Natural processes and scientific reasoning (sequentialization as the result of concurrent computation) René Vestergaard
|
|
|
- Katherine Merritt
- 5 years ago
- Views:
Transcription
1 (sequentialization as the result of concurrent computation)
2 Pivots: 1) background, 2) focus, 3) example 1 a reasoning style is (often) a model of computation 2 a model of computation is mathematics can be wrangled 3 a particular computation has problem-specific import

3 Raw data : standard molecular-biology reference Get A Genetic Switch, 3rd. Read it and read it again. Review on amazon.com by Ardent Reader
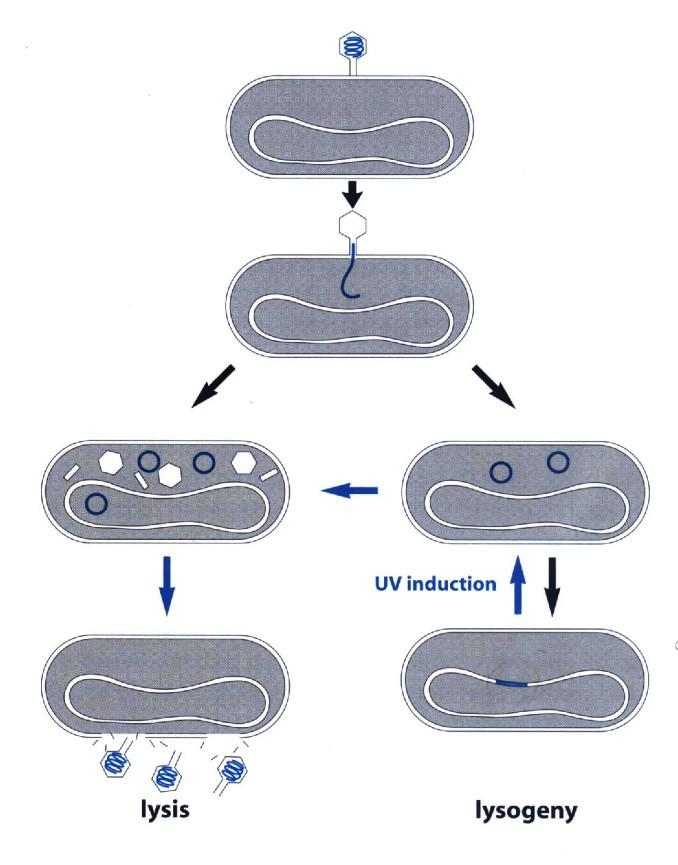




4 Informally: molecular coding of physiology TRICKY: getting it exactly right! RNA polymerase RNA polymerase repressor
 (A) (B) (E) (F) (D)")
5 Formally: start and end point (but middle is crux) (A) (B) (E) (F) (D) (A) (C)
6 Formally: intermediate coextension logic over EPPCs Key step in augmented (inverted) hierarchy of reductionist reasoning style Definition (may happen in compartment, under genotype g) R i, M i, P i, I i. g A ε B (interference) if M i. R i i Ii P i EPPC(g) i. R i M i A i. I i A = CoinhFree({R i M i Ii P i } i ) g A τ 1 B g B τ 2 C g A τ 1Bτ 2 C (sequence) B = (A \ i R i) i P i well-formed(b)


 PHENOTYPE pathway arguments")
7 CEqEA: Ptashne-style reasoning as computation demonstration + GENOTYPE PHYSIOLOGY SYNCHROTYPE form-to-function sequentialization compilation MIG/RI specification cascaded causation diagrams synchronization diagrams simulations perturbation CCP visualization (aggregation) PHENOTYPE pathway arguments pathway properties cell behaviours
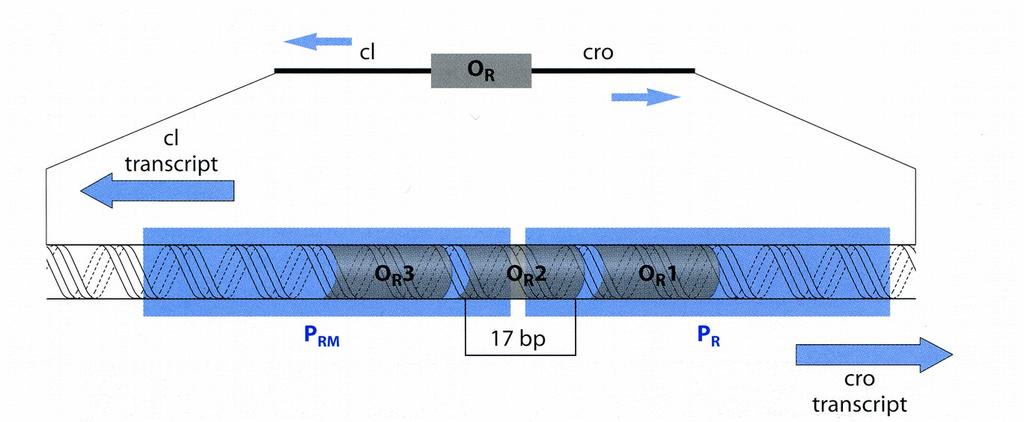
8 Coextension reasoning example Pathway argument [informal inference] Two changes result [from SOS/RecA*]. First, as repressor vacates O R {1,2} the rate of repressor synthesis drops (because repressor is required to turn on transcription of its own gene); and second, polymerase binds to P R to begin transcription of cro. [24:-7] Activity trace [formal inference, with all pertinent details] 1 [CI.l, Cro.0, RecA.*] + [DNA.ss] 2 -> [SOS] 3 x>![] -- [DNA.ss] ; [PR_tr] -- [CI.l!@[OLR12, OLR123]] ; [OLR12_CI, PRM_tr] -- [SOS] 5 [Cro.0, RecA.*] + [CI.0, DNA.ss] 6 -> [PR_tr] ; [SOS] 7 x>![] -- [DNA.ss] 9 [Cro.l, RecA.*] + [CI.0, DNA.ss] 10 -> [PR_tr] ; [SOS] 11 x>![] -- [DNA.ss]
9 λ [AGS3] ab-intra phenotype: [AGS3] pathway properties pathway argument < pathway property < cell behaviour Key: stands for absent a host SOS response and stands for its negation during a host SOS response. stands for with CII below high concentration and with CI below physiological concentration and stands for its negation with CII at high concentration or with CI at physiological concentration ; italics not in [AGS3]. 0/ lysogeny s means, when viable ( and ), take precedence over a lytic attack s 1/ CII determines λ s pathway at fresh infection : lysogeny at high vs lytic attack below 2/ the lysogenic cycle is homeostatic, i.e., is sustainable and absorbing perturbations a/ protein concentrations are CI controlled in the lysogenic cycle α/ an initial CI concentration is and must be established by high CII β/ once established, CI remains physiological and Cro non-physiological b/ a switch to the lytic attack is not effected by i/ natural or operative means this is the basis of sustainability ii/ Cro-perturbation this is the key means of homeostasis iii/ super-infection this is λ-immunity c/ a switch to the lytic attack is effected by UV-irradiation (via ) 3/ the lytic attack expires the host and, else, is sustainable but not homeostatic a/ protein concentrations are programmatic during a lytic attack α/ the lytic attack is constitutive or β/ once initiated, Cro remains physiological and CI inoperative or γ/ a lytic attack expires the host after Cro has reached high or b/ ignoring host expiration, a switch to the lysogenic cycle is not effected by i/ natural or operative means or this is the basis of sustainability ii/ CI-perturbation, although the lytic attack may be set back iii/ super-infection or not quite anti-immunity, see Chp.S5 c/ ignoring host expiration, a switch to lysogeny may be effected by ii/ CI-perturbation (and in order for the lytic attack to be viable, see 3/d/) d/ a lytic attack is unlikely and
10 Retrodiction, based also on non-observable properties Retrodiction (I) Retrodiction (II) Retrodiction (III) Retrodiction (IV) Fig.6 0/ is not considered in [AGS3] and is, in fact, contradicting superseded statements from earlier editions and printings, e.g., Cro [being transcribed from P R ] determines the course of events [24:-5] and the first Cro to be synthesized binds to O R 3 [which] prevents [RNAP] from binding to P RM and abolishes further synthesis of repressor. At this point the switch has been thrown and lytic growth ensues. [25:2a,2b,3a,3b]. Pathway property 0/ is consistent with all parts of [AGS3] that are not explicitly stated or implied to be superseded by the recent findings that prompted the 3rd edition of the monograph, see [t]his new edition is prompted by discoveries [... ] that add to, rather than reformulate, the earlier story [xi:11] and the conclusion that Cro must bind O R 3 to trigger the transition to lytic growth, although not excluded, remains uncertain [121:-4]. In particular, see If repressor were added to a phage beginning its lytic cycle, growth would be inhibited [62:-1]. More, 1/, 2/a/α/, 2/b/ii/, 3/c/ii/, and 3/d/ would not hold in their present form if 0/ did not hold, and, as we show in Sect.S5.4/Cro, failure of 0/ results in anti-immunity. [... ] Our λ [AGS3] is a concrete model of λ where Cro binding to O R 3 is not needed to trigger the switch, see [121:-4]. A main implication of the structure of the pathway properties we establish, see Fig.6, is that lysogenic cycles and lytic attacks (absent host expiration) exhibit fundamentally different stability properties, which appears to be a novel inference (of a probably-known observation, see [92:Fig.4.23:1]). [... ] A further implication of the pathway properties we establish, see Fig.6, is that the lysogenic-to-lytic switch is effected by activated-reca-mediated CI-proteolysis at the onset of the switch and only later affected also by Cro inhibition of ci transcription, see 2/c/, 3/c/ii/, and the varying of Cro s intrinsic affinities in Chp.S5. [... ] Fig.6 and Chp.S3 do not firmly establish at what point a lysogenic-to-lytic switch can be said to have been initiated, see [121:-4]. The clear suggestion is that it is at the physiologically-zero concentration of CI at which constitutive cro-transcription becomes physiological, see 0/, 2/a/α/ vs 3/a/α/, and Retrodiction (III). A consequence of this, see 2/a/ vs 3/a/β/, is the known property that there is no turning back once the lytic attack is under way, at least in the absence of physiological CII and of stochastic CI that is sufficiently strong to re-establish RNAP-recruitment at P RM, see 3/c/ii/ and 3/d/.
