Putative inorganic carbon transport and accumulation systems associated with the CO2 concentrating mechanism in Chlamydomonas reinhardtii
|
|
|
- Rodger Fitzgerald
- 5 years ago
- Views:
Transcription

1 Graduate Theses and Dissertations Iowa State University Capstones, Theses and Dissertations 2009 Putative inorganic carbon transport and accumulation systems associated with the CO2 concentrating mechanism in Chlamydomonas reinhardtii Deqiang Duanmu Iowa State University Follow this and additional works at: Part of the Cell and Developmental Biology Commons, and the Genetics and Genomics Commons Recommended Citation Duanmu, Deqiang, "Putative inorganic carbon transport and accumulation systems associated with the CO2 concentrating mechanism in Chlamydomonas reinhardtii" (2009). Graduate Theses and Dissertations This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Graduate Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact
2 Putative inorganic carbon transport and accumulation systems associated with the CO 2 concentrating mechanism in Chlamydomonas reinhardtii by Deqiang Duanmu A dissertation submitted to the graduate faculty in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY Major: Molecular, Cellular, and Developmental Biology Program of Study Committee: Martin H. Spalding, Major Professor Diane C. Bassham Basil J. Nikolau David J. Oliver Yanhai Yin Iowa State University Ames, Iowa 2009 Copyright Deqiang Duanmu, All rights reserved.
3 ii TABLE OF CONTENTS CHAPTER 1. GENERAL INTRODUCTION AND LITERATURE REVIEW 1 General Introduction 1 Literature Review 2 Dissertation Organization 11 References 12 CHAPTER 2. KNOCKDOWN OF LIMITING-CO 2 -INDUCED GENE HLA3 DECREASES HCO 3 - TRANSPORT AND PHOTOSYNTHETIC Ci-AFFINITY IN CHLAMYDOMONAS REINHARDTII 16 Abstract 16 Introduction 17 Results 19 Discussion 22 Materials and Methods 27 Acknowledgements 28 References 29 CHAPTER 3. THYLAKOID LUMEN CARBONIC ANHYDRASE (CAH3) MUTATION SUPPRESSES AIR-DIER PHENOTYPE OF LCIB MUTANT IN CHLAMYDOMONAS REINHARDTII 47 Abstract 47 Introduction 48 Results 49 Discussion 52 Materials and Methods 57 Acknowledgements 60 References 60 CHAPTER 4. INSERTIONAL SUPPRESSORS OF CHLAMYDOMONAS REINHARDTII THAT RESTORE GROWTH OF AIR-DIER LCIB MUTANTS IN LOW-CO 2 71 Abstract 71 Introduction 72 Results 74 Discussion 78 Materials and Methods 81 Acknowledgements 84 References 84 CHAPTER 5. GENERAL SUMMARY 93 ACKNOWLEDGEMENTS 96
4 1 CHAPTER 1. GENERAL INTRODUCTION AND LITERATURE REVIEW General Introduction Carbon dioxide (CO 2 ) serves both as the substrate for photosynthesis and as an important signal to regulate plant growth and development, and variable CO 2 concentrations can impact photosynthesis, growth and productivity of plants. Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) is the initial enzyme for photosynthetic carbon assimilation in plants, although this enzyme has a poor apparent affinity for carbon dioxide. To generate a higher concentration of CO 2 for Rubisco and suppress oxygenation of RuBP, a wasteful side reaction, several strategies have evolved in photosynthetic organisms, including the well known C4 and CAM (Crassulacean acid metabolism) photosynthetic pathways. C 4 plants have developed a cooperative pathway between two specialized cell types: in the mesophyll cell, CO 2 is converted to HCO - 3, which is fixed by phosphoenolpyruvate carboxylase (PEPC) to form oxaloacetate (OAA). The OAA then is converted to a second C 4 acid (malate or aspartate) and transported to a neighboring bundle sheath cell for decarboxylation and refixation by Rubisco (Buchanan et al., 2000). In the biochemically similar CAM photosynthesis, inorganic carbon (HCO - 3 ) is fixed via PEPC into OAA, which is reduced to malate. The malate is stored in the vacuole and in the subsequent light period organic acid is released, decarboxylated and the resulting CO 2 assimilated by Rubisco in the Calvin cycle. A different type of CO 2 -concentrating mechanism (CCM) is adopted in many aquatic cyanobacteria and microalgae which together account for a large proportion (~50%) of global primary productivity. Primary CCM studies focus on cyanobacteria (prokaryotic blue-green algae) and Chlamydomonas reinhardtii (eukaryotic green alga). The three most important components of the CCM in these species are the presence of active Ci (HCO - 3 and CO 2 ) uptake systems, and of carbonic anhydrases that catalyze rapid interconversion of carbon dioxide and carbonic acid, and the existence of very specific, Rubisco-containing internal compartments to provide a localized elevated CO 2 concentration for Rubisco (Moroney and Ynalvez, 2007; Wang and Spalding, 2007; Spalding, 2008; Price et al., 2008).
5 2 Interestingly, no homologs of cyanobacterial CCM genes could be identified in C. reinhardtii, even though it has a functionally similar CCM, indicating the independent evolution of CCM in these microalgae and cyanobacteria. A particular focus of the research in our lab has been understanding the limiting-co 2 - acclimation response in C. reinhardtii, including both characterization of the limiting-co 2 signal transduction pathway and identification of the functional aspects of the acclimation response. We have been using insertional mutagenesis, suppressor screening, RNAi technology, etc, to investigate potential Ci transporters and non-acclimating mutants, which should provide valuable information to advance our understanding of the CCM and its regulatory pathways. Literature Review Brief overview of the C. reinhardtii CCM Aquatic green algae, like C. reinhardtii, must obtain carbon dioxide from inorganic carbon sources (Ci: CO 2, HCO - 3, CO 2-3 ) dissolved in the aqueous environments. The supply of Ci often is limiting and aquatic photosynthetic organisms also must overcome the fact that the diffusion rate of CO 2 in water is ~10,000 times slower than the rate of diffusion in air. The first enzyme responsible for photosynthetic carbon reduction, Rubisco, also has a poor apparent affinity for CO 2, with a K m (CO 2 ) in most algae greater than 25 µmol L -1. This means that Rubisco is functioning at less than 20% of its capacity in air-equilibrated water,where the CO 2 concentration is around 10 µmol L -1 (Moroney, 2001). To overcome these limitations, in response to limiting CO 2, either air level (~0.04% CO 2 ) or very low CO 2 (~0.01% CO 2 ) conditions, representative biophysical C i pumps operate: the CCM uses active Ci transport, probably both at the plasmalemma and the chloroplast envelope, to accumulate a high concentration of bicarbonate within the chloroplast (Sültemeyer et al, 1988; Palmqvist et al, 1988). The thylakoid lumen carbonic anhydrase (CAH3) plays an essential role in the rapid dehydration of the accumulated bicarbonate to release CO 2 into the pyrenoid, a Rubisco-containing internal compartment of the chloroplast, for assimilation by Rubisco (Price et al., 2002; Spalding et al., 2002; Moroney and Ynalvez, 2007). In C. reinhardtii, the CCM is not the only acclimation-related process induced during
6 3 limiting CO 2 acclimation. Its induction is accompanied by disappearance of stromal starch granules and formation of a starch sheath around the pyrenoid within the chloroplast, transient arrest of cell cycle in G1 phase, increased vacuolization and changes in mitochondrial distribution from a central position to a peripheral position between the chloroplast envelope and the plasmalemma (Geraghty and Spalding, 1996; Spalding, 2002; Giordano et al., 2005). Also, the CCM has reciprocal interaction with other metabolic processes, such as photosynthesis, photorespiration, respiration and other metabolic processes which share common key metabolites and redox equivalents (Xue et al., 1996; Cournac et al., 2002; Giordano et al., 2003). Putative Ci transporters Perhaps the most important elements of the eukaryotic CCM are the Ci uptake systems responsible for accumulation of Ci internally to a level several fold higher than that of the external medium. On the basis of evidence from isotope disequilibrium and mass spectrometric approaches, C. reinhardtii can take up both CO 2 and HCO - 3 during steady state photosynthesis. Although specific Ci transporters have not been identified yet, many researchers have confirmed the active Ci transport occurs across both the plasma membrane and chloroplast envelope (Sültemeyer et al., 1988; 1989; Palmqvist et al., 1988). At the level of the plasma membrane, photosynthetically-driven net uptake of Ci (mainly as CO 2 and HCO - 3 ) represents the largest nutrient flux that green alga encounter, but ultimately only CO 2 can serve as the substrate for Rubisco. CO 2 availability and supply rate are often severely limiting in aquatic environments, especially at alkaline ph, so microalgae, including C. reinhardtii, have undergone stringent selection for adaptations to help maximize the efficiency of Ci uptake through usage of HCO - 3. Also, regardless of which form of Ci, CO 2 or HCO - 3, is presented to the cell, HCO - 3 is the major species accumulated internally. Because HCO - 3 is about 1000-fold less permeable to lipid membranes than the uncharged CO 2 molecule, it is reasonable to hypothesize that direct HCO - 3 uptake across the plasmalemma and chloroplast envelope would require a protein carrier. Although our understanding of Ci uptake and regulation, as well as other aspects of CCM function in C. reinhardtii is limited, a better understanding of the similar CCM in
7 4 prokaryotic organisms, specifically the cyanobacteria Synechocystis and Synechococcus, has been gained. At least five different types of Ci transporters have been identified in cyanobacteria, including three bicarbonate transporters: 1) a high affinity, low CO 2 -induced, - sodium-independent HCO 3 transporter, BCT1, encoded by the cmpabcd operon and belonging to the ABC transporter family; 2) a low affinity, constitutively expressed, sodiumdependent HCO - 3 transporter, ictb; 3) SbtA, an inducible high affinity Na + -dependent SulP type HCO - 3 transporter; and two CO 2 uptake systems which also are critical in minimizing CO 2 leakage from the carboxysomes: 4) NDH-I 4, a putative plasma membrane localized low affinity constitutive CO 2 uptake system that contains a unique protein ChpX, which is involved in net unidirectional, light-dependent CO 2 hydration reactions; 5) NDH-I 3, a thylakoid membrane CO 2 uptake system that is inducible under Ci limitation and is of higher uptake affinity, also contains a unique protein ChpY which is 44% identical to ChpX and also performs a similar function (Price et al., 2002, 2008). Since the discovery of the CCM in C. reinhardtii, significant efforts have been devoted in identifying the Ci transport systems, however, only a few candidate Ci transport genes were characterized and the functions of these gene products still remain largely unknown (the following figure depicts a proposed CCM model in C. reinhardtii; Spalding, 2009). Transport across the plasma membrane It has been established that active transport of Ci could occur at the whole cell and the cells demonstrate a preference for CO 2, although it s questionable whether or not the observed CO 2 uptake at the plasma membrane is an active transport or rapid diffusion in response to active consumption of CO 2 internally (Amoroso et al., 1998). So far only two candidate plasmalemma Ci transporters have been identified in C. reinhardtii, HLA3/MRP1 and LCI1. Im et al. (2001) identified the HLA3 gene, encoding a putative ATP-binding cassette (ABC) type transporter of the multidrug-resistance-related proteins (MRP) subfamily, whose expression is activated by both high light (under conditions in which the cells were becoming CO 2 limited) and low CO 2, for which it was shown to be under the control of zinc-finger transcription factor CIA5, a master regulator for the response of C. reinhardtii to limiting CO 2 conditions (Miura et al., 2004). HLA3 was predicted by WoLF PSORT

