The Atypical Cadherin Fat Directly Regulates Mitochondrial Function and Metabolic State
|
|
|
- Douglas Brooks
- 6 years ago
- Views:
Transcription
1 The Atypical Cadherin Fat Directly Regulates Mitochondrial Function and Metabolic State by Anson David Sing A thesis submitted in conformity with the requirements for the degree of Doctor of Philosophy Department of Molecular Genetics University of Toronto Copyright by Anson David Sing 2015
2 The Atypical Cadherin Fat Directly Regulates Mitochondrial Function and Metabolic State Abstract Anson David Sing Doctor of Philosophy Department of Molecular Genetics University of Toronto 2015 The atypical cadherin Fat plays essential roles during development to regulate tissue growth and patterning. It has also been implicated in a number of human diseases including polycystic kidney disease and some forms of cancer. Ft has been established as an input into the Hippo tumour suppressive pathway; however, the direct connections between Ft and its downstream effectors are not well understood. In this work, I have identified components of the ETC and mitochondrial import machinery as novel physical and genetic interactors of Ft. I found that mitochondrial complex I component, Ndufv2, can act in planar cell polarity and Hpo pathways. We have also carefully examined ft mutants and uncovered defects in mitochondrial morphology and complex I levels and activity that result in changes in metabolism. We outline a mechanism whereby Ft is cleaved at the cell surface, releasing a cytosolic fragment that is imported into the mitochondrial matrix to directly modulate mitochondrial function by assisting in the assembly or stability of mitochondrial complex I and V. This novel Ft function allows it to coordinate tissue growth and patterning with changing energy requirements during development, and may contribute to metabolic shifts observed in tumour progression. ii
3 Acknowledgments First and foremost, I would like to thank my supervisor Dr. Helen McNeill. From as early as my days as a roton in her lab she has always given me solid guidance and support. She has been a source of encouragement and her enthusiasm for research and scientific pursuit have been inspiring to me. Without the aid of her invaluable knowledge, experience, and advice, I would never have been able to find my way on this academic journey. I would also like to thank Helen for fostering a sense of camaraderie in the lab it was a place where I always felt comfortable sharing my joys and tribulations, scientific or otherwise. I would also like to thank my supervisory committee, Dr. CC Hui and Dr. Henry Krause. They have always challenged me to become a better scientist and have been there to keep me on track, headed towards my goals. I also want to thank them for contributing their considerable insight to the winding road that was my project. My many collaborators also need to be acknowledged. To Dr. Julie Brill and Dr. Lacramioara Fabian; Dr. Angus McQuibban and the many members of his lab that worked on this project, especially Mailis Bietenhader, Mauro Serrichio, and Riya Shanbhag; Dr. Andrea Jurisicova; Dr. Linda Penn and Dr. Peter Mullen; Dr. Marla Sokolowski I am indebted to them for all of their technical skill, assistance, scientific input and even commiseration, without which this work would not have been possible. My greatest appreciations go out to the McNeill lab members. They have made the lab feel like a second family, complete with lab mommy Yonit Tsatskis, without whom my project as well as regular lab operations would be dead in their tracks. A special acknowledgement goes to Dr. Robyn Rosenfeld, my screening buddy who started this entire endeavour together with me. I couldn t have asked for better people to spend my days with over these past years. Finally, I am forever grateful for those closest to me, my family. I would like to thank my parents, for their continuing love and encouragement. To my sister, Dr. Angela Sing, and other sister, Dr. JoE Yu, for blazing this trail for me to follow. They have truly been my unwavering supports whenever I needed them. And lastly to Stephen thank you for everything you are. iii
4 Table of Contents Acknowledgments Table of Contents List of Tables List of Figures List of Abbreviations iii iv vii viii x Chapter 1 1 Introduction Overview of the ft gene Discovery and phenotypes Ft and Ds are very large cadherins ft/ds and PCP regulation The core PCP genes The ft/ds system The Hippo pathway Vertebrate homologs of ft and human disease Mitochondria Oxidative phosphorylation Mitochondrial import Rationale and aims of thesis 24 Chapter 2 25 Identification and analysis of novel Ft interactors Introduction Materials and methods Fly stocks and genetics Tangential eye sections Wing hair analysis Antibodies Protein purification and GST pulldown Immunofluorescence EdU incorporation 28 iv
5 2.3 Results Yeast-2-Hybrid and RNAi screens mitochondrial targets identified Knockdown of several components of the mitochondrial OXPHOS system result in PCP defects, as well as broad deficits in normal tissue formation Characterizing and verifying specificity of the Ndufv2 RNAi phenotype Knockdown and overexpression of Ndufv2 in clones testing nonautonomy Direct physical interaction of Fat and mitochondrial complex proteins Genetic interactions between Ndufv2 and the ft pathway Discussion in vivo RNAi screen PCP regulators identified Establishing planar polarity a mitochondrial signal? PCP Targets under mitochondrial regulation Hpo pathway interactions with mitochondria Cryptic genetic interactions: ft and Ndufv2 77 Chapter 3 79 Ft and mitochondrial function Introduction Materials and methods Fly stocks and genetics Antibodies Cell culture and transfection Whole larvae imaging Drosophila testis preps Electron microscopy ROS levels measurements Complex I Activity Assay ATP assay Lactate assay Cell fractionation and mitochondrial protection assay Mitochondrial isolation and blue-native polyacrylamide gel electrophoresis (BN- PAGE) Mitochondrial in-gel activity assays Results 85 v
6 3.3.1 Growth of ft larvae is abnormal ft mutants show mitochondrial phenotypes Ft is cleaved to release a cytosolic fragment that is targeted to mitochondria ft mutants show loss of assembled CI holoenzyme Loss of Ft leads to defects in CV assembly The Ndufv2-binding region of Ft is necessary for normal larval growth and mitochondrial function Discussion Ft function in the mitochondria is separable from Hippo and PCP effects A rheostat model for ft function Fat cadherin Mitochondrial binding motifs and targeting sequences are ancient and conserved Ft mito and metabolism in cancer cells 115 Chapter Conclusion and future directions Summary of Thesis Results Mitochondrial components interact with ft and contribute to PCP and Hpo signaling (Chapter 2) ft is processed and targeted to mitochondria where it regulates OXPHOS and metabolic function (Chapter 3) Unresolved questions and future directions Mitochondria-mediated ft signaling The role of Ft in mitochondria Conservation of Ft mito Concluding remarks 126 References 127 vi
7 List of Tables 2.1 Phenotype summary for RNAi screen of Fat interactors in the eye 2.2 Summary of PCP phenotypes upon RNAi knockdown of mitochondrial genes 2.3 Quantification of PCP defects in Ndufv2 transgene rescue experiments 2.4 Summary of RNAi screen of Ft interactors in the wing 2.5 Summary of PCP phenotypes upon ROS level perturbations vii
8 List of Figures 1.1 ft mutants drastically overgrow and show abnormal PCP 1.2 Schematic of the atypical cadherins Ft and Ds 1.3 The core PCP pathway 1.4 Fj and Ds are expressed in gradients that bias Ft and Ds asymmetry 1.5 The Hippo pathway 1.6 Ft-ICD contains regions of high conservation important for Ft function 1.7 The oxidative phosphorylation system 1.8 Schematic of mitochondrial import via the presequence pathway 2.1 RNAi-mediated knockdown of candidate Ft interactors during Drosophila eye development results in a range of eye defects 2.2 Knockdown of candidate Ft interactors during Drosophila eye development results in a range of ommatidial phenotypes 2.3 Knockdown of Ft interactors results in ft-like PCP defects 2.4 Depletion of Ndufv2 during eye development leads to ablation of dorsal head tissue 2.5 Knockdown of candidate ft interactors induces antennal duplication 2.6 Depletion of Ndufv2 in the wing results in PCP defects 2.7 Knockdown of OXPHOS genes leads to defects in eye development and PCP 2.8 Ndufv2 depletion by multiple independent RNAi lines leads to polarity defects 2.9 Expression of Ndufv2 RNAi in the Drosophila eye gives rise to a highly variable head cuticle dysplasia phenotype 2.10 Quantification of PCP defects upon Ndufv2 depletion 2.11 Specificity of anti-ndufv2 antibody and expression levels of Ndufv2 transgenes 2.12 UAS-Ndufv2 and UAS-Ndufv2 FeS transgene expression during eye development 2.13 Perturbations of Ndufv2 levels in mitotic clones results in autonomous and nonautonomous polarity defects 2.14 Ndufv2 and CG1746 directly interact with highly conserved regions in the intracellular domain of Ft 2.15 Ft heterozygosity mildly enhances head dysplasia phenotype of Ndufv2 depletion in males 2.16 ft heterozygosity increases crossvein distance and vein 3 length in Ndufv2-depleted wings 2.17 Overexpression of Ndufv2 in ft - eye tissue causes severe degeneration of ommatidia viii
9 2.18 ft and Ndufv2 regulate PCP and Hippo pathway targets 2.19 Depletion of Ndufv2 results in increased cell death and increased proliferation 2.20 Ndufv2 does not regulate polarity via core PCP, JAK/STAT, nor AmpK signalling pathways 3.1 ft mutants experience a delay in growth and pupariation 3.2 ft larvae are not developmentally delayed as assessed by mouth hook morphology 3.3 Loss of ft or Ndufv2 results in defective mitochondria 3.4 ft affects mitochondrial morphology in imaginal discs 3.5 Depletion of mitochondrial complex I and III components increases ROS levels 3.6 ft regulates ROS independently of the Hpo pathway 3.7 The JNK pathway is activated in Ndufv2-depleted and ft mutant tissue 3.8 ft mutants have reduced CI activity and upregulated glycolysis resulting in wildtype levels of ATP 3.9 Ndufv2 colocalizes with CVα in Drosophila S2 cells 3.10 Ft is processed in vivo to release an intracellular 68 kda fragment 3.11 Predicted mitochondrial targeting sequences in the cytosolic domain of Ft 3.12 Ft contains multiple mitochondrial targeting sequences that direct mitochondrial import in transfected cell assays 3.13 Membrane association is important for mitochondrial localization of Ft 3.14 Endogenous Ft localizes to mitochondria in D11 cells 3.15 A fragment of Ft is localized to mitochondria in imaginal discs 3.16 ft larvae have altered levels of mitochondrial proteins 3.17 ft fd larvae show defecits in CI and CV assembly and activity 3.18 ft pathway mutants have increased levels of CytoC and is rescued by expressing full-length Ft in ft null larvae 3.19 Ft conserved region 2 is necessary for mitochondrial function 3.20 Model of Ft processing and function at the cell surface and in stabilizing the ETC in mitochondria 3.21 Mitochondrial-binding regions of Ft are highly conserved 3.22 Predicted mitochondrial targeting sequences in the cytosolic domain of mfat4 ix
10 List of Abbreviations AEL after egg lay Atro Atrophin BN-PAGE blue native polyacrylamide gel electrophoresis CI mitochondrial complex I CII mitochondrial complex II CIII mitochondrial complex III CIV mitochondrial complex IV CV mitochondrial complex V cdna copy DNA COS cells CV-1 (simian) in Origin and carrying the SV40 genetic material Crb Crumbs CVD cross vein distance dcia drosophila complex I assembly factor D Dachs DE dorsal enhancer, derived from mirror regulatory regions DHE dihydroethidium Ds Dachsous dsrna double stranded RNA Dsh Dishevelled DV dorsal-ventral EdU 5-Ethynyl-2'-deoxyuridine En Engrailed ETC electron transport chain Ey Eyeless Ex Expanded Fe-S iron-sulfur cluster Fj Four-jointed FLP flippase, yeast site-specific recombinase FRT flippase recombination target site Ft Fat Ft-FL full length Fat Ft-FLΔ2 full length Fat harbouring deletion of conserved region 2 Ft-FLΔ8 full length Fat harbouring deletion of conserved region 8 Ft-ICD intracellular domain of Fat Ft mito 68kDa cytosolic Fat fragment targetted to mitochondria FtΔECD Fat harbouring a deletion of nearly all of its extracellular Cadherin domains Fz Frizzled Gal4 Galactose4 GFP Green fluorescent protein GMR Glass multiple reporter GST glutathione-s-transferase HA Human influenza hemagglutinin Hpo Hippo hs heat-shock x
11 HS Hennekam s syndrome LacZ β-galactosidase MARCM mosaic analysis with a repressible cell marker Mats Mob as tumour suppressor MF morphogenetic furrow MPP mitochondrial processing protease MTS mitochondrial targeting sequence Ndufv2 NADH dehydrogenase ubiquinone flavoprotein 2 NIG National Institute of Genetics Nub Nubbin O2 - superoxide anion OXPHOS oxidative phosphorylation PCP planar cell polarity PR photoreceptor RNAi RNA interference ROS reactive oxygen species Sav Salvadore S2 cells Schneider 2 Drosophila cells UAS Gal4 upstream activation sequence VMS Van Maldergem s syndrome V3 longitudinal wing vein 3 VDRC Vienna Drosophila Resource Center wt wild type Wts Warts Y2H yeast-2-hybrid Yki Yorkie xi
12 Chapter 1 Introduction
13 2 Introduction Proper pattern formation is essential to normal development. Developing tissues require both positional as well as directional information in order to establish and maintain a reproducible pattern; the tissue needs to know its position along the body axis to form at the correct location, and it needs to determine its direction in order to be correctly oriented. Importantly, epithelial tissues must distinguish on which side to form apical versus basal domains, which are essential to their function. Epithelial tissues are also oriented in an axis perpendicular to the apico-basal axis, an axis within the plane of the epithelium. Such organization is known as planar cell polarity (PCP). PCP is readily apparent in Drosophila melanogaster and is manifest in patterned structures; for example, in almost all cuticular structures in adult Drosophila, cells produce a single actin hair that protrudes from the distal (in the wing and leg) or posterior (on the abdomen and notum) edge of the cell, resulting in parallel arrays of hairs and bristles. The Drosophila compound eye is composed of repeating units known as ommatidia. Each ommatidium is found in one of two chiral forms (Fig. 1.1B). The dorsal-ventral (DV) midline of the eye, known as the equator, serves as a boundary across which these two chiral forms are arranged in a mirror-symmetric fashion. The development of this highly organized array of cells across the eye has served as a powerful model for the study of PCP. Ommatidial PCP is dependent upon proper eye axis formation. Particularly, normal DV patterning is required for subsequent alignment of ommatidia with the DV axis. Early in eye development, wingless signaling from the posterior tip of the eye imaginal disc (the larval precursor to the adult eye) induces the expression of dorsal (e.g. mirror) and ventral (e.g. fringe) fate selectors (Cho and Choi, 1998; Heberlein et al., 1998; McNeill et al., 1997). In turn, cells on the border between these two compartments begin to express Notch, thus establishing the equator. Ommatidial chirality is then determined during 3 rd instar larval development (described in Wolff and Ready, 1993). As cells are specified, they are sequentially recruited into clusters that include photoreceptor cells. These photoreceptors are arranged as 3 pairs of outer photoreceptors (designated R2/5, R3/4, and R1/6) surrounding the inner photoreceptors R7 and R8. Initially ommatidial pre-clusters are symmetrical with the R3/4 pair on the anterior side of the cluster.
 Schematic of an adult Drosophila eye illustrates PCP organization.")
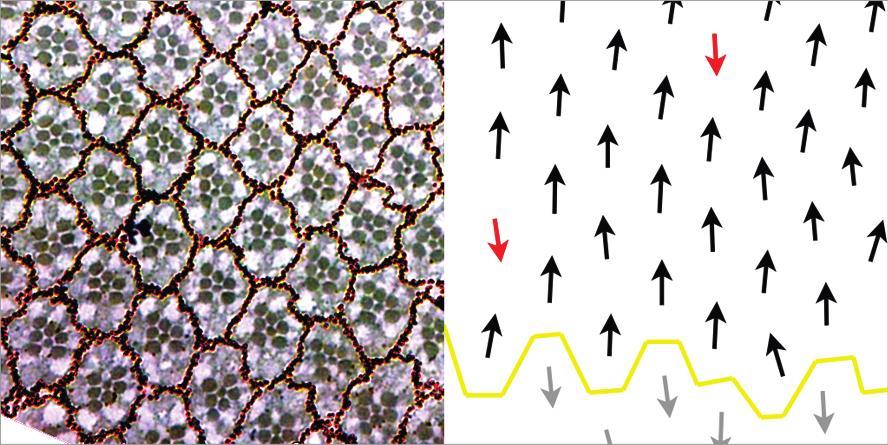
14 Figure 1.1: ft mutants drastically overgrow and show abnormal PCP (A) Wing imaginal discs from 3 rd instar wt and ft fd larvae. ft fd are clearly larger than wt and appear to have convoluted folds within the tissue. (B) Schematic of an adult Drosophila eye illustrates PCP organization. Black circles indicate the rhabdomeres of the ommatidia. Ommatidia are arranged with mirror symmetry across the equator, displaying either dorsal or ventral chirality as illustrated by black and grey arrows, respectively. (C) Micrographs of adult Drosophila eye sections and corresponding polarity schema below reveal normal polarity of ommatidia in controls (black arrows dorsal chirality), while ft mutants display polarity inversions (red arrows). 3
15 4 Then, polarity cues presumably originating from the equator further differentiate the equatorial member of the R3/4 pair into R3, and the polar cell into R4, thus breaking symmetry. Ommatidia subsequently undergo a 90º rotation to reach their final orientation, with R3 and R4 pointing towards the poles. Thus the beautifully patterned structure of the adult eye is formed. Due to these readily perceived PCP phenotypes and the amenability of Drosophila to genetic manipulation, it was in Drosophila that PCP was first studied and genes that affected PCP were isolated (Gubb and Garcia-Bellido, 1982). Another requirement during development is the restriction of growth. As tissues and organs develop, they not only need to form in the right place, but also grow to a specified size. Each organ s proportional size to the body plan, as well as the body s overall size appear to be strictly determined genetically; almost all members of an isogenic group, under similar nutritional and environmental conditions, will develop into a strikingly narrow range of sizes. In addition, there appear to be intrinsic mechanisms that define the ultimate size of organs. Transplant experiments in Drosophila and other organisms have shown that organs cultured in a host develop according to the growth plan of the donor rather than that of the host (reviewed in Bryant and Simpson, 1984). One particularly fascinating gene, fat (ft), appears to regulate both PCP and growth control, making it absolutely indispensible for normal development. In this work, I have investigated how ft might be exerting its control over these aspects of development and have found unexpected links between ft and mitochondria. 1.1 Overview of the ft gene Discovery and phenotypes The Drosophila gene fat (ft) is an essential gene that plays multiple roles during development. It was first isolated as a spontaneous mutation that affects proximal-distal patterning. These mutant flies had shorter and broader bodies, hence the name fat (Mohr, 1923). ft wings also displayed a reduced distance between anterior and posterior crossveins. Other ft alleles were later found and
16 5 were associated with massive overproliferation phenotypes in imaginal discs, the epithelial tissues found in Drosophila larvae that are the precursors to the adult body (Bryant et al., 1988; Mahoney et al., 1991). During a prolonged larval period, these tissues grow to many times larger than their wild type counterparts (Fig. 1.1A, see alsobryant et al., 1988; Mahoney et al., 1991). This overproliferation is hyperplastic, that is, cells retain their epithelial identity and remain in a continuous epithelial sheet as they grow and divide. Consequently, convoluted tissue folds are observed in ft mutant imaginal discs. It was later discovered that these phenotypes are a result of ft s influence on the tumour suppressive Hippo (Hpo) pathway (reviewed in Enderle and McNeill, 2013; see also Chapter 1.3 for detailed description). ft amorphs are late larval or early pupal lethal, presumably due to these overgrowth defects. Though genetic studies have clearly placed ft upstream of the Hpo pathway (Bennett and Harvey, 2006; Cho et al., 2006; Silva et al., 2006; Willecke et al., 2006), how signals are transduced from Ft to the Hpo pathway is uncertain. It was also noted that ft mutants had abnormalities in epidermal morphogenesis. Legs often displayed tissue evaginations/invaginations, as well as disruptions in bristle polarity (Bryant et al., 1988). Wing hairs adopted a distinctive swirling pattern, as opposed to the uniformly distaloriented array in the wild type case. Adult retinas from ft mutants were also examined for such polarity defects and were found to be similarly disorganized; ommatidia, the clusters of cells that make up individual units in the eye, had inverted chirality (Fig. 1.1C). Altogether, these phenotypes pointed towards defects in the establishment of planar cell polarity (PCP). Later studies would identify other components required for ft regulation of PCP, including dachsous (ds) and four-jointed (fj), and together these became known as the ft/ds system (Ma et al., 2003; Rawls et al., 2002; Yang et al., 2002). The ft/ds system is thought to be responsible for setting up long distance directional cues across a tissue. Important evidence that ft and ds are involved in passing of directional information derives from their non-autonomous effect observed when mitotic clones are made. In the eye, ft mutant tissue can cause inversions of neighbouring wild type ommatidia on the polar border of the clone, while ds clones have the opposite phenotype, with inversions on the equatorial border. This suggests that cells are somehow able to sense differences in Ft/Ds levels or activity, and conversely Ft/Ds activity is somehow sending such information. However, despite the significant effort expended to understand it, it remains unclear how ft activity translates into polarity information. The identity of the ft gene product has offered some insight on this front.
17 Ft and Ds are very large cadherins The ft gene product is a very large (560 kda) cell adhesion molecule of the cadherin superfamily. It is a single pass transmembrane protein that contains 34 cadherin repeats, and several EGF-like and laminin A-G motifs in its extracellular domain, while the structure of its cytosolic tail is largely uncharacterized (Fig. 1.2, Mahoney et al., 1991). Its numerous cadherin domains are notable, as this markedly contrasts with classical cadherins which contain 5 repeats. Additionally, Ft differs from classical cadherins in that classical cadherins are largely responsible for linking extracellular contacts with the cytoskeleton through interactions with β-catenin (Armadillo in Drosophila) but Ft does not appear to interact with β-catenin. Though the intracellular domain of Ft contains evolutionary remnants of β-catenin binding motifs (Clark et al., 1995), these motifs are not well-conserved and thus do not bind β-catenin. However, like classical cadherin, Ft does mediate cell-cell contacts through its extracellular cadherin repeats. It does so by interacting with a second large atypical cadherin, Dachsous (Ds). Ds presents a domain structure similar to Ft; it is also a single pass transmembrane protein with a large number of Cadherin repeats (27) and an intracellular domain with no predicted motifs (Fig. 1.2, Clark et al., 1995). Previous studies have shown that Ft on one cell heterophilically binds to Ds in a neighbouring cell in a Ca 2+ -dependent manner, and do so preferentially as this interaction mutually stabilizes both partners at the membrane (Matakatsu and Blair, 2004). Further evidence that Ft and Ds modulate cell adhesivity and cell sorting comes from experiments involving mitotic clones in imaginal discs. Wildtype clones tend to be elongated and rough edged, whereas ft and ds clones are rounder and smoother, suggesting a preference for clonal cells to adhere to one another rather than the surrounding wild type cells. ds mutants display similar phenotypes to ft animals: proximal-distal patterning is perturbed and PCP is aberrant (Adler et al., 1998; Clark et al., 1995). Both ds and ft are expressed in similar developmental times and tissues; of particular interest to this work, they both show high expression in imaginal discs throughout larval and pupal development (Clark et al., 1995; Mahoney et al., 1991). Furthermore, Ft and Ds co-localize at the surface of cells, just apically to adherens junctions (Ma et al., 2003). Through epistasis experiments, ds was found to act
18 Figure 1.2: Schematic of the atypical cadherins Ft and Ds Ft and Ds contain multiple cadherin repeats (green ellipses) in their extracellular domains. Ft also has EGF-like (blue squares) and Laminin A-G domains (orange hexagons). Ft and Ds are single pass transmembrane proteins (transmembrane domain in red). The intracellular fragments of Ft used in our Y2H screen are marked in dotted boxes. These fragments are non-overlapping. 7
19 8 upstream of ft, and also loss of ds results in a slight increase in ft levels (Yang et al., 2002). For these reasons, Ft and Ds are thought to act together as a ligand-receptor pair to mediate downstream polarity and growth signaling. 1.2 ft/ds and PCP regulation The core PCP genes Genetic studies of PCP phenomena in Drosophila have identified two main groups of genes that are required for normal polarity in all tissues tested. The first group is known as the core PCP pathway (reviewed in Carvajal-Gonzalez and Mlodzik, 2014) whose products consist of: Frizzled (Fz), a seven pass transmembrane Wg/Wnt receptor (Gubb and Garcia- Bellido, 1982; Park et al., 1994) Dishevelled (Dsh), a cytoplasmic DIX/DEP/PDZ domain containing protein (Krasnow et al., 1995) Van Gogh (Vang) also called Strabismus (Stbm), a four pass transmembrane protein (Taylor et al., 1998; Wolff and Rubin, 1998) Prickle (Pk), a cytoplasmic LIM domain protein (Gubb et al., 1999) Diego (Dgo), a cytoplasmic ankyrin repeat protein (Feiguin et al., 2001) Flamingo (Fmi) also called Starry Night (Stan), a seven-pass transmembrane atypical cadherin (Chae et al., 1999; Usui et al., 1999) In the pupal wing, these proteins form asymmetrically distributed complexes on opposing sides of the cell with Fz, Dsh and Dgo on the distal side and Vang and Pk on the proximal membrane (Fig. 1.3). The transmembrane components Fz, Vang, and Fmi (found to bridge both complexes intercellularly) stabilize each other across cell membranes while the cytoplasmic components Dsh, Dgo, and Pk set up antagonistic feedback loops within the cell to reinforce an initial asymmetric bias of Fz and Vang complexes. These polarized complexes then signal to downstream cytoskeletal rearrangements that ultimately result in the secretion of an actin hair from the distal side of the cell.
, Diego (Dgo) and Dishevelled (Dsh) form a complex that localizes to the distal membrane of the cell, while Van Gogh (Vang) and Prickled (Pk) localize to the")
20 Figure 1.3: The core PCP pathway Frizzled (Fz), Diego (Dgo) and Dishevelled (Dsh) form a complex that localizes to the distal membrane of the cell, while Van Gogh (Vang) and Prickled (Pk) localize to the proximal membrane. The cytoplasmic components destabilize each other locally, thus reinforcing asymmetric localization of the two complexes. Flamingo (Fmi) forms homodimeric bridges across cell membranes to stabilise both complexes. The distal Fz complex directs an actin hair to form at the distal side of the cell. 9
21 The ft/ds system The second group of PCP genes belong to the ft/ds system, which is of greater significance to this work. The extracellular heterophilic interactions between Ft and Ds are proposed to be crucial to the regulation of PCP. One of the results of this interaction is the transcriptional regulation of four-jointed (fj) a third PCP gene in the ft/ds system. Similarly to ft and ds, fj mutant clones show both autonomous and non-autonomous PCP defects, though wholly fj mutant animals only display mild polarity deficiencies (Zeidler et al., 1999). The relative lack of PCP defects in fj mutants suggests that there is some redundancy in fj s role in polarity establishment. Strikingly, Ds and Fj are expressed in opposing gradients in imaginal discs. In the eye, Ds levels are high near the poles and diminish towards the equator while Fj is high at the equator and low at the poles (Fig. 1.4A). These gradients provide a mechanism for the propagation of directional information across a tissue (Yang et al., 2002). Cells are somehow able to sense or compare their levels Fj and Ds with their neighbours and thus determine in which direction to polarize. When these gradients are manipulated in the eye polarity is reoriented accordingly; for example, ommatidial polarity across the eye can be nearly completely inverted by expressing Ds in a reverse gradient in an otherwise ds and fj null eye (Simon, 2004). Additionally, as the fj phenotype indicated, these two gradients do show redundancy. Each is sufficient to correctly polarize ommatidia even when the graded expression of the other is flattened (Simon, 2004). It has been proposed that Fj and Ds effect a corresponding gradient of Ft activity, and that this differential Ft activity transmits polarity information intracellularly as well as further propagating it at longer range. However, what this activity is has remained elusive. One theory arising from studies in the wing and abdomen proposes that this long-range signaling activity is simply the heterophilic intercellular binding between Ds and Ft propagating from cell to cell in a domino-like manner. Evidence supporting this model is observed at the borders of ft or ds clones. At a ft clone border, Ds in the wild type cell preferentially redistributes away from the clone border in order to form intercellular heterodimers with Ft on the surface of the next wildtype cell away from the clone. This redistribution would then propagate from one cell to the next, thus transmitting directional information (Lawrence et al., 2007). In the wildtype case, the
 Fj kinase activity increases Ft binding affinity to Ds (arrows) and reduces Ds binding affinity to Ft (blunted arrows).")
22 Figure 1.4: Fj and Ds are expressed in gradients that bias Ft and Ds asymmetry (A) Diagram of eye-antennal imaginal discs with Ds (blue) and Fj (yellow) expression patterns. (B) Fj kinase activity increases Ft binding affinity to Ds (arrows) and reduces Ds binding affinity to Ft (blunted arrows). The Fj gradient (represented by size of Fj) results in Ft and Ds with different affinities for each other across the tissue (represented by asterisks). This creates a bias for Ft-Ds interactions (dashed arrows) which leads to asymmetric localization of Ft and Ds (C) 11
23 12 endogenous Ds gradient would bias this successive redistribution along the axis of the gradient. Furthermore, asymmetric subcellular localization of both Ft and Ds has been observed (Brittle et al., 2012). Ft shows a slight bias towards localizing on polar borders of cells, while Ds is found on the equatorial side. A molecular mechanism for this apparent asymmetry stems from the earlier finding that Fj is a golgi-resident kinase that phosphorylates the extracellular cadherin domains of both Ds and Ft (Brittle et al., 2010; Simon et al., 2010). Phosphorylation of Ds decreases its affinity for Ft, while phosphorylation of Ft increases its affinity for Ds. Since fj is expressed in a gradient across the eye, differential Fj activity across a tissue would cause Ft in one cell to preferentially bind to Ds in the neighbouring cell with lower Fj activity. Conversely, Ds would preferentially bind to Ft in a cell with higher Fj activity. As a result, Ft-Ds intercellular complexes orient preferentially with Ft in the cell with higher Fj activity (Fig. 1.4B, C). In the eye, this would translate to Ft localizing to polar edges of cells, which is precisely what was observed experimentally. However, the domino model for ft/ds propagation of PCP information does not fully account for their clonal phenotypes. Our lab has shown that polarity information can still be passed from wildtype tissue into the clone which lacks ft or ds entirely (Sharma and McNeill, 2013), thus rendering redistribution impossible. An alternative model invokes the existence of an as yet undiscovered second signal that would be differentially regulated by the Ft/Ds activity gradient. This signal would be diffusible across short distances and provide an input into downstream polarity sensing systems (Fanto et al., 2003; Zeidler et al., 1999). While this model can explain the non-autonomous phenotype as well as provide a method for propagating information through non-polarized cells, there have been no other indications of this mysterious factor s existence. The question remains, then, of what exactly Ft activity is. We know very little of the downstream events in Ft polarity signalling. Some very intriguing data indicates an important role to be filled by the intracellular component of Ft; it was found that expression of a form of Ft that was missing its extracellular domain and thus unable to bind Ds, was nonetheless sufficient to substantially rescue PCP defects in a ft mutant (Matakatsu and Blair, 2006). This indicates that it is the largely uncharacterized cytosolic component of Ft that mediates PCP signalling.
24 13 To further explore the functions of the intracellular domain of Ft, our lab carried out yeast-2- hybrid (Y2H) screens to uncover physical interactors of this domain. Through these screens, we identified the nuclear receptor and transcriptional co-repressor Atrophin (Atro, also called Grunge), as a Ft interactor and PCP regulator. atro eye clones phenocopy ft clones in terms of non-autonomous PCP defects, and also in the transcriptional regulation of fj (Fanto et al., 2003), therefore atro is considered another component of the ft/ds PCP machinery. However, atro s domain of influence on PCP appears to be spatially restricted, at least in the eye. We have determined that atro is required mainly near the equator, while near the poles atro has little effect on polarity (Sharma and McNeill, 2013). Thus, there are likely other unknown Ft interactors that mediate PCP signaling in other contexts. 1.3 The Hippo pathway The Hippo pathway has emerged as a key player in developmental and cancer biology over the last decade (reviewed in Enderle and McNeill, 2013). It is a signaling pathway that primarily functions to restrict tissue growth and control organ size. The Hpo pathway was first identified in Drosophila in screens for tumour suppressor genes. The core pathway consists of the kinases Hippo (Hpo - Harvey et al., 2003; Jia et al., 2003; Pantalacci et al., 2003; Udan et al., 2003; Wu et al., 2003) and Warts (Wts - Justice et al., 1995; Xu et al., 1995). Also included are their accessory proteins Salvador (Sav - Kango-Singh et al., 2002; Tapon et al., 2002) and MOB as tumor suppressor (Mats - Lai et al., 2005). These four proteins form the central kinase cascade of the Hpo pathway (Fig. 1.5) Upstream signaling induces phosphorylation and activation of Hpo, which then phosphorylates Wts. Wts in turn, phosphorylates the transcriptional co-activator Yorkie (Yki - Huang et al., 2005), thereby creating a binding site and ultimately sequestering Yki in the cytoplasm away from the nucleus. In contrast, when the Hpo pathway is inactive and Yki is allowed to translocate to the nucleus, Yki associates with the TEAD/TEF family transcription factor Scalloped (Sd) to activate growth-promoting and anti-apoptotic genes such as CyclinE (CycE), Drosophila Inhibitor of Apoptosis 1 (diap1), and the microrna bantam (Nolo et al., 2006). Nuclear Yki also upregulates upstream Hpo-activating genes like expanded (ex) and crumbs
 which results in the phosphorylation (P) of Yki, thus sequestering Yki away from the")
25 Figure 1.5: The Hippo pathway A simplified cartoon of the Hippo pathway depicts the core kinase cascade (red dashed box) which results in the phosphorylation (P) of Yki, thus sequestering Yki away from the nucleus. In the absence of Hpo signaling, Yki translocates to the nucleus and interacts with Sd to promote the transcription of pro-growth and anti-apoptotic genes. Upstream signaling interactions are complex. Direct biochemical links are represented by solid arrows. Dashed arrows indicate interactions inferred from genetic evidence. 14
26 15 (crb) in a negative feedback loop. In this way, the Hpo pathway acts as a powerful system to restrict growth. Study of upstream regulation of the Hpo pathway has been particularly prolific in recent years. With the notable inclusion of ft among Hpo regulators, these studies have uncovered a rather intricate web of interactions, leading to the conclusion that the Hpo pathway serves as an integration point for many inputs from both inside and outside the cell. One key point that is becoming clear is that apical recruitment of core Hpo pathway components is an important activating event in Hpo control. Among the myriad factors that have been identified, Expanded (Ex) is of pertinence to this work due to its interaction with Ft. The subapically localized Ex, Merlin, and Kibra proteins form a complex which binds to Hpo, Wts, Sav, and Yki, thus bringing Hpo pathway components to the subapical membrane and in close-proximity with each other (Baumgartner et al., 2010; Cho et al., 2006; Genevet et al., 2010; Hamaratoglu et al., 2006; Yu et al., 2010). Markedly, Ft is also localized to the subapical membrane, and it is here that Ft presumably exerts its effect on the Hpo pathway. Specifically, Ft has been shown to regulate the levels and localization of Ex (Bennett and Harvey, 2006; Cho et al., 2006; Silva et al., 2006; Willecke et al., 2006), although it is not certain if this is relevant to downstream Hpo activity, as genetic evidence also indicates that Ft and Ex can act in separate parallel pathways in some contexts (Feng and Irvine, 2007). Nonetheless, Ft does affect Hpo signaling: ft overgrowth phenotypes are similar to Hpo pathway mutants, and ft regulates Yki targets. Additionally, Wts protein levels are altered in ft mutants. Genetic epistasis experiments also place ft upstream or parallel to Hpo, as overexpression of core Hpo components can rescue growth defects induced by loss of ft (Bennett and Harvey, 2006; Cho et al., 2006; Silva et al., 2006; Willecke et al., 2006). With the interactions between ft and ex as opaque as they are, other intermediaries between Ft and Hpo have been sought after. The unconventional myosin Dachs (D) was identified as such a mediator between Ft and the Hpo pathway. Loss of d appears to suppress the decrease in Wts levels observed in ft mutants (Cho et al., 2006; Mao et al., 2006). D itself is a negative regulator of the Hpo pathway and physically interacts with Wts. Together, these data outline a pathway where Ft inhibits D activity which in turn inhibits Hpo signaling at the level of Wts (Fig. 1.5). However, the biochemical link between Ft and D is still lacking. Further work will be required to delineate how Ft acts through D, and to determine if D is the sole mediator of Ft signaling to the Hpo pathway.
27 Vertebrate homologs of ft and human disease Fat cadherins have been found in a wide range of organisms from basal metazoans, like Trichoplax adhaerens, to higher vertebrates, including mice and humans (Hulpiau and van Roy, 2011). This suggests that Fat cadherins serve evolutionarily important functions in vertebrates as well as in Drosophila. Vertebrates have 4 Ft orthologs, FAT1-4. Phylogenetic analysis indicates that FAT4 is the homolog of Drosophila Ft based on similarity of their intracellular domains. Sequence alignment between Fat cadherins revealed 8 regions of high conservation in their cytosolic portions (Fig. 1.6 and Bossuyt et al., 2014; Matakatsu and Blair, 2012; Pan et al., 2013; Zhao et al., 2013). While some of these conserved regions are important for Hpo and PCP signaling in Drosophila, it is not known if they are functionally relevant in vertebrates. One particular region identified as being required for Hpo signaling in flies diverges in mouse and human (Bossuyt et al., 2014), leading investigators to conclude that Ft regulation of the Hpo pathway through this region evolved specifically in the arthropod lineage. The role that vertebrate FATs play in Hpo signaling remains controversial (Bossuyt et al., 2014; Das et al., 2013). It is known, though, that Ft regulation of PCP is conserved through mammals. Our lab has shown that Fat4 is necessary for the establishment of PCP in mice (Saburi et al., 2008). Briefly, Fat4 mutant mice exhibit defects in cochlear length and kinocilium orientation in the inner ear; they show significantly wider neural tubes suggesting defects in convergent extension cell movements. Fat4 mutant mice also display cystic kidneys as a consequence of disrupted oriented cell division. All of these phenotypes are characteristic of vertebrate PCP mutants. Our lab also uncovered genetic interactions between Fat4 and Fjx, Atro1, and Vangl2, all homologs of PCP genes in Drosophila (Saburi et al., 2012). This is consistent with Fat4 playing a role in PCP signaling in vertebrates. In humans, Fat4 mutations are associated with two genetic disorders, Van Maldergem s syndrome (VMS - Cappello et al., 2013) and the related Hennekam s syndrome (HS - Alders et al., 2014). In common to both syndromes are abnormalities in facial structure, microencephaly, reduced stature, digit anomalies, and varying levels of cognitive impairment. HS is further characterized by lymphatic edema, while individuals with VMS present with periventricular
 Matakatsu and Blair, 2012; (II) Zhao et al., 2013; (III) Bossuyt et al., 2013 ; (IV) Pan et al.")
28 Figure 1.6: Ft-ICD contains regions of high conservation important for Ft function The intracellular domain (ICD) of Ft contains 8 highly conserved regions (red boxes). Recent structure-function studies : (I) Matakatsu and Blair, 2012; (II) Zhao et al., 2013; (III) Bossuyt et al., 2013 ; (IV) Pan et al., 2013; have identified domains that are important for PCP regulation (blue lines) or Hpo pathway control (red lines) or both (PH, purple). Domains are labeled with nomenclature used in each study. Dashed lines indicate weak requirements for Ft. 17
29 18 heterotopia. Due to the high level of overlap in phenotype and association with Fat4 mutations, Alders et al. propose that HS is allelic to VMS at the Fat4 locus. However, none of these phenotypes have been observed in Fat4 mice, raising the possibility that strong Fat4 null phenotypes are masking these other defects. Indeed, our Fat4 null mice die perinatally. Alternatively, other genetic differences may be sensitizing these individuals with VMS and HS to loss of Fat4. Many of the Fat4 mutations identified in VMS occurred in individuals who were heterozygous at the Fat4 locus, yet our Fat4 heterozygous mice do not present any of the VMS or HS phenotypes either. Thus our Fat4 mouse model may not be accurately recapitulating the genetic conditions of VMS or HS. It is unclear how Fat4 may be contributing to these syndromes; a recent study removing Fat4 from facial brachiomotor neurons has suggested that migration of these neurons is a PCP process (Zakaria et al., 2014), providing a possible mechanism for the periventricular heterotopia observed in VMS. There are still numerous aspects of VMS and HS that remain unexplained. If Fat4 does prove to be causative of VMS and HS, it would indicate that Fat cadherins are crucial for more developmental processes than we have thus far exposed. Another field where Fat cadherins are emerging as significant players is cancer biology. Considered a tumour suppressor itself in Drosophila, and a regulator of the tumour suppressive Hpo pathway, ft has long been suspected to play a role in cancer progression. With the advent of genome sequencing and other genomic analyses, Fat mutations have been reported in a wide range of different cancers, from melanomas to pancreatic cancer (reviewed in Sadeqzadeh et al., 2014). One particular study on breast carcinomas revealed a loss of FAT4 expression due to promoter hypermethylation, and further testing in murine mammary cell lines implicated that this loss was causal in increased tumorigenicity (Qi et al., 2009). Moreover, reporters of Hpo pathway activity indicated disruption of Hpo signaling in these cell lines. It is tempting to postulate that loss of Fat4 leads to dysregulation of Hpo signaling and consequent overproliferation, much as in Drosophila, yet there are also conflicting data that Fat4 is upregulated in some breast cancers (Lehmann et al., 2011), or that Fat4 does not regulate Hpo signaling in other contexts (Bossuyt et al., 2014). It must be recognized that there is still much to be learned in terms of Fat cadherins and their role in cancer biology.
30 Mitochondria The mitochondrion is vital to the eukaryotic cell. It has long been studied as a bioenergetic hub responsible for managing metabolites, producing ATP, synthesizing fatty acids, nucleotides and other molecules essential to life. All of these processes occur in a neatly compartmentalized organelle, the development of which undoubtedly contributed to the evolution and success of eukaryotic organisms. In recent years, there has been a resurgence in mitochondrial research that has uncovered multiple novel roles for mitochondria in processes as diverse as apoptosis, stress signaling, cell fate determination, and cell cycle control. In this work, I describe a link between ft and mitochondria, whereby Ft impacts upon the metabolic functions of mitochondria. Following is a brief review of these mitochondrial functions and properties Oxidative phosphorylation Oxidative phosphorylation (OXPHOS) is the respiratory process carried out within mitochondria which uses a series of redox reactions to generate the energy required to ultimately phosphorylate ADP to ATP, thus replenishing the energy pool of the cell. OXPHOS is employed by aerobic organisms as one of two routes to extract the chemical potential energy from glucose; the other, glycolysis, is shared with anaerobes. As OXPHOS offers much higher efficiency OXPHOS nets 34 ATP per molecule of glucose more than glycolysis alone it is the main source of ATP in aerobic organisms. In most eukaryotic cells, OXPHOS is catalyzed by five mitochondrial complexes, termed complexes I-V (CI-CV), which are found on the inner mitochondrial membrane (Fig. 1.7 and overview in Papa et al., 2012). Electrons enter the series of complexes at CI (also called NADH dehydrogenase) which oxidizes NADH to NAD+. Electrons then pass through a series of Iron- Sulfur (Fe-S) clusters embedded in CI, ending with the reduction of ubiquinone. The energy derived from CI redox activity is coupled to the pumping of protons from the matrix into the intermembrane space, thereby generating a proton gradient across the inner mitochondrial membrane. CII (also called succinate dehydrogenase) is a second entry point, receiving electrons from succinate and transporting them to ubiquinone as well. CII does not have proton pumping activity. CIII and CIV (cytochrome C reductase and cytochrome C oxidase, respectively) continue the electron transport from ubiquinone, through CIII, to cytochrome C, then through CIV, finally resulting in the reduction of O2 to H2O. Both CIII and CIV also pump protons into
 and cytochrome C (CytC) are shown in yellow.")
31 Figure 1.7: The oxidative phosphorylation system A schematic of the five complexes of the OXPHOS system (CI-V) depicts the path of electrons (dashed arrow) from NADH or succinate to O2. Three of the complexes, CI, CIII, and CIV have proton pumping activity coupled to transfer of electrons. Intermediary electron carriers ubiquinone (Q) and cytochrome C (CytC) are shown in yellow. The proton gradient achieved this way is used by CV to phosphorylate ATP. 20
32 21 the intermembrane space, thus contributing to the proton gradient. The proton motive force generated in this way is exploited by CV (ATP synthase), whose structure resembles a molecular rotary generator; the energy of protons returning to the matrix through CV is captured to phosphorylate ADP to ATP. As elegant a system as OXPHOS is, there are imperfections and drawbacks to its operation in the cell. Electrons may leak from the transport chain, consequently, instead of reducing the intended electron carriers (ubiquinone and cytochrome C), they react with surrounding molecules to form reactive oxygen species (ROS). ROS, given high enough concentrations, can in turn react with and damage other vital components of the cell, such as DNA. Another puzzling fact is that despite its significance to energy metabolism, OXPHOS is not strictly required for cell viability (though, not surprisingly, many genetic metabolic diseases do trace back to deficiencies in OXPHOS components; reviewed in Torraco et al., 2015). In fact, in some contexts the cell will shift from OXPHOS and rely heavily on glycolysis for ATP production, despite decreased efficiency of glucose use. There is some evidence that most actively proliferating cells will undergo this switch to glycolysis (Brand and Hermfisse, 1997). These investigators speculate that if glucose supply is not limiting, then there may be advantages to this. That is, DNA undergoing replication in dividing cells are less likely to be damaged as glycolysis does not produce ROS. Glycolysis also generates pyruvate, which feeds into the citric acid cycle, thus providing a source for many of the biosynthetic intermediates necessary for anabolic pathways. Hence, the molecular building blocks necessary for proliferation and growth are supplied by increased glycolytic rates. Intriguingly, cancer cells have long been known to undergo a similar metabolic shift known as the Warburg effect (Warburg, 1956), although ROS levels are generally high in tumours, thus negating that particular benefit of reliance on glycolysis. Clearly, much remains to be learned about the subtleties of these important metabolic pathways Mitochondrial import Mitochondria are surrounded by a double membrane. As we have seen above, this compartmentalization is absolutely critical for providing an environment amenable to OXPHOS. However, this does present a challenge for the synthesis and maintenance of the mitochondrion. While mitochondria have their own genome and transcriptional/translational machinery, the
33 22 majority of proteins localized to mitochondria are encoded by the nuclear genome. Thus, most mitochondrial proteins must be targeted to and translocated across at least the outer and possibly also the inner mitochondrial membrane. Mitochondrial proteins must also be sorted according to their destinations: matrix, inner membrane, intermembrane space or outer membrane. This sorting is achieved by a complex system of signaling sequences on mitochondrial proteins in conjunction with the mitochondrial import complexes (Fig. 1.8). The key import complex that appears to be necessary for nearly all recognition and import of mitochondrial proteins is the translocase of the outer membrane (TOM) complex. At the heart of the TOM complex is TOM40, which forms β-barrel channels through the outer mitochondrial membrane, thus allowing newly synthesized peptides destined for the mitochondria to pass into the intermembrane space. TOM40 oligomerizes to form 1-3 channel groups (Model et al., 2008). In addition to TOM40, several other accessory proteins assist in recognizing and binding mitochondrial targeting presequences (MTS). The first discovered and best-described type of MTS consists of an N-terminal amphipathic α-helix of amino acids in length and is positively charged on one face (Moberg et al., 2004). This type of MTS is normally utilized by matrix localized proteins. The α-helix in the MTS is sequentially bound by TOM20, TOM22, TOM40, then the intermembrane face of TOM22 in a so-called binding chain as the unfolded peptide passes through the TOM40 channel (Komiya et al., 1998). The protein then traverses the inner mitochondrial membrane through the translocase of the inner membrane (TIM) complex which forms a channel to the matrix. The α-helix of the MTS then unravels and is recognized by mitochondrial processing protease (MPP) which cleaves off the MTS at a specific consensus cleavage site, thus releasing the mature protein into the matrix (Taylor et al., 2001). In recent years, other pathways for mitochondrial import have been discovered which utilize MTS of different forms (reviewed in Chacinska et al., 2009) and lead to protein localization at other mitochondrial compartments. There are also reports of cryptic and hybrid signal sequences that can direct proteins to mitochondria, the ER, or the nucleus depending on cellular context and physiological condition (Boopathi et al., 2008; Nargund et al., 2012). These novel mitochondrially localized proteins serve as a reminder that more studies will be necessary to decipher fully the process of mitochondrial targetting and import.
 is an amphipathic α-helix structure with positive charge on one face")
34 Figure 1.8: Schematic of mitochondrial import via the presequence pathway The presequence or mitochondrial targeting sequence (MTS) is an amphipathic α-helix structure with positive charge on one face (denoted with ++++). The MTS binds to TOM complex components in succession (arrows) to translocate across the outer mitochondrial membrane. The TIM complex then allows the protein to cross the inner mitochondrial membrane. Mitochondrial processing protease (MPP) cleaves off the MTS, releasing the mature protein into the matrix. 23
35 Rationale and aims of thesis ft is an essential gene, regulating the organization of developing tissues with respect to PCP as well as restriction of organ size. While it is known that ft and its binding partner ds are genetically upstream of the Hpo pathway and act either upstream of or in parallel to the fz PCP cassette, how ft affects these targets is not clear. The membrane tethered intracellular portion of Ft is sufficient to rescue to a significant extent the growth and PCP defects in ft animals (Bossuyt et al., 2014; Matakatsu and Blair, 2006, 2012; Pan et al., 2013; Zhao et al., 2013), suggesting the intracellular domain mediates most of Ft s signaling function. With this reasoning, interactors of the intracellular domain would serve as valuable candidates for studying how ft signals are transduced. The main aims of this thesis are to characterize Ft interactors to determine how they may be playing roles in ft signaling, ultimately to better understand how this essential gene functions during development. In Chapter 2 I outline my in vivo analysis of a previously generated list of Ft interactors obtained from Y2H screens. I survey PCP and growth phenotypes of these genes in the Drosophila eye using an RNAi approach. I then describe the physical and genetic interactions between ft and one of these interactors, Ndufv2, a component of mitochondrial Complex I. Knockdown of Ndufv2 shows remarkably similar phenotypes to ft in PCP and Hpo pathway control. These data provide evidence of a role for mitochondria in ft signaling. In Chapter 3 I describe our investigations of mitochondrial phenotypes in ft mutants. We find defects in cristae structure and a shift in metabolic pathways. We demonstrate that Ft is processed and imported into mitochondria where it promotes the assembly/stability and activity of OXPHOS complexes. Taken together, these results delineate a novel function for ft in mitochondria.
36 25 Chapter 2 Identification and analysis of novel Ft interactors Data attribution: GST pulldown experiments were carried out by Ian Hester. Hpo target analysis was completed in collaboration with Yonit Tsatskis and Robyn Rosenfeld.
37 26 Identification and analysis of novel Ft interactors 2.1 Introduction ft is essential to many processes during development; organ growth restriction, planar polarity determination and tissue patterning are all abnormal in ft mutants. It has also been implicated in a number of vertebrate developmental diseases, such as polycystic kidney disease (PKD), as well as some forms of cancer. While genetic studies have shown ft to be an upstream regulator of several pathways, most notably the Hpo tumour suppressive pathway, the direct links between ft and its downstream effects are unknown. In this chapter, I aim to identify direct interactors of Ft and to determine how they might function in ft signaling. With formerly identified candidate binding partners from a yeast-2-hybrid screen carried out in our lab, I conduct an in vivo reverse genetic RNAi candidate screen in the adult Drosophila eye. I make the surprising discovery that several mitochondrial genes participate with ft to regulate PCP and the Hpo pathway. I uncover a genetic interaction between ft and a component of mitochondrial Complex I (CI), NADH Dehydrogenase (Ubiquinone) Flavoprotein 2 (Ndufv2), in the context of proximal-distal patterning in the wing as well as in cell viability. Most remarkably, I find Ndufv2 physically binds highly conserved domains of Ft, and regulates both PCP and Hpo target genes. Altogether, these data suggest a previously unsuspected role for mitochondria in tissue patterning and development. 2.2 Materials and methods Fly stocks and genetics All wt flies are yw unless otherwise noted. Mutant alleles of ft are described in FlyBase ( briefly, ft fd and ft x13 are null alleles; the molecular nature of the hypomorphic allele ft alb is unknown; ft G-rv contains a stop codon in its 27 th Cadherin repeat, and additional rearrangements in its first intron. RNAi stocks for expression of dsrna under control of GAL4/UAS were obtained from VDRC (Dietzl et al., 2007) and the NIG-fly stock center (see Table 2.1 for a full list of RNAi lines). CG7719 served as the control RNAi. RNAi lines were driven using eygal4, GMR-Gal4, engal4, nubgal4, or yw, UAS-dicer2/Y hs-hid; eygal4, GMR-
38 27 Gal4/CyO (generously provided by Claude Desplan). For Ndufv2 knockdown, VDRC line #22194 was used unless otherwise specified. UAS-Ndufv2 transgenic flies were generated by site-specific ( C31-mediated) P-element transformation using an Ndufv2 cassette amplified from cdna. UAS-Ndufv2ΔFeS transgenic flies were generated by site-directed mutagenesis of C169S and C173S in the above Ndufv2 cassette, and was targeted to the same attp landing site used for the UAS-Ndufv2 transgene (injections done by BestGene Inc. attp site on ch. 3 at 86Fb used, strain #24749). Ndufv2 RNAi clones were generated by heatshock at 37ºC for 1 hour at ~72 hours AEL using the following stock w; act>y+>gal4, UAS-GFP/UAS-Ndufv2 RNAi ; MKRS, hsflp/+. FLP/FRT and MARCM clones for ft and Ndufv2 RNAi were generated under a similar heatshocking schedule. The fj-lacz reporter was previously described (Sopko et al., 2009). BantamGFP sensor line was obtained from Georg Halder. All other fly stocks were obtained from the Bloomington Drosophila Stock Center Tangential eye sections Adult heads were fixed with 0.25% glutaraldehyde, stained in OsO4 and embedded in Durcupan resin. Tangential eye sections were prepared as previously described (Tomlinson and Ready, 1987) and imaged on a Nikon Eclipse 80i upright microscope Wing hair analysis Adult Drosophila wings were mounted in DPX mounting media with dorsal surface upwards and imaged on a Nikon Eclipse 80i upright microscope Antibodies The following antibodies were used: mouse anti-cvα (MitoSciences), mouse anti- -Gal (Promega), mouse anti-engrailed (4G9, DHSB), rat anti-crumbs (gift from Ulrich Tepass), guinea pig anti-expanded (gift from Rick Fehon), anti-activated Caspase3 (Cell Signalling #9661S), anti-his (Sigma SAB ) anti-ndufv2 (raised in rat against full-length GSTtagged Ndufv2), and Alexa Fluor or Cy3-conjugated anti-rat and mouse IgG secondary antibodies (Invitrogen or Jackson Labs).
39 Protein purification and GST pulldown Protein purification and GST pulldowns were performed as previously described (Sambrook and Russell, 2006). Briefly, bacterial cells were transformed and induced with IPTG to allow expression of recombinant proteins. Cells were sonicated and centrifuged, and supernatant was incubated with Nickel or Glutathione beads at 4 C. Supernatant was removed after spin and protein was eluted off Nickel beads with 250mM Imidazole. Eluted protein was incubated with Glutathione-conjugated protein, washed and beads boiled before loading on SDS-PAGE for visualization with anti-his antibody Immunofluorescence To prepare samples for immunofluorescence, late 3rd instar larval eye or wing discs were washed with PBS, fixed with 4% PFA, washed again with PBS, permeabilized in PBST (PBS + 0.1% Triton X-100) and then blocked one hour in PBST + 5% goat serum before incubation with primary and secondary antibodies in PBST + 5% goat serum. Images were captured using a Nikon D-Eclipse C1 confocal microscope EdU incorporation EdU incorporation assay was performed using the Click-iT EdU AlexaFluor594 Imaging kit (Invitrogen Cat#10084) as per the manufacturer s protocol. 2.3 Results Yeast-2-Hybrid and RNAi screens mitochondrial targets identified As the mechanism(s) by which Fat signals are transduced were yet to be illuminated, our lab endeavoured to find physical interactors of Fat using a Yeast-2-Hybrid (Y2H) screen (Fanto et al., 2003). This work was completed before I began my project by Jamie Meredith, a former technician in the McNeill lab. He used two non-overlapping fragments covering the majority of the intracellular domain of Fat (Fig. 1.2) as bait and screened against a Drosophila embryonic cdna library. He identified 53 candidate interactors. I and another student in the lab, Robyn Rosenfeld, then sought to determine if any of these hits were indeed ft related; we undertook an
40 29 RNAi-based in vivo approach. We obtained RNAi lines targeted against our candidate genes from available stock collections (VDRC and NIG). These flies harbour transgenes that express under the control of the Gal4/UAS system (Brand and Perrimon, 1994) inverted repeats targetting the transcripts of specific genes for degradation by endogenous RNAi machinery. We expressed these transgenes with eye specific promoters either using eygal4, which expresses in the larval eye imaginal disc anterior to the morphogenetic furrow (MF), or using GMR-Gal4, which expresses posterior to the MF, or both Gal4 drivers in combination to express throughout larval eye development. In addition we made use of a UAS-dicer2 transgene to increase the efficiency of RNAi-mediated knockdown. We also modulated RNAi expression using temperature, as the Gal4/UAS system is maximally active at 30ºC with lesser efficiency at lower temperatures. We established conditions where these drivers would produce highest knockdown efficiency while still maintaining normal eye development in the driver-alone control (i.e. intact ommatidia and photoreceptors); conditions used in each cross, as well as a summary of the results described below are included in Table 2.1. Upon obtaining progeny from these crosses, we first examined the surface structures of the eye and head, looking for any ft-like phenotypes: PCP mutants often have mildly rough eyes, suggesting an underlying disorganization of the ommatidia, while stronger mutations in the Hippo pathway (e.g. wts) can result in overgrowth of the eye and other cuticular structures of the head. We did indeed observe several lines that resulted in rough eyes, with varying degrees of roughness. While we did not observe overgrowth of tissues that resembled Hippo pathway mutant phenotypes, we did see a range of tissue development defects, from ectopic outgrowths of cuticular tissues, to smaller eyes, to degeneration of eye tissues, to outright ablation of head structures in extreme cases (see Fig. 2.1 for examples). Occasionally we did not observe any progeny, indicating that knockdown of those particular genes induced lethality. From the crosses that did produce progeny, we took tangential sections of at least 2 eyes from different animals in order to examine more thoroughly if knockdown of these candidates produced ft-like phenotypes. Many RNAi lines produced normal, wild type eyes with properly polarized ommatidia. Of those that did show some phenotype, the most predominant phenotype we saw was abnormal ommatidial development resulting in extra, or missing, or misshapen photoreceptors (Fig. 2.2). As this confounds the interpretation of polarity organization, we did not score these eyes for PCP. Examination of the remaining lines led us to a
41 30 Table 2.1: Phenotype summary for RNAi screen of Fat interactors in the eye CG Number RNAi Line Number* Gal-4 Driver Temp (ºC) PCP Phenotype Comments ey 29 None ey 29 Weak very rare symmetrical ommatidia ey 29 Weak very rare misrotations ey 29 Weak very rare symmetrical ommatidia ey 29 None 1746** ey 29 Strong occasional dorsal-ventral inversions, misrotations and symmetrical ommatidia R-3 ey 25 Uninterpretable lethal GMR 25 Uninterpretable severely disrupted eyes R-1 GMR 25 Weak very rare misrotations ey 29 Uninterpretable lethal ey 29 Uninterpretable lethal R-3 ey 25 Weak very rare symmetrical ommatidia ey 29 None ey 29 Weak very rare misrotations and symmetrical ommatidia 3731** 3731R-1 ey 29 None (Mpp) ey 29 Strong occasional dorsal-ventral inversions ey 29 None ey 29 Uninterpretable lethal ey 29 None ey 29 None ey 29 None 5468** 5468R-2 ey 29 Strong occasional dorsal-ventral inversions (TwdlM) 5468R-3 ey 29 Strong occasional dorsal-ventral inversions GED RT Uninterpretable disrupted eyes ey 29 Weak very rare symmetrical ommatidia ey 29 None GED RT None 5703** ey 29 Strong clean dorsal-ventral inversions (Ndufv2) GED RT Uninterpretable severely disrupted eyes ey 29 Weak symmetrical ommatidia and misrotations GED RT Uninterpretable disrupted eyes
42 31 CG Number RNAi Line Number* Gal-4 Driver Temp (ºC) PCP Phenotype Comments GED RT Weak very rare misrotations GED RT Weak very rare misrotations and symmetrical ommatidia GED RT None GED RT None GED RT None GED RT Uninterpretable severely disrupted eyes GED RT Uninterpretable lethal R-3 ey 29 None 2746 GED RT Weak very rare misrotations R-1 ey 29 None GED RT None GED RT Uninterpretable severely disrupted eyes GED RT None ey 29 Weak very rare symmetrical ommatidia GED RT Weak very rare misrotations and symmetrical ommatidia R-1 ey 29 None 7719R-2 ey 25 None ey 29 None ey 29 None R-1 ey 29 Uninterpretable lethal ey 29 Weak very rare symmetrical ommatidia GED RT Uninterpretable lethal R-2 ey 25 Weak very rare symmetrical ommatidia 8332R-3 ey 29 Uninterpretable lethal GED RT Uninterpretable lethal GED RT Uninterpretable lethal GED RT Uninterpretable lethal GED RT Uninterpretable lethal ey 29 None ey 29 None GED RT Uninterpretable lethal ey 29 None ey 29 None R-1 ey 25 Weak very rare misrotations 9170R-2 ey 25 None GED RT None GED RT Weak very rare misrotations CG RNAi Gal-4 Temp PCP Phenotype Comments
43 32 Number Line Driver (ºC) Number* ey 29 None GED RT Uninterpretable severely disrupted eyes GED RT Weak very rare misrotations GED RT Weak very rare symmetrical ommatidia GED RT Weak very rare symmetrical ommatidia GED RT Weak very rare misrotations GED RT Weak very rare symmetrical ommatidia ey 29 None GED RT Uninterpretable severely disrupted eyes GED RT None GED RT Uninterpretable severely disrupted eyes GED RT Uninterpretable severely disrupted eyes GED RT Uninterpretable severely disrupted eyes GED RT Uninterpretable lethal GED RT Weak very rare symmetrical ommatidia GED RT None GED RT None GED RT Weak very rare symmetrical ommatidia ey 25 None GED RT Uninterpretable severely disrupted eyes GED RT Uninterpretable severely disrupted eyes ey 29 None GED RT Uninterpretable severely disrupted eyes GED RT Weak very rare symmetrical ommatidia GED RT None GED RT None GED RT Uninterpretable lethal GED RT Uninterpretable lethal GED RT Weak very rare misrotations GED RT None GED RT None *RNAi lines denoted with an R (e.g. 2238R-3) were obtained from the NIG-fly stock center. All other lines were obtained from the VDRC GED = UAS-dicer2/Y hshid; ey-gal4, GMR-Gal4/CyO RT = ambient temperature of ~21ºC **PCP hits: CG1746, CG3731 = Mpp, CG5468 = TwdlM, CG5703 = Ndufv2

