Regulation of epithelial sodium channel (ENaC) activity by extracellular stimuli
|
|
|
- Conrad Dalton
- 6 years ago
- Views:
Transcription

 thesis, University of Iowa, 2011. http://ir.uiowa.edu/etd/4958. Follow this and additional works at: http://ir.uiowa.edu/etd Part of the Biophysics Commons")
1 University of Iowa Iowa Research Online Theses and Dissertations 2011 Regulation of epithelial sodium channel (ENaC) activity by extracellular stimuli Daniel Mohr Collier University of Iowa Copyright 2011 Daniel Mohr Collier This dissertation is available at Iowa Research Online: Recommended Citation Collier, Daniel Mohr. "Regulation of epithelial sodium channel (ENaC) activity by extracellular stimuli." PhD (Doctor of Philosophy) thesis, University of Iowa, Follow this and additional works at: Part of the Biophysics Commons
2 REGULATION OF EPITHELIAL SODIUM CHANNEL (ENaC) ACTIVITY BY EXTRACELLULAR STIMULI by Daniel Mohr Collier An Abstract Of a thesis submitted in partial fulfillment of the requirements for the Doctor of Philosophy degree in Molecular Physiology and Biophysics in the Graduate College of The University of Iowa December 2011 Thesis Supervisor: Professor Peter M. Snyder
3 1 ABSTRACT The epithelial sodium channel, ENaC, forms the rate-limiting step for sodium reabsorption in the cortical collecting duct of the kidney. It is known that ENaC is important in maintaining fluid homeostasis and ultimately blood pressure as mutations in ENaC result in inherited forms of hyper- and hypotension (Liddle s syndrome and Pseudohypoaldosteronism (PHA type I), respectively). Thus, understanding the function and regulation of ENaC activity may provide new insight into the pathogenesis of hypertension and assist in the development of more effective treatments. ENaC is a member of the DEG/ENaC family of ion channels. Each family member is composed of multiple subunits each subunit contains two transmembrane domains, short cytoplasmic amino and carboxy termini, and a relatively large extracellular domain. ENaC is a heterotrimer of homologous subunits α-, β-, and γenac. ENaC is a constitutively active ion channel. It is not ligand gated or voltage activated. However, channel activity can be modulated by a variety of stimuli. I hypothesize that the extracellular domain functions as a sensor, allowing the channel to detect and respond to changes in extracellular conditions. To test this, we expressed human αβγenac in Xenopus oocytes and used the twoelectrode voltage clamp technique to measure changes in ENaC activity in response to changing extracellular conditions. Using this technique, I identified several novel means of regulating ENaC activity. I found that ENaC activity can be rapidly and reversibly stimulated or suppressed in response to extracellular acidification and have identified several key residues involved. I found that extracellular chloride inhibits ENaC activity through putative binding sites in the extracellular domain located at interfaces between
4 2 the α- and β-, and β- and γenac subunits. This allowed us to determine that ENaC adopts an αγβ channel architecture. Additionally, I have made progress in understanding channel movement by identifying length dependent intersubunit interactions that alter channel gating. Based on our data we conclude that the extracellular domain is integral to modulation of channel activity. The work described herein has significantly advanced the field by improving our understanding of ENaC structure and function. Abstract Approved: Thesis Supervisor Title and Department Date
5 REGULATION OF EPITHELIAL SODIUM CHANNEL (ENaC) ACTIVITY BY EXTRACELLULAR STIMULI by Daniel Mohr Collier A thesis submitted in partial fulfillment of the requirements for the Doctor of Philosophy degree in Molecular Physiology and Biophysics in the Graduate College of The University of Iowa December 2011 Thesis Supervisor: Professor Peter M. Snyder
6 Copyright by DANIEL MOHR COLLIER 2011 All Rights Reserved
7 Graduate College The University of Iowa Iowa City, Iowa CERTIFICATE OF APPROVAL PH.D. THESIS This is to certify that the Ph.D. thesis of Daniel Mohr Collier has been approved by the Examining Committee for the thesis requirement for the Doctor of Philosophy degree in Molecular Physiology and Biophysics at the December 2011 graduation. Thesis Committee: Peter M. Snyder, Thesis Supervisor Nikolai Artemyev Wayne Johnson Amy Lee DP Mohapatra Michael Welsh
8 To my friends, family, lab mates, and science teachers for tolerating and encouraging my inquisitive nature. ii
9 to learn is not to know; there are the learners and the learned. Memory makes the one, philosophy the other. Alexandre Dumas The Count of Monte Cristo iii
10 ACKNOWLEDGMENTS We thank all the current and former members of the Snyder lab, especially Diane Olson, Zeru Peterson, Caitlin Digman, Abigail Hamilton, Danielle Wentzlaf, and Kaela Kramer for assistance. We would also like to thank Dr. Chris Benson, Dr. Mamta Gautaum, and Anne Harding for their feedback and encouragement. We acknowledge the University of Iowa In Vitro Models and Cell Culture Core for providing primary airway cultures, and the University of Iowa DNA Core Facility for reagents and DNA sequencing. This work was supported by National Institutes of Health Grant HL to Dr. Peter M. Snyder. Daniel M. Collier was supported by a pre-doctoral fellowship grant from the American Heart Association from January 2010 through December iv
11 ABSTRACT The epithelial sodium channel, ENaC, forms the rate-limiting step for sodium reabsorption in the cortical collecting duct of the kidney. It is known that ENaC is important in maintaining fluid homeostasis and ultimately blood pressure as mutations in ENaC result in inherited forms of hyper- and hypotension (Liddle s syndrome and Pseudohypoaldosteronism (PHA type I), respectively). Thus, understanding the function and regulation of ENaC activity may provide new insight into the pathogenesis of hypertension and assist in the development of more effective treatments. ENaC is a member of the DEG/ENaC family of ion channels. Each family member is composed of multiple subunits each subunit contains two transmembrane domains, short cytoplasmic amino and carboxy termini, and a relatively large extracellular domain. ENaC is a heterotrimer of homologous subunits α-, β-, and γenac. ENaC is a constitutively active ion channel. It is not ligand gated or voltage activated. However, channel activity can be modulated by a variety of stimuli. I hypothesize that the extracellular domain functions as a sensor, allowing the channel to detect and respond to changes in extracellular conditions. To test this, we expressed human αβγenac in Xenopus oocytes and used the twoelectrode voltage clamp technique to measure changes in ENaC activity in response to changing extracellular conditions. Using this technique, I identified several novel means of regulating ENaC activity. I found that ENaC activity can be rapidly and reversibly stimulated or suppressed in response to extracellular acidification and have identified several key residues involved. I found that extracellular chloride inhibits ENaC activity through putative binding sites in the extracellular domain located at interfaces between v
12 the α- and β-, and β- and γenac subunits. This allowed us to determine that ENaC adopts an αγβ channel architecture. Additionally, I have made progress in understanding channel movement by identifying length dependent intersubunit interactions that alter channel gating. Based on our data we conclude that the extracellular domain is integral to modulation of channel activity. The work described herein has significantly advanced the field by improving our understanding of ENaC structure and function. vi
13 TABLE OF CONTENTS LIST OF FIGURES...x CHAPTER I. INTRODUCTION...1 ENaC: Physiology, Expression, and Biophysical Properties...1 Significance...1 ENaC and Renal Physiology...2 ENaC in the Lingual Epithelium...2 Biophysical Properties...3 Regulation of ENaC Expression...4 Hormonal Regulation...4 Degradation...5 Na + Feedback Inhibition...6 Regulation of ENaC Gating...7 Proteolysis...7 Na + Self-inhibition...8 Stoichiometry and Channel Architecture...9 II. EXTRACELLULAR PROTONS REGULATE HUMAN ENAC BY MODULATING NA + SELF-INHIBITION...16 Preface...16 Abstract...16 Introduction...17 Experimental Procedures...20 Results...22 ph Changes Alter ENaC Current...22 Species-Specificity of ph Regulation...23 ph Regulates ENaC Gating...23 Extracellular ph modulates Na + self-inhibition...24 Regulation by Extracellular ph Requires Sodium...25 Mutations that Alter Na + Self-inhibition also Affect ph Regulation...26 Discussion...26 III. EXTRACELLULAR CL - REGULATES THE EPITHELIAL SODIUM CHANNEL...44 Preface...44 Abstract...44 Introduction...45 Experimental Procedures...47 Results...49 Extracellular Anions Modulate ENaC Current...49 Cl - Inhibits ENaC in Epithelia...50 Identification of Residues that Participate in Cl - Regulation of ENaC...51 DEG mutation and proteolytic cleavage abolish response to Cl Cl - Alters Na + Self-inhibition...52 ph Modulates the Effect of Cl - on ENaC...54 vii
14 Discussion...55 IV. IDENTIFICATION OF ENAC INTER-SUBUNIT CL - INHIBITORY RESIDUES SUGGESTS A TRIMERIC αγβ CHANNEL ARCHITECTURE...71 Preface...71 Abstract...71 Introduction...72 Experimental Procedures...74 Results...76 ENaC palm domain residues participate in Cl - inhibition...76 Mutations of γenac Met-299 alter anion inhibition selectivity...77 ENaC subunits are arranged in an αγβ orientation...78 Mutations of residues in separate Cl - sites are additive...79 Different Cl - sites alter ENaC activity through distinct mechanisms...80 Discussion...82 V. IDENTIFICATION OF RESIDUES IN THE EXTRACELLULAR DOMAIN OF HUMAN ENAC THAT CONTRIBUTE TO H + REGULATION OF CHANNEL ACTIVITY PROVIDE INSIGHT INTO CONFORMATIONAL CHANGES ASSOCIATED WITH CHANNEL GATING...98 Preface...98 Abstract...98 Introduction Experimental Procedures Results Human and rat ENaC respond differently to cahnges in extracellular ph in the absence of Cl Human γenac is necessary but not sufficient for ph sensitivity Mutation of acidic residues in the extracellular domain of human γenac decrease the ph response Mutation of multiple acidic residues in the extracellular domain of human γenac specifically decreases the response to acidic ph Residues equivalent ot γe455 in α- and βenac also modulate the ph response Mutation of acidic residues in β- and γenac specifically eliminate the response to acidic ph Disruption of intersubunit interfaces modulates ENaC activity Functional effect of cross-linking opposing subunits Discussion VI. SUMMARY AND CONCLUSIONS ENaC activity is regulated by extracellular ph ENaC activity is regulated by extracellular Cl ENaC subunit architecture Conformational changes associated with ph sensing and channel gating REFERENCES viii
15 LIST OF FIGURES Figure 1.1. Sequence alignment of human α-, β-, and γenac Structure predicts function Extracellular protons modulate human ENaC current Extracellular protons modulate Na + transport in epithelia Species-specificity of ph regulation ph regulates ENaC gating Extracellular ph modulates Na + self-inhibition Regulation by extracellular ph requires Na Mutations that alter Na + self-inhibition affect ph regulation Extracellular anions modulate ENaC activity Extracellular Cl - modulates endogenous ENaC activity in epithelia Identification of residues that participate in Cl - regulation of ENaC DEG mutation and proteolytic cleavage abolish response to Cl Cl - alters Na + self-inhibition ph modulates the effect of Cl - on ENaC Sequence alignment of ASIC1a with α-, β- and γenac Effect of ENaC palm domain mutations on Cl - inhibition γenac Met-299 mutations alter selectivity of anion inhibition Potential ENaC subunit arrangements Additivity of Cl- inhibitory site mutations Differential effects of Cl - site mutations on Na + self-inhibition Human and rat ENaC respond differently to changes in extracellular ph Human γenac is necessary for ph regulation of channel activity Human γenac is not sufficient for ph regulation of channel activity Acidic residues differ in the extracellular domain of human and rat γenac ix
16 5.5 Mutation of titratable residues in the extracellular domain of γenac changes the ph response Mutation of multiple residues in γenac results in an additive decrease in the response to acidic ph Homology modeling with ASIC1a crystal structure suggests γ E455 may lie at an intersubunit interface Residues equivalent to γ E455 in α- and βenac also modulate the ph response Mutation of titratable residues in β- and γenac specifically eliminate the response to acidic ph Mutation of titratable residues in β- and γenac reduce ph sensitivity of Na + self-inhibition Electrostatic effects can be induced between interfacing residues Cross-linking reagents specifically alter α K477C β V85C γenac function in a size dependent manner Cross-linking αk477c and βv85c decreases the ph response Model of cross-linking effects x
17 1 CHAPTER I INTRODUCTION In this introductory chapter, we will begin by highlighting the physiological significance of the epithelial sodium channel (ENaC) as well as the basic biophysical properties of the channel. We will then turn our attention to the two major themes of ENaC regulation: regulation of ENaC expression and regulation of ENaC gating. Finally, we will examine potential structural similarities between ENaC and ion channels that have had structures solved by x-ray crystallography. ENaC: Physiology, Expression, and Biophysical Properties Significance The epithelial sodium channel, ENaC, is found in a variety of tight epithelia including, but not limited to, the distal nephron, airway, and distal colon (1, 2). In these tissues, ENaC forms the rate-limiting step for Na + reabsorption. Because of this, ENaC is critical in maintaining fluid homeostasis. Perturbation of ENaC activity plays a role in human diseases, such as hypertension (3). Hypertension is a common disease and the leading cause of stroke and heart attack. Recent data shows that approximately 29% of the adult population suffers from hypertension, as defined by systolic BP of 140 mm Hg or higher with diastolic BP 90 mm Hg or higher, with prevalence increasing by % over the previous ten year period (4-6). In only a small minority of patients is the cause of hypertension known. In 95% of hypertensive patients, the cause is unknown ( essential hypertension ). Thus, treatments are seldom targeted to the underlying pathogenesis, and as a result, only a minority of patients reach treatment goals. Understanding the basic molecular causes of hypertension
18 2 will lead to more targeted strategies to treat and prevent hypertension, which will reduce the burden of cardiovascular disease. ENaC and Renal Physiology Blood pressure can be maintained by controlling vascular tone or by regulating extracellular fluid volume. Our research focuses on understanding how extracellular volume is regulated by Na + transport in the distal nephron of the kidney. The kidney is critical in maintaining blood pressure. This is accomplished by maintaining the balance of sodium absorption versus sodium excretion. The epithelial Na + channel is central to the kidney s ability to regulate Na + reabsorption and to control fluid homeostasis. Patients with excessive ENaC activity, such as those with Liddle s syndrome, emphasize the critical role of ENaC in regulating blood pressure. Most of the known genetic forms of hypertension are caused by defects in ENaC regulation (3). These observations raise the possibility that defects in ENaC regulation may play an important role in the pathogenesis of essential hypertension. ENaC in the Lingual Epithelium It is clear that ENaC is critical in maintaining fluid quantity and composition in the kidney, however, a new role is emerging for ENaC activity in salt taste. The behavioral response to salt appears to separate into two opposing responses. At high sodium levels (>500 mm), the behavioral response is one of aversion, while lower sodium levels (<100 mm), especially after salt depletion, are desirable. This attractive sodium response is amiloride sensitive in mice. Furthermore, the amiloride sensitive
19 3 component of salt taste is lost in taste receptor cell specific ENaC knock out mice, while the aversive salt response remains normal (7). This suggests the possibility that ENaC may play a two-fold role in mediating sodium transport and fluid homeostasis by regulating both sodium intake and sodium excretion. It also provides a system for testing ENaC mediated physiological responses to changing extracellular conditions which cannot be achieved in the renal tubules but could be easily be manipulated on the lingual epithelium. Biophysical Properties The epithelial sodium channel (ENaC) is a member of the Degenerin/ENaC family of ion channels (8). ENaC is composed of homologous subunits termed α, β, and γ. The β- and γenac subunits share approximately 33-34% amino acid identity with αenac (2). Each subunit contains two transmembrane domains, relatively short cytoplasmic amino (~8-13% of primary structure) and carboxy termini (~15-24% of primary structure), and a large extracellular domain (~61-68% of primary structure) (Fig. 1.1 and Refs (1, 2). ENaC is expressed in a variety of epithelia where its predominant physiological role is to provide a pathway for Na + reabsorption in order to maintain fluid homeostasis. Canonical channel function can be studied in cells or tissues that endogenously express α-, β- and γenac subunits, or in heterologous cells that have been transfected with equal amounts of α-, β- and γenac. Some channel function can be measured in cells expressing αβ-, αγ- or only αenac, although, the function is ~20 fold less than that of cells expressing all three subunits (2). αβγenac is a constitutively active Na + channel
20 4 with single channel Na + conductance of 5 ps (Li + = 9 ps)(9). It is highly selective for Na + over K + (10) and ENaC mediated Na + current is eliminated by application of a diuretic, amiloride, which blocks the channel pore (11, 12). ENaC is not ligand gated or voltage sensitive and does not exhibit inward or outward rectification. However, channel activity can be modulated by a variety of extracellular stimuli. Some stimuli, such as Na +, exert reversible inhibitory effects, while others, such as proteolysis of the extracellular domain of α- and γenac, irreversibly increase ENaC activity (13). Regulation of ENaC Expression ENaC activity can be regulated in two ways; by changing channel expression or by changing channel gating. Changes in channel expression are typically seen over a time course of several minutes to hours. Changes in channel gating are comparatively immediate. Both mechanisms are import in regulating ENaC activity. We will initially focus on the regulation of channel expression through hormonal regulation, protein degradation, and Na + feedback inhibition. Hormonal Regulation ENaC protein expression can be modulated hormonally by altering transcription or by post-translational mechanisms. ENaC is sensitive to hormonal regulation by the mineralocorticoid receptor (MR) hormone, aldosterone, and antidiuretic hormone (ADH). The effects of aldosterone are two-fold. First, in the distal nephron, aldosterone regulates ENaC transcription. Aldosterone binds to basolateral mineralocorticoid receptors. The
21 5 complex is then internalized and translocated to the nucleus where aldosterone can bind transcription factors and induce ENaC transcription (13). This ultimately leads to higher ENaC protein expression at the apical membrane and increased Na + transport. Second, aldosterone has post-translational effects on ENaC expression. Aldosterone enhances transcription of serum and glucocorticoid-induced kinase (SGK). SGK binds to and phosphorylates Nedd 4-2, an E3 ubiquitin ligase that ubiquitinates ENaC, targeting it for degradation (described in next section). Phosphorylation of Nedd 4-2 disrupts its ability to bind and ubiquitinate ENaC. This also results increased Na + transport by decreasing ubiquitination and degradation of ENaC (Reviewed in (14)). Antidiuretic hormone (ADH), also known as vasopressin, has a post-translational effect on ENaC similar to that of aldosterone. ADH binds to V2 receptors at the basolateral membrane of renal epithelia. This activates adenylate cyclase, resulting in an increase in camp and activation of protein kinase A (PKA). PKA phosphorylates the same sites on Nedd 4-2 as SGK (Reviewed in (14)) - thus decreasing ENaC degradation and increasing Na + reabsorption. Degradation To balance the hormonally regulated increase in ENaC expression, ENaC endocytosis and degradation is also tightly controlled. There are a variety of factors that influence ENaC degradation. One of the best characterized is the involvement of the carboxy termini of β-, and γenac. The intracellular carboxy termini of β- and γenac contain PY motifs (15, 16) that are important in regulating ENaC surface expression. PY motifs are recognized by WW domains (17) of ubiquitin ligase proteins, such as Nedd 4-
22 6 2, an E3 ubiquitin ligase from the HECT family. Nedd 4-2 binds to ENaC and attaches a ubiquitin molecule. This targets the channel for internalization and degradation. Disruption of the carboxy terminal PY motifs of β- or γenac results in increased ENaC surface expression (18). Liddle s syndrome, an autosomal dominant form of hypertension, is the result of missense mutations or deletions in the carboxy termini of β- or γenac that disrupt or remove the PY motif. This results in aberrant ENaC cell surface expression by preventing internalization and degradation (19). This increases the amount of time ENaC resides at the cell surface, leading to increased Na + transport and ultimately severe hypertension in humans who carry the mutation in one allele. Na + Feedback Inhibition ENaC expression at the cell surface is also subject to regulation by a negative feedback loop termed Na + feedback inhibition. The mechanism of action is complex and multifactorial. First, increasing intracellular Na + decreases ENaC cell surface expression (20-24). In some reports, this is a biphasic event with fast and slow inhibitory components (22). While it is clear that Na + is necessary to trigger this event, it is also dependent on additional intracellular factors that will prevent Na + feedback inhibition if removed (23). The precise identity of the residue(s) or domains of ENaC responsible for Na + feedback inhibition are not yet known. Another layer of complexity is added because ENaC is present at the cell surface in cleaved and uncleaved states (described below). In addition to simply modulating the number of channels at the cell surface, intracellular Na + also regulates ENaC activity by
23 7 modulating proteolysis. ENaC is present at the cell surface in two pools, a fully active, cleaved state, and a lower activity, partially or uncleaved, state. When intracellular Na + is low, the majority of ENaC appears at the cell surface in the cleaved state (25). This presumably allows for maximum Na + transport when the system is challenged by low extracellular Na +. As intracellular Na + concentration increases, there is a corresponding decrease in the amount of cleaved ENaC at the cell surface (25). This provides another layer of regulation in response to intracellular Na + by regulating channel number and the balance between high activity and low activity channels. Regulation of ENaC Gating Regulation of ENaC surface expression acts as the coarse adjustment, while the fine-tuning of ENaC activity happens dynamically in response to a continuously changing extracellular environment. Extracellular stimuli modulate ENaC activity by changing channel gating properties. This is observed as a change in channel open probability (P o ) - the likelihood that a channel will be open during any given period of time. ENaC P o can be modulated by proteolysis and changes in extracellular Na + concentration. Proteolysis ENaC activity is irreversibly increased by proteolysis of the extracellular domain of α- and γenac. α- and γenac contain furin consensus cleavage sites (Fig. 1.1 and Ref. (26)). Furin, a member of the proprotein convertase family of serine proteases which localizes to the trans-golgi network, cleaves immediately after the consensus
24 8 sequence of R-X-X-R (X can be any residue, (27)). Channels can also be cleaved at the cell surface by treatment with extracellular proteases such as trypsin (28). Cleavage increases ENaC P o by approximately 20 fold (29). Mutation of these cleavage sites in α- or γenac markedly reduces ENaC activity by decreasing channel open probability (26), suggesting that cleavage is an important part of channel maturation in order to achieve normal activity. Activity of mutant channels can be restored by exogenous treatment with trypsin, which cleaves ENaC near the furin consensus sites (30). The mechanism by which cleavage activates ENaC is thought to be due to the liberation of an inhibitory peptide from extracellular domain of the α- and γenac subunits (31-34). Na + Self-inhibition In addition to the effects of intracellular Na +, increasing extracellular Na + concentration inhibits ENaC activity by altering channel gating. This response is termed Na + self-inhibition. Originally described in 1977, this process is still not completely understood (35). Na + self-inhibition is observed by rapidly changing extracellular sodium from low (1 mm) to high (120 mm) concentrations, resulting in a peak inward Na + current that relaxes to a lower steady state value over a time course of a few seconds (35-37). Na + self-inhibition is ablated by ENaC cleavage by exogenous protease treatment (36). While the location the extracellular Na + binding site is not known, it has been shown that Na + self-inhibition is not voltage sensitive, suggesting that it is not located within the transmembrane domain. Furthermore, the inhibitory effect is selective for Na + over Li + and K +, while the ion-conducting site is selective for Li + over Na + and K + (38).