8 5 (Horton et al., 2007) to be involved in the secretory pathway and possibly to be located in the plasma membrane. As HLA3 is not induced in high light under high CO 2 conditions, and not expressed in the cia5 mutant, the putative transport protein encoded by this gene may be Figure 8.3. The cell biology of the carbon concentrating mechanism. The movement of inorganic carbon (Ci) from the extracellular space to the thylakoid lumen via the cytosol and stroma is shown. The pyrenoid is a nonmembrane-bound compartment (boundary indicated as dashed line) within the stroma, and is the site of Rubisco localization. Membranes that serve as barriers for transport are shown as solid black lines. The purple ovals indicate transporters and question marks indicate unidentified, postulated carriers. Putative Ci transporters without supporting functional data are indicated without substrates. a vital component of the CCM, possibly even a Ci transporter. Interestingly, there is evidence for the inhibition of whole cell Ci uptake by the H + -ATPase inhibitor vanadate and MRP-associated transport is often sensitive to inhibition by vanadate (Karlsson et al., 1994). Based on cdna array analyses, a low-co 2 -inducible putative plasma membrane-type proton ATPase PMA2 was identified and the expression of PMA2 gene was also regulated by CIA5 (Fukuzawa, unpublished data). The functions of HLA3/MRP1 and PMA2 in Ci transport, the CCM or another aspect of limiting Ci acclimation will require further
9 6 investigations. The limiting-co 2 -inducible LCI1 gene is another candidate for plasma membrane Ci uptake. The LCI1 gene product is predicted to contain four predicted transmembrane helices and shows very little homology to any other proteins in the available database. Because the expression of LCI1 is controlled by CIA5 as well as a MYB type transcription factor LCR1, which is implicated in regulation of CCM genes, LCI1 is considered a good candidate for Ci transport (Burow et al., 1996; Yoshioka et al., 2004). However, the function of this gene product is unknown, and any putative role in Ci transport still requires further biochemical analyses. Transport into the chloroplast It is commonly accepted that chloroplasts are essential in Ci concentrating and active uptake of Ci at the chloroplast envelope has been demonstrated by using isolated intact chloroplast (Sültemeyer et al., 1988; Amoroso et al., 1998). Some promising candidate genes and proteins have been identified, and it is likely that one or more of the following proteins may be involved in active Ci uptake at chloroplast envelope. The candidate proteins include LCIA(NAR1.2), CCP1, CCP2, YCF10, LCIB and the unique LCIB protein family LCIC/LCID/LCIE. LCIA (also named NAR1.2) encodes a membrane protein belonging to the Formate/Nitrite Transporter (FNT) family of C. reinhardtii (Galvan et al., 2002; Miura et al., 2004). However, LCIA was considered to be a putative Ci transporter since the expression of LCIA is induced under limiting-co 2 conditions and is partially under the control of CIA5, a transcription factor required for induction of most other CCM genes. LCIA is predicted to be targeted to the chloroplast and has six transmembrane helices. When expressed in Xenopus oocytes, LCIA reportedly transports HCO - 3 but exhibited a very low affinity for - HCO 3 and high affinity nitrite transport, leaving its importance as Ci transporter questionable (Mariscal et al., 2006). Two other putative Ci transport proteins are CCP1 and CCP2, encoding two similar proteins with 96% identical amino acid sequences (Spalding and Jeffery, 1989; Ramazanov et al., 1993; Chen et al., 1997). These two proteins have six transmembrane domains, are associated with the chloroplast envelope, and have sequence similarity to the mitochondrial
10 7 carrier protein superfamily. This superfamily consists of many small proteins involved with transport across the mitochondrial inner membrane of various metabolites. Knockdown of CCP1 and CCP2 by RNAi exhibited similar Ci uptake characteristics as wild-type cells, although the RNAi strains grew poorly in low CO 2 levels. This might indicate that these two putative transporters are involved with transport of metabolic intermediates important in limiting CO 2, or the functions of these two proteins as Ci transporters are masked by some other Ci transport systems under the conditions tested (Pollock et al., 2005). The product of plastid gene YCF10 displays sequence similarity with the cyanobacterial CotA product which is altered in mutants defective in CO 2 transport and proton extrusion (Rolland et al., 1997). YCF10 protein is localized in the inner chloroplast envelope and disruption of ycf10 expression resulted in high light sensitivity and decreased Ci uptake affinity. However, YCF10 may not participate directly in Ci uptake and instead might play an important role in proton exchange to maintain electrical and ph homeostasis during uptake of CO 2 and HCO - 3, a similar role as the cyanobacterial CotA protein (Kaplan and Reinhold, 1999). Another limiting-co 2 -inducible, CIA5-regulated gene LCIB encodes a soluble protein which is predicted to be targeted to chloroplast (Miura et al., 2004; Wang and Spalding, 2006). Two LCIB mutants, pmp1 and ad1, which contain a nonsense mutation or a deletion mutation of LCIB gene, respectively, cannot grow in low CO 2 (air) but can grow in very low CO 2. Ci uptake in L-CO 2 -acclimated pmp1/ad1 cells was almost eliminated, suggesting an essential role of LCIB as a Ci transport system specifically in L-CO 2 physiological state. While it is unlikely that LCIB is a transporter protein itself, since it lacks any significant hydrophobic transmembrane helices, LCIB is speculated to be either a regulator of Ci transport/accumulation or an essential subunit of a unidentified Ci transport complex (Wang and Spalding, 2006). Three additional genes, LCIC, LCID and LCIE, were identified by BLAST searches as sharing sequence similarity (71~78%) in their gene products with LCIB, thus constituting an LCIB protein family. LCIC and LCID are also upregulated under limiting-co 2 conditions while expression level of LCIE was very low in L-CO 2 and VL-CO 2 acclimated cells (Wang and Spalding, 2006). Yeast two hybrid analysis indicates LCIB could physically interact
11 8 with LCIC (Wang and Spalding, unpublished). Thus it is tempting to speculate LCIB and LCIC are involved in or regulating a Ci transport complex at least in L-CO 2 conditions, while a similar complex including LCID and LCIE may function in VL-CO 2 acclimation state (Wang and Spalding 2006, 2007; Spalding, 2008). The C. reinhardtii genome also encodes two Rhesus (RHP1 and RHP2) proteins which have similarity to the Rh proteins in the human red blood membrane. Both RHP1 and RHP2 are predicted to be chloroplast membrane proteins, and the expression of RHP1 is low in limiting CO 2 but upregulated by high CO 2. Knockdown of RHP1 by RNAi resulted in a growth defect in high CO 2 but normal growth in air. RHP1 was proposed as a bidirectional CO 2 channel and may allow rapid equilibration of CO 2 under high CO 2 growth conditions to provide sufficient CO 2 for photosynthesis (Soupene et al., 2002, 2004; Kustu and Inwood, 2006). The functions, if any, of the RHP1 and RHP2 in the CCM are not clear. Transport into the thylakoid lumen Regardless of which type of Ci, CO 2 or HCO - 3, is transported across the plasma membrane and chloroplast envelope, HCO - 3 is the major Ci species accumulated within the - alkalinic chloroplast stroma. This accumulated HCO 3 serves as the source of high concentrations of CO 2 for Rubisco after CAH3-catalyzed dehydration in the acidic thylakoid lumen. However, it is still unclear how HCO - 3 is transported across the thylakoid membrane into the lumen. An active HCO - 3 transporter or a facilitated diffusion carrier is proposed, while the identities of these proteins remain to be clarified (Moroney and Ynalvez, 2007; Spalding, 2008). Carbonic anhydrases Up to now, nine carbonic anhydrase genes were identified in C. reinhardtii, including CAH1-CAH9, and some of these proteins are proposed to have possible functions in an active CCM (Moroney and Ynalvez, 2007). Two periplasmic CAs, CAH1 and CAH2, share high sequence similarity although CAH1 is induced by limiting-co 2 and the expression of CAH2 is repressed under such conditions (Moroney et al., 1985; Fukuzawa et al., 1990; Rawat and Moroney, 1991). CAH1 was proposed to convert HCO - 3 to CO 2 at the cell surface to facilitate the uptake of CO 2 at the
12 9 plasma membrane especially under high ph. However, a deletion-mutant of CAH1 gene did not demonstrate any significant growth differences from the wild-type cells across all ph conditions (ph 6 to ph 9), indicating CAH1 may not be essential for a functional CCM (Van and Spalding, 1999). CAH3 is located in the thylakoid lumen and the expression of CAH3 message is slightly upregulated under limiting-co 2 conditions (Karlsson et al., 1998). The essential role of CAH3 in the function of the CCM has been well established. C. reinhardtii strains defective in CAH3 cannot grow in limiting-co 2 conditions although a large pool of Ci, presumably HCO - - 3, is accumulated, suggesting cah3 mutants can not use accumulated internal HCO 3 efficiently for photosynthesis (Funke et al., 1997; Raven 1997; Karlsson et al., 1998; Hanson et al., 2003). CAH4 and CAH5 are two identical mitochondrial CAs that are induced under limiting- CO 2 conditions (Eriksson et al., 1998). The exact roles of these two CAs are still not clear, and multiple hypotheses have been given regarding the potential function of the two proteins, including a proposed role to buffer the mitochondrial matrix during photorespiration, or - recapturing CO 2 and supplying HCO 3 to PEP carboxylase for NH + 4 assimilation under certain conditions (Eriksson et al., 1996; Giordano et al., 2003). CAH6 is a chloroplast stroma CA, and its expression is not affected by changing CO 2 concentrations (Mitra et al., 2004, 2005). The location of CAH6 suggests it might be involved in trapping CO 2 that diffuses out of the pyrenoid and the alkaline ph of chloroplast - stroma under active photosynthesis could facilitate the conversion of CO 2 to HCO 3 (Mitra et al., 2004). Three more recently identified CA genes are CAH7, CAH8 and CAH9. CAH7 and CAH8 are constitutively expressed at the transcript and protein level and these two proteins share 63% similarity (Moroney and Ynalvez, 2007). CAH7 is predicted to be localized to chloroplast and CAH8 was found to be in the periplasmic space by immunolocalization (Ynalvez et al., 2008). Interestingly, CAH1 and CAH8 both are localized to the periplasmic space, but CAH8 resides closer to the cell s membrane while CAH1, which is more abundantly expressed under limiting-co 2, is localized farther away from the plasma membrane. The location of CAH8 suggests that the gene product might be part of the Ci
13 10 uptake system in C. reinhardtii and the discovery of CAH8 raised possibility that the observed lack of growth defects of a cah1 deletion mutation may have been caused by partial complementation by CAH8. The recently discovered CAH9 is tentatively assigned to be localized to the cytoplasm. An interesting feature of CAH9 is that its sequence is more closely related with bacterial CAs than with other C. reinhardtii CAs. The role of CAH9 in CCM remains to be determined (Mitra et al., 2005). Role of the pyrenoid The pyrenoid has recently been regarded as one of the essential components of the CCM by providing a localized elevated CO 2 concentration for Rubisco, and most eukaryotic photosynthetic algae, including C. reinhardtii, contain a pyrenoid (Moroney and Ynalvez, 2007). Prokaryotic photosynthetic cyanobacteria have a functionally similar CCM as C. reinhardtii and have a similar specific localization of Rubisco in a compartment called carboxysome within the cytoplasm. The implication of this similarity is that the pyrenoid and carboxysome are functionally similar compartments. Some cyanobacterial mutants with defects in either carboxysome carbonic anhydrase or carboxysomal shell proteins require elevated CO 2 for photosynthesis (Schwarz et al., 1995; Price and Badger, 2003). Although pyrenoid-less microalgae possessing a CCM, such as some strains of Chloromonas, have been identified, these strains have small internal Ci pools (24~31µM) compared with microalgae containing typical pyrenoids (231~252 µm; Morita et al., 1998). Multiple CO 2 -concentration-dependent acclimation states Most of the studies on CCM have focused on two acclimation states: a high CO 2 (H-CO 2 ) state (5% - 0.5% CO 2 ) and a low CO 2 (L-CO 2 ) state (air level; 0.4% % CO 2 ). Recently, another limiting-co 2 acclimation state was hypothesized based on growth, photosynthesis and gene expression characteristics, a very low CO 2 (VL-CO 2 ) state (0.01% % CO 2 ) (Vance and Spalding, 2005). Strong support for two distinct limiting-co 2 states (L-CO 2 and VL-CO 2 ) comes from the allelic HCR (high-co 2 -requiring) mutants, pmp1 and ad1, which grow as well (pmp1) or nearly as well (ad1) as wild type cells in both H-CO 2 and VL-CO 2 conditions while only dying in L-CO 2, indicating a deficient Ci transport and/or
14 11 accumulation system only in the L-CO 2 acclimation state (Wang and Spalding, 2006). Quantitative physiological analyses of L-CO 2 or VL-CO 2 acclimated wild-type cells also confirmed the existence of a multi-tiered regulatory program which is controlled by CO 2 concentrations (Vance and Spalding, 2005). Challenges and future directions for study of Ci transporters While a number of candidate Ci transport proteins have been identified in C. reinhardtii, none of them could be definitely proven to transport Ci in vivo or in vitro. One of the biggest challenges in studying the CCM is identifying the essential transport components, clarifying their substrate specificities, other regulatory components or subunits in Ci transport complexes and elucidating their mechanisms. At least several good candidates have been identified for Ci transport across the plasma membrane and the chloroplast envelope, but there still is no candidate thylakoid HCO - 3 transporter identified yet. - In cyanobacteria, a variety of active CO 2 and HCO 3 uptake systems have been characterized. These Ci transport complexes have complementary functions, and disruption of one transport complex does not result in significant growth inhibition under limiting-co 2 (Price et al., 2002, 2008). A similar scenario might exist in C. reinhardtii and would thus prevent identification of putative Ci transporters by phenotype screening. A forward genetic approach, either by knocking down candidate genes with RNAi or gain-of-function analysis in C. reinhardtii and/or other heterologous organisms, may be one way of proceeding. Transcriptional analysis of gene expression profiles, identification of proteins interacting with known CCM components, and comparative bioinformatic analysis of multiple genomes, including C. reinhardtii and other microalgae, could also be helpful in identifying key Ci transporters. Dissertation Organization The thesis is written in the format to include two published papers (Chapter 2 and 3). The first paper demonstrates the characterization of three HCO - 3 transport and accumulation systems and the second paper describes newly isolated insertional suppressor of lcib mutant that dies in air-level of CO 2 concentration. Other identified lcib suppressors were
15 12 characterized and described in chapter 4. The dissertation is concluded with a general summary in chapter 5. References Amoroso G, Sültemeyer D, Thyssen C, Fock HP (1998) Uptake of HCO 3 - and CO 2 in cells and chloroplasts from the microalgae Chlamydomonas reinhardtii and Dunaliella tertiolecta. Plant Physiol 116: Buchanan, Gruissem and Jones (2000) Biochemistry & Molecular Biology of Plants. Chapter 12.9: Burow MD, Chen ZY, Mouton TM, Moroney JV (1996) Isolation of cdna clones of genes induced upon transfer of Chlamydomonas reinhardtii cells to low CO 2. Plant Mol Biol 31: Chen ZY, Lavigne LL, Mason CB, Moroney JV (1997) Cloning and overexpression of two cdnas encoding the low-co 2 -inducible chloroplast envelope protein LIP-36 from Chlamydomonas reinhardtii. Plant Physiol 114: Cournac L, Latouche G, Cerovic Z, Redding K, Ravenel J, Peltier G (2002) In vivo interactions between photosynthesis, mitorespiration, and chlororespiration in Chlamydomonas reinhardtii. Plant Physiol 129: Eriksson M, Karlsson J, Ramazanov Z, Gardestrom P, Samuelsson G (1996) Discovery of an algal mitochondrial carbonic anhydrase: molecular cloning and characterization of a low-co 2 -induced polypeptide in Chlamydomonas reinhardtii. Proc Natl Acad Sci U S A 93: Eriksson M, Villand P, Gardestrom P, Samuelsson G (1998) Induction and regulation of expression of a low-co 2 -induced mitochondrial carbonic anhydrase in Chlamydomonas reinhardtii. Plant Physiol 116: Fukuzawa H, Fujiwara S, Yamamoyo Y, Dionisio-Sese ML, Miyachi S (1990) cdna cloning, sequence and expression of carbonic anhydrase in Chlamydomonas reinhardtii: regulation by environmental CO 2 concentration. Proc Natl Acad Sci U S A 87: Funke RP, Kovar JL, Weeks DP (1997) Intracellular carbonic anhydrase is essential to photosynthesis in Chlamydomonas reinhardtii at atmospheric levels of CO 2. Plant Physiol 114: Kaplan A, Reinhold L (1999) The CO 2 -concentrating mechanism of photosynthetic microorganisms. Annu Rev Plant Physiol Plant Mol Biol 50: Galvan A, Rexach J, Mariscal V, Fernandez E (2002) Nitrite transport to the chloroplast in Chlamydomonas reinhardtii: molecular evidence for a regulated process. J Exp Bot 53: Geraghty AM, Spalding MH (1996) Molecular and Structural Changes in Chlamydomonas under Limiting CO 2 (A Possible Mitochondrial Role in Adaptation). Plant Physiol 111: Giordano M, Norici A, Forssen M, Eriksson M, Raven JA (2003) An anaplerotic role for mitochondrial carbonic anhydrase in Chlamydomonas reinhardtii. Plant Physiol 132:
16 13 Giordano M, Beardall J, Raven JA (2005) CO 2 concentraing mechanisms in algae: Mechanisms, environmental modulation and evolution. Annu Rev Plant Biol 56: Grossman AR, Croft M, Gladyshev VN, Merchant SS, Posewitz MC, Prochnik S, Spalding MH (2007) Novel metabolism in Chlamydomonas through the lens of genomics. Curr Opin Plant Biol 10: Hanson DT, Franklin LA, Samuelsson G, Badger MR (2003) The Chlamydomonas reinhardtii cia3 mutant lacking a thylakoid lumen-localized carbonic anhydrase is limited by CO 2 supply to Rubisco and not photosystem II function in vivo. Plant Physiol 132: Horton P, Park KJ, Obayashi T, Fujita N, Harada H, Adams-Collier CJ, Nakai K (2007) WoLF PSORT: protein localization predictor. Nucleic Acids Res Jul 35 (Web Server issue): W Im CS, Grossman AR (2001) Identification and regulation of high light-induced genes in Chalmydomonas reinhardtii. Plant J 30: Jan Karlsson, Ziyadin Ramazanov, Thomas Hiltonen, Per Gardeström and Göran Samuelsson (1994) Effect of vanadate on photosynthesis and the ATP/ADP ratio in low-co 2 -adapted Chlamydomonas reinhardtii cells. Planta 192:46-51 Karlsson J, Clarke AK, Chen ZY, Hugghins SY, Youn-II P, Husic HD, Moroney JV, Samuelsson G (1998) A novel α-type carbonic anhydrase associated with the thylakoid membrane in Chlamydomonas reinhardtii is required for growth at ambient CO 2. EMBO J 17: Kustu S, Inwood W (2006) Biological gas channels for NH 3 and CO 2 : evidence that Rh (Rhesus) proteins are CO 2 channels. Transfus Clin Biol 13: Mariscal V, Moulin P, Orsel M, Miller AJ, Fernández E, Galván A (2006) Differential regulation of the Chlamydomonas Nar1 gene family by carbon and nitrogen. Protist 157: Mitra M, Lato SM, Ynalvez RA, Xiao Y, Moroney JV (2004) Identification of a new chloroplast carbonic anhydrase in Chlamydomonas reinhardtii. Plant Physiol 135: Mitra M, Mason CB, Xiao Y, Ynalvez RA, Lato SM, Moroney JV (2005) The carbonic anhydrase gene families of Chlamydomonas reinhardtii. Can J Bot 83: Miura K et al. (2004) Expression profiling-based identification of CO 2 -responsive genes regulated by CCM1 controlling a Carbon-concentrating mechanism in Chlamydomonas reinhardtii. Plant Physiol 135: Morita E, Abe T, Tsuzuki M, Fujiwara S, Sato N, Hirata A, Sonoike K, Nozaki H (1998) Presence of the CO 2 concentrating mechanism in some species of the pyrenoid-less freeliving algal genus Chloromonas (Volvocales, Chlorophyta). Planta 204: Moroney JV, Husic HD, Tolbert NE (1985) Effect of carbonic anhydrase inhibitors on inorganic carbon accumulation by Chlamydomonas reinhardtii. Plant Physiol 79: Moroney JV (2001) Algal Carbon Dioxide Concentrating Mechanisms. Encyclopedia of Life Sciences DOI: / a pub2 Moroney JV, Ynalvez RA (2007) Proposed carbon dioxide concentrating mechanism in Chlamydomonas reinhardtii. Eukaryot Cell 6: Palmqvist K, Sjöberg S, Samuelsson G (1988) Induction of inorganic carbon accumulation in the unicellular green algae Scenedesmus obliquus and Chlamydomonas reinhardtii. Plant Physiol. 87:
17 14 Pollock SV, Prout DL, Godfrey AC, Lemaire SD, Moroney JV (2004) The Chlamydomonas reinhardtii proteins Ccp1 and Ccp2 are required for long-term growth, but are not necessary for efficient photosynthesis, in a low-co 2 environment. Plant Mol Biol 91: Price GD, Maeda SI, Omata T, Badger MR (2002) Modes of active inorganic carbon uptake in the cyanobacterium, Synechococcus sp. PCC7942. Funct Plant Biol 29: Price GD, Badger MR (2003) CO 2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution. J Exp Bot 77:83-94 Price GD, Badger MR, Woodger FJ, Long BM (2008) Advances in understanding the cyanobacterial CO 2 -concentrating mechanism (CCM): functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J Exp Bot 59: Ramazanov Z, Mason CB, Geraghty AM, Spalding MH, Moroney JV (1993) The low CO 2 -inducible 36 kda protein is localized to the chloroplast envelope of Chlamydomonas reinhardtii. Physiol Plant 84: Raven JA (1997) CO 2 -concentrating mechanisms: a direct role for thylakoid lumen acidification? Plant Cell Environ 20: Rawat M, Moroney JV (1991) Partial characterization of a new isoenzyme of carbonicanhydrase isolated from Chlamydomonas reinhardtii. J Biol Chem 266: Rolland N, Dorne AJ, Amoroso G, Sultemeyer DF, Joyard J, Rochaix JD (1997) Disruption of the plastid ycf10 open reading frame affects uptake of inorganic carbon in the chloroplast of Chlamydomonas. EMBO J 16: Schwarz R, Reinhold L, Kaplan A (1995) Low activation state of ribulose-1,5- bisphosphate carboxylase/oxygenase in carboxysome-defective Synechococcus mutants. Plant Physiol 108: Soupene E, King N, Feild E, Liu P, Niyogi KK, Huang CH, Kustu S (2002) Rhesus expression in a green alga is regulated by CO 2. Proc Natl Acad Sci U S A 99: Soupene E, Inwood W, Kustu S (2004) Lack of the Rhesus protein Rh1 impairs growth of the green alga Chlamydomonas reinhardtii at high CO 2. Proc Natl Acad Sci U S A 101: Spalding MH, Jeffrey M (1989) Membrane-associated polypeptides induced in Chlamydomonas by limiting CO 2 concentrations. Plant Physiol 89: Spalding MH, Van K, Wang Y, Nakamura Y (2002) Acclimation of Chlamydomonas to changing carbon availability. Funct Plant Biol 29: Spalding MH (2008) Microalgal carbon-dioxide-concentrating mechanisms: Chlamydomonas inorganic carbon transporters. J Exp Bot 59: Spalding MH (2009) The Chlamydomonas Sourcebook Second Edition: Organellar and Metabolic Processes, volume 2, chapter 8: Sültemeyer, DF, Klöck G, Kreuzberg K, Fock HP (1988) Photosynthesis and apparent affinity for dissolved inorganic carbon by cells and chloroplasts of Chlamydomonas reinhardtii grown at high and low CO 2 concentrations. Planta 176: Sültemeyer DF, Miller AG, Espie GS, Fock HP, Canvin DT (1989) Active CO(2) Transport by the Green Alga Chlamydomonas reinhardtii. Plant Physiol 89: Van K, Spalding MH (1999) Periplasmic carbonic anhydrase (Cah1) structural gene mutant in Chlamydomonas reinhardtii. Plant Physiol 120:
18 15 Vance P, Spalding MH (2005) Growth, photosynthesis and gene expression in Chlamydomonas over a range of CO 2 concentrations and CO 2 /O 2 ratios: CO 2 regulates multiple acclimation states. Can J Bot 83: Wang Y, Spalding MH (2006) An inorganic carbon transport system responsible for acclimation specific to air levels of CO 2 in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 103: Wang Y, Spalding MH (2007) CO 2 Concentrating Mechanisms in Eukaryotic Microalgae. Funct Plant Sci Biotechnol 1: Xue X, Gauthier DA, Turpin DH, Weger HG (1996) Interactions between photosynthesis and Respiration in the green alga Chlamydomonas reinhardtii (Characterization of Light- Enhanced Dark Respiration). Plant Physiol 112: Ynalvez RA, Xiao Y, Ward AS, Cunnusamya K, Moroney JV (2008) Identification and characterization of two closely related β-carbonic anhydrases from Chlamydomonas reinhardtii. Physiol Plant 133:15-26 Yoshioka S, Taniguchi F, Miura K, Inoue T, Yamano T, Fukuzawa H (2004) The novel Myb transcription factor LCR1 regulates the CO 2 -responsive gene Cah1, encoding a periplasmic carbonic anhydrase in Chlamydomonas reinhardtii. Plant Cell 16:
19 16 CHAPTER 2. KNOCKDOWN OF LIMITING-CO 2 -INDUCED GENE HLA3 DECREASES HCO - 3 TRANSPORT AND PHOTOSYNTHETIC Ci-AFFINITY IN CHLAMYDOMONAS REINHARDTII A paper published in PNAS 106(14): (2009) Deqiang Duanmu 1, Amy R. Miller 2, Kempton M. Horken 2, Donald P. Weeks 2, and Martin H. Spalding 1,3 Abstract The CO 2 -concentrating mechanism (CCM) of Chlamydomonas reinhardtii and other microalgal species is essential for photosynthetic growth in most natural settings. A great deal has been learned regarding the CCM in cyanobacteria, including identification of inorganic carbon (Ci; CO 2 and HCO - 3 ) transporters, while specific knowledge of analogous transporters has remained elusive in eukaryotic microalgae such as C. reinhardtii. Here, we have investigated whether the limiting-co 2 -inducible, putative ABC-type transporter, HLA3 - might function as a HCO 3 transporter by evaluating the effect of ph on growth, photosynthetic Ci affinity and [ 14 C]-Ci uptake in very low CO 2 conditions following RNA interference (RNAi) knockdown of HLA3 mrna levels in wild-type and mutant cells. Although knockdown of HLA3 mrna alone resulted in only modest but high-ph-dependent decreases in photosynthetic Ci affinity and Ci uptake, the combination of nearly complete 1 Department of Genetics, Development and Cell Biology, Iowa State University, Ames, IA Department of Biochemistry, University of Nebraska, Lincoln, NE Author for correspondence Author contributions: D.D., D.P.W., and M.H.S. designed research; D.D., A.R.M., and K.M.H. performed research; D.D. and M.H.S. analyzed data; and D.D., D.P.W., and M.H.S. wrote the paper.
20 17 knockdown of HLA3 mrna with mutations in LCIB (encodes limiting-ci inducible, plastid localized protein required for normal Ci uptake or accumulation in low CO 2 conditions) and/or simultaneous, apparently off-target knockdown of LCIA mrna (encodes limiting-ci inducible, plastid envelope protein reported to transport HCO - 3 ) resulted in dramatic decreases in growth, Ci uptake and photosynthetic Ci affinity, especially at ph 9.0, where HCO - 3 is the predominant form of available Ci. Collectively, the data presented here provide compelling evidence that HLA3 is directly or indirectly involved in HCO - 3 transport and - provide additional evidence supporting a role for LCIA in chloroplast envelope HCO 3 transport and for LCIB in chloroplast Ci accumulation. Introduction Carbon dioxide (CO 2 ) is the substrate for net photosynthetic carbon assimilation via the enzyme Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) and, because of its variable availability, especially in aquatic environments, CO 2 is an important regulator of the growth and development of photosynthetic organisms. The green microalga Chlamydomonas reinhardtii responds to limitations in inorganic carbon (Ci; CO 2 and HCO - 3 ) supply by inducing a functional CO 2 -concentrating mechanism (CCM), which serves both to increase the photosynthetic rate and to suppress the wasteful oxygenation of RuBP (1, 2). The CCM uses active C i transport, both at the plasma membrane and the chloroplast envelope (3, 4), to accumulate HCO - 3 within the chloroplast stroma. The details of microalgal Ci transport remain unclear, but accumulated stromal HCO - 3 is readily dehydrated by the thylakoid lumen carbonic anhydrase (CAH3) to release CO 2 for assimilation by Rubisco (2, 5, 6). Although Ci transport and accumulation systems are critical elements of the microalgal CCM, specific C i transporters have yet to be identified. Three candidate Ci transporters identified in C. reinhardtii include LCIA, LCIB and HLA3 (7-11). LCIA (Nar1.2, predicted MW 34.8 kda), is a limiting-co 2 induced chloroplast envelope protein of the Formate/Nitrite Transporter (FNT) protein family (7, 8), and unlike other C. reinhardtii NAR1 family members, is considered a candidate Ci transporter, because its expression is
21 18 regulated by CO 2 irrespective of the nitrogen source (8), and, when expressed in Xenopus oocytes, is reported to confer HCO - 3 transport activity (12). The C. reinhardtii pmp1 mutant, originally characterized as deficient in Ci transport, grows well in both high and very low CO 2 conditions while dying only in low (air level) CO 2, indicating a deficient Ci transport or accumulation system only in low CO 2 (13, 14). The defective gene in pmp1 and an allelic mutant ad1 was identified as the limiting-co 2 inducible gene, LCIB, which encodes a soluble, plastid protein of unknown function. LCIB (predicted MW 48 kda) was postulated to represent a component of a plastid envelope Ci transporter (9) but more recently was implicated in recapture of CO 2 inside the chloroplast (15). The mutant phenotype of pmp1/ad1 distinguishes it from most other non-acclimating mutants and indicates the existence of at least three CO 2 -concentration-dependent acclimation states, subsequently defined as a high CO 2 (H-CO 2 ) state (5% - 0.5% CO 2 ), low CO 2 (L-CO 2 ) state (air level; ~0.04% CO 2 ) and very low CO 2 (VL-CO 2 ) state (~0.01% CO 2 ) (16). HLA3 (CrMRP1, predicted MW kda) is a limiting-co 2 induced member of the multidrug-resistance-related protein (MRP) subfamily of the ATP-binding cassette (ABC) transporter superfamily, best known in animals as drug efflux pumps, although plant MRP type ABC transporters are involved in diverse transport processes (10, 11, 17, 18). Seven putative MRP type ABC transporters were identified in C. reinhardtii (11), but only CrMRP1 (HLA3) expression is controlled by CIA5, a master regulator of the limiting-co 2 induced CCM (10, 19, 20). CrMRP1/HLA3 contains a full size ABC-MRP domain arrangement (10), which argues for an ATP-dependent transport function during limiting- CO 2 acclimation, possibly HCO - 3 transport across the plasma membrane, since HLA3 is predicted (WoLF PSORT; 21) to be targeted to the secretory pathway. In spite of numerous screens for H-CO 2 requiring C. reinhardtii mutants defective in Ci transport, the allelic LCIB mutants pmp1 and ad1 are the only mutants identified with any putative connection to Ci transport (2). Thus we speculated that, as in cyanobacteria (22), multiple Ci transport systems in C. reinhardtii may provide overlapping functions that complement each other, making it unlikely to identify a single Ci transporter mutant by