44 Figure 2.1: RNAi-mediated knockdown of candidate Ft interactors during Drosophila eye development results in a range of eye defects. Fly heads displaying rough eyes, cuticular outgrowths, small eyes or total ablation of eye structures. These are examples of phenotypes observed when candidate Ftinteracting genes (in brackets) are depleted by driving dsrna against them using an eye specific promoter (eygal4 or GMR-Gal4). Gal4 driver alone serves as a control (left). 33
 reveal diverse ommatidial phenotypes.")
45 Figure 2.2: Knockdown of candidate Ft interactors during Drosophila eye development results in a range of ommatidial phenotypes. Tangential sections through adult Drosophila eyes in which dsrna against various Ft interactors has been driven with eye specific drivers (eygal4 and/or GMR-Gal4) reveal diverse ommatidial phenotypes. Wildtype ommatidia contain a full complement of seven visible photoreceptors arranged in a trapezoidal pattern and oriented towards the pole (see Fig. 1.1B). Ommatidial defects observed included: misrotations; symmetric ommatidia where photoreceptors 3 and 4 are not properly differentiated; extra or missing photoreceptors; and misshappen or degenerated photoreceptors. 34
46 35 few interesting hits. Knockdown of CG9723 resulted in eyes with extra spacing between ommatidial units. This is a phenotype shared with some Hippo pathway mutants. Robyn has further pursued research on this gene, and her in-depth study of CG9723 makes up the bulk of her Ph.D. thesis work (Rosenfeld, 2014). Several interesting candidates I discovered include: CG5703 NADH dehydrogenase ubiquinone flavoprotein 2 (Ndufv2): a component of mitochondrial complex I CG3731 MPP: mitochondrial processing protease CG1746 a component of mitochondrial complex V (ATP synthase) CG5468 twdlm: a cuticle protein Knockdown of these genes resulted in dorsal-ventral inversions of ommatidia, an eye PCP phenotype specific to ft pathway mutants (ft, ds, fj, and atro mutations all cause this phenotype, see Fig 1.1 and Fig 2.3). This is in contrast to mutations in other PCP genes, such as fz and dsh, which lead to misrotations or achiral ommatidia. This strongly suggests that these four genes are related to the ft PCP pathway. Surprisingly, three of these four are mitochondrial proteins, to which neither ft nor PCP had been previously linked. Knockdown of these candidates also caused other defects. All four showed rough eye surface phenotypes; all had abnormalities in overall eye and head structures, including outgrowths of cuticular tissue from within and around the eye. Strong knockdown of Ndufv2 in particular resulted in partial to full ablation of the dorsal region of the head where ocelli normally form (Fig. 2.4). Depletion of Ndufv2, MPP, and twdlm occasionally produced antennal duplication, observed both in adult and larval stages (Fig. 2.5). This was reminiscent of perturbations in EGFR or Notch signaling that result in transdifferentiation of the eye (Kumar and Moses, 2001). Photoreceptor and ommatidial development were also variably impacted by knockdown of these four genes; markedly, depletion of Ndufv2 most reproducibly resulted in eyes (or regions of eyes) with no defects in ommatidial development, while still showing dorsal-ventral inversions. Therefore, I chose to pursue further studies of Ndufv2 and its potential interactions with ft. PCP is also evident in the unidirectional distal orientation of hairs on the Drosophila wing. I examined wings from flies in which I knocked down Ndufv2 using engal4, which drives expression in posterior compartments of the wing. I found that these animals had disruptions in normal hair polarity (Fig. 2.6); in the region distal to the posterior cross vein, hairs were misoriented, pointing in random directions in a swirling pattern. This phenotype is reminiscent of

