Long food chains are in general chaotic
|
|
|
- Marvin Morgan
- 5 years ago
- Views:
Transcription
1 OIKOS 109: 135/144, 2005 Long food chains are in general chaotic Thilo Gross, Wolfgang Ebenhöh and Ulrike Feudel Gross, T., Ebenhöh, W. and Feudel, U Long food chains are in general chaotic. / Oikos 109: 135/144. The question whether chaos exists in nature is much debated. In this paper we prove that chaotic parameter regions exist generically in food chains of length greater than three. While nonchaotic dynamics is also possible, the presence of chaotic parameter regions indicates that chaotic dynamics is likely. We show that the chaotic regions survive even at high exponents of closure. Our results have been obtained using a general food chain model that describes a large class of different food chains. The existence of chaos in models of such generality can be deduced from the presence of certain bifurcations of higher codimension. T. Gross, W. Ebenhöh and U. Feudel, Instıtut für Chemie und Biologie des Meeres, Carl von Ossietzky Universität, DE Oldenburg, Germany (thilo.gross@ physics.org). The long term dynamics of any deterministic system can either be stationary, periodic, quasiperiodic or chaotic. Although ecological models were among the first examples of chaotic dynamics (May 1976) the question whether chaos is an ecological reality remains open (May 1987, Upadhyay et al. 1998, Rai and Schaffer 2001, Cushing et al. 2002). In nature chaos is generally difficult to detect because of the presence of observational noise (Nychka et al. 1992, Ellner and Turchin 1995). Nevertheless, chaos has been found for instance in the dynamics of perennial grasses (Tilman and Wedin 1991), flour beetles (Cushing et al. 1996) and boreal rodents (Hanski et al. 1993). Many other systems seem to be in critical states at the edge of chaos (Turchin and Ellner 2000). From the theoretical point of view population dynamics should be chaotic if chaos is in principle possible in a given system and proves to be advantageous in the evolutionary context. Regarding the effect of chaos on the evolutionary fitness of species two main lines of reasoning exist. On the one hand it is argued that the seemingly random behavior that characterizes chaos can eventually cause the extinction of species (Lande 1993). On the other hand, it has been proved that chaotic fluctuations are desirable in a spatially extended environment (Allen et al. 1993, Solé and Gamarra 1998, Petrovskii et al. 2004). Such fluctuations increase the chance that populations survive periods of detrimental conditions in isolated patches. Starting from these patches the surrounding area can be repopulated once the conditions improve. Following this line of reasoning chaotic dynamics can increase the chances of species survival. Consequently, it is reasonable to expect that ecological systems could evolve towards chaotic regions in parameter space if such regions exist. While chaotic attractors have been found in many models (Hastings and Powell 1991, Boer et al. 1998) they seem to be absent from others. For instance it was shown by Ruxton and Rohani (1998) that chaotic regions exist in certain models, but disappear if the model structure is perturbed in a certain way. It has often been postulated that chaos would disappear if sufficient biological detail were taken into account (Fussmann and Heber 2002, Kondoh 2003). However, from a dynamical systems point of view one would expect that increasing the complexity of the model increases the complexity of the dynamics as well (May 1973). Consequently, the question arises whether chaos exists generically in ecological Accepted 4 October 2004 Copyright # OIKOS 2005 ISSN OIKOS 109:1 (2005) 135
2 systems. If chaos is generic feature of a given model its existence does not depend strongly on assumptions that are made in the model. In this case chaos is likely to occur in the natural system as well. In this paper we prove the existence of complex (that is, chaotic) dynamics in a simple, but very general food chain model. This model is a generalization of a large number of models that are discussed in ecological literature. A similar model has been used recently to investigate the effect of non-standard predator response functions (Gross et al. 2004). In that paper we have shown that the stability properties of the model are very sensitive to the exact shape of the response functions. In order to achieve a higher degree of generality we have extended the model to accommodate additional mortality terms and noninteger exponents of closure. The exponent of closure describes the mortality of the top predator in the food chain. It is often very sensitive to changes in environmental parameters and has a large impact on the food chain dynamics (Edwards and Yool 2000). The exponent of closure is therefore of pivotal importance for the coupling of biological and physical processes (Edwards and Bees 2001). High exponents of closure have a damping effect on food chain dynamics. In Steele and Henderson (1992) the hypothesis was raised that even limit cycles disappear at high exponents of closure. Although this was shown to be wrong Edwards and Yool (2000), high exponents of closure are still widely believed to prevent chaotic dynamics. However, in the folllowing we show that chaotic parameter regions generally exist in long food chains with high exponents of closure. To confirm the existence of chaotic regions in a general model is quite difficult. Numerically chaos can be detected by computing Lyapunov exponents, which measure the divergence of neighboring trajectories / a basic characteristic of chaos. Apart from computational problems which arise due to the large separation of time scales the numerical approach has a conceptual disadvantage. Numerically we can only deal with specific systems which makes it very difficult to draw general conclusions. By contrast, very general systems can be studied analytically. However, a direct analytical proof of the existence of chaos is only possible for very few systems. The emergence of chaotic behavior is closely connected to the existence of certain bifurcations of higher codimension (Kuznetsov 1995). We show that these bifurcation can be computed for very general models. In this way the generic existence of chaos can be proved indirectly. Our analysis reveals that in food chains of length four or more such bifurcations do generally exist. The paper is organized as follows: in A general food chain model we review and extend the general food chain model from Gross et al. (2004). The model allows us to discuss local bifurcations without restricting the predator response to a specific functional form. Some important bifurcations and their implications are discussed in Bifurcations of the general food chain model. Local bifurcation analysis is then used in Chaos in the general food chain model to discuss the effect of high exponents of closure in food chains of length three to six. The results are illustrated by numerical investigation of A specific example for the emergence of chaos. Finally, we discuss our findings. A general food chain model In order to study the existence of chaos at high exponents of closure we extend the general food chain model introduced by Gross et al. (2004). In this previous work we have shown that increasing the amount of available prey destabilizes the food chain if standard response functions (i.e. Holling functions) are used. This paradox of enrichment is well known from ecological literature. However, we have also shown that enrichment can have a stabilizing effect if the response functions have a slightly different shape. This shows that the dynamics of the model depends qualitatively on the precise shape of the response function. We therefore avoid to restrict the predator /prey interaction to any specific functional form. As a result our conclusions are valid for a whole class of different food chain models. We consider a general food chain of N trophic levels. Each of these levels is occupied by one model species. In a classical food chain the model species represents actual biological species. However, in literature many models are studied in which the model species stand for large groups of ecological species. For instance the phytoplankton/zooplankton food chain proposed by Steele and Henderson (1992) consists of two model species which denote the total abundances of phytoplankton an zooplankton respectively. In this sense the general food chain model studied here can be regarded as a generalization of a large number specific models that describe food webs as well as food chains. In the general food web model the biomass or abundance of model species n is denoted by the state variable X n : Whether abundance or biomass is used depends on the specific system under consideration. In this paper we use the two terms equivalently. We assume the primary production of biomass to be some function S of the biomass of the primary producer X 1 : All other species increase their biomass by feeding on other species. The species n feeds upon species n1 and is fed upon by species n1: We assume that the predation rates are proportional to the predator biomass and to some general response function G n of the prey biomass. Furthermore we take into account that only a fraction h n of the biomass lost by the prey can be converted into predator biomass. The top-predator 136 OIKOS 109:1 (2005)
3 mortality is given in terms of a function D of the toppredator abundance. For all other species (apart from primary producer and top-predator) we assume the mortality to be proportional to the abundance of the species. The food chain is described by the following system of ordinary differential equations (ODEs) Ẋ 1 S(X 1 )G 1 (X 1 )X 2 Ẋ n h n G n1 (X n1 )X n G n (X n )X n1 M n X n Ẋ N h N G N1 (X N-1 )X N D(X N ) (1) where n/2...n/1. We consider the dynamics around a steady state. The biomass in the steady state are denoted by X + 1 ;...; X+ N : We assume that the steady state is positive (/X + n 0 for n1...n) but not necessarily stable. Note, that there may be some cases in which no positive steady state exists. However, in most cases there is at least one positive steady state. As we show in the following most food chains have more than one positive steady state. In this case the analysis presented here can be applied to each steady state individually. We can use the steady state under consideration to normalize the ODE system. In the following we use lower case symbols for normalized functions and variables. We define x n œ X n =X + n g n (x n ) œ G n (X + n x n )=G n (X+ n ) s(x 1 ) œ S(X + 1 x 1 )=S(X+ 1 ) (2) d(x N ) œ D(X + N x n )=D(X+ N ) b n œ G n (X + n )X+ n1 =(M nx + n G n(x + n )X+ n1 ) Note that the normalized steady state is X 1... X N 1 and g n (x+ n )s(x+ 1 )d(x+ N )1: Using the normalized coordinates, the general ODE system can be written as ẋ 1 a 1 (s(x 1 )g 1 (x 1 )x 2 ) ẋ n a n (g n1 (x n1 )x n b n g n (x n )x n1 (1b n )x n ) ẋ N a N (g N1 (x N1 )x N d(x N )) (3) As a result of the normalization we get N new parameters a 1 ;...; a N : Considering the ODEs in the steady state shows that these parameters are given by a 1 S(X + 1 )=X+ 1 G 1 (X + 1 )X+ 2 =X+ 1 a n h n G n1 (X + n1 ) G n (X+ n )X+ n1 =X+ n M n a N h N G N1 (X + N1 ) D N (X+ N )=X+ N (4) We can not compute the values of a 1 ;...; a N with the chosen degree of generality. But, we can guess their values based on biological considerations. From Eq. 3 it is apparent that the parameter a n denotes the time scale on which the dynamics of species n takes place. In other words a n denotes the per-capita growth rate and the per capita death rate of individuals of species n in the steady state. In nature an allometric slowing down is observed in many food chains (Hendriks 1999). That is, the ratio r of prey and predator time scales is independent of the predator /prey pair under consideration. Furthermore, one of the time scales can always be set to one by renormalizing time. It is therefore reasonable to assume a n r n1 (5) Let us now consider the stability of the normalized steady state. The local stability of steady states depends on the eigenvalues of the Jacobian in the steady state. The Jacobian J is a NN matrix with J j (6) The Jacobian of our food chain model contains the 1 1 jx 1 x + n (x n N N jx n x + n jx N x + N f g n p Using these definitions the Jacobian in the normalized steady state can be written as r 1 : J :: B A 0 (fg 1 ) r N2 r N1 1 g 1 v ::: 2 b : 2 :: : :: g N2 v N1 b N1 g N1 (1p) 1 C A where v n b n (1g n ): Since no other parameters appear in the Jacobian, the stability of the normalized steady state is determined by r; p; f; g 1...g N1 and b 2 ;...; b N1 : Let us now examine these parameters more closely. The parameter b n denotes the fraction of the biomass flow in the steady state that is lost by species n because of predation. Consequently, 1b n is the fraction of the losses that is due to mortality (Eq. 2). The parameter r describes the separation of time scales in the food chain. For r close to zero the separation of time scales is large. In this case the dynamics of every predator is much slower than the dynamics of its prey. At r1 the dynamics of all species happens on the same timescale. In general we expect that the steady states are more stable in food chains with small r than in those with large r: However, we will see in the following that this is not always the case. (7) OIKOS 109:1 (2005) 137