11 Meta-theory guarantee 1/2: the methods are logical Definition R M I P/A { A ε (A \ R) P if R M A I A =... A ε A o/w Proposition (Unit) g, p, A. p EPPC(g)... g p/a Proposition (Recompose) g 1, g 2, p, A. ( p x, X. p x EPPC(g 1 )... g 2 p x /X) g 1 p/a... g 2 p/a
12 Meta-theory guarantee 1/2: the methods are logical Definition R M I P/A { A ε (A \ R) P if R M A I A =... A ε A o/w Proposition (Unit) g, p, A. p EPPC(g)... g p/a Proposition (Recompose) g 1, g 2, p, A. ( p x, X. p x EPPC(g 1 )... g 2 p x /X) g 1 p/a... g 2 p/a Theorem ((cut) is admissible) g p 0 /A p 0 ++g p/a g p/a (cut) if...
13 Meta-theory guarantee 2/2: the methods do something Proposition (Non-empty) g, A, B, τ. g A τ B Theorem (Non-trivial/consistent) A, B, τ. A τ B
14 Sequentialization, trace-monoid style Definition (Synchrotype [idealized]) By synchrotype, we mean an account of how a collection of physiology-change processes may interleave with each other under coextension where we do not distinguish different execution orders of processes that are independent of each other and environmental factors are retained as conditionals. Technically CCP traversals of cascaded causation diagram form a partially-commutative monoid, with composition. Synchrotype is (essentially) the trace monoid = independence factored out: leaves graph where edges are changes breaking synchronicity.
15 Independence 1/2: interpolation of observable values Axiom [informal principle] As [Cro] binds it turns off [P R ], but as the cells grow and divide, the concentration of [Cro] drops, and [P R ] turns on again. A steady state is reached at which the rate of synthesis of [Cro] just balances its rate of dilution and, presumably, a constant concentration of [Cro] is maintained. In this situation [Cro] diminishes, but does not abolish, its own synthesis. [92:-3] CII._ CII.h CI.0,CII.h ;CII.h,.RNAP.tr; -CI.l!,-CI.m!,-CI.h!;.RNAP.tr;CII.h CI.l,CII.h DNA.ds DNA.ss CI.h,Cro.h ;CI.h; CI.m,Cro.h CI.h,Cro.l ;CI.m; CI.l,Cro.h CI.m,Cro.l ;Cro.h,~CII.h; CI.l;Cro.h; ;CI.l; CI.0,Cro.h CI.l,Cro.l CI.l;; (inject),ci.0,cro.0 Cro.0 -CI.m!,-CI.h!;.RNAP.tr,-Cro.l,-Cro.h!;CI.l -Cro.h!;.RNAP.tr,-CI.l!,-CI.m!,-CI.h!; ;.RNAP.tr,-CI.l!,-CI.m!,-CI.h!; CI.l CI.h Cro.l CI.l;.RNAP.tr,-Cro.l,-Cro.h!; Cro.h;~CI.l,~CI.m,~CI.h; ;.RNAP.tr,-CI.l!,-CI.m!,-CI.h!; CI.m Cro.h CI.h,RecA.* ;RecA.*; CI.m,RecA.* CI.l;.RNAP.tr,-Cro.l,-Cro.h!; CI.l,RecA.* ;RecA.*; ;RecA.*; CI.0,RecA.* ;RecA.*; RecA.* ;-DNA.ss; RecA. DNA.ss,RecA. ;DNA.ss; DNA.ss,RecA.* CEqEA cascaded causation (v28/e39/c8; i8/x_/a_/n_) lambda[ags3]_ri ctxt: RNAP.tr
16 Independence // 1/2 (cont d): sustaining Definition (A/T/N-sustainers) States may be Always present/transitory/never present. Definition (A/T/N-sustained equilibria) Given a classification of the considered states as A/T/N sustainers, we first find all strongly-connected components over the graph without A-inhibited edges. We then classify any found component as a sustained equilibrium of the given A/T/N type if it has no out-edges in the graph without A- and/or T-inhibited edges. By default, we consider only direct inhibitors for sustaining: nested inhibition tends to express precedence. Definition (Min/Max-sustained equilibria) Sustained equilibria with all states classified as transitory are referred to as max equilibria and are annotated to cascaded causation diagrams as red boxes. Sustained equilibria with all states classified as always present are referred to as min equilibria and are annotated to cascaded causation diagrams as blue boxes.
17 Independence // 1/2 (cont d): min/max sustaining ;CII.h,.RNAP.tr; CII.h CI.0,CII.h CI.l,CII.h -CI.l!,-CI.m!,-CI.h!;.RNAP.tr;CII.h CII._ DNA.ds DNA.ss CI.l,Cro.h CI.l;Cro.h; ;Cro.h,~CII.h; ;CI.l; CI.0,Cro.h (inject),ci.0,cro.0 -CI.m!,-CI.h!;.RNAP.tr,-Cro.l,-Cro.h!;CI.l -Cro.h!;.RNAP.tr,-CI.l!,-CI.m!,-CI.h!; CI.l CI.l;.RNAP.tr,-Cro.l,-Cro.h!; CI.m CI.h,Cro.h ;CI.h; CI.m,Cro.h CI.h,Cro.l ;CI.m; CI.m,Cro.l CI.l,Cro.l CI.l;; Cro.0 ;.RNAP.tr,-CI.l!,-CI.m!,-CI.h!; CI.h Cro.l ;.RNAP.tr,-CI.l!,-CI.m!,-CI.h!; Cro.h;~CI.l,~CI.m,~CI.h; Cro.h ;RecA.*; CI.h,RecA.* ;RecA.*; CI.m,RecA.* CI.l,RecA.* ;RecA.*; ;RecA.*; CI.0,RecA.* CI.l;.RNAP.tr,-Cro.l,-Cro.h!; RecA.* ;-DNA.ss; RecA. DNA.ss,RecA. ;DNA.ss; DNA.ss,RecA.* CEqEA cascaded causation (v28/e39/c8; i_/x9/a_/n9) lambda[ags3]_ri ctxt: RNAP.tr
18 Independence 2/2: sustained-equilibria coexistability Definition (Reconcilable) Each node in a sustained equilibrium must be able to pair up with a node from each of the other sustained equilibria without having some schema instance be in two states.. Definition (Orthogonal USED HERE!) A collection of sustained equilibria are orthogonal if no schema instance is in different states in (strictly) different equilibria and if no sustained equilibrium loses strong-connectivity under inhibition by the content of the other sustained equilibria.
19 Targeted cascaded causation diagrams: exogeny CI.h ;; CI.l;Cro.h; CI.0,Cro.h CI.l,Cro.h ;Cro.h; ;; CI.m ;CI.l; ;; CI.m,Cro.h ;CI.m; -CI.m!,-CI.h!;RNAP.tr,-Cro.l,-Cro.h!;CI.l CI.l CI.l;RNAP.tr,-Cro.l,-Cro.h!; CI.h,Cro.h ;; CI.l,Cro.l (inject),ci.0,cro.0 -Cro.h!;RNAP.tr,-CI.l!,-CI.m!,-CI.h!; ;CI.h; CI.l;; CI.m,Cro.l ;; CI.h,Cro.l ;; Cro.0 ;RNAP.tr,-CI.l!,-CI.m!,-CI.h!; Cro.l Cro.h;~CI.l,~CI.m,~CI.h; Cro.h ;RNAP.tr,-CI.l!,-CI.m!,-CI.h!; CEqEA cascaded causation (v14/e26/c1; i_/x4/a_/n4) lambda[ags3]_ri ctxt#1: [CII._]; RNAP.tr CI.h CI.l;Cro.h; ;CII.h,RNAP.tr; CI.0,Cro.h CI.0 ;CII.h,RNAP.tr; -CI.l!,-CI.m!,-CI.h!;CII.h,RNAP.tr; CI.l CI.l;RNAP.tr,-Cro.l,-Cro.h!; CI.m CI.l,Cro.h ;CI.l; (inject),ci.0,cro.0 -CI.m!,-CI.h!;RNAP.tr,-Cro.l,-Cro.h!;CI.l CI.h,Cro.h ;CI.h; CI.m,Cro.h CI.h,Cro.l ;CI.m; CI.m,Cro.l CI.l,Cro.l CI.l;; Cro.0 -Cro.h!;RNAP.tr,-CI.l!,-CI.m!,-CI.h!; ;RNAP.tr,-CI.l!,-CI.m!,-CI.h!; Cro.l Cro.h;~CI.l,~CI.m,~CI.h; ;RNAP.tr,-CI.l!,-CI.m!,-CI.h!; Cro.h CEqEA cascaded causation (v15/e22/c1; i_/x3/a_/n3) lambda[ags3]_ri ctxt#2: [CII.h]; RNAP.tr
20 λ [AGS3] synchronization diagrams, w/molecular details (E) (F) (A) (B) (D) (A) (A) (B) (C) Synchronization points are groups of indistinguishable causation nodes, with expiration (red text) and channels (magenta background) lifted from the causation level. The synchro-expiration is terminal only in that sense, meaning lysis is not regulatorily terminal, see Retrodiction (II). The synchro-channel contains both inter- and intra-nodes, meaning it combines the two top nodes in Ptashne s diagram. Edges with a filled arrow head are subject to the (E) TargetRequired condition: some requisite mediator is present only in the target node. This implies that the direct route to λ-lysogeny is available only to a (F) super-infection in the considered case, namely with CII below high concentration, see 1/ and 2/b/iii/. The corresponding edge in the (D) (F) synchronization diagram with CII at high concentration is not TargetRequired, see Sect.S4.4, meaning fresh λ-lysogeny is possible in that case, see 2/a/α/. (C) Dashed edges are subject to inhibition in the target node. CI is an inhibitor for the left edge, (D) meaning only continued λ-lysogeny is possible in case of super-infection, see 2/b/iii/. For the right edge, see Retrodiction (III). Dotted edges are subject to inhibition in the source node. The inhibitor here is CI, meaning a lytic attack requires CI to become non-physiological by means other than what the λ-genotype codes for or by a (severe) stochastic event, see 2/{a,b}/ vs 2/c/. Reflexive edges indicate self-regulation. Here, of λ-lysogeny, see 2/a/. Boxes indicate synchro-sustainability, see Chp.S4. Here, of λ-lysogeny, while all other synchronization points are likely to be transient. jointly predict that the λ-lysogeny box will be relatively stable (in vacuo). Combined with the specifics of the out-edges of the intermediate synchronization point, we can further predict that λ-lysogeny will be homeostatic (in situ): if we are pushed out of the box, the only option will be to go back into the box in most circumstances.
21 super;temp;clear 0 00, {3, 2}0 {3, 2, 1}0 ; Combinatorial: pheno-over-genotype temperate phase {2, 1}... space... {2, 1} super-temperate 0 {3, 2}0 q; q q Fig.7 Phenotype phase space over intrinsic-ci-affinities genotypes phenotype classification virulent virulent/clear+ clear+ ;virulent clear+ /virulent clear+ clear temperate;clear super;temp;clear temperate super-temperate [ ] [h] 00,11 00,11 00,11 {2, 1}0 {3, 2}0 [ ] [ ] {3, 2}0 ; {3, 2}0 {2, 1}0 ; {3, 2, 1}0 ; {2, 1}0 {3, 2}0 ( ) + ( ) UV q; q q; q; @OR1_CI_int Compound phenotypes Key: The pure and compound phenotypes that may result from varying CI s int affinities see Chp.S5: 1 is for low concentration ( l ), 2 for medium ( m With / tend to left, may be(axes), right With : @OR1_CI_int
22 Main topics [Principia Mathematica]: Principles of molecular-biology reasoning: A Genetic Switch is formally correct [Curry-Howard]: Mechanisms of molecular-biology reasoning: A Genetic Switch fits together phage λ [Proof assistant]: Formally-verified cause-and-effect reasoning for emergent properties [Deep structure]: Combinatorial basis of morphogenesis: elementary processes of digitation [Computation]: Synchrotyping
arxiv: v1 [q-bio.ot] 30 Oct 2018
![arxiv: v1 [q-bio.ot] 30 Oct 2018](/thumbs/93/112987397.jpg "arxiv: v1 [q-bio.ot] 30 Oct 2018") arxiv:1811.02478v1 [q-bio.ot] 30 Oct 2018 Proofs of life: molecular-biology reasoning simulates cell behaviors from first principles René Vestergaard (no current affiliation) Emmanuel Pietriga (Inria,
arxiv:1811.02478v1 [q-bio.ot] 30 Oct 2018 Proofs of life: molecular-biology reasoning simulates cell behaviors from first principles René Vestergaard (no current affiliation) Emmanuel Pietriga (Inria,
7.32/7.81J/8.591J. Rm Rm (under construction) Alexander van Oudenaarden Jialing Li. Bernardo Pando. Rm.
 Alexander van Oudenaarden Jialing Li. Bernardo Pando. Rm.") Introducing... 7.32/7.81J/8.591J Systems Biology modeling biological networks Lectures: Recitations: ti TR 1:00-2:30 PM W 4:00-5:00 PM Rm. 6-120 Rm. 26-204 (under construction) Alexander van Oudenaarden
Introducing... 7.32/7.81J/8.591J Systems Biology modeling biological networks Lectures: Recitations: ti TR 1:00-2:30 PM W 4:00-5:00 PM Rm. 6-120 Rm. 26-204 (under construction) Alexander van Oudenaarden
Lecture 7: Simple genetic circuits I
 Lecture 7: Simple genetic circuits I Paul C Bressloff (Fall 2018) 7.1 Transcription and translation In Fig. 20 we show the two main stages in the expression of a single gene according to the central dogma.
Lecture 7: Simple genetic circuits I Paul C Bressloff (Fall 2018) 7.1 Transcription and translation In Fig. 20 we show the two main stages in the expression of a single gene according to the central dogma.
Stochastic simulations
 Stochastic simulations Application to molecular networks Literature overview Noise in genetic networks Origins How to measure and distinguish between the two types of noise (intrinsic vs extrinsic)? What
Stochastic simulations Application to molecular networks Literature overview Noise in genetic networks Origins How to measure and distinguish between the two types of noise (intrinsic vs extrinsic)? What
Analysis of Mode Transitions in Biological Networks
 Analysis of Mode Transitions in Biological Networks Nael H. El-Farra, Adiwinata Gani, and Panagiotis D. Christofides Dept. of Chemical Engineering, University of California, Los Angeles, CA 90095 DOI 10.1002/aic.10499
Analysis of Mode Transitions in Biological Networks Nael H. El-Farra, Adiwinata Gani, and Panagiotis D. Christofides Dept. of Chemical Engineering, University of California, Los Angeles, CA 90095 DOI 10.1002/aic.10499
CHAPTER : Prokaryotic Genetics
 CHAPTER 13.3 13.5: Prokaryotic Genetics 1. Most bacteria are not pathogenic. Identify several important roles they play in the ecosystem and human culture. 2. How do variations arise in bacteria considering
CHAPTER 13.3 13.5: Prokaryotic Genetics 1. Most bacteria are not pathogenic. Identify several important roles they play in the ecosystem and human culture. 2. How do variations arise in bacteria considering
Regulation of Gene Expression in Bacteria and Their Viruses
 11 Regulation of Gene Expression in Bacteria and Their Viruses WORKING WITH THE FIGURES 1. Compare the structure of IPTG shown in Figure 11-7 with the structure of galactose shown in Figure 11-5. Why is
11 Regulation of Gene Expression in Bacteria and Their Viruses WORKING WITH THE FIGURES 1. Compare the structure of IPTG shown in Figure 11-7 with the structure of galactose shown in Figure 11-5. Why is
GENETICS - CLUTCH CH.12 GENE REGULATION IN PROKARYOTES.
