Insight into Notch Signaling Steps that Involve pecanex from Dominant-Modifier
|
|
|
- Kelley Carson
- 5 years ago
- Views:
Transcription
1 Genetics: Early Online, published on May 31, 2018 as /genetics Insight into Notch Signaling Steps that Involve pecanex from Dominant-Modifier Screens in Drosophila Tomoko Yamakawa, * Yu Atsumi, Shiori Kubo, * Ami Yamagishi, * Izumi Morita, * and Kenji Matsuno * * Department of Biological Sciences, Graduate School of Science, Osaka University, 1-1 Machikaneyama, Toyonaka, Osaka , Japan Department of Biological Science and Technology, Graduate School of Industrial Science and Technology, Tokyo University of Science, Niijuku, Katsushika-ku, Tokyo , Japan 1 Copyright 2018.
2 Short running title: Genetic modifiers of pecanex KEYWORDS pecanex; Drosophila melanogaster; dominant modifier screen; Delta-Neuralized-Bearded pathway; Notch signaling Immediate Open Access: 1 Corresponding authors: Department of Biological Sciences, Graduate School of Science, Osaka University, 1-1 Machikaneyama, Toyonaka, Osaka , Japan. Tel: ; tyamakawa@bio.sci.osaka-u.ac.jp (Y.T.); Tel: ; kmatsuno@bio.sci.osaka-u.ac.jp (K.M.) 21 2
3 ABSTRACT Notch signaling plays crucial roles in intercellular communications. In Drosophila, the pecanex (pcx) gene, which encodes an evolutionarily conserved multi-pass transmembrane protein, appears to be required to activate Notch signaling in some contexts, especially during neuroblast segregation in the neuroectoderm. Although Pcx has been suggested to contribute to endoplasmic reticulum homeostasis, its functions remain unknown. Here, to elucidate these roles, we performed genetic modifier screens of pcx. We found that pcx heterozygotes lacking its maternal contribution exhibit cold-sensitive lethality, which is attributed to a reduction in Notch signaling at decreased temperatures. Using sets of deletions that uncover most of the second and third chromosomes, we identified four enhancers and two suppressors of the pcx cold-sensitive lethality. Among these, five genes encode known Notch-signaling components: big brain, Delta (Dl), neuralized (neur), Brother of Bearded A (BobA), a member of the Bearded (Brd) family, and N-ethylmaleimide-sensitive factor 2 (Nsf2). We showed that BobA suppresses Dl endocytosis during neuroblast segregation in the neuroectoderm, as Brd family genes reportedly do in the mesoderm for mesoectoderm specification. Analyses of Nsf2, a key regulator of vesicular fusion, suggested a novel role in neuroblast segregation, which is distinct from Nsf2 s previously reported role in imaginal tissues. Finally, jim lovell, which encodes a potential transcription factor, may play a role in Notch signaling during neuroblast segregation. These results reveal new research avenues for Pcx functions and Notch signaling. 41 3
4 Introduction Cell signaling via the Notch receptor is an evolutionarily conserved mechanism that mediates local cell-cell communications required for development and homeostasis in metazoans (Artavanis-Tsakonas et al. 1999; Cau and Blader 2009; Kopan and Ilagan 2009). Notch signaling plays essential roles in many biological events, including cell-fate decisions, pattern formation, and the regulation of cell physiology (Kim et al. 1996; Palmeirim et al. 1997; Bessho and Kageyama 2003; Le Bras et al Guruharsha et al. 2012). Therefore, defects in Notch signaling are associated with various human diseases, such as leukemia, cancers, cerebral infarction, Alagille syndrome, and multiple sclerosis (Ellisen et al. 1991; Joutel et al. 1997; Li et al. 1997; John et al. 2002; Nicolas et al. 2003). The molecular mechanisms of Notch signaling have been extensively studied, and its main process is well understood (Kopan and Ilagan, 2009; Guruharsha et al. 2012). However, various additional Notch-signaling components continue to be identified through a wide range of genetic and biochemical studies (Artavanis-Tsakonas et al. 1995, Bray 2016). In many cases, the specific roles of these components remain elusive, although they are required for Notch signaling activation or regulation at least in some contexts. Understanding the functions of these factors will help elucidate the detailed mechanisms of Notch signaling, and could lead to new therapeutic strategies for Notch pathway-related human diseases. The Notch receptor family consists of transmembrane proteins containing up to 36 epidermal growth factor-like repeats in their extracellular domain (Wharton et al. 1985). In the endoplasmic reticulum (ER) or Golgi, Notch undergoes post-translational modifications such as N-glycosylation, O-fucosylation, and O-glucosylation (Bruckner et al. 2000). The Notch extracellular domain is then cleaved by Furin (S1 cleavage), and the processed Notch fragments rebind to each other within the Golgi (Logeat et al. 1998; Kidd and Lieber 2002; Lake et al. 2009). At the cell surface, Notch binds to Delta- or Serrate-type ligands that are 4
5 presented on the surface of neighboring cells (Fehon et al. 1990; Rebay et al. 1991). The extracellular domain of Notch is mechanically pulled by the ligands; this pulling is achieved by ligand endocytosis, which requires ubiquitination of the ligands by Neuralized and Mindbomb (Wang and Struhl 2005; Meloty-Kapella et al. 2012). The pulling force induces a conformational change in the Notch extracellular domain, leading to its cleavage at the juxta-membrane region by Kuzbanian/ADAM10 (S2 cleavage) (Stephenson and Avis, 2012). Subsequently, the membrane-tethered Notch intracellular domain is cleaved within the transmembrane domain by γ-secretase (S3 cleavage), which releases the Notch intracellular domain from the membrane (Struhl et al. 1993; Mumm and Kopan 2000). The Notch intracellular domain is then translocated to the nucleus, where it functions as a co-activator of transcription, leading to the promoter activation of various target genes, such as Enhancer of split [E(spl)] in Drosophila (Bailey and Posakony 1995). In Drosophila, in addition to the core components of Notch signaling, several other genes have also been identified as components of the Notch signaling pathway, although the roles of their products remain to be clarified (Hori et al. 2013). One such gene is pecanex (pcx). Embryos homozygous or hemizygous for pcx lacking its maternal contribution (pcx mz ) exhibit neural hyperplasia, termed the neurogenic phenotype (Perrimon et al. 1984); this phenotype is considered evidence for the depletion of Notch signaling (Simpson, 1990). Thus, pcx is a maternal neurogenic gene. In addition, the expression of E(spl) m8 and single-minded, which are target genes of Notch signaling, is reduced in pcx mz embryos (Yamakawa et al. 2012). Therefore, pcx is essential for Notch signaling, at least in certain contexts, such as in neuroblast segregation, which occurs through lateral inhibition in the neuroectoderm. pcx encodes an ER-resident multi-pass transmembrane protein that consists of 3,433 amino acids and is highly conserved from Drosophila to humans (LaBonne et al. 1989). In addition to the neurogenic phenotype, ER enlargement is observed in a subset of cells in pcx mz embryos 5
6 (Yamakawa et al. 2012). Both the neurogenic phenotype and the ER defects of pcx mz embryos are rescued through ectopic induction of the unfolded protein response, which promotes ER functions such as protein folding (Kaufman 1999; Yamakawa et al. 2012). These findings suggest that Pcx is required for normal functions of the ER, which are essential for activating Notch signaling (Yamakawa et al. 2012). However, the biochemical functions of Pcx in Notch signaling remain unclear. We considered that a genetic modifier screen might be a useful approach for obtaining additional insights into the function of pcx. Genetic modifier screens have been used in various organisms including Drosophila to identify genes functioning in a common biochemical pathway (St Johnston 2002). Genetic modifiers identified by such screens can be informative for deducing the function of the affected gene. Such information might be particularly valuable in the case of pcx, given that Pcx does not carry any protein motifs that suggest its biochemical functions (Gilbert et al. 1992). Thus, in this study, we performed genetic screens to identify dominant modifiers of a cold-sensitive lethality that is associated with the pcx mutation condition. The dominant enhancers and suppressors of pcx identified from the screens provide insight into the steps of the Notch signaling cascade where pcx might be involved Materials and Methods Fly strains and genetics A standard Drosophila medium was used. The culture was maintained at 25 unless otherwise stated. Canton-S was used as the wild-type strain. The mutants used were: pcx 3, a hypomorphic allele (Mohler 1977; Mohler and Carroll 1984); big brain 1 (bib 1 ), a null allele (Lehmann et al. 1981); Delta 5F102 (Dl 5F102 ), a loss-of-function allele (Vӓssin and Campos-Ortega 1987); jim lovell 47 (lov 47 ), a hypomorphic allele lacking a 1.4-kb sequence 6
7 upstream of lov (a gift from Dr. Kathleen Beckingham; Bjorum et al. 2013); neuralized 1 (neur 1 ), a loss-of-function allele (Leviten and Posakony 1996); and N-ethylmaleimide-sensitive factor 2 A15 (Nsf2 A15 ), a hypomorphic allele (Mahony et al. 2006). Deficiency (Df) kits used for the initial screens were obtained from the Bloomington Drosophila Stock Center (BDSC) (Cook et al. 2012). Small-Df strains that uncovered smaller parts of the regions deleted in the original Dfs included in the Df kits were also obtained from BDSC. Furthermore, 30 additional small Dfs that uncovered the gap regions of the Df kits used for the initial screens were also provided by BDSC. Df(2R)gsb and Df(2R)GGG d13 were kindly provided by Dr. Markus Noll (He and Noll 2013) Determination of pupa and eclosed fly numbers under the cold-sensitive lethality of pcx/+ m To characterize the cold-sensitive lethality, which provides a useful assay for pcx function in our genetic screens and in our characterization of mutants identified from the screens, we scored the numbers of pupae and eclosed flies. Five pcx homozygous or wild-type (control) virgin females were mated with five wild-type males in plastic vials containing a standard Drosophila medium. The cultures were maintained at 18, 25, and 28 for eighteen days, and then the pupae and/or adult female progenies were counted. The male progenies obtained from pcx homozygous females (pcx/y m ) are known to be embryonic lethal (Yamakawa et al. 2012). The number of female wild-type pupae was deduced by assuming that half of the total pupae were female (Figure 1B). The eclosed females were identified by their female genitalia and counted (Figure 1C and Figure 5A-C). These experiments were performed in triplicate. For the temperature-shift analysis in Figure 1C and G, the parental flies were maintained at 28 until third-instar larvae appeared, and then the cultures were maintained at 18. In Figure 5A-C, to evaluate the dominant modifiers identified in the screens, genotypes of the female adults were determined by the absence of appropriate balancer chromosomes (CyO or TM3); 7
8 142 otherwise, the same procedures were used Measurement of egg numbers In Figure 1D, the number of eggs laid by wild-type and pcx homozygous females was counted. Five pcx homozygous or wild-type virgin females were mated with five wild-type males in a 50-ml tube containing a glass plate with grape agar and yeast paste. Eggs were collected for 48 hours, with four replacements of the glass plate, at 25 or Determination of female embryo percentage In Figure 1F, the percentage of female embryos was analyzed. Five pcx homozygous or wild-type virgin females were mated with five wild-type males in a 50-ml tube containing a glass plate with grape agar and yeast paste. Eggs were collected for 48 hours with four replacements of the glass plate, at 25 or 28. The vitellin membrane of the eggs was removed by heptane-methanol emulsion. Remaining vitellin membrane was removed from embryos with a tungsten needle after placing the embryos on a piece of double-sided tape covered with PBT (137 mm NaCl, 10 mm Na2HPO4, 2.7 mm KCl, 1.8 mm KH2PO4, 10% TritonX100, ph 7.4). The embryos were then stained with a female-specific anti-sexlethal (Sxl) antibody (Bopp et al. 1991), and the percentage of female embryos was determined. Experiments were performed in triplicate. Statistical significance was determined by the chi-square test Measurement of hatching rate In Figure 2A, the percentage of female eggs from which first-instar larvae successfully hatched was determined as the hatching rate. Five pcx homozygous or wild-type virgin females were mated with five wild-type males in a 50-ml tube containing a glass plate with grape agar and yeast paste. Eggs were collected for 48 hours with four replacements of the 8
9 glass plate, and then incubated for another three days, at 25 or 28. The number of female eggs was deduced by assuming that half of the total number of eggs were female. The number of wild-type female larvae was deduced using the same assumption. All of the first-instar larvae obtained from pcx homozygous females were female (pcx/+ m ) Measurement of the rate of survival to the third-instar larval stage In Figure 2B and Figure 5D-F, the rate of survival to the third-instar larval stage was determined. The rate of survival to the third-instar larval stage is the number of female third-instar larvae with the indicated genotype divided by the deduced total number of female embryos with that genotype ( 100). Five pcx homozygous virgin females were mated with five wild-type males in a 50-ml tube containing a glass plate with grape agar and yeast paste. Eggs were collected as described above and the total eggs were counted. The number of eggs with each genotype was estimated based on Mendel's laws and the sex ratio. All of the hatched larvae were transferred to the standard medium and cultured at 25 or 28. Thereafter, the sex of all larvae was determined by observing the gonad size at the third-instar larval stage. In addition, the presence of dominant modifiers in heterozygotes was determined by the absence of appropriate balancer chromosomes, judged by -Galactosidase (LacZ) staining (Kozlova et al., 1998). Experiments were performed in triplicate. The statistical significance of the survival rate to the third instar larval stage was examined by the chi-square test Measurement of pupation and eclosion rates To analyze the lethality of pcx/+ m females from the third-instar larval to pupal stages, we calculated the pupation rate, which is the number of pupating females divided by the number of collected third-instar larvae ( 100). The sex was determined at this stage based on the gonad size in Figure 2C. The pcx/+ m and wild-type larvae were collected as described 9
10 192 above To determine the lethality of pcx/+ m females during eclosion, we also calculated the eclosion rate, which was the number of eclosed females divided by the number of pupae ( 100) (Figure 2D). The sex of pupae was determined at the third-instar larval stage, as described above. Experiments were performed in triplicate. The statistical significance of the pupation and eclosion rates was examined by the chi-square test Dominant modifier screens based on the cold-sensitive phenotype of pcx/+ m Df kits of the 2nd and 3rd Drosophila chromosomes were used for the dominant modifier screens (Cook et al. 2012) (Table S1). Together, the Df kits cover approximately 98% of the genome in each chromosome (Cook et al. 2012). In addition, to fill in the gaps of the Df kits, we picked 30 available small Dfs located in the gaps of the primary Dfs (Table S2) and subjected them to the same screens. For the initial screens, five pcx homozygous virgin females were mated with five wild-type (control) or Df/Balancer males and cultured at 25 or 28. At 25, the Dfs were evaluated as positive when the subsequent generations without Balancers (pcx/+; Df/+ embryos lacking maternal pcx) eclosed (more than 10 adults), while the control progenies (pcx/+; +/+ lacking maternal pcx) were virtually embryonic lethal (no adults) under the same conditions. These deficiencies represented dominant suppressors of the pcx/+ m lethality, and potential genes encoding negative regulators of pcx might exist in the genomic regions uncovered therein. At 28, the Dfs were evaluated as positive if pcx/+; Df/+ flies lacking maternal pcx failed to eclose (no adults), while the control progenies (pcx/+; +/+ lacking maternal pcx) developed to adulthood (more than 10 adults). These deficiencies represented dominant enhancers of the lethality, and potential genes positively acting in conjunction with pcx might exist in the respective uncovered genomic regions. To identify the dominant modifiers as single loci, smaller deficiencies that uncovered 10
11 smaller parts of the genomic regions deleted in the Dfs identified in the first screens were subjected to the same analyses. After defining the minimal genomic regions that encompassed the loci responsible for the dominant modifications of the pcx lethal phenotypes, we searched for genes known to be involved in Notch signaling within these minimal regions. This mapping analysis using Dfs suggested that the Brother of bearded A (BobA) gene was a dominant suppressor; however, mutants affecting only the BobA locus were not available. Therefore, in this study, we generated a deletion mutant of BobA, BobA 1, as described below. For the lov gene, analyses using small Dfs defined the lov locus as a potential enhancer of pcx, and this possibility was confirmed using a preexisting lov mutant allele (Bjorum et al. 2013) Mutagenesis of the BobA locus A deletion mutant of BobA was generated using the FLP-FRT recombination method (Parks et al. 2004). Males carrying a P-element (PBac{RB}e00913) inserted in the 455-bp region upstream of Twin of m4 (Tom) on the third chromosome were crossed with females carrying a FLP recombinase transgene (Thibault et al. 2004). Male progeny carrying both the P-element and FLP recombinase transgene were then mated with females carrying another P-element (PBac{WH}BobA f03024 ) inserted in the 3 untranslated region of the BobA gene (Thibault et al. 2004) (see Figure S7). After 2 days, the parents and progeny were transferred into a 37 water bath and incubated for 1 hour. The cultures were then incubated at 25, and the parents were removed after 72 hours. Subsequently, the progenies were subjected to heat-shock treatments at 37 for 1 hour once a day for 4 days. The virgin females of the progeny were then collected and mated with males containing a balancer, TM6B, which carries the lacz gene, to establish stocks for further analysis Sequence analysis of the BobA 1 mutant allele 11
12 We established 14 potential lines of BobA mutants. Mutation of the BobA locus was confirmed by DNA sequence analysis. All polymerase chain reaction (PCR) assays were performed using purified genomic DNA from five embryos homozygous for each potential BobA mutant line. These homozygous embryos were selected based on the absence of X-gal staining (lacz-negative) using a standard protocol (Ashburner 1989). Genomic DNA was extracted using a standard protocol (Parks et al. 2004). PCR primers flanking the two P-elements were designed (Forward primer 1, 5 - ACA GGG AGC ACA CAA TAA CGA TC -3 ; Reverse primer 1, 5 - CAG AAG GGG TTT GAC CAG TTT C -3 ). PCR was carried out using TaKaRa Prime Script GXL and a rapid protocol according to the manufacturer s instructions: 10 sec at 98, 15 sec at 55, and 20 sec at 68 (40 cycles). We isolated three lines that carried a deletion uncovering the BobA genomic region, based on the size of the PCR product. Unincorporated primers, nucleotides, and salts were removed using a Wizard SV Gel and PCR Clean-Up System (Promega). Then, 4 μl of each eluted PCR product was added to 1 μl of Big Dye Terminator v3.1 sequencing cocktail (Applied Biosystems) containing the following sequencing primers (Reverse primer 1 and Forward primer 2, 5 - ACA GAA AAC TGC CGA ACC AA -3 ). After 25 cycles of PCR (10 sec at 96, 5 sec at 50, and 4 min at 60 ), the products were separated using an Applied Biosystems 3130xl sequencer. The results revealed that we obtained a BobA 1 allele in which the entire BobA gene and a non-coding RNA gene, CR45978, were deleted Immunohistochemistry Antibody staining of embryos was performed as previously described (Rhyu et al. 1994). The following primary antibodies were used: rat anti-elav [7E8A10, 1:500, NIH Developmental Studies Hybridoma Bank (DSHB)], mouse anti-sxl (M18, 1:10, DSHB), mouse anti-dl (C594.9B, 1:500, DSHB), guinea pig anti-dl (gp582, 1:100, a gift from Dr. 12