4 For the stability of steady states the parameters g 1 ;...; g N are very important. High values of g n indicate that the predator n1 is very sensitive to the density of its prey x n : While, at small values of g n ; the predators hardly notice variations in the prey density. If a simple response function (for instance Holling Type II) is used small values of g n occur if prey is abundant, while scarce prey corresponds to high g n : In order to present our result in a convenient way we assume g 1...g N1 G (8) If g 1 ;...; g N1 are not identical the bifurcation diagrams differ slightly from the ones shown here. However, our conclusions do not depend on the identity of the parameters in a critical way. The parameter f describes the availability of nutrients or the rate of nutrient supply. If f is 1 the gain of the primary producer is proportional to its abundance. This is typically the case in environments with unlimited carrying capacity. Smaller values of f indicate that the environment has some finite carrying capacity. At f0 the gain ofthe primary producer does not depend on its abundance at all. This behavior would be strange for a biological species of primary producers. However, we have to keep in mind that x 1 does not necessarily need to be used to describe a biological species. For instance x 1 can be used to model a nutrient which is added at a constant rate and does not leave the system unless it is consumed. In Gross et al. (2004) we have discussed the effect of f on a general food chain. In the context of that article f is called 1h: It turned out that different choices of f do not alter the behavior of the model qualitatively. Therefore we consider only the case f0:5 in this paper. In our model the most interesting parameter is p: This parameter is commonly called exponent of closure. In ecological models the closure term is often assumed to be a mononomial. That is, D(X N )cx p N (9) where c is a constant. By applying Eq. 2 and 7 it can be checked that the exponent p of the closure term from Eq. 9 is indeed the parameter p defined in Eq. 7. The value of p in any given system depends on the main cause of top predator mortality. For instance, most diseases can be modeled by assuming a biomass loss which is roughly proportional to the square of biomass density. In this case the closure is quadratic and p2: By contrast, natural mortality is proportional to the biomass density. If this is the main cause of top predator biomass loss the closure is linear and p1: Another possible cause of top predator mortality is predation by a super predator which is not explicitly modeled. In this case p depends on the feeding strategy employed by this super predator. For instance filtration feeders generally impose linear closure on the food chain. While predation by an ambush feeder can be described with a quadratic closure term. A more detailed discussion of this point is given by Edwards and Bees (2001). In nature there is often more than one cause for top predator mortality. For instance, in a given ecosystem, natural mortality as well as predation by a super predator employing ambush feeding may be of importance. Such situations can be described by non-integer exponents of closure between one and two. For instance a closure term D N (X N )c q X q N c ux u N (10) yields an exponent of closure p qc q Xq N uc u Xu N c q X q N c (11) u Xu N which is a weighted average of the individual exponents /q and u: In many systems the exponent of closure is an important link between physics and biology. Take for instance a food chain in an aquatic system. Increasing the turbulence of the water increases natural mortality (caused by shear forces). Although the total mortality increases, the linear mortality terms become relatively more important. As a result, the exponent of closure decreases. Furthermore, some super predators may change their strategy from ambush to filtration feeding, which is more efficient in a turbulent environment. Bifurcations of the general food chain model A general food chain model has been outlined in the previous section. In this section we discuss some bifurcations in general. These bifurcations play an important role in our discussion of the model s dynamics in Chaos in the general food chain model. A more detailed treatment of this subject can be found in most textbooks on the theory of dynamical systems (Argyris et al. 1994, Glendinning 1994, Kuznetsov 1995, Guckenheimer and Holmes 2002). A bifurcation is a point in parameter space in which the dynamics changes in a sudden, discontinuous and qualitative way. Bifurcations can be classified according to their codimension. The codimension of a bifurcation is the number of parameters that have to be varied in order to find the bifurcation point. We start by considering some codimension-1 bifurcations of steady states. These bifurcations are important for the formation of the bifurcations of codimension-2 on which our conclusions are based. A steady state in an ODE system is stable if all eigenvalues of its Jacobian have negative real parts. A bifurcation occurs if eigenvalues cross the imaginary axis of the complex plane. Since the Jacobian is a real matrix the eigenvalues have to be real or part of a 138 OIKOS 109:1 (2005)
5 complex conjugate pair of eigenvalues. In bifurcations of codimension-1 only a single eigenvalue or a single pair of eigenvalues can be involved. In general, the crossing of the imaginary axis by a single real eigenvalue corresponds to a saddle-node bifurcation. In this bifurcation two steady states collide in parameter space. Both states vanish in the collision. In ecological models the saddle-node bifurcation is frequently observed in a degenerate form, which is called transcritical bifurcation. In this bifurcation two steady states meet and exchange stability properties. A transcritical bifurcation of a stable steady state indicates a transition from one steady state to another one. Note that, the presence of this bifurcation proves that there has to be more than one steady state. Because of the normalization, the saddle-node and transcritical bifurcations are difficult to distinguish in the general food chain model that has been proposed in A general food chain model. In the following we refer to all bifurcations which are characterized by the presence of a single zero eigenvalue as general saddle-node bifurcations. For our analysis of the general food chain model this bifurcation is only of secondary importance. In many ecosystems the primary departure from stationary behavior occurs in a Hopf bifurcation. In this bifurcation a complex conjugate pair of eigenvalues crosses the imaginary axis. In the bifurcation the steady state becomes unstable. At the bifurcation point a stable limit cycle emerges (supercritical Hopf) or an unstable limit cycle vanishes (subcritical Hopf). In the supercritical Hopf bifurcation a transition from stationary to oscillatory behavior takes place. The subcritical Hopf bifurcation likewise gives rise to oscillations. However these are only of transient nature and disappear as the system approaches some other attractor. Since both the Hopf and the general saddle-node bifurcation are of codimension-1 we can expect the bifurcation points to form hypersurfaces in the parameter space. That is, in a three dimensional parameter space the Hopf bifurcation points are in general located on surfaces consisting entirely of such points. Likewise, the general saddle node bifurcation points form general saddle node bifurcation surfaces. Many important insights can be gained from the investigation of codimension-1 bifurcations of steady states. However, the main focus of this paper is chaos. A steady state can not evolve directly into a chaotic attractor via a codimension-1 bifurcation. Instead, the formation of a chaotic attractor often involves an infinite number of bifurcations. A route to chaos which is often observed starts with a Hopf bifurcation in which a limit cycle is formed. Subsequently, the limit cycle undergoes an infinite sequence of period doubling bifurcations. The chaotic attractor is formed from a limit cycle with infinite period. Another route from stationary to chaotic behavior involves the creation of tori. Again the first bifurcation on this route is a Hopf bifurcation in which a limit cycle is formed. As the parameters are changed further a Neĭmark-Sacker bifurcation occurs which marks a transition from periodic to quasiperiodic behavior. The quasiperiodic motion takes place on a two-torus at first. From the two-torus chaos can be reached directly or via the formation of a three-torus. In ecological systems a transition to chaos via the socalled Shil nikov mechanism is often observed (Deng and Hines 2002). This route to chaos is closely connected to homoclinic bifurcations. In a homoclinic bifurcation a limit cycle meets a saddle point. In the bifurcation a homoclinic orbit, that is a trajectory that approaches the saddle for t 0 and t 0; is formed. Under certain conditions a chaotic attractor is formed from an infinite number of saddle cycles, which emerge close to the bifurcation. The transitions to chaos presented here as well as several others have been studied in specific food chains (De Feo and Rinaldi 1998). The aim of this paper is however, to prove that chaos exists in a very general system, namely the general food chain model introduced above. Because of the generality of the model, numerical approaches can not be applied. On the other hand a direct analytical proof for the existence of chaos is not possible with the desired degree of generality. However, the existence of chaotic regions can be deduced indirectly from certain bifurcations of codimension-2. To find a codimension-2 bifurcation two parameters have to be varied. The codimension-2 bifurcations form hyper-lines in parameter space, that is they appear as lines in a three parameter bifurcation diagram. In this paper we focus on Takens-Bogdanov, Gavrilov- Guckenheimer and Double Hopf bifurcations. These bifurcations are formed on the lines in which codimension-1 bifurcation surfaces intersect. In the following we review these bifurcations briefly. A more detailed discussion of these bifurcations is given by Kuznetsov (1995). A simple bifurcation of codimension-2 is the Takens- Bogdanov (TB) point. In this bifurcation the Jacobian has a double zero eigenvalue. On the bifurcation line a Hopf bifurcation surface ends at a general saddle node bifurcation surface. It can be shown that a homoclinic bifurcation surface has to emerge from the bifurcation. This bifurcation can give rise to a chaotic attractor via the Shil nikov mechanism. A much more complicated situation is found in the Gavrilov-Guckenheimer (GG) bifurcation. Like the Takens-Bogdanov bifurcation, the GG bifurcation is found at the intersection of a Hopf bifurcation surface and a general saddle-node bifurcation surface. However, in this case the Jacobian has a single zero eigenvalue and a pair of purely imaginary eigenvalues. Consequently, the OIKOS 109:1 (2005) 139