25 9 Na + self-inhibition alters ENaC activity by changing the channel open probability (P o ) - high Na + leads to lower P o. Na + self-inhibition is emerging as a common pathway for modulation of ENaC activity in response to changes in temperature and extracellular divalent cations such as Zn 2+, Ni 2+, and Cu 2+ (37, 39-41). In Chapters II-IV, we will describe how extracellular protons and Cl - mediate ENaC activity, in part, by modulating Na + self-inhibition. Stoichiometry and Channel Architecture The recent crystallization of a closely related DEG/ENaC family member, acid sensing ion channel 1 (ASIC1), has provided excellent clues into ENaC structure and function. While the two proteins differ in some regions of the extracellular domain, structural comparisons are made possible by thirteen cysteine residues in the extracellular domain that are conserved across all DEG/ENaC family members (42, 43). The most notable functional difference between ASIC and ENaC is that ASIC channels are ligand gated while ENaC is constitutively active. Despite its constitutive activity, however, both channels are highly cation selective and share a large, highly structured, extracellular domain. This, in conjunction with work demonstrating that extracellular factors regulate ENaC activity, suggests that the extracellular domain must have retained some ability to influence channel gating, albeit not in an all-or-none fashion as seen in ASIC. Does structure predict function? Figure 1.2 shows representative ion channels that have been crystalized. Non-selective cation channels (NaK) are composed, essentially, of only transmembrane, pore forming helices (44). Inward rectifying K +
26 10 channels (Kir) have intracellular domains that allow for initiation of gating by membrane and intracellular signals (45). In contrast, channels gated by extracellular ligands, such as proton gated Na + channels (ASIC), ATP gated cation channels (P2X), and glutamate gated Cl - channels (GluCl), have large, complex, extracellular domains (43, 46-48). The similarity between ENaC and ASIC provides structural evidence that ENaC is likely a sensor of the extracellular millue. However, the identity of the residues and the mechanisms behind ENaC s response to extracellular stimuli is poorly understood. This is the focus of much of the work presented herein. Crystallization of ASIC also provided insight into the stoichiometry of ENaC and other DEG/ENaC channels. ASIC1a assembles as a trimer. Normal ENaC channels are composed of α-, β-, and γenac subunits, although, the number of each subunit needed to form a functional channel has been debated. Early reports suggested ENaC may assemble as a tetramer, with α 2 βγ stoichiometry or as a nonamer with α 3 β 3 γ 3 stoichiometry (49, 50). Recent atomic force microscopy suggests that ENaC assembles as a heterotrimer of αβγ, with some channels aggregating into hexamers and nonamers (51). The atomic force data, however, cannot determine how the three ENaC subunits assemble into a complex. We address this open question in Chapter IV and V. The goal of the work included herein is to further the field s knowledge of ENaC structure and function while focusing primarily on the regulation of ENaC activity by extracellular stimuli. Figure 1.1 shows the location of the residues of interest discussed throughout this work. First, I will describe how extracellular protons activate ENaC by relieving Na + self-inhibition (Chapter II). This work lead to the observation that extracellular Cl - inhibits ENaC activity (Chapter III). We were then able to take
27 11 advantage of the ASIC crystal structure and the putative Cl - binding site to determine ENaC subunit architecture (Chapter IV). Finally, I will describe the residues involved in activation of ENaC activity in response to extracellular protons and their potential role in extracellular conformational changes in association with channel gating (Chapter V).
28 12 Figure 1.1. Sequence alignment of human α-, β-, and γenac. Amino acid sequence alignment of human α-, β-, and γenac. Key features of the proteins are noted. Scissor icons indicate Furin and CAP1 cleavage sites in α- and γenac (none are present in βenac). Open boxes below the sequence indicate the predicted position of the first (TM1) and second (TM2) transmembrane spanning domains. The location of the DEG position (DEG) and selective filter (SEL) residues are indicated in bold in the second transmembrane domain (TM2). Residues of interest discussed throughout the text are highlighted (gold: ph sensing residues discussed in Chapter V, orange: inter-subunit interface ph sensing residues discussed in Chapter V, green: putative inter-subunit Cl - binding site residues discussed in Chapters III and IV, blue: histidine residues that modulate Na + self-inhibition discussed in Chapter II).
29 13 h. ENaC h. ENaC h. ENaC 10 MEGNKLEEQDSSPPQSTPGLMKGNKREEQGLGPEPAAPQQPTAEEEALIEFHRSYRELFEFFCNNTTIHGAIRLVCSQHN MHVKKYLLKGLHRLQKGPGY TYKELLVWYCDNTNTHGPKRIICE--G MAPGEKIKAKIKKNLPVTGPQAP TIKELMRWYCLNTNTHGCRRIVVSR-G 49 h. ENaC h. ENaC h. ENaC h. ENaC h. ENaC h. ENaC RMKTAFWAVLWLCTFGMMYWQFGLLFGEYFSYPVSLNINLNSDKLVFPAVTICTLNPYRYPEIKEELEELDRITEQTLFD 160 PKKKAMWFLLTLLFAALVCWQWGIFIRTYLSWEVSVSLSVGFKTMDFPAVTICNASPFKYSKIKHLLKDLDELMEAVLER 125 RLRRLLWIGFTLTAVALILWQCALLVFSFY--TVSVSIKVHFRKLDFPAVTICNINPYKYSTVRHLLADLEQETREALKS 127 LYKY SSFTTLVAGSRSRRDLRGTLPHPLQRLRVPPPPHGARRARSVASSLRDNNPQVDWKDWKIGFQL 228 ILAP ELSHANATRNLNFSIWNHTPLVLIDERNPHHPMVLDLFGDNHN--G-LTSSSASEKICNAHGCKMAMRL 195 LYGFPESRKRREAESWNSVSEGKQPRFSHRIPLLIFDQD-EKGKARDFFTGRKRKVGGSIIHKASNVMHIESKQVVGFQL 206 Furin Furin CAP1 Furin h. ENaC h. ENaC h. ENaC CNQNKSDCFYQTYSSGVDAVREWYRFHYINILSRLPETLPSLEEDTLGNFIFACRFNQVSCNQANYSHFHHPMYGNCYTF 308 CSLNRTQCTFRNFTSATQALTEWYILQATNIFAQVPQQELVEMSYPGEQMILACLFGAEPCNYRNFTSIFYPHYGNCYIF 275 CSNDTSDCATYTFSSGINAIQEWYKLHYMNIMAQVPLEKKINMSYSAEELLVTCFFDGVSCDARNFTLFHHPMHGNCYTF 286 h. ENaC h. ENaC h. ENaC NDKNNSNLWMSSMPGINNGLSLMLRAEQNDFIPLLSTVTGARVMVHGQDEPAFMDDGGFNLRPGVETSISMRKETLDRLG 388 NWGMTEKALPSANPGTEFGLKLILDIGQEDYVPFLASTAGVRLMLHEQRSYPFIRDEGIYAMSGTETSIGVLVDKLQRMG 355 NNRENETILSTSMGGSEYGLQVILYINEEEYNPFLVSSTGAKVIIHRQDEYPFVEDVGTEIETAMVTSIGMHLTESFKLS 366 h. ENaC h. ENaC h. ENaC GDYGDCTKNGSDVPVENLYP---SKYTQQVCIHSCFQESMIKECGCAYIFYPRPQNVEYCDYRKHSSWGYCYYKLQVDFS 465 EPYSPCTVNGSEVPVQNFYSDYNTTYSIQACLRSCFQDHMIRNCNCGHYLYPLPRGEKYCNNRDFPDWAHCYSDLQMSVA 435 EPYSQCTEDGSDVPIRNIY---NAAYSLQICLHSCFQTKMVEKCGCAQYSQPLPPAANYCNYQQHPNWMYCYYQLHRAFV 443 h. ENaC h. ENaC h. ENaC SDHLGCFTKCRKPCSVTSYQLSAGYSRWPSVTSQEWVFQMLSRQNNYTVNNKRN--GVAKVNIFFKELNYKTNSESPSVT 543 QRET-CIGMCKESCNDTQYKMTISMADWPSEASEDWIFHVLSQERDQSTNITLSRKGIVKLNIYFQEFNYRTIEESAANN 514 QEELGCQSVCKEACSFKEWTLTTSLAQWPSVVSEKWLLPVLTWDQGRQVNKKLNKTDLAKLLIFYKDLNQRSIMESPANS 523 h. ENaC h. ENaC h. ENaC MVTLLSNLGSQWSLWFGSSVLSVVEMAELVFDLLVIMFLMLLRR---FRSRYWSPGRGGRGAQEVASTLASSPPSHFCP- 619 IVWLLSNLGGQFGFWMGGSVLCLIEFGEIIIDFVWITIIKLVALAKSLRQRRAQASYAGPP-PTVAELVEAHTNFGFQPD 593 IEMLLSNFGGQLGLWMSCSVVCVIEI----IEVFFIDFFSIIARRQWQKAKEWWAWKQAPPCPEAPRSPQGQDNPALDID 599 (DEG) (SEL) h. ENaC h. ENaC h. ENaC -HPMSLSLSQ--PGPAPSPALTAPPPAYATLGPRPSPGGSAGASSSACPLGGP. TAPRSPNTGP-YPSEQALPIPGTPPPNYDSL--RLQ---PLDVIESDSEGDAI. DDLPTFNSALHLPPSLGTQVPGTPPPKYNTL--RLERA-FSNQLTDTQMLDEL. ph residues (Chap V) Inter-subunit ph residues (Chap V) Inter-subunit Cl - residues (Chap III and IV) Na + self-inhibition modulating residues (Chap II)
30 14 Figure 1.2. Structure predicts function. Crystal structures of a variety of ion channels. Horizontal lines indicate the approximate location of plasma membrane. Structures are scaled according to their transmembrane regions. From left to right: NaK (PDB ID: 2AHY) nonselective cation channel, Kir (1P7B) inward rectifying K + channel, ASIC1 (3HGC) extracellular proton gated cation channel (a DEG/ENaC family member), P2X 4 (3H9V) cationselective channels gated by extracellular ATP, GluCl (3RWH) extracellular glutamategated inhibitory anion-selective Cys-loop receptor in complex with Fab.

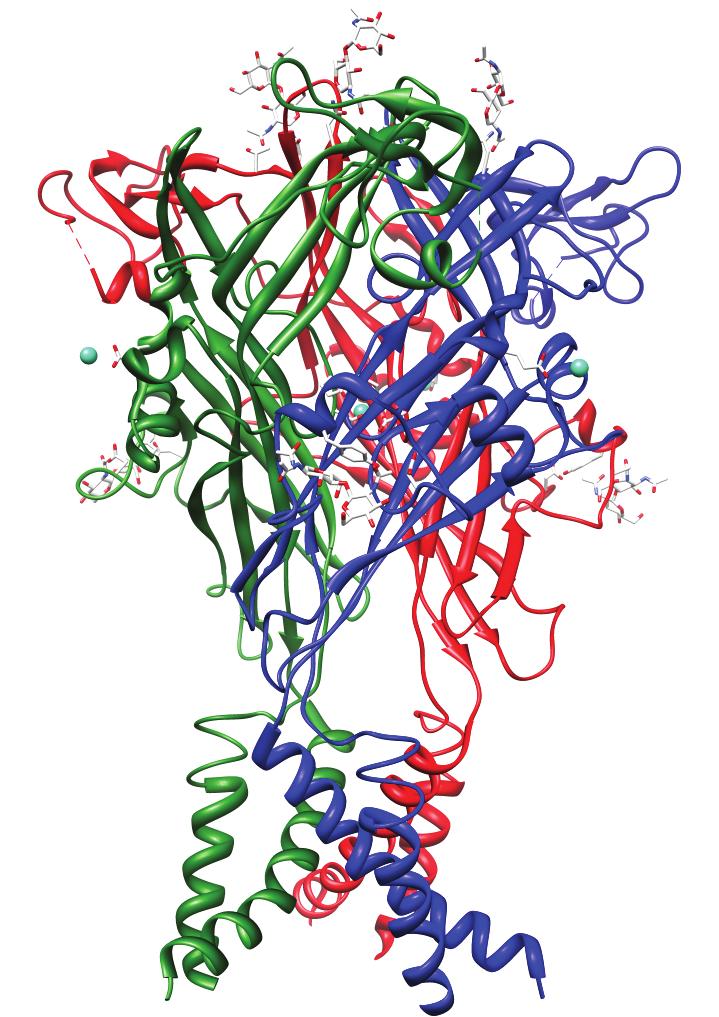


31 15 Extracellular Intracellular NaK Kir ASIC1 (DEG/ENaC) P2X4 GluCl
32 16 CHAPTER II EXTRACELLULAR PROTONS REGULATE HUMAN ENAC BY MODULATING NA + SELF-INHIBITION Preface This work appeared in the Journal of Biological Chemistry on November 6 th, I am co-author on this work with Dr. Snyder. I designed and performed the experiments, analyzed the data, and wrote the manuscript. Dr. Snyder designed experiments and wrote the manuscript. This work describes a novel means of regulating ENaC activity in response changes in extracellular ph. These experiments bring to light striking functional differences between ENaC cloned from different species and also, serendipitously, lead to the discovery that ENaC activity is regulated by changes in extracellular chloride (discussed in Chapters III and IV). Abstract The epithelial Na + channel, ENaC, is exposed to a wide range of proton concentrations in the kidney, lung, and sweat duct. We therefore tested whether ph alters ENaC activity. In Xenopus oocytes expressing human α, β, and γenac, amiloride sensitive current was altered by protons in the physiologically relevant range (ph ). Compared to ph 7.4, acidic ph increased ENaC current whereas alkaline ph decreased current (ph 50 = 7.2). Acidic ph also increased ENaC current in H441 epithelia and in human primary airway epithelia. In contrast to human ENaC, ph did not alter rat ENaC current, indicating that there are species differences in ENaC regulation by protons. This resulted predominantly from species differences in γenac. Maneuvers
33 17 that lock ENaC in a high open-probability state ( Deg mutation, proteolytic cleavage) abolished the effect of ph on human ENaC, indicating that protons alter ENaC current by modulating channel gating. Previous work showed that ENaC gating is regulated in part by extracellular Na + ( Na + self-inhibition ). Based on several observations, we conclude that protons regulate ENaC by altering Na + self-inhibition. First, protons reduced Na + self-inhibition in a dose-dependent manner. Second, ENaC regulation by ph was abolished by removing Na + from the extracellular bathing solution. Third, mutations that alter Na + self-inhibition produced corresponding changes in ENaC regulation by ph. Together, the data support a model in which protons modulate ENaC gating by relieving Na + self-inhibition. We speculate that this may be an important mechanism to facilitate epithelial Na + transport under conditions of acidosis. Introduction The epithelial Na + channel, composed of three homologous subunits (α, β, and γenac), functions as a pathway for Na + reabsorption across epithelia in the kidney collecting duct, lung, distal colon, and sweat duct (reviewed in (52, 53)). In this role, the channel is critical for the maintenance of Na + homeostasis and to control the composition and quantity of the fluid on the apical membrane of these epithelia. ENaC mutations, and defects in its regulation, cause inherited forms of hypertension and hypotension (3), and may contribute to the pathogenesis of lung disease in cystic fibrosis (54). As a member of the DEG/ENaC family of ion channels, ENaC shares common structural and functional features with channels that are gated by diverse stimuli. All DEG/ENaC subunits share a common overall structure, with two transmembrane
34 18 domains and relatively short cytoplasmic N and C termini, leaving the majority of the protein exposed as a large extracellular domain (55). Differences in these extracellular domains between DEG/ENaC family members result in dramatic functional diversity, from mechanosensitive ion channels in C. elegans (56) to ligand gated channels such as the peptide (FMRF-amide) gated FaNaCh channel in mollusks (57) and proton gated ASIC channels (58, 59). Although no analogous ligand has yet been identified for ENaC, it is clear that the extracellular domain is important in modulating ENaC activity. For example, proteolytic cleavage at two sites in the extracellular domains of α and γenac convert channels from an inactive to an active state (28, 29, 60). One population of channels is cleaved by furin in the Golgi complex (60). A second population reaches the cell surface in an uncleaved state and is susceptible to cleavage and activation by proteases at the cell surface and in the extracellular fluid (29, 61-63). Other molecules in the extracellular fluid also modulate ENaC gating. Extracellular Na + inhibits ENaC activity through a process called Na + self-inhibition, which serves as a negative feedback mechanism to regulate Na + transport (36, 39, 64). Two observations implicate a role for the extracellular domain in Na + self-inhibition. First, mutations of conserved histidine residues in the extracellular domains of α and γenac alter Na + self-inhibition (39). Second, proteolytic cleavage of the extracellular domain prevents Na + self-inhibition (65). Divalent cations including Zn 2+ and Ni 2+ are also thought to alter ENaC activity by binding to the extracellular domain (40, 66). These findings suggest that the extracellular domain might function as a sensor to allow a variety of signals in the extracellular environment to modulate ENaC activity. In
35 19 this regard, protons are a strong candidate to regulate ENaC. In its location at the apical membrane of epithelia, ENaC is exposed to extremes of ph. For instance, urine ph in the collecting duct can vary from in response to metabolic acidosis and alkalosis, as well as with changes in diet and volume status (67). Normal airway surface liquid ph is slightly alkaline (ph ) (68) but can become highly acidic (ph 4-6) with lung disease (69). Sweat ph can fall to < 6 when the production rate is low, but becomes neutral as production increases (70). Although ENaC appears poised to respond to changes in ph, previous studies investigating this possibility have been conflicting. External ph < 5 decreased shortcircuit current in frog skin (71) but increased Na + transport in toad bladder (72). In cultured Xenopus collecting duct cells (A6), acidification of the extracellular medium (ph 6.4 and 5.4) produced a slow increase in amiloride-sensitive current that peaked at ~ 30 min (73). More recent work investigated the role of external ph in regulation of rat α, β, and γenac expressed in Xenopus oocytes. These studies reported a very small transient increase (73), small decrease (ph 4.0) (74), or no change in current (75, 76). When α and βenac were coexpressed without γenac, acidic ph inhibited the channel (which has an open-probability close to 1.0) (75). Moreover, an additional subunit, δenac, was activated by acidic ph (74, 77), although this subunit is not expressed in renal epithelia. Together the data suggest that ph modulates epithelial Na + transport, but the magnitude and direction of the regulation may vary in different tissues and species. To further explore this possibility and to understand the mechanisms by which ph regulates Na + transport, we tested the effect of protons on the activity of human ENaC.
36 20 Experimental Procedures DNA Constructs cdnas for human and rat α, β, and γenac in pmt3 were cloned as previously described (1, 78). Mutations in αenac (H255R) and γenac (H233R) were generated by site-directed mutagenesis (QuickChange; Stratagene) and sequenced in the University of Iowa DNA Core. Expression and Whole-Cell Electrophysiology in Xenopus oocytes Oocytes were harvested from albino Xenopus laevis females and manually defolliculated following a one hour treatment with 0.75 mg/ml Type IV Collagenase (Sigma) in Ca 2+ free ND-96 (96 mm NaCl, 2 mm KCl, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH). Following nuclear injection of cdnas encoding human or rat α, β, and γenac (0.02 µg/µl each), cells were incubated at 18 C in modified Barth s saline (88 mm NaCl, 1 mm KCl, 0.33 mm Ca(NO 3 ) 2, 0.41 mm CaCl 2, 0.82 mm MgSO 4, 2.4 mm NaHCO 3, 10 mm HEPES, 50 µg/ml gentamycin sulfate, 10 µg/ml sodium penicillin, 10 µg/ml streptomycin sulfate, ph adjusted to 7.4 with NaOH) for hours prior to study. Oocytes were voltage clamped (two-electrode voltage clamp) and currents were amplified with an Oocyte Clamp OC-725C (Warner Instruments), digitized with a MacLab/200 interface (ADInstruments), and recorded and analyzed with Chart software (ADInstruments). Unless otherwise noted, recordings were done at 60 mv in a 116 mm NaCl solution (116 mm NaCl, 2 mm KCl, 0.4 mm CaCl 2, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH). Low Na + solutions (0 or 1 mm NaCl, 116 or 115 mm N-Methyl-D-glucamine Cl) were used as indicated in the figure legends. The ph of test solutions ( ) was adjusted with HCl or NaOH. Amiloride-sensitive current was determined by adding 10 µm amiloride to the bathing solution. The ph-induced change
37 21 in amiloride-sensitive current was calculated as the fold increase/decrease relative to the ph 7.4 baseline just prior to each test solution application. This was done to reduce the effect of time dependent current run-down. The resulting data were plotted and fit to the Hill equation using IGOR Pro software (WaveMetrics Inc.). Na + self-inhibition was measured by rapidly changing the bathing solution from low sodium (1 mm NaCl) to high sodium (116 mm NaCl) and quantitated as [(peak current (I P ) steady state current (I ss )) / peak current (I P )]. Expression and Whole-Cell Electrophysiology in H441 and primary airway epithelia H441 cells (American Type Culture Collection) were grown on 0.6 cm 2 permeable filter supports (Millipore) in RPMI with 8.5% fetal calf serum, 20 mm L- glutamine, 5 µg/ml insulin, 5 µg/ml transferrin, 5 ng/ml selenium, 100 nm dexamethasone, 100 U/ml penicillin, and 100 mg/ml streptomycin at 37 C for 5 days. Primary human airway epithelia were isolated from the trachea and bronchi of donor lungs and grown at the air-liquid interface of collagen-coated permeable filter supports, as described previously (79). Aprotinin (26 µg/ml) was present in the apical solution for 2 h prior to study. Short-circuit Na + current was measured in modified Ussing chambers (Warner Instrument Corporation) using an EC-825 Epithelial Voltage Clamp amplifier (Warner Instrument Corporation). Currents were digitized with a PowerLab interface (ADInstruments) and recorded and analyzed with Chart software (ADInstruments). The apical and basolateral surfaces were bathed in 116 mm NaCl, 2 mm KCl, 0.4 mm CaCl 2, 1 mm MgCl 2, 5 mm HEPES (ph 7.4 basolateral, 7.4 or 6.5 apical) at 37 C. Amiloridesensitive short-circuit current was determined as the difference in current with and without amiloride (10 µm) in the apical bathing solution.
38 22 Results ph Changes Alter ENaC Current To determine if changes in extracellular ph alter ENaC current, we expressed human α, β, and γenac in Xenopus oocytes and recorded Na + current at a holding potential of 60 mv. In Fig. 2.1A, we found that current was reduced when we shifted the extracellular ph from 7.4 to 8.5. Conversely, ph 6.5 increased current. The changes in current were rapidly reversible on returning to ph 7.4. When ENaC was blocked by amiloride, changes in ph did not alter current, indicating that ph regulated ENaC and not an endogenous oocyte current. Also consistent with this conclusion, ph changes from did not significantly alter current in uninjected oocytes (not shown). Fig. 2.1B shows the ph dose-response relationship for amiloride-sensitive ENaC current (relative to current at ph 7.4). Current was maximal at ph 6 (I 6 /I 7.4 = 1.4 ±.0148) and minimal at ph 8.5 (I 8.5 /I 7.4 = 0.8 ±.01), with a ph 50 of 7.2. Thus, human ENaC current is regulated by ph within the range found in the kidney collecting duct and other epithelia. In Figs. 2.2A and 2.2B, we tested the effect of ph on amiloride-sensitive current in epithelia that express endogenous ENaC. In a human lung carcinoma cell line (H441), amiloride-sensitive current was larger at ph 6.5 than at ph 7.4 (Fig. 2.2A and 2.2C). Results were similar in primary cultures of human airway epithelia (Fig. 2.2B and 2.2C). ph 6.5 increased ENaC current in both epithelia to an extent similar to that when ENaC was expressed in Xenopus oocytes (Fig. 2.2C). These data indicate that shifts in ph alter ENaC current not only in heterologous cells, but also in Na + -transporting epithelia.
39 23 Species-Specificity of ph Regulation In contrast to human ENaC, rat ENaC currents were not altered by ph changes over the range in Xenopus oocytes (Fig. 2.3A and 2.3B). This finding suggests that ph regulates ENaC in a species-specific manner. To determine which subunit(s) underlie this species-specificity, we substituted one of the human ENaC subunits in the channel complex with a rat subunit. Channels composed of human α and γenac with rat βenac had a ph dose-response relationship similar to human ENaC (Fig. 2.3B). In contrast, ph failed to alter current when rat γenac was expressed with human α and βenac (Fig. 2.3B). When rat αenac was expressed with human β and γenac, ph altered ENaC current, although the effect of ph was reduced (Fig. 2.3B). Together, the data indicate that sequence differences in γenac (and perhaps αenac) underlie the observed species variation in ENaC regulation by protons. ph Regulates ENaC Gating To determine the biophysical mechanism by which ph alters ENaC current, we took advantage of a strategy that prevents changes in ENaC gating. Covalent modification of a cysteine introduced at the DEG position in the external pore (β S520C, location noted in Fig 1.1) locks ENaC in an open conformation (open probability 0.96) (80). When β S520C was coexpressed with α and γenac in Xenopus oocytes, ph altered current to a similar extent as wild-type ENaC (Fig. 2.4A and 2.4D). In contrast, following modification of the channel with [2- (trimethylammonium)ethyl]methanethiosulfonate bromide (MTSET), changes in ph had a minimal effect on ENaC current (Fig. 2.4B and 2.4D). As a second approach to address
40 24 this question, we activated channels at the cell surface by proteolytic cleavage with trypsin, which dramatically increases ENaC open probability (29). Following incubation with trypsin, changes in ph failed to alter ENaC current (Fig. 2.4C and 2.4D). Together, the data suggest that ph regulates ENaC by altering channel gating, and argues against effects of ph on single channel conductance or channel number. Extracellular ph modulates Na + self-inhibition Previous work indicates that extracellular Na + regulates ENaC gating, a mechanism known as Na + self-inhibition. The representative traces in Fig. 2.5A illustrate this regulation. A shift in extracellular Na + from 1 mm to 116 mm induced a peak inward current (I P ) that rapidly decreased to a lower steady state level (I SS ). This decline in current reflects inhibition of ENaC by Na +. Proteolytic cleavage activates ENaC by preventing Na + self-inhibition (65). Because we found that cleavage abolished the effect of ph on ENaC, we hypothesized that ph might regulate ENaC gating by altering Na + self-inhibition. To test this hypothesis, we varied the extracellular ph and quantitated the fraction of ENaC current that was inhibited by Na + following a shift from 1 mm Na + to 116 mm Na +. We found that Na + self-inhibition was ph-dependent. At acidic ph (6.5), a smaller fraction of amiloride-sensitive current was inhibited by Na + than at alkaline ph (8.5), resulting in an increase in steady state current at ph 6.5 (Fig. 2.5A and 2.5B). In contrast to its effect on steady state current, ph had no effect on the peak current induced by the shift in Na + concentration (Fig. 2.5A and 2.5C). Protons could inhibit Na + self-inhibition by reducing binding of Na + to ENaC. To investigate this possibility, we determined dose-response relationships for Na + self-
41 25 inhibition at ph 8.5, 7.4, and 6.5 (fit to Hill equation). In Fig. 2.5D, we shifted the extracellular Na + from 0 mm to concentrations between 1 mm and 116 mm. At ph 7.4, Na + produced a dose-dependent increase in self-inhibition; half-maximal inhibition (K i ) occurred at 44.9 ± 1.6 mm Na + and the maximum fraction of current inhibited by Na + was 0.57 ± ph 6.5 decreased the maximal effect of Na + (0.49 ± 0.03) but it had no effect on the sensitivity of ENaC to Na + (K i = 44.7 ± 4.8). ph 8.5 had the opposite effect, increasing the maximal effect of Na + (0.66 ± 0.05) with no effect on sensitivity (K i = 43.5 ± 5.4). Thus, ph modulates Na + self-inhibition by altering the maximal effect of Na + on ENaC without affecting its sensitivity to Na +. Regulation by Extracellular ph Requires Sodium If ph regulates ENaC by modulating Na + self-inhibition, then the ph effect should be dependent on the presence of Na + in the extracellular solution. To test this prediction, we measured outward amiloride-sensitive currents (holding potential +30 mv) with 1 mm or 116 mm Na + in the bathing solution. In 1 mm Na +, ph had a negligible effect on ENaC current (Fig. 2.6A and 2.6C). In contrast, in 116 mm Na +, ph 8.5 reduced and ph 6.5 increased outward ENaC current (Fig. 2.6B and 2.6C), similar to the effect of ph on inward currents with 116 mm Na + (Fig. 2.6C, compare black bars to open bars). Thus, extracellular Na + is required for ph to regulate ENaC. In addition, these results indicate that ph regulates ENaC independent of voltage and the direction of Na + movement.