47 Figure 2.3: Knockdown of Ft interactors results in ft-like PCP defects. Tangential sections of adult Drosophila eyes reveals requirement for Ft interacting genes in PCP regulation. The four genes shown here were the top PCP hits from the in vivo RNAi screen. Polarity is diagrammed below each micrograph: black arrows represent normal chirality, red arrows represent polarity defects, open circles represent abnormally formed ommatidia and were not scored for polarity. dsrna against each target gene was driven with eye specific promoters (eygal4/gmr-gal4) throughout eye development. 36

48 Figure 2.4: Depletion of Ndufv2 during eye development leads to ablation of dorsal head tissue. Dorsal cuticular structures and ocelli regions of the adult head were ablated when dsrna against Ndufv2 was driven using eygal4. Compared to wildtype controls, the knockdown heads are reduced in size in both dorsal and anterior dimensions. 37
, but either fail to form (arrowhead in B) or duplicate partially (arrows in C) or completely (arrows in D) when mpp is depleted.")
49 Figure 2.5: Knockdown of candidate ft interactors induces antennal duplication. A-D: Anterior views of adult Drosophila heads. Antennae develop normally in control cases (A), but either fail to form (arrowhead in B) or duplicate partially (arrows in C) or completely (arrows in D) when mpp is depleted. E-G: Fluorescence micrographs of 3 rd instar eye-antennal discs stained with phalloidin (red) and antibodies against Bar (green) and Elav (blue). The control disc displays a single antenna (E), while an mpp-depleted antennal disc is duplicated (arrows in F). Knockdown of TwdlB results an additional ectopic antenna developing in the eye disc (arrow in G). 38

50 Figure 2.6: Depletion of Ndufv2 in the wing results in PCP defects. Micrographs of adult Drosophila wings with corresponding high magnification images below (magnified area boxed). Normal hair orientation is proximal to distal as seen in the control wing. In the Ndufv2 depleted wing (dsrna against Ndufv2 driven by engal4 which expresses in the posterior compartment of the wing), hairs are oriented randomly, resulting in a swirling pattern. These swirls were observed mainly in the region of the wing distal to the posterior cross vein. 39
51 40 hair polarity phenotypes in ft pathway mutants. This suggests that Ndufv2 functions to regulate PCP not only in the eye, but also in the wing. In addition, I knocked down MPP and twdlm in the wing, but did not observe any PCP defects. As Ndufv2 displayed more robust PCP phenotypes than MPP or twdlm in both the eye and the wing, I decided to further characterize Ndufv Knockdown of several components of the mitochondrial OXPHOS system result in PCP defects, as well as broad deficits in normal tissue formation As multiple mitochondrial proteins were identified in both the Y2H and RNAi screens, I wondered whether the PCP defects I observed upon depletion of mitochondrial proteins were specific to these particular components or if disruption of mitochondria in general led to perturbation of PCP. To distinguish these two possibilities, I selected a panel of genes that represent various mitochondrial complexes and processes then drove expression of dsrna against these candidates using eygal4 and examined adult eye sections. With the exception of CG6485 and parkin, which were indistinguishable from wildtype, all other gene knockdowns resulted in some form of disruption in eye development (see Table 2.2), thus making further interpretation of PCP difficult at best. Nonetheless, in a subset of these eyes especially those in which components of the OXPHOS complexes had been knocked down I did observe some PCP defects, including ommatidial rotations/inversions and symmetric ommatidia (Fig. 2.7). Though the presence of malformed ommatidia introduces uncertainty, these results imply a role for OXPHOS complexes in PCP regulation Characterizing and verifying specificity of the Ndufv2 RNAi phenotype There had been no previous connections between PCP and mitochondria, and so it was quite peculiar to discover that knockdown of Ndufv2 could cause such clear and distinct PCP defects. Therefore, I then sought to more carefully characterize this RNAi phenotype and validate that it was indeed due to depletion of Ndufv2 rather than a non-specific off-target effect. To this end, I used two additional transgenic RNAi lines targetting Ndufv2; the three lines used were independent insertions of different targeting constructs. Tangential sections of eyes depleted of Ndufv2 using each of these lines driven by eygal4 displayed dorsal-ventral ommatidial inversions (Fig. 2.8) making off-target effects less likely to be responsible for this
52 41 Table 2.2: Summary of PCP phenotypes upon RNAi knockdown of mitochondrial genes Mitochondrial Complex Components: CG Number Name Complex Subunit VDRC line # eygal4> phenotype CG13240 Ndufb6 I Beta DV inversions, extra/missing/abnormal PRs CG15434 Ndufa2 I Lambda extra/abnormal PRs symmetric ommatidia, extra/misshapen PRs CG2286 Ndufs1 I Lambda DV inversions, symmetric ommatidia, ommatidial rotation defects, missing/abnormal PRs DV inversions, symmetric ommatidia, ommatidial rotation defects, missing/abnormal PRs CG5703 Ndufv2 I Lambda DV inversions, ommatidial rotation defects, missing/abnormal PRs DV inversions, ommatidial rotation defects, missing/abnormal PRs 5703R-1 DV inversions, ommatidial rotation defects, missing/abnormal PRs CG4169 Core Protein II III abnormal PRs CG4769 Iron-Sulfur (Heme) III 9180 none none DV inversions, ommatidial rotation defects, missing/abnormal PRs CG14724 CO-Va IV * sub-viable 3 rd instar eye-antennal discs overgrown/duplicated CG1746 V DV inversions, symmetric ommatidia, ommatidial rotation defects, missing/abnormal PRs CG3731 Mpp N/A none DV inversions, abnormal PRs 3731R-1 none CG6485 Ndufv2 N/A none paralog none Other mitochondrial perturbations: Conditions Phenotype UAS- Gal4 T (ºC) Opa1-like RNAi (VDRC#106290) ey 25 abnormal PRs/ommatidia Parkin RNAi (VDRC#104363) ey 25 none *stock provided by Utpal Banerjee Abbreviations: DV = dorsal-ventral, PR = photoreceptor




53 Figure 2.7: Knockdown of OXPHOS genes leads to defects in eye development and PCP. Tangential sections of adult eyes expressing dsrna directed against components of mitochondrial complexes in the OXPHOS system display photoreceptor and polarity defects. In polarity diagrams, black and grey arrows indicate normal ventral and dorsal chirality respectively, red arrows represent abnormal polarity, yellow arrows indicate achiral/symmetric ommatidia, and open circles represent ommatidia with aberrant morphology that were not scored for polarity. dsrna expression was driven with eygal4 at 25 or 29ºC. 42


54 A 43
55 Figure 2.8: Ndufv2 depletion by multiple independent RNAi lines leads to polarity defects. (A) Tangential sections of adult eyes with diminished Ndufv2 expression. eygal4 driven Ndufv2 dsrna from the VDRC (lines and ) and the NIG (line 5703R-1) results in PCP defects. The NIG line also displays defects in ommatidial morphology as denoted by the black open circles; these are not scored for polarity. Yellow lines demarcate the endogenous equator. (B) Western blots reveal larval wing disc protein levels of various Ndufv2 RNAi lines driven by NubGal4 and engal4 in the wing. Actin serves as a loading control. 44
56 45 polarity phenotype. All of these also resulted in some ommatidial/photoreceptor development defects; I found that VDRC line # and NIG line 5703-R1 consistently had more of these other defects than VDRC line # This perhaps reflects a difference in efficiency of knockdown between these different lines. I confirmed that Ndufv2 protein levels were knocked down using Western blots on larval wing disc extracts in which the three RNAi lines were driven (Fig. 2.8). Indeed these lines did result in varying levels of knockdown. Due to the more moderate knockdown in line #22194, I chose to further characterize the PCP phenotype using this line to minimize other confounding defects, i.e. abnormalities in ommatidial development. However, even with this restriction, I still observed a wide range of severity in both eye surface defects and underlying ommatidial development and polarity abnormalities with Ndufv2 knockdown, likely due to the variable nature of using RNAi. In an attempt to limit this variability I first surveyed its extent; I categorized knockdown flies based on severity of surface phenotype, giving them a score from 0, indicating no phenotype, to 3, indicating the most severe phenotype, and then counted the proportions of flies in each category (see Fig. 2.9). Using this approach, I observed a pronounced shift towards more severe phenotypes in males compared to females. This is likely attributable to Ndufv2 s position on the X-chromosome. I also observed more severe phenotypes in flies raised at 29ºC over those raised at 25ºC, as expected due to stronger activity of the Gal4/UAS system at the higher temperature. I reasoned that flies with a moderate surface phenotype reflected fewer defects in ommatidial development, and thus selected those from category 2 to examine for PCP defects. Indeed this was the case; I found that in these flies, ommatidia were normally formed, while still showing some PCP defects. Specifically, male category 2 flies raised at 25ºC show on average 4.15% dorsal-ventral inverted ommatidia (Fig. 2.10). Having established this clean PCP phenotype, I sought to further confirm that it was due to specific depletion of Ndufv2. First, to verify that Ndufv2 was in fact knocked down in these RNAi experiments, I used immunofluorescence imaging of imaginal wing discs in which I drove dsrna against Ndufv2 using engal4. To visualize Ndufv2 protein levels, I generated an antibody against Ndufv2; briefly, full-length GST-tagged Ndufv2 was expressed and purified from bacteria and injected into host animals, then sera were extracted and used as a polyclonal antibody to probe for Ndufv2. Staining with this antiserum clearly showed a reduction of Ndufv2



 Dorsal and lateral views representative of adult fly heads for")



