus by coupling the transport of two Na+ ions to the transport of one glucose, the two-na+/oneglucose symporter permits cell to accumulate a very high concentration of glucose relative to the external concentration. This means that glucose present even at very low concentrations in the lumen of the intestine or in the kidney tubules can be efficiently transported into the lining cells and not lost from the body. Simultaneous binding of Na+ and glucose to the conformation with outward-facing binding sites (step 1) causes a conformational change in the protein such that the bound substrates are transiently occluded, unable to dissociate into either medium (step 2). In step 3 the protein assumes a third conformation with inward-facing sites. Dissociation of the bound Na+ and glucose into the cytosol (step 4) allows the protein to revert to its original outward-facing conformation (step 5), ready to transport additional substrate (Figure 11-26).
71
72 ph regulation


73 ph and respiration Figure Carbon dioxide transport in blood requires a Cl /HCO 3 antiporter. 1. Erythroblast 2. Reticulocyte: 핵, 세포내소기관없음, 단백질합성기구존재 3. Erythrocyte: 단백질합성기구없음
74 ph regulation 1. Two cotransporters that are activated at low ph help maintain the cytosolic ph in animal cells very close to 7.4 despite metabolic production of carbonic and latic acids. 1 One, a Na+/H+ antiporter, exports excess protons. 2 The other, a Na+HCO3-/Cl- cotransporter, importers HCO3-, which dissociates in the cytosol to yield ph-raising OH- ions. 3 A Cl-/HCO3- antiporter that is activated at high ph functions to export HCO3- when the cytosolic ph rises above normal and causes a decrease in ph. AE1, a Cl-/HCO3- antiporter in the erythrocyte membrane, increases the ability of blood to transport CO2 from tissues to the lungs. 2. In systemic capillaries, carbon dioxide gas diffuses across the erythrocyte plasma membrane and is converted into soluble HCO3- by the enzyme carbonic anhydrase; at the same time, oxygen leaves the cell and hemoglobin binds a proton. The anion antiporter AE1 catalyzes the reversible exchange of Cland HCO3- ions across the membrane. The overall reaction causes HCO3- to be released from the cell, which is essential for maximal CO2 transport from the tissues to the lungs, and for maintaining ph neutrality in the blood cell. In the lungs, where carbon dioxide is excreted, the overall reaction is reversed (Figure 11-28).
Biochemistry. Biochemistry 9/20/ Bio-Energetics. 4.2) Transport of ions and small molecules across cell membranes
 Transport of ions and small molecules across cell membranes") 9/20/15 Biochemistry Biochemistry 4. Bio-Energetics 4.2) Transport of ions and small molecules across cell membranes Aquaporin, the water channel, consists of four identical transmembrane polypeptides
9/20/15 Biochemistry Biochemistry 4. Bio-Energetics 4.2) Transport of ions and small molecules across cell membranes Aquaporin, the water channel, consists of four identical transmembrane polypeptides
Biochemistry. Biochemistry 7/11/ Bio-Energetics. 4.2) Transport of ions and small molecules across cell membranes
 Transport of ions and small molecules across cell membranes") Biochemistry Biochemistry 4. Bio-Energetics 4.2) Transport of ions and small molecules across cell membranes Aquaporin, the water channel, consists of four identical transmembrane polypeptides Key Energy
Biochemistry Biochemistry 4. Bio-Energetics 4.2) Transport of ions and small molecules across cell membranes Aquaporin, the water channel, consists of four identical transmembrane polypeptides Key Energy
ACTIVE TRANSPORT AND GLUCOSE TRANSPORT. (Chapter 14 and 15, pp and pp )
") ACTIVE TRANSPORT AND GLUCOSE TRANSPORT (Chapter 14 and 15, pp 140-143 and pp 146-151) Overview Active transport is the movement of molecules across a cell membrane in the direction against their concentration
ACTIVE TRANSPORT AND GLUCOSE TRANSPORT (Chapter 14 and 15, pp 140-143 and pp 146-151) Overview Active transport is the movement of molecules across a cell membrane in the direction against their concentration
Membranes 2: Transportation
 Membranes 2: Transportation Steven E. Massey, Ph.D. Associate Professor Bioinformatics Department of Biology University of Puerto Rico Río Piedras Office & Lab: NCN#343B Tel: 787-764-0000 ext. 7798 E-mail:
Membranes 2: Transportation Steven E. Massey, Ph.D. Associate Professor Bioinformatics Department of Biology University of Puerto Rico Río Piedras Office & Lab: NCN#343B Tel: 787-764-0000 ext. 7798 E-mail:
CELL BIOLOGY - CLUTCH CH. 9 - TRANSPORT ACROSS MEMBRANES.