42 26 Mutations that Alter Na + Self-inhibition also Affect ph Regulation Previous work identified two ENaC histidine residues critical for Na + selfinhibition (39). Mutation of His-239 in mouse γenac (His-233 in human) abolished self-inhibition, whereas mutation of the equivalent residue in αenac (mouse His-282, which is His-255 in human) increased self-inhibition. To test if these mutations alter the effect of ph on ENaC current, we expressed α H255R βγenac or αβγ H233R ENaC in Xenopus oocytes. γ H233R nearly abolished the effect of ph on ENaC current (Fig. 2.7A and 2.7C). Conversely, α H255R increased the degree of ENaC activation by acidic ph (compared to wild-type ENaC), although it had no effect at alkaline ph (Fig. 2.7B and 2.7C). Thus, mutation of γenac abolished both Na + self-inhibition and ENaC regulation by ph, whereas both properties were increased by mutation of αenac. This correlation, together with our other data, supports a model in which ph alters ENaC current by modulating Na + self-inhibition. Discussion Our data indicate that the activity of human ENaC is modulated by changes in extracellular ph; acidic ph increased activity whereas alkaline ph reduced current. ph changes altered ENaC gating; by relieving Na + self-inhibition, protons increased ENaC open-probability. How does ph alter Na + self-inhibition? We hypothesize that protons titrate one or more ENaC residues that alter the response to Na +. This could occur through two general mechanisms. First, it is possible that protons reduce Na + binding to the extracellular
43 27 domain. However, our finding that ph changes did not alter the K i for Na + self-inhibition argues against this model, since it indicates that protons do not alter the affinity of Na + for ENaC. Thus, it is unlikely that protons titrate residues that form the Na + binding site. Second, ph could function down-stream of Na + binding, altering the transduction of Na + binding into changes in ENaC gating. Consistent with this model, we found that protons increased the maximal effect of Na + on ENaC current. What residues are titrated by protons? We found that the effect of ph on ENaC current was independent of voltage and on the direction of current flow, which indicates that protons do not alter ENaC current by titrating residues in its pore. Thus, it is likely protons titrate one or more residues in extracellular domain. The recent crystal structure of ASIC1 identified H + binding sites located at the interface between the finger and thumb domains of this related channel (43). Movement of the thumb domain relative to the finger domain was proposed to underlie channel gating. Although the ASIC1 H + binding residues are not well-conserved in ENaC, we speculate that the interface between the finger and thumb domains has a conserved role in modulating channel activity, and participates in H + binding to ENaC. Consistent with this idea, residues that modulate ENaC function are located at this interface. For example, proteolytic cleavage activates ENaC by removal of part of the finger domain. Moreover, the α and γenac histidines important for Na + self-inhibition (α H255R and γ H233R ) are located at the finger-thumb interface. However, it seems unlikely that protons regulate ENaC by titrating these histidines, since the α H255R and γ H233R mutations had opposite effects on ph-modulation of ENaC current. In addition, these histidines are conserved in rat ENaC, which did not respond to changes in ph.
44 28 As a related question, which ENaC subunits mediate the ph-sensitivity of ENaC? Do protons bind to all three subunits or only a subset of subunits? Our data implicate a role for γenac, since substitution of rat in place of human γenac abolished channel regulation by protons. Substitution of the α subunit had a partial effect on ph regulation, suggesting that residues in αenac may also contribute. In contrast, substitution of βenac did not alter ENaC regulation by protons. This situation is somewhat analogous to ENaC regulation by proteolytic cleavage, where ENaC is regulated by cleavage of the extracellular domains of α and γenac, but not βenac (60). However, our current data do not exclude a role for βenac; it is possible that human and rat βenac share conserved H + binding sites. The species difference in ph regulation suggests a strategy to identify residues that contribute to this regulation (see Chapter V). The extracellular domain of human γenac contains a large number of potential targets for protons that are not present in rat ENaC (14 histidines and acidic residues) and there are 10 in αenac. Based on analogy to ASIC1 (43, 81), we speculate that titration of multiple residues contributes to ph-regulation of human ENaC - this is the focus of Chapter V. How can we reconcile our current results with previous conflicting data in the literature? First, there appear to be species differences in ENaC regulation by ph. Although ph altered human ENaC current, it had no effect on rat ENaC in the range we tested, consistent with previous work. From a comparative physiology perspective, we do not yet know why ph regulates ENaC in some species but not others. In humans, differences in diet can produce wide variations in urine ph. For example, consumption of meat produces an acidic urine, whereas vegetarians have an alkaline urine. Perhaps rats lacked selective pressure to develop or retain the ph response because of a less
45 29 varied diet. Conflicting data may also arise from differences in the proteolytic cleavage state of ENaC in various experimental systems. We found that proteolytic cleavage (trypsin) abolished the effect of ph on human ENaC. Thus, the response to ph is influenced by the cleavage state of the channel. Under conditions where the majority of ENaC is fully cleaved, the response to ph will be blunted. ENaC is exposed to extreme changes in ph. Coupled with our data showing that ph alters ENaC current, this suggests that ph may be an important regulator of epithelial Na + transport. What is the physiological role of this regulation? In the kidney collecting duct, it is well-described that there is an interdependence between Na + absorption and H + secretion; Na + absorption generates a transmembrane voltage which increases H + secretion (82, 83). Perhaps activation of ENaC by protons facilitates secretion of an acid load. In this regard, it is interesting that the Na + -absorbing principal cells are located next to intercalated cells, which are responsible for H + secretion. This suggests a potential model in which protons function as a paracrine signal to regulate Na + absorption. In the lung, it is possible that acidic airway fluid stimulates ENaC in order to remove liquid from the lung in conditions such as pneumonia and pulmonary edema.
46 30 Figure 2.1. Extracellular protons modulate human ENaC current. A, representative trace of current versus time recorded in Xenopus oocyte expressing human αβγ ENaC at holding potential of 60 mv. The extracellular bath was changed from ph 7.4 (open bars) to ph 8.5 and 6.5 (black bars) with or without 10 µm amiloride (black bars), as indicated. B, Amiloride-sensitive current for human ENaC (relative to current at ph 7.4) with bathing solution ph varied from 8.5 to 5.5 (mean ± S.E.M, n = 37; error bars are hidden by the symbols). Data are fit to Hill equation (R 2 = ).
47 31
48 32 Figure 2.2. Extracellular protons modulate Na + transport in epithelia. A and B, representative short-circuit current traces from H441 (A) and primary human airway (B) epithelia. ph changes and addition of amiloride to the apical bathing solution (10 µm) are indicated by the bars. 0.5 mv pulses were applied every 15 seconds to monitor resistance. C, percent increase in amiloride-sensitive current in response to ph 6.5 (compared to ph 7.4) for H441 epithelia (n = 6), primary airway epithelia (n = 6), and oocytes expressing human αβγ ENaC (n = 37) (mean ± S.E.M.). * indicates that the change in amiloride-sensitive current at ph 6.5 (compared to ph 7.4) is statistically significant (p < for H441, < for airway, and < for oocytes by Student s t-test).
49 33
50 34 Figure 2.3. Species-specificity of ph regulation. A, representative traces of current versus time recorded in Xenopus oocyte expressing rat αβγ ENaC at holding potential of 60 mv. The extracellular bath was changed from ph 7.4 (open bars) to ph 8.5 and 6.5 (black bars). 10 µm amiloride (black bar) was added to the bathing solution, as indicated. B, Amiloride-sensitive current for human (H) or rat (R) ENaC, or the indicated combinations of two human and one rat subunit (relative to current at ph 7.4) with bathing solution ph varied from 8.5 to 6 (mean ± S.E.M, n = 5-37; error bars are hidden by the symbols). The Hill equation was used to fit the data for human (R 2 = ), human βγ rat α (R 2 = ), and human αγ rat β (R 2 = ).
51 35
52 36 Figure 2.4. ph regulates ENaC gating. A and B, representative current traces at holding potential of 60 mv from the same Xenopus oocyte expressing human αβ S520C γ ENaC before (A) and after (B) covalent modification with 1 mm [2-(trimethylammonium)ethyl]methanethiosulfonate bromide (MTSET) for 2 min. C, representative current trace at holding potential of 60 mv from a Xenopus oocyte expressing wild-type human αβγenac after proteolytic cleavage by trypsin (2 µg/ml for 2 min). In A-C, ph changes are indicated by open (ph 7.4) or black (ph 8.5 or 6.5) bars. 10 µm amiloride was present in bathing solution, as indicated by black bars. D, fold change in amiloride-sensitive current in response to ph 8.5 or 6.5 (relative to current at ph 7.4) before and after application of MTSET (n = 5) or trypsin (n = 7) to oocytes expressing human αβ S520C γ or αβγ ENaC, respectively (mean ± S.E.M.). * indicates that the difference between the current at ph 6.5 and the current at ph 8.5 (I I 8.5 ) is statistically different between the indicated groups (p < 0.001).
53 37
54 38 Figure 2.5. Extracellular ph modulates Na + self-inhibition. A, representative current trace from Xenopus oocytes expressing human αβγ ENaC (-60 mv). The extracellular bath ph was 7.4 unless otherwise indicated by black bars. The bath was rapidly changed from 1 mm to 116 mm NaCl (at ph 8.5 or 6.5) to observe the degree of Na + self-inhibition. Peak current (I P ) and steady-state current (I SS ) are indicated. 10 µm amiloride was added to quantitate ENaC current. B, plot of Na + self-inhibition of amiloride-sensitive current [(I P I SS )/ I P ] at ph (mean ± S.E.M., n = 14). C, plot of peak amiloride-sensitive current following shift in bathing solution from 1 mm to 116 mm NaCl measured at ph (relative to peak current at ph 7.4) (mean ± S.E.M., n = 14). D, Na + self-inhibition induced by shift from 0 mm Na + to mm Na + (plotted on X-axis) at ph 8.5, ph 7.4, and ph 6.5 (mean ± S.E.M., n = 3-5). Data are fit to Hill equation; R 2 = (ph 8.5), (ph 7.4), and (ph 6.5).
55 39 A Na + (mm) Current (µa) ph I SS 6.5 I SS Time (sec) Amil I P B (IP-ISS)/IP ph C IP (rel.) D (I P -ISS)/IP ph ph [Na + ]
56 40 Figure 2.6. Regulation by extracellular ph requires Na +. A and B, representative current traces from Xenopus oocytes expressing human αβγ ENaC voltage clamped at +30 mv. Extracellular bathing solution contained 1 mm Na + (A) or 116 mm Na + (B). Extracellular ph was changed from ph 7.4 (open bars) to ph 8.5 and 6.5 (black bars), as indicated. In panel B, grey line indicates time-dependent drift in outward amiloride-insensitive current. C, plot of fold change in amiloride sensitive current at ph 8.5 and 6.5 (relative to ph 7.4, [(I ph X I ph 7.4 )/ I ph 7.4 ]. Inward currents were studied in 116 mm Na + at -60 mv (n = 37) and outward currents studied in 116 or 1 mm Na + at +30 mv (n = 9 and 14, respectively) (mean ± S.E.M., * p < versus 116 mm Na + at 60 or +30 mv by Student s t-test).
57 41
58 42 Figure 2.7. Mutations that alter Na + self-inhibition affect ph regulation. A and B, representative current traces at holding potential of 60 mv from Xenopus oocytes expressing; (A) human αβγ H233R ENaC or (B) α H255R βγ ENaC. The extracellular bath was changed from ph 7.4 (open bars) to ph 8.5 and 6.5 (black bars) with or without 10 µm amiloride (black bars), as indicated. C, amiloride-sensitive current for wild-type or mutant ENaC (relative to current at ph 7.4) with bathing solution ph varied from 8.5 to 5.5 (mean ± S.E.M, n = 21-37; some error bars are hidden by the symbols). Data are fit to Hill equation; R 2 = (α H255R βγ), (αβγ H233R ), and (wild-type).
59 43 A Current (μa) α β γ H233R ph Amil 0 α H255R β γ ph Amil Time (sec) Time (sec) B Current (µa) C IAmil (rel. to 7.4) α H255R β γ α β γ α β γ H233R ph
60 44 CHAPTER III EXTRACELLULAR CL - REGULATES THE EPITHELIAL SODIUM CHANNEL Preface This work appeared in the Journal of Biological Chemistry on August 27 th, I am co-author on this work with Dr. Snyder. I designed and performed the experiments, analyzed the data, and wrote the manuscript. Dr. Snyder designed experiments and wrote the manuscript. This work is describes the regulation of ENaC activity by extracellular chloride. Crystallization of a related protein, ASIC1a, reveled intersubunit chloride binding sites in the extracellular domain. We used the crystal structure of ASIC1a to identify potential chloride binding sites in ENaC. Abstract The extracellular domain of the epithelial sodium channel ENaC is exposed to a wide range of Cl - concentrations in the kidney and in other epithelia. We tested whether Cl - alters ENaC activity. In Xenopus oocytes expressing human ENaC, replacement of Cl - with SO 2-4, H 2 PO - 4, or SCN - produced a large increase in ENaC current, indicating that extracellular Cl - inhibits ENaC. Extracellular Cl - also inhibited ENaC in Na + - transporting epithelia. The anion selectivity sequence was SCN - < SO 2-4 < H 2 PO - 4 < F - < I - < Cl - < Br -. Crystallization of ASIC1a revealed a Cl - binding site in the extracellular domain. We found that mutation of corresponding residues in ENaC (αh418a and βr388a) disrupted the response to Cl -, suggesting that Cl - might regulate ENaC through an analogous binding site. Maneuvers that lock ENaC in an open state (a DEG mutation and trypsin) abolished ENaC regulation by Cl -. The response to Cl - was also modulated
61 45 by changes in extracellular ph; acidic ph increased and alkaline ph reduced ENaC inhibition by Cl -. Cl - regulated ENaC activity in part through enhanced Na + selfinhibition, a process by which extracellular Na + inhibits ENaC. Together, the data indicate that extracellular Cl - regulates ENaC activity, providing a potential mechanism by which changes in extracellular Cl - might modulate epithelial Na + absorption. Introduction The epithelial Na + channel ENaC is a heterotrimer of homologous α, β, and γ subunits (78, 84). ENaC functions as a pathway for Na + absorption across epithelial cells in the kidney collecting duct, lung, distal colon, and sweat duct (reviewed in (52, 53)). Na + transport is critical for the maintenance of Na + homeostasis and for the control of the composition and quantity of the fluid on the apical membrane of these epithelia. ENaC mutations, and defects in its regulation, cause inherited forms of hypertension and hypotension (3), and may contribute to the pathogenesis of lung disease in cystic fibrosis (54). ENaC is a member of the DEG/ENaC family of ion channels. A common structural feature of these channels is a large extracellular domain that plays a critical role in channel gating. For example, in ASICs, the extracellular domain function as a receptor for protons, which transiently activate the channel by titrating residues that form an acidic pocket (43). FaNaCh is a ligand-gated family member in Helix aspersa, activated by the peptide FMRFamide (57). In C. elegans MEC family members, the extracellular domain is thought to respond to mechanical signals (56).
62 46 ENaC differs from other family members since it is constitutively active in the absence of a ligand/stimulus. However, a convergence of data indicate that ENaC gating is modulated by a variety of molecules that bind to or modify its extracellular domains, including proteases (28, 29, 60), Na + (36, 39, 64), protons (Chapter II (85)), and the divalent cations Zn 2+ and Ni 2+ (40, 66). These findings suggest that the ENaC extracellular domain might regulate epithelial Na + transport by sensing and integrating diverse signals in the extracellular environment. In the current study, we tested the hypothesis that ENaC activity is regulated by changes in the extracellular Cl - concentration. Several observations suggested that Cl - might be a strong candidate to regulate the channel. First, transport of Na + and Cl - are often coupled to maintain electroneutrality. Second, ENaC is exposed to large changes in extracellular Cl - concentration. For example, in the kidney collecting duct, the urine Cl - concentration varies widely (67). As the predominant anion, its concentration parallels that of Na + in most clinical states. However, under conditions of metabolic alkalosis and metabolic acidosis, the Na + and Cl - concentrations can become dissociated as a result of increased urinary bicarbonate (alkalosis) or ammonium (acidosis) (67). Thus, ENaC is well positioned to respond to changes in Cl - concentration. Third, crystallization of ASIC1a revealed a binding site for a Cl - ion at the base of the thumb domain (43). The Cl - is coordinated by Arg-310 and Glu-314 from one subunit, and Lys-212 from an adjacent subunit. Although the functional role of Cl - binding to ASIC1a is unknown, it supports the hypothesis that extracellular Cl - might regulate the activity of DEG/ENaC ion channels.
63 47 Experimental Procedures DNA Constructs cdnas for human α, β, and γenac in pmt3 were cloned as previously described (1, 78). The mutations α H418A, β R388A, γ H396A, and γ H233R were generated by site-directed mutagenesis (QuickChange; Stratagene) and sequenced in the University of Iowa DNA Core. Expression and Whole-Cell Electrophysiology in Xenopus oocytes Oocytes were harvested from albino Xenopus laevis females and manually defolliculated following a one hour treatment with 0.75 mg/ml Type IV Collagenase (Sigma) in Ca 2+ free ND-96 (96 mm NaCl, 2 mm KCl, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH). Following nuclear injection of cdnas encoding α, β, and γenac (0.02 µg/µl each), cells were incubated at 18 C in modified Barth s saline (88 mm NaCl, 1 mm KCl, 0.33 mm Ca(NO 3 ) 2, 0.41 mm CaCl 2, 0.82 mm MgSO 4, 2.4 mm NaHCO 3, 10 mm HEPES, 50 µg/ml gentamycin sulfate, 10 µg/ml sodium penicillin, 10 µg/ml streptomycin sulfate, ph adjusted to 7.4 with NaOH) for hours prior to study. Oocytes were voltage clamped (two-electrode voltage clamp) and currents were amplified with an Oocyte Clamp OC-725C (Warner Instruments), digitized with a MacLab/200 interface (ADInstruments), and recorded and analyzed with Chart software (ADInstruments). The bathing solution was grounded with 3 M KCl-agar bridges to prevent junction potentials during changes in Cl - concentration. Unless otherwise noted, recordings were done at 60 mv in a 116 mm NaCl solution (116 mm NaCl, 2 mm KCl, 0.4 mm CaCl 2, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH). Low Na + solutions (1 mm NaCl, 115 mm N-Methyl-D-glucamine Cl or SO 4, ph adjusted with HCl or H 2 SO 4 ) were used as indicated in the figure legends. Low chloride solutions were prepared by replacing the
64 48 anion of the sodium salt with I -, Br -, F -, SCN -, H 2 PO - 4, or SO 2-4 and D-mannitol (to balance osmolarity). Amiloride-sensitive current was determined by adding 10 µm amiloride to the bathing solution. The ph and chloride-induced changes in amiloridesensitive current were calculated as the fold increase/decrease relative to the baseline current in ph 7.4, 116 NaCl solution just prior to each test solution application. This was done to reduce the effect of time-dependent current run-down. The resulting data were plotted and fit to the Hill equation using IGOR Pro software (WaveMetrics Inc.). Na + self-inhibition was measured by rapidly changing the bathing solution from low sodium (1 mm) to high sodium (116 mm) and quantitated as (peak current steady state current) / peak current. Expression and Whole-Cell Electrophysiology in H441 and primary airway epithelia H441 cells (American Type Culture Collection) were grown on 0.6 cm 2 permeable filter supports (Millipore) in RPMI with 8.5% fetal calf serum, 20 mm L- glutamine, 5 µg/ml insulin, 5 µg/ml transferrin, 5 ng/ml selenium, 100 nm dexamethasone, 100 U/ml penicillin, and 100 mg/ml streptomycin at 37 C for 5 days. Primary human airway epithelia were isolated from the trachea and bronchi of donor lungs and grown at the air-liquid interface of collagen-coated permeable filter supports, as described previously (79). Aprotinin (26 µg/ml) was present in the apical solution for 2 h prior to study. Short-circuit Na + current was measured in modified Ussing chambers (Warner Instrument Corporation) using an EC-825 Epithelial Voltage Clamp amplifier (Warner Instrument Corporation). Currents were digitized with a PowerLab interface (ADInstruments) and recorded and analyzed with Chart software (ADInstruments). The apical and basolateral surfaces were bathed in 116 mm NaCl, 2 mm KCl, 0.4 mm CaCl 2,
65 49 1 mm MgCl 2, 5 mm HEPES (ph adjusted to 7.4 with NaOH) or 58 mm Na 2 SO 4, 58 mm D-mannitol, 2 mm KCl, 0.4 mm CaCl 2, 1 mm MgCl 2, 5 mm HEPES (ph adjusted to 7.4 with NaOH) at 37 C. Amiloride-sensitive short-circuit current was determined as the difference in current with and without amiloride (10 µm) in the apical bathing solution. Results Extracellular Anions Modulate ENaC Current To determine if extracellular Cl - modulates ENaC current, we replaced Cl - with other anions and recorded ENaC currents in Xenopus oocytes expressing human α, β, and γ ENaC. When Cl - was replaced by SO 2-4, amiloride-sensitive ENaC current increased by 2.4-fold (Fig. 3.1A and 3.1B). This increase was rapid and reversible. Replacement of Cl - with H 2 PO - 4 and SCN - also increased ENaC current (Fig. 3.1B). As a control, Cl - replacement did not significantly alter current in oocytes not expressing ENaC (not shown). We also excluded the possibility that SO 2-4 and H 2 PO - 4 altered ENaC current by lowering Ca 2+ activity in the bathing solution; the effect of Cl - replacement was not reduced when Ca 2+ activity was held constant (not shown). Fig. 3.1C shows a dose-response relationship for Cl -. As extracellular Cl - increased (replacing SO 2-4 ), there was a dose-dependent decrease in ENaC current. The maximal fraction of ENaC current inhibited by Cl - was 0.73 and half-maximal inhibition occurred at a Cl - concentration of 29.5 ± 0.25 mm. The data indicate that Cl - inhibits ENaC current at concentrations found in the kidney collecting duct and other epithelia (67).