22 19 phenotype. We hypothesized that LCIB mutants are able to grow in VL-CO 2 because the lost function of LCIB in VL-CO 2 acclimated C. reinhardtii is complemented by another Ci uptake system, possibly involving HLA3. To test this hypothesis we used RNA interference (RNAi) to knockdown expression of HLA3, predicting that, even if knockdown of HLA3 mrna alone produced no apparent phenotype, HLA3 mrna knockdown in an LCIB mutant background might reveal complementary or overlapping functions between these two Ci uptake and accumulation candidates. In this paper, we demonstrate marked knockdown of HLA3 mrna levels with two distinct RNAi constructs. We also demonstrate the associated inability of HLA3 RNAi knockdown mutants, when combined either with LCIB mutations or simultaneous knockdown of LCIA mrna, to grow in VL-CO 2 at ph 9, at which the predominant form of Ci is HCO - 3. These results strongly support the hypothesis that HLA3 is directly or indirectly required for HCO - 3 transport. The data also provide additional evidence that LCIB is involved in HCO - 3 accumulation in chloroplast and that LCIA is involved in chloroplast HCO - 3 transport in C. reinhardtii. Results TR-RNAi vector decreases expression of both HLA3 and LCIA. Intrinsic direct repeats are not used as often as inverted repeats for RNAi, but also induce posttranscriptional gene silencing effectively in tobacco (23). To test the effectiveness of such tandem repeats (TR) in C. reinhardtii, we transformed the TR-RNAi plasmid AphVIII- HLA3 TR (Supporting information (SI) Fig. S1A) into wild type CC125 and pmp1, which has an air-dier phenotype resulting from a mutation in LCIB (9). Of approximately paromomycin resistant, TR-RNAi transformants of each CC125 and pmp1, approximately 1.5% showed a strong decrease in HLA3 mrna abundance. Four CC125-TR-RNAi lines (CC125TR30, CC125TR33, CC125TR64, and CC125TR225) and two pmp1-tr-rnai lines (pmp1tr29 and pmp1tr51) were selected for further analysis, along with additional transformants chosen as controls (CC125C55, CC125C80, pmp1c124 and pmp1c130). Unexpectedly, expression of another limiting-co 2 inducible gene, LCIA, also was markedly
23 20 decreased in the CC125-TR-RNAi and pmp1-tr-rnai knockdown lines but not in the control lines (Fig. 1). Message abundance for other limiting-co 2 inducible genes tested (LCIB, CCP1/2, CAH1, LCI1) appeared unaffected (data not shown). We hypothesize that this apparent co-knockdown of LCIA mrna results from moderate sequence similarity between LCIA and the HLA3 sequences used in TR-RNAi. VL-CO 2 acclimated CC125-TR-RNAi knockdown lines have severe inhibition of growth, photosynthesis and Ci accumulation at high ph. The large decrease in HLA3 and LCIA expression in CC125-TR-RNAi lines CC125TR30, CC125TR33, CC125TR64, and CC125TR225 resulted in no apparent effect on growth in VL-CO 2 at ph 6.0 or 7.3, but severe inhibition of growth at ph 9.0 (Fig. 2A and Fig. S2). Apparent photosynthetic Ci affinity of CC125TR30 and CC125TR64 also was unaffected at ph 6.0 (Table S1) but in 5 µm Ci was inhibited by 55-70% and 62-87% at ph 7.3 and ph 9.0, respectively (Table 1). Consistent with spot tests and photosynthetic affinity results, internal Ci accumulation in CC125TR30 and CC125TR64 was only slightly inhibited at ph 7.3 but was inhibited 75-85% at ph 9.0 in 50 µm Ci (Table 2). VL-CO 2 acclimated pmp1-tr-rnai knockdown lines require high CO 2 for survival. All 200 selected pmp1-tr-rnai transformants grew normally in H-CO 2, none grew in L- CO 2 (consistent with the pmp1 air-dier phenotype), and 2 transformants also could not grow in VL-CO 2, in contrast with the parent pmp1 and the control pmp1-tr-rnai lines (Fig. 2B). The large decrease in HLA3 and LCIA expression in pmp1tr29 and pmp1tr51 resulted in dramatic decreases in photosynthetic Ci affinity at ph 7.3 and ph 9.0 (Table 1) and marked inhibition of intracellular Ci accumulation (Table 2), suggesting a very low capacity for HCO - 3 usage when expression levels of both HLA3 and LCIA are suppressed. However, at odds with the lack of growth in the VL-CO 2 acclimated pmp1-tr-rnai strains (Fig. S2), no decrease in photosynthetic Ci affinity was observed at ph 6.0 (Table S1). It also is noteworthy that a loss of the H-CO 2 growth requirement of pmp1tr51 over a period of months correlated with a recovery of both HLA3 and LCIA mrna abundance (data not shown).
24 21 IR-RNAi vector decreases expression of HLA3 alone. An inverted repeat (IR) RNAi vector (AphVIII-HLA3 IR) also was constructed using the HLA3 3 UTR sequence (Fig. S1B). Of approximately 100 paromomycin resistant RNAi transformants of each cw10 and ad1, wall-less WT and LCIB-defective strains, respectively, ~10% of the putative RNAi lines showed a strong decrease in HLA3 mrna abundance. Among these, two cw10-ir- RNAi lines (cw10ir21 and cw10ir32) and three ad1-ir-rnai transformants (ad1ir53, ad1ir311 and ad1ir312) were selected for further analysis, along with control transformant lines cw10c10, cw10c20, cw10c26, cw10c38, ad1c52, and ad1c100. Unlike the TR- RNAi vector, the IR-RNAi vector appeared to target knockdown of HLA3 specifically, since the mrna abundance for all other analyzed limiting Ci inducible genes, LCIA, CAH1, CCP1/2 and LCI1, was not affected (Fig. S3B). VL-CO 2 acclimated cw10-ir-rnai and ad1-ir-rnai knockdown lines have only minor photosynthetic defects at neutral ph. No significant growth differences at ph 6.0 or 7.3 were observed with any of the cw10-ir-rnai and ad1-ir-rnai lines based on spot tests and liquid growth (Fig. S3A; liquid growth data not shown), even though HLA3 mrna abundance was almost undetectable (Fig. S3B). The apparent photosynthetic Ci affinity of the cw10-ir-rnai strains was unaffected at ph 6 and ph 7.3, except for a 25% inhibition at 20 µm Ci and ph 7.3, and that of the ad1-ir-rnai lines was unaffected at ph 6.0 but substantially inhibited at ph 7.3 (Table S2). These results are consistent with a high-ph specific inhibition of photosynthesis by knockdown of HLA3 expression in the IR-RNAi lines. Wall-less strains cw10 and ad1 were used to improve transformation efficiency, but, because of fragility of the wall-less cells during silicone oil centrifugation, Ci uptake experiments yielded highly variable data and were considered unreliable. Walled progeny of cw10-ir-rnai lines retain knockdown of HLA3. Northern analysis of selected paromomycin-resistant, walled progeny (CC620IR21.1, CC620IR21.2, CC620IR32.1 and CC620IR32.2) from crosses between cw10-ir-rnai lines, cw10ir21 and cw10ir32 with walled WT strain CC620, and of paromomycin-resistant, air-dier, walled progeny (pmp1ir21.3 and pmp1ir21.4) from crosses between cw10ir21 with pmp1
25 22 demonstrated that paromomycin resistance was co-inherited with knockdown of HLA3 mrna abundance (Figs. 3A, 3B). VL-CO 2 acclimated CC620-IR-RNAi walled progeny exhibit high ph specific growth deficiency. Spot tests showed no apparent growth inhibition of the CC620-IR- RNAi progeny in VL-CO 2 even at ph 9 (Fig. S4), but some growth inhibition of CC620IR21.1 and CC620IR32.1 was observed at ph 9.0 in liquid medium (Fig. 4). Consistent with this growth phenotype, both photosynthetic Ci affinity and internal Ci accumulation of VL-CO 2 acclimated CC620IR21.1 and CC620IR32.1 were substantially inhibited at ph 9.0, where HCO - 3 is the predominant Ci species (>99%), and even decreased slightly at ph 7.3 in 20 µm Ci (Tables 1 and 2). VL-CO 2 acclimated pmp1-ir-rnai walled progeny exhibit high ph specific growth deficiency. Spot tests showed no apparent growth inhibition of the pmp1-ir-rnai progeny pmp1ir21.3 and pmp1ir21.4 in VL-CO 2 at ph 7.3 but marked inhibition of growth at ph 9.0 (Fig. 5). Consistent with this high ph sensitive growth phenotype, both apparent photosynthetic Ci affinity and internal Ci accumulation of VL-CO 2 acclimated pmp1ir21.3 and pmp1ir21.4 were markedly inhibited at both ph 7.3 and ph 9.0 (Tables 1 and 2). Discussion Most reverse genetic approaches are technically challenging in C. reinhardtii, the exception being RNAi based knockdown of endogenous gene expression (24). In this work, two quite different RNAi constructs both were highly effective in minimizing HLA3 mrna accumulation. The IR-RNAi construct, in which the antibiotic resistance gene and HLA3 sequences were transcriptionally linked, yielded almost 10-times as many effective RNAi transformants as the TR-RNAi construct, in which the antibiotic resistance gene and HLA3 sequences were transcribed separately. Although the specific HLA3 sequences and their arrangement (TR vs IR) cannot be excluded, it is tempting to speculate that transcriptional linkage of HLA3 IR and antibiotic marker sequences increased knockdown efficiency by
26 23 minimizing transcriptional silencing. In any event, both constructs successfully decreased the HLA3 mrna abundance. As illustrated in Figure 6, there are two major inferred pathways for HCO - 3 accumulation in the chloroplast, via active HCO - 3 transport (across the plasma membrane and chloroplast envelope, possibly mediated by HLA3 and LCIA) and active CO 2 uptake (either via active transport across the plasma membrane and/or chloroplast envelope or diffusion across the same membranes in response to active depletion internally) along with rapid conversion of CO 2 to HCO - 3 in the alkaline stroma, possibly mediated by CAH6 and/or LCIB. CAH3- - catalyzed CO 2 release from accumulated HCO 3 raises the concentration of CO 2 around Rubisco, localized primarily in the pyrenoid, an internal compartment within the stroma of algae. Because the equilibrium between the weak acid CO 2 and its hydrated conjugate base, HCO - 3, is ph dependent (pka=6.37), changes in the ph and the total Ci concentration can be used to perturb the relative and absolute CO 2 and HCO - 3 concentrations, allowing insight into the Ci species utilized under given conditions. RNAi-mediated knockdown of HLA3 mrna abundance was used to explore the role of HLA3 in the C. reinhardtii CCM. Specific RNAi knockdown of HLA3 expression alone in wall-less cw10 and in walled CC620 resulted in a small decrease in growth and substantial decreases in both photosynthetic Ci affinity and Ci uptake specifically at alkaline ph, although small but significant decreases in photosynthesis and Ci uptake were observed even at neutral ph, but not at ph 6. Since the CO 2 -HCO equilibrium shifts towards HCO 3 at higher ph, the observed decrease in photosynthesis and internal Ci accumulation in the HLA3 knockdown strains only at the lower Ci concentration of 20 µm in ph 7.3 but at both 20 µm and 50 µm Ci in ph 9.0 argues strongly that the HLA3 knockdown causes a deficiency in HCO - 3 uptake. The equilibrium CO 2 concentration available for direct CO 2 uptake at the cell surface at ph 7.3 would be 5.9 µm at 50 µm Ci but only 2.4 µm at 20 µm Ci, whereas at ph 9.0 the equilibrium CO 2 concentrations would be only 0.12 µm CO 2 even at 50 µm Ci. Thus we hypothesize that the CO 2 concentration threshold for a significant - decrease in photosynthesis and Ci accumulation in the absence of HCO 3 transport lies below 5.9 µm CO 2 and that the decrease in photosynthesis and Ci accumulation below this
27 24 threshold CO 2 concentration represents a defect in the ability of the cells to utilize the more abundant HCO - 3 in the absence of substantial HLA3 expression. These data from HLA3 knockdown in cw10 and CC620 alone argue that HLA3 plays a role in the transport of HCO - 3, the predominant Ci form at alkaline ph. Convincing support for this conclusion is provided by data from the combination of HLA3 knockdown with either a mutation in LCIB, which encodes a plastid localized protein required in L-CO 2 for - normal Ci accumulation, either by trapping CO 2 released from accumulated stromal HCO 3 or by facilitating active uptake of CO 2 by chloroplasts (9, 15), or the off-target, simultaneous knockdown in CC125 of LCIA, which encodes a plastid envelope protein reported to transport HCO - 3 (14). In both cases, the combined defects resulted in dramatic decreases in growth, Ci uptake and photosynthetic Ci affinity, especially at alkaline ph. A schematic diagram of how these putative Ci uptake and accumulation components might participate in the CCM is illustrated in Figure 6. Loss of function mutations in LCIB result in loss of growth and almost complete loss of Ci accumulation in L-CO 2 acclimated cells, but have only minimal effects in cells acclimated to VL-CO 2 (9), demonstrating that an LCIB-independent Ci accumulation process masks the effect of LCIB loss in the VL-CO 2 state. Thus the inability of VL-CO 2 acclimated pmp1ir21.3 and pmp1ir21.4 to grow specifically at alkaline ph, together with the markedly decreased photosynthetic Ci affinity and Ci accumulation of the same cells at ph 9 provides compelling evidence that knockdown of HLA3 effectively removes this LCIB-independent Ci accumulation mechanism specifically at alkaline ph, where HCO - 3 is the dominant Ci species. This conclusion also is supported by decreases in photosynthesis and Ci uptake observed in pmp1ir21.3 and pmp1ir21.4 and in ad1-ir-rnai knockdowns at neutral ph, even though growth in spot tests was not significantly affected at this ph. The apparently highly specific co-suppression of both HLA3 mrna and LCIA mrna levels by the HLA3-TR-RNAi was unexpected, since few, if any, mismatches are tolerated in effective RNAi design (25). The HLA3 sequence used was screened against the C. reinhardtii genome, and its later comparison with LCIA mrna revealed the region of greatest homology contained no more than six perfectly matched sequential nucleotides in
28 25 any 23 nucleotide stretch within a sequence containing only about 70% matched nucleotides. However, with this TR-RNAi construct, severely decreased LCIA mrna abundance was always associated with HLA3 mrna knockdown, including the mrna level recovery for both genes as pmp1tr51 lost its phenotype, providing a compelling argument that, in this specific case, this homology level is sufficient to support "off target" destruction of LCIA mrna. The off-target, simultaneous knockdown of LCIA expression along with HLA3 by TR- RNAi in CC125 provided another uncovering of HLA3 function by removal of an apparent LCIA dependent Ci accumulation process. LCIA, a limiting-co 2 inducible NAR1 family member in C. reinhardtii, is a predicted chloroplast inner envelope protein (8) and is reported to transport HCO - 3 when expressed in Xenopus oocytes (12). CC125-TR-RNAi lines showed no growth inhibition at acidic and neutral ph, but cell growth, photosynthesis and Ci uptake all were dramatically decreased at alkaline ph. Since walled progeny of cw10-ir-rnai (CC620IR21.1 and CC620IR32.1) acclimated better than the CC125-TR- RNAi strains, it is clear that LCIA can partially compensate for the loss of HLA3 function in VL-CO 2 acclimated cells, at least at high ph. We have no direct evidence for the phenotype of an LCIA knockdown alone, but the high ph sensitive phenotype of the LCIA and HLA3 co-knockdown in CC125 argues strongly that LCIA and HLA3 are key synergistic or complementary components of a Ci transport pathway (Fig. 6). Combining the HLA3 knockdown with both the off-target LCIA knockdown and a LCIB mutation provided further insight and confirmation of the overlapping functions of HLA3, LCIA and LCIB. The inability of these strains to grow in VL-CO 2 at any tested ph, together with their markedly decreased photosynthetic Ci affinity and Ci uptake at neutral and alkaline ph, indicates that one or more of the three putative Ci uptake and accumulation systems also plays an important role in Ci accumulation at acidic ph in VL-CO 2. With functional expression of HLA3, LCIA and LCIB either compromised or defective in pmp1- TR-RNAi lines, their Ci uptake and photosynthetic affinity at neutral or alkaline ph decreased to the levels of the non-acclimating mutant cia5 (Tables 1 and 2) in which most limiting-co 2 inducible genes are not expressed (20). LCIB is located in the chloroplast
29 26 stroma (15) and is required for Ci accumulation specifically in L-CO 2 conditions (9), but this does not preclude LCIB from serving a redundant or overlapping function with other Ci accumulation processes in the VL-CO 2 acclimation state (15). LCIA also apparently is located in the plastid envelope, so the combination of an LCIB mutation and LCIA knockdown might explain the observed extreme phenotype in VL-CO 2 if the Ci uptake and accumulation systems associated with these two proteins together represent the bulk of Ci uptake and accumulation in the plastid. Although not yet demonstrated physically, HLA3 is predicted to be located in the plasma membrane (WoLF PSORT, 21), and the ph-dependence of the HLA3 knockdown strains phenotypes argues strongly for a plasma membrane localization. The cytosolic ph of C. reinhardtii is relatively stable to external ph changes over its optimal growth range of ph 5.5 and 8.5 (26, 27), so the observed high-ph dependent decrease in Ci affinity of HLA3 knockdown strains indirectly argues for the plasma membrane as the probable location of HLA3. The differential responses of the CC125 and pmp1 strains to TR-RNAi mediated cosilencing of HLA3 and LCIA, together with the clear differential responses of the CC620 and pmp1 strains to IR-RNAi induced knockdown of HLA3 alone, support the hypothesis that multiple limiting-co 2 induced and/or activated Ci uptake and accumulation systems function in C. reinhardtii. HLA3-dependent, LCIA-dependent and LCIB-dependent Ci uptake and/or accumulation systems appear to act at least partially in overlapping, complementary functions. It is reasonable to expect multiple Ci transport systems in C. reinhardtii, since at least four distinct Ci uptake systems are known in cyanobacteria, which have a functionally similar CCM (22). In cyanobacteria, mutation of only one Ci uptake system has little phenotypic effect, because of compensation by overlapping Ci uptake systems. So a similar phenomenon might explain the minimal phenotypic effect of HLA3 knockdown alone, as well as increased phenotypic impact of HLA3 and LCIA cosuppression or of HLA3 and LCIA co-suppression in an LCIB mutant background. In summary, knockdown of HLA3 (and LCIA) expression reported here demonstrates that the HLA3 gene product is required to provide sufficient Ci uptake and/or accumulation to
30 27 sustain an active CCM for optimal photosynthesis at high ph in limiting CO 2 environments. It is also demonstrated that the function of HLA3 is masked somewhat by other Ci uptake systems, including LCIA, a putative chloroplast HCO - 3 transporter, and LCIB, a chloroplast protein required for effective Ci accumulation in the L-CO 2 acclimation state. To our knowledge, this is the first clear link of a probable plasma membrane protein to Ci uptake function in a eukaryotic microalga. It is still not clear whether HLA3 is directly involved in HCO transport or indirectly facilitates the activity of other HCO 3 transporters, but the data - presented here strongly argue for the involvement of HLA3 in HCO 3 transport, either directly or indirectly, at the plasma membrane of C. reinhardtii. Materials and Methods C. reinhardtii strains and growth conditions. C. reinhardtii strains CC849 (cw10 mt-) and wild type CC125 were obtained from the Chlamydomonas Stock Center, Duke University, Durham, NC. The allelic LCIB-defective mutants pmp1 and ad1 have been described previously, as have media and growth conditions for C. reinhardtii strains (9, 13). RNAi vector construction, transformation and selection. For generating the AphVIII- HLA3 inverted repeat (IR) transgene, a 540-bp fragment corresponding to the 3 UTR sequence of HLA3 was amplified with primers HLA3-XbaI-S [5 - aattctagacagtgacccaaagc-3 ] and HLA3-HindIII-As [5 - tttaagctttttgtcactcgcgct-3 ]; HLA3-BamHI-S [5 - aaaggatcccaaagcaggtgagaa-3 ] and HLA3-HindIII-As [5 -gccaagcttcttttaatt CTGCAACC-3 ]. The two PCR products were ligated into XbaI/BamHI sites of plasmid psi103 (32). This HLA3 IR cassette was then excised with XbaI/KpnI and cloned into the XbaI/KpnI sites of plasmid psp124s. The AphVIII gene was further amplified with primers SacI-AphVIII [5 -ggggagctcgctgaggcttg-3 ] and XbaI-AphVIII [5 - cctctagatcagaagaactcgtccaacagc-3 ] and inserted into the XbaI/SacI sites of psp124s-hla3 IR intermediate plasmid. The final RNAi vector, in which the HLA3 inverted repeat is transcribed as part of the AphVIII transcript, was named AphVIII-HLA3 IR.
31 28 For generating tandem repeat construct, a 120-bp fragment from HLA3 exon6 was amplified by PCR with primers N-1 [5 -caaccatgg TCCATCTCCCTCTTCAACTC-3 ] and E-1 [5 - caagaattcggacgatttgcttggcgacg-3 ]; E-2 [5 - caagaattccatctccctcttcaactccg -3 ] and P-2 [5 - caactgcagggacgatttgcttggcgacg-3 ]. The two PCR products were inserted into NcoI/PstI sites of plasmid p300 (35). The intermediate plasmid was PCR amplified with primers P-3 [5 -caactgcagtccatctccctcttcaactc-3 ] and X-3[5 - caactcgagggacgatttgcttggcgacg-3 ]. The PCR product was ligated into PstI/XhoI site of intermediate plasmid and thus named 4 HLA3 plasmid. The 4 HLA3 construct was then excised with XbaI/BamHI and inserted into XbaI/BamHI sites of vector pgend-ble (33). An AphVIII cassette was excised with HindIII/KpnI from the plasmid psi103 and inserted into HindIII/KpnI sites of the p53 vector. The chimeric plasmid was double digested with HindIII/NdeI to eliminate the Ble gene and re-ligated. The resulting RNAi plasmid (containing 4 HLA3 cdna tandem repeat and AphVIII gene) was named AphVIII-HLA3 TR. For transformation, cells were collected and resuspended in autolysin for approximately 1h (autolysin treatment only for walled cells CC125 and pmp1) to remove the cell walls and glass bead transformations were performed as previously described (16). Paromomycin resistant transformants were selected as RNAi lines. Spot tests, Photosynthetic O 2 exchange and Ci uptake measurements. Cells grown to logarithmic phase were serially diluted in minimal medium, spotted onto agar plates with specified ph and kept at H-CO 2, L-CO 2 and VL-CO 2 levels for 9 days. Plates were buffered with 25 mm MES-KOH for ph 6, 25 mm MOPS-KOH for ph 7.3, and 25 mm AMPSO- KOH for ph 9. Photosynthetic O 2 evolution was measured as described previously (11). The Ci uptake by C. reinhardtii cells at ph 7.3 or 9.0 was estimated by the silicone oil filtration technique (30). ACKNOWLEDGMENTS. This work was supported by the ISU College of Agriculture and Life Sciences and the ISU College of Liberal Arts and Sciences, as well as by a grant from the USDA to M.H.S (grant no ).
32 29 References 1. Giordano M, Beardall J, Raven JA (2005) CO 2 concentrating mechanisms in algae: mechanisms, environmental modulation, and evolution. Annu Rev Plant Biol 56: Spalding MH (2008) Microalgal carbon-dioxide-concentrating mechanisms: Chlamydomonas inorganic carbon transporters. J Exp Bot 59(7): Palmqvist K, Sjoberg S, Samuelsson G (1988) Induction of inorganic carbon accumulation in the unicellular green algae Scenedesmus obliquus and Chlamydomonas reinhardtii. Plant Physiol 87: Sültemeyer DF, Klöck G, Kreuzberg K, Fock HP (1988) Photosynthesis and apparent affinity for dissolved inorganic carbon by cells and chloroplasts of Chlamydomonas reinhardtii grown at high and low CO 2 concentrations. Planta 176: Funke RP, Kovar JL, Weeks DP (1997) Intracellular carbonic anhydrase is essential to photosynthesis in Chlamydomonas reinhardtii at atmospheric levels of CO 2. Plant Physiol 114: Karlsson J, et al. (1998) A novel α-type carbonic anhydrase associated with the thylakoid membrane in Chlamydomonas reinhardtii is required for growth at ambient CO 2. EMBO J 17: Rexach J, Fernández E, Galván A (2000) The Chlamydomonas reinhardtii Nar1 gene encodes a chloroplast membrane protein involved in nitrite transport. Plant Cell 12: Miura K, et al. (2004) Expression profiling-based identification of CO 2 -responsive genes regulated by CCM1 controlling a carbon-concentrating mechanism in Chlamydomonas reinhardtii. Plant Physiol 135: Wang Y, Spalding MH (2006) An inorganic carbon transport system responsible for acclimation specific to air levels of CO 2 in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 103: Im CS, Grossman AR (2001) Identification and regulation of high light-induced genes in Chlamydomonas reinhardtii. Plant J 30: Hanikenne M, Krämer U, Demoulin V, Baurain D (2005) A comparative inventory of metal transporters in the green alga Chlamydomonas reinhardtii and the red alga Cyanidioschizon merolae. Plant Physiol 137: Mariscal V, et al. (2006) Differential regulation of the Chlamydomonas Nar1 gene family by carbon and nitrogen. Protist 157: Spalding MH, Spreitzer RJ, Ogren WL (1983) Reduced inorganic carbon transport in a CO 2 -requiring mutant of Chlamydomonas reinhardtii. Plant Physiol 73: Van K, Wang Y, Nakamura Y, Spalding MH (2001) Insertional mutants of Chlamydomonas reinhardtii that require elevated CO 2 for survival. Plant Physiol 127: Duanmu D, Wang Y, Spalding MH (2009) Thylakoid lumen carbonic anhydrase (CAH3) mutation suppresses air-dier phenotype of LCIB mutant in Chlamydomonas reinhardtii. Plant Physiol 149:
33 Vance P, Spalding MH (2005) Growth, photosynthesis and gene expression in Chlamydomonas over a range of CO 2 concentrations and CO 2 /O 2 ratios: CO 2 regulates multiple acclimation states. Can J Bot 83: Kruh GD, Belinsky MG (2003) The MRP family of drug efflux pumps. Oncogene 22: Klein M, Burla B, Martinoia E (2006) The multidrug resistance-associated protein (MRP/ABCC) subfamily of ATP-binding cassette transporters in plants. FEBS Lett 580: Fukuzawa H, et al. (2001) Ccm1, a regulatory gene controlling the induction of a carbon-concentrating mechanism in Chlamydomonas reinhardtii by sensing CO 2 availability. Proc Natl Acad Sci USA 98: Xiang Y, Zhang J, Weeks DP (2001) The cia5 gene controls formation of the carbon concentrating mechanism in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 98: Paul Horton, et al. (2007) WoLF PSORT: protein localization predictor. Nucleic Acids Res doi: /nar/gkm Price GD, Maeda SI, Omata T, Badger MR (2002) Modes of active inorganic carbon uptake in the cyanobacterium, Synechococcus sp. PCC7942. Funct Plant Biol 29: Ma C, Mitra A (2002) Intrinsic direct repeats generate consistent post-transcriptional gene silencing in tobacco. Plant J 31: Schroda M (2006) RNA silencing in Chlamydomonas: mechanisms and tools. Curr Genet 49: Ma Y, Creanga A, Lum L, Beachy PA (2006) Prevalence of off-target effects in Drosophila RNA interference screens. Nature 443: Lustigman B, Lee LH, Magasic CW (1995) Effects of cobalt and ph on the growth of Chlamydomonas reinhardtii. Bull Environ Contam Toxicol 55: Messerli MA, et al. (2005) Life at acidic ph imposes an increased energetic cost for a eukaryotic acidophile. J Exp Biol 208: Sizova I, Fuhrmann M, Hegemann P (2001) A Streptomyces rimosus aphviii gene encoding for a new type phosphotransferase provides stable antibiotic resistance to Chlamydomonas reinhardtii. Gene 277: Fischer N, Rochaix JD (2001) The flanking regions of PsaD drive efficient gene expression in the nucleus of the green alga Chlamydomonas reinhardtii. Mol Genet Genomics 265: Badger MR, Kaplan A, Berry JA (1980) Internal inorganic carbon pool of Chlamydomonas reinhardtii: evidence for a carbon dioxide concentrating mechanism. Plant Physiol 66:
34 31 Figure Legends. Figure 1. Northern blot analysis of TR-RNAi strains derived from (A) pmp1 and (B) wildtype CC125. Cells were acclimated to VL-CO 2 for 4h and 16h. Total RNA was separated and hybridized to HLA3 and LCIA coding sequences. Figure 2. Effect of HLA3 TR-RNAi transgene on the growth of (A) CC125 RNAi transformants and (B) pmp1 RNAi transformants on minimal plates in H-CO 2 or VL-CO 2. Cells grown to logarithmic phase were diluted to the indicated cell numbers per 5 µl, spotted on plates and incubated for 9 days under dim light. Figure 3. Northern blot analysis of (A) wild type CC620 and CC620 cw10-rnai-progeny and (B) pmp1 and pmp1 cw10-ir21 progeny, all acclimated to VL-CO 2 for 24 hours. Figure 4. Liquid growth curves for wild type CC620 ( ) and CC620 cw10-rnai-progeny IR21.1 ( ) and IR32.1( ) in ph 7.3 or ph 9.0 minimal medium and VL-CO 2. Growth curves shown are averages of three independent measurements, and the grey bars indicate maximum standard errors for each time point. Figure 5. Spot test growth of walled pmp1 cw10-ir21 progeny (pmp1ir21.3 and pmp1ir21.4) on neutral and alkaline minimal plates in H-CO 2 or VL-CO 2. Figure 6. Schematic diagram illustrating the suggested roles of HLA3, LCIA and LCIB, as well as of the carbonic anhydrases CAH1, CAH3 and CAH6, in the C. reinhardtii CCM. Other potential Ci transport and accumulation components indicated by question marks only include: pale green circles - potential active CO 2 transport or passive CO 2 diffusion sites; purple ovals - potential additional HCO 3 - transporters; purple circle - predicted HCO 3 - facilitated diffusion across the thylakoid membrane. Predicted sites of disruption by mutations ad1, cia5, and pmp1, and of IR-RNAi and TR-RNAi mediated gene expression knockdowns are indicated in red print. Figure S1. Schematic diagram of the RNAi constructs (A) AphVIII-HLA3 TR, in which a 120-bp fragment of the HLA3 coding sequence (exon 6) was cloned into the vector as four tandem repeats under control of the RBCS2 promoter and terminator, and (B) AphVIII- HLA3 IR, in which the 540-bp HLA3 3 UTR was cloned in the forward and reverse
35 32 orientations behind the AphVIII selectable marker to generate an AphVIII/HLA3-IR cistronic transcription unit under control of the HSP70/RBCS2 promoter and terminator. Restriction sites: B, BamHI; E, EcoRI; H, HindIII; K, KpnI; N, NcoI; P, PstI; S, SacI; Xb, XbaI; Xh, XhoI. Figure S2. Effect of HLA3 TR RNAi transgene expression on the growth of CC125-TR- RNAi and pmp1-tr-rnai transformants on minimal plates at ph 6 in H-CO 2 or VL-CO 2 chambers. Cells grown to logarithmic phase were diluted to the indicated cell numbers per 5 µl, spotted on plates and incubated for 9 days under dim lights. Figure S3. Effects of HLA3 IR transgene on growth and HLA3 expression in wall-less cw10- IR-RNAi and ad1-ir-rnai strains. (A) Growth of cw10 and ad1 RNAi transformants on ph 7.3 minimal plates in H-CO 2 or VL-CO 2 chambers. Cells grown to logarithmic phase were diluted to the indicated numbers per 5µl, spotted on plates and incubated for 9 days under dim lights. (B) Northern blot analysis showing the effects of the HLA3 IR-RNAi transgene on the expression of HLA3 and other limiting-ci-regulated genes (LCIA, CAH1, CCP1/2, and LCI1) in wall-less strains cw10 and ad1 using the coding sequences of these genes as probes. Cells were acclimated to VL-CO 2 conditions for 24 hours. Figure S4. Growth of walled progeny of CC620 cw10-ir-rnai on neutral and alkaline minimal plates in H-CO 2 or VL-CO 2 chambers.
36 33 Table 1. Photosynthetic Ci affinity of wild type (CC125 and CC620), pmp1, cia5, and TR- RNAi or IR-RNAi strains. ph 7.3 ph 9.0 Strains P5 P20 P50 P5 P20 P50 No RNAi CC ± ± ± ±0.02 CC ± ± ± ±0.03 pmp1 0.24± ± ± ± ± ±0.09 cia5 0.04± ± ± ±0.01 TR-RNAi CC125TR ± ± ± ±0.09 CC125TR ± ± ± ±0.08 CC125C ± ± ± ±0.03 pmp1tr ± ± ± ±0.04 pmp1tr ± ± ± ±0.02 pmp1c ± ± ± ±0.08 IR-RNAi CC620IR ± ± ± ±0.04 CC620IR ± ± ± ±0.03 pmp1ir ± ± ± ±0.05 pmp1ir ± ± ± ±0.05 Cells acclimated 1d to VL-CO 2 and O 2 evolution rates determined in ph 7.3 and 9.0 at 5 µm (v5), 20 µm (v20), 50 µm (v50), and 4000 µm (v4000) NaHCO 3. Relative Ci affinity calculated as ratios: P5=v5/v4000; P20=v20/v4000; P50=v50/v4000. Chl was 25 µg/ml.
37 34 Table 2. Internal Ci concentration (mm) of wild type (CC125 and CC620), pmp1, cia5, and TR-RNAi or IR-RNAi strains. ph 7.3 ph 9.0 Strains 20 µm 50 µm 20 µm 50 µm No RNAi CC B B0.33 CC B B B B0.19 pmp1 1.16B B B B0.34 cia5 0.30B B0.08 TR-RNAi CC125TR B B0.18 CC125TR B B0.21 CC125C B B0.28 pmp1tr B B0.04 pmp1tr B B0.10 pmp1c B B0.21 IR-RNAi CC620IR B B0.34 CC620IR B B B B B B0.18 pmp1ir B B B B0.11 pmp1ir B B B B0.11 Cells were acclimated to VL-CO 2 for 1 day and internal Ci accumulation (mm Ci after 80 s) determined in ph 7.3 and ph 9.0 at initial Ci Concentrations of 20 µm and 50 µm [ 14 C]-NaHCO 3. Chl was 25 µg/ml.
38 35 Supplemental Table 1. Relative photosynthetic Ci affinity in wildtype CC125, pmp1, cia5 and derivative RNAi strains at ph 6.0. ph 6.0 Strain P5 P50 V4000 CC B B ± 12 CC125TR B B ± 8 CC125TR B B ± 7 CC125C B B ± 12 pmp1 0.20B B ± 6 pmp1tr B B ± 19 pmp1tr B B ± 15 pmp1c B B ± 11 cia5 0.09B B ± 8 Cells were acclimated to VL-CO 2 for 1 day and rates of O 2 evolution (µmol mg Chl -1 h -1 ) determined in ph 6.0 (MES-KOH) buffer at 5 µm (v5), 50 µm (v50), and 4000 µm (v4000) NaHCO 3. Relative Ci affinity was calculated as ratios: P5=v5/v4000 or P50=v50/v4000.
39 36 Supplemental Table 2. Effect of ph on the photosynthetic Ci affinities of wall-less wild type (cw10), ad1 and RNAi strains. ph 6.0 ph 7.3 Strain P20 P50 V4000 P20 P50 V4000 cw ± ± ± ± ± ±7 cw10ir ± ± ± ± ± ±12 cw10ir ± ± ± ± ± ±10 cw10c ± ± ± ± ± ±14 ad1 0.23± ± ± ± ± ±8 ad1ir ± ± ± ± ± ±9 ad1ir ± ± ± ± ± ±9 ad1ir ± ± ± ± ± ±7 ad1c ± ± ± ± ± ±12 Cells were acclimated to VL-CO 2 for 1 day and rates of O 2 evolution (µmol mg Chl -1 h -1 ) determined in ph 6.0 (MES-KOH) and ph 7.3 (MOPS-KOH) buffers at 20 µm (v20), 50 µm (v50), and 4000 µm (v4000) NaHCO 3. Relative Ci affinity was calculated as ratios: P20=v20/v4000 or P50=v50/v4000. Chl was 25 µg/ml.