 !! www.clutchprep.com K + K + K + K + CELL BIOLOGY - CLUTCH CONCEPT: PRINCIPLES OF TRANSMEMBRANE TRANSPORT Membranes and Gradients Cells must be able to communicate across their membrane barriers to materials
!! www.clutchprep.com K + K + K + K + CELL BIOLOGY - CLUTCH CONCEPT: PRINCIPLES OF TRANSMEMBRANE TRANSPORT Membranes and Gradients Cells must be able to communicate across their membrane barriers to materials
Membrane transport 1. Summary
 Membrane transport 1. Summary A. Simple diffusion 1) Diffusion by electrochemical gradient no energy required 2) No channel or carrier (or transporter protein) is needed B. Passive transport (= Facilitated
Membrane transport 1. Summary A. Simple diffusion 1) Diffusion by electrochemical gradient no energy required 2) No channel or carrier (or transporter protein) is needed B. Passive transport (= Facilitated
Advanced Higher Biology. Unit 1- Cells and Proteins 2c) Membrane Proteins
 Membrane Proteins") Advanced Higher Biology Unit 1- Cells and Proteins 2c) Membrane Proteins Membrane Structure Phospholipid bilayer Transmembrane protein Integral protein Movement of Molecules Across Membranes Phospholipid
Advanced Higher Biology Unit 1- Cells and Proteins 2c) Membrane Proteins Membrane Structure Phospholipid bilayer Transmembrane protein Integral protein Movement of Molecules Across Membranes Phospholipid
TRANSPORT ACROSS MEMBRANE
 TRANSPORT ACROSS MEMBRANE The plasma membrane functions to isolate the inside of the cell from its environment, but isolation is not complete. A large number of molecules constantly transit between the
TRANSPORT ACROSS MEMBRANE The plasma membrane functions to isolate the inside of the cell from its environment, but isolation is not complete. A large number of molecules constantly transit between the
Lecture 3 13/11/2018
 Lecture 3 13/11/2018 1 Plasma membrane ALL cells have a cell membrane made of proteins and lipids. protein channel Cell Membrane Layer 1 Layer 2 lipid bilayer protein pump Lipid bilayer allows water, carbon
Lecture 3 13/11/2018 1 Plasma membrane ALL cells have a cell membrane made of proteins and lipids. protein channel Cell Membrane Layer 1 Layer 2 lipid bilayer protein pump Lipid bilayer allows water, carbon
Membrane Physiology. Dr. Hiwa Shafiq Oct-18 1
 Membrane Physiology Dr. Hiwa Shafiq 22-10-2018 29-Oct-18 1 Chemical compositions of extracellular and intracellular fluids. 29-Oct-18 2 Transport through the cell membrane occurs by one of two basic processes:
Membrane Physiology Dr. Hiwa Shafiq 22-10-2018 29-Oct-18 1 Chemical compositions of extracellular and intracellular fluids. 29-Oct-18 2 Transport through the cell membrane occurs by one of two basic processes:
Electrical Properties of the Membrane
 BIOE 2520 Electrical Properties of the Membrane Reading: Chapter 11 of Alberts et al. Stephen Smith, Ph.D. 433 Biotech Center shs46@pitt.edu Permeability of Lipid membrane Lipid bilayer is virtually impermeable
BIOE 2520 Electrical Properties of the Membrane Reading: Chapter 11 of Alberts et al. Stephen Smith, Ph.D. 433 Biotech Center shs46@pitt.edu Permeability of Lipid membrane Lipid bilayer is virtually impermeable
Chapter 10. Thermodynamics of Transport. Thermodynamics of Transport, con t. BCH 4053 Summer 2001 Chapter 10 Lecture Notes. Slide 1.
 BCH 4053 Summer 2001 Chapter 10 Lecture Notes 1 Chapter 10 Membrane Transport 2 3 Thermodynamics of Transport Free Energy change is given by difference in electrochemical potential and the quantity transported
BCH 4053 Summer 2001 Chapter 10 Lecture Notes 1 Chapter 10 Membrane Transport 2 3 Thermodynamics of Transport Free Energy change is given by difference in electrochemical potential and the quantity transported
CELL SIGNALLING and MEMBRANE TRANSPORT. Mark Louie D. Lopez Department of Biology College of Science Polytechnic University of the Philippines
 CELL SIGNALLING and MEMBRANE TRANSPORT Mark Louie D. Lopez Department of Biology College of Science Polytechnic University of the Philippines GENERIC SIGNALLING PATHWAY CELL RESPONSE TO SIGNALS CELL RESPONSE
CELL SIGNALLING and MEMBRANE TRANSPORT Mark Louie D. Lopez Department of Biology College of Science Polytechnic University of the Philippines GENERIC SIGNALLING PATHWAY CELL RESPONSE TO SIGNALS CELL RESPONSE
Active Transport * OpenStax. 1 Electrochemical Gradient
 OpenStax-CNX module: m44418 1 Active Transport * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 4.0 By the end of this section, you will be able
OpenStax-CNX module: m44418 1 Active Transport * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 4.0 By the end of this section, you will be able
Chem Lecture 9 Pumps and Channels Part 1
 Chem 45 - Lecture 9 Pumps and Channels Part 1 Question of the Day: What two factors about a molecule influence the change in its free energy as it moves across a membrane? Membrane proteins function as
Chem 45 - Lecture 9 Pumps and Channels Part 1 Question of the Day: What two factors about a molecule influence the change in its free energy as it moves across a membrane? Membrane proteins function as
Membrane Protein Pumps
 Membrane Protein Pumps Learning objectives You should be able to understand & discuss: Active transport-na + /K + ATPase ABC transporters Metabolite transport by lactose permease 1. Ion pumps: ATP-driven
Membrane Protein Pumps Learning objectives You should be able to understand & discuss: Active transport-na + /K + ATPase ABC transporters Metabolite transport by lactose permease 1. Ion pumps: ATP-driven
MEMBRANE STRUCTURE. Lecture 9. Biology Department Concordia University. Dr. S. Azam BIOL 266/
 MEMBRANE STRUCTURE Lecture 9 BIOL 266/4 2014-15 Dr. S. Azam Biology Department Concordia University RED BLOOD CELL MEMBRANE PROTEINS The Dynamic Nature of the Plasma Membrane SEM of human erythrocytes
MEMBRANE STRUCTURE Lecture 9 BIOL 266/4 2014-15 Dr. S. Azam Biology Department Concordia University RED BLOOD CELL MEMBRANE PROTEINS The Dynamic Nature of the Plasma Membrane SEM of human erythrocytes
Ch. 3: Cells & Their Environment
 Ch. 3: Cells & Their Environment OBJECTIVES: 1. Understand cell membrane permeability 2. To recognize different types of cellular transport (passive vs active) 3. To understand membrane potential and action
Ch. 3: Cells & Their Environment OBJECTIVES: 1. Understand cell membrane permeability 2. To recognize different types of cellular transport (passive vs active) 3. To understand membrane potential and action
Main idea of this lecture:
 Ac#ve Transport Main idea of this lecture: How do molecules, big and small, get in OR out of a cell? 2 Main ways: Passive Transport (Does not require energy) Lecture 1 Ac=ve Transport (Requires energy)
Ac#ve Transport Main idea of this lecture: How do molecules, big and small, get in OR out of a cell? 2 Main ways: Passive Transport (Does not require energy) Lecture 1 Ac=ve Transport (Requires energy)
Neurophysiology. Danil Hammoudi.MD
 Neurophysiology Danil Hammoudi.MD ACTION POTENTIAL An action potential is a wave of electrical discharge that travels along the membrane of a cell. Action potentials are an essential feature of animal
Neurophysiology Danil Hammoudi.MD ACTION POTENTIAL An action potential is a wave of electrical discharge that travels along the membrane of a cell. Action potentials are an essential feature of animal
Molecular Biology of the Cell
 Alberts Johnson Lewis Morgan Raff Roberts Walter Molecular Biology of the Cell Sixth Edition Chapter 11 Membrane Transport of Small Molecules and the Electrical Properties of Membranes Copyright Garland
Alberts Johnson Lewis Morgan Raff Roberts Walter Molecular Biology of the Cell Sixth Edition Chapter 11 Membrane Transport of Small Molecules and the Electrical Properties of Membranes Copyright Garland
Part I => CARBS and LIPIDS. 1.5 MEMBRANE TRANSPORT 1.5a Passive Transport 1.5b Facilitated Transport 1.5c Active Transport
 Part I => CARBS and LIPIDS 1.5 MEMBRANE TRANSPORT 1.5a Passive Transport 1.5b Facilitated Transport 1.5c Active Transport Section 1.5a: Passive Transport Synopsis 1.5a - Passive transport (or passive diffusion)
Part I => CARBS and LIPIDS 1.5 MEMBRANE TRANSPORT 1.5a Passive Transport 1.5b Facilitated Transport 1.5c Active Transport Section 1.5a: Passive Transport Synopsis 1.5a - Passive transport (or passive diffusion)
Energy and Cells. Appendix 1. The two primary energy transformations in plants are photosynthesis and respiration.