13 Artavanis-Tsakonas), chicken anti- -Galactosidase (ab9361, 1:500, Abcam), rabbit anti-rab11 (1:5000, a gift from Dr. Nakamura; Tanaka and Nakamura 2008), and rabbit anti-rhomboid (1:500, a gift from Dr. Bier; Sturtevant et al. 1996) antibodies. The following secondary antibodies were used: Cy3-conjugated donkey anti-mouse (1:500, Jackson ImmunoResearch), Cy3-conjugated donkey anti-guinea pig (1:500, Jackson ImmunoResearch), Alexa488-conjugated donkey anti-rabbit (1:500, Jackson ImmunoResearch), Alexa488-conjugated donkey anti-rat (1:500, Jackson ImmunoResearch), Cy5-conjugated donkey anti-chick (1:500, Jackson ImmunoResearch), and Cy5-conjugated goat anti-mouse (1:500, Jackson ImmunoResearch). Confocal microscopy images were captured using an LSM 700 (Zeiss) and analyzed on an LSM Image Browser, ZEN2010 (Zeiss) Data availability Strains are available upon request. File S1 contains all supplemental figures, tables and reagent table. Table S1 and S2 contains lists of all D. melanogaster strains used for this screen. All supplementary materials were uploaded to figshare (DOIs: /m9.figshare ) Results pcx exhibits a cold-sensitive lethal phenotype pcx is a maternal neurogenic gene, given that embryos homozygous or hemizygous for pcx without a maternal contribution of pcx (referred to as pcx mz in this study), exhibit the neurogenic phenotype, which is a benchmark implicating a deficiency in Notch signaling (Perrimon et al. 1984; LaBonne et al. 1989). Furthermore, the maternal neurogenic phenotype can be paternally rescued, as the introduction of a paternal pcx locus into pcx 13
14 heterozygotes lacking maternal pcx results in a much weaker neurogenic phenotype than that of pcx mz (pcx/y m in males) at 25 (optimal conditions for Drosophila) (LaBonne et al. 1989; Yamakawa et al. 2012) (Figure 1A). Here, we refer to the heterozygote for pcx lacking maternal pcx (but containing paternal pcx) as pcx/+ m. However, in a previous study, we discovered that pcx/+ m is lethal at 25 in a standard Drosophila medium (Yamakawa et al. 2012). To analyze the lethality of pcx/+ m further, in this study, we mated five pcx homozygous or wild-type virgin females with five wild-type males at 25, and maintained the cultures at 25 (Figure 1A). Wild type showed normal development to pupae and adults at this temperature (Figure 1B, C). However, only a small fraction of pcx/+ m individuals reached the pupal stage, and none reached adulthood, under the same condition (Figure 1B, C). Thus, the lethal phenotype associated with pcx mz was not paternally rescued at 25. For some mutants of genes, including those encoding Notch-signaling components, the associated phenotypes are sensitive to a change in temperature (Leonardi et al. 2011; Shimizu et al. 2014; Ishio et al. 2015). The heat- or cold-sensitive properties of such mutations may be useful for performing genetic analyses, including genetic modifier screens, because the gene activity can be controlled by temperature shifts to reveal the phenotype (O'Connell et al. 1998; Dellinger et al. 2000; Lindsay et al. 2008). Thus, in the current study, we examined the lethal phenotypes of pcx/+ m at different temperatures. As performed at 25, we mated five pcx homozygous or wild-type virgin females with five wild-type males at 18 and 28 (Figure 1A). The cultures were maintained at each temperature, and the average number of female pupae and adults obtained from each culture vial was counted (Figure 1B, C). We found that pcx/+ m developed to pupae and adults at 28 but not at 18, while wild type reached the adult stage at both temperatures (Figure. 1B, C). In particular, the numbers of pcx/+ m pupae and adults increased markedly at 28, compared with 25. This difference was not attributed to temperature-dependent changes in the number of eggs laid by females of parental crosses at 14
15 these two temperature, because we did not observe a marked difference in the numbers of eggs laid by wild-type and pcx homozygous females at 25 versus 28 (Figure 1D). pcx/y m is reported to exhibit a neurogenic phenotype and embryonic lethality at 25 (Yamakawa, et al. 2012). However, because we did not determine the sex in Figure 1B it was possible that a portion of the pcx/y m embryos became viable at 28 until the pupal stage, to explain the increase in pupae observed. Therefore, to confirm that the pcx/y m embryos were embryonic lethal at 28, we examined whether all of the male embryos cultured at this temperature showed a neurogenic phenotype. Male embryos (pcx/y m ) were sorted out based on the absence of anti-sexlethal (Sxl) antibody staining, and their neuronal cells were detected by anti-elav antibody staining (Figure 1E) (Bopp et al. 1991). The repetitive structure of the ladder-like nervous system was observed in wild-type embryos (Figure 1E, +/Y), while the pcx/y m embryos exhibited a severe neurogenic phenotype in all cases examined at 28 (Figure 1E, pcx/y m ). Thus, we concluded that pcx/y m males did not contribute to the observed change in the lethality, and that pcx/+ m exhibited a cold-sensitive lethal phenotype. Given that pcx/+ m exhibited a clear cold-sensitive lethal phenotype, a change in the sex ratio could also have influenced the number of pupae and adults in our experiments. However, we found that the females of parental crosses laid almost the same percentage of female eggs (+/+ and pcx/+ m, respectively) at 25 and 28 (Figure 1F). In addition, the percentage of pcx/+ m females in the total embryos (pcx/+ m plus pcx/y m ) laid by pcx homozygote mothers was almost the same at 25 and 28 (Figure 1F). Therefore, the sex ratio of the eggs laid by pcx homozygous females did not show a significant difference at these two temperatures. These results excluded the possibility that a change in the sex ratio contributed to the cold-sensitive lethality. To determine the developmental stage responsible for the cold-sensitive lethality, we next compared the survival rate of pcx/+ m and wild type between these two temperatures at 15
16 different developmental stages. First, we analyzed the hatching rates of wild-type and pcx/+ m embryos. To obtain the hatching rate, the number of female first-instar larvae (+/+ or pcx/+ m ) was divided by the number of total female embryos, which was deduced by assuming that half of the total embryos were female. For the wild-type control, the number of female first-instar larvae was also deduced from the sex ratio. We found that the hatching rate of pcx/+ m was significantly increased at 28 compared with 25 (Figure 2A). In contrast, the hatching rate of wild type was slightly reduced at 28 compared with 25, probably due to the nonoptimal temperature (Figure 2A). Therefore, we concluded that pcx/+ m shows cold-sensitive lethality during its embryonic stage. We also determined the rate of survival to the third-instar larval stage (Figure 2B). We calculated the percentage of deduced female wild-type and pcx/+ m embryos based on the sex ratio that reached the third-instar larval stage. The survival rate of pcx/+ m to the third-instar larval stage was significantly greater at 28 than at 25 (Figure 2B). Therefore, the cold-sensitive lethality of pcx/+ m could also be detected as the rate of survival to the third-instar larval stage, which was similar to the cold-sensitive hatching rate of the pcx/+ m embryos. Thus, the survival rate to the third-instar larval stage could be used to analyze the cold-sensitive lethality of pcx/+ m in combination with other mutants in the latter part of this study, which was useful because the sex and genotypes could be determined more easily at this stage, compared with those of first-instar larvae. We also examined whether other developmental stages, such as pupation and eclosion, were involved in the cold-sensitivity of pcx/+ m lethality. To determine the pupation (percentage of third-instar larvae that became pupae) and eclosion (percentage of pupae that successfully eclosed) rates, we counted the female larvae at the third instar (pcx/+ m and +/+ in each experiment) based on gonad size and cultured them until pupation and eclosion. However, the pupation rates of pcx/+ m and wild-type females were not markedly different at 16
17 versus 28 (Figure 2C). Therefore, once pcx/+ m embryos reached the third-instar larval stage, most of them developed to the pupal stage. We found that the eclosion rate of the pcx/+ m pupae was significantly greater at 28 than at 25 (Figure 2D). However, the eclosion rate of pcx/+ m versus wild-type was not significantly different at 28, probably because the eclosion rate of wild type was slightly lower at 28 than at 25 (Figure 2D). Thus, although the eclosion rate has some influence on the cold-sensitive lethality of pcx/+ m, it may not have a major effect. Collectively, these results showed that the cold-sensitive lethality of pcx/+ m was mostly associated with a reduction in the hatching rate. The pcx/+ m that were cultured continuously at 28 reached adulthood, and these adult flies did not exhibit detectable defects in their overall morphology (data not shown). Thus, to detect the potential requirement of pcx during the pupal development, we also performed a stage-specific temperature-shift experiment from 28 to 25 during the larval and pupal stages. Such a temperature shift could cause some developmental defects in adulthood, because pcx/+ m individuals showed a slightly reduced eclosion rate at 25 compared to 28, which suggested potential defects in their pupal development. We cultured pcx/+ m at 28 until the third-instar larval stage, then shifted the culture temperature to 18. However, we did not observe a marked reduction in the number of eclosed flies under this temperature-shift condition (Figure 1C). Furthermore, the adult flies that experienced the temperature shift did not exhibit detectable defects, in particular those suggesting a reduction in Notch signaling, such as wing notch, wing-vein thickening, or rough eye (Figure 1G) (Schellenbarger and Mohler 1978; de Celis and Garcia-Bellido 1994; Go et al. 1998). Thus, the cold-sensitive lethality of pcx/+ m did not appear to be directly attributable to a decline in Notch signaling after the third-instar larval stage. This idea is consistent with our observation that the cold-sensitive lethality of pcx/+ m was mostly observed as a reduction in the hatching rate (Figure 2A). 17
18 The cold-sensitive phenotypes of pcx is due to a reduction in Notch signaling at lower temperatures We next examined whether the cold-sensitive lethality of pcx/+ m was caused by a further reduction in Notch signaling at the lower temperature during embryogenesis. To test this possibility, we examined whether the cold-sensitive lethality of pcx/+ m embryos coincided with the extent of the neurogenic phenotype, detected by anti-elav antibody staining, even though the neurogenic phenotype of pcx/+ m embryos was less prominent owing to paternal rescue (weak neurogenic phenotype), compared with that of pcx mz embryos (strong neurogenic phenotype) at 25 (Figure 3A-D) (Yamakawa et al. 2012). Wild-type embryos exhibited a normal central and peripheral nervous system at stage 14 (100%, n = 47) (Figure 3A). As previously reported, a strong neurogenic phenotype, characterized by neural hyperplasia of the central and peripheral nervous systems, was observed in the pcx mz (pcx/y m ) embryos, in which the zygotic and maternal pcx function was absent (100%, n = 19) at 25 (Figure 3B) (Yamakawa et al., 2012). We also observed the paternal rescue of the neurogenic phenotype in pcx/+ m embryos, in which the strong neurogenic phenotype was suppressed in all cases examined at 18 (n = 47), 20 (n = 52), 22 (n = 102), 25 (n = 44), 28 (n = 51), and 30 (n = 34), and the strong neurogenic phenotype was not observed at all (Figure 3C-E). Among the paternally rescued embryos, we identified two classes of phenotypes in the embryonic nervous system: one exhibited a mostly normal nervous system as determined by its overview structure (quenched neurogenic phenotype) (Figure 3C, E), whereas the other still showed a weak neurogenic phenotype, especially in the central nervous system, although the neural hyperplasia of the peripheral nervous system was effectively suppressed (Figure 3D, E). We observed that the percentage of embryos classified as weak neurogenic increased, and the percentage of quenched neurogenic embryos decreased, as the culture temperature 18
19 was decreased (Figure 3E). The percentage of embryos classified as weak neurogenic was 80.9, 69.2, 68.6, 52.3, 31.4, and 14.7% at 18, 20, 22, 25, 28, and 30, respectively (Figure 3E). These results suggested that the Notch signaling activity in pcx/+ m embryos decreased with the decline in temperature. Therefore, we concluded that the cold-sensitive lethality could be attributed, at least in part, to a reduction in Notch signaling in pcx/+ m embryos Genetic screens to identify mutations that dominantly modify the cold-sensitive phenotype of pcx Using the cold-sensitive phenotype of pcx/+ m, we next conducted dominant modifier screens. First, we screened for dominant enhancers of the cold-sensitive lethality (Figure 4A). At 28, pcx/+ m animals survived until adulthood, whereas this genotype was virtually lethal before adulthood at 25 (Figure 1B, C). Therefore, we searched for second-site mutations that ameliorated the viability of pcx/+ m adults, thereby resulting in lethality prior to adulthood in a dominant manner at 28. Second, we screened for dominant suppressors of the lethality that was observed in pcx/+ m at 25 (Figure 1B). In this screen, we looked for second-site mutations that resulted in the emergence of pcx/+ m adult flies at this temperature. In both screens, we used Df kits that carried chromosomal deletions of the second and third chromosomes on a uniform genetic background (see Materials and Methods and Figure 4A). We screened 189 and 179 deficiency lines on the second and third chromosomes, respectively (Table S1). These screens covered approximately 98% of the Drosophila genome (Cook et al. 2012) Identification of dominant enhancers of pcx/+ m In our dominant enhancer screen, we identified seven deficiencies that enhanced the 19
20 cold-sensitive lethality of pcx/+ m in a dominant manner at 28 (Figure 4B). To identify the genetic loci responsible for the dominant enhancement, we narrowed down the genomic regions using smaller Dfs that uncovered smaller parts of the deleted regions in the seven positive Dfs identified in our first screen (Figure 4B and Figure S1-S6, S8). Df(2L)BSC50 and Df(2L)BSC689 were positive Dfs carrying deletions that partially overlapped (Figure 4B and Figure S1). Among the smaller Dfs uncovering the genomic regions deleted in Df(2L)BSC50 and Df(2L)BSC689, we found that Df(2L)BSC205 and Df(2L)BSC251, but not other Dfs, enhanced the pcx/+ m lethality (Figure 4B and Figure S1). The genomic deletions in Df(2L)BSC205 and Df(2L)BSC251 partially overlapped, and the bib locus exists in the overlapping region (Figure S1). bib is a classic neurogenic gene that encodes a component of the Notch signaling pathway (Lehmann et al. 1983; Doherty et al. 1997). The deduced product of bib has high homology to mammalian aquaporin-4, although a water channel activity of Bib was not detected in previous studies (Kaufmann et al. 2005; Park et al. 2008). In the current study, we found that bib 1, an existing allele of bib, dominantly enhanced the cold-sensitive lethality of pcx/+ m (Figure 5A, pcx/+ m ; bib 1 /+). However, the heterozygote for bib 1 showed normal development to adulthood at 28, demonstrating a synergistic interaction between pcx and bib (Figure 5A, +/+; bib 1 /+). From these results, bib was identified as a dominant enhancer of pcx. Using the same approach, we identified the genetic loci that were uncovered in each positive deficiency and responsible for dominant enhancement of the cold-sensitive lethality of pcx/+ m (Figure S2 S6, S8). Among the small deficiencies overlapping Df(3R)ED5938, we found that Df(3R)BSC850 and Df(3R)ED5942, which partially overlapped each other, enhanced the lethality of pcx/+ m (Figure 4B and Figure S2). Notably, the overlapping region contained the Dl locus (Figure S2). Dl encodes a ligand for the Notch receptor (Vässin et al. 1987; Kopczynski et al. 1988). We found that Dl 5F102, a preexisting allele of Dl, dominantly 20
21 enhanced the lethality of pcx/+ m (Figure 4B and Figure 5A, pcx/+ m ; Dl 5F102 /+). In contrast, heterozygotes for Dl 5F102 developed normally to adulthood at 28 (Figure 5A, +/+; Dl 5F102 /+). Thus, we identified Dl as another dominant enhancer of pcx. Another of the seven deficiency lines identified by our screens was Df(3R)ED5330, which contained the neur locus within its deleted genomic region (Figure 4B and Figure S3). We found that neur 1, a preexisting allele of neur, dominantly enhanced the lethality of pcx/+ m (pcx/+ m ; neur 1 /+), although neur 1 /+ flies developed normally to adulthood at 28 (Figure 4B and Figure 5A). Thus, neur was also identified as a dominant enhancer of pcx. Thus, our enhancer screen of the cold-sensitive lethality of pcx/+ m at 28 identified three genes, bib, Dl, and neur, all of which encode known components of the Notch signaling pathway, as dominant enhancers of pcx. On the other hand, in the genomic regions deleted in Df(2R)Kr10, which was also identified as dominant enhancer of pcx, we failed to find a gene that was known to be involved in Notch signaling (Figure 4B and Figure S4). However, it was possible that gene(s) encoding novel component(s) of the Notch signaling pathway exist in these regions, given that three genes involved in Notch signaling were successfully identified by our enhancer screen as described above. Therefore, using small Dfs, we attempted to narrow down the genomic region containing the dominant enhancer to single loci. The genomic deletion in Df(2R)Kr10 partially overlapped with two small Dfs, Df(2R)GGG d13 and Df(2R)gsb (Figure 4B and Figure S4). We found that Df(2R)gsb, which overlapped with the lov locus, enhanced the cold-sensitive lethality of pcx/+ m at 28, but Df(2R)GGG d13, which did not overlap with the lov locus, did not (Figure S4). These results collectively suggested that lov may be responsible for the dominant enhancement of lethality (Figure S4). We found that an available lov allele, lov 47, dominantly enhanced the lethality of pcx/+ m at 28 (Figure 5B, pcx/+ m ; lov 47 /+). However, lov 47 /+ flies developed normally to adulthood at 28 (Figure 5B, +/+; lov 47 /+). Thus, we concluded that lov is also a dominant enhancer of pcx, and 21
22 demonstrated for the first time that lov may be involved in the Notch signaling pathway. Finally, we also identified two partially overlapping Dfs, Df(2R)X58-12 and Df(2R)X58-8, from the dominant enhancer screen (Figure 4B and Figure S5). In this case, we failed to identify a preexisting mutation(s) that enhanced the lethality of pcx/+ m in the overlapping region of these two Dfs, although we excluded CG3927, Vacuolar protein sorting 20 (Vps20), bonsai, and Ribosomal protein 624 (Rp624) from the candidates for the modifier gene (Figure S5). Therefore, these Dfs were not analyzed further in this study. We found that the cold-sensitive lethality of pcx/+ m could be easily assessed as a reduction in the rate of survival to the third-instar larval stage (Figure 2B). Thus, we evaluated the effect of these dominant enhancers on the viability of pcx/+ m based on the survival rate at this stage. We found that all four enhancers, bib, Dl, neur, and lov, significantly reduced the survival rates in a dominant manner, when each was combined with pcx/+ m at 28 (Figure 5D, E). However, bib 1 /+ and neur 1 /+ did not show significantly different survival rates from wild type at 28 (Figure 5D). In contrast, the survival rates of Dl 5F102 /+ and lov 47 /+ were reduced to about half that of wild type (Figure 5D, E). However, the survival rates observed in these heterozygous embryos were not as low as those of embryos with these enhancers combined with pcx/+ m (Figure 5D, E). Therefore, we concluded that bib, Dl, neur, and lov dominantly enhanced the lethality of pcx/+ m from the third-instar larval stage to the embryonic stage Identification of dominant suppressors of pcx/+ m At 25, pcx/+ m failed to develop to adulthood. We next screened for Dfs that suppressed the cold-sensitive lethality of pcx/+ m in a dominant manner at 25 (Figure 4A and Table S1), and identified Df(3L)ED217. The genomic region deleted in Df(3L)ED217 was partially uncovered in six small deficiencies, Df(3L)BSC441, Df(3L)Exe16125, Df(3L)Exe16126, 22