 GEETICS - CLUTCH CH.12 GEE REGULATIO I PROKARYOTES!! www.clutchprep.com GEETICS - CLUTCH CH.12 GEE REGULATIO I PROKARYOTES COCEPT: LAC OPERO An operon is a group of genes with similar functions that are
GEETICS - CLUTCH CH.12 GEE REGULATIO I PROKARYOTES!! www.clutchprep.com GEETICS - CLUTCH CH.12 GEE REGULATIO I PROKARYOTES COCEPT: LAC OPERO An operon is a group of genes with similar functions that are
Models of transcriptional regulation
 Models of transcriptional regulation We have already discussed four simple mechanisms of transcriptional regulation, nuclear exclusion nuclear concentration modification of bound activator redirection
Models of transcriptional regulation We have already discussed four simple mechanisms of transcriptional regulation, nuclear exclusion nuclear concentration modification of bound activator redirection
Lecture 4: Transcription networks basic concepts
 Lecture 4: Transcription networks basic concepts - Activators and repressors - Input functions; Logic input functions; Multidimensional input functions - Dynamics and response time 2.1 Introduction The
Lecture 4: Transcription networks basic concepts - Activators and repressors - Input functions; Logic input functions; Multidimensional input functions - Dynamics and response time 2.1 Introduction The
Biological Pathways Representation by Petri Nets and extension
 Biological Pathways Representation by and extensions December 6, 2006 Biological Pathways Representation by and extension 1 The cell Pathways 2 Definitions 3 4 Biological Pathways Representation by and
Biological Pathways Representation by and extensions December 6, 2006 Biological Pathways Representation by and extension 1 The cell Pathways 2 Definitions 3 4 Biological Pathways Representation by and
Lysogeny maintenance: a matter of looping. Laura Finzi
 Lysogeny maintenance: a matter of looping Laura Finzi λ is a temperate phage Two possible modes LYSOGENIC MODE (passive replication) LYTIC MODE (active replication) Lytic mode Lysogenic mode replication
Lysogeny maintenance: a matter of looping Laura Finzi λ is a temperate phage Two possible modes LYSOGENIC MODE (passive replication) LYTIC MODE (active replication) Lytic mode Lysogenic mode replication
REGULATION OF GENE EXPRESSION. Bacterial Genetics Lac and Trp Operon
 REGULATION OF GENE EXPRESSION Bacterial Genetics Lac and Trp Operon Levels of Metabolic Control The amount of cellular products can be controlled by regulating: Enzyme activity: alters protein function
REGULATION OF GENE EXPRESSION Bacterial Genetics Lac and Trp Operon Levels of Metabolic Control The amount of cellular products can be controlled by regulating: Enzyme activity: alters protein function
3.B.1 Gene Regulation. Gene regulation results in differential gene expression, leading to cell specialization.
 3.B.1 Gene Regulation Gene regulation results in differential gene expression, leading to cell specialization. We will focus on gene regulation in prokaryotes first. Gene regulation accounts for some of
3.B.1 Gene Regulation Gene regulation results in differential gene expression, leading to cell specialization. We will focus on gene regulation in prokaryotes first. Gene regulation accounts for some of
Is chaos possible in 1d? - yes - no - I don t know. What is the long term behavior for the following system if x(0) = π/2?
 = π/2?") Is chaos possible in 1d? - yes - no - I don t know What is the long term behavior for the following system if x(0) = π/2? In the insect outbreak problem, what kind of bifurcation occurs at fixed value
Is chaos possible in 1d? - yes - no - I don t know What is the long term behavior for the following system if x(0) = π/2? In the insect outbreak problem, what kind of bifurcation occurs at fixed value
APGRU6L2. Control of Prokaryotic (Bacterial) Genes
 Genes") APGRU6L2 Control of Prokaryotic (Bacterial) Genes 2007-2008 Bacterial metabolism Bacteria need to respond quickly to changes in their environment STOP u if they have enough of a product, need to stop production
APGRU6L2 Control of Prokaryotic (Bacterial) Genes 2007-2008 Bacterial metabolism Bacteria need to respond quickly to changes in their environment STOP u if they have enough of a product, need to stop production
CHAPTER 10. Gentzen Style Proof Systems for Classical Logic
 CHAPTER 10 Gentzen Style Proof Systems for Classical Logic Hilbert style systems are easy to define and admit a simple proof of the Completeness Theorem but they are difficult to use. By humans, not mentioning
CHAPTER 10 Gentzen Style Proof Systems for Classical Logic Hilbert style systems are easy to define and admit a simple proof of the Completeness Theorem but they are difficult to use. By humans, not mentioning
Efficient Circuit Analysis
 Efficient Circuit Analysis Chris Myers, Nathan Barker, Kevin Jones, Hiroyuki Kuwahara, Scott Little, Curtis Madsen, Nam Nguyen, Robert Thacker, David Walter University of Utah CE Junior Seminar September
Efficient Circuit Analysis Chris Myers, Nathan Barker, Kevin Jones, Hiroyuki Kuwahara, Scott Little, Curtis Madsen, Nam Nguyen, Robert Thacker, David Walter University of Utah CE Junior Seminar September
ANALYSIS OF BIOLOGICAL NETWORKS USING HYBRID SYSTEMS THEORY. Nael H. El-Farra, Adiwinata Gani & Panagiotis D. Christofides
 ANALYSIS OF BIOLOGICAL NETWORKS USING HYBRID SYSTEMS THEORY Nael H El-Farra, Adiwinata Gani & Panagiotis D Christofides Department of Chemical Engineering University of California, Los Angeles 2003 AIChE
ANALYSIS OF BIOLOGICAL NETWORKS USING HYBRID SYSTEMS THEORY Nael H El-Farra, Adiwinata Gani & Panagiotis D Christofides Department of Chemical Engineering University of California, Los Angeles 2003 AIChE
Control of Gene Expression in Prokaryotes
 Why? Control of Expression in Prokaryotes How do prokaryotes use operons to control gene expression? Houses usually have a light source in every room, but it would be a waste of energy to leave every light
Why? Control of Expression in Prokaryotes How do prokaryotes use operons to control gene expression? Houses usually have a light source in every room, but it would be a waste of energy to leave every light
Name Period The Control of Gene Expression in Prokaryotes Notes
 Bacterial DNA contains genes that encode for many different proteins (enzymes) so that many processes have the ability to occur -not all processes are carried out at any one time -what allows expression
Bacterial DNA contains genes that encode for many different proteins (enzymes) so that many processes have the ability to occur -not all processes are carried out at any one time -what allows expression
Welcome to Class 21!
 Welcome to Class 21! Introductory Biochemistry! Lecture 21: Outline and Objectives l Regulation of Gene Expression in Prokaryotes! l transcriptional regulation! l principles! l lac operon! l trp attenuation!
Welcome to Class 21! Introductory Biochemistry! Lecture 21: Outline and Objectives l Regulation of Gene Expression in Prokaryotes! l transcriptional regulation! l principles! l lac operon! l trp attenuation!
Written Exam 15 December Course name: Introduction to Systems Biology Course no
 Technical University of Denmark Written Exam 15 December 2008 Course name: Introduction to Systems Biology Course no. 27041 Aids allowed: Open book exam Provide your answers and calculations on separate
Technical University of Denmark Written Exam 15 December 2008 Course name: Introduction to Systems Biology Course no. 27041 Aids allowed: Open book exam Provide your answers and calculations on separate
Bacterial Genetics & Operons
 Bacterial Genetics & Operons The Bacterial Genome Because bacteria have simple genomes, they are used most often in molecular genetics studies Most of what we know about bacterial genetics comes from the
Bacterial Genetics & Operons The Bacterial Genome Because bacteria have simple genomes, they are used most often in molecular genetics studies Most of what we know about bacterial genetics comes from the
Control of Gene Expression
 Control of Gene Expression Mechanisms of Gene Control Gene Control in Eukaryotes Master Genes Gene Control In Prokaryotes Epigenetics Gene Expression The overall process by which information flows from
Control of Gene Expression Mechanisms of Gene Control Gene Control in Eukaryotes Master Genes Gene Control In Prokaryotes Epigenetics Gene Expression The overall process by which information flows from
Propositional Logic. Fall () Propositional Logic Fall / 30
 Propositional Logic Fall / 30") Propositional Logic Fall 2013 () Propositional Logic Fall 2013 1 / 30 1 Introduction Learning Outcomes for this Presentation 2 Definitions Statements Logical connectives Interpretations, contexts,... Logically
Propositional Logic Fall 2013 () Propositional Logic Fall 2013 1 / 30 1 Introduction Learning Outcomes for this Presentation 2 Definitions Statements Logical connectives Interpretations, contexts,... Logically
Propositional Logic. Spring Propositional Logic Spring / 32
 Propositional Logic Spring 2016 Propositional Logic Spring 2016 1 / 32 Introduction Learning Outcomes for this Presentation Learning Outcomes... At the conclusion of this session, we will Define the elements
Propositional Logic Spring 2016 Propositional Logic Spring 2016 1 / 32 Introduction Learning Outcomes for this Presentation Learning Outcomes... At the conclusion of this session, we will Define the elements
Basic Synthetic Biology circuits
 Basic Synthetic Biology circuits Note: these practices were obtained from the Computer Modelling Practicals lecture by Vincent Rouilly and Geoff Baldwin at Imperial College s course of Introduction to
Basic Synthetic Biology circuits Note: these practices were obtained from the Computer Modelling Practicals lecture by Vincent Rouilly and Geoff Baldwin at Imperial College s course of Introduction to
arxiv: v1 [q-bio.mn] 14 May 2009
![arxiv: v1 [q-bio.mn] 14 May 2009](/thumbs/75/72878987.jpg "arxiv: v1 [q-bio.mn] 14 May 2009") , DNA looping provides stability and robustness to the bacteriophage λ switch Marco J. Morelli, 1,2 Pieter Rein ten Wolde, 1 and Rosalind J. Allen 3 1 FOM Institute for Atomic and Molecular Physics, arxiv:0905.2329v1
, DNA looping provides stability and robustness to the bacteriophage λ switch Marco J. Morelli, 1,2 Pieter Rein ten Wolde, 1 and Rosalind J. Allen 3 1 FOM Institute for Atomic and Molecular Physics, arxiv:0905.2329v1
Modeling Multiple Steady States in Genetic Regulatory Networks. Khang Tran. problem.
 Modeling Multiple Steady States in Genetic Regulatory Networks Khang Tran From networks of simple regulatory elements, scientists have shown some simple circuits such as the circadian oscillator 1 or the
Modeling Multiple Steady States in Genetic Regulatory Networks Khang Tran From networks of simple regulatory elements, scientists have shown some simple circuits such as the circadian oscillator 1 or the
Bi 8 Lecture 11. Quantitative aspects of transcription factor binding and gene regulatory circuit design. Ellen Rothenberg 9 February 2016
 Bi 8 Lecture 11 Quantitative aspects of transcription factor binding and gene regulatory circuit design Ellen Rothenberg 9 February 2016 Major take-home messages from λ phage system that apply to many
Bi 8 Lecture 11 Quantitative aspects of transcription factor binding and gene regulatory circuit design Ellen Rothenberg 9 February 2016 Major take-home messages from λ phage system that apply to many
Chapter 15 Active Reading Guide Regulation of Gene Expression
 Name: AP Biology Mr. Croft Chapter 15 Active Reading Guide Regulation of Gene Expression The overview for Chapter 15 introduces the idea that while all cells of an organism have all genes in the genome,
Name: AP Biology Mr. Croft Chapter 15 Active Reading Guide Regulation of Gene Expression The overview for Chapter 15 introduces the idea that while all cells of an organism have all genes in the genome,
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
A Simple Protein Synthesis Model
 A Simple Protein Synthesis Model James K. Peterson Department of Biological Sciences and Department of Mathematical Sciences Clemson University September 3, 213 Outline A Simple Protein Synthesis Model
A Simple Protein Synthesis Model James K. Peterson Department of Biological Sciences and Department of Mathematical Sciences Clemson University September 3, 213 Outline A Simple Protein Synthesis Model
Designer gene networks: Towards fundamental cellular control
 CHAOS VOLUME 11, NUMBER 1 MARCH 2001 Designer gene networks: Towards fundamental cellular control Jeff Hasty a) and Farren Isaacs Center for BioDynamics and Department of Biomedical Engineering, Boston
CHAOS VOLUME 11, NUMBER 1 MARCH 2001 Designer gene networks: Towards fundamental cellular control Jeff Hasty a) and Farren Isaacs Center for BioDynamics and Department of Biomedical Engineering, Boston
Simulation of Gene Regulatory Networks
 Simulation of Gene Regulatory Networks Overview I have been assisting Professor Jacques Cohen at Brandeis University to explore and compare the the many available representations and interpretations of
Simulation of Gene Regulatory Networks Overview I have been assisting Professor Jacques Cohen at Brandeis University to explore and compare the the many available representations and interpretations of
Principles of Synthetic Biology: Midterm Exam
 Principles of Synthetic Biology: Midterm Exam October 28, 2010 1 Conceptual Simple Circuits 1.1 Consider the plots in figure 1. Identify all critical points with an x. Put a circle around the x for each
Principles of Synthetic Biology: Midterm Exam October 28, 2010 1 Conceptual Simple Circuits 1.1 Consider the plots in figure 1. Identify all critical points with an x. Put a circle around the x for each
Computational Modelling in Systems and Synthetic Biology
 Computational Modelling in Systems and Synthetic Biology Fran Romero Dpt Computer Science and Artificial Intelligence University of Seville fran@us.es www.cs.us.es/~fran Models are Formal Statements of
Computational Modelling in Systems and Synthetic Biology Fran Romero Dpt Computer Science and Artificial Intelligence University of Seville fran@us.es www.cs.us.es/~fran Models are Formal Statements of
Computational Genomics. Systems biology. Putting it together: Data integration using graphical models
 02-710 Computational Genomics Systems biology Putting it together: Data integration using graphical models High throughput data So far in this class we discussed several different types of high throughput
02-710 Computational Genomics Systems biology Putting it together: Data integration using graphical models High throughput data So far in this class we discussed several different types of high throughput
SPECIES OF ARCHAEA ARE MORE CLOSELY RELATED TO EUKARYOTES THAN ARE SPECIES OF PROKARYOTES.
 THE TERMS RUN AND TUMBLE ARE GENERALLY ASSOCIATED WITH A) cell wall fluidity. B) cell membrane structures. C) taxic movements of the cell. D) clustering properties of certain rod-shaped bacteria. A MAJOR
THE TERMS RUN AND TUMBLE ARE GENERALLY ASSOCIATED WITH A) cell wall fluidity. B) cell membrane structures. C) taxic movements of the cell. D) clustering properties of certain rod-shaped bacteria. A MAJOR
Intrinsic Noise in Nonlinear Gene Regulation Inference
 Intrinsic Noise in Nonlinear Gene Regulation Inference Chao Du Department of Statistics, University of Virginia Joint Work with Wing H. Wong, Department of Statistics, Stanford University Transcription
Intrinsic Noise in Nonlinear Gene Regulation Inference Chao Du Department of Statistics, University of Virginia Joint Work with Wing H. Wong, Department of Statistics, Stanford University Transcription
New Computational Methods for Systems Biology
 New Computational Methods for Systems Biology François Fages, Sylvain Soliman The French National Institute for Research in Computer Science and Control INRIA Paris-Rocquencourt Constraint Programming
New Computational Methods for Systems Biology François Fages, Sylvain Soliman The French National Institute for Research in Computer Science and Control INRIA Paris-Rocquencourt Constraint Programming
Boolean models of gene regulatory networks. Matthew Macauley Math 4500: Mathematical Modeling Clemson University Spring 2016
 Boolean models of gene regulatory networks Matthew Macauley Math 4500: Mathematical Modeling Clemson University Spring 2016 Gene expression Gene expression is a process that takes gene info and creates
Boolean models of gene regulatory networks Matthew Macauley Math 4500: Mathematical Modeling Clemson University Spring 2016 Gene expression Gene expression is a process that takes gene info and creates
Introduction. ECE/CS/BioEn 6760 Modeling and Analysis of Biological Networks. Adventures in Synthetic Biology. Synthetic Biology.
 Introduction ECE/CS/BioEn 6760 Modeling and Analysis of Biological Networks Chris J. Myers Lecture 17: Genetic Circuit Design Electrical engineering principles can be applied to understand the behavior
Introduction ECE/CS/BioEn 6760 Modeling and Analysis of Biological Networks Chris J. Myers Lecture 17: Genetic Circuit Design Electrical engineering principles can be applied to understand the behavior
Design principles for elementary gene circuits: Elements, methods, and examples
 CHAOS VOLUME 11, NUMBER 1 MARCH 2001 Design principles for elementary gene circuits: Elements, methods, and examples Michael A. Savageau a) Department of Microbiology and Immunology, University of Michigan
CHAOS VOLUME 11, NUMBER 1 MARCH 2001 Design principles for elementary gene circuits: Elements, methods, and examples Michael A. Savageau a) Department of Microbiology and Immunology, University of Michigan
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Tuesday, December 27, 16
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
GLOBEX Bioinformatics (Summer 2015) Genetic networks and gene expression data
 Genetic networks and gene expression data") GLOBEX Bioinformatics (Summer 2015) Genetic networks and gene expression data 1 Gene Networks Definition: A gene network is a set of molecular components, such as genes and proteins, and interactions between
GLOBEX Bioinformatics (Summer 2015) Genetic networks and gene expression data 1 Gene Networks Definition: A gene network is a set of molecular components, such as genes and proteins, and interactions between
Deducing Interactions in Partially Unspecified Biological Systems
 Deducing Interactions in Partially Unspecified Biological Systems P. Baldan 1 A. Bracciali 2 L. Brodo 3 R. Bruni 2 1 Università di Padova 2 Università di Pisa 3 Università di Sassari Algebraic Biology
Deducing Interactions in Partially Unspecified Biological Systems P. Baldan 1 A. Bracciali 2 L. Brodo 3 R. Bruni 2 1 Università di Padova 2 Università di Pisa 3 Università di Sassari Algebraic Biology
Lecture 18 June 2 nd, Gene Expression Regulation Mutations
 Lecture 18 June 2 nd, 2016 Gene Expression Regulation Mutations From Gene to Protein Central Dogma Replication DNA RNA PROTEIN Transcription Translation RNA Viruses: genome is RNA Reverse Transcriptase
Lecture 18 June 2 nd, 2016 Gene Expression Regulation Mutations From Gene to Protein Central Dogma Replication DNA RNA PROTEIN Transcription Translation RNA Viruses: genome is RNA Reverse Transcriptase
86 Part 4 SUMMARY INTRODUCTION
 86 Part 4 Chapter # AN INTEGRATION OF THE DESCRIPTIONS OF GENE NETWORKS AND THEIR MODELS PRESENTED IN SIGMOID (CELLERATOR) AND GENENET Podkolodny N.L. *1, 2, Podkolodnaya N.N. 1, Miginsky D.S. 1, Poplavsky
86 Part 4 Chapter # AN INTEGRATION OF THE DESCRIPTIONS OF GENE NETWORKS AND THEIR MODELS PRESENTED IN SIGMOID (CELLERATOR) AND GENENET Podkolodny N.L. *1, 2, Podkolodnaya N.N. 1, Miginsky D.S. 1, Poplavsky
Modeling and Systems Analysis of Gene Regulatory Networks
 Modeling and Systems Analysis of Gene Regulatory Networks Mustafa Khammash Center for Control Dynamical-Systems and Computations University of California, Santa Barbara Outline Deterministic A case study:
Modeling and Systems Analysis of Gene Regulatory Networks Mustafa Khammash Center for Control Dynamical-Systems and Computations University of California, Santa Barbara Outline Deterministic A case study:
SPA for quantitative analysis: Lecture 6 Modelling Biological Processes
 1/ 223 SPA for quantitative analysis: Lecture 6 Modelling Biological Processes Jane Hillston LFCS, School of Informatics The University of Edinburgh Scotland 7th March 2013 Outline 2/ 223 1 Introduction
1/ 223 SPA for quantitative analysis: Lecture 6 Modelling Biological Processes Jane Hillston LFCS, School of Informatics The University of Edinburgh Scotland 7th March 2013 Outline 2/ 223 1 Introduction
Division Ave. High School AP Biology
 Control of Prokaryotic (Bacterial) Genes 20072008 Bacterial metabolism n Bacteria need to respond quickly to changes in their environment u if they have enough of a product, need to stop production n why?
Control of Prokaryotic (Bacterial) Genes 20072008 Bacterial metabolism n Bacteria need to respond quickly to changes in their environment u if they have enough of a product, need to stop production n why?
Control of Prokaryotic (Bacterial) Gene Expression. AP Biology
 Gene Expression. AP Biology") Control of Prokaryotic (Bacterial) Gene Expression Figure 18.1 How can this fish s eyes see equally well in both air and water? Aka. Quatro ojas Regulation of Gene Expression: Prokaryotes and eukaryotes
Control of Prokaryotic (Bacterial) Gene Expression Figure 18.1 How can this fish s eyes see equally well in both air and water? Aka. Quatro ojas Regulation of Gene Expression: Prokaryotes and eukaryotes
Simplicity is Complexity in Masquerade. Michael A. Savageau The University of California, Davis July 2004
 Simplicity is Complexity in Masquerade Michael A. Savageau The University of California, Davis July 2004 Complexity is Not Simplicity in Masquerade -- E. Yates Simplicity is Complexity in Masquerade One
Simplicity is Complexity in Masquerade Michael A. Savageau The University of California, Davis July 2004 Complexity is Not Simplicity in Masquerade -- E. Yates Simplicity is Complexity in Masquerade One
Living Mechanisms as Complex Systems
 Living Mechanisms as Complex Systems The Traditional Mechanistic Perspective Decompose the organism into parts and operations Understand each on its own Link the whole together (sequentially, linearly)
Living Mechanisms as Complex Systems The Traditional Mechanistic Perspective Decompose the organism into parts and operations Understand each on its own Link the whole together (sequentially, linearly)
GENE REGULATION AND PROBLEMS OF DEVELOPMENT
 GENE REGULATION AND PROBLEMS OF DEVELOPMENT By Surinder Kaur DIET Ropar Surinder_1998@ yahoo.in Mob No 9988530775 GENE REGULATION Gene is a segment of DNA that codes for a unit of function (polypeptide,
GENE REGULATION AND PROBLEMS OF DEVELOPMENT By Surinder Kaur DIET Ropar Surinder_1998@ yahoo.in Mob No 9988530775 GENE REGULATION Gene is a segment of DNA that codes for a unit of function (polypeptide,
Networks in systems biology
 Networks in systems biology Matthew Macauley Department of Mathematical Sciences Clemson University http://www.math.clemson.edu/~macaule/ Math 4500, Spring 2017 M. Macauley (Clemson) Networks in systems
Networks in systems biology Matthew Macauley Department of Mathematical Sciences Clemson University http://www.math.clemson.edu/~macaule/ Math 4500, Spring 2017 M. Macauley (Clemson) Networks in systems
Deducing Interactions in Partially Unspecified Biological Systems
 Deducing Interactions in Partially Unspecified Biological Systems P. Baldan 1 A. Bracciali 2 L. Brodo 3 R. Bruni 2 1 Università di Padova 2 Università di Pisa 3 Università di Sassari Algebraic Biology
Deducing Interactions in Partially Unspecified Biological Systems P. Baldan 1 A. Bracciali 2 L. Brodo 3 R. Bruni 2 1 Università di Padova 2 Università di Pisa 3 Università di Sassari Algebraic Biology
Prokaryotic Gene Expression (Learning Objectives)
") Prokaryotic Gene Expression (Learning Objectives) 1. Learn how bacteria respond to changes of metabolites in their environment: short-term and longer-term. 2. Compare and contrast transcriptional control
Prokaryotic Gene Expression (Learning Objectives) 1. Learn how bacteria respond to changes of metabolites in their environment: short-term and longer-term. 2. Compare and contrast transcriptional control
Understanding Science Through the Lens of Computation. Richard M. Karp Nov. 3, 2007
 Understanding Science Through the Lens of Computation Richard M. Karp Nov. 3, 2007 The Computational Lens Exposes the computational nature of natural processes and provides a language for their description.
Understanding Science Through the Lens of Computation Richard M. Karp Nov. 3, 2007 The Computational Lens Exposes the computational nature of natural processes and provides a language for their description.
Unit 3: Control and regulation Higher Biology
 Unit 3: Control and regulation Higher Biology To study the roles that genes play in the control of growth and development of organisms To be able to Give some examples of features which are controlled
Unit 3: Control and regulation Higher Biology To study the roles that genes play in the control of growth and development of organisms To be able to Give some examples of features which are controlled
UNIT 6 PART 3 *REGULATION USING OPERONS* Hillis Textbook, CH 11
 UNIT 6 PART 3 *REGULATION USING OPERONS* Hillis Textbook, CH 11 REVIEW: Signals that Start and Stop Transcription and Translation BUT, HOW DO CELLS CONTROL WHICH GENES ARE EXPRESSED AND WHEN? First of
UNIT 6 PART 3 *REGULATION USING OPERONS* Hillis Textbook, CH 11 REVIEW: Signals that Start and Stop Transcription and Translation BUT, HOW DO CELLS CONTROL WHICH GENES ARE EXPRESSED AND WHEN? First of
Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
Chapter 18 Regulation of Gene Expression PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
3 The Semantics of the Propositional Calculus
 3 The Semantics of the Propositional Calculus 1. Interpretations Formulas of the propositional calculus express statement forms. In chapter two, we gave informal descriptions of the meanings of the logical
3 The Semantics of the Propositional Calculus 1. Interpretations Formulas of the propositional calculus express statement forms. In chapter two, we gave informal descriptions of the meanings of the logical
CHAPTER 2 THE CHEMICAL BASIS OF LIFE
 CHAPTER 2 THE CHEMICAL BASIS OF LIFE CHAPTER OVERVIEW This chapter introduces very basic concepts of chemistry, emphasizing the structure of atoms and how they combine (form bonds). The types of bonds,
CHAPTER 2 THE CHEMICAL BASIS OF LIFE CHAPTER OVERVIEW This chapter introduces very basic concepts of chemistry, emphasizing the structure of atoms and how they combine (form bonds). The types of bonds,
4. Why not make all enzymes all the time (even if not needed)? Enzyme synthesis uses a lot of energy.
? Enzyme synthesis uses a lot of energy.") 1 C2005/F2401 '10-- Lecture 15 -- Last Edited: 11/02/10 01:58 PM Copyright 2010 Deborah Mowshowitz and Lawrence Chasin Department of Biological Sciences Columbia University New York, NY. Handouts: 15A
1 C2005/F2401 '10-- Lecture 15 -- Last Edited: 11/02/10 01:58 PM Copyright 2010 Deborah Mowshowitz and Lawrence Chasin Department of Biological Sciences Columbia University New York, NY. Handouts: 15A
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
REVIEW SESSION. Wednesday, September 15 5:30 PM SHANTZ 242 E
 REVIEW SESSION Wednesday, September 15 5:30 PM SHANTZ 242 E Gene Regulation Gene Regulation Gene expression can be turned on, turned off, turned up or turned down! For example, as test time approaches,
REVIEW SESSION Wednesday, September 15 5:30 PM SHANTZ 242 E Gene Regulation Gene Regulation Gene expression can be turned on, turned off, turned up or turned down! For example, as test time approaches,
Chapters AP Biology Objectives. Objectives: You should know...
 Objectives: You should know... Notes 1. Scientific evidence supports the idea that evolution has occurred in all species. 2. Scientific evidence supports the idea that evolution continues to occur. 3.
Objectives: You should know... Notes 1. Scientific evidence supports the idea that evolution has occurred in all species. 2. Scientific evidence supports the idea that evolution continues to occur. 3.
Supporting Information
 Supporting Information S. Discrete Chemical Master Equation Here we first discuss the chemical master equation on a discrete state space. The CME describes the gain and loss of probability associated with
Supporting Information S. Discrete Chemical Master Equation Here we first discuss the chemical master equation on a discrete state space. The CME describes the gain and loss of probability associated with
AP Biology Gene Regulation and Development Review
 AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
natural development from this collection of knowledge: it is more reliable to predict the property
 1 Chapter 1 Introduction As the basis of all life phenomena, the interaction of biomolecules has been under the scrutiny of scientists and cataloged meticulously [2]. The recent advent of systems biology
1 Chapter 1 Introduction As the basis of all life phenomena, the interaction of biomolecules has been under the scrutiny of scientists and cataloged meticulously [2]. The recent advent of systems biology
Chapter 11: Automated Proof Systems
 Chapter 11: Automated Proof Systems SYSTEM RS OVERVIEW Hilbert style systems are easy to define and admit a simple proof of the Completeness Theorem but they are difficult to use. Automated systems are
Chapter 11: Automated Proof Systems SYSTEM RS OVERVIEW Hilbert style systems are easy to define and admit a simple proof of the Completeness Theorem but they are difficult to use. Automated systems are
16 CONTROL OF GENE EXPRESSION
 16 CONTROL OF GENE EXPRESSION Chapter Outline 16.1 REGULATION OF GENE EXPRESSION IN PROKARYOTES The operon is the unit of transcription in prokaryotes The lac operon for lactose metabolism is transcribed
16 CONTROL OF GENE EXPRESSION Chapter Outline 16.1 REGULATION OF GENE EXPRESSION IN PROKARYOTES The operon is the unit of transcription in prokaryotes The lac operon for lactose metabolism is transcribed
56:198:582 Biological Networks Lecture 8
 56:198:582 Biological Networks Lecture 8 Course organization Two complementary approaches to modeling and understanding biological networks Constraint-based modeling (Palsson) System-wide Metabolism Steady-state
56:198:582 Biological Networks Lecture 8 Course organization Two complementary approaches to modeling and understanding biological networks Constraint-based modeling (Palsson) System-wide Metabolism Steady-state
K i + [R i ] n = [Rtot i ] (2) ] = β i [M i ] δ i [Ri tot ] (5) + α m j (6) 1 + r n i. dr i dt = βm i δr i (7) m j = βm i = δr i (9) 1 + r n + α.
![K i + [R i ] n = [Rtot i ] (2) ] = β i [M i ] δ i [Ri tot ] (5) + α m j (6) 1 + r n i. dr i dt = βm i δr i (7) m j = βm i = δr i (9) 1 + r n + α.](/thumbs/85/92758342.jpg "K i + [R i ] n = [Rtot i ] (2) ] = β i [M i ] δ i [Ri tot ] (5) + α m j (6) 1 + r n i. dr i dt = βm i δr i (7) m j = βm i = δr i (9) 1 + r n + α.") The lamba phage will remain in the lysogenic state if ci proteins preominate, but will be transforme into the lytic cycle if cro proteins preominate. cro: (Control of Repressor's Operator) Transcription
The lamba phage will remain in the lysogenic state if ci proteins preominate, but will be transforme into the lytic cycle if cro proteins preominate. cro: (Control of Repressor's Operator) Transcription
Equalities and Uninterpreted Functions. Chapter 3. Decision Procedures. An Algorithmic Point of View. Revision 1.0
 Equalities and Uninterpreted Functions Chapter 3 Decision Procedures An Algorithmic Point of View D.Kroening O.Strichman Revision 1.0 Outline Decision Procedures Equalities and Uninterpreted Functions
Equalities and Uninterpreted Functions Chapter 3 Decision Procedures An Algorithmic Point of View D.Kroening O.Strichman Revision 1.0 Outline Decision Procedures Equalities and Uninterpreted Functions
the noisy gene Biology of the Universidad Autónoma de Madrid Jan 2008 Juan F. Poyatos Spanish National Biotechnology Centre (CNB)
") Biology of the the noisy gene Universidad Autónoma de Madrid Jan 2008 Juan F. Poyatos Spanish National Biotechnology Centre (CNB) day III: noisy bacteria - Regulation of noise (B. subtilis) - Intrinsic/Extrinsic
Biology of the the noisy gene Universidad Autónoma de Madrid Jan 2008 Juan F. Poyatos Spanish National Biotechnology Centre (CNB) day III: noisy bacteria - Regulation of noise (B. subtilis) - Intrinsic/Extrinsic
2.2: Logical Equivalence: The Laws of Logic
 Example (2.7) For primitive statement p and q, construct a truth table for each of the following compound statements. a) p q b) p q Here we see that the corresponding truth tables for two statement p q
Example (2.7) For primitive statement p and q, construct a truth table for each of the following compound statements. a) p q b) p q Here we see that the corresponding truth tables for two statement p q
System Biology - Deterministic & Stochastic Dynamical Systems
 System Biology - Deterministic & Stochastic Dynamical Systems System Biology - Deterministic & Stochastic Dynamical Systems 1 The Cell System Biology - Deterministic & Stochastic Dynamical Systems 2 The
System Biology - Deterministic & Stochastic Dynamical Systems System Biology - Deterministic & Stochastic Dynamical Systems 1 The Cell System Biology - Deterministic & Stochastic Dynamical Systems 2 The
Chapter 17. From Gene to Protein. Biology Kevin Dees
 Chapter 17 From Gene to Protein DNA The information molecule Sequences of bases is a code DNA organized in to chromosomes Chromosomes are organized into genes What do the genes actually say??? Reflecting
Chapter 17 From Gene to Protein DNA The information molecule Sequences of bases is a code DNA organized in to chromosomes Chromosomes are organized into genes What do the genes actually say??? Reflecting
I. COMPOSITIONALITY, STOCHASTICITY AND COOPERATIVITY IN DY- NAMIC MODELS OF GENE REGULATION (SUPPLEMENTARY MATERIAL)
") I. COMPOSITIONALITY, STOCHASTICITY AND COOPERATIVITY IN DY- NAMIC MODELS OF GENE REGULATION (SUPPLEMENTARY MATERIAL) A. A primer on the stochastic π-calculus An ordinary function is a method of computing
I. COMPOSITIONALITY, STOCHASTICITY AND COOPERATIVITY IN DY- NAMIC MODELS OF GENE REGULATION (SUPPLEMENTARY MATERIAL) A. A primer on the stochastic π-calculus An ordinary function is a method of computing
Hybrid Model of gene regulatory networks, the case of the lac-operon
 Hybrid Model of gene regulatory networks, the case of the lac-operon Laurent Tournier and Etienne Farcot LMC-IMAG, 51 rue des Mathématiques, 38041 Grenoble Cedex 9, France Laurent.Tournier@imag.fr, Etienne.Farcot@imag.fr
Hybrid Model of gene regulatory networks, the case of the lac-operon Laurent Tournier and Etienne Farcot LMC-IMAG, 51 rue des Mathématiques, 38041 Grenoble Cedex 9, France Laurent.Tournier@imag.fr, Etienne.Farcot@imag.fr
2. What was the Avery-MacLeod-McCarty experiment and why was it significant? 3. What was the Hershey-Chase experiment and why was it significant?
 Name Date Period AP Exam Review Part 6: Molecular Genetics I. DNA and RNA Basics A. History of finding out what DNA really is 1. What was Griffith s experiment and why was it significant? 1 2. What was
Name Date Period AP Exam Review Part 6: Molecular Genetics I. DNA and RNA Basics A. History of finding out what DNA really is 1. What was Griffith s experiment and why was it significant? 1 2. What was
Decoherence and the Classical Limit
 Chapter 26 Decoherence and the Classical Limit 26.1 Introduction Classical mechanics deals with objects which have a precise location and move in a deterministic way as a function of time. By contrast,
Chapter 26 Decoherence and the Classical Limit 26.1 Introduction Classical mechanics deals with objects which have a precise location and move in a deterministic way as a function of time. By contrast,
More Protein Synthesis and a Model for Protein Transcription Error Rates
 More Protein Synthesis and a Model for Protein James K. Peterson Department of Biological Sciences and Department of Mathematical Sciences Clemson University October 3, 2013 Outline 1 Signal Patterns Example
More Protein Synthesis and a Model for Protein James K. Peterson Department of Biological Sciences and Department of Mathematical Sciences Clemson University October 3, 2013 Outline 1 Signal Patterns Example
A Weak Bisimulation for Weighted Automata
 Weak Bisimulation for Weighted utomata Peter Kemper College of William and Mary Weighted utomata and Semirings here focus on commutative & idempotent semirings Weak Bisimulation Composition operators Congruence
Weak Bisimulation for Weighted utomata Peter Kemper College of William and Mary Weighted utomata and Semirings here focus on commutative & idempotent semirings Weak Bisimulation Composition operators Congruence
2. Mathematical descriptions. (i) the master equation (ii) Langevin theory. 3. Single cell measurements
 the master equation (ii) Langevin theory. 3. Single cell measurements") 1. Why stochastic?. Mathematical descriptions (i) the master equation (ii) Langevin theory 3. Single cell measurements 4. Consequences Any chemical reaction is stochastic. k P d φ dp dt = k d P deterministic
1. Why stochastic?. Mathematical descriptions (i) the master equation (ii) Langevin theory 3. Single cell measurements 4. Consequences Any chemical reaction is stochastic. k P d φ dp dt = k d P deterministic
Enduring understanding 1.A: Change in the genetic makeup of a population over time is evolution.
 The AP Biology course is designed to enable you to develop advanced inquiry and reasoning skills, such as designing a plan for collecting data, analyzing data, applying mathematical routines, and connecting
The AP Biology course is designed to enable you to develop advanced inquiry and reasoning skills, such as designing a plan for collecting data, analyzing data, applying mathematical routines, and connecting
Supplementary Material for MTH 299 Online Edition
 Supplementary Material for MTH 299 Online Edition Abstract This document contains supplementary material, such as definitions, explanations, examples, etc., to complement that of the text, How to Think
Supplementary Material for MTH 299 Online Edition Abstract This document contains supplementary material, such as definitions, explanations, examples, etc., to complement that of the text, How to Think
Argumentation with Abduction
 Argumentation with Abduction Neophytos Demetriou Dept. of Computer Science University of Cyprus Nicosia CY-1678, Cyprus nkd@cs.ucy.ac.cy Tel.: (+357) 22892673 Antonis Kakas Dept. of Computer Science University
Argumentation with Abduction Neophytos Demetriou Dept. of Computer Science University of Cyprus Nicosia CY-1678, Cyprus nkd@cs.ucy.ac.cy Tel.: (+357) 22892673 Antonis Kakas Dept. of Computer Science University
CELL CYCLE AND DIFFERENTIATION
 CELL CYCLE AND DIFFERENTIATION Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id WHAT IS CELL CYCLE? 09/12/14 d.purnomosari@ugm.ac.id
CELL CYCLE AND DIFFERENTIATION Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id WHAT IS CELL CYCLE? 09/12/14 d.purnomosari@ugm.ac.id
A synthetic oscillatory network of transcriptional regulators
 A synthetic oscillatory network of transcriptional regulators Michael B. Elowitz & Stanislas Leibler, Nature, 403, 2000 igem Team Heidelberg 2008 Journal Club Andreas Kühne Introduction Networks of interacting
A synthetic oscillatory network of transcriptional regulators Michael B. Elowitz & Stanislas Leibler, Nature, 403, 2000 igem Team Heidelberg 2008 Journal Club Andreas Kühne Introduction Networks of interacting
AN INTRODUCTION TO SEPARATION LOGIC. 2. Assertions
 AN INTRODUCTION TO SEPARATION LOGIC 2. Assertions John C. Reynolds Carnegie Mellon University January 7, 2011 c 2011 John C. Reynolds Pure Assertions An assertion p is pure iff, for all stores s and all
AN INTRODUCTION TO SEPARATION LOGIC 2. Assertions John C. Reynolds Carnegie Mellon University January 7, 2011 c 2011 John C. Reynolds Pure Assertions An assertion p is pure iff, for all stores s and all
Analysing the microrna-17-92/myc/e2f/rb Compound Toggle Switch by Theorem Proving
 Analysing the microrna-17-92/myc/e2f/rb Compound Toggle Switch by Theorem Proving Giampaolo Bella and Pietro Liò @ Catania and Cambridge Under the auspices of British Council's grant ``Computer-assisted
Analysing the microrna-17-92/myc/e2f/rb Compound Toggle Switch by Theorem Proving Giampaolo Bella and Pietro Liò @ Catania and Cambridge Under the auspices of British Council's grant ``Computer-assisted
A systems approach to biology
 A systems approach to biology SB200 Lecture 5 30 September 2008 Jeremy Gunawardena jeremy@hms.harvard.edu Recap of Lecture 4 matrix exponential exp(a) = 1 + A + A2/2 +... + Ak/k! +... dx/dt = Ax matrices
A systems approach to biology SB200 Lecture 5 30 September 2008 Jeremy Gunawardena jeremy@hms.harvard.edu Recap of Lecture 4 matrix exponential exp(a) = 1 + A + A2/2 +... + Ak/k! +... dx/dt = Ax matrices
Lab 2 Worksheet. Problems. Problem 1: Geometry and Linear Equations
 Lab 2 Worksheet Problems Problem : Geometry and Linear Equations Linear algebra is, first and foremost, the study of systems of linear equations. You are going to encounter linear systems frequently in
Lab 2 Worksheet Problems Problem : Geometry and Linear Equations Linear algebra is, first and foremost, the study of systems of linear equations. You are going to encounter linear systems frequently in
chapter 12 MORE MATRIX ALGEBRA 12.1 Systems of Linear Equations GOALS
 chapter MORE MATRIX ALGEBRA GOALS In Chapter we studied matrix operations and the algebra of sets and logic. We also made note of the strong resemblance of matrix algebra to elementary algebra. The reader
chapter MORE MATRIX ALGEBRA GOALS In Chapter we studied matrix operations and the algebra of sets and logic. We also made note of the strong resemblance of matrix algebra to elementary algebra. The reader
Noisy Attractors and Ergodic Sets in Models. of Genetic Regulatory Networks
 Noisy Attractors and Ergodic Sets in Models of Genetic Regulatory Networks Andre S. Ribeiro Institute for Biocomplexity and Informatics, Univ. of Calgary, Canada Department of Physics and Astronomy, Univ.
Noisy Attractors and Ergodic Sets in Models of Genetic Regulatory Networks Andre S. Ribeiro Institute for Biocomplexity and Informatics, Univ. of Calgary, Canada Department of Physics and Astronomy, Univ.