40 Figure 1. 37


41 Figure 2. 38

42 Figure 3. 39
43 Figure 4. 40

44 Figure 5. 41
45 Figure 6. 42


46 Figure S1. 43

47 Figure S2. 44



48 Figure S3. 45

49 Figure S4. 46
50 47 CHAPTER 3. THYLAKOID LUMEN CARBONIC ANHYASE (CAH3) MUTATION SUPPRESSES AIR-DIER PHENOTYPE OF lcib MUTANT IN CHLAMYDOMONAS REINHARDTII A paper published in Plant Physiology 149: (2009) Deqiang Duanmu 1, Yingjun Wang 1, and Martin H. Spalding 1,2 Abstract An active CO 2 concentrating mechanism (CCM) is induced when Chlamydomonas reinhardtii acclimates to limiting inorganic carbon (Ci), either low-co 2 (air level; ~0.04% CO 2 ) or very low-co 2 (~0.01% CO 2 ) conditions. A mutant, ad1, which is defective in the limiting-co 2 -inducible, plastid localized LCIB, can grow in high-co 2 or very low-co 2 conditions but dies in low-co 2, indicating a deficiency in a low-co 2 -specific Ci uptake and accumulation system. In this study, we identified two ad1 suppressors that can grow in low- CO 2 but die in very low-co 2. Molecular analyses revealed that both suppressors have mutations in the CAH3 gene, which encodes a thylakoid lumen localized carbonic anhydrase. Photosynthetic rates of low-co 2 acclimated suppressors under acclimation CO 2 concentrations were more than two fold higher than ad1, apparently resulting from a more than 20 fold increase in the intracellular concentration of Ci as measured by direct Ci uptake. However, photosynthetic rates of very low-co 2 acclimated cells under acclimation CO 2 concentrations were too low to support growth in spite of a significantly elevated intracellular Ci concentration. We conclude that LCIB functions downstream of CAH3 in 1 Department of Genetics, Development and Cell Biology, Iowa State University, Ames, IA Author for correspondence Author contributions: D.D. and M.H.S. designed research; D.D. and Y.W. performed research; D.D. and M.H.S. analyzed data; and D.D. and M.H.S. wrote the paper. The data from Y.W. was removed from this thesis.
51 48 the CCM and probably plays a role in trapping CO 2 released by CAH3 dehydration of accumulated Ci. Apparently dehydration by the chloroplast stromal carbonic anhydrase CAH6 of the very high internal Ci caused by the defect in CAH3 provides Rubisco sufficient CO 2 to support growth in low-co 2 acclimated cells, but not in very low-co 2 acclimated cells, even in the absence of LCIB. Introduction Carbon dioxide (CO 2 ) serves both as the substrate for photosynthesis and as an important signal to regulate plant growth and development, so variable CO 2 concentrations can impact photosynthesis, growth and productivity of plants. Terrestrial C4 plants have developed a CO 2 -concentrating mechanism (CCM) involving anatomical and biochemical adaptations to accumulate a higher concentration of CO 2 as substrate for Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) and to suppress oxygenation of RuBP, a wasteful side reaction. In contrast, a different type of CCM is induced in the unicellular green microalga Chlamydomonas reinhardtii when the supply of dissolved inorganic carbon (Ci; CO 2 and HCO - 3 ) for photosynthesis is limited (Beardall and Giordano, 2002; Giordano et al., 2005; Spalding, 2008; Moroney and Ynalvez, 2007). In response to limiting CO 2, representative biophysical C i pumps operate: the CCM uses active Ci transport, both at the plasma - membrane and the chloroplast envelope, to accumulate a high concentration of HCO 3 within the chloroplast (Sültemeyer et al., 1988; Palmqvist et al., 1988). The thylakoid lumen carbonic anhydrase (CAH3) plays an essential role in the rapid dehydration of the - accumulated HCO 3 to release CO 2 into the pyrenoid, a Rubisco-containing internal compartment of the chloroplast, for assimilation by Rubisco (Price et al., 2002; Spalding et al., 2002). While a number of genes and proteins essential to the operation of the CCM in C. reinhardtii have been identified, our understanding of Ci uptake and its regulation, as well as other aspects of CCM function is limited. A better understanding of the similar CCM in prokaryotic organisms, specifically the cyanobacteria Synechocystis and Synechococcus, has been gained. At least five different types of Ci transporters have been identified in cyanobacteria, including three HCO - 3 transporters and two active CO 2 uptake systems (Price
52 49 et al., 2002; 2004). Recently, at least three distinct CO 2 -regulated acclimation states were identified in C. reinhardtii based on growth, photosynthesis and gene expression characteristics, a high CO 2 (H-CO 2 ) state (5% - 0.5% CO 2 ), low CO 2 (L-CO 2 ) state (air level; 0.4% % CO 2 ) and very low CO 2 (VL-CO 2 ) state (0.01% % CO 2 ) (Vance and Spalding, 2005). Two allelic HCR (high-co 2 -requiring) mutants, pmp1 and ad1, grow as well (pmp1) or nearly as well (ad1) as wild type cells in both H-CO 2 and VL-CO 2 conditions while only dying in L- CO 2, indicating a deficient Ci transport and/or accumulation system only in the L-CO 2 acclimation state (Spalding et al., 1983b; 2002). The defective gene responsible for the pmp1/ad1 phenotype was identified as LCIB, a limiting CO 2 -inducible gene, the product of which is predicted to be located in the chloroplast stroma and proposed to be involved with chloroplast Ci uptake in L-CO 2 conditions (Wang and Spalding, 2006). The LCIB gene product is a member of a small gene family so far only found in a few microalgae species (Spalding, 2008). To investigate the roles of LCIB in eukaryotic photosynthetic organisms and identify other functional components involved in chloroplast Ci accumulation in C. reinhardtii, we used an insertional mutagenesis approach to select suppressors of the air-dier phenotype of the LCIB mutant ad1. In the current study, we describe two ad1 suppressors, ad-su6 and adsu7, that grow normally in L-CO 2 but, unlike ad1, die in VL-CO 2. This report also presents data suggesting that the air-dier phenotype of ad1 is suppressed by increased intracellular Ci concentrations in the two suppressors, and suggesting a possible role for LCIB as a CO 2 trap rather than having any direct role in chloroplast envelope Ci transport. Results Identification and Genetic Analysis of ad1 Suppressors The C. reinhardtii strain chosen for this study was ad1, containing a deletion mutation of LCIB (Wang and Spalding, 2006). This strain can not grow in L-CO 2 but can grow either in H-CO 2 or in VL-CO 2. To isolate and identify suppressors which can grow in L-CO 2, we performed insertional mutagenesis using a Par R -containing plasmid (psi103) to transform ad1 (Fig. 2A). From ~10 6 transformants, three displayed the suppression phenotype, and two
53 50 of these, ad-su6 and ad-su7, are described here. These two suppressors exhibited suboptimal growth in L-CO 2 where the parental strain ad1 could not grow at all (Fig. 1). Unexpectedly, neither suppressor could survive in VL-CO 2, indicating that although the second site suppressor mutations could suppress the L-CO 2 lethal, air-dier phenotype of the LCIB mutation in ad1, growth of the two suppressors in VL-CO 2 was completely abolished. The ad-su6 and ad-su7 strains were crossed with wild-type strain CC620 to determine whether the suppression phenotype cosegregated with the inserted Par R gene. More than 150 Zeo R (zeocin-resistance, conferred by the Ble R insert responsible for the LCIB mutation) random progeny from each cross were screened for their growth in different levels of CO 2 and their resistance to paromomycin. In the ad-su6 cross, all 90 random progeny with the suppressor phenotype were paromomycin resistant, while all paromomycin-sensitive progeny exhibited an ad1-like growth phenotype in L-CO 2 and VL-CO 2, indicating cosegregation of the suppressor phenotype with the Par R insert. Southern analysis with probe specific for the Par R gene indicated a single insert present in ad-su6 (Fig. 2B). Although ad-su7 also contains only one Par R insert, genetic analysis showed the suppressor phenotype was not linked to the insert (data not shown). Inverse PCR was employed to identify the flanking DNA in ad-su6. This flanking sequence was used in a BLAST search against the C. reinhardtii genome ( and the insertion site was shown to be located between exon 5 and intron 5 of CAH3 (Fig. 2C). Further PCR and DNA gel blot analyses revealed that only 186 nucleotides of CAH3 (63 nt of exon 5 and 123 nt of intron 5) were deleted in ad-su6. Since ad-su6 has the same growth phenotype as ad-su7, we PCR-amplified and sequenced CAH3 genomic DNA from ad-su7 and found that two nucleotides were deleted downstream of the ATG translation initiation codon (ATG CGC TCA GCC GTT CTA CAA CGC GGC CAG GCG CGG CGA GTG TCT TGC CGG GTG AGT GAA, underline indicates deletion mutation in ad-su7), which predicts a premature stop codon (ATG CGC TCA GCC GCT ACA ACG CGG CCA GGC GCG GCG AGT GTC TTG CCG GGT GAG TGA, underline indicates stop codon).
54 51 Expression Patterns of Limiting-CO 2 -inducible Genes in ad-su6 Northern-blot analysis showed that ad-su6 apparently is a hypomorphic mutant for CAH3. In ad1 cells, a transient, slight increase in the CAH3 transcript abundance was observed when cells were shifted from a H-CO 2 to either L-CO 2 or VL-CO 2 atmosphere, and, after a longer acclimation time (14 hours), the CAH3 message abundance was decreased (Fig. 3A). In the CAH3 hypomorphic mutant ad-su6, CAH3 transcript was undetectable throughout all CO 2 conditions. Western-blot analysis using polyclonal antiserum raised against CAH3 from C. reinhardtii also failed to detect the CAH3 protein in the suppressor mutant ad-su6 cultures (Fig. 3B). Message accumulations of several limiting CO 2 inducible genes also were analyzed. Among all the genes tested, including LCIC, CCP, LCI1, and LCIA, patterns of expression in the suppressor ad-su6 relative to the original LCIB mutant ad1 were not found to be significantly different. The expression level of chloroplast stromal carbonic anhydrase (CAH6) was not affected by CO 2 conditions, and the transcript abundance of CAH6 in adsu6 under limiting-co 2 conditions was comparable to that in ad1 (Fig. 3A). Complementation of ad-su6 To confirm whether the CAH3 mutation is responsible for the suppressor phenotype, we transformed a genomic DNA fragment containing a wild-type copy of CAH3 into the suppressors ad-su6 and ad-su7 and selected complemented lines that could survive in VL- CO 2. Complemented ad-su6 (su6-c1 and su6-c3) and ad-su7 lines showed the same growth phenotype as ad1, growth in VL-CO 2 and no growth in L-CO 2 (Fig. 1, ad-su7 data not shown). In addition, western blot analysis showed that complemented ad-su6 lines recovered the expression of CAH3 protein (Fig. 3B). Complementation of the ad-su6 VL-CO 2 lethal phenotype also was achieved by expressing CAH3 cdna under control of the constitutive PsaD promoter and terminator (Fischer and Rochaix, 2001; data not shown). Photosynthetic Affinity for Ci and Direct Ci-uptake Characteristics Photosynthetic O 2 evolution in response to Ci concentrations for L-CO 2 and VL-CO 2 acclimated wild-type and various mutant cells was compared (Table I). Walled progeny of
55 52 mutants were generated for physiological measurements, including the CAH3 mutation of ad-su6 in a WT background (wt-su6) or in an ad1 background (ad-su6-1). Consistent with the ad1 air-dier phenotype, the LCIB-defective ad1-1 mutant cells (walled progeny of ad1) acclimated in L-CO 2 showed dramatically decreased photosynthetic affinity for Ci compared with wild-type cells acclimated under the same conditions (~11% of WT). In contrast, when acclimated to VL-CO 2, photosynthetic affinity of ad1-1 was increased to 57~61% of WT strain (P20 and P50, estimated at 20 µm and 50 µm total Ci, respectively). Photosynthetic affinity of L-CO 2 acclimated ad-su6-1 was significantly higher than ad1-1 at 50 µm total Ci (P50: 0.14±0.02 versus 0.07±0.02, p-value <0.05), while VL-CO 2 acclimated ad-su6-1 had a lower relative affinity than ad1-1 (P20: 0.07±0.03 versus 0.27±0.03, p <0.05) at 20 µm total Ci, both of which are consistent with the phenotypes of these two strains. The aberrant photosynthetic affinity of ad-su6-1 was caused by the CAH3 mutation, since wt-su6 showed the same pattern of photosynthetic affinity in both L-CO 2 and VL-CO 2 conditions as ad-su6-1. Intracellular Ci accumulation in L-CO 2 acclimated ad1-1 cells was similar to that of the non-acclimating mutant cia5 (0.26±0.08 mm versus 0.19±0.05 mm), in which the CCM presumably does not function (Table II). An active Ci accumulation mechanism was regained in VL-CO 2 acclimated ad1-1 cells, although not as high as the WT strain (0.80±0.15 mm versus 1.65±0.25 mm, measured with 20 µm total Ci). The measured intracellular Ci pool in the LCIB-CAH3 double mutant ad-su6-1 was increased more than 20 fold over that of the LCIB single mutant ad1-1 when acclimated in L-CO 2 conditions (6.15±1.15 mm versus 0.26±0.08 mm), and the increased Ci accumulation could be attributed to the CAH3 mutation, because CAH3 single mutant wt-su6 accumulated the same level of intracellular [Ci] as ad-su6-1 (7.60±1.25 mm versus 6.15±1.15 mm). In VL-CO 2 acclimated strains, intracellular [Ci] in ad-su6-1 was only 2.5 fold higher than that in ad1-1 (2.85±0.32 mm versus 0.80±0.15 mm, measured with 20 µm total Ci). Discussion Rubisco, the primary enzyme for photosynthetic CO 2 assimilation, is an inefficient catalyst with a low affinity for atmospheric CO 2. For most algae, the Rubisco K m (CO 2 ) is
56 53 greater than 25 µm, so Rubisco is functioning at < 20% of capacity at 10 µm CO 2 in an airequilibrated aquatic environment. Many photosynthetic organisms have an inducible CCM that raises the CO 2 concentration around the active site of Rubisco several fold higher than the environmental level, thus improving the efficiency of CO 2 assimilation. In the eukaryotic microalga C. reinhardtii, active Ci uptake (mainly as CO 2 and HCO - 3 ) at either the plasma membrane or the inner chloroplast envelope is an essential component of the CCM. Although the products of several limiting CO 2 -inducible genes have been identified as putative Ci transporters on the chloroplast envelope, including LCIA, LCI1, CCP1/2, and Ycf10, or on the plasma membrane, including HLA3, none of the respective gene products have been definitively determined to transport Ci species (Burow et al., 1996; Chen et al., 1997; Rolland et al., 1997; Im and Grossman, 2001; Pollock et al., 2004; Miura et al., 2004; Mariscal et al., 2006), leaving Ci transport largely a mystery. Mutants with lesions in the limiting-co 2 -induced gene LCIB shed some light on the nature of Ci uptake and accumulation in chloroplasts. Two conditional lethal mutants, pmp1 and ad1, with lesions in LCIB, are defective in Ci accumulation when acclimated in L-CO 2 but not when acclimated in VL-CO 2, indicating the existence of multiple Ci uptake and accumulation pathways in C. reinhardtii corresponding to multiple limiting-co 2 acclimation states (Vance and Spalding, 2005). LCIB was proposed to be involved in active Ci transport (Spalding et al., 1983b; Wang and Spalding, 2006), although, since it is predicted to be a soluble protein with no significant transmembrane regions, it was acknowledged to be unlikely that LCIB itself is directly involved in transport. Therefore, it was argued that LCIB might be a subunit of a Ci transport complex or might have a regulatory role in Ci uptake and/or accumulation. In this paper, we identified two independent, second-site suppressors of the LCIB mutant air-dier phenotype that restored growth to ad1 in L-CO 2 but, unlike ad1 itself, were unable to grow in VL-CO 2. Both suppressors have lesions in CAH3, encoding a thylakoid lumen located, α-type carbonic anhydrase. The requirement of a thylakoidal CA and an acidic compartment in a functional CCM was first suggested by Pronina and colleagues and later CAH3 was identified and proposed to catalyze the rapid conversion of HCO - 3 to CO 2 in the acidic thylakoid lumen, with the CO 2 then diffusing to the pyrenoid to supply elevated
57 54 substrate CO 2 concentrations for Rubisco (Pronina et al., 1981, 1990; Funke et al., 1997; Raven 1997; Karlsson et al., 1998; Hanson et al., 2003; Moroney and Ynalvez, 2007). Consistent with previous reports (Spalding et al., 1983a; Moroney et al., 1987), the L-CO 2 acclimated CAH3 single mutant wt-su6, generated in a cross between the suppressed line and WT, accumulated a very high internal Ci concentration but was unable to use Ci efficiently for photosynthesis. However, the photosynthetic Ci affinity of this CAH3 single gene mutant wt-su6 still is significantly higher than that of the L-CO 2 acclimated LCIB mutant ad1, which accumulates only very low internal Ci levels and cannot grow at all in L- CO 2, both traits agreeing with prior reports of the characteristics of the allelic LCIB mutant pmp1 and various CAH3 single mutants (Spalding et al., 1983a, b; Moroney et al., 1987; Suzuki and Spalding, 1989). The LCIB-CAH3 double mutant suppressor ad-su6-1 accumulated the same level of intracellular Ci as the CAH3 single-gene mutant wt-su6, strongly arguing against LCIB as being required for or solely responsible for Ci transport into the chloroplast. This effect of combining lesions in CAH3 and LCIB has been reported previously (Spalding et al., 1983c), before the genes themselves were identified and before the full significance of the observation could be appreciated. It was recognized at the time that the lesion in CAH3 was apparently epistatic to the lesion in LCIB, and that the double mutant exhibited a leaky CO 2 -requiring phenotype in air, similar to the CAH3 single mutant. Our current work definitively demonstrates that a CAH3 loss-of-function mutation can suppress the air-dier phenotype of LCIB mutants, as well as revealing a novel, VL-CO 2 lethal phenotype of the double mutant suppressors. The suppressors exhibited the same levels of photosynthetic Ci affinity and internal Ci accumulation, regardless of whether they were acclimated to L-CO 2 or VL-CO 2. No significant difference in intracellular Ci accumulation was detected between the L-CO 2 acclimated suppressor (LCIB-CAH3 double mutant) and the CAH3 single mutant, indicating the LCIB mutation had no influence on Ci accumulation in the absence of a functional CAH3, which is contrary to the proposed direct involvement of LCIB in Ci transport in L-CO 2 acclimated cells (Spalding et al., 1983b; Wang and Spalding, 2006), but is consistent with the reported epistatic interaction between these two mutations (Spalding et al., 1983c).
58 55 The position of LCIB in the biochemical pathway of Ci uptake and accumulation in C. reinhardtii is informed by the epistatic interaction between the CAH3 and LCIB mutations. It is clear that CAH3 functions to dehydrate HCO - 3 accumulated in the stroma, although it is not known how HCO - 3 gains access to the thylakoid lumen. Because CAH3 mutations are epistatic over LCIB mutations, LCIB must act downstream of CAH3, meaning it must act after the accumulated stromal HCO - 3 is dehydrated to CO 2. The localization of LCIB also is inconsistent with a direct role in Ci transport, regardless whether it is diffusely distributed throughout the stroma or concentrated around the pyrenoid. The combination of LCIB localization and the epistatic interaction between LCIB and CAH3 mutations, together with the clear demonstration that LCIB mutants can transport and accumulate Ci in the absence of functional CAH3, provide a very compelling argument against a direct role for LCIB in Ci transport. Since it is very unlikely LCIB is involved in Ci transport, it appears most likely LCIB is involved in preventing the loss of CO 2 from the chloroplast (Fig. 5). One possibility is that LCIB is involved in sequestering or trapping excess CO 2 from CAH3-catalyzed - dehydration of HCO 3 that might otherwise diffuse out of the chloroplast and be lost. Functionally similar CCM also exists in cyanobacterial (blue-green algae) species and interestingly, Price et al. (2002) have proposed that two unique hydrophilic proteins, ChpX and ChpY (also named CupA and CupB), are critical components of thylakoid-based NDH- 1 CO 2 -uptake complex and aid in the recapture or recycling of CO 2 leakage from carboxysomes by CO 2 hydration activity in the light in the model cyanobacterium, Synechococcus sp. PCC7942 (Maeda et al., 2002; Shibata et al., 2001, 2002; Price et al., 2002). Thus LCIB is possibly part of a novel CO 2 pump/trap and may function mainly to prevent depletion of the HCO - 3 pool by trapping CO 2 from CAH3 dehydration back into the stromal HCO - 3 pool, or by providing a barrier to CO 2 diffusion back out of the pyrenoid. Although it is clear that CAH3 mutations are epistatic to LCIB mutations and thus that LCIB must function downstream of CAH3, it is more complicated to explain mechanistically how the characteristics of the individual mutants and the double mutants explain this epistatic interaction. Dehydration, either uncatalyzed or catalyzed by the stromal carbonic anhydrase CAH6, of the very high stromal HCO - 3 concentration in the LCIB-CAH3 double mutant ad-su6-1 may provide a direct supply of CO 2 to Rubisco sufficient to support