57 46 A Phenotype severity 0 - None 1 - Mild 2 - Moderate 3 - Severe B 100.0% 90.0% 80.0% 70.0% 60.0% 50.0% 40.0% 30.0% 3 - Severe 2 - Moderate 1 - Mild 0 - None 20.0% 10.0% 0.0% Ndufv2 KD - 25º Ndufv2 KD - 25º Ndufv2 KD - 29º Ndufv2 KD - 29º Figure 2.9: Expression of Ndufv2 RNAi in the Drosophila eye gives rise to a highly variable head cuticle dysplasia phenotype. (A) Dorsal and lateral views representative of adult fly heads for each of the following categories: 0 no phenotype; 1 mild phenotype, missing some ocelli structures on dorsal side of head, some abnormal bristles, little to no roughness of the eye surface; 2 moderate phenotype, missing ocelli, abnormal bristles, moderate eye roughness; 3 severe phenotype, cleft in head cuticle instead of ocelli, abnormal bristles and antennae, severe roughness/bulging/cuticular growths in eye. (B) Graph scoring all progeny, separated by sex, from two individual crosses at the temperatures indicated.
58 Figure 2.10: Quantification of PCP defects upon Ndufv2 depletion Polarity diagrams created from tangential sections of adult Drosophila eyes in which Ndufv2 RNAi (line #22194) is driven with eygal4 at 25ºC. Eyes were from male flies of moderate surface phenotype (category 2, see Fig. 2.9A). Green and red arrows represent ommatidia with dorsal and ventral chirality respectively. Yellow arrows represent achiral ommatidia, not included in this quantification. Black line represents the endogenous equator separating the dorsal (above equator) and ventral (below equator) halves of the eye. Numbers below each diagram indicate the number of DV inversions (red arrows on the dorsal side or green arrows on the ventral side) divided by the total number of ommatidia to obtain a percentage frequency of DV inversions. 47
59 48 levels in posterior compartments of wing discs where dsrna against Ndufv2 had been expressed (Fig. 2.11). Thus, I confirmed that Ndufv2 RNAi lines do knock down Ndufv2 levels, and also that the antibody I generated against Ndufv2 is specifically reactive. To fully establish the specificity of the Ndufv2 knockdown PCP phenotype, I attempted a rescue experiment. I generated transgenic fly lines harbouring full-length Ndufv2 cdna under the control of UAS. I tested for expression of this transgene by driving it with engal4 and found that Ndufv2 levels were significantly increased in posterior compartments of wing discs (Fig. 2.11). Notably, this staining strongly co-localizes with CVα, an established mitochondrial marker. I then co-expressed the UAS-Ndufv2 transgene simultaneously with dsrna against Ndufv2 in the eye using eygal4 and compared to eyes that expressed either transgene alone. I found that expression of the UAS-Ndufv2 transgene was able to fully rescue the RNAi phenotype; where the Ndufv2 knockdown animals had eye and head tissue hypoplasia, the rescued eyes were completely normal on the surface. I then scored for PCP in sections and observed a complete rescue of PCP defects as well (Fig. 2.12). These data confirm that the PCP defects I observed in the Ndufv2 RNAi experiments are specific to the depletion of Ndufv2 and thus support a role for Ndufv2 in the regulation of PCP. Of interest here, eyes in which I drove only UAS-Ndufv2 with eygal4 were also mostly normal on the surface as well in terms of ommatidial development; however, I did see rare polarity defects. One of four eyes sectioned contained ommatidia with reversed AP polarity (Fig. 2.12). Many other PCP genes also cause polarity defects when misexpressed. Therefore, these data further implicate Ndufv2 as a PCP regulator. Ndufv2 is a highly conserved member of CI, which provides an entry point for electrons in the ETC in eukaryotes. Ndufv2 has a highly conserved iron-sulfur (Fe-S) cluster that has been hypothesized to act as an antioxidant to regulate the generation of ROS (Brandt, 2006; Hirst, 2013). I tested if the Fe-S cluster of Ndufv2 is essential for normal PCP by mutating two conserved cysteine residues that coordinate binding of the Fe-S cluster, generating UAS- Ndufv2ΔFeS. This transgene was inserted into the same ϕc31 recombination site I had used to generate the UAS-Ndufv2 transgene, thus ensuring similar expression levels. I confirmed this by expressing UAS-Ndufv2ΔFeS with engal4 and saw comparable levels of expression to UAS-
 engal4-driven expression of Ndufv2 dsrna in the posterior compartment of the wing imaginal disc (as marked by")
60 Figure 2.11: Specificity of anti-ndufv2 antibody and expression levels of Ndufv2 transgenes (A) engal4-driven expression of Ndufv2 dsrna in the posterior compartment of the wing imaginal disc (as marked by αen in red) demonstrates both efficacy of Ndufv2 knockdown (VDRC line 22194) as well as specificity of Ndufv2 antibody. UAS-Ndufv2 and UAS-Ndufv2 FeS transgenes are ectopically expressed by engal4 at similar levels. (B) Ndufv2 co-localizes with CVα in wildtype wing discs. Boxed region shown in higher magnification in lower panels. 49