66 50 We considered the possibility that removal of extracellular Cl - could alter ENaC current by reducing the concentration of intracellular Cl - (86). To exclude this possibility, we voltage-clamped cells at +30 mv to prevent movement of Cl - out of the cell. Under these conditions, replacement of Cl - with SO 2-4 still increased amiloridesensitive current (Fig. 3.1D). This result indicates that changes in extracellular Cl - are sufficient to alter ENaC current. We asked whether other anions could substitute for Cl -. Compared to SO 2-4, I -, Br -, and F - each decreased ENaC current. F - and I - decreased current to a smaller extent than Cl -, and Br - produced a larger decrease in current (Fig. 3.1E). Thus, the anion selectivity sequence for ENaC inhibition is SCN - < SO 2-4 < H 2 PO - 4 < F - < I - < Cl - < Br -. Cl - Inhibits ENaC in Epithelia To test the effect of extracellular chloride on native ENaC in epithelial cells, we measured short-circuit current. In a human lung carcinoma cell line (H441), we measured amiloride-sensitive current when the apical and basolateral membranes were bathed in NaCl (Fig. 3.2A). When Cl - was replaced with SO 2-4 on the apical and basolateral membranes (to eliminate a Cl - gradient), there was an increase in amiloridesensitive current (Fig. 3.2A and quantitated in Fig. 3.2B). Results were similar in primary cultures of human airway epithelia (Fig 3.2B). Thus, Cl - regulates ENaC both in Xenopus oocytes and in Na + -transporting epithelia.
67 51 Identification of Residues that Participate in Cl - Regulation of ENaC We hypothesized that Cl - might regulate ENaC by binding to a site analogous to the Cl - binding site observed in the crystal structure of ASIC1a (43). To identify candidate residues, we lined up the ASIC1a and ENaC sequences. In general, sequence conservation in the extracellular domains is low. We therefore focused on a portion of the thumb domain where the lineup was facilitated by the presence of four highly conserved cysteine residues (Fig. 3.3A). In ASIC1a, the positively charged side chain of Arg-310 participates in the coordination of Cl -. In βenac, an arginine is located one position down-stream (β-arg-388). The equivalent residues in αenac and γenac are histidine (418 and 396, respectively), which when protonated carry a positive charge that could coordinate Cl -. To test their role in Cl - regulation of ENaC, we mutated each of these residues to alanine. In Fig. 3.3B, we expressed wild-type or mutant (α H418A β R388A γ H396A ) ENaC in Xenopus oocytes and varied the extracellular Cl - concentration. Compared to wild-type ENaC, the mutations reduced Cl - -dependent inhibition by more than 50%. We also tested the effect of mutations in individual ENaC subunits. ENaC currents in cells expressing α H418A βγenac or αβ R388A γenac were also less responsive to Cl -, whereas Cl - inhibited αβγ H396A channels similar to wild-type (Fig. 3.3B). Together, these results suggest that α- His-418 and β-arg-388 participate in ENaC regulation by Cl -. Although we cannot exclude a role for γ-his-396, this residue is not required for Cl - to inhibit ENaC. Because mutation of these residues did not completely abolish the effect of Cl - on ENaC, it seems likely that additional residues also contribute.
68 52 DEG mutation and proteolytic cleavage abolish response to Cl - Because Cl - removal or re-addition altered ENaC current over a rapid time-course (seconds), it seemed likely that Cl - affected channel activity rather than changing ENaC trafficking. To more directly test this idea, we used two strategies to prevent changes in ENaC activity. First, we covalently modified a cysteine introduced at the DEG position (Fig 1.1) at the extracellular end of the pore in βenac (β S520C ), a maneuver that locks ENaC in the open state (open probability = 0.96) (80). Fig. 3.4A shows an oocyte expressing αβ S520C γ ENaC before (black line) and after (gray line) modification with MTSET. To emphasize the Cl - -dependent effect, the traces were normalized to the peak currents and superimposed. Before modification, Cl - decreased current similar to wildtype ENaC (Fig. 3.4A, black line, and 3.4B). In contrast, Cl - had no effect on ENaC current after the introduced cysteine was modified with MTSET (Fig. 3.4A, gray line, and 3.4B). As a second strategy, we took advantage of the observation that ENaC gating is disrupted by proteolytic cleavage of the extracellular domain, which also increases open probability (29). Treatment of wild-type ENaC with trypsin (2 µg/ml for 3 min) abolished the effect of Cl - on current (Fig. 3.4C, gray line, and 3.4D). Together, the data indicate that Cl - is incapable of inhibiting ENaC channels that are locked in an open conformation or that are proteolytically cleaved and activated by trypsin. Cl - Alters Na + Self-inhibition ENaC is regulated in part by changes in extracellular Na + (Na + self-inhibition) (36, 39, 64). We hypothesized that Cl - might inhibit ENaC by modulating the effect of
69 53 Na + on ENaC. To test this, we quantitated Na + self-inhibition with Cl - or SO 2-4 as the predominant anion in the extracellular bathing solution. In Xenopus oocytes expressing ENaC, a rapid increase in extracellular Na + (from 1 to 116 mm) produced an inward Na + current (downward deflection) that rapidly decreased to a lower steady-state level, as a result of Na + self-inhibition (Fig. 3.5A). When Cl - was the anion, the fraction of amiloride-sensitive current inhibited by Na + was 0.58 ± 0.02 (Fig. 3.5B). When Cl - was replaced by SO 2-4 or SCN -, a smaller fraction of ENaC current was inhibited by Na + (0.23 ± 0.06 and 0.25 ± 0.02, respectively) (Fig. 3.5A and 3.5B). Mutation of the putative Cl - binding site in all three subunits (α H418A β R388A γ H396A ) reduced Na + self-inhibition to a similar extent as Cl - removal (Fig. 3.5C and 3.5D). Na + self-inhibition was also reduced by mutation of the α subunit alone, but not by mutation of β or γenac (Fig. 3.5D). The data suggest that Cl - inhibits ENaC in part by increasing Na + self-inhibition. If correct, then removal of Na + from the extracellular solution should disrupt the effect of anion substitution on ENaC current. To test this prediction, we replaced extracellular Na + with NMDG and measured outward amiloride-sensitive current. Under these conditions, substitution of extracellular Cl - with SO 2-4 increased ENaC current much less (I SO4 /I Cl = 1.46 ± 0.06) than when Na + was present in the extracellular solution (I SO4 /I Cl = 2.38 ± 0.12) (Fig. 3.5E). In contrast, when the putative Cl - binding sites were mutated, the residual effect of anion substitution was not altered by changes in extracellular Na + (Fig. 3.5E). As a further test of the role of Na + self-inhibition in ENaC regulation by Cl -, we mutated a residue required for Na + self-inhibition. Previous work found that mutation of His-239 in the extracellular domain of mouse γenac abolished Na + self-inhibition (40,
70 54 66). Mutation of the equivalent residue in human γenac (His-233) had a similar effect (85). We tested whether this mutation (αβγ H233R ) would disrupt the effect of Cl - on ENaC. In contrast to wild-type ENaC, changes in Cl - concentration had little effect on the mutant channel (Fig. 3.5F). Together, the data suggest that Cl - regulates ENaC in part by modulating Na + self-inhibition. ph Modulates the Effect of Cl - on ENaC If the histidines we identified in Fig. 3.3 are Cl - binding sites, then we might expect ENaC regulation by Cl - to be altered by changes in extracellular ph. The imidazole side chain of histidine has a relatively neutral pka (6.0), although this value can shift depending on its local environment. At alkaline ph, the side chain is uncharged, but at acidic ph it is protonated and carries a positive charge, which could enhance Cl - binding through electrostatic effects. To test this prediction, we varied the ph in the bathing solution and replaced Cl - with SO 2-4. Fig. 3.6A shows a representative current trace. At ph 6.5, Cl - removal produced a large increase in amiloride-sensitive current. At ph 8.5, Cl - removal also increased current, but the increase was smaller than at ph 6.5. Fig. 3.6B shows Cl - dose-response relationships at ph 6.5, 7.4, and 8.5. Compared to ph 7.4, Cl - inhibition of ENaC was increased at acidic ph and reduced at alkaline ph. Mutation of the putative Cl - binding residues (α H418A β R388A γ H396A ) abolished the effect of ph on ENaC inhibition by Cl - (Fig. 3.6C). Thus, ENaC regulation by Cl - is modulated by changes in extracellular ph. Because protons increase ENaC inhibition by Cl -, this effect should tend to reduce ENaC current at acidic ph. However, in Chapter II we observed the opposite; acidic ph
71 55 increased ENaC current (85) (in Fig. 3.6A, compare current at ph 6.5, 7.4, and 8.5 with Cl - bathing solution). This resulted from a decrease in Na + self-inhibition at acidic ph ((85) and Fig. 3.6D Cl - ). We hypothesized that protons have two opposing effects on Na + self-inhibition; one alters Cl - binding and increases self-inhibition (which decreases current), whereas the other is independent of Cl - and reduces self-inhibition (which increases current). To eliminate the Cl - -dependent component, we removed Cl - from the bathing solution and tested the effect of protons on Na + self-inhibition. Under these conditions, the inhibitory effect of protons on Na + self-inhibition was greatly enhanced; acidic ph decreased Na + self-inhibition to a larger extent than when Cl - was the extracellular anion (Fig. 3.6D). Thus, although protons tend to decrease ENaC current through their effect on Cl - binding, this is countered by an independent inhibitory effect of protons on Na + self-inhibition. The net result is increased ENaC current at acidic ph. Discussion Our findings in heterologous cells and in Na + -transporting epithelia indicate that ENaC is regulated by changes in extracellular Cl -. As the concentration of extracellular Cl - increased, we observed a dose-dependent reduction in ENaC Na + current. Cl - could regulate ENaC directly by binding to the channel, or indirectly through interactions with lipids in the plasma membrane. Based on two observations, we favor direct binding of Cl - to ENaC. First, the crystal structure of ASIC1a identified a Cl - binding site in its extracellular domain, raising the possibility that an analogous site is present in ENaC. Second, Cl - inhibition of ENaC was reduced by mutation of residues in αenac (His-418) or βenac (Arg-388) that correspond to the ASIC1a Cl - binding site.
72 56 Thus, it seems likely that these residues contribute to Cl - binding in ENaC. However, mutation of these residues did not completely abolish the effect of Cl - on ENaC current. In ASIC1, Cl - is coordinated by three residues (43), so it is possible that there is residual Cl - binding to other ENaC residues. Alternatively, it is possible that Cl - regulates ENaC in part through binding to a second site or through a mechanism independent of channel binding. Our data indicate that Cl - inhibits ENaC through a change in channel activity. Maneuvers that locked the channel in a high open probability state (a DEG mutation and trypsin) abolished the effect of Cl - on ENaC current. This finding is most consistent with a model in which Cl - regulates ENaC predominately through a change in channel gating, although we can t exclude additional effects on ENaC trafficking or single channel conductance. How might this occur? Previous work indicates that ENaC gating is regulated in part by extracellular Na +, which inhibits the channel (Na + self-inhibition) (36, 39, 64). We found that Na + self-inhibition was reduced in the absence of extracellular Cl -. This finding supports a model in which Cl - inhibits ENaC gating by stabilizing the Na + self-inhibited state. Perhaps the binding of Na + to ENaC is enhanced when the channel is bound to Cl -. However, changes in Na + self-inhibition do not completely explain the effects of Cl - on ENaC current. For example, upon removal of Na + from the extracellular bathing solution, the effect of Cl - on ENaC was dramatically reduced, but not abolished (Fig. 3.5E). Moreover, mutation of the putative Cl - binding site in βenac reduced ENaC regulation by Cl -, but it had no effect on Na + self-inhibition (Fig. 3.5D). These observations suggest that in addition to its effect on Na + self-
73 57 inhibition, Cl - may have a direct effect on ENaC activity that is independent of its effect on Na + self-inhibition. We found that extracellular protons alter the effect of Cl - on ENaC; acidic ph enhanced inhibition, whereas alkaline ph reduced inhibition. Together with the data presented in Chapter II, this indicates that protons modulate ENaC through at least two independent mechanisms. We propose a model in which titration of the histidine residues that form the Cl - binding site (His-418 in αenac and possibly His-396 in γenac) increase Cl - binding, resulting in reduced ENaC current at acidic ph. This effect is opposed by titration of additional residues which decreases Na + self-inhibition, resulting in increased current at acidic ph (85). Thus, the net effect of ph on ENaC current is determined by the relative contribution of these two mechanisms. As a result, ENaC regulation by ph might vary under different physiological and pathophysiological conditions. For example, when the Cl - concentration is low (e.g. volume depletion), acidic ph would have a greater stimulatory effect on ENaC than when the Cl - concentration is high (e.g. volume expansion). ENaC is not the only cation channel that is modulated by extracellular Cl -. For example, Cl - and other anions shift the voltage dependence of voltage-gated Na + channels (87). The selectivity sequence for voltage-gated Na + channels approximated the lyotropic series (SO 2-4 < F - < Cl - < NO 3 < Br - < I - < SCN - ), where the anion effect is primarily related to the energy required for its dehydration, rather than the energy of interaction between the anion and its binding site. This situation is observed when the binding site has a relatively low affinity for anions (88). With ENaC, the anion selectivity sequence (SCN - < SO 2-4 < H 2 PO - 4 < F - < I - < Cl - < Br - ) also corresponded generally to the lyotropic
74 58 series, although SCN - and I - were displaced from their predicted positions. Together with the Cl - dose-response relationship, the data indicate that the ENaC binding site has a relatively low affinity for anions. This feature is important since it allows ENaC to be regulated by Cl - concentrations found in urine and airway surface liquid. What is the physiological role of ENaC regulation by Cl -? In the kidney collecting duct, the urine Na + and Cl - concentrations can vary over a wide range (67). Previous work indicates that changes in Na + concentration regulate the rate of Na + transport through two distinct mechanisms; Na + self-inhibition (36, 39, 64) and Na + feedback inhibition (89-94). We speculate that Cl - regulates Na + transport in an analogous manner. For example, a drop in collecting duct Cl - concentration under conditions of hypovolemia would facilitate Na + absorption, whereas increases in Cl - concentration in hypervolemia would reduce Na + absorption. Similar to ENaC regulation by Na +, this mechanism may function to maintain Na + and volume homeostasis. Regulation of ENaC by Cl - may also serve to indirectly regulate Cl - absorption, since the transport of Cl - is coupled to Na + transport to maintain electroneutrality. Together with previous work, our findings support a model in which the extracellular domains of ENaC function as sensors by which a variety of extracellular signals modulate epithelial Na + absorption.
75 59 Figure 3.1. Extracellular anions modulate ENaC activity. A, representative trace of current versus time from a Xenopus oocyte expressing human αβγ ENaC clamped at -60 mv. The bathing solution contained Cl - or SO 2-4, as indicated. Amiloride ( Amil, 10 µm) was present in the bathing solution, as indicated by black bars. B, amiloride-sensitive current with the indicated anion in the bathing solution (relative to Cl - ) (mean +/- S.E., n = 5-23, *p <.01). C, fraction of amiloridesensitive current inhibited versus extracellular Cl - concentration (total anion concentration held constant with SO 2-4 ) (mean +/- S.E., n = 9, error bars are hidden by the data points). D, amiloride-sensitive current with Cl - or SO 2-4 in the bathing solution (relative to Cl - ) at a holding potential of -60 mv (inward current) or +30 mv (outward current) (mean +/- S.E., n = 10-11, *p <.01. E, amiloride-sensitive current when Cl - in the bathing solution was replaced with the indicated anion (relative to Cl - ) (mean +/- S.E., n = 9-23, *p <.01).
76 60
77 61 Figure 3.2. Extracellular Cl - modulates endogenous ENaC activity in epithelia. A, representative short-circuit current trace in H441 cells. The apical and basolateral bathing solutions contained Cl - or SO 2-4, as indicated. Amiloride ( Amil, 10 µm) was added to the apical bathing solution, as indicated by the gray bars. 0.5 mv pulses were applied every 15 seconds to monitor resistance. B, amiloride-sensitive current with Cl - or SO 2-4 in the bathing solution (relative to Cl - ) in H441 and primary airway epithelial cells (mean +/- S.E., n = 5-6, *p <.01).
78 62
79 63 Figure 3.3. Identification of residues that participate in Cl - regulation of ENaC. A, sequence alignment of human α, β, and γenac and chicken ASIC1a. Boxes indicate residues that bind Cl - in ASIC1 and potential equivalent residue in ENaC. Arrowheads indicate conserved cysteine residues. B, fraction of amiloride sensitive current inhibited by Cl - (replacing SO 2-4 ) in oocytes expressing the indicated ENaC subunits (mean +/- S.E., n = 6-9). Some of the error bars are hidden by the data points.
80 64
81 65 Figure 3.4. DEG mutation and proteolytic cleavage abolish response to Cl -. A, representative current traces at a holding potential of -60 mv in the same Xenopus oocyte expressing human αβ S520C γ ENaC before (black trace) and after (gray trace) modification with MTSET ( ET ). The bathing solution contained SO 2-4 or Cl -, as indicated. 10 µm amiloride ( Amil ) was present in the bathing solution, as indicated by the gray bars. B, amiloride-sensitive current in oocytes expressing αβ S520C γ ENaC with Cl - or SO 2-4 in the bathing solution (relative to Cl - ) before and after treatment of with MTSET (mean +/- S.E., n = 3, *p <.01). C, representative current traces at a holding potential of -60 mv in the same Xenopus oocyte expressing human αβγ ENaC before (black trace) and after (gray trace) treatment with trypsin (2 µg/ml). D, amiloridesensitive current in oocytes expressing αβγ ENaC with Cl - or SO 2-4 in the bathing solution (relative to Cl - ) before and after treatment with trypsin (mean +/- S.E., n = 4, *p <.001).
82 66
83 67 Figure 3.5. Cl - alters Na + self-inhibition. A, representative current trace showing Na + self-inhibition in Xenopus oocytes expressing human αβγ ENaC with Cl -, SO 2-4, or SCN - in the bathing solution. The extracellular Na + concentration was 1 mm (white bars) or 116 mm (black bars), as indicated. 10 µm amiloride ( Amil ) was present in the bathing solution, as indicated by the gray bar, and current in the presence of amiloride by the gray line. B, Summary data of Na + self-inhibition [(peak current steady state current) / peak current ] with Cl -, SO 2-4, or SCN - bathing solutions (mean +/- S.E., n = 6-7, *p <.001). C, representative current traces showing Na + self-inhibition in Xenopus oocytes expressing wild-type or mutant (α H418A β R388A γ H396A ) ENaC in Cl - or SO 2-4 bathing solution, as indicated. The traces are scaled to the peak amiloride-sensitive currents. The vertical scale bar is 1.7 µa (αβγ Cl - ), 0.6 µa (αβγ SO 4 ), and 1.9 µa (α H418A β R388A γ H396A Cl - ). D, Na + selfinhibition in oocytes expressing αβγ ENaC, wild type ( wt ) or the indicated mutant subunits (α H418A, β R388A, or γ H396A ). Bathing solution contained Cl - or SO 2-4, as indicated. Data are mean +/- S.E., n = 4-16, *p <.01 versus wild-type in Cl -. E, ratio of amiloridesensitive current in SO 2-4 versus Cl - bathing solution in oocytes expressing wild-type or mutant ENaC, and incubated in 116 mm Na + ( + ) or 1 mm Na + ( - ) (mean +/- S.E., n = 3-14, *p <.01). Holding potential was -60 mv. Inward current was recorded for cells in 116 mm Na + and outward current in 1 mm Na +. F, fraction of amiloride-sensitive current inhibited versus extracellular Cl - concentration for wild-type or mutant (αβγ H233R ) ENaC (mean +/- S.E., n = 3-9, error bars are hidden by the data points).
84 68
85 69 Figure 3.6. ph modulates the effect of Cl - on ENaC. A, representative current trace at a holding potential of -60 mv in a Xenopus oocyte expressing human αβγ ENaC. ph of bathing solution was 8.5, 7.4, or 6.5, and it contained Cl - or SO 2-4, as indicated. 10 µm amiloride ( Amil ) was present in the bathing solution, as indicated by the gray bar. B and C, fraction of amiloride sensitive current inhibited by Cl - (replacing SO 2-4 ) in oocytes expressing wild-type (B) or α H418A β R388A γ H396A (C) ENaC (mean +/- S.E., n = 3-9). D, ph dose-response relationship for Na + self-inhibition of αβγ ENaC in Cl - or SO 2-4 bathing solution (mean +/- S.E., n = 4-14, *p <.01 versus Cl - ). In panels B-D, some of the error bars are hidden by the data points.
86 70
87 71 CHAPTER IV IDENTIFICATION OF ENAC INTER-SUBUNIT CL - INHIBITORY RESIDUES SUGGESTS A TRIMERIC αγβ CHANNEL ARCHITECTURE Preface This work appeared in the Journal of Biological Chemistry on February 25 th, I am co-author on this work with Dr. Snyder. I designed and performed the experiments, analyzed the data, and wrote the manuscript. Dr. Snyder designed experiments and wrote the manuscript. This work is a follow-up to the original description of chloride inhibition of ENaC activity. Herein we describe, in detail the putative chloride binding site and use these findings to determine the subunit architecture of heterotrimeric human ENaC. This work was the first in a line of experiments exploring the importance of ENaC intersubunit interactions. Abstract The extracellular domain of the epithelial Na + channel (ENaC) is exposed to a wide range of anion concentrations in the kidney. We have previously demonstrated that extracellular Cl - inhibits ENaC activity. To identify residues involved in Cl - inhibition, we mutated residues in the extracellular domain of α-, β- and γenac that are homologous to the Cl - binding site in ASIC1a and tested the effect of Cl - on the activity of ENaC expressed in Xenopus oocytes. We identified two Cl - inhibitory sites in ENaC. One is formed by residues in the thumb domain of αenac and the palm domain of βenac. Mutation of residues at this interface decreased Cl - inhibition and decreased Na + self-inhibition. The second site is formed by residues at the interface of the thumb
88 72 domain of βenac and the palm domain of γenac. Mutation of these residues also decreased Cl - inhibition, yet had no effect on Na + self-inhibition. In contrast, mutations in the thumb domain of γenac and palm of αenac had little or no effect on Cl - inhibition or Na + self-inhibition. The data demonstrate that Cl - inhibits ENaC activity by two distinct Na + dependent and Na + independent mechanisms that correspond to the two functional Cl - inhibitory sites. Furthermore, based on the effects of mutagenesis on Cl - inhibition, the additive nature of mutations, and on differences in the mechanisms of Cl - inhibition, the data support a model in which ENaC subunits assemble in an αγβ orientation (listed clockwise when viewed from the top). Introduction The epithelial Na + channel, composed of three homologous subunits (α-, β-, and γenac), functions as a pathway for Na + reabsorption across epithelia in the kidney collecting duct, lung, distal colon, and sweat duct (53, 95). In this role, the channel is critical for the maintenance of Na + homeostasis and control of the composition and quantity of fluid on the apical membrane of these epithelia. ENaC mutations, and defects in its regulation, cause inherited forms of hypertension and hypotension (3, 95), and may contribute to the pathogenesis of lung disease in cystic fibrosis (10). At the apical membrane of epithelia, ENaC is exposed to highly varied extracellular environments. Through its large extracellular domain, ENaC detects concentration changes in a number of molecules, producing changes in ENaC gating. This mechanism may fine-tune ENaC activity to respond to diverse challenges. For example, the urine Na + concentration in the kidney collecting duct ranges between ~1-
89 mm under conditions of volume depletion and volume excess. Binding of Na + to ENaC extracellular domains inhibits channel gating through a process known as Na + selfinhibition (13, 39, 51). ph also fluctuates widely (e.g in kidney collecting duct (96)); by modulating Na + self-inhibition, acidic ph increases and alkaline ph reduces ENaC activity (85). In Chapter III, we demonstrated that extracellular Cl - also modulates ENaC activity (97). As the predominant anion in the extracellular fluid, the concentration of Cl - varies over a wide range (1-100 mm in kidney collecting duct), in parallel with changes in Na + (96). Within this concentration range, Cl - inhibits ENaC gating through two distinct mechanisms (97). First, Cl - enhances Na + self-inhibition of the channel. Second, Cl - inhibits ENaC through a direct effect that is independent of extracellular Na +. Crystallization of ASIC1a has begun to provide structural insights into the DEG/ENaC family of ion channels. ASIC1a assembles as a homotrimer (43, 46), suggesting that other members of the family are also trimers. Surprisingly, the structure revealed three Cl - ions bound to the extracellular domains (43). The Cl - binding sites are located at each of the three interfaces between the subunits, formed by residues at the base of the thumb domain of one subunit (Arg-310, Glu-314) and the palm domain of the opposing subunit (Fig. 1, A and B show ASIC1a looking down from the top). Recent work found that Cl - binding slowed the rate of ASIC1a desensitization when the channel was activated by protons (98). Because ASIC1a and ENaC share a high degree of sequence similarity, including a number of conserved cysteine residues that form disulfide bonds, the ASIC1a crystal structure has been used as a starting point to investigate structure-function relationships
90 74 in the extracellular domain of ENaC (97, 99). Using this approach in Chapter III, we identified homologous residues in the thumb domains of α- (His-418) and βenac (Arg- 388) that contribute to ENaC inhibition by Cl - (97). Importantly, the equivalent residue in γenac (His-396) did not contribute. These findings support the possibility that Cl - regulates ENaC through sites that are analogous to the Cl - binding sites identified in ASIC1a. Here, we test whether ENaC palm domain residues also contribute to Cl - inhibition. Furthermore, because the residues that form the Cl - inhibitory sites are predicted to lie at the interface between two neighboring subunits, our approach provides insight into the subunit arrangement of the ENaC trimeric channel complex. Experimental Procedures DNA Constructs cdnas for human α-, β-, and γenac in pmt3 were cloned as previously described (2, 100). Mutations were generated by site-directed mutagenesis (QuickChangeII; Stratagene) and sequenced in the University of Iowa DNA Core. Expression and Whole-Cell Electrophysiology in Xenopus oocytes Oocytes were harvested from albino Xenopus laevis females and manually defolliculated following a one-hour treatment with 0.75 mg/ml Type IV collagenase (Sigma) in Ca 2+ free ND-96 (96 mm NaCl, 2 mm KCl, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH). Following nuclear injection of cdnas encoding α-, β-, and γenac (0.02 µg/µl each), cells were incubated at 18 C in modified Barth s saline (88 mm NaCl, 1 mm KCl, 0.33 mm Ca(NO 3 ) 2, 0.41 mm CaCl 2, 0.82 mm MgSO 4, 2.4 mm NaHCO 3, 10 mm HEPES, 50 µg/ml gentamycin sulfate, 10 µg/ml sodium penicillin, 10 µg/ml streptomycin sulfate, ph adjusted to 7.4 with NaOH) for 20-24h prior to study. Oocytes were voltage clamped
91 75 (two-electrode voltage clamp) and currents were amplified with an Oocyte Clamp OC- 725C (Warner Instruments), digitized with a MacLab/200 interface (ADInstruments), and recorded and analyzed with Chart software (ADInstruments). Unless otherwise noted, recordings were done at 60 mv in a 116 mm NaCl solution (116 mm NaCl, 2 mm KCl, 0.4 mm CaCl 2, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH). Low Na + solutions (1 mm NaCl, 115 mm N-Methyl-D-glucamine Cl, ph adjusted with HCl) were used as indicated in the figure legends. Low Cl - solutions were prepared by replacing the anion of the sodium salt with SO 2-4 and D-mannitol (to balance osmolarity), SCN -, I -, or Br - ; the composition of the low Cl - solutions was otherwise identical to that of 116 NaCl Ringers. Amiloride-sensitive current was determined by adding 10 µm amiloride to the bathing solution. The Cl - -induced changes in amiloride-sensitive current were calculated as the fold increase/decrease relative to the baseline current in 116 mm NaCl just prior to each test solution application. This was done to reduce the effect of time dependent current run-down. Na + self-inhibition was measured by rapidly changing the bathing solution from low sodium (1 mm NaCl) to high sodium (116 mm NaCl) and quantitated as [(peak current steady state current) / peak current]. To account for day-to-day variation, values were normalized to values recorded from cells expressing wild-type ENaC on the same day. All values are reported as averages +/- S.E. Statistical significance was calculated using a two-tailed Students T-test (p<0.05).
