Biological response to iron fertilization in the eastern equatorial Pacific (IronEx II). I. Microplankton community abundances and biomass
|
|
|
- Eustace Cole
- 6 years ago
- Views:
Transcription
1 MARINE ECOLOGY PROGRESS SERIES Vol. 201: 27 42, 2000 Published August 9 Mar Ecol Prog Ser Biological response to iron fertilization in the eastern equatorial Pacific (IronEx II). I. Microplankton community abundances and biomass M. R. Landry 1, *, M. E. Ondrusek 2, S. J. Tanner 3, S. L. Brown 1, J. Constantinou 1, R. R. Bidigare 1, 4, K. H. Coale 3, S. Fitzwater 5 1 Department of Oceanography, University of Hawai i at Manoa, Honolulu, Hawai i 96822, USA 2 DOC/NOAA/NESDIS, E/RA30, 5200 Auth Rd. Camp Springs, Maryland 20746, USA 3 Moss Landing Marine Laboratories, PO Box 450, Moss Landing, California 96039, USA 4 Hawai i Institute of Marine Biology, PO Box 1346, Kaneohe, Hawai i 96744, USA 5 MBARI, PO Box 628, Moss Landing, California 95039, USA ABSTRACT: During the IronEx II experiment in the eastern equatorial Pacific (May to June 1995), the response of the microplankton community to mesoscale iron fertilization was followed using a combination of marker-pigment, microscopical and flow cytometric techniques. Phytoplankton standing stock bloomed dramatically over a period of 6 d following 3 iron additions of 2 and 1 nm, respectively. Carbon biomass in the patch increased by a factor of 4, chlorophyll a by about a factor of 16 and diatoms by >70-fold relative to contemporaneous levels in the ambient community. The bloom then plateaued sharply and remained at a more or less constant level for 4 d, despite the addition of more iron (1 nm) and physiological indices (low C:chl a ratio and elevated photochemical quantum efficiency) suggesting that the cells were healthy and growing rapidly. Relatively large pennate diatoms (Nitzschia spp., median length 20 to 24 µm) dominated the patch bloom, with smaller pennate species and centric diatoms declining in relative importance. Heterotrophic bacteria increased at a slow rate (0.08 d 1 ) for >10 d during the experiment, as did heterotrophic nanoflagellates. There were also indications of enhanced cell size, cellular pigment content and possibly growth rates of small phytoplankton. Nonetheless, little difference was observed between the ambient community and the peak patch bloom with respect to the size composition of auto- and heterotrophic populations <10 µm in cell size. The relative constancy of the smaller size fractions, the sharp curtailment of net growth of the bloom after 6 d, and >3-fold increase in large heterotrophic dinoflagellates and ciliates suggest that protistan grazers may have played an active role in controlling the phytoplankton response to increased iron availability. KEY WORDS: Diatom Bloom Phytoplankton pigments Carbon:chlorophyll Size structure Heterotrophic protists Resale or republication not permitted without written consent of the publisher INTRODUCTION The eastern equatorial Pacific is one of 3 major oceanic ecosystems, including the subarctic Pacific and Antarctic circumpolar waters, in which the inability of phytoplankton to fully utilize high ambient levels of macronutrients is believed to be due to iron limitation (Martin 1990, Martin et al. 1991). Consistently high concentrations of macronutrients, low levels of * landry@soest.hawaii.edu phytoplankton chlorophyll, and dominance by autotrophic pico- and nanoplankton are common characteristics of these HNLC (high-nutrient, low-chlorophyll) systems. Initially, the case for iron limitation rested on observations of iron-stimulated chlorophyll increase and nutrient utilization in bottle experiments. Interpretations of these experiments were controversial, however, because bottles excluded important components and processes of the natural system (e.g. large zooplankton, sinking and mixing) and because the responses occurred only after several days of con- Inter-Research 2000
2 28 Mar Ecol Prog Ser 201: 27 42, 2000 tainment with dramatic shifts in community composition to otherwise rare organisms (e.g. Banse 1990, Cullen 1991). Interpretations were further confounded by the fact that phytoplankton growth and nutrient uptake also occurred in control bottles without (intentional) iron additions. The first mesoscale iron-enrichment experiment, IronEx I (October 1993), was a major step in taking the iron hypothesis from discussions of bottle artifacts to unequivocal effects on natural ecosystems. It demonstrated the feasibility of fertilizing a patch of the ocean with a micronutrient, of marking the patch with an inert gaseous tracer (SF 6 ), and of tracking and resampling the patch in time. It also documented significant enhancements in physiological condition, biomass, and productivity of phytoplankton in the enriched patch, thus providing strong supporting evidence for the hypothesis that waters of the equatorial Pacific were iron-limited (Greene et al. 1994, Kolber et al. 1994, Martin et al. 1994). Despite these achievements, many questions remained unresolved by this experiment; in particular, the responses of the system in terms of chlorophyll growth, nutrient utilization and inorganic carbon drawdown were not as dramatic as expected (Martin et al. 1994, Watson et al. 1994). In addition, the patch was prematurely subducted under more buoyant waters, and the potential roles of grazers in modulating the phytoplankton iron response were not addressed (Banse 1995, Cullen 1995). The IronEx II experiment (May to June, 1995) was designed to be a longer and more detailed investigation of geochemical, physiological, and ecological responses to iron enrichment. The massive phytoplankton bloom that resulted provided dramatic and unqualified proof of iron limitation in the equatorial Pacific (Coale et al. 1996), and its success has been the basis for the planning and implementation of similar experiments in other HNLC regions. Although the geochemical results and implications of the IronEx II study have been presented in some detail (e.g. Coale et al. 1996, Cooper et al. 1996, Turner et al. 1996, Bidigare et al. 1999), information on the biological responses has been sparse and in some cases anecdotal. The goal of the present paper is to provide an integrated, quantitative overview of the microplankton community response to iron fertilization from complementary assessments of phytoplankton pigments, flow cytometric and microscopically based population abundances, carbon estimates and community size structure. In the 2 companion papers in this issue, we describe the population and community responses of mesozooplankton (Rollwagen Bollens & Landry 2000, in this issue) and taxa-specific rates of phytoplankton growth and microzooplankton grazing (Landry et al. 2000, in this issue). MATERIALS AND METHODS Experimental design and sampling. The basic designs of the mesoscale iron-enrichment experiments in the eastern equatorial Pacific have been described in detail elsewhere (Martin et al. 1994, Coale et al. 1996). Briefly, the main IronEx II experiment (May to June, 1995) began at about 4 40 S, 105 W with the infusion of 72 km 2 of ocean sea surface with a constant ratio mixture of FeSO 4 (ph 2) and the inert tracer SF 6. A total of 225 kg of iron was added to the area as the ship steamed through a rectangular grid pattern, giving an initial mixed-layer concentration of about 2 nm. This initial fertilization took place between 10:00 h (GMT) on 29 May and 04:00 h on 30 May, the midpoint (19:00 h, Julian Day [JD] 149) being the nominal starting point (t = 0) of the experiment. Two additional iron infusions of about 1 nm each were conducted from 15:00 h to 24:00 h on JD 152 (Day 3) and from 18:00 h on JD 157 to 01:00 h on JD 158 (Day 7). To facilitate navigation for the iron infusions and as a point of reference for repeated sampling of the ironenriched patch, the center of the patch was marked with an instrumented buoy drogued at 15 m depth. A second buoy marked the location of a control site with no iron addition. As indicated by Coale et al. (1996), the patch buoy drifted at an average speed of 2.8 km h 1 throughout the experiment, ending up 1500 km south-southwest of its initial position after 19 d. Distributions of seawater properties related to the patch development (e.g. phytoplankton pigments, SF 6, iron, NO 3 and pco 2 ) were mapped approximately daily from JDs 151 to 159 during nighttime grid surveys using water collected at 6 m depth from the ship s PVC pumping system. This strategy was used to follow the patch through time and to locate the area of strongest patch response relative to the marker buoy for more detailed sampling and experimental work each day. In the present study, discrete seawater samples for population pigment and microscopical analyses were collected in the central region of the patch and near the control buoy using a CTD rosette with 30 l Go-Flo bottles. HPLC pigment analyses. Seawater samples (2.05 l) for pigment analyses were positive-pressure filtered (<0.3 bar) through 25 mm Whatman GF/F glass fiber filters. Size-fractioned pigment samples were prepared by passing 2.05 l samples through a series of 25 mm Gelman in-line filter holders containing Poretics polycarbonate membrane filters of decreasing porosity (18, 10, 5 and 2 and 0.8 µm), followed by a GF/F filter for the smallest fraction. The filters were wrapped in aluminum foil, immediately frozen in liquid nitrogen and subsequently stored at 85 C until analysis. Filters were sonicated (0 C, dark) in 3 ml of 100% acetone
3 Landry et al.: Microplankton response to iron fertilization 29 with 50 µl of canthaxanthin in acetone added as an internal standard and extracted in the dark at 20 C for 24 h. Prior to analysis, pigment extracts were vortexed and centrifuged to remove cellular debris. Subsamples of 200 µl of a mixture of 1.0 ml pigment extract plus 0.3 ml H 2 O were injected into a Spectra- Physics SP8800 HPLC system equipped with autosampler, autosampler cooler (4 C), Dynamax 3 µm C-8 analytical and guard columns, and a Waters 440 absorbance (436 nm) detector. Separations followed the binary solvent protocol described by Goericke & Repeta (1993). Peak identifications were made by comparing the retention times of eluting peaks with those of pure standards and cultured organisms with known pigment composition (Latasa et al. 1996). HPLC analysis provided concentration estimates for chlorophyll a (chl a) as well as group-specific accessory marker pigments such as divinyl chl a (prochlorophytes), peridinin (dinoflagellates), 19 -butanoyloxyfucoxanthin (pelagophytes and prymnesiophytes), fucoxanthin (diatoms, pelagophytes and prymnesiophytes), 19 -hexanoyloxyfucoxanthin (prymnesiophytes), prasinoxanthin (prasinophytes), and zeaxanthin (cyanobacteria and prochlorophytes). These basic pigment data were used to determine the contributions of the different phytoplankton groups to total chl a using the CHEMTAX program developed by Mackey et al. (1996, 1997). Population abundances and biomass. All samples for enumeration of plankton population abundances were taken from the middle of the mixed layer at 15 m depth, typically in the morning after nighttime convective mixing. Samples (3 ml) for analyses of bacteria and pico-phytoplankton were preserved with 0.9% paraformaldehyde (final concentration), frozen in liquid nitrogen and stored at 85 C until analysis (Vaulot et al. 1989). Frozen samples were thawed and stained in the dark with Hoechst (1 µg ml 1 ) for 1 h (Monger & Landry 1993), and subsamples of 100 µl were enumerated with a Coulter EPICS 753 flow cytometer equipped with dual 5W argon lasers, MSDS II automatic sampling, a Biosense flow cell and CICERO Cytomation software. Standard beads (0.57 and 0.98 µm YG Fluoresbrite microspheres for visible excitation and 0.46 µm UV-excitable beads, Polysciences) were added to each sample as a basis for normalizing fluorescence signals. The dual lasers were aligned co-linearly with the first tuned to the UV range at 200 mw to excite Hoechst-stained DNA. The second was tuned to 488 nm at 1.3 W to excite the pigment molecules of autotrophic cells (Landry et al. 1996). Forward angle light scatter (FALS), right angle light scatter (RALS) and 4 fluorescence signals were collected for each cell, stored in list mode files, and analyzed using CYTOPC software (Vaulot 1989). Populations of heterotrophic bacteria (HBACT), Prochlorococcus (PRO), Synechococcus (SYN) and pico-eukaryotic cells (PEUK) were distinguished by characteristic autofluorescence, size (RALS) and presence of DNA. SYN were identified principally from their orange fluorescence (575 ± 40 nm; phycoerythrin), but they were also separate from other populations in scatter plots of red (680 ± 40 nm; chl a) versus blue (450 ± 40 nm; DNA) fluorescence. Among cells of smaller size, PRO were distinguished from HBACT by the presence of red autofluorescence. Eukaryotic algae had substantially larger RALS signals (a function of size and refractive index) and red chlorophyll florescence compared to PRO. Enumerations were corrected for volume analyzed and counting efficiency as a function of cell density using E. coli cells enumerated with an Elzone/Cellscope 80XY particle analyzer. Samples (20 to 50 ml) for analyses of phytoplankton and heterotrophic nanoplankton were preserved with gluteraldehyde (1%), concentrated on 0.2 µm, 25 nm black polycarbonate membrane filters, and stained with DAPI (5 µg ml 1 ) for 3 to 4 min in the final stages of filtration. The filters were mounted on glass slides with immersion oil and frozen at 20 C prior to analysis. At least 2000 cells were enumerated at 400 to 1000 magnification for each slide using a Zeiss or Olympus BH2 epifluorescence microscope. Biovolumes were computed for each enumerated group from the closest geometric shape, the 2 visible (length and width) dimensions of the cells on the filter and the estimated third dimension according to Chavez et al. (1991). Samples (250 ml) for analyses of ciliated protists and >20 µm dinoflagellates were preserved with 5% acid Lugol s fixative and stored in the dark. Subsamples of 100 ml were settled 48 h in Utermöhl chambers, and all of the cells in the sample were counted at 400 using a Zeiss inverted microscope. Biomass estimates and size distributions are based on the measurement of at least 50 cells from each sample. Total abundance and biomass of >20 µm dinoflagellates from Lugol s preserved samples were divided into autotrophic and heterotrophic categories based on the percentages of pigmented and nonpigmented cells in the same size fraction in gluteraldehyde-preserved samples analyzed by epifluorescence microscopy. Cellular carbon contents of diatoms were estimated from biovolumes following Strathmann (1967), with a 0.54 correction factor as suggested by Silver & Gowing (1991) and Chavez et al. (1991). This correction accounts for the volumes of the diatom frustrules and internal vacuoles, which are relatively large with respect to total measured volumes of the slender pennate diatoms that dominated the phytoplankton assemblage. Predicted cellular carbon content from these calculations agreed closely with measured carbon for a culture of Pseudonitzschia sp. with comparable
4 30 Mar Ecol Prog Ser 201: 27 42, 2000 dimensions to equatorial Pacific dominants (S. Tanner pers. obs.). For consistency with previous Joint Global Ocean Flux Study (JGOFS) work in the equatorial Pacific (Verity et al. 1996), we used comparable carbon conversion factors of 220 fg C µm 3 for nanoflagellates (Børshein & Bratbak 1987) and 240 and 140 fg C µm 3 for <20 µm and >20 µm dinoflagellates (Verity et al and Lessard 1991, respectively). On average, the biomass estimates using these factors were slightly higher (4% higher for nanoflagellates and 18% higher for dinoflagellates) than those from the uncorrected Strathmann (1967) equations. Ciliate biovolumes were converted to carbon equivalents using a mean factor of 190 fg C µm 3 for Lugol s preserved cells (Putt & Stoecker 1989). Comparable cell abundances of PEUKs were observed in flow cytometric and microscopical analyses; therefore, abundance estimates from both techniques were converted to carbon using the mean microscopical size measurement and the nanoflagellate conversion factor (2 µm cell diameter, 920 fg C cell 1 ). PRO and SYN were both observed to increase in cellular biovolumes by about 50% during the IronEx II enrichment (Cavender-Bares et al. 1999). To account for these changes, we used carbon estimates of 32 and 101 fg C cell 1, respectively, for PRO and SYN cells in the ambient community (Shalapyonok et al. in press), increasing both by 50% (48 and 152 fg C cell 1 ) following the temporal pattern described by Cavender-Bares et al. (1999) (i.e., highest biovolumes occurred from Days 3 to 6). Given large uncertainties in bacterial carbon conversions and unmeasured cell biovolumes in the present experiment, we report only HBACT abundances. RESULTS Pigment distributions Phytoplankton pigment responses to the ironenriched patch were evident in mapping surveys beginning 1 d after the initial fertilization. Monovinyl chl a (MV chl a), the major chlorophyll molecule in all phytoplankton except PRO, was enriched >2-fold over ambient concentrations (100 ng l 1 ) on JD 151, and it doubled again on JD 152 (Fig. 1). MV chl a also increased by approximately another factor of 2 between JDs 152 and 154, the day following the second iron-enrichment, and reached a maximum subsurface concentration of about 16 ambient levels following the third enrichment. The patch was abandoned to pursue other experimental work after MV chl a had begun to decline on JD 159. Subsequent efforts to relocate the patch on JDs 163 and 166 identified waters with enhanced fluorometic chl a and depleted concentrations of nitrate and CO 2 to the south of the central buoy (see Fig. 2 in Coale et al. 1996), but these distributions were not clearly resolved with the reduced number of samples analyzed for HPLC pigments. Overall, the patch slipped relative to the central marker buoy such that a plume of enhanced phytoplankton pigments extended to the south and west as the waters around the buoy were periodically re-fertilized with iron (Fig. 1). Water-column sampling in the central region of the patch also revealed that the depth distribution of pigment enhanced water expanded as the mixed-layer deepened from 25 to 50 m (Fig. 2). During the peak of the phytoplankton bloom on JDs 157 to 158, the maximum MV chl a concentration was at about 20 m. Algal marker pigments showed different responses to the iron-enrichment. For example, fucoxanthin (FUCO), the dominant accessory pigment in diatoms, was initially present in low concentrations but responded rapidly and ultimately dominated the observed distribution patterns in MV chl a (Figs. 1 & 2). In contrast, 19 - hexanoyloxyfucoxanthin (19 -HEX), a diagnostic pigment for prymnesiophytes and the dominant accessory pigment in the ambient seawater, responded rapidly at first, from <40 to >200 ng l 1, but subsequently declined to a lower level (~140 ng l 1 ) after JD butanoyloxyfucoxanthin (19 -BUT), a major pigment in pelagophytes, increased gradually to about 4 times ambient concentrations by JD 158 (Fig. 2). Divinyl chl a (DV chl a), found only in PRO, showed no consistent pattern in enrichment relative to background concentrations (not shown). However, the accessory pigment zeaxanthin (ZEAX), present in both PRO and SYN, showed declining concentrations in the patch as the bloom of eukaryotic phytoplankton reached its peak (Fig. 2). CHEMTAX analysis of the phytoplankton pigments from the patch surface maps suggested that most of the response in community chl a was attributable to prymnesiophytes initially, but ultimately to diatoms as the bloom developed (Fig. 3). By JD 158, the chl a associated with diatoms had increased by more than 2 orders of magnitude above initial concentrations. Other phytoplankton groups, pelagophytes, dinoflagellates and prasinophytes, increased by 5 during the same period, each accounting for as much or more of total phytoplankton chl a as PRO by the peak of the patch bloom (Fig. 3). Phytoplankton abundance and biomass As noted above, the phytoplankton bloom response was marked by a 16-fold increase in MV chl a over the 5 d period from JDs 150 to 155 (Fig. 4). The corre-













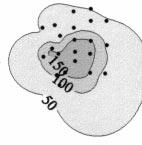
5 Landry et al.: Microplankton response to iron fertilization 31 Fig. 1. Evolution of phytoplankton pigment distributions in surface waters of the IronEx II patch. Distances are relative to the central patch buoy


 from JDs 155 to 158.")


 Fucoxanthin (ng/l) 19")
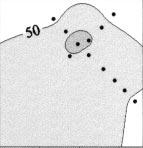
6 32 Mar Ecol Prog Ser 201: 27 42, 2000 sponding increase in microscopically estimated phytoplankton carbon biomass was about a factor of 4 over the same period. Both measures of phytoplankton community biomass remained at more-or-less constant levels (~1600 ng chl a and 120 µg C l 1 ) from JDs 155 to 158. Thus, in terms of net community response, phytoplankton chlorophyll and carbon increased markedly after the first and second iron additions. The third addition had virtually no effect on phytoplankton standing stocks in the patch. MV-Chlorophyll a (ng/l) Fucoxanthin (ng/l) 19 -hexanoyloxyfucoxanthin (ng/l) 19 -butanoyloxyfucoxanthin (ng/l) Zeaxanthin (ng/l) Fig. 2. Time-depth distributions of phytoplankton pigments in the iron-fertilized patch










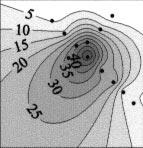

7 Landry et al.: Microplankton response to iron fertilization 33 Fig. 3. Distributions of chlorophyll a (chl a) associated with different phytoplankton groups in surface waters of the iron-fertilized patch. Group-specific chl a based on CHEMTAX analyses of HPLC measured pigments. Distances are relative to the central patch buoy
8 34 Mar Ecol Prog Ser 201: 27 42, 2000 Fig. 4. Phytoplankton chl a and carbon biomass in the ironfertilized patch and in ambient control seawater during the IronEx II experiment. All samples from 15 m depth Pico-phytoplankton populations responded little to iron-enrichment (Fig. 5). Patch abundances of PRO were typically at or below the concentrations in contemporaneous control samples, and they fell somewhat below control estimates at the peak of the bloom (as shown more clearly in the detailed sampling of Cavender-Bares et al. 1999). Given the inferred increases in PRO cell size during the bloom, however, there was little difference between patch and control estimates of population biomass throughout the experiment. SYN abundances in the patch increased comparably to those in the ambient seawater, but the inferred increase in cell size from flowcytometric light scattering suggested a slight (<2 ) enhancement in SYN biomass between JDs 153 and 156. Overall, the abundances and biomass of pico-eukaryotic phytoplankton (PEUK) in the patch were similar to or slightly below those at the control site. Prymnesiophytes, the most abundant of the larger phytoplankton in the ambient community, showed a modest (1.6 ) increase in the patch between JDs 152 and 154. Their abundances were otherwise close to control levels (Fig. 6). The response of dinoflagellates was similar (1.4 increase). Diatoms differed from all other taxa in showing a clear and dramatic response to iron fertilization. Diatom abundances increased by a factor of 30 from initial estimates of 100 to 3000 cells ml 1 at the peak of the bloom. Diatom carbon estimates increased by about 100 to >90 µg C l 1 during the same time period. Whether large or small, all measured accumulations of phytoplankton cells in the patch are, of course, minimal estimates of the taxon s true response to iron-enrichment, since they do not include dilution due to the spatial expansion of the patch or other loss processes. All categories of diatoms enumerated in microscopical analyses increased in absolute abundances and biomass in the iron enriched patch, but the increase was disproportionately greater for pennates (Table 1). Nitzschia bicapitata, the most abundant individual species in control waters, increased by less than a factor of 2 in the patch and decreased markedly in relative importance from 41 to 3% of diatom abundance and from 21 to 0.6% of biomass. Nonetheless, Nitzschia species dominated the bloom overall, accounting for 97% of diatom cell abundance and 70% of peak carbon biomass. Centric diatoms declined in relative importance in the patch, representing 38% of total diatom carbon in ambient waters but only 4% at the peak of the patch bloom. Fig. 5. Abundance and carbon biomass of picophytoplankton in patch and control waters during the IronEx II study
9 Landry et al.: Microplankton response to iron fertilization 35 Fig. 6. Abundance and carbon biomass of prymnesiophytes, pigmented dinoflagellates and diatoms in patch and control waters during the IronEx II study Carbon:chlorophyll ratios The partitioning of chl a among the various phytoplankton groups with the CHEMTAX algorithm (Fig. 7) gives a somewhat different impression of net population changes compared to abundance and carbon biomass estimates (Figs. 5 & 6). For instance, there appeared to be a substantial decrease in SYN chl a in the latter stages of the patch bloom, even though cell-abundances and biomass of SYN were similar to those in ambient waters. Pelagophytes were not enumerated in microscopical analyses, but pelagophyteassociated chl a from CHEMTAX increased markedly with the added iron. Among other eukaryotic taxa, prymnesiophytes and dinoflagellates showed substantially higher enhancement in taxon-specific chl a than noted for cell abundances. However, the dramatic increase in diatom-associated chl a in the patch was consistent with abundance and carbon changes. Fig. 8 shows carbon:chlorophyll ratios (C:chl a, w:w) computed for all control and sampling stations where microscopical estimates of phytoplankton community carbon and CHEMTAX chlorophyll assessments were obtained from the same water bottles or closely spaced hydrocasts. The data suggest that community C:chl a decreased from >150 at the start of the experiment to about 70 at the bloom peak. C:chl a of PRO was generally in the range of 70 to 80, with little or no difference between patch and control water throughout the experiment. Diatom C:chl a declined to similar values during the patch bloom, and also apparently in some control samples. The C:chl a of prymnesiophytes varied from 65 to 85 in the ambient community and from 20 to 36 in the patch, with most of the difference having occurred before the first sampling (JD 150). Patch C:chl a estimates for cyanobacteria and dinoflagellates exhibited more gradual, but substantial, departures from control estimates over the course of the experiment. Dinoflagellate C:chl a was unrealistically high, dropping from an initial value of about 1500 to a low of 270 on JDs 155 and 156. In contrast to estimates for other taxa, the C:chl a ratio for SYN in the patch increased relative to initial and control ratios. This latter trend is the opposite to that indicated by cellular fluorescence from flow cytometric analyses (Fig. 9). While the fluorescence characteristics of PRO cells were entirely consistent with a slight patch increase in cell size with little difference between control and patch C:chl a, fluorescence emissions by SYN pigments (red = chlorophyll, orange = phycoerythrin) showed Table 1. Percentage composition of the diatom community in and out of the iron-fertilized patch. Control estimates are the mean of 3 samplings of the ambient community; Patch estimates are the mean of 4 samplings during peak concentrations in the fertilized patch (Julian Days 155 to 158) Taxon Diatom Abundance (%) Carbon (%) Control Patch Control Patch Nitzschia bicapitata Nitzschia spp Pseudonitzschia spp. < < Tropidoneis/Haslea spp Thallasiothrix spp. < < Other pennates < < Total pennate diatoms Chaetoceros spp Thalasiosira spp. < < Other centrics Total centric diatoms
10 36 Mar Ecol Prog Ser 201: 27 42, 2000 bloom. Dinoflagellate biomass was also enhanced about 2-fold in the patch, although there is some indication that the patch site contained higher concentrations of heterotrophic dinoflagellates initially. Population abundances of heterotrophic bacteria were relatively constant in the ambient environment, varying only between 0.77 and cells ml 1 from JDs 149 to 159 (Fig. 11). In contrast, there was a gradual, but significant bacterial increase in the iron-enriched patch. Cell abundances in the patch reached ml 1 by JD 157. When the patch was revisited 6 d later (JD 163), bacterial densities had increased to > cells ml 1. However, no evidence of enhanced bacterial concentrations was evident at the final sampling on JD 166. Community size structure Fig. 7. Taxon-specific chl a associated with various phytoplankton groups in patch and control waters during the IronEx II study Comparison of phytoplankton community size structure from filter-fractionated significant enhancement in the patch. These observations suggest that C:pigment ratios for SYN should have decreased for cellular carbon increases of <3-fold. Heterotrophic populations Nanoflagellates were numerical dominants in the ambient community of protistan grazers (Fig. 10). However, ciliates and non-pigmented dinoflagellates comprised about half of the biomass on average. Choanoflagellates appeared in low numbers in most samples, and cryptomonads were enumerated occasionally, but neither of these groups accounted for much of the protistan grazer biomass. Within the variability of measurements, heterotrophic nanoflagellates showed little difference in or out of the patch, but a slight increasing trend is suggested from JDs 150 to 163 (Fig. 10). Ciliate abundance and carbon were about 3-fold higher in the patch than in the ambient environment during the peak of the Fig. 8. Estimated C:chl a ratios in patch and control waters during the IronEx II study
11 Landry et al.: Microplankton response to iron fertilization 37 patch) (Fig. 12). The major difference between patch and control assemblages occurred among cells >20 µm in length. Among heterotrophic protists, the 2 to 5 and 5 to 10 µm size categories also remained relatively unchanged in the patch (Fig. 13). Ciliates and dinoflagellates in each of the larger size categories were at least a factor of 2 higher in the patch at the peak of the bloom. In particular, the patch was marked by a significant assemblage of > 40 µm protistan grazers. Net growth rates Fig. 9. Cellular fluorescence characteristics of PRO and SYN in patch and control waters during the IronEx II study. Cell fluorescence from flow cytometric analyses in arbitrary units relative to fluorescent microspheres During the 5 d period of the IronEx II bloom increase (JDs 150 to 155), net growth rates of diatoms surpassed all other measured populations averaging 0.8 d 1 in terms of pigments and cell abundances and 0.62 d 1 as carbon biomass (Table 2). For relevant populations, the pigment data sets, with higher sampling density, typi- pigment samples and microscopical analyses give similar general conclusions <2 µm cells were the most important size category in control waters while larger (>18 µm) phytoplankton, particularly diatoms, dominated in the patch (Fig. 12). Due to unrealistically high estimates of dinoflagellate C:chl a as noted above, the relative contributions of dinoflagellates to community biomass stand out as markedly different in terms of pigments and carbon. Also different in these comparisons is the strong influence of diatoms in all pigment size fractions in the patch. This difference is presumably a consequence of size sorting the microscopical cell counts according to cell length, while the relatively narrow pennate diatoms (or cell fragments and chloroplasts) might easily pass through the membrane filter pores in the pigment sizefractions. Microscopical carbon estimates from patch and control samples showed relatively little difference in biomass structure of phytoplankton size classes <20 µm, and only modest differences in taxonomic composition (e.g. some Phaeocystis spp. and more diatoms in 5 to 20 µm size fractions from the Fig. 10. Abundance and carbon biomass of heterotrophic protists in patch and control waters during the IronEx II study
12 38 Mar Ecol Prog Ser 201: 27 42, 2000 Fig. 11. Abundance of heterotrophic bacteria in patch (closed symbols) and control (open symbols) waters during the IronEx II study cally gave higher rate estimates and narrower confidence intervals than corresponding biomass estimates (except for SYN). Among the autotrophs, only diatoms and total phytoplankton showed significantly positive net growth rates by all variables measured. Among the heterotrophs, only ciliates, bacteria and total heterotrophic biomass increased at statistically significant net rates. Unlike most other populations, which plateaued or declined after the peak bloom biomass on JD 155, bacteria continued to accumulate at a steady rate of 0.08 d 1 until JD 163. Over this extended time period heterotrophic nanoflagellates, which presumably feed on bacteria, also increased at a comparable rate. Fig. 12. Size-structure of the phytoplankton community in patch and control waters during the IronEx II study. Chl a size distributions are from filter fractioned samples on JD 163 (control) and JD 155 (patch). Carbon distributions are the means of 3 microscopically analyzed samples for controls (JDs 152, 157 and 158) and 4 patch samples from the peak of bloom biomass (JDs 155, 156, 157 and 158). PRO: Prochlorococcus spp., CYAN: cyanobacteria, PELAG: pelagophytes, PRYM: prymnesiophytes, DIAT: diatoms, DINO: dinoflagellates, SYN: Synechococcus spp., PEUK: pico-eukaryotes, and PHAEO: Phaeocystis spp. DISCUSSION Possible deficiencies in the CHEMTAX algorithm Microscopical enumeration and pigment biomarkers have both been used extensively for assessing phytoplankton standing stocks in the oceans. Even so, when one considers uncertainties associated with pigmentbiomass ratios and biovolume-biomass conversions, neither is entirely reliable for assessing taxon-specific contributions to total community biomass. Methodological assumptions and uncertainties are particularly problematic under changing environmental conditions, such as the patch boom. Thus, it is reassuring that the inferences about community changes in the present study were in generally good agreement from both approaches. Our results do, however, bring into question possible problems with the CHEMTAX pigment algorithm that may be important when that is used as the main indicator of phytoplankton community structure in an open ocean system (e.g. Meyer- Harms et al. 1999). First, it seems unlikely that the cells counted as autotrophic dinoflagellates had natural C:chl a ratios of 1500 or more. When viewed under epifluorescence microscopy, such cells have a deep red fluorescence indicative of significant chlorophyll content. Bidigare & Ondrusek (1996) and Andersen et al. (1996) have also observed that HPLC pigments underestimate the relative contribution of dinoflagellates compared to microscopical assessments. One possible explanation is that oceanic dinoflagellates and the coastal species used in the algorithm construction differ in accessory pigmentation. There are several distinct pigment types in the dinoflagellate group, including those dominated by peridinin, fucoxanthin or 19 - hexanoyloxyfucoxanthin and those possessing phycobilipigments (Bjørnland & Liaan- Jensen 1989). The dinoflagellate Gymnodinium breve, for example, lacks peridinin and possesses fucoxanthin-related carotenoids (Millie et al. 1993). Chavez et al. (1990) reported that most of the autotrophic dinoflagellates collected from equatorial waters were gymnodinoid in shape. Since it was not possible to distinguish peridinin and non-peridinin containing dinoflagellates from our microscopical counts, we cannot say conclusively that many or most lacked this assumed diagnostic pigment. Nonetheless, it seems likely that this was the case and that much of the chlorophyll residing in dinoflagellates was computationally assigned to other groups.
13 Landry et al.: Microplankton response to iron fertilization 39 Even if not numerically accurate, the patterns in dinoflagellate C:chl a are somewhat satisfying in being consistent with the general declining trend for the phytoplankton community under iron-enrichment. In contrast, the CHEMTAX-based C:chl a trend for SYN (Fig. 8) was at odds with the independent assessment of cellular fluorescence from flow cytometry (Fig. 9). We attribute this discrepancy to SYN being a relatively minor component of the phytoplankton community and sharing an accessory pigment, zeaxanthin (ZEAX), with the more abundant PRO Consequently, small errors in the assignment of DV chl a:zeax to PRO can lead to large errors in the assignment of chl a to SYN in the CHEMTAX program. We are also aware of a case where the application of the CHEMTAX algorithm to a pigment data set from the Southern Ocean suggested a large standing stock of PRO, which were not observed in contemporaneous FCM analyses (R. Letelier pers. com.). In this particular circumstance, direct assessment of DV chl a was unavailable, and the CHEMTAX algorithm was allowed to quantify PRO from other accessory pigments (chl b and ZEAX). The obvious lesson from these examples is that a priori assumptions about the possible taxa present and their diagnostic properties strongly influence the results of the CHEMTAX algorithm. If this tool is to yield meaningful quantitative information on phytoplankton distributions in the oceans, we need to better understand how groupspecific pigment ratios vary with environmental conditions. Fig. 13. Size-structure of the protistan grazer assemblage in patch and control waters during the IronEx II study. Control distributions are the means of 3 microscopically analyzed samples (JDs 152, 157 and 158). Patch distributions are the means of 4 sample analyses during the time of peak patch phytoplankton biomass (JDs 155, 156, 157 and 158) The IronEx II bloom Several complementary lines of evidence, including temporal and spatial evolution of phytoplankton pigment distributions, population abundances, biomass and community size composition, provide a relatively Table 2. Net growth rate summary for autotrophic and heterotrophic populations during the IronEx II bloom. Pigments are from CHEMTAX analyses of group-specific chl a. Abundances and biomass are from flow cytometric or microscopical enumeration and carbon-biovolume conversions. Mean rate estimates are from least squares linear regressions of LN (standing stock) versus Time (Julian Day [JD]). All rates are for the period of fastest bloom response (JDs 150 to 155), unless otherwise noted. 95% confidence limits of mean rate estimates in parentheses Population Net rate of increase (d 1 ) Pigment Abundance Biomass Autotrophic (pigmented) cells Prochlorococcus 0.12 (0.10, 0.15) 0.05 ( 0.04, 0.14) 0.12 (0.01, 0.22) Synechococcus 0.04 ( 0.18, 0.14) 0.17 (0.09, 0.25) 0.24 (0.15, 0.33) Pico-eukaryotes 0.06 ( 0.05, 0.17) 0.15 (0.06, 0.24) Prymnesiophytes 0.16 (0.06, 0.26) 0.05 ( 0.21, 0.32) 0.09 ( 0.17, 0.34) Pelagophytes 0.34 (0.27, 0.41) Dinoflagellates 0.36 (0.25, 0.47) 0.02 ( 0.11, 0.16) 0.08 ( 0.10, 0.26) Diatoms 0.80 (0.76, 0.84) 0.80 (0.55, 1.06) 0.62 (0.20, 1.04) Total phytoplankton 0.43 (0.39, 0.47) 0.30 (0.24, 0.37) Heterotrophic (nonpigmented) cells Nanoflagellates 0.03 ( 0.06, 0.12) 0.05 ( 0.06, 0.15) Choanoflagellates 0.07 ( 0.23, 0.38) 0.24 ( 0.11, 0.59) Dinoflagellates 0.03 ( 0.04, 0.11) 0.02 ( 0.03, 0.07) Ciliates 0.12 (0.05, 0.20) 0.16 (0.03, 0.29) Total heterotrophs 0.05 (0.04, 0.10) Heterotrophic bacteria 0.08 (0.07, 0.09) HBACT (JD ) 0.08 (0.00, 0.15) HFLAG (JD ) 0.08 (0.03, 0.14)
14 40 Mar Ecol Prog Ser 201: 27 42, 2000 coherent picture of the plankton community response to iron fertilization in the HNLC equatorial Pacific. The greening (Barber 1992) of equatorial waters occurred as approximately equal parts of increased phytoplankton biomass and increased pigment content per unit biomass. The latter effect was broadly distributed among the eukaryotic algae. The former was strongly dominated by diatoms, which increased by about 70-fold over a 6 d period, compared to typically less than 2-fold changes for other plankton groups. The diatom response was notable in being comprised largely of pennate forms, similar though somewhat larger than the dominant species in the ambient environment. Centric forms, typically the most important forms in coastal blooms, actually declined in relative importance. Among both autotrophic and heterotrophic protists, iron fertilization had little impact on the composition and biomass structure of <10 µm cells. However, some stimulation of the microbial loop pathway, either directly from iron addition (e.g. Barbeau et al. 1996, Tortell et al. 1996) or indirectly from enhanced availability of dissolved organic substrates, is evident in the gradual increases of HBACT and HFLAG bacterivores. The increasing abundance and biomass of large protistan consumers (heterotrophic dinoflagellates and ciliates) indicate additional food web responses. Several control mechanisms must operate to explain all of the observations described above. First and foremost, the effects of iron are clearly demonstrated by the explosive increase in diatom abundance and biomass following fertilization. Prior to any biomass accumulation, however, the photochemical quantum efficiency of the phytoplankton photosystem II apparatus approximately doubled during the first 24 h after iron addition (Behrenfeld et al. 1996). The decreasing C:chl a ratios in most phytoplankton groups were another likely cellular-level response to increased iron availability. Although centric diatoms were present in the background community in appreciable quantities (10% of diatom abundance and 38% of biomass), their surprising decrease in relative importance during the patch bloom suggests that not all taxa benefited equally from the added iron. The winners, larger versions of the long slender pennate cells that dominate the ambient community, must have enjoyed a competitive advantage over centric cells in iron uptake (high surface area:volume) and growth rates or suffered reduced losses due to zooplankton grazing and cell sinking. Among Nitzschia species, the shift to cells of larger size suggests selective grazing on the smaller forms, which are presumed to have an inherent advantage in nutrient uptake capability and reduced cell sinking. This would imply that protistan grazers, such as the larger ciliates and dinoflagellates that increased during the bloom, may have been significant consumers of diatoms. The relative constancy of small size fractions during the patch bloom provides circumstantial evidence for closely balanced phytoplankton growth and loss processes at lower levels of the food web. Previous experimental studies have indicated that grazer regulation of small phytoplankton is a general feature of open ocean HNLC and oligotrophic regions (Landry et al. 1995, 1997, 1998, Calbet & Landry 1999). The present results suggest that this control mechanism can be remarkably efficient even in the face of substantial environmental perturbation. What is not apparent from the present results, however, is the extent to which grazer regulation was tested by increased growth rates of small phytoplankton taxa in the iron-enriched patch. The present evidence, including size increases of pico-phytoplankton inferred from flow cytometric analyses (Cavender-Bares et al. 1999) and enhanced cellular pigment levels in other taxa, implies some increase in growth rates. At least partial escape of pennate diatoms from grazers and other loss processes is evident in their pronounced biomass accumulation in the iron-fertilized patch. Nonetheless, the leveling off of plankton stocks between JDs 155 and 159 could mean that grazing balance was reestablished after the first 6 d of net positive growth. The alternative explanation is that the bloom was attenuated by the depletion of an essential macroor micro-nutrient (e.g. N, P, Si, Fe, Zn). In this regard, we note that the extended period of peak bloom biomass began 2 d before the third and final iron addition. Microscopical observations during this time showed no indication of phytoplankton physiological stress, and C:chl a ratios remained low through the bloom plateau. More tellingly, Fast-Repetition-Rate fluorometer estimates of quantum efficiency remained 2-fold higher than the ambient community and comparable to measurements on Days 1 to 3 of the experiment (Behrenfeld et al. 1996). Nonetheless, there were some indications that iron may not have been sufficiently available for optimal phytoplankton growth (e.g. Erdner & Anderson 1999), and macro-nutrients were depleted during the bloom from 10 to 6 µm nitrate and from 5 to 1 µm silicate. If micro- or macro-nutrients limited the patch bloom exclusively, we would expect to see evidence of this in sharply reduced phytoplankton growth rates during the stagnant biomass phase. On the other hand, if grazing balance was involved in the control of the bloom peak, we would expect high phytoplankton growth rates to continue through the bloom plateau, with essential nutrients returned to phytoplankton via an efficient remineralization loop (Landry et al. 1997), as well as advective and mixing processes. These issues are explored more thoroughly by Landry et al. (2000).