59 56 growth and suppress the air-dier phenotype of ad1-1 in L-CO 2 acclimated cells. However, the stromal HCO - 3 concentration of VL-CO 2 acclimated ad-su6-1 also is fairly high, yet apparently fails to provide Rubisco sufficient CO 2 to support growth in VL-CO 2 conditions. The predicted carboxylation rates for Rubisco (64 µmol CO 2 mg -1 Chl h -1 and 41 µmol CO 2 mg -1 Chl h -1 for L-CO 2 and VL-CO 2 acclimated cells, respectively) assuming complete equilibration at ph of 8.5 between HCO - 3 and CO 2 at the observed internal Ci concentrations of 6.15 mm (50 µm external Ci) and 2.9 mm (20 µm external Ci) for L-CO 2 and VL-CO 2 acclimated cells, respectively, substantially overestimates the observed photosynthetic rates of 18 µmol CO 2 mg -1 Chl h -1 and 5.8 µmol CO 2 mg -1 Chl h -1 for L-CO 2 and VL-CO 2 acclimated cells, respectively. On the other hand, calculations assuming an uncatalyzed rate of HCO - 3 dehydration (CO 2 supply rates of 8 µmol CO 2 mg -1 Chl h -1 and 2.9 µmol CO 2 mg -1 Chl h -1 for L-CO 2 and VL-CO 2, respectively) underestimate the actual photosynthetic rates (see Spalding and Portis, 1985). Thus the observed photosynthetic rates argue for only limited CAH6-catalyzed dehydration of the accumulated HCO - 3, which may explain why an internal Ci concentration of 2.9 mm in VL-CO 2 acclimated cells is unable to support growth under VL-CO 2 conditions. A key question is why an observed photosynthetic rate of 18.2 µmol CO 2 mg -1 Chl h -1 in L-CO 2 allows for substantial growth of the double mutant suppressor but a rate of 5.8 µmol CO 2 mg -1 Chl h -1 in VL-CO 2 does not. A photosynthetic rate of 5.8 µmol CO 2 mg -1 Chl h -1 may simply be below the threshold for survival, a conclusion supported by the observed rates of photosynthesis of three other strains unable to grow, L-CO 2 acclimated LCIB mutant ad1 at 50 µm external Ci and VL-CO 2 acclimated WT-su6 (CAH3 mutation alone) and cia5 at 20 µm external Ci, which were 7.4, 7.2 and 4.3 µmol CO 2 mg -1 Chl h -1, respectively, and by the minimum photosynthetic rate observed for any strain able to grow, which was 11.8 µmol CO 2 mg -1 Chl h -1 (L-CO 2 acclimated cia5 at 50 µm external Ci). Based on the reported effect of photon flux density on Ci accumulation (Spalding, 1990), we - also would expect substantially lower stromal HCO 3 accumulation and thus a lower photosynthetic rate under light conditions used for growth (~100 µmol photons m -2 s -1 ), which is much lower than that used for photosynthesis and Ci uptake measurements (~800 µmol photons m -2 s -1 ).
60 57 Even though the data presented here help identify the position of LCIB in the pathway for Ci uptake and accumulation by placing its function downstream of CAH3, the actual function of LCIB remains a mystery, as does how the single mutation in LCIB eliminates almost all Ci accumulation to the same extent as the cia5 mutant, in which almost no limiting-co 2 inducible genes are expressed (Xiang et al., 2001; Fukuzawa et al., 2001). LCIB is essential in L-CO 2 conditions, and we hypothesize that LCIB might be involved in somehow preventing the loss of CAH3-generated CO 2 that is not captured by Rubisco in the pyrenoid, either by preventing its diffusion from the pyrenoid or by recapturing any CO 2 escaping from the pyrenoid or from thylakoids outside the pyrenoid. The physiological observations for the single and double mutants are consistent with the role in CO 2 trapping, and the immunolocalization of LCIB in the stroma or surrounding the pyrenoid is more consistent with CO 2 trapping than with Ci transport. The constitutively expressed chloroplast stromal CAH6 might normally also help retain Ci in the stroma by trapping it as HCO - 3. It is tempting to speculate that LCIB and CAH6 might cooperate in trapping CO 2 released from the pyrenoid and/or from thylakoids, especially since CAH6 reportedly is differentially concentrated around the pyrenoid (Mitra et al., 2004), as is LCIB, at least under some conditions. It will be interesting to investigate the potential co-localization of LCIB and CAH6, and the generation of CAH6 mutants by either an RNAi approach or by insertional mutagenesis could help to clarify the physiological roles of and any potential interactions between LCIB and CAH6 in the L-CO 2 acclimation state. Materials and Methods Cell Strains and Culture Conditions C. reinhardtii strains CC125 and CC620 were obtained from the Chlamydomonas Stock Center, Duke University, Durham, NC. The LCIB-defective mutant ad1 was generated by insertional mutagenesis (Wang and Spalding, 2006). And the cia5 mutant was a gift from Dr Donald P. Weeks (University of Nebraska, Lincoln). Media and growth conditions for C. reinhardtii strains have been previously described (Wang and Spalding, 2006). All strains were maintained on CO 2 minimal plates and kept in high CO 2 (air enriched with 5% vol/vol CO 2 ) chambers at room temperature, under
61 58 continuous illumination (50 µmol photons m -2 s -1 ). Liquid cultures were grown on a gyratory shaker under aeration in white light (approximately 100 µmol photons m -2 s -1 ). For experiments in which cells were shifted from high to limiting CO 2 (low CO 2, ppm or very low CO 2, ppm) conditions, cells were cultured in CO 2 minimal medium aerated with 5% CO 2 to a density of cells/ml and then shifted to aeration with the appropriate limiting CO 2 for various times. Very low CO 2 was obtained by mixing normal air with compressed, CO 2 -free air. Isolation of Suppressors, Growth Spot Tests, and Genetic Analysis Glass bead transformations were performed as described previously (Van and Spalding, 1999). Air-dier mutant ad1 was transformed with linearized plasmid psi103 (Sizova et al., 2001) and spread onto CO 2 minimal plates containing 15 µg/ml paromomycin. Plates were placed in air for 6 weeks, and surviving colonies selected as putative suppressors were verified by spot tests. For spot test of growth, actively growing cells were serially diluted to similar cell densities in minimal medium and spotted (5 µl/spot) onto minimal agar plates, and grown in various CO 2 concentrations for around 9 days. Genetic crosses and random progeny analyses were performed as described by Harris (1989). DNA, RNA and Protein-Blot Analysis Southern analyses were performed as described by Van and Spalding (1999). Northern blot hybridizations were performed by standard procedures. Total RNA was prepared by the acid guanidinium thiocyanate method (Chomczynski and Sacchi, 2006). RNA (~15 µg) were separated on 0.9% formaldehyde - agarose gels and blotted onto nylon transfer membranes (Osmonics Inc.). PCR amplified cdna sequences of corresponding genes were used as templates and all probes were generated by randomly primed labeling with [α- 32 P]dCTP following instructions of the manufacturer. Each northern blot was analyzed by phosphorimager scanning (BioRad, CA). For total protein analyses, cells were harvested and resuspended in a buffer containing 10 mm Tris-HCl ph 7.5, 1 mm EDTA, 10 mm NaCl, 1 mm PMSF, 1 mm benzamidine and 5
62 59 mm DTT. Protein concentrations were measured using Bio-Rad protein assay kit (Bio-Rad Catalog Number ). Proteins were separated on 12% polyacrylamide gels as described previously (Laemmli, 1970). Immunoblotting was performed as described in the protocol from Bio-Rad Laboratories. The CAH3 antiserum was a gift from James V. Moroney (Louisiana State University, USA) and was generated using a synthetic peptide derived from an internal amino acid sequence of CAH3 (residues ) as antigen. Isolation of Sequences Flanking the Plasmid Insert in ad-su6 by Inverse PCR Based on information from Southern blot analysis, SacI was used to digest the genomic DNA isolated from ad-su6 to produce a fragment with a size ~4.7 kb, including part of the inserted psi103 vector and its 3 flanking genomic DNA. The SacI-digested ad-su6 genomic DNA (1 µg) was circularized with 1 unit of T4 DNA ligase (Invitrogen), precipitated, and the circularized product was used as a template for inverse PCR by using standard PCR procedures. Two pairs of primers were designed, with each pair complementing the psi103 sequence in opposite orientations. Both primer pairs produced PCR products with the correct predicted sizes, and amplified DNA from one primer pair (5 - GGTCTGACGCTCAGTGGAACGA-3 and 5 -CGC AAC GCA TCG TCC ATG CTT C-3 ) was sequenced to determine the sequences flanking the insert. Photosynthetic O 2 Exchange and Ci Uptake Measurements Low CO 2 -induced or very low CO 2 -induced cells (24 hour induction) were collected by centrifugation and resuspended in 25 mm MOPS-KOH buffer to a final chlorophyll concentration of 20~25 µg/ml for analysis of photosynthesis and Ci uptake. Photosynthetic O 2 evolution was measured at 25 C with a Clark-type oxygen electrode (Rank Brothers, Cambridge, UK). Under constant illumination (800 µmol photons/m 2 /s), cells were transferred to the electrode chamber and allowed to exhaust endogenous Ci until net O 2 exchange was zero. Measurements were initiated by addition of various concentrations of NaHCO 3. Oxygen evolution rates were recorded as V20 or V50 when 20 µm or 50 µm NaHCO 3 were used, respectively. The maximum O 2 evolution rate, V4000, was obtained by using 4000 µm NaHCO 3. Relative affinity for Ci was calculated as the ratios P20 =
63 60 V20/V4000 and P50 = V50/V4000. Ci uptake by C. reinhardtii cells at 20 µm or 50 µm total Ci was estimated by the silicone oil filtration technique (Badger et al., 1980). The cell volume was measured by using 14 C-sorbitol and 3 H 2 O as previously described (Wirtz et al., 1980). The intracellular Ci concentration was calculated by using cell volume and the acid labile 14 C data. ACKNOWLEDGMENTS Here we thank Prof. Donald P. Weeks (Biochemistry Department, University of Nebraska) for providing CAH3 genomic DNA plasmid, James V. Moroney (Department of Biological Sciences, Louisiana State University) for the anti-cah3 antiserum. References Badger MR, Kaplan A, Berry JA (1980) Internal inorganic carbon pool of Chlamydomonas reinhardtii: evidence for a carbon dioxide concentrating mechanism. Plant Physiol 66(3): Burow MD, Chen ZY, Mouton TM, Moroney JV (1996) Isolation of cdna clones of genes induced upon transfer of Chlamydomonas reinhardtii to low CO 2. Plant Mol Biol 31: Beardall J, Giordano M (2002) Ecological implications of microalgal and cyanobacterial CO 2 concentrating mechanisms and their regulation. Funct Plant Biol 29: Chen ZY, Lavigne LL, Mason CB, Moroney JV (1997) Cloning and overexpression of two cdnas encoding the low-co 2 -inducible chloroplast envelope protein LIP-36 from Chlamydomonas reinhardtii. Plant Physiol 114: Chomczynski P, Sacchi N (2006) The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: twenty-something years on. Nat Protol 1(2): Cole DG, Diener DR, Himelblau AL, Beech PL, Fuster JC, Rosenbaum JL (1998) Chlamydomonas kinesin-ii dependent intraflagellar transport (IFT): IFT particles contain proteins required for ciliary assembly in Caenorhabditis elegans sensory neurons. J Cell Biol 141(4): Fischer N, Rochaix JD (2001) The flanking regions of PsaD drive efficient gene expression in the nucleus of the green alga Chlamydomonas reinhardtii. Mol Genet Genomics 265: Funke RP, Kovar JL, Weeks DP (1997) Intracellular carbonic anhydrase is essential to photosynthesis in Chlamydomonas reinhardtii at atmospheric levels of CO 2. Plant Physiol 114:
64 61 Fukuzawa H, Miura K, Ishizaki K, Kucho KI, Saito T, Kohinata T, Ohyama K (2001) Ccm1, a regulatory gene controlling the induction of a carbon-concentrating mechanism in Chlamydomonas reinhardtii by sensing CO 2 availability. Proc Natl Acad Sci USA 98(9): Giordano M, Beardall J, Raven JA (2005) CO 2 concentrating mechanisms in algae: mechanisms, environmental modulatioin, and evolution. Annu Rev Plant Biol 56: Hanson DT, Franklin LA, Samuelsson G, Badger MR (2003) The Chlamydomonas reinhardtii cia3 mutant lacking a thylakoid lumen-localized carbonic anhydrase is limited by CO 2 supply to Rubisco and not photosystem II function in vivo. Plant Physiol 132: Harris EH (1989) The Chlamydomonas Source Book: a comprehensive guide to biology and laboratory use. Academic Press Inc, San Diego, pp IM CS, Grossman AR (2001) Identification and regulation of high light-induced genes in Chlamydomonas reinhardtii. Plant J 30(3): Karlsson J, Clarke AK, Chen ZY, Hugghins SY, Youn-II P, Husic HD, Moroney JV, Samuelsson G (1998) A novel α-type carbonic anhydrase associated with the thylakoid membrane in Chlamydomonas reinhardtii is required for growth at ambient CO 2. EMBO J 17(5): Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: Maeda S, Badger MR, Price GD (2002) Novel gene products associated with NdhD3/D4- containing NDH-1 complexes are involved in photosynthetic CO 2 hydration in the cyanobacterium, Synechococcus sp. PCC7942. Mol Microbiol 43: Mariscal V, Moulin P, Orsel M, Miller AJ, Fernández E, Galván A (2006) Differential regulation of the Chlamydomonas Nar1 gene family by carbon and nitrogen. Protist 157(4): Mitra M, Lato SM, Ynalvez RA, Xiao Y, Moroney JV (2004) Identification of a new chloroplast carbonic anhydrase in Chlamydomonas reinhardtii. Plant Physiol 135: Miura K, Yamano T, Yoshioka S, Kohinata T, Inoue Y, Taniguchi F, Asamizu E, Nakaura Y, Tabata S, Yamato KT, Ohyama K, Fukuzawa H (2004) Expression profiling-based identification of CO 2 -responsive genes regulated by CCM1 controlling a carbon-concentrating mechanism in Chlamydomonas reinhardtii. Plant Physiol 135: Moroney JV, Togasaki RK, Husic HD, Tolbert NE (1987) Evidence that an internal carbonic anhydrase is present in 5% CO 2 -grown and air-grown Chlamydomonas. Plant Physiol 84(3): Moroney JV, Ynalvez RA (2007) Proposed carbon dioxide concentrating mechanism in Chlamydomonas reinhardtii. Eukaryot Cell 6(8): Palmqvist K, Sjoberg S, Samuelsson G (1988) Induction of inorganic carbon accumulation in the unicellular green algae Scenedesmus obliquus and Chlamydomonas reinhardtii. Plant Physiol 87(2): Pollock SV, Prout DL, Godfrey AC, Lemaire SD, Moroney JV (2004) The Chlamydomonas reinhardtii proteins Ccp1 and Ccp2 are required for long-term growth,
65 62 but are not necessary for efficient photosynthesis, in a low-co 2 environment. Plant Mol Biol 56: Price GD, Maeda SI, Omata T, Badger MR (2002) Modes of active inorganic carbon uptake in the cyanobacterium, Synechococcus sp. PCC7942. Funct Plant Biol 29: Price GD, Woodger FJ, Badger MR, Howitt SM, Tucker L (2004) Identification of a SulP-type bicarbonate transporter in marine cyanobacteria. Proc Natl Acad Sci USA 101(52): Pronina NA, Ramazanov ZM, Semenenko VE (1981) Carbonic-anhydrase activity of Chlorella cells as a function of CO 2 concentration. Sov Plant Physiol 28: Pronina NA, Semenenko VE (1990) Membrane-bound carbonic anhydrase takes part in CO 2 concentration in algal cells. Curr Res Photosynth 4: Raven JA (1997) CO 2 -concentrating mechanisms: a direct role for thylakoid lumen acidification? Plant Cell Environ 20: Rolland N, Dorne AJ, Amoroso G, Sultemeyer DF, Joyard J, Rochaix JD (1997) Disruption of the plastid ycf10 open reading frame affects uptake of inorganic carbon in the chloroplast of Chlamydomonas. EMBO J 16(22): Sanders MA, Salisbury JL (1995) Immunofluorescence microscopy of cilia and flagella. Methods Cell Biol. 47: Spalding MH, Spreitzer RJ, Ogren WL (1983a) Carbonic anhydrase-deficient mutant of Chlamydomonas reinhardtii requires elevated carbon dioxide concentration for photoautotrophic growth. Plant Physiol 73: Spalding MH, Spreitzer RJ, Ogren WL (1983b) Reduced inorganic carbon transport in a CO 2 -requiring mutant of Chlamydomonas reinhardtii. Plant Physiol 73: Spalding MH, Spreitzer RJ, Ogren WL (1983c) Genetic and physiological analysis of the CO 2 -concentrating system of Chlamydomonas reinhardtii. Planta 159: Spalding MH, Portis AR (1985) A model of carbon dioxide assimilation in Chlamydomonas reinhardii. Planta 164: Spalding MH (1990) Effect of photon flux density on inorganic carbon accumulation and net CO 2 exchange in a high-co 2 -requiring mutant of Chlamydomonas reinhardtii. Photosynth Res 24: Spalding MH, Van K, Wang Y, Nakamura Y (2002) Acclimation of Chlamydomonas to changing carbon availability. Funct Plant Biol 29: Spalding MH (2008) Microalgal carbon-dioxide-concentrating mechanisms: Chlamydomonas inorganic carbon transporters. J Exp Bot 59(7): Shibata M, Ohkawa H, Kaneko T, Fukuzawa H, Tabata S, Kaplan A,Ogawa T (2001) Distinct constitutive and low-co 2 -induced CO 2 uptake systems in cyanobacteria: genes involved and their phylogenetic relationship with homologous genes in other organisms. Proc Natl Acad Sci USA 98: Shibata M, Ohkawa H, Katoh H, Shimoyama M, Ogawa T (2002) Two CO 2 uptake systems in cyanobacteria: four systems for inorganic carbon acquisition in Synechocystis sp. strain CC6803. Funct Plant Biol 29: Sizova I, Fuhrmann M, Hegemann P (2001) A Streptomyces rimosus aphviii gene encoding for a new type phosphotransferase provides stable antibiotic resistance to Chlamydomonas reinhardtii. Gene 277:
66 63 Sültemeyer DF, Klöck G, Kreuzberg K, Fock HP (1988) Photosynthesis and apparent affinity for dissolved inorganic carbon by cells and chloroplasts of Chlamydomonas reinhardtii grown at high and low CO 2 concentrations. Planta 176: Suzuki K, Spalding MH (1989) Adaptation of Chlamydomonas reinhardtii high-co 2 - requiring mutants to limiting CO 2. Plant Physiol 90: Vance P, Spalding MH (2005) Growth, photosynthesis and gene expression in Chlamydomonas over a range of CO 2 concentrations and CO 2 /O 2 ratios: CO 2 regulates multiple acclimation states. Can J Bot 83: Van K, Spalding MH (1999) Periplasmic carbonic anhydrase structural gene (Cah1) mutant in Chlamydomonas reinhardtii. Plant Physiol 120: Wang Y, Spalding MH (2006) An inorganic carbon transport system responsible for acclimation specific to air levels of CO 2 in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 103: Wirtz W, Stitt M, Heldt HW (1980) Enzymic determination of metabolites in the subcellular compartments of Spinach protoplasts. Plant Physiol 66(1): Xiang Y, Zhang J, Weeks DP (2001) The cia5 gene controls formation of the carbon concentrating mechanism in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 98:
67 64 Figure Legends Figure 1. Growth of ad1 suppressors and CAH3 genomic DNA complemented ad-su6 on minimal plates in H-CO 2, L-CO 2 and VL-CO 2 chambers. Cells grown to logarithmic phase were diluted to the indicated numbers ( 10 3 ) per 5 µl, spotted on plates and incubated for 9 days under dim lights. Figure 2. Southern blot analysis of ad-su6 and ad-su7 probed with a 880-bp PCR fragment of the Par R gene. (A) Map of the Par R portion of the psi103 plasmid used for insertional mutagenesis. Arrows above and below the map indicate the primers used to make the probe. (B) Southern analysis of two suppressors, with linearized plasmid as control. Genomic DNA was digested with the indicated restriction enzymes. (C) Map of the CAH3 genomic region in ad-su6. Black arrows, exons; grey arrow, psi103 insertion; grey bars, introns. Figure 3. (A) Northern blot analysis of ad1 and ad-su6. Cells were adapted to L-CO 2 or VL-CO 2 conditions for 4h, 14h respectively and 10 µg of total RNA was used. (B) Immunoblot analysis of whole cell fractions. Cells were either H-CO 2 grown or induced 24 hours under L-CO 2 (L24) or VL-CO 2 (VL24) conditions. 100 µg total protein were analyzed with CAH3 antibody. su6-c1, su6-c3 are two CAH3 genomic DNA-complemented ad-su6 transformants as described in Figure 1. Figure 4. Simple schematic of the C. reinhardtii CCM, showing the relative position of CAH3 and LCIB in the pathway of Ci transport, HCO - 3 accumulation, and the subsequent - dehydration of HCO 3 to produce CO 2.
68 65 Table 1. Relative affinity of photosynthetic O 2 evolution in cells acclimated to L-CO 2 or VL-CO 2 for 1 day. L-CO 2 VL-CO 2 Strains P20 P50 V max P20 P50 V max CC ± ± ± ± ± ±18 ad ± ± ± ± ± ±8 wt-su6 0.08± ± ± ± ± ±9 ad-su ± ± ± ± ± ±10 cia5 0.08± ± ± ± ± ±9 Rates of O 2 evolution (µmol mg Chl -1 h -1 ) were determined in ph 7.3 (MOPS-KOH) buffer at 20 µm (V20), 50 µm (V50), and 4000 µm (V4000) NaHCO 3. Relative Ci affinity was calculated as ratios: P20 = V20/V4000 and P50 = V50/V4000. V max = V4000.
69 66 Table 2. Intracellular Ci accumulation in cells acclimated to L-CO 2 or VL-CO 2 for 1 day. L-CO 2 VL-CO 2 Strains 50µM Ci 20 µm Ci 50µM Ci CC ± ± ±0.24 ad ± ± ±0.22 wt-su6 7.60± ± ±1.45 ad-su ± ± ±0.58 cia5 0.19± ± ±0.15 Internal Ci accumulation (mm Ci after 80 s) was determined by silicone oil centrifugation in ph 7.3 (MOPS-KOH) buffer at initial external Ci concentrations of 20 µm and 50 µm [ 14 C]-NaHCO 3 and a Chl concentration of 25 µg/ml.