6 dynamics near the bifurcation are more difficult to analyze than in the TB case. In general, a homoclinic bifurcation and a Neĭmark-Sacker bifurcation emerge from the GG bifurcation. The homoclinic bifurcation indicates the presence of a homoclinic orbit, which may again give rise to Shil nikov chaos. At the Neĭmark- Sacker bifurcation the limit cycle which has been created in the Hopf bifurcation becomes a torus, in many (but not all) cases the torus only exists close to the GG bifurcation, before it evolves into a chaotic attractor. The TB and GG bifurcations indicate in many cases (but not always) the existence of a chaotic region. Nevertheless we may say that chaotic dynamics are likely (but not certain) to exist near these bifurcations. In this paper we draw our conclusions mainly from the presence of Double Hopf (DH) bifurcations. Double Hopf bifurcations are formed at the intersection of two Hopf bifurcations. In the DH bifurcation the Jacobian has two purely imaginary pairs of complex conjugate eigenvalues. The analysis of the dynamics is in this case even more complicated. Nevertheless, it has been shown that a chaotic region has to exist close to a generic DH bifurcation (Kuznetsov 1995). Only in degenerate cases (i.e. Lotka/Volterra food chains) the creation of a chaotic attractor can be avoided. Degeneracies are very special exceptions to a general rule. In models degeneracies often appear because of the simple assumptions on which many models are based. However, the dynamics encountered in nature is in general generic (that is, nondegenerate). The probability that a system observed in nature is by chance degenerate is zero. We can summarize the contents of this section by formulating the following approach. The generic existence of a chaotic region in the food chain model from A general food chain model can be proved by showing that a Double Hopf bifurcation exists. If the Double Hopf bifurcation extends to high exponents of closure, the chaotic region extends to high exponent of closure as well. In the model Double Hopf bifurcations can be found by computing Hopf bifurcation surfaces. Plotting the Hopf bifurcations in three-parameter bifurcation diagrams reveals the Double Hopf bifurcations as intersection lines of Hopf bifurcation surfaces. Chaos in the general food chain model In the previous section we have formulated an indirect approach by which chaos can be proved to exist. In this section we apply this approach to the general food chain model introduced in A general food chain model. We start our analysis by computing the codimension-1 bifurcations of the steady states. The general saddlenode bifurcations can be easily computed by demanding the determinant of the Jacobian to vanish. For the computation of Hopf bifurcations we use the method of resultants (Guckenheimer et al. 1997, Gross and Feudel 2004). This method yields an analytical testfunction. For long food chains the testfunctions become rather lengthy. In order to obtain bifurcation diagrams shown in this paper testfunctions have been computed and solved using the symbolic algebra software Maple. Let us consider the case in which the mortality terms for all species except for the top predator and primary producer can be neglected (/b 2...b N1 1): Three parameter bifurcation diagrams for food chains of different length are shown in Fig. 1. The three parameter axes are the relative time scale r; the sensitivity to prey density G and the exponent of closure p: Every point in the bifurcation diagram is related to a specific system with the corresponding parameter values. The surfaces shown in the diagram consist of codimension-1 bifurcations of steady states. Surfaces of general saddle-node bifurcations are shown in blue. All other colors correspond to Hopf bifurcations. High sensitivity to prey abundance, that is high G always stabilizes the normalized steady state. Therefore the normalized steady state is always stable in the topmost volume of the diagrams, that is above all bifurcation surfaces. As parameter values are varied Hopf bifurcations may be crossed, leaving the topmost volume. At this point the normalized steady state becomes unstable. More bifurcations may be encountered if parameters are changed further. However, the steady state is unstable everywhere except in the topmost volume of parameter space. In the remainder of this section we discuss each of the bifurcation diagrams individually. We focus on the Double Hopf bifurcations, which indicate the existence of chaos. In addition we point out some other results regarding the effect of the time scale separation r and the exponent of closure p on steady state stability. Let us start by considering the tri-trophic (three level) food chain. The corresponding bifurcation diagram is shown in the top left corner of Fig. 1. The diagram shows a Hopf (red) and a general saddle-node (blue) bifurcation surface. The primary loss of stability occurs as the Hopf bifurcation is crossed from above. In the bifurcation diagram, it can be seen that high exponents of closure stabilize the food chain as expected. As p is increased the critical value of G at which the bifurcation occurs decreases. Therefore the area in which the steady state is stable grows. Higher codimension bifurcations seem to be absent from the bifurcation diagram of the tri-trophic chain. However, there is a single point (/r0; p1; G0:5) in which the general saddle-node bifurcation meets the Hopf bifurcation. This point is part of a line of Gavrilov-Guckenheimer bifurcation points which extends to lower exponents of closure. As we increase the exponent of closure, the general saddlenode bifurcation surface folds back. At higher exponents of closure no further Gavrilov-Guckenheimer bifurcations are possible. It is reasonableto assume that chaos is 140 OIKOS 109:1 (2005)
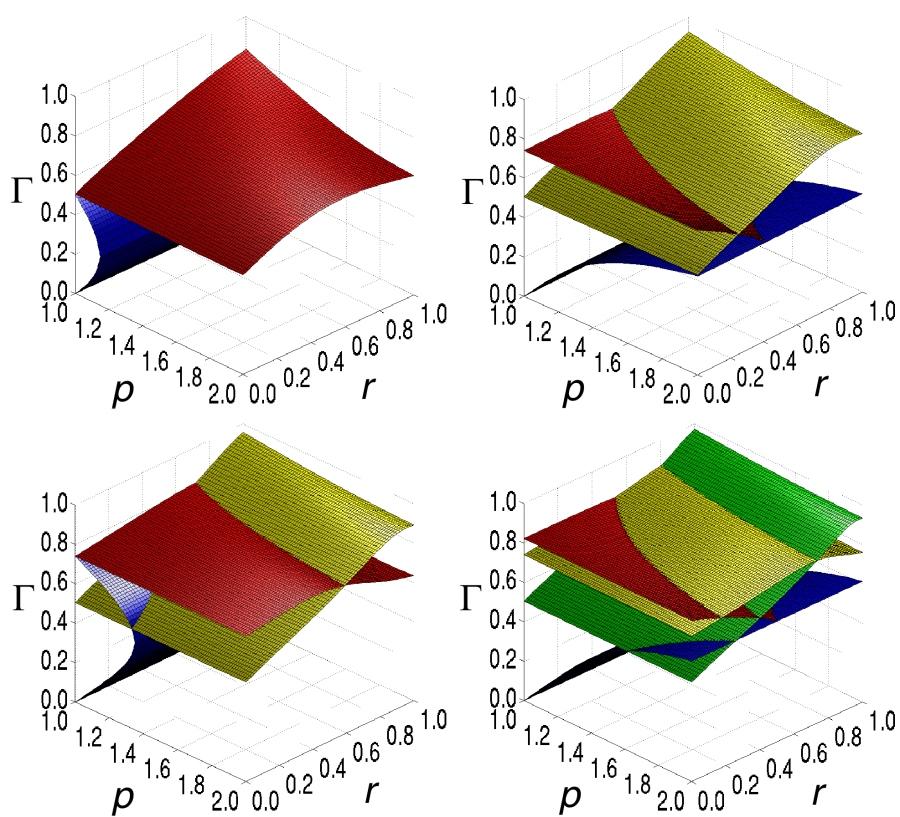
7

8 tri- or the four-trophic case. Which of the two possible shapes is found depends on whether N is even or odd. Takens-Bogdanov and Gavrilov-Guckenheimer bifurcations occur therefore at high exponents of closure in food chains of even length and at low exponents of closure in food chains of odd length. Most importantly, Double Hopf bifurcation lines are present in every food chain with N/3. Their presence proves that chaotic regions generically exist in all long food chains. It may be argued that the chaotic regions disappear if additional mortality terms are introduced. To prove that this is not the case we consider the four-trophic food chain again. In Eq. 2 we have denoted the fraction of the biomass loss of species n that occurs because of predation by b n. So far we have assumed that for species 2 and 3 predation is the only cause of biomass loss, that is b 2 /b 3 /1. If additional mortality terms are introduced the fraction of the biomass that is lost because of predation decreases. For the sake of simplicity we consider the case in which b 2 /b 3 /b. Fig. 2 shows a three-parameter bifurcation diagram at p/1.9. As b decreases the Double Hopf bifurcation occurs at a larger separation of timescales (lower r). This shows that even with strong mortality chaotic regions generically exist at high exponents of closure. These regions are located in the biologically interesting parameter range, where timescale separation is large. Fig. 2. Bifurcations of the four-trophic food chain with high exponent of closure (p/1.9). Depending on the relative separation of timescales r, the fraction of the biomass loss of species 2 and 3 that is due to predation b, and the species sensitivity to prey density G. As the mortality becomes more important (decreasing b), the double Hopf bifurcation that is formed at the intersection of the two Hopf bifurcation surfaces (grey surfaces) occurs at larger separation of timescales. In addition there is a general saddle node bifurcation surface (white surface). A specific example for the emergence of chaos In the previous section we have shown that complex dynamics can generally be expected to occur in long food chains. The existence of chaotic regions was deduced from the presence of a Double Hopf bifurcation of the steady state. By using this indirect approach we did not have to restrict ourselves to a specific model but discussed the general case. However, the creation of the chaotic attractor itself, involves more complicated bifurcations. These bifurcations can not be computed with the same degree of generality. Let us therefore illustrate our results by investigating the dynamics of a specific model. We consider a food chain which is defined analogously to Eq. 1. In this specific model we chose the food chain with four trophic levels (N/4). We assume logistic growth of the primary producer and set S(X 1 )A 0 (CX 1 )X 1 (12) where A 0 is some constant growth factor and C is the carrying capacity. The interaction between species is assumed to be of Holling type II. Furthermore, we neglect all mortality terms except the closure term which is assumed to be of mononomial form. This yields the following the differential equations Ẋ 1 A 0 (CX 1 )X 1 A 1 X 1 X 2 K 1 X 1 Ẋ 2 B 1 X 1 X 2 K 1 X 1 A 2X 2 X 3 K 2 X 2 Ẋ 3 B 2 X 2 X 3 K 2 X 2 A 3 X 3 X (13) 4 K 3 X 3 Ẋ 4 B 3 X 3 X 4 K 3 X 3 MX p 4 The parameters A 0,...,A 3, B 1,...,B 3, and M which appear in these equations depend on the specific system under consideration. Since our aim is to discuss some arbitrary example we can choose the parameters in a convenient way. We set A 0 1=(C1) (14) A n r n1 (K n 1); n1...3 (15) B n r n (K n 1); n1...3 (16) Mr 3 (17) In this case X 1...X 4 1 is a steady state and X n /x n for all n. This saves us the work of performing the normalization. For simplicity we choose K 1... K N1 K: The general parameters r and p already appear in our model. We set C to 3, which corresponds to f/0.5. The remaining general parameter G is related to the specific parameter K by G K (18) K 1 which has been obtained using Eq. 7 and 8. We can now study the dynamics of this specific model by numerical computation of Lyapunov exponents. The 142 OIKOS 109:1 (2005)

9 Lyapunov exponents measure the rate at which the distance between neighboring trajectories changes. Negative Lyapunov exponents indicate approaching trajectories, while positive Lyapunov exponents correspond to diverging trajectories. The nature of the dynamics can therefore be deduced from the number of positive, negative and zero Lyapunov exponents. In a stable steady state all Lyapunov exponents are negative. On all other attractors (in a system of ODEs) there is always at least one zero Lyapunov exponents and one negative Lyapunov exponent. Take for instance a stable limit cycle. The Lyapunov exponent which corresponds to the motion along the cycle is zero while all others are negative. Quasiperiodic motion takes place on tori. In this case we have a zero Lyapunov exponent for every dimension of the surface of the torus. That is, two zero Lyapunov exponents on a two-torus and three on a three-torus. Higher dimensional tori are rarely encountered, according to the Ruelle /Takens theorem (Ruelle and Takens 1971). On a torus attractor all other Lyapunov exponent are negative. Positive Lyapunov exponents on an attractor always correspond to chaotic motion. Figure 3 shows a two parameter bifurcation diagram for the specific model. The Double Hopf bifurcation is formed at the intersection of the two Hopf bifurcation lines. The colored areas indicate regions of different dynamical behavior. In the diagram we have distinguished between periodic, quasiperiodic and chaotic regions based on the computed Lyapunov exponents. In order to make a clear distinction between periodic and stationary behavior simulations have been used in some areas. There is one area in which all Lyapunov exponents are very small. Here we can not decide whether the attractor is chaotic or quasiperiodic on the basis of numerical computation. This is due to the very slow convergence of the algorithms for computation of Lyapunov exponents. Still, it can be seen that the chaotic region extends to high exponents of closure. From Fig. 1 it is obvious that the Double Hopf bifurcation would occur at even higher exponents of closure if a lower value of r had been chosen. In this case the chaotic region could be expected to extend to higher exponents of closure as well. However, the computation of Lyapunov exponents becomes more difficult as time scale separation grows. Discussion In this paper we have introduced a very general food chain model. By looking at higher codimension bifurcations of steady states we were able to show that chaotic regions generically exist in food chains with more than three trophic levels. Although high exponents of closure Fig. 3. Qualitative behavior of a specific four-trophic food chain, depending on the exponent of closure p and the sensitivity to prey density G. Top: local bifurcations computed analytically. The two Hopf bifurcations meet in a codimension-2 double Hopf bifurcation. Bottom: results from the numerical computation of Lyapunov exponents. The attractor is a steady state (pattern), a limit cycle (white), a two-torus (medium grey), a three-torus (light grey) or a chaotic attractor (black). In the dark grey region all Lyapunov exponents are small. In this region the nature of the attractor could not be determined. have a stabilizing effect on steady states they can not generally prevent chaotic dynamics. The fact that high exponents of closure stabilize steady states, but do not prevent complex dynamics conveys an important message. When talking of stabilizing and destabilizing effects one should be very careful to indicate the considered type of dynamical behavior. An effect or an influence stabilizing steady states may at the same time destabilize or destroy cycles or tori. Our findings describe the generic case. In certain models (like Lotka /Volterra food chains) degeneracies may exist which prevent the formation of chaotic regions. However, these degeneracies should disappear if the structure of the specific models is disturbed slightly. It is therefore reasonable to assume that chaotic parameter regions exist in natural food chains. The generic existence of chaotic regions does not imply that the dynamics of long food chains has to be OIKOS 109:1 (2005) 143
