mrna and Genetic code standard
|
|
|
- Gary Nelson
- 5 years ago
- Views:
Transcription
1 Synthesis and Processing of the Proteome. (mrna and Genetic code: standard and variations; codon anti codon interactions), The role of trna in protein synthesis, Role of Ribosome in Protein synthesis (Ribosome structure, The players... mrna, trna, activating enzymes, initiation of translation in bacteria and eukaryotes, elongation of polypeptide in bacteria and eukaryotes and termination of polypepetide synthesis in bacteria and eukayrotes), Post translation modifications (Protein folding, Proteolysis, Protein chemical modification) Protein trafficking and degradation, inhibition of protein synthesis (antibiotic). Mitesh Shrestha

2 mrna and Genetic code standard
3 The Genetic Code Codons: 3 base code for the production of a specific amino acid, sequence of three of the four different nucleotides Since there are 4 bases and 3 positions in each codon, there are 4 x 4 x 4 = 64 possible codons 64 codons but only 20 amino acids, therefore most have more than 1 codon 3 of the 64 codons are used as STOP signals; they are found at the end of every gene and mark the end of the protein One codon is used as a START signal: it is at the start of every protein Universal: in all living organisms
4 Variations In 1979, researchers studying human mitochondrial genes discovered they used an alternative code Small variants such as translation of the codon UGA as tryptophan in Mycoplasma species, and translation of CUG as a serine rather than a leucine in yeasts of the "CTG clade" (Candida albicans is member of this group). In bacteria and archaea, GUG and UUG are common start codons, but in rare cases, certain proteins may use alternative start codons not normally used by that species. In certain proteins, non-standard amino acids are substituted for standard stop codons, depending on associated signal sequences in the messenger RNA. For example, UGA can code for selenocysteine and UAG can code for pyrrolysine. Selenocysteine is now viewed as the 21st amino acid, and pyrrolysine is viewed as the 22nd.
5 The Vertebrate Mitochondrial Code: Differences from the Standard Code: Codes Standard AGA Ter * Arg R AGG Ter * Arg R AUA Met M Ile I UGA Trp W Ter *
6 The Yeast Mitochondrial Code Differences from the Standard Code: Codes Standard AUA Met M Ile I CUU Thr T Leu L CUC Thr T Leu L CUA Thr T Leu L CUG Thr T Leu L UGA Trp W Ter * CGA absent Arg R CGC absent Arg R
7 trnas nt in length Structure of trna The acceptor arm is formed by seven base pairs between the 5 and 3 ends of the molecule. The amino acid is attached to the extreme 3 end of the trna, to the adenosine of the invariant CCA terminal sequence. The D arm, named after the modified nucleoside dihydrouridine, which is always present in this structure. The anticodon arm contains the triplet of nucleotides called the anticodon which base-pair with the mrna during translation. The V loop contains 3 5 nucleotides in Class 1 trnas or nucleotides in Class 2 trnas. The TΨC arm, named after the sequence thymidine-pseudouridine-cytosine, which is always present.


8 Structure of trna
9 Structure of trna The cloverleaf structure can be formed by virtually all trnas, the main exceptions being the trnas used in vertebrate mitochondria, which are coded by the mitochondrial genome and which sometimes lack parts of the structure. An example is the human mitochondrial trna Ser, which has no D arm. As well as the conserved secondary structure, the identities of nucleotides at some positions are completely invariant (always the same nucleotide) or semi-invariant (always a purine or always a pyrimidine), and the positions of the modified nucleotides are almost always the same.
10 The role of trna in protein synthesis Transfer RNAs play the central role in translation. Adaptor molecules, which form the link between the mrna and the polypeptide that is being synthesized. A physical link, trnas binding to both the mrna and the growing polypeptide, and an informational link, trnas ensuring that the polypeptide being synthesized has the amino acid sequence that is denoted, via the genetic code, by the sequence of nucleotides in the mrna. trnas play this dual role through aminoacylation, the process by which the correct amino acid is attached to each trna, and codon-anticodon recognition, the interaction between trna and mrna.
11
12 Aminoacylation Process of adding an aminoacyl group to a compound. It produces a trna molecule with its CCA 3' end covalently linked to an amino acid. Each trna is aminoacylated (or charged) with a specific amino acid by an aminoacyl trna synthetase. There is normally a single aminoacyl trna synthetase for each amino acid, despite the fact that there can be more than one trna, and more than one anticodon, for an amino acid. Recognition of the appropriate trna by the synthetases is not mediated solely by the anticodon, and the acceptor stem often plays a prominent role. As only 20 amino acids are designated by the genetic code, this means that all organisms have at least some isoaccepting trnas, different trnas that are specific for the same amino acid. The terminology used when describing trnas is to indicate the amino acid specificity with a superscript suffix, using the numbers 1, 2, etc., to distinguish different isoacceptors: for example, two trnas specific for glycine would be written as trna Gly1 and trna Gly2.
13 Aminoacylation The attachment of amino acids to trnas - charging in molecular biology jargon - is the function of the group of enzymes called aminoacyl-trna synthetases. The chemical reaction that results in aminoacylation occurs in two steps. An activated amino acid intermediate is first formed by reaction between the amino acid and ATP, and then the amino acid is transferred to the 3 end of the trna, the link being formed between the -COOH group of the amino acid and the -OH group attached to either the 2 or 3 carbon on the sugar of the last nucleotide, which is always an A
14 Aminoacylation The result of aminoacylation by a Class II aminoacyl-trna synthetase is shown, the amino acid being attached via its -COOH group to the 3 -OH of the terminal nucleotide of the trna. A Class I aminoacyl-trna synthetase attaches the amino acid to the 2 -OH group.
15 Features of aminoacyl-trna synthetases Feature Class I enzymes Class II enzymes Structure of the enzyme active site Interaction with the trna Orientation of the bound trna Amino acid attachment Enzymes for * Parallel β-sheet Minor groove of the acceptor stem V loop faces away from the enzyme To the 2 -OH of the terminal nucleotide of the trna Arg, Cys, Gln, Glu, Ile, Leu, Lys1, Met, Trp, Tyr, Val Antiparallel β-sheet Major groove of the acceptor stem V loop faces the enzyme To the 3 -OH of the terminal nucleotide of the trna Ala, Asn, Asp, Gly, His, LysII, Phe, Pro, Thr, Ser
16 Aminoacylation With a few exceptions, organisms have 20 aminoacyltrna synthetases, one for each amino acid. This means that groups of isoaccepting trnas are aminoacylated by a single enzyme. Although the basic chemical reaction is the same for each amino acid, the 20 aminoacyl-trna synthetases fall into two distinct groups, Class I and Class II, with several important differences between them. In particular, Class I enzymes attach the amino acid to the 2 -OH group of the terminal nucleotide of the trna, whereas Class II enzymes attach the amino acid to the 3 -OH group
17 Building a Molecule of trna A specific enzyme called an aminoacyl-trna synthetase joins each amino acid to the correct trna Amino acid Aminoacyl-tRNA synthetase (enzyme) P P P ATP Adenosine 1 Active site binds the amino acid and ATP. 2 ATP loses two P groups and joins amino acid as AMP. Pyrophosphate P P i P Adenosine 3 Appropriate trna covalently Bonds to amino Acid, displacing AMP. Phosphates P i P i trna P Adenosine AMP 4 Activated amino acid is released by the enzyme. Aminoacyl trna (an activated amino acid )
18 Unusual types of aminoacylation


19 Codon Anti codon interactions Codon anticodon pairing is unstable in the absence of the ribosome.
20 Codon Anti codon interactions A straightforward process involving base-pairing between the anticodon of the trna and a codon in the mrna
21 Wobble hypothesis Francis Crick The first two bases in the codon create the coding specificity, for they form strong Watson-Crick base pairs and bond strongly to the anticodon of the trna. When reading 5' to 3, the first nucleotide in the anticodon (which is on the trna and pairs with the last nucleotide of the codon on the mrna) determines how many nucleotides the trna actually distinguishes. If the first nucleotide in the anticodon is a C or an A pairing is specific and acknowledges original Watson-Crick pairing, that is only one specific codon can be paired to that trna. If the first nucleotide is U or G, the pairing is less specific and in fact two bases can be interchangeably recognized by the trna. Inosine displays the true qualities of wobble, in that if that is the first nucleotide in the anticodon then any of three bases in the original codon can be matched with the trna. Due to the specificity inherent in the first two nucleotides of the codon, if one amino acid codes for multiple anticodons and those anticodons differ in either the second or third position (first or second position in the codon) then a different trna is required for that anticodon. The minimum requirement to satisfy all possible codons (61 excluding three stop codons) is 32 trnas. That is 31 trna's for the amino acids and one initiation codon.
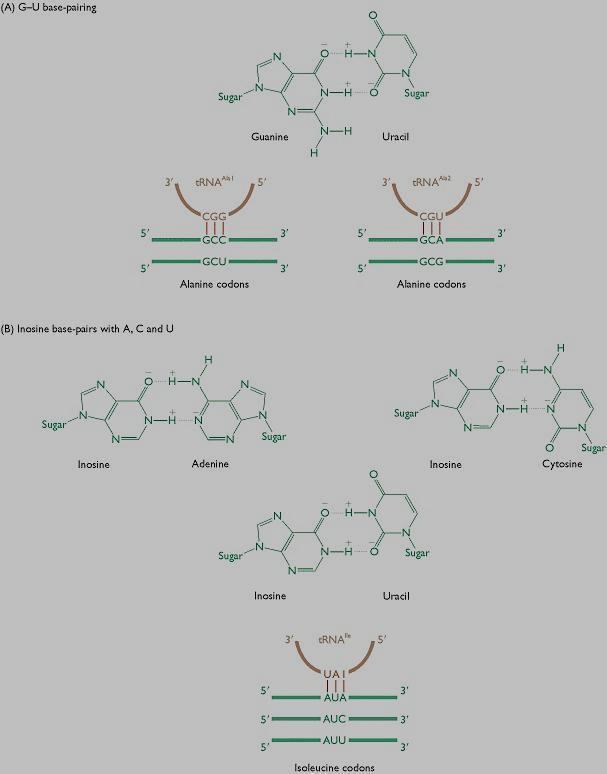
22 Wobble hypothesis

23
24 Codon Anti codon interactions The specificity of aminoacylation ensures that the trna carries the amino acid denoted by the codon that it pairs with, and the ribosome controls the topology of the interaction in such a way that only a single triplet of nucleotides is available for pairing.
25 Role of the Ribosome in Protein Synthesis Two active roles in protein synthesis Ribosomes coordinate protein synthesis by placing the mrna, aminoacyl-trnas and associated protein factors in their correct positions relative to one another. Components of ribosomes, including the rrnas, catalyze at least some of the chemical reactions occurring during translation.

