Nerve Excitation. L. David Roper This is web page
|
|
|
- Violet Austin
- 6 years ago
- Views:
Transcription
 is shown in Figure 1. Figure 1. A typical nerve axon action potential (http://en.wikipedia.")
1 Nerve Excitation L. David Roper This is web page Chapter 1. The Action Potential A typical action potential (inside relative to outside) is shown in Figure 1. Figure 1. A typical nerve axon action potential ( 1
2 The explanation for the action potential, which once generated travels down the nerve axon with a speed of 10 to 100 meters/second, is as follows: A resting electric potential of about -70 millivolts (mv) inside relative to outside exists due to approximate concentration differences across the squid giant-axon lipid membrane as follows: o 400 millimoles/liter (mm/l) inside and 20 mm/l outside for + ions. o 50 mm/l inside and 440 mm/l outside for Na + ions. o About 100 mm/l inside and 560 mm/l outside for Cl - ions. o There are other ions present, such as Ca ++, with concentration gradients across the membrane. o The main cause of the -65 mv resting potential is that the membrane is mainly permeable to + ions at rest, so that some + ions migrate to the outside setting up the potential. An applied positive transient electric potential inside relative to outside above a threshold value (about 40 mv) triggers a brief opening of a channel that allows Na + ions to pass into the axon, causing the membrane potential to transiently move positively. That is followed by a brief opening of a channel that allows a greater flow of + ions outside the axon, which typically causes an overshoot below the resting potential Eventually the membrane potential restores to the resting potential, unless the stimulating potential is maintained, in which case, after a refractory period, it is repeated. The time dependence of the action potential across the excitable membrane of the axon of a nerve cell can be represented by a combination of several hyperbolic tangents. (See and The opening of the two ion channels can be represented mathematically by two double hyperbolic tangents: 1 t t Na1 t t Na 2 1 t t 1 t t 2 V () t = Vr + CNa tanh tanh + C tanh tanh, 2 wna1 wna 2 2 w1 w2 where V r = + resting potential. (1.1) The hyperbolic-tangent function is a natural function to use for situations such as nerve excitation where a system changes from one state to another. In this case both the sodium and potassium ions have molecular channels across the axon membrane which change from one conductance state to another and then back to the original conductance state. The time interval between the turning on and off of the middle state may be so small that the full conductance is never realized because there are time constants (inverse of the rates) for turning on and off a conductance state. That is, there is a time constant for all channels changing from one state to another. It is more physical to use the hyperbolic-tangent function than the exponential function for state changes, because an exponential rise or fall cannot continue forever. Eventually there must be an asymptote, which the hyperbolic tangent provides. 2

3 An example of a calculated action potential is shown in Figure 2. Figure 2. Example of a calculated action potential using two double hyperbolic tangents for the two ion channels. The two double hyperbolic tangents are shown along with the sum of them. The parameters for this curve are given in Table 1. Table 1. Action potential fit parameters. V r = -70 C t 1 w 1 t 2 w 2 Na parameters: parameters: This indicates that the and Na ion-channels transitions that cause the nerve-membrane action potential can be well represented by the hyperbolic tangent. 3

4 One can create many different shapes for action potentials by varying the ten parameters in the equation. Figure 3 shows an example of a different shape. Figure 3. Example of a calculated action potential using two double hyperbolic tangents for the two ion channels. The two double hyperbolic tangents are shown along with the sum of them. The parameters for this curve are given in Table 2. Table 2. Action potential parameters. V r = -70 C t 1 w 1 t 2 w 2 Na parameters: parameters: These curves do not contain the transient stimulus electric potential of about +40 mv. It can be subtracted out from the measurement. It could be added to the equation as another double hyperbolic tangent. If more ions are involved with channels opening, such as Ca ++, they can be added as another double hyperbolic tangent. It would be interesting to try to obtain an approximation to the Hodgkin-Huxley equations ( for the action potential by mathematically manipulating the equation given here. 4
5 Chapter 2. Voltage-Clamp Data Experiments can be arranged for the squid giant axon that fix the electric potential (voltage) across the membrane at different values. ( I use the data shown in Figure 7 on p. 36 of Hille (Hille, 1984). The data are for a membrane resting potential of 60 mv, opposite of the convention used in the action-potential calculation above. As Hille describes, it is well known that the current density (current/area = ma/cm 2 ) consists of a potassium current density (J ) and a sodium current density (J Na ). Here the former is parameterized by a hyperbolic tangent and the latter by two hyperbolic tangents, similar to what was used in the actionpotential calculation above: ( ) = ( ) ( ) + ( ) ( ) ( ) J t, V J V tanh r V t JNa V tanh rna1 V t tanh rna2 V t. (1.2) (Note that the initial time is t=0 and tanh(at)=0 when t=0.) The voltage dependence of the parameters is indicated by, for example, r ( ) V. That is, the current density changes from one value to another and the Na current density transiently changes negatively, but then goes back to its original value. Another way to say it is that more channels in the membrane open up and Na channels transiently open. Seven different voltages were fitted with this equation with five types of parameters, yielding the 35 parameters given in Table 3. Table 3. Fit parameters at the different voltages for Eq. (1.2). Potential -30 mv -10 mv 10 mv 30 mv 50 mv 70 mv 90 mv J ( ) V r ( ) V J ( ) Na V r ( ) Na1 V r ( ) Na2 V Chi Sq The total Chi Square is for fitting 224 data with 35 parameters. The fit was done by using the Solver routine in the Excel spreadsheet to minimize least squares between the data array and the array calculated using the fitting parameters. 5

6 The fits are shown in Figure 4. Figure 4. Voltage-clamp data for the squid giant axon for seven voltages and fits to them. These currents are recorded as positive going out of the axon, opposite the convention for the action potential data given above. 6


7 The individual voltage fits are shown in Figure 5. 7
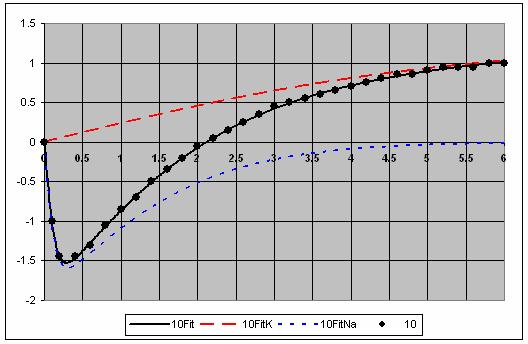
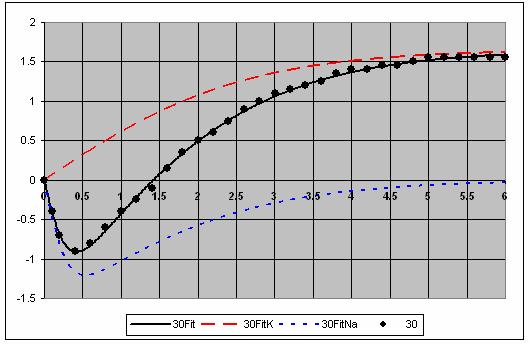
8 8
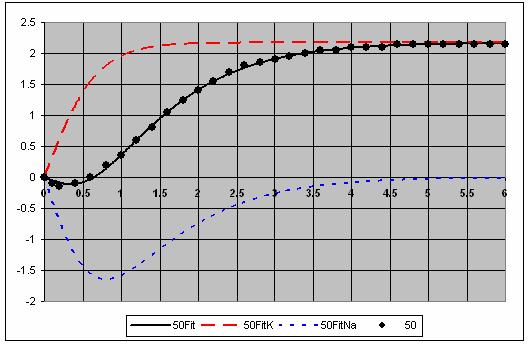
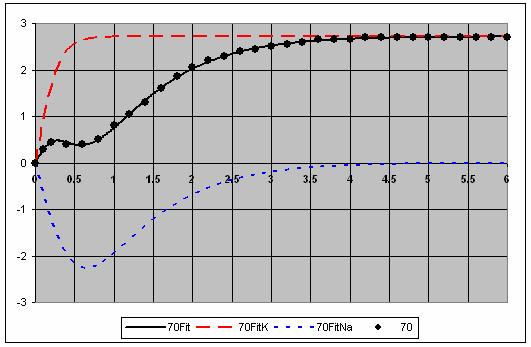
9 9

10 Figure 5. Fits for each voltage showing the and Na components. This is a very good fit to the data. 10
 points out that the fit parameters are more mathematically stable as a function of V if one factors ( V V ) out of the potassium current and ( V V Na ) out of the")
11 The 35 fit parameters as a function of the clamping voltage are shown in Figure 6. Figure 6. Fit parameters versus clamping voltage. The Nfactor ( J Na ) scale is on the right. However, Cronin (Cronin, 1987) points out that the fit parameters are more mathematically stable as a function of V if one factors ( V V ) out of the potassium current and ( V V Na ) out of the sodium current, where V is the potassium Nernst potential ( 72 mv ) and V Na is the sodium Nernst potential ( + 55 mv ), outside relative to inside. Then the equation for the current density is: J( t, V) = J ( ){ ( ) ( ) ( )( ( ) V tanh r V t V + tanh r V t t V ) }( V V) tanh r ( ) ( )( ( ) Na1 V tn1 tanh rna 1 V t tn1 V ) ( ) + + JNa V ( V VNa ). tanh r ( ) ( )( ( ) Na2 V tn 2 tanh rna2 V t tn 2 V ) The voltage dependence of the parameters is indicated by, for example, r ( ) V. The two hyperbolictangent terms insure that the and Na currents are zero at t = 0 and that the behavior at t = 0 can be exponential. (See Appendix 1.) This is a slight improvement over Eq. (1.2). (1.3) The leakage current is neglected because it is very small after about 0.05 milliseconds. (Malmivuo, 1995 Ch. 4.) 11
12 The values of the eight types of fit parameters for seven voltages are given in Table 4. Table 4. The 56 fit parameters at the different voltages in Eq. (1.3). Potential J ( ) V r ( ) V t ( ) V J ( ) Na V r ( ) N1 V t ( ) N1 V r ( ) N2 V t ( ) N2 V The χ 2 of the least-squares fit is for fitting 224 data with 56 parameters. For those who might think that it easy to fit the 224 data with 56 parameters, I assure you that it is not easy! There are many local minima that must be negotiated around. 12


13 The 56 fit parameters as a function of the clamping voltage are shown in Figure 7. Figure 7. Fit parameters versus clamping voltage. Right arrows indicate parameters for the right-hand scale. These parameters as a function of voltage appear to be changing through several states. 13

14 It is interesting to see the fit to the data when assuming that there is no voltage dependence for the 35 parameters. The fit parameters are given in Table 5 and the fit is shown in Figure 8. Table 5. The 8 fit parameters assuming no voltage dependence. J r t J Na r N1 t N1 r N2 t N Figure 8. Fit of the data by 35 parameters that are not voltage dependent. 2 The fit χ is 18.13, obviously not a very good fit. Voltage dependence besides the ( V V ) and ( V V ) factors is Eq. (1.3) is required. Na 14
15 The voltage dependence of the five types of fit parameters shown in Figure 7 is intriguing. I fitted them very precisely with functions of the voltage by varying 80 parameters as given in Table 6. J r t JNa rna1 tna1 rna2 tna2 ( V ) ( V ) ( V ) ( V ) ( V ) ( V ) ( V ) ( V ) Table 6. Voltage-dependent equations for the 80 parameters of Eq. (1.3). ( ) ( ) ( ) a + b1 tanh r1 V V1 + b2 tanh r2 V V2 + b3 tanh r3 V V3 (10 parameters) c + d1 tanh r4( V V4) + d2 tanh r5( V V5) (7 parameters) e + f1 tanh r6( V V6) + f2 tanh r7( V V7) + f3 tanh r8( V V8) (10 parameters) an + bn1 tanh rn1( V VN1) + bn2 tanh rn2( V VN2) + bn3 tanh rn3( V VN3) + bn4 tanh rn4( V VN4) (13 parameters) ( ) tanh ( ) cn1+ dn1 tanh rn5 V VN5 + dn2 rn6 V VN6 (7 parameters) ( ) ( ) ( ) en1 + fn1 tanh rn8 V VN8 + fn2 tanh rn9 V VN9 + fn3 tanh rn10 V VN10 (10 parameters) cn2 + dn4 tanh rn11( V VN11) + dn5 tanh rn12( V VN12) ( ) tanh ( ) + dn6 tanh rn13 V VN13 + dn7 rn14 V VN14 (13 parameters) ( ) ( ) ( ) en2 + fn4 tanh rn15 V VN15 + fn5 tanh rn16 V VN16 + fn6 tanh rn17 V VN17 (10 parameters) There are 80 parameters to be varied to fit the data. I do not use the definition of voltage relative to the resting potential, as do Hodgkin and Huxley; it just changes the value of the transitions positions by the value of the resting voltage (-60 volts). The justification for using the hyperbolic-tangent function for the voltage dependence is that it is expected that the parameters (conductance magnitude, rate and voltage position) for either ion s channels changes from one state to another as a function of voltage. There may be several states for a given molecular channel and/or there may be several different types of molecular channels for either ion. The parameters for turning on and off a state may be different. 15
.")
16 Then I fitted the original data by varying the 80 parameters to yield the fit shown in Figure 9. Figure 9. Fits to the voltage-clamp data by varying the 80 voltage-dependent parameters. The voltage-dependent parameters of the fit are given in Table 7. Table 7. The 80 fit parameters at the different voltages in Eq. (1.3). J ( V ) r ( V ) t ( V ) J ( V ) r ( V ) t ( V ) r ( V ) t ( V ) Na constant factor position rate factor position rate E factor position rate factor position rate Na1 Na1 Na2 Na2 The 2 χ is for fitting 224 data with 80 parameters. Note the several position parameters that have nearly the same value, marked with colors. Also, some of the rate parameters have nearly the same value. Later we will set some of them equal and redo the fit. 16
17 The parameters values are given above. This is an alternate time and voltage dependence parameterization of the squid giant-axon membrane excitability to the usual Hodgkin-Huxley parameterization that involves differential equations. ( Undoubtedly, to some approximation of the hyperbolic functions used it is the same as the Hodgkin-Huxley model. The fact that many of the parameters in Table 7 have very nearly the same value is an expected result because the same channels may be active for the different voltage-dependent parameters. One can eliminate 34 variable parameters by equating and eliminating some of the parameters, leaving only 46 2 variable parameters. A higher χ = occurs when this is done and all the parameters are re-fitted, still a reasonably good fit to the data. The equated parameters are shown by colors in Table 8. Table 8. Voltage-dependent fit parameters after 34 parameters are eliminated, leaving 46 variable parameters. The colors indicate those of the original 50 parameters that have been equalized in the fit. J ( V ) r ( V ) t ( V ) J ( V ) r ( V ) t ( V ) r ( V ) t ( V ) Na constant factor position rate factor position rate factor position rate factor position rate factor position rate factor position rate The fit to the data is shown in Figure 10. Na1 Na1 Na2 Na2 17
18 Figure 10. The fit to the voltage-clamp data when 31 parameters are eliminated, leaving 49 parameters. The equation now is: (, ) (, ) (, ) J t V = J t V + J t V where Na and (, ) = ( ){ ( ) ( ) ( )( ( ) tanh + tanh ) }( ) J t V J V r V t V r V t t V V V ( ) + tanh ( )( ( )) ( ) tanh ( )( ( )) ( ) ( ) tanh rna1 V tn1 rna 1 V t tn1 V JNa t, V = JNa V ( V VNa ). tanh rna2 V tn 2 rna2 V t tn 2 V 18
19 The voltage-dependent parameters are [ ] [ ( )] [ ( )] J ( V) = tanh 1.183( V ) tanh V tanh V [ ] [ ( )] r ( V) = tanh 1.183( V ) tanh V [ ] [ ( )] t ( V) = tanh 1.183( V ) tanh V [ ] [ ( )] [ ( )] [ ( )] J ( V) = tanh 1.183( V ) tanh V Na tanh V tanh V [ ] [ ( )] r ( ) ( ) Na1 V = tanh V tanh V [ ] [ ] [ ( )] t ( ) ( ) Na1 V = tanh V tanh 2.519( V 9.189) 7.252tanh V [ ] [ ( )] t ( ) ( ) Na2 V = tanh V tanh V tanh[ ( V 56.38) ]-23.90tanh[ 2.377( V 95.17) ] [ ] [ ] [ ( )] t ( ) ( ) Na2 V = tanh V tanh ( V 9.189) 2.776tanh V


20 The following graphs show the fit parameters as a function of voltage: The transition centered at volts is the most important one. The other two transitions at volts and volts are nearly linear behavior and are necessary for a good fit to the data. There appear to be three states negotiated by these three transitions. Perhaps only two transitions would do the job reasonably well. The transition centered at volts is the most important one. The other transition at volts is just linear behavior and is necessary for a good fit to the data. There appear to be three states negotiated by these two transitions. 20


21 The transition centered at volts is the most important one. The other transition at volts is just linear behavior and is necessary for a good fit to the data. There appear to be three states negotiated by these two transitions. The two transitions centered at volts and volts are the most important ones. The other two transitions at volts and volts are necessary for a good fit to the data. There appear to be five states negotiated by these four transitions, although four states and three transitions might do the job reasonably well. 21


22 The two transitions centered at volts and 56.8 volts are nearly just linear behavior. There appear to be two states negotiated by these two transitions. It appears that one transition might do the job. The two transitions centered at volts and volts are the most important ones. The other transition at volts is just linear behavior and is necessary for a good fit to the data.. There are four states negotiated by these three transitions. 22


23 The two transitions centered at volts and volts are the most important ones. The other two transitions are just linear behavior and are necessary for a good fit to the data. There appear to be two states negotiated by these two transitions. It appears that one transition might do the job reasonably well. All three transitions centered at volts, volts and volts are important. There are four states negotiated by these three transitions. 23
24 However, note that many of the rates [ r ( ) n V ] are quite small. For small argument [ r( x p) < π /2] the hyperbolic tangent, tanh r( x p), is well approximated by a straight line with slope r and intercept p. That is, tanh r( x p) r( x p). Using this in the parameters equations and refitting the data 2 yields a somewhat better fit ( χ = ) with the following parameters: [ ] [ ] [ ( )] J ( ) ( ) V = tanh V tanh ( V 9.189) V [ ] [ ( )] r ( V) = tanh 1.183( V ) V [ ] [ ( )] t ( V) = tanh 1.183( V ) V [ ] [ ( )] J ( ) ( ) Na V = tanh V tanh V tanh[ ( V 56.80) ] [ ( V 66.57) ] [ ] [ ( )] r ( ) ( ) Na1 V = tanh V tanh V [ ] [ ] [ ( )] t ( ) ( ) Na1 V = tanh V tanh 2.806( V 9.189) V [ ] [ ( )] t ( ) ( ) Na2 V = tanh V V tanh[ ( V 56.80) ]-24.53tanh[ 2.377( V 95.17) ] [ ] [ ] [ ( )] t ( ) ( ) Na2 V = tanh V ( V 9.189) tanh V


25 The voltage-dependent parameters are as shown in the following graphs: The arrows pointing toward the right for some curves indicate that the scale is on the right. 25
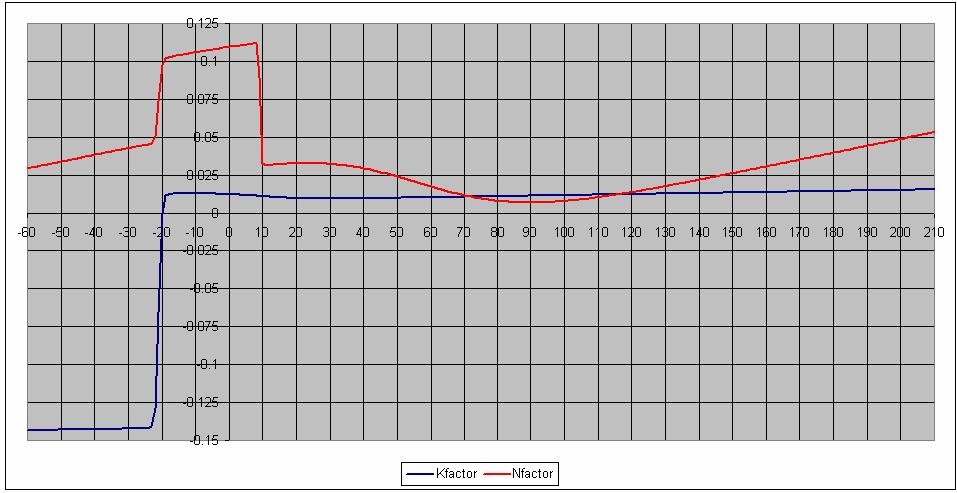
26 26
27 Appendix 1: Hyperbolic Tangent The hyperbolic tangent s definition ( is 1 2 1tanhx + ( ) x x tanh ( x) tanh x e e ex+ e x. = = (1.4) (e is the irrational number See Figure 11 is a graph of the hyperbolic tangent. Figure 11. The hyperbolic-tangent function. 27
28 The hyperbolic-tangent has interesting asymptotic properties: ( x) 1 x 2x 0> x 1 e 1 ex e x ex+ e x x 0 0 2x 0< x + 1 e 1 x tanh. = (1.5) 2x Figure 12 shows the hyperbolic tangent and the( e 1) function plotted together. Figure 12. The hyperbolic-tangent function and the exp( x) 1 function. 28
29 So we see that the following holds: ( x) x 1 2x x 1 2 e ex 1 x e x + x x x e 1+ tanh =. (1.6) 1 2 e x This function is graphed in Figure Figure 13. The + ( ) 1 tanh x 2 function. Note that the function of Figure 13 has the very nice property of being zero for large negative x and 1 for large positive x, having gone through zero at x = zero. One can multiply this function by any number to model many natural phenomena which start at zero for some parameter 1 for a large negative value of parameter 2 and then exponentially rise near zero for parameter 2 and then asymptotically approach the multiplying constant for parameter 1 at large parameter 2. A more general function is needed for real-world fits to natural phenomena, as given in the following equation. ( ;,, ) Q x C p w C 2 1 tanh x p = + (1.7) 2w 29
30 For example, tanh x + is plotted in Figure Figure 14. Plot of tanh x 5 We see why the parameter symbols w and p were chosen in the more general hyperbolic tangent in Eq. (1.3) when we take the derivative of the function. d C x p C x p P x C p w dx 2 2w 4w 2w 1+ tanh = 1 tanh2 = ( ;,, ). (1.8) 2 2 Figure 15 is a plot of 1 1 tanh x
31 Figure 15. Plot of x 2 1 tanh. 5 We see that the equation s parameters have the following meanings: p = peak position of the function. w = a measure of the width of the function; also the exponential time constant near x= p. C = height of the function. 4w (1.9) Sometimes natural data are in the form of an asymptotic curve such as Eq. (1.4) and sometimes natural data are in the form of a peaked curve, such as Eq. (1.5). As shown above, they are related by a calculus derivative, or inversely a calculus integral. 31
32 The hyperbolic tangent can be zero at x = 0 and have an exponential growth toward y near x = 0 or an exponential growth toward x near y = 0 as follows: p x p tanh + tanh. 2w 2w Examples: 2 2 tanh + tanh x for exponential growth in y as a function of x: Figure 16. Plot of tanh + tanh x
33 2 2 tanh + tanh x + for exponential growth in x as a function of y: 2 2 Figure 17. Plot of 2 2 tanh + tanh x The expansion of the hyperbolic tangent for small argument is ( So, tanh r( x p) r( x p) 1 r( x p) 3 = 3 + L for small x. 33
34 For turning on and then off a state the following function works quite well: t i c t t i t t o tanh tanh, where 2 wi wo = transition time position for going into the state and t o = transition time position for leaving the state; w = width for going into the state and i w o = width for leaving the state. For example, Error! Reference source not found. shows the two transitions represented by 1 t 2 t 10 tanh tanh Figure 18. Plot of 1 t tanh 2 t tanh
35 Appendix 2: Hodgkin-Huxley Model References Cronin, 1987: Mathematical Aspects of Hodgkin-Huxley Neural Theory, Jane Cronin, Cambridge University Press, Hille, 1984: Ionic Channels of Excitable Membranes, Bertil Hille, Sinauer Assoc. Inc, Malmivuo, 1995: Bioelectromagnetism: Principles and Applications of Bioelectric and Biomagnetic Fields, Jaakko Malmivuo and Robert Plonsey, Oxford Univ. Press,
Voltage-clamp and Hodgkin-Huxley models
 Voltage-clamp and Hodgkin-Huxley models Read: Hille, Chapters 2-5 (best) Koch, Chapters 6, 8, 9 See also Clay, J. Neurophysiol. 80:903-913 (1998) (for a recent version of the HH squid axon model) Rothman
Voltage-clamp and Hodgkin-Huxley models Read: Hille, Chapters 2-5 (best) Koch, Chapters 6, 8, 9 See also Clay, J. Neurophysiol. 80:903-913 (1998) (for a recent version of the HH squid axon model) Rothman
Voltage-clamp and Hodgkin-Huxley models
 Voltage-clamp and Hodgkin-Huxley models Read: Hille, Chapters 2-5 (best Koch, Chapters 6, 8, 9 See also Hodgkin and Huxley, J. Physiol. 117:500-544 (1952. (the source Clay, J. Neurophysiol. 80:903-913
Voltage-clamp and Hodgkin-Huxley models Read: Hille, Chapters 2-5 (best Koch, Chapters 6, 8, 9 See also Hodgkin and Huxley, J. Physiol. 117:500-544 (1952. (the source Clay, J. Neurophysiol. 80:903-913
Resting membrane potential,
 Resting membrane potential Inside of each cell is negative as compared with outer surface: negative resting membrane potential (between -30 and -90 mv) Examination with microelectrode (Filled with KCl
Resting membrane potential Inside of each cell is negative as compared with outer surface: negative resting membrane potential (between -30 and -90 mv) Examination with microelectrode (Filled with KCl
BIOELECTRIC PHENOMENA
 Chapter 11 BIOELECTRIC PHENOMENA 11.3 NEURONS 11.3.1 Membrane Potentials Resting Potential by separation of charge due to the selective permeability of the membrane to ions From C v= Q, where v=60mv and
Chapter 11 BIOELECTRIC PHENOMENA 11.3 NEURONS 11.3.1 Membrane Potentials Resting Potential by separation of charge due to the selective permeability of the membrane to ions From C v= Q, where v=60mv and
Transmission of Nerve Impulses (see Fig , p. 403)
") How a nerve impulse works Transmission of Nerve Impulses (see Fig. 12.13, p. 403) 1. At Rest (Polarization) outside of neuron is positively charged compared to inside (sodium ions outside, chloride and
How a nerve impulse works Transmission of Nerve Impulses (see Fig. 12.13, p. 403) 1. At Rest (Polarization) outside of neuron is positively charged compared to inside (sodium ions outside, chloride and
Lecture 10 : Neuronal Dynamics. Eileen Nugent
 Lecture 10 : Neuronal Dynamics Eileen Nugent Origin of the Cells Resting Membrane Potential: Nernst Equation, Donnan Equilbrium Action Potentials in the Nervous System Equivalent Electrical Circuits and
Lecture 10 : Neuronal Dynamics Eileen Nugent Origin of the Cells Resting Membrane Potential: Nernst Equation, Donnan Equilbrium Action Potentials in the Nervous System Equivalent Electrical Circuits and
STUDENT PAPER. Santiago Santana University of Illinois, Urbana-Champaign Blue Waters Education Program 736 S. Lombard Oak Park IL, 60304
 STUDENT PAPER Differences between Stochastic and Deterministic Modeling in Real World Systems using the Action Potential of Nerves. Santiago Santana University of Illinois, Urbana-Champaign Blue Waters
STUDENT PAPER Differences between Stochastic and Deterministic Modeling in Real World Systems using the Action Potential of Nerves. Santiago Santana University of Illinois, Urbana-Champaign Blue Waters
Single-Compartment Neural Models
 Single-Compartment Neural Models BENG/BGGN 260 Neurodynamics University of California, San Diego Week 2 BENG/BGGN 260 Neurodynamics (UCSD) Single-Compartment Neural Models Week 2 1 / 18 Reading Materials
Single-Compartment Neural Models BENG/BGGN 260 Neurodynamics University of California, San Diego Week 2 BENG/BGGN 260 Neurodynamics (UCSD) Single-Compartment Neural Models Week 2 1 / 18 Reading Materials
BME 5742 Biosystems Modeling and Control
 BME 5742 Biosystems Modeling and Control Hodgkin-Huxley Model for Nerve Cell Action Potential Part 1 Dr. Zvi Roth (FAU) 1 References Hoppensteadt-Peskin Ch. 3 for all the mathematics. Cooper s The Cell
BME 5742 Biosystems Modeling and Control Hodgkin-Huxley Model for Nerve Cell Action Potential Part 1 Dr. Zvi Roth (FAU) 1 References Hoppensteadt-Peskin Ch. 3 for all the mathematics. Cooper s The Cell
Biomedical Instrumentation
 ELEC ENG 4BD4: Biomedical Instrumentation Lecture 5 Bioelectricity 1. INTRODUCTION TO BIOELECTRICITY AND EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
ELEC ENG 4BD4: Biomedical Instrumentation Lecture 5 Bioelectricity 1. INTRODUCTION TO BIOELECTRICITY AND EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
Membrane Potentials, Action Potentials, and Synaptic Transmission. Membrane Potential
 Cl Cl - - + K + K+ K + K Cl - 2/2/15 Membrane Potentials, Action Potentials, and Synaptic Transmission Core Curriculum II Spring 2015 Membrane Potential Example 1: K +, Cl - equally permeant no charge
Cl Cl - - + K + K+ K + K Cl - 2/2/15 Membrane Potentials, Action Potentials, and Synaptic Transmission Core Curriculum II Spring 2015 Membrane Potential Example 1: K +, Cl - equally permeant no charge
they give no information about the rate at which repolarization restores the
 497 J. Physiol. (1952) ii6, 497-506 THE DUAL EFFECT OF MEMBRANE POTENTIAL ON SODIUM CONDUCTANCE IN THE GIANT AXON OF LOLIGO BY A. L. HODGKIN AND A. F. HUXLEY From the Laboratory of the Marine Biological
497 J. Physiol. (1952) ii6, 497-506 THE DUAL EFFECT OF MEMBRANE POTENTIAL ON SODIUM CONDUCTANCE IN THE GIANT AXON OF LOLIGO BY A. L. HODGKIN AND A. F. HUXLEY From the Laboratory of the Marine Biological
Membrane Potentials. Why are some cells electrically active? Model 1: The Sodium/Potassium pump. Critical Thinking Questions
 Membrane Potentials Model 1: The Sodium/Potassium pump Why are some cells electrically active? 1. What ion is being moved out of the cell according to model 1? a. How many of these are being moved out?
Membrane Potentials Model 1: The Sodium/Potassium pump Why are some cells electrically active? 1. What ion is being moved out of the cell according to model 1? a. How many of these are being moved out?
Overview Organization: Central Nervous System (CNS) Peripheral Nervous System (PNS) innervate Divisions: a. Afferent
 Peripheral Nervous System (PNS) innervate Divisions: a. Afferent") Overview Organization: Central Nervous System (CNS) Brain and spinal cord receives and processes information. Peripheral Nervous System (PNS) Nerve cells that link CNS with organs throughout the body.
Overview Organization: Central Nervous System (CNS) Brain and spinal cord receives and processes information. Peripheral Nervous System (PNS) Nerve cells that link CNS with organs throughout the body.
On Parameter Estimation for Neuron Models
 On Parameter Estimation for Neuron Models Abhijit Biswas Department of Mathematics Temple University November 30th, 2017 Abhijit Biswas (Temple University) On Parameter Estimation for Neuron Models November
On Parameter Estimation for Neuron Models Abhijit Biswas Department of Mathematics Temple University November 30th, 2017 Abhijit Biswas (Temple University) On Parameter Estimation for Neuron Models November
Electrophysiology of the neuron
 School of Mathematical Sciences G4TNS Theoretical Neuroscience Electrophysiology of the neuron Electrophysiology is the study of ionic currents and electrical activity in cells and tissues. The work of
School of Mathematical Sciences G4TNS Theoretical Neuroscience Electrophysiology of the neuron Electrophysiology is the study of ionic currents and electrical activity in cells and tissues. The work of
Lecture 11 : Simple Neuron Models. Dr Eileen Nugent
 Lecture 11 : Simple Neuron Models Dr Eileen Nugent Reading List Nelson, Biological Physics, Chapter 12 Phillips, PBoC, Chapter 17 Gerstner, Neuronal Dynamics: from single neurons to networks and models
Lecture 11 : Simple Neuron Models Dr Eileen Nugent Reading List Nelson, Biological Physics, Chapter 12 Phillips, PBoC, Chapter 17 Gerstner, Neuronal Dynamics: from single neurons to networks and models
Sample HHSim Exercises
 I. Equilibrium Potential II. Membrane Potential III. The Action Potential IV. The Fast Sodium Channel V. The Delayed Rectifier VI. Voltage-Gated Channel Parameters Part I: Equilibrium Potential Sample
I. Equilibrium Potential II. Membrane Potential III. The Action Potential IV. The Fast Sodium Channel V. The Delayed Rectifier VI. Voltage-Gated Channel Parameters Part I: Equilibrium Potential Sample
ACTION POTENTIAL. Dr. Ayisha Qureshi Professor MBBS, MPhil
 ACTION POTENTIAL Dr. Ayisha Qureshi Professor MBBS, MPhil DEFINITIONS: Stimulus: A stimulus is an external force or event which when applied to an excitable tissue produces a characteristic response. Subthreshold
ACTION POTENTIAL Dr. Ayisha Qureshi Professor MBBS, MPhil DEFINITIONS: Stimulus: A stimulus is an external force or event which when applied to an excitable tissue produces a characteristic response. Subthreshold
Neurons and the membrane potential. N500 John Beggs 23 Aug, 2016
 Neurons and the membrane potential N500 John Beggs 23 Aug, 2016 My background, briefly Neurons Structural elements of a typical neuron Figure 1.2 Some nerve cell morphologies found in the human
Neurons and the membrane potential N500 John Beggs 23 Aug, 2016 My background, briefly Neurons Structural elements of a typical neuron Figure 1.2 Some nerve cell morphologies found in the human
Action Potential Propagation
 Action Potential Propagation 2 Action Potential is a transient alteration of transmembrane voltage (or membrane potential) across an excitable membrane generated by the activity of voltage-gated ion channels.
Action Potential Propagation 2 Action Potential is a transient alteration of transmembrane voltage (or membrane potential) across an excitable membrane generated by the activity of voltage-gated ion channels.
Neural Modeling and Computational Neuroscience. Claudio Gallicchio
 Neural Modeling and Computational Neuroscience Claudio Gallicchio 1 Neuroscience modeling 2 Introduction to basic aspects of brain computation Introduction to neurophysiology Neural modeling: Elements
Neural Modeling and Computational Neuroscience Claudio Gallicchio 1 Neuroscience modeling 2 Introduction to basic aspects of brain computation Introduction to neurophysiology Neural modeling: Elements
General Physics. Nerve Conduction. Newton s laws of Motion Work, Energy and Power. Fluids. Direct Current (DC)
") Newton s laws of Motion Work, Energy and Power Fluids Direct Current (DC) Nerve Conduction Wave properties of light Ionizing Radiation General Physics Prepared by: Sujood Alazzam 2017/2018 CHAPTER OUTLINE
Newton s laws of Motion Work, Energy and Power Fluids Direct Current (DC) Nerve Conduction Wave properties of light Ionizing Radiation General Physics Prepared by: Sujood Alazzam 2017/2018 CHAPTER OUTLINE
Converting energy into nerve impulses, resting potentials and action potentials Sensory receptors
 D 1.3 s Converting energy into nerve impulses, resting potentials and action potentials Sensory receptors A receptor converts an external or internal stimulus into an electrical signal. Sensory receptors
D 1.3 s Converting energy into nerve impulses, resting potentials and action potentials Sensory receptors A receptor converts an external or internal stimulus into an electrical signal. Sensory receptors
The Membrane Potential
 The Membrane Potential Graphics are used with permission of: Pearson Education Inc., publishing as Benjamin Cummings (http://www.aw-bc.com) ** It is suggested that you carefully label each ion channel
The Membrane Potential Graphics are used with permission of: Pearson Education Inc., publishing as Benjamin Cummings (http://www.aw-bc.com) ** It is suggested that you carefully label each ion channel
Neural Conduction. biologyaspoetry.com
 Neural Conduction biologyaspoetry.com Resting Membrane Potential -70mV A cell s membrane potential is the difference in the electrical potential ( charge) between the inside and outside of the cell. The
Neural Conduction biologyaspoetry.com Resting Membrane Potential -70mV A cell s membrane potential is the difference in the electrical potential ( charge) between the inside and outside of the cell. The
From neuronal oscillations to complexity
 1/39 The Fourth International Workshop on Advanced Computation for Engineering Applications (ACEA 2008) MACIS 2 Al-Balqa Applied University, Salt, Jordan Corson Nathalie, Aziz Alaoui M.A. University of
1/39 The Fourth International Workshop on Advanced Computation for Engineering Applications (ACEA 2008) MACIS 2 Al-Balqa Applied University, Salt, Jordan Corson Nathalie, Aziz Alaoui M.A. University of
Rahaf Nasser mohammad khatatbeh
 7 7... Hiba Abu Hayyeh... Rahaf Nasser mohammad khatatbeh Mohammad khatatbeh Brief introduction about membrane potential The term membrane potential refers to a separation of opposite charges across the
7 7... Hiba Abu Hayyeh... Rahaf Nasser mohammad khatatbeh Mohammad khatatbeh Brief introduction about membrane potential The term membrane potential refers to a separation of opposite charges across the
3.3 Simulating action potentials
 6 THE HODGKIN HUXLEY MODEL OF THE ACTION POTENTIAL Fig. 3.1 Voltage dependence of rate coefficients and limiting values and time constants for the Hodgkin Huxley gating variables. (a) Graphs of forward
6 THE HODGKIN HUXLEY MODEL OF THE ACTION POTENTIAL Fig. 3.1 Voltage dependence of rate coefficients and limiting values and time constants for the Hodgkin Huxley gating variables. (a) Graphs of forward
4. Active Behavior of the Cell Membrane 4.1 INTRODUCTION
 1 of 50 10/17/2014 10:48 PM 4.1 INTRODUCTION When a stimulus current pulse is arranged to depolarize the resting membrane of a cell to or beyond the threshold voltage, then the membrane will respond with
1 of 50 10/17/2014 10:48 PM 4.1 INTRODUCTION When a stimulus current pulse is arranged to depolarize the resting membrane of a cell to or beyond the threshold voltage, then the membrane will respond with
The Membrane Potential
 The Membrane Potential Graphics are used with permission of: adam.com (http://www.adam.com/) Benjamin Cummings Publishing Co (http://www.aw.com/bc) ** It is suggested that you carefully label each ion
The Membrane Potential Graphics are used with permission of: adam.com (http://www.adam.com/) Benjamin Cummings Publishing Co (http://www.aw.com/bc) ** It is suggested that you carefully label each ion
Mathematical Foundations of Neuroscience - Lecture 3. Electrophysiology of neurons - continued
 Mathematical Foundations of Neuroscience - Lecture 3. Electrophysiology of neurons - continued Filip Piękniewski Faculty of Mathematics and Computer Science, Nicolaus Copernicus University, Toruń, Poland
Mathematical Foundations of Neuroscience - Lecture 3. Electrophysiology of neurons - continued Filip Piękniewski Faculty of Mathematics and Computer Science, Nicolaus Copernicus University, Toruń, Poland
Fundamentals of the Nervous System and Nervous Tissue
 Chapter 11 Part B Fundamentals of the Nervous System and Nervous Tissue Annie Leibovitz/Contact Press Images PowerPoint Lecture Slides prepared by Karen Dunbar Kareiva Ivy Tech Community College 11.4 Membrane
Chapter 11 Part B Fundamentals of the Nervous System and Nervous Tissue Annie Leibovitz/Contact Press Images PowerPoint Lecture Slides prepared by Karen Dunbar Kareiva Ivy Tech Community College 11.4 Membrane
Chapter 1 subtitles Ion gradients
 CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 1 subtitles Ion gradients Introduction In this first chapter, I'll explain the basic knowledge required to understand the electrical signals generated
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 1 subtitles Ion gradients Introduction In this first chapter, I'll explain the basic knowledge required to understand the electrical signals generated
Lecture Notes 8C120 Inleiding Meten en Modelleren. Cellular electrophysiology: modeling and simulation. Nico Kuijpers
 Lecture Notes 8C2 Inleiding Meten en Modelleren Cellular electrophysiology: modeling and simulation Nico Kuijpers nico.kuijpers@bf.unimaas.nl February 9, 2 2 8C2 Inleiding Meten en Modelleren Extracellular
Lecture Notes 8C2 Inleiding Meten en Modelleren Cellular electrophysiology: modeling and simulation Nico Kuijpers nico.kuijpers@bf.unimaas.nl February 9, 2 2 8C2 Inleiding Meten en Modelleren Extracellular
Simulation of Cardiac Action Potentials Background Information
 Simulation of Cardiac Action Potentials Background Information Rob MacLeod and Quan Ni February 7, 2 Introduction The goal of assignments related to this document is to experiment with a numerical simulation
Simulation of Cardiac Action Potentials Background Information Rob MacLeod and Quan Ni February 7, 2 Introduction The goal of assignments related to this document is to experiment with a numerical simulation
FRTF01 L8 Electrophysiology
 FRTF01 L8 Electrophysiology Lecture Electrophysiology in general Recap: Linear Time Invariant systems (LTI) Examples of 1 and 2-dimensional systems Stability analysis The need for non-linear descriptions
FRTF01 L8 Electrophysiology Lecture Electrophysiology in general Recap: Linear Time Invariant systems (LTI) Examples of 1 and 2-dimensional systems Stability analysis The need for non-linear descriptions
All-or-None Principle and Weakness of Hodgkin-Huxley Mathematical Model
 All-or-None Principle and Weakness of Hodgkin-Huxley Mathematical Model S. A. Sadegh Zadeh, C. Kambhampati International Science Index, Mathematical and Computational Sciences waset.org/publication/10008281
All-or-None Principle and Weakness of Hodgkin-Huxley Mathematical Model S. A. Sadegh Zadeh, C. Kambhampati International Science Index, Mathematical and Computational Sciences waset.org/publication/10008281
LESSON 2.2 WORKBOOK How do our axons transmit electrical signals?