70 Figure 1. 67

71 Figure 2. 68




72 Figure 3. 69



73 Figure 4. 70
74 71 CHAPTER 4. INSERTIONAL SUPPRESSORS OF CHLAMYDOMONAS REINHARDTII THAT RESTORE GROWTH OF AIR-DIER lcib MUTANTS IN LOW-CO 2 Abstract C. reinhardtii and other microalgae show adaptive changes to limiting CO 2 conditions by induction of CO 2 -concentrating mechanisms (CCM). Despite recent progress in the understanding of the CCM and identification of several limiting-co 2 -regulated genes, the molecular basis of intracellular Ci uptake and accumulations are largely unknown. The limiting-co 2 -inducible gene, LCIB, encodes an apparently soluble plastid protein and is proposed to play a role in trapping CO 2 released by CAH3 dehydration of accumulated Ci, especially in low CO 2 (L-CO 2 ; ~ 0.04% CO 2 ) conditions. In order to gain further insight into the mechanisms of Ci uptake and accumulation in L-CO 2 acclimated C. reinhardtii, we performed an insertional mutagenesis screen to isolate extragenic suppressors that restore the growth of lcib mutants (pmp1 and ad1) in L-CO 2. Four independent suppressors (su1,su4,su5 and su8) are described here and classified by their photosynthetic affinities for inorganic carbon and expression patterns of known limiting-co 2 -inducible transcripts. All four suppressors have single inserts, although genetic linkage between phenotype and insert was established only in ad-su8. Genetic analysis of the four suppressors identified 2 allelic, dominant suppressors (pmp-su4 and pmp-su5), and two recessive suppressors (pmp-su1 and ad-su8). Consistent with the suppression phenotype, both the relative affinities of photosynthetic O 2 evolution and internal Ci accumulation in all 4 suppressors were substantially increased relative to pmp1/ad1 in L-CO 2 acclimated cells. Interestingly, the relative affinity of pmp-su4/su5 was intermediate between pmp-su1 and pmp1. Our results reveal a complex LCIB-dependent and independent Ci uptake/accumulation systems and therefore might be useful to provide new clues about the roles played by LCIB in limiting Ci acclimation.
75 72 Introduction Although atmospheric CO 2 has increased from a pre-industrial concentration of about 280 ppm to a globally averaged concentration of approximately 387 ppm at present, this level of CO 2 concentration is still limiting to photoautotrophic growth of most plants and photosynthetic microorganisms, largely due to the inefficiency of ribulose bisphosphate carboxylase-oxygenase (Rubisco) enzyme, which has both oxygenase as well as the carboxylase function and a low CO 2 affinity (Long et al., 2004). A number of photosynthetic organisms have developed strategies to increase the level of CO 2 at the location site of Rubisco, including well known C 4 photosynthesis and Crassulacean acid metabolism (CAM) in terrestrial vascular plants. In contrast, a different type of CO 2 concentrating mechanism (CCM) is found in some C 3 -type aquatic photosynthetic microorganisms, which also face a challenge of ~10,000 times slower diffusion rate of dissolved CO 2 than the rate of diffusion in air. Much knowledge of the mechanisms of the CCM has been gained in the wellinvestigated eukaryotic organisms as Chlamydomonas reinhardtii (Giordano et al., 2005; Moroney and Ynalvez, 2007; Spalding, 2008). When the availability of Ci (inorganic carbon, CO 2 and HCO - 3 ) is limiting, the CCM is induced via active Ci uptake systems on both plasma membrane and chloroplast envelope to increase internal Ci accumulation, and various carbonic anhydrases function to dehydrate accumulated bicarbonate and provide elevated internal CO 2 concentrations to the pyrenoid, a Rubisco-containing microcompartment of the chloroplast (Palmqvist et al., 1988; Sültemeyer et al., 1988; Funke et al., 1997; Karlsson et al., 1998; Moroney and Ynalvez, 2007). Recently, at least three distinct CO 2 level dependent acclimation states were identified in C. reinhardtii, including: high CO 2 (H-CO 2, 5% - 0.5% CO 2 ), low CO 2 (L-CO 2, 0.4% % CO 2 ) and very low CO 2 (VL-CO 2, 0.01% % CO 2 ) (Vance and Spalding, 2005). Over the last decade, significant progress has been made in understanding the functionally similar prokaryotic CCMs in cyanobacteria, and at least five distinct although functionally redundant modes of Ci uptake systems have been identified in Synechocystis and Synechococcus, including three bicarbonate transporters and two active CO 2 uptake systems associated with NDH-1 complexes (Price et al., 2002, 2004). Despite extensive efforts to identify Ci transport systems in C. reinhardtii, little information other than
76 73 physiological evidence is available regarding active Ci transport in whole cells and isolated chloroplasts (Palmqvist et al., 1988; Sültemeyer et al., 1988). While a number of promising candidate genes and proteins have been identified, none of them have been definitely confirmed to be Ci transporters (Pollock et al., 2004; Mariscal et al., 2006; Spalding, 2008; Duanmu et al., 2009). Several labs screened chemically or insertionally mutagenized cells for growth defects in low or very low CO 2 (Van et al., 2001; Colombo et al., 2002; Thyssen et al., 2003). This approach has yielded mutants in CA activity (Spalding et al., 1983a; Moroney et al., 1986; Funke et al., 1997), the photorespiratory pathway (Suzuki, 1995; Nakamura et al., 2005), regulatory transcription factors (Xiang et al., 2001; Fukuzawa et al., 2001) and unknown proteins. Only two allelic mutants, pmp1 and ad1, were characterized as deficient in Ci transport and were further identified as conditional lethal in air levels of CO 2, but not in VL- CO 2, indicating an L-CO 2 specific Ci accumulation component was disrupted (Van et al., 2001; Spalding et al., 2002). The defective gene in pmp1 and ad1 was identified as LCIB, a limiting-co 2 -upregulated gene, with the gene product targeted to the chloroplast stroma and the function of this soluble protein proposed to be involved in minimizing leakage of CAH3- generated CO 2 from the pyrenoid (Wang and Spalding, 2006; Duanmu et al., 2009). Three additional genes in the C. reinhardtii genome share high similarities to LCIB (62~65%) in deduced amino acid sequences, including a similar CO 2 -responsive gene LCIC and two previously unreported genes LCID and LCIE. This small, unique LCIB gene family are also identified in several other green algae as evidenced by bioinformatic analysis of several databases (Wang and Spalding, 2007; Spalding, 2008), and more recently, some cyanobacteria and bacteria are also found to contain LCIB homologs (Wang and Spalding, unpublished results). To understand the function of LCIB in L-CO 2 specific Ci-uptake and accumulation processes and identify other functional components involved in chloroplast Ci accumulation in C. reinhardtii, we performed suppressor screens using an insertional mutagenesis approach. In this paper, we describe the identification and characterization of three pmp1 suppressors and one ad1 suppressor, that grow normally in all CO 2 conditions. Our data suggest that mutations in several genes/loci could mask the L-CO 2 conditional lethal
77 74 phenotype of lcib mutants. Results Isolation of Insertionally Generated Suppressors The C. reinhardtii strains chosen for this study were pmp1 and ad1, containing a point nonsense mutation or a deletion mutation of LCIB, respectively (Spalding, 1983b; Wang and Spalding, 2006). These two allelic strains cannot grow in L-CO 2 but can grow either in H- CO 2 or in VL-CO 2. In an effort to identify suppressors which can restore the growth of lcib mutants in L-CO 2, we performed insertional mutagenesis using a Par R -containing plasmid (psi103; Fig.1A) to transform pmp1 and ad1. From ~10 6 transgenic lines, eight (5 pmp1 and 3 ad1) colonies capable of growth under L-CO 2 were isolated. The suppressor phenotypes of two transformants (pmp-su2, pmp-su3) were lost over time and it has been shown that two allelic ad1 suppressors (ad-su6 and ad-su7) were caused by defects in the CAH3 gene, which encodes a thylakoid lumen targeted carbonic anhydrase (Duanmu et al., 2009). The other four suppressors, including three pmp1 suppressors (pmp-su1, pmp-su4 and pmp-su5) and one ad1 suppressor (ad-su8) are described in this study. The three pmp- suppressors exhibited a near-wild type growth phenotype in L-CO 2 and VL-CO 2 (Fig. 2). These pmp-suppressors also showed resistance to the antibiotic paromomycin and were found by Southern analysis to contain only one copy of the Par R insert (Fig. 1B). Genetic analysis was performed by crossing the suppressors to wild-type strain CC-620, and the meiotic progeny demonstrated that paromomycin resistance conferred by the Par R insert did not cosegregate with the suppressor phenotype. Suppressor ad-su8 demonstrated suboptimal growth in L-CO 2 where the parental strain ad1 could not grow at all (Fig. 2). Southern analysis with probe specific for the Par R gene indicated a single insert present in ad-su8 (Fig. 1B). The ad-su8 strain also was crossed with CC620 to determine whether the suppression phenotype was linked to the inserted Par R gene. More than 130 Zeo R (zeocin-resistance, caused by the Ble R insert responsible for the LCIB deletion mutation) random progeny were screened for their growth in different levels of CO 2 and resistance to paromomycin. All 95 random progeny with the suppressor phenotype were paromomycin resistant, while all paromomycin-sensitive progeny exhibited
78 75 an ad1-like growth phenotype in L-CO 2 and VL-CO 2, demonstrating linkage of the suppressor phenotype to the Par R insert. The genetic linkage indicates that the functional Par R insert is located in, or close to the disrupted gene responsible for the suppressor phenotype. Genetic Characteristics of Suppressors The four suppressors are derived from two recipient strains, pmp1 and ad1 (all strains used in this study are listed in Table 1). To test if pmp-su1, pmp-su4 and pmp-su5 could also suppress ad1 air-dier phenotype and correspondingly, if ad-su8 could restore growth of pmp1 in L-CO 2, mt+ progeny of pmp-su1/4/5 were crossed with ad1 and also ad-su8 with mt+ pmp1, and the phenotypes of meiotic progeny were analyzed by random progeny selection or tetrad analysis. In all crosses, su1/4/5/8 could suppress both pmp1 and ad1 mutant alleles of LCIB gene in L-CO 2. Suppressors pmp-su1/4/5 and ad-su8 were crossed with wild-type strain CC620 and by tetrad identification, progeny with su1/4/5/8 mutations in a WT background were selected and the growth of these strains in L- and VL-CO 2 was investigated in spot tests. No observable growth defects of these strains compared with WT strain CC620 were identified (data not shown), indicating the suppressor mutations themselves are not detrimental for limiting-co 2 growth of C. reinhardtii. Complementation analysis was performed to test allelism and to place the various suppressor mutations into different complementation groups. Only the pmp-su4 pmp-su5 cross failed to generate pmp1-phenotype colonies. Suppressors pmp-su4 and pmp-su5 are independently isolated suppressors, and Southern blot analysis demonstrated they were not siblings (Fig. 1B). Thus pmp-su1 and ad-su8 each defines a separate, new suppressor locus while the same gene/locus appears to be affected in pmp-su4 and pmp-su5. Diploid analysis was performed to ascertain the dominant/recessive nature of the suppressor mutations. Based on growth phenotypes of the heterozygous diploids, su1 mutation was determined to be recessive (Table 2). General Characteristics of Suppressors Intracellular Ci accumulation and the response of photosynthetic O 2 evolution to external Ci concentrations at ph 7.3 were compared for wild-type, two lcib mutants, pmp1
79 76 and ad1, and the four suppressor lines (Tables 3 and 4). Consistent with the air-dier phenotype, L-CO 2 acclimated pmp1 and ad1 showed dramatically decreased photosynthetic Ci affinities relative to wild-type cells acclimated under the same conditions. Intracellular Ci accumulation in L-CO 2 acclimated pmp1 and ad1-1 cells (walled progeny of ad1) also was similar to the nonacclimating mutant cia5, in which almost none of the limiting-co 2 - inducible genes are expressed (Xiang et al., 2001; Fukuzawa et al., 2001; Miura et al., 2004). In contrast, when acclimated to L-CO 2, photosynthetic affinities of all suppressors were significantly higher than pmp1 or ad1 and correspondingly, a much higher internal Ci accumulation was observed in all the suppressors as evidenced by direct [ 14 C] NaHCO 3 uptake analysis. Interestingly, both pmp-su1 and ad-su8 demonstrated similar level of affinities as wild-type control CC620, while the relative affinities of pmp-su4 and pmp-su5 were intermediate between pmp-su1 and pmp1 (Table 3). The pmp1 and ad1 mutants are deficient in Ci transport and accumulation only in L- CO 2 but not in VL-CO 2 growth conditions. True to this air-dier phenotype, VL-CO 2 acclimated pmp1 and ad1 regained a higher photosynthetic Ci affinity, up to approximately 49% to 54% of the wild-type strains. In contrast, VL-CO 2 acclimated pmp-su1 exhibited similar photosynthetic Ci-affinities as pmp1, while pmp-su4/5 showed slightly lower affinities than pmp1, and ad-su8 demonstrated WT phenotype of photosynthetic Ci affinities (Table 3). The selected suppressors in WT background are named wt-su1, wt-su5, and wt-su8. Although no obvious growth differences are observed when the wt-suppressors were spot tested in all CO 2 conditions, high-co 2 -grown wt-suppressors were associated with higher photosynthetic Ci affinities than wild-type CC620 when measured at 50µM total Ci, but not at 20µM total Ci. Unexpectedly, high-co 2 -adapted pmp- or ad- suppressors demonstrated similar photosynthetic Ci affinities to pmp1/ad1 at both 20 and 50µM total Ci (Table 3). Accumulation of Low CO 2 -inducible Transcripts Along with induction of a functional CCM, acclimation to limiting CO 2 also results in upregulation or down-regulation of specific genes/proteins (Miura et al., 2004). Transcript abundance of several limiting-co 2 -inducible genes also were analyzed, including periplasmic carbonic anhydrase CAH1, a probable plasma membrane ABC-MRP type
80 77 transporter HLA3, chloroplast soluble protein LCIC, and putative plastid envelope Ci transport candidates LCIA, LCI1, CCP1&CCP2 (for review of these proteins/genes, Spalding, 2008). Strong induction or up-regulation of the above genes was observed in both pmp1 and ad1 after 4h of L-CO 2 acclimation, and the message levels of HLA3, LCI1, LCIC reproducibly decreased after 14h of acclimation (Fig. 3A). A similar pattern of expression for these seven genes was detected in pmp-su4 and pmp-su5 (data only shown for pmp-su5). Suppressors pmp-su1 and ad-su8, which showed WT-like phenotype of photosynthetic Ci affinities when acclimated in L-CO 2, retained (pmp-su1) or induced (ad-su8) much higher level expression of HLA3 mrna after 14 h of ambient CO 2 induction. Over-accumulation of the HLA3 transcript after 14h L-CO 2 induction was also observed in wt-su1 and ad-su1 (Fig. 3B). Inverse PCR to Identify Genomic DNA Disrupted in ad-su8 and Rough Mapping of pmp-su5 Genetic linkage analysis demonstrated that the single insert was linked with the suppressor phenotype in ad-su8. We employed inverse PCR to identify the flanking DNA in ad-su8 by taking advantage of the unique sequences of the par R gene originally from Streptomyces rimosus (Sizova et al., 2001). Genomic DNA fragments flanking the 5 and 3 of the par R insert were PCR-amplified following PvuII or SacI digestion and religation, and the flanking sequences were used in a BLAST search against the C. reinhardtii version 4.0 genome ( The two insertion sites were determined to be located on chromosome_16: and chromosome_16: , respectively. Genomic DNA from ad-su8 was also digested with ApaI and by plasmid rescue, the two flanking regions adjacent to the plasmid insert were confirmed. Chromosome_16 contains around 6.6 Mb nucleotides according to the Version 4.0 genome database. Further PCR analysis of chromosome_16 revealed the deletion of at least four separated DNA fragments totaling around 100kb (data not shown), suggesting complicated DNA rearrangements in ad-su8. Several predicted ORFs included in the ~100 kb deletion in ad-su8 were identified, including the Acyl-CoA dehydrogenase gene at one insertion site and some hypothetical genes. Reverse genetic approach by RNAi-knockdown of candidate genes in ad1 is being evaluated to identify the gene responsible for suppressor phenotype of
81 78 ad-su8. To develop a population for rough mapping of pmp-su5, the mt+ progeny of pmp-su5 was crossed to polymorphic strain S1-D2 and air-dier meiotic progeny were randomly selected. Since wt-su5 demonstrated WT growth phenotype in L-CO 2, these progeny of the cross carry the lcib point mutation and the S1-D2 derived wild-type SU5 allele. We collected ~120 such progeny from six different crosses to minimize analysis of duplicate recombination events and we conclude there are more than 60 independent colonies by PCR analysis of several markers included in the mapping kit (data not shown). With a population of > 60 independent progeny, a 20-cM (Kosambi units) resolution could be expected with a confidence level of 95%. At this resolution, 80% of the progeny should contain the wildtype SU5 allele. The IDA7 marker on chromosome 1 shows the highest linkage, for which 104/120 (87%) progeny showed the PCR product size associated with S1-D2 parent, while all other markers demonstrated linkage frequency of less than 70%. Thus we could unambiguously assign the su5 mutant locus to chromosome 1. Discussion Random insertional mutagenesis has proven to be a useful tool to disrupt and tag the nuclear genes in the unicellular green alga Chlamydomonas reinhardtii. In this study, by simply vortexing pmp1 or ad1 cells and linearized psi103 plasmid with glass beads (Kindle, 1990), four suppressors were identified with single inserts in each, but only one suppressor (ad-su8) demonstrated genetic linkage of the phenotype to the insert. On average, it was found in several labs that most insertionally generated mutants have single copy inserts, while cosegregation of the phenotype with the insert was observed in ~30 % of the transformants, both characters agreeing with the findings in this report (Spalding et al., 2002; Colombo et al., 2002). In the microalgal CCM model system Chlamydomonas reinhardtii, the optimized acquisition of Ci from environment is an essential component of a limiting-co 2 -inducible CCM, and active Ci uptake across both the plasma membrane and chloroplast envelope are well supported experimentally, yet the identity of Ci transport/accumulation systems still remains illusive (Sültemeyer et al., 1989, 1991; Palmqvist et al., 1994). Recently, the
82 79 existence of multiple Ci uptake and accumulation systems were proposed, including putative plasma membrane HCO - 3 transporter HLA3 protein, chloroplast envelope LCIA protein and chloroplast stromal LCIB protein (Duanmu et al., 2009). LCIB is a member of a small gene family in Chlamydomonas, including three additional genes LCIC, LCID and LCIE, and homologies of LCIB could be found in several other green microalgae and some bacterial and cyanobacterial species (Spalding 2008; Wang and Spalding, unpublished data). The conditional lethal, air-dieing phenotype of two lcib mutants, pmp1 and ad1, clearly indicates that LCIB-associated Ci uptake/accumulation system is essential for L-CO 2 acclimation while dispensable under H-CO 2 or VL-CO 2 conditions (Wang and Spalding, 2006). Compared with wild-type CC620, much lower photosynthetic Ci affinities in H-CO 2 and VL-CO 2 acclimated pmp1/ad1 mutants were observed in this study, suggesting both LCIBdependent and independent Ci uptake/accumulation processes operating in VL-CO 2 condition and the basal level of active Ci uptake/accumulation activities in H-CO 2 is impaired in pmp1/ad1 mutants. Consistent with the growth phenotype of suppressors, both apparent photosynthetic Ci affinities and direct Ci uptake are significantly increased in L-CO 2 acclimated suppressor mutnts relative to pmp1/ad1 mutants, indicating restoration of active Ci transport into the cell is essential for suppression of the air dier phenotype of pmp1/ad1. L-CO 2 -acclimated pmp-su5 showed only slightly increased photosynthetic Ci affinities compared with pmp1, while pmp-su1 and ad-su8 demonstrated wild-type level of affinities. Mutations of all three suppressor genes/loci (su1, su4/5, and su8) alone did not result in significant decrease of photosynthetic Ci affinities in L-CO 2 and VL-CO 2 -acclimated cells, while a striking feature shared by all three type suppressors is their increased photosynthetic Ci affinities in H-CO 2 state at 50 µm total Ci, but not at 20 µm total Ci. The increase of photosynthetic Ci affinities in H-CO 2 state at 50 µm total Ci was also masked by lcib mutations, since the combination of lcib mutations and suppressor mutations all showed similar level of affinities as pmp1/ad1 and cia5, in which LCIB gene product was either disrupted or not induced. We propose in the H-CO 2 acclimation state that active Ci uptake/accumulation activities are largely dependent on either LCIB itself or other proteins/complexes regulated by LCIB, and the measured photosynthetic affinities in lcib mutants, either lcib mutations alone or
83 80 combined with suppressor mutations, are mainly due to the passive diffusion of external CO 2. The air dier phenotype of lcib mutants in L-CO 2 and suboptimal photosynthetic Ci affinities in VL-CO 2 suggests that LCIB-dependent Ci uptake/accumulation systems account for most of the active Ci concentrating in L-CO 2 acclimated cells, while LCIB-independent Ci uptake/accumulation systems play major roles in the VL-CO 2 acclimation state. However, LCIB-associated systems still contribute to part of active Ci uptake/accumulations in the VL-CO 2 state as evidenced by the significant differences of VL-CO 2 grown wild-type and lcib mutants. When acclimated under VL-CO 2, suppressor ad-su8 showed WT phenotype of photosynthetic Ci affinities and suppressor pmp-su1 demonstrated a similar level of photosynthetic Ci affinity as that of pmp1/ad1, but the affinities were significantly reduced in pmp-su4/5, indicating the mutations in su4/5 decreased the Ci uptake/accumulation in VL-CO 2. Active Ci uptake/accumulations in all three suppressors (su1, su4/5, su8) are likely to be de-repressed or re-activated in H-CO 2 state, since the photosynthetic Ci affinities in such condition are all significantly higher than wild-type control. Based on allelism tests, it is reasonable to hypothesize that different Ci concentrating systems or regulatory components might be mutated in the three suppressor loci so that the active Ci uptake and accumulation is restored in L-CO 2 growth condition. Active uptake of Ci appears to take place at both plasma membrane and chloroplast envelope and it is commonly accepted that chloroplast is the major site of accumulating intracellular Ci pool. Duanmu et al., (2009) proposed that the putative plasma membrane bicarbonate transporter HLA3 and chloroplast envelope protein LCIA are key synergistic or complementary components of a Ci transport pathway. Northern analysis in the suppressors of HLA3, LCIA and several other limiting-co 2 -inducible genes demonstrated higher level of HLA3 transcript abundance in pmp-su1 and ad-su8, and this pattern of HLA3 expression was also retained in wt-su1. Interestingly, the HLA3 expression in pmp-su4/5 was not significantly different from pmp1, which provides additional evidence that different Ci uptake/accumulation components might be mutated in the three suppressor genes/loci and it is tempting to ascribe the mutations in su1 and su8 to HLA3-dependent Ci uptake system. Since genetic analysis indicates that two different genes/loci were mutated in su1 and su8, it
84 81 remains interesting to investigate why the transcript level of HLA3 was increased in both suppressors. Suppressor mutations in su1 and su4/5 were not linked to the inserts and in su8 which demonstrated cosegregation of the phenotype with plasmid insert, approximately 100kb deletion including more than 14 putative genes was detected. Map-based positional cloning of su1, su4/5 and reverse genetic approaches are being evaluated to identify the genes responsible for suppression phenotypes in these lines. Meanwhile, generation of additional tagged pmp1/ad1 suppressors might also facilitate identification of the key components involved in active Ci uptake/accumulation processes in either L-CO 2 or VL-CO 2 acclimation states. Materials and Methods Cell Strains and Culture Conditions C. reinhardtii strains CC86 (nr-u-2-1, mt+, kanamycin resistant), CC118 (sr-u-2-60, mt+, streptomycin resistant), CC620 and CC2290 (S1-D2 mt-) were obtained from the Chlamydomonas Stock Center, Duke University, Durham, NC. The LCIB-defective mutants pmp1 and ad1 were generated by ethyl methanesulfonate treatment and insertional mutagenesis, respectively (Spalding et al., 1983b; Wang and Spalding, 2006). The cia5 mutant was a gift from Dr Donald P. Weeks (University of Nebraska, Lincoln). Media and growth conditions for C. reinhardtii strains have been previously described (Wang and Spalding, 2006). All strains were maintained on CO 2 minimal plates and grown photoautotrophically at 50 ~ 80 µe m -2 s -1 with CO 2 enrichment (5% CO 2 in air). Liquid cultures were grown on a gyratory shaker under aeration in white light (~ 100 µe m -2 s -1 ). For biochemical or physiological analysis, cells were shifted from high to limiting CO 2 (L- CO 2, ppm or VL-CO 2, ppm) conditions for various hours. Cultures were in the log phase of growth when CO 2 enrichment was removed. VL-CO 2 was obtained by mixing normal air with compressed, CO 2 -free air. Generation and Isolation of Suppressors and Genetic Analysis Air-dier mutant pmp1 and ad1 was transformed with linearized plasmid psi103 (Sizova
85 82 et al., 2001) by the glass bead method and transformed cells were kept in ambient CO 2 and selected on minimal medium plates supplemented with 15 µg/ml paromomycin. Phenotypes of surviving colonies selected as putative suppressors were verified by serially diluted spot tests. Genetic crosses and random progeny analyses were performed as described by Harris (1989). For diploid analysis, chloroplast markers were first introduced into suppressors and pmp1 by crossing them with CC118 (an A C change at nucleotide 474 in the chloroplast 16S rrna confers streptomycin resistance) and CC86 (kanamycin resistance caused by mutation at the 3 end of chloroplast 16S rrna), respectively. Heterozygous vegetative diploids were then generated by crossing the streptomycin resistant pmp/ad-suppressors with kanamycin resistant pmp1 progeny and selected by their resistance to both antibiotics. In each cross, 10 to 20 diploids were collected and the presence of both mt+ and mt- loci were confirmed by a PCR-based approach (Zamora et al., 2004). Nucleic Acid Extractions and Analysis Genomic DNA was isolated and purified as described previously (Van and Spalding, 1999). Southern analyses were performed by standard procedures. Total cellular RNA was isolated by the acid guanidinium thiocyanate method (Chomczynski and Sacchi, 2006). RNA (15~20µg) samples were separated in 1% formaldehyde containing agarose gels and blotted onto Hybond nylon transfer membranes (Osmonics Inc.). DNA probes were radiolabeled by random priming using [α- 32 P]dCTP (Promega, WI) and hybridizations were performed at 65 0 C. Each northern blot was analyzed by phosphorimager scanning (BioRad, CA). Isolation of Sequences Flanking the Plasmid Insert in ad-su8 by Inverse PCR Based on information from Southern blot analysis, PvuII was used to digest the genomic DNA isolated from ad-suy to produce a fragment with a size ~3.4 kb, including part of the inserted psi103 vector and its 3 flanking genomic DNA. The PvuII-digested ad-su8 genomic DNA (1 µg) was circularized with 1 unit of T4 DNA ligase (Invitrogen), precipitated, and the circularized product was used as a template for inverse PCR by using
86 83 standard PCR procedures. Two pairs of primers were designed, with each pair complementing the psi103 sequence in opposite orientations. Both primer pairs produced PCR products with the correct predicted sizes, and amplified DNA from one primer pair (5 - GCCCTCATAGCCCGCCAAATC -3 and 5 - GTTGGTTGTAACGATCCTCCGTTG -3 ) was sequenced to determine the sequences flanking the 3 of plasmid insert. A similar approach was used to identify the genomic DNA flanking the 5 of insert by circularizing the ~4.5 kb SacI-digested ad-su8 genomic DNA. Amplified DNA from primer pair (5 - GCACCCAACTGATCTTCAGCATC -3 and 5 - GCGAGTCAGTGAGCGAGGA-3 ) was sequenced to determine the flanking genomic region. Rough Mapping of pmp-su5 An interfertile strain known as Chlamydomonas grossii, S1-D2 or its culture collection designation of CC-2290, was transformed with psi103 plasmid to be resistant to the antibiotic paromomycin. Suppressor pmp-su5 was backcrossed to a wild-type laboratory strain to ensure that the suppressor phenotype was caused by single, nuclear mutation and the su5 mutation is not linked with the plasmid insert. A paromomycin sensitive pmp-su5 progeny was chosen to cross to S1-D2 and several modified S1-D2 derivatives. Out of 6 different crosses, approximately 120 paromomycin resistant, air-dying meiotic progeny were selected as a mapping population, so these progeny should carry the lcib point mutation and the S1-D2 wild-type SU5 allele. A PCR-based rough mapping was performed to analyze 53 STS markers which cover the entire chromosomes at 20- to 30-cM resolution. The marker kit is available via the stock center and a complete set of amplification instructions and actual results can be found at the Chlamydomonas genome web site ( Photosynthetic O 2 Exchange and Ci Uptake Measurements H-CO 2, L-CO 2 or VL-CO 2 -acclimated cells (24 hours induction) were collected and resuspended in 25 mm MOPS-KOH buffer to a final chlorophyll concentration of 25 µg/ml for analysis of photosynthesis and Ci uptake characteristics. Photosynthetic O 2 evolution
87 84 rates were measured at 25 C with a Clark-type oxygen electrode (Rank Brothers, Cambridge, UK) as previously described (Duanmu et al., 2009). Oxygen evolution rates were recorded as V20 or V50 when 20 µm or 50 µm NaHCO 3 were used, respectively, and the maximum O 2 evolution rate, V4000, was obtained by using 4000 µm NaHCO 3. Relative affinities for Ci were calculated as the ratios P20 = V20/V4000 and P50 = V50/V4000. Ci uptake by L-CO 2 -adapted C. reinhardtii cells at 50 µm total Ci was measured by the silicone oil filtration technique (Badger et al., 1980). The cell volume was estimated by using 14 C-sorbitol and 3 H 2 O as previously described (Wirtz et al., 1980). The intracellular Ci concentration was calculated by using cell volume and the acid labile 14 C data. Acknowledgments This work was supported by the U. S. Department of Agriculture National Research Initiative (grant no to M.H.S.), as well as by the College of Agriculture and Life Sciences and the College of Liberal Arts and Sciences at Iowa State University. References Amoroso G, Sültemeyer DF, Thyssen C, Fock HP (1998) Uptake of HCO 3 - and CO 2 in cells and chloroplasts from the microalgae Chlamydomonas reinhardtii and Dunaliella tertiolecta. Plant Physiol 116: Badger MR, Kaplan A, Berry JA (1980) Internal inorganic carbon pool of Chlamydomonas reinhardtii: evidence for a carbon dioxide concentrating mechanism. Plant Physiol 66: Chomczynski P, Sacchi N (2006) The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: twenty-something years on. Nat Protol 1(2): Colombo SL, Pollock SV, Eger KA, Godfrey AC, Adams JE, Mason CB, Moroney JV (2002) Use of the bleomycin resistance gene to generate tagged insertional mutants of Chlamydomonas reinhardtii that require elevated CO 2 for optimal growth. Funct Plant Biol 29: Duanmu D, Wang Y, Spalding MH (2009) Thylakoid Lumen Carbonic Anhydrase (CAH3) Mutation Suppresses Air-Dier Phenotype of LCIB Mutant in Chlamydomonas reinhardtii. Plant Physiol 149: Fukuzawa H, Miura K, Ishizaki K, Kucho KI, Saito T, Kohinata T, Ohyama K (2001) Ccm1, a regulatory gene controlling the induction of a carbon-concentrating mechanism in Chlamydomonas reinhardtii by sensing CO 2 availability. Proc Natl Acad Sci USA 98(9): Funke RP, Kovar JL, Weeks DP (1997) Intracellular carbonic anhydrase is essential to
88 85 photosynthesis in Chlamydomonas reinhardtii at atmospheric levels of CO 2. Plant Physiol 114: Giordano M, Beardall J, Raven JA (2005) CO 2 concentrating mechanisms in algae: mechanisms, environmental modulatioin, and evolution. Annu Rev Plant Biol 56: Harris EH (1989) The Chlamydomonas Source Book: a comprehensive guide to biology and laboratory use. Academic Press Inc, San Diego, pp Karlsson J, Clarke AK, Chen ZY, Hugghins SY, Youn-II P, Husic HD, Moroney JV, Samuelsson G (1998) A novel α-type carbonic anhydrase associated with the thylakoid membrane in Chlamydomonas reinhardtii is required for growth at ambient CO 2. EMBO J 17(5): Kindle KL (1990) High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 87(3): Long SP, Ainsworth EA, Rogers A, Ort DR (2004) Rising Atmospheric Carbon Dioxide: Plants Face the Future. Annu Rev Plant Biol 55: Mariscal V, Moulin P, Orsel M, Miller AJ, Fernández E, Galván A (2006) Differential regulation of the Chlamydomonas Nar1 gene family by carbon and nitrogen. Protist 157(4): Miura K, Yamano T, Yoshioka S, Kohinata T, Inoue Y, Taniguchi F, Asamizu E, Nakaura Y, Tabata S, Yamato KT, Ohyama K, Fukuzawa H (2004) Expression profiling-based identification of CO 2 -responsive genes regulated by CCM1 controlling a carbon-concentrating mechanism in Chlamydomonas reinhardtii. Plant Physiol 135: Moroney JV, Tolbert NE, Sears BB. (1986) Complementation analysis of the inorganic carbon concentrating mechanism of Chlamydomonas reinhardtii. Mol Gen Genet 204: Moroney JV, Ynalvez RA (2007) Proposed carbon dioxide concentrating mechanism in Chlamydomonas reinhardtii. Eukaryot Cell 6(8): Palmqvist K, Sjoberg S, Samuelsson G (1988) Induction of inorganic carbon accumulation in the unicellular green algae Scenedesmus obliquus and Chlamydomonas reinhardtii. Plant Physiol 87(2): Palmqvist K, Yu J-W, Badger MR (1994) Carbonic anhydrase activity and inorganic carbon fluxes in low and high Ci of Chlamydomonas reinhardtii and Scenedesmus obliquus. Physiol Plant 90: Pollock SV, Prout DL, Godfrey AC, Lemaire SD, Moroney JV (2004) The Chlamydomonas reinhardtii proteins Ccp1 and Ccp2 are required for long-term growth, but are not necessary for efficient photosynthesis, in a low-co 2 environment. Plant Mol Biol 56: Price GD, Maeda SI, Omata T, Badger MR (2002) Modes of active inorganic carbon uptake in the cyanobacterium, Synechococcus sp. PCC7942. Funct Plant Biol 29: Price GD, Woodger FJ, Badger MR, Howitt SM, Tucker L (2004) Identification of a SulP-type bicarbonate transporter in marine cyanobacteria. Proc Natl Acad Sci USA 101: Sizova I, Fuhrmann M, Hegemann P (2001) A Streptomyces rimosus aphviii gene encoding for a new type phosphotransferase provides stable antibiotic resistance to Chlamydomonas reinhardtii. Gene 277:
89 86 Spalding MH, Spreitzer RJ, Ogren WL (1983a) Carbonic anhydrase-deficient mutant of Chlamydomonas reinhardtii requires elevated carbon dioxide concentration for photoautotrophic growth. Plant Physiol 73: Spalding MH, Spreitzer RJ, Ogren WL (1983b) Reduced inorganic carbon transport in a CO 2 -requiring mutant of Chlamydomonas reinhardtii. Plant Physiol 73: Spalding MH, Van K, Wang Y, Nakamura Y (2002) Acclimation of Chlamydomonas to changing carbon availability. Funct Plant Biol 29: Spalding MH (2008) Microalgal carbon-dioxide-concentrating mechanisms: Chlamydomonas inorganic carbon transporters. J Exp Bot 59(7): Sültemeyer DF, Klöck G, Kreuzberg K, Fock HP (1988) Photosynthesis and apparent affinity for dissolved inorganic carbon by cells and chloroplasts of Chlamydomonas reinhardtii grown at high and low CO 2 concentrations. Planta 176: Sültemeyer DF, Miller AG, Espie GS, Fock HP, Canvin DT (1989) Active CO 2 transport by the green alga Chlamydomonas reinhardtii. Plant Physiol 89: Sültemeyer DF, Fock HP, Canvin DT (1991) Active uptake of inorganic carbon by Chlamydomonas reinhardtii: evidence for simultaneous transport of HCO 3 - and CO 2 and characterization of active CO 2 transport. Can J Bot 69: Suzuki K. (1995) Phosphoglycolate phosphatase-deficient mutants of Chlamydomonas reinhardtii capable of growth under air. Plant Cell Physiol 36: Thyssen C, Hermes M, Sültemeyer D (2003) Isolation and characterization of Chlamydomonas reinhardtii mutants with an impaired CO 2 -concentrating mechanism. Planta 217: Vance P, Spalding MH (2005) Growth, photosynthesis and gene expression in Chlamydomonas over a range of CO 2 concentrations and CO 2 /O 2 ratios: CO 2 regulates multiple acclimation states. Can J Bot 83: Van K, Wang Y, Nakamura Y, Spalding MH (2001) Insertional mutants of Chlamydomonas reinhardtii that require elevated CO 2 for survival. Plant Physiol 127: Van K, Spalding MH (1999) Periplasmic carbonic anhydrase structural gene (Cah1) mutant in Chlamydomonas reinhardtii. Plant Physiol 120: Wang Y, Spalding MH (2006) An inorganic carbon transport system responsible for acclimation specific to air levels of CO 2 in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 103: Wang Y, Spalding MH (2007) CO 2 Concentrating Mechanisms in Eukaryotic Microalgae. Funct Plant Sci Biotechnol 1(1): Wirtz W, Stitt M, Heldt HW (1980) Enzymic determination of metabolites in the subcellular compartments of Spinach protoplasts. Plant Physiol 66(1): Xiang Y, Zhang J, Weeks DP (2001) The cia5 gene controls formation of the carbon concentrating mechanism in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 98: Yoshiko N, Saradadevi K, Kyujung V, Wei H, Spalding MH. (2005) Disruption of the glycolate dehydrogenase gene in the high-co 2 -requiring mutant HCR89 of Chlamydomonas reinhardtii. Can J Bot 83: Zamora I, Feldman JL, Marshall WF (2004) PCR-based assay for mating type and diploidy in Chlamydomonas. BioTechniques 37:
90 87 Figure Legends Figure 1. Southern blot analysis of suppressors probed with a 880-bp PCR fragment of the Par R gene. (A) Map of the Par R portion of the psi103 plasmid used for insertional mutagenesis. Arrows above and below the map indicate the primers used to make the probe. (B) Southern analysis of four suppressors. Genomic DNA was digested with the indicated restriction enzymes. Figure 2. Growth of ad- and pmp- suppressors on minimal plates in H-CO 2, L-CO 2 and VL- CO 2 chambers. Cells grown to logarithmic phase were diluted to the indicated numbers ( 10 3 ) per 5 µl, spotted on plates and incubated for 9 days under dim lights. Figure 3. (A) Northern blot analysis of suppressors. H-CO 2 grown cells (0h) were acclimated to L-CO 2 for 4h, 14h respectively and 15 µg of total RNA was used. (B) Total RNA from L-CO 2 acclimated wt-su1, ad-su1 was separated and hybridized to HLA3- and LCI1-coding sequences. Same amount of RNA (~15 µg) from ad1-1 and CC620 was also included for Northern blot.
91 88 Table 1. C. reinhardtii strains used in the study Strains Genotype Description Reference CC86 nr-u-2-1 mt + Chloroplast mutation resistant to kanamycin Harris (1989) CC118 sr-u-2-60 mt + Chloroplast mutation resistant to streptomycin Harris (1989) CC620 mt + Wild type (137c) Harris (1989) CC1860 pmp1-1 mt - Deficient in Ci transport, no growth in air Van et al. (2001) Generated by CC1860 CC124 pmp-su1 mt - pmp1-1 suppressor This report pmp-su4 mt - pmp1-1 suppressor This report pmp-su5 mt - pmp1-1 suppressor This report CC2702 cia-5 mt + No acclimation to limiting CO 2 Moroney et al. (1989) CC4154 ad1 mt - air dier phenotype, deficient in Ci transport Wang and Spalding (2006) ad1-1 mt - Walled progeny of ad1 mutant, generated by CC4154 CC620 Wang and Spalding (2006) ad-su8 mt - ad1 suppressor This report Table 2. Characteristics of suppressors Mutants Growth in L-CO 2 Par R Insert Cosegregation with Par R Insert Diploid Analysis pmp1/ad1 No NA a NA Recessive pmp-su1 Yes 1 No Recessive pmp-su4 Yes 1 No Dominant pmp-su5 Yes 1 No Dominant ad-su8 Yes 1 Yes Recessive a NA, Not applicable.
92 89 Table 3. Photosynthetic Ci affinity of wild type CC620, pmp1, ad1-1, cia5, and suppressor strains acclimated to H-CO 2, L-CO 2 or VL-CO 2 for 1 d. Relative affinity were ratios of the O 2 evolution rates (µmol mg Chl -1 h -1 ) at 20 or 50 µm Ci over the maximal rate at 4000 µm Ci. H-CO 2 L-CO 2 VL-CO 2 Strains P20 P50 P20 P50 P20 P50 CC ± ± ± ± ± ±0.09 pmp1 0.04± ± ± ± ± ±0.03 ad ± ± ± ± ± ±0.04 pmp-su1 0.06± ± ± ± ± ±0.03 pmp-su5 0.06± ± ± ± ± ±0.02 ad-su8 0.07± ± ± ± ± ±0.05 wt-su1 0.28± ± ± ± ± ±0.02 wt-su5 0.22± ± ± ± ± ±0.08 wt-su8 0.31± ± ± ± ± ±0.03 cia5 0.04± ± ± ± ± ±0.03 Table 4. Intracellular Ci accumulation in cells acclimated to L-CO 2 for 1 d. Internal Ci accumulation (mm Ci after 80 s) was determined by silicone oil centrifugation in ph 7.3 (MOPS-KOH) buffer at initial external Ci concentration of 50 µm [ 14 C]NaHCO 3 and a Chl concentration of 25 µg/ml. Strains mm internal Ci CC ±0.35 pmp1 0.21±0.10 ad ±0.16 pmp-su1 1.48±0.44 pmp-su5 1.65±0.30 ad-su8* 2.43±0.42 cia ±0.13 *a walled progeny of ad-su8.

