G3: Genes Genomes Genetics Early Online, published on March 3, 2017 as doi: /g
|
|
|
- Hilary Warren
- 5 years ago
- Views:
Transcription
1 G3: Genes Genomes Genetics Early Online, published on March 3, 2017 as doi: /g Planar cell polarity effector Fritz interacts with Dishevelled and has multiple functions in regulating PCP Ying Wang, Victor F. Naturale and Paul N. Adler Biology Department and Cell Biology Department University of Virginia Charlottesville, VA 22903, USA 1 The Author(s) Published by the Genetics Society of America.
2 8 The role of frtz in Planar Cell Polarity Keywords: Drosophila; Planar Polarity Effector; Planar Cell Polarity; fritz; inturned 13 2
3 14 Abstract The Planar cell Polarity Effector (PPE) genes inturned, fuzzy and fritz are downstream components in the frizzled/starry night signaling pathway, and their function is instructed by upstream Planar Cell Polarity (PCP) core genes such as frizzled and dishevelled. PPE proteins accumulate asymmetrically in wing cells and function in a protein complex mediated by direct interactions between In and Frtz and In and Fy. How the PCP proteins instruct the accumulation of PPE protein is unknown. We found a likely direct interaction between Dishevelled and Fritz and Dishevelled and Fuzzy that could play a role in this. We previously found that mild over expression of frtz rescued a weak in allele. To determine if this was due to extra Frtz stabilizing mutant In or due to Frtz being able to bypass the need for In we generate a precise deletion of the inturned gene (in PD ). We found that mild overexpression of Fritz partially rescued in PD, indicating that fritz has In independent activity in PCP. Previous studies of PPE proteins used fixed tissues, and did not provide any insights into the dynamic properties of PPE proteins. We used CRISPR/Cas9 genome editing technology to edit the fritz gene to add a green fluorescent protein tag. fritz mneongreen provides complete rescue activity and works well for in vivo imaging. Our data showed that Fritz is very dynamic in epidermal cells and preferentially distributed to discrete membrane subdomains ( puncta ). Surprisingly, we found it in stripes in developing bristles. 3
4 32 Introduction Many tissues in animals display Planar Cell Polarity (PCP). The coordinate alignment of bristles and cuticular hairs in insects, scales in fish, feathers in birds and hairs in mammals are easily visualized examples of PCP (JENNY AND MLODZIK 2006). PCP also regulates the development of internal organs and tissues. For example, stereocilia in the mammalian inner ear and ommatidia in the Drosophila eye show PCP in their uniform orientation (ZHENG et al. 1995; WOLFF AND RUBIN 1998; DABDOUB AND KELLEY 2005). Convergent extension, aspects of kidney development and several other developmental processes are also controlled by PCP (KELLER et al. 2000; WALLINGFORD et al. 2000; WALLINGFORD et al. 2002; BABAYEVA et al. 2013; GOGGOLIDOU 2014). Over the past several decades the importance of the frizzled (fz)/starry night (stan) signaling pathway in controlling PCP in both vertebrates and invertebrates has been intensively studied (ZHENG et al. 1995; WALLINGFORD et al. 2000; WANG et al. 2005; WANG AND NATHANS 2007; GOODRICH AND STRUTT 2011; ADLER 2012; GOGGOLIDOU 2014). Early research on Drosophila identified several groups of genes including the upstream PCP core genes and the downstream Planar Polarity Effector (PPE) genes. This pathway has been most intensively studied on the fly wing, where in wild type each cell forms a single distally pointing hair (WONG AND ADLER 1993) (ADLER 2012). Mutations in the upstream genes result in most cells producing a hair with abnormal polarity. Rarely, cells form two or three hairs. Mutations in the downstream PPE genes result in similar abnormal hair polarity but notably many cells form more than one hair (usually two or three) (WONG AND ADLER 1993) (ADLER 2012). The protein products of all the genes in the pathway accumulate asymmetrically in epithelial cells and this is thought to play a key role in their function (WONG AND ADLER 1993; USUI et al. 1999; AXELROD 2001; FEIGUIN et al. 2001; STRUTT 2001; TREE et al. 2002; BASTOCK et al. 2003; JENNY et al. 2003; ADLER et al. 2004; DAS et al. 2004; COLLIER et al. 2005; STRUTT AND WARRINGTON 2008). The PPE group of gene has not been as well studied as the upstream genes. The group includes three genes: inturned (in), fuzzy (fy) and fritz (frtz). The asymmetric accumulation of these proteins is dependent on 4
5 and instructed by the upstream PCP proteins (ADLER et al. 2004; STRUTT AND WARRINGTON 2008). In PCP mutants the PPE proteins do not preferentially accumulate on the proximal edge of wing cells but they retain some activity as the frequent multiple hair cells seen in PPE mutants are not seen in PCP mutants. The mechanism by which the PCP proteins instruct the accumulation of PPE proteins is unknown although in mammals the upstream Dishevelled (Dsh) protein has been shown to bind the Fuz PPE protein (fuz is the mammalian homolog of fy) (ZILBER et al. 2013). We found this interaction to be conserved in Drosophila and that the Frtz PPE protein can also bind Dsh. A variety of information has supported the idea that the 3 Drosophila PPE genes and proteins function together as a unit in fly PCP. This was first suggested due to the similarity of the mutant phenotypes for these genes and because double mutants for strong alleles of any of the PPE genes show a similar phenotype to each single mutant (WONG AND ADLER 1993; ADLER et al. 2004; COLLIER et al. 2005). Further double mutants of weak alleles of any two PPE genes resulted in a synergistic interaction to produce a strong phenotype (COLLIER et al. 2005). It is also known that the accumulation of each of the PPE proteins is dependent on the other members of the group although there is some complexity to this interaction (ADLER et al. 2004; STRUTT AND WARRINGTON 2008; WANG et al. 2014). Loss of function mutations in any of these genes results in a decrease in the level of endogenous Frtz and In (WANG et al. 2014) (ADLER et al. 2004)(STRUTT AND WARRINGTON 2008). We also previously reported that moderate overexpression of frtz increased the accumulation of endogenous In while over expression of fy decreased the accumulation of endogenous In (WANG et al. 2014). We suggested this was responsible for the ability of frtz over expression to suppress and the ability of fy over expression to enhance a temperature sensitive hypomorphic allele of in (in II53 ) (WANG et al. 2014). However we could not rule out the possibility that these genetic interactions were due to over expressed frtz or fy being able to bypass the need for functional in. To determine if all of the phenotypic interactions could simply be explained by frtz or fy stabilizing or destabilizing mutant In we generated a precise deletion of the in gene, in PD, using the CRISPR/Cas9 genome editing technique. We found overexpression of frtz could partially 5
6 82 83 rescue in PD and that the over expression of fy could enhance in PD establishing that frtz and fy can function partly in PCP in an In independent manner To study the Frtz protein in living cells when expressed at normal levels we used CRISPR/Cas9 to add a green fluorescent protein, mneongreen, to the carboxyl terminus of frtz gene. This edited gene displayed normal frtz function. In vivo imaging of the Frtz mneongreen showed that Frtz accumulated asymmetrically in wing, thorax, abdominal and arista cells. Using time-lapse in vivo imaging, we found Frtz was very dynamic and preferentially distributed to discrete membrane subdomains. Fluorescence recovery after photo-bleaching (FRAP) studies established that the dynamics of Frtz accumulation was similar in the several cell types studied. We also found that Frtz accumulated in stripes in bristles and the pattern was dependent on the actin cytoskeleton Materials and Methods 94 Fly genetics All flies were raised at 25 C unless otherwise stated. Mutant stocks were either obtained from the Bloomington Drosophila stock center at Indiana University, from the VDRC, generated in our lab or were generous gifts from J. Axelrod, D. Strutt, or T. Uemura. To direct transgene expression, we used the Gal4/UAS system (BRAND AND PERRIMON 1993). Dsh-myc stock was obtained from Bloomington Drosophila Stock Center at Indiana University (stock number 25385). We also generated transgenic flies that expressed myc-frtz-gfp from the ubiquitin promoter/enhancer (ubi-myc-frtz-gfp) (SEKELSKY et al. 1995; WANG et al. 2014). This transgene provided complete rescue of frtz phenotypic null alleles. 103 Antibodies 6
7 Anti-Myc antibody was obtained from Cell Signaling Technology. Anti-GFP antibody was obtained from Molecular Probes. Alexa 488- and Alexa 568-conjugated secondary antibodies were purchased from Molecular Probes. Anti-Frtz antibody was generated in rats using a Frtz protein made in E. coli and His tagged to facilitate purification. The expressed protein was found in inclusion bodies and solubilized and we do not know if it is in a native conformation. It was not able to detect the endogenous protein. The anti-in monoclonal antibody was generated in our laboratory and described in a previous paper (ADLER et al. 2004). 111 Immunostaining A standard paraformaldehyde fixation and staining procedure was used (ADLER et al. 2004). Briefly, fly pupae were fixed in 4% paraformaldehyde, PBS for 1-2 hr at 4 C. After fixation, wings were dissected, rinsed, blocked and then stained overnight at 4 C with primary antibodies in PBS, 0.3% Triton X-100 and 10% goat-serum. Secondary antibodies were applied for 2-3 hr at room temperature Isolation of flies carrying edited genes In all injections to edit genes, two types of plasmids were injected into embryos. Plasmids that drive expression of the grnas from the U6 promoter (U6:1 or U6:3 promoter) and homology dependent repair templates. The details of the construction of these plasmids can be found in the File S1. The DNA injection into embryos was performed at Rainbow Transgenic Flies, Inc in PD : Both grna expressing plasmids and HDR repair template were injected into Nos- cas9 attp40 embryos. The grnas were targeted to sequences located 5 and 3 to the in gene. 7
8 We obtained 30 adults from this injection and these were crossed to in IH56 /TM3 flies. Four out of the 30 G0 flies gave rise to in mutant G1 progeny, all of which were subsequently found to be the result of a precise deletion of the in gene (in PD ). Stocks were established and confirmed by genomic DNA PCR followed by DNA sequencing of PCR products. One of the lines expressed Ds-Red and was due to homology dependent repair. The other 3 lines did not express Ds-Red and were presumably due to non-homologous end joining Ruby-in: Both grna plasmid and HDR template were injected into Nos-cas9 attp40 (18055). The G1 progeny carrying the inserts were selected by Ds-Red fluorescence, stocks established and confirmed by genomic DNA PCR followed by DNA sequencing of PCR products. The Ds-Red segment was removed by crossing to a hs-cre stock frtz-mneongreen: There are seven exons in frtz gene. The two grna constructs were designed to specifically cut in the 7 th exon of frtz. The homology directed repair (HDR) template included a mneongreen sequence, a DS-Red sequence to screen for edit containing flies and two 1.5kb homology arms flanking both ends of the grnas cuts and the frtz 7 th exon (Figure 5A). Figure 5A shows the frtz gene map, the two grnas targeting sites and the design of the HDR template (Figure 5A). Both the grna plasmid and HDR template were injected into a nos-cas9 attp2 (21104) stock. The G0 flies were crossed to w; CyO/Gla flies and the G1 progeny screened for Ds-Red fluorescence. Genomic DNA was extracted from adult flies with Ds-Red fluorescence and PCR done to validate edited genes. 50% of G0 progeny gave rise to G1 progeny that displayed Ds-Red fluorescence and 20% of those were confirmed to have an insertion of the mneongreen sequence at the correct genomic location. The Ds-Red segment was removed by crossing to a hs-cre stock. 8
9 147 Yeast two hybrid assays: For the yeast two hybrid assays, we used yeast strain AH109 and Matchmaker Two- Hybrid System 3 from Clontech. Briefly, full length or truncated cdna was sub-cloned into pgadt7 or pgbkt7 vector using specific restriction enzyme sites or Gibson assembly. To test the interaction of defined protein partners, pgadt7 and pgbkt7 plasmids containing one of the two protein partners were co-transformed into yeasts. Transformed yeasts were selected by growing on two-marker dropout medium (SD/-Leu/-Trp). These two markers select for the presence of both plasmids. Colonies from the previous dropout medium were transferred to a four-marker dropout medium (SD/-Ade/-His/-Leu/-Trp) with X-alpha-Gal. Only colonies growing on four-marker plates were considered as evidence for a positive interaction Quick Preparation of Genomic DNA from Drosophila Buffer A (store at room temperature): 100 mm Tris-Cl (ph 7.5); 100 mm EDTA; 100 mm NaCl; 0.5% SDS Buffer B (store at 4ºC): 200 ml potassium acetate (5 M); 500 ml lithium chloride (6 M) About 30 anesthetized flies were collected in a 1.5-mL micro-centrifuge tube and ground in 200ul buffer A with a disposable tissue grinder. An additional 200 µl of buffer A was added (to a total volume of 400 µl) and continue grinding until only cuticles remain. Samples were incubated at 65ºC for 30 min and 800ul buffer B was added to each sample and mix well by inverting the tube multiple times, and then incubate on ice for at least 10 min and up to a few hours. Centrifuge at 12,000 rpm for 15 min at room temperature. Transfer 1 ml of the 9
10 supernatant into a new micro-centrifuge and add add 600 µl of isopropanol to each sample, and mix well by inverting the tube several times. Centrifuge at 12,000 rpm for 15 min at room temperature. Discard the supernatant. Wash the pellet with 70% ethanol, air-dry, and re-suspend in 150 µl of TE buffer. 172 Coimmunoprecipitation and western blotting 173 The following genotypes were used in these experiments: 174 w; ptc-gal4 dsh-myc ; UAS-fy-GFP 175 w; ptc-gal4 dsh-myc ;UAS-(myc) 6 -frtz wing discs of transgenic flies were dissected from third instar larvae and homogenized in pre-chilled lysis buffer containing protease inhibitors. The lysis buffer (and other reagents) came from a kit (Roche ) and contained 50mM Tris HCl, ph7.5; 150 mm NaCl; 1% Nonidet P40; 0.5% deoxycholate). The extract was then spun to remove cell debris. Protein A agarose beads (Roche) were added to the extract and incubated for at least 3 hr to reduce the background of non-specific binding. The extract was spun again and 7 10 µl desired antibody was added to the supernatant and incubated for 3 4 hr. New protein A agarose beads were then added and the sample was incubated overnight. The beads were pelleted and washed. Protein was released from the beads and analyzed by standard western blot procedures. 185 Imaging 10
11 Adult wings were mounted in Euparal and bright field images obtained using a Spot RT camera and a Zeiss Axiskop II microscope. In most cases a 40X Apochromatic objective was used (oil, 1.4 NA) Most live imaging was done using a Zeiss 780 confocal microscope and a 63x or 40x oil immersion lens. One panel was obtained using a Zeiss 880 confocal microscope equipped with an Airy Scanner. Fixed and stained samples were imaged using a Zeiss 780 confocal microscope 192 and a 63x or 40x oil immersion lens. Final panels were assembled using Adobe Illustrator or Microsoft PowerPoint. Frtz mneongreen, ubi-myc-frtz-gfp, ubi-myc-venus-in and ubi-myc-frtzmcherry white pupae were collected then aged at 25 C for either 30 hr for imaging wing, thorax and arista cells or for about 42 hr for imaging abdominal cells. Aged pupae were placed on double-side tape attached on microscope slides. The pupal cases were peeled back over the region to be imaged. Two silicon rubber spacers were used to build a chamber and one small drop of halocarbon oil was applied to the cover slip to create a good optical interface For FRAP experiments we routinely bleached a part of one or more cells that contained an edge where the fluorescent protein preferentially accumulated. Recovery was typically followed for from 5-10 minutes with images taken from every 30 seconds to every 2 minutes. The age of the pupae examined depended on the tissue being examined and we used time points where the fluorescence was optimal. For example on the wing we used hr awp pupae. 204 Data Policy Key fly lines described for the first time in this paper (e.g. frtz NeonGreen ) will be sent to the Drosophila stock center in Bloomington to insure wide availability to the community. Other 11
12 stocks will be provided by the PI (PNA) upon request. Plasmids and aliquots of antibodies will also be provided upon request. Substantial data and methods information is provided in the Supplemental Material (File S1). The PI is happy to provide additional data upon request Results 212 Reanalyzing Frtz Structure In the original characterization of the frtz gene and protein the SMART annotation tool (SCHULTZ et al. 1998; COLLIER et al. 2005) identified a pair of tandem WD40 repeats from amino acids 296 to 375. We re-analyzed the Frtz protein structure using WD40-repeat protein Structure Predictor (WDSP), which was developed specifically to identify WD40 repeats and predict the secondary structures of WD40 domains (WU et al. 2012; WANG et al. 2015). Six WD40 repeats were predicted in the protein using WDSP (Table S1A). Similarly, seven WD40 repeats were predicted in the human Frtz (hfrtz) protein, which is five more than previously suggested (CUI et al. 2013)(Table S1B). We previously established that the ability of Frtz to bind to In as assayed by the yeast two hybrid system was associated with a fragment that contained the previously identified WD40 domains (WANG et al. 2014). This fragment contained aa 296 to aa 375 in Frtz. Since fragments containing the other putative WD40 domains did not give a positive reaction it appears that In cannot bind to a generic WD40 motif but rather to specific surfaces in WD40-4 and WD-5 of Frtz. We attempted to identify residues critical for the interaction of Frtz and In by using the analysis of the Frtz WD4 and WD-5 repeats from WDSP and CN3D. We mutated to alanine the two WDSP predicted hotspots on the top surface and 10 additional amino acid residues predicted by CN3D to be on the top, inner or outer peripheral 12
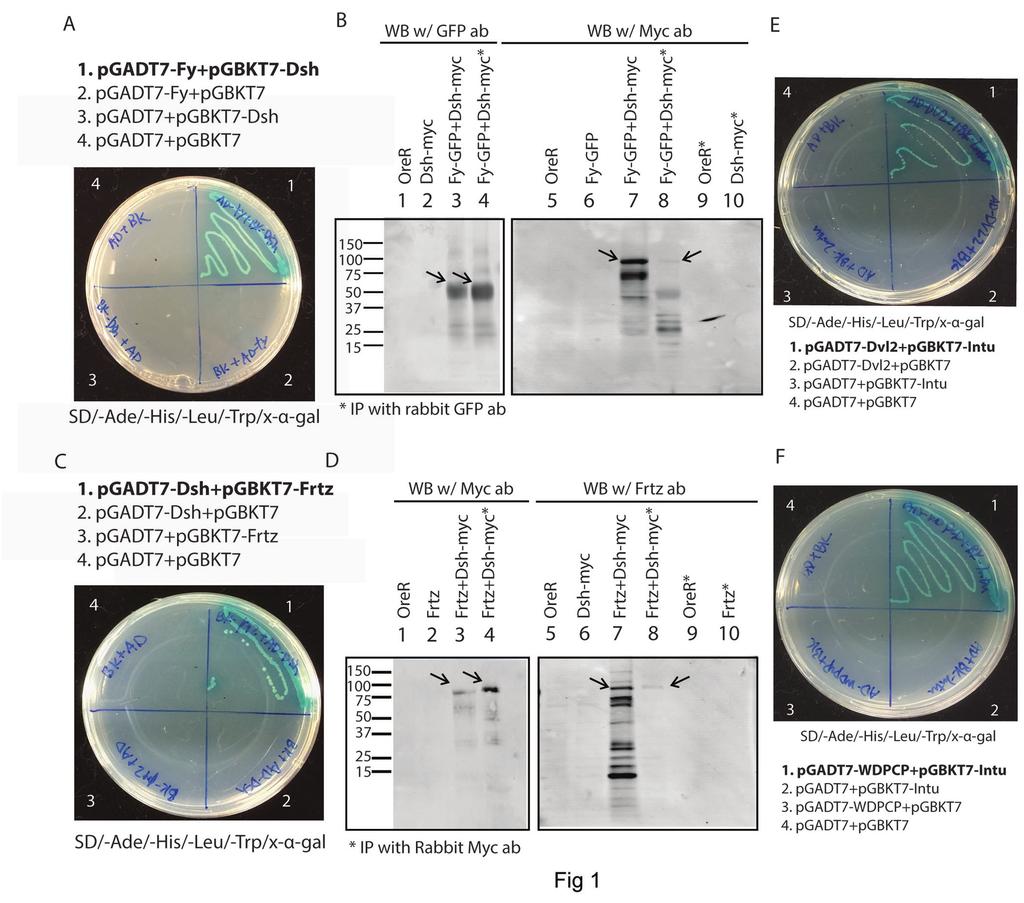
13 surfaces of the WD-4 and WD-5 repeats, except for A343 and A377 which were mutated to Glycine (Table S2). All 12 mutations were introduced into a single Drosophila Frtz WD40 fragment including the WD-4 and WD-5 repeats. Surprisingly, this highly mutated fragment still interacted strongly with In in the yeast-two-hybrid system. We conclude from this result that an alternative surface (e.g. bottom) in WD-4 and 5 is likely responsible for mediating the binding of Frtz to In Dsh interacts with Fy and Frtz The In, Fy and Frtz proteins accumulate on the proximal side of wing cells (Adler et al., 2004; Collier et al., 2005; Strutt and Warrington, 2008; Wang et al., 2014) and function together in a protein complex (WANG et al. 2014). Early studies also showed the accumulation of PPE proteins Frtz and In on the proximal edge of wing cells was decreased in fz and Vang mutant clones (ADLER et al. 2004; STRUTT AND WARRINGTON 2008). Together it suggests one or more PCP core proteins are required for the asymmetric recruitments of PPE proteins or protein complex to the proximal edge. Using co-immunoprecipitation and the yeast-two-hybrid system, we found that the distally localized protein Dsh interacted with both Frtz and Fy (Figure 1A- D)(Table S3). Since an interaction between the mammalian Dsh homolog Dvl2 and the Fy homolog Fuz has been demonstrated (ZILBER et al. 2013) we emphasized studying the interaction between Frtz and Dsh. In order to determine what part of Frtz was responsible for the interaction with Dsh, several Frtz fragments were generated (Figure S1A), but only full length Frtz interacted with Dsh, suggesting the interaction came from multiple locations all of which are required or that the protein as a whole is needed to fold into a conformation that allows the interaction with Dsh. In 13
14 order to test these hypotheses, we generated a set of internally deleted frtz genes where fragments 300 bp or 450 bp were deleted from the full length cdna. These alterations resulted in 100 or 150 amino acid deletions (Figure S1B). Each deletion was tested for an interaction with full length Dsh in the yeast-two-hybrid assay. The yeast-two-hybrid data showed that Frtz proteins where aa , , , or were deleted still interacted with Dsh. In contrast Frtz proteins where aa or were deleted failed to interact with Dsh. The data argued that these two regions mediated the interaction between Frtz and Dsh and that both regions are required. WD40 domains 1 to 3 are found in the fragment and no WD40 domains are present in aa Thus, the WD40 domains are not sufficient for the interaction and other sequences are also required. To determine if these interaction are conserved for their mammalian homologs, human intu (in homolog), wdpcp (frtz homolog) and dvl2 (dsh homolog) were sub-cloned into yeasttwo-hybrid vectors and protein interaction assays were performed (Table S3). We found Intu interacted with both Dvl2 and WDPCP (Figure 1EF). Thus, the interaction between In and Frtz detected in our studies on the fly proteins appears to be conserved. However we did not see an interaction between In and Dsh, thus for these two proteins the fly model does not appear to mimic the human system. We also did not see an interaction between Dvl2 and WDPCP. Thus, the interaction between Frtz and Dsh that we detected with fly proteins may not be conserved in the human proteins. It remains possible that WDPCP interacts with another one of the Dvl isoforms, such as Dvl3. Generation of a precise deletion of in (in PD ) using CRIPSR/Cas9 Previously, we described that the mild overexpression of Frtz almost completely suppressed the mutant phenotype of the temperature sensitive hypomorphic allele in II53 and the 14
15 over expression of Fy enhanced the in II53 phenotype (WANG et al. 2014). More recently we found the mild over expression of Frtz could suppress the strong in IH56 allele (Figure S2) This allele is due to a nonsense mutation as are most in alleles (ADLER et al. 1994). Genes containing nonsense mutations can produce protein by read-through (STENEBERG AND SAMAKOVLIS 2001; WANG et al. 2001), and it seemed possible that the suppression of in IH56 by Frtz over expression could be due to read through and the resulting small amount of In protein being stabilized by the extra Frtz (WANG et al. 2014). Alternatively, it is possible that over expressed Frtz can bypass the requirement for In. To distinguish between these models we needed to isolate an in mutation that we could be confident was a null allele. To generate an in null mutation, we used the CRISPR/Cas 9 genome editing technique (GRATZ et al. 2013a; GRATZ et al. 2013b; RAN et al. 2013). Figure 2A shows a genomic map of in and the location of the two grna targets and the HDR repair template. As described in the Methods we obtained 4 precise deletions of the in gene. Since the in gene is completely deleted in in PD, no In protein can be produced in the mutant. In a number of wing region in PD appeared to have a stronger mutant phenotype than in IH56 (Figure 2B,C). We quantified the frequency of cells that formed multiple hairs in two such regions of the wing and found the in PD wings had a stronger phenotype (Figure 2D-G), which suggests the in IH56 nonsense mutation produces some protein by read-through. This is consistent with the hypothesis that the frtz suppression of in alleles is due to the stabilizing of mutant In by over-expressed Frtz. The hair polarity phenotype also appeared somewhat stronger in in PD, but this phenotype was not quantified. We then tested if mild overexpression of frtz could suppress the mutant phenotype of in PD, similarly to in II53 and in IH56. apterous-gal4 (ap-gal4) was used to drive the expression of mycfrtz in the dorsal layer of the wing. We analyzed the phenotype in two wing regions by 15
16 observing hair polarity and by counting the number of multiple hair cells (Figure 3B-D ). Both hair polarity and hair numbers in in PD were weakly, but significantly rescued by overexpression of frtz (Figure 3G). We also observed a partial suppression of the abdominal bristle phenotype of in PD with over expression of frtz (Figure S3). We carried out an independent second test for the ability of frtz to suppress in PD using a fusion protein encoding gene (UAS-frtz-cherry-pk). The rationale for this experiment was that in the absence of In, Frtz would not localize to the proximal side of wing cells and that would hamper its ability to function in wing PCP. By fusing Frtz to Pk, which localizes proximally (TREE et al. 2002; JENNY et al. 2003), we hoped to drive Frtz to the proximal edge of wing cells independent of In. We established that the fusion protein had both Pk and In activity, as when expressed by act-gal4 it provided partial rescue of a frtz mutation (Figure S4BC). Further, when its expression was driven by the stronger ptc-gal4 driver a typical pk gain of function was seen (Figure S4EF). Again the in PD phenotype was weakly but significantly rescued by expressing the Frtz-Cherry-Pk fusion protein (Figure S4G-I). However, the rescue was not better than that seen with UAS-myc-frtz. This could be due to the relatively poor accumulation of the fusion protein, perhaps due to folding problems. The partial rescue of the in PD mutant phenotype in these experiments indicates that Frtz can function in part independently of In in PCP in Drosophila. Unlike the situation with in II53 and in IH56, the in PD mutant phenotype was only weakly rescued consistent with Frtz having important In dependent functions in PCP as well We previously found that the overexpression of Fy using ap-gal4 significantly enhanced the phenotype of in II53 and suggested this was due to Fy destabilizing the mutant In protein (WANG et al. 2014). Alternatively, over expressed Fy could be impacting PCP in an In independent manner. To distinguish between these hypotheses we tested if over expressed Fy 16
17 could enhance the in PD phenotype. We found that it could. Over expressing Fy using apgal4 the percentage of cell forming multiple hairs was significantly increased in both the anterior and central domain of the wing (Figure 3E-F,H). This established that Fy can impact PCP in an In independent manner Live imaging of transgenic Frtz protein The previous experiments on stained fixed wing tissues showed the Frtz protein asymmetrically localized at the proximal edge of wing cells (STRUTT AND WARRINGTON 2008). However, those experiments did not provide any insight into the dynamic property of the Frtz protein. To understand how the Frtz protein functioned in live cells we first utilized flies that carried the Ubi-myc-frtz-GFP transgene. We observed the tagged protein in the wings of living pupae and observed that Frtz protein accumulated in the zig-zag pattern reported in experiments with fixed cells (Figure 4A) (STRUTT AND WARRINGTON 2008). It was clear that the Frtz protein accumulated in discrete membrane subdomains ( puncta ) as is seen for the protein products of the upstream PCP genes (STRUTT et al. 2011). The puncta were very dynamic with several types of movement observed including but not limited to puncta movement along the membrane, puncta fusion, puncta splitting into two, puncta fusing with and leaving the membrane. Figure 4B-D shows several examples of these With the goal of simultaneously imaging multiple PPE proteins in vivo we generated a set of constructs and transgenic flies where different colored fluorescent proteins were linked to PPE proteins (see Supplemental File S1 for details) at the same PhiC31 landing site. Unfortunately, none of transgenes tested resulted in as good imaging as the P insertions of Ubimyc-frtz-GFP (see Figure 4). Using the attp1 landing site we were able to detect the zigzag 17
18 accumulation of Frtz and In (Table S4, Figure S5A & B). The in transgene resulted in a higher number of puncta than we had seen for previously the endogenous protein by immunostaining experiments (compare Figure S5B and C) (ADLER et al. 2004). It was unclear if the high number of puncta was due to the protein behaving abnormally due to being tagged or to the ubiquitin 349 enhancer/promoter resulting in an abnormal level of gene expression. In none of our experiments with Fy did we see asymmetric accumulation consistent with our previous attempts to detect tagged transgenic Fy using a transgene that contained the fy enhancer/promoter (Yan, 2008). All of the tagged transgenes provided full rescue of our strongest frtz, in and fy alleles so the tagged proteins had activity and were expressed at a level that was compatible with in vivo function. However, the tagged PPE transgenes were less than ideal as the proteins were not expressed at the endogenous level and in some tissues, such as the developing adult abdominal epidermis our ability to image the tagged protein was compromised by a higher level of expression in other cell types that were closely juxtaposed to the epidermis. With the development of CRISPR/Cas9 technology (GRATZ et al. 2013a; GRATZ et al. 2013b; RAN et al. 2013) the superior approach of tagging the endogenous gene became feasible and we took that approach We used CRISPR/Cas9 to add a green fluorescent protein, mneongreen (Allele Biotechnology), to the carboxy terminus of the endogenous frtz gene (see Methods and Figure 5A). The edited frtz mneongreen gene completely rescued a frtz loss-of-function mutant and was wild type when homozygous suggesting the edited gene and fusion protein is fully functional (Figure 5B). We observed the asymmetric accumulation of Frtz mneongreen protein in both wing and thorax cells at 28-30hr apf (after puparium formation) (Figure 6AB). 18
19 We also used the CRISPR/Cas9 to add the red fluorescent protein Ruby to the amino terminal end of in. The ruby-in gene provided rescue of in function and was wild type when homozygous. However, when we did in vivo imaging we observed a large number of puncta (Figure S5D), reminiscent of what we had observed with ubi-venus-in. This suggests the large number of puncta is a consequence of the fluorescent protein tag. Our best results in imaging Ruby-In came in experiments on the abdominal epithelial cells of 52 hr pupae and in these cells we could detect the asymmetric accumulation of Ruby-In, although there was substantial variation between individuals (see for example Figure S6EF). We did not examine this tissue in ubi-venus-in pupae. Further experiments will be required to find a location where a tag can be added without perturbing In dynamics in most cell types Frtz protein functions and asymmetrically accumulates in the abdomen. The adult abdomen stands out due to the phenotypic additivity of mutations in genes that are part of the fz/stan and ds/ft pathways (CASAL et al. 2006; DONOUGHE AND DINARDO 2011). It is known that PPE mutations alter bristle and hair polarity on the abdomen (GUBB AND GARCIA- BELLIDO 1982) but this aspect of PPE gene function has not been studied in depth. It is interesting that mutations in fz/stan PCP genes only result in grossly abnormal hair polarity over part of each abdominal segment (CASAL et al. 2006) (Figure 7B) although in our hands the proteins appear to accumulate asymmetrically throughout the segment (Figure S6AB). This has also been seen by others in the embryonic, larval and pupal abdomen (PRICE et al. 2006; DONOUGHE AND DINARDO 2011; SHARP AND AXELROD 2016). In contrast mutations in frtz, in and fy produce strong hair phenotypes in all regions of abdominal segments where cells produce hairs (Figure 7E-G). As in other body regions they consist of cells forming extra hairs and 19
20 abnormal hair polarity. In double mutants between fz and any of the PPE mutations the hair phenotype resembled the PPE phenotype (Figure 7I-K, I -K vs B, B and E-G, E -G ). Thus in terms of the abdominal hair phenotype the PPE mutations are epistatic to fz mutations, as is the case for the wing hair phenotype (WONG AND ADLER 1993). Although we did not study it in as much detail we also observed that in double mutants between ds and any of the PPE mutants the hair phenotype resembled the PPE phenotype (Figure 7 M-O, M -O compare with C-D, C -D ). As is the case for the PPE mutant wing hair phenotype the phenotype of PPE single and double mutants were similar (e.g. Figure 7 E-G, E -G vs LP and L P ). In contrast to the wing where the phenotype of mwh is stronger than the PPE mutants on the abdomen it was weaker than that of the PPE mutants (compare Figure 7 E-G vs H, E -G and H ). The dorsal abdominal bristle polarity of PPE mutants stands out due to bristles remaining aligned with their neighbors and a tendency to point to the midline (GUBB AND GARCIA-BELLIDO 1982). That differs from the more randomized bristle polarity of other PCP mutations. The bristle polarity phenotype of PPE, fz double mutants on the abdomen was intermediate between fz and PPE single mutants. Thus, in terms of the bristle polarity phenotype the PPE genes are not epistatic to the PCP genes. This is also true in the thorax. The basis for this is unclear. Around 43 hr apf, we observed the asymmetric accumulation of Frtz protein in cells throughout the developing adult abdomen (Figure 6C). We were unable to detect Frtz mneongreen in dsh mutants. This could be due to a lack of localization, reduced stability or both. This is similar to what has been seen in the wing (STRUTT AND WARRINGTON 2008). Similarly, we found that we could not detect Frtz mneongreen in in PD homozygous flies either, consistent with the requirement for in for the accumulation of endogenous Frtz (STRUTT AND WARRINGTON 2008). 20
21 These data suggest that the PPE proteins function downstream of the PCP proteins in abdominal development for specifying hair number and polarity Frtz protein functions and asymmetrically accumulates in the arista. Previous research in our lab showed the fz/stan signaling pathway regulates the development of Drosophila arista. Mutations in the PPE genes in and fy resulted in production of multipled and split laterals that were not evenly distributed along the central shaft (HE AND ADLER 2002). We found that mutations in frtz also produced such a phenotype (Figure S7). We also previously observed that the Fz protein accumulated asymmetrically in the cells that form the central core of the arista (HE AND ADLER 2002). Using frtz mneongreen fly, we detected the asymmetric accumulation of Frtz in arista central core cells (Figure 6D) consistent with the asymmetric accumulation of Frtz being important for arista morphogenesis and for the pattern of Frtz protein being instructed by the upstream PCP proteins in this tissue Stability of localized Frtz protein. In order to determine if there is a difference in the dynamic properties of the Frtz protein in different tissues, FRAP analysis was done on wing, thorax and abdominal cells. In these experiments we routinely bleached a part of one or more cells over the region where 431 Frtz mneongreen preferentially accumulated. We then followed over time the reappearance of fluorescence in the region of preferential accumulation. After bleaching the cells, the fluorescence substantially recovered in all three tissues within 60s (Figure 6E-J) and the estimated recovery half time is 70s. However, over the course of these experiments (5 mins) we never detected a complete recovery of fluorescence. One explanation for this is that the cells 21
22 contain of both stable and less stable pools of localized Frtz protein. The stable protein pool would either not recover or recover slowly, while the unstable protein pool would recover rapidly. An alternative possibility is that the recovery came from Frtz mneongreen protein dispersed in non-bleached regions of the cell and that there was not enough Frtz mneongreen in this pool to provide for complete recovery. 441 Accumulation of Frtz mneongreen in hairs and sensory bristles Our live imaging of Frtz mneongreen demonstrated the asymmetric accumulation of Frtz in various tissues. To our surprise, we also observed the accumulation of Frtz in Drosophila sensory bristles and hairs (Figure S6C). We also observed this in fixed preparations. Frtz mneongreen started to accumulate in the proximal part of sensory bristles around 43h apf (Figure 8A, arrows), and by about 45 h apf it reached its maximum intensity and accumulated throughout the entire bristles in a pattern of stripes parallel to the long axis of the bristle shaft (Figure 8D, arrows) To further determine the localization of Frtz in bristles, we co-localized Frtz and F-actin in sensory bristles at 45h apf (Figure 8D-F, arrows). Developing bristle shaft cells are characterized by a set of large bundles of F-actin (TILNEY et al. 1995). The stripes of F-actin did not co-localize with the Frtz stripes, rather the Frtz stripes were localized to the region between the actin stripes. The Frtz stripes did not fill the space between the actin bundles and did not appear to contact the bundles. To determine if the bundles of F-actin found in the bristles were important for restricting the accumulation of Frtz we examined Frtz accumulation in a sn 3 f 36a double mutant where the large actin bundles are missing (Figure 8G-I) (TILNEY et al. 1995). In the mutant bristles we still observed Frtz accumulation in stripes but they were no longer well 22
23 organized and appeared increased in numbers indicating that in some way the actin bundles mediate the localization of Frtz In the experiments where we imaged Frtz mneongreen in bristles we noticed strong staining in what appeared to be socket cells. In a 3-D reconstruction and from examining orthogonal views we observed that the high level of Frtz mneongreen accumulation was likely in the socket cell where it juxtaposed to the shaft cell (Figure S8, File S2). The significance of this localization remains to be established. 465 Discussion The interaction of Dsh with Fy and Frtz The formation of the proximal and distal complexes of PCP proteins involves positive intercellular feedback and negative intracellular feedback. The proximal and distally localized cytoplasmic proteins are thought to be recruited both by a positive interaction with a similarly localized membrane protein (e.g. between Fz and Dsh) (CHEN et al. 2008; STRUTT AND STRUTT 2008; WU AND MLODZIK 2008) and negative interactions between proximal and distally cytoplasmic proteins (e.g. Pk and Dsh) (TREE et al. 2002; AMONLIRDVIMAN et al. 2005). Based on this example we expected that the proximal localization of PPE proteins would involve a positive recruitment by a proximally localized PCP protein and perhaps a negative interaction with a distally localized PCP protein. We detected interactions between both Fy and Dsh and Frtz and Dsh. An interaction between the mammalian homologs Fuz and Dvl was described previously (ZILBER et al. 2013) and our experiments established that this interaction is conserved. We did not see such an interaction between the mammalian homologs of Frtz (WDPCP) and Dsh (Dvl2) so this interaction may not be conserved but such a negative result must be interpreted 23
24 with caution. The interaction between the distally localized Dsh and the proximally localized Frtz and Fy is likely to be a negative one, perhaps analogous to the one seen between Dsh and Pk (TREE et al. 2002). That could result in incorrectly localized Frtz or Fy being degraded (STRUTT et al. 2013; CHO et al. 2015). Previous experiments established that the function of the upstream core PCP genes was essential for the proximal localization of In and Frtz to the proximal side of wing cells (ADLER et al. 2004; STRUTT AND WARRINGTON 2008). Based on those observations it seemed likely that one or more of the PPE proteins would bind to one or more of the proximally localized PCP proteins. We did not obtain convincing and reproducible evidence for such an interaction. That does not mean that such an interaction does not take place. For example, it is possible that the recruitment of one of the PPE proteins such as Frtz or In to the proximal side of wing cells could require an interaction with both Pk and Vang, an interaction with a modified protein or an interaction with an as yet unidentified proximal protein. It could also be weak enough for us not to be able to detect it. Our experiments with the Pk-Cherry-Frtz fusion protein establish that Pk and Frtz can function in close proximity Not all PPE gene function requires the presence of all 3 members of the group. Most, but not all previous data on the PPE genes in Drosophila suggested that these genes and proteins functioned as a unit in PCP and each were required for the function of the others. In this paper we found that the over expression of frtz could partially suppress and the over expression of fy could weakly enhance the phenotype of a fly that completely lacked the in gene. Thus, in these experiments both Frtz and Fy were able to influence PCP in an In independent manner. Since the ability to influence the planar cell polarity phenotype required over 24
25 expression it is not clear that in a wild type fly frtz and/or fy function in an in independent manner but our results indicate the potential is there. This is consistent with previous results where we found the PCP gain of function phenotype that resulted from the over expression of frtz or frtz-gfp was not eliminated by a lack of in function, but those experiments utilized an in nonsense mutation and a gain of function frtz phenotype (WANG et al. 2014). That both Frtz and Fy could impact PCP in the absence of any In suggests that it s principal function might be to mediate the accumulation and localization of these proteins. When over expressed enough Frtz and Fy might be present to allow a partial bypass of that function Frtz and the actin cytoskeleton In studying the expression and localization of Frtz using the frtz mneongreen gene we discovered that Frtz accumulated in stripes along the proximal/distal axis of sensory bristles in the space between the large actin bundles found in bristles. In the absence of the F actin bundles this pattern became disorganized and there appeared to be an increased number of Frtz stripes. This indicates that the patterning of the Frtz stripes is regulated by the actin cytoskeleton, although since the two did not overlap the regulation seems likely to be indirect. We previously found that the over expression of frtz resulted in a delay in the activation the actin cytoskeleton that drives wing hair morphogenesis (WANG et al. 2014). Further, the morphology of the actin cytoskeleton and the resulting hair were quite abnormal. Taken together it suggests that the actin cytoskeleton and Frtz mutually interact and regulate each other s localization and activity. If and how this relates to the function of Frtz in PCP will require further research. It is interesting to note that in vertebrates the WDPCP mutant phenotype differs from that of intu and fuz (KIM et al. 25
26 ; CUI et al. 2013) and that WDPCP mutants appear to have an actin cytoskeleton phenotype (CUI et al. 2013) The PPE genes and PCP in the abdomen. We found that mutations in Drosophila PPE genes produced a strong multiple hair phenotype and a hair polarity phenotype in all regions of the dorsal abdomen. This differs from that seen with mutations in upstream fz/stan pathway genes where a strong hair polarity phenotype is only seen in part of each segment (CASAL et al. 2006), even though PCP proteins accumulate asymmetrically in cells throughout segment. The ds/ft pathway also plays a key role in abdomen PCP and double mutants where both pathways are inactivated show an additive phenotype with a strong phenotype in all regions (CASAL et al. 2006). This raised the possibility that the PPE genes functioned downstream of both pathways in the abdomen and that the region specific phenotypes of fz/stan and ds/ft could be due to two upstream pathways being dominant with regard to instructing the accumulation of PPE proteins in different regions of the segment. Consistent with the PPE genes functioning downstream of fz/stan in the abdomen double mutants of fz with either frtz, fy or in resembled the PPE hair phenotype. We found that both In and Frtz accumulated asymmetrically in abdominal cells throughout each segment consistent with that being important for their function. However, this asymmetric accumulation was lost throughout 543 each segment in a dsh 1 mutant background. Thus, having a functional ds/ft pathway is insufficient to promote asymmetric accumulation of PPE proteins in any part of the segment. It is worth noting that in the abdomen the PPE proteins retain substantial function even when not properly localized as the strong PPE multiple hair phenotype is not seen in a dsh 1 mutant. This mirrors what is seen in the wing (WONG AND ADLER 1993; ADLER et al. 2004). 26
27 Two pools of Frtz protein within the cells We analyzed the dynamics of Frtz in several body regions using FRAP on epithelial cells that expressed the edited frtz mneongreen gene. Similar results were obtained in all body regions. We found the recovery of most of the fluorescence was relatively fast (~70 seconds), however the recovery did not go to completion over the time period (~5-10 minutes) followed. The recovery was limited to approximately 80% of the starting fluorescence. This could be due to two pools of Frtz protein within cells that vary in stability. Alternatively, it could be due to a lack of sufficient unbleached protein in the cell to allow complete recovery of fluorescence. We observed a greater fraction of recovery than was seen by Strutt and colleagues in their FRAP analysis on the PCP core proteins Fz and Stan (Strutt et al., 2011). Those two proteins are transmembrane proteins and that might be part of the reason for a larger fraction of stable and non-recovering protein Tagged Endogenous Genes vs transgenes The ability to edit the endogenous frtz gene provided us with a tool that was clearly superior to our previous generation Ubi-myc-frtz-GFP transgenes as we could be confident that the former was providing normal levels and developmental expression pattern. It also avoided the sometimes confounding effects of the artificial promoter/enhancer that resulted in myc-frtz- GFP being abnormally expressed in tissues that were closely juxtaposed to the epidermis which hampered imaging. However, the Ubi-myc-frtz-GFP transgenes remain quite useful as they provide function and a tagged protein that is encoded at other locations in the genome (e.g. 3 rd 27
28 chromosome). The flexibility that this provides is advantageous for some experiments that involve complicated genetics The ability to precisely delete the in gene using CRISPR/Cas9 allowed us to determine that small amounts of In protein activity were confounding some of our gene interaction experiments. In our case our nonsense allele of in proved not to be a null allele. It has long been true in some systems (e.g. bacteria and yeast) that single gene deletions were easy to generate and that is now the case generally and the advantages of such alleles are obvious Acknowledgements We thank FlyBase and the stock centers at Indiana University and the VDRC and ADDGENE where we obtained many stocks and plasmids used in the paper. We also thank G. Guild, J. 581 Axelrod, D. Strutt and M. Mlodzik for reagents. We also acknowledge the Keck Imaging Center where the confocal microscopy was done. This research was supported by a grant from the National Institutes of Health (GM037163) Literature Cited Adler, P. N., 2012 The frizzled/stan pathway and planar cell polarity in the Drosophila wing. Curr Top Dev Biol 101: Adler, P. N., J. Charlton and W. J. Park, 1994 The Drosophila tissue polarity gene inturned functions prior to wing hair morphogenesis in the regulation of hair polarity and number. Genetics 137: Adler, P. N., C. Zhu and D. Stone, 2004 Inturned localizes to the proximal side of wing cells under the instruction of upstream planar polarity proteins. Curr Biol 14: Amonlirdviman, K., N. A. Khare, D. R. Tree, W. S. Chen, J. D. Axelrod et al., 2005 Mathematical modeling of planar cell polarity to understand domineering nonautonomy. Science 307:
29 Axelrod, J. D., 2001 Unipolar membrane association of Dishevelled mediates Frizzled planar cell polarity signaling. Genes Dev 15: Babayeva, S., B. Rocque, L. Aoudjit, Y. Zilber, J. Li et al., 2013 Planar cell polarity pathway regulates nephrin endocytosis in developing podocytes. J Biol Chem 288: Bastock, R., H. Strutt and D. Strutt, 2003 Strabismus is asymmetrically localised and binds to Prickle and Dishevelled during Drosophila planar polarity patterning. Development 130: Brand, A. H., and N. Perrimon, 1993 Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118: Casal, J., P. A. Lawrence and G. Struhl, 2006 Two separate molecular systems, Dachsous/Fat and Starry night/frizzled, act independently to confer planar cell polarity. Development 133: Chen, W. S., D. Antic, M. Matis, C. Y. Logan, M. Povelones et al., 2008 Asymmetric homotypic interactions of the atypical cadherin flamingo mediate intercellular polarity signaling. Cell 133: Cho, B., G. Pierre-Louis, A. Sagner, S. Eaton and J. D. Axelrod, 2015 Clustering and negative feedback by endocytosis in planar cell polarity signaling is modulated by ubiquitinylation of prickle. PLoS Genet 11: e Collier, S., H. Lee, R. Burgess and P. Adler, 2005 The WD40 repeat protein fritz links cytoskeletal planar polarity to frizzled subcellular localization in the Drosophila epidermis. Genetics 169: Cui, C., B. Chatterjee, T. P. Lozito, Z. Zhang, R. J. Francis et al., 2013 Wdpcp, a PCP protein required for ciliogenesis, regulates directional cell migration and cell polarity by direct modulation of the actin cytoskeleton. PLoS Biol 11: e Dabdoub, A., and M. W. Kelley, 2005 Planar cell polarity and a potential role for a Wnt morphogen gradient in stereociliary bundle orientation in the mammalian inner ear. J Neurobiol 64: Das, G., A. Jenny, T. J. Klein, S. Eaton and M. Mlodzik, 2004 Diego interacts with Prickle and Strabismus/Van Gogh to localize planar cell polarity complexes. Development 131: Donoughe, S., and S. DiNardo, 2011 dachsous and frizzled contribute separately to planar polarity in the Drosophila ventral epidermis. Development 138: Feiguin, F., M. Hannus, M. Mlodzik and S. Eaton, 2001 The ankyrin repeat protein Diego mediates Frizzled-dependent planar polarization. Dev Cell 1: Goggolidou, P., 2014 Wnt and planar cell polarity signaling in cystic renal disease. Organogenesis 10: Goodrich, L. V., and D. Strutt, 2011 Principles of planar polarity in animal development. Development 138: Gratz, S. J., A. M. Cummings, J. N. Nguyen, D. C. Hamm, L. K. Donohue et al., 2013a Genome engineering of Drosophila with the CRISPR RNA-guided Cas9 nuclease. Genetics 194: Gratz, S. J., J. Wildonger, M. M. Harrison and K. M. O'Connor-Giles, 2013b CRISPR/Cas9-mediated genome engineering and the promise of designer flies on demand. Fly (Austin) 7: Gubb, D., and A. Garcia-Bellido, 1982 A genetic analysis of the determination of cuticular polarity during development in Drosophila melanogaster. J Embryol Exp Morphol 68: He, B., and P. N. Adler, 2002 The frizzled pathway regulates the development of arista laterals. BMC Dev Biol 2: 7. Jenny, A., R. S. Darken, P. A. Wilson and M. Mlodzik, 2003 Prickle and Strabismus form a functional complex to generate a correct axis during planar cell polarity signaling. EMBO J 22: Jenny, A., and M. Mlodzik, 2006 Planar cell polarity signaling: a common mechanism for cellular polarization. Mt Sinai J Med 73: Keller, R., L. Davidson, A. Edlund, T. Elul, M. Ezin et al., 2000 Mechanisms of convergence and extension by cell intercalation. Philos Trans R Soc Lond B Biol Sci 355:
30 Kim, S. K., A. Shindo, T. J. Park, E. C. Oh, S. Ghosh et al., 2010 Planar cell polarity acts through septins to control collective cell movement and ciliogenesis. Science 329: Perkins, L. A., L. Holderbaum, R. Tao, Y. Hu, R. Sopko et al., 2015 The Transgenic RNAi Project at Harvard Medical School: Resources and Validation. Genetics 201: Price, M. H., D. M. Roberts, B. M. McCartney, E. Jezuit and M. Peifer, 2006 Cytoskeletal dynamics and cell signaling during planar polarity establishment in the Drosophila embryonic denticle. J Cell Sci 119: Ran, F. A., P. D. Hsu, J. Wright, V. Agarwala, D. A. Scott et al., 2013 Genome engineering using the CRISPR-Cas9 system. Nat Protoc 8: Schultz, J., F. Milpetz, P. Bork and C. P. Ponting, 1998 SMART, a simple modular architecture research tool: identification of signaling domains. Proc Natl Acad Sci U S A 95: Sekelsky, J. J., S. J. Newfeld, L. A. Raftery, E. H. Chartoff and W. M. Gelbart, 1995 Genetic characterization and cloning of mothers against dpp, a gene required for decapentaplegic function in Drosophila melanogaster. Genetics 139: Sharp, K. A., and J. D. Axelrod, 2016 Prickle isoforms control the direction of tissue polarity by microtubule independent and dependent mechanisms. Biol Open 5: Steneberg, P., and C. Samakovlis, 2001 A novel stop codon readthrough mechanism produces functional Headcase protein in Drosophila trachea. EMBO Rep 2: Strutt, D., and S. J. Warrington, 2008 Planar polarity genes in the Drosophila wing regulate the localisation of the FH3-domain protein Multiple Wing Hairs to control the site of hair production. Development 135: Strutt, D. I., 2001 Asymmetric localization of frizzled and the establishment of cell polarity in the Drosophila wing. Mol Cell 7: Strutt, H., and D. Strutt, 2008 Differential stability of flamingo protein complexes underlies the establishment of planar polarity. Curr Biol 18: Strutt, H., V. Thomas-MacArthur and D. Strutt, 2013 Strabismus promotes recruitment and degradation of farnesylated prickle in Drosophila melanogaster planar polarity specification. PLoS Genet 9: e Strutt, H., S. J. Warrington and D. Strutt, 2011 Dynamics of core planar polarity protein turnover and stable assembly into discrete membrane subdomains. Dev Cell 20: Tilney, L. G., M. S. Tilney and G. M. Guild, 1995 F actin bundles in Drosophila bristles. I. Two filament cross-links are involved in bundling. J Cell Biol 130: Tree, D. R., J. M. Shulman, R. Rousset, M. P. Scott, D. Gubb et al., 2002 Prickle mediates feedback amplification to generate asymmetric planar cell polarity signaling. Cell 109: Usui, T., Y. Shima, Y. Shimada, S. Hirano, R. W. Burgess et al., 1999 Flamingo, a seven-pass transmembrane cadherin, regulates planar cell polarity under the control of Frizzled. Cell 98: Wallingford, J. B., S. E. Fraser and R. M. Harland, 2002 Convergent extension: the molecular control of polarized cell movement during embryonic development. Dev Cell 2: Wallingford, J. B., B. A. Rowning, K. M. Vogeli, U. Rothbacher, S. E. Fraser et al., 2000 Dishevelled controls cell polarity during Xenopus gastrulation. Nature 405: Wang, J., S. Mark, X. Zhang, D. Qian, S. J. Yoo et al., 2005 Regulation of polarized extension and planar cell polarity in the cochlea by the vertebrate PCP pathway. Nat Genet 37: Wang, W., K. Czaplinski, Y. Rao and S. W. Peltz, 2001 The role of Upf proteins in modulating the translation read-through of nonsense-containing transcripts. EMBO J 20: Wang, Y., X. J. Hu, X. D. Zou, X. H. Wu, Z. Q. Ye et al., 2015 WDSPdb: a database for WD40-repeat proteins. Nucleic Acids Res 43: D
31 Wang, Y., and J. Nathans, 2007 Tissue/planar cell polarity in vertebrates: new insights and new questions. Development 134: Wang, Y., J. Yan, H. Lee, Q. Lu and P. N. Adler, 2014 The proteins encoded by the Drosophila Planar Polarity Effector genes inturned, fuzzy and fritz interact physically and can re-pattern the accumulation of "upstream" Planar Cell Polarity proteins. Dev Biol 394: Wolff, T., and G. M. Rubin, 1998 Strabismus, a novel gene that regulates tissue polarity and cell fate decisions in Drosophila. Development 125: Wong, L. L., and P. N. Adler, 1993 Tissue polarity genes of Drosophila regulate the subcellular location for prehair initiation in pupal wing cells. J Cell Biol 123: Wu, J., and M. Mlodzik, 2008 The frizzled extracellular domain is a ligand for Van Gogh/Stbm during nonautonomous planar cell polarity signaling. Dev Cell 15: Wu, X. H., Y. Wang, Z. Zhuo, F. Jiang and Y. D. Wu, 2012 Identifying the hotspots on the top faces of WD40-repeat proteins from their primary sequences by beta-bulges and DHSW tetrads. PLoS One 7: e Zheng, L., J. Zhang and R. W. Carthew, 1995 frizzled regulates mirror-symmetric pattern formation in the Drosophila eye. Development 121: Zilber, Y., S. Babayeva, J. H. Seo, J. J. Liu, S. Mootin et al., 2013 The PCP effector Fuzzy controls cilial assembly and signaling by recruiting Rab8 and Dishevelled to the primary cilium. Mol Biol Cell 24: Figure Legends Figure 1. Protein interactions between PPE and PCP core proteins. (A) and (C) are yeast-twohybrid plates for interaction between Fy and Dsh (A), Frtz and Dsh (C). The plates are 4 amino acid (Ade/-His/-Leu/-Trp) drop out plates containing x-±-gal. Only the colonies with interacting protein pairs grow and turn blue. (B) and (D) are co-immunoprecipitation results for Fy and Dsh (B) Frtz and Dsh (D), respectively. (B) The co-immunoprecipitation of Dsh and Fy. This experiment used UAS-Fy-GFP driven by ptc-gal4 and Dsh-myc transgenes (Bloomington Drosophila Stock Center stock number 25385). Wing disc samples were immunoprecipitated using rabbit GFP antibody then detected with mouse GFP and Myc antibody for western blotting. Arrows point to Fy-GFP and Dsh-Myc on the Western blots. (D) shows the coimmunoprecipitation of Dsh and Frtz. This experiment used UAS-frtz driven by ptc-gal4 and 31
32 Dsh-myc transgenes. Wing disc samples were immunoprecipitated using rabbit Myc antibody then detected with rat Frtz and mouse Myc antibody for western bloting. Arrows point to Dsh- Myc and Frtz on the Western blots. (E) and (F) are yeast-two-hybrid plates for interactions between Dvl2 and Intu (E) and WDPCP and Intu (F) Figure 2. (A) The genomic region surrounding the in gene, the locations targeted by the grnas, and the homology fragment. (B). The phenotype of in PD and in IH56 in the anterior region of the wing. Twenty wings were scored for each genotype to quantify the multiple hair cell phenotype. (C). The phenotype of in PD and in IH56 in a central region of the wing. Twenty wings were scored for each genotype to quantify the multiple hair cell phenotype. (D). An image of the anterior region of the wing in in IH56. (E). An image of the central region of the wing in in IH56. (F). An image of the anterior region of the wing in in PD. (G). An image of the central region of the wing in in PD. Note the stronger phenotypes in in PD Figure 3. The mild over expression of myc-frtz and fy-flag-ollas partially suppresses and enhances the phenotype of in PD, respectively. (A, A ) The regions of the wing scored are shown. (B, B ) Images of ap>myc-frtz; in PD in the two wing regions. (C, C ) Images of UAS-myc-frtz/+; in PD in the two wing regions. (D, D ) Images of in PD in the two wing regions. (E, E ) Images of ap>fy-flag-ollas; in PD in the two wing regions. (F, F ) Images of fy-flag-ollas/+; in PD in the two wing regions. (G) Quantitation of the multiple hair cell phenotype of the genotypes shown in panels B-D. The numbers on the Y axis indicate the fraction of multiple hair cells. Significant differences are indicated with an asterisk. (H) Quantitation of the multiple hair cell phenotype of the genotypes shown in panels E-F. The numbers on the Y axis indicate the fraction of 32
33 multiple hair cells. Significant differences are indicated with an asterisk. pwalium is a phic31 UAS germ line transformation vector ((PERKINS et al. 2015) Figure 4. The ubi-myc-frtz-gfp transgene works well to show the asymmetric accumulation of the Frtz protein by in vivo imaging of pupae. (A) An image of a pupal wing. (B) A time series showing the fusion of a Frtz-GFP puncta. (C) A time series showing a puncta appearing to leave the membrane and move to the cytoplasm. (D) A time series showing a puncta that splits into 2. The wings in these experiments were from hr pupal wings Figure 5. The CRISPR/Cas9 mediated editing of the endogenous frtz gene. (A) The genomic region around the frtz gene is shown along with the location of the grnas and the homology dependent repair template. (B) Adult wings that show the frtz mneongreen edited gene is functional and provides complete rescue of a strong frtz allele Figure 6. The edited frtz mneongreen gene is a very useful tool for in vivo imaging of the Frtz protein. (A) A 30 hr apf (after white prepupae formation) wing. (B) A 30 hr apf thorax. (C) A 43 hr apf abdomen. (D) A 28 hr apf antenna. (E) Quantitation of FRAP experiments on wing, thorax and abdomen. (F-J) Time points from a FRAP experiment on the abdomen Figure 7. The phenotype of PPE mutants in the adult dorsal abdomen (tergite). All images are from a region just lateral to the midline of the 3rd abdominal segment. A higher magnification image from a region of each lower magnification is shown on the right and labelled with a. (A) 33
34 Oregon-R. (B) fz P21 /fz K21 (C) ptc>ds-rnai (D) ds UAO71 /ds YAC41 (E) in IH56 (F) fy 2 (G) frtz 30 (H) mwh 1 (I) fz 1 in 1 (J) fy 2 ; fz 1 (K) frtz 30 ; fz 1 (L) fy 2 ; in IH56 (M) ds UAO71 /ds YAC41 ; in IH56 (N) fy 2 ; ptc>ds- i ; (O) frtz 30 ; ptc>ds-i (P) frtz 30 ; in IH Figure 8. Frtz-NG accumulates in bristles and hairs. (ABC) Immunostained frtz mneongreen pupal head (43 hr after puparium formation). Frtz mneongreen immunostaining in green and F-actin in red. The arrows point to the accumulation of Frtz mneongreen in the proximal part of bristles. (DEF). Immunostained frtz-ng pupal head (45 hr after puparium formation). Frtz mneongreen immunostaining in green and F-actin in red. The arrows point to the accumulation of Frtz mneongreen in stripes down the bristles. From the merged image it is clear that the Frtz mneongreen is in between large bundles of F-actin. (GHI) ). Immunostained sn 3 f 36a ; frtz mneongreen pupal head (45 hr after puparium formation). Frtz mneongreen immunostaining in green and F-actin in red. The arrows point to the accumulation of Frtz mneongreen in disorganized stripes in the bristles. The actin staining image I shows the lack of large bundles of F-actin as expected for the sn 3 f 36a mutant
35

36

37

38

39

40

41

42
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
THE epidermis of many animals is polarized within
 Copyright Ó 2008 by the Genetics Society of America DOI: 10.1534/genetics.108.091314 The multiple-wing-hairs Gene Encodes a Novel GBD FH3 Domain-Containing Protein That Functions Both Prior to and After
Copyright Ó 2008 by the Genetics Society of America DOI: 10.1534/genetics.108.091314 The multiple-wing-hairs Gene Encodes a Novel GBD FH3 Domain-Containing Protein That Functions Both Prior to and After
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway
 Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
THE formation of properly differentiated organs
 Copyright Ó 2010 by the Genetics Society of America DOI: 10.1534/genetics.109.112060 Drosophila Rab23 Is Involved in the Regulation of the Number and Planar Polarization of the Adult Cuticular Hairs Csilla
Copyright Ó 2010 by the Genetics Society of America DOI: 10.1534/genetics.109.112060 Drosophila Rab23 Is Involved in the Regulation of the Number and Planar Polarization of the Adult Cuticular Hairs Csilla
Drosophila Rab23 is involved in the regulation of the number and planar. polarization of the adult cuticular hairs
 Genetics: Published Articles Ahead of Print, published on February 1, 2010 as 10.1534/genetics.109.112060 Drosophila Rab23 is involved in the regulation of the number and planar polarization of the adult
Genetics: Published Articles Ahead of Print, published on February 1, 2010 as 10.1534/genetics.109.112060 Drosophila Rab23 is involved in the regulation of the number and planar polarization of the adult
NucView TM 488 Caspase-3 Assay Kit for Live Cells
 NucView TM 488 Caspase-3 Assay Kit for Live Cells Catalog Number: 30029 (100-500 assays) Contact Information Address: Biotium, Inc. 3423 Investment Blvd. Suite 8 Hayward, CA 94545 USA Telephone: (510)
NucView TM 488 Caspase-3 Assay Kit for Live Cells Catalog Number: 30029 (100-500 assays) Contact Information Address: Biotium, Inc. 3423 Investment Blvd. Suite 8 Hayward, CA 94545 USA Telephone: (510)
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290
 PLoS Biol 4(9): e290") Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
OESTREICH LAB CHIP PROTOCOL
 OESTREICH LAB CHIP PROTOCOL Buffers (all starred (*) buffers must be autoclaved before use) 10X Cell Lysis Buffer*: 100mL 0.214g Potassium Acetate (KOAc) = 10mM 0.147g Magnesium Acetate (MgAc) = 15mM 10mL
OESTREICH LAB CHIP PROTOCOL Buffers (all starred (*) buffers must be autoclaved before use) 10X Cell Lysis Buffer*: 100mL 0.214g Potassium Acetate (KOAc) = 10mM 0.147g Magnesium Acetate (MgAc) = 15mM 10mL
U.S. Patent No. 9,051,563 and other pending patents. Ver
 INSTRUCTION MANUAL Direct-zol 96 RNA Catalog Nos. R2054, R2055, R2056 & R2057 Highlights Quick, 96-well purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and all other
INSTRUCTION MANUAL Direct-zol 96 RNA Catalog Nos. R2054, R2055, R2056 & R2057 Highlights Quick, 96-well purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and all other
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila
 Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
2. Yeast two-hybrid system
 2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
NOVABEADS FOOD 1 DNA KIT
 NOVABEADS FOOD 1 DNA KIT NOVABEADS FOOD DNA KIT is the new generation tool in molecular biology techniques and allows DNA isolations from highly processed food products. The method is based on the use
NOVABEADS FOOD 1 DNA KIT NOVABEADS FOOD DNA KIT is the new generation tool in molecular biology techniques and allows DNA isolations from highly processed food products. The method is based on the use
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
A reaction-diffusion, partial differential equation model of planar cell polarity signaling
 1 Mathematical modeling of planar cell polarity to understand domineering non-autonomy Keith Amonlirdviman 1, Narmada A. Khare 2, David R.P. Tree 2, Wei-Shen Chen 2, Jeffrey D. Axelrod 2* and Claire J.
1 Mathematical modeling of planar cell polarity to understand domineering non-autonomy Keith Amonlirdviman 1, Narmada A. Khare 2, David R.P. Tree 2, Wei-Shen Chen 2, Jeffrey D. Axelrod 2* and Claire J.
downstream (0.8 kb) homologous sequences to the genomic locus of DIC. A DIC mutant strain (ro- 6
 homologous sequences to the genomic locus of DIC. A DIC mutant strain (ro- 6") A B C D ts Figure S1 Generation of DIC- mcherry expressing N.crassa strain. A. N. crassa colony morphology. When a cot1 (top, left panel) strain is grown at permissive temperature (25 C), it exhibits straight
A B C D ts Figure S1 Generation of DIC- mcherry expressing N.crassa strain. A. N. crassa colony morphology. When a cot1 (top, left panel) strain is grown at permissive temperature (25 C), it exhibits straight
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
mrna Isolation Kit for Blood/Bone Marrow For isolation mrna from blood or bone marrow lysates Cat. No
 For isolation mrna from blood or bone marrow lysates Cat. No. 1 934 333 Principle Starting material Application Time required Results Key advantages The purification of mrna requires two steps: 1. Cells
For isolation mrna from blood or bone marrow lysates Cat. No. 1 934 333 Principle Starting material Application Time required Results Key advantages The purification of mrna requires two steps: 1. Cells
Supplemental Information. The Mitochondrial Fission Receptor MiD51. Requires ADP as a Cofactor
 Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype by altering frizzled signaling
 Development 125, 959-968 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV7601 959 Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype
Development 125, 959-968 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV7601 959 Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype
A complementation test would be done by crossing the haploid strains and scoring the phenotype in the diploids.
 Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
Problem set H answers 1. To study DNA repair mechanisms, geneticists isolated yeast mutants that were sensitive to various types of radiation; for example, mutants that were more sensitive to UV light.
SUPPLEMENTARY INFORMATION
 med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
Component Product # Product # Cell Lysis Reagent 100 ml 500 ml Product Insert 1 1
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Cell Lysis Reagent Product # 18800 (100 ml) Product # 18801 (500
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Cell Lysis Reagent Product # 18800 (100 ml) Product # 18801 (500
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Dachsous-Dependent Asymmetric Localization of Spiny-Legs Determines Planar Cell Polarity Orientation in Drosophila
 Cell Reports Article Dachsous-Dependent Asymmetric Localization of Spiny-Legs Determines Planar Cell Polarity Orientation in Drosophila Tomonori Ayukawa, 1,2,3 Masakazu Akiyama, 4 Jennifer L. Mummery-Widmer,
Cell Reports Article Dachsous-Dependent Asymmetric Localization of Spiny-Legs Determines Planar Cell Polarity Orientation in Drosophila Tomonori Ayukawa, 1,2,3 Masakazu Akiyama, 4 Jennifer L. Mummery-Widmer,
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1 Sns and Duf co-localise in embryonic nephrocytes a-c, Wild-type stage 11 (a),14 (b) and 16 (c) embryos stained with anti-duf (green) and anti-sns (red). Higher magnification images
Supplementary Figure 1 Sns and Duf co-localise in embryonic nephrocytes a-c, Wild-type stage 11 (a),14 (b) and 16 (c) embryos stained with anti-duf (green) and anti-sns (red). Higher magnification images
U.S. Patent No. 9,051,563 and other pending patents. Ver
 INSTRUCTION MANUAL Direct-zol RNA MiniPrep Catalog Nos. R050, R05, R05, & R053 Highlights Quick, spin column purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and other
INSTRUCTION MANUAL Direct-zol RNA MiniPrep Catalog Nos. R050, R05, R05, & R053 Highlights Quick, spin column purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and other
Mag-Bind Soil DNA Kit. M preps M preps M preps
 Mag-Bind Soil DNA Kit M5635-00 5 preps M5635-01 50 preps M5635-02 200 preps January 2013 Mag-Bind Soil DNA Kit Table of Contents Introduction and Overview...2 Kit Contents/Storage and Stability...3 Preparing
Mag-Bind Soil DNA Kit M5635-00 5 preps M5635-01 50 preps M5635-02 200 preps January 2013 Mag-Bind Soil DNA Kit Table of Contents Introduction and Overview...2 Kit Contents/Storage and Stability...3 Preparing
Prickle Mediates Feedback Amplification to Generate Asymmetric Planar Cell Polarity Signaling
 Cell, Vol. 109, 371 381, May 3, 2002, Copyright 2002 by Cell Press Prickle Mediates Feedback Amplification to Generate Asymmetric Planar Cell Polarity Signaling David R.P. Tree, 1,2 Joshua M. Shulman,
Cell, Vol. 109, 371 381, May 3, 2002, Copyright 2002 by Cell Press Prickle Mediates Feedback Amplification to Generate Asymmetric Planar Cell Polarity Signaling David R.P. Tree, 1,2 Joshua M. Shulman,
7.06 Cell Biology EXAM #3 April 21, 2005
 7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
Genomes and Their Evolution
 Chapter 21 Genomes and Their Evolution PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
Chapter 21 Genomes and Their Evolution PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from
EST1 Homology Domain. 100 aa. hest1a / SMG6 PIN TPR TPR. Est1-like DBD? hest1b / SMG5. TPR-like TPR. a helical. hest1c / SMG7.
 hest1a / SMG6 EST1 Homology Domain 100 aa 853 695 761 780 1206 hest1 / SMG5 -like? -like 109 145 214 237 497 165 239 1016 114 207 212 381 583 hest1c / SMG7 a helical 1091 Sc 57 185 267 284 699 Figure S1:
hest1a / SMG6 EST1 Homology Domain 100 aa 853 695 761 780 1206 hest1 / SMG5 -like? -like 109 145 214 237 497 165 239 1016 114 207 212 381 583 hest1c / SMG7 a helical 1091 Sc 57 185 267 284 699 Figure S1:
Protocol for 2D-E. Protein Extraction
 Protocol for 2D-E Protein Extraction Reagent 1 inside the ReadyPrep TM Sequential Extraction kit (in powder form) 50ml of deionized water is used to dissolve all the Reagent 1. The solution is known as
Protocol for 2D-E Protein Extraction Reagent 1 inside the ReadyPrep TM Sequential Extraction kit (in powder form) 50ml of deionized water is used to dissolve all the Reagent 1. The solution is known as
Supplementary Materials for
 www.sciencemag.org/cgi/content/full/science.1244624/dc1 Supplementary Materials for Cytoneme-Mediated Contact-Dependent Transport of the Drosophila Decapentaplegic Signaling Protein Sougata Roy, Hai Huang,
www.sciencemag.org/cgi/content/full/science.1244624/dc1 Supplementary Materials for Cytoneme-Mediated Contact-Dependent Transport of the Drosophila Decapentaplegic Signaling Protein Sougata Roy, Hai Huang,
Reading: Chapter 5, pp ; Reference chapter D, pp Problem set F
 Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Protease Inhibitor Cocktail A (1 tablet / 7 10 ml, Roche Cat# ) Protease inhibitor Cocktail B (0.5ml per 250ml, Calbiochem Cat# )
 Protease inhibitor Cocktail B (0.5ml per 250ml, Calbiochem Cat# )") Protocol for Western Blotting Tissue/Cell Sample Preparation Lysis Buffer 1 (ph8.0) o 50mM Tris-Cl o 150mM NaCl o 1% v/v NP40 o protease inhibitor cocktail A/B Lysis Buffer 2 (RIPA) (ph 8.0) o 50mM Tris-Cl
Protocol for Western Blotting Tissue/Cell Sample Preparation Lysis Buffer 1 (ph8.0) o 50mM Tris-Cl o 150mM NaCl o 1% v/v NP40 o protease inhibitor cocktail A/B Lysis Buffer 2 (RIPA) (ph 8.0) o 50mM Tris-Cl
7.06 Problem Set
 7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
Bypass and interaction suppressors; pathway analysis
 Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Nature Biotechnology: doi: /nbt Supplementary Figure 1. Overexpression of YFP::GPR-1 in the germline.
 Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
camp Direct Immunoassay Kit
 camp Direct Immunoassay Kit Catalog Number KA0886 100 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
camp Direct Immunoassay Kit Catalog Number KA0886 100 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Supplementary Information
 Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
DNA can be extracted from the following sample types using this procedure: Archived
 Sample types Principle Safety Equipment and supplies DNA can be extracted from the following sample types using this procedure: concentrated DNA samples (e.g., blood, saliva, non-contact samples) hair
Sample types Principle Safety Equipment and supplies DNA can be extracted from the following sample types using this procedure: concentrated DNA samples (e.g., blood, saliva, non-contact samples) hair
Phenol-Chloroform reagents. Selection guide. OH ; MW : High quality reagents for use in nucleic acid purification.
 Phenol-Chloroform reagents Extraction with phenol and phenol/chloroform mixtures is a universal method for purification of DNA and RNA. Proteins and restriction enzymes are removed by phenol and chloroform
Phenol-Chloroform reagents Extraction with phenol and phenol/chloroform mixtures is a universal method for purification of DNA and RNA. Proteins and restriction enzymes are removed by phenol and chloroform
Supplementary Figure 1. Phenotype of the HI strain.
 Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Penghui Li, Beibei Chen, Gaoyang Zhang, Longxiang Chen, Qiang Dong, Jiangqi Wen, Kirankumar S. Mysore and Jian Zhao
 New Phytologist Supporting Information Regulation of anthocyanin and proanthocyanidin biosynthesis by Medicago truncatula bhlh transcription factor MtTT8 Penghui Li, Beibei Chen, Gaoyang Zhang, Longxiang
New Phytologist Supporting Information Regulation of anthocyanin and proanthocyanidin biosynthesis by Medicago truncatula bhlh transcription factor MtTT8 Penghui Li, Beibei Chen, Gaoyang Zhang, Longxiang
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
MCB 141 Midterm I Feb. 19, 2009
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Evolution of Transcription factor function: Homeotic (Hox) proteins
 proteins") Evolution of Transcription factor function: Homeotic (Hox) proteins Hox proteins regulate morphology in cellular zones on the anterior-posterior axis of embryos via the activation/repression of unknown
Evolution of Transcription factor function: Homeotic (Hox) proteins Hox proteins regulate morphology in cellular zones on the anterior-posterior axis of embryos via the activation/repression of unknown
Data Sheet. Azide Cy5 RNA T7 Transcription Kit
 Cat. No. Size 1. Description PP-501-Cy5 10 reactions à 40 µl For in vitro use only Quality guaranteed for 12 months Store all components at -20 C. Avoid freeze and thaw cycles. DBCO-Sulfo-Cy5 must be stored
Cat. No. Size 1. Description PP-501-Cy5 10 reactions à 40 µl For in vitro use only Quality guaranteed for 12 months Store all components at -20 C. Avoid freeze and thaw cycles. DBCO-Sulfo-Cy5 must be stored
allosteric cis-acting DNA element coding strand dominant constitutive mutation coordinate regulation of genes denatured
 A B C D E F G H I J K L M N O P Q R S T U V W X Y Z AA BB CC DD EE FF GG HH II JJ KK LL MM NN OO PP QQ RR SS TT UU VV allosteric cis-acting DNA element coding strand codominant constitutive mutation coordinate
A B C D E F G H I J K L M N O P Q R S T U V W X Y Z AA BB CC DD EE FF GG HH II JJ KK LL MM NN OO PP QQ RR SS TT UU VV allosteric cis-acting DNA element coding strand codominant constitutive mutation coordinate
RNA Transport. R preps R preps
 RNA Transport R0527-00 5 preps R0527-01 50 preps July 2014 RNA Transport Table of Contents Introduction...2 Kit Contents/Storage and Stability...3 Protocol...4 Storage Procedure...4 Recovery Procedure...5
RNA Transport R0527-00 5 preps R0527-01 50 preps July 2014 RNA Transport Table of Contents Introduction...2 Kit Contents/Storage and Stability...3 Protocol...4 Storage Procedure...4 Recovery Procedure...5
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
Separating the adhesive and signaling functions of the Fat and Dachsous protocadherins
 RESEARCH ARTICLE 2315 Development 133, 2315-2324 (2006) doi:10.1242/dev.02401 Separating the adhesive and signaling functions of the Fat and Dachsous protocadherins Hitoshi Matakatsu and Seth S. Blair*
RESEARCH ARTICLE 2315 Development 133, 2315-2324 (2006) doi:10.1242/dev.02401 Separating the adhesive and signaling functions of the Fat and Dachsous protocadherins Hitoshi Matakatsu and Seth S. Blair*
Review. Planar Polarity and Tissue Morphogenesis. Leading Edge
 Leading Edge Review Planar Polarity and Tissue Morphogenesis Jennifer A. Zallen 1, * 1 Developmental Biology Program, Sloan-Kettering Institute, New York, NY 10021, USA *Correspondence: zallenj@mskcc.org
Leading Edge Review Planar Polarity and Tissue Morphogenesis Jennifer A. Zallen 1, * 1 Developmental Biology Program, Sloan-Kettering Institute, New York, NY 10021, USA *Correspondence: zallenj@mskcc.org
7.06 Cell Biology EXAM #3 KEY
 7.06 Cell Biology EXAM #3 KEY May 2, 2006 This is an OPEN BOOK exam, and you are allowed access to books, a calculator, and notes BUT NOT computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 KEY May 2, 2006 This is an OPEN BOOK exam, and you are allowed access to books, a calculator, and notes BUT NOT computers or any other types of electronic devices. Please write
Role of Mitochondrial Remodeling in Programmed Cell Death in
 Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Preparing Colloidal Gold for Electron Microscopy
 Corporate Headquarters 400 Valley Road Warrington, PA 18976 1-800-523-2575 FAX 1-800-343-3291 Email: info@polysciences.com www.polysciences.com Europe - Germany Polysciences Europe GmbH Handelsstr. 3 D-69214
Corporate Headquarters 400 Valley Road Warrington, PA 18976 1-800-523-2575 FAX 1-800-343-3291 Email: info@polysciences.com www.polysciences.com Europe - Germany Polysciences Europe GmbH Handelsstr. 3 D-69214
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Honors Biology Reading Guide Chapter 11
 Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Cyproheptadine ELISA Test Kit
 Cyproheptadine ELISA Test Kit Catalog No. LSY-10041 1. Principal This test kit is based on the competitive enzyme immunoassay for the detection of Cyproheptadine in urine, tissue sample. The coupling antigens
Cyproheptadine ELISA Test Kit Catalog No. LSY-10041 1. Principal This test kit is based on the competitive enzyme immunoassay for the detection of Cyproheptadine in urine, tissue sample. The coupling antigens
cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody
 Based on Monoclonal Anti-cGMP Antibody") (FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
(FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
Tandem repeat 16,225 20,284. 0kb 5kb 10kb 15kb 20kb 25kb 30kb 35kb
 Overview Fosmid XAAA112 consists of 34,783 nucleotides. Blat results indicate that this fosmid has significant identity to the 2R chromosome of D.melanogaster. Evidence suggests that fosmid XAAA112 contains
Overview Fosmid XAAA112 consists of 34,783 nucleotides. Blat results indicate that this fosmid has significant identity to the 2R chromosome of D.melanogaster. Evidence suggests that fosmid XAAA112 contains
Supporting Online Material. On-Chip Dielectrophoretic Co-Assembly of Live Cells and. Particles into Responsive Biomaterials
 Supporting Online Material On-Chip Dielectrophoretic Co-Assembly of Live Cells and Particles into esponsive Biomaterials Shalini Gupta, ossitza G. Alargova, Peter K. Kilpatrick and Orlin D. Velev* Description
Supporting Online Material On-Chip Dielectrophoretic Co-Assembly of Live Cells and Particles into esponsive Biomaterials Shalini Gupta, ossitza G. Alargova, Peter K. Kilpatrick and Orlin D. Velev* Description
GFP GAL bp 3964 bp
 Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
Supporting Information
 Supporting Information Mullins et al. 10.1073/pnas.0906781106 SI Text Detection of Calcium Binding by 45 Ca 2 Overlay. The 45 CaCl 2 (1 mci, 37 MBq) was obtained from NEN. The general method of 45 Ca 2
Supporting Information Mullins et al. 10.1073/pnas.0906781106 SI Text Detection of Calcium Binding by 45 Ca 2 Overlay. The 45 CaCl 2 (1 mci, 37 MBq) was obtained from NEN. The general method of 45 Ca 2
Bis sulfone Reagents. Figure 1.
 Bis sulfone Reagents An intact IgG molecule has four accessible inter chain disulfide bonds that can be reduced to form eight free cysteine thiols, which can serve as sites for conjugation. The reaction
Bis sulfone Reagents An intact IgG molecule has four accessible inter chain disulfide bonds that can be reduced to form eight free cysteine thiols, which can serve as sites for conjugation. The reaction
Crime Prep Adem-Kit (Cat #06210) Instruction for manual protocol
 Instruction for manual protocol") (Cat #06210) Instruction for manual protocol ADEMTECH SA Parc scientifique Unitec 1 4, allée du Doyen G. Brus 33600 PESSAC France Tel: +33557020201 Fax +3355702020 Visit our Web site: www.ademtech.com
(Cat #06210) Instruction for manual protocol ADEMTECH SA Parc scientifique Unitec 1 4, allée du Doyen G. Brus 33600 PESSAC France Tel: +33557020201 Fax +3355702020 Visit our Web site: www.ademtech.com
ADH Activity Assay Kit
 ADH Activity Assay Kit Catalog Number KA3713 100 assays Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
ADH Activity Assay Kit Catalog Number KA3713 100 assays Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Transport of glucose across epithelial cells: a. Gluc/Na cotransport; b. Gluc transporter Alberts
 Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
RayBio Rat TNF-alpha ELISA Kit (For Lysates)
") RayBio Rat TNF-alpha ELISA Kit (For Lysates) Catalog #: ELR-TNFa-CL User Manual Last revised April 15, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite
RayBio Rat TNF-alpha ELISA Kit (For Lysates) Catalog #: ELR-TNFa-CL User Manual Last revised April 15, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite
Genetic Lab 3. Drosophila Fly
 Genetic Lab 3 Drosophila Fly An Introduction to fruit or vinegar fly Drosophila Melanogaster Is a small (about 3mm long), common fly found near unripe and rotted fruit, so that it called fruit or vinegar
Genetic Lab 3 Drosophila Fly An Introduction to fruit or vinegar fly Drosophila Melanogaster Is a small (about 3mm long), common fly found near unripe and rotted fruit, so that it called fruit or vinegar
Biology 112 Practice Midterm Questions
 Biology 112 Practice Midterm Questions 1. Identify which statement is true or false I. Bacterial cell walls prevent osmotic lysis II. All bacterial cell walls contain an LPS layer III. In a Gram stain,
Biology 112 Practice Midterm Questions 1. Identify which statement is true or false I. Bacterial cell walls prevent osmotic lysis II. All bacterial cell walls contain an LPS layer III. In a Gram stain,
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Human papillomavirus,hpv ELISA Kit
 Human papillomavirus,hpv ELISA Kit Catalog No: E0787h 96 Tests Operating instructions www.eiaab.com FOR RESEARCH USE ONLY; NOT FOR THERAPEUTIC OR DIAGNOSTIC APPLICATIONS! PLEASE READ THROUGH ENTIRE PROCEDURE
Human papillomavirus,hpv ELISA Kit Catalog No: E0787h 96 Tests Operating instructions www.eiaab.com FOR RESEARCH USE ONLY; NOT FOR THERAPEUTIC OR DIAGNOSTIC APPLICATIONS! PLEASE READ THROUGH ENTIRE PROCEDURE
AP Biology Unit 6 Practice Test 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8
 AP Biology Unit 6 Practice Test Name: 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8 picograms of DNA per nucleus. How many picograms
AP Biology Unit 6 Practice Test Name: 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8 picograms of DNA per nucleus. How many picograms
Frizzled signalling and cell polarisation in Drosophila and vertebrates
 Review 4501 Frizzled signalling and cell polarisation in Drosophila and vertebrates David Strutt Centre for Developmental Genetics, Department of Biomedical Science, University of Sheffield, Western Bank,
Review 4501 Frizzled signalling and cell polarisation in Drosophila and vertebrates David Strutt Centre for Developmental Genetics, Department of Biomedical Science, University of Sheffield, Western Bank,
RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit
 ELISA Kit") RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit Catalog #: PEL-EIF2A-S52 User Manual Last revised August 10, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway
RayBio Human Phospho-eIF- 2alpha (Ser52) ELISA Kit Catalog #: PEL-EIF2A-S52 User Manual Last revised August 10, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway
SUPPLEMENTARY INFORMATION
 Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Functional Genomics Research Stream
 Functional Genomics Research Stream http://fc09.deviantart.net/fs70/i/2010/214/2/f/dna_heart_by_micche.jpg http://www.ryersondesigns.com/skanndelus/dnaheart.jpg Research Meeting: February 14, 2012 Nucleic
Functional Genomics Research Stream http://fc09.deviantart.net/fs70/i/2010/214/2/f/dna_heart_by_micche.jpg http://www.ryersondesigns.com/skanndelus/dnaheart.jpg Research Meeting: February 14, 2012 Nucleic
Lab I: Three-Point Mapping in Drosophila melanogaster
 Lab I: Three-Point Mapping in Drosophila melanogaster Makuo Aneke Partner: Christina Hwang BIO 365-004: Genetics with Laboratory TA: Dr. Hongmei Ma February 18, 2016 Abstract The purpose of this experiment
Lab I: Three-Point Mapping in Drosophila melanogaster Makuo Aneke Partner: Christina Hwang BIO 365-004: Genetics with Laboratory TA: Dr. Hongmei Ma February 18, 2016 Abstract The purpose of this experiment
MQCA ELISA Test Kit. Catalog No. LSY-10046
 MQCA ELISA Test Kit Catalog No. LSY-10046 1. Principle This test kit is based on the competitive enzyme immunoassay for the detection of MQCA in the sample. The coupling antigens are pre-coated on the
MQCA ELISA Test Kit Catalog No. LSY-10046 1. Principle This test kit is based on the competitive enzyme immunoassay for the detection of MQCA in the sample. The coupling antigens are pre-coated on the
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
 DOI: 10.1038/ncb2819 Gαi3 / Actin / Acetylated Tubulin Gαi3 / Actin / Acetylated Tubulin a a Gαi3 a Actin Gαi3 WT Gαi3 WT Gαi3 WT b b Gαi3 b Actin Gαi3 KO Gαi3 KO Gαi3 KO # # Figure S1 Loss of protein
DOI: 10.1038/ncb2819 Gαi3 / Actin / Acetylated Tubulin Gαi3 / Actin / Acetylated Tubulin a a Gαi3 a Actin Gαi3 WT Gαi3 WT Gαi3 WT b b Gαi3 b Actin Gαi3 KO Gαi3 KO Gαi3 KO # # Figure S1 Loss of protein
Supplemental Materials Molecular Biology of the Cell
 Supplemental Materials Molecular iology of the Cell Figure S1 Krüger et al. Arabidopsis Plasmodium H. sapiens* 1) Xenopus* 1) Drosophila C.elegans S.cerevisae S.pombe L.major T.cruzi T.brucei DCP5 CITH
Supplemental Materials Molecular iology of the Cell Figure S1 Krüger et al. Arabidopsis Plasmodium H. sapiens* 1) Xenopus* 1) Drosophila C.elegans S.cerevisae S.pombe L.major T.cruzi T.brucei DCP5 CITH
Full-length GlpG sequence was generated by PCR from E. coli genomic DNA. (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
,") Supplementary Methods Protein expression and purification Full-length GlpG sequence was generated by PCR from E. coli genomic DNA (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
Supplementary Methods Protein expression and purification Full-length GlpG sequence was generated by PCR from E. coli genomic DNA (with two sequence variations, D51E/L52V, from the gene bank entry aac28166),
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms.
 Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
dachsous and frizzled contribute separately to planar polarity in the Drosophila ventral epidermis
 RESEARCH ARTICLE 2751 Development 138, 2751-2759 (2011) doi:10.1242/dev.063024 2011. Published by The Company of Biologists Ltd dachsous and frizzled contribute separately to planar polarity in the Drosophila
RESEARCH ARTICLE 2751 Development 138, 2751-2759 (2011) doi:10.1242/dev.063024 2011. Published by The Company of Biologists Ltd dachsous and frizzled contribute separately to planar polarity in the Drosophila
Center for Cell Imaging Department of Cell Biology
 Center for Cell Imaging Department of Cell Biology Contents Preparation of Colloidal Gold Conjugates Coupling the Protein A to the Gold Particles Purification of the protein A-gold. Storage Influence of
Center for Cell Imaging Department of Cell Biology Contents Preparation of Colloidal Gold Conjugates Coupling the Protein A to the Gold Particles Purification of the protein A-gold. Storage Influence of