 Energy and Cells Appendix 1 Energy transformations play a key role in all physical and chemical processes that occur in plants. Energy by itself is insufficient to drive plant growth and development. Enzymes
Energy and Cells Appendix 1 Energy transformations play a key role in all physical and chemical processes that occur in plants. Energy by itself is insufficient to drive plant growth and development. Enzymes
Dr. Ketki Assistant Professor Department of Biochemistry Heritage IMS, Varanasi
 TRANSPORT MECHANISMS Dr. Ketki Assistant Professor Department of Biochemistry Heritage IMS, Varanasi Membrane selectivity allows adjustments of cell composition and function If plasma membrane is relatively
TRANSPORT MECHANISMS Dr. Ketki Assistant Professor Department of Biochemistry Heritage IMS, Varanasi Membrane selectivity allows adjustments of cell composition and function If plasma membrane is relatively
Renal handling of substances. Dr.Charushila Rukadikar Assistance Professor Physiology
 Renal handling of substances Dr.Charushila Rukadikar Assistance Professor Physiology GENERAL PRINCIPLES OF RENAL TUBULAR TRANSPORT Transport mechanisms across cell membrane 1) Passive transport i. Diffusion
Renal handling of substances Dr.Charushila Rukadikar Assistance Professor Physiology GENERAL PRINCIPLES OF RENAL TUBULAR TRANSPORT Transport mechanisms across cell membrane 1) Passive transport i. Diffusion
Membrane Protein Channels
 Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
b) What is the gradient at room temperature? Du = J/molK * 298 K * ln (1/1000) = kj/mol
 What is the gradient at room temperature? Du = J/molK * 298 K * ln (1/1000) = kj/mol") Chem350 Practice Problems Membranes 1. a) What is the chemical potential generated by the movement of glucose by passive diffusion established by a 1000 fold concentration gradient at physiological temperature?
Chem350 Practice Problems Membranes 1. a) What is the chemical potential generated by the movement of glucose by passive diffusion established by a 1000 fold concentration gradient at physiological temperature?
(Be sure to clearly state the principles addressed in your discussion.)
") CELL QUESTION 1992: AP BIOLOGY A laboratory assistant prepared solutions of 0.8 M, 0.6 M, 0.4 M, and 0.2 M sucrose, but forgot to label them. After realizing the error, the assistant randomly labeled the
CELL QUESTION 1992: AP BIOLOGY A laboratory assistant prepared solutions of 0.8 M, 0.6 M, 0.4 M, and 0.2 M sucrose, but forgot to label them. After realizing the error, the assistant randomly labeled the
f) Adding an enzyme does not change the Gibbs free energy. It only increases the rate of the reaction by lowering the activation energy.
 Adding an enzyme does not change the Gibbs free energy. It only increases the rate of the reaction by lowering the activation energy.") Problem Set 2-Answer Key BILD1 SP16 1) How does an enzyme catalyze a chemical reaction? Define the terms and substrate and active site. An enzyme lowers the energy of activation so the reaction proceeds
Problem Set 2-Answer Key BILD1 SP16 1) How does an enzyme catalyze a chemical reaction? Define the terms and substrate and active site. An enzyme lowers the energy of activation so the reaction proceeds
Transporters and Membrane Motors Nov 15, 2007
 BtuB OM vitamin B12 transporter F O F 1 ATP synthase Human multiple drug resistance transporter P-glycoprotein Transporters and Membrane Motors Nov 15, 2007 Transport and membrane motors Concentrations
BtuB OM vitamin B12 transporter F O F 1 ATP synthase Human multiple drug resistance transporter P-glycoprotein Transporters and Membrane Motors Nov 15, 2007 Transport and membrane motors Concentrations
Chapter 3 Part 1! 10 th ed.: pp ! 11 th ed.: pp !! Cellular Transport Mechanisms! The Cell Cycle!
 Chapter 3 Part 1! 10 th ed.: pp. 87 105! 11 th ed.: pp. 90 107!! Cellular Transport Mechanisms! The Cell Cycle! Transport Processes: Passive and Active (1 of 2)! 1. Passive transport! Does not use ATP!
Chapter 3 Part 1! 10 th ed.: pp. 87 105! 11 th ed.: pp. 90 107!! Cellular Transport Mechanisms! The Cell Cycle! Transport Processes: Passive and Active (1 of 2)! 1. Passive transport! Does not use ATP!