92 76 Results ENaC palm domain residues participate in Cl - inhibition To identify ENaC palm domain residues that contribute to inhibition by Cl -, we did a sequence alignment between ASIC1a and α-, β-, and γenac in the region of ASIC1a Lys-212 (Fig. 4.1C). The alignment was facilitated by adjacent conserved Gly residues as well as a highly conserved upstream Cys-containing motif. Lys-212 is not conserved in ENaC. The corresponding residues are Asn-288 in βenac, Met-321 in αenac, and Met-299 in γenac. To test their potential role in ENaC regulation by extracellular Cl -, we mutated these residues and quantitated ENaC inhibition by Cl - in Xenopus oocytes. We first focused on βenac. Fig. 4.2A shows representative traces from cells expressing human α-, β-, and γenac. For wild-type ENaC (gray trace), addition of Cl - to the extracellular bathing solution (replacing SO 2-4, which does not inhibit ENaC (97)) decreased amiloride-sensitive current 55.4%, consistent with the work presented in Chapter III (97). Mutation of Asn-288 to Ala in βenac (coexpressed with wild-type α- and γenac) reduced ENaC inhibition by Cl - (Fig. 4.2A-4.2C). Mutation of this residue to Cys or Lys decreased Cl - inhibition to a similar extent (Fig. 4.2C). These findings support two conclusions. First, Asn-288 in βenac contributes to inhibition by extracellular Cl -. Second, at this position, lysine (the amino acid at the equivalent position in ASIC1a) is not a functionally conservative substitution. To test the role of the equivalent palm domain residues in α- and γenac, we mutated them to Lys (αm321k and γm299k). In γenac, mutation of Met-299 to Lys resulted in a significant decrease in Cl - inhibition (Fig. 4.2D and 4.2F). In contrast,
93 77 mutation of αenac Met-321 to Lys produced only a small decrease in Cl - inhibition (Fig. 4.2E and 4.2F). These results indicate that residues located in the palm domains of β- and γenac contribute to Cl - inhibition, whereas the palm domain of αenac plays a lesser role. In Chapter III, we identified thumb domain residues in α- and β-, but not γenac, that contribute to Cl - inhibition (97). Together, these findings suggest that Cl - inhibits ENaC predominately through two Cl - inhibitory sites, in contrast to the three sites identified in the ASIC1a crystal structure. Mutations of γenac Met-299 alter anion inhibition selectivity If palm domain residues contribute to anion binding, analogous to the equivalent residues in ASIC1a, then mutation of these residues might alter the anion selectivity of ENaC inhibition. To test this possibility, we focused on Met-299 in γenac. Fig. 4.3A and 4.3B show the effects of anion substitutions on wild-type ENaC current. Br - inhibited ENaC to the greatest extent (compared to SO 2-4 ), followed by I - and Cl -, whereas SCN - did not inhibit ENaC (97). Thus, the selectivity sequence for inhibition of wild-type ENaC is Br - > I - > Cl - >> SCN -. Mutation of Met-299 to Lys reduced ENaC inhibition by Cl -, I -, and Br - (Fig. 4.3B and 4.3D). In addition, this mutation altered anion selectivity of inhibition, such that Br - and I - decreased ENaC current to an equivalent degree (Br - = I - > Cl - >> SCN - ) (Fig. 4.3C and 4.3D). Mutation of Met-299 to another basic residue, Arg, had the same effect as Lys on relative Br - /I - selectivity (Fig. 4.3E and 4.3F). However, this mutation had two additional effects. First, it allowed SCN - to
94 78 inhibit ENaC (I SCN- < I SO4-, p<.001), albeit less than Cl - (Br - = I - > Cl - > SCN - ). Second, it did not reduce, but slightly enhanced ENaC inhibition by Cl -, Br -, and I - (Fig. 4.3D and 4.3F). Together, these data suggest that both electrostatic and steric factors at residue 299 influence selectivity of anion inhibition, which is consistent with a role for this residue in anion binding. ENaC subunits are arranged in an αγβ orientation Based on our findings and the ASIC1a crystal structure, we hypothesized that the Cl - inhibitory sites are positioned at the interfaces between ENaC subunits, with palm domain residues from one subunit paired with thumb domain residues from the adjacent subunit. A key question is which ENaC subunit provides the palm domain and which subunit provides the thumb domain at each Cl - site? There are two possible structural arrangements for the three ENaC subunits. When viewed from the top, they could adopt an αγβ (Fig. 4.4A) or an αβγ (Fig. 4.4B) orientation (listed clockwise). To determine the subunit orientation, we made two assumptions. First, we assumed that the palm domain residues that contribute to Cl - inhibition would be paired with thumb domain residues that also contribute to Cl - inhibition. Second, we assumed that the palm domain and thumb domain that had little or no effect on Cl - regulation would be paired together. Using these assumptions, we examined the pairing of palm and thumb domain residues predicted by the two models. We started with the βenac palm domain residue Asn-288 which, when mutated, reduced Cl - inhibition (Fig. 4.2A and 4.2B). In the αγβ model, this residue is paired with the αenac thumb domain residue His-418 (Fig. 4.4A). We previously reported that mutation of this residue reduced Cl - inhibition (97).
95 79 Conversely, in the αβγ model, Asn-288 pairs with the γenac thumb domain residue His- 396 (Fig. 4.4B), which had no effect on ENaC Cl - inhibition (97). Thus, these data support the αγβ orientation. At a second interface in the αγβ model, Met-299 in γenac is paired with Arg-388 in βenac (Fig. 4.4A). Consistent with this model, mutation of both residues reduced Cl - inhibition (Fig. 4.2 and (97)). The αγβ model also places the residues that had little or no effect on Cl - inhibition (α Met-321 and γ His-396) together at the third interface (Fig. 4.4A). Contrary to the data, the αβγ model would pair these non-functional residues together with residues that contribute to Cl - inhibition (Fig. 4.4B). Taken together, the data suggest that ENaC assembles in an αγβ orientation. Mutations of residues in separate Cl - sites are additive To further test whether the ENaC subunits form an αγβ heterotrimer, we combined mutations at the two Cl - inhibitory sites. If each site has an independent effect on ENaC activity, we predict that combining a mutation from the α-β interface with a mutation in the β-γ interface will produce an additive decrease in Cl - inhibition. Fig. 4.5A shows two examples. In β N288K,R388A, we simultaneously mutated the β subunit palm domain (predicted location at the α-β interface) and thumb domain (predicted location at the β-γ interface). When co-expressed with wild-type α- and γenac, this double mutation greatly decreased the effect of extracellular Cl - on ENaC current (Fig. 4.5A and 4.5B). A similar decrease in Cl - inhibition was observed when we combined mutations in the thumb domain of the α-β interface (α H418A ) with the palm domain of the
96 80 β-γ interface (γ M299K ) (Fig. 4.5A and 4.5B). In Fig. 4.5B, we quantitated the results for each of the single and double mutations at the α-β and β-γ interfaces; single mutations are indicated by white circles (α-β interface) and black circles (β-γ interface), the double mutations are indicated by grey bars, and predicted additive values are indicated by vertical black lines. When mutations from the α-β and β-γ interfaces were combined, there was an additive reduction in Cl - inhibition relative to the corresponding individual mutations (Fig 4.5B, compare grey bars to vertical black lines). In contrast, if we mutated two residues predicted to lie at the same interface (α-β interface), αh418a and βn288k, the effects were not additive (Fig. 4.5B). These data indicate that the two Cl - inhibitory sites have independent effects on ENaC regulation and provide support for the αγβ subunit model. Different Cl - sites alter ENaC activity through distinct mechanisms As a third test of the αγβ model, we examined the mechanisms by which Cl - inhibits ENaC. In Chapter III, we found that inhibition occurs through two different mechanisms. One is dependent on extracellular Na + (Cl - enhanced Na + self-inhibition) whereas the second is independent of Na + (97). We hypothesized that these two mechanisms might correspond to the two Cl - inhibitory sites, one site modulating Na + self-inhibition and the other site regulating ENaC in a Na + -independent manner. To test this hypothesis, we investigated the effect of Cl - site mutations on Na + self-inhibition. Fig. 4.6A shows representative current traces for mutations predicted to be in the α-β interface. We measured ENaC Na + self-inhibition and Cl - inhibition in the same
97 81 cells. At the start of the trace, addition of Na + to the extracellular bathing solution generated a peak current that rapidly decreased to a lower steady-state level, as a result of Na + self-inhibition. In Fig. 4.6B, we plotted the fraction of current inhibited by Na + (relative to wild-type ENaC). α H418A and β N288K each reduced Na + self-inhibition ( / and / relative to wild-type, respectively). In the second part of the trace, Cl - in the bathing solution was replaced with SO 2-4 to measure Cl - inhibition (Fig. 4.6A). Both mutations decreased ENaC Cl - inhibition, similar to our data in Fig Because mutation of α H418A and β N288K had congruent effects on Na + self-inhibition and Cl - inhibition, the data suggest that they are located at the same interface. In Figs. 4.6C and 4.6D, we tested mutations predicted to be located at the β-γ interface. Although β R388A and γ M299K each reduced ENaC inhibition by Cl -, they had no effect on Na + self-inhibition ( / and / , respectively). This supports the idea that β R388A and γ M299K are located at a common interface. Likewise, mutations at the γ-α interface, α M321K and γ H396A (mutations which had minimal effect on Cl - inhibition), did not alter Na + self-inhibition ( / and / , respectively, Fig. 4.6E). Together, the data indicate that the two Cl - sites inhibit ENaC through different mechanisms. At the α-β interface, Cl - enhances Na + self-inhibition of ENaC, whereas at the β-γ interface, Cl - inhibits ENaC through a mechanism that is independent of Na + self-inhibition. In addition to providing mechanistic insights, these results also confirm our hypothesis that ENaC adopts an αγβ channel architecture. This is the only model that pairs the residues that reduced Na + self-inhibition (α H418A with β N288K ) and the residues that had no effect on Na + self-inhibition (β R388A and γ M299K ). In the alternative αβγ
98 82 structure, α H396, a residue that functions through Na + self-inhibition, would be paired with γ M299, a residue that is Na + -independent (Fig. 4.4B and Fig. 4.6), a possibility not supported by the data. Discussion In this work, we identified palm domain residues that participate in ENaC inhibition by extracellular Cl -. One residue is located in βenac (Asn-288) and the second is at the equivalent position in γenac (Met-299). In Chapter III, we identified thumb domain residues in α (H418) and β (R388) ENaC that also contribute to Cl - inhibition. Together, the data suggest that ENaC has two predominant Cl - inhibitory sites which are positioned at the interfaces between ENaC subunits. The conclusion that the Cl - inhibitory sites are formed by residues in two adjacent subunits is consistent with the previously reported ASIC1a crystal structure (43). We exploited this observation to deduce the subunit orientation of the ENaC channel structure. Based on the effects of mutagenesis on Cl - inhibition, the additivity of mutations, and on differences in the mechanisms of Cl - inhibition, the data support an αγβ orientation (listed clockwise when viewed from the top). Moreover, the data suggest that assembly of the ENaC subunits is not stochastic, but occurs in a favored configuration. Although unlikely, we can t completely exclude the existence of a population of αβγ channels that are not modulated by changes in extracellular Cl -. In the ASIC1a crystal structure, Cl - is coordinated in part through its interaction with the positively charged side chain of Lys-212 in the palm domains (43, 98). It is interesting that the amino acids at the equivalent positions in ENaC (β N288 and γ M299 ) do
99 83 not carry a positive charge. Although it is clear from our mutagenesis data that these residues are critical for ENaC inhibition by Cl -, their specific role in this regulation is unclear. They may participate directly in Cl - binding. Direct interactions could occur on the backbone, as demonstrated by structural data from other halide binding sites (9, 101) or, analogous to their role in ASIC1a, direct interactions could occur with the residue side chain. Consistent with this possibility, we found that γ M299 mutations altered the selectivity sequence for anion inhibition. In particular, mutation to Arg increased Cl - inhibition and created a binding site for SCN -. This is analogous to previous work on deoxyhemoglobin, where mutation of Trp-37 to Arg created a novel anion-binding site (27). Our data, in combination with structural data from the literature, suggest that palm domain residues directly participate in Cl - binding. However, in the absence of an ENaC crystal structure, we cannot completely exclude the possibility that the residues we identified influence anion binding indirectly. In Chapter III, we demonstrated that Cl - inhibited ENaC through two different mechanisms (97). First, Cl - reduced ENaC current by facilitating Na + self-inhibition of the channel. Second, Cl - also reduced ENaC current through a direct mechanism that was independent of extracellular Na +. Our current studies provide a structural basis for these two mechanisms. We found that Cl - modulation of Na + self-inhibition occurred at the α-β interface, whereas Cl - inhibited ENaC through a direct effect at the β-γ interface. It is interesting that residues in βenac contribute to both Cl - inhibitory sites. This raises the possibility that the Cl - -induced conformational change might be restricted to the β subunit. However, we think it is more likely that conformational changes are distributed throughout all three subunits. This notion is supported by our findings that Cl - regulates
100 84 ENaC through two distinct mechanisms, coupled with the additive effect of mutations at the two sites. Although the three ENaC subunits share sequence similarity, there is a distinct lack of symmetry in the contribution of each subunit to ENaC regulation by Cl -. Of the three interfaces between ENaC subunits, two play a predominant role in this regulation. Additionally, at these two interfaces, the mechanisms by which Cl - regulates ENaC differ. There are several other examples of asymmetry in the regulation of ENaC gating. For example, ENaC is activated by proteolytic cleavage of α- and γ-, but not βenac (12, 28, 102). Also, mutation of an extracellular histidine in γenac decreases Na + selfinhibition, but mutation of the equivalent histidine in αenac has the opposite effect (39). Mutations at the DEG position also have varying effects when introduced in each ENaC subunit (103, 104). Such asymmetry might expand the range of conformational changes available to fine-tune channel gating. Even though ENaC is a constitutively active channel, there is a growing list of molecules that modulate its function including, but not limited to, Na +, Cl -, H +, Zn 2+, and Ni 2+ (13, 40, 85, 97). In this regard, it is clear that the extracellular domain functions as a sensor to detect the concentrations of these molecules in a variety of fluids, including urine, airway surface liquid, and sweat. This may allow ENaC to rapidly respond to a variety of challenges that threaten to disrupt Na + homeostasis. Our current findings explain in part the mechanisms by which ENaC gating is modulated by extracellular Cl -. Moreover, they provide a mechanistic basis to explain functional interactions between ENaC regulation by Cl - and Na +, the two principal components of extracellular fluids.
101 85 We speculate that defects in this regulation may contribute to the pathogenesis of hypertension and other disorders of Na + transport.
102 86 Figure 4.1. Sequence alignment of ASIC1a with α-, β- and γenac. A, ribbon structure of chicken ASIC1a (2QTS) shown perpendicular to the plane of the membrane from an extracellular prospective. B, model of ASIC1a subunit arrangement. Dotted lines represent the thumb domain, dashed lines represent the palm domain. Approximate location of Cl - is depicted by an open circle located between the thumb and palm domains of two adjacent subunits. C, partial sequence alignment of the palm and thumb domains of chicken ASIC1a with human α-, β-, and γenac using the Clustal W method. ASIC1a Cl - coordinating residues are indicated with an asterisk. Potential ENaC Cl - coordinating residues identified by sequence alignment are in bold.

103 87
104 88 Figure 4.2. Effect of ENaC palm domain mutations on Cl - inhibition. A, D and E, representative recordings of current versus time from Xenopus oocytes expressing αβγenac (grey trace, A, C and D, scale bars: vertical 1 µa, horizontal 10 sec), αβ N288A γ (A, black trace, scale bars: vertical 1.5 µa, horizontal 15 sec), αβγ M299K (D, black trace, scale bars: vertical 1.7 µa, horizontal 15 sec), α M321K βγ (E, black trace, scale bars: vertical 1.7 µa, horizontal 14 sec). Cl - Inhibition was measured by exchanging SO 2-4 for Cl - in extracellular bathing solution. 10 µm amiloride was added at the end of the recording to determine the amount of ENaC current. All traces are normalized to amiloride sensitive current in the presence of Na 2 SO 4. B, summary data of current relative to SO 2-4 for cells expressing Wt or β N288A ENaC (n = 6, *, p < 0.001). C, summary data of fraction of amiloride-sensitive current inhibited by Cl - (relative to SO 2-4 ) for various βenac palm domain mutations relative to wild-type ENaC (mean ± S.E., n = 6-11, *, p < 0.001). F, summary data of Cl - inhibition for α M321K (mean ± S.E., n = 13, *, p < 0.001).
105 89
106 90 Figure 4.3. γenac Met-299 mutations alter selectivity of anion inhibition. A, C and E, representative recordings of current versus time from Xenopus oocytes expressing αβγenac (A), αβγ M299K (C), or αβγ M299R (E). Anion selectivity was measured by replacing extracellular Cl - with SCN -, SO 2-4, I -, or Br -. Traces are scaled to amiloride sensitive current for wild-type ENaC in the presence of Cl -. B, D and F, summary data of fold change in amiloride sensitive current for the indicated anion relative to SO 2-4 for cells expressing αβγenac (B), αβγ M299K (D), or αβγ M299R (F). Data are mean ± S.E. (n = 6-9, *, p < 0.001).
107 91
108 92 Figure 4.4. Potential ENaC subunit arrangements. A trimeric assembly could result in an αγβ (A) or αβγ (β) channel (listed clockwise, viewing channel from the top). Dotted lines represent the thumb domain interface. Dashed lines represent the palm domain interface. Approximate locations of putative Cl - inhibitory residues are labeled accordingly; αh = His-418, αm = Met-321, γh = His-396, γm = Met-299, βr = Arg-388, βn = Asn-288. Approximate locations of chloride ions are identified with closed circles. Residues that had large effects on Cl - inhibition are identified with an open box.
109 93
110 94 Figure 4.5. Additivity of Cl - inhibitory site mutations. A, representative recordings of current versus time from Xenopus oocytes expressing αβγenac (grey trace, scale bars: vertical 1 µa, horizontal 10 sec), αβ N288K,R388A γ (black trace, scale bars: vertical 1.6 µa, horizontal 13 sec) or α H418A βγ M299K (black trace, scale bars: vertical 1.75 µa, horizontal 13 sec). Cl - Inhibition was measured by replacing SO 2-4 in the bathing solution with Cl µm amiloride was added at the end of the recording to quantitate ENaC current. All traces are normalized to amiloride-sensitive current in the presence of SO 2-4. B, summary data showing relative Cl - inhibition for all possible combinations of double mutations (grey bars) at the α-β and β-γ interfaces. Individual α-β and β-γ interface mutations shown as white or black circles, respectively. Predicted values for perfectly additive double mutations are indicated with vertical black lines. Data are mean ± S.E. (n = 4-14). All values for single or double mutations are statistically significant when compared to wild-type ENaC (p < 0.001). αh418a βn288k double mutation is statistically significantly different from the predicted additive value (n = 4, *, p = 0.006).
111 95
112 96 Figure 4.6. Differential effects of Cl - site mutations on Na + self-inhibition. A and C, representative recordings of current versus time from Xenopus oocytes expressing αβγenac (A and C, light grey traces, scale bars: vertical 1 µa, horizontal 10 sec), αβ N288K γ (A, dark grey trace, scale bars: vertical.3 µa, horizontal 9 sec), α H418A βγ (A, black trace, scale bars: vertical 1 µa, horizontal 10 sec), αβ R388A γ (C, dark grey trace, scale bars: vertical 1.22 µa, horizontal 8 sec) or αβγ M299K (C, black trace, scale bars: vertical 1 µa, horizontal 9 sec). Na + self-inhibition was measured in the presence of extracellular Cl - by rapidly changing from low (1 mm Na +, 115 mm NMDG) to high Na + (116 mm Na + ) extracellular solutions. Cl - Inhibition was measured by replacing extracellular Cl - with SO 2-4. Traces are scaled relative to amiloride sensitive peak current. B, D, and E, summary data showing Na + self-inhibition relative to wild-type ENaC for single mutations at the predicted α-β interface (B), β-γ interface (D), and γ-α interface (E). Data are mean ± S.E. (n = 3-10, *, p < 0.001).
113 97
114 98 CHAPTER V IDENTIFICATION OF RESIDUES IN THE EXTRACELLULAR DOMAIN OF HUMAN ENAC THAT CONTRIBUTE TO H + REGULATION OF CHANNEL ACTIVITY PROVIDE INSIGHT INTO CONFORMATIONAL CHANGES ASSOCIATED WITH CHANNEL GATING Preface I designed and performed the experiments, analyzed the data, and wrote the manuscript. Dr. Snyder aided in designing experiments. This work is a follow-up to the original description of ph regulation of ENaC activity. Herein, we describe residues in the extracellular domains of α-, β-, and γenac that contribute to the ph sensitivity of human ENaC. In doing so, we determined that a subset of these residues are located at the interface of adjacent subunits. The apparent intersubunit interactions agree with our previous finding that ENaC preferentially forms an αγβ heterotrimer. Moreover, identification of these intersubunit interfaces provides insight into extracellular domain conformational changes associated with channel gating. Abstract A growing body of evidence suggests that the extracellular domain of ENaC functions as a sensor that fine tunes channel activity in response to changes in the extracellular environment. We previously demonstrated that H + have dual effects on ENaC activity; they increase ENaC activity by decreasing Na + self-inhibition, and inhibit ENaC activity by increasing Cl - inhibition. In the current work, we investigated the mechanisms by which H + stimulate ENaC. We found that rat ENaC is less sensitive to
115 99 H + than human ENaC, an effect mediated by the γ subunit. Our strategy was to mutate acidic residues in the extracellular domain of human γenac that are not conserved in rat γenac. We expressed the mutant γenac cdnas (with wild type α- and βenac) in Xenopus oocytes and tested the effect of ph changes on amiloride-sensitive Na + current (by TEVC at -60 mv). We identified a group of 7 residues in the extracellular domain of γenac (D164, Q165, D166, E292, E335, H439, and E455) that, when individually mutated to Ala, decreased H + activation of ENaC. Based on the ASIC1 crystal structures (2QTS and 3HGC), a subset of these residues appear to be located at an intersubunit interface. Mutating the sites equivalent to one of these residues, γ E455, in αenac (α K477 ) and βenac (β E446 ) increased and decreased the response to acidic ph, respectively. Combining the seven mutations in γenac with β E446A generated a channel that was not activated by acidic ph. The data demonstrate that residues in human β- and γenac are required for regulation by ph. To further understand the mechanism of H + activation, we identified the residues at the opposing interface. Based on the crystal structure, we predict that α K477, β E446, and γ E455 interact with residues in the wrist domain of adjacent subunits; β V85, γ V87 and α L120, respectively. To test this, we substituted these residues with Cys. We found that covalent modification of Cys mutations at β V85 and α L120 with MTSET and MTSES altered ENaC activity. Furthermore, treatment with long cysteine reactive cross-linking reagents increased current in cells expressing α K477C β V85C γenac. Intriguingly, treatment with either long or short cross-linking reagents disrupted channel gating, although short reagents do not increase current. The data provides a potential link between ph sensing, extracellular domain conformational changes, and channel gating.
116 100 Introduction The epithelial Na + channel is composed of three homologous subunits (α-, β-, and γenac). Each subunit has relatively short cytoplasmic N- and C-termini, leaving the bulk of the protein exposed to the extracellular environment. ENaC functions as a pathway for Na + reabsorption across epithelia in the kidney collecting duct, lung, distal colon, and sweat duct (53, 95). In this role, the channel is critical for the maintenance of Na + homeostasis and control of the composition and quantity of fluid on the apical membrane of these epithelia. ENaC mutations, and defects in its regulation, cause inherited forms of hypertension and hypotension (3, 95), and may contribute to the pathogenesis of lung disease in cystic fibrosis (10). The extracellular domain of ENaC is exposed to highly variable environments. A growing body of evidence suggests that the large extracellular domain detects concentration changes in a number of molecules, producing changes in ENaC gating. As a complement to long-term regulation by changes in channel expression, this mechanism may define normal physiology by providing a fast, fine-tuning of ENaC activity in response to a relentless assault of diverse extracellular challenges. For example, urine Na +, Cl - and ph change dynamically. Na + concentration in the kidney collecting duct ranges between ~1-150 mm under conditions of volume depletion and volume excess, respectively. Exposure of the ENaC extracellular domain to high Na + concentrations inhibits channel gating through a process known as Na + self-inhibition (13, 39, 51). Cl - concentration in the collecting duct ranges between ~1-120 mm. Changes in Cl - generally fluctuate along with changes in Na + concentration. Under conditions of hypovolemia, luminal Cl - is low, while under conditions of hypervolemia, luminal Cl - is
117 101 high. Increasing extracellular Cl - decreases ENaC activity through a mechanism that is, in part, convergent with Na + self-inhibition (Chapter II and III, (97)). Luminal H + concentration changes between ph in kidney collecting duct (96). Contrary to changes in Na + and Cl -, high H + concentration is associated with hypovolemia while low H + concentrations are associated with hypervolemia. Decreasing extracellular ph has dual, opposing, effects on ENaC activity depending on the balance of Na + and Cl - (described in Chapters II and III). Acidic ph decreases Na + self-inhibition (85) (which increases ENaC activity) and increases Cl - inhibition (97) (which decreases ENaC activity). Intriguingly, these changes in Na +, Cl -, and H + concentrations alter ENaC activity in the direction that would be predicted to restore normal volume conditions increasing ENaC activity under hypovolemic conditions and decreasing ENaC activity under hypervolemic conditions. In Chapter II, we demonstrated that extracellular ph regulates human ENaC activity, while rat ENaC is unresponsive to ph in the presence of high extracellular NaCl (85). In the current work, we take advantage of the species differences and focus on the stimulatory effects of H + (by performing experiments under low extracellular Cl - conditions). We use site directed mutagenesis and electrophysiological techniques to investigate titratable residues in the extracellular domain of human γenac that are not present in the extracellular domain of rat γenac. We hypothesize that mutation of these residues in human γenac will decrease activation of human αβγenac in response to acidic ph conditions.
118 102 Experimental Procedures DNA Constructs cdnas for human α-, β-, and γenac in pmt3 were cloned as previously described (2, 100). Mutations were generated by site-directed mutagenesis (QuickChangeII; Stratagene) and sequenced in the University of Iowa DNA Core. Expression and Whole-Cell Electrophysiology in Xenopus oocytes Oocytes were harvested from albino Xenopus laevis females and manually defolliculated following a one-hour treatment with 2.4 units FALGPA activity per ml Type IV collagenase (Sigma) in Ca 2+ free ND-96 (96 mm NaCl, 2 mm KCl, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH). Following nuclear injection of cdnas encoding α-, β-, and γenac (0.02 µg/µl each), cells were incubated at 18 C in modified Barth s saline (88 mm NaCl, 1 mm KCl, 0.33 mm Ca(NO 3 ) 2, 0.41 mm CaCl 2, 0.82 mm MgSO 4, 2.4 mm NaHCO 3, 10 mm HEPES, 50 µg/ml gentamycin sulfate, 10 µg/ml sodium penicillin, 10 µg/ml streptomycin sulfate, ph adjusted to 7.4 with NaOH) for 20-24h prior to study. Oocytes were voltage clamped (two-electrode voltage clamp) and currents were amplified with an Oocyte Clamp OC-725C (Warner Instruments), digitized with a MacLab/200 interface (ADInstruments), and recorded and analyzed with Chart software (ADInstruments). Unless otherwise noted, recordings were performed at -60 mv in a 58 mm Na 2 SO 4 solution (58 mm Na 2 SO 4, 58 mm D-mannitol, 2 mm KCl, 0.4 mm CaCl 2, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH). High Na + (116 mm NaCl, 2 mm KCl, 0.4 mm CaCl 2, 1 mm MgCl 2, 5 mm HEPES, ph adjusted to 7.4 with NaOH) and Low Na + solutions (1 mm NaCl, 115 mm N-Methyl-D-glucamine, 2 mm KCl, 0.4 mm CaCl 2, 1 mm MgCl 2, 5 mm HEPES, ph adjusted with HCl) were used as indicated in the figure legends. Amiloride-sensitive current was determined by adding 10 µm amiloride to the
119 103 bathing solution. The ph-induced changes in amiloride-sensitive current were calculated as the fold increase/decrease relative to the baseline current in ph 7.4 Na 2 SO 4 Ringers just prior to each test solution application. This was done to reduce the effect of time dependent current run-down. Na + self-inhibition was measured by rapidly changing the bathing solution from low sodium (1 mm NaCl) to high sodium (116 mm NaCl) and quantitated as [(peak current steady state current) / peak current]. Cl - inhibition was measured by rapidly changing the bathing solution from high chloride (116 mm NaCl) to low chloride (58 mm Na 2 SO 4 ) and quantitated as fold change in amiloride sensitive current. To account for day-to-day variation, values were normalized to values recorded from cells expressing wild-type ENaC on the same day. All values are reported as averages +/- S.E. Statistical significance was calculated using a two-tailed Students T- test (p<0.05). Results Human and rat ENaC respond differently to changes in extracellular ph in the absence of Cl - To test the hypothesis that titratable residues in the extracellular domain of ENaC contribute to increased channel activity in response to low ph, we took advantage of the species dependent differences in ph sensitivity (Initially described in Chapter II, (85)). We measured the change in activity of human and rat ENaC in response to extracellular ph under low extracellular Cl - conditions. This experimental system allows us to simplify the response to acidic ph and focus only on the Cl - -independent increase in ENaC activity in response to low extracellular ph (Chapter III, (97)). We measured
120 104 changes in ENaC activity over a ph range from 9.0 to 5.25 and found, consistent with previous results, that alkaline ph decreased human ENaC activity (Fig 5.1A, black trace, compare ph 7.4 baseline to ph 9.0 or 8.5) and acidic ph increased human ENaC activity (Fig 5.1A, black trace, compare ph 7.4 baseline to ph 7.0, 6.5, 5.5 or 5.25). However, rat ENaC activity did not change in response to extracellular ph over a range of ph 9.0 to ph 6.5 (Fig 5.1A, grey trace, compare ph 7.4 baseline to ph 9.0, 8.5, 7.0 or 6.5). Acidic conditions below ph 5.5 increased rat ENaC activity, albeit to a lesser degree than human ENaC (Fig 5.1A, grey trace, compare ph 7.4 baseline to ph 5.5 and 5.25). Figure 5.1B shows the change in ENaC activity in response to changes in extracellular ph relative to channel activity at ph 7.4. At ph 9.0, rat ENaC activity is not decreased while human ENaC activity is decreased by approximately 29%. At ph 5.25, human ENaC activity is increased by 91% relative to ph 7.4, while rat ENaC activity is only increased by 55%. The ph dose reponse curve for rat ENaC is strongly shifted to the right (Fig 5.1B). The difference in ph sensitivity between human and rat ENaC provides a strategy to identify the residues that contribute to changes in channel activity in response to changes in extracellular ph. Human γenac is necessary but not sufficient for ph sensitivity Normal ENaC function requires expression of α-, β-, and γenac subunits. To narrow our search for the residues that contribute to human ENaC ph sensitivity, we measured the change in ENaC activity in response to ph in cells expressing a combination of human and rat ENaC subunits. Given the decreased ph sensitivity of rat
121 105 ENaC, we predicted that co-expression of rat ENaC subunits with human ENaC subunits would decrease ph sensitivity relative to a channel composed of only human ENaC subunits. We began by expressing rat βenac with human α- and γenac. Figure 5.2A shows representative traces of current versus time recorded from Xenopus oocytes. Replacing human βenac with rat βenac had no effect on the ph dose response relative to cells expressing human αβγenac (Fig 5.2A & 5.2B). When human αenac was replaced with rat αenac, acidic ph produced a larger increase in current than cells expressing only human ENaC subunits (Fig 5.2C & 5.2D). However, when human γenac was replaced with rat γenac, ENaC activation by acidic ph was effectively eliminated (Fig 5.2E & 5.2F). This data demonstrates that human γenac is necessary for human αβγenac channels to respond to changes in ph. To test if human γenac is sufficient for the ph response, we expressed rat α- and βenac with human γenac. The ph dose response relationship for channels expressing this combination of two rat and one human subunit was very similar to channels expressing only rat αβγenac (Fig 5.3A & 5.3B). This demonstrates that human γenac was not sufficient to restore ph sensitivity in rat ENaC. Therefore, the data suggest that more than one subunit may be involved in the ph response seen in human αβγenac. Mutation of acidic residues in the extracellular domain of human γenac decrease the ph response We focused our efforts on identifying the residues necessary for ph sensitivity in human ENaC. Because human γenac is necessary for for ph sensitivity, we hypothesize that differences between the extracellular domains of human and rat ENaC contribute to
122 106 the decreased response to extracellular acidification seen in cells expressing rat γ- and human αβenac. Figure 5.4 shows a partial sequence alignment of the extracellular domains of human and rat γenac. Residues that differ between the two subunits are identified with an open box. In total, 50 residues differ between the two species in the extracellular domains of γenac. Of these residues, we focused on 15 residues that H + might titrate within the physiologically relevant ph range (acidic residues and histidines, Fig 5.4, grey boxes). To test the hypothesis that acidic and histidine residues in the extracellular domain of human γenac contribute to ph activation of ENaC activity, we individually mutated each of the 15 acidic residues that differ between human and rat ENaC to alanine. We predicted that mutation of these residues would decrease the response to acidic ph. Figure 5.5A shows a representative trace of current versus time recorded from a Xenopus oocyte expressing wild type αβγenac or αβγ D166A ENaC. Relative to wild type, γ D166A increased ENaC current less in response to acidic ph. We performed the same experiment for each of the 15 mutant channels. Figure 5.5B shows the total change in ENaC activity between ph 9.0 and ph 5.25 for each mutation relative to wild type ENaC. We found γ D335A, γ Q165A, γ H439A, γ D164A, γ E455A, γ E292A or γ D166A each decreased ENaC ph response (Fig. 5.5B). In contrast, the other mutations had no effect or increased (γ H112A ) ph response (Fig. 5.5B). Because no single mutation abolished the response to ph, the data suggest that multiple residues in the extracellular domain of γenac are involved in ph sensing.
123 107 Mutation of multiple acidic residues in the extracellular domain of human γenac specifically decreases the response to acidic ph To further test the role of the identified residues, we generated a γenac construct containing mutations at the seven positions that decrease the ph response when individually mutated to alanine (γ D164A, Q165A, D166A, E292A, D335A, H439A, E455A ; labeled γ 7Ala ). Cells expressing human αβγ 7Ala ENaC had a greater reduction in their response to acidic ph than any single mutation alone (Fig 5.6A, 5.6B). The response to alkaline conditions (ph 7.4 to ph 9.0) was similar to wild type (Fig 5.6B). The data suggests that mutation of acidic residues preferentially reduces the response to acidic ph. However, mutation of these residues in γenac could cause a nonspecific change in function that prevents increases in activity for the mutant channel. To further confirm the specificity of the γ 7Ala mutations, we measured an additional channel property, Cl - inhibition. While Cl - inhibition is sensitive to extracellular ph (Chapter III), none of the residues mutated in the γ 7Ala construct are believed to participate in the putative Cl - binding site. We, therefore, expected that mutation of these residues (γ D164A, Q165A, D166A, E292A, D335A, H439A, E455A) would not significantly disrupt Cl - inhibition. Compared to wild type ENaC, cells expressing αβγ 7Ala ENaC did not have significantly decreased Cl - inhibition (.89 fold, Fig 5.6D and E). This demonstrates that the decrease in the response to acidic ph seen with the γ 7Ala mutations is a specific effect, as opposed to a broad disruption of channel function.
124 108 Residues equivalent to γe455 in α- and βenac also modulate the ph response Because human γenac is not sufficient to restore ph sensitivity in rat ENaC and because mutation of all seven γenac residues didn t abolish the ph response, we predict that multiple subunits play a role in ph sensing. How could this information be integrated into a concerted response? We hypothesize that this may occur at an intersubunit interface. We examined residues predicted by the ASIC1a crystal structures (2QTS and 3HGC) to lie at the interface of adjacent subunits. Of the seven γenac residues that decreased proton activation when mutated to alanine, four residues are predicted to lie at an intersubunit interface; γ D164, γ Q165, γ D166, and γ E455. The region around γ is not well conserved between ASIC1 and ENaC. Therefore, we focused on the γ E455 position (and its equivalent residues in α- and βenac) because of its location between two conserved cysteine residues (Fig 5.7A). We hypothesize that titration of α K477, β E446 and γ E455 may alter intersubunit interactions important in modulating channel gating. Figure 5.7A shows a partial sequence alignment of human αβγenac and chicken ASIC1a. Based on sequence alignment, γenac Glu 455 is homologous to ASIC1a Met 362. ASIC1a Met 362 forms an interface with ASIC1a Val 81 from an adjacent subunit. The homologous residue to ASIC1a Val 81 is γenac Val 87. In αenac, the equivalent residues are Lys 477 and Leu 120 and Glu 446 and Val 85 in βenac, respectively. The predicted location of these residues in α-, β-, and γenac is shown in Figure 5.7B. We mutated residues equivalent to γ E455 in α- (α K477 ) and βenac (β E446 ) to alanine. Figure 5.8A shows a representative trace of current versus time for cells
125 109 expressing αβγenac, α K477A βγenac or αβ E446A γenac. In cells expressing β E446A, changing extracellular ph from 7.4 to 5.25 increased Na + current significantly less than wild type ENaC. In contrast α K477A increased the response to ph 5.25 (Fig 5.8A). Figure 5.8B shows averaged data for the full ph dose range. α K477A increased, and β E446A decreased the response to acidic ph (Fig 5.8B). Thus, charged residues at equivalent positions in each subunit are involved in the ability of the channel to respond to changes in extracellular ph. We hypothesize that titration of negatively charged residues (γ E455 and β E446 ) increases ENaC activity. We speculate that uncharged residues, such as alanine and cysteine, are functionally equivalent to protonated, neutralized, acidic residues. To determine if negative charge is sufficient to restore ph activation of ENaC, we generated a cysteine mutation at β E446 and modified the residue with a negatively charged cysteine reactive compound, MTSES. Cells expressing β E446C have decreased activation by acidic ph similar to that seen with β E446A (Fig 8C). Modification of β E446C with MTSES partially restored ph activation (Fig 5.8C). The data provide further evidence that side chain charge at this position is an important regulator of ENaC function. Mutation of acidic residues in β- and γenac eliminate the response to acidic ph We have shown that mutations of seven acidic residues in γenac (Fig 5.6B, γ7ala ) and one in βenac (Fig 5.8B, βe446a ), on their own, significantly decrease the response to acidic ph. However, the response to acidic ph was not eliminated. In Fig 5.9A & B, we combined γ 7Ala with β E446A (coexpressed with wild type αenac). This
126 110 abolished the response to acidic ph, while the response to alkaline ph was unchanged (Fig 5.9B). In contrast, β E446A and γ 7Ala did not have decreased Cl - inhibition (Fig 5.9C) again suggesting that mutating these residues specifically decreases the ability of the channel to respond changes in extracellular ph. This data demonstrates that a subset of residues in β- and γenac are necessary for human αβγenac to be able to respond to changes in extracellular ph. In Chapter II, we demonstrated that protons increase ENaC activity, in part, by decreasing Na + self-inhibition (85). We, therefore, hypothesized that mutation of these residues in β- and γenac would decrease Na + self-inhibition and decrease the sensitivity of Na + self-inhibition to ph. We expressed wild type human αβγenac, αβγ 7Ala ENaC, or αβ E446A γ 7Ala ENaC and measured Na + self-inhibition at ph 7.4, and 5.5 under conditions of low extracellular Cl - (Fig 5.10A & B). As previously described, Na + self-inhibition of wild type ENaC was reduced at acidic ph ((85), and Fig 5.10A and C). Consistent with our hypothesis, cells expressing αβγ 7Ala ENaC or αβ E446A γ 7Ala ENaC had decreased Na + self-inhibition at ph 7.4 compared to Wt ENaC (Fig 5.10C). Furthermore, the ph dependence (slope) of Na + self-inhibition was also decreased for the mutant groups relative to wild type; extracellular acidification produced a marginal decrease in Na + selfinhibition (Fig 5.10C). Intriguingly, Na + self-inhibition for Wt ENaC and the mutant β- and γenac subunits appears to converge to one value at ph 5.5 (Fig 5.10C). We speculate that neutralization of the acidic residues by protonation or mutagenesis have equivalent effects on Na + self-inhibition.
127 111 Disruption of intersubunit interfaces modulates ENaC activity We have demonstrated that mutation of seven acidic residues in γenac and one residue in βenac abolishes the activation of ENaC activity in response to extracellular acidification. We hypothesized that protonation of α K477, β E446 and γ E455 may alter intersubunit interfaces important in modulating channel gating. To test this hypothesis, we mutated residues predicted by the ASIC1a crystal structures (2QTS and 3HGC) to lie at the opposing interface to α K477, β E446 and γ E466. In Chapter IV, we demonstrated that ENaC adopts an αγβ channel architecture (105). Using an αγβenac channel architecture, homology modeling to the ASIC1a structure predicts that α K477 forms an interface with β V85, β E446 interfaces with γ V87, and γ E466 interfaces with α L120 (Fig 5.7B). If these sets of residues form an interface we make two predictions. First, mutation of the uncharged interfacing residues can have electrostatic effects on α K477, β E455, and γ E446. Second, we predict that we will be able to specifically cross-link residues located at the same interface. To test for an interface between these residues, we engineered cysteine mutations at α L120, β V85, and γ V87. We hypothesized that modification of these residues might disrupt the intersubunit interface and alter channel function. To test this possibility we modified each mutant channel with positively (MTSET) or negatively (MTSES) charged cysteine reactive compounds. When oocytes expressing αβ V85C γenac were treated with MTSET, current irreversibly increased 2.19 fold relative to wild type ENaC, while treatment with MTSES irreversibly decreased current 0.61 fold relative to wild type ENaC (Fig 5.11A and B). Current in cells expressing α L120C βγenac was irreversibly decreased 0.45 fold by MTSET and irreversibly increased 1.14 fold by MTSES (Fig
128 C). Neither MTSES nor MTSET had significant functional effects on cells expressing αβγ V87C ENaC (data not shown). The data demonstrates that α L120 and β V85 are accessible to MTS compounds and that they are functionally important as modification of these residues change ENaC activity. Furthermore, the data indicate that at both interfaces, repulsive electrostatic interactions (α K477+ and β V85C-MTSET+ or γ E446- and α L120C-MTSES- ) increase ENaC activity whereas attractive electrostatic interactions (α K477+ and β V85C-MTSES- or γ E446- and α L120C-MTSET+ ) decrease ENaC activity. Functional effect of cross-linking opposing subunits By manipulating electrostatic interactions, we have demonstrated that α K477 and β V85 play an important role in modulating ENaC activity and may form an interface. To further test if these two residues are located in close proximity to each other, we introduced cysteines at both positions and tested whether they could be cross-linked with bifunctional cysteine reactive compounds of various lengths (5.2 to 20.8Å in length (22)). To assay for cross-linking, we monitored changes in ENaC steady state current as well as changes in Na + self-inhibition. After treatment with MTS-14-MTS for 60 seconds in the presence of amiloride (to prevent non-specific inhibition of Wt ENaC activity, data not shown), cells expressing α K477C β V85C γenac had a 421% increase in steady state current (I ss ) and a 38% decrease in Na + self-inhibition (Fig 5.13A, C and D). As a negative control, we measured steady state (I ss ) current and Na + self-inhibition in cells expressing either cysteine mutation alone (αβ V85C γenac or α K477C βγenac). Current and Na + self-inhibition recorded from these cells after treatment with MTS-14-
129 113 MTS was similar to Wt ENaC (Fig 5.13C and D). This indicates that both cysteins were required, suggesting they were cross-linked by MTS-14-MTS. We also tested the effects of shorter cross-linkers. Treatment with MTS-11-MTS (16.9 Å) also increased steady state current (370%) and decreased Na + self-inhibition (42%, Figure 5.12C & D). However, treatment with short cross-linking reagents did not increase current (relative to wild type ENaC), but still decreased Na + self-inhibition. MTS-6-MTS (10.4Å), MTS-4-MTS (7.8Å) and MTS-2-MTS (5.2Å) decreased Na + selfinhibition 47%, 41% and 75%, respectively (Fig 5.12B, C and D). Again, treatment of either cysteine mutation alone was not significantly different from wild type ENaC (Fig 5.12C and D). This demonstrates that short cross-linking reagents modify these channels despite the lack of an effect on steady state current. Regardless of their length, each reagent cross-linked ENaC and disrupted channel gating. However, only the long reagents (MTS-14-MTS and MTS-11-MTS) increased ENaC steady state current. Thus, the data indicate that the functional effect of cross-linking is dependent on the length of the cross-linker. We hypothesized that cross-linking ENaC residues would disrupt its regulation by ph. We expressed α K477C β V85C γenac in Xenopus oocytes and determined the ph dose response relationship before and after treatment with MTS-14-MTS. MTS-14-MTS decreased ph activation of ENaC activity across all ph solutions tested (Fig 5.13A and B). MTS-14-MTS treatment had no significant effect on the ph dose response of wild type ENaC (data not shown).
130 114 Discussion In this work, we identified residues that participate in regulation of ENaC activity by extracellular acidification. Furthermore, we are beginning to make key insights into how movements of the extracellular domain of the channel are tied to changes in channel gating. Interpretation of our data is based on putative structural similarity between ASIC1a, which has been crystalized, and ENaC. ASIC1a crystalized as a trimer. At least four independent lines of evidence support conservation of channel stoichiometry between ASIC1a and αβγenac. In Chapter IV, we demonstrated a conserved Cl - inhibitory site between ASIC1a (101) and ENaC (97). Identification of this Cl - inhibitory site allowed us to determine that ENaC adopts an αγβ channel architecture (105). The work described in Chapter V, also agrees with an αγβenac architecture. This architecture has also been confirmed by Chen et al in their work examining an intersubunit Cu 2+ binding site (98). Recently, atomic force microscopy data has revealed that ENaC assembles as a heterotrimer (or multiple heterotrimers), although the distinction between αγβ or αβγenac could not be determined (51). While certain regions and sub domains of ASIC and ENaC are variable, it appears that subunit organization is conserved. The current work adds to growing body of evidence suggesting that modulation of ENaC activity occurs at the interfaces of the extracellular domains of adjacent subunits. Ligand binding at subunit-subunit interfaces is important in modulation of a variety of channels and receptors (23, ). In particular, binding of glycine to Glycine Receptors (GlyR) initiates channel opening by binding at the interface of two subunits
131 115 (41). Crystallization of ASIC1 revealed a Cl - binding site composed of residues from adjacent subunits. In ENaC, we have previously demonstrated that mutation of putative Cl - binding site residues at the interface between α- and βenac and β- and γenac decreases inhibition by extracellular Cl - (Chapter IV, (97)). Chen et al. have recently demonstrated that mutations to intersubunit residues decreases ENaC inhibition by extracellular Cu 2+ (24). To visualize hypothetical subunit movements, we have developed a two state model of ENaC activity (Fig 5.14). In Chapter II, we demonstrated that ENaC activity is increased when the extracellular proton concentration increases. What could be the mechanism? Based on our data, we believe that residues located at the interface of two opposing subunits may be interacting. Manipulation of these interfacing residues (α K477 and β V85 ) by introducing repulsive electrostatic interactions or by treating with a long cross-linking reagent specifically increases ENaC activity and disrupts channel gating (as assayed as a decrease in Na + self-inhibition). Manipulation of these interfacing residues by introducing attractive electrostatic interactions or by treating with a short cross-linking reagent specifically decreases ENaC activity and disrupts channel gating. This disruption also dramatically decreases ph activation of ENaC activity. Furthermore, the data demonstrates that cross-linking is length dependent. From this, we speculate that ENaC is in a closed conformation when α K477 and β V85 are close together ( Å) and in an open conformation when α K477 and β V85 are farther apart ( Å). We predict that physiologically relevant ph fluctuations alter the distance between subunits at this interface.