26 Structure of Ribosome
 Schematic model showing binding sites. A ribosome has an mrna binding site and three trna binding sites, known as the A, P, and E sites.")
27 Ribosome The ribosome has three binding sites for trna The P site The A site The E site P site (Peptidyl-tRNA binding site) E site (Exit site) E P A A site (AminoacyltRNA binding site) Large subunit mrna binding site Small subunit (b) Schematic model showing binding sites. A ribosome has an mrna binding site and three trna binding sites, known as the A, P, and E sites. This schematic ribosome will appear in later diagrams.
28 Ribosome in protein synthesis. In E. coli, ribosome is assembled on mrna at initiation codon w/translation initiation factor IF-3 (prevents premature dissociation); 3 of 16S rrna attached to ribosome binding site (Shine-Dalgarno sequence).
29 Translation The process in which cellular ribosomes create proteins. Translation proceeds in three phases: Initiation: The ribosome assembles around the target mrna. The first trna is attached at the start codon. Elongation: The trna transfers an amino acid to the trna corresponding to the next codon. The ribosome then moves (translocates) to the next mrna codon to continue the process, creating an amino acid chain. Termination: When a stop codon is reached, the ribosome releases the polypeptide.
30 BACTERIAL TRANSLATION Factor Function Initiation factors IF-1 Unclear; X-ray crystallography studies show that binding of IF-1 blocks the A site (see page 329), so its function may be to prevent premature entry of trnas into the A site. Alternatively IF-1 may cause conformational changes that prepare the small subunit for attachment to the large subunit IF-2 IF-3 Elongation factors EF-Tu EF-Ts EF-G Release factors RF-1 RF-2 RF-3 Directs the initiator trnai Met to its correct position in the initiation complex Prevents premature reassociation of the large and small subunits of the ribosome Directs the next trna to its correct position in the ribosome Regenerates EF-Tu after the latter has yielded the energy contained in its attached GTP molecule Mediates translocation Recognizes the termination codons 5 -UAA-3 and 5 -UAG-3 Recognizes 5 -UAA-3 and 5 -UGA-3 Stimulates dissociation of RF1 and RF2 from the ribosome after termination Ribosome recycling factor RRF Responsible for disassociating the ribosome subunits after translation has terminated
31 Initiation of translation The main difference between initiation of translation in bacteria and eukaryotes is that in bacteria the translation initiation complex is built up directly at the initiation codon, the point at which protein synthesis will begin, whereas eukaryotes use a more indirect process for locating the initiation point.

32 Initiation of Translation The initiation stage of translation brings together mrna, trna bearing the first amino acid of the polypeptide, and two subunits of a ribosome 3 5 U A C A U G 5 3 P site Large ribosomal subunit Initiator trna mrna GTP GDP E A 5 Start codon mrna binding site Small ribosomal subunit Translation initiation complex 1 A small ribosomal subunit binds to a molecule of mrna. In a prokaryotic cell, the mrna binding site on this subunit recognizes a specific nucleotide sequence on the mrna just upstream of the start codon. An initiator trna, with the anticodon UAC, base-pairs with the start codon, AUG. This trna carries the amino acid methionine (Met). 2 The arrival of a large ribosomal subunit completes the initiation complex. Proteins called initiation factors (not shown) are required to bring all the translation components together. GTP provides the energy for the assembly. The initiator trna is in the P site; the A site is available to the trna bearing the next amino acid.
33 Initiation of protein synthesis in E. coli requires initiation factors IF-1, IF-2, & IF-3. IF-3 binds to the 30S ribosomal subunit, freeing it from its complex with the 50S subunit. IF-1 assists binding of IF-3 to the 30S ribosomal subunit. IF-1 also occludes the A site of the small ribosomal subunit, helping insure that the initiation aa-trna fmet-trna fmet can bind only in the P site & that no other aa-trna can bind in the A site during initiation. IF-2 is a small GTP-binding protein. IF-2-GTP binds the initiator fmet-trna fmet & helps it to dock with the small ribosome subunit.
34 As mrna binds, IF-3 helps to correctly position the complex such that the trna fmet interacts via base pairing with the mrna initiation codon (AUG). A region of mrna upstream of the initiation codon, the Shine-Dalgarno sequence, base pairs with the 3' end of the 16S rrna. This positions the 30S ribosomal subunit in relation to the initiation codon. As the large ribosomal subunit joins the complex, GTP on IF-2 is hydrolyzed, leading to dissociation of IF-2-GDP and dissociation of IF-1. A domain of the large ribosomal subunit serves as GAP (GTPase activating protein) for IF-2. Once the two ribosomal subunits come together, the mrna is threaded through a curved channel that wraps around the "neck" region of the small subunit.

35
36 Initiation of translation in eukaryotes Initiation of protein synthesis is much more complex in eukaryotes, & requires a large number of protein factors. Some eukaryotic initiation factors (e.g., eif3 & eif4g) serve as scaffolds, with multiple domains that bind other proteins during assembly of large initiation complexes.
37 Eukaryotic translation factors Factor Function Initiation factors eif-1 Component of the pre-initiation complex eif-1a Component of the pre-initiation complex eif-2 Binds to the initiator trna Met within the ternary complex component of the pre-initiation complex; phosphorylation of eif-2 results in a global repression of translation eif-3 Component of the pre-initiation complex; makes direct contact with eif-4g and so forms the link with the cap binding complex eif-4a eif-4b eif-4e eif-4f eif-4g eif-5 eif-6 Component of the cap binding complex; a helicase that aids scanning by breaking intramolecular base pairs in the mrna Aids scanning, possibly by acting as a helicase that breaks intramolecular base pairs in the mrna Component of the cap binding complex, possibly the component that makes direct contact with the cap structure at the 5 end of the mrna The cap binding complex, comprising eif-4a, eif-4e and eif-4g, which makes the primary contact with the cap structure at the 5 end of the mrna Component of the cap binding complex; forms a bridge between the cap binding complex and eif-3 in the pre-initiation complex; in at least some organisms, eif-4g also forms an association with the poly(a) tail, via the polyadenylate-binding protein Aids release of the other initiation factors at the completion of initiation Associated with the large subunit of the ribosome; prevents large subunits from attaching to small subunits in the cytoplasm Elongation factors eef-1 Complex of four subunits (eef-1a, eef-1b, eef-1d and eef-1g); directs the next trna to its correct position in the ribosome eef-2 Mediates translocation Release factors erf-1 Recognizes the termination codon erf-3 Possibly stimulates dissociation of erf-1 from the ribosome after termination; possibly causes the ribosome subunits to disassociate after termination of translation
38 Initiation of translation in eukaryotes The first step involves assembly of the pre-initiation complex. This structure comprises the 40S subunit of the ribosome, a ternary complex made up of the initiation factor eif-2 bound to the initiator trnamet and a molecule of GTP, and three additional initiation factors, eif- 1, eif-1a, and eif-3. As in bacteria, the initiator trna is distinct from the normal trnamet that recognizes internal 5 -AUG-3 codons but, unlike bacteria, it is aminoacylated with normal methionine, not the formylated version.
39 Initiation of translation in eukaryotes After assembly, the pre-initiation complex associates with the 5 end of the mrna. This step requires the cap binding complex (sometimes called eif-4f), which comprises the initiation factors eif-4a, eif-4e and eif-4g. The contact with the cap might be made by eif-4e alone or might involve a more general interaction with the cap binding complex. The factor eif-4g acts as a bridge between eif-4e, bound to the cap, and eif-3, attached to the pre-initiation complex. The result is that the pre-initiation complex becomes attached to the 5 region of the mrna. Attachment of the pre-initiation complex to the mrna is also influenced by the poly(a) tail, at the distant 3 end of the mrna. This interaction is thought to be mediated by the polyadenylate-binding protein (PADP), which is attached to the poly(a) tail. In yeast and plants it has been shown that PADP can form an association with eif-4g, this association requiring that the mrna bends back on itself. With artificially uncapped mrnas, the PADP interaction is sufficient to load the pre-initiation complex onto the 5 end of the mrna, but under normal circumstances the cap structure and poly(a) tail probably work together. The poly(a) tail could have an important regulatory role, as the length of the tail appears to be correlated with the extent of initiation that occurs with a particular mrna.
40 Initiation of translation in eukaryotes After becoming attached to the 5 end of the mrna, the initiation complex, as it is now called, has to scan along the molecule and find the initiation codon. The leader regions of eukaryotic mrnas can be several tens, or even hundreds, of nucleotides in length and often contain regions that form hairpins and other base-paired structures. These are probably removed by a combination of eif-4a and eif-4b. eif-4a, and possibly also eif-4b, has a helicase activity and so is able to break intramolecular base pairs in the mrna, freeing the passage for the initiation complex. The initiation codon, which is usually 5 -AUG-3 in eukaryotes, is recognizable because it is contained in a short consensus sequence, 5 -ACCAUGG-3, referred to as the Kozak consensus.
41 Initiation of translation in eukaryotes Once the initiation complex is positioned over the initiation codon, the large subunit of the ribosome attaches. As in bacteria, this requires hydrolysis of GTP and leads to release of the initiation factors. Two final initiation factors are involved at this stage: eif-5, which aids release of the other factors, and eif-6, which is associated with the unbound large subunit and prevents it from attaching to a small subunit in the cytoplasm.
42 Initiation of translation in eukaryotes

43 Initiation of translation in eukaryotes Translation initiation in eukaryote. Preinitiation complex scans along mrna until it reaches the initiation codon (a few tens or hundreds nt downstream & located within Kozak consensus sequence) large subunits then attach.
44 Initiation of eukaryotic translation without scanning The scanning system for initiation of translation does not apply to every eukaryotic mrna. This was first recognized with the picornaviruses, a group of viruses with RNA genomes which includes the human poliovirus and rhinovirus, the latter being responsible for the common cold. Transcripts from these viruses are not capped but instead have an internal ribosome entry site (IRES) which is similar in function to the ribosome binding site of bacteria, although the sequences of IRESs and their positions relative to the initiation codon are more variable than the bacterial versions. The presence of IRESs on their transcripts means that picornaviruses can block protein synthesis in the host cell by inactivating the cap binding complex, without affecting translation of their own transcripts, although this is not a normal part of the infection strategy of all picornaviruses. Examples are the mrnas for the mammalian immunoglobulin heavy-chain binding protein and the Drosophila Antennapedia protein
45 Usually a pre-initiation complex forms, including: several initiation factors the small ribosomal subunit the loaded initiator trna, Met-tRNA i Met. This then binds to a separate complex that includes: mrna initiation factors including ones that interact with the 5' methylguanosine cap & the 3' poly-a tail, structures unique to eukaryotic mrna. Within this complex mrna is thought to circularize via interactions between factors that associate with the 5' cap & with a poly-a binding protein..
46 After the initiation complex assembles, it translocates along the mrna in a process called scanning, until the initiation codon is reached. Scanning is facilitated by eukaryotic initiation factor eif4a, which functions as an ATP-dependent helicase to unwind mrna secondary structure while releasing bound proteins. A short sequence of bases adjacent to the AUG initiation codon may aid in recognition of the start site. After the initiation codon is recognized, there is hydrolysis of GTP and release of initiation factors, as the large ribosomal subunit joins the complex and elongation commences.
47 Eukaryotic Translation Translation of mrna is highly regulated in multi-cellular eukaryotic organisms, whereas in prokaryotes regulation occurs mainly at the level of transcription. There is global regulation of protein synthesis. E.g., protein synthesis may be regulated in relation to the cell cycle or in response to cellular stresses such as starvation or accumulation of unfolded proteins in the endoplasmic reticulum. Mechanisms include regulation by signal-activated phosphorylation or dephosphorylation of initiation and elongation factors.
48 Regulation of translation initiation. Global regulation (e.g. under stressful conditions) by eif-2 phosphorylation prevents GTP binding, therefore represses translation; Transcript specific regulation by feedback inhibition or feedback activation mechanisms (left) Figure 13.17a Genomes 3 ( Garland Science 2007)
49 Translation of particular mrnas may be inhibited by small single-stranded microrna molecules about nucleotides long. MicroRNAs bind via base-pairing to 3' un-translated regions of mrna along with a protein complex RISC (RNAinduced silencing complex), inhibiting translation and in some cases promoting mrna degradation. Tissue-specific expression of particular genomeencoded micrornas is an essential regulatory mechanism controlling embryonic development. Some forms of cancer are associated with altered expression of micrornas that regulate synthesis of proteins relevant to cell cycle progression or apoptosis.
50 Elongation of translation Attachment of the large subunit results in two sites at which aminoacyl-trnas can bind. The first of these, the P or peptidyl site, is already occupied by the initiator trna i Met, charged with N-formylmethionine or methionine, and base-paired with the initiation codon. The second site, the A or aminoacyl site, covers the second codon in the open reading frame