61 Figure 2.12: UAS-Ndufv2 and UAS-Ndufv2 FeS transgene expression during eye development Tangential eye sections taken from progeny of the indicated crosses. Polarity is diagrammed below each micrograph. Black arrows represent normal polarity, red arrow represents DV inversion, blue arrow represents AP inversion. 50
62 51 Ndufv2 (Fig. 2.11). Further, Ndufv2ΔFeS appeared to localize properly to mitochondria. I then drove UAS-Ndufv2ΔFeS and Ndufv2 dsrna in the eye, but I was unable to fully rescue the PCP defects induced by knockdown of Ndufv2 (Fig and Table 2.3). Thus, the Fe-S cluster of Ndufv2 is necessary for normal PCP Knockdown and overexpression of Ndufv2 in clones testing nonautonomy Given that ft pathway mutants have non-autonomous PCP defects, I postulated that Ndufv2 might also phenocopy ft in this respect. To test this, I drove expression of dsrna against Ndufv2 in heat-shock induced mitotic clones using the FLP-out system (Struhl and Basler, 1993). I also expressed RNAi against w to mark the boundaries of the clone in the adult eye. When I examined sections from eyes containing such clones, I found that there were occasional, mild autonomous PCP defects: approximately 10% of clones examined had misrotated ommatidia (Fig. 2.13). I did not observe any non-autonomous effects in these experiments. In addition to this PCP phenotype, I often saw disruptions in normal photoreceptor formation ommatidia had extra, missing or misshapen rhabdomeres. Moreover, I found that large Ndufv2 knockdown clones were difficult to recover, often resulting in eyes with only very small or no clones at all. Taking into consideration Ndufv2 s role in metabolism, along with its effects on cuticle development I had seen with eygal4 driven knockdown, these data indicate an essential function for Ndufv2 in cell viability or possibly in cell competition. As I had observed some mild disruptions in PCP upon global misexpression of Ndufv2, I reasoned that ectopic expression of Ndufv2 might also produce PCP defects and uncover any potential non-autonomous effects. I drove UAS-Ndufv2 in mitotic clones similarly to the RNAi experiment above. I examined five such clones and found that overexpression of Ndufv2 in clones does indeed induce both autonomous and non-autonomous polarity defects: two clones displayed misrotated ommatidia, one clone had an autonomous DV inversion, while the last clone, remarkably, contained one autonomous and two non-autonomous DV inversions on its equatorial border (Fig. 2.13). Intriguingly, stronger PCP defects (DV inversions) were located farther from the equator (>10 rows of ommatidia) while the misrotations were nearer to the equator (within three rows). This indicates a possible positional effect of Ndufv2 on PCP. Intriguingly, both ds and atro (members of the ft PCP pathway) also show positional effects in
63 52 Table 2.3: Quantification of PCP defects in Ndufv2 transgene rescue experiments Genotype eygal4>uas-ndufv2rnai DV inverted ommatidia Total ommatidia Percent inversion eygal4>uas-ndufv2rnai UAS-Ndufv eygal4>uas-ndufv2rnai UAS-Ndufv2ΔFeS
 or UAS-Ndufv2 transgene.")
64 Figure 2.13: Perturbations of Ndufv2 levels in mitotic clones results in autonomous and non-autonomous polarity defects. Tangential sections taken from adult Drosophila eyes in which mitotic clones have been generated that either express Ndufv2 RNAi (left) or UAS-Ndufv2 transgene. Polarity is diagrammed below each micrograph. Clone tissue in white, surrounding wild type tissue is shaded. Black arrows represent normal polarity, red arrows represent DV inversion, blue arrow represents misrotation, open circles represent ommatidia with abnormally formed photoreceptors. 53
65 54 PCP regulation (Sharma and McNeill, 2013; Strutt and Strutt, 2002) suggesting that Ndufv2 acts on PCP in a similar manner to the ft pathway, or is itself within the ft pathway. Besides this striking PCP phenotype, I also observed an unexpected effect on cell viability when I overexpressed Ndufv2 in clones. As with RNAi-mediated knockdown of Ndufv2 in clones, ectopic expression also resulted primarily in small clones or no clones at all, suggesting either these cells are dying or being competed out. While this effect remains unexplained, it does indicate that proper levels of Ndufv2 are essential for cell survival Direct physical interaction of Fat and mitochondrial complex proteins The Y2H and RNAi screens identified several mitochondrial proteins as potential interactors of Ft. In order to confirm these interactions and additionally test if they are physical and direct, we used GST pull-down assays with purified fragments of the Ft cytoplasmic domain and Histagged Ndufv2 (Fig. 2.14B) or CG1746 (Fig. 2.14C). In each of these assays, we saw both Ndufv2 and CG1746 co-precipitate with Ft fragments, but not with GST alone controls. We also dissected the intracellular fragment of Ft to further characterize its interaction with mitochondrial proteins and found that Ndufv2 and CG1746 bind directly to the highly conserved regions 1 and 2 of Ft (Fig. 2.14A). Strikingly, deletion of four amino acids of Region 2 abrogates interaction with Ndufv2. We found that Region 1 also contributes to binding stability between Ft and Ndufv2, but is not sufficient to confer binding in this assay. These data indicate that the highly conserved region 2 is critical for direct binding between Ft and Ndufv2 in vitro Genetic interactions between Ndufv2 and the ft pathway Ndufv2 depletion in a ft heterozygous genetic background Having established that Ft and Ndufv2 physically interact, I then attempted to determine if Ndufv2 and ft also interact genetically. Accordingly, I expressed dsrna against Ndufv2 using eygal4 or nubgal4 in a ft heterozygous genetic background (ft fd /+), then examined these animals wings and eyes for indications of genetic enhancement/suppression. ft heterozygous animals themselves appear wholly wild type in terms of PCP phenotypes in the eye. Therefore any modification in the severity of the Ndufv2 eye phenotype in this background would suggest a genetic interaction with ft.
66 Figure 2.14: Ndufv2 and CG1746 directly interact with highly conserved regions in the intracellular domain of Ft. (A) Schematic of the intracellular domain of Ft with eight highly conserved domains numbered in red boxes. Constructs used in binding studies are shown below, with peptide sequence for the C-terminus of the last three constructs shown in inset. Binding data is summarized to the right. (B and C) In-vitro-GST pull-down using purified His-Ndufv2 (B) or His-CG1746 (C) indicates interaction with highly conserved regions 1 and 2. Ponceau S serves as loading control. 55
67 56 I first assessed the surface dysplasia phenotype in the eygal4-driven case. I found that in male flies, removing a copy of ft slightly worsens the phenotype compared to Ndufv2 knockdown alone, while in females, there is a minor improvement (Fig. 2.15). However, neither of these differences are nearly as dramatic as the shift towards more severe phenotypes I observed when flies were raised at higher temperatures (Fig. 2.9). In addition, there was variability between different trials even under identical conditions (compare males depleted of Ndufv2 alone raised at 25ºC in each of these independent trials Fig. 2.9 and Fig. 2.15); thus it is difficult to conclude if these minimal changes observed are due to ft heterozygosity or to variability of the RNAi phenotype itself. I then examined PCP in these same eygal4-driven Ndufv2 knockdown animals by looking at adult eye sections. To reduce variability, I only used male flies with moderate levels of eye/head tissue dysplasia as I had done during quantification of Ndufv2 RNAi PCP defects (see 2.3.3). I found that overall percentage of PCP defects were similar regardless of the presence of 1 or 2 copies of ft I saw 2.03% (± 4.62%, n=6 eyes) DV inversions in the ft heterozygotes, versus 2.66% (± 3.10%, n=8 eyes) in the wild type background. Another readout of ft pathway activity is wing patterning; the cross vein distance (CVD) between the anterior and posterior crossvein is appreciably shorter in ft mutants and can be rescued by expression of full-length ft (Pan et al., 2013). To test if Ndufv2 is also involved in these processes, I drove dsrna against Ndufv2 in the wing pouch using nubgal4, and measured CVD as well as the length of wing vein 3 (V3) (see Fig. 2.16A for example measurements). I found that both CVD and V3 length were significantly shorter in nubgal4-driven Ndufv2 RNAi wings than in nubgal4, ft fd /+ control wings (Fig. 2.16B and C note, I have not compared CVD or V3 lengths of nubgal4, ft fd /+ wings to wild type wings, thus I do not know if ft heterozygotes themselves have abnormal CVD and V3 length). This reduction in CVD and V3 in Ndufv2-depleted wings may be a consequence of disrupted ft signaling; however, it could also simply be perturbations in normal wing development resulting in an overall smaller wing. Indeed, many of these wings were either crumpled or fragile, displaying some morphological defects such as extra-pigmented regions or disrupted veins and wing margins (Fig. 2.16A).
68 % 90.0% 80.0% 70.0% 60.0% 50.0% 40.0% 30.0% 3 - Severe 2 - Moderate 1 - Mild 0 - None 20.0% 10.0% 0.0% Ndufv2 KD - 25º Ndufv2 KD - 25º Ndufv2 KD, fd/+ - 25º Ndufv2 KD, fd/+ - 25º Figure 2.15: ft heterozygosity mildly enhances head dysplasia phenotype of Ndufv2 depletion in males Graph scoring proportions of progeny, separated by sex, in each category (see Fig. 2.9 for category descriptions) from an Ndufv2 knockdown cross (eygal4 X UAS-Ndufv2- RNAi-22194) either in a wildtype or ft heterozygous background (ft fd /+).
 Scatter plots of CVD (B) and V3 (C) measurements. Line represents mean.")
69 Figure 2.16: ft heterozygosity increases crossvein distance and vein 3 length in Ndufv2-depleted wings. (A) Micrographs of adult wings from indicated genotypes. Ndufv2 RNAi was expressed in the wing pouch with nubgal4 at 25ºC. Red line marks CVD, blue line marks V3 length. (B and C) Scatter plots of CVD (B) and V3 (C) measurements. Line represents mean. Removal of one copy of ft results in a significant rescue of CVD and V3 length (**p=0.0085, *p=0.032, 2-tailed t test). 58
70 59 Confusingly, when I removed a copy of ft from these Ndufv2-depleted wings, I observed a significant increase in both CVD and V3 length over Ndufv2 RNAi alone (p = for CVD and p = for V3 length). This is puzzling as loss of ft or other ft pathway components normally leads to a decrease in CVD. This result suggests that ft could be interacting with Ndufv2 in complex ways; alternately, ft heterozygotes may behave differently from ft nulls in terms of CVD phenotypes Overexpression of Ndufv2 in ft tissue results in synthetic cell lethality I had previously observed that misexpression of Ndufv2 resulted in perturbations in PCP; I wondered if this effect required ft function. To test this, I drove overexpression of Ndufv2 in a ft clone using the MARCM system. I made such clones in the eye and examined tangential sections from newly eclosed adults. Unexpectedly, these sections displayed an acute degeneration phenotype; the photoreceptors within clones appeared to have severely deteriorated, leaving large vacuolated areas in the eye (Fig. 2.17A). Though ft clones in the eye do experience some ommatidial degeneration, it is far less severe and also only occurs in older flies. Widespread expression of Ndufv2 did not result in any visible disruptions to photoreceptor morphology or survival (Fig. 2.17B), but I have already noted that overexpression clones were difficult to recover; taken together, these data indicate that local misexpression of Ndufv2 leads to cell lethality. Furthermore, ft clones do not display this severe degeneration phenotype, nor did the small Ndufv2 overexpression clones that were ultimately recovered; thus, there appears to be a synergistic effect between these two genes in terms of cell viability. Unfortunately, photoreceptor degeneration within the clone precluded analysis of PCP in these eyes, and thus I was unable to determine if ft is necessary for Ndufv2 s PCP phenotype. Nonetheless, these experiments suggest a genetic interaction between ft and Ndufv2.
. (B) Overexpression of Ndufv2 in the dorsal half of the eye using DE-Gal4 has no illeffects on photoreceptor/ommatidial development.")
71 Figure 2.17: Overexpression of Ndufv2 in ft - eye tissue causes severe degeneration of ommatidia. (A) Tangential eye section containing MARCM ft - clones expressing UAS-Ndufv2 transgene. Clone tissue is marked by absence of pigment (white RNAi). (B) Overexpression of Ndufv2 in the dorsal half of the eye using DE-Gal4 has no illeffects on photoreceptor/ommatidial development. 60
72 Loss of Ndufv2 leads to activation of Hippo and PCP target genes in larval wings and eyes Because we found that Ndufv2 binds directly to Ft, and that loss of Ndufv2 alters PCP, we examined if Ndufv2 knockdown affects Ft pathway target expression. A well-characterized PCP target of Ft is four-jointed (fj). Loss of ft leads to upregulation of fj in mitotic clones, as assessed by fj-lacz (Fig. 2.18A and (Yang et al., 2002). To test whether Ndufv2 also affects fj expression, I generated clones of cells expressing Ndufv2 dsrna in eye discs from flies containing a fj-lacz reporter. fj-lacz expression was clearly increased in Ndufv2 dsrna clones, indicating that Ndufv2 regulates expression of the Ft-PCP target fj in vivo (Fig. 2.18B). As ft has been shown to be upstream of the Hippo pathway, we wondered if Ndufv2 also played a role in Hippo signaling. Activation of the Hippo pathway can be observed in vivo by monitoring expression of Yki target genes such as Crumbs (Crb), Bantam and Expanded (Ex). Expression of Ndufv2 dsrna in the posterior compartment of wing discs with engal4 resulted in marked upregulation of Crb (Fig. 2.18C) and Ex (Fig. 2.18D). Clones of cells expressing Ndufv2 dsrna also showed upregulation of Ex (Fig. 2.18E). Bantam-GFP, which is inhibited by Hippo signaling, was repressed by depletion of Ndufv2 (Fig. 2.18F). Taken together, these data show that loss of Ndufv2 alters expression of Hippo targets. Presumably changes to Hippo targets could result in alterations in growth regulation. While Ndufv2 knockdown does not show classical Hippo-like overgrowth phenotypes, I did observe perturbations in head cuticle growth. I wondered if these effects were accompanied by changes in proliferation or apoptosis. I first tested whether proliferation was affected by carrying out an EdU incorporation experiment. I drove dsrna against Ndufv2 in the posterior compartment of third instar wing discs and observed a marked increase in proliferation (Fig. 2.19B), consistent with disruption of the Hpo pathway. I then examined activated Caspase staining in Ndufv2-depleted wing discs, and revealed that strong knockdown of Ndufv2 led to increased apoptosis indicating disruption of growth regulation (Fig. 2.19A). Hippo pathway mutations generally result in increased proliferation but suppressed apoptosis. Increase in apoptosis in the case of Ndufv2 knockdown could be caused by deficits in other mitochondrial functions certainly, mitochondria s role in apoptosis has been well characterized and so, a Hippo growth phenotype might be obscured by these other effects.
 Hippo targets Crumbs (Crb C) and Expanded (Ex D) are upregulated in the posterior compartments of larval wing discs expressing Ndufv2 dsrna driven with engal4.")
73 Figure 2.18: ft and Ndufv2 regulate PCP and Hippo pathway targets. (A-B) fj-lacz is upregulated in both ft fd clones (A) and clones expressing Ndufv2 dsrna (B) in the larval eye. (C-D) Hippo targets Crumbs (Crb C) and Expanded (Ex D) are upregulated in the posterior compartments of larval wing discs expressing Ndufv2 dsrna driven with engal4. Engrailed (En) marks the posterior compartment. (E) Ex is also upregulated in Ndufv2 knockdown clones in the wing. (F) Bantam-GFP, a Hippo sensor is reduced in Ndufv2 knockdown tissue. ft - clones marked by lack of GFP, Ndufv2 knockdown clones positive for GFP. 62
 causes increased apoptosis, as shown by αcaspase staining (A), and")
74 Figure 2.19: Depletion of Ndufv2 results in increased cell death and increased proliferation. Expression of Ndufv2 dsrna in posterior compartments of wing discs (marked by αen) causes increased apoptosis, as shown by αcaspase staining (A), and increased proliferation, as shown by EdU incorporation (B). 63
75 Discussion Ft is essential for distinct and genetically separable processes during development, namely growth control and planar cell polarity. Much effort has been made to study how Ft functions in each of these, however few physical interactors of Ft had been identified to define a mechanism by which Ft acts in these pathways. Here, I have made the unexpected discovery that Ft directly interacts with components of the mitochondrial import machinery and OXPHOS system. Further, I have shown that depletion of mitochondrial proteins, especially Ndufv2, leads to disruptions in normal PCP regulation. I have also shown that Ndufv2 genetically interacts with ft in wing patterning and cell viability. Remarkably, I have found that loss of Ndfuv2 also impacts ft PCP and Hpo targets. No previous studies had implicated mitochondria in these processes, and so these findings shed new insight on how Ft may be functioning during development in vivo RNAi screen Through our Y2H screen, we produced a list of candidate Ft interactors; however, we still needed to verify if any of these candidates were actually involved in ft signaling. With the advent of readily available RNAi stocks targeting the D. melanogaster genome, we were able to systematically test each of these candidate genes for ft-like phenotypes in vivo. In this way we identified one gene potentially involved in growth control: CG9723 or speggless, the subject of my colleague, Robyn Rosenfeld s PhD work. We also identified four genes that regulate PCP, which will be discussed further below. The other candidates from the Y2H proved to show no phenotype, very mild PCP deficiencies, or a cell/organism lethality phenotype (see Table 2.1 for summary). Our candidates that resulted in no phenotype when knocked down could indicate that these genes are either false positives from the Y2H screen (i.e. they do not interact with Ft) or that their interaction with Ft is biologically irrelevant to the system we interrogated, namely, growth and PCP in the eye. One other possibility is that the RNAi stocks used were not effective, or did not sufficiently deplete the target to manifest a phenotype. While we tried to ensure that these dsrna were indeed expressed by using various conditions and gal4 drivers, we cannot exclude the possibility that we did not in fact knock down each candidate, as verifying each knock down was not feasible in this screen. To address these issues, colleagues in our laboratory have since
76 65 completed a second round of screening by expressing these RNAi lines in the context of adult Drosophila wings; their data have largely confirmed our eye screen results. Similarly, the set of genes that produced very mild PCP defects, occasional symmetric or misrotated ommatidia, could also be due to insufficient knockdown, and thus we saw weaker phenotypes. Alternately, these genes were adequately depleted, yet still displayed weak phenotypes. Either way, these mild defects could be pointing to disturbances in other processes during eye development: symmetric ommatidia can result from disruptions in photoreceptor fate determination. Differences in Notch signaling between prospective receptors 3 and 4 are the determining factor for asymmetric development of the photoreceptor cluster (Fanto and Mlodzik, 1999; Tomlinson and Struhl, 1999). Ommatidial misrotation is indicative of PCP defects; indeed mutations in Core PCP genes often result in such misrotations. However, eye specific downstream effectors of PCP may also be responsible here; for example, the kinase encoded by nemo is essential for proper ommatidial rotation, as are the cell adhesion molecules Echinoid and Friend-of-Echinoid. Our candidate genes that displayed these mild phenotypes may be participating in any of these pathways; however, their relatively impenetrant phenotypes precluded simple analysis. More detailed work will be necessary to determine if these genes are in fact involved with ft or PCP signaling. The last group of genes in our screen resulted in eye phenotypes so severe that they masked any observation of potential growth or PCP effects. Knockdown of some of these genes induced lethality. It may seem unusual for localized expression of these RNAi constructs to have such an effect, as we used eye specific Gal4 drivers; while it is possible that these drivers are not completely restricted in their expression domains and consequently leaky expression of dsrna disrupted normal development in other tissues, we find that the identity of these particular genes that caused lethality when depleted provide a more likely explanation: most of these candidates encoded components of the translational machinery. CG2238 is an elongation factor (ef2b), and CG3203, CG5271, CG7808, CG8332 are all ribosomal subunits. It is not surprising that knockdown of these genes had such adverse effects. Whether these interact with Ft or ft-related pathways remains unexplored. The remaining non-lethal genes when depleted caused severely disrupted eyes; ommatidia and photoreceptors were not properly formed. These candidate interactors could be involved in any number of other functions essential to eye development: cell proliferation, specification, patterning, etc. These defects all render interpretation of polarity
77 66 impossible, and as such, the adult eye has proven to be a poor system to study these specific genes. The wing screen undertaken by my colleagues has helped to address some of these concerns. Briefly, they surveyed for Hpo-like phenotypes by measuring overall wing size and distance between anterior and posterior crossveins. They also examined wing hairs and ridges for PCP defects. These results are summarized in Table PCP regulators identified Despite some of the barriers to finding ft interactors mentioned above, our RNAi screen did identify several hits: CG5703 (Ndufv2), CG3731 (MPP), CG1746 (subunit of V0-ATP Synthase), and CG5468 (twdlm). These four genes produced strikingly similar PCP phenotypes when knocked down they all displayed DV inverted ommatidia. This is quite remarkable in that this phenotype is specific to mutations in the Ft/Ds PCP regulatory pathway. This pathway is thought to coordinate longer range, tissue-wide organization of polarity, in contrast to the Fz/Dsh core pathway, which is responsible for, at least in the eye, reading the Ft/Ds global signal then establishing and reinforcing local polarity between neighbouring cells. As a result, mutants of these two PCP pathways manifest in the eye in different characteristic ways. Eyes deficient in the core pathway tend to display symmetric ommatidia (photoreceptors R3 and R4 are improperly specified), or misrotated ommatidia (asymmetric ommatidia that do not respond or respond incorrectly to the global polarity cue). In contrast, Ft/Ds pathway mutants have normally formed ommatidia, but randomly specify dorsal-ventral chirality due to a lack of tissue-wide polarity information, ultimately resulting in DV inversions across the eye field. As this characteristic DV inversion phenotype was the main polarity phenotype observed for the four hits from our RNAi screen, these four candidates are likely operating to establish global planar polarity cues during eye development. Moreover, as these are also candidate Ft interactors from our Y2H screen, these four genes are also likely to be working in the Ft pathway. One of the four hits, twdlm, is particularly baffling. The tweedle family of proteins were first isolated in screens for altered body shape and were found to be cuticle related proteins (Guan et al., 2006). These proteins appear to be insect specific. A second tweedle protein, TwdlB (CG6478), was also identified in our Y2H screen, though it only gave very mild PCP defects upon knockdown (slight misrotations and rare symmetric ommatidia, see Table 2.1). It is unclear how secreted cuticle structure proteins would be interacting with Ft to affect PCP, as no other
78 67 Table 2.4: Summary of RNAi screen of Ft interactors in the wing Genes with growth/proximal-distal patterning defects: Crossvein distance Wing area (% of wt) (% of wt) CG# Gene name/description female male female male CG2849 ras-like protein A (rala) CG12052 longitudinals lacking (lola) CG15023 novel; no known function CG14598 novel; no known function CG6715 KP78a CG13387 embargoed (emb) CG8651 trithorax (trx) CG3530 novel; tyrosine phosphatase activity CG7137 novel; methyltransefrase activity CG10645 lamina ancestor (lama) CG31477 ATP synthase, ε subunit-like (ATPsynεL) *Shaded boxes represent >5% deviations from wt in both males and females. Genes with wing PCP phenotypes (ridge/hair misalignment): CG# CG7660 CG9593 CG9723 Gene name/description peroxinectin-like (Pxt) novel; fibrinogen-like spegless (speg)
79 68 previous data have drawn this connection. One possible explanation for these PCP phenotypes is that they reflect off-target effects; all of the RNAi lines targeted to these two genes do have predicted off-targets, as the tweedle genes are all similar in sequence, thus RNAi against one of them is likely to affect other tweedles as well. In addition to the other tweedles, several other developmentally important genes are predicted off-targets, including suppressor of Hairless and trithorax/trithorax-like. Knockdown of these genes could have far-ranging consequences, perhaps impacting on other PCP determinants. While the twdlm phenotype was similar to ft, these other confounding factors made it difficult to study, and thus I chose to pursue the other hits from our RNAi screen Establishing planar polarity a mitochondrial signal? Perhaps the most extraordinary discovery of our RNAi screen is that three out of four of the hits from the screen are mitochondrial in nature. Until this study, mitochondria had never been implicated in PCP regulation, thus it was quite astonishing when I observed such precise ft-like PCP phenotypes in Ndufv2 depleted eyes. The two other mitochondrial genes from the screen, MPP and CG1746, also showed DV inverted ommatidia, though these were more often accompanied by defects in PR/ommatidial development. Furthermore, when I tested other mitochondrial genes for PCP phenotypes I found that several of these also induced DV inversions upon knockdown, albeit also with accompanying abnormal and malformed ommatidia. These other defects in eye development are perhaps foreseeable, given that mitochondria play well-studied roles in energy metabolism and apoptosis one would expect some detrimental effects to cell functions upon disrupting such processes. However, the DV inversion phenotype is highly specific to perturbations in eye field and pattern establishment pathways (Mirror/Fringe, Fat/Ds). My observations indicate that mitochondria are somehow working in one of these pathways that ultimately establish global polarity signals. This raises the question of how this might be occurring. My data have not elucidated what this mechanism is, but a few possibilities emerge Polar mitochondrial PCP signals In PCP regulation, gradients of PCP determinants are proposed to give global directional information across a tissue. Both fj and ds exist as gradients in imaginal discs, which are then
80 69 thought to create a gradient of ft activity. This would predict that the PCP phenotypes of mitotic clones in the eye would be sensitive to their position along these gradients. Our lab has found that the transcriptional co-repressor and ft interactor atrophin (atro) does in fact behave this way in the eye: atro clones have a much stronger effect nearer the equator indicating that atro is responsible for mediating PCP effects close to the equator (Sharma and McNeill, 2013). However, PCP is still normally established near the poles where atro does not have a strong effect, thus there must be additional mediators of PCP there. The positional effect I observe in Ndufv2 overexpression clones may be implicating Ndufv2 as a polar PCP factor OXPHOS and PCP One unresolved question early in this study was whether PCP was specifically regulated by Ndufv2 or more generally by mitochondria. While exploring this question, I came across another potential mechanism for mitochondrial PCP establishment. I surveyed many mitochondrial genes for PCP phenotypes. Strikingly, PCP defects were more pronounced in eyes depleted of OXPHOS genes (see Table several CI, one CIII and one CV component) in contrast to other mitochondrial genes (e.g. Parkin and OPA1-like, with no indications of polarity defects). This suggests that the OXPHOS complexes are specifically required for normal PCP establishment. The OXPHOS complexes are responsible for catalyzing the redox reactions of the electron transport chain. These are coupled to active pumping of protons across the inner mitochondrial membrane, resulting in the proton motive force. This proton gradient is ultimately harnessed by CV (ATP synthase) to phosphorylate ADP, thus producing an energy supply for the cell. Is PCP regulation linked to this process in some way? Previous data from our lab have shown that PCP establishment is largely unresponsive to energy levels AMP kinase clones in starved pupal retinas display normal polarity (Fig. 2.20C). Furthermore, ATP levels in ft mutants are not significantly altered (see Chapter ). Thus it is unlikely that the direct product of OXPHOS, ATP, controls PCP. Nonetheless, intact complexes with functioning catalytic cores are required for normal polarity, as I found that Ndufv2 with a mutated Fe-S cluster-binding pocket is unable to rescue the effects of Ndufv2 depletion by RNAi. Fe-S clusters serve as electron acceptors and donors in the ETC; the sensitivity of PCP regulation to the presence of Fe-S clusters may signify a requirement for normal redox chemistry in polarity determination, though what the relationship between these
 Fmi antibody staining (blue on left, white on right) is not affected in Ndufv2 RNAi clones (GFP positive on left, outlined in red on right) in 3 rd instar eye discs.")
81 Figure 2.20: Ndufv2 does not regulate polarity via core PCP, JAK/STAT, nor AmpK signalling pathways. (A) Fmi antibody staining (blue on left, white on right) is not affected in Ndufv2 RNAi clones (GFP positive on left, outlined in red on right) in 3 rd instar eye discs. First six rows of ommatidial clusters emerging from the morphogenetic furrow (MF) are represented here MF to the left of image (B) STAT92E-GFP reporter is not affected in 3 rd instar wing discs in which Ndufv2 RNAi is driven in the posterior using engal4 posterior to the right (C) Polarity is unaffected in ampk clones (GFP negative, clone border marked with dashed line) as assessed by polarity markes Arm and apkc in pupal retinas. 70
82 71 two processes is remains unclear. One attractive possibility for such an intermediary is reactive oxygen species (ROS) Are ROS PCP signals? ROS, particularly the superoxide anion (O2 - ), are produced in mitochondria when electrons leak from the ETC during respiration. At high levels, these reactive species have a negative effect on cellular function; yet intriguingly, a complete lack of ROS is also detrimental to the cell as they are crucial signaling molecules in a number of regulatory pathways. O2 - itself acts on a rather limited range, as it is highly reactive and has a short half-life, but through either spontaneous reaction or via the action of superoxide dismutases (SOD), O2 - is reduced to H2O2 which is more readily diffusible (Handy and Loscalzo, 2012). Previously established models of PCP determination in the eye have intimated the existence of a diffusible second signal to account for its long range action; could ROS serve as this signal? CI and CIII are the major sites of mitochondrial ROS production; indeed, when I depleted CI and CIII components in wing discs, I observed an increase in ROS (see Fig. 3.5C, D). Knockdown of these same genes led to PCP defects in the eye (Table 2.2), consistent with my model of ROS as a PCP signal. Furthermore, structural studies of the Fe-S cluster of Ndufv2 have shown that its position within CI excludes it from direct participation in the ETC (Sazanov and Hinchliffe, 2006). Rather, it has been proposed to be an acceptor or temporary sink for stray electrons, thus serving as a ROS regulator. Taken together with Ndufv2ΔFeS s inability to rescue PCP defects, ROS are well positioned to be a crucial component of PCP regulation. Two more observations I made also suggested a relationship between ft, ROS and PCP. The first was that ROS exists as a gradient in the eye disc (see Fig. 3.5E). The ROS gradient is reminiscent of the fj expression gradient in the eye, thus raising the possibility that it may serve as a directional cue across the tissue. The second observation that supported a role for ROS in ft signaling was that ft mutant tissue dysregulates ROS (see Fig. 3.6A, B). Since ft is able to modulate ROS levels, it may also be responsible for setting up the ROS gradient. For these reasons, I exhaustively carried out numerous experiments perturbing ROS levels in vivo, assaying for PCP disruption. Much to my disappointment I could not convincingly show that ROS were acting as PCP signals; I only observed very rare and mild PCP defects with this
83 72 line of inquiry (see Table 2.5). I did meet some technical limitations in these experiments (e.g. complicated genetics, efficient drug delivery, lack of adequate apparatus for larval immobilization, difficulty achieving temporal/spatial precision or degree of expression of ROS perturbing tools, etc.). Nevertheless, I had expected more robust PCP phenotypes, given my previous results suggesting ROS involvement. While the negative results summarized on Table 2.5 cannot completely exclude ROS as an actor in PCP regulation, they imply that ROS are neither solely nor primarily responsible for normal PCP determination. What other mitochondrially-derived signals then could be mediating Ndufv2 s effect on PCP? In addition to its role as a nexus for metabolic processes and pathways, the mitochondrion also serves as a hub for many crucial signaling functions, including oxygen sensing, regulation of apoptosis and the redox state of the cell. These signaling pathways provide several other candidates for a PCP signal originating in mitochondria. One possibility is nitric oxide (NO ), a relative to ROS and also a reactive radical. NO is a small lipophilic molecule that readily diffuses through plasma membranes, thus allowing it to act on neighbouring cells. It has been shown to function as a second messenger in a multitude of processes, ranging from vasodilation of cardiac vessels to neuronal patterning in the eye (reviewed in Hamanaka and Chandel, 2010). It acts by reversibly modifying thiol groups, particularly cysteine residues in enzymatic active sites, thus producing a change in protein function. NO is synthesized by nitric oxide synthase (NOS), one isoform of which has been found to reside within mitochondria. NOS staining in eye imaginal discs has revealed an intriguing gradient pattern resembling that of fj stronger at the equator and weaker towards the poles (Kuzin et al., 1996), potentially providing directional information across the tissue. NO can interact with Cytochrome C Oxidase (Complex IV) to modulate throughput of the ETC (Handy and Loscalzo, 2012), but it is not immediately apparent if or how Ndufv2 and other OXPHOS complexes affect NO dynamics. Still, NO s properties lend itself to being a good effector signal for PCP, thus warranting further study. Another signaling molecule that could potentially serve as a PCP signal is Ca 2+. Ca 2+ is well studied as a second messenger in numerous pathways. Of note, Ca 2+ is known to play important roles in Wnt signaling and has been suggested to mediate crosstalk between different branches of this pathway, including the Fz/PCP branch (reviewed in Kohn and Moon, 2005). As mitochondria are regulators of Ca 2+ homeostasis in the cell acting as Ca 2+ buffers (reviewed in Berridge et al., 2003), changes in mitochondrial levels of Ca 2+ may have an impact on PCP
84 73 Table 2.5: Summary of PCP phenotypes upon ROS level perturbations ROS Scavenging Conditions PCP Phenotype UAS- Gal4 T (ºC) Cat ey 25 rare PCP defects GED RT none Cat in ft fd clone MARCM 25 no clear rescue of ft clone phenotype Cat+Sod2 ey 29 none hsod1 GED RT none Sod2 GED RT none Sod2 in ft fd clone MARCM 25 no clear rescue of ft clone phenotype hsod1+sod2 ey 29 none GPX GED RT none GPX+Cat GED RT none ey 29 none GPX+Sod2 ey 29 none AOX ey 29 none Ndi ey 29 none AOX+Ndi ey 29 none Vitamin E feeding 0-40 µg/ml none some abnormal PRs Vitamin E fed ft fd clone 0-40 µg/ml no clear rescue of ft clone phenotype some abnormal PRs ROS Inducing Conditions PCP Phenotype UAS- Gal4 T (ºC) MitoKillerRed ey/gmr 25 rare DV inversions with many abnormal PRs/ommatidia CytoKillerRed ey/gmr 25 abnormal PRs/ommatidia Paraquot feeding ft fd /+ haploinsufficiency 0-1 mm none Other mitochondrial perturbations Conditions Phenotype UAS- Gal4 T (ºC) mucp1 ey 25 none mucp3 ey 25 none hucp2 ey 25 none Abbreviations: DV = dorsal-ventral PR = photoreceptor GED = UAS-dicer2/Y hshid; ey-gal4, GMR-Gal4/CyO MARCM = hsflp, w 1118 ; tubgal80, FRT40A; tubgal4/sm6-tm6
85 74 pathways. Therefore it would be interesting to see if Ca 2+ is at all affected in ft or Ndufv2 mutant cells PCP Targets under mitochondrial regulation While the specific mitochondrially-derived PCP signal remains elusive, my data have shed some light on what it may be acting upon. As fj is a PCP gene itself and is the only known target regulated by ft/ds PCP signaling, it was an obvious candidate to test for mitochondrial regulation. Importantly, I observed that the fj-lacz transcriptional reporter does respond to Ndufv2 depletion. Interestingly, it appears to be a repressive interaction, similar to ft s relation to fj; both loss of ft activity (e.g. in ft mutant clones) and depletion of Ndufv2 lead to increased expression of fj, indicating that they normally act to repress fj transcription. Given that Ft and Ndufv2 physically interact and that their PCP phenotypes are similar (though not identical), a reasonable hypothesis would be that they are acting in concert in the same pathway to regulate fj and consequently PCP. My data do not strictly show this to be the case though. Regrettably the classical genetic epistasis experiments to establish ft and Ndufv2 s placement in the same pathway could not be performed due to a lack of null alleles for Ndufv2. Incidentally, strong knockdown of Ndufv2 led to confounding stochastic effects on development. Therefore a null allele may not have been useful in any case. Nonetheless, that a mitochondrial gene is impacting a known PCP target is undoubtedly remarkable. It is, however, unclear whether modification of fj alone would account for all of the PCP phenotypes observed in Ndufv2 knockdowns as their phenotypes are not identical; for example, fj clones show non-autonomous PCP defects, whereas Ndufv2 knockdown clones do not. Certainly, one of the rationales for seeking more ft interactors was to elucidate downstream PCP targets other than fj. Previous studies demonstrated that fj is only partly responsible for ft and ds s PCP phenotypes for several reasons. PCP defects in fj wholly mutant animals are rather mild compared to ft mutants (Zeidler et al., 1999). fj appears to be redundant with ds in some contexts (Simon, 2004). And work from our lab has shown that the non-autonomous propagation of polarity information in and around ft or ds clones is not dependent on fj, suggesting the existence of other PCP targets (Sharma and McNeill, 2013). What, then, could these targets be? My initial experiments to interrogate known PCP pathways has tentatively eliminated two prominent candidates the Fz/Dsh pathway and JAK-STAT signaling (Fig. 2.20A and B). First,
86 75 Fmi s characteristic asymmetric localization during ommatidial precluster development, a hallmark for normal Fz/Dsh PCP signaling, is not appreciably altered in Ndufv2 RNAi clones. Second, depleting Ndufv2 in wing discs has no obvious effect on the JAK-STAT reporter 10xSTAT-GFP. A third PCP establishing event can be found earlier during eye patterning eye field and boundary formation. DV inverted ommatidia can also be caused by anomalous or ectopic equators. Such a mechanism was put forward to explain ft s effect on PCP (Rawls et al., 2002) though our lab has discounted this hypothesis we do not observe changes in equatorial markers (e.g. serlacz or Wg), nor do we see sharp inverse equators being formed in ft clones (Sharma and McNeill, 2013). However, I cannot reject the possibility that Ndufv2 might diverge from ft in how it regulates PCP. In fact, I have observed what could potentially represent inverse equators, as Ndufv2 knockdown induced DV inversions tend to occur in contained patches (Fig. 2.10). Also, the as yet unexplained antennal duplication phenotype (Fig. 2.5) may serve to cause such a disruption to normal eye field establishment. Marker analysis of Ndufv2 depleted eye discs, using Mirror, Fringe or serlacz for example, will be necessary to determine if eye fields are improperly established. Regardless, my finding that mitochondria are regulating PCP in any manner whatsoever has brought novel concepts to consider in the study of both mitochondria and PCP Hpo pathway interactions with mitochondria My data have uncovered not only a connection between mitochondria and PCP signaling, but also an interaction with Hpo pathway targets. I found that several Hpo target genes, ex, crb, bantam, all of which have been widely used in the literature as readouts of Hpo pathway activity, were misregulated in Ndufv2 depleted tissue. Markedly, these effects were internally consistent in that they all indicated Yki activation: activated Yki translocates to the nucleus and promotes the expression of ex, crb, and the micro RNA bantam (which represses the bantamgfp reporter). Thus, my data indicate that Ndufv2 normally acts to repress these genes, presumably by repressing Yki activity through the Hpo pathway. As ft affects these same Hpo targets in such a similar manner, I propose that Ndufv2 is cooperating with ft to signal to the Hpo pathway. Curiously, the region of Ft we identified as being crucial for its binding to Ndufv2 is both highly conserved and has been previously associated with weak PCP and Hpo phenotypes (Matakatsu and Blair, 2012). I have shown that mitochondria affect both PCP and Hpo pathways, thus, a part
87 76 of the ft phenotype could be explained by its association with mitochondria. But why would such an association be advantageous for PCP or Hpo signaling? Unlike in the case of PCP, a connection has previously been made between Hpo and mitochondria. Nagaraj et al. (2012) found that Yki, together with its coactivator Scalloped (Sd), binds to and activates specific enhancer sequences upstream of several mitochondrial genes, including CI components, mitochondrial fusion genes, and antioxidant enzymes. Accordingly, they observed that overexpression of yki leads to more robust and lengthened mitochondrial networks due to the upregulation of mitochondrial fusion genes opa1 and Marf. Furthermore, overgrowth resulting from yki overexpression can be suppressed by reducing or blocking mitochondrial fusion, suggesting that precise modification of the mitochondrial network is a requirement for proliferation downstream of Yki activation. It was a previously unconsidered outcome that a growth control pathway could also act upon mitochondrial dynamics, though upon deeper contemplation perhaps not entirely surprising: regulation of growth is necessarily complex, requiring coordination of a multitude of processes like DNA replication, apoptosis, cell cycle control, etc. The mitochondrion is also crucial to many of these, and so is a sensible target for growth control regulation. My data reflect a similar though inverse connection mitochondria are somehow regulating the Hpo pathway. It is entirely possible that this apparent activation of Yki targets is secondary to the many impacts of perturbing mitochondria; however, in light of what has been discussed above, I suspect that this effect reflects a kind of feedback loop. The Hpo pathway directs mitochondrial dynamics, and in turn the mitochondrial network can communicate its current condition and status back to the Hpo pathway to further refine this regulation. Additionally, mitochondrial stress may also be a reason for this feedback signal. One consequence of mitochondrial fusion is the relief of stress through complementation of damaged mitochondrial genomes, or the proposed transmission of membrane potential through mitochondrial networks to allow energy production in oxygen-poor regions of the cell (Hales, 2010; Skulachev, 2001). Perhaps stressed mitochondria are promoting mitochondrial fusion through the Hpo pathway in this feedback system. It is unclear what might be mediating this effect; I have not determined whether it is the same signal working in mitochondrial-pcp signaling or something else entirely. More investigation will be required to characterize these signaling events.
88 77 While I have shown that mitochondria are certainly having an effect on the Hpo pathway, I have also noted that the phenotype observed in Ndufv2-depleted tissues is not entirely consistent with loss of Hpo signaling: I do not observe Hpo-like overgrowth, and also I see an increase rather than a suppression of apoptosis (Fig. 2.19). A simple explanation would be that perturbing mitochondrial components triggers the release of cytochrome C leading to apoptosis. In this way, Ndufv2-depleted cells promote both the apoptotic pathway and Yki-activated proliferation, resulting in the increase in proliferation and increased cleaved-caspase3 that I observe. This may also explain the lack of overgrowth, as increased apoptosis could balance overproliferation. It would be interesting to see if blocking apoptosis in Ndufv2-depleted tissue, via expression of p35 for example, would uncover a Hpo-like overgrowth phenotype. Alternately, if Ndufv2 s effect on Hpo is genetically separable from its other functions, experiments with a specifically Hpodeficient isoform of Ndufv2 would reveal whether there are underlying characteristic Hpo-like defects in the Ndufv2 phenotype Cryptic genetic interactions: ft and Ndufv2 As alleles for Ndufv2 were unavailable and I could not easily produce one (the extant Ndufv2 proximal P-element insertion was refractory to imprecise excision), I was unable to use classical genetic analyses to study the interaction between ft and Ndufv2. Instead, I relied upon RNAi mediated knockdown of Ndufv2 in various genetic backgrounds to interrogate these interactions. Regrettably, despite my attempts to minimize it, the variability inherent to RNAi proved to be problematic in interpreting the results of these experiments: in terms of eye PCP, variance was an order of magnitude larger than the difference between experimental and control conditions. Still, examination of wing phenotypes did uncover an interesting potential interaction between ft and Ndufv2. Crossvein distance (CVD) is widely used in the literature as a readout of ft pathway disruption ft, ds, fj, approximated (app), and dachs (d) viable mutants all show a shortening in CVD (Mao et al., 2009). I observed that Ndufv2 knockdown in the wing also produced this phenotype, supporting its assignment to the ft pathway. I had expected that further dampening of ft signaling by removing a copy of ft in Ndufv2 knockdown wings would result in further shoretning of the CVD. Contrarily, CVD was increased in the ft/+ background. Typically, when a mutation ameliorates the phenotype of another mutant, there is an inhibitory relationship between the two genes, such that the loss of one compensates for the loss of the other. For
89 78 example, if Ndufv2 were an inhibitor of ft, loss of Ndufv2 would result in increased ft levels or activity, producing the observed phenotype. If ft dosage were then reduced in the Ndufv2 mutant, then ft levels or activity would be restored nearer to wt, resulting in rescue. However, genes that interact in this manner normally have opposite phenotypes in the single mutants; ft and Ndufv2 appear to have the same phenotype not only in CVD, but also in regulation of target genes (Ex, Crb, Bantam, Fj). Thus it is unlikely that ft and Ndufv2 are interacting in an inhibitory manner. What, then might be causing the rescue I observed? One possibility is that CVD is sensitive to the relative gene dosage of ft and Ndufv2. For instance, in both single gene disruptions, their levels relative to each other are disturbed, producing a shorter CVD. In the Ndufv2 knockdown ft-heterozygous wings, the ratio of ft to Ndufv2 is closer to the wt case, resulting in the partial rescue. Notably, mathematical models have predicted that the relative levels of Ft and Ds are causative for some of their growth phenotypes (Mani et al., 2013). Perhaps the same is true between ft and Ndufv2. Further support for the significance of ft and Ndufv2 gene balance was observed in an attempted rescue experiment. With the hypothesis that loss of ft would lead to loss of Ndufv2, I overexpressed Ndufv2 in ft clones to test if this could restore ft signaling in the clone and perhaps rescue PCP phenotypes (Fig. 2.17). Remarkably, I observed a synthetic lethality phenotype causing severe tissue degeneration. This is a novel phenotype for ft and it is still unknown what the nature of this degeneration is. Nonetheless, in this experiment Ndufv2 expression was increased, leading to a greater imbalance between the two genes than in the case of a ft clone alone, and I saw an enhanced phenotype. This is consistent with the notion that tissues are sensitive to the relative gene dosage of ft and Ndufv2. Additional investigation into these cryptic interactions between ft and Ndufv2 will surely be necessary to uncover how they are working with each other to regulate growth, PCP, and possibly cell survival.
90 79 Chapter 3 Ft and mitochondrial function These data, figures (with the exception of Fig. 3.2), results, and discussions were published in Sing et al. (2014). Cell 158, Data attribution: Drosophila testes analysis was carried out by Lacramioara Fabian. Complex I activity and lactate assays were performed in collaboration with Riya Shabhag. Pulse-chase Ft expression experiment was performed by Lesley Clayton. MTS constructs were made and tested in COS cells by Maïlis Bietenhader. D11 cell staining was performed in collaboration with Robyn Rosenfeld. Biochemical and metabolic analysis of S2, D11 and larval tissues were performed in collaboration with Yonit Tsatskis.
91 80 Ft and mitochondrial function 3.1 Introduction Ft and Ds have been well-studied in their roles as cell surface cadherins and upstream members of Hpo and PCP signaling; though how signals are transduced from these membrane embedded molecules to intracellular events is not fully understood. In the previous chapter, I described mitochondrial genes that interact with ft, and that appear to regulate both Hpo and PCP branches of ft signaling. As this was a novel and unexpected connection between ft and mitochondria, I aimed to investigate more thoroughly how they are working together. Mitochondria are wellknown as the metabolic heart of the cell a vast array of processes occur within mitochondria, not the least of which is oxidative phosphorylation (OXPHOS) which results in the production of energy stores for the cell in the form of ATP. The ft interacting mitochondrial genes that I found were all a part of or related to the OXPHOS system; therefore, I speculated that ft might be affecting mitochondrial function, especially OXPHOS activities. Another unresolved matter is how Ft at the cell surface could be physically interacting with mitochondria-resident proteins (Ndufv2, MPP, and CG1746) as implied by both our Y2H and GST-pulldown experiments. Ndufv2 and CG1746 are part of CI and CV respectively, with Ndufv2 protruding into the matrix and CG1746 embedded in the inner mitochondrial membrane. Orthologs of CG3731/MPP exist as either a soluble heterodimer in the matrix, or may possibly be incorporated into CIII as core protein 1. It is unclear how CG3731 is situated in Drosophila, but, either case would place it within mitochondria, far-removed from the cell membrane where Ft is thought to reside. In this chapter I survey ft mutants for mitochondrial phenotypes and find that mitochondrial ultrastructure is abnormal, accompanied by defects in OXPHOS activity. I observe that ft cells favour glycolysis over OXPHOS, perhaps as a form of metabolic compensation. I find that ROS and the ROS-responsive JNK pathway are upregulated in ft tissue. Further, I establish a mechanism whereby cell surface Ft is processed to release a cytosolic fragment that is capable of translocating into mitochondria where it interacts with CI to promote its assembly/stability and function. Importantly, I determine that a highly conserved Ndufv2-binding domain found in Ft is
92 81 necessary for these mitochondrial functions. In sum, these data highlight a completely novel mode of ft activity in cellular metabolism and signaling. 3.2 Materials and methods Fly stocks and genetics All wt flies are yw unless otherwise noted. Gal4 drivers used were: engal4, bamgal4. GSTD1- GFP was provided by Dirk Bohmann. ft and ampkα 3 clones were generated using FLP-FRTmediated recombination (Xu and Rubin, 1993) with hsflp; ft fd FRT40A/Ubi-GFP FRT40A, or ampkα 3, FRT101/ubiGFP, FRT101; hsflp/+. Complex I assembly mutant (dcia30 ex80 /TM6) was provided by David Walker and described previously (Cho et al., 2012). ft fd mutants were rescued with ft fd /CyO, GFP; P[acman]- ft /TM6B (expressing full-length Ft - FL, or versions lacking region 2 - FL 2 or region 8 - FL 8) flies obtained from Kenneth Irvine as previously described (Pan et al., 2013). P[A92]pucE69/TM6B,Se was obtained from Utpal Bannerjee and is available from DGRC-Kyoto (stock #109029) Antibodies The following antibodies were used: mouse M2 anti-flag (Sigma), mouse anti-cvα (MitoSciences), mouse anti-porin (MitoSciences), mouse anti-pdh (MitoSciences), mouse anti- Ndufs3 (MitoSciences), mouse anti-lamin (DSHB), anti- -tubulin (DSHB), mouse anti-actin (Millipore, clone C4), rabbit anti-cytochrome C (Boster Immunoleader), rabbit anti-calnexin (abcam), rabbit anti-pex5 (Sigma SAB ), mouse anti-mmp1 (1:1:1 cocktail of 3A6B4, 3B8D12, 5H7B11, DSHB), anti-ndufv2 (raised in rat against full-length GST-tagged Ndufv2), anti-fat (raised in rat against a GST-tagged intracellular fragment of Fat consisting of residues ), horseradish peroxidase-conjugated anti-rat and mouse IgG secondary antibodies (Jackson Labs), and Alexa Fluor or Cy3-conjugated anti-rat and mouse IgG secondary antibodies (Invitrogen or Jackson Labs).
93 Cell culture and transfection S2 and D11 cells were cultured in M3 media supplemented with 10% FBS. D11 culture media was supplemented with 10 µg/ml of insulin. S2 cells were transfected using FuGENE 6 (Roche) or Effectene Transfection Kit (Qiagen) Whole larvae imaging Larvae were reared from overnight (~16 hr) egg lays on grape juice plates and maintained at 25ºC until appropriate time points. Larvae were then selected (either against GFP or against tubby marker) and washed in PBS, then briefly (~15 sec) boiled in water before dry mounting on glass slides for image capture on a Nikon AZ100 microscope Drosophila testis preps Adult and larval testes were dissected in TIB (testes isolation buffer) (Casal et al., 1990) with Hoechst (1:2000 or 8.3 µg/ml) (Sigma, St. Louis, MO) and live squashed preparations of Drosophila male germ cells were made as previously described (Regan and Fuller, 1990). Preparations were examined on an epifluorescence microscope (Zeiss Axioplan 2, Axiocam black and white camera, Axiovision software from Carl Zeiss). Germline clones were made using the hsflp-frt system. Two day-old larvae were heatshocked for 1h at 37 o C (in an incubator) or for 2h at 37 o C (in a water bath), for 2-3 consecutive days. For RNAi experiments, VDRC line #22194 line was crossed with bamgal4, as described (Chen and McKearin, 2003) Electron microscopy Adult and 3 rd instar larval testes (wt and ft mutants) were dissected in 0.1M phosphate buffer (PB) (ph=7.4) on ice and immediately subjected to high-pressure freezing (Leica HPM100). Samples were then subjected to freeze substitution (Leica AFS2) in 1% osmium tetroxide and 0.1% uranyl acetate in HPLC grade acetone. Samples were washed several times in acetone and embedded in a Quetol-Spurr resin mixture. Alternatively, testes were fixed immediately after dissection in a freshly prepared solution of 2.5% glutaraldehyde + 2% paraformaldehyde in PB. Samples were further prepared following a previously described protocol (Fabian et al., 2010) and embedded in Quetol-Spurr resin. 100nm sections were cut on an RMC MT6000 ultramicrotome, stained with 2% aqueous uranyl acetate and 0.2% lead citrate and viewed in a JEOL JTE transmission electron microscope (JEOL, Peabody, MA; The Hospital for
94 83 Sick Children Electron Microscopy Facility). Images were captured with AMTv542 software (Advanced Microscopy Techniques, Danvers, MA) ROS levels measurements 3x10 6 cells were collected and incubated with or without 5 µm Antimycin A at 25 C for 2.5 hours. 5 µm MitoSOX Red was added, and incubated at 25 C for 30 minutes then washed with media. Fluorescence was read at 510 nm excitation, 580 nm emission. ROS levels were monitored in vivo using the superoxide indicator dihydroethidium (DHE) as described by Owusu-Ansah et al. (2008a). Imaginal discs were dissected in S2 media and stained in 30 µm DHE for 5 min then fixed in 4% PFA for 5 min Complex I Activity Assay Mitochondria purified from 5 female larvae were resuspended in 100 µl of mitochondrial isolation medium (MIM, 250 mm sucrose, 10 mm Tris (ph 7.4), 0.15 mm MgCl2), and 5 µl were used in a colorimetric complex I activity assay as previously described (Cho et al., 2012). NADH: ubiquinone oxidoreductase activity was monitored as a decrease in absorbance at 600 nm, for 1 hour, with no inhibitor and in the presence of rotenone (2 um). The measured activities were normalized to citrate synthase activity ATP assay Lysates for ATP assays were prepared from whole larvae as previously described (Monserrate et al., 2012). Lysates were diluted 1:100 and ATP concentration was determined using the CellTiter-Glo kit (Promega product#g7571) as per manufacturer s protocol Lactate assay 30 wt larvae and an equivalent volume of ft fd larvae were homogenized in 100 µl of PBS with 30 strokes of a Kontes pestle. For protein determination, 5 µl were taken for a BCA assay (BCA Protein Assay Kit, Pierce, 23227). The remaining samples were heat shocked at 60 C for 15 minutes and centrifuged at g for 5 minutes; 5-10 µl of the supernatant were used for lactate determination using a Lactate Assay Kit (Biovision, K , Milpitas, CA, USA). All lactate readings were normalized to protein content.
95 Cell fractionation and mitochondrial protection assay Mitochondria from D11 cells or eye/brain complexes from wt, ds UAO71 and ft alb /ft fd larvae were isolated using the Qiagen Qproteome mitochondria isolation kit (#37612). Aliquots of mitochondria were treated with 5, 6 or 10 g/ml proteinase K at 4 C for 10 min. If needed, 1% Triton X-100 was included during the proteinase K digestion. After digestion, proteinase K was quenched by addition of 1X Complete Protease Inhibitor (Roche Applied Science) followed by incubation at 4 C for 10 min Mitochondrial isolation and blue-native polyacrylamide gel electrophoresis (BN-PAGE) Mitochondria for BN-PAGE were purified from larvae by differential centrifugation using mitochondrial isolation medium (MIM, 250 mm sucrose, 10 mm Tris (ph 7.4), 0.15 mm MgCl2, with protease inhibitor). Mitochondria were resuspended in 1x Native PAGE Sample buffer (Invitrogen) with 1% digitonin and protease inhibitors, and incubated for 15 min on ice. Samples were centrifuged at x g for 30min at 4 C and supernatant was resuspended with G250 sample additive and Native PAGE Sample Buffer (Invitrogen). Mitochondria were then visualized with the Novex NativePAGE Bis Tris Gel system (Invitrogen) using 3-12% Bis-Tris Native PAGE gels as previously described (Cho et al., 2012). For further immunodetection, samples were transferred to PVDF membranes with transfer buffer containing 20% methanol and no SDS. Membranes were blocked in 5% skim milk powder in Tris-buffered saline for 1hr and incubated with primary and secondary antibodies. Chemiluminescence detection was performed on a Versadoc imager (Bio-Rad) Mitochondrial in-gel activity assays Mitochondria from Drosophila larvae were isolated as described (Walker et al., 2006), resuspended in lysis buffer (50 mm Bis-Tris, ph 7, 750 mm 6-aminocaproic acid, 2 % (w/v) digitonin, protease inhibitor cocktail), cleared by centrifugation at x g at 4 C and supplemented with glycerol to a final concentration of 4 %. 40 μg of mitochondrial protein was resolved on a 4-13 % native gel using a cathode buffer containing 0.05 % (w/v) deoxycholate and 0.01 % (w/v) n-dodecyl β-d-maltoside. In-gel activity assays were performed as described (Wittig et al., 2007). Briefly, complex I activity was measured in a NADH:NTB reductase assay. Gel strips from clear-native gels above were incubated in assay buffer (2.5 mg/ml NADH, 0.1
96 85 mg/ml NTB in 5mM Tris/HCl, ph 7.4) for 5 min, then fixed in 50% methanol, 10% acetic acid. Similarly, assays for complex II and IV activity were carried out in complex II assay buffer (200 µm sodium succinate, 200 µm phenazine methosulfate, 0.1 mg/ml NTB in 5mM Tris/HCl, ph 7.4) or complex IV assay buffer (0.5 mg/ml diaminobenzidine, 50 µm horse heart cytochrome c in 50 mm sodium phosphate, ph 7.2), incubated for min then fixed. Complex V activity was monitored by following ATP hydrolysis activity. Gel strips were pre-incubated in 35 mm Tris, 270 mm glycine, ph 8.3, then transferred to assay buffer (35 mm Tris, 270 mm glycine, 14 mm MgSO4, 0.2% Pb(NO3)2, 8 mm ATP, ph 8.3). Gel strips were then fixed in 50% methanol for 30 min to stop the reaction. 3.3 Results Growth of ft larvae is abnormal While previous studies of ft mutants have focused on the Hippo-mediated overgrowth of tissues one phenotype exhibited by these larvae, though noted (Bryant et al., 1988; Mahoney et al., 1991), has largely been ignored. That is, ft null mutant larvae are significantly smaller than their age-matched wildtype counterparts (Fig. 3.1B). It is only after an extended late larval stage that these overgrown imaginal discs are observed. Indeed, ft larvae do not pupariate until up to 12 days AEL (Fig. 3.1C). We first wondered whether this initial deficit in growth and extended larval stage was a delay in development. By examining mouth hooks of these larvae and comparing to age-matched controls, we found that ft larvae do reach each larval stage (1 st, 2 nd and 3 rd instar) at appropriate time points (Fig. 3.2). Thus, the overgrowth of ft larvae occurs during an extended 3 rd instar stage. Interestingly, we found that larvae harbouring a mutation in Complex I (CI) Assembly Factor (dcia) also displayed a deficit in growth similar to ft larvae (Fig. 3.1B). They are also significantly smaller than age-matched wildtype controls, and experience an extended larval stage. In contrast, they do not overgrow. Considering our discovery that Ft interacts with the CI component Ndufv2, these data suggest that growth deficits in ft larvae may be due to defects in CI.
 Whole mount 3 rd instar larvae reveal a growth delay in ft fd and dcia compared to wt.")
97 Figure 3.1: ft mutants experience a delay in growth and pupariation (A) Whole mount 3 rd instar larvae reveal a growth delay in ft fd and dcia compared to wt. (B) Pupariation counts of 50 1 st instar larvae reveals a pupariation delay of approximately 7 days in ft fd compared to wt. AEL = after egg lay. 86
 show normal progression through larval moults of ft fd")
98 Figure 3.2: ft larvae are not developmentally delayed as assessed by mouth hook morphology. Light micrographs of larval mouth hooks at the indicated time points (hours after egg lay ±2 hrs) show normal progression through larval moults of ft fd larvae. All micrographs are taken at the same magnification. 87
99 ft mutants show mitochondrial phenotypes ft mutants show defects in mitochondrial morphology Since the cytoplasmic domain of Ft binds mitochondrial proteins, we wondered if overall mitochondrial structure is affected in ft mutants. We examined mitochondria in developing Drosophila sperm as they undergo dramatic remodeling during spermatid differentiation, allowing us to closely examine mitochondrial morphogenesis and ultrastructure. All the mitochondria in each early round spermatid aggregate and fuse to form a spherical mitochondrial derivative called the Nebenkern. During elongation, the Nebenkern splits and extends along the developing flagellum (Fig. 3.3Ai-iv). Analysis of germline clones of ft mutants revealed defects in Nebenkern morphology. In early round spermatids, Nebenkerne are lobate and broken, with vacuolated areas (Fig. 3.3Av). In elongating spermatids, Nebenkerne are aberrant, with areas of uneven condensation (Figure 3Avi). Similar mitochondrial defects were seen in ft larval testes (Fig. 3.3Av, vi insets). Thus, ft is necessary for normal mitochondrial morphogenesis during sperm development. Expression of Ndufv2 dsrna resulted in similar Nebenkern defects (Fig. 3.3Avii, viii), highlighting requirement for both ft and Ndufv2 in mitochondrial morphogenesis. We then used transmission electron microscopy to analyze mitochondrial ultrastructure of ft mutants in maturing sperm and surrounding cyst cells (Fig. 3.3B). Defects in mitochondrial morphogenesis and ultrastructure were observed in ft mutant spermatids (Fig. 3.3Biii). Mitochondria in wildtype somatic cyst cells (which surround developing sperm) are elongated and exhibited regularly spaced, flattened cristae (Fig. 3.3Bii). In contrast, mitochondria in ft mutant cyst cells were round and with swollen and irregular cristae (Fig. 3.3Biv and C). Similar defects in cristae morphology were seen in eye and wing discs obtained from ft fd and ft alb larvae (Fig. 3.4), indicating this is not a tissue- or allele-specific effect. These dramatic alterations in mitochondrial architecture in ft mutants support a role for Ft in mitochondria, and raised the possibility that loss of Ft affects mitochondrial function.
 Spermatids in onion stage with fused mitochondria that form the Nebenkern. (Aii) Elongated spermatid cysts.")
100 Figure 3.3: Loss of ft or Ndufv2 results in defective mitochondria. (Ai-ii) Schematics of spermatid differentiation. n=nucleus. (Ai) Spermatids in onion stage with fused mitochondria that form the Nebenkern. (Aii) Elongated spermatid cysts. (Aiiiviii) Overlays of phase-contrast and epifluorescence micrographs of live preparations stained with Hoechst to reveal DNA (blue) show defects in Nebenkern morphology in ft fd mutant clones (GFP-negative) in early spermatids (Av) and elongated spermatid cysts; aberrant clumping occurs along the Nebenkern (Avi). Insets show spermatids from ft fd larval testes. (Avii, viii) Early and elongating spermatids expressing Ndufv2 dsrna show defects in Nebenkern morphology. Inset: high magnification detail of Nebenkern (arrows) defect. Arrowheads indicate nuclei. Scale bars: 20 µm. (B) Transmission electron micrographs of early elongated spermatids and cyst cell mitochondria. (Bi) Wildtype spermatids show axonemes (arrowhead) and Nebenkerne (arrows) of comparable size. (Biii) In ft fd mutants the two halves of each Nebenkern show large variation in size (arrow). Scale bar 500 nm. (Bii) Wildtype cyst cell mitochondria are elongated and show regularly spaced, flattened cristae. (Biv) In ft mutants, mitochondria (arrows) appear round or oval and have swollen cristae Insets show mitochondria from wildtype and ft mutant cyst cells at high magnification. Scale bar 500 nm. Percentage of defective mitochondria is quantified in (C) (*p= , Student s t-test. wt, n=39. ft fd, n=34). 89