Chapter 3 Part 1! 10 th ed.: pp ! 11 th ed.: pp !! Cellular Transport Mechanisms! The Cell Cycle!
 Chapter 3 Part 1! 10 th ed.: pp. 87 105! 11 th ed.: pp. 90 107!! Cellular Transport Mechanisms! The Cell Cycle! Transport Processes: Passive and Active (1 of 2)! 1. Passive transport! Does not use ATP!
Chapter 3 Part 1! 10 th ed.: pp. 87 105! 11 th ed.: pp. 90 107!! Cellular Transport Mechanisms! The Cell Cycle! Transport Processes: Passive and Active (1 of 2)! 1. Passive transport! Does not use ATP!
Cellular Transport. 1. Transport to and across the membrane 1a. Transport of small molecules and ions 1b. Transport of proteins
 Transport Processes Cellular Transport 1. Transport to and across the membrane 1a. Transport of small molecules and ions 1b. Transport of proteins 2. Vesicular transport 3. Transport through the nuclear
Transport Processes Cellular Transport 1. Transport to and across the membrane 1a. Transport of small molecules and ions 1b. Transport of proteins 2. Vesicular transport 3. Transport through the nuclear
Biochemistry Prof. S. Dasgupta Department of Chemistry. Indian Institute of Technology Kharagpur. Lecture - 15 Nucleic Acids III
 Biochemistry Prof. S. Dasgupta Department of Chemistry. Indian Institute of Technology Kharagpur Lecture - 15 Nucleic Acids III In the last two classes we spoke about lipids and membranes. Now, what we
Biochemistry Prof. S. Dasgupta Department of Chemistry. Indian Institute of Technology Kharagpur Lecture - 15 Nucleic Acids III In the last two classes we spoke about lipids and membranes. Now, what we
Cells. A. The iodine diffused into the bag. B. The starch was changed to sugar.
 Name: Date: 1. A student filled a bag of dialysis tubing with a milky-white starch solution and placed the bag in a beaker of iodine-water as shown in the diagram. An hour later, the student observed that
Name: Date: 1. A student filled a bag of dialysis tubing with a milky-white starch solution and placed the bag in a beaker of iodine-water as shown in the diagram. An hour later, the student observed that
Basic Chemistry. Chemistry Review. Bio 250: Anatomy & Physiology
 Basic Chemistry Bio 250: Anatomy & Physiology Chemistry Review It is going to be your responsibility to review the basic principles of chemistry you learned in BIO 101 This basic set of notes will help
Basic Chemistry Bio 250: Anatomy & Physiology Chemistry Review It is going to be your responsibility to review the basic principles of chemistry you learned in BIO 101 This basic set of notes will help
6 Mechanotransduction. rotation
 rotation inflow outflow Figure 6.3: Circumferential and uniaxial flow devices applying shear stress to the cell culture. They are stimulated through a circumferential fluid flow generating by a rotating
rotation inflow outflow Figure 6.3: Circumferential and uniaxial flow devices applying shear stress to the cell culture. They are stimulated through a circumferential fluid flow generating by a rotating
Chapt. 12, Movement Across Membranes. Chapt. 12, Movement through lipid bilayer. Chapt. 12, Movement through lipid bilayer
 Chapt. 12, Movement Across Membranes Two ways substances can cross membranes Passing through the lipid bilayer Passing through the membrane as a result of specialized proteins 1 Chapt. 12, Movement through
Chapt. 12, Movement Across Membranes Two ways substances can cross membranes Passing through the lipid bilayer Passing through the membrane as a result of specialized proteins 1 Chapt. 12, Movement through
2. In regards to the fluid mosaic model, which of the following is TRUE?
 General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
Chapter 7-3 Cells and Their Environment
 Chapter 7-3 Cells and Their Environment 7-3 Passive Transport Passive transport-the movement of substances across the cell membrane without using NRG Concentration Gradient-difference in concentration
Chapter 7-3 Cells and Their Environment 7-3 Passive Transport Passive transport-the movement of substances across the cell membrane without using NRG Concentration Gradient-difference in concentration
Basic Chemistry. Chapter 2 BIOL1000 Dr. Mohamad H. Termos
 Basic Chemistry Chapter 2 BIOL1000 Dr. Mohamad H. Termos Chapter 2 Objectives Following this chapter, you should be able to describe: - Atoms, molecules, and ions - Composition and properties - Types of
Basic Chemistry Chapter 2 BIOL1000 Dr. Mohamad H. Termos Chapter 2 Objectives Following this chapter, you should be able to describe: - Atoms, molecules, and ions - Composition and properties - Types of
3.1 Cell Theory. KEY CONCEPT Cells are the Basic unit of life.
 3.1 Cell Theory KEY CONCEPT Cells are the Basic unit of life. 3.1 Cell Theory The cell theory grew out of the work of many scientists and improvements in the microscope. Many scientists contributed to
3.1 Cell Theory KEY CONCEPT Cells are the Basic unit of life. 3.1 Cell Theory The cell theory grew out of the work of many scientists and improvements in the microscope. Many scientists contributed to
2002NSC Human Physiology Semester Summary
 2002NSC Human Physiology Semester Summary Griffith University, Nathan Campus Semester 1, 2014 Topics include: - Diffusion, Membranes & Action Potentials - Fundamentals of the Nervous System - Neuroanatomy
2002NSC Human Physiology Semester Summary Griffith University, Nathan Campus Semester 1, 2014 Topics include: - Diffusion, Membranes & Action Potentials - Fundamentals of the Nervous System - Neuroanatomy
Pathways that Harvest and Store Chemical Energy
 6 Pathways that Harvest and Store Chemical Energy Energy is stored in chemical bonds and can be released and transformed by metabolic pathways. Chemical energy available to do work is termed free energy
6 Pathways that Harvest and Store Chemical Energy Energy is stored in chemical bonds and can be released and transformed by metabolic pathways. Chemical energy available to do work is termed free energy
BIOLOGY 10/11/2014. An Introduction to Metabolism. Outline. Overview: The Energy of Life
 8 An Introduction to Metabolism CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes
8 An Introduction to Metabolism CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes
Class Work 31. Describe the function of the Golgi apparatus? 32. How do proteins travel from the E.R. to the Golgi apparatus? 33. After proteins are m
 Eukaryotes Class Work 1. What does the word eukaryote mean? 2. What is the one major difference between eukaryotes and prokaryotes? 3. List the different kingdoms of the eukaryote domain in the order in
Eukaryotes Class Work 1. What does the word eukaryote mean? 2. What is the one major difference between eukaryotes and prokaryotes? 3. List the different kingdoms of the eukaryote domain in the order in
The following question(s) were incorrectly answered.
 were incorrectly answered.") Name: Marcie Joseph Module: Cells & chemistry Test topic/animation: My animations/all animations Test type: Multiple choice Score: 48/50 Percent correct: 96% The following question(s) were incorrectly
Name: Marcie Joseph Module: Cells & chemistry Test topic/animation: My animations/all animations Test type: Multiple choice Score: 48/50 Percent correct: 96% The following question(s) were incorrectly
لجنة الطب البشري رؤية تنير دروب تميزكم
 1) Hyperpolarization phase of the action potential: a. is due to the opening of voltage-gated Cl channels. b. is due to prolonged opening of voltage-gated K + channels. c. is due to closure of the Na +
1) Hyperpolarization phase of the action potential: a. is due to the opening of voltage-gated Cl channels. b. is due to prolonged opening of voltage-gated K + channels. c. is due to closure of the Na +
Biology Midterm Review
 Biology Midterm Review Unit 1 Keystone Objectives: A.1.1, A.1.2, B.4.1.1 1.1 Biology explores life from the global to the microscopic level. Put the levels of organization in order, starting with subatomic
Biology Midterm Review Unit 1 Keystone Objectives: A.1.1, A.1.2, B.4.1.1 1.1 Biology explores life from the global to the microscopic level. Put the levels of organization in order, starting with subatomic
Aerobic Cellular Respiration
 Aerobic Cellular Respiration Under aerobic conditions (oxygen gas is available), cells will undergo aerobic cellular respiration. The end products of aerobic cellular respiration are carbon dioxide gas,
Aerobic Cellular Respiration Under aerobic conditions (oxygen gas is available), cells will undergo aerobic cellular respiration. The end products of aerobic cellular respiration are carbon dioxide gas,
I. MEMBRANE POTENTIALS
 I. MEMBRANE POTENTIALS Background to Nerve Impulses We have all heard that nerve impulses are electrical impulses. Stimuli at one end of a nerve cell are communicated to the far end of the nerve cell through
I. MEMBRANE POTENTIALS Background to Nerve Impulses We have all heard that nerve impulses are electrical impulses. Stimuli at one end of a nerve cell are communicated to the far end of the nerve cell through
AP Biology Chapter 36
 Chapter 36 Chapter 36 Transport in Plants 2006-2007 Transport in plants - Overview H2O & minerals transport in xylem transpiration evaporation, adhesion & cohesion negative pressure Sugars transport in
Chapter 36 Chapter 36 Transport in Plants 2006-2007 Transport in plants - Overview H2O & minerals transport in xylem transpiration evaporation, adhesion & cohesion negative pressure Sugars transport in
9/25/2011. Outline. Overview: The Energy of Life. I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V.
 Chapter 8 Introduction to Metabolism Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes Overview: The Energy of Life Figure 8.1 The living cell is a miniature
Chapter 8 Introduction to Metabolism Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes Overview: The Energy of Life Figure 8.1 The living cell is a miniature
Contains ribosomes attached to the endoplasmic reticulum. Genetic material consists of linear chromosomes. Diameter of the cell is 1 m
 1. (a) Complete each box in the table, which compares a prokaryotic and a eukaryotic cell, with a tick if the statement is correct or a cross if it is incorrect. Prokaryotic cell Eukaryotic cell Contains
1. (a) Complete each box in the table, which compares a prokaryotic and a eukaryotic cell, with a tick if the statement is correct or a cross if it is incorrect. Prokaryotic cell Eukaryotic cell Contains
MOLECULAR CELL BIOLOGY
 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
Cellular Respiration: Harvesting Chemical Energy. 9.1 Catabolic pathways yield energy by oxidizing organic fuels
 Cellular Respiration: Harvesting Chemical Energy 9.1 Catabolic pathways yield energy by oxidizing organic fuels 9.2 Glycolysis harvests chemical energy by oxidizing glucose to pyruvate 9.3 The citric acid
Cellular Respiration: Harvesting Chemical Energy 9.1 Catabolic pathways yield energy by oxidizing organic fuels 9.2 Glycolysis harvests chemical energy by oxidizing glucose to pyruvate 9.3 The citric acid
Life is Cellular Section 7.1
 Life is Cellular Section 7.1 Objectives Understand Cell theory Distinguish between prokaryotes and eukaryotes Understand different types of microscopy, and how they work in more detail What is a Cell?
Life is Cellular Section 7.1 Objectives Understand Cell theory Distinguish between prokaryotes and eukaryotes Understand different types of microscopy, and how they work in more detail What is a Cell?