51 Elongation of translation
52 Elongation of translation The A site becomes filled with the appropriate aminoacyl-trna, which in E. coli is brought into position by the elongation factor EF-Tu, which ensures that only trnas that carry the correct amino acid are able to enter the ribosome, mischarged trnas being rejected at this point. EF-Tu is an example of a G protein, meaning that it binds a molecule of GTP which it can hydrolyze to release energy. In eukaryotes the equivalent factor is called eef-1, which is a complex of four subunits: eef-1a, eef-1b, eef-1d and eef-1g. The first of these exists in at least two forms, eef-1a1 and eef-1a2, which are highly similar proteins that probably have equivalent functions in different tissues. Specific contacts between the trna, mrna and the 16S rrna within the A site ensure that only the correct trna is accepted. These contacts are able to discriminate between a codon-anticodon interaction in which all three base pairs have formed, and one in which one or more mis-pairs are present, the latter signaling that the wrong trna is present. This is probably just one part of a series of safeguards that ensure the accuracy of the translation process
53 Elongation of translation When the aminoacyl-trna has entered the A site, a peptide bond is formed between the two amino acids. This involves a peptidyl transferase enzyme, which releases the amino acid from the initiator trnai Met and then forms a peptide bond between this amino acid and the one attached to the second trna. In bacteria, the peptidyl transferase activity resides in the 23S rrna of the large subunit, and so is an example of a ribozyme. The reaction is energy dependent and requires hydrolysis of the GTP attached to EF-Tu (eef-1 in eukaryotes). This inactivates EF-Tu, which is ejected from the ribosome and regenerated by EF-Ts. A eukaryotic equivalent of EF-Ts has not been identified, and it is possible that one of the subunits of eef-1 has the regenerative activity
54 P site trna A site trna Transpeptidation O O (peptide bond O P O CH Adenine O P O CH Adenine 2 2 O O formation) involves O H H O H H H H H H nucleophilic attack O OH O OH O C O C of the amino N of HC R HC R the amino acid NH : NH 2 linked to the 3'OH of the terminal O C adenosine of the HC R + NH trna in the A site 3 on the carbonyl C of the amino acid (with attached nascent polypeptide) in ester linkage to the trna in the P site. The reaction is promoted by the geometry of the active site consisting solely of residues of the 23S rrna of the large ribosomal subunit. No protein is found at the active site.
55 O trna O P O O CH 2 H H O O C HC P site NH Adenine O H H OH R O trna O P O O CH 2 O Adenine H H H O H OH O C HC R NH 2 : A site O C HC R NH 3 + The 23S rrna may be considered a "ribozyme." As part of the reaction a proton (H + ) is extracted from the attacking amino N.
56 O trna O P O O P site CH 2 O Adenine H H H OH H OH O trna O P O O A site CH 2 O Adenine H H H O H OH O C HC R NH O C HC NH R O C HC R NH 3 + This H + is then donated to the hydroxyl of the trna in the P site, as the ester linkage is cleaved.
57 It had been proposed that a ring N of a highly conserved adenosine at the active site might act as a catalyst mediating this H + transfer. However, on the basis of recent structural and mutational evidence it has been concluded that the active site adenine is essential only as part of the structure of the active site that positions the substrates correctly. H + shuttling is attributed instead to the adjacent ribose 2' hydroxyl group of the P-site peptidyl-trna, along with ribose hydroxyls of active site rrna residues & structured water molecules that collectively form a H-bonded network at the active site.
58 The nascent polypeptide, one residue longer, is now linked to the A-site trna. O trna O P O O P site CH 2 O Adenine H H H OH H OH O trna O P O A site O CH 2 O Adenine H H H O H OH O C HC R NH O C HC R NH O C HC R NH 3 +
59 Translocation Now the dipeptide corresponding to the first two codons in the open reading frame is attached to the trna in the A site. The next step is translocation, during which three things happen at once: The ribosome moves along three nucleotides, so the next codon enters the A site. The dipeptide-trna in the A site moves to the P site. The deacylated trna in the P site moves to a third position, the E or exit site, in bacteria or, in eukaryotes, is simply ejected from the ribosome.
60 Translocation Translocation requires hydrolysis of a molecule of GTP and is mediated by EF-G in bacteria and by eef-2 in eukaryotes. Electron microscopy of ribosomes at different intermediate stages in translocation suggests that the two subunits rotate slightly in opposite directions, opening up the space between them and enabling the ribosome to slide along the mrna. Translocation results in the A site becoming vacant, allowing a new aminoacyl-trna to enter. The elongation cycle is now repeated, and continues until the end of the open reading frame is reached.

61 trna grey, EF-Tu red, EF-G blue The unloaded trna in the P site will shift to the E (exit) site during translocation. Translocation of the ribosome relative to mrna involves the GTP-binding protein EF-G. The size & shape of EF-G are comparable to that of the complex of EF-Tu with an aa-trna. Structural studies & molecular dynamics indicate that EF-G-GTP binding in the vicinity of the A site causes a ratchet-like motion of the small ribosomal subunit against the large subunit.

62 large subunit trna EF-G small subunit mrna location Figure provided by Dr. J. Frank, Wadsworth Center. The trna with attached nascent polypeptide is pushed from the A site to the P site. Unloaded trna that was in the P site shifts to the E site. Since trnas are linked to mrna by codon-anticodon base pairing, the mrna moves relative to the ribosome.
63 Additionally, it has been postulated that translocation is spontaneous after peptide bond formation because: the deacylated trna in the P site has a higher affinity for the E site, & the peptidyl-trna in the A site has a higher affinity for the P site. Interaction with the ribosome, which acts as GAP (GTPase activating protein) for EF-G, causes EF-G to hydrolyze its bound GTP to GDP + P i. EF-G-GDP then dissociates from the ribosome. A domain of EF-G functions as its own GEF (guanine nucleotide exchange factor) to regenerate EF-G-GTP.
64 The continued codon-anticodon base paring of the trna in the E site is postulated to have a role in preventing potentially serious frame-shift errors, e.g., such as would occur if the trnas were to able to shift laterally by one base pair. Normally the empty trna is released from the E site after binding of the correct aminoacyl-trna at the causes a decreased affinity for trna in the E site. only A site

65 Translation: Elongation Elongation: the ribosome moves down the messenger RNA, adding new amino acids to the growing polypeptide chain. The ribosome has 2 sites for binding transfer RNA. The first RNA with its attached amino acid binds to the first site, and then the transfer RNA corresponding to the second codon bind to the second site.

66 Translation: Elongation The ribosome then removes the amino acid from the first transfer RNA and attaches it to the second amino acid. At this point, the first transfer RNA is empty: no attached amino acid, and the second transfer RNA has a chain of 2 amino acids attached to it.