23 Df(3L)BSC837, Df(3L)ED218, and Df(3L)Exe16262 (Figure 4B and Figure S6). We found that among these six small Dfs, only Df(3L)BSC441 dominantly suppressed the lethality of pcx/+ m at 25 (Figure 4B, Figure S6). Five genes, CG17839, CG34245, DCX-EMAP, CG13465, and BobA existed in the genomic region encompassed by Df(3L)BSC441, which contained the locus responsible for the dominant suppression. Among these, DCX-EMAP was the only gene with an available mutant (Bechstedt et al. 2010). However, DCX-EMAP did not suppress the lethality, which excluded this gene as the dominant suppressor. To identify the dominant suppressor as a single locus, we next generated a BobA mutant allele, because BobA is a member of the Bearded (Brd) family, which is known to negatively regulate Notch signaling (Lai et al. 2000). Brd family members bind to and inhibit Neur, an E3 ubiquitin ligase that ubiquitinates Dl (Bardin and Schweisguth 2006). The ubiquitination of Dl is required for Dl s endocytosis, which facilitates the signaling activity of Dl (Lai et al. 2001; Le Borgne et al. 2005). However, the specific functions of BobA have remained unknown, because a mutation affecting only the BobA locus has not been available. In addition, the previous study was based on deletion mutants uncovering multiple Brd family genes, including BobA (Bardin and Schweisguth 2006). Thus, in the current study, we generated a mutant allele of BobA, designated as BobA 1, using the FLP-FRT recombination method (Figure S7). Thus, BobA 1 is the first mutant in which a single Brd family gene was disrupted. We found that BobA 1 dominantly suppressed the lethality of pcx/+ m at 25 (Figure 4B and Figure 5C, pcx/+ m ; BobA 1 /+). In addition, a gene encoding a non-coding RNA, CR45978, was also deleted in BobA 1 (see Figure S7) (Young et al. 2012). However, it is unlikely that CR45978 is the dominant suppressor of pcx/+ m lethality, because CR45978 expression has only been detected in adult males (Young et al. 2012). Thus, our results suggested that BobA is a dominant suppressor of pcx. Notably, the Df kits used in our screens have small gaps in the coverage of deleted 23
24 genomic regions, corresponding to approximately 2% of the chromosomes (Cook et al. 2012). Thus, the genes located within these gaps had escaped from our screens. To minimize this occurrence, we examined an additional 30 preexisting small Dfs that uncovered the gap regions on the second and third chromosomes (Table S2). Among these, Df(3R)ED5662 dominantly suppressed the cold-sensitive lethality of pcx/+ m at 25 (Figure 4B and Table S2). We found that this Df uncovered the N-ethylmaleimide-sensitive factor 2 (Nsf2) gene, which was previously identified as a modifier of Notch signaling (Figure S8) (Stewart et al. 2001). We here showed that a preexisting allele of Nsf2, Nsf2 A15 dominantly suppressed the cold-sensitive lethality of pcx/+ m at 25 (Figure 4B and Figure 5C, pcx/+ m ; Nsf2 A15 /+). This result identified Nsf2 as a dominant suppressor of pcx. Based on the above findings, we speculated that the dominant suppression of the cold-sensitive lethality of pcx/+ m might be due to an increase in the survival rate to the third-instar larval stage. We found that BobA 1 and Nsf2 A15 significantly increased this survival rate in a dominant manner when combined with pcx/+ m at 25 (Figure 5F, pcx/+ m ; BobA 1 /+ and pcx/+ m ; Nsf2 A15 /+, respectively). BobA 1 /+ by itself showed a slightly decreased survival rate (Figure 5F, +/+; BobA 1 /+). However, we might be able to ignore this effect, because the number of eclosed BobA 1 /+ flies was comparable to that of wild type at 25 (Figure 5C). These results suggested that BobA 1 and Nsf2 A15 dominantly suppressed the lethality of pcx/+ m from the embryonic to the third-instar larval stage pcx modifiers influence Notch signaling activity Our genetic screens identified four dominant enhancers and two dominant suppressors of the cold-sensitive lethality of pcx/+ m. As described above, in pcx/+ m, we observed a correlation between the extent of cold-sensitive lethality and the neurogenic phenotype, which is an indicator of reduced Notch signaling activity (Simpson 1990). In addition, we found that the 24
25 dominant enhancers and suppressors reduced and increased the survival rate by the third-instar larval stage, respectively, suggesting that these modifiers affect embryonic development, such as the neuroblast segregation controlled by Notch signaling. Therefore, we next examined whether these enhancers and suppressors also modified the neurogenic phenotype of pcx/+ m. At 28, 31.4% of the pcx/+ m embryos showed a weak neurogenic phenotype, whereas the rest of the embryos exhibited a largely normal nervous system (quenched neurogenic phenotype), based on the criteria described in Figure 3. However, in combination with pcx/+ m, all of the dominant enhancers identified by our screen; i.e., bib, neur, Dl, and lov, increased the percentage of embryos showing a weak neurogenic phenotype to 75.9, 78.5, 81.8, and 76.3%, respectively, in a dominant manner at 28 (Figure 6A). We also found that bib 1 /+, Dl 5F102 /+, neur 1 /+, and lov 47 /+ exhibited a normal nervous system (normal) upon anti-elav antibody staining at 28, suggesting that a synergistic genetic interaction, rather than a simple additive effect, occurred between pcx and each mutant (Figure 6A). Under the same conditions, an arbitrarily selected deficiency, Df(2R)DE2457, did not markedly affect the frequency of the pcx/+ m phenotypes (Figure 6A). These results suggested that the identified dominant enhancers further reduced the Notch signaling in pcx/+ m embryos. We next examined whether the dominant suppressors identified by our screen suppressed the neurogenic phenotype of pcx/+ m. At 25, 46.7% of the pcx/+ m embryos showed a weak neurogenic phenotype, while the rest showed a quenched neurogenic phenotype (n = 15) (Figure 6B). However, the two dominant suppressors, BobA 1 (6.1%, n = 33) and Nsf2 A15 (0%, n = 21), almost completely suppressed the neurogenic phenotype of pcx/+ m in a dominant manner at 25, at which almost all of these embryos exhibited a quenched neurogenic phenotype (Figure 6B). Conversely, under the same conditions, an arbitrarily selected deficiency, Df(2R)DE2457, did not markedly affect the frequency of the weak neurogenic 25
26 phenotype (64.7%, n = 34) (Figure 6B). These results suggested that mutations of the BobA and Nsf2 genes augmented the Notch signaling activity in pcx/+ m embryos. We also found that BobA 1 /+ and Nsf2 A15 /+ exhibited a normal nervous system (normal) upon anti-elav antibody staining at 25, demonstrating that BobA 1 and Nsf2 A15 showed a synergistic genetic interaction with pcx (Figure 6B) BobA may have a similar function as other Brd family genes in neuroblast segregation as well as in mesoderm specification The endocytosis of Dl shows dorsal-ventral polarity in the cellular blastoderm at stage 5 (Bardin and Schweisguth 2006). Dl endocytosis is active in the ventral part (mesoderm) of embryos, depends on Neur, and is required for the spatially controlled activation of Notch signaling in the mesectoderm (Bardin and Schweisguth 2006). It was previously shown that neur activity is suppressed in the dorsal region of embryos by Brd family genes, so Dl trafficking is upregulated in this region in mutants of Brd family genes (Bardin and Schweisguth 2006; De Renzis et al. 2006). Thus, the Brd family proteins suppress Neur activity, leading to reduced Dl trafficking in the early embryo. In these experiments, however, multiple Brd family genes were simultaneously removed by a deletion mutation. In the current study, we found that the BobA 1 mutant, in which only the BobA gene was deleted among the Brd family genes, suppressed the neurogenic phenotype (6.1%, n = 33) associated with pcx/+ m, compared with the wild-type BobA gene (46.7%, n = 15) (Figure 6B). In addition, although the role of the Dl-Neur-Brd pathway in specification of the mesoectoderm has been extensively analyzed (Bardin and Schweisguth, 2006), and it was proposed that the inhibition of Neur by Brd regulates the epithelial polarity (Chanet and Schweisguth 2012), its role in neuroblast segregation in the neuroectoderm has not been studied. Therefore, in this study, it was important to determine whether Dl trafficking was also upregulated in the BobA 1 26
27 mutant embryos, especially in the neuroectoderm. The distribution of Dl was analyzed by anti-dl antibody staining. As previously shown, Dl was detected in recycling endosomes, which were specifically labeled by anti-rab11 antibody staining, in the ventral but not the dorsal side of wild-type embryos at stage 5 in all cases examined (n=5) (Figure 7) (Wang and Struhl 2004; Emery et al. 2005). However, Dl was ectopically detected in Rab11-positive vesicles in the dorsal side of BobA 1 homozygous embryos at stage 5 in all cases examined (n = 8) (Figure 7). This observation was consistent with a previous finding that Brd family genes suppress the neur-dependent endocytosis of Dl in the dorsal but not the ventral side (Bardin and Schweisguth 2006). Importantly, we detected the D1-containing ectopic vesicles in the neuroectoderm of BobA 1 mutant embryos (n=7), which was visualized by specific staining with an anti-rhomboid antibody (left side of each panel in Figure 8; boundaries of Rhomboid-expressing and non-expressing regions are indicated by a white broken line) (Sturtevant et al. 1996). In addition, notably, we found that BobA 1 homozygous embryos showed an anti-neurogenic phenotype at 41.2% (n=17) frequency at 28, as revealed by a reduction in neuronal cells detected by anti-elav antibody staining, whereas wild-type embryos did not show this phenotype at this temperature in any case examined (n=27). Thus, our results suggest that the Dl-Neur-Brd pathway also regulates Dl endocytosis in the neuroectoderm during neuroblast segregation. On the other hand, we did not detect a notable difference in the number of Dl-containing vesicles in the ventral region of these embryos (Figure 7 and right sided of each panel in Figure 8), probably because the Brd genes are not expressed in the ventral region of wild-type embryos (Chanet and Schweisguth 2012). Based on these results, BobA appears to have a similar activity to that of other Brd family proteins. This idea is consistent with the previous finding that the majority of Brd family proteins, including BobA, bind to Neur (Bardin and Schweisguth 2006). Conversely, our 27
28 results also showed that the function of Brd family genes is not completely redundant regarding the suppression of Dl endocytosis and the ectopic activation of Notch signaling in the neuroectoderm. We predicted that if pcx is also directly involved in the Neur-mediated endocytosis of Dl, the number of Dl-positive vesicles should decrease in the mesoderm of pcx m/z embryos, as previously found in embryos homozygous for neur or overexpressing a Brd family gene (Bardin and Schweisguth 2006). However, we did not observe any abnormality in Dl-positive vesicles in the pcx m/z embryos in any case examined (n = 5) (Figure 7). We also examined whether there was a synergistic effect between pcx and BobA on the Dl trafficking by combining pcx/+ m and BobA/+, in each of which the D1-containing vesicles did not show any detectable abnormality. However, we did not observe any alteration in the Dl-positive vesicles in pcx/+ m ; BobA/+ (Figure 9). Therefore, at this point, the molecular mechanisms underlying the genetic connection between the Dl-Neur-Brd pathway and Pcx remain unknown Discussion Genes encoding known and novel components of Notch signaling were identified as dominant modifiers of pcx Our dominant modifier screens of pcx identified four enhancers and two suppressors (Figure 4B). Among these, four genes encode well-characterized components of the Notch signaling pathway, suggesting that our procedure efficiently screened for Notch-signaling components. Although our primary screens were based on the enhancement and suppression of the cold-sensitive lethality demonstrated in pcx/+ m embryos, such modifications of this lethality coincided well with the enhancement and suppression of the neurogenic phenotype, implying that the lethality is related to a depletion of Notch signaling. Thus, the primary cause of the cold-sensitive lethality was probably a further reduction of Notch signaling with the decrease 28
29 in temperature, and the modifications of the lethal phenotype reflected up- or down-regulations of Notch signaling in the heterozygous combination with each modifier. We predicted that the phenotypes of pcx/+ m could have resulted from an alteration in the initial timing of the zygotic expression of pcx. However, in our screen, mutations that might have generally changed the onset of zygotic gene expression in the early embryo, such as chromatin remodeling factors, were not identified. Conversely, most of the identified genes were found to encode factors that directly contribute to the Notch signaling cascade. The highly specific genetic interactions between pcx and these genes support the idea that Pcx is a critical component of Notch signaling, at least during the period of neuroblast segregation in the neuroectoderm. This idea is also consistent with our previous finding that the expression of Notch signaling target genes, including Enhancer of split and single-minded, is severely reduced in pcx mz embryos (Yamakawa et al. 2012) The Dl-Neur-BobA pathway functions not only in mesoectoderm specification but also in neuroblast segregation Our screens identified the Dl, neur, and BobA genes, which have been shown to function in a common biochemical process that regulates the dorsal-ventral pattern of Dl endocytosis, as dominant modifiers of pcx (Bardin and Schweisguth 2006; De Renzis et al. 2006). The activity of Dl as a ligand for Notch requires the endocytosis of Dl, which depends on the ubiquitination of Dl s cytoplasmic domain by Neur, a RING domain-containing E3 ubiquitin ligase (Lai et al. 2001; Pavlopoulos et al. 2001). During early embryogenesis around stage 5, the endocytosis of Dl is particularly active in the mesoderm located in the most ventral part of the embryo, although Dl endocytosis is much less active in the neuroectoderm that is dorsally adjacent to the mesoderm. Therefore, the high Dl activity in the mesoderm induces the Notch signaling-dependent transcription of sim in single rows of cells next to the mesoderm 29
30 (mesectoderm) (Morel and Schweisguth 2000). In the absence of the Brd complex, in which Oche, Tom, BobA, BobB, and BobC are clustered, the active endocytosis of Dl is observed in the entire embryo, indicating that the Brd family genes normally inhibit Neur activity in the dorsal region of the embryo (Bardin and Schweisguth 2006). Under the Brd-deleted condition, the sim expression is expanded (Bardin and Schweisguth 2006). To regulate sim expression, Brd proteins bind to Neur and interfere with the physical interaction between Neur and Dl (Lai et al. 2001; Bardin and Schweisguth 2006). However, unlike this mechanism of mesoectoderm specification, the contribution of the Dl-Neur-BobA pathway to the regulation of Notch signaling during neuroblast segregation in the neuroectoderm has not been well studied. In the current study, we found that the neurogenic phenotype associated with the pcx mutation was dominantly enhanced by neur and Dl and suppressed by BobA. Consistent with these observations in pcx/+ m ; BobA 1 /+ embryos, Dl-containing vesicles were increased in the neuroectoderm of BobA 1 homozygous embryos, as well as in their dorsal side (Figure 8A). Moreover, we found that BobA homozygotes exhibited an anti-neurogenic phenotype at 28, implying that Notch signaling was augmented during neuroblast segregation in these embryos (Figure 8B). Thus, our results suggest that the Dl-Neur-BobA pathway also functions in the regulation of Notch signaling during neuroblast segregation in the neuroectoderm. Although the Dl endocytosis was hyperactivated in the neuroectoderm of BobA 1 mutant embryos, we did not observe an attenuation of Dl endocytosis even in the complete absence of pcx function (pcx m/z embryo) (Figure 7). In addition, we did not observe synergistic effects between pcx and BobA on the Dl endocytosis (Figure 8). Therefore, at this point, it is unclear whether Pcx has a direct role in the Dl-Neur-BobA pathway. We are currently investigating the potential connections between Pcx and Dl endocytosis
31 lov transcribed in the early embryo may contribute to the activation of Notch signaling during neuroblast segregation In this study, we identified the lov gene as a potential component of the Notch signaling pathway for the first time. lov encodes a Bric-a-Brac/Tramtrack/Broad/Pox virus and Zinc finger (BTB/POZ) domain-containing transcription factor and is expressed in various neurons of the central and peripheral nervous system and in the trachea during Drosophila development (Albagli et al. 1995; Armstrong et al. 2006). A loss-of-function mutant of lov demonstrates multiple behavioral defects in the larval and adult stages, including gravity sensing (Bjorum et al. 2013). These behavioral defects were previously attributed to the abnormal specifications of neurons, given that an absence of lov expression in a subset of neurons in mutants of a particular lov allele was correlated with the behavioral defects (Bjorum et al. 2013). The expression of lov was also observed in specific cells of the central nervous system (CNS) midline in embryos (Wheeler et al. 2008). Notably, Notch signaling regulates gene expression in these CNS midline cells, which defines the fates of these cells (Wheeler et al. 2008). However, a detailed analysis of lov expression revealed that the lov gene is not a target of Notch signaling, at least in the ventral unpaired median neurons of the CNS midline (Wheeler et al. 2008). Although it is still possible that lov is involved in Notch signaling activation as a component of the pathway, the contribution of lov to Notch signaling appears to be highly context-dependent, redundant, or subtle, because the defects in the cell-fate specification of neurons found in lov mutants are much milder than those of classic Notch pathway genes (Bjorum et al. 2013). It is reported that the expression of lov begins in the cellular blastoderm stage and continues during the development of the CNS and peripheral nervous system (Weiszmann et al. 2009; Kearney et al. 2004). Furthermore, the Lov protein is detected in the neuroectoderm during the stage of neuroblast segregation, as well as in other parts of embryos (Bjorum et al. 31
32 ). Therefore, lov may control the transcription of genes involved in Notch signaling during the periods of neuroblast segregation through lateral inhibition, although this possibility remains to be investigated. In any case, we speculate that lov s function is not essential for Notch signaling during neuroblast segregation, given that lov homozygous embryos did not exhibit a neurogenic phenotype (data now shown). In addition, a maternal expression of lov was not observed, suggesting that lov does not have a maternal function (Coté et al. 1987). Therefore, it is unlikely that lov plays an indispensable role in lateral inhibition in the embryo, although our results suggest that it makes some contribution to it A novel role of Nsf2 supports the contribution of Pcx to ER homeostasis In this study, we identified Nsf2 as a dominant suppressor of pcx. Drosophila has two Nsf genes, comatose (Nsf1) and Nsf2 (Pallanck et al. 1995a). The Nsf genes encode homohexameric AAA ATPases that are involved in the membrane fusion required for the transfer of membrane vesicles from one membrane compartment to another (Wilson et al. 1989; Söllner et al. 1993a). Nsf1 is mainly expressed in neurons (Ordway et al. 1994), and its loss causes a disruption in synaptic transmission (Pallanck et al. 1995b; Gokhale et al. 2015). To ensure the specificity of membrane fusion, SNARE proteins on vesicles and target membranes form a tight complex (Söllner et al. 1993a). In this process, Nsf promotes the dissociation of SNARE complexes once membrane fusion has been completed, using ATP hydrolysis as the energy source (Sollner et al. 1993a). The dissociation of SNAREs is required for their recycling for further rounds of membrane fusion events (Söllner et al. 1993b). Thus, the absence of Nsf results in an attenuation of membrane trafficking (Beckers et al. 1989). These studies suggested that Nsf1 functions to disassemble SNARE complexes after vesicle docking, which is required to maintain the readily releasable pool of synaptic vesicles (Muller et al. 2002; Gokhale et al. 2015). 32