10 chaotic, but that it could be chaotic if this is advantageous from an evolutionary point of view. Although this point is much debated, some evidence seems to suggest that this is the case (Solé and Gamarra 1998). Other findings indicate that the dynamics at the edge of chaos is particularly advantageous (Gragnani et al. 1998, Turchin and Ellner 2000). In this paper we have focused on the dynamics of general food chains. However, the model species that form these food chains can be interpreted as groups of similar biological species. In this way our general food chain model describes not only a large class of food chains but also certain food webs. In order to decide whether chaos is indeed an ecological reality (or may be even an ecological necessity) even more general models should be studied. Models that take extra mortality terms, food web structure, nutrient recycling and evolutionary processes into account can be formulated and studied essentially along the same lines as the class of models discussed here. Other properties of natural ecosystems, like the effect of a spatially extended environment or external forcing may be more difficult to deal with and will probably require a different approach. Acknowledgements / The authors would like to thank A. Edwards, G. Fussmann and B. Kooi for valuable discussions. This work was supported by the Deutsche Forschungsgemeinschaft (FE 359/6). References Allen, J. C., Schaffer, W. M. and Rosko, D Chaos reduces species extinction by amplifying local population noise. / Nature 364: 229/232. Argyris, J. H., Faust, G. and Haase, M An exploration of chaos: an introduction for natural scientists and engineers. / North-Holland. Boer, M. P., Kooi, B. W. and Kooijman, S. A. L. M Food chain dynamics in the chemostat. / Math. Biosci. 150: 43/62. Cushing, J. M., Constantino, R. M. and Dennis, B Chaos in ecology: experimental nonlinear dynamics. / Academic Press. Cushing, J. M., Dennis, B., Desharnais, R. A. et al An interdisciplinary approach to understanding nonlinear ecological dynamics. / Ecol. Modell. 92: 111/119. De Feo, O. and Rinaldi, S Singular homoclinic bifurcations in tritrophic food chains. / Math. Biosci. 148: 7 /20. Deng, B. and Hines, G Food chain chaos due to Shilnikov s orbit. / Chaos 12: 533/538. Edwards, A. M. and Bees, M. A Generic dynamics of a simple plankton population model with a non-integer exponent of closure. / Chaos, Solitons and Fractals 12: 289/300. Edwards, A. M. and Yool, A The role of higher predation in plankton population models. / J. Plankton Res. 22: 1085/1112. Ellner, S. and Turchin, P Chaos in a noisy world: new methods and evidence from time-series analysis. / Am. Nat. 145: 343/375. Fussmann, G. F. and Heber, G Food web complexity and chaotic population dynamics. / Ecol. Lett. 5: 394/401. Glendinning, P Stability, instability and chaos: an introduction to the theory of nonlinear differential equations. / Cambridge Univ. Press. Gragnani, A., De Feo, O. and Rinaldi, S Food chains in the chemostat: relationships between mean yield and complex dynamics. / Bull. Math. Biol. 60: 703/719. Gross, T. and Feudel, U Analytical search for bifurcation surfaces in parameter space. / Physica D 195: 292/302. Gross, T., Ebenhöh, W. and Feudel, U Enrichment and foodchain stability: the impact of different forms of predator-prey interaction. / J. Theor. Biol. 227: 349/358. Guckenheimer, J. and Holmes, P Nonlinear oscillations, dynamical systems, and bifurcations of vector fields. / Springer. Guckenheimer, J., Myers, M. and Sturmfels, B Computing Hopf bifurcations I. / SIAM J. Numer. Anal. 34: 1 /21. Hanski, I., Turchin, P., Korpimäki, E. et al Population oscillations of boreal rodents: regulation by mustelid predators leads to chaos. / Nature 364: 232/235. Hastings, A. and Powell, T Chaos in the three-species food chain. / Ecology 72: 896/903. Hendriks, A. J Allometric scaling of rate, age and density parameters in ecological models. / Oikos 86: 293/310. Kondoh, M Foraging adaptation and the relationship between food-web complexity and stability. / Science 299: 1388/1391. Kuznetsov, Y. A Elements of applied bifurcation theory. / Springer. Lande, R Risks of population extinction from demographic and environmental stochasticity and random catastrophes. / Am. Nat. 142: 911/922. May, R. M Stability and complexity in model ecosystems. / Princeton Univ. Press. May, R. M Simple mathematical models with very complicated dynamics. / Nature 261: 459/467. May, R. M Chaos and the dynamics of biological populations. / Proc. R. Soc. Lond. A 413: 27/44. Nychka, D., Ellner, S., McCaffrey, D. et al Finding chaos in noisy systems. / J. R. Stat. Soc. B 54: 399/426. Petrovskii, S., Li, B. and Malchow, H Transition to spatiotemporal chaos can resolve the paradox of enrichment. / Ecol. Complexity 1: 37/47. Rai, V. and Schaffer, W. M Chaos in ecology. / Chaos, Solitons and Fractals 12: 197/203. Ruelle, D. and Takens, F On the nature of turbulence. / Commun. Math. Phys. 20: 167/192. Ruxton, G. D. and Rohani, P Population floors and persistence of chaos in population models. / Theor. Popul. Biol. 53: 175/183. Solé, R. V. and Gamarra, J. G. P Chaos, dispersal and extinction in coupled ecosystems. / J. Theor. Biol. 193: 539/541. Steele, J. H. and Henderson, E. W The role of predation in plankton models. / J. Plankton Res. 14: 157/172. Tilman, D. and Wedin, D Oscillations and chaos in the dynamics of perennial grasses. / Nature 353: 653/655. Turchin, P. and Ellner, S. P Living on the edge of chaos: population dynamics of Fennoscandian voles. / Ecology 81: 3099/3116. Upadhyay, R. K., Iyengar, S. R. K. and Rai, V Chaos: an ecological reality? / Int. J. Bifurcation Chaos 8: 1325/ OIKOS 109:1 (2005)
Computing 3D Bifurcation Diagrams
 Computing 3D Bifurcation Diagrams Dirk Stiefs a Ezio Venturino b and U. Feudel a a ICBM, Carl von Ossietzky Universität, PF 2503, 26111 Oldenburg, Germany b Dipartimento di Matematica,via Carlo Alberto
Computing 3D Bifurcation Diagrams Dirk Stiefs a Ezio Venturino b and U. Feudel a a ICBM, Carl von Ossietzky Universität, PF 2503, 26111 Oldenburg, Germany b Dipartimento di Matematica,via Carlo Alberto
The effect of emigration and immigration on the dynamics of a discrete-generation population
 J. Biosci., Vol. 20. Number 3, June 1995, pp 397 407. Printed in India. The effect of emigration and immigration on the dynamics of a discrete-generation population G D RUXTON Biomathematics and Statistics
J. Biosci., Vol. 20. Number 3, June 1995, pp 397 407. Printed in India. The effect of emigration and immigration on the dynamics of a discrete-generation population G D RUXTON Biomathematics and Statistics
Dynamics of Modified Leslie-Gower Predator-Prey Model with Predator Harvesting
 International Journal of Basic & Applied Sciences IJBAS-IJENS Vol:13 No:05 55 Dynamics of Modified Leslie-Gower Predator-Prey Model with Predator Harvesting K. Saleh Department of Mathematics, King Fahd
International Journal of Basic & Applied Sciences IJBAS-IJENS Vol:13 No:05 55 Dynamics of Modified Leslie-Gower Predator-Prey Model with Predator Harvesting K. Saleh Department of Mathematics, King Fahd
BIFURCATION PHENOMENA Lecture 4: Bifurcations in n-dimensional ODEs
 BIFURCATION PHENOMENA Lecture 4: Bifurcations in n-dimensional ODEs Yuri A. Kuznetsov August, 2010 Contents 1. Solutions and orbits: equilibria cycles connecting orbits compact invariant manifolds strange
BIFURCATION PHENOMENA Lecture 4: Bifurcations in n-dimensional ODEs Yuri A. Kuznetsov August, 2010 Contents 1. Solutions and orbits: equilibria cycles connecting orbits compact invariant manifolds strange
Bifurcations and chaos in a predator prey system with the Allee effect
 Received 11 February Accepted 5 February Published online May Bifurcations and chaos in a predator prey system with the Allee effect Andrew Morozov 1,, Sergei Petrovskii 1,* and Bai-Lian Li 1 1 Ecological
Received 11 February Accepted 5 February Published online May Bifurcations and chaos in a predator prey system with the Allee effect Andrew Morozov 1,, Sergei Petrovskii 1,* and Bai-Lian Li 1 1 Ecological
NUMERICAL SIMULATION DYNAMICAL MODEL OF THREE-SPECIES FOOD CHAIN WITH LOTKA-VOLTERRA LINEAR FUNCTIONAL RESPONSE
 Journal of Sustainability Science and Management Volume 6 Number 1, June 2011: 44-50 ISSN: 1823-8556 Universiti Malaysia Terengganu Publisher NUMERICAL SIMULATION DYNAMICAL MODEL OF THREE-SPECIES FOOD
Journal of Sustainability Science and Management Volume 6 Number 1, June 2011: 44-50 ISSN: 1823-8556 Universiti Malaysia Terengganu Publisher NUMERICAL SIMULATION DYNAMICAL MODEL OF THREE-SPECIES FOOD
Introduction to Dynamical Systems Basic Concepts of Dynamics
 Introduction to Dynamical Systems Basic Concepts of Dynamics A dynamical system: Has a notion of state, which contains all the information upon which the dynamical system acts. A simple set of deterministic
Introduction to Dynamical Systems Basic Concepts of Dynamics A dynamical system: Has a notion of state, which contains all the information upon which the dynamical system acts. A simple set of deterministic
NOTES ON CHAOS. Chaotic dynamics stem from deterministic mechanisms, but they look very similar to random fluctuations in appearance.
 NOTES ON CHAOS. SOME CONCEPTS Definition: The simplest and most intuitive definition of chaos is the extreme sensitivity of system dynamics to its initial conditions (Hastings et al. 99). Chaotic dynamics
NOTES ON CHAOS. SOME CONCEPTS Definition: The simplest and most intuitive definition of chaos is the extreme sensitivity of system dynamics to its initial conditions (Hastings et al. 99). Chaotic dynamics
Chaos and adaptive control in two prey, one predator system with nonlinear feedback
 Chaos and adaptive control in two prey, one predator system with nonlinear feedback Awad El-Gohary, a, and A.S. Al-Ruzaiza a a Department of Statistics and O.R., College of Science, King Saud University,
Chaos and adaptive control in two prey, one predator system with nonlinear feedback Awad El-Gohary, a, and A.S. Al-Ruzaiza a a Department of Statistics and O.R., College of Science, King Saud University,
arxiv:nlin/ v1 [nlin.cd] 29 Jan 2006
![arxiv:nlin/ v1 [nlin.cd] 29 Jan 2006](/thumbs/78/78035309.jpg "arxiv:nlin/ v1 [nlin.cd] 29 Jan 2006") Generalized models as a universal approach to the analysis of nonlinear dynamical systems Thilo Gross AG Nichtlineare Dynamik, Universität Potsdam, am Neuen Palais 10, 14469 Potsdam, Germany Ulrike Feudel
Generalized models as a universal approach to the analysis of nonlinear dynamical systems Thilo Gross AG Nichtlineare Dynamik, Universität Potsdam, am Neuen Palais 10, 14469 Potsdam, Germany Ulrike Feudel
Mathematical Foundations of Neuroscience - Lecture 7. Bifurcations II.