132 116 Although ENaC is a constitutively active channel, a wide variety of extracellular stimuli can modulate its activity, including Na +, Cl -, H +, Cu 2+, Zn 2+, and Ni 2+, shear stress and proteases (13, 27, 40, 85, 97, 98, 109). In this regard, it is clear that the extracellular domain functions as a sensor to detect changes in the composition and quantity of the highly variable extracellular millue in which ENaC resides. We speculate that this rapid tuning of channel activity in response to changing extracellular conditions may underpin normal physiology by allowing ENaC to respond to a variety of challenges that threaten to disrupt Na + transport and fluid homeostasis. We demonstrated that human ENaC can be rendered insensitive to extracellular acidification by mutation of acidic residues in the extracellular domain. This is achieved by mutation of one residue in βenac (Glu-446) and seven residues in γenac (Asp-164, Gln-165, Asp-166, Glu-292, Asp-335, His-439 and Glu-455). The data suggest these eight residues are necessary for human αβγenac to respond to acidification of the extracellular milieu. Although ENaC subunits share a high degree of sequence similarity, there is a growing body of evidence to suggest that the functional roles of each subunit are strikingly different. Several examples exist. In Chapter IV, we demonstrated that the functional role of extracellular Cl - is dependent on the subunit involved (105). Mutation of the putative Cl - binding site composed of the thumb domain of αenac and palm domain of βenac decreased Cl - inhibition and Na + self-inhibition. Mutation of the equivalent Cl - site composed of the thumb domain of βenac and the palm domain of γenac decreased Cl - inhibition and did not affect Na + self-inhibition, while the Cl - site composed of residues from the thumb domain of γenac and the palm domain of αenac
133 117 had no effect on Cl - inhibition or Na + self-inhibition. Although the extracellular Na + binding site remains unknown, Sheng et al. have shown that mutation of equivalent histidine residues in α- and γenac have opposite effects on Na + self-inhibition (39). ENaC is activated by proteolytic cleavage of α- and γ-, but not βenac (12, 28, 102). Mutations at the DEG position also have varying effects when introduced in each ENaC subunit (103, 104). Here we demonstrated that mutation of an equivalent residue in αenac increases the ph response while mutation of the equivalent residues in β- and γenac decreased the ph response. The intermolecular interactions with neighboring residues with α K477 and β E446 or γ E455 must be different, however, the details will not be clear until the channel is crystalized. Such functional asymmetry of equivalent residues in different subunits of the channel may underlie the homeostatic nature of ENaC opposing subunits exert opposing effects to balance channel activity and maintain fluid homeostasis. We speculate that this intra-channel regulation expands the range of conformational and functional changes available and allows fine-tuning of channel activity in response to dynamic extracellular environments found in the kidney, colon and lingual epithelia. The key to understanding how ENaC responds to its dynamic extracellular environment will be determining how intra-subunit interactions change in response to extracellular stimuli. We have contributed to this endeavor by linking ph sensing to extracellular domain movements and changes in channel gating.
134 118 Figure 5.1. Human and rat ENaC respond differently to changes in extracellular ph. A, representative trace of current versus time recorded at -60mV from a Xenopus oocyte expressing human αβγenac (black) or rat αβγenac (grey). Currents were recorded under conditions of high Na + (116mM) and low extracellular Cl - (~5mM) over a range of ph from 9.0 to ENaC dependent inward Na + current was determined by application of amiloride at the end of the experiment. B, summary data of change in ENaC activity in response to extracellular ph relative to ph 7.4 (Ave +/- SEM, n = 6).
135 119 A Amil 0 0 I ( A) h r. h Time (Sec) -2-4 I ( A) r. B Change from ph % h. 50% 0% r. -50% ph
136 120 Figure 5.2. Human γenac is necessary for ph regulation of channel activity. A,C,E, representative traces of current versus time recorded at -60mV from Xenopus oocytes expressing a combination of human and rat ENaC subunits (A, rat β human αγenac; C, rat α human βγenac; E, rat γ human αβenac). Currents were recorded under conditions of high Na + (116mM) and low extracellular Cl - (~5mM) over a range of ph from 9.0 to Traces show ENaC activity at ph 7.4 (open bar) and ph 5.25 (black bar). ENaC dependent inward Na + current was determined by application of amiloride at the end of the experiment. Summary data of change in ENaC activity in response to extracellular ph relative to ph 7.4 obtained from cells expressing: B, rat βenac and human α- and γenac (Ave +/- SEM, n = 6). D, rat αenac and human β- and γenac (Ave +/- SEM, n = 3). F, rat γenac and human α- and βenac (Ave +/- SEM, n = 9).
137 121 A A 5s C E A 5s 5.25 Amil r. h h Amil h. r. h B Change from ph 7.4 D Change from ph 7.4 F 200% 100% 0% -100% ph 200% 100% h. h. r. h. r. h. 0% -100% ph h. h. r h. r h A 5s 5.25 Amil r. h h. Change from ph % 100% r. h. h. h. 0% h. r -100% ph
138 122 Figure 5.3. Human γenac is not sufficient for ph regulation of channel activity. A, representative traces of current versus time recorded at -60 mv from Xenopus oocytes expressing rat αβγ (light grey trace), rat αβ- and human γenac (medium grey trace), or human αβγenac (black trace). Currents were recorded under conditions of high Na + (116mM) and low extracellular Cl - (~5mM) over a range of ph from 9.0 to Trace shows ENaC activity at ph 7.4 (open bar) and ph 5.25 (black bar). ENaC dependent inward Na + current was determined by application of amiloride at the end of the experiment. B, summary data of change in ENaC activity in response to extracellular ph relative to ph 7.4 obtained from cells expressing rat αβγ (grey, Ave +/- SEM, n = 6), rat αβ- and human γenac (white, Ave +/- SEM, n = 6), or human αβγenac (black, Ave +/- SEM, n = 6).
139 123 A A 5s 5.25 Amil r. r. h h. B Change from ph % 100% 0% r. h. r. -100% ph h. r. r. h
140 124 Figure 5.4. Acidic residues differ in the extracellular domain of human and rat γenac. A, partial sequence alignment of the extracellular domains of human and rat γenac. Residues that differ between the two subunits are indicated with an open box. Acidic residues (Asp, Glu, His) that differ between the two subunits are indicated with a grey box.
141 125 A h. ENaC r. ENaC VSVSIKVHFRKLDFPAVTICNINPYKYSTVRHLLADLEQETREALKSLYGFPESRKRREAESWNSVSEGKQPRFSHRIPL 160 VSVSIKVHFQKLDFPAVTICNINPYKYSAVSDLLTDLDSETKQALLSLYGVKESRKRREAGSMPSTLEGTPPRFFKLIPL 160 h. ENaC r. ENaC LIFDQDEKGKARDFFTGRKRKVGGSIIHKASNVMHI-ESKQVVGFQLCSNDTSDCATYTFSSGINAIQEWYKLHYMNIMA 239 LVFNENEKGKARDFFTGRKRKISGKIIHKASNVMHVHESKKLVGFQLCSNDTSDCATYTFSSGINAIQEWYKLHYMNIMA 240 h. ENaC r. ENaC QVPLEKKINMSYSAEELLVTCFFDGVSCDARNFTLFHHPMHGNCYTFNNRENETILSTSMGGSEYGLQVILYINEEEYNP 319 QVPLEKKINMSYSAEELLVTCFFDGMSCDARNFTLFHHPMYGNCYTFNNKENATILSTSMGGSEYGLQVILYINEDEYNP 320 h. ENaC r. ENaC FLVSSTGAKVIIHRQDEYPFVEDVGTEIETAMVTSIGMHLTESFKLSEPYSQCTEDGSDVPIRNIYNAAYSLQICLHSCF 399 FLVSSTGAKVLIHQQNEYPFIEDVGMEIETAMSTSIGMHLTESFKLSEPYSQCTEDGSDVPVTNIYNAAYSLQICLYSCF 400 h. ENaC r. ENaC QTKMVEKCGCAQYSQPLPPAANYCNYQQHPNWMYCYYQLHRAFVQEELGCQSVCKEACSFKEWTLTTSLAQWPSVVSEKW 479 QTKMVEKCGCAQYSQPLPPAANYCNYQQHPNWMYCYYQLYQAFVREELGCQSVCKQSCSFKEWTLTTSLAQWPSEASEKW 480
142 126 Figure 5.5. Mutation of titratable residues in the extracellular domain of γenac change the ph response. A, representative trace of current versus time recorded at -60mV from a Xenopus oocyte expressing human αβγenac or human αβ- and γ D166A ENaC. Currents were recorded under conditions of high Na + (116mM) and low extracellular Cl - (~5mM) over a range of ph from 9.0 to 5.25 (Only ph 5.25 is shown). ENaC dependent inward Na + current was determined by application of amiloride at the end of the experiment. B, summary data of the total ph response (calculated as the difference in ENaC activity between ph 9.0 and 5.25) relative to wt ENaC (Ave +/- SEM, n = 3-8).
143 127 A Amil h. D166A B Rel (ph ) * H112A E123A H396A E118A E455A D164A H439A Q165A D335A H280A H156A h. E315A E141A * * * * * * * D166A E292A 1 A 5s h.
144 128 Figure 5.6. Mutation of multiple residues in γenac results in an additive decrease in the response to acidic ph. A, representative trace of current versus time recorded -60mV from a Xenopus oocyte expressing human αβγenac or human αβ- and γ D164A, Q165A, D166A, E292A, D335A, H439A, E455A ENaC ( γ7ala mut, grey trace). Currents were recorded under conditions of high Na + (116mM) and low extracellular Cl - (~5mM) over a range of ph from 9.0 to 5.25 (Only ph 5.25 is shown). ENaC dependent inward Na + current was determined by application of amiloride at the end of the experiment. B, summary data of change in ENaC activity in response to extracellular ph relative to ph 7.4 for Wt and γ7ala mutation (Ave +/- SEM, n = 11). C, representative trace of Cl - inhibition recorded from oocytes expressing Wt (black) or γ D164A, Q165A, D166A, E292A, D335A, H439A, E455A ENaC ( γ7ala mut, grey trace). D, summary data showing Cl - inhibition relative to Wt ENaC (Ave +/- SEM, n = 11).
145 129 A C 1µA 1µA 0 5s s 5mM Cl Amil γ7ala h.αβγ Amil γ 7Ala h.αβγ B Change from ph 7.4 D Rel Cl - Inhibition 100% 50% 0% h.αβγ -50% γ 7Ala h.αβγ ph γ 7Ala
146 130 Figure 5.7. Homology modeling with ASIC1a crystal structure suggests γ E455 may lie at an intersubunit interface. A, partial sequence alignment of the extracellular domains of human αβγenac and chicken ASIC1a. γ E455 and equivalent positions in α- and βenac and chicken ASIC1a are indicated on the left with a gold box. Interfacing residues are highlighted in the sequence alignment on the right. B, based on the ASIC crystal structure, γ E455A is predicted to form an interface with α L120, β E446 with γ V87 and α K477 with β V85. On the left, regions of interest the ASIC1a crystal structure (3HGC) are color coded to indicate the relative location of α- (red), γ- (green) and βenac (blue). On the right, a 90 degree rotation of the structure looking down through the pore. Only the areas of interest are shown. Other regions of the structure were hidden for clarity.

147 131 A ENaC ENaC ENaC casic1a CRKPC 477 CKESC 446 CKEAC 455 CEMPC 362 ENaC PVSLNINLNSDKLVFPAVTIC 120 ENaC EVSVSLSVGFKTMDFPAVTIC 85 ENaC TVSVSIKVHFRKLDFPAVTIC 87 casic1a HVTKLDEVAATRLTFPAVTFC 81 B 90 o V85 K477 E446 L120 V87 E455
Overview of ion channel proteins. What do ion channels do? Three important points:
 Overview of ion channel proteins Protein Structure Membrane proteins & channels Specific channels Several hundred distinct types Organization Evolution We need to consider 1. Structure 2. Functions 3.
Overview of ion channel proteins Protein Structure Membrane proteins & channels Specific channels Several hundred distinct types Organization Evolution We need to consider 1. Structure 2. Functions 3.
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
External Nickel Inhibits Epithelial Sodium Channel by Binding to Histidine Residues within the Extracellular Domains of
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 51, Issue of December 20, pp. 50098 50111, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. External Nickel
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 51, Issue of December 20, pp. 50098 50111, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. External Nickel
Renal handling of substances. Dr.Charushila Rukadikar Assistance Professor Physiology
 Renal handling of substances Dr.Charushila Rukadikar Assistance Professor Physiology GENERAL PRINCIPLES OF RENAL TUBULAR TRANSPORT Transport mechanisms across cell membrane 1) Passive transport i. Diffusion
Renal handling of substances Dr.Charushila Rukadikar Assistance Professor Physiology GENERAL PRINCIPLES OF RENAL TUBULAR TRANSPORT Transport mechanisms across cell membrane 1) Passive transport i. Diffusion
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016
 Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Patrick: An Introduction to Medicinal Chemistry 5e Chapter 04
 01) Which of the following statements is not true about receptors? a. Most receptors are proteins situated inside the cell. b. Receptors contain a hollow or cleft on their surface which is known as a binding
01) Which of the following statements is not true about receptors? a. Most receptors are proteins situated inside the cell. b. Receptors contain a hollow or cleft on their surface which is known as a binding
Transmembrane Domains (TMDs) of ABC transporters
 of ABC transporters") Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Dr. Ketki Assistant Professor Department of Biochemistry Heritage IMS, Varanasi
 TRANSPORT MECHANISMS Dr. Ketki Assistant Professor Department of Biochemistry Heritage IMS, Varanasi Membrane selectivity allows adjustments of cell composition and function If plasma membrane is relatively
TRANSPORT MECHANISMS Dr. Ketki Assistant Professor Department of Biochemistry Heritage IMS, Varanasi Membrane selectivity allows adjustments of cell composition and function If plasma membrane is relatively
Inhibition of S532C by MTSET at intracellular ph 6.8 indicates accessibility in the closed
 Supplementary Text Inhibition of S532C by MTSET at intracellular ph 6.8 indicates accessibility in the closed state It is difficult to examine accessibility of cysteine-substituted mutants in the fully
Supplementary Text Inhibition of S532C by MTSET at intracellular ph 6.8 indicates accessibility in the closed state It is difficult to examine accessibility of cysteine-substituted mutants in the fully
NIH/3T3 CFTR. Cystic fibrosis transmembrane receptors. Application Report:
 Application Report: NIH/3T3 CFTR Cystic fibrosis transmembrane receptors Cystic fibrosis transmembrane conductance regulator (CFTR) functions as a chloride channel and controls the regulation of other
Application Report: NIH/3T3 CFTR Cystic fibrosis transmembrane receptors Cystic fibrosis transmembrane conductance regulator (CFTR) functions as a chloride channel and controls the regulation of other
Advanced Higher Biology. Unit 1- Cells and Proteins 2c) Membrane Proteins
 Membrane Proteins") Advanced Higher Biology Unit 1- Cells and Proteins 2c) Membrane Proteins Membrane Structure Phospholipid bilayer Transmembrane protein Integral protein Movement of Molecules Across Membranes Phospholipid
Advanced Higher Biology Unit 1- Cells and Proteins 2c) Membrane Proteins Membrane Structure Phospholipid bilayer Transmembrane protein Integral protein Movement of Molecules Across Membranes Phospholipid
Acids, Bases, Salts, Buffers
 Acids, Bases, Salts, Buffers Acids, Bases, Salts, Buffers An acid is any solute that dissociates in a solution and releases hydrogen ions, thereby lowering ph Since a hydrogen ion consist solely of a proton,
Acids, Bases, Salts, Buffers Acids, Bases, Salts, Buffers An acid is any solute that dissociates in a solution and releases hydrogen ions, thereby lowering ph Since a hydrogen ion consist solely of a proton,
Reception The target cell s detection of a signal coming from outside the cell May Occur by: Direct connect Through signal molecules
 Why Do Cells Communicate? Regulation Cells need to control cellular processes In multicellular organism, cells signaling pathways coordinate the activities within individual cells that support the function
Why Do Cells Communicate? Regulation Cells need to control cellular processes In multicellular organism, cells signaling pathways coordinate the activities within individual cells that support the function
Overview of Physiology & Homeostasis. Biological explanations Levels of organization Homeostasis
 Overview of Physiology & Homeostasis 1 Biological explanations Levels of organization Homeostasis 2 Biological Explanations Proximate Proximate causation: an explanation of an animal's behavior based on
Overview of Physiology & Homeostasis 1 Biological explanations Levels of organization Homeostasis 2 Biological Explanations Proximate Proximate causation: an explanation of an animal's behavior based on
Ion Channel Structure and Function (part 1)
") Ion Channel Structure and Function (part 1) The most important properties of an ion channel Intrinsic properties of the channel (Selectivity and Mode of Gating) + Location Physiological Function Types
Ion Channel Structure and Function (part 1) The most important properties of an ion channel Intrinsic properties of the channel (Selectivity and Mode of Gating) + Location Physiological Function Types
Membrane Protein Channels
 Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
Host-Pathogen Interaction. PN Sharma Department of Plant Pathology CSK HPKV, Palampur
 Host-Pathogen Interaction PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 PATHOGEN DEFENCE IN PLANTS A BIOLOGICAL AND MOLECULAR VIEW Two types of plant resistance response to potential
Host-Pathogen Interaction PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 PATHOGEN DEFENCE IN PLANTS A BIOLOGICAL AND MOLECULAR VIEW Two types of plant resistance response to potential
STEIN IN-TERM EXAM -- BIOLOGY FEBRUARY 12, PAGE 1 of 7
 STEIN IN-TERM EXAM -- BIOLOGY 3058 -- FEBRUARY 12, 2009 -- PAGE 1 of 7 There are 25 questions in this Biology 3058 exam. All questions are "A, B, C, D, E, F, G, H" questions worth one point each. There
STEIN IN-TERM EXAM -- BIOLOGY 3058 -- FEBRUARY 12, 2009 -- PAGE 1 of 7 There are 25 questions in this Biology 3058 exam. All questions are "A, B, C, D, E, F, G, H" questions worth one point each. There
Raghuram et al PNAS 100:9620
 ION CHANNL MACROMOLCULAR COMPLXS A V I R W c g I S L S R d L L LLL I VA S f L LS HP a b D G C S1 S2 S3 S4 S5 I e V N S6 COOH NH 2 PDZ domains; recognition of short peptides with a COOH terminal hydrophobic
ION CHANNL MACROMOLCULAR COMPLXS A V I R W c g I S L S R d L L LLL I VA S f L LS HP a b D G C S1 S2 S3 S4 S5 I e V N S6 COOH NH 2 PDZ domains; recognition of short peptides with a COOH terminal hydrophobic
Transport of glucose across epithelial cells: a. Gluc/Na cotransport; b. Gluc transporter Alberts
 Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Principles Of Acid-Base Balance
 Principles Of Acid-Base Balance I. Introduction A. For normal body function the free H+ concentration [H+] or ph must be kept within a narrow normal range. Some reasons why: 1. The proton "pump" within
Principles Of Acid-Base Balance I. Introduction A. For normal body function the free H+ concentration [H+] or ph must be kept within a narrow normal range. Some reasons why: 1. The proton "pump" within
Potassium channel gating and structure!
