The molecular mechanism of Noxa-induced mitochondrial dysfunction in p53- mediated cell death
|
|
|
- Brandon Payne
- 6 years ago
- Views:
Transcription
1 The molecular mechanism of Noxa-induced mitochondrial dysfunction in p53- mediated cell death Young-Woo Seo 2, 8, Jin na Shin 1, 8, Kang Hee Ko 1, 8, Jong Hee Cha 1, Jae Yoon Park 1, Byoung Rai Lee 1, Cheol-won Yun 3, Young Myeong Kim 4, Dai-wu Seol 5, Dong-wook Kim 6, Xiao-Ming Yin 7, Tae-Hyoung Kim 1, 9 1 Department of Biochemistry, Chosun University School of Medicine, 375 Seosuk-Dong, Dong-Gu, Gwangju, , Korea; 2 Korea Basic Science Institute Gwang-Ju branch, Chonnam National University 300 Yongbong-Dong, Buk-Gu, Gwang-Ju, , Korea; 3 Department of Microbiology, Pukyong National University, Daeyeon 3 Dong, Nam- Gu, Pusan, , Korea; 4 Kangwon National University Vascular System Research Center, Hyoja 2 Dong, Chunchon, Kangwon-Do, , Korea; 5 Surgery Department, University of Pittsburgh School of Medicine, Pittsburgh, PA15261, USA; 6 Molecular Neurobiology Laboratory, Mclean Hospital Harvard Medical School, Belmont, MA 02478, USA; 7 Pathology Department, University of Pittsburgh School of Medicine, Pittsburgh, PA15261, USA; 8 These authors are equally contributed to this work. 9 Correspondence should be addressed to: Dr. Tae-Hyoung Kim Room # 2205, 2 nd Floor of New Medical Building, Department of Biochemistry Chosun University School of Medicine 375 Seosuk-Dong, Dong-Gu, Gwangju, , Korea Phone: Fax: thkim65@mail.chosun.ac.kr
2 Abstract Genotoxic stresses stabilize the p53 tumor suppressor protein which, in turn, transactivates target genes to cause apoptosis. Although Noxa, a BH3-only member of the Bcl-2 family, was shown to be a target of p53-mediated transactivation and to function as a mediator of p53-dependent apoptosis through mitochondrial dysfunction, the molecular mechanism by which Noxa causes mitochondrial dysfunction is largely unknown. Here we show that two domains (BH3 domain and mitochondrial targeting domain) in Noxa are essential for release of cytochrome c from mitochondria. Noxa-induced cytochrome c release is inhibited by permeability transition (PT) pore inhibitors such as CsA or MgCl 2, and Noxa induces an ultra-structural change of mitochondria yielding swollen mitochondria that are unlike changes induced by tbid. This indicates that Noxa may activate the PT-related pore to release cytochrome c from mitochondria into cytosol. Moreover, Bak-oligomerization, which is an essential event for tbid-induced cytochrome c release in the extrinsic death signaling pathway, is not associated with Noxa-induced cytochrome c release. This finding suggests that the pathway of Noxa-induced mitochondrial dysfunction is distinct from the one of tbid-induced mitochondrial dysfunction. Thus, we propose that there are at least two different pathways of mitochondrial dysfunction; one mediated through Noxa in response
3 to genotoxic stresses and the other through tbid in response to death ligands. Key words: Apoptosis/Bcl-2 family/noxa/mitochondrial dysfunction/p53-induced cell death
4 Introduction The p53 mutation is found in more than half of all human cancer patients. Cancers with loss of p53 function are often resistant to chemotherapeutic agents, mainly due to the absence of p53-dependent apoptosis (1-3). The p53-dependent apoptosis largely depends on the capability of p53 to function as a transcription factor although recent reports show that the transcription-independent function of p53 plays a role in this process. There are many target genes (e.g., Apaf-1, DR5, p53aip, PUMA, and Noxa) transactivated by p53 that are proposed to mediate p53-dependent apoptosis in various cancer cells (4). A recent report shows a close correlation between p53 level and Noxa induction that could be a key determinant of chemically-induced hepatocellular carcinoma in mice, suggesting that Noxa is a crucial mediator of p53-dependent apoptosis in the liver (5). Noxa induced by p53 is up-regulated by x-ray irradiation, and is heavily dependent on p53 and its homologue p73 (6). Noxa belongs to the BH3-only branch of the Bcl-2 protein family although mouse Noxa has two BH3 domains, whilte other BH3-only proteins have a single BH3 domain. Like many Bcl-2 family proteins that translocate to mitochondria and modulate mitochondrial function, Noxa translocates to mitochondria, and then leads to cytochrome c release and caspase-9 activation (7). Most of the BH3-only proteins such as Bid and Bim
5 appear to induce efflux of mitochondrial death-stimulating proteins through Bax or Bak homo-oligomerization leading to the permeabilization of the mitochondrial outer membrane (8,9). However, the detailed molecular mechanism by which Noxa induces mitochondrial dysfunction remains to be solved. In these studies, we characterize Noxainduced mitochondrial dysfunction in order to provide insight on the molecular mechanism by which p53 induces apoptosis. We show that Noxa causes mitochondrial dysfunction by a distinct pathway in mediating p53-dependent apoptosis, unlike mitochondrial dysfunction induced by tbid in response to FasL or TNF-.
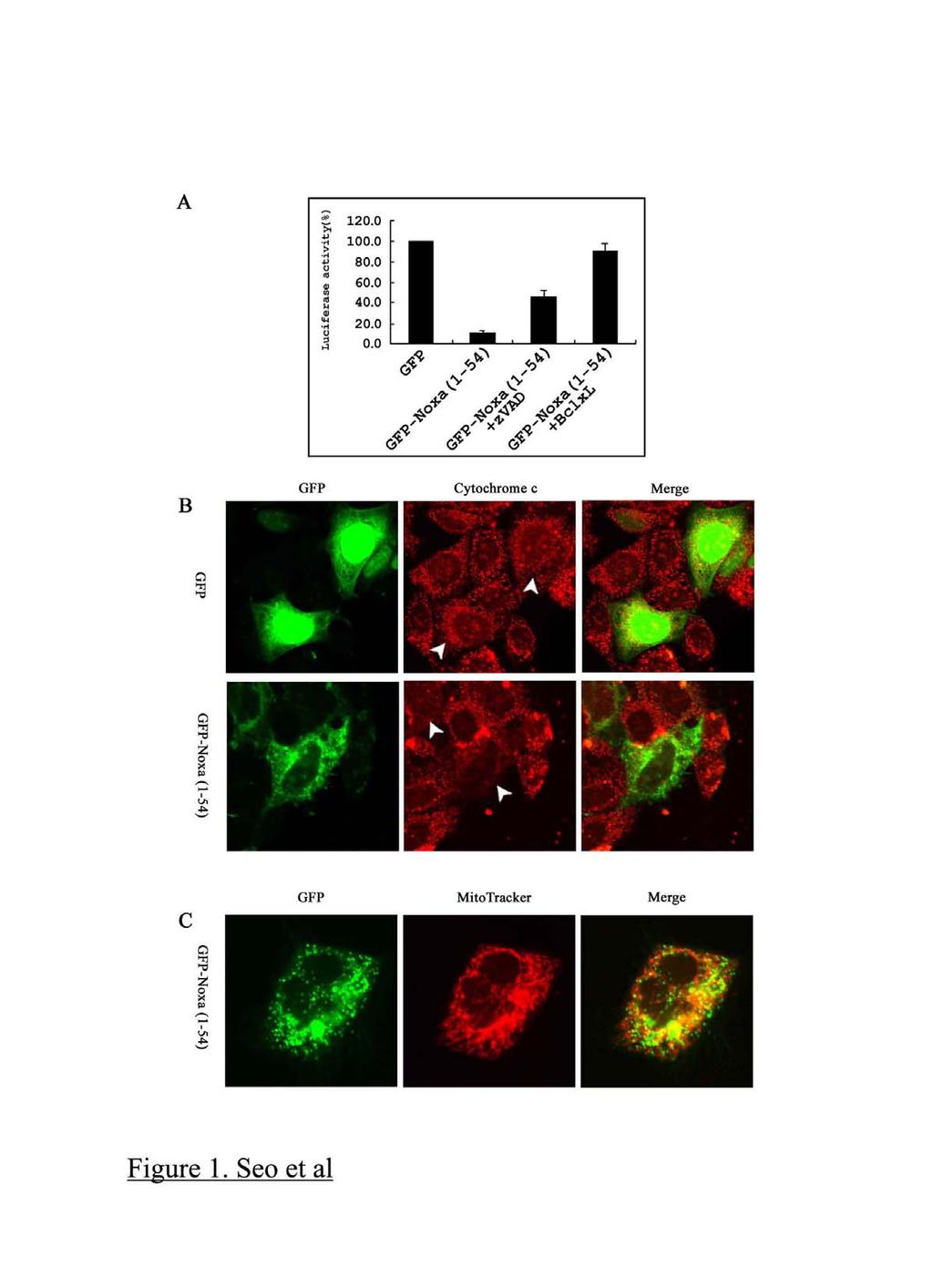
6 Results It has been reported that mouse Noxa has two BH3 domains and induces apoptosis, whereas human Noxa has only one BH3 domain. First, to determine whether human Noxa is able to induce cell death in HeLa cells, GFP-Noxa (1-54) was transfected into HeLa cells. As shown in Fig. 1A, GFP-Noxa (1-54) induced cell death, and Bcl-x L blocked Noxainduced cell death. In addition, the pan-caspase inhibitor z-vad-fmk inhibited Noxainduced cell death, indicating that caspases play a crucial role in Noxa-induced cell death. These results indicate that one BH3 domain in human Noxa is sufficient to induce apoptosis. To determine whether cytochrome c release accompanies human Noxa-induced cell death, GFP-Noxa (1-54) was transfected into HeLa cells, and the cells were immunostained using anti-cytochrome c antibody. HeLa cells transfected with human Noxa showed diffuse cytochrome c distribution throughout the cytosol, but control cells (transfected with GFP alone) maintained the typical mitochondrial distribution of cytochrome c (Fig. 1B), indicating that cytochrome c release from mitochondria is an event induced during human Noxa-mediated cell death. To further confirm that human Noxa targets to mitochondria and causes mitochondrial dysfunction, we checked whether GFP- Noxa (1-54) was present in mitochondria following transfection. GFP-Noxa (1-54) showed
7 a punctate distribution, typical of mitochondria (green color, left panel) and mitochondrial localization of Noxa was confirmed by overlapping staining (yellow color, right panel) with Mitotracker (red color, middle panel, Fig. 1C). This finding is in agreement with a previous report showing the mitochondrial localization of mouse Noxa when using an adenovirus expressing mouse Noxa and suggests that human Noxa is functionally identical to mouse Noxa despite having a single BH3 domain (7). Although the data indicated that human Noxa causes the release of cytochrome c from mitochondria in HeLa cells, it was not clear whether Noxa is able to directly trigger efflux of death-stimulating proteins from the inter-membrane space of mitochondria or whether another, non-mitochondrial, factor is also necessary. To address this question, we adopted the in vitro system of Noxa-induced mitochondrial dysfunction. In this system, recombinant human Noxa protein was able to induce cytochrome c release directly from isolated mitochondria. Noxa-induced cytochrome c release was inhibited by Bcl-x L TM (Fig. 2 A, B). These results strongly suggest that the presence of human Noxa in mitochondria is enough to induce mitochondrial dysfunction. It has been shown that a death-ligand activated BH3-only protein, tbid, targets to the mitochondria and that the DEF helices of tbid play a crucial role in this localization. We
8 hypothesized that human Noxa may have a mitochondrial targeting domain (MTD), similar to tbid. To determine the MTD of Noxa, various expression plasmids of Noxa fused to GFP were transfected into HeLa cells (Fig. 3A). In Fig.1 we showed that Noxa (1-54) displays the punctate distribution that is typical of a mitochondrial staining pattern and mitochondrial distribution of Noxa (1-54) was confirmed by overlapped staining with Mitotracker. This same punctate distribution disappeared in Noxa (1-30) and Noxa (1-40); however, Noxa (21-54) and Noxa (31-54) maintained a similar pattern, indicating that Noxa (31-54) contains the MTD. To define the MTD of Noxa, Noxa (31-54) was dissected as shown in Fig. 3B. Noxa (31-40) and Noxa (31-45) showed a dispersed cytoplasmic distribution, whereas Noxa (41-54) and Noxa (41-50) displayed the punctate pattern. These results demonstrate that human Noxa (41-50) contains the entire MTD. This MTD appears to be conserved between human, mouse, cow, pig and chicken (Fig. 3C), suggesting that the MTD of Noxa is functionally significant for induction of cell death in p53-dependent apoptosis. To verify whether human Noxa needs the MTD to kill cells, plasmids containing various GFP-Noxa constructs and a reporter gene were co-transfected into HeLa cells. Noxa (1-54) and Noxa (21-54), which have intact BH3 and MTD domains, were able to induce cell
9 death (Fig. 3D). We further confirmed that the cell killing induced by Noxa (1-54) and Noxa (21-54) was associated with an efflux of cytochrome c from mitochondria (Fig. 4). Interestingly, Noxa (11-40) and Noxa (21-40), which have the BH3 domain but not the MTD domain, failed to induce cell death (Fig. 3D). In addition, Noxa (31-54) and Noxa (41-54), which have the MTD domain but harbor a disrupted BH3 domain or no BH3 domain, respectively, failed to induce apoptosis, indicating that the MTD domain is necessary but not enough to induce cell death (Fig. 3D). These results demonstrate that the killing activity of the Noxa BH3 domain is functional only when it targets to mitochondria, and this mitochondria targeting is mediated by the MTD. It is known that after engagement of death ligands to death receptors, caspase-8-activated tbid binds to Cardiolipin, which exists exclusively in the mitochondrial membrane and is enriched at the inner membrane and the mitochondrial contact sites. The interaction of tbid with Cardiolipin is a critical step for tbid targeting to mitochondria (10,11). To understand the molecular mechanism by which Noxa targets to mitochondria in p53-dependent apoptosis, we examined whether Noxa also targets to Cardiolipin like tbid. It is unlikely that Noxa binds to Cardiolipin (data not shown), and it implies that p53-mediated mitochondrial dysfunction through Noxa might be distinct from mitochondria dysfunction
10 triggered by death ligands that is mediated through tbid. To examine whether Noxa triggers efflux of mitochondrial proteins in a distinct manner from tbid, we examined the inhibitory activity of CsA in a cytochrome c release assay. CsA showed no significant change in the release of cytochrome c induced by tbid, consistent with a previous report (12) (Fig. 5A). On the other hand, calcium, which is a strong activator of the PT pore, was able to release cytochrome c from isolated mitochondria and, as expected, was inhibited by CsA. Surprisingly, CsA also blocked Noxa-induced cytochrome c release. In addition, Mg 2+, another inhibitor of the PT pore, was also able to inhibit Noxa-induced cytochrome c release, indicating that Noxa in p53-dependent apoptosis is likely to cause efflux of mitochondrial proteins through a PT-related pore in a manner that is distinct from the tbidinduced pathway activated by death ligands. Bak-oligomerization is a key event of tbid-induced cytochrome c release. In fact, tbid is unable to release cytochrome c from Bak-deficient mitochondria (8,9). A recent report showed that unlike mouse liver, rat liver mitochondria display little or no cytochrome c release in response to tbid, due to either insufficient or absent Bak in their mitochondria (13). Therefore, we analyzed Bak-oligomerization using mouse and rat liver mitochondria to determine whether Bak-oligomerization is an important component of Noxa-induced
11 cytochrome c release. The cross-linker BMH or, DMSO as a control, has been used for Bak-oligomerization analyses. As expected, tbid induced significant Bak-oligomerization from mouse but not rat mitochondria. Noxa did not cause significant Bak-oligomerization in either case, although it was able to effect cytochrome c release (Fig. 5B). Furthermore, our results confirmed that rat liver mitochondria only release a small amount of cytochrome c in response to tbid (Fig. 5B). On the other hand, Noxa was able to release a comparable amount of cytochrome c from both rat and mouse mitochondria, without Bakoligomerization. These results reveal that Bak-oligomerization is not required for cytochrome c release in response to Noxa. However, we do not rule out the possibility that Bax may be involved in the Noxa-induced cytochrome c release because Bax is able to compensate for Bak-deficiency. Because CsA and Mg 2+ are blocking agents of the PT pore and inhibit Noxa-induced cytochrome c release (Fig. 5A), we assumed that Noxa is likely to activate the PT pore, resulting in mitochondrial swelling. We next measured mitochondrial swelling by light scatter after Noxa treatment; however, we were unable to detect any significant high amplitude swelling (data not shown). Recently, Scorrano et. al. reported that tbid causes cristae-remodeling of the mitochondrial structure, resulting in the movement of cytochrome
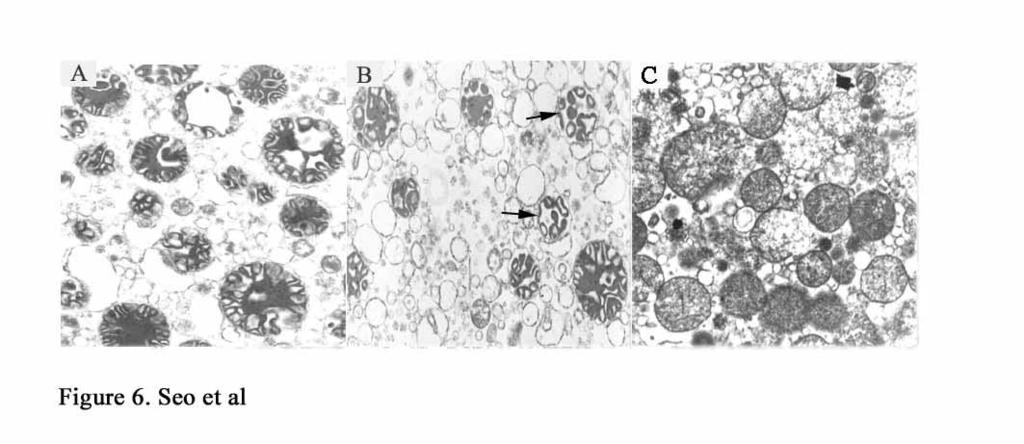
12 c stores (14). We hypothesized that Noxa might also cause ultra-structural changes in the mitochondria without high-amplitude swelling, and this structural change might be distinct from that induced by tbid. To examine the hypothesis, morphological changes of mitochondria treated with tbid, Noxa, or calcium were observed using electron microscopy. As shown in Fig. 6A, freshly isolated mitochondria showed an electron-dense matrix of the isolated mitochondria, the typical condensed state described by Hackenbrock (15). When tbid was used to treat the mitochondria, the electron-dense matrix was reorganized to have mitochondrial features showing a sausage-shaped electron-dense matrix (Fig. 6B) as described by Scorrano et. al (14). Noxa-treated mitochondria, however, showed an expanded and dilute matrix, and small matrical granules with tubular-shaped cristae were widely dispersed (Fig. 6C). These morphological changes are characteristics of swollen mitochondria as designated by Hackenbrock. It is likely that these morphological changes take place without large-amplitude mitochondrial swelling because no significant changes in mitochondrial size were observed. Thus, Noxa seems to cause changes in the mitochondrial ultra-structure characteristic of swollen mitochondria without inducing large-amplitude swelling.
13 Discussion Mitochondrial dysfunction in response to FasL or TNF- appears to be mediated through tbid activated by caspase-8. This process requires either Bak or Bax-oligomerization to release cytochrome c and Smac without the opening of PT pore (8,9,16). However, Bid may not be essential for mitochondrial dysfunction in stress-activated death stimuli such as irradiation because Bid-deficient hepatocytes are able to lose membrane potential of mitochondria in response to these death stimuli (17). This indicates that there is an alternative pathway leading to mitochondrial dysfunction in stress-activated death stimuli. Noxa may be the candidate protein for induction of this alternative pathway because Noxa is transactivated by p53 in response to genotoxic agents, and Noxa causes mitochondrial dysfunction (7). Indeed, our present results show the differences of cytochrome c releases from mitochondria induced by tbid or Noxa. First, Noxa-induced cytochrome c release is not dependent on Bak-oligomerization, whereas Bax- or Bak-oligomerization is requisite for tbid-induced cytochrome c release. Although our data indicates that Noxa-induced mitochondrial dysfunction is not associated with Bak-oligomerization, we do not rule out the possibility of Bax participation in this process. The fact that thymocytes isolated from Bax -/- and Bak -/- mice are resistant to etoposide-induced apoptosis suggests that Bax might
14 be associated with Noxa-induced mitochondrial dysfunction. However, this remains to be answered. Second, PT-related components might, at least in part, participate in the Noxainduced permeabilization of the mitochondrial outer membrane because the PT pore inhibitors CsA and Mg 2+ are able to inhibit Noxa-induced cytochrome c release. This is further supported by a Noxa-induced swollen electron-dilute matrix, an indicator of the opening of the PT pore. This supports the notion that stress-activated p53 utilizes Noxa which leads to mitochondrial dysfunction in a distinct pathway through PT-related pore. Together, it suggests that there are two different pathways in the permeabilization of the mitochondrial outer membrane, which are activated by different death stimuli. This notion might provide an explanation for enhanced apoptotic response of some cancer cells to combinatory treatment of genotoxic agents and death ligand. Bax-deficient colorectal cancer cells, for instance, are resistant to TRAIL alone mainly due to the lack of cytochrome c release; however, these cells are sensitive to TRAIL and the topoisomerase I inhibitor camptothecin. It is possible that Bax-deficient cells have the defective pathway of mitochondrial death signaling induced by tbid; however, the camptothecin might restore the mitochondrial death signaling through Noxa-mediated pathway.
15 Materials and Methods Cell Culture and Mitochondrial Localization HeLa cells were seeded onto an 8 well Lab-Tek Chamber Slide Glass (Nunc, USA) and were grown in DMEM supplemented with 10 % FBS (GibcoBRL, USA). The indicated GFP-Noxa plasmids were transfected into HeLa cells using Effectene (Qiagen, Germany) according to the manufacturer's instructions and cultured for two additional days. In some experiments, Mitotracker Red CMXROS (Molecular Probes Inc., USA) was added to the medium at a final concentration of 200 nm, and cells were further incubated for 10 min. In these experiments, the dye was visualized without a fixation step. After washing with PBS, fresh medium was added. Mitochondrial localization was analyzed with a Laser Scanning Confocal Microscope (Leica Microsystems TCS NT, Leica ltd, Germany). Immunostaining of Cytochrome c HeLa cells were rinsed with PBS three times and fixed with ice-cold 2% buffered paraformaldehyde (ph 7.4) for 10 min. After washing with PBS and blocking with a buffer containing 0.1 % saponin and 0.05 % BSA in PBS, ph 7.4, for 30 min, the cells were
16 incubated with anti-cytochrome c antibody (dilution 1:250, BD Biosciences, USA) for 1 hour at 30C, and then washed with the blocking buffer. Tetramethylrhodamine labeled anti-mouse IgG antibody (dilution 1:250, BD Biosciences, USA) was added to the cells and they were incubated for 1 hour more. After washing with PBS, cells were examined with a Laser Scanning Confocal Microscope. Apoptosis Assay HeLa cells were cultured in 24-well plates and transfected with the indicated plasmid constructs. Each well was harvested after 48 hours of culture and lysates were prepared using the luciferase assay system (Promega, USA). Luciferase activities were determined using Micro Lumat plus LB96V (Berthold Technologies, USA) and were normalized against control -galactosidase activities. Purification of recombinant human Noxa Recombinant human Noxa protein was expressed in E. coli BL21(DE3) cells transformed with pet30(+)a-human Noxa (1-54). It was induced by addition of 0.8 mm IPTG for three hours. Purification was carried out according to the company s instruction (Novagen, USA).
17 Cytochrome c release assay and Assay of Endonuclease G activity Isolated mitochondria treated with recombinant Noxa protein were incubated at 30 C for 1 hour in buffer containing 250 mm sucrose, 10 mm HEPES-NaOH, ph 7.5, 2 mm KH 2 PO 4, 5 mm sodium succinate, and 25 µm EGTA. After centrifugation at 10,000 x g for 10 minutes, supernatants were subjected to SDS-PAGE, and released cytochrome c was visualized by western blot analysis. Analysis of Bak-oligomerization Mitochondria were isolated from either mouse or rat liver and treated with either tbid or Noxa before incubation for 1 hour at 30 C. Following centrifugation, mitochondrial pellets were resuspended in buffer B containing 250 mm sucrose, 10 mm HEPES-NaOH, ph 7.5, 2 mm KH 2 PO 4, 5 mm sodium succinate, and 25 µm EGTA. A cross-linking agent of BMH (10µM) or alternatively, DMSO as a control, was then added. After further incubation for 30 minutes at room temperature with shaking, mitochondrial pellets were harvested by centrifugation and then analyzed by western blot analyses using anti-bak antibody.
18 Alignment of EST clones of Noxa homologues EST clones of cow, chicken, and pig Noxa were identified by a Blast search at the NCBI web site using human Noxa/APR as the reference. GenBank Accession numbers of identified EST clones containing cow, chicken, and pig homologues of Noxa are BF075905, AJ450393, and AF319660, respectively.
19 Acknowledgements We thank Dr. Wendy Mars in the University of Pittsburgh for critical reading of the manuscript and KBSI Gwangju branch for technical assistance of confocal microscope. This work was supported by Korea Research Foundation KRF CP0404 (to T-H Kim) and Vascular System Research Center of KOSEF (to Y-M Kim).
20 References 1. Clarke, A. R., Purdie, C. A., Harrison, D. J., Morris, R. G., Bird, C. C., Hooper, M. L., and Wyllie, A. H. (1993) Nature 362, Lowe, S. W., Schmitt, E. M., Smith, S. W., Osborne, B. A., and Jacks, T. (1993) Nature 362, Lowe, S. W., Bodis, S., McClatchey, A., Remington, L., Ruley, H. E., Fisher, D. E., Housman, D. E., and Jacks, T. (1994) Science 266, El-Deiry, W. S. (2001) Cell Death Differ 8, Eferl, R., Ricci, R., Kenner, L., Zenz, R., David, J. P., Rath, M., and Wagner, E. F. (2003) Cell 112, Flores, E. R., Tsai, K. Y., Crowley, D., Sengupta, S., Yang, A., McKeon, F., and Jacks, T. (2002) Nature 416, Oda, E., Ohki, R., Murasawa, H., Nemoto, J., Shibue, T., Yamashita, T., Tokino, T., Taniguchi, T., and Tanaka, N. (2000) Science 288, Wei, M. C., Zong, W. X., Cheng, E. H., Lindsten, T., Panoutsakopoulou, V., Ross, A. J., Roth, K. A., MacGregor, G. R., Thompson, C. B., and Korsmeyer, S. J. (2001) Science 292,
21 9. Wei, M. C., Lindsten, T., Mootha, V. K., Weiler, S., Gross, A., Ashiya, M., Thompson, C. B., and Korsmeyer, S. J. (2000) Genes Dev 14, Lutter, M., Fang, M., Luo, X., Nishijima, M., Xie, X., and Wang, X. (2000) Nat Cell Biol 2, Lutter, M., Perkins, G. A., and Wang, X. (2001) BMC Cell Biol 2, Kim, T. H., Zhao, Y., Barber, M. J., Kuharsky, D. K., and Yin, X. M. (2000) J Biol Chem 275, Brustovetsky, N., Dubinsky, J. M., Antonsson, B., and Jemmerson, R. (2003) J Neurochem 84, Scorrano, L., Ashiya, M., Buttle, K., Weiler, S., Oakes, S. A., Mannella, C. A., and Korsmeyer, S. J. (2002) Dev Cell 2, Hackenbrock, C. R. (1966) J Cell Biol 30, Wang, X. (2001) Genes Dev 15, Yin, X. M., Wang, K., Gross, A., Zhao, Y., Zinkel, S., Klocke, B., Roth, K. A., and Korsmeyer, S. J. (1999) Nature 400,
22 Figure Legends Fig. 1. Human Noxa induces cell death through mitochondrial dysfunction. (A) GFPhuman Noxa (1-54) (200 ng per well) and luciferase (100 ng per well) expression plasmids were transfected into HeLa cells in the presence of z-vad-fmk (20 µm) or a Bcl-x L (300 ng per well) expression plasmid. Luciferase activity was measured to determine the rate of cell death. (B) Plasmids expressing GFP (200ng per well) or GFP-Noxa (1-54) (200ng per well) were transfected into HeLa cells. After 2 days in culture, cytochrome c was immunostained and observed with a laser scanning confocal microscope. (C) Alternatively, GFP in the cells was visualized in the presence of Mitotracker Red CMXROS (200 nm). Arrows indicate cytochrome c staining (diffuse or obvious) in the cells where transfection has occurred. Fig. 2. Noxa is able to release cytochrome c from isolated mitochondria. (A) Recombinant tbid (0.1 µg ml -1 ) or recombinant human Noxa (1, 2.5, 5, 10, 25, 50 µg ml -1 ) was incubated with isolated mitochondria at 30 C for 1 hour. Following centrifugation, supernatants were subjected to western blot analyses with anti-cytochrome c antibody. (B) Isolated mitochondria were treated with tbid (0.1 µg ml -1 ) or Noxa (50 µg ml -1 ) in the
23 presence of Bcl-x L TM (30 µg ml -1 ), and incubated at 30 C for 1 hour. Cytochrome c was visualized in the supernatants by western blot analyses. Fig. 3. C-terminal region of Noxa contains the mitochondrial targeting domain. (A, B) Schematic diagrams of plasmid constructs expressing various region of Noxa fused to GFP are presented. Plasmid constructs containing the indicated regions of Noxa fused to GFP were transfected into HeLa cells, and cultured for 2 days. Cells were visualized for colocalization of Mitotracker CMXROS (200 nm) and GFP using a confocal microscope. (C) Sequence alignment of Noxa from human, mouse, cow, chicken, and pig. The BH3 domain and the MTD are indicated. (D) Plasmids expressing the indicated regions of Noxa fused to GFP (200 ng per well) were transfected into HeLa cells with the reporter plasmid, luciferase (100 ng per well), and further cultured for 2 days before measuring the luciferase transfected into HeLa cells. After 2 days in culture, cytochrome c was stained with anti- activity. Fig. 4. Cytochrome c release induced by Noxa needs BH3 and MTD domains. Plasmids expressing the indicated regions of Noxa fused to GFP (200 ng per well) were
24 cytochrome c antibody. All data are representative of three separate experiments. Arrows indicate cytochrome c staining (diffuse or obvious) in the cells where transfection has occurred. Fig. 5. Differential mitochondrial dysfunction between tbid and Noxa. (A) Supernatants were analyzed by western blot using anti-cytochrome c antibody. Isolated mitochondria were treated with tbid (0.1 µg ml -1 ), Noxa (50 µg ml -1 ), CaCl 2 (50µM) or MgCl 2 (4mM) in the presence or absence of CsA (50µM). Cytochrome c in the supernatant was analyzed by western blot. (B) Isolated mitochondria from either mouse or rat livers were treated with either tbid (0.1 µg ml -1 ) or Noxa (50 µg ml -1 ) and then incubated for 1 hour at 30 C. Following centrifugation, mitochondrial pellets were resuspended and either the cross-linking agent BMH (10µM) or DMSO as a control buffer was added. The supernatants were analyzed by western blot using anti-cytochrome c antibody. After further incubation with shaking for 30 minutes at room temperature, mitochondrial pellets were harvested by centrifugation and then analyzed by western blot using anti-bak antibody. Equivalent protein loading was verified by western blot using anti-cytochrome c oxidase IV
25 antibodies. Fig. 6. Ultra-structural changes of mitochondria induced by tbid or Noxa. Freshly isolated mitochondria (A) were treated with tbid (B), and Noxa (C), and the incubated for 1 hour at 30 C. Arrows indicate the matrix re-organized mitochondria. After centrifugation, mitochondrial pellets were processed for electron microscopy. Mitochondrial images were visualized with Hitachi's Transmission Electron Microscope (TEM). All images were taken with 7,000 magnification.
26 Running heads Noxa-induced mitochondrial dysfunction
27

28

29
30

31
32
33 The molecular mechanism of Noxa-induced mitochondrial dysfunction in p53-mediated cell death Young-Woo Seo, Jin na Shin, Kang Hee Ko, Jong Hee Cha, Jae Yoon Park, Byoung Rai Lee, Cheol-won Yun, Young Myeong Kim, Dai-wu Seol, Dong-wook Kim, Xiao-Ming Yin and Tae-Hyoung Kim J. Biol. Chem. published online September 18, 2003 Access the most updated version of this article at doi: /jbc.M Alerts: When this article is cited When a correction for this article is posted Click here to choose from all of JBC's alerts
Supplemental Figures S1 S5
 Beyond reduction of atherosclerosis: PON2 provides apoptosis resistance and stabilizes tumor cells Ines Witte (1), Sebastian Altenhöfer (1), Petra Wilgenbus (1), Julianna Amort (1), Albrecht M. Clement
Beyond reduction of atherosclerosis: PON2 provides apoptosis resistance and stabilizes tumor cells Ines Witte (1), Sebastian Altenhöfer (1), Petra Wilgenbus (1), Julianna Amort (1), Albrecht M. Clement
TNFα 18hr. Control. CHX 18hr. TNFα+ CHX 18hr. TNFα: 18 18hr (KDa) PARP. Cleaved. Cleaved. Cleaved. Caspase3. Pellino3 shrna. Control shrna.
 PARP. Cleaved. Cleaved. Cleaved. Caspase3. Pellino3 shrna. Control shrna.") Survival ( %) a. TNFα 18hr b. Control sirna Pellino3 sirna TNFα: 18 18hr c. Control shrna Pellino3 shrna Caspase3 Actin Control d. Control shrna Pellino3 shrna *** 100 80 60 CHX 18hr 40 TNFα+ CHX 18hr
Survival ( %) a. TNFα 18hr b. Control sirna Pellino3 sirna TNFα: 18 18hr c. Control shrna Pellino3 shrna Caspase3 Actin Control d. Control shrna Pellino3 shrna *** 100 80 60 CHX 18hr 40 TNFα+ CHX 18hr
RayBio CaspGLOW TM Fluorescein Active Caspase-3 Staining Kit
 RayBio CaspGLOW TM Fluorescein Active Caspase-3 Staining Kit User Manual Version 1.0 May 10 th, 2015 RayBio Caspase-3 Fluorometric Assay Kit Protocol (Cat#: 68FLS-Casp3-S) RayBiotech, Inc. We Provide You
RayBio CaspGLOW TM Fluorescein Active Caspase-3 Staining Kit User Manual Version 1.0 May 10 th, 2015 RayBio Caspase-3 Fluorometric Assay Kit Protocol (Cat#: 68FLS-Casp3-S) RayBiotech, Inc. We Provide You
NucView TM 488 Caspase-3 Assay Kit for Live Cells
 NucView TM 488 Caspase-3 Assay Kit for Live Cells Catalog Number: 30029 (100-500 assays) Contact Information Address: Biotium, Inc. 3423 Investment Blvd. Suite 8 Hayward, CA 94545 USA Telephone: (510)
NucView TM 488 Caspase-3 Assay Kit for Live Cells Catalog Number: 30029 (100-500 assays) Contact Information Address: Biotium, Inc. 3423 Investment Blvd. Suite 8 Hayward, CA 94545 USA Telephone: (510)
Role of Mitochondrial Remodeling in Programmed Cell Death in
 Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Cytochrome c, cytochrome b 5 and electron transfer
 Cytochrome c, cytochrome b 5 and electron transfer Covalent bonds Cytochrome c is a major player in membrane associated electron transport systems in bacteria and mitochondria. Cytochrome c has two axial
Cytochrome c, cytochrome b 5 and electron transfer Covalent bonds Cytochrome c is a major player in membrane associated electron transport systems in bacteria and mitochondria. Cytochrome c has two axial
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
 http://cwp.embo.org/w09-13/index.html Roads to Ruin is s o t p o ap healthy cell autophagic cell death ne cr os is Thanks to Seamus Martin! Evolution of apoptosis signalling cascades Inhibitor Adopted
http://cwp.embo.org/w09-13/index.html Roads to Ruin is s o t p o ap healthy cell autophagic cell death ne cr os is Thanks to Seamus Martin! Evolution of apoptosis signalling cascades Inhibitor Adopted
BH-3-only BIK Functions at the Endoplasmic Reticulum to Stimulate Cytochrome c Release from Mitochondria*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 20, Issue of May 17, pp. 18053 18060, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. BH-3-only BIK Functions
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 20, Issue of May 17, pp. 18053 18060, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. BH-3-only BIK Functions
Effects of the BH3-only protein human Noxa on mitochondrial dynamics
 FEBS Letters 583 (2009) 2349 2354 journal homepage: www.febsletters.org Effects of the BH3-only protein human Noxa on mitochondrial dynamics Ha-Na Woo a,c,e, Young-Woo Seo b, Ae Ran Moon c, Seon-Yong Jeong
FEBS Letters 583 (2009) 2349 2354 journal homepage: www.febsletters.org Effects of the BH3-only protein human Noxa on mitochondrial dynamics Ha-Na Woo a,c,e, Young-Woo Seo b, Ae Ran Moon c, Seon-Yong Jeong
Enterovirus 71 2B induces cell apoptosis by directly inducing the conformational activation of the pro-apoptotic protein Bax
 JVI Accepted Manuscript Posted Online 24 August 2016 J. Virol. doi:10.1128/jvi.01499-16 Copyright 2016, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
JVI Accepted Manuscript Posted Online 24 August 2016 J. Virol. doi:10.1128/jvi.01499-16 Copyright 2016, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Supplementary Figure 1. AnnexinV FITC and Sytox orange staining in wild type, Nlrp3 /, ASC / and casp1/11 / TEC treated with TNF /CHX.
 Supplementary Figure 1. AnnexinV FITC and Sytox orange staining in wild type, Nlrp3 /, ASC / and casp1/11 / TEC treated with TNF /CHX. Phase contrast and widefield fluorescence microscopy (20x magnification).
Supplementary Figure 1. AnnexinV FITC and Sytox orange staining in wild type, Nlrp3 /, ASC / and casp1/11 / TEC treated with TNF /CHX. Phase contrast and widefield fluorescence microscopy (20x magnification).
Hybrid Gold Superstructures: Synthesis and. Specific Cell Surface Protein Imaging Applications
 Supporting Information Hybrid Gold Nanocube@Silica@Graphene-Quantum-Dot Superstructures: Synthesis and Specific Cell Surface Protein Imaging Applications Liu Deng, Ling Liu, Chengzhou Zhu, Dan Li and Shaojun
Supporting Information Hybrid Gold Nanocube@Silica@Graphene-Quantum-Dot Superstructures: Synthesis and Specific Cell Surface Protein Imaging Applications Liu Deng, Ling Liu, Chengzhou Zhu, Dan Li and Shaojun
Nature Medicine: doi: /nm.3776
 C terminal Hsp90 inhibitors restore glucocorticoid sensitivity and relieve a mouse allograft model of Cushing s disease Mathias Riebold, Christian Kozany, Lee Freiburger, Michael Sattler, Michael Buchfelder,
C terminal Hsp90 inhibitors restore glucocorticoid sensitivity and relieve a mouse allograft model of Cushing s disease Mathias Riebold, Christian Kozany, Lee Freiburger, Michael Sattler, Michael Buchfelder,
SUPPLEMENTARY INFORMATION
 GP2 Type I-piliated bacteria FAE M cell M cell pocket idc T cell mdc Generation of antigenspecific T cells Induction of antigen-specific mucosal immune response Supplementary Figure 1 Schematic diagram
GP2 Type I-piliated bacteria FAE M cell M cell pocket idc T cell mdc Generation of antigenspecific T cells Induction of antigen-specific mucosal immune response Supplementary Figure 1 Schematic diagram
4) Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.
 Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.") Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Apoptosis in Mammalian Cells
 Apoptosis in Mammalian Cells 7.16 2-10-05 Apoptosis is an important factor in many human diseases Cancer malignant cells evade death by suppressing apoptosis (too little apoptosis) Stroke damaged neurons
Apoptosis in Mammalian Cells 7.16 2-10-05 Apoptosis is an important factor in many human diseases Cancer malignant cells evade death by suppressing apoptosis (too little apoptosis) Stroke damaged neurons
The cytotoxicity of gold nanoparticles is dispersitydependent
 Electronic Supplementary Material (ESI) for Dalton Transactions. This journal is The Royal Society of Chemistry 2015 The cytotoxicity of gold nanoparticles is dispersitydependent Dengtong Huang, Hualu
Electronic Supplementary Material (ESI) for Dalton Transactions. This journal is The Royal Society of Chemistry 2015 The cytotoxicity of gold nanoparticles is dispersitydependent Dengtong Huang, Hualu
Production of Recombinant Annexin V from plasmid pet12a-papi
 Tait Research Laboratory Page 1 of 5 Principle Production of Recombinant Annexin V from plasmid pet12a-papi Annexin V is expressed cytoplasmically in BL21(DE3) E. coli (Novagen) with the pet vector system
Tait Research Laboratory Page 1 of 5 Principle Production of Recombinant Annexin V from plasmid pet12a-papi Annexin V is expressed cytoplasmically in BL21(DE3) E. coli (Novagen) with the pet vector system
Research Communication
 IUBMB Life, 60(7): 473 480, July 2008 Research Communication Hepatitis B Virus X Protein Induces Apoptosis by Enhancing Translocation of Bax to Mitochondria Hye Jin Kim, Sang Yong Kim, Jinchul Kim, Heemin
IUBMB Life, 60(7): 473 480, July 2008 Research Communication Hepatitis B Virus X Protein Induces Apoptosis by Enhancing Translocation of Bax to Mitochondria Hye Jin Kim, Sang Yong Kim, Jinchul Kim, Heemin
Proteome-wide High Throughput Cell Based Assay for Apoptotic Genes
 John Kenten, Doug Woods, Pankaj Oberoi, Laura Schaefer, Jonathan Reeves, Hans A. Biebuyck and Jacob N. Wohlstadter 9238 Gaither Road, Gaithersburg, MD 2877. Phone: 24.631.2522 Fax: 24.632.2219. Website:
John Kenten, Doug Woods, Pankaj Oberoi, Laura Schaefer, Jonathan Reeves, Hans A. Biebuyck and Jacob N. Wohlstadter 9238 Gaither Road, Gaithersburg, MD 2877. Phone: 24.631.2522 Fax: 24.632.2219. Website:
Embedded together: The life and death consequences of interaction of the Bcl-2 family with membranes
 DOI 10.1007/s10495-007-0746-4 Embedded together: The life and death consequences of interaction of the Bcl-2 family with membranes Brian Leber Jialing Lin David W. Andrews Published online: 15 February
DOI 10.1007/s10495-007-0746-4 Embedded together: The life and death consequences of interaction of the Bcl-2 family with membranes Brian Leber Jialing Lin David W. Andrews Published online: 15 February
SEPTEMBER 10, 2010 VOLUME 285 NUMBER 37 JOURNAL OF BIOLOGICAL CHEMISTRY 28749
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 37, pp. 28749 28763, September 10, 2010 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Bcl-2 and Bax Interact
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 37, pp. 28749 28763, September 10, 2010 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Bcl-2 and Bax Interact
Supporting Information. Carbon Imidazolate Framework-8 Nanoparticles for
 Supporting Information Carbon Nanodots@Zeolitic Imidazolate Framework-8 Nanoparticles for Simultaneous ph-responsive Drug Delivery and Fluorescence Imaging Liu He, a Tingting Wang, b Jiping An, c Xiaomeng
Supporting Information Carbon Nanodots@Zeolitic Imidazolate Framework-8 Nanoparticles for Simultaneous ph-responsive Drug Delivery and Fluorescence Imaging Liu He, a Tingting Wang, b Jiping An, c Xiaomeng
Apoptosis: Death Comes for the Cell
 Apoptosis: Death Comes for the Cell Joe W. Ramos joeramos@hawaii.edu From Ingmar Bergman s The Seventh Seal 1 2 Mutations in proteins that regulate cell proliferation, survival and death can contribute
Apoptosis: Death Comes for the Cell Joe W. Ramos joeramos@hawaii.edu From Ingmar Bergman s The Seventh Seal 1 2 Mutations in proteins that regulate cell proliferation, survival and death can contribute
Supplementary Information
 Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Direct Interaction between Survivin and Smac/DIABLO Is Essential for the Anti-apoptotic Activity of Survivin during Taxol-induced Apoptosis*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 25, Issue of June 20, pp. 23130 23140, 2003 2003 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Direct Interaction
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 25, Issue of June 20, pp. 23130 23140, 2003 2003 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Direct Interaction
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1
 and Ksp/Cre;Pkd1") Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
Supplementary Figure 1: To test the role of mir-17~92 in orthologous genetic model of ADPKD, we generated Ksp/Cre;Pkd1 F/F (Pkd1-KO) and Ksp/Cre;Pkd1 F/F ;mir-17~92 F/F (Pkd1-miR-17~92KO) mice. (A) Q-PCR
Intracellular GSH Assay Kit
 ab112132 Intracellular GSH Assay Kit Instructions for Use For detecting Intracellular GSH in cells by using our proprietary green fluorescence probe This product is for research use only and is not intended
ab112132 Intracellular GSH Assay Kit Instructions for Use For detecting Intracellular GSH in cells by using our proprietary green fluorescence probe This product is for research use only and is not intended
Bio-Plex Pro RBM Apoptosis Assays
 Acute Phase Apoptosis Cancer Cardiovascular Disease Cytokines Chemokines, Growth Factors Diabetes Gene Expression Genotyping Immunoglobulin Isotyping MicroRNA Expression Bio-Plex Pro RBM Apoptosis Assays
Acute Phase Apoptosis Cancer Cardiovascular Disease Cytokines Chemokines, Growth Factors Diabetes Gene Expression Genotyping Immunoglobulin Isotyping MicroRNA Expression Bio-Plex Pro RBM Apoptosis Assays
Supporting Information
 Supporting Information Wang et al. 10.1073/pnas.0804871105 SI Materials and Methods Cell Culture and Transfection. Human neuroblastoma M17 cells were grown as described before (1). Transfection was performed
Supporting Information Wang et al. 10.1073/pnas.0804871105 SI Materials and Methods Cell Culture and Transfection. Human neuroblastoma M17 cells were grown as described before (1). Transfection was performed
SUPPLEMENTARY INFORMATION
 Cell viability rate 0.8 0.6 0 0.05 0.1 0.2 0.3 0.4 0.5 0.7 1 Exposure duration (s) Supplementary Figure 1. Femtosecond laser could disrupt and turn off GFP without photons at 473 nm and keep cells alive.
Cell viability rate 0.8 0.6 0 0.05 0.1 0.2 0.3 0.4 0.5 0.7 1 Exposure duration (s) Supplementary Figure 1. Femtosecond laser could disrupt and turn off GFP without photons at 473 nm and keep cells alive.
Opa1-Mediated Cristae Opening Is Bax/Bak and BH3 Dependent, Required for Apoptosis, and Independent of Bak Oligomerization
 Article Opa1-Mediated Cristae Opening Is Bax/Bak and BH3 Dependent, Required for Apoptosis, and Independent of Bak Oligomerization Ryuji Yamaguchi, 1,4 Lydia Lartigue, 1 Guy Perkins, 2 Ray T. Scott, 2
Article Opa1-Mediated Cristae Opening Is Bax/Bak and BH3 Dependent, Required for Apoptosis, and Independent of Bak Oligomerization Ryuji Yamaguchi, 1,4 Lydia Lartigue, 1 Guy Perkins, 2 Ray T. Scott, 2
Supplemental Information. The Mitochondrial Fission Receptor MiD51. Requires ADP as a Cofactor
 Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
JCB. Mitochondrial release of apoptosis-inducing factor occurs downstream of cytochrome c release in response to several proapoptotic stimuli.
 JCB Published Online: 16 December, 2002 Supp Info: http://doi.org/10.1083/jcb.200207071 Downloaded from jcb.rupress.org on July 3, 2018 Report Mitochondrial release of apoptosis-inducing factor occurs
JCB Published Online: 16 December, 2002 Supp Info: http://doi.org/10.1083/jcb.200207071 Downloaded from jcb.rupress.org on July 3, 2018 Report Mitochondrial release of apoptosis-inducing factor occurs
Cell Death & Trophic Factors II. Steven McLoon Department of Neuroscience University of Minnesota
 Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Apoptosis EXTRA REQUIREMETS
 Apoptosis Introduction (SLIDE 1) Organisms develop from a single cell, and in doing so an anatomy has to be created. This process not only involves the creation of new cells, but also the removal of cells
Apoptosis Introduction (SLIDE 1) Organisms develop from a single cell, and in doing so an anatomy has to be created. This process not only involves the creation of new cells, but also the removal of cells
Regulation of mitochondrial fission and apoptosis by the mitochondrial outer membrane protein hfis1
 Research Article 4141 Regulation of mitochondrial fission and apoptosis by the mitochondrial outer membrane protein hfis1 Tianzheng Yu 1, Randall J. Fox 1, Lindsay S. Burwell 2 and Yisang Yoon 1, * 1 Department
Research Article 4141 Regulation of mitochondrial fission and apoptosis by the mitochondrial outer membrane protein hfis1 Tianzheng Yu 1, Randall J. Fox 1, Lindsay S. Burwell 2 and Yisang Yoon 1, * 1 Department
ELECTRONIC SUPPLEMENTARY INFORMATION (ESI) variable light emission created via direct ultrasonic exfoliation of
 variable light emission created via direct ultrasonic exfoliation of") Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2015 ELECTRONIC SUPPLEMENTARY INFORMATION (ESI) High quantum-yield luminescent MoS 2 quantum dots
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2015 ELECTRONIC SUPPLEMENTARY INFORMATION (ESI) High quantum-yield luminescent MoS 2 quantum dots
Biochimica et Biophysica Acta
 Biochimica et Biophysica Acta 1832 (2013) 453 474 Contents lists available at SciVerse ScienceDirect Biochimica et Biophysica Acta journal homepage: www.elsevier.com/locate/bbadis PSAP induces a unique
Biochimica et Biophysica Acta 1832 (2013) 453 474 Contents lists available at SciVerse ScienceDirect Biochimica et Biophysica Acta journal homepage: www.elsevier.com/locate/bbadis PSAP induces a unique
Center for Cell Imaging Department of Cell Biology
 Center for Cell Imaging Department of Cell Biology Contents Preparation of Colloidal Gold Conjugates Coupling the Protein A to the Gold Particles Purification of the protein A-gold. Storage Influence of
Center for Cell Imaging Department of Cell Biology Contents Preparation of Colloidal Gold Conjugates Coupling the Protein A to the Gold Particles Purification of the protein A-gold. Storage Influence of
Death Ligand- Death Receptor. Annexin V. Fas PI MKK7 PIP3. pro-caspase-3. Granzyme B. Caspase Activation. Bid. Bcl-2 Caspase-3 Caspase-12
 International Corporation life. science. discovery. { Apoptosis Apoptosis Kits International Corporation Apoptosis Signal Cascade and Detection Kits Annexin V Kits MEBCYTO Apoptosis Kit Annexin V Apoptosis
International Corporation life. science. discovery. { Apoptosis Apoptosis Kits International Corporation Apoptosis Signal Cascade and Detection Kits Annexin V Kits MEBCYTO Apoptosis Kit Annexin V Apoptosis
TRAIL-induced apoptosis requires Bax-dependent mitochondrial release of Smac/DIABLO
 TRAIL-induced apoptosis requires Bax-dependent mitochondrial release of Smac/DIABLO Yibin Deng, Yahong Lin, and Xiangwei Wu 1 Huffington Center on Aging and Department of Molecular and Cellular Biology,
TRAIL-induced apoptosis requires Bax-dependent mitochondrial release of Smac/DIABLO Yibin Deng, Yahong Lin, and Xiangwei Wu 1 Huffington Center on Aging and Department of Molecular and Cellular Biology,
 DOI: 10.1038/ncb2819 Gαi3 / Actin / Acetylated Tubulin Gαi3 / Actin / Acetylated Tubulin a a Gαi3 a Actin Gαi3 WT Gαi3 WT Gαi3 WT b b Gαi3 b Actin Gαi3 KO Gαi3 KO Gαi3 KO # # Figure S1 Loss of protein
DOI: 10.1038/ncb2819 Gαi3 / Actin / Acetylated Tubulin Gαi3 / Actin / Acetylated Tubulin a a Gαi3 a Actin Gαi3 WT Gαi3 WT Gαi3 WT b b Gαi3 b Actin Gαi3 KO Gαi3 KO Gαi3 KO # # Figure S1 Loss of protein
Supplemental material
 Supplemental material THE JOURNAL OF CELL BIOLOGY Mourier et al., http://www.jcb.org/cgi/content/full/jcb.201411100/dc1 Figure S1. Size and mitochondrial content in Mfn1 and Mfn2 knockout hearts. (A) Body
Supplemental material THE JOURNAL OF CELL BIOLOGY Mourier et al., http://www.jcb.org/cgi/content/full/jcb.201411100/dc1 Figure S1. Size and mitochondrial content in Mfn1 and Mfn2 knockout hearts. (A) Body
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11419 Supplementary Figure 1 Schematic representation of innate immune signaling pathways induced by intracellular Salmonella in cultured macrophages. a, During the infection Salmonella
doi:10.1038/nature11419 Supplementary Figure 1 Schematic representation of innate immune signaling pathways induced by intracellular Salmonella in cultured macrophages. a, During the infection Salmonella
Kyung-Won LEE, a,b Hyun-Ju JUNG, c Hee-Juhn PARK, c Deog-Gon KIM, d Jin-Yong LEE, d and Kyung-Tae LEE*,a,b
 854 Biol. Pharm. Bull. 28(5) 854 859 (2005) Vol. 28, No. 5 b -D-Xylopyranosyl-(1 3)-b -D-glucuronopyranosyl Echinocystic Acid Isolated from the Roots of Codonopsis lanceolata Induces Caspase-Dependent
854 Biol. Pharm. Bull. 28(5) 854 859 (2005) Vol. 28, No. 5 b -D-Xylopyranosyl-(1 3)-b -D-glucuronopyranosyl Echinocystic Acid Isolated from the Roots of Codonopsis lanceolata Induces Caspase-Dependent
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
APOPTOSIS REGULATOR BCL-2
 Papers on Anthropology Apoptosis regulator XXII, 2013, BCL-2 pp. 63 67 P. Hussar, M. Žuravskaja, M. Kärner APOPTOSIS REGULATOR BCL-2 Piret Hussar 1, Maria Žuravskaja 2, Martin Kärner 2 1 Institute of Anatomy,
Papers on Anthropology Apoptosis regulator XXII, 2013, BCL-2 pp. 63 67 P. Hussar, M. Žuravskaja, M. Kärner APOPTOSIS REGULATOR BCL-2 Piret Hussar 1, Maria Žuravskaja 2, Martin Kärner 2 1 Institute of Anatomy,
Vibrio cholerae porin OmpU induces caspase-independent programmed cell death upon translocation to the host cell mitochondria
 JBC Papers in Press. Published on November 11, 2015 as Manuscript M115.670182 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m115.670182 OmpU induces caspase-independent programmed cell
JBC Papers in Press. Published on November 11, 2015 as Manuscript M115.670182 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m115.670182 OmpU induces caspase-independent programmed cell
Apoptosis Detection Using the BD Accuri C6 Flow Cytometer
 Apoptosis Detection Using the BD Accuri C6 Flow Cytometer Stacey Roys, Marketing Applications Specialist, BD Biosciences Cyndy Lane, Senior Product Manager, BD Biosciences 23-14519-00 Outline What is apoptosis?
Apoptosis Detection Using the BD Accuri C6 Flow Cytometer Stacey Roys, Marketing Applications Specialist, BD Biosciences Cyndy Lane, Senior Product Manager, BD Biosciences 23-14519-00 Outline What is apoptosis?
Mechanisms. Cell Death. Carol M. Troy, MD PhD August 24, 2009 PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT 8/20/2009
 Mechanisms of Cell Death Carol M. Troy, MD PhD August 24, 2009 PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT A. MORPHOGENESIS: SCULPTING/SHAPING STRUCTURES CREATION OF CAVITIES AND TUBES FUSION OF TISSUE
Mechanisms of Cell Death Carol M. Troy, MD PhD August 24, 2009 PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT A. MORPHOGENESIS: SCULPTING/SHAPING STRUCTURES CREATION OF CAVITIES AND TUBES FUSION OF TISSUE
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
Technical Bulletin. Caspase 9 Fluorescein (FLICA) Assay. Catalog Number CS0300 Storage Temperature 2-8 C
 Assay. Catalog Number CS0300 Storage Temperature 2-8 C") 1 Caspase 9 Fluorescein (FLICA) Assay Catalog Number CS0300 Storage Temperature 2-8 C Technical Bulletin Product Description Caspases are detected by immunoprecipitation, immunoblotting with caspase specific
1 Caspase 9 Fluorescein (FLICA) Assay Catalog Number CS0300 Storage Temperature 2-8 C Technical Bulletin Product Description Caspases are detected by immunoprecipitation, immunoblotting with caspase specific
In Situ Gelation-Induced Death of Cancer Cells Based on Proteinosomes
 Supporting information for In Situ Gelation-Induced Death of Cancer Cells Based on Proteinosomes Yuting Zhou, Jianmin Song, Lei Wang*, Xuting Xue, Xiaoman Liu, Hui Xie*, and Xin Huang* MIIT Key Laboratory
Supporting information for In Situ Gelation-Induced Death of Cancer Cells Based on Proteinosomes Yuting Zhou, Jianmin Song, Lei Wang*, Xuting Xue, Xiaoman Liu, Hui Xie*, and Xin Huang* MIIT Key Laboratory
Supporting Information
 Supporting Information A Multifunctional PB@mSiO 2 -PEG/DOX Nanoplatform for Combined Photothermal-Chemotherapy of Tumor Yun Yan Su a#, Zhaogang Teng a#, Hui Yao b, Shou Ju Wang a, Ying Tian a, Yun Lei
Supporting Information A Multifunctional PB@mSiO 2 -PEG/DOX Nanoplatform for Combined Photothermal-Chemotherapy of Tumor Yun Yan Su a#, Zhaogang Teng a#, Hui Yao b, Shou Ju Wang a, Ying Tian a, Yun Lei
Modeling a Snap-Action, Variable-Delay Switch Controlling Extrinsic Cell Death
 Modeling a Snap-Action, Variable-Delay Switch Controlling Extrinsic Cell Death John G. Albeck 1[, John M. Burke 1,2[, Sabrina L. Spencer 1,3, Douglas A. Lauffenburger 2,3, Peter K. Sorger 1,2* PLoS BIOLOGY
Modeling a Snap-Action, Variable-Delay Switch Controlling Extrinsic Cell Death John G. Albeck 1[, John M. Burke 1,2[, Sabrina L. Spencer 1,3, Douglas A. Lauffenburger 2,3, Peter K. Sorger 1,2* PLoS BIOLOGY
Caspase Substrate Assay Kit (Colorimetric)
") Caspase Substrate Assay Kit (Colorimetric) Catalog Number KA3694 7 x 25 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Caspase Substrate Assay Kit (Colorimetric) Catalog Number KA3694 7 x 25 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Caspase 8 (active) Red Staining Kit
 Red Staining Kit") ab65618 Caspase 8 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 8 in living cells This product is for research use only and is not
ab65618 Caspase 8 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 8 in living cells This product is for research use only and is not
RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit
 ELISA Kit") RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit Catalog #: PEL-EIF2A-S52 User Manual Last revised August 10, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway
RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit Catalog #: PEL-EIF2A-S52 User Manual Last revised August 10, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway
The on-line version of this article (available at contains supplemental
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 21, pp. 14764 14775, May 26, 2006 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Auto-activation of the
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 21, pp. 14764 14775, May 26, 2006 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Auto-activation of the
Programmed Cell Death
 Programmed Cell Death Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id What is apoptosis? a normal component of the development
Programmed Cell Death Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id What is apoptosis? a normal component of the development
Supplementary Figure 1. Biochemical and sequence alignment analyses the
 Supplementary Figure 1. Biochemical and sequence alignment analyses the interaction of OPTN and TBK1. (a) Analytical gel filtration chromatography analysis of the interaction between TBK1 CTD and OPTN(1-119).
Supplementary Figure 1. Biochemical and sequence alignment analyses the interaction of OPTN and TBK1. (a) Analytical gel filtration chromatography analysis of the interaction between TBK1 CTD and OPTN(1-119).
Monte Carlo study elucidates the type 1/type 2 choice in apoptotic death signaling in normal and cancer cells
 Cells 2013, 2, 1-x manuscripts; doi:10.3390/cells20x000x OPEN ACCESS cells ISSN 2073-4409 www.mdpi.com/journal/cells Article Monte Carlo study elucidates the type 1/type 2 choice in apoptotic death signaling
Cells 2013, 2, 1-x manuscripts; doi:10.3390/cells20x000x OPEN ACCESS cells ISSN 2073-4409 www.mdpi.com/journal/cells Article Monte Carlo study elucidates the type 1/type 2 choice in apoptotic death signaling
Supporting Information
 Supporting Information Phenyl-Modified Carbon Nitride Quantum Dots with Distinct Photoluminescence Behavior Qianling Cui, Jingsan Xu,* Xiaoyu Wang, Lidong Li,* Markus Antonietti, and Menny Shalom anie_201511217_sm_miscellaneous_information.pdf
Supporting Information Phenyl-Modified Carbon Nitride Quantum Dots with Distinct Photoluminescence Behavior Qianling Cui, Jingsan Xu,* Xiaoyu Wang, Lidong Li,* Markus Antonietti, and Menny Shalom anie_201511217_sm_miscellaneous_information.pdf
(17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON
 CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON") (17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON Jim Burton 1 and Marianne Pedersen Abstract. Cyclanilide [1-(2,4-dichlorophenylaminocarbonyl)-cyclopropane carboxylic
(17) CYCLANILIDE: MECHANISM OF ACTION AND USES AS A PLANT GROWTH REGULATOR IN COTTON Jim Burton 1 and Marianne Pedersen Abstract. Cyclanilide [1-(2,4-dichlorophenylaminocarbonyl)-cyclopropane carboxylic
Chlamydia trachomatis Infection Inhibits Both Bax and Bak Activation Induced by Staurosporine
 INFECTION AND IMMUNITY, Sept. 2004, p. 5470 5474 Vol. 72, No. 9 0019-9567/04/$08.00 0 DOI: 10.1128/IAI.72.9.5470 5474.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Chlamydia
INFECTION AND IMMUNITY, Sept. 2004, p. 5470 5474 Vol. 72, No. 9 0019-9567/04/$08.00 0 DOI: 10.1128/IAI.72.9.5470 5474.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Chlamydia
Chapter 4. Regulation of caspase-3 gene expression in MCF-7 cells by doxorubicin
 Chapter 4 Regulation of caspase-3 gene expression in MCF-7 cells by doxorubicin 4.1 Introduction The tumor suppressor p53 plays a major role in regulating the cellular response to various stresses and
Chapter 4 Regulation of caspase-3 gene expression in MCF-7 cells by doxorubicin 4.1 Introduction The tumor suppressor p53 plays a major role in regulating the cellular response to various stresses and
The Caspase System: a potential role in muscle proteolysis and meat quality? Tim Parr
 The Caspase System: a potential role in muscle proteolysis and meat quality? Tim Parr Caroline Kemp, Ron Bardsley,, Peter Buttery Division of Nutritional Sciences, School of Biosciences, University of
The Caspase System: a potential role in muscle proteolysis and meat quality? Tim Parr Caroline Kemp, Ron Bardsley,, Peter Buttery Division of Nutritional Sciences, School of Biosciences, University of
For the rapid, sensitive and accurate detection of active Caspase 9 in living cells
 ab65615 Caspase 9 (active) FITC Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of active Caspase 9 in living cells This product is for research use only and is not intended
ab65615 Caspase 9 (active) FITC Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of active Caspase 9 in living cells This product is for research use only and is not intended
Bio 3411, Fall 2006, Lecture 19-Cell Death.
 Types of Cell Death Questions : Apoptosis (Programmed Cell Death) : Cell-Autonomous Stereotypic Rapid Clean (dead cells eaten) Necrosis : Not Self-Initiated Not Stereotypic Can Be Slow Messy (injury can
Types of Cell Death Questions : Apoptosis (Programmed Cell Death) : Cell-Autonomous Stereotypic Rapid Clean (dead cells eaten) Necrosis : Not Self-Initiated Not Stereotypic Can Be Slow Messy (injury can
Apoptosis and Cancer. Carol M. Troy, MD, PhD October 26, 2016
 Apoptosis and Cancer Carol M. Troy, MD, PhD October 26, 2016 PHENOMENOLOGY OF CELL DEATH -1 I. DEVELOPMENT A. MORPHOGENESIS: SCULPTING/SHAPING STRUCTURES FUSION OF TISSUE MASSES (PALATE/NEURAL TUBE) CREATION
Apoptosis and Cancer Carol M. Troy, MD, PhD October 26, 2016 PHENOMENOLOGY OF CELL DEATH -1 I. DEVELOPMENT A. MORPHOGENESIS: SCULPTING/SHAPING STRUCTURES FUSION OF TISSUE MASSES (PALATE/NEURAL TUBE) CREATION
7.06 Problem Set
 7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
Supporting Information
 Supporting Information A Low-Temperature Solid-Phase Method to Synthesize Highly Fluorescent Carbon Nitride Dots with Tunable Emission Juan Zhou, Yong Yang, and Chun-yang Zhang* Single-Molecule Detection
Supporting Information A Low-Temperature Solid-Phase Method to Synthesize Highly Fluorescent Carbon Nitride Dots with Tunable Emission Juan Zhou, Yong Yang, and Chun-yang Zhang* Single-Molecule Detection
7.06 Cell Biology EXAM #3 April 21, 2005
 7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
Mechanisms of Cell Death
 Mechanisms of Cell Death CELL DEATH AND FORMATION OF THE SEMICIRCULAR CANALS Carol M. Troy, MD PhD August 24, 2009 FROM: Fekete et al., Development 124: 2451 (1997) PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT
Mechanisms of Cell Death CELL DEATH AND FORMATION OF THE SEMICIRCULAR CANALS Carol M. Troy, MD PhD August 24, 2009 FROM: Fekete et al., Development 124: 2451 (1997) PHENOMENOLOGY OF CELL DEATH I. DEVELOPMENT
IAPs. Structure of BIR2 Zn finger domain from XIAP
 IAPs The only known endogenous caspase inhibitors are members of a second family of proteins identified by apoptosis research, the IAP (inhibitor of apoptosis proteins) family. IAPs were originally described
IAPs The only known endogenous caspase inhibitors are members of a second family of proteins identified by apoptosis research, the IAP (inhibitor of apoptosis proteins) family. IAPs were originally described
For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates.
 ab65608 Caspase 9 Assay Kit (Colorimetric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates. This product is for research use only
ab65608 Caspase 9 Assay Kit (Colorimetric) Instructions for Use For the rapid, sensitive and accurate measurement of Caspase 9 activity in cell and tissue lysates. This product is for research use only
Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing
 Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing 0.1% SDS without boiling. The gel was analyzed by a fluorescent
Supplementary Figure 1. SDS-PAGE analysis of GFP oligomer variants with different linkers. Oligomer mixtures were applied to a PAGE gel containing 0.1% SDS without boiling. The gel was analyzed by a fluorescent
Measuring Mitochondrial Membrane Potential with JC-1 Using the Cellometer Vision Image Cytometer
 Measuring Mitochondrial Membrane Potential with JC-1 Using the Cellometer Vision Image Cytometer Nexcelom Bioscience LLC. 360 Merrimack Street, Building 9 Lawrence, MA 01843 T: 978.327.5340 F: 978.327.5341
Measuring Mitochondrial Membrane Potential with JC-1 Using the Cellometer Vision Image Cytometer Nexcelom Bioscience LLC. 360 Merrimack Street, Building 9 Lawrence, MA 01843 T: 978.327.5340 F: 978.327.5341
Bax and Bak Coalesce into Novel Mitochondria-associated Clusters during Apoptosis
 Bax and Bak Coalesce into Novel Mitochondria-associated Clusters during Apoptosis Amotz Nechushtan,* Carolyn L. Smith, Itschak Lamensdorf, Soo-Han Yoon,* and Richard J. Youle* *Biochemistry Section, Surgical
Bax and Bak Coalesce into Novel Mitochondria-associated Clusters during Apoptosis Amotz Nechushtan,* Carolyn L. Smith, Itschak Lamensdorf, Soo-Han Yoon,* and Richard J. Youle* *Biochemistry Section, Surgical
Multifunctional polyphosphazene-coated multi-walled carbon. nanotubes for the synergistic treatment of redox-responsive
 Electronic Supplementary Material (ESI) for Polymer Chemistry. This journal is The Royal Society of Chemistry 2017 Supporting information for Multifunctional polyphosphazene-coated multi-walled carbon
Electronic Supplementary Material (ESI) for Polymer Chemistry. This journal is The Royal Society of Chemistry 2017 Supporting information for Multifunctional polyphosphazene-coated multi-walled carbon
BCL-2, BCL-X L Sequester BH3 Domain-Only Molecules Preventing BAX- and BAK-Mediated Mitochondrial Apoptosis
 Molecular Cell, Vol. 8, 705 711, September, 2001, Copyright 2001 by Cell Press BCL-2, BCL-X L Sequester BH3 Domain-Only Molecules Preventing BAX- and BAK-Mediated Mitochondrial Apoptosis Emily H.-Y. A.
Molecular Cell, Vol. 8, 705 711, September, 2001, Copyright 2001 by Cell Press BCL-2, BCL-X L Sequester BH3 Domain-Only Molecules Preventing BAX- and BAK-Mediated Mitochondrial Apoptosis Emily H.-Y. A.
targets. clustering show that different complex pathway
 Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Caspase 3 (active) Red Staining Kit
 Red Staining Kit") ab65617 Caspase 3 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 3 in living cells. This product is for research use only and is not
ab65617 Caspase 3 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 3 in living cells. This product is for research use only and is not
Co-localization Analysis of Bak-Drp1 on Mitochondria Using Super-resolution Microscopy
 Co-localization Analysis of Bak-Drp1 on Mitochondria Using Super-resolution Microscopy Danzi Song University of Tokyo Research Internship Program Ozawa Laboratory Abstract The mechanistic connection between
Co-localization Analysis of Bak-Drp1 on Mitochondria Using Super-resolution Microscopy Danzi Song University of Tokyo Research Internship Program Ozawa Laboratory Abstract The mechanistic connection between
Caspase 3 (active) Red Staining Kit
 Red Staining Kit") ab65617 Caspase 3 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 3 in living cells. This product is for research use only and is not
ab65617 Caspase 3 (active) Red Staining Kit Instructions for Use For the rapid, sensitive and accurate detection of activated Caspase 3 in living cells. This product is for research use only and is not
Lab 2A: Sub-Cellular Fractionation
 Lab 2A: Sub-Cellular Fractionation A response is required for each item marked: (# ). Your grade for the lab 2 report (2A and 2B combined) will be the fraction of correct responses on a 50 point scale[(#
Lab 2A: Sub-Cellular Fractionation A response is required for each item marked: (# ). Your grade for the lab 2 report (2A and 2B combined) will be the fraction of correct responses on a 50 point scale[(#
Biology. Expression of BID in SH-SY5Y Cells After Acute Exposure to Rotenone
 26 Biology Expression of BID in SH-SY5Y Cells After Acute Exposure to Rotenone Sponsoring Faculty Member: Dr. Melinda Pomeroy-Black Hunter Connell and Melinda Pomeroy-Black Introduction Parkinson s Disease
26 Biology Expression of BID in SH-SY5Y Cells After Acute Exposure to Rotenone Sponsoring Faculty Member: Dr. Melinda Pomeroy-Black Hunter Connell and Melinda Pomeroy-Black Introduction Parkinson s Disease
Contains ribosomes attached to the endoplasmic reticulum. Genetic material consists of linear chromosomes. Diameter of the cell is 1 m
 1. (a) Complete each box in the table, which compares a prokaryotic and a eukaryotic cell, with a tick if the statement is correct or a cross if it is incorrect. Prokaryotic cell Eukaryotic cell Contains
1. (a) Complete each box in the table, which compares a prokaryotic and a eukaryotic cell, with a tick if the statement is correct or a cross if it is incorrect. Prokaryotic cell Eukaryotic cell Contains
Supplementary Figure 1.
 Supplementary Figure 1. Characterisation of IHG-1 overexpressing and knockdown cell lines. (A) Total cellular RNA was prepared from HeLa cells stably overexpressing IHG-1 or mts-ihg-1. IHG-1 mrna was quantified
Supplementary Figure 1. Characterisation of IHG-1 overexpressing and knockdown cell lines. (A) Total cellular RNA was prepared from HeLa cells stably overexpressing IHG-1 or mts-ihg-1. IHG-1 mrna was quantified
Matrix-assisted laser desorption ionization/time of flight (MALDI/TOF) mass spectra were obtained on
 mass spectra were obtained on") MALDI-TOF Mass Spectroscopy Matrix-assisted laser desorption ionization/time of flight (MALDI/TOF) mass spectra were obtained on Ultraflextreme (Bruker Daltonics, Germany) mass spectrometer using sinapinic
MALDI-TOF Mass Spectroscopy Matrix-assisted laser desorption ionization/time of flight (MALDI/TOF) mass spectra were obtained on Ultraflextreme (Bruker Daltonics, Germany) mass spectrometer using sinapinic
Death signaling. Color PDF file of handouts can be found at Wu lab web-page:
 Death signaling Hao Wu References 1. Wang, X. The expanding role of mitochondria in apoptosis. Genes Dev 15, 2922-2933. (2001). 2. Fesik, S. W. Insights into programmed cell death through structural biology.
Death signaling Hao Wu References 1. Wang, X. The expanding role of mitochondria in apoptosis. Genes Dev 15, 2922-2933. (2001). 2. Fesik, S. W. Insights into programmed cell death through structural biology.
Biochimica et Biophysica Acta
 Biochimica et Biophysica Acta 1843 (2014) 2100 2113 Contents lists available at ScienceDirect Biochimica et Biophysica Acta journal homepage: www.elsevier.com/locate/bbamcr Review Regulating cell death
Biochimica et Biophysica Acta 1843 (2014) 2100 2113 Contents lists available at ScienceDirect Biochimica et Biophysica Acta journal homepage: www.elsevier.com/locate/bbamcr Review Regulating cell death
Nexcelom ViaStain Live Caspase 3/7 Detection for 2D/3D Culture
 Nexcelom ViaStain Live Caspase 3/7 Detection for 2D/3D Culture Product Numbers: CSK-V0002-1, CSK-V0003-1 This product is for RESEARCH USE ONLY and is not approved for diagnostic or therapeutic use. Table
Nexcelom ViaStain Live Caspase 3/7 Detection for 2D/3D Culture Product Numbers: CSK-V0002-1, CSK-V0003-1 This product is for RESEARCH USE ONLY and is not approved for diagnostic or therapeutic use. Table
Spherotech, Inc Irma Lee Circle, Unit 101, Lake Forest, Illinois PARTICLE COATING PROCEDURES
 SPHERO TM Technical Note STN-1 Rev C. 041106 Introduction Currently, there are several methods of attaching biological ligands to polystyrene particles. These methods include adsorption to plain polystyrene
SPHERO TM Technical Note STN-1 Rev C. 041106 Introduction Currently, there are several methods of attaching biological ligands to polystyrene particles. These methods include adsorption to plain polystyrene
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins
 13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
Preparing Colloidal Gold for Electron Microscopy
 Corporate Headquarters 400 Valley Road Warrington, PA 18976 1-800-523-2575 FAX 1-800-343-3291 Email: info@polysciences.com www.polysciences.com Europe - Germany Polysciences Europe GmbH Handelsstr. 3 D-69214
Corporate Headquarters 400 Valley Road Warrington, PA 18976 1-800-523-2575 FAX 1-800-343-3291 Email: info@polysciences.com www.polysciences.com Europe - Germany Polysciences Europe GmbH Handelsstr. 3 D-69214