101 Figure 3.4: ft affects mitochondrial morphology in imaginal discs. Transmission electron micrographs of wildtype mitochondria show regularly spaced, flattened cristae. In ft alb and ft fd mutants, mitochondria have swollen and irregular cristae. Scale bars, 200 nm. 90
102 ROS are upregulated in ft mutant tissue and Ndufv2-depleted tissue ROS are a byproduct of OXPHOS and are primarily produced in CI and CIII in mitochondria. As Ft interacts with Ndufv2, a component of CI, I wondered if ROS levels are perturbed in ft and Ndufv2 mutants. Structural analysis of CI suggested the Ndufv2 Fe-S cluster may have a role in ROS regulation (Sazanov and Hinchliffe, 2006), and my rescue analysis indicates a critical role for the Fe-S cluster in PCP. We measured ROS levels by staining with dihydroethidium (DHE) which becomes more fluorescent at higher superoxide levels. We first examined the effect of reduction of Ndufv2 on ROS levels using dsrna in Drosophila S2 cells, and found that loss of Ndufv2 led to increased levels of ROS in tissue culture cells (Fig. 3.5A, B). Reduction of Ndufv2 in the posterior compartment of the wing by expressing dsrna driven by engal4, also led to dramatically increased ROS levels in vivo (Fig. 3.5C). Of note, different RNAi lines resulted in varying degrees of ROS generation, perhaps reflecting differential efficiency of knockdown between the three lines tested. CIII is also a site of mitochondrial ROS generation, so I tested if depleting CIII components impacted ROS levels as well. I drove dsrna against CIII components Core 2 (CG4169) and Heme protein (CG4769) and found marked upregulation of ROS in both cases (Fig. 3.5D) To test if loss of ft also affected ROS levels, I generated mitotic clones of ft mutant tissue in eye discs. I found that ft mutant clones have increased DHE staining, indicating loss of ft leads to increased ROS levels (Fig. 3.6A). To confirm that ROS are upregulated in ft tissue I used GSTD1-GFP, a reporter of oxidative stress (Sykiotis and Bohmann, 2008). Loss of ft led to clear increases in GSTD1-GFP expression (Fig. 3.6B), indicating that ft mutant tissue undergoes oxidative stress, consistent with increased ROS levels. In the course of these experiments, I observed an intriguing pattern of ROS in the eye disc. DHE staining in wildtype eye discs revealed a gradient of ROS stronger at the equator and weaker towards the poles (Fig. 3.5E). This pattern is highly reminiscent of the fj expression gradient which is partly responsible for PCP signaling. This suggests that ROS could also provide PCP information via its graded pattern.
 S2 cells treated with dsrna against Ndufv2 produce more ROS as detected by DHE fluorescence.")
103 Figure 3.5: Depletion of mitochondrial complex I and III components increases ROS levels. (A) Lysates of S2 cells left untreated or treated with dsrna against Ndufv2 were probed using Ndufv2 antibody. Ndufv2 protein levels are appreciably reduced upon dsrna treatment. (B) S2 cells treated with dsrna against Ndufv2 produce more ROS as detected by DHE fluorescence. Error bars represent standard error. (C) ROS are appreciably increased in the posterior compartment of 3rd instar larval wing discs expressing Ndufv2 dsrna driven with engal4 as compared to control (en>gfp). The effect is stronger in the VDRC RNAi lines than in the NIG RNAi line. ROS detected by DHE. Posterior right. (D) engal4 driven knockdown of CIII components CG4169 and CG4769, also reveals strong upregulation of ROS in the posterior of the wing. (E) 3 rd instar eye disc shows a graded pattern of fj expression (left). DHE stain also reveals a similar gradient of ROS (right). 92
 ROS are upregulated in ft fd mitotic clones (marked by lack of GFP (A) or β-gal (B) staining) as detected by DHE (A) and GSTD1-GFP (B).")
104 Figure 3.6: ft regulates ROS independently of the Hpo pathway. (A-B) ROS are upregulated in ft fd mitotic clones (marked by lack of GFP (A) or β-gal (B) staining) as detected by DHE (A) and GSTD1-GFP (B). (C-D) Overexpression of Yki S168A -GFP (a phospho-deficient and thus activated form of Yki) driven with engal4 in posterior wing compartments (C) or wts e1 clones in eye discs (D clone marked by lack of GFP) do not show changes in ROS as detected by DHE. 93
105 94 I then wondered how ft might be regulating ROS. I speculated that the increase in ROS in ft tissue could be due to ft s influence on the Hpo pathway, as Nagaraj et al. (2012) had observed that activation of YAP, the mammalian Yki homolog, leads to changes in ROS levels in human breast cancer cell lines. Overexpression of yki, or loss of the core Hippo kinase wts, in Drosophila wing discs does not lead to increased ROS, indicating that the increased ROS in ft clones is not due to loss of Hpo regulation by ft (Fig. 3.6C, D) Loss of Ft or Ndufv2 leads to activation of the ROS target JNK ROS activate the JNK pathway (Hamanaka and Chandel, 2010; Owusu-Ansah and Banerjee, 2009; Owusu-Ansah et al., 2008b), which has been suggested to be a regulator of PCP (reviewed in McNeill, 2010). To test if JNK signaling is affected in ft mutants, I examined the downstream JNK target puckered (puc) by using the puc-lacz reporter. Expression of Ndufv2 dsrna using engal4 led to strong activation of puc-lacz (Fig. 3.7A), consistent with increased ROS leading to activation of JNK signaling. Strikingly, ft clones also display strong increases in puc-lacz expression (Fig. 3.7B), indicating loss of ft leads to activation of JNK signaling. This activation is more pronounced in clones anterior to the MF. Additionally, higher levels of the JNK target MMP1 are also revealed by antibody staining of ft clones in wing discs (Fig. 3.7C). Thus, the ROS target JNK is upregulated in Ndufv2 and ft depleted tissue. Curiously, JNK activation as assayed by both puc-lacz and MMP1 staining appeared to be stronger in mutant tissue that is immediately adjacent to wt tissue at the borders of ft clones, suggesting that sharp differences in ft or ROS levels signal more robustly to the JNK pathway Loss of Ft leads to reduced CI activity and alterations in cell metabolism I had observed that loss of ft or knockdown of Ndufv2 led to increased ROS. As Complex I is a major site of ROS generation in the cell, I wondered if ft might regulate CI activity. We tested if CI activity was affected in ft mutants by measuring the reduction of 2,6-dichloroindophenol (DCIP) in a spectrophotometric assay (Abs600nm) on larval mitochondrial extracts. Significantly, DCIP turnover was much slower in ft mutant extracts suggesting that loss of ft leads to a dramatic loss of CI activity (Fig. 3.8A).
 The JNK pathway activity reporter puc-lacz is upregulated in Ndufv2-depleted tissue (A Ndufv2 dsrna driven with engal4) as well as in ft fd clones in eye imaginal discs.")
106 Figure 3.7: The JNK pathway is activated in Ndufv2-depleted and ft mutant tissue. (A-B) The JNK pathway activity reporter puc-lacz is upregulated in Ndufv2-depleted tissue (A Ndufv2 dsrna driven with engal4) as well as in ft fd clones in eye imaginal discs. (C) Antibody staining of ft clones in 3 rd instar wing discs reveals elevated levels of MMP1, a downstream target of JNK signalling. Clones are GFP negative in (B) and (C). 95
107 Figure 3.8: ft mutants have reduced CI activity and upregulated glycolysis resulting in wildtype levels of ATP. (A) Complex I (CI) activity is severely reduced in ft mutant larvae (error bars represent standard deviation. *p<0.003, Student s t-test, n=3). (B) ATP levels in whole larval extracts from ft fd mutants show no reproducible difference from wildtype control (error bars represent standard deviation, n=6 samples of >10 larvae each). (C) Lactate levels are increased in extracts from both age-matched and size-matched ft fd mutant larvae compared to controls (error bars represent standard deviation. *p<0.05 **p<0.002, Student s t-test, n=3). 96
108 97 Considering CI is a major entry point into the electron transport chain, and thus its normal functioning is essential for efficient energy production, I asked if ft mutants were deficient in cellular energy stores. ATP levels can be monitored through chemiluminescence produced by luciferase, as the luciferase reaction requires ATP. I measured ATP levels in whole larval extracts from ft mutants and found that ATP levels are not appreciably altered (Fig. 3.8B). The concurrence of low CI activity with normal levels of ATP led us to hypothesize that there was an alteration in metabolism in ft mutants these cells might be compensating for reduced OXPHOS activity by increasing rates of glycolysis. Therefore, we measured levels of lactate, a readout of glycolytic activity. First, we compared size-matched ft and wt larvae and found a significant increase in lactate levels (Fig. 3.8C). We reflected that ft larvae might not be at the same developmental stage as wt larvae of the same size, as size-matching may select for older ft larvae due to the delayed growth of ft mutants. Therefore we also compared age-matched larvae. Strikingly, ft mutants have significantly increased levels of lactate in age-matched larvae as well, reflecting increased glycolysis (Fig. 3.8C) Ft is cleaved to release a cytosolic fragment that is targeted to mitochondria Ft functions at the cell surface in cadherin-mediated adhesion (Ma et al., 2003), whereas Ndufv2 is a core component of mitochondrial CI. We had found that Ft binds directly to Ndufv2 in vitro; however in vivo, Ft and Ndufv2 could potentially interact either at the cell surface or in mitochondria. I had previously generated antibodies to Ndufv2 and confirmed their specificity (see Fig. 2.11A). Co-staining for the mitochondrial protein CVα demonstrated that Ndufv2 localized to mitochondria in S2 cells (Fig. 3.9) and in Drosophila discs (see Fig. 2.11B). Ndufv2 was not detected at the cell surface in cultured cells nor in Drosophila imaginal discs. I hypothesized that Ft might interact with Ndufv2 inside mitochondria. Ft is post-translationally processed to generate a 450kDa extracellular fragment and a 110kDa transmembrane fragment (Feng and Irvine, 2009; Sopko et al., 2009). In addition, we detected a more labile ~68 kda fragment of ft in a pulse-chase experiment (Fig. 3.10A), suggesting a second cleavage step that releases a soluble cytoplasmic fragment. I wondered if this fragment could translocate to the mitochondria. In silico analysis of the cytoplasmic domain of Ft with MitoProt II detected two