33 On the other hand, Nsf2 is broadly expressed in embryonic and imaginal tissues, as well as in the nervous system (Boulianne and Trimble 1995). Therefore, Nsf2 is likely to encode an Nsf isoform that contributes to vesicular trafficking in non-neuronal cells, including the neuroectoderm of the embryo (Boulianne and Trimble 1995). It was previously shown that the expression of a dominant-negative Nsf2 (Nsf2 E/Q ) suppresses Notch signaling in wing imaginal discs (Stewart et al. 2001). Genetic interaction between Nsf2 E/Q and mutants of other genes encoding Notch-signaling components, including Dl, Serrate, fringe, and bib, has also been reported (Stewart et al. 2001). Notably, bib was also identified as a dominant enhancer of the pcx/+ m lethality in the present study (Figure 4B, Figure S1). In addition, it was shown that Notch protein accumulates in the intracellular vesicles in epithelial cells misexpressing Nsf2 E/Q (Stewart et al. 2001). The contribution of Nsf2 to Notch signaling was further supported by the identification of Nsf2 as a dominant enhancer of Presenilin hypomorphic mutants (Mahoney et al. 2006). These results all suggested that Nsf2 positively contributes to Notch signaling (Mahoney et al. 2006). In contrast, in our screen, Nsf2 was identified as a suppressor of pcx, demonstrating that the wild-type Nsf2 gene suppresses Notch signaling activity under the conditions of our experiment. Thus, we speculate that Nsf2 may have different roles in Notch signaling between imaginal tissues and embryonic neuroectoderm, which has not been noted before. This idea is consistent with the finding that various sets of SNAREs control the membrane fusion of different intracellular compartments under the control of an Nsf protein (Söllner et al. 1993). Furthermore, we previously reported that the ER is enlarged in a subset of cells in pcx mz embryos (Yamakawa et al. 2012). In mammalian and yeast cells, Nsf is involved in ER membrane fusion and required for ER reassembly (Uchiyama et al. 2002; Kano et al. 2005). Thus, given that the ER enlargement in the absence of pcx function may be due to excessive assembly of the ER, a loss-of-function mutation of Nsf2 may reduce the excess ER 33
34 reassembly occurring in the absence of pcx. This scenario is consistent with Nsf2 s behavior as a dominant suppressor of pcx/+ m lethality. This mechanism is currently under active investigation in our laboratory. The genetic screens performed in the current study identified dominant modifiers of the cold-sensitive lethality associated with a pcx mutation condition. In particular, although the genetic interactions of pcx with neur or BobA may not reflect a direct relationship with respect to the regulation of Dl endocytosis, the results obtained from our genetic screens revealed intimate relationships between Pcx and the Notch signaling pathway and implied a close relationship between pcx function and vesicular transport during neuroblast segregation in the neuroectoderm. Furthermore, our analysis uncovered a potential role of lov and a previously unknown function of Nsf2 in Notch signaling. There results contribute to our understanding of the complex regulation of Notch signaling
35 Figure Legends Figure 1 pcx exhibits cold-sensitive lethality. (A) Mating scheme to analyze the cold-sensitive lethality of pcx/+ m. Five pcx homozygous females were mated with wild-type males. All females of the F1 progeny were pcx heterozygotes lacking maternal pcx (pcx/+ m ) and exhibited cold-sensitive lethality. All males of the F1 progeny were pcx hemizygotes lacking maternal pcx (pcx/y m ) and exhibited embryonic lethality. (B) Numbers of wild-type female (+/+, 50% of pupae were predicted to be female) and pcx/+ m (F1 female) pupae cultured at 18, 25, or 28. Standard deviations are indicated. (C) Numbers of wild-type female (+/+) and pcx/+ m (F1 female) adults obtained from cultures at 18, 25, or 28 or cultures with a temperature shift from 28 to 18. In the temperature-shift experiment, the cultures were maintained at 28 until third-instar larvae appeared, and then they were maintained at 18 for eighteen days in total. Standard deviations are indicated. (D) Numbers of wild-type female (+/+) and pcx/+ m (F1 female) embryos at 25 and 28. Sex of embryos was determined by anti-sxl antibody staining. Standard deviations are indicated. (E) Lateral views of wild-type (+/Y) and pcx mz (pcx/y m ) embryos cultured at 28 and immunostained with a neuron-specific antibody (anti-elav antibody) at stage These embryos were identified as males based on the absence of anti-sxl antibody staining. (F) Phenotypes of wild-type (wt) and pcx/+ m adults cultured with the temperature shift from 28 to 18. From top to bottom, adult females, wings, eyes, and thoraxes. (G) Percentage of total embryos cultured at 25 or 28 that were female embryos (+/+ and pcx/y m ). The number of total embryos examined is shown as (N) at the top. * and ** indicate p>0.01 and p<0.01, respectively Figure 2 Survival rate of pcx/+ m is decreased at lower temperature. (A) Hatching rate of wild-type (+/+) and pcx/+ m females by the first-instar larval stage cultured at 25 or
36 Percentages of total female embryos that became hatched female first-instar larvae are shown. For the calculation, half of the total embryos and hatched first-instar larvae were assumed to be female. The number of female embryos examined is shown as (N) at the top. (B) Survival rate of wild-type (+/+) and pcx/+ m females to the third-instar larval stage, cultured at 25 or 28. Percentages of total female embryos that became surviving female third-instar larvae are shown. For the calculation, half of the total embryos were assumed to be female. The number of female embryos examined is shown as (N) at the top. (C) Pupation rate of wild-type (+/+) and pcx/+ m females cultured at 25 or 28. The percentages of female third instar larvae that became pupae are shown. The number of female larvae examined is shown as (N) at the top. (D) Eclosion rate of wild type (+/+) females and pcx/+ m cultured at 25 or 28. Percentages of pupae (sex was determined at the third-instar larval stage) that successfully eclosed as adults are shown. The number of female pupae examined is shown as (N) at the top. In A and B, we rounded the deduced N down. All experiments were performed in triplicate. * and ** indicate p>0.01 and p<0.01, respectively Figure 3 pcx/+ m exhibits a cold-sensitive neurogenic phenotype. (A-D) Lateral views of embryos cultured at 25 and immunostained with a neuron-specific antibody (anti-elav antibody) at stage (A) Wild type (+/+). (B) pcx mz (pcx/y m ) embryo showing a strong neurogenic phenotype (100%, n=19) ( Strong ). (C) pcx/+ m embryo showing a mostly normal nervous system structure (quenched neurogenic phenotype) (47.7%, n=44) ( Quenched ). (D) pcx/+ m embryo showing a weak neurogenic phenotype (52.3%, n=44) ( Weak ). In A-D, scale bars are 50 m. (E) Percentage of pcx/+ m embryos showing a mostly normal nervous system (quenched, green) and a weak neurogenic phenotype (weak, yellow) at the indicated temperatures. Number of embryos examined in each experiment (N) is indicated at the top. 36
37 Figure 4 Genetic screen procedure and summary of the dominant modifiers of pcx. (A) Five pcx homozygous females were mated with five males of each Deficiency (Df) mutant on the second or third chromosome at 25 or 28. The genotype of F1 females without balancers was identified as pcx/+ m ; Df/+. After eighteen days, the numbers of F1 progeny were counted. pcx/+ m individuals survived until adulthood at 28. In the enhancer screen, the Df lines that showed reduced viability at 28 (i.e., those in which F1 did not survive until adulthood) were identified. pcx/+ m individuals were lethal by the pupal stage at 25. In the suppressor screen, Df lines that suppressed the lethality at 25 (i.e., those in which F1 survived to adulthood) were identified. (B) Tables summarizing the dominant enhancers [28 lethal (Enhancer)] and suppressors [25 viable (Suppressor)] of pcx/+ m. Df lines (Symbol No.) identified as dominant modifiers of pcx/+ m in the first screens and the deleted genomic region (Deleted region) in each Df line are listed in the left columns (Df). The small Df lines (Symbol No.) uncovering smaller parts of the original Dfs and containing the potential loci of the modifiers are listed in the middle columns (Small Df) with the deleted genomic regions (Deleted region). Dominant modifiers identified as single genes are indicated in the right columns (Tested allele) with information about the alleles (Allele name), molecular functions of the gene products (Molecular function), and potential associated cellular processes (Cellular process). - indicates inapplicable Figure 5 Dominant modification of the cold-sensitive lethality of pcx/+ m by pcx modifiers identified as single loci. (A) The numbers of female adult flies of wild type (+/+), pcx/+ m, heterozygotes of each enhancer (bib 1, Dl 5F102, or neur 1 ), and pcx/+ m combined with bib 1 /+, Dl 5F102 /+, or neur 1 /+ at 28 are shown. Genotypes were determined by sex and marker phenotypes associated with balancer chromosomes. (B) Numbers of female adult flies of wild 37
38 type (+/+), pcx/+ m, heterozygote of lov 47, and pcx/+ m combined with lov 47 /+ at 28 are shown. Genotypes were determined by sex and marker phenotypes associated with balancer chromosomes. (C) Numbers of female adult flies of wild type (+/+), pcx/+ m, heterozygotes of each suppressor (BobA 1 and Nsf2 A15 ), or pcx/+ m combined with BobA 1 /+ and Nsf2 A15 /+ at 25. Genotypes were determined by sex and marker phenotypes associated with balancer chromosomes. (D) Survival rate of wild-type (+/+) females, pcx/+ m, and heterozygous females of each enhancer (bib 1, Dl 5F102, or neur 1 ), and pcx/+ m combined with bib 1 /+, Dl 5F102 /+, or neur 1 /+ to the third-instar larval stage at 28. Percentages of surviving female third-instar larvae with the indicated genotype in the deduced total number of female embryos with that genotype are shown. (E) Survival rate of wild-type (+/+) females, pcx/+ m, heterozygous females of lov 47, and pcx/+ m combined with lov 47 /+, to the third-instar larval stage at 28. Percentages of surviving female third-instar larvae with the indicated genotype in the deduced total number of female embryos with that genotype are shown. (F) Survival rate of wild-type (+/+) females, pcx/+ m, heterozygous females of each suppressor (BobA 1 and Nsf2 A15 ), or pcx/+ m combined with BobA 1 /+ and Nsf2 A15 /+, to the third-instar larval stage at 25. Percentages of surviving female third-instar larvae with the indicated genotype in the deduced total number of female embryos with that genotype are shown. In D-F, genotypes were determined by sex and marker phenotypes associated with balancer chromosomes at the third-instar larval stage. In D-F, for the calculation, the numbers of embryos with the indicated genotypes were predicted based on Mendel's laws, and half of the embryos were assumed to be female. In D-F, the number of female embryos examined is shown as (N) at the top of each graph. We rounded the deduced N down. All experiments were performed in triplicate. In A-C, standard deviations are indicated. In D-F, * and ** indicate p>0.01 and p<0.01, respectively
39 Figure 6 Dominant modification of the neurogenic phenotype of pcx/+ m by pcx modifiers. (A) Percentages of embryos showing a normal nervous system (normal, blue bars), quenched neurogenic phenotype (quenched, green bars), and weak neurogenic phenotype (weak, yellow bars) at 28. Embryos of pcx/+ m, pcx/+ m combined with Df(2R)ED2475/+ (an arbitrarily selected deletion as a control), bib 1 /+, Dl 5F102 /+, lov 47 /+, or neur 1 /+, and heterozygotes of each enhancer (bib 1 /+, Dl 5F102 /+, lov 47 /+, or neur 1 /+) were analyzed. Embryos heterozygous for all the enhancers were scored as normal; none of them showed a dominant neurogenic phenotype at 28. (B) Percentages of embryos showing normal nervous system (normal, blue bars), quenched neurogenic phenotype (quenched, green bars), and weak neurogenic phenotype (weak, yellow bars) at 25. Embryos of pcx/+ m, pcx/+ m combined with Df(2R)ED2475/+ (an arbitrarily selected deletion as a control), BobA 1 /+, or Nsf2 A15 /+, and heterozygotes of each suppressor (BobA 1 /+ or Nsf2 A15 /+) were analyzed. Embryos heterozygous for each suppressor were scored as normal; none of them showed a dominant neurogenic or anti-neurogenic phenotype at 25. The number of embryos examined in each experiment (N) is indicated at the top of the graph Figure 7 BobA suppresses Dl endocytosis in the dorsal region of embryos. Subcellular localization of Dl (magenta, left and right panels) and Rab11 (green, middle and right panels), a marker of recycling endosomes, in the dorsal (upper panels) and ventral (lower panels) epithelium of wild-type (wt), BobA 1 /BobA 1 (BobA), and pcx mz embryos at stage 5. Right panels are merged images of the left and middle panels. White arrowheads indicate vesicles where Dl and Rab11 are colocalized. Percentage of embryos showing the represented phenotypes is indicated at the upper right. The number of embryos examined is in parentheses. Scale bar is 5 m
40 Figure 8 Dl recycling and Notch signaling are hyperactivated in the neuroectoderm of the BobA mutant. (A) Subcellular localization of Dl (magenta, left and right panels) and Rhomboid (Rho) (green, middle and right panels), a neuroectoderm marker, in wild-type (wt) or BobA 1 /BobA 1 (BobA) embryos at stage 5. Boundary between the neuroectoderm and mesoderm is indicated by a white broken line, and the regions left (Rho-positive) and right (Rho-negative) of the broken lines are neuroectoderm and mesoderm, respectively. The percentage of embryos showing the represented phenotype is indicated at the upper right. The number of embryos examined is in parentheses. Scale bar is 5 m. (B) Lateral views of wild-type (wt) and BobA 1 /BobA 1 (BobA) embryos cultured at 28 and immunostained with a neuron-specific (anti-elav) antibody at stage Percentage of embryos showing the represented phenotype in each panel is shown at the bottom right, and the number of embryos examined is in parentheses. Scale bar is 50 m Figure 9 No synergistic interaction between pcx/+ m and BobA/+ in the Dl-trafficking 944 phenotype. Subcellular localization of Dl (magenta, left and right panels) and Rab11 (green, 945 middle and right panels), a marker of recycling endosomes, in the dorsal (upper panels) and 946 ventral (lower panels) epithelium of pcx/+ m, BobA/+, and pcx/+ m ; BobA/+ embryos at stage Right panels are merged images of the left and middle panels. White arrowheads indicate 948 vesicles where Dl and Rab11 are colocalized. Percentage of embryos showing the represented 949 phenotype is indicated at the upper right. The number of embryos examined is in parentheses. 950 Scale bar is 5 m
41 952 Table S1 Results of the primary screens to identify dominant modifiers of the cold-sensitive 953 lethality associated with pcx/+ m. The tested Df lines (stock No. and Symbol No.) used in the 954 first screens are listed in the left columns with the respective deleted genomic regions 955 (Deleted region). The results of mating between each Df line and pcx homozygous virgin 956 females cultured at 25 or 28 are indicated in the right columns (25 and 28 ). CyO/+ and 957 TM3/+ represent F1 progenies that carried the balancers but not Dfs. Df/+ represents F1 958 progenies that carried Dfs. - indicates a category that did not yield any F1 progeny indicates a category that yielded more than ten F1 progenies. If fewer than ten F1 progenies 960 were obtained, the number of F1 progenies is shown. At 25, Dfs were evaluated as positive 961 when more than ten F1 progenies without balancers (pcx/+; Df/+ embryos lacking maternal 962 pcx) eclosed (+), whereas those with balancers (pcx/+; balancers /+ lacking maternal pcx) did 963 not eclose (-), given that the control progeny (pcx/+; +/+ lacking maternal pcx) were lethal at 964 this temperature. At 28, Dfs were evaluated as positive if the F1 progeny without balancers 965 failed to eclose (-), while those with balancers developed to adulthood (+), given that the 966 control progeny developed to adulthood at this temperature. All positive Df lines are shown 967 in yellow Table S2 Results of screens involving small deficiencies that uncovered the gaps found in the 970 Df kits. The Df kits used in the screens have small gaps in the coverage of deleted genomic 41
42 971 regions, corresponding to approximately 2% of the chromosomes (Cook et al. 2012). To fill 972 in the gaps of the Df kits, 30 available small Dfs that were selected and screened are shown. 973 All categories in the list are the same as in Table S Figure S1 Identification of bib as a dominant enhancer of pcx. Schematic of the genomic 976 region encompassing the bib locus (magenta rectangle) and the deletions used to map bib as 977 an enhancer of pcx. The genomic region that was collectively shown to include the enhancer 978 of pcx is shown in pink with boundaries indicated by red broken lines Figure S2 Identification of Dl as a dominant enhancer of pcx. Schematic of the genomic 981 region encompassing the Dl locus (magenta rectangle) and deletions used to map Dl as an 982 enhancer of pcx. The genomic region that was collectively shown to include the enhancer of 983 pcx is shown in pink with boundaries indicated by red broken lines Figure S3 Identification of neur as a dominant enhancer of pcx. Schematic of the genomic 986 region encompassing neur locus (magenta rectangle) and deletions used to map neur as an 987 enhancer of pcx. The genomic region that was collectively shown to include the enhancer of 988 pcx is shown in pink with boundaries indicated by red broken lines
43 990 Figure S4 Identification of lov as a dominant enhancer of pcx. Schematic of the genomic 991 region encompassing the lov locus (magenta rectangle) and deletions used to map lov as an 992 enhancer of pcx. The genomic region that was collectively shown to include the enhancer of 993 pcx is shown in pink with boundaries indicated by red broken lines Figure S5 Potential region that includes a dominant enhancer of pcx, although we failed to 996 map it as a single locus. Schematic diagram of the genomic region encompassing a potential 997 enhancer of pcx and the deletions used to map it. The genomic region that was collectively 998 shown to include the enhancer of pcx is shown in pink with boundaries indicated by red 999 broken lines. Genes that were genetically excluded from the candidates for the enhancer are 1000 indicated by red Xs Figure S6 Identification of BobA as a dominant suppressor of pcx. Schematic of the genomic 1003 region encompassing the BobA locus (magenta rectangle) and the deletions used to map BobA 1004 as a suppressor of pcx. The genomic region that was collectively shown to include the 1005 suppressor of pcx is shown in pink with boundaries indicated by red broken lines. The 1006 DCX-EMAP gene, which was genetically excluded from the candidates for the enhancer is 1007 indicated by a red X
44 1009 Figure S7 Schematic of the procedure used to generate a deletion mutant of BobA, BobA Two P-element insertions, PBac{WH}BobA f03024 and pbac{rb}e00913, were combined in 1011 cis by FLP-mediated recombination. In the BobA 1 allele, a 23-kb genomic fragment 1012 (indicated by the double-headed arrow) was deleted Figure S8 Identification of Nsf2 as a dominant suppressor of pcx. Schematic of the genomic 1015 region encompassing the Nsf2 locus (magenta rectangle) and the deletion used to map Nsf2 as 1016 a suppressor of pcx. The genomic region that was collectively shown to include the 1017 suppressor of pcx is shown in pink with boundaries indicated by red broken lines