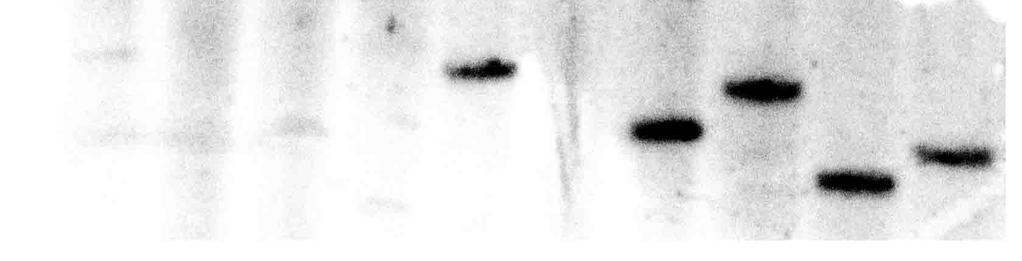
93 Figure 1. 90

94 Figure 2. 91



95 Figure 3. 92
The Role of Inorganic Carbon Transport and Accumulation in the CO 2 -Concentrating Mechanism and CO 2 Assimilation in Chlamydomonas
 The Role of Inorganic Carbon Transport and Accumulation in the CO 2 -Concentrating Mechanism and CO 2 Assimilation in Chlamydomonas Is there a Role for the CCM in Increasing Biological CO 2 Capture? Generalized
The Role of Inorganic Carbon Transport and Accumulation in the CO 2 -Concentrating Mechanism and CO 2 Assimilation in Chlamydomonas Is there a Role for the CCM in Increasing Biological CO 2 Capture? Generalized
CO 2 Concentrating Mechanisms in Eukaryotic Microalgae
 Functional Plant Science and Biotechnology 2007 Global Science Books CO 2 Concentrating Mechanisms in Eukaryotic Microalgae Yingjun Wang Martin H. Spalding * Department of Genetics, Development and Cell
Functional Plant Science and Biotechnology 2007 Global Science Books CO 2 Concentrating Mechanisms in Eukaryotic Microalgae Yingjun Wang Martin H. Spalding * Department of Genetics, Development and Cell
Knockdown of limiting-co 2 induced gene HLA3 decreases HCO 3 transport and photosynthetic Ci affinity in Chlamydomonas reinhardtii
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Biochemistry -- Faculty Publications Biochemistry, Department of 4-7-2009 Knockdown of limiting- induced gene HLA3 decreases
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Biochemistry -- Faculty Publications Biochemistry, Department of 4-7-2009 Knockdown of limiting- induced gene HLA3 decreases
MINIREVIEWS. Proposed Carbon Dioxide Concentrating Mechanism in Chlamydomonas reinhardtii. James V. Moroney* and Ruby A. Ynalvez
 EUKARYOTIC CELL, Aug. 2007, p. 1251 1259 Vol. 6, No. 8 1535-9778/07/$08.00 0 doi:10.1128/ec.00064-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. MINIREVIEWS Proposed Carbon
EUKARYOTIC CELL, Aug. 2007, p. 1251 1259 Vol. 6, No. 8 1535-9778/07/$08.00 0 doi:10.1128/ec.00064-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. MINIREVIEWS Proposed Carbon
Lecture 9: Photosynthesis
 Lecture 9: Photosynthesis I. Characteristics of Light A. Light is composed of particles that travel as waves 1. Comprises a small part of the electromagnetic spectrum B. Radiation varies in wavelength
Lecture 9: Photosynthesis I. Characteristics of Light A. Light is composed of particles that travel as waves 1. Comprises a small part of the electromagnetic spectrum B. Radiation varies in wavelength
RuBP has 5 carbons and is regenerated in the Calvin cycle. In the Calvin cycle, carbon is conserved, ATP is used and NADPH is used.
 Carbon Reactions: CO 2 is fixed by Rubisco located in the stroma. The molecule that is carboxylated is RuBP. RuBP has 5 carbons and is regenerated in the Calvin cycle. In the Calvin cycle, carbon is conserved,
Carbon Reactions: CO 2 is fixed by Rubisco located in the stroma. The molecule that is carboxylated is RuBP. RuBP has 5 carbons and is regenerated in the Calvin cycle. In the Calvin cycle, carbon is conserved,
Photosynthesis. Chapter 8
 Photosynthesis Chapter 8 Photosynthesis Overview Energy for all life on Earth ultimately comes from photosynthesis 6CO 2 + 12H 2 O C 6 H 12 O 6 + 6H 2 O + 6O 2 Oxygenic photosynthesis is carried out by
Photosynthesis Chapter 8 Photosynthesis Overview Energy for all life on Earth ultimately comes from photosynthesis 6CO 2 + 12H 2 O C 6 H 12 O 6 + 6H 2 O + 6O 2 Oxygenic photosynthesis is carried out by
cytosol stroma Photorespiration: Ribulose bisphosphate carboxylase/oxygenase (Rubisco) Ribulose bisphosphate carboxylase/oxygenase (Rubisco)
 Ribulose bisphosphate carboxylase/oxygenase (Rubisco)") Carbon Reactions: CO 2 is fixed by Rubisco located in the stroma. The molecule that is carboxylated is RuBP. RuBP has 5 carbons and is regenerated in the Calvin cycle. In the Calvin cycle, carbon is conserved,
Carbon Reactions: CO 2 is fixed by Rubisco located in the stroma. The molecule that is carboxylated is RuBP. RuBP has 5 carbons and is regenerated in the Calvin cycle. In the Calvin cycle, carbon is conserved,
Chapter 5: Photosynthesis: The Energy of Life pg : Alternative Mechanisms of Carbon Fixation pg
 UNIT 2: Metabolic Processes Chapter 5: Photosynthesis: The Energy of Life pg. 210-240 5.4: Alternative Mechanisms of Carbon Fixation pg. 231 234 Photosynthesis requires reactants; CO 2 and H 2 O, to produce
UNIT 2: Metabolic Processes Chapter 5: Photosynthesis: The Energy of Life pg. 210-240 5.4: Alternative Mechanisms of Carbon Fixation pg. 231 234 Photosynthesis requires reactants; CO 2 and H 2 O, to produce
C-4 PATHWAY USMAN SUMO FRIEND TAMBUNAN ARLI ADITYA PARIKESIT EVI KRISTIN WULANDARI
 C-4 PATHWAY USMAN SUMO FRIEND TAMBUNAN ARLI ADITYA PARIKESIT EVI KRISTIN WULANDARI BIOINFORMATICS GROUP DEPARTMENT OF CHEMISTRY FACULTY OF MATHEMATICS AND SCIENCE UNIVERSITY OF INDONESIA C-4 Plants The
C-4 PATHWAY USMAN SUMO FRIEND TAMBUNAN ARLI ADITYA PARIKESIT EVI KRISTIN WULANDARI BIOINFORMATICS GROUP DEPARTMENT OF CHEMISTRY FACULTY OF MATHEMATICS AND SCIENCE UNIVERSITY OF INDONESIA C-4 Plants The
8.1 Photosynthesis and Energy
 BIOL 100 Ch. 8 1 8.1 Photosynthesis and Energy Photosynthesis and Energy Photosynthesis Making food from light energy Photoautotrophs Use CO2 and water to make sugars Made life possible as we know it Provides
BIOL 100 Ch. 8 1 8.1 Photosynthesis and Energy Photosynthesis and Energy Photosynthesis Making food from light energy Photoautotrophs Use CO2 and water to make sugars Made life possible as we know it Provides
Photosynthesis Definition and Superficial Overview
 Photosynthesis Photosynthesis Definition and Superficial Overview Photosynthesis is the process used by plants to convert light energy from the sun into chemical energy that can be later released to fuel
Photosynthesis Photosynthesis Definition and Superficial Overview Photosynthesis is the process used by plants to convert light energy from the sun into chemical energy that can be later released to fuel
9/8/2010. Chapter 4. Structures Internal to the Cell Wall. The Plasma Membrane. Functional Anatomy of Prokaryotic and Eukaryotic Cells
 Chapter 4 Functional Anatomy of Prokaryotic and Eukaryotic Cells Johana Meléndez Part II slides 39-87 Lectures prepared by Christine L. Case Structures Internal to the Cell Wall Learning Objectives 4-8
Chapter 4 Functional Anatomy of Prokaryotic and Eukaryotic Cells Johana Meléndez Part II slides 39-87 Lectures prepared by Christine L. Case Structures Internal to the Cell Wall Learning Objectives 4-8
Chapter 10: PHOTOSYNTHESIS
 Chapter 10: PHOTOSYNTHESIS 1. Overview of Photosynthesis 2. Light Absorption 3. The Light Reactions 4. The Calvin Cycle 1. Overview of Photosynthesis Chapter Reading pp. 185-190, 206-207 What is Photosynthesis?
Chapter 10: PHOTOSYNTHESIS 1. Overview of Photosynthesis 2. Light Absorption 3. The Light Reactions 4. The Calvin Cycle 1. Overview of Photosynthesis Chapter Reading pp. 185-190, 206-207 What is Photosynthesis?
Chapter 10 Photosynthesis
 Chapter 10 Photosynthesis Overview: The Process That Feeds the Biosphere Photosynthesis is the process that converts solar energy into chemical energy Photosynthesis occurs in plants, algae, certain other
Chapter 10 Photosynthesis Overview: The Process That Feeds the Biosphere Photosynthesis is the process that converts solar energy into chemical energy Photosynthesis occurs in plants, algae, certain other
2015 AP Biology PRETEST Unit 3: Cellular Energetics Week of October
 Name: Class: _ Date: _ 2015 AP Biology PRETEST Unit 3: Cellular Energetics Week of 19-23 October Multiple Choice Identify the choice that best completes the statement or answers the question. 1) Which
Name: Class: _ Date: _ 2015 AP Biology PRETEST Unit 3: Cellular Energetics Week of 19-23 October Multiple Choice Identify the choice that best completes the statement or answers the question. 1) Which
Questions for Biology IIB (SS 2006) Wilhelm Gruissem
 Wilhelm Gruissem") Questions for Biology IIB (SS 2006) Plant biology Wilhelm Gruissem The questions for my part of Biology IIB, Plant Biology, are provided for self-study and as material for the exam. Please note that the
Questions for Biology IIB (SS 2006) Plant biology Wilhelm Gruissem The questions for my part of Biology IIB, Plant Biology, are provided for self-study and as material for the exam. Please note that the
Vital metabolism for survival of life in the earth. Prof Adinpunya Mitra Agricultural & Food Engineering Department
 Vital metabolism for survival of life in the earth Prof Adinpunya Mitra Agricultural & Food Engineering Department THE SUN: MAIN SOURCE OF ENERGY FOR LIFE ON EARTH THE BASICS OF PHOTOSYNTHESIS Almost all
Vital metabolism for survival of life in the earth Prof Adinpunya Mitra Agricultural & Food Engineering Department THE SUN: MAIN SOURCE OF ENERGY FOR LIFE ON EARTH THE BASICS OF PHOTOSYNTHESIS Almost all
Big Idea 2: Biological systems utilize free energy and molecular building blocks to grow, to reproduce and to maintain dynamic homeostasis.
 Big Idea 2: Biological systems utilize free energy and molecular building blocks to grow, to reproduce and to maintain dynamic homeostasis. Enduring understanding 2.B: Growth, reproduction and dynamic
Big Idea 2: Biological systems utilize free energy and molecular building blocks to grow, to reproduce and to maintain dynamic homeostasis. Enduring understanding 2.B: Growth, reproduction and dynamic
PHOTOSYNTHESIS Student Packet SUMMARY
 PHOTOSYNTHESIS Student Packet SUMMARY LIVING SYSTEMS REQUIRE ENERGY AND MATTER TO MAINTAIN ORDER, GROW AND REPRODUCE Energy input must exceed energy lost and used by an organism. Organisms use various
PHOTOSYNTHESIS Student Packet SUMMARY LIVING SYSTEMS REQUIRE ENERGY AND MATTER TO MAINTAIN ORDER, GROW AND REPRODUCE Energy input must exceed energy lost and used by an organism. Organisms use various
Life on Earth is solar powered. Photosynthesis => conversion of light energy to chemical energy (stored in sugars and other organic molecules).
.") Photosynthesis Life on Earth is solar powered. Photosynthesis => conversion of light energy to chemical energy (stored in sugars and other organic molecules). Organisms obtain organic compounds by one
Photosynthesis Life on Earth is solar powered. Photosynthesis => conversion of light energy to chemical energy (stored in sugars and other organic molecules). Organisms obtain organic compounds by one
GACE Biology Assessment Test I (026) Curriculum Crosswalk
 Curriculum Crosswalk") Subarea I. Cell Biology: Cell Structure and Function (50%) Objective 1: Understands the basic biochemistry and metabolism of living organisms A. Understands the chemical structures and properties of biologically
Subarea I. Cell Biology: Cell Structure and Function (50%) Objective 1: Understands the basic biochemistry and metabolism of living organisms A. Understands the chemical structures and properties of biologically
TALE activation and functional analysis of endogenous genes in Chlamydomonas reinhardtii
 Graduate Theses and Dissertations Iowa State University Capstones, Theses and Dissertations 2014 TALE activation and functional analysis of endogenous genes in Chlamydomonas reinhardtii Han Gao Iowa State
Graduate Theses and Dissertations Iowa State University Capstones, Theses and Dissertations 2014 TALE activation and functional analysis of endogenous genes in Chlamydomonas reinhardtii Han Gao Iowa State
Energy Conversions. Photosynthesis. Plants. Chloroplasts. Plant Pigments 10/13/2014. Chapter 10 Pg
 Energy Conversions Photosynthesis Chapter 10 Pg. 184 205 Life on Earth is solar-powered by autotrophs Autotrophs make their own food and have no need to consume other organisms. They are the ultimate source
Energy Conversions Photosynthesis Chapter 10 Pg. 184 205 Life on Earth is solar-powered by autotrophs Autotrophs make their own food and have no need to consume other organisms. They are the ultimate source
Chapter 8 PHOTOSYNTHESIS Chapter # Chapter Title PowerPoint Image Slideshow
 COLLEGE BIOLOGY PHYSICS Chapter 8 PHOTOSYNTHESIS Chapter # Chapter Title PowerPoint Image Slideshow Figure 8.0 Photosynthesis Figure 8.1 Earth s distribution of photosynthesis as seen via chlorophyll a
COLLEGE BIOLOGY PHYSICS Chapter 8 PHOTOSYNTHESIS Chapter # Chapter Title PowerPoint Image Slideshow Figure 8.0 Photosynthesis Figure 8.1 Earth s distribution of photosynthesis as seen via chlorophyll a
Photosynthesis. Excitation of chlorophyll in a chloroplast
 Photosynthesis The process of photosynthesis begins with light-absorbing pigments in plant cells. A pigment molecule is able to absorb the energy from light only within a narrow range of wavelengths. In
Photosynthesis The process of photosynthesis begins with light-absorbing pigments in plant cells. A pigment molecule is able to absorb the energy from light only within a narrow range of wavelengths. In
1 P a g e h t t p s : / / w w w. c i e n o t e s. c o m / Photosynthesis (chapter 12):
:") 1 P a g e h t t p s : / / w w w. c i e n o t e s. c o m / Photosynthesis (chapter 12): Photosynthesis is the fixation of CO 2 and its subsequent reduction to carbohydrate, using hydrogen from water, taking
1 P a g e h t t p s : / / w w w. c i e n o t e s. c o m / Photosynthesis (chapter 12): Photosynthesis is the fixation of CO 2 and its subsequent reduction to carbohydrate, using hydrogen from water, taking
Photosynthesis (Chapter 7 Outline) A. For life based on organic compounds, two questions can be raised:
 A. For life based on organic compounds, two questions can be raised:") Photosynthesis (Chapter 7 Outline) Sun, Rain, and Survival A. For life based on organic compounds, two questions can be raised: 1. Where does the carbon come from? 2. Where does the energy come from to
Photosynthesis (Chapter 7 Outline) Sun, Rain, and Survival A. For life based on organic compounds, two questions can be raised: 1. Where does the carbon come from? 2. Where does the energy come from to
REVIEW 3: METABOLISM UNIT RESPIRATION & PHOTOSYNTHESIS. A. Top 10 If you learned anything from this unit, you should have learned:
 Period Date REVIEW 3: METABOLISM UNIT RESPIRATION & PHOTOSYNTHESIS A. Top 10 If you learned anything from this unit, you should have learned: 1. Energy production through chemiosmosis a. pumping of H+
Period Date REVIEW 3: METABOLISM UNIT RESPIRATION & PHOTOSYNTHESIS A. Top 10 If you learned anything from this unit, you should have learned: 1. Energy production through chemiosmosis a. pumping of H+
Photosynthesis: Carbon Reactions. Dr. Obaidur Rahman
 Photosynthesis: Carbon Reactions Dr. Obaidur Rahman Topics: Part I THE CALVIN CYCLE The Calvin Cycle Has Three Stages: Carboxylation, Reduction, and Regeneration The Carboxylation of Ribulose Bisphosphate
Photosynthesis: Carbon Reactions Dr. Obaidur Rahman Topics: Part I THE CALVIN CYCLE The Calvin Cycle Has Three Stages: Carboxylation, Reduction, and Regeneration The Carboxylation of Ribulose Bisphosphate
Lecture Series 13 Photosynthesis: Energy from the Sun
 Lecture Series 13 Photosynthesis: Energy from the Sun Photosynthesis: Energy from the Sun A. Identifying Photosynthetic Reactants and Products B. The Two Pathways of Photosynthesis: An Overview C. Properties
Lecture Series 13 Photosynthesis: Energy from the Sun Photosynthesis: Energy from the Sun A. Identifying Photosynthetic Reactants and Products B. The Two Pathways of Photosynthesis: An Overview C. Properties
Chapter 5: Photosynthesis: The Energy of Life pg : Pathways of Photosynthesis pg
 UNIT 2: Metabolic Processes Chapter 5: Photosynthesis: The Energy of Life pg. 210-240 5.2: Pathways of Photosynthesis pg. 220-228 Light Dependent Reactions Photosystem II and I are the two light capturing
UNIT 2: Metabolic Processes Chapter 5: Photosynthesis: The Energy of Life pg. 210-240 5.2: Pathways of Photosynthesis pg. 220-228 Light Dependent Reactions Photosystem II and I are the two light capturing
Oxidative Phosphorylation versus. Photophosphorylation
 Photosynthesis Oxidative Phosphorylation versus Photophosphorylation Oxidative Phosphorylation Electrons from the reduced cofactors NADH and FADH 2 are passed to proteins in the respiratory chain. In eukaryotes,
Photosynthesis Oxidative Phosphorylation versus Photophosphorylation Oxidative Phosphorylation Electrons from the reduced cofactors NADH and FADH 2 are passed to proteins in the respiratory chain. In eukaryotes,
9- #60 5. Photosynthesis. Sixth edition. D. O. Hall. and. K. K. Rao. Published in association with the Institute of Biology CAMBRIDGE UNIVERSITY PRESS
 9- #60 5 Photosynthesis Sixth edition D. O. Hall and K. K. Rao Published in association with the Institute of Biology CAMBRIDGE UNIVERSITY PRESS Contents General preface to the series Preface to the sixth
9- #60 5 Photosynthesis Sixth edition D. O. Hall and K. K. Rao Published in association with the Institute of Biology CAMBRIDGE UNIVERSITY PRESS Contents General preface to the series Preface to the sixth
THE BASICS OF PHOTOSYNTHESIS
 THE BASICS OF PHOTOSYNTHESIS Almost all plants are photosynthetic autotrophs, as are some bacteria and protists Autotrophs generate their own organic matter through photosynthesis Sunlight energy is transformed
THE BASICS OF PHOTOSYNTHESIS Almost all plants are photosynthetic autotrophs, as are some bacteria and protists Autotrophs generate their own organic matter through photosynthesis Sunlight energy is transformed
Metabolism 2 Photosynthesis
 Metabolism 2 Photosynthesis Light energy is trapped in the form of high energy electrons. High energy electrons are used to synthesize ATP and reduce CO 2 to form carbohydrates. Oxygen is produced as a
Metabolism 2 Photosynthesis Light energy is trapped in the form of high energy electrons. High energy electrons are used to synthesize ATP and reduce CO 2 to form carbohydrates. Oxygen is produced as a
Section A2: The Pathways of Photosynthesis
 CHAPTER 10 PHOTOSYNTHESIS Section A2: The Pathways of Photosynthesis 4. The Calvin cycle uses ATP and NADPH to convert CO2 to sugar: a closer look 5. Alternative mechanisms of carbon fixation have evolved
CHAPTER 10 PHOTOSYNTHESIS Section A2: The Pathways of Photosynthesis 4. The Calvin cycle uses ATP and NADPH to convert CO2 to sugar: a closer look 5. Alternative mechanisms of carbon fixation have evolved
Where does most of our society s energy come from (think of fossil fuels), how does that energy become fixed for human use?
, how does that energy become fixed for human use?") Where does most of our society s energy come from (think of fossil fuels), how does that energy become fixed for human use? The Photosynthesis equation 6 CO 2 + 12 H 2 O + Light energy C 6 H 12 O 6 +
Where does most of our society s energy come from (think of fossil fuels), how does that energy become fixed for human use? The Photosynthesis equation 6 CO 2 + 12 H 2 O + Light energy C 6 H 12 O 6 +
8.2 Photosynthesis Draw and label a diagram showing the structure of a chloroplast as seen in electron micrographs
 8.2 Photosynthesis 8.2.1 - Draw and label a diagram showing the structure of a chloroplast as seen in electron micrographs double membrane starch grain grana thylakoid internal membrane - location of the
8.2 Photosynthesis 8.2.1 - Draw and label a diagram showing the structure of a chloroplast as seen in electron micrographs double membrane starch grain grana thylakoid internal membrane - location of the
Photosynthesis in Higher Plants
 Photosynthesis in Higher Plants Very Short Answers Questions: 1. Name the processes which take place in the grana and stroma regions of chloroplasts? A: Grana Light reactions. Trapping light energy, synthesizing
Photosynthesis in Higher Plants Very Short Answers Questions: 1. Name the processes which take place in the grana and stroma regions of chloroplasts? A: Grana Light reactions. Trapping light energy, synthesizing
CHAPTER 13 : PHOTOSYNTHESIS IN HIGHER PLANTS K C MEENA PGT BIOLOGY KV VIKASPURI II SHIFT
 CHAPTER 13 : PHOTOSYNTHESIS IN HIGHER PLANTS K C MEENA PGT BIOLOGY KV VIKASPURI II SHIFT Photosynthesis is a Physic o chemical process, uses light energy to synthesis organic compounds (sugar). Importance
CHAPTER 13 : PHOTOSYNTHESIS IN HIGHER PLANTS K C MEENA PGT BIOLOGY KV VIKASPURI II SHIFT Photosynthesis is a Physic o chemical process, uses light energy to synthesis organic compounds (sugar). Importance
Photosynthesis Overview. Photosynthesis Overview. Photosynthesis Overview. Photosynthesis
 Photosynthesis Photosynthesis Overview Chapter 8 Energy for all life on Earth ultimately comes from photosynthesis. 6CO2 + 12H2O C6H12O6 + 6H2O + 6O2 Oxygenic photosynthesis is carried out by: cyanobacteria,
Photosynthesis Photosynthesis Overview Chapter 8 Energy for all life on Earth ultimately comes from photosynthesis. 6CO2 + 12H2O C6H12O6 + 6H2O + 6O2 Oxygenic photosynthesis is carried out by: cyanobacteria,
Importance of Protein sorting. A clue from plastid development
 Importance of Protein sorting Cell organization depend on sorting proteins to their right destination. Cell functions depend on sorting proteins to their right destination. Examples: A. Energy production
Importance of Protein sorting Cell organization depend on sorting proteins to their right destination. Cell functions depend on sorting proteins to their right destination. Examples: A. Energy production
THIS IS. In photosynthesis A) Carbon gets oxidized B) Carbon gets reduced C) Carbon gets metabolized D) Carbon gets digested
 Carbon gets oxidized B) Carbon gets reduced C) Carbon gets metabolized D) Carbon gets digested") THIS IS With Your Host... table Column A Column B Column C Column D Column E Column F 100 100 100 100 100 100 200 200 200 200 200 200 300 300 300 300 300 300 400 400 400 400 400 400 In photosynthesis A)
THIS IS With Your Host... table Column A Column B Column C Column D Column E Column F 100 100 100 100 100 100 200 200 200 200 200 200 300 300 300 300 300 300 400 400 400 400 400 400 In photosynthesis A)
METABOLISM. What is metabolism? Categories of metabolic reactions. Total of all chemical reactions occurring within the body
 METABOLISM What is metabolism? METABOLISM Total of all chemical reactions occurring within the body Categories of metabolic reactions Catabolic reactions Degradation pathways Anabolic reactions Synthesis
METABOLISM What is metabolism? METABOLISM Total of all chemical reactions occurring within the body Categories of metabolic reactions Catabolic reactions Degradation pathways Anabolic reactions Synthesis
Biochemistry: A Review and Introduction
 Biochemistry: A Review and Introduction CHAPTER 1 Chem 40/ Chem 35/ Fundamentals of 1 Outline: I. Essence of Biochemistry II. Essential Elements for Living Systems III. Classes of Organic Compounds IV.
Biochemistry: A Review and Introduction CHAPTER 1 Chem 40/ Chem 35/ Fundamentals of 1 Outline: I. Essence of Biochemistry II. Essential Elements for Living Systems III. Classes of Organic Compounds IV.
PHOTOSYNTHESIS. Light Reaction Calvin Cycle
 PHOTOSYNTHESIS Light Reaction Calvin Cycle Photosynthesis Purpose: use energy from light to convert inorganic compounds into organic fuels that have stored potential energy in their carbon bonds Carbon
PHOTOSYNTHESIS Light Reaction Calvin Cycle Photosynthesis Purpose: use energy from light to convert inorganic compounds into organic fuels that have stored potential energy in their carbon bonds Carbon
Biology: Life on Earth
 Teresa Audesirk Gerald Audesirk Bruce E. Byers Biology: Life on Earth Eighth Edition Lecture for Chapter 4 Cell Structure and Function Copyright 2008 Pearson Prentice Hall, Inc. Chapter 4 Outline 4.1 What