15 Landry et al.: Microplankton response to iron fertilization 41 In summary, the present study documents a substantial diatom bloom and a shift to larger autotrophic and heterotrophic microplankton in response to iron fertilization of HNLC equatorial waters. The temporal patterns in biomass and size-structure response suggest several ways in which grazing processes and/or nutrient limitation might be implicated in the regulation of phytoplankton stocks at the peak of the IronEx II patch bloom. In companion studies, we investigate the extent to which mesozooplankton populations respond to and contribute to grazing of the patch phytoplankton community (Rollwagen Bollens & Landry 2000). Following that, we provide a dynamic interpretation of phytoplankton growth and microzooplankton grazing rate interactions during the IronEx II bloom (Landry et al. 2000). Acknowledgements. The planning and implementation of the IronEx fertilization experiments required the efforts and logistical support of many. We gratefully acknowledge the inspiration of John Martin and the dedication of all those in the IronEx science team and RV Melville crew who contributed to the success of this study. The present work was supported by US Office of Naval Research Grant N to KHC and by National Science Foundation Grants OCE and to MRL, to RRB and to KHC. Contributions No from the School of Ocean and Earth Science and Technology, University of Hawai i at Manoa, Honolulu, Hawai i 96822, USA, and No. 559 from the US JGOFS Program. LITERATURE CITED Andersen RA, Bidigare RR, Keller MD, Latasa M (1996) A comparison of HPLC pigment signatures and electron microscopic observations for oligotrophic waters of the North Atlantic and Pacific Oceans. Deep-Sea Res II 43: Banse K (1990) Does iron really limit phytoplankton production in the offshore subarctic Pacific? Limnol Oceanogr 35: Banse K (1995) Community response to IRONEX. Nature 375: 112 Barbeau, K, Moffett JW, Caron DA, Croot PL, Erdner DL (1996) Role of protozoan grazing in relieving iron limitation of phytoplankton. Nature 380:61 64 Barber RT (1992) Introduction to the WEC88 cruise: an investigation into why the equator is not greener. J Geophys Res 97: Behrenfeld MJ, Bale AJ, Kolber ZS, Aiken J, Falkowski PG (1996) Confirmation of iron limitation of phytoplankton photosynthesis in the equatorial Pacific Ocean. Nature 383: Bidigare RR, Ondrusek ME (1996) Spatial and temporal variability of phytoplankton pigment distributions in the central equatorial Pacific Ocean. Deep-Sea Res II 43: Bidigare RR, Hanson KL, Buesseler K, Wakeham SG, Freeman KH, Pancost RD, Millero FJ, Steinberg P, Popp BN, Latasa M, Landry MR, Laws EA (1999) Iron-stimulated changes in 13 C fractionation and export by equatorial Pacific phytoplankton: toward a paleo-growth rate proxy. Paleoceanogr 14: Bjørnland T, Liaaen-Jensen S (1989) Distribution of carotenoids in relation to chromophyte phylogen and systematics. In: Green JC, Leadbeater BSC, Diver WL (eds) The chromophyte algae: problems and perspectives. Oxford Science Press, Oxford, p Børsheim KY, Bratbak G (1987) Cell volume to cell carbon conversion factors for a bacterivorous Monas sp. enriched from seawater. Mar Ecol Prog Ser 36: Calbet A, Landry MR (1999) Mesozooplankton influences on the microbial food web: direct and indirect trophic interactions in the oligotrophic open-ocean. Limnol Oceanogr 44: Cavender-Bares KK, Mann EL, Chisholm SW, Ondrusek ME, Bidigare RR (1999) Differential response of equatorial Pacific phytoplankton to iron fertilization. Limnol Oceanogr 44: Chavez FP, Buck KR, Barber RT (1990) Phytoplankton taxa in relation to primary production in the equatorial Pacific. Deep-Sea Res 37: Chavez FP, Buck KR, Coale KH, Martin JH, DiTullio GR, Welschmeyer NA (1991) Growth rates, grazing, sinking, and iron limitation of equatorial Pacific phytoplankton. Limnol Oceanogr 36: Coale KH, Johnson KS, Fitzwater SE, Gordon RM, Tanner S, Chavez FP, Ferioli L, Sakamoto C, Rogers P, Millero F, Steinberg P, Nightingale P, Cooper D, Cochlan WP, Landry MR, Constantinou J, Rollwagen G, Trasvina A, Kudela R (1996) A massive phytoplankton bloom induced by an ecosystem-scale iron fertilization experiment in the equatorial Pacific Ocean. Nature 383: Cooper DJ, Watson AJ, Nightingale RD (1996) Large decrease in ocean-surface CO 2 fugacity in response to in situ iron fertilization. Nature 383: Cullen JJ (1991) Hypotheses to explain high-nutrient conditions in the open sea. Limnol Oceanogr 40: Cullen JJ (1995) Status of the iron hypothesis after the Open- Ocean Enrichment Experiment. Limnol Oceanogr 40: Erdner DL, Anderson DM (1999) Ferredoxin and flavodoxin as biochemical indicators if iron limitation during openocean iron-enrichments. Limnol Oceanogr 44: Goericke R, Repeta DJ (1993) Chlorophylls a and b and divinyl chlorophylls a and b in the open subtropical North Atlantic Ocean. Mar Ecol Prog Ser 101: Greene RM, Kolber ZS, Swift DG, Tindale NW, Falkowski PG (1994) Physiological limitation of phytoplankton photosynthesis in the Eastern Equatorial Pacific determined from variability of the quantum yield of fluorescence. Limnol Oceanogr 39: Kolber ZS, Barber RT, Coale KH, Fitzwater SE, Greene RM, Johnson KS, Lindley S, Falkowski PG (1994) Iron limitation of phytoplankton photosynthesis in the equatorial Pacific Ocean. Nature 731: Landry MR, Constantinou J, Kirshtein J (1995) Microzooplankton grazing in the central equatorial Pacific during February and August Deep-Sea Res II 42: Landry MR, Kirshtein J, Constantinou J (1996) Abundances and distributions of picoplankton populations in the central equatorial Pacific from 12 N to 12 S, 140 W. Deep- Sea Res II 43: Landry MR, Barber RT, Bidigare RR, Chai F, Coale KH, Dam HG, Lewis MR, Lindley ST, McCarthy JJ, Roman MR, Stoecker DK, Verity PG, White JR (1997) Iron and grazing constraints on primary production in the central equatorial Pacific: an EqPac synthesis. Limnol Oceanogr 42:
Microbial Grazers Lab
 Microbial Grazers Lab Objective: Measure the rate at which bacteria are consumed by predators. Overview Size based food webs Microbial loop concepts Bacterial predators Methods to assess microbial grazing
Microbial Grazers Lab Objective: Measure the rate at which bacteria are consumed by predators. Overview Size based food webs Microbial loop concepts Bacterial predators Methods to assess microbial grazing
Microbial Grazers Lab
 Microbial Grazers Lab Objective: Measure the rate at which bacteria are consumed by predators. Overview Size based food webs Microbial loop concepts acterial predators Methods to assess microbial grazing
Microbial Grazers Lab Objective: Measure the rate at which bacteria are consumed by predators. Overview Size based food webs Microbial loop concepts acterial predators Methods to assess microbial grazing
1/24/2011. Why do we care about biomass? Direct aggregation and pulsed export is also important
 Food web structure is a key determinant on carbon fluxes: 1) Cell size and geometry influence sinking rate 2) Zooplankton repackage material and vertically migrate 3) Small cells support longer food webs
Food web structure is a key determinant on carbon fluxes: 1) Cell size and geometry influence sinking rate 2) Zooplankton repackage material and vertically migrate 3) Small cells support longer food webs
Marine Ecology I: Phytoplankton and Primary production
 Marine Ecology I: Phytoplankton and Primary production Osvaldo Ulloa University of Concepcion, Chile oulloa@profc.udec.cl From SOLAS Science Plan Phytoplankton, biogeochemistry and climate I Uptake (through
Marine Ecology I: Phytoplankton and Primary production Osvaldo Ulloa University of Concepcion, Chile oulloa@profc.udec.cl From SOLAS Science Plan Phytoplankton, biogeochemistry and climate I Uptake (through
The effects of iron fertilization on carbon sequestration in the Southern Ocean
 The effects of iron fertilization on carbon sequestration in the Southern Ocean Ken O. Buesseler, John E. Andrews, Steven M. Pike and Matthew A. Charette Department of Marine Chemistry and Geochemistry
The effects of iron fertilization on carbon sequestration in the Southern Ocean Ken O. Buesseler, John E. Andrews, Steven M. Pike and Matthew A. Charette Department of Marine Chemistry and Geochemistry
Effects of Long-Term Sample Preservation on Flow Cytometric Analysis of Natural Populations of Pico- and Nanophytoplankton
 Journal of Oceanography, Vol. 62, pp. 903 to 908, 2006 Short Contribution Effects of Long-Term Sample Preservation on Flow Cytometric Analysis of Natural Populations of Pico- and Nanophytoplankton MITSUHIDE
Journal of Oceanography, Vol. 62, pp. 903 to 908, 2006 Short Contribution Effects of Long-Term Sample Preservation on Flow Cytometric Analysis of Natural Populations of Pico- and Nanophytoplankton MITSUHIDE
Size scaling deviation in phytoplankton photosynthesis and the energy flow through a
 ICES CM2004/Q:04 Size scaling deviation in phytoplankton photosynthesis and the energy flow through a coastal ecosystem. Pedro Cermeño, Emilio Marañón, Jaime Rodríguez, Emilio Fernández, Francisco Jiménez
ICES CM2004/Q:04 Size scaling deviation in phytoplankton photosynthesis and the energy flow through a coastal ecosystem. Pedro Cermeño, Emilio Marañón, Jaime Rodríguez, Emilio Fernández, Francisco Jiménez
Fluorometry Project Chlorophyll Temperature Time Series
 Fluorometry Project Ocean Institute + Scripps Institution of Oceanography Chlorophyll Temperature Time Series The California Current Long Term Ecological Research (CCE LTER) Phytoplankton Phytoplankton
Fluorometry Project Ocean Institute + Scripps Institution of Oceanography Chlorophyll Temperature Time Series The California Current Long Term Ecological Research (CCE LTER) Phytoplankton Phytoplankton
Phytoplankton iron limitation in the Humboldt Current and Peru Upwelling
 Limnol. Oceanogr., 47(4),, 997 111, by the American Society of Limnology and Oceanography, Inc. Phytoplankton iron limitation in the Humboldt Current and Peru Upwelling D. A. Hutchins, 1 C. E. Hare, R.
Limnol. Oceanogr., 47(4),, 997 111, by the American Society of Limnology and Oceanography, Inc. Phytoplankton iron limitation in the Humboldt Current and Peru Upwelling D. A. Hutchins, 1 C. E. Hare, R.
Grazing by the calanoid copepod Neocalanus cristatus on the microbial food web in the coastal Gulf of Alaska
 JOURNAL OF PLANKTON RESEARCH j VOLUME 27 j NUMBER 7 j PAGES 647 662 j 25 Grazing by the calanoid copepod Neocalanus cristatus on the microbial food web in the coastal Gulf of Alaska HONGBIN LIU 1, MICHAEL
JOURNAL OF PLANKTON RESEARCH j VOLUME 27 j NUMBER 7 j PAGES 647 662 j 25 Grazing by the calanoid copepod Neocalanus cristatus on the microbial food web in the coastal Gulf of Alaska HONGBIN LIU 1, MICHAEL
manuscript associated with each of your comments are described below. Your comments are in bold
 First of all thank you for your detailed analysis of our article. All your comments and suggestions were taken into account to improve our manuscript. The corrections of the manuscript associated with
First of all thank you for your detailed analysis of our article. All your comments and suggestions were taken into account to improve our manuscript. The corrections of the manuscript associated with
Phytoplankton. Zooplankton. Nutrients
 Phytoplankton Zooplankton Nutrients Patterns of Productivity There is a large Spring Bloom in the North Atlantic (temperate latitudes remember the Gulf Stream!) What is a bloom? Analogy to terrestrial
Phytoplankton Zooplankton Nutrients Patterns of Productivity There is a large Spring Bloom in the North Atlantic (temperate latitudes remember the Gulf Stream!) What is a bloom? Analogy to terrestrial
Composition of ultraphytoplankton in the central North Atlantic
 MARINE ECOLOGY PROGRESS SERIES Mar Ecol Prog Ser Published June 15 Composition of ultraphytoplankton in the central North Atlantic W. K. W. Li Biological Oceanography Division. Bedford Institute of Oceanography,
MARINE ECOLOGY PROGRESS SERIES Mar Ecol Prog Ser Published June 15 Composition of ultraphytoplankton in the central North Atlantic W. K. W. Li Biological Oceanography Division. Bedford Institute of Oceanography,
Flow cytometry and methods to count aquatic viruses and assess viral-induced induced mortality of bacteria
 Viruses Bacteria Flow cytometry and methods to count aquatic viruses and assess viral-induced induced mortality of bacteria Personnic S 1, Duhamel S 1, Sime-Ngando T 2, Domaizon I 1 & Jacquet S 1 (1) UMR
Viruses Bacteria Flow cytometry and methods to count aquatic viruses and assess viral-induced induced mortality of bacteria Personnic S 1, Duhamel S 1, Sime-Ngando T 2, Domaizon I 1 & Jacquet S 1 (1) UMR
Lesson: Primary Production
 Lesson: Primary Production By Keith Meldahl Corresponding to Chapter 14: Primary Producers Microscopic phytoplankton -- tiny single-celled plants that float at the ocean s surface, are the ultimate food
Lesson: Primary Production By Keith Meldahl Corresponding to Chapter 14: Primary Producers Microscopic phytoplankton -- tiny single-celled plants that float at the ocean s surface, are the ultimate food
Taxonomic recognition of plankton using optics
 Taxonomic recognition of plankton using optics Emmanuel Boss, University of Maine With much help from: Mike Sieracki, Bigelow Laboratory Collin Roesler, Bigelow Laboratory Oscar Schofield, Rutgers University
Taxonomic recognition of plankton using optics Emmanuel Boss, University of Maine With much help from: Mike Sieracki, Bigelow Laboratory Collin Roesler, Bigelow Laboratory Oscar Schofield, Rutgers University
(review) Organization of life
 Organization of life") Marine life: the plankton Production & Energy Transfer Part of Chapter 12, Chapter 13 (review) Organization of life Prokaryotes (usually no nucleus simple life forms) Domain Archaea: : most are extremophiles
Marine life: the plankton Production & Energy Transfer Part of Chapter 12, Chapter 13 (review) Organization of life Prokaryotes (usually no nucleus simple life forms) Domain Archaea: : most are extremophiles
EBENE: A JGOFS investigation of plankton variability and trophic interactions in the equatorial Pacific (180 )
") JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 108, NO. C12, 8136, doi:10.1029/2001jc001252, 2003 EBENE: A JGOFS investigation of plankton variability and trophic interactions in the equatorial Pacific (180 ) Robert
JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 108, NO. C12, 8136, doi:10.1029/2001jc001252, 2003 EBENE: A JGOFS investigation of plankton variability and trophic interactions in the equatorial Pacific (180 ) Robert
COMPARISON OF REFRACTIVE INDEX ESTIMATED FROM SINGLE-CELL AND BULK OPTICAL PROPERTIES. St. John s, NF A1C 5S7 Canada
 COMPARISON OF REFRACTIVE INDEX ESTIMATED FROM SINGLE-CELL AND BULK OPTICAL PROPERTIES Rebecca E. Green 1, Heidi M. Sosik 1, Michele D. DuRand 1,2, and Robert J. Olson 1 1 Woods Hole Oceanographic Institution,
COMPARISON OF REFRACTIVE INDEX ESTIMATED FROM SINGLE-CELL AND BULK OPTICAL PROPERTIES Rebecca E. Green 1, Heidi M. Sosik 1, Michele D. DuRand 1,2, and Robert J. Olson 1 1 Woods Hole Oceanographic Institution,
Primary Producers. Key Ideas
 Primary Producers Kelp forests are one of the ocean s most productive habitats. 1 Key Ideas Energy flows through living systems, but matter is recycled. Primary producers (autotrophs) synthesize glucose
Primary Producers Kelp forests are one of the ocean s most productive habitats. 1 Key Ideas Energy flows through living systems, but matter is recycled. Primary producers (autotrophs) synthesize glucose
SEAWIFS VALIDATION AT THE CARIBBEAN TIME SERIES STATION (CATS)
") SEAWIFS VALIDATION AT THE CARIBBEAN TIME SERIES STATION (CATS) Jesús Lee-Borges* and Roy Armstrong Department of Marine Science, University of Puerto Rico at Mayagüez, Mayagüez, Puerto Rico 00708 Fernando
SEAWIFS VALIDATION AT THE CARIBBEAN TIME SERIES STATION (CATS) Jesús Lee-Borges* and Roy Armstrong Department of Marine Science, University of Puerto Rico at Mayagüez, Mayagüez, Puerto Rico 00708 Fernando
Marine Primary Producers. Primary production
 Marine Primary Producers OCN 201 Biology Lecture 4 http://micro.magnet.fsu.edu/moviegallery/ Photo: Fuhrman Lab; University of Southern California Primary production Incorporation of new organic matter
Marine Primary Producers OCN 201 Biology Lecture 4 http://micro.magnet.fsu.edu/moviegallery/ Photo: Fuhrman Lab; University of Southern California Primary production Incorporation of new organic matter
Look For the Following Key Ideas
 Look For the Following Key Ideas * Energy flows through living systems, but matter is recycled. * Primary producers, called autotrophs, synthesize glucose by the process of photosynthesis or chemosynthesis.
Look For the Following Key Ideas * Energy flows through living systems, but matter is recycled. * Primary producers, called autotrophs, synthesize glucose by the process of photosynthesis or chemosynthesis.
MARINE PELAGIC ECOLOGY
 SOLAS SUMMER SCHOOL 2011 Cargèse, Corsica, France (August 29th to Septembre 7th 2011) MARINE PELAGIC ECOLOGY Maurice Levasseur Université Laval (Québec Océan), Québec, Canada Maurice.levasseur@bio.ulaval.ca
SOLAS SUMMER SCHOOL 2011 Cargèse, Corsica, France (August 29th to Septembre 7th 2011) MARINE PELAGIC ECOLOGY Maurice Levasseur Université Laval (Québec Océan), Québec, Canada Maurice.levasseur@bio.ulaval.ca
INSIGHTS INTO PARTICLE FORMATION AND REMINERALIZATION USING THE SHORT-LIVED RADIONUCLIDE, THORUIM-234 LA Woods Hole, MA 02543
 1 1 2 3 4 INSIGHTS INTO PARTICLE FORMATION AND REMINERALIZATION USING THE SHORT-LIVED RADIONUCLIDE, THORUIM-234 Kanchan Maiti 1,2, Claudia R. Benitez-Nelson 3 and Ken O. Buesseler 2 5 6 7 8 9 10 1 Department
1 1 2 3 4 INSIGHTS INTO PARTICLE FORMATION AND REMINERALIZATION USING THE SHORT-LIVED RADIONUCLIDE, THORUIM-234 Kanchan Maiti 1,2, Claudia R. Benitez-Nelson 3 and Ken O. Buesseler 2 5 6 7 8 9 10 1 Department
WHAT ARE PHYTOPLANKTON?
 WHAT ARE PHYTOPLANKTON? Richard Kirby IVONA CETINIĆ NASA GSFC / USRA @teuta PHYTOPLANKTON drifting plants, 1887 DINOFLAGELLATE DIATOM COCCOLITHOPHORE SILICOFLAGELLATE 2 Ljubesic & Bosak GREAT GENETIC DIVERSITY
WHAT ARE PHYTOPLANKTON? Richard Kirby IVONA CETINIĆ NASA GSFC / USRA @teuta PHYTOPLANKTON drifting plants, 1887 DINOFLAGELLATE DIATOM COCCOLITHOPHORE SILICOFLAGELLATE 2 Ljubesic & Bosak GREAT GENETIC DIVERSITY
Microbial food web structure in a changing Arctic
 Microbial food web structure in a changing Arctic Tatiana M Tsagaraki, Jorun K Egge, Gunnar Bratbak, Øystein Leikness, T. Frede Thingstad, Lise Øvreås, Ruth-Anne Sandaa, Elzbieta A. Petelenz-Kurdziel,
Microbial food web structure in a changing Arctic Tatiana M Tsagaraki, Jorun K Egge, Gunnar Bratbak, Øystein Leikness, T. Frede Thingstad, Lise Øvreås, Ruth-Anne Sandaa, Elzbieta A. Petelenz-Kurdziel,
Figure 14 p.385 5/11/2016. Plankton Production and Food Webs. Plankton Production and Food Webs
 Plankton Production and Food Webs (Chapter 12) What is Plankton? Phytoplankton and Zooplankton Food Web: All the feeding relationships of a community including production, consumption, decomposition and
Plankton Production and Food Webs (Chapter 12) What is Plankton? Phytoplankton and Zooplankton Food Web: All the feeding relationships of a community including production, consumption, decomposition and
Tracking El Niño using optical indices of phytoplankton dynamics in the equatorial Pacific
 Abstract Tracking El Niño using optical indices of phytoplankton dynamics in the equatorial Pacific Joel Craig 1, Pete Strutton 2, Wiley Evans 2 1. College of Earth and Atmospheric Science, Georgia Institute
Abstract Tracking El Niño using optical indices of phytoplankton dynamics in the equatorial Pacific Joel Craig 1, Pete Strutton 2, Wiley Evans 2 1. College of Earth and Atmospheric Science, Georgia Institute
Seasonal cycle of phytoplankton community composition off Newport, Oregon, in 2009
 Seasonal cycle of phytoplankton community composition off Newport, Oregon, in 29 Xiuning Du 1, William Peterson 2 1 College of Environmental science and Engineering, Ocean University of China, Qingdao,
Seasonal cycle of phytoplankton community composition off Newport, Oregon, in 29 Xiuning Du 1, William Peterson 2 1 College of Environmental science and Engineering, Ocean University of China, Qingdao,
Chlorophyll determination of microalgae and cyanobacteria
 14th Webinar Some considerations on the determination of an important biological parameter Chlorophyll determination of microalgae and cyanobacteria bbe Moldaenke GmbH, Preetzer Chaussee 177, 24222 Schwentinental,
14th Webinar Some considerations on the determination of an important biological parameter Chlorophyll determination of microalgae and cyanobacteria bbe Moldaenke GmbH, Preetzer Chaussee 177, 24222 Schwentinental,
THE CONTRIBUTION OF PHYTOPLANKTON AND NON-PHYTOPLANKTON PARTICLES TO INHERENT AND APPARENT OPTICAL PROPERTIES IN NEW ENGLAND CONTINENTAL SHELF WATERS
 THE CONTRIBUTION OF PHYTOPLANKTON AND NON-PHYTOPLANKTON PARTICLES TO INHERENT AND APPARENT OPTICAL PROPERTIES IN NEW ENGLAND CONTINENTAL SHELF WATERS ABSTRACT Rebecca E. Green, Heidi M. Sosik, and Robert
THE CONTRIBUTION OF PHYTOPLANKTON AND NON-PHYTOPLANKTON PARTICLES TO INHERENT AND APPARENT OPTICAL PROPERTIES IN NEW ENGLAND CONTINENTAL SHELF WATERS ABSTRACT Rebecca E. Green, Heidi M. Sosik, and Robert
Bottom-up and Top-down Controls on the Microzooplankton. Community in the Sargasso Sea. Megan Wolverton
 Bottom-up and Top-down Controls on the Microzooplankton Community in the Sargasso Sea by Megan Wolverton A Thesis Presented in Partial Fulfillment of the Requirements for the Degree Master of Science Approved
Bottom-up and Top-down Controls on the Microzooplankton Community in the Sargasso Sea by Megan Wolverton A Thesis Presented in Partial Fulfillment of the Requirements for the Degree Master of Science Approved
- vertical and horizontal segregation Univ. Washington - case studies (Fe and N) (10/29/01)
 (10/29/01)") Chapter 10: Biolimiting Elements James W. Murray - vertical and horizontal segregation Univ. Washington - case studies (Fe and N) (10/29/01) By definition, biolimiting elements are those: necessary to
Chapter 10: Biolimiting Elements James W. Murray - vertical and horizontal segregation Univ. Washington - case studies (Fe and N) (10/29/01) By definition, biolimiting elements are those: necessary to
Low temperature constrains growth rates but not short-term ingestion rates of Antarctic ciliates
 Polar Biol (2013) 36:645 659 DOI 10.1007/s00300-013-1291-y ORIGINAL PAPER Low temperature constrains growth rates but not short-term ingestion rates of Antarctic ciliates Julie M. Rose Elizabeth Fitzpatrick
Polar Biol (2013) 36:645 659 DOI 10.1007/s00300-013-1291-y ORIGINAL PAPER Low temperature constrains growth rates but not short-term ingestion rates of Antarctic ciliates Julie M. Rose Elizabeth Fitzpatrick
Parting the Red Seas: The Optics of Red Tides
 Parting the Red Seas: The Optics of Red Tides H.M. Dierssen 1*, Kudela, R.M. 2, Ryan, J.P. 3 1 University of Connecticut, Department of Marine Science, Groton, CT 06340. 2 University of California, Ocean
Parting the Red Seas: The Optics of Red Tides H.M. Dierssen 1*, Kudela, R.M. 2, Ryan, J.P. 3 1 University of Connecticut, Department of Marine Science, Groton, CT 06340. 2 University of California, Ocean
Phytoplankton. The Biological Pump. Nutrient Cycling and the Marine Biological Pump. Phytoplankton and Zooplankton. CSU ATS Sco9 Denning 1
 Nutrient Cycling and the Marine Biological Pump Readings: SelecGons from Williams & Follows (2011) Sabine et al (2004): Ocean Sink for Anthropogenic CO 2 Phytoplankton Diameter: < 1 mm to over 100 mm ConcentraGon:
Nutrient Cycling and the Marine Biological Pump Readings: SelecGons from Williams & Follows (2011) Sabine et al (2004): Ocean Sink for Anthropogenic CO 2 Phytoplankton Diameter: < 1 mm to over 100 mm ConcentraGon:
Primary Productivity. Global Net PP. Chapter 5- The Microbial World. Fill in the blank. Bacteria Unicellular algae Protists The Microbial Loop
 Chapter 5- The Microbial World Bacteria Unicellular algae Protists The Microbial Loop Megaplankton Macroplankton Mesoplankton Microplankton Nanoplankton Picoplankton Fill in the blank A. Femtoplankton
Chapter 5- The Microbial World Bacteria Unicellular algae Protists The Microbial Loop Megaplankton Macroplankton Mesoplankton Microplankton Nanoplankton Picoplankton Fill in the blank A. Femtoplankton
Amanda Murby University of New Hampshire. Cyanobacteria Monitoring and Analysis Workshop June 26, Cyanobacteria. Importance of Toxins and Size
 Amanda Murby University of New Hampshire Cyanobacteria Monitoring and Analysis Workshop June 26, 2013 Cyanobacteria Importance of Toxins and Size Single-cells breaking off of the Microcystis? Aphanizomenon
Amanda Murby University of New Hampshire Cyanobacteria Monitoring and Analysis Workshop June 26, 2013 Cyanobacteria Importance of Toxins and Size Single-cells breaking off of the Microcystis? Aphanizomenon
Does the Iron Cycle Regulate Atmospheric CO2?
 Does the Iron Cycle Regulate Atmospheric CO2? Mick Follows, Dec 2005 http://ocean.mit.edu/~mick What regulates atmospheric CO2 on glacial-interglacial timescales? Role of ocean biology? Does the iron cycle
Does the Iron Cycle Regulate Atmospheric CO2? Mick Follows, Dec 2005 http://ocean.mit.edu/~mick What regulates atmospheric CO2 on glacial-interglacial timescales? Role of ocean biology? Does the iron cycle
FISHERIES AND AQUACULTURE Vol. V Microzooplankton, Key Organisms in the Pelagic Food Web - Albert Calbet and Miquel Alcaraz
 MICROZOOPLANKTON, KEY ORGANISMS IN THE PELAGIC FOOD WEB Albert Calbet and Institut de Ciències del Mar, CSIC, Barcelona, Spain Keywords: microzooplankton, marine, food webs, phytoplankton, grazing, ciliate,
MICROZOOPLANKTON, KEY ORGANISMS IN THE PELAGIC FOOD WEB Albert Calbet and Institut de Ciències del Mar, CSIC, Barcelona, Spain Keywords: microzooplankton, marine, food webs, phytoplankton, grazing, ciliate,
The North Atlantic Bloom: Species composition and vertical fluxes
 The North Atlantic Bloom: Species composition and vertical fluxes T. Rynearson Graduate School of Oceanography, University of Rhode Island North Atlantic-Arctic ecocsystems Develop a process-based understanding
The North Atlantic Bloom: Species composition and vertical fluxes T. Rynearson Graduate School of Oceanography, University of Rhode Island North Atlantic-Arctic ecocsystems Develop a process-based understanding
Observation system for early warning of HAB events
 Observation system for early warning of HAB events Vera L. Trainer, NOAA Fisheries Northwest Fisheries Science Center Marine Biotoxins Program Seattle, Washington, USA Juan de Fuca eddy Regional HAB OOS
Observation system for early warning of HAB events Vera L. Trainer, NOAA Fisheries Northwest Fisheries Science Center Marine Biotoxins Program Seattle, Washington, USA Juan de Fuca eddy Regional HAB OOS
VEGETATION PROCESSES IN THE PELAGIC: A MODEL FOR ECOSYSTEM THEORY
 Colin S. Reynolds VEGETATION PROCESSES IN THE PELAGIC: A MODEL FOR ECOSYSTEM THEORY Introduction (Otto Kinne) Colin S. Reynolds: A Laudatio (William D. Williams) Publisher: Ecology Institute Nordbunte
Colin S. Reynolds VEGETATION PROCESSES IN THE PELAGIC: A MODEL FOR ECOSYSTEM THEORY Introduction (Otto Kinne) Colin S. Reynolds: A Laudatio (William D. Williams) Publisher: Ecology Institute Nordbunte
Substantial energy input to the mesopelagic ecosystem from the seasonal mixed-layer pump
 SUPPLEMENTARY INFORMATION DOI: 10.1038/NGEO2818 Substantial energy input to the mesopelagic ecosystem from the seasonal mixed-layer pump Giorgio Dall Olmo, James Dingle, Luca Polimene, Robert J. W. Brewin
SUPPLEMENTARY INFORMATION DOI: 10.1038/NGEO2818 Substantial energy input to the mesopelagic ecosystem from the seasonal mixed-layer pump Giorgio Dall Olmo, James Dingle, Luca Polimene, Robert J. W. Brewin
Iron biogeochemistry & the HNLC condition. Philip Boyd Institute for Marine & Antarctic Studies
 Iron biogeochemistry & the HNLC condition Philip Boyd Institute for Marine & Antarctic Studies 2014 Outline HNLC waters definition and implications What causes the HNLC condition? Everyday life in HNLC
Iron biogeochemistry & the HNLC condition Philip Boyd Institute for Marine & Antarctic Studies 2014 Outline HNLC waters definition and implications What causes the HNLC condition? Everyday life in HNLC
LESSON THREE Time, Temperature, Chlorophyll a Does sea surface temperature affect chlorophyll a concentrations?
 STUDENT PAGES LESSON THREE A partnership between California Current Ecosystem Long Term Ecological Research (CCE LTER) and Ocean Institute (OI) Beth Simmons, Education and Outreach Coordinator, CCE LTER,
STUDENT PAGES LESSON THREE A partnership between California Current Ecosystem Long Term Ecological Research (CCE LTER) and Ocean Institute (OI) Beth Simmons, Education and Outreach Coordinator, CCE LTER,
Variations in phytoplankton dynamics and primary production associated with ENSO cycle in the western and central equatorial Pacific during
 JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 116,, doi:10.1029/2010jc006845, 2011 Variations in phytoplankton dynamics and primary production associated with ENSO cycle in the western and central equatorial Pacific
JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 116,, doi:10.1029/2010jc006845, 2011 Variations in phytoplankton dynamics and primary production associated with ENSO cycle in the western and central equatorial Pacific
Vancouver Lake Biotic Assessment
 Vancouver Lake Biotic Assessment Washington State University Vancouver Aquatic Ecology Laboratory Dr. Stephen M. Bollens Dr. Gretchen Rollwagen-Bollens Co-Directors Problem: Noxious cyanobacteria blooms
Vancouver Lake Biotic Assessment Washington State University Vancouver Aquatic Ecology Laboratory Dr. Stephen M. Bollens Dr. Gretchen Rollwagen-Bollens Co-Directors Problem: Noxious cyanobacteria blooms
The Microbial World. Microorganisms of the Sea
 The Microbial World Microorganisms of the Sea Microorganisms Smallest, simplest marine organisms. Very important in evolutionary history of life on Earth. Important primary producers. Include prokaryotes
The Microbial World Microorganisms of the Sea Microorganisms Smallest, simplest marine organisms. Very important in evolutionary history of life on Earth. Important primary producers. Include prokaryotes
Carbon and Nutrient Cycles on Continental Shelves - Current Advances and Thoughts for Future Research
 Carbon and Nutrient Cycles on Continental Shelves - Current Advances and Thoughts for Future Research 1. Brief summary of N-based productivity measures. 2. Thoughts for future studies of the ocean margin
Carbon and Nutrient Cycles on Continental Shelves - Current Advances and Thoughts for Future Research 1. Brief summary of N-based productivity measures. 2. Thoughts for future studies of the ocean margin
Silicate to Nitrate Ratio of the Upper Sub-Arctic Pacific and the Bering Sea Basin in Summer: Its Implication for Phytoplankton Dynamics
 Journal of Oceanography, Vol. 57, pp. 253 to 260, 2001 Silicate to Nitrate Ratio of the Upper Sub-Arctic Pacific and the Bering Sea Basin in Summer: Its Implication for Phytoplankton Dynamics ISAO KOIKE*,
Journal of Oceanography, Vol. 57, pp. 253 to 260, 2001 Silicate to Nitrate Ratio of the Upper Sub-Arctic Pacific and the Bering Sea Basin in Summer: Its Implication for Phytoplankton Dynamics ISAO KOIKE*,
for CESM Jessica Luo, Matthew Long, Keith Lindsay, Mike Levy NCAR Climate and Global Dynamics OMWG / BGC Working Group Meeting, Jan 12, 2018
 Constructing a sizestructured plankton model for CESM Jessica Luo, Matthew Long, Keith Lindsay, Mike Levy NCAR Climate and Global Dynamics OMWG / BGC Working Group Meeting, Jan 12, 2018 Challenge: predicting
Constructing a sizestructured plankton model for CESM Jessica Luo, Matthew Long, Keith Lindsay, Mike Levy NCAR Climate and Global Dynamics OMWG / BGC Working Group Meeting, Jan 12, 2018 Challenge: predicting
Picoplankton community structure in the subarctic Pacific Ocean and the Bering Sea during summer 1999
 MARINE ECOLOGY PROGRESS SERIES Vol. 237: 1 14, 2002 Published July 18 Mar Ecol Prog Ser Picoplankton community structure in the subarctic Pacific Ocean and the Bering Sea during summer 1999 Hongbin Liu
MARINE ECOLOGY PROGRESS SERIES Vol. 237: 1 14, 2002 Published July 18 Mar Ecol Prog Ser Picoplankton community structure in the subarctic Pacific Ocean and the Bering Sea during summer 1999 Hongbin Liu
Variable Fluorescence 4/13/10. Fluorescence HEAT
 Fluorescence HEAT Variable Fluorescence 1 F Start with a pulse of weak light--this will cause weak (background) fluorescence and is called the probe flash 0 Time 1 Variable Fluorescence 1 F 0 Time Turn
Fluorescence HEAT Variable Fluorescence 1 F Start with a pulse of weak light--this will cause weak (background) fluorescence and is called the probe flash 0 Time 1 Variable Fluorescence 1 F 0 Time Turn
Biogeochemical modelling and data assimilation: status in Australia
 Biogeochemical modelling and data assimilation: status in Australia Richard Matear, Andrew Lenton, Matt Chamberlain, Mathieu Mongin, Emlyn Jones, Mark Baird www.cmar.csiro.au/staff/oke/ Biogeochemical
Biogeochemical modelling and data assimilation: status in Australia Richard Matear, Andrew Lenton, Matt Chamberlain, Mathieu Mongin, Emlyn Jones, Mark Baird www.cmar.csiro.au/staff/oke/ Biogeochemical
How the Ecumenical Iron Hypothesis Emerged from Ocean Station P Data. Charlie Miller Prof. Emeritus of Oceanography Oregon State University
 How the Ecumenical Iron Hypothesis Emerged from Ocean Station P Data Charlie Miller Prof. Emeritus of Oceanography Oregon State University In High-Nitrate-Low-Chlorophyll oceanic ecosystems, phytoplankton
How the Ecumenical Iron Hypothesis Emerged from Ocean Station P Data Charlie Miller Prof. Emeritus of Oceanography Oregon State University In High-Nitrate-Low-Chlorophyll oceanic ecosystems, phytoplankton
Primary Production using Ocean Color Remote Sensing. Watson Gregg NASA/Global Modeling and Assimilation Office
 Primary Production using Ocean Color Remote Sensing Watson Gregg NASA/Global Modeling and Assimilation Office watson.gregg@nasa.gov Classification of Ocean Color Primary Production Methods Carr, M.-E.,
Primary Production using Ocean Color Remote Sensing Watson Gregg NASA/Global Modeling and Assimilation Office watson.gregg@nasa.gov Classification of Ocean Color Primary Production Methods Carr, M.-E.,
1.3 Scientist in charge of the project (include CV and passport photo): Dr. Michael Landry*
: Dr. Michael Landry*") Application for Consent to Conduct Marine Scientific Research in Areas Under National Jurisdiction of Costa Rica Date (fecha): 10 diciembre 2009 1. General Information 1.1 Cruise name and/or #: MV1007
Application for Consent to Conduct Marine Scientific Research in Areas Under National Jurisdiction of Costa Rica Date (fecha): 10 diciembre 2009 1. General Information 1.1 Cruise name and/or #: MV1007
Parallel Measurements of Light Scattering and Characterization of Marine Particles in Water: An Evaluation of Methodology
 Parallel Measurements of Light Scattering and Characterization of Marine Particles in Water: An Evaluation of Methodology Dariusz Stramski Marine Physical Laboratory Scripps Institution of Oceanography
Parallel Measurements of Light Scattering and Characterization of Marine Particles in Water: An Evaluation of Methodology Dariusz Stramski Marine Physical Laboratory Scripps Institution of Oceanography
Density estimation of plankton size spectra: a reanalysis of IronEx II data
 Density estimation of plankton size spectra: a reanalysis of IronEx II data MARKUS SCHARTAU 1 *, MICHAEL R. LANDRY 2 AND ROBERT A. ARMSTRONG 3 1 GKSS FORSCHUNGSZENTRUM GEESTHACHT, INSTITUT FÜR KÜSTENFORSCHUNG,
Density estimation of plankton size spectra: a reanalysis of IronEx II data MARKUS SCHARTAU 1 *, MICHAEL R. LANDRY 2 AND ROBERT A. ARMSTRONG 3 1 GKSS FORSCHUNGSZENTRUM GEESTHACHT, INSTITUT FÜR KÜSTENFORSCHUNG,
Modeling Marine Microbes: Past, Present and Prospects Mick Follows MIT
 Modeling Marine Microbes: Past, Present and Prospects Mick Follows MIT What do we mean by models? Concepts Statistical relationships Mathematical descriptions Numerical models Why theory and numerical
Modeling Marine Microbes: Past, Present and Prospects Mick Follows MIT What do we mean by models? Concepts Statistical relationships Mathematical descriptions Numerical models Why theory and numerical
Announcements. Third problem site due November 30 Review calculations
 Announcements Geology Primary Fields of Oceanography Chemistry Third problem site due November 30 Review calculations Next Thursday quiz on Kirchman reading (microbes) Moving from physical to biological
Announcements Geology Primary Fields of Oceanography Chemistry Third problem site due November 30 Review calculations Next Thursday quiz on Kirchman reading (microbes) Moving from physical to biological
Improving estimations of phytoplankton class abundances using CHEMTAX
 MARINE ECOLOGY PROGRESS SERIES Vol. 329: 13 21, 27 Published January 11 Mar Ecol Prog Ser Improving estimations of phytoplankton class abundances using CHEMTAX Mikel Latasa* Institut Ciències del Mar (CSIC),
MARINE ECOLOGY PROGRESS SERIES Vol. 329: 13 21, 27 Published January 11 Mar Ecol Prog Ser Improving estimations of phytoplankton class abundances using CHEMTAX Mikel Latasa* Institut Ciències del Mar (CSIC),
Next-generation Imaging Flow Cytometry
 1 4/21/13 Next-generation Imaging Flow Cytometry New instruments, such as the Amnis system, combine flow cytometry with imaging you get the advantage of having a microscope-like image combined with lasers,
1 4/21/13 Next-generation Imaging Flow Cytometry New instruments, such as the Amnis system, combine flow cytometry with imaging you get the advantage of having a microscope-like image combined with lasers,
Life on Earth
 Life on Earth By feeding, i.e. source of energy a) Autotrophs, self-feeding, e.g. plants (phyto-) b) Heterotrophs, eat others, e.g. animals (zoo-) By feeding, i.e. source of energy a) Autotrophs b)
Life on Earth By feeding, i.e. source of energy a) Autotrophs, self-feeding, e.g. plants (phyto-) b) Heterotrophs, eat others, e.g. animals (zoo-) By feeding, i.e. source of energy a) Autotrophs b)
Capturing Evolution and Ecology in a Global Ocean Model Tim Lenton, Stuart Daines, James Clark, Hywel Williams
 Capturing Evolution and Ecology in a Global Ocean Model Tim Lenton, Stuart Daines, James Clark, Hywel Williams College of Life and Environmental Sciences, University of Exeter, UK t.m.lenton@exeter.ac.uk
Capturing Evolution and Ecology in a Global Ocean Model Tim Lenton, Stuart Daines, James Clark, Hywel Williams College of Life and Environmental Sciences, University of Exeter, UK t.m.lenton@exeter.ac.uk
2001 State of the Ocean: Chemical and Biological Oceanographic Conditions in the Newfoundland Region
 Stock Status Report G2-2 (2) 1 State of the Ocean: Chemical and Biological Oceanographic Conditions in the Background The Altantic Zone Monitoring Program (AZMP) was implemented in 1998 with the aim of
Stock Status Report G2-2 (2) 1 State of the Ocean: Chemical and Biological Oceanographic Conditions in the Background The Altantic Zone Monitoring Program (AZMP) was implemented in 1998 with the aim of
Southern Ocean Iron Enrichment Experiment: Carbon Cycling in High- and Low-Si Waters
 28. W. W. Cochran, M. Wikelski, in Birds of Two Worlds, P. Marra, R. Greenberg, Eds. (Johns Hopkins Press, Baltimore, 2004), in press. 29. R. Sandberg, et al., Auk 119, 201 (2002). 30. S. T. Emlen, Anim.
28. W. W. Cochran, M. Wikelski, in Birds of Two Worlds, P. Marra, R. Greenberg, Eds. (Johns Hopkins Press, Baltimore, 2004), in press. 29. R. Sandberg, et al., Auk 119, 201 (2002). 30. S. T. Emlen, Anim.
Pigment Fluorescence Signatures as an Index to the Taxonomic Structure of Phytoplankton Communities
 Pigment Fluorescence Signatures as an Index to the Taxonomic Structure of Phytoplankton Communities Dr. Gary Hitchcock Marine Biology and Fisheries Rosenstiel School of Marine and Atmospheric Science 4600
Pigment Fluorescence Signatures as an Index to the Taxonomic Structure of Phytoplankton Communities Dr. Gary Hitchcock Marine Biology and Fisheries Rosenstiel School of Marine and Atmospheric Science 4600
Variability in pigment composition and optical characteristics of phytoplankton in the Labrador Sea and the Central North Atlantic
 MARINE ECOLOGY PROGRESS SERIES Vol. 260: 1 18, 2003 Published September 30 Mar Ecol Prog Ser Variability in pigment composition and optical characteristics of phytoplankton in the Labrador Sea and the
MARINE ECOLOGY PROGRESS SERIES Vol. 260: 1 18, 2003 Published September 30 Mar Ecol Prog Ser Variability in pigment composition and optical characteristics of phytoplankton in the Labrador Sea and the
INTRODUCTION MATERIALS AND METHODS. Alexandrium tamarense (strain ATKR ) used in the present study was isolated in Kure Bay,
 used in the present study was isolated in Kure Bay,") Nitrate Availability for the Accumulation of Shinorine, Palythine, and Porphyra-334 (Mycosporine-like Amino Acids) in Dinoflagellate Alexandrium tamarense Nobuyuki Kobashi 1, Ai Murata 1, Hitomi Taira
Nitrate Availability for the Accumulation of Shinorine, Palythine, and Porphyra-334 (Mycosporine-like Amino Acids) in Dinoflagellate Alexandrium tamarense Nobuyuki Kobashi 1, Ai Murata 1, Hitomi Taira
MARINE ECOLOGY PROGRESS SERIES Mar. Ecol. Prog. Ser.
 Vol. 68: 121-127, 1990 MARINE ECOLOGY PROGRESS SERIES Mar. Ecol. Prog. Ser. Published December 15 Vertical distribution and pigment composition of a picoplanktonic prochlorophyte in the subtropical North
Vol. 68: 121-127, 1990 MARINE ECOLOGY PROGRESS SERIES Mar. Ecol. Prog. Ser. Published December 15 Vertical distribution and pigment composition of a picoplanktonic prochlorophyte in the subtropical North
Time-series observations in the Northern Indian Ocean V.V.S.S. Sarma National Institute of Oceanography Visakhapatnam, India
 The Second GEOSS Asia-Pacific Symposium, Tokyo, 14-16 th April 28 Time-series observations in the Northern Indian Ocean V.V.S.S. Sarma National Institute of Oceanography Visakhapatnam, India Seasonal variations
The Second GEOSS Asia-Pacific Symposium, Tokyo, 14-16 th April 28 Time-series observations in the Northern Indian Ocean V.V.S.S. Sarma National Institute of Oceanography Visakhapatnam, India Seasonal variations
Copepod community growth rates in relation to body size, temperature, and food availability in the East China Sea: A test of Metabolic Theory
 Copepod community growth rates in relation to body size, temperature, and food availability in the East China Sea: A test of Metabolic Theory Kuan-Yu Lin, Akash Sastri, Gwo-Ching Gong, and Chih-hao Hsieh
Copepod community growth rates in relation to body size, temperature, and food availability in the East China Sea: A test of Metabolic Theory Kuan-Yu Lin, Akash Sastri, Gwo-Ching Gong, and Chih-hao Hsieh
Year Two Annual Report (March 2008 February 2009) Introduction. Background
 Introduction. Background") Plankton Monitoring and Zooplankton Grazing Assessment in Vancouver Lake, WA Stephen Bollens and Gretchen Rollwagen-Bollens Washington State University Vancouver Year Two Annual Report (March 28 February
Plankton Monitoring and Zooplankton Grazing Assessment in Vancouver Lake, WA Stephen Bollens and Gretchen Rollwagen-Bollens Washington State University Vancouver Year Two Annual Report (March 28 February
THE ARABIAN SEA: A NATURAL EXPERIMENT IN PHYTOPLANKTON BIOGEOGRAPHY
 THE ARABIAN SEA: A NATURAL EXPERIMENT IN PHYTOPLANKTON BIOGEOGRAPHY LONG TERM GOALS A. Michelle Wood Department of Biology University of Oregon Eugene, Oregon 97403 541-346-0454 email:miche@darkwing.,uoregon.edu
THE ARABIAN SEA: A NATURAL EXPERIMENT IN PHYTOPLANKTON BIOGEOGRAPHY LONG TERM GOALS A. Michelle Wood Department of Biology University of Oregon Eugene, Oregon 97403 541-346-0454 email:miche@darkwing.,uoregon.edu
Meat and two veg? Determining feeding selectivity of bivalve larvae in the Western English Channel with traditional and molecular techniques.
 Meat and two veg? Determining feeding selectivity of bivalve larvae in the Western English Channel with traditional and molecular techniques. Pennie Lindeque, Elaine Fileman, Claudia Halsband-Lenk, Helen
Meat and two veg? Determining feeding selectivity of bivalve larvae in the Western English Channel with traditional and molecular techniques. Pennie Lindeque, Elaine Fileman, Claudia Halsband-Lenk, Helen
Climate Variability Studies in the Ocean
 Climate Variability Studies in the Ocean Topic 1. Long-term variations of vertical profiles of nutrients in the western North Pacific Topic 2. Biogeochemical processes related to ocean carbon cycling:
Climate Variability Studies in the Ocean Topic 1. Long-term variations of vertical profiles of nutrients in the western North Pacific Topic 2. Biogeochemical processes related to ocean carbon cycling:
Microbial Grazers Lab
 Microbial Grazers Lab Objective: Measure the rate at which bacteria are consued by predators. Overview Size based food webs Microbial loop concepts acterial predators Methods to assess icrobial grazing
Microbial Grazers Lab Objective: Measure the rate at which bacteria are consued by predators. Overview Size based food webs Microbial loop concepts acterial predators Methods to assess icrobial grazing
Size distribution and abundance of phytoplankton in the Pacific equatorial upwelling
 ~ Aubert La mer 29: 175-179, 1991 Sociktk franco-japonaise d'ockanographie, Tokyo Size distribution and abundance of phytoplankton in the Pacific equatorial upwelling Le B OUTE I LLER* and Jean B LANGHOT*
~ Aubert La mer 29: 175-179, 1991 Sociktk franco-japonaise d'ockanographie, Tokyo Size distribution and abundance of phytoplankton in the Pacific equatorial upwelling Le B OUTE I LLER* and Jean B LANGHOT*
Protists: Algae Lecture 5 Spring 2014
 Protists: Algae Lecture 5 Spring 2014 Meet the algae 1 Protist Phylogeny Algae - Not monophyletic What unites them as a group? Range from unicellular to multicellular From phytoplankton to kelp forests
Protists: Algae Lecture 5 Spring 2014 Meet the algae 1 Protist Phylogeny Algae - Not monophyletic What unites them as a group? Range from unicellular to multicellular From phytoplankton to kelp forests
Protists: Algae Lecture 5 Spring Protist Phylogeny. Meet the algae. Primary & Secondary Endosymbiosis. Endosymbiosis. Secondary Endosymbiosis
 Meet the algae Protists: Algae Lecture 5 Spring 2014 Protist Phylogeny 1 Primary & Secondary Endosymbiosis 2 Algae - Not monophyletic What unites them as a group? Range from unicellular to multicellular
Meet the algae Protists: Algae Lecture 5 Spring 2014 Protist Phylogeny 1 Primary & Secondary Endosymbiosis 2 Algae - Not monophyletic What unites them as a group? Range from unicellular to multicellular
Bacteria, Friends or Foes?
 Bacteria, Friends or Foes? This unit integrates molecular biology techniques with the role of bacteria in our environment, specifically in the marine environment. The unit starts with introductory activities
Bacteria, Friends or Foes? This unit integrates molecular biology techniques with the role of bacteria in our environment, specifically in the marine environment. The unit starts with introductory activities
Phytoplankton and iron: validation of a global three-dimensional ocean biogeochemical model
 Deep-Sea Research II 50 (2003) 3143 3169 Phytoplankton and iron: validation of a global three-dimensional ocean biogeochemical model Watson W. Gregg a, *, Paul Ginoux b, Paul S. Schopf c, Nancy W. Casey
Deep-Sea Research II 50 (2003) 3143 3169 Phytoplankton and iron: validation of a global three-dimensional ocean biogeochemical model Watson W. Gregg a, *, Paul Ginoux b, Paul S. Schopf c, Nancy W. Casey
Marine Primary Producers
 Marine Primary Producers OCN 201 Biology Lecture 4 http://video.conncoll.edu/f/pasiv/lucid/cyanophora-900.html http://micro.magnet.fsu.edu/moviegallery/ http://www.cellsalive.com/howbig.htm Photo: Fuhrman
Marine Primary Producers OCN 201 Biology Lecture 4 http://video.conncoll.edu/f/pasiv/lucid/cyanophora-900.html http://micro.magnet.fsu.edu/moviegallery/ http://www.cellsalive.com/howbig.htm Photo: Fuhrman
XI. the natural carbon cycle. with materials from J. Kasting (Penn State)
") XI. the natural carbon cycle with materials from J. Kasting (Penn State) outline properties of carbon the terrestrial biological cycle of carbon the ocean cycle of carbon carbon in the rock cycle overview
XI. the natural carbon cycle with materials from J. Kasting (Penn State) outline properties of carbon the terrestrial biological cycle of carbon the ocean cycle of carbon carbon in the rock cycle overview
PHOTOSYNTHESIS Student Packet SUMMARY
 PHOTOSYNTHESIS Student Packet SUMMARY LIVING SYSTEMS REQUIRE ENERGY AND MATTER TO MAINTAIN ORDER, GROW AND REPRODUCE Energy input must exceed energy lost and used by an organism. Organisms use various
PHOTOSYNTHESIS Student Packet SUMMARY LIVING SYSTEMS REQUIRE ENERGY AND MATTER TO MAINTAIN ORDER, GROW AND REPRODUCE Energy input must exceed energy lost and used by an organism. Organisms use various
Lesson 2. Antarctic Oceanography: Component I - Ice/Glaciers Component II - Marine Snow
 Lesson 2. Antarctic Oceanography: Component I - Ice/Glaciers Component II - Marine Snow Lesson Objectives: Introduces students to the different kinds of ice found in Antarctica, Students will become familiar
Lesson 2. Antarctic Oceanography: Component I - Ice/Glaciers Component II - Marine Snow Lesson Objectives: Introduces students to the different kinds of ice found in Antarctica, Students will become familiar
Biogeochemical Impacts due to Mesoscale Eddy Activity in the Sargasso Sea as Measured at the Bermuda Atlantic Time-series Study (BATS)
") Biogeochemical Impacts due to Mesoscale Eddy Activity in the Sargasso Sea as Measured at the Bermuda Atlantic Time-series Study (BATS) Erin N. Sweeney a, Dennis J. McGillicuddy, Jr. *b and Ken O. Buesseler
Biogeochemical Impacts due to Mesoscale Eddy Activity in the Sargasso Sea as Measured at the Bermuda Atlantic Time-series Study (BATS) Erin N. Sweeney a, Dennis J. McGillicuddy, Jr. *b and Ken O. Buesseler
Phytoplankton and Primary Production. Marine habitats
 Phytoplankton and Primary Production (www.microbiological garden) Marine habitats High tide Supralitoral Low tide Pelagic zone Litoral Sublitoral neritic Bathyal oceanic Epipelagic Mesopelagic Bathypelagic
Phytoplankton and Primary Production (www.microbiological garden) Marine habitats High tide Supralitoral Low tide Pelagic zone Litoral Sublitoral neritic Bathyal oceanic Epipelagic Mesopelagic Bathypelagic
1 Carbon - Motivation
 1 Carbon - Motivation Figure 1: Atmospheric pco 2 over the past 400 thousand years as recorded in the ice core from Vostok, Antarctica (Petit et al., 1999). Figure 2: Air-sea flux of CO 2 (mol m 2 yr 1
1 Carbon - Motivation Figure 1: Atmospheric pco 2 over the past 400 thousand years as recorded in the ice core from Vostok, Antarctica (Petit et al., 1999). Figure 2: Air-sea flux of CO 2 (mol m 2 yr 1
Copepod grazing impact on the trophic structure of the microbial assemblage of the San Pedro Channel, California
 JOURNAL OF PLANKTON RESEARCH j VOLUME 27 j NUMBER 1 j PAGES 99 971 j 2 Copepod grazing impact on the trophic structure of the microbial assemblage of the San Pedro Channel, California ASTRID SCHNETZER*
JOURNAL OF PLANKTON RESEARCH j VOLUME 27 j NUMBER 1 j PAGES 99 971 j 2 Copepod grazing impact on the trophic structure of the microbial assemblage of the San Pedro Channel, California ASTRID SCHNETZER*
Liverpool NEMO Shelf Arctic Ocean modelling
 St. Andrews 22 July 2013 ROAM @ Liverpool NEMO Shelf Arctic Ocean modelling Maria Luneva, Jason Holt, Sarah Wakelin NEMO shelf Arctic Ocean model About 50% of the Arctic is shelf sea (
St. Andrews 22 July 2013 ROAM @ Liverpool NEMO Shelf Arctic Ocean modelling Maria Luneva, Jason Holt, Sarah Wakelin NEMO shelf Arctic Ocean model About 50% of the Arctic is shelf sea (
CHAPTER 5 WARM UPS. Mrs. Hilliard
 CHAPTER 5 WARM UPS Mrs. Hilliard CHAPTER 5 VOCABULARY 1. Photosynthesis 2. Cellular respiration 3. Producer 4. Consumer 5. Decomposer 6. Food chain 7. Food web 8. Trophic level 9. Carbon cycle 10. Nitrogen-fixing
CHAPTER 5 WARM UPS Mrs. Hilliard CHAPTER 5 VOCABULARY 1. Photosynthesis 2. Cellular respiration 3. Producer 4. Consumer 5. Decomposer 6. Food chain 7. Food web 8. Trophic level 9. Carbon cycle 10. Nitrogen-fixing
INTRODUCTION prokaryotic eukaryotic pigments
 INTRODUCTION This exercise is intended for you to get familiar and comfortable with using a microscope as well as identifying common microbial groups. Thus, we will observe representatives of all microbes
INTRODUCTION This exercise is intended for you to get familiar and comfortable with using a microscope as well as identifying common microbial groups. Thus, we will observe representatives of all microbes
Productivity in a Changing Southern Ocean. Kevin R. Arrigo Stanford University
 Productivity in a Changing Southern Ocean Kevin R. Arrigo Stanford University 1 Productivity in a Changing Southern Ocean A Paleo-perspective Satellite view of the Southern Ocean Role of ice and iron Controls
Productivity in a Changing Southern Ocean Kevin R. Arrigo Stanford University 1 Productivity in a Changing Southern Ocean A Paleo-perspective Satellite view of the Southern Ocean Role of ice and iron Controls
Arctic Ocean Biology. from the surface to the deep sea
 Arctic Ocean Biology from the surface to the deep sea Christina Bienhold Helmholtz Max Planck Research Group for Deep Sea Ecology and Technology cbienhol@mpi-bremen.de ACCESS Summerschool, Bremen, Germany
Arctic Ocean Biology from the surface to the deep sea Christina Bienhold Helmholtz Max Planck Research Group for Deep Sea Ecology and Technology cbienhol@mpi-bremen.de ACCESS Summerschool, Bremen, Germany
The beam attenuation coefficient and its spectra. (also known as beam-c or extinction coefficient ). Emmanuel Boss, U. of Maine
. Emmanuel Boss, U. of Maine") The beam attenuation coefficient and its spectra (also known as beam-c or extinction coefficient ). Emmanuel Boss, U. of Maine Review: IOP Theory Collin s lecture 2: F o F t Incident Radiant Flux Transmitted
The beam attenuation coefficient and its spectra (also known as beam-c or extinction coefficient ). Emmanuel Boss, U. of Maine Review: IOP Theory Collin s lecture 2: F o F t Incident Radiant Flux Transmitted
Phytoplankton pigment analysis by HPLC and its application in algal community investigations
 Li Bot. et Bull. al. Acad. Pigment Sin. (2002) analysis 43: and 283-290 algal community investigations 283 Phytoplankton pigment analysis by HPLC and its application in algal community investigations Hsiu-Ping
Li Bot. et Bull. al. Acad. Pigment Sin. (2002) analysis 43: and 283-290 algal community investigations 283 Phytoplankton pigment analysis by HPLC and its application in algal community investigations Hsiu-Ping