 Reading: Potassium channel gating and structure Hille (3rd ed.) chapts 10, 13, 17 Doyle et al. The Structure of the Potassium Channel: Molecular Basis of K1 Conduction and Selectivity. Science 280:70-77
Reading: Potassium channel gating and structure Hille (3rd ed.) chapts 10, 13, 17 Doyle et al. The Structure of the Potassium Channel: Molecular Basis of K1 Conduction and Selectivity. Science 280:70-77
Zool 3200: Cell Biology Exam 5 4/27/15
 Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Rahaf Nasser mohammad khatatbeh
 7 7... Hiba Abu Hayyeh... Rahaf Nasser mohammad khatatbeh Mohammad khatatbeh Brief introduction about membrane potential The term membrane potential refers to a separation of opposite charges across the
7 7... Hiba Abu Hayyeh... Rahaf Nasser mohammad khatatbeh Mohammad khatatbeh Brief introduction about membrane potential The term membrane potential refers to a separation of opposite charges across the
Physiology Lecture 2 and 27 Chapter 2: Acids, Bases and Salts REVIEW:
 Physiology Lecture 2 and 27 Chapter 2: Acids, Bases and Salts REVIEW: 1. Salts an ionic compounds containing other than H + or OH - ; can dissociate in water to form electrolytes. Electrolytes can conduct
Physiology Lecture 2 and 27 Chapter 2: Acids, Bases and Salts REVIEW: 1. Salts an ionic compounds containing other than H + or OH - ; can dissociate in water to form electrolytes. Electrolytes can conduct
Neurophysiology. Danil Hammoudi.MD
 Neurophysiology Danil Hammoudi.MD ACTION POTENTIAL An action potential is a wave of electrical discharge that travels along the membrane of a cell. Action potentials are an essential feature of animal
Neurophysiology Danil Hammoudi.MD ACTION POTENTIAL An action potential is a wave of electrical discharge that travels along the membrane of a cell. Action potentials are an essential feature of animal
NURS1004 Week 12 Lecture 1 Acid Base Balance Prepared by Didy Button
 NURS1004 Week 12 Lecture 1 Acid Base Balance Prepared by Didy Button The Role of Amino Acids in Protein Buffer Systems 2-7 ph and Homeostasis ph The concentration of hydrogen ions (H + ) in a solution
NURS1004 Week 12 Lecture 1 Acid Base Balance Prepared by Didy Button The Role of Amino Acids in Protein Buffer Systems 2-7 ph and Homeostasis ph The concentration of hydrogen ions (H + ) in a solution
Name: TF: Section Time: LS1a ICE 5. Practice ICE Version B
 Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
MOLECULAR DRUG TARGETS
 MOLECULAR DRUG TARGETS LEARNING OUTCOMES At the end of this session student shall be able to: List different types of druggable targets Describe forces involved in drug-receptor interactions Describe theories
MOLECULAR DRUG TARGETS LEARNING OUTCOMES At the end of this session student shall be able to: List different types of druggable targets Describe forces involved in drug-receptor interactions Describe theories
LABETTE COMMUNITY COLLEGE BRIEF SYLLABUS. ANATOMY AND PHYSIOLOGY, lecture and lab
 LABETTE COMMUNITY COLLEGE BRIEF SYLLABUS SPECIAL NOTE: This brief syllabus is not intended to be a legal contract. A full syllabus will be distributed to students at the first class session. TEXT AND SUPPLEMENTARY
LABETTE COMMUNITY COLLEGE BRIEF SYLLABUS SPECIAL NOTE: This brief syllabus is not intended to be a legal contract. A full syllabus will be distributed to students at the first class session. TEXT AND SUPPLEMENTARY
6 Mechanotransduction
 6.1 Motivation The process of converting physical forces into biochemical signals and integrating these signals into the cellular response is referred to as mechnotransduction [11, 20]. To fully understand
6.1 Motivation The process of converting physical forces into biochemical signals and integrating these signals into the cellular response is referred to as mechnotransduction [11, 20]. To fully understand
Biology September 2015 Exam One FORM G KEY
 Biology 251 17 September 2015 Exam One FORM G KEY PRINT YOUR NAME AND ID NUMBER in the space that is provided on the answer sheet, and then blacken the letter boxes below the corresponding letters of your
Biology 251 17 September 2015 Exam One FORM G KEY PRINT YOUR NAME AND ID NUMBER in the space that is provided on the answer sheet, and then blacken the letter boxes below the corresponding letters of your
Biology September 2015 Exam One FORM W KEY
 Biology 251 17 September 2015 Exam One FORM W KEY PRINT YOUR NAME AND ID NUMBER in the space that is provided on the answer sheet, and then blacken the letter boxes below the corresponding letters of your
Biology 251 17 September 2015 Exam One FORM W KEY PRINT YOUR NAME AND ID NUMBER in the space that is provided on the answer sheet, and then blacken the letter boxes below the corresponding letters of your
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
Signal Transduction Phosphorylation Protein kinases. Misfolding diseases. Protein Engineering Lysozyme variants
 Signal Transduction Phosphorylation Protein kinases Misfolding diseases Protein Engineering Lysozyme variants Cells and Signals Regulation The cell must be able to respond to stimuli Cellular activities
Signal Transduction Phosphorylation Protein kinases Misfolding diseases Protein Engineering Lysozyme variants Cells and Signals Regulation The cell must be able to respond to stimuli Cellular activities
Lecture 3 13/11/2018
 Lecture 3 13/11/2018 1 Plasma membrane ALL cells have a cell membrane made of proteins and lipids. protein channel Cell Membrane Layer 1 Layer 2 lipid bilayer protein pump Lipid bilayer allows water, carbon
Lecture 3 13/11/2018 1 Plasma membrane ALL cells have a cell membrane made of proteins and lipids. protein channel Cell Membrane Layer 1 Layer 2 lipid bilayer protein pump Lipid bilayer allows water, carbon
BA, BSc, and MSc Degree Examinations
 Examination Candidate Number: Desk Number: BA, BSc, and MSc Degree Examinations 2017-8 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes
Examination Candidate Number: Desk Number: BA, BSc, and MSc Degree Examinations 2017-8 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes
Hole s Human Anatomy and Physiology Tenth Edition. Chapter 2
 PowerPoint Lecture Outlines to accompany Hole s Human Anatomy and Physiology Tenth Edition Shier w Butler w Lewis Chapter 2 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction
PowerPoint Lecture Outlines to accompany Hole s Human Anatomy and Physiology Tenth Edition Shier w Butler w Lewis Chapter 2 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction
Equivalent Circuit of the Membrane Connected to the Voltage Clamp. I mon. For Large Depolarizations, Both I Na and I K Are Activated
 VoltageGated Ion Channels and the Action Potential jdk3 Principles of Neural Science, chaps 8&9 VoltageGated Ion Channels and the Action Potential The Action Potential Generation Conduction VoltageGated
VoltageGated Ion Channels and the Action Potential jdk3 Principles of Neural Science, chaps 8&9 VoltageGated Ion Channels and the Action Potential The Action Potential Generation Conduction VoltageGated
V m = the Value of the Na Battery Plus the Voltage Drop Across g Na. I Na is Isolated By Blocking I K. and g K
 VoltageGated Ion Channels and the Action Potential VoltageGated Ion Channels and the Action Potential jdk3 Principles of Neural Science, chaps 8&9 The Action Potential Generation Conduction VoltageGated
VoltageGated Ion Channels and the Action Potential VoltageGated Ion Channels and the Action Potential jdk3 Principles of Neural Science, chaps 8&9 The Action Potential Generation Conduction VoltageGated
Practical: Acid-base balance
 Practical: Acid-base balance Definition All the processes inside the body which keep the H+ concentration within normal values. It is a result of the hydro-electrolytic balance and a sum of mechanisms
Practical: Acid-base balance Definition All the processes inside the body which keep the H+ concentration within normal values. It is a result of the hydro-electrolytic balance and a sum of mechanisms
Ch. 3: Cells & Their Environment
 Ch. 3: Cells & Their Environment OBJECTIVES: 1. Understand cell membrane permeability 2. To recognize different types of cellular transport (passive vs active) 3. To understand membrane potential and action
Ch. 3: Cells & Their Environment OBJECTIVES: 1. Understand cell membrane permeability 2. To recognize different types of cellular transport (passive vs active) 3. To understand membrane potential and action
SUMMARY OF THE EVENTS WHICH TRIGGER AN ELECTRICAL IMPUSLE IN NERVE CELLS (see figures on the following page)
") Anatomy and Physiology/AP Biology ACTION POTENTIAL SIMULATION BACKGROUND: The plasma membrane of cells is a selectively permeable barrier, which separates the internal contents of the cell from the surrounding
Anatomy and Physiology/AP Biology ACTION POTENTIAL SIMULATION BACKGROUND: The plasma membrane of cells is a selectively permeable barrier, which separates the internal contents of the cell from the surrounding
Endocrine Physiology. Introduction to Endocrine Principles
 Endocrine Physiology Introduction to Endocrine Principles There are TWO major groups of hormones Peptide and protein hormones Amine hormones Peptide and protein hormones act through cell membrane receptors
Endocrine Physiology Introduction to Endocrine Principles There are TWO major groups of hormones Peptide and protein hormones Amine hormones Peptide and protein hormones act through cell membrane receptors
ADAM FAMILY. ephrin A INTERAZIONE. Eph ADESIONE? PROTEOLISI ENDOCITOSI B A RISULTATO REPULSIONE. reverse. forward
 ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
ADAM FAMILY - a family of membrane-anchored metalloproteases that are known as A Disintegrin And Metalloprotease proteins and are key components in protein ectodomain shedding Eph A INTERAZIONE B ephrin
Physiology lecture (8): Acid Base regulation
: Acid Base regulation") Physiology lecture (8): Acid Base regulation If we add hydrogen, we have three lines of defense against a mild change in ph: 1) Buffers, instantaneous, within a fraction of milliseconds. 2) The lung, takes
Physiology lecture (8): Acid Base regulation If we add hydrogen, we have three lines of defense against a mild change in ph: 1) Buffers, instantaneous, within a fraction of milliseconds. 2) The lung, takes
Cardiac cell-cell Communication Part 1 Alonso P. Moreno D.Sc. CVRTI, Cardiology
 Bioengineering 6003 Cellular Electrophysiology and Biophysics Cardiac cell-cell Communication Part 1 Alonso P. Moreno D.Sc. CVRTI, Cardiology moreno@cvrti.utah.edu November 2010 poster Physiological Relevance
Bioengineering 6003 Cellular Electrophysiology and Biophysics Cardiac cell-cell Communication Part 1 Alonso P. Moreno D.Sc. CVRTI, Cardiology moreno@cvrti.utah.edu November 2010 poster Physiological Relevance
CELL BIOLOGY - CLUTCH CH. 9 - TRANSPORT ACROSS MEMBRANES.
 !! www.clutchprep.com K + K + K + K + CELL BIOLOGY - CLUTCH CONCEPT: PRINCIPLES OF TRANSMEMBRANE TRANSPORT Membranes and Gradients Cells must be able to communicate across their membrane barriers to materials
!! www.clutchprep.com K + K + K + K + CELL BIOLOGY - CLUTCH CONCEPT: PRINCIPLES OF TRANSMEMBRANE TRANSPORT Membranes and Gradients Cells must be able to communicate across their membrane barriers to materials
Channels can be activated by ligand-binding (chemical), voltage change, or mechanical changes such as stretch.
, voltage change, or mechanical changes such as stretch.") 1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic
 NEUROPHYSIOLOGY NOTES L1 WHAT IS NEUROPHYSIOLOGY? NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic receptor
NEUROPHYSIOLOGY NOTES L1 WHAT IS NEUROPHYSIOLOGY? NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic receptor
Gene Control Mechanisms at Transcription and Translation Levels
 Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
Gene Control Mechanisms at Transcription and Translation Levels Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by MHRD Page 1 of 9
MEMBRANE STRUCTURE. Lecture 9. Biology Department Concordia University. Dr. S. Azam BIOL 266/
 MEMBRANE STRUCTURE Lecture 9 BIOL 266/4 2014-15 Dr. S. Azam Biology Department Concordia University RED BLOOD CELL MEMBRANE PROTEINS The Dynamic Nature of the Plasma Membrane SEM of human erythrocytes
MEMBRANE STRUCTURE Lecture 9 BIOL 266/4 2014-15 Dr. S. Azam Biology Department Concordia University RED BLOOD CELL MEMBRANE PROTEINS The Dynamic Nature of the Plasma Membrane SEM of human erythrocytes
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
LESSON 2.2 WORKBOOK How do our axons transmit electrical signals?
 LESSON 2.2 WORKBOOK How do our axons transmit electrical signals? This lesson introduces you to the action potential, which is the process by which axons signal electrically. In this lesson you will learn
LESSON 2.2 WORKBOOK How do our axons transmit electrical signals? This lesson introduces you to the action potential, which is the process by which axons signal electrically. In this lesson you will learn
10/16/17 ACIDS AND BASES, DEFINED WATER IS AMPHOTERIC OUTLINE. 9.1 Properties of Acids and Bases. 9.2 ph. 9.3 Buffers
 ACIDS AND BASES, DEFINED A hydrogen atom contains a proton and an electron, thus a hydrogen ion (H + ) is a proton: Acids: Proton (H + ) transfer between molecules is the basis of acid/base chemistry Ø
ACIDS AND BASES, DEFINED A hydrogen atom contains a proton and an electron, thus a hydrogen ion (H + ) is a proton: Acids: Proton (H + ) transfer between molecules is the basis of acid/base chemistry Ø
Receptors and Ion Channels
 Receptors and Ion Channels Laurie Kellaway Senior Lecturer Department of Human Biology Laurie@curie.uct.ac.za Tel. +27 +21 4066 271 What are the two types of Neurotransmitter receptors Ionotropic receptors
Receptors and Ion Channels Laurie Kellaway Senior Lecturer Department of Human Biology Laurie@curie.uct.ac.za Tel. +27 +21 4066 271 What are the two types of Neurotransmitter receptors Ionotropic receptors
Ion Channels. 6 th December, Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology
 Ion Channels 6 th December, 2016 Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology Introduction The ion channels of nerve cells are optimally
Ion Channels 6 th December, 2016 Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology Introduction The ion channels of nerve cells are optimally
7.013 Problem Set
 7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as shown below.
ADVANCED PLACEMENT BIOLOGY
 ADVANCED PLACEMENT BIOLOGY Description Advanced Placement Biology is designed to be the equivalent of a two-semester college introductory course for Biology majors. The course meets seven periods per week
ADVANCED PLACEMENT BIOLOGY Description Advanced Placement Biology is designed to be the equivalent of a two-semester college introductory course for Biology majors. The course meets seven periods per week
Chem Lecture 10 Signal Transduction
 Chem 452 - Lecture 10 Signal Transduction 111202 Here we look at the movement of a signal from the outside of a cell to its inside, where it elicits changes within the cell. These changes are usually mediated
Chem 452 - Lecture 10 Signal Transduction 111202 Here we look at the movement of a signal from the outside of a cell to its inside, where it elicits changes within the cell. These changes are usually mediated
Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.
 Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.441/HST522J INTEGRINS I.V. Yannas, Ph.D. and M. Spector, Ph.D. Regulator
Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/BE.441/HST522J INTEGRINS I.V. Yannas, Ph.D. and M. Spector, Ph.D. Regulator
Physiology. Biol 219 Lec 1 Fall The Science of Body Function. Themes of Physiology. Themes of Physiology
 Physiology The Science of Body Function Themes of Physiology 1. Physical-chemical basis of body function Scientific method to study and understand the body Descriptive and quantitative Focus on processes
Physiology The Science of Body Function Themes of Physiology 1. Physical-chemical basis of body function Scientific method to study and understand the body Descriptive and quantitative Focus on processes
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Identification of Amino Acid Residues in the,, and Subunits of the Epithelial Sodium Channel (ENaC) Involved in Amiloride Block and Ion Permeation
 Involved in Amiloride Block and Ion Permeation") Identification of Amino Acid Residues in the,, and Subunits of the Epithelial Sodium Channel (ENaC) Involved in Amiloride Block and Ion Permeation Laurent Schild, Estelle Schneeberger, Ivan Gautschi, and
Identification of Amino Acid Residues in the,, and Subunits of the Epithelial Sodium Channel (ENaC) Involved in Amiloride Block and Ion Permeation Laurent Schild, Estelle Schneeberger, Ivan Gautschi, and
Biol403 - Receptor Serine/Threonine Kinases
 Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Renal potassium transport: mechanisms and regulation
 AJP Centennial Renal potassium transport: mechanisms and regulation GERHARD GIEBISCH Department of Cellular and Molecular Physiology, Yale University School of Medicine, New Haven, Connecticut 06520-8026
AJP Centennial Renal potassium transport: mechanisms and regulation GERHARD GIEBISCH Department of Cellular and Molecular Physiology, Yale University School of Medicine, New Haven, Connecticut 06520-8026
CHAPTER 1 THE STRUCTURAL BIOLOGY OF THE FGF19 SUBFAMILY
 CHAPTER 1 THE STRUCTURAL BIOLOGY OF THE FGF19 SUBFAMILY Andrew Beenken and Moosa Mohammadi* Department of Pharmacology, New York University School of Medicine, New York, New York, USA. *Corresponding Author:
CHAPTER 1 THE STRUCTURAL BIOLOGY OF THE FGF19 SUBFAMILY Andrew Beenken and Moosa Mohammadi* Department of Pharmacology, New York University School of Medicine, New York, New York, USA. *Corresponding Author:
III. Voltage Independence of Basolateral Membrane Na - Efflux
 Na' and K+ Transport at Basolateral Membranes of Epithelial Cells III. Voltage Independence of Basolateral Membrane Na - Efflux THOMAS C. COX and SANDY I. HELMAN From the Department of Physiology and Biophysics,
Na' and K+ Transport at Basolateral Membranes of Epithelial Cells III. Voltage Independence of Basolateral Membrane Na - Efflux THOMAS C. COX and SANDY I. HELMAN From the Department of Physiology and Biophysics,
Acid-Base balance. Acid : is the substance that donates H+, base : is the substance which receives or removes that H+.
 Acid-Base balance Introduction : We usually suffer from metabolic acidosis more than alkalosis, acidosis means that there is an increase in H+ concentration in plasma and thus interstial fluid ;because
Acid-Base balance Introduction : We usually suffer from metabolic acidosis more than alkalosis, acidosis means that there is an increase in H+ concentration in plasma and thus interstial fluid ;because
SUPPLEMENTARY INFORMATION. doi: /nature07461
 Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
The Potassium Ion Channel: Rahmat Muhammad
 The Potassium Ion Channel: 1952-1998 1998 Rahmat Muhammad Ions: Cell volume regulation Electrical impulse formation (e.g. sodium, potassium) Lipid membrane: the dielectric barrier Pro: compartmentalization
The Potassium Ion Channel: 1952-1998 1998 Rahmat Muhammad Ions: Cell volume regulation Electrical impulse formation (e.g. sodium, potassium) Lipid membrane: the dielectric barrier Pro: compartmentalization
Action potentials. Conductances channels
 Action potentials Conductances channels Cole and Curtis AC Wheatstone bridge resistance decreased during action potential R1 & R2 divide one path, Rv (variable) and Ru divide the other Galvanometer between
Action potentials Conductances channels Cole and Curtis AC Wheatstone bridge resistance decreased during action potential R1 & R2 divide one path, Rv (variable) and Ru divide the other Galvanometer between
Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a
 To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a") Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a series of tmfret-pairs comprised of single cysteine mutants
Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a series of tmfret-pairs comprised of single cysteine mutants
The circle and the basics of signal transduction. Course Outline. Topic #! Topic lecture! Silverthorn! Membranes (pre-requisite material)" "
") Homeostasis 03 The goal of this lecture is to discuss the concept of homeostasis and to introduce basic signal transduction mechanisms involved in homeostatic regulation The sections for this lecture are:
Homeostasis 03 The goal of this lecture is to discuss the concept of homeostasis and to introduce basic signal transduction mechanisms involved in homeostatic regulation The sections for this lecture are:
Nervous System Organization
 The Nervous System Nervous System Organization Receptors respond to stimuli Sensory receptors detect the stimulus Motor effectors respond to stimulus Nervous system divisions Central nervous system Command
The Nervous System Nervous System Organization Receptors respond to stimuli Sensory receptors detect the stimulus Motor effectors respond to stimulus Nervous system divisions Central nervous system Command
Delivery. Delivery Processes. Delivery Processes: Distribution. Ultimate Toxicant
 Delivery Ultimate Toxicant The chemical species that reacts with the endogenous target. Toxicity depends on the concentration (dose) of the ultimate toxicant at the target site Delivery Processes Absorption
Delivery Ultimate Toxicant The chemical species that reacts with the endogenous target. Toxicity depends on the concentration (dose) of the ultimate toxicant at the target site Delivery Processes Absorption
Membranes 2: Transportation
 Membranes 2: Transportation Steven E. Massey, Ph.D. Associate Professor Bioinformatics Department of Biology University of Puerto Rico Río Piedras Office & Lab: NCN#343B Tel: 787-764-0000 ext. 7798 E-mail:
Membranes 2: Transportation Steven E. Massey, Ph.D. Associate Professor Bioinformatics Department of Biology University of Puerto Rico Río Piedras Office & Lab: NCN#343B Tel: 787-764-0000 ext. 7798 E-mail:
Regulation of gene expression. Premedical - Biology
 Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Signaling to the Nucleus by an L-type Calcium Channel- Calmodulin Complex Through the MAP Kinase Pathway
 Signaling to the Nucleus by an L-type Calcium Channel- Calmodulin Complex Through the MAP Kinase Pathway Ricardo E. Dolmetsch, Urvi Pajvani, Katherine Fife, James M. Spotts, Michael E. Greenberg Science
Signaling to the Nucleus by an L-type Calcium Channel- Calmodulin Complex Through the MAP Kinase Pathway Ricardo E. Dolmetsch, Urvi Pajvani, Katherine Fife, James M. Spotts, Michael E. Greenberg Science
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Interpreting and evaluating biological NMR in the literature. Worksheet 1
 Interpreting and evaluating biological NMR in the literature Worksheet 1 1D NMR spectra Application of RF pulses of specified lengths and frequencies can make certain nuclei detectable We can selectively
Interpreting and evaluating biological NMR in the literature Worksheet 1 1D NMR spectra Application of RF pulses of specified lengths and frequencies can make certain nuclei detectable We can selectively
Gene regulation I Biochemistry 302. Bob Kelm February 25, 2005
 Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility
 proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility") (P&S Ch 5; Fer Ch 2, 9; Palm Ch 10,11; Zub Ch 9) A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility B.
(P&S Ch 5; Fer Ch 2, 9; Palm Ch 10,11; Zub Ch 9) A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility B.
Cells to Tissues. Peter Takizawa Department of Cell Biology
 Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Module Membrane Biogenesis and Transport Lecture 15 Ion Channels Dale Sanders
 Module 0220502 Membrane Biogenesis and Transport Lecture 15 Ion Channels Dale Sanders 9 March 2009 Aims: By the end of the lecture you should understand The principles behind the patch clamp technique;
Module 0220502 Membrane Biogenesis and Transport Lecture 15 Ion Channels Dale Sanders 9 March 2009 Aims: By the end of the lecture you should understand The principles behind the patch clamp technique;
Membrane Potentials, Action Potentials, and Synaptic Transmission. Membrane Potential
 Cl Cl - - + K + K+ K + K Cl - 2/2/15 Membrane Potentials, Action Potentials, and Synaptic Transmission Core Curriculum II Spring 2015 Membrane Potential Example 1: K +, Cl - equally permeant no charge
Cl Cl - - + K + K+ K + K Cl - 2/2/15 Membrane Potentials, Action Potentials, and Synaptic Transmission Core Curriculum II Spring 2015 Membrane Potential Example 1: K +, Cl - equally permeant no charge
The body has three primary lines of defense against changes in hydrogen ion concentration in the body fluids.
 ph and Nucleic acids Hydrogen Ion (H+) concentration is precisely regulated. The H+ concentration in the extracellular fluid is maintained at a very low level, averaging 0.00000004Eq/L. normal variations
ph and Nucleic acids Hydrogen Ion (H+) concentration is precisely regulated. The H+ concentration in the extracellular fluid is maintained at a very low level, averaging 0.00000004Eq/L. normal variations
The EGF Signaling Pathway! Introduction! Introduction! Chem Lecture 10 Signal Transduction & Sensory Systems Part 3. EGF promotes cell growth
 Chem 452 - Lecture 10 Signal Transduction & Sensory Systems Part 3 Question of the Day: Who is the son of Sevenless? Introduction! Signal transduction involves the changing of a cell s metabolism or gene
Chem 452 - Lecture 10 Signal Transduction & Sensory Systems Part 3 Question of the Day: Who is the son of Sevenless? Introduction! Signal transduction involves the changing of a cell s metabolism or gene
Introduction to electrophysiology. Dr. Tóth András
 Introduction to electrophysiology Dr. Tóth András Topics Transmembran transport Donnan equilibrium Resting potential Ion channels Local and action potentials Intra- and extracellular propagation of the
Introduction to electrophysiology Dr. Tóth András Topics Transmembran transport Donnan equilibrium Resting potential Ion channels Local and action potentials Intra- and extracellular propagation of the
introduction Endothelin-1 Inhibits Inward Rectifier K+ Channels in Rabbit Coronary Arterial Smooth Muscle Cells Through Protein Kinase C
 Current Topic 6 ก มภาพ นธ 2549 เวลา 9.00-12.00 น. น าเสนอโดย น.ส.สายวร ณ ศร ร ง 4836203 อาจารย ท ปร กษา อ.นพ.ล อชา บ ญทว ก ล Endothelin-1 Inhibits Inward Rectifier K+ Channels in Rabbit Coronary Arterial
Current Topic 6 ก มภาพ นธ 2549 เวลา 9.00-12.00 น. น าเสนอโดย น.ส.สายวร ณ ศร ร ง 4836203 อาจารย ท ปร กษา อ.นพ.ล อชา บ ญทว ก ล Endothelin-1 Inhibits Inward Rectifier K+ Channels in Rabbit Coronary Arterial
[Part II] Chemistry and Physiology
![[Part II] Chemistry and Physiology](/thumbs/74/70281995.jpg "[Part II] Chemistry and Physiology") [Part II] Quantitative Acid-Base Chemistry and Physiology [Chapter 10] Normal [SID] Peter Wilkes 10.1 Introduction This chapter will discuss issues surrounding the measurement and conceptualization of
[Part II] Quantitative Acid-Base Chemistry and Physiology [Chapter 10] Normal [SID] Peter Wilkes 10.1 Introduction This chapter will discuss issues surrounding the measurement and conceptualization of
ClC-1 inhibition by low ph and ATP INHIBITION OF SKELETAL MUSCLE ClC-1 CHLORIDE CHANNELS BY LOW INTRACELLULAR ph AND ATP
 JBC Papers in Press. Published on August 10, 2007 as Manuscript M703259200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m703259200 INHIBITION OF SKELETAL MUSCLE ClC-1 CHLORIDE CHANNELS
JBC Papers in Press. Published on August 10, 2007 as Manuscript M703259200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m703259200 INHIBITION OF SKELETAL MUSCLE ClC-1 CHLORIDE CHANNELS
Identification of a region of strong discrimination in the pore of CFTR
 Am J Physiol Lung Cell Mol Physiol 281: L852 L867, 2001. Identification of a region of strong discrimination in the pore of CFTR NAEL A. MCCARTY AND ZHI-REN ZHANG Departments of Physiology and Pediatrics,
Am J Physiol Lung Cell Mol Physiol 281: L852 L867, 2001. Identification of a region of strong discrimination in the pore of CFTR NAEL A. MCCARTY AND ZHI-REN ZHANG Departments of Physiology and Pediatrics,
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
REVIEW 2: CELLS & CELL COMMUNICATION. A. Top 10 If you learned anything from this unit, you should have learned:
 Name AP Biology REVIEW 2: CELLS & CELL COMMUNICATION A. Top 10 If you learned anything from this unit, you should have learned: 1. Prokaryotes vs. eukaryotes No internal membranes vs. membrane-bound organelles
Name AP Biology REVIEW 2: CELLS & CELL COMMUNICATION A. Top 10 If you learned anything from this unit, you should have learned: 1. Prokaryotes vs. eukaryotes No internal membranes vs. membrane-bound organelles
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1
 and Ksp/Cre;Pkd1") Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
12 Ion Channels in Epithelial Cells
 12 Ion Channels in Epithelial Cells Lawrence G. Palmer 12.1 Ion Channels and Epithelial Function Ion channels in epithelial cells serve to move ions, and in some cases fluid, between compartments of the
12 Ion Channels in Epithelial Cells Lawrence G. Palmer 12.1 Ion Channels and Epithelial Function Ion channels in epithelial cells serve to move ions, and in some cases fluid, between compartments of the
Quantitative Electrophysiology
 ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Tuesday, September 18, 2012 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Tuesday, September 18, 2012 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
Hole s Human Anatomy and Physiology Eleventh Edition. Chapter 2
 Hole s Human Anatomy and Physiology Eleventh Edition Shier Butler Lewis Chapter 2 1 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. CHAPTER 2 CHEMICAL BASIS OF
Hole s Human Anatomy and Physiology Eleventh Edition Shier Butler Lewis Chapter 2 1 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. CHAPTER 2 CHEMICAL BASIS OF