45 Acknowledgements We thank Dr. Kathleen Beckingham and Dr. Markus Noll for providing fly stocks. We thank Dr. Akira Nakamura, Dr. Ethan Bier and Dr. Spyros Artavanis-Tsakonas for providing antibodies. We also thank the KYOTO Stock Center (Kyoto), the Bloomington Drosophila Stock Center (Indiana), and the Developmental Studies Hybridoma Bank (University of Iowa). This work was supported by JSPS KAKENHI Grant Number JP and by a Japan Prize Foundation 2014 Grant Summary Notch signaling plays essential roles in many biological processes in development and homeostasis through intercellular communications. Here, we performed genetic screens in Drosophila to identify genes that could modify the neurogenic phenotype and embryonic lethality associated with a maternal deficit of pecanex, which encodes a Notch-signaling component. The screens identified dominant enhancers and suppressors of pecanex, including potential novel components of Notch signaling, and analyses of these molecules uncovered new regulatory mechanisms of Notch signaling. Identifying the functions of these factors will improve our understanding of Notch signaling, and may lead to new therapeutic strategies for Notch-pathway-related diseases References 1039 Albagli, O., P. Dhordain, C. Deweindt, G. Lecocq, and D. Leprince, 1995 The BTB/POZ 1040 domain: a new protein-protein interaction motif common to DNA- and actin-binding 1041 proteins. Cell Growth Differ. 6: Armstrong, J. D., M. J. Texada, R. Munjaal, D. A. Baker, and K. M. Beckingham,
46 1043 Gravitaxis in Drosophila melanogaster: a forward genetic screen. Genes Brain Behav. 5: Artavanis-Tsakonas S., K. Matsuno, and M. E. Fortini, 1995 Notch signaling. Science 268: : Artavanis-Tsakonas S., M. D. Rand, and R. J. Lake, 1999 Notch signaling: cell fate control 1048 and signal integration in development. Science 284: Ashburner M, 1989 Drosophila: A Laboratory Manual. Cold Spring Harbor Laboratory Press, 1050 Cold Spring Harbor Bailey A. M., and J. W. Posakony, 1995 Suppressor of hairless directly activates transcription 1052 of enhancer of split complex genes in response to Notch receptor activity. Genes Dev. 9: Bardin A. J., and F. Schweisguth, 2006 Bearded family members inhibit Neuralized-mediated 1055 endocytosis and signaling activity of Delta in Drosophila. Dev. Cell 10: Bechstedt S., J. T. Albert, D. P. Kreil, T. Müller-Reichert, M. C. Göpfert et al., 2010 A 1057 doublecortin containing microtubule-associated protein is implicated in 1058 mechanotransduction in Drosophila sensory cilia. Nat. Commun. 1: Beckers C. J., M. R. Block, B. S. Glick, J. E. Rothman, and W. E. Balch, 1989 Vesicular 1060 transport between the endoplasmic reticulum and the Golgi stack requires the 1061 NEM-sensitive fusion protein. Nature 339:
47 1062 Bessho Y., and R. Kageyama, 2003 Oscillations, clocks and segmentation. Curr. Opin. Genet Dev. 13: Bjorum S. M., R. A. Simonette, R. Alanis Jr., J. E. Wang, B. M. Lewis et al., 2013 The 1065 Drosophila BTB domain protein jim lovell has roles in multiple larval and adult behaviors PLoS One 8: e Bopp D., L. R. Bell, T. W. Cline, and P. Schedl, 1991 Developmental distribution of 1068 female-specific sex-lethal proteins in 1069 Drosophila melanogaster. Genes & Dev. 5: Boulianne G. L., and W. S. Trimble, 1995 Identification of a second homolog of 1071 N-ethylmaleimide-sensitive fusion protein that is expressed in the nervous system and 1072 secretory tissues of Drosophila. Proc. Natl. Acad. Sci. U.S.A. 92: Bray S. J., 2016 Notch signalling in context. Nat. Rev. Mol. Cell Biol. 17: Bruckner K., L. Perez, H. Clausen, and S. Cohen, 2000 Glycosyltransferase activity of Fringe 1075 modulates Notch-Delta interactions. Nature 406: Cau E., and P. Blader, 2009 Notch activity in the nervous system: to switch or not switch? 1077 Neural Dev. 4: Chanet S., F. Schweisguth, 2012 Regulation of epithelial polarity by the E3 ubiquitin ligase 1079 Neuralized and the Bearded inhibitors in Drosophila. Nat. Cell Biol. 14: Cook R. K., S. J. Christensen, J. A. Deal, R. A. Coburn, M. E. Deal et al., 2012 The 47
48 1081 generation of chromosomal deletions to provide extensive coverage and subdivision of 1082 the Drosophila melanogaster genome. Genome Biol. 3: R Coté S., A. Preiss, J. Haller, R. Schuh, A. Kienlin et al., 1987 The gooseberry-zipper region of 1084 Drosophila: five genes encode different spatially restricted transcripts in the embryo EMBO J. 6: De Celis J. F. and A. García-Bellido, 1994 Roles of the Notch gene in Drosophila wing 1087 morphogenesis. Mech. Dev. 46: De Renzis S., J. Yu, R. Zinzen, and E. Wieschaus, 2006 Dorsal-ventral pattern of Delta 1089 trafficking is established by a Snail-Tom-Neuralized pathway. Dev. Cell 10: Dellinger B., R. Felling, and R. W. Ordway, 2000 Genetic modifiers of the Drosophila NSF 1091 mutant, comatose, include a temperature-sensitive paralytic allele of the calcium channel 1092 alpha1-subunit gene, cacophony. Genetics 155: Doherty D., L. Y. Jan, and Y. N. Jan, 1997 The Drosophila neurogenic gene big brain, which 1094 encodes a membrane-associated protein, acts cell autonomously and can act 1095 synergistically with Notch and Delta. Development 124: Ellisen L. W., J. Bird, D. C. West, A. L. Soreng, T. C. Reynolds et al., 1991 TAN-1, the 1097 human homolog of the Drosophila notch gene, is broken by chromosomal translocations 1098 in T lymphoblastic neoplasms. Cell 66: Emery G., A. Hutterer, D. Berdnik, B. Mayer, F. Wirtz-Peitz, M. G. Gaitan and J. A. Knoblich, 48
49 Asymmetric Rab 11 endosomes regulate delta recycling and specify cell fate in the 1101 Drosophila nervous system. Cell 122: Fehon R. G., P. J. Kooh, I. Rebay, C. L. Regan, T. Xu et al., 1990 Molecular interactions 1103 between the protein products of the neurogenic loci Notch and Delta, two 1104 EGF-homologous genes in Drosophila. Cell 61: Gilbert T. L., B. A. Haldeman, E. Mulvihill, and P. J. O'Hara, 1992 A mammalian homologue 1106 of a transcript from the Drosophila pecanex locus. J. Neurogenet. 8: Go M. J., D. S. Eastman, S. Artavanis-Tsakonas, 1998 Cell proliferation control by Notch 1108 signaling in Drosophila development. Development 125: Gokhale A., A. P. Mullin, S. A. Zlatic, C. A. Easley 4th, M. E. Merritt, et al., 2015 The 1110 N-ethylmaleimide-sensitive factor and dysbindin interact to modulate synaptic plasticity J. Neurosci. 35: Guruharsha K. G., M. W. Kankel, and S. Artavanis-Tsakonas, 2012 The Notch signalling 1113 system: recent insights into the complexity of a conserved pathway. Nat. Rev. Genet. 13: He H., and M. Noll, 2013 Differential and redundant functions of gooseberry and gooseberry 1116 neuro in the central nervous system and segmentation of the Drosophila embryo. Dev Biol. 382: Hori K., A. Sen, S. Artavanis-Tsakonas, 2013 Notch signaling at a glance. J. Cell Sci. 126: 49
50 Ishio A., T. Sasamura, T. Ayukawa, J. Kuroda, H. O. Ishikawa et al., 2015 O-fucose 1121 monosaccharide of Drosophila Notch has a temperature-sensitive function and cooperates 1122 with O-glucose glycan in Notch transport and Notch signaling activation. J. Biol. Chem : John G. R., S. L. Shankar, B. Shafit-Zagardo, A. Massimi, S. C. Lee et al., 2002 Multiple 1125 sclerosis: re-expression of a developmental pathway that restricts oligodendrocyte 1126 maturation. Nat. Med. 8: Joutel A., K. Vahedi, C. Corpechot, A. Troesch, H. Chabriat et al., 1997 Strong clustering and 1128 stereotyped nature of Notch3 mutations in CADASIL patients. Lancet 350: Kano F., H. Kondo, A. Yamamoto, Y. Kaneko, K. Uchiyama et al., 2005 NSF/SNAPs and 1130 p97/p47/vcip135 are sequentially required for cell cycle-dependent reformation of the 1131 ER network. Genes Cells 10: Kaufman R. J., 1999 Stress signaling from the lumen of the endoplasmic reticulum: 1133 coordination of gene transcriptional and translational controls. Genes Dev. 13: Kaufmann N., J. C. Mathai, W. G. Hill, J. A. Dow, M. L. Zeidel et al., 2005 Developmental 1135 expression and biophysical characterization of a Drosophila melanogaster aquaporin. Am J. Physiol. Cell Physiol. 289: C397-C Kearney J. B., S. R. Wheeler, P. Estes, B. Parente, and S. T. Crews, 2004 Gene expression 50
51 1138 profiling of the developing Drosophila CNS midline cells. Dev. Biol. 275: Kidd S., and T. Lieber, 2002 Furin cleavage is not a requirement for Drosophila Notch 1140 function. Mech. Dev. 115: Kim J., A. Sebring, J. J. Esch, M. E. Kraus, K. Vorwerk et al., 1996 Integration of positional 1142 signals and regulation of wing formation and identity by Drosophila vestigial gene Nature 382: Kooh P. J., R. G. Fehon, and M. A. Muskavitch, 1993 Implications of dynamic patterns of 1145 Delta and Notch expression for cellular interactions during Drosophila development Development 117: Kopan R. and M. X. Ilagan, 2009 The canonical Notch signaling pathway: unfolding the 1148 activation mechanism. Cell 137: Kopczynski C. C., A. K. Alton, K. Fechtel, P. J. Kooh and M. A. Muskavitch, 1988 Delta, a 1150 Drosophila neurogenic gene, is transcriptionally complex and encodes a protein related to 1151 blood coagulation factors and epidermal growth factor of vertebrates. Genes Dev. 2: Kozlova T., G. V. Pokholkova, G. Tzertzinis, J. D. Sutherland, I. F. Zhimulev and F. C Kafatos, 1998 Drosophila hormone receptor 38 functions in metamorphosis: a role in 1155 adult cuticle formation. Genetics. 149: LaBonne S. G., I. Sunitha, A. P. Mahowald, 1989 Molecular genetics of pecanex, a 51
52 1157 maternal-effect neurogenic locus of Drosophila melanogaster that potentially encodes a 1158 large transmembrane protein. Dev. Biol. 136: Lai E. C., R. Bodner, J. Kavaler, G. Freschi, and J. W. Posakony, 2000 Antagonism of Notch 1160 signaling activity by members of a novel protein family encoded by the Bearded and 1161 Enhancer of split gene complexes. Development 127: Lai E. C., G. A. Deblandre, C. Kintner, and G. M. Rubin, 2001 Drosophila neuralized is a 1163 ubiquitin ligase that promotes the internalization and degradation of delta. Dev. Cell 1: Lake R. J., L. M. Grimm, A. Veraksa, A. Banos, and S. Artavanis-Tsakonas In vivo 1166 analysis of the Notch receptor S1 cleavage. PLoS One 4: e Le Borgne R., S. Remaud, S. Hamel, and F. Schweisguth, 2005 Two distinct E3 ubiquitin 1168 ligases have complementary functions in the regulation of delta and serrate signaling in 1169 Drosophila. PLoS Biol. 3: e Le Bras S., N. Loyer and R. Le Borgne, 2011 The multiple facets of ubiquitination in the 1171 regulation of Notch signaling pathway. Traffic 12: Lehmann R., U. Dietrich, F. Jimenez, and J. A. Campos-Ortega, 1981 Mutations of early 1173 neurogenesis in Drosophila. Roux Arch. Dev. Biol. 190: Lehmann R., F. Jimenez, U. Dietrich, and J. A. Campos-Ortega, 1983 On the phenotype and 1175 development of mutants of early neurogenesis in Drosophila melanogaster. Roux Arch. 52
53 1176 Dev. Biol. 192: Leonardi J., R. Fernandez-Valdivia, Y. D. Li, A. A. Simcox, and H. Jafar-Nejad, Multiple O-glucosylation sites on Notch function as a buffer against 1179 temperature-dependent loss of signaling. Development 138: Leviten M. W., and J. W. Posakony, 1996 Gain-of-function alleles of Bearded interfere with 1181 alternative cell fate decisions in Drosophila adult sensory organ development. Dev. Biol : Li L., I. D. Krantz, Y. Deng, A. Genin, A. B. Banta et al., 1997 Alagille syndrome is caused 1184 by mutations in human Jagged1, which encodes a ligand for Notch1. Nat. Genet. 16: Lindsay H. A., R. Baines, R. ffrench-constant, K. Lilley, H. T. Jacobs et al., 2008 The 1187 dominant cold-sensitive Out-cold mutants of Drosophila melanogaster have novel 1188 missense mutations in the voltage-gated sodium channel gene paralytic. Genetics 180: Logeat F., C. Bessia, C. Brou, O. LeBail, S. Jarriault et al., 1998 The Notch1 receptor is 1191 cleaved constitutively by a furin-like convertase. Proc. Natl. Acad. Sci. U. S. A. 95: Mahoney M. B., A. L. Parks, D. A. Ruddy, S. Y. Tiong, H. Esengil et al., Presenilin-based genetic screens in Drosophila melanogaster identify novel Notch 53
54 1195 pathway modifiers. Genetics 172: Meloty-Kapella L., B. Shergill, J. Kuon, E. Botvinick, and G. Weinmaster, 2012 Notch ligand 1197 endocytosis generates mechanical pulling force dependent on dynamin, epsins, and actin Dev. Cell 22: Mohler J. D., 1977 Developmental genetics of the Drosophila egg. I. Identification of sex-linked cistrons with maternal effects on embryonic development. Genetics 85: Mohler D., and A. Carroll, 1984 Report of new mutants. Dros. Inf. Serv. 60: Morel V., and F. Schweisguth, 2000 Repression by suppressor of hairless and activation by 1204 Notch are required to define a single row of single-minded expressing cells in the 1205 Drosophila embryo. Genes Dev. 14: Morel V., R. Le Borgne, and F. Schweisguth, 2003 Snail is required for Delta endocytosis and 1207 Notch-dependent activation of single-minded expression. Dev. Genes Evol. 213: Muller J. M., J. Shorter, R. Newman, K. Deinhardt, Y. Sagiv et al., 2002 Sequential SNARE 1209 disassembly and GATE-16-GOS-28 complex assembly mediated by distinct NSF 1210 activities drives Golgi membrane fusion. J. Cell Biol. 157: Mumm J. S., and R. Kopan, 2000 Notch signaling: from the outside in. Dev. Biol. 228: Nagiec E. E., A. Bernstein, and S. W. Whiteheart, 1995 Each domain of the 54
55 1214 N-ethylmaleimide-sensitive fusion protein contributes to its transport activity. J. Biol Chem. 270: Nicolas M., A. Wolfer, K. Raj, J. A. Kummer, P. Mill et al., 2003 Notch1 functions as a tumor 1217 suppressor in mouse skin. Nat. Genet. 33: O'Connell K. F., C. M. Leys, and J. G. White, 1998 A genetic screen for temperature-sensitive 1219 cell-division mutants of Caenorhabditis elegans. Genetics 149: Ordway R., L. Pallanck, and B. Ganetzky, 1994 Neurally expressed Drosophila genes 1221 encoding homologs of the NSF and SNAP secretory proteins. Proc. Natl. Acad. Sci. U. S A. 91: Pallanck L., R. W. Ordway, M. Ramaswami, W. Y. Chi, K. S. Krishnan et al., 1995a Distinct 1224 roles for N-ethylmaleimide-sensitive fusion protein (NSF) suggested by the identification 1225 of a second Drosophila NSF homolog. J. Biol. Chem. 270: Pallanck L., R. W. Ordway, and B. Ganetzky, 1995b A Drosophila NSF mutant. Nature 376: Palmeirim I., D. Henrique, D. Ish-Horowicz, and O. Pourquié, 1997 Avian hairy gene 1229 expression identifies a molecular clock linked to vertebrate segmentation and 1230 somitogenesis. Cell 91: Park J. S., S. R. Kim, S. Y. Park, D. J. Yang, S. H. Lee et al., 2008 Big brain, a Drosophila 1232 homologue of mammalian aquaporin, is regulated by the DRE/DREF system. Biochim. 55
56 1233 Biophys. Acta 1779: Parks A. L., K. R. Cook, M. Belvin, N. A. Dompe, R. Fawcett et al., 2004 Systematic 1235 generation of high-resolution deletion coverage of the Drosophila melanogaster genome Nat. Genet. 36: Pavlopoulos E., C. Pitsouli, K. M. Klueg, M. A. Muskavitch, N. K. Moschonas et al., neuralized encodes a peripheral membrane protein involved in delta signaling and 1239 endocytosis. Dev. Cell 1: Perrimon N., L. Engstrom, and A. P. Mahowald, 1984 Developmental genetics of the 2E-F 1241 region of the Drosophila X chromosome: a region rich in developmentally important 1242 genes. Genetics 108: Rebay I., R. J. Fleming, R. G. Fehon, L. Cherbas, P. Cherbas et al., 1991 Specific EGF 1244 repeats of Notch mediate interactions with Delta and Serrate: implications for Notch as a 1245 multifunctional receptor. Cell 67: Rhyu M. S., L. Y. Jan, and Y. N. Jan, 1994 Asymmetric distribution of numb protein during 1247 division of the sensory organ precursor cell confers distinct fates to daughter cells. Cell : Roote J and S. Russell, 2012 Toward a complete Drosophila deficiency kit. Genome Biol. 13: Shellenbarger D. L. and J. D. Mohler, 1978 Temperature-sensitive periods and autonomy of 56
57 1252 pleiotropic effects of l(1)nts1, a conditional notch lethal in Drosophila. Dev. Biol. 62: Shimizu H., S. A. Woodcock, M. B. Wilkin, B. Trubenová, N. A. Monk et al., Compensatory flux changes within an endocytic trafficking network maintain thermal 1256 robustness of Notch signaling. Cell 157: Simpson P., 1990 Lateral inhibition and the development of the sensory bristles of the adult 1258 peripheral nervous system of Drosophila. Development 109: Söllner T., S. W. Whiteheart, M. Brunner, H. Erdjument-Bromage, S. Geromanos et al., 1993a 1260 SNAP receptors implicated in vesicle targeting and fusion. Nature 362: Söllner T., M. K. Bennett, S. W. Whiteheart, R. H. Scheller, and J. E. Rothman, 1993b A 1262 protein assembly-disassembly pathway in vitro that may correspond to sequential steps of 1263 synaptic vesicle docking, activation and fusion. Cell 75: Stephenson N. L., and J. M. Avis, 2012 Direct observation of proteolytic cleavage at the S site upon forced unfolding of the Notch negative regulatory region. Proc. Natl. Acad. Sci U. S. A. 109: E Stewart B. A., M. Mohtashami, L. Zhou, W. S. Trimble and G. L. Boulianne, SNARE-dependent signaling at the Drosophila wing margin. Dev. Biol. 234: St Johnston D., 2002 The art and design of genetic screens: Drosophila melanogaster. Nat Rev. Genet. 3:
58 1271 Struhl G., K. Fitzgerald, and I. Greenwald, 1993 Intrinsic activity of the Lin12 and Notch 1272 intracellular domains in vivo. Cell 74: Sturtevant M. A., M. Roark, J. W. O'Neill, B. Biehs, N. Colley and E. Bier, 1996 The 1274 Drosophila rhomboid protein is concentrated in patches at the apical cell surface. Dev Biol. 174: Tanaka T., and A. Nakamura, 2008 The endocytic pathway acts downstream of Oskar in 1277 Drosophila germ plasm assembly. Development 135: Thibault S. T., M. A. Singer, W. Y. Miyazaki, B. Milash, N. A. Dompe et al., 2004 A 1279 complementary transposon tool kit for Drosophila melanogaster using P and piggybac Nat. Genet. 36: Uchiyama K., E. Jokitalo, F. Kano, M. Murata, X. Zhang et al., 2002 VCIP135, a novel 1282 essential factor for p97/p47-mediated membrane fusion, is required for Golgi and ER 1283 assembly in vivo. J. Cell Biol. 159: Vӓssin H., K. A. Bremer, E. Knust, and J. A. Campos-Ortega, 1987 The neurogenic Gene 1285 Delta of Drosophila melanogaster is expressed in neurogenic territories and encodes a 1286 putative transmembrane protein with EGF-like repeats. EMBO J. 6: Vӓssin H., and J. A. Campos-Ortega, 1987 Genetic analysis of Delta, a neurogenic gene of 1288 Drosophila melanogaster. Genetics 116: Wang W., and G. Struhl, 2004 Drosophila Epsin mediates a select endocytic pathway that 58
59 1290 DSL ligands must enter to activate Notch. Development 131: Wang W., and G. Struhl, 2005 Distinct roles for Mind bomb, Neuralized and Epsin in 1292 mediating DSL endocytosis and signaling in Drosophila. Development 132: Weiszmann R., A. S. Hammonds, and S. E. Celniker, 2009 Determination of gene expression 1294 patterns using high-throughput RNA in situ hybridization to whole-mount Drosophila 1295 embryos. Nat. Protoc. 4: Wharton K. A., K. M. Johansen, T. Xu, and S. Artavanis-Tsakonas, 1985 Nucleotide sequence 1297 from the neurogenic locus notch implies a gene product that shares homology with 1298 proteins containing EGF-like repeats. Cell 43: Wheeler S. R., S. B. Stagg, and S. T. Crews, 2008 Multiple Notch signaling events control 1300 Drosophila CNS midline neurogenesis, gliogenesis and neuronal identity. Development : Whiteheart S. W., K. Rossnagel, S. A. Buhrow, M. Brunner, R. Jaenicke et al., N-ethylmaleimide-sensitive fusion protein: A trimeric ATPase whose hydrolysis of ATP is 1304 required for membrane fusion. J. Cell Biol., 126: Wilson D. W., C. A. Wilcox, G. C. Flynn, E. Chen, W. J. Kuang et al., 1989 A fusion protein 1306 required for vesicle-mediated transport in both mammalian cells and yeast. Nature 339: Yamakawa T., K. Yamada, T. Sasamura, N. Nakazawa, M. Kanai et al., 2012 Deficient Notch 59
60 1309 signaling associated with neurogenic pecanex is compensated for by the unfolded protein 1310 response in Drosophila. Development 139: Young R. S., A. C. Marques, C. Tibbit, W. Haerty, A. R. Bassett et al., 2012 Identification and 1312 properties of 1,119 candidate lincrna loci in the Drosophila melanogaster genome Genome Biol. Evol. 4: Zhao M., S. Wu, Q. Zhou, S. Vivona, D. J. Cipriano et al., 2015 Mechanistic insights into the 1315 recycling machine of the SNARE complex. Nature 518:



. C + Y 18 25 28 18 25 28 28")

 pcx/y m (28 ) F +/+")

 * * * * 100% (n=6) Figure.")
61 A B The number of pupae pcx pcx X All females are zygotic pcx heterozygous flies lacking its maternal contribution (pcx/+ m ). All males are zygotic pcx hemizygous flies lacking its maternal contribution (pcx/y m ). C + Y The number of eclosed flies D The number of eggs E +/Y (28 ) 75.9% (n=29) pcx/y m (28 ) F +/ pcx/+ m G The percentages of female embryos in total embryos (N) * * * * 100% (n=6) Figure. 1 pcx shows cold-sensitive lethality.
 C The pupariation rate (larvae to")
 D * (N) Figure.")
62 A The hatching rate (N) ** ** ** ** ** ** ** ** B The survival rate by third instar larvae (N) C The pupariation rate (larvae to pupae) (N) * * * ** ** ** ** The eclosion rate (pupae to adults) D * (N) Figure. 2 Survival rate of pcx/+ m is decreased at lower temperature.
 C 100% (n=47) pcx/+ m")
 Elav 47.")
 Elav E % different")
 18 20 22 25")

63 A +/+ (Wild-type) B pcx/y m (Strong) C 100% (n=47) pcx/+ m (Quenched) Elav D 100% (n=19) pcx/+ m (Weak) Elav 47.7% (n=44) Elav 52.3% (n=44) Elav E % different extent of neural hyperplasia (N) Temperature Quenched Weak Figure. 3 pcx/+ m shows cold-sensitive neurogenic phenotype.
64 A pcx + Deficiency (Df) pcx X Y ; Balancers (2nd or 3rd) All females without balancers are zygotic pcx heterozygous flies lacking its maternal contribution and one copy of Df (pcx/+ m ; Df/+). Viable Df are screened at 25 degree. Lethal Df are screened at 28 degree. B 28 lethal (Enhancer) Df Small Df Tested allele Symbol No. Deleted region Symbol No. Deleted region Allele name Molecular function Cellular processes Df(2L)BSC50 30F4-31B1 Df(2L)BSC205 30F5-31A2 bib 1, Aquaporin Notch signaling Df(2L)BSC689 30F5-31B1 Df(2L)BSC251 30F5-31A1 Df(3R)ED D4-92A11 Df(3R)BSC850 91F11-92A6 Df(3R)ED F12-92B3 Dl 5F102 Notch ligand Notch signaling Df(2R)X D1-59A Df(2R)X B3-59A1 Df(2R)Kr10 60E10-60F5 Df(3R)ED A5-85D1 25 viable (Suppressor) Df(2R)gsb 60E12-60F4 lov 47 DNA binding Gravitaxis neur 1 E3 ubiquitin ligase Df Small Df Tested allele Symbol No. Deleted region Symbol No. Deleted region Allele name Molecular function Notch signaling Cellular processes Df(3L)ED217 70F4-71E1 Df(3L)BSC441 71A1-71B4 BobA 1 Bearded family Notch signaling Df(3R)ED F10-88A4 Nsf2 A6, Nsf2 A15 Nsf2 urd-2 ATPase SNARE-complex disassembly Figure. 4 Procedure of genetic screens and the summary of dominant modifiers of pcx.
65 A B The number of eclosed flies The number of eclosed flies C The number of eclosed flies 25 D The survival rate by third instar larvae (N) 260 * * ** ** ** ** E The survival rate by third instar larvae (N) 393 ** ** ** F The survival rate by third instar larvae (N) ** * ** Figure. 5 Dominant modification of the cold-sensitive lethality of pcx/+ m by the pcx modifiers identified as single loci.
 15 34 42")

66 A 28 (Enhancers) (N) % different extent of neural hyperplasia Normal Quenched Weak B 25 (Suppressors) (N) % different extent of neural hyperplasia Figure. 6 Dominant modification of the neurogenic phenotype of pcx/+ m by the pcx modifiers.
 Dl")
")

 Dl")



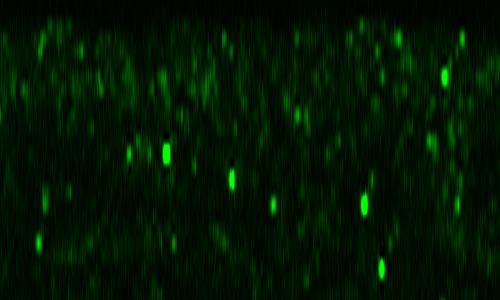










67 wt 100% (n=5) Dl 100% (n=5) Rab11 Dorsal BobA 100% (n=8) Dl 100% (n=8) Rab11 pcx mz 100% (n=5) Dl 100% (n=5) Rab11 wt 100% (n=5) Dl 100% (n=5) Rab11 Ventral BobA 100% (n=8) Dl 100% (n=8) Rab11 pcx mz 100% (n=5) Dl 100% (n=5) Rab11 Figure. 7 BobA suppresses Dl endocytosis in the dorsal region of embryos.
 Dl")

 Rho B")
")
")



68 A wt 100% (n=11) Dl 100% (n=11) Rho BobA 100% (n=7) Dl 100% (n=7) Rho B wt BobA 100% (n=27) Elav 41.2% (n=17) Elav Figure. 8 In the neuroectoderm of BobA mutant, Dl recycling and Notch signaling is hyperactivated.
")

")

")