67 Elongation of the Polypeptide Chain In the elongation stage, amino acids are added one by one to the preceding amino acid TRANSCRIPTION TRANSLATION DNA mrna Ribosome Polypeptide Ribosome ready for next aminoacyl trna Amino end of polypeptide mrna 5 E P A site site 2 3 GTP 1 Codon recognition. The anticodon of an incoming aminoacyl trna base-pairs with the complementary mrna codon in the A site. Hydrolysis of GTP increases the accuracy and efficiency of this step. 2 GDP E E P A P A 3 Translocation. The ribosome translocates the trna in the A site to the P site. The empty trna in the P site is moved to the E site, where it is released. The mrna moves along with its bound trnas, bringing the next codon to be translated into the A site. GDP GTP E P A 2 Peptide bond formation. An rrna molecule of the large subunit catalyzes the formation of a peptide bond between the new amino acid in the A site and the carboxyl end of the growing polypeptide in the P site. This step attaches the polypeptide to the trna in the A site.
68 Frameshifting and other unusual events during elongation The straightforward codon-by-codon translation of an mrna is looked upon as the standard way in which proteins are synthesized. But an increasing number of unusual elongation events are being discovered. One of these is frameshifting, which occurs when a ribosome pauses in the middle of an mrna, moves back one nucleotide or, less frequently, forward one nucleotide, and then continues translation. The result is that the codons that are read after the pause are not contiguous with the preceding set of codons: they lie in a different reading frame. Spontaneous frameshifts occur randomly and are deleterious because the polypeptide synthesized after the frameshift has the incorrect amino acid sequence. But not all frameshifts are spontaneous: a few mrnas utilize programmed frameshifting to induce the ribosome to change frame at a specific point within the transcript.
69 Programmed frameshifting Occurs in all types of organism, from bacteria through to humans, as well as during expression of a number of viral genomes. An example occurs during synthesis of DNA polymerase III in E. coli, the main enzyme involved in replication of DNA. Two of the DNA polymerase III subunits, γ and τ, are coded by a single gene, dnax. Subunit τ is the full-length translation product of the dnax mrna, and subunit γ is a shortened version. Synthesis of γ involves a frameshift in the middle of the dnax mrna, the ribosome encountering a termination codon immediately after the frameshift and so producing the truncated γ version of the translation product.
70 Programmed frameshifting It is thought that the frameshift is induced by three features of the dnax mrna: A hairpin loop, located immediately after the frameshift position, which stalls the ribosome; A sequence similar to a ribosome binding site immediately upstream of the frameshift position, which is thought to base-pair with the 16S rrna (as does an authentic ribosome binding site), again causing the ribosome to stall; The codon 5 -AAG-3 at the frameshift position. The presence of a modified nucleotide at the wobble position of the trna Lys that decodes 5 -AAG-3 means that the codonanticodon interaction is relatively weak at this position, enabling the frameshift to occur.
71 Translational Slippage Enables a single ribosome to translate an mrna that contains copies of two or more genes. This means that, for example, a single ribosome can synthesize each of the five proteins coded by the mrna transcribed from the tryptophan operon of E. coli. When the ribosome reaches the end of one series of codons it releases the protein it has just made, slips to the next initiation codon, and begins synthesizing the next protein.
72 Translational bypassing A form of slippage in which a large part of an mrna is skipped during translation, elongation of the original protein continuing after the bypassing event. The bypass starts and ends either at two identical codons or at two codons that can be translated by the same trna by wobble. This suggests that the jump is controlled by the trna attached to the growing polypeptide, which scans the mrna as the ribosome tracks along, and halts the bypass when a new codon to which it can base-pair is reached.
73 Translational bypassing Translational bypassing of 44 nucleotides occurs in E. coli during translation of the mrna for gene 60 of T4 bacteriophage, which codes for a DNA topoisomerase subunit. Similar events have also been identified in a variety of other bacteria. Bypassing could result in two different proteins being synthesized from one mrna - one protein from normal translation and one from bypassing - but whether this is its general function is not yet known.
74 Frameshifting and other unusual events during elongation
 Growing polypeptides End of mrna (3 end) Completed polypeptide An mrna molecule is generally translated simultaneously by several ribosomes in")
75 Polyribosomes A number of ribosomes can translate a single mrna molecule simultaneously forming a polyribosome Polyribosomes enable a cell to make many copies of a polypeptide very quickly (a) Incoming ribosomal subunits Start of mrna (5 end) Growing polypeptides End of mrna (3 end) Completed polypeptide An mrna molecule is generally translated simultaneously by several ribosomes in clusters called polyribosomes. Ribosomes mrna 0.1 µm This micrograph shows a large polyribosome in a prokaryotic cell (TEM).
76 Termination of translation Protein synthesis ends when one of the three termination codons is reached. The A site is now entered not by a trna but by a protein release factor. Bacteria have three of these: RF-1 which recognizes the termination codons 5 -UAA-3 and 5 -UAG-3, RF-2 which recognizes 5 -UAA-3 and 5 -UGA-3, and RF-3 which stimulates release of RF1 and RF2 from the ribosome after termination, in a reaction requiring energy from the hydrolysis of GTP. Eukaryotes have just two release factors: erf-1, which recognizes the termination codon, and erf-3, which might play the same role as RF-3 although this has not been proven. The structure of erf-1 has been solved by X-ray crystallography, showing that the shape of this protein is very similar to that of a trna. This gives an indication of how the release factor is able to enter the A site when the termination codon is reached.
77 Termination of translation The release factors terminate translation but they do not appear to be responsible for disassociation of the ribosomal subunits, at least not in bacteria. This is the function of an additional protein called ribosome recycling factor (RRF) which, like erf-1, has a trna-like structure. RRF probably enters the P or A site and unlocks the ribosome. Disassociation requires energy, which is released from GTP by EF-G, one of the elongation factors, and also requires the initiation factor IF-3 to prevent the subunits from attaching together again. A eukaryotic equivalent of RRF has not been identified, and this may be one of the functions of erf-3. The disassociated ribosome subunits enter the cytoplasmic pool, where they remain until used again in another round of translation.

78 Termination of translation

79 Post-translational Processing of Proteins
 tendency to form insoluble aggregates; (2) more stable alternative folding")
80 Protein folding Four levels of protein structure; need correct tertiary structure to be activated; a dynamic process; for large proteins, renaturation is not always spontaneous due to (1) tendency to form insoluble aggregates; (2) more stable alternative folding pathways.

81 Protein folding Protein folding is assisted by molecular chaperons (to hold proteins in an open conformation for folding) & chaperonins (a protein complex to promote folding through a cavity & proofread incorrectly folded proteins into correct folding).
 or polyprotein processing (to cut into small pieces of functional proteins).")
82 Proteolytic cleavage Protein cutting is either endprocessing (to cut off N or C terminals to make functional proteins) or polyprotein processing (to cut into small pieces of functional proteins).
83 Proteolytic cleavage An example of endprocessing is pre-pro-insulin. Step 1. Cut off 24 amino acids from N terminal to give pro--insulin; step 2. Cut internal B chain to give insulin.

84 Proteolytic cleavage An example of poly-protein processing used as a way to reduce size of genomes w/ a single gene & 1 promoter & 1 terminator; can be spliced in various ways in different cells.
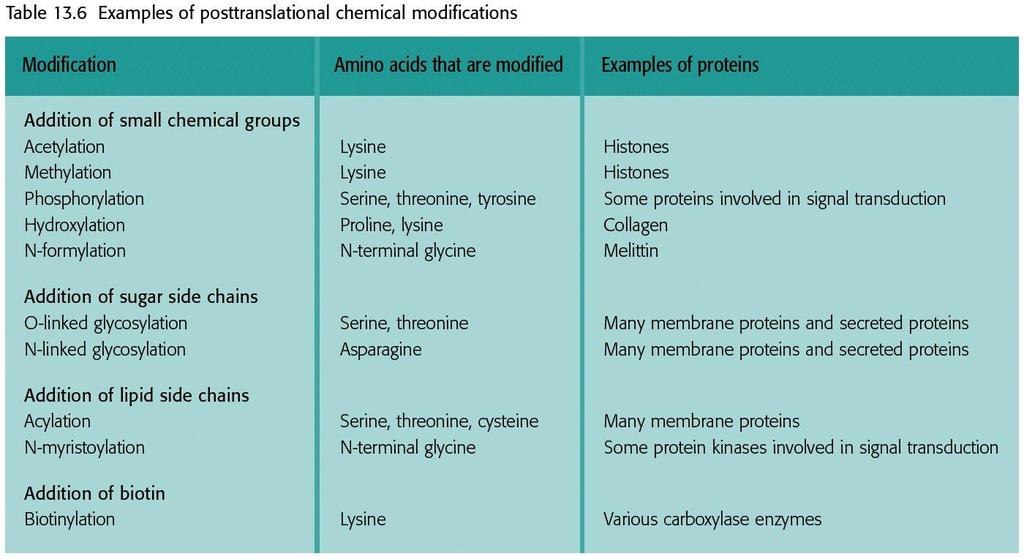
85 Chemical modification
86 Chemical modification More complex modification is glycosylation used to add large carbohydrate side chains to Serine (O-linked) or Asparagine (N-linked).
87 Intein splicing A protein version of RNA splicing (vs. extein); first discovered in yeast in 1990; also found in bacteria & archaea; most ~150 aa & self-catalyzed; intein homing (convert a intein- gene into a intein+ gene; used as a mechanism to propagate).
88 Targeting Polypeptides to Specific Locations Completed proteins are targeted to specific sites in the cell Two populations of ribosomes are evident in cells: free ribsomes (in the cytosol) and bound ribosomes (attached to the ER) Free ribosomes mostly synthesize proteins that function in the cytosol Bound ribosomes make proteins of the endomembrane system and proteins that are secreted from the cell Ribosomes are identical and can switch from free to bound
 binds to the signal peptide The SRP brings the signal peptide and its ribosome to")
89 Targeting Polypeptides to Specific Locations Polypeptide synthesis always begins in the cytosol Synthesis finishes in the cytosol unless the polypeptide signals the ribosome to attach to the ER Polypeptides destined for the ER or for secretion are marked by a signal peptide A signal-recognition particle (SRP) binds to the signal peptide The SRP brings the signal peptide and its ribosome to the ER Ribosomes mrna Signal peptide Signalrecognition particle SRP (SRP) receptor protein CYTOSOL Signal peptide removed ER membrane Protein ER LUMEN Translocation complex
90 Protein Degradation The concept that the proteome of a cell can change over time requires not only de novo protein synthesis but also the removal of proteins whose functions are no longer required. This removal must be highly selective so that only the correct proteins are degraded, and must also be rapid in order to account for the abrupt changes that occur under certain conditions, for example during key transitions in the cell cycle
91 Protein Degradation A link between ubiquitin and protein degradation was first established in 1975 when it was shown that this abundant 76-amino-acid protein is involved in energy-dependent proteolysis reactions in rabbit cells Subsequent research identified a series of three enzymes that attach ubiquitin molecules, singly or in chains, to lysine amino acids in proteins that are targeted for breakdown.
92 Protein Degradation Whether or not a protein becomes ubiquitinated depends on the presence or absence within it of amino acid motifs that act as degradation-susceptibility signals. These signals have not been completely characterized but there are thought to be at least ten different types in S. cerevisiae, including: The N-degron, a sequence element present at the N terminus of a protein; PEST sequences, internal sequences that are rich in proline (P), glutamic acid (E), serine (S) and threonine (T). These sequences are permanent features of the proteins that contain them and so cannot be straightforward degradation signals : if they were then these proteins would be broken down as soon as they are synthesized. Instead, they must determine susceptibility to degradation and hence the general stability of a protein in the cell. How this might be linked to the controlled breakdown of selected proteins at specific times, for instance during the cell cycle, is not yet clear.
93 Protein Degradation The second component of the ubiquitin-dependent degradation pathway is the proteasome, the structure within which ubiquitinated proteins are broken down. In eukaryotes the proteasome is a large, multi-subunit structure with a sedimentation coefficient of 26S, comprising a hollow cylinder of 20S and two caps of 19S. Archaea also have proteasomes of about the same size but these are less complex, being composed of multiple copies of just two proteins; eukaryotic proteasomes contain 14 different types of protein subunit. The entrance into the cavity within the proteasome is narrow, and a protein must be unfolded before it can enter. This unfolding probably occurs through an energy-dependent process and may involve structures similar to chaperonins but with unfolding rather than folding activity. After unfolding, the protein can enter the proteasome within which it is cleaved into short peptides 4 10 amino acids in length. These are released back into the cytoplasm where they are broken down into individual amino acids which can be re-utilized in protein synthesis.

94
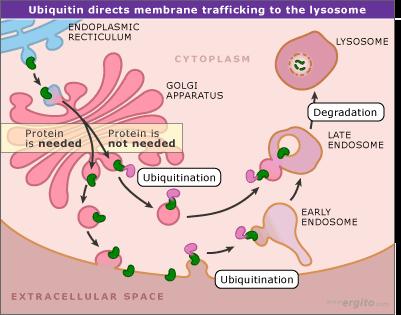
95 Protein Degradation
 is spliced and modified to produce mrna, which moves from the nucleus to the cytoplasm.")
96 A summary of transcription and translation in a eukaryotic cell TRANSCRIPTION 1 RNA is transcribed from a DNA template. 3 DNA 5 RNA transcript RNA polymerase RNA PROCESSING Exon 2 In eukaryotes, the RNA transcript (premrna) is spliced and modified to produce mrna, which moves from the nucleus to the cytoplasm. NUCLEUS RNA transcript (pre-mrna) Intron Aminoacyl-tRNA synthetase CYTOPLASM Amino FORMATION OF acid INITIATION COMPLEX AMINO ACID ACTIVATION trna 3 After leaving the 4 Each amino acid nucleus, mrna attaches attaches to its proper trna to the ribosome. with the help of a specific enzyme and ATP. mrna Growing polypeptide Activated amino acid Ribosomal subunits 5 TRANSLATION Figure E A A A A U G G U U U A U G Ribosome Codon Anticodon 5 A succession of trnas add their amino acids to the polypeptide chain as the mrna is moved through the ribosome one codon at a time. (When completed, the polypeptide is released from the ribosome.)
Molecular Biology (9)
") Molecular Biology (9) Translation Mamoun Ahram, PhD Second semester, 2017-2018 1 Resources This lecture Cooper, Ch. 8 (297-319) 2 General information Protein synthesis involves interactions between three
Molecular Biology (9) Translation Mamoun Ahram, PhD Second semester, 2017-2018 1 Resources This lecture Cooper, Ch. 8 (297-319) 2 General information Protein synthesis involves interactions between three
CHAPTER4 Translation
 CHAPTER4 Translation 4.1 Outline of Translation 4.2 Genetic Code 4.3 trna and Anticodon 4.4 Ribosome 4.5 Protein Synthesis 4.6 Posttranslational Events 4.1 Outline of Translation From mrna to protein
CHAPTER4 Translation 4.1 Outline of Translation 4.2 Genetic Code 4.3 trna and Anticodon 4.4 Ribosome 4.5 Protein Synthesis 4.6 Posttranslational Events 4.1 Outline of Translation From mrna to protein
9 The Process of Translation
 9 The Process of Translation 9.1 Stages of Translation Process We are familiar with the genetic code, we can begin to study the mechanism by which amino acids are assembled into proteins. Because more
9 The Process of Translation 9.1 Stages of Translation Process We are familiar with the genetic code, we can begin to study the mechanism by which amino acids are assembled into proteins. Because more
Gene Expression: Translation. transmission of information from mrna to proteins Chapter 5 slide 1
 Gene Expression: Translation transmission of information from mrna to proteins 601 20000 Chapter 5 slide 1 Fig. 6.1 General structural formula for an amino acid Peter J. Russell, igenetics: Copyright Pearson
Gene Expression: Translation transmission of information from mrna to proteins 601 20000 Chapter 5 slide 1 Fig. 6.1 General structural formula for an amino acid Peter J. Russell, igenetics: Copyright Pearson
Advanced Topics in RNA and DNA. DNA Microarrays Aptamers
 Quiz 1 Advanced Topics in RNA and DNA DNA Microarrays Aptamers 2 Quantifying mrna levels to asses protein expression 3 The DNA Microarray Experiment 4 Application of DNA Microarrays 5 Some applications
Quiz 1 Advanced Topics in RNA and DNA DNA Microarrays Aptamers 2 Quantifying mrna levels to asses protein expression 3 The DNA Microarray Experiment 4 Application of DNA Microarrays 5 Some applications
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
GENETICS - CLUTCH CH.11 TRANSLATION.
 !! www.clutchprep.com CONCEPT: GENETIC CODE Nucleotides and amino acids are translated in a 1 to 1 method The triplet code states that three nucleotides codes for one amino acid - A codon is a term for
!! www.clutchprep.com CONCEPT: GENETIC CODE Nucleotides and amino acids are translated in a 1 to 1 method The triplet code states that three nucleotides codes for one amino acid - A codon is a term for
Protein synthesis II Biochemistry 302. Bob Kelm February 25, 2004
 Protein synthesis II Biochemistry 302 Bob Kelm February 25, 2004 Two idealized views of the 70S ribosomal complex during translation 70S cavity Fig. 27.25 50S tunnel View with 30S subunit in front, 50S
Protein synthesis II Biochemistry 302 Bob Kelm February 25, 2004 Two idealized views of the 70S ribosomal complex during translation 70S cavity Fig. 27.25 50S tunnel View with 30S subunit in front, 50S
-14. -Abdulrahman Al-Hanbali. -Shahd Alqudah. -Dr Ma mon Ahram. 1 P a g e
 -14 -Abdulrahman Al-Hanbali -Shahd Alqudah -Dr Ma mon Ahram 1 P a g e In this lecture we will talk about the last stage in the synthesis of proteins from DNA which is translation. Translation is the process
-14 -Abdulrahman Al-Hanbali -Shahd Alqudah -Dr Ma mon Ahram 1 P a g e In this lecture we will talk about the last stage in the synthesis of proteins from DNA which is translation. Translation is the process
Translation. A ribosome, mrna, and trna.
 Translation The basic processes of translation are conserved among prokaryotes and eukaryotes. Prokaryotic Translation A ribosome, mrna, and trna. In the initiation of translation in prokaryotes, the Shine-Dalgarno
Translation The basic processes of translation are conserved among prokaryotes and eukaryotes. Prokaryotic Translation A ribosome, mrna, and trna. In the initiation of translation in prokaryotes, the Shine-Dalgarno
Section 7. Junaid Malek, M.D.
 Section 7 Junaid Malek, M.D. RNA Processing and Nomenclature For the purposes of this class, please do not refer to anything as mrna that has not been completely processed (spliced, capped, tailed) RNAs
Section 7 Junaid Malek, M.D. RNA Processing and Nomenclature For the purposes of this class, please do not refer to anything as mrna that has not been completely processed (spliced, capped, tailed) RNAs
BCH 4054 Spring 2001 Chapter 33 Lecture Notes
 BCH 4054 Spring 2001 Chapter 33 Lecture Notes Slide 1 The chapter covers degradation of proteins as well. We will not have time to get into that subject. Chapter 33 Protein Synthesis Slide 2 Prokaryotic
BCH 4054 Spring 2001 Chapter 33 Lecture Notes Slide 1 The chapter covers degradation of proteins as well. We will not have time to get into that subject. Chapter 33 Protein Synthesis Slide 2 Prokaryotic
Reading Assignments. A. Genes and the Synthesis of Polypeptides. Lecture Series 7 From DNA to Protein: Genotype to Phenotype
 Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Chapter
 Chapter 17 17.4-17.6 Molecular Components of Translation A cell interprets a genetic message and builds a polypeptide The message is a series of codons on mrna The interpreter is called transfer (trna)
Chapter 17 17.4-17.6 Molecular Components of Translation A cell interprets a genetic message and builds a polypeptide The message is a series of codons on mrna The interpreter is called transfer (trna)
Lecture 9 Translation.
 1 Translation Summary of important events in translation. 2 Translation Reactions involved in peptide bond formation. Lecture 9 3 Genetic code Three types of RNA molecules perform different but complementary
1 Translation Summary of important events in translation. 2 Translation Reactions involved in peptide bond formation. Lecture 9 3 Genetic code Three types of RNA molecules perform different but complementary
TRANSLATION: How to make proteins?
 TRANSLATION: How to make proteins? EUKARYOTIC mrna CBP80 NUCLEUS SPLICEOSOME 5 UTR INTRON 3 UTR m 7 GpppG AUG UAA 5 ss 3 ss CBP20 PABP2 AAAAAAAAAAAAA 50-200 nts CYTOPLASM eif3 EJC PABP1 5 UTR 3 UTR m 7
TRANSLATION: How to make proteins? EUKARYOTIC mrna CBP80 NUCLEUS SPLICEOSOME 5 UTR INTRON 3 UTR m 7 GpppG AUG UAA 5 ss 3 ss CBP20 PABP2 AAAAAAAAAAAAA 50-200 nts CYTOPLASM eif3 EJC PABP1 5 UTR 3 UTR m 7
Translation and the Genetic Code
 Chapter 11. Translation and the Genetic Code 1. Protein Structure 2. Components required for Protein Synthesis 3. Properties of the Genetic Code: An Overview 4. A Degenerate and Ordered Code 1 Sickle-Cell
Chapter 11. Translation and the Genetic Code 1. Protein Structure 2. Components required for Protein Synthesis 3. Properties of the Genetic Code: An Overview 4. A Degenerate and Ordered Code 1 Sickle-Cell
Chapter 19 Overview. Protein Synthesis. for amino acid. n Protein Synthesis genetic info encoded in nucleic acids translated into standard amino acids
 Chapter 19 Overview Protein Synthesis n Protein Synthesis genetic info encoded in nucleic acids translated into standard amino acids n Genetic code dictionary defining meaning for base sequence n Codon
Chapter 19 Overview Protein Synthesis n Protein Synthesis genetic info encoded in nucleic acids translated into standard amino acids n Genetic code dictionary defining meaning for base sequence n Codon
Introduction to the Ribosome Overview of protein synthesis on the ribosome Prof. Anders Liljas
 Introduction to the Ribosome Molecular Biophysics Lund University 1 A B C D E F G H I J Genome Protein aa1 aa2 aa3 aa4 aa5 aa6 aa7 aa10 aa9 aa8 aa11 aa12 aa13 a a 14 How is a polypeptide synthesized? 2
Introduction to the Ribosome Molecular Biophysics Lund University 1 A B C D E F G H I J Genome Protein aa1 aa2 aa3 aa4 aa5 aa6 aa7 aa10 aa9 aa8 aa11 aa12 aa13 a a 14 How is a polypeptide synthesized? 2
Biochemistry Prokaryotic translation
 1 Description of Module Subject Name Paper Name Module Name/Title Dr. Vijaya Khader Dr. MC Varadaraj 2 1. Objectives 2. Understand the concept of genetic code 3. Understand the concept of wobble hypothesis
1 Description of Module Subject Name Paper Name Module Name/Title Dr. Vijaya Khader Dr. MC Varadaraj 2 1. Objectives 2. Understand the concept of genetic code 3. Understand the concept of wobble hypothesis
Information Content in Genetics:
 Information Content in Genetics: DNA, RNA and protein mrna translation into protein (protein synthesis) Francis Crick, 1958 [Crick, F. H. C. in Symp. Soc. Exp. Biol., The Biological Replication of Macromolecules,
Information Content in Genetics: DNA, RNA and protein mrna translation into protein (protein synthesis) Francis Crick, 1958 [Crick, F. H. C. in Symp. Soc. Exp. Biol., The Biological Replication of Macromolecules,
L I F E S C I E N C E S
 1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 NOVEMBER 7, 2006
1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 NOVEMBER 7, 2006
From Gene to Protein
 From Gene to Protein Gene Expression Process by which DNA directs the synthesis of a protein 2 stages transcription translation All organisms One gene one protein 1. Transcription of DNA Gene Composed
From Gene to Protein Gene Expression Process by which DNA directs the synthesis of a protein 2 stages transcription translation All organisms One gene one protein 1. Transcription of DNA Gene Composed
Laith AL-Mustafa. Protein synthesis. Nabil Bashir 10\28\ First
 Laith AL-Mustafa Protein synthesis Nabil Bashir 10\28\2015 http://1drv.ms/1gigdnv 01 First 0 Protein synthesis In previous lectures we started talking about DNA Replication (DNA synthesis) and we covered
Laith AL-Mustafa Protein synthesis Nabil Bashir 10\28\2015 http://1drv.ms/1gigdnv 01 First 0 Protein synthesis In previous lectures we started talking about DNA Replication (DNA synthesis) and we covered
Types of RNA. 1. Messenger RNA(mRNA): 1. Represents only 5% of the total RNA in the cell.
: 1. Represents only 5% of the total RNA in the cell.") RNAs L.Os. Know the different types of RNA & their relative concentration Know the structure of each RNA Understand their functions Know their locations in the cell Understand the differences between prokaryotic
RNAs L.Os. Know the different types of RNA & their relative concentration Know the structure of each RNA Understand their functions Know their locations in the cell Understand the differences between prokaryotic
Translation. Genetic code
 Translation Genetic code If genes are segments of DNA and if DNA is just a string of nucleotide pairs, then how does the sequence of nucleotide pairs dictate the sequence of amino acids in proteins? Simple
Translation Genetic code If genes are segments of DNA and if DNA is just a string of nucleotide pairs, then how does the sequence of nucleotide pairs dictate the sequence of amino acids in proteins? Simple
From gene to protein. Premedical biology
 From gene to protein Premedical biology Central dogma of Biology, Molecular Biology, Genetics transcription replication reverse transcription translation DNA RNA Protein RNA chemically similar to DNA,
From gene to protein Premedical biology Central dogma of Biology, Molecular Biology, Genetics transcription replication reverse transcription translation DNA RNA Protein RNA chemically similar to DNA,
Conceptofcolinearity: a continuous sequence of nucleotides in DNA encodes a continuous sequence of amino acids in a protein
 Translation Conceptofcolinearity: a continuous sequence of nucleotides in DNA encodes a continuous sequence of amino acids in a protein Para além do fenómeno do wobble, há que considerar Desvios ao código
Translation Conceptofcolinearity: a continuous sequence of nucleotides in DNA encodes a continuous sequence of amino acids in a protein Para além do fenómeno do wobble, há que considerar Desvios ao código
BCMB Chapters 39 & 40 Translation (protein synthesis)
") BCMB 3100 - Chapters 39 & 40 Translation (protein synthesis) Translation Genetic code trna Amino acyl trna Ribosomes Initiation Elongation Termination How is the nucleotide code translated into a protein
BCMB 3100 - Chapters 39 & 40 Translation (protein synthesis) Translation Genetic code trna Amino acyl trna Ribosomes Initiation Elongation Termination How is the nucleotide code translated into a protein
BCMB Chapters 39 & 40 Translation (protein synthesis)
") BCMB 3100 - Chapters 39 & 40 Translation (protein synthesis) Translation Genetic code trna Amino acyl trna Ribosomes Initiation Elongation Termination How is the nucleotide code translated into a protein
BCMB 3100 - Chapters 39 & 40 Translation (protein synthesis) Translation Genetic code trna Amino acyl trna Ribosomes Initiation Elongation Termination How is the nucleotide code translated into a protein
Chapter 12. Genes: Expression and Regulation
 Chapter 12 Genes: Expression and Regulation 1 DNA Transcription or RNA Synthesis produces three types of RNA trna carries amino acids during protein synthesis rrna component of ribosomes mrna directs protein
Chapter 12 Genes: Expression and Regulation 1 DNA Transcription or RNA Synthesis produces three types of RNA trna carries amino acids during protein synthesis rrna component of ribosomes mrna directs protein
NO!!!!! BCMB Chapters 39 & 40 Translation (protein synthesis) BCMB Chapters 39 & 40 Translation (protein synthesis)
 BCMB Chapters 39 & 40 Translation (protein synthesis)") BCMB 3100 - Chapters 39 & 40 Translation How is the nucleotide code translated into a protein code? translation DNA RNA protein transcription 5 UCA 3 NH 2 Ser COO -????? Adapter Molecule Hypothesis (Crick,
BCMB 3100 - Chapters 39 & 40 Translation How is the nucleotide code translated into a protein code? translation DNA RNA protein transcription 5 UCA 3 NH 2 Ser COO -????? Adapter Molecule Hypothesis (Crick,
Degeneracy. Two types of degeneracy:
 Degeneracy The occurrence of more than one codon for an amino acid (AA). Most differ in only the 3 rd (3 ) base, with the 1 st and 2 nd being most important for distinguishing the AA. Two types of degeneracy:
Degeneracy The occurrence of more than one codon for an amino acid (AA). Most differ in only the 3 rd (3 ) base, with the 1 st and 2 nd being most important for distinguishing the AA. Two types of degeneracy:
Molecular Biology - Translation of RNA to make Protein *
 OpenStax-CNX module: m49485 1 Molecular Biology - Translation of RNA to make Protein * Jerey Mahr Based on Translation by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative
OpenStax-CNX module: m49485 1 Molecular Biology - Translation of RNA to make Protein * Jerey Mahr Based on Translation by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative
ومن أحياها Translation 1. Translation 1. DONE BY :Maen Faoury
 Translation 1 DONE BY :Maen Faoury 0 1 ومن أحياها Translation 1 2 ومن أحياها Translation 1 In this lecture and the coming lectures you are going to see how the genetic information is transferred into proteins
Translation 1 DONE BY :Maen Faoury 0 1 ومن أحياها Translation 1 2 ومن أحياها Translation 1 In this lecture and the coming lectures you are going to see how the genetic information is transferred into proteins
ومن أحياها Translation 2. Translation 2. DONE BY :Nisreen Obeidat
 Translation 2 DONE BY :Nisreen Obeidat Page 0 Prokaryotes - Shine-Dalgarno Sequence (2:18) What we're seeing here are different portions of sequences of mrna of different promoters from different bacterial
Translation 2 DONE BY :Nisreen Obeidat Page 0 Prokaryotes - Shine-Dalgarno Sequence (2:18) What we're seeing here are different portions of sequences of mrna of different promoters from different bacterial
Lecture 13: PROTEIN SYNTHESIS II- TRANSLATION
 http://smtom.lecture.ub.ac.id/ Password: https://syukur16tom.wordpress.com/ Password: Lecture 13: PROTEIN SYNTHESIS II- TRANSLATION http://hyperphysics.phy-astr.gsu.edu/hbase/organic/imgorg/translation2.gif
http://smtom.lecture.ub.ac.id/ Password: https://syukur16tom.wordpress.com/ Password: Lecture 13: PROTEIN SYNTHESIS II- TRANSLATION http://hyperphysics.phy-astr.gsu.edu/hbase/organic/imgorg/translation2.gif
Molecular Genetics Principles of Gene Expression: Translation
 Paper No. : 16 Module : 13 Principles of gene expression: Translation Development Team Principal Investigator: Prof. Neeta Sehgal Head, Department of Zoology, University of Delhi Paper Coordinator: Prof.
Paper No. : 16 Module : 13 Principles of gene expression: Translation Development Team Principal Investigator: Prof. Neeta Sehgal Head, Department of Zoology, University of Delhi Paper Coordinator: Prof.
GCD3033:Cell Biology. Transcription
 Transcription Transcription: DNA to RNA A) production of complementary strand of DNA B) RNA types C) transcription start/stop signals D) Initiation of eukaryotic gene expression E) transcription factors
Transcription Transcription: DNA to RNA A) production of complementary strand of DNA B) RNA types C) transcription start/stop signals D) Initiation of eukaryotic gene expression E) transcription factors
Lesson Overview. Ribosomes and Protein Synthesis 13.2
 13.2 The Genetic Code The first step in decoding genetic messages is to transcribe a nucleotide base sequence from DNA to mrna. This transcribed information contains a code for making proteins. The Genetic
13.2 The Genetic Code The first step in decoding genetic messages is to transcribe a nucleotide base sequence from DNA to mrna. This transcribed information contains a code for making proteins. The Genetic
Molecular Biology of the Cell
 Alberts Johnson Lewis Morgan Raff Roberts Walter Molecular Biology of the Cell Sixth Edition Chapter 6 (pp. 333-368) How Cells Read the Genome: From DNA to Protein Copyright Garland Science 2015 Genetic
Alberts Johnson Lewis Morgan Raff Roberts Walter Molecular Biology of the Cell Sixth Edition Chapter 6 (pp. 333-368) How Cells Read the Genome: From DNA to Protein Copyright Garland Science 2015 Genetic
Chapter 17. From Gene to Protein. Biology Kevin Dees
 Chapter 17 From Gene to Protein DNA The information molecule Sequences of bases is a code DNA organized in to chromosomes Chromosomes are organized into genes What do the genes actually say??? Reflecting
Chapter 17 From Gene to Protein DNA The information molecule Sequences of bases is a code DNA organized in to chromosomes Chromosomes are organized into genes What do the genes actually say??? Reflecting
Chapter 17 The Mechanism of Translation I: Initiation
 Chapter 17 The Mechanism of Translation I: Initiation Focus only on experiments discussed in class. Completely skip Figure 17.36 Read pg 521-527 up to the sentence that begins "In 1969, Joan Steitz..."
Chapter 17 The Mechanism of Translation I: Initiation Focus only on experiments discussed in class. Completely skip Figure 17.36 Read pg 521-527 up to the sentence that begins "In 1969, Joan Steitz..."
ATP. P i. trna. 3 Appropriate trna covalently bonds to amino acid, displacing AMP. Computer model Hydrogen bonds
 mino acid attachment site nticodon Hydrogen bonds mino acid T i denosine i i denosine minoacyl-trn synthetase (enzyme) trn 1 ctive site binds the amino acid and T. 2 T loses two groups and bonds to the
mino acid attachment site nticodon Hydrogen bonds mino acid T i denosine i i denosine minoacyl-trn synthetase (enzyme) trn 1 ctive site binds the amino acid and T. 2 T loses two groups and bonds to the
TRANSLATION: How to make proteins?
 TRANSLATION: How to make proteins? EUKARYOTIC mrna CBP80 NUCLEUS SPLICEOSOME 5 UTR INTRON 3 UTR m 7 GpppG AUG UAA 5 ss 3 ss CBP20 PABP2 AAAAAAAAAAAAA 50-200 nts CYTOPLASM eif3 EJC PABP1 5 UTR 3 UTR m 7
TRANSLATION: How to make proteins? EUKARYOTIC mrna CBP80 NUCLEUS SPLICEOSOME 5 UTR INTRON 3 UTR m 7 GpppG AUG UAA 5 ss 3 ss CBP20 PABP2 AAAAAAAAAAAAA 50-200 nts CYTOPLASM eif3 EJC PABP1 5 UTR 3 UTR m 7
Protein synthesis I Biochemistry 302. February 17, 2006
 Protein synthesis I Biochemistry 302 February 17, 2006 Key features and components involved in protein biosynthesis High energy cost (essential metabolic activity of cell Consumes 90% of the chemical energy
Protein synthesis I Biochemistry 302 February 17, 2006 Key features and components involved in protein biosynthesis High energy cost (essential metabolic activity of cell Consumes 90% of the chemical energy
Part IV => DNA and RNA. 4.6 RNA Translation 4.6a Genetic Code 4.6b Translational Machinery
 Part IV => DNA and RNA 4.6 RNA Translation 4.6a Genetic Code 4.6b Translational Machinery Section 4.6a: Genetic Code Synopsis 4.6a - In order to translate the genetic information (or genetic code) carried
Part IV => DNA and RNA 4.6 RNA Translation 4.6a Genetic Code 4.6b Translational Machinery Section 4.6a: Genetic Code Synopsis 4.6a - In order to translate the genetic information (or genetic code) carried
RNA & PROTEIN SYNTHESIS. Making Proteins Using Directions From DNA
 RNA & PROTEIN SYNTHESIS Making Proteins Using Directions From DNA RNA & Protein Synthesis v Nitrogenous bases in DNA contain information that directs protein synthesis v DNA remains in nucleus v in order
RNA & PROTEIN SYNTHESIS Making Proteins Using Directions From DNA RNA & Protein Synthesis v Nitrogenous bases in DNA contain information that directs protein synthesis v DNA remains in nucleus v in order
Organic Chemistry Option II: Chemical Biology
 Organic Chemistry Option II: Chemical Biology Recommended books: Dr Stuart Conway Department of Chemistry, Chemistry Research Laboratory, University of Oxford email: stuart.conway@chem.ox.ac.uk Teaching
Organic Chemistry Option II: Chemical Biology Recommended books: Dr Stuart Conway Department of Chemistry, Chemistry Research Laboratory, University of Oxford email: stuart.conway@chem.ox.ac.uk Teaching
Ribosome readthrough
 Ribosome readthrough Starting from the base PROTEIN SYNTHESIS Eukaryotic translation can be divided into four stages: Initiation, Elongation, Termination and Recycling During translation, the ribosome
Ribosome readthrough Starting from the base PROTEIN SYNTHESIS Eukaryotic translation can be divided into four stages: Initiation, Elongation, Termination and Recycling During translation, the ribosome
Translational Initiation
 Translational Initiation Lecture Outline 1. Process of Initiation. Alternative mechanisms of Initiation 3. Key Experiments on Initiation 4. Regulation of Initiation Translation is a process with three
Translational Initiation Lecture Outline 1. Process of Initiation. Alternative mechanisms of Initiation 3. Key Experiments on Initiation 4. Regulation of Initiation Translation is a process with three
Energy and Cellular Metabolism
 1 Chapter 4 About This Chapter Energy and Cellular Metabolism 2 Energy in biological systems Chemical reactions Enzymes Metabolism Figure 4.1 Energy transfer in the environment Table 4.1 Properties of
1 Chapter 4 About This Chapter Energy and Cellular Metabolism 2 Energy in biological systems Chemical reactions Enzymes Metabolism Figure 4.1 Energy transfer in the environment Table 4.1 Properties of
1. In most cases, genes code for and it is that
 Name Chapter 10 Reading Guide From DNA to Protein: Gene Expression Concept 10.1 Genetics Shows That Genes Code for Proteins 1. In most cases, genes code for and it is that determine. 2. Describe what Garrod
Name Chapter 10 Reading Guide From DNA to Protein: Gene Expression Concept 10.1 Genetics Shows That Genes Code for Proteins 1. In most cases, genes code for and it is that determine. 2. Describe what Garrod
UNIT 6 PART 3 *REGULATION USING OPERONS* Hillis Textbook, CH 11
 UNIT 6 PART 3 *REGULATION USING OPERONS* Hillis Textbook, CH 11 REVIEW: Signals that Start and Stop Transcription and Translation BUT, HOW DO CELLS CONTROL WHICH GENES ARE EXPRESSED AND WHEN? First of
UNIT 6 PART 3 *REGULATION USING OPERONS* Hillis Textbook, CH 11 REVIEW: Signals that Start and Stop Transcription and Translation BUT, HOW DO CELLS CONTROL WHICH GENES ARE EXPRESSED AND WHEN? First of
What is the central dogma of biology?
 Bellringer What is the central dogma of biology? A. RNA DNA Protein B. DNA Protein Gene C. DNA Gene RNA D. DNA RNA Protein Review of DNA processes Replication (7.1) Transcription(7.2) Translation(7.3)
Bellringer What is the central dogma of biology? A. RNA DNA Protein B. DNA Protein Gene C. DNA Gene RNA D. DNA RNA Protein Review of DNA processes Replication (7.1) Transcription(7.2) Translation(7.3)
UNIT 5. Protein Synthesis 11/22/16
 UNIT 5 Protein Synthesis IV. Transcription (8.4) A. RNA carries DNA s instruction 1. Francis Crick defined the central dogma of molecular biology a. Replication copies DNA b. Transcription converts DNA
UNIT 5 Protein Synthesis IV. Transcription (8.4) A. RNA carries DNA s instruction 1. Francis Crick defined the central dogma of molecular biology a. Replication copies DNA b. Transcription converts DNA
Lecture 25: Protein Synthesis Key learning goals: Be able to explain the main stuctural features of ribosomes, and know (roughly) how many DNA and
 how many DNA and") Lecture 25: Protein Synthesis Key learning goals: Be able to explain the main stuctural features of ribosomes, and know (roughly) how many DNA and protein subunits they contain. Understand the main functions
Lecture 25: Protein Synthesis Key learning goals: Be able to explain the main stuctural features of ribosomes, and know (roughly) how many DNA and protein subunits they contain. Understand the main functions
Videos. Bozeman, transcription and translation: https://youtu.be/h3b9arupxzg Crashcourse: Transcription and Translation - https://youtu.
 Translation Translation Videos Bozeman, transcription and translation: https://youtu.be/h3b9arupxzg Crashcourse: Transcription and Translation - https://youtu.be/itsb2sqr-r0 Translation Translation The
Translation Translation Videos Bozeman, transcription and translation: https://youtu.be/h3b9arupxzg Crashcourse: Transcription and Translation - https://youtu.be/itsb2sqr-r0 Translation Translation The
Multiple Choice Review- Eukaryotic Gene Expression
 Multiple Choice Review- Eukaryotic Gene Expression 1. Which of the following is the Central Dogma of cell biology? a. DNA Nucleic Acid Protein Amino Acid b. Prokaryote Bacteria - Eukaryote c. Atom Molecule
Multiple Choice Review- Eukaryotic Gene Expression 1. Which of the following is the Central Dogma of cell biology? a. DNA Nucleic Acid Protein Amino Acid b. Prokaryote Bacteria - Eukaryote c. Atom Molecule
Regulation of Transcription in Eukaryotes
 Regulation of Transcription in Eukaryotes Leucine zipper and helix-loop-helix proteins contain DNA-binding domains formed by dimerization of two polypeptide chains. Different members of each family can
Regulation of Transcription in Eukaryotes Leucine zipper and helix-loop-helix proteins contain DNA-binding domains formed by dimerization of two polypeptide chains. Different members of each family can
Chapters 12&13 Notes: DNA, RNA & Protein Synthesis
 Chapters 12&13 Notes: DNA, RNA & Protein Synthesis Name Period Words to Know: nucleotides, DNA, complementary base pairing, replication, genes, proteins, mrna, rrna, trna, transcription, translation, codon,
Chapters 12&13 Notes: DNA, RNA & Protein Synthesis Name Period Words to Know: nucleotides, DNA, complementary base pairing, replication, genes, proteins, mrna, rrna, trna, transcription, translation, codon,
15.2 Prokaryotic Transcription *
 OpenStax-CNX module: m52697 1 15.2 Prokaryotic Transcription * Shannon McDermott Based on Prokaryotic Transcription by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons
OpenStax-CNX module: m52697 1 15.2 Prokaryotic Transcription * Shannon McDermott Based on Prokaryotic Transcription by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons
Quiz answers. Allele. BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA)
 Lecture 17: The Quiz (and back to Eukaryotic DNA)") BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA) http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Quiz answers Kinase: An enzyme
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA) http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Quiz answers Kinase: An enzyme
BME 5742 Biosystems Modeling and Control
 BME 5742 Biosystems Modeling and Control Lecture 24 Unregulated Gene Expression Model Dr. Zvi Roth (FAU) 1 The genetic material inside a cell, encoded in its DNA, governs the response of a cell to various
BME 5742 Biosystems Modeling and Control Lecture 24 Unregulated Gene Expression Model Dr. Zvi Roth (FAU) 1 The genetic material inside a cell, encoded in its DNA, governs the response of a cell to various
GENE ACTIVITY Gene structure Transcription Transcript processing mrna transport mrna stability Translation Posttranslational modifications
 1 GENE ACTIVITY Gene structure Transcription Transcript processing mrna transport mrna stability Translation Posttranslational modifications 2 DNA Promoter Gene A Gene B Termination Signal Transcription
1 GENE ACTIVITY Gene structure Transcription Transcript processing mrna transport mrna stability Translation Posttranslational modifications 2 DNA Promoter Gene A Gene B Termination Signal Transcription
Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p
 Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p.110-114 Arrangement of information in DNA----- requirements for RNA Common arrangement of protein-coding genes in prokaryotes=
Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p.110-114 Arrangement of information in DNA----- requirements for RNA Common arrangement of protein-coding genes in prokaryotes=
Name: SBI 4U. Gene Expression Quiz. Overall Expectation:
 Gene Expression Quiz Overall Expectation: - Demonstrate an understanding of concepts related to molecular genetics, and how genetic modification is applied in industry and agriculture Specific Expectation(s):
Gene Expression Quiz Overall Expectation: - Demonstrate an understanding of concepts related to molecular genetics, and how genetic modification is applied in industry and agriculture Specific Expectation(s):
Eukaryotic vs. Prokaryotic genes
 BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 18: Eukaryotic genes http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Eukaryotic vs. Prokaryotic genes Like in prokaryotes,
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 18: Eukaryotic genes http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Eukaryotic vs. Prokaryotic genes Like in prokaryotes,
Biophysics Lectures Three and Four
 Biophysics Lectures Three and Four Kevin Cahill cahill@unm.edu http://dna.phys.unm.edu/ 1 The Atoms and Molecules of Life Cells are mostly made from the most abundant chemical elements, H, C, O, N, Ca,
Biophysics Lectures Three and Four Kevin Cahill cahill@unm.edu http://dna.phys.unm.edu/ 1 The Atoms and Molecules of Life Cells are mostly made from the most abundant chemical elements, H, C, O, N, Ca,
Prokaryotic Regulation
 Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
9/2/17. Molecular and Cellular Biology. 3. The Cell From Genes to Proteins. key processes
 Molecular and Cellular Biology Animal Cell ((eukaryotic cell) -----> compare with prokaryotic cell) ENDOPLASMIC RETICULUM (ER) Rough ER Smooth ER Flagellum Nuclear envelope Nucleolus NUCLEUS Chromatin
Molecular and Cellular Biology Animal Cell ((eukaryotic cell) -----> compare with prokaryotic cell) ENDOPLASMIC RETICULUM (ER) Rough ER Smooth ER Flagellum Nuclear envelope Nucleolus NUCLEUS Chromatin
9/11/18. Molecular and Cellular Biology. 3. The Cell From Genes to Proteins. key processes
 Molecular and Cellular Biology Animal Cell ((eukaryotic cell) -----> compare with prokaryotic cell) ENDOPLASMIC RETICULUM (ER) Rough ER Smooth ER Flagellum Nuclear envelope Nucleolus NUCLEUS Chromatin
Molecular and Cellular Biology Animal Cell ((eukaryotic cell) -----> compare with prokaryotic cell) ENDOPLASMIC RETICULUM (ER) Rough ER Smooth ER Flagellum Nuclear envelope Nucleolus NUCLEUS Chromatin
Protein Synthesis. Unit 6 Goal: Students will be able to describe the processes of transcription and translation.
 Protein Synthesis Unit 6 Goal: Students will be able to describe the processes of transcription and translation. Types of RNA Messenger RNA (mrna) makes a copy of DNA, carries instructions for making proteins,
Protein Synthesis Unit 6 Goal: Students will be able to describe the processes of transcription and translation. Types of RNA Messenger RNA (mrna) makes a copy of DNA, carries instructions for making proteins,
(Lys), resulting in translation of a polypeptide without the Lys amino acid. resulting in translation of a polypeptide without the Lys amino acid.
, resulting in translation of a polypeptide without the Lys amino acid. resulting in translation of a polypeptide without the Lys amino acid.") 1. A change that makes a polypeptide defective has been discovered in its amino acid sequence. The normal and defective amino acid sequences are shown below. Researchers are attempting to reproduce the
1. A change that makes a polypeptide defective has been discovered in its amino acid sequence. The normal and defective amino acid sequences are shown below. Researchers are attempting to reproduce the
Computational Biology: Basics & Interesting Problems
 Computational Biology: Basics & Interesting Problems Summary Sources of information Biological concepts: structure & terminology Sequencing Gene finding Protein structure prediction Sources of information
Computational Biology: Basics & Interesting Problems Summary Sources of information Biological concepts: structure & terminology Sequencing Gene finding Protein structure prediction Sources of information
Molecular Biology of the Cell
 Alberts Johnson Lewis Raff Roberts Walter Molecular Biology of the Cell Fifth Edition Chapter 6 How Cells Read the Genome: From DNA to Protein Copyright Garland Science 2008 Figure 6-1 Molecular Biology
Alberts Johnson Lewis Raff Roberts Walter Molecular Biology of the Cell Fifth Edition Chapter 6 How Cells Read the Genome: From DNA to Protein Copyright Garland Science 2008 Figure 6-1 Molecular Biology
Protein Synthesis. Unit 6 Goal: Students will be able to describe the processes of transcription and translation.
 Protein Synthesis Unit 6 Goal: Students will be able to describe the processes of transcription and translation. Protein Synthesis: Protein synthesis uses the information in genes to make proteins. 2 Steps
Protein Synthesis Unit 6 Goal: Students will be able to describe the processes of transcription and translation. Protein Synthesis: Protein synthesis uses the information in genes to make proteins. 2 Steps
Translation Part 2 of Protein Synthesis
 Translation Part 2 of Protein Synthesis IN: How is transcription like making a jello mold? (be specific) What process does this diagram represent? A. Mutation B. Replication C.Transcription D.Translation
Translation Part 2 of Protein Synthesis IN: How is transcription like making a jello mold? (be specific) What process does this diagram represent? A. Mutation B. Replication C.Transcription D.Translation
Gene regulation II Biochemistry 302. Bob Kelm February 28, 2005
 Gene regulation II Biochemistry 302 Bob Kelm February 28, 2005 Catabolic operons: Regulation by multiple signals targeting different TFs Catabolite repression: Activity of lac operon is restricted when
Gene regulation II Biochemistry 302 Bob Kelm February 28, 2005 Catabolic operons: Regulation by multiple signals targeting different TFs Catabolite repression: Activity of lac operon is restricted when
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Protein synthesis I Biochemistry 302. Bob Kelm February 23, 2004
 Protein synthesis I Biochemistry 302 Bob Kelm February 23, 2004 Key features of protein synthesis Energy glutton Essential metabolic activity of the cell. Consumes 90% of the chemical energy (ATP,GTP).
Protein synthesis I Biochemistry 302 Bob Kelm February 23, 2004 Key features of protein synthesis Energy glutton Essential metabolic activity of the cell. Consumes 90% of the chemical energy (ATP,GTP).
Translation and Operons
 Translation and Operons You Should Be Able To 1. Describe the three stages translation. including the movement of trna molecules through the ribosome. 2. Compare and contrast the roles of three different
Translation and Operons You Should Be Able To 1. Describe the three stages translation. including the movement of trna molecules through the ribosome. 2. Compare and contrast the roles of three different
Translation - Prokaryotes
 1 Translation - Prokaryotes Shine-Dalgarno (SD) Sequence rrna 3 -GAUACCAUCCUCCUUA-5 mrna...ggagg..(5-7bp)...aug Influences: Secondary structure!! SD and AUG in unstructured region Start AUG 91% GUG 8 UUG
1 Translation - Prokaryotes Shine-Dalgarno (SD) Sequence rrna 3 -GAUACCAUCCUCCUUA-5 mrna...ggagg..(5-7bp)...aug Influences: Secondary structure!! SD and AUG in unstructured region Start AUG 91% GUG 8 UUG
1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine.
 Protein Synthesis & Mutations RNA 1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine. RNA Contains: 1. Adenine 2.
Protein Synthesis & Mutations RNA 1. Contains the sugar ribose instead of deoxyribose. 2. Single-stranded instead of double stranded. 3. Contains uracil in place of thymine. RNA Contains: 1. Adenine 2.
From DNA to protein, i.e. the central dogma
 From DNA to protein, i.e. the central dogma DNA RNA Protein Biochemistry, chapters1 5 and Chapters 29 31. Chapters 2 5 and 29 31 will be covered more in detail in other lectures. ph, chapter 1, will be
From DNA to protein, i.e. the central dogma DNA RNA Protein Biochemistry, chapters1 5 and Chapters 29 31. Chapters 2 5 and 29 31 will be covered more in detail in other lectures. ph, chapter 1, will be
Ranjit P. Bahadur Assistant Professor Department of Biotechnology Indian Institute of Technology Kharagpur, India. 1 st November, 2013
 Hydration of protein-rna recognition sites Ranjit P. Bahadur Assistant Professor Department of Biotechnology Indian Institute of Technology Kharagpur, India 1 st November, 2013 Central Dogma of life DNA
Hydration of protein-rna recognition sites Ranjit P. Bahadur Assistant Professor Department of Biotechnology Indian Institute of Technology Kharagpur, India 1 st November, 2013 Central Dogma of life DNA
TRANSLATION: How to make proteins?
 TRANSLATION: How to make proteins? EUKARYOTIC mrna CBP80 NUCLEUS SPLICEOSOME 5 UTR INTRON 3 UTR m 7 GpppG AUG UAA 5 ss 3 ss CBP20 PABP2 AAAAAAAAAAAAA 50-200 nts CYTOPLASM eif3 EJC PABP1 5 UTR 3 UTR m 7
TRANSLATION: How to make proteins? EUKARYOTIC mrna CBP80 NUCLEUS SPLICEOSOME 5 UTR INTRON 3 UTR m 7 GpppG AUG UAA 5 ss 3 ss CBP20 PABP2 AAAAAAAAAAAAA 50-200 nts CYTOPLASM eif3 EJC PABP1 5 UTR 3 UTR m 7
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Regulation of gene expression. Premedical - Biology
 Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Controlling Gene Expression
 Controlling Gene Expression Control Mechanisms Gene regulation involves turning on or off specific genes as required by the cell Determine when to make more proteins and when to stop making more Housekeeping
Controlling Gene Expression Control Mechanisms Gene regulation involves turning on or off specific genes as required by the cell Determine when to make more proteins and when to stop making more Housekeeping
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Molecular Biology of the Cell
 Alberts Johnson Lewis Raff Roberts Walter Molecular Biology of the Cell Fifth Edition Chapter 6 How Cells Read the Genome: From DNA to Protein Copyright Garland Science 2008 Figure 6-1 Molecular Biology
Alberts Johnson Lewis Raff Roberts Walter Molecular Biology of the Cell Fifth Edition Chapter 6 How Cells Read the Genome: From DNA to Protein Copyright Garland Science 2008 Figure 6-1 Molecular Biology
Central Dogma. DNA is the genetic material within the nucleus. The process of replication creates new copies of DNA.
 Protein Synthesis Outline Central dogma Genetic code Ribosome Structure and Assembly Mechanics of Protein Synthesis Protein Synthesis in Eukaryotes Inhibitors of Protein Synthesis Postranslation modification
Protein Synthesis Outline Central dogma Genetic code Ribosome Structure and Assembly Mechanics of Protein Synthesis Protein Synthesis in Eukaryotes Inhibitors of Protein Synthesis Postranslation modification
Gene regulation II Biochemistry 302. February 27, 2006
 Gene regulation II Biochemistry 302 February 27, 2006 Molecular basis of inhibition of RNAP by Lac repressor 35 promoter site 10 promoter site CRP/DNA complex 60 Lewis, M. et al. (1996) Science 271:1247
Gene regulation II Biochemistry 302 February 27, 2006 Molecular basis of inhibition of RNAP by Lac repressor 35 promoter site 10 promoter site CRP/DNA complex 60 Lewis, M. et al. (1996) Science 271:1247
CHAPTER 3. Cell Structure and Genetic Control. Chapter 3 Outline
 CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
Honors Biology Reading Guide Chapter 11
 Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
PROTEIN SYNTHESIS: TRANSLATION AND THE GENETIC CODE
 PROTEIN SYNTHESIS: TRANSLATION AND THE GENETIC CODE HLeeYu Jsuico Junsay Department of Chemistry School of Science and Engineering Ateneo de Manila University 1 Nucleic Acids are important for their roles
PROTEIN SYNTHESIS: TRANSLATION AND THE GENETIC CODE HLeeYu Jsuico Junsay Department of Chemistry School of Science and Engineering Ateneo de Manila University 1 Nucleic Acids are important for their roles
7.05 Spring 2004 February 27, Recitation #2
 Recitation #2 Contact Information TA: Victor Sai Recitation: Friday, 3-4pm, 2-132 E-mail: sai@mit.edu ffice ours: Friday, 4-5pm, 2-132 Unit 1 Schedule Recitation/Exam Date Lectures covered Recitation #2
Recitation #2 Contact Information TA: Victor Sai Recitation: Friday, 3-4pm, 2-132 E-mail: sai@mit.edu ffice ours: Friday, 4-5pm, 2-132 Unit 1 Schedule Recitation/Exam Date Lectures covered Recitation #2