Teresa Audesirk Gerald Audesirk Bruce E. Byers Biology: Life on Earth Eighth Edition Lecture for Chapter 4 Cell Structure and Function Copyright 2008 Pearson Prentice Hall, Inc. Chapter 4 Outline 4.1 What
PLANT PHYSIOLOGY. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 PLANT PHYSIOLOGY Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 Carbon reactions of the photosynthesis Photosynthetic activity and the environmental factors Overview 1. Carbon
PLANT PHYSIOLOGY Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 Carbon reactions of the photosynthesis Photosynthetic activity and the environmental factors Overview 1. Carbon
Metabolism Review. A. Top 10
 A. Top 10 Metabolism Review 1. Energy production through chemiosmosis a. pumping of H+ ions onto one side of a membrane through protein pumps in an Electron Transport Chain (ETC) b. flow of H+ ions across
A. Top 10 Metabolism Review 1. Energy production through chemiosmosis a. pumping of H+ ions onto one side of a membrane through protein pumps in an Electron Transport Chain (ETC) b. flow of H+ ions across
WERE FE(II) OXIDIZING PHOTOAUTOTROPHS INVOLVED IN THE DEPOSITION OF PRECAMBRIAN BANDED IRON
 OXIDIZING PHOTOAUTOTROPHS INVOLVED IN THE DEPOSITION OF PRECAMBRIAN BANDED IRON") 19 1. Introduction WERE FE(II) OXIDIZING PHOTOAUTOTROPHS INVOLVED IN THE DEPOSITION OF PRECAMBRIAN BANDED IRON FORMATIONS? Banded Iron Formations (BIFs) are ancient sedimentary rocks characterized by laminations
19 1. Introduction WERE FE(II) OXIDIZING PHOTOAUTOTROPHS INVOLVED IN THE DEPOSITION OF PRECAMBRIAN BANDED IRON FORMATIONS? Banded Iron Formations (BIFs) are ancient sedimentary rocks characterized by laminations
NOTES: CH 10, part 3 Calvin Cycle (10.3) & Alternative Mechanisms of C-Fixation (10.4)
 & Alternative Mechanisms of C-Fixation (10.4)") NOTES: CH 10, part 3 Calvin Cycle (10.3) & Alternative Mechanisms of C-Fixation (10.4) 10.3 - The Calvin cycle uses ATP and NADPH to convert CO 2 to sugar The Calvin cycle, like the citric acid cycle,
NOTES: CH 10, part 3 Calvin Cycle (10.3) & Alternative Mechanisms of C-Fixation (10.4) 10.3 - The Calvin cycle uses ATP and NADPH to convert CO 2 to sugar The Calvin cycle, like the citric acid cycle,
Located in the thylakoid membranes. Chlorophyll have Mg + in the center. Chlorophyll pigments harvest energy (photons) by absorbing certain
 by absorbing certain") a review Located in the thylakoid membranes. Chlorophyll have Mg + in the center. Chlorophyll pigments harvest energy (photons) by absorbing certain wavelengths (blue-420 nm and red-660 nm are most important).
a review Located in the thylakoid membranes. Chlorophyll have Mg + in the center. Chlorophyll pigments harvest energy (photons) by absorbing certain wavelengths (blue-420 nm and red-660 nm are most important).
The summary equation of photosynthesis including the source and fate of the reactants and products. How leaf and chloroplast anatomy relates to
 1 The summary equation of photosynthesis including the source and fate of the reactants and products. How leaf and chloroplast anatomy relates to photosynthesis. How photosystems convert solar energy to
1 The summary equation of photosynthesis including the source and fate of the reactants and products. How leaf and chloroplast anatomy relates to photosynthesis. How photosystems convert solar energy to
Scenario: Mapping Photosynthesis
 Scenario Mapping Photosynthesis Scenario: Mapping Photosynthesis Photosynthesis is arguably the most important biological process on the Earth. All of the energy for the planet can be traced to the sun,
Scenario Mapping Photosynthesis Scenario: Mapping Photosynthesis Photosynthesis is arguably the most important biological process on the Earth. All of the energy for the planet can be traced to the sun,
Photosynthesis. Nearly all of the usable energy on this planet came, at one time or another, from the sun by the process of photosynthesis
 Photosynthesis Nearly all of the usable energy on this planet came, at one time or another, from the sun by the process of photosynthesis Photosynthesis 6CO 2 + 12H 2 O C 6 H 12 O 6 + 6O 2 + 6H 2 O Pigments
Photosynthesis Nearly all of the usable energy on this planet came, at one time or another, from the sun by the process of photosynthesis Photosynthesis 6CO 2 + 12H 2 O C 6 H 12 O 6 + 6O 2 + 6H 2 O Pigments
Using Light Energy to Make Organic Molecules
 OpenStax-CNX module: m44449 1 Using Light Energy to Make Organic Molecules OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the
OpenStax-CNX module: m44449 1 Using Light Energy to Make Organic Molecules OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the
Host-Pathogen Interaction. PN Sharma Department of Plant Pathology CSK HPKV, Palampur
 Host-Pathogen Interaction PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 PATHOGEN DEFENCE IN PLANTS A BIOLOGICAL AND MOLECULAR VIEW Two types of plant resistance response to potential
Host-Pathogen Interaction PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 PATHOGEN DEFENCE IN PLANTS A BIOLOGICAL AND MOLECULAR VIEW Two types of plant resistance response to potential
Photosynthesis is the main route by which that energy enters the biosphere of the Earth.
 Chapter 5-Photosynthesis Photosynthesis is the main route by which that energy enters the biosphere of the Earth. To sustain and power life on Earth, the captured energy has to be released and used in
Chapter 5-Photosynthesis Photosynthesis is the main route by which that energy enters the biosphere of the Earth. To sustain and power life on Earth, the captured energy has to be released and used in
Photosynthesis Thursday, July 7, 2011
 Photosynthesis Photosynthesis in Overview Process by which plants and other autotrophs store the energy of sunlight into sugars. Requires sunlight, water, and carbon dioxide. Overall equation: 6 CO 2
Photosynthesis Photosynthesis in Overview Process by which plants and other autotrophs store the energy of sunlight into sugars. Requires sunlight, water, and carbon dioxide. Overall equation: 6 CO 2
CHLOROPLASTS, CALVIN CYCLE, PHOTOSYNTHETIC ELECTRON TRANSFER AND PHOTOPHOSPHORYLATION (based on Chapter 19 and 20 of Stryer )
") CHLOROPLASTS, CALVIN CYCLE, PHOTOSYNTHETIC ELECTRON TRANSFER AND PHOTOPHOSPHORYLATION (based on Chapter 19 and 20 of Stryer ) Photosynthesis Photosynthesis Light driven transfer of electron across a membrane
CHLOROPLASTS, CALVIN CYCLE, PHOTOSYNTHETIC ELECTRON TRANSFER AND PHOTOPHOSPHORYLATION (based on Chapter 19 and 20 of Stryer ) Photosynthesis Photosynthesis Light driven transfer of electron across a membrane
Photosynthesis and Cellular Respiration
 Photosynthesis and Cellular Respiration Photosynthesis and Cellular Respiration All cellular activities require energy. Directly or indirectly nearly all energy for life comes from the sun. Autotrophs:
Photosynthesis and Cellular Respiration Photosynthesis and Cellular Respiration All cellular activities require energy. Directly or indirectly nearly all energy for life comes from the sun. Autotrophs:
Metabolismo Biología de 12º
 DEPARTAMENTO DE CIENCIAS NATURALES Metabolismo Biología de 12º Nombre y Apellidos FOTOSÍNTESIS 1) Organisms that can exist with light as an energy source and an inorganic form of carbon and other raw materials
DEPARTAMENTO DE CIENCIAS NATURALES Metabolismo Biología de 12º Nombre y Apellidos FOTOSÍNTESIS 1) Organisms that can exist with light as an energy source and an inorganic form of carbon and other raw materials
2. In regards to the fluid mosaic model, which of the following is TRUE?
 General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-13 PHOTOSYNTHESIS IN HIGHER PLANTS
 CHAPTER-13 PHOTOSYNTHESIS IN HIGHER PLANTS") CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-13 PHOTOSYNTHESIS IN HIGHER PLANTS Photosynthesis is an enzyme regulated anabolic process of manufacture of organic compounds inside the chlorophyll
CBSE Quick Revision Notes (Class-11 Biology) CHAPTER-13 PHOTOSYNTHESIS IN HIGHER PLANTS Photosynthesis is an enzyme regulated anabolic process of manufacture of organic compounds inside the chlorophyll
Photosynthesis Overview
 Photosynthesis 1 2 Photosynthesis Overview Energy for all life on Earth ultimately comes from photosynthesis 6CO 2 + 12H 2 O C 6 H 12 O 6 + 6H 2 O + 6O 2 Oxygenic photosynthesis is carried out by Cyanobacteria
Photosynthesis 1 2 Photosynthesis Overview Energy for all life on Earth ultimately comes from photosynthesis 6CO 2 + 12H 2 O C 6 H 12 O 6 + 6H 2 O + 6O 2 Oxygenic photosynthesis is carried out by Cyanobacteria
Photosynthesis and Life
 7-1 Chapter 7 Photosynthesis and Life During photosynthesis Organisms use the energy of light to build highenergy organic molecules. Plants, algae, and some bacteria can do this. Can make their own food
7-1 Chapter 7 Photosynthesis and Life During photosynthesis Organisms use the energy of light to build highenergy organic molecules. Plants, algae, and some bacteria can do this. Can make their own food
BIOL 1030 Introduction to Biology: Organismal Biology. Spring 2011 Section A. Steve Thompson:
 BIOL 1030 Introduction to Biology: Organismal Biology. Spring 2011 Section A Steve Thompson: stthompson@valdosta.edu http://www.bioinfo4u.net 1 First off for something completely different, before you
BIOL 1030 Introduction to Biology: Organismal Biology. Spring 2011 Section A Steve Thompson: stthompson@valdosta.edu http://www.bioinfo4u.net 1 First off for something completely different, before you
1 (a) Fig. 1.1 is a diagram representing a three-dimensional view of a chloroplast. space B. Fig (i) Name parts A to C in Fig A... B...
 Fig. 1.1 is a diagram representing a three-dimensional view of a chloroplast. space B. Fig (i) Name parts A to C in Fig A... B...") 1 (a) Fig. 1.1 is a diagram representing a three-dimensional view of a chloroplast. A space B C Fig. 1.1 (i) Name parts A to C in Fig. 1.1. A... B... C... [3] (ii) Describe two ways in which the structure
1 (a) Fig. 1.1 is a diagram representing a three-dimensional view of a chloroplast. A space B C Fig. 1.1 (i) Name parts A to C in Fig. 1.1. A... B... C... [3] (ii) Describe two ways in which the structure
Energy Converion: Mitochondria and Chloroplasts. Pınar Tulay, Ph.D.
 Energy Converion: Mitochondria and Chloroplasts Pınar Tulay, Ph.D. pintulay@gmail.com Energy Conversion Prokaryotes use plasma membrane to produce adenosine triphosphate (ATP) used in the cell function
Energy Converion: Mitochondria and Chloroplasts Pınar Tulay, Ph.D. pintulay@gmail.com Energy Conversion Prokaryotes use plasma membrane to produce adenosine triphosphate (ATP) used in the cell function
AP Bio-Ms.Bell Unit#3 Cellular Energies Name
 AP Bio-Ms.Bell Unit#3 Cellular Energies Name 1. Base your answer to the following question on the image below. 7. Base your answer to the following question on Which of the following choices correctly
AP Bio-Ms.Bell Unit#3 Cellular Energies Name 1. Base your answer to the following question on the image below. 7. Base your answer to the following question on Which of the following choices correctly
Name: Class: _ Date: ID: A. AP Photosynthesis Test 2012
 Name: Class: _ Date: ID: A AP Photosynthesis Test 2012 Multiple Choice (3 polnts each) _ Directions: Each of the questions or incomplete statements below is followed by four suggested answers or completions.
Name: Class: _ Date: ID: A AP Photosynthesis Test 2012 Multiple Choice (3 polnts each) _ Directions: Each of the questions or incomplete statements below is followed by four suggested answers or completions.
Chemistry of Life Cells & Bioprocesses CRT Review
 Chemistry of Life Cells & Bioprocesses CRT Review Chapter 2: The Chemistry of Life macromolecules - The four types of macromolecules are carbohydrates, lipids, nucleic acids, and proteins Types of Macromolecules
Chemistry of Life Cells & Bioprocesses CRT Review Chapter 2: The Chemistry of Life macromolecules - The four types of macromolecules are carbohydrates, lipids, nucleic acids, and proteins Types of Macromolecules
AP Biology. Chloroplasts: sites of photosynthesis in plants
 The summary equation of photosynthesis including the source and fate of the reactants and products. How leaf and chloroplast anatomy relates to photosynthesis. How photosystems convert solar energy to
The summary equation of photosynthesis including the source and fate of the reactants and products. How leaf and chloroplast anatomy relates to photosynthesis. How photosystems convert solar energy to
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
Follow this and additional works at: Part of the Agriculture Commons, Genetics Commons, and the Plant Biology Commons
 Graduate Theses and Dissertations Iowa State University Capstones, Theses and Dissertations 2015 The CO2 concentrating mechanism and nitrogen starvation effects under photoautotrophic culture conditions
Graduate Theses and Dissertations Iowa State University Capstones, Theses and Dissertations 2015 The CO2 concentrating mechanism and nitrogen starvation effects under photoautotrophic culture conditions
Page 1. Name: UNIT: PHOTOSYNTHESIS AND RESPIRATION TOPIC: PHOTOSYNTHESIS
 Name: 4667-1 - Page 1 UNIT: PHOTOSYNTHESIS AND RESPIRATION TOPIC: PHOTOSYNTHESIS 1) The diagram below illustrates the movement of materials involved in a process that is vital for the energy needs of organisms.
Name: 4667-1 - Page 1 UNIT: PHOTOSYNTHESIS AND RESPIRATION TOPIC: PHOTOSYNTHESIS 1) The diagram below illustrates the movement of materials involved in a process that is vital for the energy needs of organisms.
Light form the sun is composed of a range of wavelengths (colors). The visible spectrum to the left illustrates the wavelengths and associated color
. The visible spectrum to the left illustrates the wavelengths and associated color") Photosynthesis Englemann Experiment In 1883, Thomas Engelmann of Germany used a combination of aerobic bacteria and a filamentous alga to study the effect of various colors of the visible light spectrum
Photosynthesis Englemann Experiment In 1883, Thomas Engelmann of Germany used a combination of aerobic bacteria and a filamentous alga to study the effect of various colors of the visible light spectrum
Biology: Life on Earth
 Biology: Life on Earth Eighth Edition Lecture for Chapter 7 Capturing Solar Energy: Photosynthesis Chapter 7 Outline 7.1 What Is Photosynthesis? p. 118 7.2 Light-Dependent Reactions: How Is Light Energy
Biology: Life on Earth Eighth Edition Lecture for Chapter 7 Capturing Solar Energy: Photosynthesis Chapter 7 Outline 7.1 What Is Photosynthesis? p. 118 7.2 Light-Dependent Reactions: How Is Light Energy
Isoenzymic forms of carbonic anhydrase in the red macroalga Porphyra leucosticta
 PHYSIOLOGIA PLANTARUM 106: 69 74. 1999 Copyright Physiologia Plantarum 1999 Printed in Ireland all rights reser ed ISSN 0031-9317 Isoenzymic forms of carbonic anhydrase in the red macroalga Porphyra leucosticta
PHYSIOLOGIA PLANTARUM 106: 69 74. 1999 Copyright Physiologia Plantarum 1999 Printed in Ireland all rights reser ed ISSN 0031-9317 Isoenzymic forms of carbonic anhydrase in the red macroalga Porphyra leucosticta
PHOTOSYNTHESIS Chapter 6
 PHOTOSYNTHESIS Chapter 6 5.1 Matter and Energy Pathways in Living Systems Chapter 5 Photosynthesis & Cellular Respiration 1 2 5.1 Matter and Energy Pathways in Living Systems In this section you will:
PHOTOSYNTHESIS Chapter 6 5.1 Matter and Energy Pathways in Living Systems Chapter 5 Photosynthesis & Cellular Respiration 1 2 5.1 Matter and Energy Pathways in Living Systems In this section you will:
PHOTOSYNTHESIS. Chapter 10
 PHOTOSYNTHESIS Chapter 10 Modes of Nutrition Autotrophs self-feeders Capture free energy from physical sources in the environment Photosynthetic organisms = sunlight Chemosynthetic organisms = small inorganic
PHOTOSYNTHESIS Chapter 10 Modes of Nutrition Autotrophs self-feeders Capture free energy from physical sources in the environment Photosynthetic organisms = sunlight Chemosynthetic organisms = small inorganic
Photosynthesis. Diffusion. Basic Properties of Molecules in Motion. Osmosis- passive transport of water across a membrane
 The detailed structure of an animal cell s plasma membrane, in cross section Photosynthesis Basic Properties of Molecules in Motion Diffusion: the random movement of molecules from a region of high concentration
The detailed structure of an animal cell s plasma membrane, in cross section Photosynthesis Basic Properties of Molecules in Motion Diffusion: the random movement of molecules from a region of high concentration
1/25/2018. Bio 1101 Lec. 5, Part A Chapter 6: Cellular Respiration
 1 2 3 4 5 Bio 1101 Lec. 5, Part A Chapter 6: Cellular Respiration Energy is needed by cells to do work Chemical energy, a form of potential energy, is stored in bonds of food molecules (such as glucose)
1 2 3 4 5 Bio 1101 Lec. 5, Part A Chapter 6: Cellular Respiration Energy is needed by cells to do work Chemical energy, a form of potential energy, is stored in bonds of food molecules (such as glucose)
PHOTOSYNTHESIS: converts light energy to the chemical energy of food 6CO 2 + 6H 2 O + light energy C 6 H 12 O 6 + 6O 2
 Photosynthesis Life on Earth is solar powered Photosynthesis nourishes almost all the living world directly or indirectly All organisms use organic compounds for energy and for carbon skeletons. Organisms
Photosynthesis Life on Earth is solar powered Photosynthesis nourishes almost all the living world directly or indirectly All organisms use organic compounds for energy and for carbon skeletons. Organisms
Class XI Chapter 8 Cell The Unit of Life Biology
 Question 1: Which of the following is not correct? (a) Robert Brown discovered the cell. (b) Schleiden and Schwann formulated the cell theory. (c) Virchow explained that cells are formed from pre-existing
Question 1: Which of the following is not correct? (a) Robert Brown discovered the cell. (b) Schleiden and Schwann formulated the cell theory. (c) Virchow explained that cells are formed from pre-existing
Honors Biology Fall Final Exam Study Guide
 Honors Biology Fall Final Exam Study Guide Helpful Information: Exam has 100 multiple choice questions. Be ready with pencils and a four-function calculator on the day of the test. Review ALL vocabulary,
Honors Biology Fall Final Exam Study Guide Helpful Information: Exam has 100 multiple choice questions. Be ready with pencils and a four-function calculator on the day of the test. Review ALL vocabulary,
Chapter 10 Photosynthesis
 Chapter 10 Photosynthesis Photosynthesis Process by which plants use light energy to make food. A reduction process that makes complex organic molecules from simple molecules. Ps General Equation 6 CO
Chapter 10 Photosynthesis Photosynthesis Process by which plants use light energy to make food. A reduction process that makes complex organic molecules from simple molecules. Ps General Equation 6 CO
2/6/2011. Essentials of Biology. 6.1 Overview of Photosynthesis. Investigating Photosynthesis
 Investigating Photosynthesis Essentials of Biology Sylvia S. Mader One of the first questions. When a tiny seedling grows into a tall tree with a mass of several tons, where does all that mass come from?
Investigating Photosynthesis Essentials of Biology Sylvia S. Mader One of the first questions. When a tiny seedling grows into a tall tree with a mass of several tons, where does all that mass come from?
Heterotrophs: Organisms that depend on an external source of organic compounds
 Heterotrophs: Organisms that depend on an external source of organic compounds Autotrophs: Organisms capable of surviving on CO2 as their principle carbon source. 2 types: chemoautotrophs and photoautotrophs
Heterotrophs: Organisms that depend on an external source of organic compounds Autotrophs: Organisms capable of surviving on CO2 as their principle carbon source. 2 types: chemoautotrophs and photoautotrophs
 Question 1: Which of the following is not correct? (a) Robert Brown discovered the cell. (b) Schleiden and Schwann formulated the cell theory. (c) Virchow explained that cells are formed from pre-existing
Question 1: Which of the following is not correct? (a) Robert Brown discovered the cell. (b) Schleiden and Schwann formulated the cell theory. (c) Virchow explained that cells are formed from pre-existing
Organelles & Cells Student Edition. A. chromosome B. gene C. mitochondrion D. vacuole
 Name: Date: 1. Which structure is outside the nucleus of a cell and contains DNA? A. chromosome B. gene C. mitochondrion D. vacuole 2. A potato core was placed in a beaker of water as shown in the figure
Name: Date: 1. Which structure is outside the nucleus of a cell and contains DNA? A. chromosome B. gene C. mitochondrion D. vacuole 2. A potato core was placed in a beaker of water as shown in the figure
VOCABULARY COMPTETENCIES. Students, after mastering the materials of Plant Physiology course, should be able to:
 1 VOCABULARY Forget not, exam includes ENGLISH WORDS 1. Involve 2. Bundle 3. Sheath 4. Subsequent 5. Ambient 6. Stick together 7. Determine 8. Evolution 9. Thrive 10. Allow COMPTETENCIES Students, after
1 VOCABULARY Forget not, exam includes ENGLISH WORDS 1. Involve 2. Bundle 3. Sheath 4. Subsequent 5. Ambient 6. Stick together 7. Determine 8. Evolution 9. Thrive 10. Allow COMPTETENCIES Students, after
2.A.2- Capture and Storage of Free Energy
 2.A.2- Capture and Storage of Free Energy Big Idea 2: Biological systems utilize free energy and molecular building blocks to grow, to reproduce, and to maintain dynamic homeostasis. EU 2.A- Growth, reproduction
2.A.2- Capture and Storage of Free Energy Big Idea 2: Biological systems utilize free energy and molecular building blocks to grow, to reproduce, and to maintain dynamic homeostasis. EU 2.A- Growth, reproduction
Forms of stored energy in cells
 Forms of stored energy in cells Electrochemical gradients Covalent bonds (ATP) Reducing power (NADH) During photosynthesis, respiration and glycolysis these forms of energy are converted from one to another
Forms of stored energy in cells Electrochemical gradients Covalent bonds (ATP) Reducing power (NADH) During photosynthesis, respiration and glycolysis these forms of energy are converted from one to another
Scale in the biological world
 Scale in the biological world 2 A cell seen by TEM 3 4 From living cells to atoms 5 Compartmentalisation in the cell: internal membranes and the cytosol 6 The Origin of mitochondria: The endosymbion hypothesis
Scale in the biological world 2 A cell seen by TEM 3 4 From living cells to atoms 5 Compartmentalisation in the cell: internal membranes and the cytosol 6 The Origin of mitochondria: The endosymbion hypothesis
MOLECULAR ACTIVITIES OF PLANT CELLS
 MOLECULAR ACTIVITIES OF PLANT CELLS An introduction to plant biochemistry JOHN W. ANDERSON BAgrSc, PhD Reader, Botany Department, School of Biological Sciences, La Trobe University, Bundoora, Victoria,
MOLECULAR ACTIVITIES OF PLANT CELLS An introduction to plant biochemistry JOHN W. ANDERSON BAgrSc, PhD Reader, Botany Department, School of Biological Sciences, La Trobe University, Bundoora, Victoria,
Transport of glucose across epithelial cells: a. Gluc/Na cotransport; b. Gluc transporter Alberts
 Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Active Learning Exercise 6. Photosynthesis
 Name Biol 211 - Group Number Active Learning Exercise 6. Photosynthesis Reference: Chapter 10 (Biology by Campbell/Reece, 8 th ed.) Note: See the last page of this ALE for a diagram that summarizes the
Name Biol 211 - Group Number Active Learning Exercise 6. Photosynthesis Reference: Chapter 10 (Biology by Campbell/Reece, 8 th ed.) Note: See the last page of this ALE for a diagram that summarizes the
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression