1 Single neuron computation and the separation of processing and signaling
|
|
|
- May Stevenson
- 6 years ago
- Views:
Transcription
1 Physics 178/278 - David Kleinfeld - Winter Week 1 1 Single neuron computation and the separation of processing and signaling This overview serves to connect simple notions of statistical physics with neuronal signal integration, decision making, and communication. 1.1 Ions and Voltages A good place to start is with an overview of how sensitive and robust nervous systems are. We consider a fundamental issue, the scale of the potentials across cell membranes relative to the thermal noise floor. We further discuss how these potentials are used to form pulses, or action potentials, that allow one cell to communicate with another. We shall see that the Boltzman energy serves to define a band of voltages that can be used to integrate and process nervous activity, and that the same energy acts as a barrier to isolate integration of inputs from output and communication. 1.2 The Nernst Potential Bags comprised of lipid walls provide a means to separate high concentrations from low concentrations of ions. Why is this useful? If we recall a little bit of thermodynamics, we realize that a difference in ion concentration leads to a potential, and thus work can be done. Thus, one could conceivably build a device that is capable of communicating to all agents along the surface of a bag, or some topological equivalent, that a disturbance has occurred at a point. Let s run through this. Consider a world that consists of two compartments, labeled in and out. The compartments are separated by a thin wall. To make this situation somewhat real, we will fill both compartments with water and consider the diffusion of ions in the water. We take these ions to be Na + and Cl, the ions that result from dissolving ordinary table salt. On the inside of the box, the concentration of ions is denoted [Na + ] in and [Cl ] in and on the outside they are denoted [Na + ] out and [Cl ] out, where concentration is in units of molecules/cm 3 or, in chemistry, in units of moles per liter or Molar(M). To get a feel for the scale of moles/liter, lets put it into terms relevant for the size of a cell, i.e., ions per cubic micrometer. In a biological cell, the ion concentration is about 0.15 M, so we have about 10 8 ions/µm 3 in a cell. Suppose we set the box up so that, initially, [Na + ] in = [Cl ] in and [Na + ] out = [Cl ] out. This insures that individual compartments are electrically neutral. Further, we impose [Na + ] out > [Na + ] out, so that the ion concentrations are initially greater on the outside than the inside. Since there is a wall between the two sides, there is clearly no interaction between the two compartments. 1
2 Suppose we now remove the wall in entirety. What happens? The Na + ions will flow down their concentration difference, so that ultimately [Na + ] is the same on both sides of the membrane. Similarly, the Cl ions will flow down their gradient. Thus after some time the concentration of ions is the same on both sides, so that no energy is extracted from the concentration difference. A side remark is that there is a transient potential difference if the mobility of Na + and Cl are different. What is the trick necessary to get the concentration difference across the two sides of the wall to turn into a sustained electrical potential? The idea is to make a hole that allows only one kind of ion to pass. This is called an ion selective pore. To be concrete, we open up a hole that allows [Na + ] ions, but not [Cl ] ions, to pass. This is a Na + selective pore, or Na + selective channel. Recall that [Na + ] is higher outside than inside. The process works as follows: Initially, the [Na + ] moves down its concentration gradient, driven by diffusion. As Na + ions move across the wall, the solutions in the two compartments are no longer electrically neutral. Positive charge (from the Na + ) leaves the outside and builds up on the inside. This leads to an electric field across the wall. The electric fields points from the inside to the outside. direction that opposes motion of additional Na + ions. This field is the In time, the electric field caused by the initial movement of ions points from the inside to the outside. This field is the direction that opposes motion of additional Na + ions and will prevent any more Na + ions from moving. As this point the system is in equilibrium. The result is that the concentration difference in Na + ions across the wall has been used to form a difference in electrical potential across the cell. The relation between concentration differences and electrical potential is given by the Nernst equation, which equates the electrical potential, ev, with the chemical potential, µ, caused by the concentration difference, i.e., µ = ( ) F N T,V = k B T lnz N = k BT ln ξn N! N = k BT lnn + constant (1.1) for N >> 1 so that we can approximate N! by Sterling s formula. Thus V = k BT e ln[na+ ] out [Na + ] in (1.2) We see immediately that V is on the order of k BT 25mV. Before we worry e about details, let s think about how big this is on the scale of the electrical noise expected for this process. 2
3 1.3 Thermal Fluctuations in Voltage Because there is a pore, there is a conductance G across the cell. This leads to a fluctuation in the potential, known as the Johnson noise, of size δv = 4kB T f G = kb T C = ( kb T ) ( L e ɛ m ) e 4πa = 1 2 2a ( kb T ) ( ) e 1 e c m π (1.3) where we used f = 0 df (2πf(C/G)) 2 = G 4C. (1.4) Further, C = ɛ m (area/thickness), so that for a thin dielectric sphere of thickness L and radius a, C = ɛ m 4πa 2 L. For most all cells, the ratio ɛm L is c m ɛ m L F 1.0x10 14 µm. (1.5) 2 For a neuron of radius a = 20µm, the noise level is found to be δv 20µV. The important result that for cells the noise level is much less than the thermal voltage k B T/e, where k B T 25mV (1.6) e Only at the smallest structure, the synaptic vesicle with radius a 10 2 µm = 10nm, will the noise level approach the thermal voltage, so that for a 10 2 µm we have δv 10mV. Actually, the case of vesicles is interesting, in that we can turn the argument around and look at the fluctuation in the number of ions across the cell. In synaptic vesicles, the membrane potential V is set by a hydrogen ion, or ph gradient. Then V = k BT e ] out ln[h+ = k BT [H + ] in e (ph in ph out ). (1.7) Typically, ph in 5 and ph out 7.5. The variance in the transmembrane voltage in terms of ion concentration is δv = V δ[h + ] [H + in = k BT δ[h + ] in (1.8) ] in e [H + ] in We equate noise level this with the expression for Johnson noise to get δ[h + ] in [H + ] in = 4e 2 ( k B T C = e k B T ) 1 c m e πa 2 (1.9) An interesting number is the value of the radius a for which the fluctuations in ion concentration are of order unity, i.e., δ[h+ ] in [H + ] in 1. We call this a crit, where 3
4 a crit = ( ) 1 e e 30nm (1.10) π k B T c m This is on the small side of the observed radius of vesicles, or synaptosomes, which is not too bad as an estimate of the smallest cell. A slightly different way to look at the noise issue is to recall that Q = Ne = C V, where N is the number of ions responsible for the net charge imbalance across the membrane. If we substitute this expression for C into the formula for Johnson noise, the Signal-to-Noise ratio for the cell is Signal Noise V δ ( V ) = 1 2 N V k B T/e (1.11) The dependence on the square root of N is intuitive. Note that the above expressions need to be modified if the conductance is not set by a static pore, but a pore that fluctuates open and closed. In the latter case, the fluctuations cut off at some high temporal frequency. But the major story holds. In the larger picture, the cell membrane may have selectively permeable channels to more than one ion. Thus the final potential may represent a steady state rather than an equilibrium value. In fact, nature uses two ions, K + and Na +, to set two potentials, one low and one high. The presence of a voltage-dependent (nonlinear) conductance, one of the major findings of the century, allows the cell to switch between these two levels. Let s review the gist of this - the details will be the topic of another lecture. In the simplest, or Hodgkin Huxley cell, there are three major conductances, those due to K +, Na +, and Cl. The equilibrium potential for each of these ions is V K + = k BT ln [K+ ] out e [K + ] in 75mV V Na + = k BT ln [Na+ ] out e [Na + ] in V Cl = k BT ln [Cl ] out e [Cl ] in +50mV 55mV The steady state potential for a system with these three ions and associated membrane permeabilities P Na +, P K +, and P Cl, is V SS = k BT e lnp K+[K + ] out + P Na+[Na + ] out + P Cl [Cl ] in P K +[K + ] in + P Na +[Na + ] in + P Cl [Cl ] out 50mV (1.12) This relation is derived by considering the diffusion equation for the total flux of ions through the cell membrane and is found is standard textbooks an the Nerst- Plank equation. The important point is that the steady state potential is dominated by the ion that has the maximal conductance. The resting level is dominated by the K + conductance. 4
5 1.4 Fundamentals of the Action Potential The essential ingredient that allows signaling is that the conductance for Na + is voltage dependent. This is reminiscent of the drain-to-source conductance in a FET, which is a nonlinear function of the voltage to the gate. A nice way to think of this, which we will discuss in detail, is that the currentvoltage relationship has an essential nonlinearity. Thus for small disturbances of the membrane potential, the cell returns to the resting potential. However, for current injections beyond some critical value, the potential will jump to a new equilibrium point. A simplified model makes use of a voltage dependent change in the conductance for one of two ions. To be concrete, we take a cell with a solely Ohmic potassium current, G K +, and a voltage dependent sodium conductance, G Na +(V ), that has a value of zero below a threshold potential, V th, and that is constant above V th with value G Na +(V ). Thus we have a current-voltage relation given by I(V ) = { GK +V G K +V K + if V < V th (G K +G Na +(V )) V (G K +V K + + G Na +(V )V Na +) if V > V th where, in this approximation, V Na + and V K + are the Na + and K + Nernst potential for sodium and potassium, respectfully. This relation is discontinuous at V th and Ohmic below and above this potential. We consider a pulse of current that causes the cell to change from the lower to the upper curve. This represents the front of the action potential. The shift in potential from V = V K + to V = G K + V K + +G Na + (V ) G K + +G Na + (V ) occurs in roughly 10 4 s. On the longer time scale of 10 3 s, relaxation processes associated with the Na + current and the activation of an additional voltage dependent K + current cause the front to decay, so we are left with a pulse. The critical lesson is that neurons use two voltage levels, and at least one voltage dependent conductance, to shift between the two levels. We now know how neurons signal, and that the signaling, at least in principle, operates well above the physical limits to membrane noise. We put off the issue of variability in the transmission of spikes between cells - suffice it to say that at some synapses, e.g., those formed as calyces, the failure rate is very low indeed. 1.5 Separation of Dendritic Integration and Communication Our final point concerns how a neuron performs logic, which is to say how it separates the integration of synaptic inputs from the decision making that leads to the production of an action potential. We require a band of voltages over which the cell can integrate, that is summate, synaptic inputs. The range of this band must clearly be larger than the scale of thermal noise and also large compared to the activation of the Na + -based action potential. Given the experimental fact that the Na + channel turns on through the action of 4 charges, the range of synaptic integration 5
6 is expected to exceed V > k B T/4e 6 mv. This range corresponds to the difference between the K + reversal potential (the lowest voltage for inhibitory inputs) and the activation of the Na + -based action potential, a range of about 1-1/2-times k B T/e 35 mv. Others may argue that the level is the reversal potential for Cl, the dominant inhibitory input in mammals. In this case the range is about 1-times k B T/e 25 mv. Either way, we see that the scale for integration is always of order k B T/e and is always large compared to the noise level across the membrane. How large are the post-synaptic potentials that impinge on the cell? This distribution has been measured by a number of investigators in pair-electrode measurements, where current is injected into the presynaptic cell to induce an action potential and measured in the post-synaptic. cell. The typical values are around 0.5 mv or less, or a very small fraction of k B T/4e. But a small percentage, maybe 5 %, come in at a few millivolts. Thus coactivation of a small number of inputs can, in principle, drive a neuron to spike. The issue is an open research question; see handout on Mrsic-Flogel data. The above argument suggest that the cell has headroom for integration without firing an action potential for insignificant changes in input. We now consider how a cell can isolate the synapse from integration. The idea is that the action potential must be large enough activate a process whose turn-on occurs far from the range of synaptic activation. This implies that the activation of synaptic transition, which occurs over a range voltage, must be separated from the range of voltage V > k B T/4e 6 mv that governs the activation of the inward Na + current. Synaptic activation depends on a chemical cascade that is initiated by the activation of a high-threshold voltage gated Ca 2+ current. This current peaks at intracellular potentials of about + 5 mv, less than V Na + of course Thus there is headroom of order of 2k B T/e 50 mv that separates the turn-on of the action potential from the turn-on of synaptic transmission, so that dendritic integration per se cannot lead to synaptic release, or communication, unit the threshold for spiking is crossed. 6
 Single-synapse analysis of a diverse synapse population: Proteomic imaging methods and markers.")
7 Micheva KD, Smith SJ (2007) Array tomography: A new tool for imaging the molecular architecture and ultrastructure of neural circuits. Neuron 55: Micheva KD, Busse B, Weiler NC, O'Rourke N, Smith SJ (2010) Single-synapse analysis of a diverse synapse population: Proteomic imaging methods and markers. Neuron 68:
8

9 Equivalent Circuit Model of a Neuron è


10 Resting Potential: Electrochemical Equilibrium!!!!!!!"#!!!!"!!!"

11 Common Ionic Concentrations E K =!75 mv E Na = +55 mv E Cl =!70 mv E Ca " +150 mv
: I K =!")
12 Out of Equilibrium For V > V X current flows into cell: I X > 0 For V < V X current flows out of cell: I X < 0 I-V curve (current-voltage): I K =! K ( V " V ) K

13 Multiple Kinds of Ions Several I-V curves in parallel: New equivalent circuit:
14 Compact Cell Model (isopotential voltage) C dv dt ( Vm V R m = I r ) inj + V r + I inj R V r I inj t
15 Real Data from Lobster Axon
16
17
18 chapt_1_neoctx_pyramidal.eps
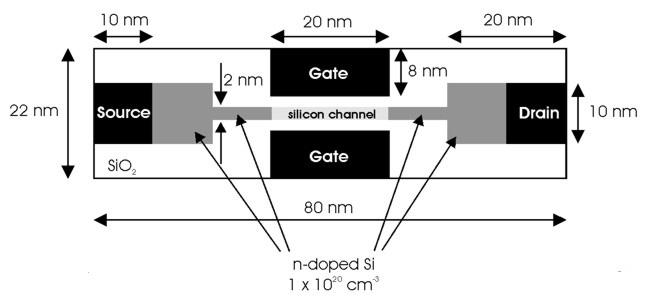
19 chapt_1_nanofet.eps


20 Voltage gated Potassium Channel Analysis of MacKinnon data by Shaw
21 N-type calcium current in presynaptic terminals initiates synaptic vessel fusion

22
23
Action Potential Propagation
 Action Potential Propagation 2 Action Potential is a transient alteration of transmembrane voltage (or membrane potential) across an excitable membrane generated by the activity of voltage-gated ion channels.
Action Potential Propagation 2 Action Potential is a transient alteration of transmembrane voltage (or membrane potential) across an excitable membrane generated by the activity of voltage-gated ion channels.
Cell membrane resistance and capacitance
 Cell membrane resistance and capacitance 1 Two properties of a cell membrane gives rise to two passive electrical properties: Resistance: Leakage pathways allow inorganic ions to cross the membrane. Capacitance:
Cell membrane resistance and capacitance 1 Two properties of a cell membrane gives rise to two passive electrical properties: Resistance: Leakage pathways allow inorganic ions to cross the membrane. Capacitance:
9 Generation of Action Potential Hodgkin-Huxley Model
 9 Generation of Action Potential Hodgkin-Huxley Model (based on chapter 12, W.W. Lytton, Hodgkin-Huxley Model) 9.1 Passive and active membrane models In the previous lecture we have considered a passive
9 Generation of Action Potential Hodgkin-Huxley Model (based on chapter 12, W.W. Lytton, Hodgkin-Huxley Model) 9.1 Passive and active membrane models In the previous lecture we have considered a passive
Neurons and the membrane potential. N500 John Beggs 23 Aug, 2016
 Neurons and the membrane potential N500 John Beggs 23 Aug, 2016 My background, briefly Neurons Structural elements of a typical neuron Figure 1.2 Some nerve cell morphologies found in the human
Neurons and the membrane potential N500 John Beggs 23 Aug, 2016 My background, briefly Neurons Structural elements of a typical neuron Figure 1.2 Some nerve cell morphologies found in the human
Electrophysiology of the neuron
 School of Mathematical Sciences G4TNS Theoretical Neuroscience Electrophysiology of the neuron Electrophysiology is the study of ionic currents and electrical activity in cells and tissues. The work of
School of Mathematical Sciences G4TNS Theoretical Neuroscience Electrophysiology of the neuron Electrophysiology is the study of ionic currents and electrical activity in cells and tissues. The work of
Action Potentials and Synaptic Transmission Physics 171/271
 Action Potentials and Synaptic Transmission Physics 171/271 Flavio Fröhlich (flavio@salk.edu) September 27, 2006 In this section, we consider two important aspects concerning the communication between
Action Potentials and Synaptic Transmission Physics 171/271 Flavio Fröhlich (flavio@salk.edu) September 27, 2006 In this section, we consider two important aspects concerning the communication between
BME 5742 Biosystems Modeling and Control
 BME 5742 Biosystems Modeling and Control Hodgkin-Huxley Model for Nerve Cell Action Potential Part 1 Dr. Zvi Roth (FAU) 1 References Hoppensteadt-Peskin Ch. 3 for all the mathematics. Cooper s The Cell
BME 5742 Biosystems Modeling and Control Hodgkin-Huxley Model for Nerve Cell Action Potential Part 1 Dr. Zvi Roth (FAU) 1 References Hoppensteadt-Peskin Ch. 3 for all the mathematics. Cooper s The Cell
LESSON 2.2 WORKBOOK How do our axons transmit electrical signals?
 LESSON 2.2 WORKBOOK How do our axons transmit electrical signals? This lesson introduces you to the action potential, which is the process by which axons signal electrically. In this lesson you will learn
LESSON 2.2 WORKBOOK How do our axons transmit electrical signals? This lesson introduces you to the action potential, which is the process by which axons signal electrically. In this lesson you will learn
2.6 The Membrane Potential
 2.6: The Membrane Potential 51 tracellular potassium, so that the energy stored in the electrochemical gradients can be extracted. Indeed, when this is the case experimentally, ATP is synthesized from
2.6: The Membrane Potential 51 tracellular potassium, so that the energy stored in the electrochemical gradients can be extracted. Indeed, when this is the case experimentally, ATP is synthesized from
BIOELECTRIC PHENOMENA
 Chapter 11 BIOELECTRIC PHENOMENA 11.3 NEURONS 11.3.1 Membrane Potentials Resting Potential by separation of charge due to the selective permeability of the membrane to ions From C v= Q, where v=60mv and
Chapter 11 BIOELECTRIC PHENOMENA 11.3 NEURONS 11.3.1 Membrane Potentials Resting Potential by separation of charge due to the selective permeability of the membrane to ions From C v= Q, where v=60mv and
Lecture 10 : Neuronal Dynamics. Eileen Nugent
 Lecture 10 : Neuronal Dynamics Eileen Nugent Origin of the Cells Resting Membrane Potential: Nernst Equation, Donnan Equilbrium Action Potentials in the Nervous System Equivalent Electrical Circuits and
Lecture 10 : Neuronal Dynamics Eileen Nugent Origin of the Cells Resting Membrane Potential: Nernst Equation, Donnan Equilbrium Action Potentials in the Nervous System Equivalent Electrical Circuits and
Membrane Potentials, Action Potentials, and Synaptic Transmission. Membrane Potential
 Cl Cl - - + K + K+ K + K Cl - 2/2/15 Membrane Potentials, Action Potentials, and Synaptic Transmission Core Curriculum II Spring 2015 Membrane Potential Example 1: K +, Cl - equally permeant no charge
Cl Cl - - + K + K+ K + K Cl - 2/2/15 Membrane Potentials, Action Potentials, and Synaptic Transmission Core Curriculum II Spring 2015 Membrane Potential Example 1: K +, Cl - equally permeant no charge
Structure and Measurement of the brain lecture notes
 Structure and Measurement of the brain lecture notes Marty Sereno 2009/2010!"#$%&'(&#)*%$#&+,'-&.)"/*"&.*)*-'(0&1223 Neurons and Models Lecture 1 Topics Membrane (Nernst) Potential Action potential/voltage-gated
Structure and Measurement of the brain lecture notes Marty Sereno 2009/2010!"#$%&'(&#)*%$#&+,'-&.)"/*"&.*)*-'(0&1223 Neurons and Models Lecture 1 Topics Membrane (Nernst) Potential Action potential/voltage-gated
3 Detector vs. Computer
 1 Neurons 1. The detector model. Also keep in mind this material gets elaborated w/the simulations, and the earliest material is often hardest for those w/primarily psych background. 2. Biological properties
1 Neurons 1. The detector model. Also keep in mind this material gets elaborated w/the simulations, and the earliest material is often hardest for those w/primarily psych background. 2. Biological properties
Neuron. Detector Model. Understanding Neural Components in Detector Model. Detector vs. Computer. Detector. Neuron. output. axon
 Neuron Detector Model 1 The detector model. 2 Biological properties of the neuron. 3 The computational unit. Each neuron is detecting some set of conditions (e.g., smoke detector). Representation is what
Neuron Detector Model 1 The detector model. 2 Biological properties of the neuron. 3 The computational unit. Each neuron is detecting some set of conditions (e.g., smoke detector). Representation is what
Passive Membrane Properties
 Passive Membrane Properties Communicating through a leaky garden hose... Topics I Introduction & Electrochemical Gradients Passive Membrane Properties Action Potentials Voltage-Gated Ion Channels Topics
Passive Membrane Properties Communicating through a leaky garden hose... Topics I Introduction & Electrochemical Gradients Passive Membrane Properties Action Potentials Voltage-Gated Ion Channels Topics
Channels can be activated by ligand-binding (chemical), voltage change, or mechanical changes such as stretch.
, voltage change, or mechanical changes such as stretch.") 1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
Basic elements of neuroelectronics -- membranes -- ion channels -- wiring
 Computing in carbon Basic elements of neuroelectronics -- membranes -- ion channels -- wiring Elementary neuron models -- conductance based -- modelers alternatives Wires -- signal propagation -- processing
Computing in carbon Basic elements of neuroelectronics -- membranes -- ion channels -- wiring Elementary neuron models -- conductance based -- modelers alternatives Wires -- signal propagation -- processing
Mathematical Foundations of Neuroscience - Lecture 3. Electrophysiology of neurons - continued
 Mathematical Foundations of Neuroscience - Lecture 3. Electrophysiology of neurons - continued Filip Piękniewski Faculty of Mathematics and Computer Science, Nicolaus Copernicus University, Toruń, Poland
Mathematical Foundations of Neuroscience - Lecture 3. Electrophysiology of neurons - continued Filip Piękniewski Faculty of Mathematics and Computer Science, Nicolaus Copernicus University, Toruń, Poland
Supratim Ray
 Supratim Ray sray@cns.iisc.ernet.in Biophysics of Action Potentials Passive Properties neuron as an electrical circuit Passive Signaling cable theory Active properties generation of action potential Techniques
Supratim Ray sray@cns.iisc.ernet.in Biophysics of Action Potentials Passive Properties neuron as an electrical circuit Passive Signaling cable theory Active properties generation of action potential Techniques
2002NSC Human Physiology Semester Summary
 2002NSC Human Physiology Semester Summary Griffith University, Nathan Campus Semester 1, 2014 Topics include: - Diffusion, Membranes & Action Potentials - Fundamentals of the Nervous System - Neuroanatomy
2002NSC Human Physiology Semester Summary Griffith University, Nathan Campus Semester 1, 2014 Topics include: - Diffusion, Membranes & Action Potentials - Fundamentals of the Nervous System - Neuroanatomy
Ch. 5. Membrane Potentials and Action Potentials
 Ch. 5. Membrane Potentials and Action Potentials Basic Physics of Membrane Potentials Nerve and muscle cells: Excitable Capable of generating rapidly changing electrochemical impulses at their membranes
Ch. 5. Membrane Potentials and Action Potentials Basic Physics of Membrane Potentials Nerve and muscle cells: Excitable Capable of generating rapidly changing electrochemical impulses at their membranes
The Membrane Potential
 The Membrane Potential Graphics are used with permission of: adam.com (http://www.adam.com/) Benjamin Cummings Publishing Co (http://www.aw.com/bc) ** It is suggested that you carefully label each ion
The Membrane Potential Graphics are used with permission of: adam.com (http://www.adam.com/) Benjamin Cummings Publishing Co (http://www.aw.com/bc) ** It is suggested that you carefully label each ion
PNS Chapter 7. Membrane Potential / Neural Signal Processing Spring 2017 Prof. Byron Yu
 PNS Chapter 7 Membrane Potential 18-698 / 42-632 Neural Signal Processing Spring 2017 Prof. Byron Yu Roadmap Introduction to neuroscience Chapter 1 The brain and behavior Chapter 2 Nerve cells and behavior
PNS Chapter 7 Membrane Potential 18-698 / 42-632 Neural Signal Processing Spring 2017 Prof. Byron Yu Roadmap Introduction to neuroscience Chapter 1 The brain and behavior Chapter 2 Nerve cells and behavior
The Membrane Potential
 The Membrane Potential Graphics are used with permission of: Pearson Education Inc., publishing as Benjamin Cummings (http://www.aw-bc.com) ** It is suggested that you carefully label each ion channel
The Membrane Potential Graphics are used with permission of: Pearson Education Inc., publishing as Benjamin Cummings (http://www.aw-bc.com) ** It is suggested that you carefully label each ion channel
Neurons and Nervous Systems
 34 Neurons and Nervous Systems Concept 34.1 Nervous Systems Consist of Neurons and Glia Nervous systems have two categories of cells: Neurons, or nerve cells, are excitable they generate and transmit electrical
34 Neurons and Nervous Systems Concept 34.1 Nervous Systems Consist of Neurons and Glia Nervous systems have two categories of cells: Neurons, or nerve cells, are excitable they generate and transmit electrical
/639 Final Solutions, Part a) Equating the electrochemical potentials of H + and X on outside and inside: = RT ln H in
 Equating the electrochemical potentials of H + and X on outside and inside: = RT ln H in") 580.439/639 Final Solutions, 2014 Question 1 Part a) Equating the electrochemical potentials of H + and X on outside and inside: RT ln H out + zf 0 + RT ln X out = RT ln H in F 60 + RT ln X in 60 mv =
580.439/639 Final Solutions, 2014 Question 1 Part a) Equating the electrochemical potentials of H + and X on outside and inside: RT ln H out + zf 0 + RT ln X out = RT ln H in F 60 + RT ln X in 60 mv =
Compartmental Modelling
 Modelling Neurons Computing and the Brain Compartmental Modelling Spring 2010 2 1 Equivalent Electrical Circuit A patch of membrane is equivalent to an electrical circuit This circuit can be described
Modelling Neurons Computing and the Brain Compartmental Modelling Spring 2010 2 1 Equivalent Electrical Circuit A patch of membrane is equivalent to an electrical circuit This circuit can be described
Biomedical Instrumentation
 ELEC ENG 4BD4: Biomedical Instrumentation Lecture 5 Bioelectricity 1. INTRODUCTION TO BIOELECTRICITY AND EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
ELEC ENG 4BD4: Biomedical Instrumentation Lecture 5 Bioelectricity 1. INTRODUCTION TO BIOELECTRICITY AND EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
Ionic basis of the resting membrane potential. Foundations in Neuroscience I, Oct
 Ionic basis of the resting membrane potential Foundations in Neuroscience I, Oct 3 2017 The next 4 lectures... - The resting membrane potential (today) - The action potential - The neural mechanisms behind
Ionic basis of the resting membrane potential Foundations in Neuroscience I, Oct 3 2017 The next 4 lectures... - The resting membrane potential (today) - The action potential - The neural mechanisms behind
Control and Integration. Nervous System Organization: Bilateral Symmetric Animals. Nervous System Organization: Radial Symmetric Animals
 Control and Integration Neurophysiology Chapters 10-12 Nervous system composed of nervous tissue cells designed to conduct electrical impulses rapid communication to specific cells or groups of cells Endocrine
Control and Integration Neurophysiology Chapters 10-12 Nervous system composed of nervous tissue cells designed to conduct electrical impulses rapid communication to specific cells or groups of cells Endocrine
CELL BIOLOGY - CLUTCH CH. 9 - TRANSPORT ACROSS MEMBRANES.
 !! www.clutchprep.com K + K + K + K + CELL BIOLOGY - CLUTCH CONCEPT: PRINCIPLES OF TRANSMEMBRANE TRANSPORT Membranes and Gradients Cells must be able to communicate across their membrane barriers to materials
!! www.clutchprep.com K + K + K + K + CELL BIOLOGY - CLUTCH CONCEPT: PRINCIPLES OF TRANSMEMBRANE TRANSPORT Membranes and Gradients Cells must be able to communicate across their membrane barriers to materials
Biosciences in the 21st century
 Biosciences in the 21st century Lecture 1: Neurons, Synapses, and Signaling Dr. Michael Burger Outline: 1. Why neuroscience? 2. The neuron 3. Action potentials 4. Synapses 5. Organization of the nervous
Biosciences in the 21st century Lecture 1: Neurons, Synapses, and Signaling Dr. Michael Burger Outline: 1. Why neuroscience? 2. The neuron 3. Action potentials 4. Synapses 5. Organization of the nervous
Membrane Protein Channels
 Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
Nerve Signal Conduction. Resting Potential Action Potential Conduction of Action Potentials
 Nerve Signal Conduction Resting Potential Action Potential Conduction of Action Potentials Resting Potential Resting neurons are always prepared to send a nerve signal. Neuron possesses potential energy
Nerve Signal Conduction Resting Potential Action Potential Conduction of Action Potentials Resting Potential Resting neurons are always prepared to send a nerve signal. Neuron possesses potential energy
Phys498BIO; Prof. Paul Selvin Hw #9 Assigned Wed. 4/18/12: Due 4/25/08
 1. Ionic Movements Across a Permeable Membrane: The Nernst Potential. In class we showed that if a non-permeable membrane separates a solution with high [KCl] from a solution with low [KCl], the net charge
1. Ionic Movements Across a Permeable Membrane: The Nernst Potential. In class we showed that if a non-permeable membrane separates a solution with high [KCl] from a solution with low [KCl], the net charge
Lojayn Salah. Zaid R Al Najdawi. Mohammad-Khatatbeh
 7 Lojayn Salah Zaid R Al Najdawi Mohammad-Khatatbeh Salam everyone, I made my best to make this sheet clear enough to be easily understood let the party begin :P Quick Revision about the previous lectures:
7 Lojayn Salah Zaid R Al Najdawi Mohammad-Khatatbeh Salam everyone, I made my best to make this sheet clear enough to be easily understood let the party begin :P Quick Revision about the previous lectures:
Introduction and the Hodgkin-Huxley Model
 1 Introduction and the Hodgkin-Huxley Model Richard Bertram Department of Mathematics and Programs in Neuroscience and Molecular Biophysics Florida State University Tallahassee, Florida 32306 Reference:
1 Introduction and the Hodgkin-Huxley Model Richard Bertram Department of Mathematics and Programs in Neuroscience and Molecular Biophysics Florida State University Tallahassee, Florida 32306 Reference:
Neural Modeling and Computational Neuroscience. Claudio Gallicchio
 Neural Modeling and Computational Neuroscience Claudio Gallicchio 1 Neuroscience modeling 2 Introduction to basic aspects of brain computation Introduction to neurophysiology Neural modeling: Elements
Neural Modeling and Computational Neuroscience Claudio Gallicchio 1 Neuroscience modeling 2 Introduction to basic aspects of brain computation Introduction to neurophysiology Neural modeling: Elements
Basic elements of neuroelectronics -- membranes -- ion channels -- wiring. Elementary neuron models -- conductance based -- modelers alternatives
 Computing in carbon Basic elements of neuroelectronics -- membranes -- ion channels -- wiring Elementary neuron models -- conductance based -- modelers alternatives Wiring neurons together -- synapses
Computing in carbon Basic elements of neuroelectronics -- membranes -- ion channels -- wiring Elementary neuron models -- conductance based -- modelers alternatives Wiring neurons together -- synapses
Integration of synaptic inputs in dendritic trees
 Integration of synaptic inputs in dendritic trees Theoretical Neuroscience Fabrizio Gabbiani Division of Neuroscience Baylor College of Medicine One Baylor Plaza Houston, TX 77030 e-mail:gabbiani@bcm.tmc.edu
Integration of synaptic inputs in dendritic trees Theoretical Neuroscience Fabrizio Gabbiani Division of Neuroscience Baylor College of Medicine One Baylor Plaza Houston, TX 77030 e-mail:gabbiani@bcm.tmc.edu
Cells have an unequal distribution of charge across their membrane: more postiive charges on the outside; more negative charges on the inside.
 Resting Membrane potential (V m ) or RMP Many cells have a membrane potential (Vm) that can be measured from an electrode in the cell with a voltmeter. neurons, muscle cells, heart cells, endocrine cells...
Resting Membrane potential (V m ) or RMP Many cells have a membrane potential (Vm) that can be measured from an electrode in the cell with a voltmeter. neurons, muscle cells, heart cells, endocrine cells...
MEMBRANE POTENTIALS AND ACTION POTENTIALS:
 University of Jordan Faculty of Medicine Department of Physiology & Biochemistry Medical students, 2017/2018 +++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ Review: Membrane physiology
University of Jordan Faculty of Medicine Department of Physiology & Biochemistry Medical students, 2017/2018 +++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ Review: Membrane physiology
COGNITIVE SCIENCE 107A
 COGNITIVE SCIENCE 107A Electrophysiology: Electrotonic Properties 2 Jaime A. Pineda, Ph.D. The Model Neuron Lab Your PC/CSB115 http://cogsci.ucsd.edu/~pineda/cogs107a/index.html Labs - Electrophysiology
COGNITIVE SCIENCE 107A Electrophysiology: Electrotonic Properties 2 Jaime A. Pineda, Ph.D. The Model Neuron Lab Your PC/CSB115 http://cogsci.ucsd.edu/~pineda/cogs107a/index.html Labs - Electrophysiology
Quantitative Electrophysiology
 ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Tuesday, September 18, 2012 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Tuesday, September 18, 2012 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in 1780s
Overview Organization: Central Nervous System (CNS) Peripheral Nervous System (PNS) innervate Divisions: a. Afferent
 Peripheral Nervous System (PNS) innervate Divisions: a. Afferent") Overview Organization: Central Nervous System (CNS) Brain and spinal cord receives and processes information. Peripheral Nervous System (PNS) Nerve cells that link CNS with organs throughout the body.
Overview Organization: Central Nervous System (CNS) Brain and spinal cord receives and processes information. Peripheral Nervous System (PNS) Nerve cells that link CNS with organs throughout the body.
Nervous Systems: Neuron Structure and Function
 Nervous Systems: Neuron Structure and Function Integration An animal needs to function like a coherent organism, not like a loose collection of cells. Integration = refers to processes such as summation
Nervous Systems: Neuron Structure and Function Integration An animal needs to function like a coherent organism, not like a loose collection of cells. Integration = refers to processes such as summation
Resting membrane potential,
 Resting membrane potential Inside of each cell is negative as compared with outer surface: negative resting membrane potential (between -30 and -90 mv) Examination with microelectrode (Filled with KCl
Resting membrane potential Inside of each cell is negative as compared with outer surface: negative resting membrane potential (between -30 and -90 mv) Examination with microelectrode (Filled with KCl
Quantitative Electrophysiology
 ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Wednesday, September 13, 2006 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in
ECE 795: Quantitative Electrophysiology Notes for Lecture #1 Wednesday, September 13, 2006 1. INTRODUCTION TO EXCITABLE CELLS Historical perspective: Bioelectricity first discovered by Luigi Galvani in
Neurons. The Molecular Basis of their Electrical Excitability
 Neurons The Molecular Basis of their Electrical Excitability Viva La Complexity! Consider, The human brain contains >10 11 neurons! Each neuron makes 10 3 (average) synaptic contacts on up to 10 3 other
Neurons The Molecular Basis of their Electrical Excitability Viva La Complexity! Consider, The human brain contains >10 11 neurons! Each neuron makes 10 3 (average) synaptic contacts on up to 10 3 other
Neurons, Synapses, and Signaling
 Chapter 48 Neurons, Synapses, and Signaling PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Chapter 48 Neurons, Synapses, and Signaling PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
NEURONS, SENSE ORGANS, AND NERVOUS SYSTEMS CHAPTER 34
 NEURONS, SENSE ORGANS, AND NERVOUS SYSTEMS CHAPTER 34 KEY CONCEPTS 34.1 Nervous Systems Are Composed of Neurons and Glial Cells 34.2 Neurons Generate Electric Signals by Controlling Ion Distributions 34.3
NEURONS, SENSE ORGANS, AND NERVOUS SYSTEMS CHAPTER 34 KEY CONCEPTS 34.1 Nervous Systems Are Composed of Neurons and Glial Cells 34.2 Neurons Generate Electric Signals by Controlling Ion Distributions 34.3
3 Action Potentials - Brutal Approximations
 Physics 172/278 - David Kleinfeld - Fall 2004; Revised Winter 2015 3 Action Potentials - Brutal Approximations The Hodgkin-Huxley equations for the behavior of the action potential in squid, and similar
Physics 172/278 - David Kleinfeld - Fall 2004; Revised Winter 2015 3 Action Potentials - Brutal Approximations The Hodgkin-Huxley equations for the behavior of the action potential in squid, and similar
Balance of Electric and Diffusion Forces
 Balance of Electric and Diffusion Forces Ions flow into and out of the neuron under the forces of electricity and concentration gradients (diffusion). The net result is a electric potential difference
Balance of Electric and Diffusion Forces Ions flow into and out of the neuron under the forces of electricity and concentration gradients (diffusion). The net result is a electric potential difference
9 Generation of Action Potential Hodgkin-Huxley Model
 9 Generation of Action Potential Hodgkin-Huxley Model (based on chapter 2, W.W. Lytton, Hodgkin-Huxley Model) 9. Passive and active membrane models In the previous lecture we have considered a passive
9 Generation of Action Potential Hodgkin-Huxley Model (based on chapter 2, W.W. Lytton, Hodgkin-Huxley Model) 9. Passive and active membrane models In the previous lecture we have considered a passive
لجنة الطب البشري رؤية تنير دروب تميزكم
 1) Hyperpolarization phase of the action potential: a. is due to the opening of voltage-gated Cl channels. b. is due to prolonged opening of voltage-gated K + channels. c. is due to closure of the Na +
1) Hyperpolarization phase of the action potential: a. is due to the opening of voltage-gated Cl channels. b. is due to prolonged opening of voltage-gated K + channels. c. is due to closure of the Na +
SECOND PUBLIC EXAMINATION. Honour School of Physics Part C: 4 Year Course. Honour School of Physics and Philosophy Part C C7: BIOLOGICAL PHYSICS
 2757 SECOND PUBLIC EXAMINATION Honour School of Physics Part C: 4 Year Course Honour School of Physics and Philosophy Part C C7: BIOLOGICAL PHYSICS TRINITY TERM 2013 Monday, 17 June, 2.30 pm 5.45 pm 15
2757 SECOND PUBLIC EXAMINATION Honour School of Physics Part C: 4 Year Course Honour School of Physics and Philosophy Part C C7: BIOLOGICAL PHYSICS TRINITY TERM 2013 Monday, 17 June, 2.30 pm 5.45 pm 15
STUDENT PAPER. Santiago Santana University of Illinois, Urbana-Champaign Blue Waters Education Program 736 S. Lombard Oak Park IL, 60304
 STUDENT PAPER Differences between Stochastic and Deterministic Modeling in Real World Systems using the Action Potential of Nerves. Santiago Santana University of Illinois, Urbana-Champaign Blue Waters
STUDENT PAPER Differences between Stochastic and Deterministic Modeling in Real World Systems using the Action Potential of Nerves. Santiago Santana University of Illinois, Urbana-Champaign Blue Waters
Lecture 04, 04 Sept 2003 Chapters 4 and 5. Vertebrate Physiology ECOL 437 University of Arizona Fall instr: Kevin Bonine t.a.
 Lecture 04, 04 Sept 2003 Chapters 4 and 5 Vertebrate Physiology ECOL 437 University of Arizona Fall 2003 instr: Kevin Bonine t.a.: Bret Pasch Vertebrate Physiology 437 1. Membranes (CH4) 2. Nervous System
Lecture 04, 04 Sept 2003 Chapters 4 and 5 Vertebrate Physiology ECOL 437 University of Arizona Fall 2003 instr: Kevin Bonine t.a.: Bret Pasch Vertebrate Physiology 437 1. Membranes (CH4) 2. Nervous System
Neurophysiology. Danil Hammoudi.MD
 Neurophysiology Danil Hammoudi.MD ACTION POTENTIAL An action potential is a wave of electrical discharge that travels along the membrane of a cell. Action potentials are an essential feature of animal
Neurophysiology Danil Hammoudi.MD ACTION POTENTIAL An action potential is a wave of electrical discharge that travels along the membrane of a cell. Action potentials are an essential feature of animal
Nervous Lecture Test Questions Set 2
 Nervous Lecture Test Questions Set 2 1. The role of chloride in a resting membrane potential: a. creates resting potential b. indirectly causes repolarization c. stabilization of sodium d. it has none,
Nervous Lecture Test Questions Set 2 1. The role of chloride in a resting membrane potential: a. creates resting potential b. indirectly causes repolarization c. stabilization of sodium d. it has none,
Neuroscience: Exploring the Brain
 Slide 1 Neuroscience: Exploring the Brain Chapter 3: The Neuronal Membrane at Rest Slide 2 Introduction Action potential in the nervous system Action potential vs. resting potential Slide 3 Not at rest
Slide 1 Neuroscience: Exploring the Brain Chapter 3: The Neuronal Membrane at Rest Slide 2 Introduction Action potential in the nervous system Action potential vs. resting potential Slide 3 Not at rest
Chapter 48 Neurons, Synapses, and Signaling
 Chapter 48 Neurons, Synapses, and Signaling Concept 48.1 Neuron organization and structure reflect function in information transfer Neurons are nerve cells that transfer information within the body Neurons
Chapter 48 Neurons, Synapses, and Signaling Concept 48.1 Neuron organization and structure reflect function in information transfer Neurons are nerve cells that transfer information within the body Neurons
Neuroscience 201A Exam Key, October 7, 2014
 Neuroscience 201A Exam Key, October 7, 2014 Question #1 7.5 pts Consider a spherical neuron with a diameter of 20 µm and a resting potential of -70 mv. If the net negativity on the inside of the cell (all
Neuroscience 201A Exam Key, October 7, 2014 Question #1 7.5 pts Consider a spherical neuron with a diameter of 20 µm and a resting potential of -70 mv. If the net negativity on the inside of the cell (all
Chapter 2 Cellular Homeostasis and Membrane Potential
 Chapter 2 Cellular Homeostasis and Membrane Potential 2.1 Membrane Structure and Composition The human cell can be considered to consist of a bag of fluid with a wall that separates the internal, or intracellular,
Chapter 2 Cellular Homeostasis and Membrane Potential 2.1 Membrane Structure and Composition The human cell can be considered to consist of a bag of fluid with a wall that separates the internal, or intracellular,
3.3 Simulating action potentials
 6 THE HODGKIN HUXLEY MODEL OF THE ACTION POTENTIAL Fig. 3.1 Voltage dependence of rate coefficients and limiting values and time constants for the Hodgkin Huxley gating variables. (a) Graphs of forward
6 THE HODGKIN HUXLEY MODEL OF THE ACTION POTENTIAL Fig. 3.1 Voltage dependence of rate coefficients and limiting values and time constants for the Hodgkin Huxley gating variables. (a) Graphs of forward
The Nervous System and the Sodium-Potassium Pump
 The Nervous System and the Sodium-Potassium Pump 1. Define the following terms: Ion: A Student Activity on Membrane Potentials Cation: Anion: Concentration gradient: Simple diffusion: Sodium-Potassium
The Nervous System and the Sodium-Potassium Pump 1. Define the following terms: Ion: A Student Activity on Membrane Potentials Cation: Anion: Concentration gradient: Simple diffusion: Sodium-Potassium
Introduction to Neural Networks U. Minn. Psy 5038 Spring, 1999 Daniel Kersten. Lecture 2a. The Neuron - overview of structure. From Anderson (1995)
") Introduction to Neural Networks U. Minn. Psy 5038 Spring, 1999 Daniel Kersten Lecture 2a The Neuron - overview of structure From Anderson (1995) 2 Lect_2a_Mathematica.nb Basic Structure Information flow:
Introduction to Neural Networks U. Minn. Psy 5038 Spring, 1999 Daniel Kersten Lecture 2a The Neuron - overview of structure From Anderson (1995) 2 Lect_2a_Mathematica.nb Basic Structure Information flow:
From neuronal oscillations to complexity
 1/39 The Fourth International Workshop on Advanced Computation for Engineering Applications (ACEA 2008) MACIS 2 Al-Balqa Applied University, Salt, Jordan Corson Nathalie, Aziz Alaoui M.A. University of
1/39 The Fourth International Workshop on Advanced Computation for Engineering Applications (ACEA 2008) MACIS 2 Al-Balqa Applied University, Salt, Jordan Corson Nathalie, Aziz Alaoui M.A. University of
Chapter 1 subtitles Ion gradients
 CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 1 subtitles Ion gradients Introduction In this first chapter, I'll explain the basic knowledge required to understand the electrical signals generated
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 1 subtitles Ion gradients Introduction In this first chapter, I'll explain the basic knowledge required to understand the electrical signals generated
General Physics. Nerve Conduction. Newton s laws of Motion Work, Energy and Power. Fluids. Direct Current (DC)
") Newton s laws of Motion Work, Energy and Power Fluids Direct Current (DC) Nerve Conduction Wave properties of light Ionizing Radiation General Physics Prepared by: Sujood Alazzam 2017/2018 CHAPTER OUTLINE
Newton s laws of Motion Work, Energy and Power Fluids Direct Current (DC) Nerve Conduction Wave properties of light Ionizing Radiation General Physics Prepared by: Sujood Alazzam 2017/2018 CHAPTER OUTLINE
Nervous Tissue. Neurons Electrochemical Gradient Propagation & Transduction Neurotransmitters Temporal & Spatial Summation
 Nervous Tissue Neurons Electrochemical Gradient Propagation & Transduction Neurotransmitters Temporal & Spatial Summation What is the function of nervous tissue? Maintain homeostasis & respond to stimuli
Nervous Tissue Neurons Electrochemical Gradient Propagation & Transduction Neurotransmitters Temporal & Spatial Summation What is the function of nervous tissue? Maintain homeostasis & respond to stimuli
Universality of sensory-response systems
 excite.org(anism): Electrical Signaling Universality of sensory-response systems Three step process: sensation-integration-response Bacterial chemotaxis Madigan et al. Fig. 8.24 Rick Stewart (CBMG) Human
excite.org(anism): Electrical Signaling Universality of sensory-response systems Three step process: sensation-integration-response Bacterial chemotaxis Madigan et al. Fig. 8.24 Rick Stewart (CBMG) Human
Electrical Properties of the Membrane
 BIOE 2520 Electrical Properties of the Membrane Reading: Chapter 11 of Alberts et al. Stephen Smith, Ph.D. 433 Biotech Center shs46@pitt.edu Permeability of Lipid membrane Lipid bilayer is virtually impermeable
BIOE 2520 Electrical Properties of the Membrane Reading: Chapter 11 of Alberts et al. Stephen Smith, Ph.D. 433 Biotech Center shs46@pitt.edu Permeability of Lipid membrane Lipid bilayer is virtually impermeable
Nernst Equilibrium Potential. p. 1
 Nernst Equilibrium Potential p. 1 Diffusion The conservation law for a compound with concentration c: rate change of c = local production + accumulation due to transport. Model: d c dv = p dv J n da dt
Nernst Equilibrium Potential p. 1 Diffusion The conservation law for a compound with concentration c: rate change of c = local production + accumulation due to transport. Model: d c dv = p dv J n da dt
Lecture 11 : Simple Neuron Models. Dr Eileen Nugent
 Lecture 11 : Simple Neuron Models Dr Eileen Nugent Reading List Nelson, Biological Physics, Chapter 12 Phillips, PBoC, Chapter 17 Gerstner, Neuronal Dynamics: from single neurons to networks and models
Lecture 11 : Simple Neuron Models Dr Eileen Nugent Reading List Nelson, Biological Physics, Chapter 12 Phillips, PBoC, Chapter 17 Gerstner, Neuronal Dynamics: from single neurons to networks and models
Nervous Tissue. Neurons Neural communication Nervous Systems
 Nervous Tissue Neurons Neural communication Nervous Systems What is the function of nervous tissue? Maintain homeostasis & respond to stimuli Sense & transmit information rapidly, to specific cells and
Nervous Tissue Neurons Neural communication Nervous Systems What is the function of nervous tissue? Maintain homeostasis & respond to stimuli Sense & transmit information rapidly, to specific cells and
Biophysical Foundations
 Biophysical Foundations BENG/BGGN 260 Neurodynamics University of California, San Diego Week 1 BENG/BGGN 260 Neurodynamics (UCSD Biophysical Foundations Week 1 1 / 15 Reading Material B. Hille, Ion Channels
Biophysical Foundations BENG/BGGN 260 Neurodynamics University of California, San Diego Week 1 BENG/BGGN 260 Neurodynamics (UCSD Biophysical Foundations Week 1 1 / 15 Reading Material B. Hille, Ion Channels
Side View with Rings of Charge
 1 Ion Channel Biophysics Describe the main biophysical characteristics of at least one type of ionic channel. How does its biophysical properties contribute to its physiological function. What is thought
1 Ion Channel Biophysics Describe the main biophysical characteristics of at least one type of ionic channel. How does its biophysical properties contribute to its physiological function. What is thought
Bo Deng University of Nebraska-Lincoln UNL Math Biology Seminar
 Mathematical Model of Neuron Bo Deng University of Nebraska-Lincoln UNL Math Biology Seminar 09-10-2015 Review -- One Basic Circuit By Kirchhoff's Current Law 0 = I C + I R + I L I ext By Kirchhoff s Voltage
Mathematical Model of Neuron Bo Deng University of Nebraska-Lincoln UNL Math Biology Seminar 09-10-2015 Review -- One Basic Circuit By Kirchhoff's Current Law 0 = I C + I R + I L I ext By Kirchhoff s Voltage
Decoding. How well can we learn what the stimulus is by looking at the neural responses?
 Decoding How well can we learn what the stimulus is by looking at the neural responses? Two approaches: devise explicit algorithms for extracting a stimulus estimate directly quantify the relationship
Decoding How well can we learn what the stimulus is by looking at the neural responses? Two approaches: devise explicit algorithms for extracting a stimulus estimate directly quantify the relationship
Chapter 9. Nerve Signals and Homeostasis
 Chapter 9 Nerve Signals and Homeostasis A neuron is a specialized nerve cell that is the functional unit of the nervous system. Neural signaling communication by neurons is the process by which an animal
Chapter 9 Nerve Signals and Homeostasis A neuron is a specialized nerve cell that is the functional unit of the nervous system. Neural signaling communication by neurons is the process by which an animal
Single neuron models. L. Pezard Aix-Marseille University
 Single neuron models L. Pezard Aix-Marseille University Biophysics Biological neuron Biophysics Ionic currents Passive properties Active properties Typology of models Compartmental models Differential
Single neuron models L. Pezard Aix-Marseille University Biophysics Biological neuron Biophysics Ionic currents Passive properties Active properties Typology of models Compartmental models Differential
Synaptic dynamics. John D. Murray. Synaptic currents. Simple model of the synaptic gating variable. First-order kinetics
 Synaptic dynamics John D. Murray A dynamical model for synaptic gating variables is presented. We use this to study the saturation of synaptic gating at high firing rate. Shunting inhibition and the voltage
Synaptic dynamics John D. Murray A dynamical model for synaptic gating variables is presented. We use this to study the saturation of synaptic gating at high firing rate. Shunting inhibition and the voltage
ACTION POTENTIAL. Dr. Ayisha Qureshi Professor MBBS, MPhil
 ACTION POTENTIAL Dr. Ayisha Qureshi Professor MBBS, MPhil DEFINITIONS: Stimulus: A stimulus is an external force or event which when applied to an excitable tissue produces a characteristic response. Subthreshold
ACTION POTENTIAL Dr. Ayisha Qureshi Professor MBBS, MPhil DEFINITIONS: Stimulus: A stimulus is an external force or event which when applied to an excitable tissue produces a characteristic response. Subthreshold
The Neuron - F. Fig. 45.3
 excite.org(anism): Electrical Signaling The Neuron - F. Fig. 45.3 Today s lecture we ll use clickers Review today 11:30-1:00 in 2242 HJ Patterson Electrical signals Dendrites: graded post-synaptic potentials
excite.org(anism): Electrical Signaling The Neuron - F. Fig. 45.3 Today s lecture we ll use clickers Review today 11:30-1:00 in 2242 HJ Patterson Electrical signals Dendrites: graded post-synaptic potentials
Dendrites - receives information from other neuron cells - input receivers.
 The Nerve Tissue Neuron - the nerve cell Dendrites - receives information from other neuron cells - input receivers. Cell body - includes usual parts of the organelles of a cell (nucleus, mitochondria)
The Nerve Tissue Neuron - the nerve cell Dendrites - receives information from other neuron cells - input receivers. Cell body - includes usual parts of the organelles of a cell (nucleus, mitochondria)
Physiology Unit 2. MEMBRANE POTENTIALS and SYNAPSES
 Physiology Unit 2 MEMBRANE POTENTIALS and SYNAPSES Neuron Communication Neurons are stimulated by receptors on dendrites and cell bodies (soma) Ligand gated ion channels GPCR s Neurons stimulate cells
Physiology Unit 2 MEMBRANE POTENTIALS and SYNAPSES Neuron Communication Neurons are stimulated by receptors on dendrites and cell bodies (soma) Ligand gated ion channels GPCR s Neurons stimulate cells
Problem Set No. 4 Due: Monday, 11/18/10 at the start of class
 Department of Chemical Engineering ChE 170 University of California, Santa Barbara Fall 2010 Problem Set No. 4 Due: Monday, 11/18/10 at the start of class Objective: To understand the thermodynamic and
Department of Chemical Engineering ChE 170 University of California, Santa Barbara Fall 2010 Problem Set No. 4 Due: Monday, 11/18/10 at the start of class Objective: To understand the thermodynamic and
Voltage-clamp and Hodgkin-Huxley models
 Voltage-clamp and Hodgkin-Huxley models Read: Hille, Chapters 2-5 (best) Koch, Chapters 6, 8, 9 See also Clay, J. Neurophysiol. 80:903-913 (1998) (for a recent version of the HH squid axon model) Rothman
Voltage-clamp and Hodgkin-Huxley models Read: Hille, Chapters 2-5 (best) Koch, Chapters 6, 8, 9 See also Clay, J. Neurophysiol. 80:903-913 (1998) (for a recent version of the HH squid axon model) Rothman
Equivalent Circuit Model of the Neuron
 Generator Potentials, Synaptic Potentials and Action Potentials All Can Be Described by the Equivalent Circuit Model of the Membrane Equivalent Circuit Model of the Neuron PNS, Fig 211 The Nerve (or Muscle)
Generator Potentials, Synaptic Potentials and Action Potentials All Can Be Described by the Equivalent Circuit Model of the Membrane Equivalent Circuit Model of the Neuron PNS, Fig 211 The Nerve (or Muscle)
NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic
 NEUROPHYSIOLOGY NOTES L1 WHAT IS NEUROPHYSIOLOGY? NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic receptor
NEUROPHYSIOLOGY NOTES L1 WHAT IS NEUROPHYSIOLOGY? NEURONS Excitable cells Therefore, have a RMP Synapse = chemical communication site between neurons, from pre-synaptic release to postsynaptic receptor
Kirchhoff s Rules. Survey available this week. $ closed loop. Quiz on a simple DC circuit. Quiz on a simple DC circuit
 RC Circuits. Start Magnetic Fields Announcement on MTE 1 This Lecture: RC circuits Membrane electrical currents Magnetic Fields and Magnets Wednesday Oct. 4, slightly later start time:5:45 pm - 7:15 pm
RC Circuits. Start Magnetic Fields Announcement on MTE 1 This Lecture: RC circuits Membrane electrical currents Magnetic Fields and Magnets Wednesday Oct. 4, slightly later start time:5:45 pm - 7:15 pm
6.3.4 Action potential
 I ion C m C m dφ dt Figure 6.8: Electrical circuit model of the cell membrane. Normally, cells are net negative inside the cell which results in a non-zero resting membrane potential. The membrane potential
I ion C m C m dφ dt Figure 6.8: Electrical circuit model of the cell membrane. Normally, cells are net negative inside the cell which results in a non-zero resting membrane potential. The membrane potential
BIOLOGY 11/10/2016. Neurons, Synapses, and Signaling. Concept 48.1: Neuron organization and structure reflect function in information transfer
 48 Neurons, Synapses, and Signaling CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick Concept 48.1: Neuron organization
48 Neurons, Synapses, and Signaling CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick Concept 48.1: Neuron organization
Introduction to cardiac electrophysiology 1. Dr. Tóth András 2018
 Introduction to cardiac electrophysiology 1. Dr. Tóth ndrás 2018 Topics Transmembran transport Donnan equilibrium Resting potential 1 Transmembran transport Major types of transmembran transport J: net
Introduction to cardiac electrophysiology 1. Dr. Tóth ndrás 2018 Topics Transmembran transport Donnan equilibrium Resting potential 1 Transmembran transport Major types of transmembran transport J: net
Propagation& Integration: Passive electrical properties
 Fundamentals of Neuroscience (NSCS 730, Spring 2010) Instructor: Art Riegel; email: Riegel@musc.edu; Room EL 113; time: 9 11 am Office: 416C BSB (792.5444) Propagation& Integration: Passive electrical
Fundamentals of Neuroscience (NSCS 730, Spring 2010) Instructor: Art Riegel; email: Riegel@musc.edu; Room EL 113; time: 9 11 am Office: 416C BSB (792.5444) Propagation& Integration: Passive electrical
Vertebrate Physiology 437 EXAM I NAME, Section (circle): am pm 23 September Exam is worth 100 points. You have 75 minutes.
: am pm 23 September Exam is worth 100 points. You have 75 minutes.") 1 Vertebrate Physiology 437 EXAM I NAME, Section (circle): am pm 23 September 2004. Exam is worth 100 points. You have 75 minutes. True or False (write true or false ; 10 points total; 1 point each) 1.
1 Vertebrate Physiology 437 EXAM I NAME, Section (circle): am pm 23 September 2004. Exam is worth 100 points. You have 75 minutes. True or False (write true or false ; 10 points total; 1 point each) 1.
All-or-None Principle and Weakness of Hodgkin-Huxley Mathematical Model
 All-or-None Principle and Weakness of Hodgkin-Huxley Mathematical Model S. A. Sadegh Zadeh, C. Kambhampati International Science Index, Mathematical and Computational Sciences waset.org/publication/10008281
All-or-None Principle and Weakness of Hodgkin-Huxley Mathematical Model S. A. Sadegh Zadeh, C. Kambhampati International Science Index, Mathematical and Computational Sciences waset.org/publication/10008281
NE 204 mini-syllabus (weeks 4 8)
") NE 24 mini-syllabus (weeks 4 8) Instructor: John Burke O ce: MCS 238 e-mail: jb@math.bu.edu o ce hours: by appointment Overview: For the next few weeks, we will focus on mathematical models of single neurons.
NE 24 mini-syllabus (weeks 4 8) Instructor: John Burke O ce: MCS 238 e-mail: jb@math.bu.edu o ce hours: by appointment Overview: For the next few weeks, we will focus on mathematical models of single neurons.