109 Figure 3.9: Ndufv2 colocalizes with CVα in Drosophila S2 cells. Immunofluorescence images of an S2 cell stained with α-ndufv2 and mitochondrial marker α-cvα reveals mitochondrial localization of Ndufv2. 98
 Heatshock induced expression of a full-length HA-tagged ft transgene in Drosophila embryos reveals multiple forms of Ft: α-ha (left) or α-ft (right) antibodies detect 560, 110 and")
110 Figure 3.10: Ft is processed in vivo to release an intracellular 68 kda fragment. (A) Heatshock induced expression of a full-length HA-tagged ft transgene in Drosophila embryos reveals multiple forms of Ft: α-ha (left) or α-ft (right) antibodies detect 560, 110 and ~68kDa bands. α-tubulin (lower left) serves as a loading control. (B) Schematic of Ft cleavage products. 99
111 100 mitochondrial targeting sequences (MTS) and their corresponding MPP cleavage sites (Fig. 3.11). To test if these putative MTS can direct mitochondrial localization, we generated constructs expressing Ft-MTS fragments tagged with a FLAG epitope (Fig. 3.12A). Immunofluorescence of COS cells demonstrated that fragments of Ft containing the predicted MTS sequences coupled to FLAG localize to mitochondria (Fig. 3.12B). Our studies also indicate there is an additional MTS signal, located after predicted MTS1 and MTS2 that is not identified by in silico analysis. This is not surprising because MitoProt II does not recognize all mitochondrial-targeting motifs. Together these in silico and in vitro data indicate that the cytoplasmic domain of Ft contains multiple regions that could direct mitochondrial localization Exogenous FtΔECD is detected in mitochondria from cultured cells To determine if Ft can be detected in mitochondria, we first used cultured cell overexpression experiments. Since a 110kDa transmembrane form of Ft (FtΔECD) can rescue most defects of ft mutants (Matakatsu and Blair, 2006), we transfected Drosophila S2 cells with FtΔECD and immunostained with antibodies to the FLAG epitope and Ndufv2. A portion of Flag-tagged Ft co-localized with Ndufv2 in S2 cells, indicating mitochondrial localization (Fig. 3.13A). Interestingly, expression of the entire cytosolic domain of Ft (Fat-IC ~74kDa) was not sufficient to confer mitochondrial localization (Fig. 3.13B), suggesting that processing at the plasma membrane facilitates production of a Ft fragment that is competent to enter mitochondria. Consistent with this, soluble FtIC cannot rescue ft mutants, while a membrane tethered FtIC can (Bossuyt et al., 2014; Matakatsu and Blair, 2012; Pan et al., 2013; Zhao et al., 2013). We confirmed this result biochemically by mitochondrial fractionation of transfected S2 cells the processed 68kDa fragment derived from FtΔECD is present in mitochondrial fractions (Fig. 3.13A), whereas the FtIC is only detected in total extracts (Fig. 3.13B) Endogenous Ft is detected in mitochondria from cultured cells To determine if endogenous Ft localizes to mitochondria, we examined a Drosophila cell line, D11, that expresses full-length Ft. Staining of D11 cells and Drosophila tissue showed colocalization of Ft staining with the mitochondrial marker CVα (Fig. 3.14A). In addition, cell

112 101 A Fat RFRGKQEKIGSLSCGVPGFKIKHPGGPVTQSQVDHVLVRNLHPSEAPSPPVGAGDHMRPPVGSHHLVGPELLTKKFK EPTAEMPQPQQQQQRPQRPDIIERESPLIREDHHLPIPPLHPLPLEHASSVDMGSEYPEHYDLENASSIAPSDIDIV YHYKGYREAAGLRKYKASVPPVSAYTHHKHQNSGSQQQQQQHRHTAPFVTRNQGGQPPPPPTSASRTHQSTPLARL* SPSSELSSQQPRILTLHDISGKPLQSALLATTSSSGGVGKDVHSNSERSLNSPVMSQLSGQSSSASRQ*KPGVPQQQ AQQTSMGLTAEEIERLNGRPRTCSLISTLDAVSSSSEAPRVSSSALHMSLGGDVDAHSSTSTDESGNDSFTCSEIEY DNNSLSGDGKYSTSKSLLDGRSPVSRALSGGETSRNPPTTVVKTPPIPPHAYDGFESSFRGSLSTLVASDDDIANHL SGIYRKANGAASPSATTLGWEYLLNWGPSYENLMGVFKDIAELPDTNGPSQQQQQQTQVVSTLRMPSSNGPAAPEEY V B MTS1 MTS1: 89% C MTS2 MTS2: 87% Figure 3.11: Predicted mitochondrial targeting sequences in the cytosolic domain of Ft. (A) The peptide sequence of the intracellular fragment of D. melanogaster Ft contains predicted mitochondrial targeting sequences (MTS). The two targeting sequences with highest probability of mitochondrial import are marked in bold. Red asterisks denote predicted cleavage sites for MPP. Sample readouts from MitoProtII are shown in (B) and (C).
 Schematic and summary of Ft intracellular fragments used in mitochondrial")
113 Figure 3.12: Ft contains multiple mitochondrial targeting sequences that direct mitochondrial import in transfected cell assays (A) Schematic and summary of Ft intracellular fragments used in mitochondrial localization assays. Localization data summarized to the right. (B) Ft fragments a-d partially colocalize with MitoGFP in COS cells, whereas e and f do not, as detected by immunofluorescence (IF). 102
 FtIC does not colocalize with mitochondrial markers in FtIC transfected COS cells.")
114 Figure 3.13: Membrane association is important for mitochondrial localization of Ft. (A) Ndufv2 and Ft colocalize in mitochondria of S2 cells transfected with FtΔECD-FLAG (Ft lacking the ECD). (B) FtIC does not colocalize with mitochondrial markers in FtIC transfected COS cells. (C, D) Immunoblots of fractionated S2 cells transfected with FtΔECD (C) or FtIC (D) show that Ft ECD is processed into 110 and 68kDa forms, whereas FtIC runs as a discrete 74kDa band. Notably the processed 68 kda fragment is detected in the mitochondrial fraction while the FtIC fragment is not. T total extract, M mitochondrial fraction. 103
 Ft mito is specifically enriched in mitochondrial fractions from D11 cells, along with Porin, but not α-tubulin.")
115 Figure 3.14: Endogenous Ft localizes to mitochondria in D11 cells. (A) Endogenous Ft colocalizes with mitochondrial marker CVα in D11 cells. (B) Ft mito is specifically enriched in mitochondrial fractions from D11 cells, along with Porin, but not α-tubulin. Asterisk denotes a 50kDa Ft fragment found in the total and heavy membrane fractions. (C) In D11 cells, Ft mito is protected from protease digestion by Proteinase K, as is CVα, whereas the 110kDa fragment is degraded. Red arrows indicate the 68kDa Ft mito fragment, black arrowheads indicate the 110kDa fragment. T total, C cytoplasmic, H heavy membranes, M mitochondrial fraction. 104
116 105 fractionation experiments revealed that a ~68kDa fragment of the endogenous Ft intracellular domain is enriched in mitochondrial fractions from D11 cells (Fig. 3.14B). To determine if the ~68kDa fragment of Ft is located within mitochondria, we conducted protease protection assays on intact mitochondria isolated from D11 cells. The 68kDa fragment is resistant to protease digestion in intact organelles (Fig. 3.14C). In contrast, the 110kDa fragment was digested by protease treatment, suggesting that the transmembrane form of Ft is associated with mitochondria, but is not imported into mitochondria. Importantly, addition of Triton X-100 to permeabilize mitochondrial membranes resulted in digestion of the 68kDa fragment, verifying that this fragment of Ft is susceptible to digestion by proteinase K, but is protected from proteolysis until membranes are solubilized. Taken together, these data indicate that a 68kDa Ft fragment is imported into mitochondria. We refer to this 68kDa Ft fragment as Ft mito Ft is detected in mitochondria in vivo To determine if Ft processing occurs in vivo, we isolated eye discs from wildtype and ft larvae and conducted fractionation and protease digestion experiments (Figure 4H and S4C). In total larval extracts, the 110kDa form of Ft was the most abundant form. Isolation of mitochondrial fractions of wildtype larvae revealed enrichment of the endogenous ~68kDa form of Ft. Both the 110kDa and 68kDa fragments were missing in immunoblots of mitochondrial extracts obtained from ft larvae (Fig. 3.15A), confirming that the 68kDa fragment represents mitochondrial Ft (Ft mito ). As in cultured cells, Ft mito is resistant to protease digestion in larval extracts, confirming its localization inside mitochondria (Fig. 3.15B). Interestingly, Ft mito appears as a doublet in disc extracts, suggesting additional processing may occur in vivo. Discs and fat body (Fig. 3.15C, D) display clear co-localization of Ft and mitochondrial markers; however, interestingly the staining was often unevenly distributed, suggesting that Ft mito may be restricted to specific subdomains in mitochondria. Ds binding to the extracellular domain of Ft is important for the regulation of Hippo and PCP signaling. We tested if loss of ds altered cleavage or import of Ft into mitochondria. We found that Ft mito is clearly detectable in ds mutant larval discs (Fig. 3.16A). Thus mitochondrial localization of Ft occurs in the absence of Ds binding.
 Ft mito is enriched in vivo in mitochondrial fractions from wildtype and ds - larval discs, but is absent in ft alb/fd disc extracts.")
. T total, M mitochondrial fraction.")
117 Figure 3.15: A fragment of Ft is localized to mitochondria in imaginal discs. (A) Ft mito is enriched in vivo in mitochondrial fractions from wildtype and ds - larval discs, but is absent in ft alb/fd disc extracts. *NS non-specific fragment detected by Ft antibody. (B) Ft mito, CV and Porin are protected from protease digestion in wildtype and ds - larval discs. Ft mito fragment is indicated by red arrows, black arrowheads indicate the 110kDa fragment in (A, B). T total, M mitochondrial fraction. (C, D) Structured Illumination Microscopy (SIM) of Drosophila larval tissue shows partial costaining of endogenous Ft and mitochondrial marker CV 106

118 Figure 3.16: ft larvae have altered levels of mitochondrial proteins. Immunoblots against lysates from wildtype versus ft fd larval eye/brain complexes show a slight decrease in Ndufv2, increase in CytoC, and no change in PDH, CVα and Porin protein levels. α-actin serves as a loading control. 107
119 ft mutants show loss of assembled CI holoenzyme We wondered if Ft regulates the stability of Ndufv2, but we found only mild changes in Ndufv2 total levels in SDS PAGE analysis of ft larvae (Fig. 3.16). We hypothesized that Ft might affect the assembly or stability of the CI holoenzyme, which would result in loss of CI activity. CI forms a complex of >900kDa that can be resolved on non-denaturing, blue nativepolyacrylamide gel electrophoresis (BN-PAGE). BN-PAGE analysis of mitochondria from wt larvae shows a clear band for fully assembled CI at ~980kDa, as well as complex V (CV) monomers at ~800kDa and CV dimers at ~1100kDa (Fig. 3.17A). As previously published, loss of a well-characterized CI assembly factor (dcia) results in loss of CI, with no effects on the assembly of CV monomer (CV1) or CV dimer (CV2) (Fig. 3.17A and Cho et al., 2012). Strikingly, BN-PAGE of mitochondria isolated from ft mutants shows loss of assembled CI, suggesting Ft is needed for assembly or stability of CI (Fig. 3.17A). Western blot analysis with antibodies to CI components Ndufv2 and Ndufs3 confirmed loss of assembled CI in ft fd mutants (Fig. 3.17B). Overexposure of gels reveals there is some CI present, but at drastically reduced levels. In-gel assays confirm that CI activity is dramatically reduced in ft mutants (Fig. 3.17C). There is also a consistent increase in CII activity, and mobility shifts in CIV and CV activity. Western blot analysis demonstrated that a null ft allele ft fd, shows a dramatic loss of CI, while the hypomorphic ft allele, ft alb, does not significantly alter CI assembly or stability (Fig. 3.18A). Importantly, western blotting for Ndufv2 revealed that defects in CI can be rescued by expression of full-length Ft on a BAC transgene. Impaired CI assembly provides an explanation for the reduced CI activity seen in ft fd mutants. Interestingly, ft fd mutants as well as dcia mutants show an increase in complex-associated CytoC, possibly reflecting metabolic compensation (Fig A, B). Mutations of Hippo pathway components, wts and ds, do not affect CI levels but do show an increase in CytoC, possibly due to regulation of mitochondrial dynamics (Nagaraj et al., 2012). We hypothesized that the 68kDa Ft mito might be a component of mitochondrial complexes. Immunoblotting of larval mitochondrial extracts separated on BN-PAGE gels with affinitypurified antibodies to Ft revealed that Ft mito is part of the CI holoenzyme (Fig. 3.17B). Importantly, the Ft signal is lost in CI assembly mutants, which also lose Ndufv2 staining as well

120 Figure 3.17: ft fd larvae show defecits in CI and CV assembly and activity. (A) BN-PAGE of crude mitochondrial isolates from larval extracts reveal CI deficiencies in ft fd and dcia ex80. Wedges indicate increasing amounts of extract. Arrow and asterisk denote CI; arrowheads indicate CV bands. (B) Immunoblot of BN-PAGE confirms absence of CI in ft fd and dcia ex80 and Ft staining indicates Ft associates with CI. Assembly of CV is also disrupted in ft fd mutants, with a CVα-containing fragment of ~500kDa. (C) ClearNative-PAGE (bottom, for loading control) and in-gel activity assays (top) reveal strong decreases in CI activity, increase in CII activity, and a shift in CIV mobility. CV activity is perturbed. 109
 Immunoblots of crude mitochondrial extracts run on native PAGE (A) The hypomorphic allele ft alb shows defects in CV assembly, while CI is unaffected as shown by Ndufv2 levels.")
121 Figure 3.18: ft pathway mutants have increased levels of CytoC and are rescued by expressing full-length Ft in ft null larvae. (A-C) Immunoblots of crude mitochondrial extracts run on native PAGE (A) The hypomorphic allele ft alb shows defects in CV assembly, while CI is unaffected as shown by Ndufv2 levels. Expression of two copies of a full-length ft BAC transgene (ft FL ) rescues all of these defects in a ft fd background. (B) Complex-associated Cytochrome C is upregulated in both ft fd and dcia ex80. (C) The ft fd/x13 transheterozygote phenocopies ft fd in both CI and CV defects. wts and ds mutants retain intact CI and CV, but upregulate CytoC. 110
122 111 as other CI components. No signal was detected in CV complexes. Together, these data indicate that Ft mito is incorporated into CI, and is needed for CI assembly or stability in vivo Loss of Ft leads to defects in CV assembly. Immunoblotting with antibodies to CVα reveals normally assembled CV1 and CV2 in wildtype larvae and CI assembly mutants (Fig. 3.17B, 3.18B). Strikingly, however, CV is incorrectly assembled in both ft fd and ft alb mutants, with reduction of CV dimers, and the appearance of a novel prominent band at ~500kDa (Fig. 3.17B, 3.18A, B). ft alb appears to have defects specifically in CV while CI remains intact. Taken together with the fact that CV is unaffected in dcia mutants despite the deficits in CI, we infer that these observed CV assembly defects are separable from loss of CI. Furthermore, normal assembly of CV in ft mutants can be rescued by expression of full length Ft (Ft-FL) provided by a BAC transgene (Fig. 3.18A). These data suggest that Ft mito is also needed for proper assembly and/or stability of CV. Our Y2H screens had indicated that Ft interacts with CG1746, a component of CV, and my RNAi screens had demonstrated that loss of CG1746 leads to PCP defects (Fig. 2.3 and Table 2.1). GST pull-down assays confirmed that CG1746 binds directly to the cytoplasmic domain of Ft (Fig. 2.14). We do not detect Ft mito on assembled CV in BN-PAGE analysis, suggesting this interaction may be weak or transient (Fig. 3.17B). However, the obvious defects in CV assembly in ft mutants suggest that the Ft-CG1746 interaction is biologically relevant. Importantly, ftdependent defects in CV are not secondary to reduced CI, as loss of CI (through mutation of a CI assembly factor, dcia), does not lead to accumulation of the aberrant ~500kDa CV band (Fig. 3.17B, 3.18B). As CV dimerization is essential for cristae formation (Paumard et al., 2002), one interesting possibility is that the cristae morphology defects in ft mutants may be due to loss of interaction between CG1746 and Ft The Ndufv2-binding region of Ft is necessary for normal larval growth and mitochondrial function We reasoned that if Ft binding to Ndufv2 is essential for mitochondrial complex assembly and OXPHOS, blocking this interaction should result in CI and CV defects. We first tested if BAC transgenes containing full length Ft can rescue the early growth defects of ft mutant larva. Fulllength Ft can rescue the early growth defect, confirming that the growth delay is due to Ft loss
123 112 (Fig. 3.19B, C). Additionally, the increased level of lactate in ft mutants is also rescued by fulllength Ft (Fig. 3.19D). Importantly, BN-PAGE analysis shows that full length Ft can rescue CI and CV assembly (Fig. 3.19E). Previous studies have defined 8 highly conserved regions in the intracellular domain of Ft (Fig. 3.19A), and tested the requirement for these regions in Ft function. Pan et al. (2013) have shown that BAC transgenes expressing a full-length Ft lacking region 2 (FLΔ2) can fully rescue Hippo and PCP signaling, implying this region is not necessary for Hippo or PCP signaling. In contrast, rescuing ft mutants with a BAC transgene expressing Ft with a deletion of region 8 (FLΔ8) rescues Hippo pathway defects but not PCP defects, implying this region is essential for PCP activity. We have defined region 2 as the domain that binds Ndufv2 (see chapter and Fig. 2.14). Therefore, if binding between Ft and Ndufv2 is important for normal mitochondrial function, FLΔ2 should not be able to rescue mitochondrial defects, whereas deletion of an unrelated region, e.g. FLΔ8, would still retain mitochondrial functions. Significantly, FLΔ2 fails to rescue larval growth, lactic acid production, and CI or CV assembly, while FLΔ8 does rescue (Fig B-E). Thus the region of Ft that binds Ndufv2 is essential for mitochondrial complex stability, and mitochondrial ETC activity.
 FL and FLΔ8 rescue the delay in growth of ft fd larvae, while FLΔ2 does not. (D) Lactate levels are elevated in FLΔ2-expressing ft fd. Levels normalized to FL-rescued lactate content.")
124 Fig 3.19: Ft conserved region 2 is necessary for mitochondrial function. (A) Schematic of FL, FLΔ2 and FLΔ8 rescue constructs. (B-C) FL and FLΔ8 rescue the delay in growth of ft fd larvae, while FLΔ2 does not. (D) Lactate levels are elevated in FLΔ2-expressing ft fd. Levels normalized to FL-rescued lactate content. One sample Student s t-test used to test significance : µ0=1, n=4, p= (E-F) Immunoblotting of crude mitochondrial extracts run on native-page reveals that CI and CV defects in ft fd larvae are partially rescued by FLΔ8 but not FLΔ2 (E) and that the rescue of CI and CV deficiencies in ft fd larvae is ft FL dosage-dependent (F). 113
125 Discussion Here, I have shown an unexpected and direct role for Ft in regulating mitochondrial morphology and metabolism. I find that Ft controls ROS production and promotes OXPHOS activity during Drosophila development. Ft is cleaved, generating a 68kDa fragment that is imported into mitochondria, where it binds Ndufv2, promotes CI and CV assembly and/or stability, and enhances the activity of the ETC. To my knowledge this is the first example of a cell surface protein that functions to directly stabilize the mitochondrial ETC and promote OXPHOS Ft function in the mitochondria is separable from Hippo and PCP effects. Previous studies indicated that the Hippo and PCP functions of Ft map to distinct regions of the Ft cytoplasmic domain (Bossuyt et al., 2014; Matakatsu and Blair, 2012; Pan et al., 2013; Zhao et al., 2013). We show here that the regulation of CI by Ft is separable from its roles in Hippo signaling. While loss of ft leads to increased ROS, loss of wts or overexpression of yki does not increase ROS (Fig. 3.6C, D). In addition, BN-PAGE analysis demonstrates that CI and CV levels are not altered in wts and ds mutants (Fig. 3.18C), further indicating the mitochondrial effects of Ft to promote CI and CV levels are independent of Hippo signaling. Importantly, in the context of full length Ft expressed under the control of its endogenous promoter, deletion of Regions 1 and 2 have no detectable effect on PCP or Hippo activity. Interestingly, in experiments in which truncated versions of Ft are overexpressed via the UAS/Gal4 system, these regions do have weak PCP and Hippo activity (Matakatsu and Blair, 2012). As discussed in chapter 2, loss of mitochondrial integrity can lead to PCP and Hippo defects. I propose that the weak Hippo and PCP effects seen in overexpression experiments in Matakatsu et al. (2013) may be in part due to feedback from disruption of the mitochondrial functions of Ft A rheostat model for ft function Thus far, Ft has been presumed to transduce extracellular signals from the cell surface to activate the Hippo kinase pathway. Activation of the Hippo kinase pathway ultimately sequesters Yki away from the nucleus, thus repressing the transcription of cell proliferation and anti-apoptotic genes. I have now shown that Ft can be cleaved and targeted to mitochondria to regulate metabolic activity. The proteolytic cleavage that generates Ft mito would remove Ft from the cell surface, rendering it unable to interact with Hpo pathway components, and therefore no longer
126 115 active in the inhibition of cell proliferation. Instead, Ft mito enters the mitochondrion, where it binds Ndufv2 and stabilizes CI and CV, thus promoting OXPHOS. I propose that cleavage of Ft can function as a switch mechanism to coordinate cell cycle and metabolism (Fig. 3.20). Altering the levels of Ft mito could allow an organism to directly and incrementally adjust metabolic rates in accordance with the dynamic energy requirements of the cell as it enters and exits proliferative states. In support of this rheostat model, we find that defects in ETC assembly are rescued in a dose-dependent manner by one and two copies of the Ft transgene (Fig. 3.19F). I do not yet know the identity of the protease that cleaves Ft releasing Ft mito, but I hypothesize that regulation of this cleavage would provide a potent mechanism for coordinating growth and metabolism Fat cadherin Mitochondrial binding motifs and targeting sequences are ancient and conserved The cytoplasmic domain of Ft has several regions that are evolutionarily conserved. Heretofore, the function of some of the most highly conserved residues of Ft has been unclear. Ft binding to CI and stabilization of CI depends on these highly conserved residues (Fig and 3.21), suggesting the urbilaterian ancestor had a form of Ft mito and that an ancient function of Ft is to regulate mitochondrial activity. Consistent with this model, mammalian Fat4 has two highconfidence mitochondrial targeting sequences, similar to Ft (Fig. 3.22), supporting the conservation of a mitochondrial role for Fat4. The role of Ft in PCP (Saburi et al., 2008; Saburi et al., 2012), and possibly Hippo activity (Cappello et al., 2013), is conserved in mammals. This deep conservation of CI binding sequences suggests there is also conservation of the mitochondrial functions of Fat cadherins Ft mito and metabolism in cancer cells As CV is a major source of cellular ATP, I measured ATP levels in ft mutant larvae. Loss of ft lead to no significant effect on total ATP levels (Fig. 3.8B). Defects in OXPHOS can be compensated for by increased glycolysis. Analysis of ft mutant larva showed greatly increased levels of lactic acid (Fig. 3.8C), consistent with a shift in metabolism of ft mutants from OXPHOS to glycolysis. This is intriguing, as ft is a classic Drosophila tumor suppressor gene, and a hallmark of cancer cells in mammals is a shift from oxidative metabolism to aerobic glycolysis, known as the Warburg effect (reviewed in Lunt and Vander Heiden, 2011). ft mutants show a long delay before discs begin to overgrow. Our data suggest that the mitochondrial

127 Fig 3.20: Model of Ft processing and function at the cell surface and in stabilizing the ETC in mitochondria. Ft is processed to a 450 and 110kDa receptor at the cell membrane where it coordinates PCP and restricts growth via the Hpo pathway. Ft is further cleaved and transported to mitochondria, relieving repression of growth by the Hpo pathway. Once inside mitochondria Ft interacts with CI and CV to promote OXPHOS. In cells lacking Ft, both PCP and Hpo pathways are disregulated leading to PCP defects and unrestricted growth. CI and CV are destabilized leading to release of ROS and impaired OXPHOS, resulting in a metabolic shift to glycolysis, resulting in higher levels of lactate 116

128 Fig 3.21: Mitochondrial-binding regions of Ft are highly conserved. Regions of the Ft intracellular domain have been shown to function in processes including PCP and Hippo. Regions 1 and 2, demarcated by red boxes, are highly conserved among bilaterians and are found in this study to bind the mitochondrial Ndufv2 and CG1746 proteins. DmFat D. melanogaster, AgFat Anopheles gambiae, TcFat Tribolium castaneum, DpFat Daphnia pulex, BfFat Branchiostoma floridae, MmFat4 M. musculus and HsFat4 H. sapiens. See Matakatsu and Blair (2012) for definitions of PCP and Hippo N regions. 117
The Atypical Cadherin Fat Directly Regulates Mitochondrial Function and Metabolic State
 The Atypical Cadherin Fat Directly Regulates Mitochondrial Function and Metabolic State Anson Sing, 1,2,5 Yonit Tsatskis, 1,5 Lacramioara Fabian, 3 Ian Hester, 1 Robyn Rosenfeld, 1,2 Mauro Serricchio,
The Atypical Cadherin Fat Directly Regulates Mitochondrial Function and Metabolic State Anson Sing, 1,2,5 Yonit Tsatskis, 1,5 Lacramioara Fabian, 3 Ian Hester, 1 Robyn Rosenfeld, 1,2 Mauro Serricchio,
Conclusions. The experimental studies presented in this thesis provide the first molecular insights
 C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Separating the adhesive and signaling functions of the Fat and Dachsous protocadherins
 RESEARCH ARTICLE 2315 Development 133, 2315-2324 (2006) doi:10.1242/dev.02401 Separating the adhesive and signaling functions of the Fat and Dachsous protocadherins Hitoshi Matakatsu and Seth S. Blair*
RESEARCH ARTICLE 2315 Development 133, 2315-2324 (2006) doi:10.1242/dev.02401 Separating the adhesive and signaling functions of the Fat and Dachsous protocadherins Hitoshi Matakatsu and Seth S. Blair*
Regulation of PCP by the Fat signaling pathway
 Downloaded from genesdev.cshlp.org on October 18, 2013 - Published by Cold Spring Harbor Laboratory Press Regulation of PCP by the Fat signaling pathway Maja Matis and Jeffrey D. Axelrod Genes Dev. 2013
Downloaded from genesdev.cshlp.org on October 18, 2013 - Published by Cold Spring Harbor Laboratory Press Regulation of PCP by the Fat signaling pathway Maja Matis and Jeffrey D. Axelrod Genes Dev. 2013
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway
 Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
What are mitochondria?
 What are mitochondria? What are mitochondria? An intracellular organelle. There are 100 to 1000s of mitochondria/cell. Most mitochondria come from the mother. Mitochondria have their own DNA Mitochondria
What are mitochondria? What are mitochondria? An intracellular organelle. There are 100 to 1000s of mitochondria/cell. Most mitochondria come from the mother. Mitochondria have their own DNA Mitochondria
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Cells to Tissues. Peter Takizawa Department of Cell Biology
 Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Cell Death & Trophic Factors II. Steven McLoon Department of Neuroscience University of Minnesota
 Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Metabolism. Fermentation vs. Respiration. End products of fermentations are waste products and not fully.
 Outline: Metabolism Part I: Fermentations Part II: Respiration Part III: Metabolic Diversity Learning objectives are: Learn about respiratory metabolism, ATP generation by respiration linked (oxidative)
Outline: Metabolism Part I: Fermentations Part II: Respiration Part III: Metabolic Diversity Learning objectives are: Learn about respiratory metabolism, ATP generation by respiration linked (oxidative)
Biochemical bases for energy transformations. Biochemical bases for energy transformations. Nutrition 202 Animal Energetics R. D.
 Biochemical bases for energy transformations Biochemical bases for energy transformations Nutrition 202 Animal Energetics R. D. Sainz Lecture 02 Energy originally from radiant sun energy Captured in chemical
Biochemical bases for energy transformations Biochemical bases for energy transformations Nutrition 202 Animal Energetics R. D. Sainz Lecture 02 Energy originally from radiant sun energy Captured in chemical
M i t o c h o n d r i a
 M i t o c h o n d r i a Dr. Diala Abu-Hassan School of Medicine dr.abuhassand@gmail.com Mitochondria Function: generation of metabolic energy in eukaryotic cells Generation of ATP from the breakdown of
M i t o c h o n d r i a Dr. Diala Abu-Hassan School of Medicine dr.abuhassand@gmail.com Mitochondria Function: generation of metabolic energy in eukaryotic cells Generation of ATP from the breakdown of
TCA Cycle. Voet Biochemistry 3e John Wiley & Sons, Inc.
 TCA Cycle Voet Biochemistry 3e Voet Biochemistry 3e The Electron Transport System (ETS) and Oxidative Phosphorylation (OxPhos) We have seen that glycolysis, the linking step, and TCA generate a large number
TCA Cycle Voet Biochemistry 3e Voet Biochemistry 3e The Electron Transport System (ETS) and Oxidative Phosphorylation (OxPhos) We have seen that glycolysis, the linking step, and TCA generate a large number
Supplementary Figures
 Supplementary Figures Supplementary Fig. S1: Normal development and organization of the embryonic ventral nerve cord in Platynereis. (A) Life cycle of Platynereis dumerilii. (B-F) Axonal scaffolds and
Supplementary Figures Supplementary Fig. S1: Normal development and organization of the embryonic ventral nerve cord in Platynereis. (A) Life cycle of Platynereis dumerilii. (B-F) Axonal scaffolds and
Lecture 7 Cell Biolog y ٢٢٢ ١
 Lecture 7 ١ Mitochondria ٢ Mitochondria Mitochondria are the energy factories of the cells. The energy currency for the work that animals must do is the energy-rich molecule adenosine triphosphate (ATP).
Lecture 7 ١ Mitochondria ٢ Mitochondria Mitochondria are the energy factories of the cells. The energy currency for the work that animals must do is the energy-rich molecule adenosine triphosphate (ATP).
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
Lectures by Kathleen Fitzpatrick
 Chapter 10 Chemotrophic Energy Metabolism: Aerobic Respiration Lectures by Kathleen Fitzpatrick Simon Fraser University Figure 10-1 Figure 10-6 Conversion of pyruvate The conversion of pyruvate to acetyl
Chapter 10 Chemotrophic Energy Metabolism: Aerobic Respiration Lectures by Kathleen Fitzpatrick Simon Fraser University Figure 10-1 Figure 10-6 Conversion of pyruvate The conversion of pyruvate to acetyl
AP Bio-Ms.Bell Unit#3 Cellular Energies Name
 AP Bio-Ms.Bell Unit#3 Cellular Energies Name 1. Base your answer to the following question on the image below. 7. Base your answer to the following question on Which of the following choices correctly
AP Bio-Ms.Bell Unit#3 Cellular Energies Name 1. Base your answer to the following question on the image below. 7. Base your answer to the following question on Which of the following choices correctly
Aerobic Cellular Respiration
 Aerobic Cellular Respiration Under aerobic conditions (oxygen gas is available), cells will undergo aerobic cellular respiration. The end products of aerobic cellular respiration are carbon dioxide gas,
Aerobic Cellular Respiration Under aerobic conditions (oxygen gas is available), cells will undergo aerobic cellular respiration. The end products of aerobic cellular respiration are carbon dioxide gas,
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
A reaction-diffusion, partial differential equation model of planar cell polarity signaling
 1 Mathematical modeling of planar cell polarity to understand domineering non-autonomy Keith Amonlirdviman 1, Narmada A. Khare 2, David R.P. Tree 2, Wei-Shen Chen 2, Jeffrey D. Axelrod 2* and Claire J.
1 Mathematical modeling of planar cell polarity to understand domineering non-autonomy Keith Amonlirdviman 1, Narmada A. Khare 2, David R.P. Tree 2, Wei-Shen Chen 2, Jeffrey D. Axelrod 2* and Claire J.
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis
 18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
Cellular Respiration: Harvesting Chemical Energy. 9.1 Catabolic pathways yield energy by oxidizing organic fuels
 Cellular Respiration: Harvesting Chemical Energy 9.1 Catabolic pathways yield energy by oxidizing organic fuels 9.2 Glycolysis harvests chemical energy by oxidizing glucose to pyruvate 9.3 The citric acid
Cellular Respiration: Harvesting Chemical Energy 9.1 Catabolic pathways yield energy by oxidizing organic fuels 9.2 Glycolysis harvests chemical energy by oxidizing glucose to pyruvate 9.3 The citric acid
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
CHAPTER 1 INTRODUCTION TO CELLS 2009 Garland Science Publishing 3 rd Edition
 Unity and Diversity of Cells 1-1 The smallest unit of life is a(n) (a) DNA molecule. (b) cell. (c) organelle. (d) virus. (e) protein. CHAPTER 1 INTRODUCTION TO CELLS 2009 Garland Science Publishing 3 rd
Unity and Diversity of Cells 1-1 The smallest unit of life is a(n) (a) DNA molecule. (b) cell. (c) organelle. (d) virus. (e) protein. CHAPTER 1 INTRODUCTION TO CELLS 2009 Garland Science Publishing 3 rd
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290
 PLoS Biol 4(9): e290") Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Cell Biology Review. The key components of cells that concern us are as follows: 1. Nucleus
 Cell Biology Review Development involves the collective behavior and activities of cells, working together in a coordinated manner to construct an organism. As such, the regulation of development is intimately
Cell Biology Review Development involves the collective behavior and activities of cells, working together in a coordinated manner to construct an organism. As such, the regulation of development is intimately
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
2015 AP Biology PRETEST Unit 3: Cellular Energetics Week of October
 Name: Class: _ Date: _ 2015 AP Biology PRETEST Unit 3: Cellular Energetics Week of 19-23 October Multiple Choice Identify the choice that best completes the statement or answers the question. 1) Which
Name: Class: _ Date: _ 2015 AP Biology PRETEST Unit 3: Cellular Energetics Week of 19-23 October Multiple Choice Identify the choice that best completes the statement or answers the question. 1) Which
20. Electron Transport and Oxidative Phosphorylation
 20. Electron Transport and Oxidative Phosphorylation 20.1 What Role Does Electron Transport Play in Metabolism? Electron transport - Role of oxygen in metabolism as final acceptor of electrons - In inner
20. Electron Transport and Oxidative Phosphorylation 20.1 What Role Does Electron Transport Play in Metabolism? Electron transport - Role of oxygen in metabolism as final acceptor of electrons - In inner
Lecture Series 9 Cellular Pathways That Harvest Chemical Energy
 Lecture Series 9 Cellular Pathways That Harvest Chemical Energy Reading Assignments Review Chapter 3 Energy, Catalysis, & Biosynthesis Read Chapter 13 How Cells obtain Energy from Food Read Chapter 14
Lecture Series 9 Cellular Pathways That Harvest Chemical Energy Reading Assignments Review Chapter 3 Energy, Catalysis, & Biosynthesis Read Chapter 13 How Cells obtain Energy from Food Read Chapter 14
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Bio102 Problems Photosynthesis
 Bio102 Problems Photosynthesis 1. Why is it advantageous for chloroplasts to have a very large (in surface area) thylakoid membrane contained within the inner membrane? A. This limits the amount of stroma
Bio102 Problems Photosynthesis 1. Why is it advantageous for chloroplasts to have a very large (in surface area) thylakoid membrane contained within the inner membrane? A. This limits the amount of stroma
Developmental Biology
 Developmental Biology 350 (2011) 255 266 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology The Salvador/Warts/Hippo pathway controls
Developmental Biology 350 (2011) 255 266 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology The Salvador/Warts/Hippo pathway controls
All organisms require a constant expenditure of energy to maintain the living state - "LIFE".
 CELLULAR RESPIRATION All organisms require a constant expenditure of energy to maintain the living state - "LIFE". Where does the energy come from and how is it made available for life? With rare exception,
CELLULAR RESPIRATION All organisms require a constant expenditure of energy to maintain the living state - "LIFE". Where does the energy come from and how is it made available for life? With rare exception,
Transport between cytosol and nucleus
 of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
Lesson Overview. Gene Regulation and Expression. Lesson Overview Gene Regulation and Expression
 13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
Energy Converion: Mitochondria and Chloroplasts. Pınar Tulay, Ph.D.
 Energy Converion: Mitochondria and Chloroplasts Pınar Tulay, Ph.D. pintulay@gmail.com Energy Conversion Prokaryotes use plasma membrane to produce adenosine triphosphate (ATP) used in the cell function
Energy Converion: Mitochondria and Chloroplasts Pınar Tulay, Ph.D. pintulay@gmail.com Energy Conversion Prokaryotes use plasma membrane to produce adenosine triphosphate (ATP) used in the cell function
3/8/ Complex adaptations. 2. often a novel trait
 Chapter 10 Adaptation: from genes to traits p. 302 10.1 Cascades of Genes (p. 304) 1. Complex adaptations A. Coexpressed traits selected for a common function, 2. often a novel trait A. not inherited from
Chapter 10 Adaptation: from genes to traits p. 302 10.1 Cascades of Genes (p. 304) 1. Complex adaptations A. Coexpressed traits selected for a common function, 2. often a novel trait A. not inherited from
Extranuclear Inheritance
 Extranuclear Inheritance Extranuclear Inheritance The past couple of lectures, we ve been exploring exceptions to Mendel s principles of transmission inheritance. Scientists have observed inheritance patterns
Extranuclear Inheritance Extranuclear Inheritance The past couple of lectures, we ve been exploring exceptions to Mendel s principles of transmission inheritance. Scientists have observed inheritance patterns
Principles of Cellular Biology
 Principles of Cellular Biology آشنایی با مبانی اولیه سلول Biologists are interested in objects ranging in size from small molecules to the tallest trees: Cell Basic building blocks of life Understanding
Principles of Cellular Biology آشنایی با مبانی اولیه سلول Biologists are interested in objects ranging in size from small molecules to the tallest trees: Cell Basic building blocks of life Understanding
BIOH111. o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system
 BIOH111 o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 Textbook
BIOH111 o Cell Biology Module o Tissue Module o Integumentary system o Skeletal system o Muscle system o Nervous system o Endocrine system Endeavour College of Natural Health endeavour.edu.au 1 Textbook
The Mitochondrion. Definition Structure, ultrastructure Functions
 The Mitochondrion Definition Structure, ultrastructure Functions Organelle definition Etymology of the name Carl Benda (1903): (mitos) thread; (khondrion) granule. Light microscopy identification First
The Mitochondrion Definition Structure, ultrastructure Functions Organelle definition Etymology of the name Carl Benda (1903): (mitos) thread; (khondrion) granule. Light microscopy identification First
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
GSBHSRSBRSRRk IZTI/^Q. LlML. I Iv^O IV I I I FROM GENES TO GENOMES ^^^H*" ^^^^J*^ ill! BQPIP. illt. goidbkc. itip31. li4»twlil FIFTH EDITION
 FIFTH EDITION IV I ^HHk ^ttm IZTI/^Q i I II MPHBBMWBBIHB '-llwmpbi^hbwm^^pfc ' GSBHSRSBRSRRk LlML I I \l 1MB ^HP'^^MMMP" jflp^^^^^^^^st I Iv^O FROM GENES TO GENOMES %^MiM^PM^^MWi99Mi$9i0^^ ^^^^^^^^^^^^^V^^^fii^^t^i^^^^^
FIFTH EDITION IV I ^HHk ^ttm IZTI/^Q i I II MPHBBMWBBIHB '-llwmpbi^hbwm^^pfc ' GSBHSRSBRSRRk LlML I I \l 1MB ^HP'^^MMMP" jflp^^^^^^^^st I Iv^O FROM GENES TO GENOMES %^MiM^PM^^MWi99Mi$9i0^^ ^^^^^^^^^^^^^V^^^fii^^t^i^^^^^
Ev e ry living c e l l needs a source of
 12 Photosynthesis and Cellular Respiration Ev e ry living c e l l needs a source of energy. Without energy, metabolism all of the chemical reactions that occur within cells will not occur. In this activity,
12 Photosynthesis and Cellular Respiration Ev e ry living c e l l needs a source of energy. Without energy, metabolism all of the chemical reactions that occur within cells will not occur. In this activity,
Role of Mitochondrial Remodeling in Programmed Cell Death in
 Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Supplementary Information
 Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
Life 21 - Aerobic respiration Raven & Johnson Chapter 9 (parts)
") 1 Life 21 - Aerobic respiration Raven & Johnson Chapter 9 (parts) Objectives 1: Describe the overall action of the Krebs cycle in generating ATP, NADH and FADH 2 from acetyl-coa 2: Understand the generation
1 Life 21 - Aerobic respiration Raven & Johnson Chapter 9 (parts) Objectives 1: Describe the overall action of the Krebs cycle in generating ATP, NADH and FADH 2 from acetyl-coa 2: Understand the generation
METABOLISM CHAPTER 04 BIO 211: ANATOMY & PHYSIOLOGY I. Dr. Lawrence G. Altman Some illustrations are courtesy of McGraw-Hill.
 BIO 211: ANATOMY & PHYSIOLOGY I CHAPTER 04 1 Please wait 20 seconds before starting slide show. Mouse click or Arrow keys to navigate. Hit ESCAPE Key to exit. CELLULAR METABOLISM Dr. Lawrence G. Altman
BIO 211: ANATOMY & PHYSIOLOGY I CHAPTER 04 1 Please wait 20 seconds before starting slide show. Mouse click or Arrow keys to navigate. Hit ESCAPE Key to exit. CELLULAR METABOLISM Dr. Lawrence G. Altman
CELL METABOLISM OVERVIEW Keep the big picture in mind as we discuss the particulars!
 BIO 211: ANATOMY & PHYSIOLOGY I CHAPTER 04 CELLULAR METABOLISM 1 Please wait 20 seconds before starting slide show. Mouse click or Arrow keys to navigate. Hit ESCAPE Key to exit. Dr. Lawrence G. Altman
BIO 211: ANATOMY & PHYSIOLOGY I CHAPTER 04 CELLULAR METABOLISM 1 Please wait 20 seconds before starting slide show. Mouse click or Arrow keys to navigate. Hit ESCAPE Key to exit. Dr. Lawrence G. Altman
Scale in the biological world
 Scale in the biological world 2 A cell seen by TEM 3 4 From living cells to atoms 5 Compartmentalisation in the cell: internal membranes and the cytosol 6 The Origin of mitochondria: The endosymbion hypothesis
Scale in the biological world 2 A cell seen by TEM 3 4 From living cells to atoms 5 Compartmentalisation in the cell: internal membranes and the cytosol 6 The Origin of mitochondria: The endosymbion hypothesis
Photosynthesis and Cellular Respiration
 Name Date Class CHAPTER 5 TEST PREP PRETEST Photosynthesis and Cellular Respiration In the space provided, write the letter of the term or phrase that best completes each statement or best answers each
Name Date Class CHAPTER 5 TEST PREP PRETEST Photosynthesis and Cellular Respiration In the space provided, write the letter of the term or phrase that best completes each statement or best answers each
Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.
 Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.441/HST522J INTEGRINS I.V. Yannas, Ph.D. and M. Spector, Ph.D. Regulator
Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.441/HST522J INTEGRINS I.V. Yannas, Ph.D. and M. Spector, Ph.D. Regulator
Chapter Cells and the Flow of Energy A. Forms of Energy 1. Energy is capacity to do work; cells continually use energy to develop, grow,
 Chapter 6 6.1 Cells and the Flow of Energy A. Forms of Energy 1. Energy is capacity to do work; cells continually use energy to develop, grow, repair, reproduce, etc. 2. Kinetic energy is energy of motion;
Chapter 6 6.1 Cells and the Flow of Energy A. Forms of Energy 1. Energy is capacity to do work; cells continually use energy to develop, grow, repair, reproduce, etc. 2. Kinetic energy is energy of motion;
Eukaryotic Cells. Figure 1: A mitochondrion
 Eukaryotic Cells Figure 1: A mitochondrion How do cells accomplish all their functions in such a tiny, crowded package? Eukaryotic cells those that make up cattails and apple trees, mushrooms and dust
Eukaryotic Cells Figure 1: A mitochondrion How do cells accomplish all their functions in such a tiny, crowded package? Eukaryotic cells those that make up cattails and apple trees, mushrooms and dust
Gene Control Mechanisms at Transcription and Translation Levels
 Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1
 and Ksp/Cre;Pkd1") Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
Review. Planar Polarity and Tissue Morphogenesis. Leading Edge
 Leading Edge Review Planar Polarity and Tissue Morphogenesis Jennifer A. Zallen 1, * 1 Developmental Biology Program, Sloan-Kettering Institute, New York, NY 10021, USA *Correspondence: zallenj@mskcc.org
Leading Edge Review Planar Polarity and Tissue Morphogenesis Jennifer A. Zallen 1, * 1 Developmental Biology Program, Sloan-Kettering Institute, New York, NY 10021, USA *Correspondence: zallenj@mskcc.org
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Sara Khraim. Shaymaa Alnamos ... Dr. Nafeth
 10 Sara Khraim Shaymaa Alnamos... Dr. Nafeth *Requirement of oxidative phosphorylation: 1- Source and target for electrons(nadh+fadh2 >> O2). 2- Electron carriers. 3- Enzymes, like oxidoreductases and
10 Sara Khraim Shaymaa Alnamos... Dr. Nafeth *Requirement of oxidative phosphorylation: 1- Source and target for electrons(nadh+fadh2 >> O2). 2- Electron carriers. 3- Enzymes, like oxidoreductases and
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Cellular Neuroanatomy I The Prototypical Neuron: Soma. Reading: BCP Chapter 2
 Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
Reading: Chapter 5, pp ; Reference chapter D, pp Problem set F
 Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
5.1 Cell Division and the Cell Cycle
 5.1 Cell Division and the Cell Cycle Lesson Objectives Contrast cell division in prokaryotes and eukaryotes. Identify the phases of the eukaryotic cell cycle. Explain how the cell cycle is controlled.
5.1 Cell Division and the Cell Cycle Lesson Objectives Contrast cell division in prokaryotes and eukaryotes. Identify the phases of the eukaryotic cell cycle. Explain how the cell cycle is controlled.
Center for Academic Services & Advising
 March 2, 2017 Biology I CSI Worksheet 6 1. List the four components of cellular respiration, where it occurs in the cell, and list major products consumed and produced in each step. i. Hint: Think about
March 2, 2017 Biology I CSI Worksheet 6 1. List the four components of cellular respiration, where it occurs in the cell, and list major products consumed and produced in each step. i. Hint: Think about
Cellular Respiration Stage 4: Electron Transport Chain
 Cellular Respiration Stage 4: Electron Transport Chain 2006-2007 Cellular respiration What s the point? The point is to make ATP! ATP 2006-2007 ATP accounting so far Glycolysis 2 ATP Kreb s cycle 2 ATP
Cellular Respiration Stage 4: Electron Transport Chain 2006-2007 Cellular respiration What s the point? The point is to make ATP! ATP 2006-2007 ATP accounting so far Glycolysis 2 ATP Kreb s cycle 2 ATP
2. Cellular and Molecular Biology
 2. Cellular and Molecular Biology 2.1 Cell Structure 2.2 Transport Across Cell Membranes 2.3 Cellular Metabolism 2.4 DNA Replication 2.5 Cell Division 2.6 Biosynthesis 2.1 Cell Structure What is a cell?
2. Cellular and Molecular Biology 2.1 Cell Structure 2.2 Transport Across Cell Membranes 2.3 Cellular Metabolism 2.4 DNA Replication 2.5 Cell Division 2.6 Biosynthesis 2.1 Cell Structure What is a cell?
Dachsous-Dependent Asymmetric Localization of Spiny-Legs Determines Planar Cell Polarity Orientation in Drosophila
 Cell Reports Article Dachsous-Dependent Asymmetric Localization of Spiny-Legs Determines Planar Cell Polarity Orientation in Drosophila Tomonori Ayukawa, 1,2,3 Masakazu Akiyama, 4 Jennifer L. Mummery-Widmer,
Cell Reports Article Dachsous-Dependent Asymmetric Localization of Spiny-Legs Determines Planar Cell Polarity Orientation in Drosophila Tomonori Ayukawa, 1,2,3 Masakazu Akiyama, 4 Jennifer L. Mummery-Widmer,
ΔG o' = ηf ΔΕ o' = (#e ( V mol) ΔΕ acceptor
 ΔΕ acceptor") Reading: Sec. 19.1 Electron-Transfer Reactions in Mitochondria (listed subsections only) 19.1.1 Electrons are Funneled to Universal Electron Acceptors p. 692/709 19.1.2 Electrons Pass through a Series
Reading: Sec. 19.1 Electron-Transfer Reactions in Mitochondria (listed subsections only) 19.1.1 Electrons are Funneled to Universal Electron Acceptors p. 692/709 19.1.2 Electrons Pass through a Series
Zool 3200: Cell Biology Exam 5 4/27/15
 Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use.
 Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use. This chapter was originally published in the book Invertebrate and Vertebrate Eye Development,
Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use. This chapter was originally published in the book Invertebrate and Vertebrate Eye Development,
Three Distinct Roles for Notch in Drosophila R7 Photoreceptor Specification
 Three Distinct Roles for Notch in Drosophila R7 Photoreceptor Specification Andrew Tomlinson 1. *, Yannis Emmanuel Mavromatakis 1., Gary Struhl 1,2 1 Department of Genetics and Development, College of
Three Distinct Roles for Notch in Drosophila R7 Photoreceptor Specification Andrew Tomlinson 1. *, Yannis Emmanuel Mavromatakis 1., Gary Struhl 1,2 1 Department of Genetics and Development, College of
Advanced Higher Biology. Unit 1- Cells and Proteins 2c) Membrane Proteins
 Membrane Proteins") Advanced Higher Biology Unit 1- Cells and Proteins 2c) Membrane Proteins Membrane Structure Phospholipid bilayer Transmembrane protein Integral protein Movement of Molecules Across Membranes Phospholipid
Advanced Higher Biology Unit 1- Cells and Proteins 2c) Membrane Proteins Membrane Structure Phospholipid bilayer Transmembrane protein Integral protein Movement of Molecules Across Membranes Phospholipid
An intracellular partitioning-based framework for tissue cell polarity in plants and animals
 HYPOTHESIS 2061 Development 140, 2061-2074 (2013) doi:10.1242/dev.062984 2013. Published by The ompany of iologists Ltd n intracellular partitioning-based framework for tissue cell polarity in plants and
HYPOTHESIS 2061 Development 140, 2061-2074 (2013) doi:10.1242/dev.062984 2013. Published by The ompany of iologists Ltd n intracellular partitioning-based framework for tissue cell polarity in plants and
Mitochondria Mitochondria were first seen by kollicker in 1850 in muscles and called them sarcosomes. Flemming (1882) described these organelles as
 described these organelles as") Mitochondria Mitochondria were first seen by kollicker in 1850 in muscles and called them sarcosomes. Flemming (1882) described these organelles as filia Altmann (1890) observed these structures and named
Mitochondria Mitochondria were first seen by kollicker in 1850 in muscles and called them sarcosomes. Flemming (1882) described these organelles as filia Altmann (1890) observed these structures and named
Gene regulation I Biochemistry 302. Bob Kelm February 25, 2005
 Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Nature Neuroscience: doi: /nn.2662
 Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
ADVANCED PLACEMENT BIOLOGY
 ADVANCED PLACEMENT BIOLOGY Description Advanced Placement Biology is designed to be the equivalent of a two-semester college introductory course for Biology majors. The course meets seven periods per week
ADVANCED PLACEMENT BIOLOGY Description Advanced Placement Biology is designed to be the equivalent of a two-semester college introductory course for Biology majors. The course meets seven periods per week
Bypass and interaction suppressors; pathway analysis
 Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of