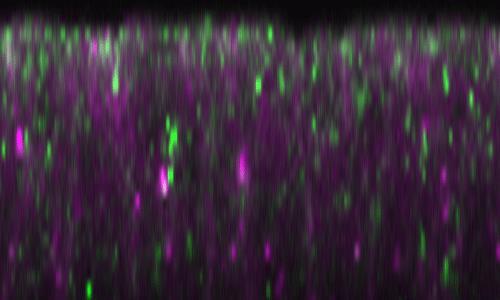





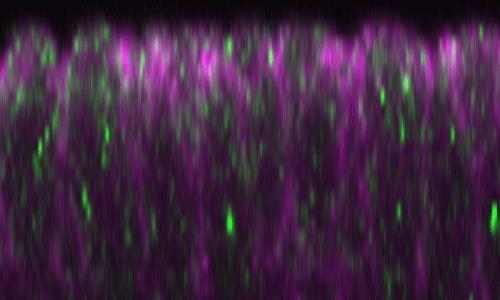



69 pcx/+ m 100% (n=3) Dl 100% (n=3) Rab11 Dorsal BobA/+ 100% (n=8) Dl 100% (n=8) Rab11 pcx/+ m ; BobA/+ 100% (n=5) Dl 100% (n=5) Rab11 pcx/+ m 100% (n=3) Dl 100% (n=3) Rab11 Ventral BobA/+ 100% (n=8) Dl 100% (n=8) Rab11 pcx/+ m ; BobA/+ 100% (n=5) Dl 100% (n=5) Rab11 Figure. 9 No synergistic interaction is observed between pcx/+ m and BobA/+ in Dl-trafficking phenotype.
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
SUPPLEMENTARY INFORMATION
 Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
allosteric cis-acting DNA element coding strand dominant constitutive mutation coordinate regulation of genes denatured
 A B C D E F G H I J K L M N O P Q R S T U V W X Y Z AA BB CC DD EE FF GG HH II JJ KK LL MM NN OO PP QQ RR SS TT UU VV allosteric cis-acting DNA element coding strand codominant constitutive mutation coordinate
A B C D E F G H I J K L M N O P Q R S T U V W X Y Z AA BB CC DD EE FF GG HH II JJ KK LL MM NN OO PP QQ RR SS TT UU VV allosteric cis-acting DNA element coding strand codominant constitutive mutation coordinate
1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms.
 Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Supporting Information
 Supporting Information Cao et al. 10.1073/pnas.1306220110 Gram - bacteria Hemolymph Cytoplasm PGRP-LC TAK1 signalosome Imd dfadd Dredd Dnr1 Ikk signalosome P Relish Nucleus AMP and effector genes Fig.
Supporting Information Cao et al. 10.1073/pnas.1306220110 Gram - bacteria Hemolymph Cytoplasm PGRP-LC TAK1 signalosome Imd dfadd Dredd Dnr1 Ikk signalosome P Relish Nucleus AMP and effector genes Fig.
Designer Genes C Test
 Northern Regional: January 19 th, 2019 Designer Genes C Test Name(s): Team Name: School Name: Team Number: Rank: Score: Directions: You will have 50 minutes to complete the test. You may not write on the
Northern Regional: January 19 th, 2019 Designer Genes C Test Name(s): Team Name: School Name: Team Number: Rank: Score: Directions: You will have 50 minutes to complete the test. You may not write on the
(Write your name on every page. One point will be deducted for every page without your name!)
") POPULATION GENETICS AND MICROEVOLUTIONARY THEORY FINAL EXAMINATION (Write your name on every page. One point will be deducted for every page without your name!) 1. Briefly define (5 points each): a) Average
POPULATION GENETICS AND MICROEVOLUTIONARY THEORY FINAL EXAMINATION (Write your name on every page. One point will be deducted for every page without your name!) 1. Briefly define (5 points each): a) Average
Nature Biotechnology: doi: /nbt Supplementary Figure 1. Overexpression of YFP::GPR-1 in the germline.
 Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Full file at CHAPTER 2 Genetics
 CHAPTER 2 Genetics MULTIPLE CHOICE 1. Chromosomes are a. small linear bodies. b. contained in cells. c. replicated during cell division. 2. A cross between true-breeding plants bearing yellow seeds produces
CHAPTER 2 Genetics MULTIPLE CHOICE 1. Chromosomes are a. small linear bodies. b. contained in cells. c. replicated during cell division. 2. A cross between true-breeding plants bearing yellow seeds produces
Supplementary Figure 1. Nature Genetics: doi: /ng.3848
 Supplementary Figure 1 Phenotypes and epigenetic properties of Fab2L flies. A- Phenotypic classification based on eye pigment levels in Fab2L male (orange bars) and female (yellow bars) flies (n>150).
Supplementary Figure 1 Phenotypes and epigenetic properties of Fab2L flies. A- Phenotypic classification based on eye pigment levels in Fab2L male (orange bars) and female (yellow bars) flies (n>150).
Chromosome duplication and distribution during cell division
 CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
Chromosome Chr Duplica Duplic t a ion Pixley
 Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
Green Fluorescent Protein (GFP) Today s Nobel Prize in Chemistry
 Today s Nobel Prize in Chemistry") In the news: High-throughput sequencing using Solexa/Illumina technology The copy number of each fetal chromosome can be determined by direct sequencing of DNA in cell-free plasma from pregnant women Confession:
In the news: High-throughput sequencing using Solexa/Illumina technology The copy number of each fetal chromosome can be determined by direct sequencing of DNA in cell-free plasma from pregnant women Confession:
A complementation test would be done by crossing the haploid strains and scoring the phenotype in the diploids.
 Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
Chapter 13 Meiosis and Sexual Reproduction
 Biology 110 Sec. 11 J. Greg Doheny Chapter 13 Meiosis and Sexual Reproduction Quiz Questions: 1. What word do you use to describe a chromosome or gene allele that we inherit from our Mother? From our Father?
Biology 110 Sec. 11 J. Greg Doheny Chapter 13 Meiosis and Sexual Reproduction Quiz Questions: 1. What word do you use to describe a chromosome or gene allele that we inherit from our Mother? From our Father?
Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section B and ONE question from Section C.
 UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2017-2018 GENETICS BIO-5009A Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section
UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2017-2018 GENETICS BIO-5009A Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section
Biology 322 Fall 2009 Wasp Genetics: Genetic Heterogeneity and Complementation Revisted
 Biology 322 Fall 2009 Wasp Genetics: Genetic Heterogeneity and Complementation Revisted Required Reading: Deaf by Design Nature 431: 894-896 October 21, 2004 http://fire.biol.wwu.edu/trent/trent/naturedeafdesign.pdf
Biology 322 Fall 2009 Wasp Genetics: Genetic Heterogeneity and Complementation Revisted Required Reading: Deaf by Design Nature 431: 894-896 October 21, 2004 http://fire.biol.wwu.edu/trent/trent/naturedeafdesign.pdf
Genetics 275 Notes Week 7
 Cytoplasmic Inheritance Genetics 275 Notes Week 7 Criteriafor recognition of cytoplasmic inheritance: 1. Reciprocal crosses give different results -mainly due to the fact that the female parent contributes
Cytoplasmic Inheritance Genetics 275 Notes Week 7 Criteriafor recognition of cytoplasmic inheritance: 1. Reciprocal crosses give different results -mainly due to the fact that the female parent contributes
Bypass and interaction suppressors; pathway analysis
 Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
2. Der Dissertation zugrunde liegende Publikationen und Manuskripte. 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera)
") 2. Der Dissertation zugrunde liegende Publikationen und Manuskripte 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera) M. Hasselmann 1, M. K. Fondrk², R. E. Page Jr.² und
2. Der Dissertation zugrunde liegende Publikationen und Manuskripte 2.1 Fine scale mapping in the sex locus region of the honey bee (Apis mellifera) M. Hasselmann 1, M. K. Fondrk², R. E. Page Jr.² und
BIS &003 Answers to Assigned Problems May 23, Week /18.6 How would you distinguish between an enhancer and a promoter?
 Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Solutions to Problem Set 4
 Question 1 Solutions to 7.014 Problem Set 4 Because you have not read much scientific literature, you decide to study the genetics of garden peas. You have two pure breeding pea strains. One that is tall
Question 1 Solutions to 7.014 Problem Set 4 Because you have not read much scientific literature, you decide to study the genetics of garden peas. You have two pure breeding pea strains. One that is tall
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Cell Division: the process of copying and dividing entire cells The cell grows, prepares for division, and then divides to form new daughter cells.
 Mitosis & Meiosis SC.912.L.16.17 Compare and contrast mitosis and meiosis and relate to the processes of sexual and asexual reproduction and their consequences for genetic variation. 1. Students will describe
Mitosis & Meiosis SC.912.L.16.17 Compare and contrast mitosis and meiosis and relate to the processes of sexual and asexual reproduction and their consequences for genetic variation. 1. Students will describe
Biology I Level - 2nd Semester Final Review
 Biology I Level - 2nd Semester Final Review The 2 nd Semester Final encompasses all material that was discussed during second semester. It s important that you review ALL notes and worksheets from the
Biology I Level - 2nd Semester Final Review The 2 nd Semester Final encompasses all material that was discussed during second semester. It s important that you review ALL notes and worksheets from the
Constructing a Pedigree
 Constructing a Pedigree Use the appropriate symbols: Unaffected Male Unaffected Female Affected Male Affected Female Male carrier of trait Mating of Offspring 2. Label each generation down the left hand
Constructing a Pedigree Use the appropriate symbols: Unaffected Male Unaffected Female Affected Male Affected Female Male carrier of trait Mating of Offspring 2. Label each generation down the left hand
- interactions between alleles. - multiple phenotypic effects of one gene. - phenotypic variability in single genes. - interactions between genes
 Genotype Phenotype - interactions between alleles - dominance - multiple phenotypic effects of one gene - phenotypic variability in single genes - interactions between genes amount of product threshold
Genotype Phenotype - interactions between alleles - dominance - multiple phenotypic effects of one gene - phenotypic variability in single genes - interactions between genes amount of product threshold
Transport between cytosol and nucleus
 of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila
 Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Objectives. Announcements. Comparison of mitosis and meiosis
 Announcements Colloquium sessions for which you can get credit posted on web site: Feb 20, 27 Mar 6, 13, 20 Apr 17, 24 May 15. Review study CD that came with text for lab this week (especially mitosis
Announcements Colloquium sessions for which you can get credit posted on web site: Feb 20, 27 Mar 6, 13, 20 Apr 17, 24 May 15. Review study CD that came with text for lab this week (especially mitosis
Biol. 303 EXAM I 9/22/08 Name
 Biol. 303 EXAM I 9/22/08 Name -------------------------------------------------------------------------------------------------------------- This exam consists of 40 multiple choice questions worth 2.5
Biol. 303 EXAM I 9/22/08 Name -------------------------------------------------------------------------------------------------------------- This exam consists of 40 multiple choice questions worth 2.5
Genetic Lab 3. Drosophila Fly
 Genetic Lab 3 Drosophila Fly An Introduction to fruit or vinegar fly Drosophila Melanogaster Is a small (about 3mm long), common fly found near unripe and rotted fruit, so that it called fruit or vinegar
Genetic Lab 3 Drosophila Fly An Introduction to fruit or vinegar fly Drosophila Melanogaster Is a small (about 3mm long), common fly found near unripe and rotted fruit, so that it called fruit or vinegar
Bio/Life: Cell Biology
 Bio/Life: Cell Biology 1a The fundamental life processes of plants and animals depend on a variety of chemical reactions that occur in specialized areas of the organism's cells. As a basis for understanding
Bio/Life: Cell Biology 1a The fundamental life processes of plants and animals depend on a variety of chemical reactions that occur in specialized areas of the organism's cells. As a basis for understanding
F1 Parent Cell R R. Name Period. Concept 15.1 Mendelian inheritance has its physical basis in the behavior of chromosomes
 Name Period Concept 15.1 Mendelian inheritance has its physical basis in the behavior of chromosomes 1. What is the chromosome theory of inheritance? 2. Explain the law of segregation. Use two different
Name Period Concept 15.1 Mendelian inheritance has its physical basis in the behavior of chromosomes 1. What is the chromosome theory of inheritance? 2. Explain the law of segregation. Use two different
Mapping of the l(3)dts4 Gene and Analysis of its Role in the Notch Pathway. Jessica Keating Bachelor of Science George Mason University 2005
dts4 Gene and Analysis of its Role in the Notch Pathway. Jessica Keating Bachelor of Science George Mason University 2005") Mapping of the l(3)dts4 Gene and Analysis of its Role in the Notch Pathway A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science at George Mason University By
Mapping of the l(3)dts4 Gene and Analysis of its Role in the Notch Pathway A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science at George Mason University By
Q2 (4.6) Put the following in order from biggest to smallest: Gene DNA Cell Chromosome Nucleus. Q8 (Biology) (4.6)
 Put the following in order from biggest to smallest: Gene DNA Cell Chromosome Nucleus. Q8 (Biology) (4.6)") Q1 (4.6) What is variation? Q2 (4.6) Put the following in order from biggest to smallest: Gene DNA Cell Chromosome Nucleus Q3 (4.6) What are genes? Q4 (4.6) What sort of reproduction produces genetically
Q1 (4.6) What is variation? Q2 (4.6) Put the following in order from biggest to smallest: Gene DNA Cell Chromosome Nucleus Q3 (4.6) What are genes? Q4 (4.6) What sort of reproduction produces genetically
Reading: Chapter 5, pp ; Reference chapter D, pp Problem set F
 Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Darwinian Selection. Chapter 7 Selection I 12/5/14. v evolution vs. natural selection? v evolution. v natural selection
 Chapter 7 Selection I Selection in Haploids Selection in Diploids Mutation-Selection Balance Darwinian Selection v evolution vs. natural selection? v evolution ² descent with modification ² change in allele
Chapter 7 Selection I Selection in Haploids Selection in Diploids Mutation-Selection Balance Darwinian Selection v evolution vs. natural selection? v evolution ² descent with modification ² change in allele
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
7.06 Problem Set
 7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
Lab I: Three-Point Mapping in Drosophila melanogaster
 Lab I: Three-Point Mapping in Drosophila melanogaster Makuo Aneke Partner: Christina Hwang BIO 365-004: Genetics with Laboratory TA: Dr. Hongmei Ma February 18, 2016 Abstract The purpose of this experiment
Lab I: Three-Point Mapping in Drosophila melanogaster Makuo Aneke Partner: Christina Hwang BIO 365-004: Genetics with Laboratory TA: Dr. Hongmei Ma February 18, 2016 Abstract The purpose of this experiment
Cellular Neuroanatomy I The Prototypical Neuron: Soma. Reading: BCP Chapter 2
 Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
BILD7: Problem Set. 2. What did Chargaff discover and why was this important?
 BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
GSBHSRSBRSRRk IZTI/^Q. LlML. I Iv^O IV I I I FROM GENES TO GENOMES ^^^H*" ^^^^J*^ ill! BQPIP. illt. goidbkc. itip31. li4»twlil FIFTH EDITION
 FIFTH EDITION IV I ^HHk ^ttm IZTI/^Q i I II MPHBBMWBBIHB '-llwmpbi^hbwm^^pfc ' GSBHSRSBRSRRk LlML I I \l 1MB ^HP'^^MMMP" jflp^^^^^^^^st I Iv^O FROM GENES TO GENOMES %^MiM^PM^^MWi99Mi$9i0^^ ^^^^^^^^^^^^^V^^^fii^^t^i^^^^^
FIFTH EDITION IV I ^HHk ^ttm IZTI/^Q i I II MPHBBMWBBIHB '-llwmpbi^hbwm^^pfc ' GSBHSRSBRSRRk LlML I I \l 1MB ^HP'^^MMMP" jflp^^^^^^^^st I Iv^O FROM GENES TO GENOMES %^MiM^PM^^MWi99Mi$9i0^^ ^^^^^^^^^^^^^V^^^fii^^t^i^^^^^
Related Courses He who asks is a fool for five minutes, but he who does not ask remains a fool forever.
 CSE 527 Computational Biology http://www.cs.washington.edu/527 Lecture 1: Overview & Bio Review Autumn 2004 Larry Ruzzo Related Courses He who asks is a fool for five minutes, but he who does not ask remains
CSE 527 Computational Biology http://www.cs.washington.edu/527 Lecture 1: Overview & Bio Review Autumn 2004 Larry Ruzzo Related Courses He who asks is a fool for five minutes, but he who does not ask remains
Chapter 2: Extensions to Mendel: Complexities in Relating Genotype to Phenotype.
 Chapter 2: Extensions to Mendel: Complexities in Relating Genotype to Phenotype. please read pages 38-47; 49-55;57-63. Slide 1 of Chapter 2 1 Extension sot Mendelian Behavior of Genes Single gene inheritance
Chapter 2: Extensions to Mendel: Complexities in Relating Genotype to Phenotype. please read pages 38-47; 49-55;57-63. Slide 1 of Chapter 2 1 Extension sot Mendelian Behavior of Genes Single gene inheritance
SIGNALING through the Notch receptor is widely
 Copyright Ó 2009 by the Genetics Society of America DOI: 10.1534/genetics.109.105023 Genome Engineering-Based Analysis of Bearded Family Genes Reveals Both Functional Redundancy and a Nonessential Function
Copyright Ó 2009 by the Genetics Society of America DOI: 10.1534/genetics.109.105023 Genome Engineering-Based Analysis of Bearded Family Genes Reveals Both Functional Redundancy and a Nonessential Function
9/4/2015 INDUCTION CHAPTER 1. Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology. Fig 1.
 INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
The Drosophila neurogenic gene big brain, which encodes a membraneassociated protein, acts cell autonomously and can act synergistically with
 Development 124, 3881-3893 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV8422 3881 The Drosophila neurogenic gene big brain, which encodes a membraneassociated protein, acts
Development 124, 3881-3893 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV8422 3881 The Drosophila neurogenic gene big brain, which encodes a membraneassociated protein, acts
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
BIOLOGY 111. CHAPTER 5: Chromosomes and Inheritance
 BIOLOGY 111 CHAPTER 5: Chromosomes and Inheritance Chromosomes and Inheritance Learning Outcomes 5.1 Differentiate between sexual and asexual reproduction in terms of the genetic variation of the offspring.
BIOLOGY 111 CHAPTER 5: Chromosomes and Inheritance Chromosomes and Inheritance Learning Outcomes 5.1 Differentiate between sexual and asexual reproduction in terms of the genetic variation of the offspring.
Extranuclear Inheritance
 Extranuclear Inheritance Extranuclear Inheritance The past couple of lectures, we ve been exploring exceptions to Mendel s principles of transmission inheritance. Scientists have observed inheritance patterns
Extranuclear Inheritance Extranuclear Inheritance The past couple of lectures, we ve been exploring exceptions to Mendel s principles of transmission inheritance. Scientists have observed inheritance patterns
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
SUPPLEMENTARY INFORMATION
 med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
Lesson Overview Meiosis
 11.4 As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located. They expected genes to be carried on structures inside the cell, but which structures? What cellular
11.4 As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located. They expected genes to be carried on structures inside the cell, but which structures? What cellular
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Natural Selection. Population Dynamics. The Origins of Genetic Variation. The Origins of Genetic Variation. Intergenerational Mutation Rate
 Natural Selection Population Dynamics Humans, Sickle-cell Disease, and Malaria How does a population of humans become resistant to malaria? Overproduction Environmental pressure/competition Pre-existing
Natural Selection Population Dynamics Humans, Sickle-cell Disease, and Malaria How does a population of humans become resistant to malaria? Overproduction Environmental pressure/competition Pre-existing
1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine.
 Protein Synthesis & Mutations RNA 1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine. RNA Contains: 1. Adenine 2.
Protein Synthesis & Mutations RNA 1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine. RNA Contains: 1. Adenine 2.
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
The phenotype of this worm is wild type. When both genes are mutant: The phenotype of this worm is double mutant Dpy and Unc phenotype.
 Series 2: Cross Diagrams - Complementation There are two alleles for each trait in a diploid organism In C. elegans gene symbols are ALWAYS italicized. To represent two different genes on the same chromosome:
Series 2: Cross Diagrams - Complementation There are two alleles for each trait in a diploid organism In C. elegans gene symbols are ALWAYS italicized. To represent two different genes on the same chromosome:
Sequence analysis and comparison
 The aim with sequence identification: Sequence analysis and comparison Marjolein Thunnissen Lund September 2012 Is there any known protein sequence that is homologous to mine? Are there any other species
The aim with sequence identification: Sequence analysis and comparison Marjolein Thunnissen Lund September 2012 Is there any known protein sequence that is homologous to mine? Are there any other species
Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain
 Development 127, 367-379 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV5342 367 Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain Wei Du
Development 127, 367-379 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV5342 367 Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain Wei Du
Name KEY. Biology Developmental Biology Winter Quarter Midterm 1
 Name KEY 100 Total Points Open Book Biology 411 - Developmental Biology Winter Quarter 2008 Midterm 1 25 questions - 4 pts each 1 extra-credit question - 4 points Provide answers using full sentences,
Name KEY 100 Total Points Open Book Biology 411 - Developmental Biology Winter Quarter 2008 Midterm 1 25 questions - 4 pts each 1 extra-credit question - 4 points Provide answers using full sentences,
Lesson Overview 11.4 Meiosis
 Lesson Overview 11.4 Meiosis THINK ABOUT IT As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located. They expected genes to be carried on structures inside the
Lesson Overview 11.4 Meiosis THINK ABOUT IT As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located. They expected genes to be carried on structures inside the
Axis determination in flies. Sem 9.3.B.5 Animal Science
 Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
KEY: Chapter 9 Genetics of Animal Breeding.
 KEY: Chapter 9 Genetics of Animal Breeding. Answer each question using the reading assigned to you. You can access this information by clicking on the following URL: https://drive.google.com/a/meeker.k12.co.us/file/d/0b1yf08xgyhnad08xugxsnfvba28/edit?usp=sh
KEY: Chapter 9 Genetics of Animal Breeding. Answer each question using the reading assigned to you. You can access this information by clicking on the following URL: https://drive.google.com/a/meeker.k12.co.us/file/d/0b1yf08xgyhnad08xugxsnfvba28/edit?usp=sh
PRACTICE EXAM. 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos.
 PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
Objective 3.01 (DNA, RNA and Protein Synthesis)
") Objective 3.01 (DNA, RNA and Protein Synthesis) DNA Structure o Discovered by Watson and Crick o Double-stranded o Shape is a double helix (twisted ladder) o Made of chains of nucleotides: o Has four types
Objective 3.01 (DNA, RNA and Protein Synthesis) DNA Structure o Discovered by Watson and Crick o Double-stranded o Shape is a double helix (twisted ladder) o Made of chains of nucleotides: o Has four types
Follow this and additional works at: Part of the Medical Sciences Commons
 Bucknell University Bucknell Digital Commons Master s Theses Student Theses 2010 The overexpression of homeotic complex gene Ultrabithorax in the post-embryonic neuronal lineages of the ventral nervous
Bucknell University Bucknell Digital Commons Master s Theses Student Theses 2010 The overexpression of homeotic complex gene Ultrabithorax in the post-embryonic neuronal lineages of the ventral nervous
Which of these best predicts the outcome of the changes illustrated in the diagrams?
 1. The diagrams below show two different scenarios for a pair of homologous chromosomes, known as a tetrad, undergoing a change where segments of DNA switch on parts of the chromosomes. In each scenario,
1. The diagrams below show two different scenarios for a pair of homologous chromosomes, known as a tetrad, undergoing a change where segments of DNA switch on parts of the chromosomes. In each scenario,
Unit 3 - Molecular Biology & Genetics - Review Packet
 Name Date Hour Unit 3 - Molecular Biology & Genetics - Review Packet True / False Questions - Indicate True or False for the following statements. 1. Eye color, hair color and the shape of your ears can
Name Date Hour Unit 3 - Molecular Biology & Genetics - Review Packet True / False Questions - Indicate True or False for the following statements. 1. Eye color, hair color and the shape of your ears can
7) In an organism with 52 chromosomes, how many bivalents would be expected to form during meiosis? A) 13 B) 104 C) 26 D) 208 E) 52 Answer: C
 In an organism with 52 chromosomes, how many bivalents would be expected to form during meiosis? A) 13 B) 104 C) 26 D) 208 E) 52 Answer: C") Essentials of Genetics 9th Edition by William S. Klug, Michael R. Cummings, Charlotte A. Spencer, Michael A. Palladino Test Bank MULTIPLE CHOICE. Choose the one alternative that best completes the statement
Essentials of Genetics 9th Edition by William S. Klug, Michael R. Cummings, Charlotte A. Spencer, Michael A. Palladino Test Bank MULTIPLE CHOICE. Choose the one alternative that best completes the statement
- mutations can occur at different levels from single nucleotide positions in DNA to entire genomes.
 February 8, 2005 Bio 107/207 Winter 2005 Lecture 11 Mutation and transposable elements - the term mutation has an interesting history. - as far back as the 17th century, it was used to describe any drastic
February 8, 2005 Bio 107/207 Winter 2005 Lecture 11 Mutation and transposable elements - the term mutation has an interesting history. - as far back as the 17th century, it was used to describe any drastic
BIOLOGY STANDARDS BASED RUBRIC
 BIOLOGY STANDARDS BASED RUBRIC STUDENTS WILL UNDERSTAND THAT THE FUNDAMENTAL PROCESSES OF ALL LIVING THINGS DEPEND ON A VARIETY OF SPECIALIZED CELL STRUCTURES AND CHEMICAL PROCESSES. First Semester Benchmarks:
BIOLOGY STANDARDS BASED RUBRIC STUDENTS WILL UNDERSTAND THAT THE FUNDAMENTAL PROCESSES OF ALL LIVING THINGS DEPEND ON A VARIETY OF SPECIALIZED CELL STRUCTURES AND CHEMICAL PROCESSES. First Semester Benchmarks:
AP Biology Gene Regulation and Development Review
 AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
MCB 141 Midterm I Feb. 19, 2009
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
TGF-β/BMP superfamily members, Gbb-60A and Dpp, cooperate to provide pattern information and establish cell identity in the Drosophila wing
 Development 125, 2723-2734 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5189 2723 TGF-β/BMP superfamily members, Gbb-60A and Dpp, cooperate to provide pattern information and
Development 125, 2723-2734 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5189 2723 TGF-β/BMP superfamily members, Gbb-60A and Dpp, cooperate to provide pattern information and
THINK ABOUT IT. Lesson Overview. Meiosis. As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located.
 Notes THINK ABOUT IT As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located. They expected genes to be carried on structures inside the cell, but which structures?
Notes THINK ABOUT IT As geneticists in the early 1900s applied Mendel s laws, they wondered where genes might be located. They expected genes to be carried on structures inside the cell, but which structures?
Multiple Choice Review- Eukaryotic Gene Expression
 Multiple Choice Review- Eukaryotic Gene Expression 1. Which of the following is the Central Dogma of cell biology? a. DNA Nucleic Acid Protein Amino Acid b. Prokaryote Bacteria - Eukaryote c. Atom Molecule
Multiple Choice Review- Eukaryotic Gene Expression 1. Which of the following is the Central Dogma of cell biology? a. DNA Nucleic Acid Protein Amino Acid b. Prokaryote Bacteria - Eukaryote c. Atom Molecule
Essentials of Genetics, 8e (Klug) Chapter 2 Mitosis and Meiosis
 Chapter 2 Mitosis and Meiosis") Essentials of Genetics, 8e (Klug) Chapter 2 Mitosis and Meiosis 1) During interphase of the cell cycle,. A) DNA recombines B) sister chromatids move to opposite poles C) the nuclear membrane disappears
Essentials of Genetics, 8e (Klug) Chapter 2 Mitosis and Meiosis 1) During interphase of the cell cycle,. A) DNA recombines B) sister chromatids move to opposite poles C) the nuclear membrane disappears
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
Systematic Study of Drosophila MicroRNA Functions Using a Collection of Targeted Knockout Mutations
 Resource Systematic Study of Drosophila MicroRNA Functions Using a Collection of Targeted Knockout Mutations Ya-Wen Chen, 1 Shilin Song, 1 Ruifen Weng, 1 Pushpa Verma, 1 Jan-Michael Kugler, 1,4 Marita
Resource Systematic Study of Drosophila MicroRNA Functions Using a Collection of Targeted Knockout Mutations Ya-Wen Chen, 1 Shilin Song, 1 Ruifen Weng, 1 Pushpa Verma, 1 Jan-Michael Kugler, 1,4 Marita
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
CHAPTER 23 THE EVOLUTIONS OF POPULATIONS. Section C: Genetic Variation, the Substrate for Natural Selection
 CHAPTER 23 THE EVOLUTIONS OF POPULATIONS Section C: Genetic Variation, the Substrate for Natural Selection 1. Genetic variation occurs within and between populations 2. Mutation and sexual recombination
CHAPTER 23 THE EVOLUTIONS OF POPULATIONS Section C: Genetic Variation, the Substrate for Natural Selection 1. Genetic variation occurs within and between populations 2. Mutation and sexual recombination
Lesson 4: Understanding Genetics
 Lesson 4: Understanding Genetics 1 Terms Alleles Chromosome Co dominance Crossover Deoxyribonucleic acid DNA Dominant Genetic code Genome Genotype Heredity Heritability Heritability estimate Heterozygous
Lesson 4: Understanding Genetics 1 Terms Alleles Chromosome Co dominance Crossover Deoxyribonucleic acid DNA Dominant Genetic code Genome Genotype Heredity Heritability Heritability estimate Heterozygous
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION
 MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
BS 50 Genetics and Genomics Week of Oct 3 Additional Practice Problems for Section. A/a ; B/B ; d/d X A/a ; b/b ; D/d
 BS 50 Genetics and Genomics Week of Oct 3 Additional Practice Problems for Section 1. In the following cross, all genes are on separate chromosomes. A is dominant to a, B is dominant to b and D is dominant
BS 50 Genetics and Genomics Week of Oct 3 Additional Practice Problems for Section 1. In the following cross, all genes are on separate chromosomes. A is dominant to a, B is dominant to b and D is dominant
Complex Function and Expression of Delta During Drosophila Oogenesis
 Copyright 0 1993 by the Genetics Society of America Complex Function and Expression of Delta During Drosophila Oogenesis Laurel B. Bender, Pamela J. Kooh' and Marc A. T. Muskavitch' Program in Genetics,
Copyright 0 1993 by the Genetics Society of America Complex Function and Expression of Delta During Drosophila Oogenesis Laurel B. Bender, Pamela J. Kooh' and Marc A. T. Muskavitch' Program in Genetics,
VCE BIOLOGY Relationship between the key knowledge and key skills of the Study Design and the Study Design
 VCE BIOLOGY 2006 2014 Relationship between the key knowledge and key skills of the 2000 2005 Study Design and the 2006 2014 Study Design The following table provides a comparison of the key knowledge (and
VCE BIOLOGY 2006 2014 Relationship between the key knowledge and key skills of the 2000 2005 Study Design and the 2006 2014 Study Design The following table provides a comparison of the key knowledge (and
AP Biology Unit 6 Practice Test 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8
 AP Biology Unit 6 Practice Test Name: 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8 picograms of DNA per nucleus. How many picograms
AP Biology Unit 6 Practice Test Name: 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8 picograms of DNA per nucleus. How many picograms