 LESSON 2.2 WORKBOOK How do our axons transmit electrical signals? This lesson introduces you to the action potential, which is the process by which axons signal electrically. In this lesson you will learn
LESSON 2.2 WORKBOOK How do our axons transmit electrical signals? This lesson introduces you to the action potential, which is the process by which axons signal electrically. In this lesson you will learn
Νευροφυσιολογία και Αισθήσεις
 Biomedical Imaging & Applied Optics University of Cyprus Νευροφυσιολογία και Αισθήσεις Διάλεξη 5 Μοντέλο Hodgkin-Huxley (Hodgkin-Huxley Model) Response to Current Injection 2 Hodgin & Huxley Sir Alan Lloyd
Biomedical Imaging & Applied Optics University of Cyprus Νευροφυσιολογία και Αισθήσεις Διάλεξη 5 Μοντέλο Hodgkin-Huxley (Hodgkin-Huxley Model) Response to Current Injection 2 Hodgin & Huxley Sir Alan Lloyd
Quantitative Electrophysiology
 ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Wednesday, September 13, 2006 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in
ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Wednesday, September 13, 2006 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in
Transport of ions across plasma membranes
 Transport of ions across plasma membranes Plasma Membranes of Excitable tissues Ref: Guyton, 13 th ed: pp: 61-71. 12 th ed: pp: 57-69. 11th ed: p57-71, Electrical properties of plasma membranes Part A:
Transport of ions across plasma membranes Plasma Membranes of Excitable tissues Ref: Guyton, 13 th ed: pp: 61-71. 12 th ed: pp: 57-69. 11th ed: p57-71, Electrical properties of plasma membranes Part A:
6.3.4 Action potential
 I ion C m C m dφ dt Figure 6.8: Electrical circuit model of the cell membrane. Normally, cells are net negative inside the cell which results in a non-zero resting membrane potential. The membrane potential
I ion C m C m dφ dt Figure 6.8: Electrical circuit model of the cell membrane. Normally, cells are net negative inside the cell which results in a non-zero resting membrane potential. The membrane potential
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Supplementary Figure S1. Pulses >3mJ reduce membrane resistance in HEK cells. Reversal potentials in a representative cell for IR-induced currents with laser pulses of 0.74 to
SUPPLEMENTARY INFORMATION Supplementary Figure S1. Pulses >3mJ reduce membrane resistance in HEK cells. Reversal potentials in a representative cell for IR-induced currents with laser pulses of 0.74 to
! Depolarization continued. AP Biology. " The final phase of a local action
 ! Resting State Resting potential is maintained mainly by non-gated K channels which allow K to diffuse out! Voltage-gated ion K and channels along axon are closed! Depolarization A stimulus causes channels
! Resting State Resting potential is maintained mainly by non-gated K channels which allow K to diffuse out! Voltage-gated ion K and channels along axon are closed! Depolarization A stimulus causes channels
Curtis et al. Il nuovo Invito alla biologia.blu BIOLOGY HIGHLIGHTS KEYS
 BIOLOGY HIGHLIGHTS KEYS Watch the videos and download the transcripts of this section at: online.scuola.zanichelli.it/curtisnuovoinvitoblu/clil > THE HUMAN NERVOUS SYSTEM 2. WARM UP a) The structures that
BIOLOGY HIGHLIGHTS KEYS Watch the videos and download the transcripts of this section at: online.scuola.zanichelli.it/curtisnuovoinvitoblu/clil > THE HUMAN NERVOUS SYSTEM 2. WARM UP a) The structures that
Introduction and the Hodgkin-Huxley Model
 1 Introduction and the Hodgkin-Huxley Model Richard Bertram Department of Mathematics and Programs in Neuroscience and Molecular Biophysics Florida State University Tallahassee, Florida 32306 Reference:
1 Introduction and the Hodgkin-Huxley Model Richard Bertram Department of Mathematics and Programs in Neuroscience and Molecular Biophysics Florida State University Tallahassee, Florida 32306 Reference:
Ch. 5. Membrane Potentials and Action Potentials
 Ch. 5. Membrane Potentials and Action Potentials Basic Physics of Membrane Potentials Nerve and muscle cells: Excitable Capable of generating rapidly changing electrochemical impulses at their membranes
Ch. 5. Membrane Potentials and Action Potentials Basic Physics of Membrane Potentials Nerve and muscle cells: Excitable Capable of generating rapidly changing electrochemical impulses at their membranes
Quantitative Electrophysiology
 ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Tuesday, September 18, 2012 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Tuesday, September 18, 2012 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
2401 : Anatomy/Physiology
 Dr. Chris Doumen Week 6 2401 : Anatomy/Physiology Action Potentials NeuroPhysiology TextBook Readings Pages 400 through 408 Make use of the figures in your textbook ; a picture is worth a thousand words!
Dr. Chris Doumen Week 6 2401 : Anatomy/Physiology Action Potentials NeuroPhysiology TextBook Readings Pages 400 through 408 Make use of the figures in your textbook ; a picture is worth a thousand words!
9 Generation of Action Potential Hodgkin-Huxley Model
 9 Generation of Action Potential Hodgkin-Huxley Model (based on chapter 12, W.W. Lytton, Hodgkin-Huxley Model) 9.1 Passive and active membrane models In the previous lecture we have considered a passive
9 Generation of Action Potential Hodgkin-Huxley Model (based on chapter 12, W.W. Lytton, Hodgkin-Huxley Model) 9.1 Passive and active membrane models In the previous lecture we have considered a passive
Electrical Signaling. Lecture Outline. Using Ions as Messengers. Potentials in Electrical Signaling
 Lecture Outline Electrical Signaling Using ions as messengers Potentials in electrical signaling Action Graded Other electrical signaling Gap junctions The neuron Using Ions as Messengers Important things
Lecture Outline Electrical Signaling Using ions as messengers Potentials in electrical signaling Action Graded Other electrical signaling Gap junctions The neuron Using Ions as Messengers Important things
Thermo-electrical equivalents for simulating the electro-mechanical behavior of biological tissue
 Ilaria Cinelli Thermo-electrical equivalents for simulating the electro-mechanical behavior of biological tissue Cao, H., Migliazza, J., Liu, X. C., & Dominick, D. (2012). Elia, S., Lamberti, P., & Tucci,
Ilaria Cinelli Thermo-electrical equivalents for simulating the electro-mechanical behavior of biological tissue Cao, H., Migliazza, J., Liu, X. C., & Dominick, D. (2012). Elia, S., Lamberti, P., & Tucci,
Topics in Neurophysics
 Topics in Neurophysics Alex Loebel, Martin Stemmler and Anderas Herz Exercise 2 Solution (1) The Hodgkin Huxley Model The goal of this exercise is to simulate the action potential according to the model
Topics in Neurophysics Alex Loebel, Martin Stemmler and Anderas Herz Exercise 2 Solution (1) The Hodgkin Huxley Model The goal of this exercise is to simulate the action potential according to the model
Neurons, Synapses, and Signaling
 Chapter 48 Neurons, Synapses, and Signaling PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Chapter 48 Neurons, Synapses, and Signaling PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Nervous System: Part II How A Neuron Works
 Nervous System: Part II How A Neuron Works Essential Knowledge Statement 3.E.2 Continued Animals have nervous systems that detect external and internal signals, transmit and integrate information, and
Nervous System: Part II How A Neuron Works Essential Knowledge Statement 3.E.2 Continued Animals have nervous systems that detect external and internal signals, transmit and integrate information, and
2.6 The Membrane Potential
 2.6: The Membrane Potential 51 tracellular potassium, so that the energy stored in the electrochemical gradients can be extracted. Indeed, when this is the case experimentally, ATP is synthesized from
2.6: The Membrane Potential 51 tracellular potassium, so that the energy stored in the electrochemical gradients can be extracted. Indeed, when this is the case experimentally, ATP is synthesized from
Neurochemistry 1. Nervous system is made of neurons & glia, as well as other cells. Santiago Ramon y Cajal Nobel Prize 1906
 Neurochemistry 1 Nervous system is made of neurons & glia, as well as other cells. Santiago Ramon y Cajal Nobel Prize 1906 How Many Neurons Do We Have? The human brain contains ~86 billion neurons and
Neurochemistry 1 Nervous system is made of neurons & glia, as well as other cells. Santiago Ramon y Cajal Nobel Prize 1906 How Many Neurons Do We Have? The human brain contains ~86 billion neurons and
Physiology Unit 2. MEMBRANE POTENTIALS and SYNAPSES
 Physiology Unit 2 MEMBRANE POTENTIALS and SYNAPSES Neuron Communication Neurons are stimulated by receptors on dendrites and cell bodies (soma) Ligand gated ion channels GPCR s Neurons stimulate cells
Physiology Unit 2 MEMBRANE POTENTIALS and SYNAPSES Neuron Communication Neurons are stimulated by receptors on dendrites and cell bodies (soma) Ligand gated ion channels GPCR s Neurons stimulate cells
STUDIES OF CHANGING MEMBRANE POTENTIAL * : 1. BASIC ELECTRICAL THEORY, 2. GRADED AND ACTION POTENTIALS 3. THE VOLTAGE CLAMP AND MEMBRANE POTENTIALS
 STUDIES OF CHANGING MEMBRANE POTENTIAL * : 1. BASIC ELECTRICAL THEORY, 2. GRADED AND ACTION POTENTIALS 3. THE VOLTAGE CLAMP AND MEMBRANE POTENTIALS I. INTRODUCTION A. So far we have only considered the
STUDIES OF CHANGING MEMBRANE POTENTIAL * : 1. BASIC ELECTRICAL THEORY, 2. GRADED AND ACTION POTENTIALS 3. THE VOLTAGE CLAMP AND MEMBRANE POTENTIALS I. INTRODUCTION A. So far we have only considered the
Electronics 101 Solving a differential equation Incorporating space. Numerical Methods. Accuracy, stability, speed. Robert A.
 Numerical Methods Accuracy, stability, speed Robert A. McDougal Yale School of Medicine 21 June 2016 Hodgkin and Huxley: squid giant axon experiments Top: Alan Lloyd Hodgkin; Bottom: Andrew Fielding Huxley.
Numerical Methods Accuracy, stability, speed Robert A. McDougal Yale School of Medicine 21 June 2016 Hodgkin and Huxley: squid giant axon experiments Top: Alan Lloyd Hodgkin; Bottom: Andrew Fielding Huxley.
Nonlinear Observer Design and Synchronization Analysis for Classical Models of Neural Oscillators
 Nonlinear Observer Design and Synchronization Analysis for Classical Models of Neural Oscillators Ranjeetha Bharath and Jean-Jacques Slotine Massachusetts Institute of Technology ABSTRACT This work explores
Nonlinear Observer Design and Synchronization Analysis for Classical Models of Neural Oscillators Ranjeetha Bharath and Jean-Jacques Slotine Massachusetts Institute of Technology ABSTRACT This work explores
Effects of Betaxolol on Hodgkin-Huxley Model of Tiger Salamander Retinal Ganglion Cell
 Effects of Betaxolol on Hodgkin-Huxley Model of Tiger Salamander Retinal Ganglion Cell 1. Abstract Matthew Dunlevie Clement Lee Indrani Mikkilineni mdunlevi@ucsd.edu cll008@ucsd.edu imikkili@ucsd.edu Isolated
Effects of Betaxolol on Hodgkin-Huxley Model of Tiger Salamander Retinal Ganglion Cell 1. Abstract Matthew Dunlevie Clement Lee Indrani Mikkilineni mdunlevi@ucsd.edu cll008@ucsd.edu imikkili@ucsd.edu Isolated
Lecture goals: Learning Objectives
 Title: Membrane Potential in Excitable Cells 1 Subtitle: Voltage-Gated Ion Channels and the basis of the Action Potential Diomedes E. Logothetis, Ph.D. Lecture goals: This first of two lectures will use
Title: Membrane Potential in Excitable Cells 1 Subtitle: Voltage-Gated Ion Channels and the basis of the Action Potential Diomedes E. Logothetis, Ph.D. Lecture goals: This first of two lectures will use
Math 345 Intro to Math Biology Lecture 20: Mathematical model of Neuron conduction
 Math 345 Intro to Math Biology Lecture 20: Mathematical model of Neuron conduction Junping Shi College of William and Mary November 8, 2018 Neuron Neurons Neurons are cells in the brain and other subsystems
Math 345 Intro to Math Biology Lecture 20: Mathematical model of Neuron conduction Junping Shi College of William and Mary November 8, 2018 Neuron Neurons Neurons are cells in the brain and other subsystems
BIOL Week 5. Nervous System II. The Membrane Potential. Question : Is the Equilibrium Potential a set number or can it change?
 Collin County Community College BIOL 2401 Week 5 Nervous System II 1 The Membrane Potential Question : Is the Equilibrium Potential a set number or can it change? Let s look at the Nernst Equation again.
Collin County Community College BIOL 2401 Week 5 Nervous System II 1 The Membrane Potential Question : Is the Equilibrium Potential a set number or can it change? Let s look at the Nernst Equation again.
BRIEF COMMUNICATIONS COMPUTATION OF AXON GATING CURRENTS FROM DIPOLE MOMENT CHANGES IN CHANNEL SUBUNITS
 BRIEF COMMUNICATIONS COMPUTATION OF AXON GATING CURRENTS FROM DIPOLE MOMENT CHANGES IN CHANNEL SUBUNITS E. LEVITANand Y. PALTI From the Department of Physiology and Biophysics, Technion School of Medicine,
BRIEF COMMUNICATIONS COMPUTATION OF AXON GATING CURRENTS FROM DIPOLE MOMENT CHANGES IN CHANNEL SUBUNITS E. LEVITANand Y. PALTI From the Department of Physiology and Biophysics, Technion School of Medicine,
Resting Membrane Potential
 Resting Membrane Potential Fig. 12.09a,b Recording of Resting and It is recorded by cathode ray oscilloscope action potentials -70 0 mv + it is negative in polarized (resting, the membrane can be excited)
Resting Membrane Potential Fig. 12.09a,b Recording of Resting and It is recorded by cathode ray oscilloscope action potentials -70 0 mv + it is negative in polarized (resting, the membrane can be excited)
Channels can be activated by ligand-binding (chemical), voltage change, or mechanical changes such as stretch.
, voltage change, or mechanical changes such as stretch.") 1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
- the flow of electrical charge from one point to the other is current.
 Biology 325, Fall 2004 Resting membrane potential I. Introduction A. The body and electricity, basic principles - the body is electrically neutral (total), however there are areas where opposite charges
Biology 325, Fall 2004 Resting membrane potential I. Introduction A. The body and electricity, basic principles - the body is electrically neutral (total), however there are areas where opposite charges
Decoding. How well can we learn what the stimulus is by looking at the neural responses?
 Decoding How well can we learn what the stimulus is by looking at the neural responses? Two approaches: devise explicit algorithms for extracting a stimulus estimate directly quantify the relationship
Decoding How well can we learn what the stimulus is by looking at the neural responses? Two approaches: devise explicit algorithms for extracting a stimulus estimate directly quantify the relationship
POTASSIUM PERMEABILITY IN
 SLOW CHANGES OF POTASSIUM PERMEABILITY IN THE SQUID GIANT AXON GERALD EHRENSTEIN and DANIEL L. GILBERT From the National Institutes of Health, Bethesda, Maryland, and the Marine Biological Laboratory,
SLOW CHANGES OF POTASSIUM PERMEABILITY IN THE SQUID GIANT AXON GERALD EHRENSTEIN and DANIEL L. GILBERT From the National Institutes of Health, Bethesda, Maryland, and the Marine Biological Laboratory,
PNS Chapter 7. Membrane Potential / Neural Signal Processing Spring 2017 Prof. Byron Yu
 PNS Chapter 7 Membrane Potential 18-698 / 42-632 Neural Signal Processing Spring 2017 Prof. Byron Yu Roadmap Introduction to neuroscience Chapter 1 The brain and behavior Chapter 2 Nerve cells and behavior
PNS Chapter 7 Membrane Potential 18-698 / 42-632 Neural Signal Processing Spring 2017 Prof. Byron Yu Roadmap Introduction to neuroscience Chapter 1 The brain and behavior Chapter 2 Nerve cells and behavior
Action Potentials and Synaptic Transmission Physics 171/271
 Action Potentials and Synaptic Transmission Physics 171/271 Flavio Fröhlich (flavio@salk.edu) September 27, 2006 In this section, we consider two important aspects concerning the communication between
Action Potentials and Synaptic Transmission Physics 171/271 Flavio Fröhlich (flavio@salk.edu) September 27, 2006 In this section, we consider two important aspects concerning the communication between
Biological membranes and bioelectric phenomena
 Lectures on Medical Biophysics Dept. Biophysics, Medical faculty, Masaryk University in Brno Biological membranes and bioelectric phenomena A part of this lecture was prepared on the basis of a presentation
Lectures on Medical Biophysics Dept. Biophysics, Medical faculty, Masaryk University in Brno Biological membranes and bioelectric phenomena A part of this lecture was prepared on the basis of a presentation
Dynamics and complexity of Hindmarsh-Rose neuronal systems
 Dynamics and complexity of Hindmarsh-Rose neuronal systems Nathalie Corson and M.A. Aziz-Alaoui Laboratoire de Mathématiques Appliquées du Havre, 5 rue Philippe Lebon, 766 Le Havre, FRANCE nathalie.corson@univ-lehavre.fr
Dynamics and complexity of Hindmarsh-Rose neuronal systems Nathalie Corson and M.A. Aziz-Alaoui Laboratoire de Mathématiques Appliquées du Havre, 5 rue Philippe Lebon, 766 Le Havre, FRANCE nathalie.corson@univ-lehavre.fr
Signal processing in nervous system - Hodgkin-Huxley model
 Signal processing in nervous system - Hodgkin-Huxley model Ulrike Haase 19.06.2007 Seminar "Gute Ideen in der theoretischen Biologie / Systembiologie" Signal processing in nervous system Nerve cell and
Signal processing in nervous system - Hodgkin-Huxley model Ulrike Haase 19.06.2007 Seminar "Gute Ideen in der theoretischen Biologie / Systembiologie" Signal processing in nervous system Nerve cell and
Nervous Lecture Test Questions Set 2
 Nervous Lecture Test Questions Set 2 1. The role of chloride in a resting membrane potential: a. creates resting potential b. indirectly causes repolarization c. stabilization of sodium d. it has none,
Nervous Lecture Test Questions Set 2 1. The role of chloride in a resting membrane potential: a. creates resting potential b. indirectly causes repolarization c. stabilization of sodium d. it has none,
The Nervous System and the Sodium-Potassium Pump
 The Nervous System and the Sodium-Potassium Pump 1. Define the following terms: Ion: A Student Activity on Membrane Potentials Cation: Anion: Concentration gradient: Simple diffusion: Sodium-Potassium
The Nervous System and the Sodium-Potassium Pump 1. Define the following terms: Ion: A Student Activity on Membrane Potentials Cation: Anion: Concentration gradient: Simple diffusion: Sodium-Potassium
Ch 7. The Nervous System 7.1 & 7.2
 Ch 7 The Nervous System 7.1 & 7.2 SLOs Describe the different types of neurons and supporting cells, and identify their functions. Identify the myelin sheath and describe how it is formed in the CNS and
Ch 7 The Nervous System 7.1 & 7.2 SLOs Describe the different types of neurons and supporting cells, and identify their functions. Identify the myelin sheath and describe how it is formed in the CNS and
MEMBRANE POTENTIALS AND ACTION POTENTIALS:
 University of Jordan Faculty of Medicine Department of Physiology & Biochemistry Medical students, 2017/2018 +++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ Review: Membrane physiology