 Mathematical Foundations of Neuroscience - Lecture 7. Bifurcations II. Filip Piękniewski Faculty of Mathematics and Computer Science, Nicolaus Copernicus University, Toruń, Poland Winter 2009/2010 Filip
Mathematical Foundations of Neuroscience - Lecture 7. Bifurcations II. Filip Piękniewski Faculty of Mathematics and Computer Science, Nicolaus Copernicus University, Toruń, Poland Winter 2009/2010 Filip
Simple approach to the creation of a strange nonchaotic attractor in any chaotic system
 PHYSICAL REVIEW E VOLUME 59, NUMBER 5 MAY 1999 Simple approach to the creation of a strange nonchaotic attractor in any chaotic system J. W. Shuai 1, * and K. W. Wong 2, 1 Department of Biomedical Engineering,
PHYSICAL REVIEW E VOLUME 59, NUMBER 5 MAY 1999 Simple approach to the creation of a strange nonchaotic attractor in any chaotic system J. W. Shuai 1, * and K. W. Wong 2, 1 Department of Biomedical Engineering,
DYNAMICS OF A PREDATOR-PREY INTERACTION IN CHEMOSTAT WITH VARIABLE YIELD
 Journal of Sustainability Science Management Volume 10 Number 2, December 2015: 16-23 ISSN: 1823-8556 Penerbit UMT DYNAMICS OF A PREDATOR-PREY INTERACTION IN CHEMOSTAT WITH VARIABLE YIELD SARKER MD SOHEL
Journal of Sustainability Science Management Volume 10 Number 2, December 2015: 16-23 ISSN: 1823-8556 Penerbit UMT DYNAMICS OF A PREDATOR-PREY INTERACTION IN CHEMOSTAT WITH VARIABLE YIELD SARKER MD SOHEL
Application demonstration. BifTools. Maple Package for Bifurcation Analysis in Dynamical Systems
 Application demonstration BifTools Maple Package for Bifurcation Analysis in Dynamical Systems Introduction Milen Borisov, Neli Dimitrova Department of Biomathematics Institute of Mathematics and Informatics
Application demonstration BifTools Maple Package for Bifurcation Analysis in Dynamical Systems Introduction Milen Borisov, Neli Dimitrova Department of Biomathematics Institute of Mathematics and Informatics
MATH 415, WEEK 11: Bifurcations in Multiple Dimensions, Hopf Bifurcation
 MATH 415, WEEK 11: Bifurcations in Multiple Dimensions, Hopf Bifurcation 1 Bifurcations in Multiple Dimensions When we were considering one-dimensional systems, we saw that subtle changes in parameter
MATH 415, WEEK 11: Bifurcations in Multiple Dimensions, Hopf Bifurcation 1 Bifurcations in Multiple Dimensions When we were considering one-dimensional systems, we saw that subtle changes in parameter
8.1 Bifurcations of Equilibria
 1 81 Bifurcations of Equilibria Bifurcation theory studies qualitative changes in solutions as a parameter varies In general one could study the bifurcation theory of ODEs PDEs integro-differential equations
1 81 Bifurcations of Equilibria Bifurcation theory studies qualitative changes in solutions as a parameter varies In general one could study the bifurcation theory of ODEs PDEs integro-differential equations
Example of a Blue Sky Catastrophe
 PUB:[SXG.TEMP]TRANS2913EL.PS 16-OCT-2001 11:08:53.21 SXG Page: 99 (1) Amer. Math. Soc. Transl. (2) Vol. 200, 2000 Example of a Blue Sky Catastrophe Nikolaĭ Gavrilov and Andrey Shilnikov To the memory of
PUB:[SXG.TEMP]TRANS2913EL.PS 16-OCT-2001 11:08:53.21 SXG Page: 99 (1) Amer. Math. Soc. Transl. (2) Vol. 200, 2000 Example of a Blue Sky Catastrophe Nikolaĭ Gavrilov and Andrey Shilnikov To the memory of
Workshop on Theoretical Ecology and Global Change March 2009
 2022-3 Workshop on Theoretical Ecology and Global Change 2-18 March 2009 Stability Analysis of Food Webs: An Introduction to Local Stability of Dynamical Systems S. Allesina National Center for Ecological
2022-3 Workshop on Theoretical Ecology and Global Change 2-18 March 2009 Stability Analysis of Food Webs: An Introduction to Local Stability of Dynamical Systems S. Allesina National Center for Ecological
Asynchronous and Synchronous Dispersals in Spatially Discrete Population Models
 SIAM J. APPLIED DYNAMICAL SYSTEMS Vol. 7, No. 2, pp. 284 310 c 2008 Society for Industrial and Applied Mathematics Asynchronous and Synchronous Dispersals in Spatially Discrete Population Models Abdul-Aziz
SIAM J. APPLIED DYNAMICAL SYSTEMS Vol. 7, No. 2, pp. 284 310 c 2008 Society for Industrial and Applied Mathematics Asynchronous and Synchronous Dispersals in Spatially Discrete Population Models Abdul-Aziz
CHALMERS, GÖTEBORGS UNIVERSITET. EXAM for DYNAMICAL SYSTEMS. COURSE CODES: TIF 155, FIM770GU, PhD
 CHALMERS, GÖTEBORGS UNIVERSITET EXAM for DYNAMICAL SYSTEMS COURSE CODES: TIF 155, FIM770GU, PhD Time: Place: Teachers: Allowed material: Not allowed: January 14, 2019, at 08 30 12 30 Johanneberg Kristian
CHALMERS, GÖTEBORGS UNIVERSITET EXAM for DYNAMICAL SYSTEMS COURSE CODES: TIF 155, FIM770GU, PhD Time: Place: Teachers: Allowed material: Not allowed: January 14, 2019, at 08 30 12 30 Johanneberg Kristian
BIFURCATIONS AND CHAOS IN A PERIODICALLY FORCED PROTOTYPE ADAPTIVE CONTROL SYSTEM 1
 KYBERNETIKA VOLUME 3 0 (1994), NUMBER 2, PAG ES 121-128 BIFURCATIONS AND CHAOS IN A PERIODICALLY FORCED PROTOTYPE ADAPTIVE CONTROL SYSTEM 1 YURI A. KUZNETSOV AND CARLO PICCARDI An adaptive control system
KYBERNETIKA VOLUME 3 0 (1994), NUMBER 2, PAG ES 121-128 BIFURCATIONS AND CHAOS IN A PERIODICALLY FORCED PROTOTYPE ADAPTIVE CONTROL SYSTEM 1 YURI A. KUZNETSOV AND CARLO PICCARDI An adaptive control system
Models Involving Interactions between Predator and Prey Populations
 Models Involving Interactions between Predator and Prey Populations Matthew Mitchell Georgia College and State University December 30, 2015 Abstract Predator-prey models are used to show the intricate
Models Involving Interactions between Predator and Prey Populations Matthew Mitchell Georgia College and State University December 30, 2015 Abstract Predator-prey models are used to show the intricate
A Producer-Consumer Model With Stoichiometry
 A Producer-Consumer Model With Stoichiometry Plan B project toward the completion of the Master of Science degree in Mathematics at University of Minnesota Duluth Respectfully submitted by Laura Joan Zimmermann
A Producer-Consumer Model With Stoichiometry Plan B project toward the completion of the Master of Science degree in Mathematics at University of Minnesota Duluth Respectfully submitted by Laura Joan Zimmermann
3.5 Competition Models: Principle of Competitive Exclusion
 94 3. Models for Interacting Populations different dimensional parameter changes. For example, doubling the carrying capacity K is exactly equivalent to halving the predator response parameter D. The dimensionless
94 3. Models for Interacting Populations different dimensional parameter changes. For example, doubling the carrying capacity K is exactly equivalent to halving the predator response parameter D. The dimensionless
Feedback-mediated oscillatory coexistence in the chemostat
 Feedback-mediated oscillatory coexistence in the chemostat Patrick De Leenheer and Sergei S. Pilyugin Department of Mathematics, University of Florida deleenhe,pilyugin@math.ufl.edu 1 Introduction We study
Feedback-mediated oscillatory coexistence in the chemostat Patrick De Leenheer and Sergei S. Pilyugin Department of Mathematics, University of Florida deleenhe,pilyugin@math.ufl.edu 1 Introduction We study
Equilibriumizing all food chain chaos through reproductive efficiency
 CHAOS 16 043125 2006 Equilibriumizing all food chain chaos through reproductive efficiency Bo Deng Department of Math and Statistics University of Nebraska Lincoln Nebraska 68588-0323 Received 31 May 2006;
CHAOS 16 043125 2006 Equilibriumizing all food chain chaos through reproductive efficiency Bo Deng Department of Math and Statistics University of Nebraska Lincoln Nebraska 68588-0323 Received 31 May 2006;
Dynamical Systems: Ecological Modeling
 Dynamical Systems: Ecological Modeling G Söderbacka Abstract Ecological modeling is becoming increasingly more important for modern engineers. The mathematical language of dynamical systems has been applied
Dynamical Systems: Ecological Modeling G Söderbacka Abstract Ecological modeling is becoming increasingly more important for modern engineers. The mathematical language of dynamical systems has been applied
1.Introduction: 2. The Model. Key words: Prey, Predator, Seasonality, Stability, Bifurcations, Chaos.
 Dynamical behavior of a prey predator model with seasonally varying parameters Sunita Gakkhar, BrhamPal Singh, R K Naji Department of Mathematics I I T Roorkee,47667 INDIA Abstract : A dynamic model based
Dynamical behavior of a prey predator model with seasonally varying parameters Sunita Gakkhar, BrhamPal Singh, R K Naji Department of Mathematics I I T Roorkee,47667 INDIA Abstract : A dynamic model based
2D-Volterra-Lotka Modeling For 2 Species
 Majalat Al-Ulum Al-Insaniya wat - Tatbiqiya 2D-Volterra-Lotka Modeling For 2 Species Alhashmi Darah 1 University of Almergeb Department of Mathematics Faculty of Science Zliten Libya. Abstract The purpose
Majalat Al-Ulum Al-Insaniya wat - Tatbiqiya 2D-Volterra-Lotka Modeling For 2 Species Alhashmi Darah 1 University of Almergeb Department of Mathematics Faculty of Science Zliten Libya. Abstract The purpose
Predator-Prey Model with Ratio-dependent Food
 University of Minnesota Duluth Department of Mathematics and Statistics Predator-Prey Model with Ratio-dependent Food Processing Response Advisor: Harlan Stech Jana Hurkova June 2013 Table of Contents
University of Minnesota Duluth Department of Mathematics and Statistics Predator-Prey Model with Ratio-dependent Food Processing Response Advisor: Harlan Stech Jana Hurkova June 2013 Table of Contents
2 Discrete growth models, logistic map (Murray, Chapter 2)
") 2 Discrete growth models, logistic map (Murray, Chapter 2) As argued in Lecture 1 the population of non-overlapping generations can be modelled as a discrete dynamical system. This is an example of an
2 Discrete growth models, logistic map (Murray, Chapter 2) As argued in Lecture 1 the population of non-overlapping generations can be modelled as a discrete dynamical system. This is an example of an
7 Two-dimensional bifurcations
 7 Two-dimensional bifurcations As in one-dimensional systems: fixed points may be created, destroyed, or change stability as parameters are varied (change of topological equivalence ). In addition closed
7 Two-dimensional bifurcations As in one-dimensional systems: fixed points may be created, destroyed, or change stability as parameters are varied (change of topological equivalence ). In addition closed
CHALMERS, GÖTEBORGS UNIVERSITET. EXAM for DYNAMICAL SYSTEMS. COURSE CODES: TIF 155, FIM770GU, PhD
 CHALMERS, GÖTEBORGS UNIVERSITET EXAM for DYNAMICAL SYSTEMS COURSE CODES: TIF 155, FIM770GU, PhD Time: Place: Teachers: Allowed material: Not allowed: August 22, 2018, at 08 30 12 30 Johanneberg Jan Meibohm,
CHALMERS, GÖTEBORGS UNIVERSITET EXAM for DYNAMICAL SYSTEMS COURSE CODES: TIF 155, FIM770GU, PhD Time: Place: Teachers: Allowed material: Not allowed: August 22, 2018, at 08 30 12 30 Johanneberg Jan Meibohm,
Bifurcation and Stability Analysis of a Prey-predator System with a Reserved Area
 ISSN 746-733, England, UK World Journal of Modelling and Simulation Vol. 8 ( No. 4, pp. 85-9 Bifurcation and Stability Analysis of a Prey-predator System with a Reserved Area Debasis Mukherjee Department
ISSN 746-733, England, UK World Journal of Modelling and Simulation Vol. 8 ( No. 4, pp. 85-9 Bifurcation and Stability Analysis of a Prey-predator System with a Reserved Area Debasis Mukherjee Department
Parameter Sensitivity In A Lattice Ecosystem With Intraguild Predation
 Parameter Sensitivity In A Lattice Ecosystem With Intraguild Predation N. Nakagiri a, K. Tainaka a, T. Togashi b, T. Miyazaki b and J. Yoshimura a a Department of Systems Engineering, Shizuoka University,
Parameter Sensitivity In A Lattice Ecosystem With Intraguild Predation N. Nakagiri a, K. Tainaka a, T. Togashi b, T. Miyazaki b and J. Yoshimura a a Department of Systems Engineering, Shizuoka University,
8 Ecosystem stability
 8 Ecosystem stability References: May [47], Strogatz [48]. In these lectures we consider models of populations, with an emphasis on the conditions for stability and instability. 8.1 Dynamics of a single
8 Ecosystem stability References: May [47], Strogatz [48]. In these lectures we consider models of populations, with an emphasis on the conditions for stability and instability. 8.1 Dynamics of a single
Simplest Chaotic Flows with Involutional Symmetries
 International Journal of Bifurcation and Chaos, Vol. 24, No. 1 (2014) 1450009 (9 pages) c World Scientific Publishing Company DOI: 10.1142/S0218127414500096 Simplest Chaotic Flows with Involutional Symmetries
International Journal of Bifurcation and Chaos, Vol. 24, No. 1 (2014) 1450009 (9 pages) c World Scientific Publishing Company DOI: 10.1142/S0218127414500096 Simplest Chaotic Flows with Involutional Symmetries
Population Dynamics II
 Population Dynamics II In this class, we shall analyze behavioral patterns of ecosystems, in which more than two species interact with each other. Such systems frequently exhibit chaotic behavior. Chaotic
Population Dynamics II In this class, we shall analyze behavioral patterns of ecosystems, in which more than two species interact with each other. Such systems frequently exhibit chaotic behavior. Chaotic
CHALMERS, GÖTEBORGS UNIVERSITET. EXAM for DYNAMICAL SYSTEMS. COURSE CODES: TIF 155, FIM770GU, PhD
 CHALMERS, GÖTEBORGS UNIVERSITET EXAM for DYNAMICAL SYSTEMS COURSE CODES: TIF 155, FIM770GU, PhD Time: Place: Teachers: Allowed material: Not allowed: January 08, 2018, at 08 30 12 30 Johanneberg Kristian
CHALMERS, GÖTEBORGS UNIVERSITET EXAM for DYNAMICAL SYSTEMS COURSE CODES: TIF 155, FIM770GU, PhD Time: Place: Teachers: Allowed material: Not allowed: January 08, 2018, at 08 30 12 30 Johanneberg Kristian
Stability and bifurcation in a two species predator-prey model with quintic interactions
 Chaotic Modeling and Simulation (CMSIM) 4: 631 635, 2013 Stability and bifurcation in a two species predator-prey model with quintic interactions I. Kusbeyzi Aybar 1 and I. acinliyan 2 1 Department of
Chaotic Modeling and Simulation (CMSIM) 4: 631 635, 2013 Stability and bifurcation in a two species predator-prey model with quintic interactions I. Kusbeyzi Aybar 1 and I. acinliyan 2 1 Department of
COMPLEX DYNAMICS AND CHAOS CONTROL IN DUFFING-VAN DER POL EQUATION WITH TWO EXTERNAL PERIODIC FORCING TERMS
 International J. of Math. Sci. & Engg. Appls. (IJMSEA) ISSN 0973-9424, Vol. 9 No. III (September, 2015), pp. 197-210 COMPLEX DYNAMICS AND CHAOS CONTROL IN DUFFING-VAN DER POL EQUATION WITH TWO EXTERNAL
International J. of Math. Sci. & Engg. Appls. (IJMSEA) ISSN 0973-9424, Vol. 9 No. III (September, 2015), pp. 197-210 COMPLEX DYNAMICS AND CHAOS CONTROL IN DUFFING-VAN DER POL EQUATION WITH TWO EXTERNAL
Numerical Bifurcation Analysis of Dynamical Systems. Bob W. Kooi Faculty of Earth and Life Sciences,
 MRI Master Class 2009/2010: Numerical Bifurcation Analysis of Dynamical Systems WEEK 15, Apr 13 & 15: Bob W. Kooi Faculty of Earth and Life Sciences, Department of Theoretical Biology, Vrije University,
MRI Master Class 2009/2010: Numerical Bifurcation Analysis of Dynamical Systems WEEK 15, Apr 13 & 15: Bob W. Kooi Faculty of Earth and Life Sciences, Department of Theoretical Biology, Vrije University,
Lecture 6. Lorenz equations and Malkus' waterwheel Some properties of the Lorenz Eq.'s Lorenz Map Towards definitions of:
 Lecture 6 Chaos Lorenz equations and Malkus' waterwheel Some properties of the Lorenz Eq.'s Lorenz Map Towards definitions of: Chaos, Attractors and strange attractors Transient chaos Lorenz Equations
Lecture 6 Chaos Lorenz equations and Malkus' waterwheel Some properties of the Lorenz Eq.'s Lorenz Map Towards definitions of: Chaos, Attractors and strange attractors Transient chaos Lorenz Equations
Nonlinear dynamics & chaos BECS
 Nonlinear dynamics & chaos BECS-114.7151 Phase portraits Focus: nonlinear systems in two dimensions General form of a vector field on the phase plane: Vector notation: Phase portraits Solution x(t) describes
Nonlinear dynamics & chaos BECS-114.7151 Phase portraits Focus: nonlinear systems in two dimensions General form of a vector field on the phase plane: Vector notation: Phase portraits Solution x(t) describes
Transition to spatiotemporal chaos can resolve the paradox of enrichment
 Ecological Complexity 1 (24) 37 47 Transition to spatiotemporal chaos can resolve the paradox of enrichment Sergei Petrovskii a,, Bai-Lian Li b, Horst Malchow c a Shirshov Institute of Oceanology, Russian
Ecological Complexity 1 (24) 37 47 Transition to spatiotemporal chaos can resolve the paradox of enrichment Sergei Petrovskii a,, Bai-Lian Li b, Horst Malchow c a Shirshov Institute of Oceanology, Russian
SUPPLEMENTARY INFORMATION
 High-amplitude fluctuations and alternative dynamical states of midges in Lake Myvatn Anthony R. Ives 1, Árni Einarsson 2, Vincent A. A. Jansen 3, and Arnthor Gardarsson 2 1 Department of Zoology, UW-Madison,
High-amplitude fluctuations and alternative dynamical states of midges in Lake Myvatn Anthony R. Ives 1, Árni Einarsson 2, Vincent A. A. Jansen 3, and Arnthor Gardarsson 2 1 Department of Zoology, UW-Madison,
Fundamentals of Dynamical Systems / Discrete-Time Models. Dr. Dylan McNamara people.uncw.edu/ mcnamarad
 Fundamentals of Dynamical Systems / Discrete-Time Models Dr. Dylan McNamara people.uncw.edu/ mcnamarad Dynamical systems theory Considers how systems autonomously change along time Ranges from Newtonian
Fundamentals of Dynamical Systems / Discrete-Time Models Dr. Dylan McNamara people.uncw.edu/ mcnamarad Dynamical systems theory Considers how systems autonomously change along time Ranges from Newtonian
Relating generalized and specific modeling in population dynamical systems
 Relating generalized and specific modeling in population dynamical systems Acknowledgments: The research in this thesis has partly been supported by the DAAD-VIGONI program, and was carried out at the
Relating generalized and specific modeling in population dynamical systems Acknowledgments: The research in this thesis has partly been supported by the DAAD-VIGONI program, and was carried out at the
Stability Analysis of Uzawa-Lucas Endogenous Growth Model
 Abstract: Stability Analysis of Uzawa-Lucas Endogenous Growth Model William A. Barnett* University of Kansas, Lawrence and Center for Financial Stability, NY City and Taniya Ghosh Indira Gandhi Institute
Abstract: Stability Analysis of Uzawa-Lucas Endogenous Growth Model William A. Barnett* University of Kansas, Lawrence and Center for Financial Stability, NY City and Taniya Ghosh Indira Gandhi Institute
arxiv: v1 [nlin.cd] 20 Jul 2010
![arxiv: v1 [nlin.cd] 20 Jul 2010](/thumbs/92/109497388.jpg "arxiv: v1 [nlin.cd] 20 Jul 2010") Invariant manifolds of the Bonhoeffer-van der Pol oscillator arxiv:1007.3375v1 [nlin.cd] 20 Jul 2010 R. Benítez 1, V. J. Bolós 2 1 Departamento de Matemáticas, Centro Universitario de Plasencia, Universidad
Invariant manifolds of the Bonhoeffer-van der Pol oscillator arxiv:1007.3375v1 [nlin.cd] 20 Jul 2010 R. Benítez 1, V. J. Bolós 2 1 Departamento de Matemáticas, Centro Universitario de Plasencia, Universidad
One Dimensional Dynamical Systems
 16 CHAPTER 2 One Dimensional Dynamical Systems We begin by analyzing some dynamical systems with one-dimensional phase spaces, and in particular their bifurcations. All equations in this Chapter are scalar
16 CHAPTER 2 One Dimensional Dynamical Systems We begin by analyzing some dynamical systems with one-dimensional phase spaces, and in particular their bifurcations. All equations in this Chapter are scalar
... it may happen that small differences in the initial conditions produce very great ones in the final phenomena. Henri Poincaré
 Chapter 2 Dynamical Systems... it may happen that small differences in the initial conditions produce very great ones in the final phenomena. Henri Poincaré One of the exciting new fields to arise out
Chapter 2 Dynamical Systems... it may happen that small differences in the initial conditions produce very great ones in the final phenomena. Henri Poincaré One of the exciting new fields to arise out
A Novel Three Dimension Autonomous Chaotic System with a Quadratic Exponential Nonlinear Term
 ETASR - Engineering, Technology & Applied Science Research Vol., o.,, 9-5 9 A Novel Three Dimension Autonomous Chaotic System with a Quadratic Exponential Nonlinear Term Fei Yu College of Information Science
ETASR - Engineering, Technology & Applied Science Research Vol., o.,, 9-5 9 A Novel Three Dimension Autonomous Chaotic System with a Quadratic Exponential Nonlinear Term Fei Yu College of Information Science
Complex dynamics of a three species food-chain model with Holling type IV functional response
 Nonlinear Analysis: Modelling and Control, 211, Vol. 16, No. 3, 353 374 353 Complex dynamics of a three species food-chain model with Holling type IV functional response Ranjit Kumar Upadhyay, Sharada
Nonlinear Analysis: Modelling and Control, 211, Vol. 16, No. 3, 353 374 353 Complex dynamics of a three species food-chain model with Holling type IV functional response Ranjit Kumar Upadhyay, Sharada
Physics: spring-mass system, planet motion, pendulum. Biology: ecology problem, neural conduction, epidemics
 Applications of nonlinear ODE systems: Physics: spring-mass system, planet motion, pendulum Chemistry: mixing problems, chemical reactions Biology: ecology problem, neural conduction, epidemics Economy:
Applications of nonlinear ODE systems: Physics: spring-mass system, planet motion, pendulum Chemistry: mixing problems, chemical reactions Biology: ecology problem, neural conduction, epidemics Economy:
B5.6 Nonlinear Systems
 B5.6 Nonlinear Systems 5. Global Bifurcations, Homoclinic chaos, Melnikov s method Alain Goriely 2018 Mathematical Institute, University of Oxford Table of contents 1. Motivation 1.1 The problem 1.2 A
B5.6 Nonlinear Systems 5. Global Bifurcations, Homoclinic chaos, Melnikov s method Alain Goriely 2018 Mathematical Institute, University of Oxford Table of contents 1. Motivation 1.1 The problem 1.2 A
Chaos. Dr. Dylan McNamara people.uncw.edu/mcnamarad
 Chaos Dr. Dylan McNamara people.uncw.edu/mcnamarad Discovery of chaos Discovered in early 1960 s by Edward N. Lorenz (in a 3-D continuous-time model) Popularized in 1976 by Sir Robert M. May as an example
Chaos Dr. Dylan McNamara people.uncw.edu/mcnamarad Discovery of chaos Discovered in early 1960 s by Edward N. Lorenz (in a 3-D continuous-time model) Popularized in 1976 by Sir Robert M. May as an example
STABILITY. Phase portraits and local stability
 MAS271 Methods for differential equations Dr. R. Jain STABILITY Phase portraits and local stability We are interested in system of ordinary differential equations of the form ẋ = f(x, y), ẏ = g(x, y),
MAS271 Methods for differential equations Dr. R. Jain STABILITY Phase portraits and local stability We are interested in system of ordinary differential equations of the form ẋ = f(x, y), ẏ = g(x, y),
On the Takens-Bogdanov Bifurcation in the Chua s Equation
 1722 PAPER Special Section on Nonlinear Theory and Its Applications On the Takens-Bogdanov Bifurcation in the Chua s Equation Antonio ALGABA, Emilio FREIRE, Estanislao GAMERO, and Alejandro J. RODRÍGUEZ-LUIS,
1722 PAPER Special Section on Nonlinear Theory and Its Applications On the Takens-Bogdanov Bifurcation in the Chua s Equation Antonio ALGABA, Emilio FREIRE, Estanislao GAMERO, and Alejandro J. RODRÍGUEZ-LUIS,
DIFFERENT TYPES OF CRITICAL BEHAVIOR IN CONSERVATIVELY COUPLED HÉNON MAPS
 DIFFERENT TYPES OF CRITICAL BEHAVIOR IN CONSERVATIVELY COUPLED HÉNON MAPS D.V. Savin, *, A.P. Kuznetsov,, A.V. Savin and U. Feudel 3 Department of Nonlinear Processes, ernyshevsky Saratov State University,
DIFFERENT TYPES OF CRITICAL BEHAVIOR IN CONSERVATIVELY COUPLED HÉNON MAPS D.V. Savin, *, A.P. Kuznetsov,, A.V. Savin and U. Feudel 3 Department of Nonlinear Processes, ernyshevsky Saratov State University,
2 One-dimensional models in discrete time
 2 One-dimensional models in discrete time So far, we have assumed that demographic events happen continuously over time and can thus be written as rates. For many biological species with overlapping generations
2 One-dimensional models in discrete time So far, we have assumed that demographic events happen continuously over time and can thus be written as rates. For many biological species with overlapping generations
Bifurcation Analysis, Chaos and Control in the Burgers Mapping
 ISSN 1749-3889 print, 1749-3897 online International Journal of Nonlinear Science Vol.4007 No.3,pp.171-185 Bifurcation Analysis, Chaos and Control in the Burgers Mapping E. M. ELabbasy, H. N. Agiza, H.
ISSN 1749-3889 print, 1749-3897 online International Journal of Nonlinear Science Vol.4007 No.3,pp.171-185 Bifurcation Analysis, Chaos and Control in the Burgers Mapping E. M. ELabbasy, H. N. Agiza, H.
Nonlinear Dynamics. Moreno Marzolla Dip. di Informatica Scienza e Ingegneria (DISI) Università di Bologna.
 Università di Bologna.") Nonlinear Dynamics Moreno Marzolla Dip. di Informatica Scienza e Ingegneria (DISI) Università di Bologna http://www.moreno.marzolla.name/ 2 Introduction: Dynamics of Simple Maps 3 Dynamical systems A dynamical
Nonlinear Dynamics Moreno Marzolla Dip. di Informatica Scienza e Ingegneria (DISI) Università di Bologna http://www.moreno.marzolla.name/ 2 Introduction: Dynamics of Simple Maps 3 Dynamical systems A dynamical
Student Background Readings for Bug Dynamics: The Logistic Map
 Student Background Readings for Bug Dynamics: The Logistic Map Figure 1 The population as a function of a growth rate parameter observed in the experiment of [Bugs]. Graphs such as this are called bifurcation
Student Background Readings for Bug Dynamics: The Logistic Map Figure 1 The population as a function of a growth rate parameter observed in the experiment of [Bugs]. Graphs such as this are called bifurcation
Introduction to Applied Nonlinear Dynamical Systems and Chaos
 Stephen Wiggins Introduction to Applied Nonlinear Dynamical Systems and Chaos Second Edition With 250 Figures 4jj Springer I Series Preface v L I Preface to the Second Edition vii Introduction 1 1 Equilibrium
Stephen Wiggins Introduction to Applied Nonlinear Dynamical Systems and Chaos Second Edition With 250 Figures 4jj Springer I Series Preface v L I Preface to the Second Edition vii Introduction 1 1 Equilibrium
SPATIOTEMPORAL CHAOS IN COUPLED MAP LATTICE. Itishree Priyadarshini. Prof. Biplab Ganguli
 SPATIOTEMPORAL CHAOS IN COUPLED MAP LATTICE By Itishree Priyadarshini Under the Guidance of Prof. Biplab Ganguli Department of Physics National Institute of Technology, Rourkela CERTIFICATE This is to
SPATIOTEMPORAL CHAOS IN COUPLED MAP LATTICE By Itishree Priyadarshini Under the Guidance of Prof. Biplab Ganguli Department of Physics National Institute of Technology, Rourkela CERTIFICATE This is to
Invariant manifolds of the Bonhoeffer-van der Pol oscillator
 Invariant manifolds of the Bonhoeffer-van der Pol oscillator R. Benítez 1, V. J. Bolós 2 1 Dpto. Matemáticas, Centro Universitario de Plasencia, Universidad de Extremadura. Avda. Virgen del Puerto 2. 10600,
Invariant manifolds of the Bonhoeffer-van der Pol oscillator R. Benítez 1, V. J. Bolós 2 1 Dpto. Matemáticas, Centro Universitario de Plasencia, Universidad de Extremadura. Avda. Virgen del Puerto 2. 10600,
Stabilization through spatial pattern formation in metapopulations with long-range dispersal
 Stabilization through spatial pattern formation in metapopulations with long-range dispersal Michael Doebeli 1 and Graeme D. Ruxton 2 1 Zoology Institute, University of Basel, Rheinsprung 9, CH^4051 Basel,
Stabilization through spatial pattern formation in metapopulations with long-range dispersal Michael Doebeli 1 and Graeme D. Ruxton 2 1 Zoology Institute, University of Basel, Rheinsprung 9, CH^4051 Basel,
Dynamical Systems and Chaos Part II: Biology Applications. Lecture 6: Population dynamics. Ilya Potapov Mathematics Department, TUT Room TD325
 Dynamical Systems and Chaos Part II: Biology Applications Lecture 6: Population dynamics Ilya Potapov Mathematics Department, TUT Room TD325 Living things are dynamical systems Dynamical systems theory
Dynamical Systems and Chaos Part II: Biology Applications Lecture 6: Population dynamics Ilya Potapov Mathematics Department, TUT Room TD325 Living things are dynamical systems Dynamical systems theory
Lattice models of habitat destruction in a prey-predator system
 22nd International Congress on Modelling and Simulation, Hobart, Tasmania, Australia, 3 to 8 December 2017 mssanz.org.au/modsim2017 Lattice models of habitat destruction in a prey-predator system Nariiyuki
22nd International Congress on Modelling and Simulation, Hobart, Tasmania, Australia, 3 to 8 December 2017 mssanz.org.au/modsim2017 Lattice models of habitat destruction in a prey-predator system Nariiyuki
arxiv: v2 [q-bio.pe] 20 Feb 2009
![arxiv: v2 [q-bio.pe] 20 Feb 2009](/thumbs/83/88707739.jpg "arxiv: v2 [q-bio.pe] 20 Feb 2009") Joint effects of nutrients and contaminants on the dynamics of a food chain in marine ecosystems arxiv:0803.0624v2 [q-bio.pe] 20 Feb 2009 Flora S. Bacelar, a Sibylle Dueri, b Emilio Hernández-García, a,
Joint effects of nutrients and contaminants on the dynamics of a food chain in marine ecosystems arxiv:0803.0624v2 [q-bio.pe] 20 Feb 2009 Flora S. Bacelar, a Sibylle Dueri, b Emilio Hernández-García, a,
Dynamical Systems and Chaos Part I: Theoretical Techniques. Lecture 4: Discrete systems + Chaos. Ilya Potapov Mathematics Department, TUT Room TD325
 Dynamical Systems and Chaos Part I: Theoretical Techniques Lecture 4: Discrete systems + Chaos Ilya Potapov Mathematics Department, TUT Room TD325 Discrete maps x n+1 = f(x n ) Discrete time steps. x 0
Dynamical Systems and Chaos Part I: Theoretical Techniques Lecture 4: Discrete systems + Chaos Ilya Potapov Mathematics Department, TUT Room TD325 Discrete maps x n+1 = f(x n ) Discrete time steps. x 0
Lecture 20/Lab 21: Systems of Nonlinear ODEs
 Lecture 20/Lab 21: Systems of Nonlinear ODEs MAR514 Geoffrey Cowles Department of Fisheries Oceanography School for Marine Science and Technology University of Massachusetts-Dartmouth Coupled ODEs: Species
Lecture 20/Lab 21: Systems of Nonlinear ODEs MAR514 Geoffrey Cowles Department of Fisheries Oceanography School for Marine Science and Technology University of Massachusetts-Dartmouth Coupled ODEs: Species
arxiv: v1 [nlin.ao] 19 May 2017
![arxiv: v1 [nlin.ao] 19 May 2017](/thumbs/74/70757831.jpg "arxiv: v1 [nlin.ao] 19 May 2017") Feature-rich bifurcations in a simple electronic circuit Debdipta Goswami 1, and Subhankar Ray 2, 1 Department of Electrical and Computer Engineering, University of Maryland, College Park, MD 20742, USA
Feature-rich bifurcations in a simple electronic circuit Debdipta Goswami 1, and Subhankar Ray 2, 1 Department of Electrical and Computer Engineering, University of Maryland, College Park, MD 20742, USA
Reliable Computation of Equilibrium States and Bifurcations in Food Chain Models
 Reliable Computation of Equilibrium States and Bifurcations in Food Chain Models C. Ryan Gwaltney, Mark P. Styczynski, and Mark A. Stadtherr * Department of Chemical and Biomolecular Engineering University
Reliable Computation of Equilibrium States and Bifurcations in Food Chain Models C. Ryan Gwaltney, Mark P. Styczynski, and Mark A. Stadtherr * Department of Chemical and Biomolecular Engineering University
Additive resonances of a controlled van der Pol-Duffing oscillator
 Additive resonances of a controlled van der Pol-Duffing oscillator This paper has been published in Journal of Sound and Vibration vol. 5 issue - 8 pp.-. J.C. Ji N. Zhang Faculty of Engineering University
Additive resonances of a controlled van der Pol-Duffing oscillator This paper has been published in Journal of Sound and Vibration vol. 5 issue - 8 pp.-. J.C. Ji N. Zhang Faculty of Engineering University
Spatiotemporal pattern formation in a prey-predator model under environmental driving forces
 Home Search Collections Journals About Contact us My IOPscience Spatiotemporal pattern formation in a prey-predator model under environmental driving forces This content has been downloaded from IOPscience.
Home Search Collections Journals About Contact us My IOPscience Spatiotemporal pattern formation in a prey-predator model under environmental driving forces This content has been downloaded from IOPscience.
Bifurcation and chaos in simple jerk dynamical systems
 PRAMANA c Indian Academy of Sciences Vol. 64, No. 1 journal of January 2005 physics pp. 75 93 Bifurcation and chaos in simple jerk dynamical systems VINOD PATIDAR and K K SUD Department of Physics, College
PRAMANA c Indian Academy of Sciences Vol. 64, No. 1 journal of January 2005 physics pp. 75 93 Bifurcation and chaos in simple jerk dynamical systems VINOD PATIDAR and K K SUD Department of Physics, College
Numerical bifurcation analysis of a tri-trophic food web with omnivory
 Mathematical Biosciences 177&178 (2002) 201 228 www.elsevier.com/locate/mbs Numerical bifurcation analysis of a tri-trophic food web with omnivory B.W. Kooi *, L.D.J. Kuijper, M.P. Boer 1, S.A.L.M. Kooijman
Mathematical Biosciences 177&178 (2002) 201 228 www.elsevier.com/locate/mbs Numerical bifurcation analysis of a tri-trophic food web with omnivory B.W. Kooi *, L.D.J. Kuijper, M.P. Boer 1, S.A.L.M. Kooijman
The influence of noise on two- and three-frequency quasi-periodicity in a simple model system
 arxiv:1712.06011v1 [nlin.cd] 16 Dec 2017 The influence of noise on two- and three-frequency quasi-periodicity in a simple model system A.P. Kuznetsov, S.P. Kuznetsov and Yu.V. Sedova December 19, 2017
arxiv:1712.06011v1 [nlin.cd] 16 Dec 2017 The influence of noise on two- and three-frequency quasi-periodicity in a simple model system A.P. Kuznetsov, S.P. Kuznetsov and Yu.V. Sedova December 19, 2017
Nonlinear Dynamics and Chaos Summer 2011
 67-717 Nonlinear Dynamics and Chaos Summer 2011 Instructor: Zoubir Benzaid Phone: 424-7354 Office: Swart 238 Office Hours: MTWR: 8:30-9:00; MTWR: 12:00-1:00 and by appointment. Course Content: This course
67-717 Nonlinear Dynamics and Chaos Summer 2011 Instructor: Zoubir Benzaid Phone: 424-7354 Office: Swart 238 Office Hours: MTWR: 8:30-9:00; MTWR: 12:00-1:00 and by appointment. Course Content: This course
A Simple Model of Evolution with Variable System Size
 A Simple Model of Evolution with Variable System Size Claus Wilke and Thomas Martinetz Institut für Neuroinformatik Ruhr-Universität Bochum (Submitted: ; Printed: September 28, 2001) A simple model of
A Simple Model of Evolution with Variable System Size Claus Wilke and Thomas Martinetz Institut für Neuroinformatik Ruhr-Universität Bochum (Submitted: ; Printed: September 28, 2001) A simple model of
Towards a Global Theory of Singularly Perturbed Dynamical Systems John Guckenheimer Cornell University
 Towards a Global Theory of Singularly Perturbed Dynamical Systems John Guckenheimer Cornell University Dynamical systems with multiple time scales arise naturally in many domains. Models of neural systems
Towards a Global Theory of Singularly Perturbed Dynamical Systems John Guckenheimer Cornell University Dynamical systems with multiple time scales arise naturally in many domains. Models of neural systems
Difference Resonances in a controlled van der Pol-Duffing oscillator involving time. delay
 Difference Resonances in a controlled van der Pol-Duffing oscillator involving time delay This paper was published in the journal Chaos, Solitions & Fractals, vol.4, no., pp.975-98, Oct 9 J.C. Ji, N. Zhang,
Difference Resonances in a controlled van der Pol-Duffing oscillator involving time delay This paper was published in the journal Chaos, Solitions & Fractals, vol.4, no., pp.975-98, Oct 9 J.C. Ji, N. Zhang,
Continuous Threshold Policy Harvesting in Predator-Prey Models
 Continuous Threshold Policy Harvesting in Predator-Prey Models Jon Bohn and Kaitlin Speer Department of Mathematics, University of Wisconsin - Madison Department of Mathematics, Baylor University July
Continuous Threshold Policy Harvesting in Predator-Prey Models Jon Bohn and Kaitlin Speer Department of Mathematics, University of Wisconsin - Madison Department of Mathematics, Baylor University July
Lesson 4: Non-fading Memory Nonlinearities
 Lesson 4: Non-fading Memory Nonlinearities Nonlinear Signal Processing SS 2017 Christian Knoll Signal Processing and Speech Communication Laboratory Graz University of Technology June 22, 2017 NLSP SS
Lesson 4: Non-fading Memory Nonlinearities Nonlinear Signal Processing SS 2017 Christian Knoll Signal Processing and Speech Communication Laboratory Graz University of Technology June 22, 2017 NLSP SS
Antagonistic and Synergistic Interactions Among Predators
 Bulletin of Mathematical Biology 2007 69: 2093 2104 DOI 10.1007/s11538-007-9214-0 ORIGINAL ARTICLE Antagonistic and Synergistic Interactions Among Predators Gary R. Huxel Department of Biological Sciences,
Bulletin of Mathematical Biology 2007 69: 2093 2104 DOI 10.1007/s11538-007-9214-0 ORIGINAL ARTICLE Antagonistic and Synergistic Interactions Among Predators Gary R. Huxel Department of Biological Sciences,
Turing and Non-Turing patterns in diffusive plankton model
 Computational Ecology and Software 05 5(): 6-7 Article Turing and Non-Turing patterns in diffusive plankton model N. K. Thakur R. Gupta R. K. Upadhyay National Institute of Technology Raipur India Indian
Computational Ecology and Software 05 5(): 6-7 Article Turing and Non-Turing patterns in diffusive plankton model N. K. Thakur R. Gupta R. K. Upadhyay National Institute of Technology Raipur India Indian
High-Dimensional Dynamics in the Delayed Hénon Map
 EJTP 3, No. 12 (2006) 19 35 Electronic Journal of Theoretical Physics High-Dimensional Dynamics in the Delayed Hénon Map J. C. Sprott Department of Physics, University of Wisconsin, Madison, WI 53706,
EJTP 3, No. 12 (2006) 19 35 Electronic Journal of Theoretical Physics High-Dimensional Dynamics in the Delayed Hénon Map J. C. Sprott Department of Physics, University of Wisconsin, Madison, WI 53706,
CONTROL OF POWER SYSTEMS WITH FACTS DEVICES CONSIDERING DIFFERENT LOAD CHARACTERISTICS
 CONTROL OF POWER SYSTEMS WITH FACTS DEVICES CONSIDERING DIFFERENT LOAD CHARACTERISTICS Ingo Winzenick *, Michael Fette **, Joachim Horn * * Helmut-Schmidt-University / University of the Federal Armed Forces
CONTROL OF POWER SYSTEMS WITH FACTS DEVICES CONSIDERING DIFFERENT LOAD CHARACTERISTICS Ingo Winzenick *, Michael Fette **, Joachim Horn * * Helmut-Schmidt-University / University of the Federal Armed Forces
Behaviour of simple population models under ecological processes
 J. Biosci., Vol. 19, Number 2, June 1994, pp 247 254. Printed in India. Behaviour of simple population models under ecological processes SOMDATTA SINHA* and S PARTHASARATHY Centre for Cellular and Molecular
J. Biosci., Vol. 19, Number 2, June 1994, pp 247 254. Printed in India. Behaviour of simple population models under ecological processes SOMDATTA SINHA* and S PARTHASARATHY Centre for Cellular and Molecular
Edward Lorenz. Professor of Meteorology at the Massachusetts Institute of Technology
 The Lorenz system Edward Lorenz Professor of Meteorology at the Massachusetts Institute of Technology In 1963 derived a three dimensional system in efforts to model long range predictions for the weather
The Lorenz system Edward Lorenz Professor of Meteorology at the Massachusetts Institute of Technology In 1963 derived a three dimensional system in efforts to model long range predictions for the weather
MULTISTABILITY IN A BUTTERFLY FLOW
 International Journal of Bifurcation and Chaos, Vol. 23, No. 12 (2013) 1350199 (10 pages) c World Scientific Publishing Company DOI: 10.1142/S021812741350199X MULTISTABILITY IN A BUTTERFLY FLOW CHUNBIAO
International Journal of Bifurcation and Chaos, Vol. 23, No. 12 (2013) 1350199 (10 pages) c World Scientific Publishing Company DOI: 10.1142/S021812741350199X MULTISTABILITY IN A BUTTERFLY FLOW CHUNBIAO
Semi-analytical solutions for cubic autocatalytic reaction-diffusion equations; the effect of a precursor chemical
 ANZIAM J. 53 (EMAC211) pp.c511 C524, 212 C511 Semi-analytical solutions for cubic autocatalytic reaction-diffusion equations; the effect of a precursor chemical M. R. Alharthi 1 T. R. Marchant 2 M. I.
ANZIAM J. 53 (EMAC211) pp.c511 C524, 212 C511 Semi-analytical solutions for cubic autocatalytic reaction-diffusion equations; the effect of a precursor chemical M. R. Alharthi 1 T. R. Marchant 2 M. I.
Stability of Ecosystem Induced by Mutual Interference between Predators
 Available online at www.sciencedirect.com Procedia Environmental Sciences (00) 4 48 International Society for Environmental Information Sciences 00 Annual Conference (ISEIS) Stability of Ecosystem Induced
Available online at www.sciencedirect.com Procedia Environmental Sciences (00) 4 48 International Society for Environmental Information Sciences 00 Annual Conference (ISEIS) Stability of Ecosystem Induced
Recent new examples of hidden attractors
 Eur. Phys. J. Special Topics 224, 1469 1476 (2015) EDP Sciences, Springer-Verlag 2015 DOI: 10.1140/epjst/e2015-02472-1 THE EUROPEAN PHYSICAL JOURNAL SPECIAL TOPICS Review Recent new examples of hidden
Eur. Phys. J. Special Topics 224, 1469 1476 (2015) EDP Sciences, Springer-Verlag 2015 DOI: 10.1140/epjst/e2015-02472-1 THE EUROPEAN PHYSICAL JOURNAL SPECIAL TOPICS Review Recent new examples of hidden
Is chaos possible in 1d? - yes - no - I don t know. What is the long term behavior for the following system if x(0) = π/2?
 = π/2?") Is chaos possible in 1d? - yes - no - I don t know What is the long term behavior for the following system if x(0) = π/2? In the insect outbreak problem, what kind of bifurcation occurs at fixed value
Is chaos possible in 1d? - yes - no - I don t know What is the long term behavior for the following system if x(0) = π/2? In the insect outbreak problem, what kind of bifurcation occurs at fixed value
Lotka Volterra Predator-Prey Model with a Predating Scavenger
 Lotka Volterra Predator-Prey Model with a Predating Scavenger Monica Pescitelli Georgia College December 13, 2013 Abstract The classic Lotka Volterra equations are used to model the population dynamics
Lotka Volterra Predator-Prey Model with a Predating Scavenger Monica Pescitelli Georgia College December 13, 2013 Abstract The classic Lotka Volterra equations are used to model the population dynamics
Dynamics on a General Stage Structured n Parallel Food Chains
 Memorial University of Newfoundland Dynamics on a General Stage Structured n Parallel Food Chains Isam Al-Darabsah and Yuan Yuan November 4, 2013 Outline: Propose a general model with n parallel food chains
Memorial University of Newfoundland Dynamics on a General Stage Structured n Parallel Food Chains Isam Al-Darabsah and Yuan Yuan November 4, 2013 Outline: Propose a general model with n parallel food chains
Bifurcation in a Discrete Two Patch Logistic Metapopulation Model
 Bifurcation in a Discrete Two Patch Logistic Metapopulation Model LI XU Tianjin University of Commerce Department of Statistics Jinba road, Benchen,Tianjin CHINA beifang xl@63.com ZHONGXIANG CHANG Tianjin
Bifurcation in a Discrete Two Patch Logistic Metapopulation Model LI XU Tianjin University of Commerce Department of Statistics Jinba road, Benchen,Tianjin CHINA beifang xl@63.com ZHONGXIANG CHANG Tianjin