Chapter Cells and the Flow of Energy A. Forms of Energy 1. Energy is capacity to do work; cells continually use energy to develop, grow,
 Chapter 6 6.1 Cells and the Flow of Energy A. Forms of Energy 1. Energy is capacity to do work; cells continually use energy to develop, grow, repair, reproduce, etc. 2. Kinetic energy is energy of motion;
Chapter 6 6.1 Cells and the Flow of Energy A. Forms of Energy 1. Energy is capacity to do work; cells continually use energy to develop, grow, repair, reproduce, etc. 2. Kinetic energy is energy of motion;
Transport of glucose across epithelial cells: a. Gluc/Na cotransport; b. Gluc transporter Alberts
 Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Name # Class Date Regents Review: Cells & Cell Transport
 Name # Class Date Regents Review: Cells & Cell Transport 1. All of the following are true regarding cells except? A) All cells have genetic material B) All cells have cell walls C) All cells have plasma
Name # Class Date Regents Review: Cells & Cell Transport 1. All of the following are true regarding cells except? A) All cells have genetic material B) All cells have cell walls C) All cells have plasma
Lectures by Kathleen Fitzpatrick
 Chapter 10 Chemotrophic Energy Metabolism: Aerobic Respiration Lectures by Kathleen Fitzpatrick Simon Fraser University Figure 10-1 Figure 10-6 Conversion of pyruvate The conversion of pyruvate to acetyl
Chapter 10 Chemotrophic Energy Metabolism: Aerobic Respiration Lectures by Kathleen Fitzpatrick Simon Fraser University Figure 10-1 Figure 10-6 Conversion of pyruvate The conversion of pyruvate to acetyl
Name: TF: Section Time: LS1a ICE 5. Practice ICE Version B
 Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
Cell Organelles. a review of structure and function
 Cell Organelles a review of structure and function TEKS and Student Expectations (SE s) B.4 Science concepts. The student knows that cells are the basic structures of all living things with specialized
Cell Organelles a review of structure and function TEKS and Student Expectations (SE s) B.4 Science concepts. The student knows that cells are the basic structures of all living things with specialized
Questions: Properties of excitable tissues Transport across cell membrane Resting potential Action potential Excitability change at excitation
 Questions: Properties of excitable tissues Transport across cell membrane Resting potential Action potential Excitability change at excitation EXCITABLE TISSUES The tissues can change the properties under
Questions: Properties of excitable tissues Transport across cell membrane Resting potential Action potential Excitability change at excitation EXCITABLE TISSUES The tissues can change the properties under
REVIEW 2: CELLS & CELL COMMUNICATION. A. Top 10 If you learned anything from this unit, you should have learned:
 Name AP Biology REVIEW 2: CELLS & CELL COMMUNICATION A. Top 10 If you learned anything from this unit, you should have learned: 1. Prokaryotes vs. eukaryotes No internal membranes vs. membrane-bound organelles
Name AP Biology REVIEW 2: CELLS & CELL COMMUNICATION A. Top 10 If you learned anything from this unit, you should have learned: 1. Prokaryotes vs. eukaryotes No internal membranes vs. membrane-bound organelles
THE CELL 3/15/15 HUMAN ANATOMY AND PHYSIOLOGY I THE CELLULAR BASIS OF LIFE
 HUMAN ANATOMY AND PHYSIOLOGY I Lecture: M 6-9:30 Randall Visitor Center Lab: W 6-9:30 Swatek Anatomy Center, Centennial Complex Required Text: Marieb 9 th edition Dr. Trevor Lohman DPT (949) 246-5357 tlohman@llu.edu
HUMAN ANATOMY AND PHYSIOLOGY I Lecture: M 6-9:30 Randall Visitor Center Lab: W 6-9:30 Swatek Anatomy Center, Centennial Complex Required Text: Marieb 9 th edition Dr. Trevor Lohman DPT (949) 246-5357 tlohman@llu.edu
An Introduction to Metabolism
 Chapter 8 1 An Introduction to Metabolism PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
Chapter 8 1 An Introduction to Metabolism PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
Multiple Choice Identify the letter of the choice that best completes the statement or answers the question.
 Exam 1 Multiple Choice Identify the letter of the choice that best completes the statement or answers the question. 1. The smallest units of life in all living things are a. cells. c. cytoplasm. b. mitochondria.
Exam 1 Multiple Choice Identify the letter of the choice that best completes the statement or answers the question. 1. The smallest units of life in all living things are a. cells. c. cytoplasm. b. mitochondria.
Chapter 2 Cellular Homeostasis and Membrane Potential
 Chapter 2 Cellular Homeostasis and Membrane Potential 2.1 Membrane Structure and Composition The human cell can be considered to consist of a bag of fluid with a wall that separates the internal, or intracellular,
Chapter 2 Cellular Homeostasis and Membrane Potential 2.1 Membrane Structure and Composition The human cell can be considered to consist of a bag of fluid with a wall that separates the internal, or intracellular,
- the flow of electrical charge from one point to the other is current.
 Biology 325, Fall 2004 Resting membrane potential I. Introduction A. The body and electricity, basic principles - the body is electrically neutral (total), however there are areas where opposite charges
Biology 325, Fall 2004 Resting membrane potential I. Introduction A. The body and electricity, basic principles - the body is electrically neutral (total), however there are areas where opposite charges
Hole s Human Anatomy and Physiology Tenth Edition. Chapter 2
 PowerPoint Lecture Outlines to accompany Hole s Human Anatomy and Physiology Tenth Edition Shier w Butler w Lewis Chapter 2 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction
PowerPoint Lecture Outlines to accompany Hole s Human Anatomy and Physiology Tenth Edition Shier w Butler w Lewis Chapter 2 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction
The diagram below represents levels of organization within a cell of a multicellular organism.
 STATION 1 1. Unlike prokaryotic cells, eukaryotic cells have the capacity to a. assemble into multicellular organisms b. establish symbiotic relationships with other organisms c. obtain energy from the
STATION 1 1. Unlike prokaryotic cells, eukaryotic cells have the capacity to a. assemble into multicellular organisms b. establish symbiotic relationships with other organisms c. obtain energy from the
CELL BIOLOGY. Which of the following cell structures does not have membranes? A. Ribosomes B. Mitochondria C. Chloroplasts D.
 1 CELL BIOLOGY PROKARYOTIC and EUKARYOTIC SP/1. SP/2. SP/4. Plant and animal cells both have A. ribosomes, cell walls and mitochondria. B. Golgi apparatus, chromosomes and mitochondria. C. Golgi apparatus,
1 CELL BIOLOGY PROKARYOTIC and EUKARYOTIC SP/1. SP/2. SP/4. Plant and animal cells both have A. ribosomes, cell walls and mitochondria. B. Golgi apparatus, chromosomes and mitochondria. C. Golgi apparatus,
Chapter 5. The Working Cell. PowerPoint Lectures for Campbell Biology: Concepts & Connections, Seventh Edition Reece, Taylor, Simon, and Dickey
 Chapter 5 The Working Cell PowerPoint Lectures for Campbell Biology: Concepts & Connections, Seventh Edition Reece, Taylor, Simon, and Dickey 2012 Pearson Education, Inc. Lecture by Edward J. Zalisko Introduction
Chapter 5 The Working Cell PowerPoint Lectures for Campbell Biology: Concepts & Connections, Seventh Edition Reece, Taylor, Simon, and Dickey 2012 Pearson Education, Inc. Lecture by Edward J. Zalisko Introduction
(Na++ K +)-ATPase in artificial lipid vesicles: influence of the concentration of mono- and divalent cations on the pumping rate
-ATPase in artificial lipid vesicles: influence of the concentration of mono- and divalent cations on the pumping rate") 254 Biochimica et Biophysica Acta 862 (1986) 254-264 Elsevier BBA 72961 (Na++ K +)-ATPase in artificial lipid vesicles: influence of the concentration of mono- and divalent cations on the pumping rate
254 Biochimica et Biophysica Acta 862 (1986) 254-264 Elsevier BBA 72961 (Na++ K +)-ATPase in artificial lipid vesicles: influence of the concentration of mono- and divalent cations on the pumping rate
Channels can be activated by ligand-binding (chemical), voltage change, or mechanical changes such as stretch.
, voltage change, or mechanical changes such as stretch.") 1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
Chapter 5. The Working Cell. PowerPoint Lectures for Campbell Biology: Concepts & Connections, Seventh Edition Reece, Taylor, Simon, and Dickey
 Chapter 5 The Working Cell PowerPoint Lectures for Campbell Biology: Concepts & Connections, Seventh Edition Reece, Taylor, Simon, and Dickey Lecture by Edward J. Zalisko Lesson Plans Flipped Classroom
Chapter 5 The Working Cell PowerPoint Lectures for Campbell Biology: Concepts & Connections, Seventh Edition Reece, Taylor, Simon, and Dickey Lecture by Edward J. Zalisko Lesson Plans Flipped Classroom
NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic
 NEUROPHYSIOLOGY NOTES L1 WHAT IS NEUROPHYSIOLOGY? NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic receptor
NEUROPHYSIOLOGY NOTES L1 WHAT IS NEUROPHYSIOLOGY? NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic receptor
Introduction to electrophysiology. Dr. Tóth András
 Introduction to electrophysiology Dr. Tóth András Topics Transmembran transport Donnan equilibrium Resting potential Ion channels Local and action potentials Intra- and extracellular propagation of the
Introduction to electrophysiology Dr. Tóth András Topics Transmembran transport Donnan equilibrium Resting potential Ion channels Local and action potentials Intra- and extracellular propagation of the
Unit 2: Cellular Chemistry, Structure, and Physiology Module 4: Cellular Physiology
 Unit 2: Cellular Chemistry, Structure, and Physiology Module 4: Cellular Physiology NC Essential Standard: 1.2.1 Explain how homeostasis is maintained in a cell and within an organism in various environments
Unit 2: Cellular Chemistry, Structure, and Physiology Module 4: Cellular Physiology NC Essential Standard: 1.2.1 Explain how homeostasis is maintained in a cell and within an organism in various environments
An Introduction to Metabolism
 Chapter 8 An Introduction to Metabolism Dr. Wendy Sera Houston Community College Biology 1406 Key Concepts in Chapter 8 1. An organism s metabolism transforms matter and energy, subject to the laws of
Chapter 8 An Introduction to Metabolism Dr. Wendy Sera Houston Community College Biology 1406 Key Concepts in Chapter 8 1. An organism s metabolism transforms matter and energy, subject to the laws of
Biology September 2015 Exam One FORM G KEY
 Biology 251 17 September 2015 Exam One FORM G KEY PRINT YOUR NAME AND ID NUMBER in the space that is provided on the answer sheet, and then blacken the letter boxes below the corresponding letters of your
Biology 251 17 September 2015 Exam One FORM G KEY PRINT YOUR NAME AND ID NUMBER in the space that is provided on the answer sheet, and then blacken the letter boxes below the corresponding letters of your
Biology September 2015 Exam One FORM W KEY
 Biology 251 17 September 2015 Exam One FORM W KEY PRINT YOUR NAME AND ID NUMBER in the space that is provided on the answer sheet, and then blacken the letter boxes below the corresponding letters of your
Biology 251 17 September 2015 Exam One FORM W KEY PRINT YOUR NAME AND ID NUMBER in the space that is provided on the answer sheet, and then blacken the letter boxes below the corresponding letters of your
2.6 The Membrane Potential
 2.6: The Membrane Potential 51 tracellular potassium, so that the energy stored in the electrochemical gradients can be extracted. Indeed, when this is the case experimentally, ATP is synthesized from
2.6: The Membrane Potential 51 tracellular potassium, so that the energy stored in the electrochemical gradients can be extracted. Indeed, when this is the case experimentally, ATP is synthesized from
Unit 2: Cells Guided Reading Questions (55 pts total)
") Name: AP Biology Biology, Campbell and Reece, 7th Edition Adapted from chapter reading guides originally created by Lynn Miriello Chapter 6 A Tour of the Cell Unit 2: Cells Guided Reading Questions (55
Name: AP Biology Biology, Campbell and Reece, 7th Edition Adapted from chapter reading guides originally created by Lynn Miriello Chapter 6 A Tour of the Cell Unit 2: Cells Guided Reading Questions (55
Cells have an unequal distribution of charge across their membrane: more postiive charges on the outside; more negative charges on the inside.
 Resting Membrane potential (V m ) or RMP Many cells have a membrane potential (Vm) that can be measured from an electrode in the cell with a voltmeter. neurons, muscle cells, heart cells, endocrine cells...
Resting Membrane potential (V m ) or RMP Many cells have a membrane potential (Vm) that can be measured from an electrode in the cell with a voltmeter. neurons, muscle cells, heart cells, endocrine cells...
CELLS NOT YOUR CELL PHONE HOMEOSTASIS: LESSON 5 OVERVIEW TEKS
 Lesson 5: Active Transport Protein Pumps Objectives: In this lesson the student will: CELLS NOT YOUR CELL PHONE HOMEOSTASIS: LESSON 5 OVERVIEW 1. Identify how the unique structure of the cell membrane
Lesson 5: Active Transport Protein Pumps Objectives: In this lesson the student will: CELLS NOT YOUR CELL PHONE HOMEOSTASIS: LESSON 5 OVERVIEW 1. Identify how the unique structure of the cell membrane
An Introduction to Metabolism
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 8 An Introduction to Metabolism
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 8 An Introduction to Metabolism
Chapter 3: Cells. Lectures by Mark Manteuffel, St. Louis Community College
 Chapter 3: Cells Lectures by Mark Manteuffel, St. Louis Community College Learning Objectives Be able to describe: what a cell is & two main classes of cells. structure & functions of cell membranes. how
Chapter 3: Cells Lectures by Mark Manteuffel, St. Louis Community College Learning Objectives Be able to describe: what a cell is & two main classes of cells. structure & functions of cell membranes. how
Honors Biology-CW/HW Cell Biology 2018
 Class: Date: Honors Biology-CW/HW Cell Biology 2018 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Hooke s discovery of cells was made observing a. living
Class: Date: Honors Biology-CW/HW Cell Biology 2018 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Hooke s discovery of cells was made observing a. living
2. Cellular and Molecular Biology
 2. Cellular and Molecular Biology 2.1 Cell Structure 2.2 Transport Across Cell Membranes 2.3 Cellular Metabolism 2.4 DNA Replication 2.5 Cell Division 2.6 Biosynthesis 2.1 Cell Structure What is a cell?
2. Cellular and Molecular Biology 2.1 Cell Structure 2.2 Transport Across Cell Membranes 2.3 Cellular Metabolism 2.4 DNA Replication 2.5 Cell Division 2.6 Biosynthesis 2.1 Cell Structure What is a cell?
Energy Transformation. Metabolism = total chemical reactions in cells.
 Energy Transformation Metabolism = total chemical reactions in cells. metabole = change Metabolism is concerned with managing the material and energy resources of the cell -Catabolism -Anabolism -Catabolism
Energy Transformation Metabolism = total chemical reactions in cells. metabole = change Metabolism is concerned with managing the material and energy resources of the cell -Catabolism -Anabolism -Catabolism
An Introduction to Metabolism
 CAMPBELL BIOLOGY IN FOCUS Urry Cain Wasserman Minorsky Jackson Reece 6 An Introduction to Metabolism Lecture Presentations by Kathleen Fitzpatrick and Nicole Tunbridge Overview: The Energy of Life The
CAMPBELL BIOLOGY IN FOCUS Urry Cain Wasserman Minorsky Jackson Reece 6 An Introduction to Metabolism Lecture Presentations by Kathleen Fitzpatrick and Nicole Tunbridge Overview: The Energy of Life The
Unit 2: Cells Guided Reading Questions (60 pts total)
") Name: AP Biology Biology, Campbell and Reece, 7th Edition Adapted from chapter reading guides originally created by Lynn Miriello Chapter 6 A Tour of the Cell Unit 2: Cells Guided Reading Questions (60
Name: AP Biology Biology, Campbell and Reece, 7th Edition Adapted from chapter reading guides originally created by Lynn Miriello Chapter 6 A Tour of the Cell Unit 2: Cells Guided Reading Questions (60
Organelles & Cells Student Edition. A. chromosome B. gene C. mitochondrion D. vacuole
 Name: Date: 1. Which structure is outside the nucleus of a cell and contains DNA? A. chromosome B. gene C. mitochondrion D. vacuole 2. A potato core was placed in a beaker of water as shown in the figure
Name: Date: 1. Which structure is outside the nucleus of a cell and contains DNA? A. chromosome B. gene C. mitochondrion D. vacuole 2. A potato core was placed in a beaker of water as shown in the figure
CELL PRACTICE TEST
 Name: Date: 1. As a human red blood cell matures, it loses its nucleus. As a result of this loss, a mature red blood cell lacks the ability to (1) take in material from the blood (2) release hormones to
Name: Date: 1. As a human red blood cell matures, it loses its nucleus. As a result of this loss, a mature red blood cell lacks the ability to (1) take in material from the blood (2) release hormones to
Overview of Cells. Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory
 Overview of Cells Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory Prokaryotic Cells Archaea Bacteria Come in many different shapes and sizes.5 µm 2 µm, up to 60 µm long Have large
Overview of Cells Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory Prokaryotic Cells Archaea Bacteria Come in many different shapes and sizes.5 µm 2 µm, up to 60 µm long Have large
Overview of ion channel proteins. What do ion channels do? Three important points:
 Overview of ion channel proteins Protein Structure Membrane proteins & channels Specific channels Several hundred distinct types Organization Evolution We need to consider 1. Structure 2. Functions 3.
Overview of ion channel proteins Protein Structure Membrane proteins & channels Specific channels Several hundred distinct types Organization Evolution We need to consider 1. Structure 2. Functions 3.
8/25/ Opening Questions: Are all living things made of cells? What are at least five things you know about cells?
 Chapter 3 The Cell: Module Hyperlinks 3.1 Cells are the fundamental units of life 3.2 Plant vs. animal cells 3.3 Membranes: structure 3.4 Membranes: function 3.5 The nucleus 3.6 Organelles in protein production
Chapter 3 The Cell: Module Hyperlinks 3.1 Cells are the fundamental units of life 3.2 Plant vs. animal cells 3.3 Membranes: structure 3.4 Membranes: function 3.5 The nucleus 3.6 Organelles in protein production
All organisms require a constant expenditure of energy to maintain the living state - "LIFE".
 CELLULAR RESPIRATION All organisms require a constant expenditure of energy to maintain the living state - "LIFE". Where does the energy come from and how is it made available for life? With rare exception,
CELLULAR RESPIRATION All organisms require a constant expenditure of energy to maintain the living state - "LIFE". Where does the energy come from and how is it made available for life? With rare exception,