University of Jordan Faculty of Medicine Department of Physiology & Biochemistry Medical students, 2017/2018 +++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ Review: Membrane physiology
Peripheral Nerve II. Amelyn Ramos Rafael, MD. Anatomical considerations
 Peripheral Nerve II Amelyn Ramos Rafael, MD Anatomical considerations 1 Physiologic properties of the nerve Irritability of the nerve A stimulus applied on the nerve causes the production of a nerve impulse,
Peripheral Nerve II Amelyn Ramos Rafael, MD Anatomical considerations 1 Physiologic properties of the nerve Irritability of the nerve A stimulus applied on the nerve causes the production of a nerve impulse,
3 Action Potentials - Brutal Approximations
 Physics 172/278 - David Kleinfeld - Fall 2004; Revised Winter 2015 3 Action Potentials - Brutal Approximations The Hodgkin-Huxley equations for the behavior of the action potential in squid, and similar
Physics 172/278 - David Kleinfeld - Fall 2004; Revised Winter 2015 3 Action Potentials - Brutal Approximations The Hodgkin-Huxley equations for the behavior of the action potential in squid, and similar
Multi-loop Circuits and Kirchoff's Rules
 1 of 8 01/21/2013 12:50 PM Multi-loop Circuits and Kirchoff's Rules 7-13-99 Before talking about what a multi-loop circuit is, it is helpful to define two terms, junction and branch. A junction is a point
1 of 8 01/21/2013 12:50 PM Multi-loop Circuits and Kirchoff's Rules 7-13-99 Before talking about what a multi-loop circuit is, it is helpful to define two terms, junction and branch. A junction is a point
1 Hodgkin-Huxley Theory of Nerve Membranes: The FitzHugh-Nagumo model
 1 Hodgkin-Huxley Theory of Nerve Membranes: The FitzHugh-Nagumo model Alan Hodgkin and Andrew Huxley developed the first quantitative model of the propagation of an electrical signal (the action potential)
1 Hodgkin-Huxley Theory of Nerve Membranes: The FitzHugh-Nagumo model Alan Hodgkin and Andrew Huxley developed the first quantitative model of the propagation of an electrical signal (the action potential)
9 Generation of Action Potential Hodgkin-Huxley Model
 9 Generation of Action Potential Hodgkin-Huxley Model (based on chapter 2, W.W. Lytton, Hodgkin-Huxley Model) 9. Passive and active membrane models In the previous lecture we have considered a passive
9 Generation of Action Potential Hodgkin-Huxley Model (based on chapter 2, W.W. Lytton, Hodgkin-Huxley Model) 9. Passive and active membrane models In the previous lecture we have considered a passive
Physiology Unit 2. MEMBRANE POTENTIALS and SYNAPSES
 Physiology Unit 2 MEMBRANE POTENTIALS and SYNAPSES In Physiology Today Ohm s Law I = V/R Ohm s law: the current through a conductor between two points is directly proportional to the voltage across the
Physiology Unit 2 MEMBRANE POTENTIALS and SYNAPSES In Physiology Today Ohm s Law I = V/R Ohm s law: the current through a conductor between two points is directly proportional to the voltage across the
Neuron Structure. Why? Model 1 Parts of a Neuron. What are the essential structures that make up a neuron?
 Why? Neuron Structure What are the essential structures that make up a neuron? Cells are specialized for different functions in multicellular organisms. In animals, one unique kind of cell helps organisms
Why? Neuron Structure What are the essential structures that make up a neuron? Cells are specialized for different functions in multicellular organisms. In animals, one unique kind of cell helps organisms
Using Sigmoid and Double-Sigmoid Functions for Earth-States Transitions. 1. L. David Roper. ABSTRACT
 Using Sigmoid and Double-Sigmoid Functions for Earth-States Transitions. 1 by L. David Roper roperld@vt.edu ABSTRACT The sigmoid and double-sigmoid functions are very useful for representing earth-states
Using Sigmoid and Double-Sigmoid Functions for Earth-States Transitions. 1 by L. David Roper roperld@vt.edu ABSTRACT The sigmoid and double-sigmoid functions are very useful for representing earth-states
Cell membrane resistance and capacitance
 Cell membrane resistance and capacitance 1 Two properties of a cell membrane gives rise to two passive electrical properties: Resistance: Leakage pathways allow inorganic ions to cross the membrane. Capacitance:
Cell membrane resistance and capacitance 1 Two properties of a cell membrane gives rise to two passive electrical properties: Resistance: Leakage pathways allow inorganic ions to cross the membrane. Capacitance:
Errata for Bioelectricity: A Quantitative Approach, 3rd edition by Robert Plonsey and Roger C. Barr (Springer 2007)
") Errata for Bioelectricity: A Quantitative Approach, 3rd edition by Robert Plonsey and Roger C. Barr (Springer 2007) Frederick J. Vetter Department of Electrical, Computer and Biomedical Engineering University
Errata for Bioelectricity: A Quantitative Approach, 3rd edition by Robert Plonsey and Roger C. Barr (Springer 2007) Frederick J. Vetter Department of Electrical, Computer and Biomedical Engineering University
Nerve Signal Conduction. Resting Potential Action Potential Conduction of Action Potentials
 Nerve Signal Conduction Resting Potential Action Potential Conduction of Action Potentials Resting Potential Resting neurons are always prepared to send a nerve signal. Neuron possesses potential energy
Nerve Signal Conduction Resting Potential Action Potential Conduction of Action Potentials Resting Potential Resting neurons are always prepared to send a nerve signal. Neuron possesses potential energy
Nervous Systems: Neuron Structure and Function
 Nervous Systems: Neuron Structure and Function Integration An animal needs to function like a coherent organism, not like a loose collection of cells. Integration = refers to processes such as summation
Nervous Systems: Neuron Structure and Function Integration An animal needs to function like a coherent organism, not like a loose collection of cells. Integration = refers to processes such as summation
Neurons, Synapses, and Signaling
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 48 Neurons, Synapses, and Signaling
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 48 Neurons, Synapses, and Signaling
Modeling Action Potentials in Cell Processes
 Modeling Action Potentials in Cell Processes Chelsi Pinkett, Jackie Chism, Kenneth Anderson, Paul Klockenkemper, Christopher Smith, Quarail Hale Tennessee State University Action Potential Models Chelsi
Modeling Action Potentials in Cell Processes Chelsi Pinkett, Jackie Chism, Kenneth Anderson, Paul Klockenkemper, Christopher Smith, Quarail Hale Tennessee State University Action Potential Models Chelsi
NeuroPhysiology and Membrane Potentials. The Electrochemical Gradient
 NeuroPhysiology and Membrane Potentials Communication by neurons is based on changes in the membrane s permeability to ions This depends on the presence of specific membrane ion channels and the presence
NeuroPhysiology and Membrane Potentials Communication by neurons is based on changes in the membrane s permeability to ions This depends on the presence of specific membrane ion channels and the presence
Electrophysiological Modeling of Membranes and Cells
 Bioeng 6460 Electrophysiology and Bioelectricity Electrophysiological Modeling of Membranes and Cells Frank B. Sachse fs@cvrti.utah.edu Overview Motivation and Principles Electrical Modeling of Membranes
Bioeng 6460 Electrophysiology and Bioelectricity Electrophysiological Modeling of Membranes and Cells Frank B. Sachse fs@cvrti.utah.edu Overview Motivation and Principles Electrical Modeling of Membranes
1 Single neuron computation and the separation of processing and signaling
 Physics 178/278 - David Kleinfeld - Winter 2017 - Week 1 1 Single neuron computation and the separation of processing and signaling This overview serves to connect simple notions of statistical physics
Physics 178/278 - David Kleinfeld - Winter 2017 - Week 1 1 Single neuron computation and the separation of processing and signaling This overview serves to connect simple notions of statistical physics
Probabilistic Modeling of Action Potential Generation by Neurons
 Probabilistic Modeling of Action Potential Generation by Neurons Student: Bobby Sena under the direction of Professors Helen Wearing and Eric Toolson University of New Mexico bsena@unm.edu May 6, 21 Abstract
Probabilistic Modeling of Action Potential Generation by Neurons Student: Bobby Sena under the direction of Professors Helen Wearing and Eric Toolson University of New Mexico bsena@unm.edu May 6, 21 Abstract
Lojayn Salah. Zaid R Al Najdawi. Mohammad-Khatatbeh
 7 Lojayn Salah Zaid R Al Najdawi Mohammad-Khatatbeh Salam everyone, I made my best to make this sheet clear enough to be easily understood let the party begin :P Quick Revision about the previous lectures:
7 Lojayn Salah Zaid R Al Najdawi Mohammad-Khatatbeh Salam everyone, I made my best to make this sheet clear enough to be easily understood let the party begin :P Quick Revision about the previous lectures: