Comparative study on memory phases in the parasitic wasps Cotesia glomerata and Cotesia rubecula
|
|
|
- Julia Bridges
- 5 years ago
- Views:
Transcription

1 Department of Zoology Institute of animal ecology Prof. Dr. Johannes Steidle University of Hohenheim (220c) Stuttgart, Germany Laboratory of Entomology Dr. Hans Smid University of Wageningen The Netherlands Comparative study on memory phases in the parasitic wasps Cotesia glomerata and Cotesia rubecula Diploma Thesis This work was financially supported by the Herzog-Carl-Foundation, University of Hohenheim Submitted by Silja Tribuhl (07.05) Hohenheim, Stuttgart, March 2007 Faculty of Agricultural Sciences
2 Table of contents I Table of contents Comparative study on memory phases in the parasitic wasps Cotesia glomerata and Cotesia rubecula List of figures III List of tables IV 1. Summary 1 2. Introduction Insects and learning Associative learning Learning and memory formation Cotesia glomerata and Cotesia rubecula Memory phases in Cotesia glomerata and Cotesia rubecula Material and Methods Insects Plants Odour source Wind tunnel Experimental design Wind tunnel assay Statistical analysis Results Results of different learning trials in Cotesia glomerata Results of single learning trials in C. glomerata and C. rubecula after 4 hours Discussion Cotesia glomerata Cotesia rubecula Variations in learning during the experiments Costs and benefits of learning Cotesia glomerata and Cotesia rubecula why are they so different in learning? References 28
3 Table of contents II Appendix: Transition from solitary to gregarious development in parasitic wasps 1. Introduction Solitary and gregarious development Transition from solitary to gregarious development Cotesia glomerata and Cotesia rubecula Material and Methods Experimental setup Statistical analysis Results Discussion References 43
4 List of figures III List of figures Comparative study on memory phases in the parasitic wasps Cotesia glomerata and Cotesia rubecula Fig. 1: Putative pathway of memory formation. 4 Fig. 2: Putative pathway of memory formation. 5 Fig. 3: Life cycle of parasitic wasps. 6 Fig. 4: Memory retention in C. glomerata and C. rubecula. 8 Fig. 5a: Memory retention in C. rubecula after three spaced trials. 9 Fig. 5b: Memory retention in C. glomerata after three spaced trials. 9 Fig. 6: Memory retention in C. glomerata and C. rubecula 4 hours after single training. 10 Fig. 7: Wind tunnel set-up. 13 Fig. 8: Wind tunnel. 14 Fig. 9: Cotesia glomerata: Choice for cabbage or nasturtium after no, one single or three spaced learning trials. 16 Fig. 10: C. glomerata and C. rubecula: Choice for nasturtium depending on the treatment. 18 Appendix: Transition from solitary to gregarious development in parasitic wasps Fig. 1: Mean numbers of twisting and folding in mymariform larvae of the solitary A. victus and the gregarious A. listronoti. 35 Fig. 2: Mean numbers of lateral bending of the solitary C. rubecula and the gregarious C. glomerata. 37 Fig. 3: Larvae of C. glomerata and C. rubecula 6 days after oviposition. 37 Fig. 4: Mouthparts of larvae of C. glomerata and C. rubecula 5 to 8 days after oviposition. 38 Fig. 5: The evolution of gregariousness. 41
5 List of tables IV List of tables Comparative study on memory phases in the parasitic wasps Cotesia glomerata and Cotesia rubecula Table 1: C. glomerata: Choice for nasturtium after different learning trials. 17 Table 2: C. glomerata: Choice for nasturtium 4 hours after one learning trial. 19 Table 3: C. rubecula: Choice for nasturtium 4 hours after one learning trial. 19 Appendix: Transition from solitary to gregarious development in parasitic wasps Table 1: Mean numbers of bending in C. glomerata and C. rubecula. 37
6 1. Summary 1 Comparative study on memory phases in the parasitic wasps Cotesia glomerata and Cotesia rubecula 1. Summary The parasitic wasps Cotesia glomerata and Cotesia rubecula are closely related and live in the same habitat in the Netherlands. The wasps lay their eggs in caterpillars of Pieris butterflies. To find their hosts the wasps are innately attracted by volatiles emitted by infested cabbage plants. If hosts are encountered on plants different than cabbage the wasps learn to associate the odour of this plant species with the presence of suitable hosts. Although the wasps are closely related, clear differences in learning and memory formation occur. C. glomerata forms long-term memory (LTM) already after one successful egg laying experience whereas C. rubecula needs three spaced experiences. Former experiments assumed that after three spaced conditioning trials LTM is consolidated directly out of anesthesia sensitive memory (ASM) in C. glomerata and no anesthesia resistant memory (ARM) is present. In C. rubecula two memory traces coexist after three spaced trials, ARM and LTM. To test if ARM is present in C. glomerata a classical conditioning setup is used, in combination with cooling to inhibit ASM and with a translation inhibitor (anisomycin) that blocks LTM. Both species were tested with one conditioning trial. In C. glomerata both, cooling and anisomycin inhibited 4 hour memory retention, which shows that ARM seems to be absent in C. glomerata after 4 hours. ASM is directly consolidated into LTM in this species suggesting that LTM induction prevents formation of ARM. In C. rubecula there was no effect of cooling on 4 hour memory retention, and previous results showed that 4 hour memory retention in this species cannot be inhibited by anisomycin. This shows that 4 hour memory retention is constituted exclusively by ARM. Looking at learning and memory formation in the context of the natural behaviour of the wasps, it becomes clearer why these differences in memory formation might occur. For C. glomerata finding a host on a special plant species is reliable information due to the egg laying performance of its host P. brassicae which chooses a stand of plants of the same species. If C. glomerata associates the odour of the new encountered plant with the presence of suitable hosts it can be sure to find enough hosts for its offspring. Therefore new memory is stored directly into LTM. For C. rubecula finding a host is not reliable information because
7 2. Introduction 2 its host P. rapae spreads its eggs widely on different host plants. For the wasp it would be maladaptive to change its innate behaviour and search for the new encountered plant species. Still associations are formed in C. rubecula but this memory is stored in ASM and ARM which wane within one day and are a form of low cost memories, demanding less energetic costs than the formation of LTM which requires new gene expression. 2. Introduction 2.1 Insects and learning When you think of insects you think of tiny little animals flying or crawling around maybe disturbing you. The idea that these little insects are able to learn and that they react in a flexible way to their environment would maybe never come to your mind. Do insects have a brain? is a frequently asked question when I tell people about my research. Of course the brains of insects look a little bit different than brains in higher animals or humans but the way they function is not very different. Insects are able to learn and this fact has been of great interest for a lot of researchers all over the world. Are there any advantages for the insects if they are able to learn? Do they all learn in the same way, with the same speed? How long are they able to remember learned information? Which genes are involved in memory formation? Especially the fruit fly Drosophila has been well-investigated concerning these questions but also parasitic wasps are used to know more about different forms of learning behaviour. Knowing more about learning in insects enables us to know more about our own learning behaviour. 2.2 Associative learning To know more about learning, experiments with all kinds of animals have been performed until now. Famous experiments have been carried out by PAVLOV (1927). In his experiments Pavlov always rang a bell directly before he fed a dog. After a while the dog started salivating when it heard the sound of the bell alone. This association between a neutral stimulus (called conditioned stimulus or CS) the bell, and a meaningful stimulus (called unconditioned stimulus or US) the food, is called pavlovian or classical conditioning and is a form of associative learning. Classical conditioning will only take place if the CS is directly followed by the US. Associative learning also plays an important role in memory formation in insects.
8 2. Introduction 3 Another form of associative learning is operant conditioning (THORNDICKE 1901, SKINNER 1938). In operant conditioning an association is formed between the response to a CS and the reinforcer (US). In one of his experiments Thorndicke put a cat in a puzzle box with a lockable door which could be opened from the inside. He observed the behaviour of the cat and noted the time the cat needed to escape from the box. After several trials the cat learned to unlock the door and escape from the box very quickly. The cat associated its specific behaviour (unlock the door) with the reinforcer (US), the freedom. This means that in operant conditioning the important stimulus (reinforcer) occurs after the response and not before it like in classical conditioning (LUTZ 2005). Operant learning is also found in parasitic wasps like Cotesia glomerata (L.) and Cotesia rubecula (Marshall) (Hymenoptera: Braconidae). C. glomerata and C. rubecula lay their eggs in caterpillars of Pieris butterflies. The larvae of the wasps develop in the caterpillars until they finally leave their dying host and pupate. To find their hosts the wasp smells a damaged plant with caterpillars feeding on it, flies towards the plant, lands on it and finds suitable hosts for oviposition. The odour of the damaged plant represents the CS, the wasp responds by landing on this plant where it is rewarded with suitable hosts (US) (SMID 2006). An association is formed between the flight response and the reward, which is represented by the host. 2.3 Learning and memory formation If insects learn, the information has to be stored somehow so memory is formed. Instead of one general memory form, different memory phases can be distinguished. JAMES (1890) first separated memory into primary and secondary memory, which are now known as short-term (STM) and long-term (LTM) memory. Memory phases can be classified by their duration but this is a criterion which differs considerably between animal species including humans. Another possible criterion to distinguish is the biochemical processes which are critically involved in forming these different memory phases (ROSENZWEIG et al. 1993). The biochemical pathways are not completely understood until now, most of all chemical substances or treatments are found which inhibit the different phases without really knowing why. STM is sensitive to amnestic treatments (e.g. cooling, narcosis with CO 2 or N 2 ), and is therefore also called anesthesia sensitive memory (ASM) (e.g. TULLY et al. 1994). LTM is sensitive to protein synthesis inhibitors showing that LTM requires new gene expression (TULLY et al. 1994).
9 2. Introduction 4 The formation of memory phases is time and event dependent meaning that different learning experiences depending on their chronology cause different memory phases. After single learning experiences STM occurs but also another form of memory appears which is called anesthesia resistant memory (ARM). ARM is resistant to amnestic treatments (e.g. TULLY et al. 1994, MARGULIES et al. 2005, SMID 2006), and is also formed after massed (repeated experiences without a break) and spaced (several experiences with breaks in between) learning experiences. LTM normally occurs only after spaced learning. Spaced learning experiences ensure that only important information is stored in long-term memory. For animals it is important not to learn too fast to prevent the storage of wrong information (SMID 2006). A lot of research about memory and memory formation has been done in the fruit fly Drosophila. The flies are trained with classical conditioning with different learning experiences (single, massed, spaced). In Drosophila STM appears immediately after training and fades away within 60 minutes. ARM appears slowly, shows its highest level two hours after training and can last for three days after massed training. LTM appears 4-5 hours after spaced learning trials (10 sessions with a 15 minute rest interval in between) lasting at least one week (TULLY et al. 1994, MARGULIES et al. 2005). In literature you find contradictive opinions about the way these different memory phases occur, if they appear sequentially, meaning one phase consolidating into the other, or also in parallel. MARGULIES et al. (2005) state in their review that ARM and LTM are genetically distinct, formed independently of each other and function in an additive way (Fig. 1). ARM Acquisition ASM LTM Fig. 1: Putative pathway of memory formation: ARM and LTM are genetically distinct and formed in parallel (adapted from TULLY et al. 1994). These statements are based on studies of TULLY et al. (1994) and YIN et al. (1994) that showed that LTM but not ARM is protein synthesis dependent. This means that ARM is not affected by protein synthesis inhibitors and there has to be a different underlying mechanism.
10 2. Introduction 5 ISABEL et al. (2004) presented a different hypothesis; in their research ARM and LTM do not coexist in Drosophila but LTM replaces ARM (Fig. 2). Drosophila mutants in which LTM is deficient showed almost no memory retention 5 hours after spaced conditioning. Normally ARM should be present 5 hours after spaced training and last for several days. It is suggested that ARM is blocked during LTM-specific training. The assumption is that in wild-type flies LTM is formed after spaced conditioning and replaces ARM which is only formed after massed conditioning. This mechanism is thought to ensure that only information with a high predictive value is stored in LTM. Acquisition ASM ARM LTM Fig. 2: Putative pathway of memory formation: LTM replaces ARM during spaced learning experiences. 2.4 Cotesia glomerata and Cotesia rubecula C. glomerata and C. rubecula are two closely related parasitic wasps coexisting in the same habitat in the Netherlands and are indigenous to Europe. They belong to the endoparasitoids, they lay their eggs in young larvae (first to third larval instar) of Pieris butterflies. The larvae of the wasps leave the caterpillar approximately after 2 weeks by eating through its cuticle and thereafter the caterpillar dies. The larvae spin a cocoon and develop to the pupal stage (Fig. 3) (SMID 2006). C. glomerata is a generalist, it can parasitize Pieris rapae (L.), Pieris brassicae (L.) and Pieris napi (L.) but in the Netherlands it parasitizes mainly the large cabbage white P. brassicae (GEERVLIET et al. 2000a). C. glomerata is a gregarious species; approximately 20 eggs are laid in a single host (VOS & VET 2004). The solitary wasp C. rubecula is a specialist on the small cabbage white P. rapae; only one egg is laid in a single host (GEERVLIET et al. 2000b).

11 2. Introduction 6 Fig. 3: Life cycle of parasitic wasps (TUMLINSON et al. 1993). The wasps emerging from the cocoons have to find food, a mate and a host to lay their eggs. To find a suitable host is quite a difficult task, the environment of the wasps is full of different odours emitted by plants and non-hosts. Also the young caterpillars are very small; they are well camouflaged and do not emit attractants which are detectable from a distance (SMID 2006). So, how are the wasps able to find their hosts? Caterpillars of Pieris butterflies are restricted to plants containing glucosinolates like most of the cruciferous plants (CHEW 1980). The wasps could orientate itself from distance towards the plant odours but still this stimulus is not very reliable (VET & DICKE 1992) because the plant odours by themselves do not promise the presence of suitable hosts. GEERVLIET et al. (1994) showed that volatiles of cabbage plants on which caterpillars are feeding are highly attractive to the wasps. This means plants change their volatile composition when they are damaged by feeding caterpillars. The wasps orientate at first towards the volatiles which are emitted by the damaged plants and not towards the odour of the caterpillars. The wasps do not have to learn to recognize the plant odour of cabbage; naïve wasps already have this preference (GEERVLIET et al. 1996). After landing on a host plant with caterpillars, the wasps starts local searching behaviour by using host-derived stimuli like frass or silk. Also non-cruciferous plants containing glucosinolates, e.g. nasturtium (Tropaeolum major L.), are used as a host plant by Pieris butterflies (FELTWELL 1982, GEERVLIET 1997). If the wasp encounters a host e.g. on nasturtium it can learn to associate the odour of this different plant species with the presence of a suitable host. The wasp changes its innate
12 2. Introduction 7 behaviour and starts searching for hosts on nasturtium. C. glomerata changes its preference already after one oviposition experience on a plant different than cabbage and forms a longlasting memory of at least 2-3 days. C. rubecula also learns to associate the odours of other plant species with the presence of a suitable host but this memory already wanes within one day (BLEEKER 2005, SMID submitted). Thus there is a difference in host-searching strategies between the generalist C. glomerata and the specialist C. rubecula (GEERVLIET et al. 1998, SMID 2006). In the case of C. glomerata and C. rubecula the difference in preference behaviour might be due to their different host distributions (BLEEKER 2006). P. brassicae females lay their eggs in clusters of approximately 150 eggs per plant. The caterpillars are conditioned on the first species they feed on and need to find more plants of the same species. P. brassicae therefore selects a stand of host plants of the same species to ensure enough food for their offspring (ROTHSCHILD 1987, LE MASURIER 1994). C. glomerata will find enough hosts for its own offspring if it orientates itself towards the odour of the specific plant species. Therefore it is useful for the wasp to associate the odour of the newly encountered plant with the presence of its host. This change in behaviour ensures C. glomerata to find enough hosts to lay all its eggs. P. rapae usually lays only one egg per plant and spreads its eggs widely in their habitat on different host plant species (ROOT & KAREIVA 1984). For C. rubecula the presence of one caterpillar does not indicate the presence of other suitable hosts so it would be a disadvantage to change its preference towards the newly encountered plant (BLEEKER 2006). C. glomerata and C. rubecula are two closely related wasps; they live in the same habitat and show clear differences in learning behaviour. Therefore these two wasps are a good model for comparative investigations about different memory phases and duration of memory retention. 2.5 Memory phases in Cotesia glomerata and Cotesia rubecula In C. glomerata and C. rubecula 3 memory phases are distinguished until now. We find ASM, ARM and LTM but there seem to be differences in the two parasitic wasps. SMID et al. (submitted) trained the wasps with classical conditioning on nasturtium plants (infested with Pieris caterpillars) and tested them for memory retention in a dual choice test in the wind tunnel. The wasps had a choice between infested cabbage and nasturtium plants. Memory retention could be seen on the basis of the preference shift from cabbage to nasturtium. If the wasps receive one learning trial both wasps show high memory retention after one and four hours (Fig. 4); around 70 % of the wasps chose for nasturtium in the wind
.")
13 2. Introduction 8 tunnel. After 24 hours there is no memory left in C. rubecula whereas in C. glomerata there is still high memory retention. C. glomerata was also fed with the translation blocker anisomycin which inhibits LTM. After one conditioning trial 24 hour memory retention was completely inhibited showing that the wasp forms long-term memory already after one oviposition experience (SMID et al. submitted). This phenomenon has also been shown for e.g. appetitive conditioning in the snail Lymnea (FULTON et al. 2005), fear conditioning in rats (IGAZ et al. 2002), and oviposition learning in parasitic wasps (COLLATZ et al. 2006). In C. rubecula no LTM is formed after one learning trial (SMID et al. submitted). Choice for nasturtium (%) Time (h) Fig. 4: Memory retention in C. glomerata and C. rubecula after 1, 4, and 24 hours after one learning trial (SMID submitted). Both wasp species also received three learning trials (spaced training with 10 minutes break in between), and were fed with sucrose solution alone or with anisomycin. Again there are differences between the two species. In C. rubecula, fed with sucrose, there are still high levels of memory retention after 5 days. Fed with anisomycin the wasp still shows memory retention after 24 hours (Fig. 5a). Memory is only partially inhibited by anisomycin showing that 24 hour retention is composed of a protein-synthesis independent component (ARM) and of a protein-synthesis dependent component (LTM).
.")
14 2. Introduction 9 Time (h) Fig. 5a: Memory retention in C. rubecula after three spaced trials. Bright bars: Wasps were fed with 3 % sucrose solution. There is still memory retention after 5 days. Dark bars: Wasps were fed with anisomycin in 3 % sucrose solution. Although LTM is inhibited by anisomycin there is still memory retention left after 24 and 48 hours (SMID submitted). Choice for nasturtium (%) Choice for nasturtium (%) Time (h) Fig. 5b: Memory retention in C. glomerata after three spaced trials. Bright bars: Wasps were fed with 3 % sucrose solution. There is still memory retention after 5 days. Dark bars: Wasps were fed with anisomycin in 3 % sucrose solution. After 4 hours there is no memory retention left (SMID submitted). Feeding C. glomerata with anisomycin the picture looks different (Fig. 5b). After 4 hours there is no memory retention left which shows that from 4 hours on memory retention is
15 2. Introduction 10 composed completely of a protein-synthesis dependent component (LTM). This result leads to the question if ARM might be absent in C. glomerata (SMID submitted). To get more information about ASM also experiments with cooling were carried out by WANG (unpublished). Cooling inhibits ASM without blocking ARM and LTM. The wasps were trained with classical conditioning as explained before, received a single training and were cooled in ice for two minutes to inhibit ASM. Cooling was carried out directly, 20 min, 1, 2 and 3 hours after training (Fig. 6). The wasps were tested after 4 hours in the wind tunnel. choice for nasturtium (%) , Cotesia glomerata Cotesia rubecula cooling time after oviposition (h) Fig. 6: Memory retention in C. glomerata and C. rubecula 4 hours after single training. The wasps were cooled directly, 20 min, 1, 2 and 3 hours after oviposition (WANG unpublished). Direct cooling after training had no effect on memory retention but cooling after 20 min seems to have a major effect because in both species there is no memory retention left after 4 hours. The later the wasps were cooled the less effect is seen on memory retention showing that ASM is consolidating into ARM and/or LTM which are not sensitive to cooling. There seems to be a sensible phase 20 min after training concerning the consolidation of ARM and LTM. This could be explained if ARM and LTM are consolidated out of ASM because inhibiting ASM seems to influence the formation of ARM and LTM. This thesis will focus on some remaining questions of the studies mentioned above. Comparing the memory phases of C. glomerata and C. rubecula you see that in C. rubecula ARM seems to be present and formed in parallel with LTM (SMID submitted). This is comparable with the situation that MARGULIES et al. (2005) described for Drosophila (Fig. 1).
16 3. Materials and methods 11 In C. glomerata ARM seems to be absent and LTM might be directly consolidated out of ASM. This would support the suggestions of ISABEL et al. (2004) that memory phases are mutually exclusive and one phase replaces the other. This research study will try to find out if ARM is present in C. glomerata 4 hours after one conditioning trial. 4 hours after one trial both species show high memory retention (Fig. 4). This memory does not seem to consist of ASM because cooling 3 hours after training had no effect on memory retention after 4 hours any more (Fig. 6) and ASM seems to be consolidated into ARM or LTM by that time. If C. glomerata is cooled after 3 hours (to inhibit ASM) and fed with anisomycin (to block LTM) there should be no memory retention left after 4 hours if ARM is missing. The same experiments will be carried out with C. rubecula where ARM seems to be present. Another question is if LTM consolidates out of ASM and at which point in time LTM starts in C. glomerata. Data of WANG (unpublished) show, that cooling seems to have an influence on the consolidation of LTM. Cooling should normally only inhibit ASM. In this study wasps will be trained and tested in the wind tunnel after different points in time and also cooled after different times to state more precisely when LTM starts and if it is formed out of ASM. 3. Materials and methods 3.1 Insects C. glomerata and C. rubecula originated from individuals collected on cabbage fields near Wageningen, The Netherlands. The parasitoids were maintained on P. brassicae and P. rapae larvae respectively, feeding on cabbage plants in a climatic room at C, % r.h. and a L16:D8 regime as described by GEERVLIET et al. (1994). The cocoons were kept in cages under the same conditions and the emerging parasitoids were supplied with water and honey. P. brassicae and P. rapae larvae were maintained on cabbage plants under the conditions described above. For the experiments first instar larvae were used. 3.2 Plants Cabbage (Brassica oleracea gemmifera cv. Cyrus) and nasturtium plants (Tropaleum majus L. cv. Glorious Gleam) were reared in a greenhouse at C, % r.h. and a L16:D8 regime.
17 3. Materials and methods Odour source Cabbage and nasturtium plants of approximately the same size were chosen. On two leaves of each plant 20 early first instar larvae were transferred and were allowed to feed for 24 hours to induce feeding damage. For the oviposition experience nasturtium plants were treated the same way. 3.4 Wind tunnel The wind tunnel (Fig. 7) used in these experiments measures 200 x 60 x 60 cm. The flightcompartment is made out of polyacryl; both ends are covered with fine mesh copper gauze. At the observer s side there are three little gates which can be opened to put in the odour source and the wasps. During the experiments outside air is filtered over glass wool and led over a humidity controlled air humidifier. The air is then pressurised, filtered over an active charcoal filter and passes through a temperature controlled heating system. Wind speed was constantly at 18 cm/s. The temperature varied between C, relative humidity fluctuated between 40 and 60 %. Above the flight-compartment there is one half of a concave metal cylinder with 8 fluorescent high-frequency tubes and 4 bulbs as a light source. The inner side of the cylinder is covered with aluminium plates for light dispersion. Light intensity during the tests was 1700 lux on the bottom, increasing to 2000 lux at the ceiling. The wasps were released in a horizontal glass cylinder (30 cm long, 15 cm diameter) 40 cm in distance of the odour source (GEERVLIET et al. 1994).
18 3. Materials and methods 13 Fig. 7: Wind tunnel set-up: AI = air inlet, CF = charcoal filter, HS = heating system, RP = parasitoid release point, FT = high-frequency fluorescent tubes, B = bulbs, X = position of odour source, HM = hygrometer, G = gate (GEERVLIET et al. 1994). 3.5 Experimental design 5-8 days old females of C. glomerata and C. rubecula were deprived from water and honey over night and fed the next day with 0,5 l of a solution containing 5 mm anisomycin (2-pmethoxyphenylmethyl-3-acetoxy-4-hydroxypyrrolidine) in 3 % sucrose. The wasps were kept in vials for 2.5 hours until they consumed the entire solution. After this they were transferred for one hour to a glass cage with honey and water. The wasps were trained by giving them a single oviposition experience on first instar larvae of P. brassicae and P. rapae, which had been feeding on nasturtium plants for 24 hours. In these experiments the wasps are trained with classical conditioning to standardize the experiments. The wasps are placed directly on the leaf (odour = CS) and the taste/odour of the host or host-derived products represents the US. An association is formed between the neutral stimulus (plant odour) which is directly followed by a meaningful stimulus, the suitable host. The test for memory formation in the wind tunnel represents an operant context because the reinforcing stimulus (suitable host) occurs after the response (flight) to the conditioned stimulus (plant odour). The wasps are able to transfer the information from the classical conditioning to the operant context.


19 3. Materials and methods 14 Three hours after oviposition the wasps were cooled for 2 minutes in ice. Four hours after oviposition the wasps were tested in the wind tunnel for their preferences, infested cabbage or nasturtium. Forty individuals were tested per group. Control groups were given the same treatment as explained above. Control group 1: Control group 2: Control group 3: Control group 4: training + anisomycin training + cooling cooling + anisomycin training 3.6 Wind tunnel assay The wind tunnel as described in GEERVLIET et al. (1994) was used for all experiments. The wasps had the choice between odour sources of cabbage and nasturtium plants, both infested with P. brassicae or P. rapae larvae for 24 hours (Fig. 8). Each wasp was released 40 cm downwind from the plants. Every test lasted a maximum time of five minutes. Landing on one of the plants was recorded as a positive response. Wasps showing no preference or landing somewhere else within 5 min were recorded as a no response. After testing five wasps the position of the plants was switched. All wasps were used only once. Fig. 8: Wind tunnel (REMCO VAN POECKE). Learning is defined in this study as the preference shift from cabbage to nasturtium. If there is a significant difference between naïve and trained wasps in their preference for nasturtium, learning takes place.
20 4. Results Statistical analysis Generalized linear modelling (GLM) procedures using procedure GENMOD in SAS version 8.02 (Proc GENMOD, SAS Inc, Chicago, IL, U.S.A.) were used for data with a binominal distribution of error variance and a logit-link function. When main effects or their interactions were found significantly different, further separation of the treatment levels was carried out by acquiring specific contrasts for particular comparisons, which are given in the text. Data collected on different experimental days were considered as replicates. We tested whether treatments (4 levels: training; training + anisomycin; training + cooling; training + anisomycin + cooling) and species (2 levels: C. glomerata and C. rubecula) had an effect on the response level to nasturtium plants (GLM treatment ² 6 = 41.42, P < ). Choices between odour sources were analysed with a two-tailed binominal test with continuity correction. 4. Results 4.1 Results of different learning trials in C. glomerata Experiments from SMID et al. (submitted) and WANG (unpublished) were carried out with a different population than the population that was used in this study. Every year a new population of Cotesia wasps is collected from the field and established to avoid inbreeding. Years of rearing showed that it is normal to observe a decrease in yield in the beginning of the new breed. Also this time low yields were obtained and high mortality rates. In addition, it has been found that the learning capability of the wasps was very low. Normally the wasps show a pronounced preference shift towards nasturtium hours after classical conditioning, % of the wasps choose nasturtium instead of cabbage. Only 36 % of the wasps chose for nasturtium 4 hours after training (Fig. 9) in the beginning of this study. The wasps were tested after different times (1, 4, 24 hours) and with different learning protocols (single, spaced) in the wind tunnel to check if memory retention changed dependent on different learning experiences (Fig. 9). Data were not statistically analyzed because not enough replicates were taken. Still it can be seen that there was a low preference shift from cabbage to nasturtium also after three spaced trials which should normally lead to a high level of memory retention. In contrast, control experiments with C. rubecula showed normal memory retention in an identical setup, showing that the experimental setup was functioning well. Unfortunately also
21 4. Results 16 in the rearing of C. rubecula low yields and high mortality rates were obtained. Therefore only 30 individuals were tested in the Training and Training + Cooling - groups and around 10 individuals in the Training + Anisomycin and Training + Anisomycin + Cooling Groups. It was not possible to include a naïve control group due to the low availability of the wasps. C. glomerata was tested every week to see if the mortality rates became normal in successive generations and learning performance improved to values comparable to results obtained by previous studies. Meanwhile an alternative study was performed which is described in the appendix. After several generations, batches of C. glomerata came available that showed adequate levels of memory retention again so the experiments could be started. In this study single and spaced trials were planned but finally only single training trials could be tested due to lack of individuals with adequate learning capabilities. The wasps were only tested after 4 hours and not after 5 and 6 hours as planned before. Nasturtium Cabbage naïve after 4 hours 1 hour after single trial 4 hours after single trial 24 hours after three spaced trials Choice (%) Fig. 9: Cotesia glomerata: Choice for cabbage or nasturtium after no, one single or three spaced learning trials. Test in the wind tunnel 1, 4 and 24 hours after oviposition.
22 4. Results 17 Table 1: C. glomerata: Choice for nasturtium after different learning trials, 1, 4 and 24 hours after training. Treatment Hours after oviposition Nasturtium Cabbage N N responding naïve n.a single trial single trial spaced trial Results of single learning trials in C. glomerata and C. rubecula after 4 hours A new batch of C. glomerata which showed adequate levels of memory retention was taken for the final experiments. Naïve wasps fed with anisomycin and cooled after 3 hours flew mostly to cabbage (94 %), showing their natural preference for this plant. The wasps behaved normally and showed low no-response levels (22 %) which indicates that feeding them with anisomycin and cooling has no detrimental effect on the behaviour of the wasps. In trained C. glomerata a significant preference shift from cabbage to nasturtium plants could be observed compared to naïve wasps (GLM treatment ² 1 = 11.91, P = ) (Fig. 10). 61 % of the wasps chose for nasturtium in the wind tunnel which was a result not as high as observed in previous studies (SMID submitted), normally % chooses for nasturtium. Still it shows that the wasps were able to learn and that they showed memory retention after 4 hours. Cooling trained wasps after 3 hours reduced memory retention significantly from trained wasps (61 %) to 30 % (GLM treatment ² 1 = 4.40, P = ) but the memory retention level of cooled wasps was still significantly different from naïve control wasps (GLM treatment ² 1 = 5.36, P = ) (Fig. 10). Feeding wasps with anisomycin before training reduced memory retention from trained wasps (61 %) to 38 % but this reduction was not significant (GLM treatment ² 1 = 2.44, P = ) and the memory retention level of anisomycin fed wasps was significantly different from naïve control wasps (GLM treatment ² 1 = 6.97, P = ) (Fig. 10). In trained wasps which were fed with anisomycin and were cooled after 3 hours, memory retention was reduced significantly compared to trained wasps (GLM treatment ² 1 = 4.43, P = ) (Fig. 10).
23 4. Results 18 Choice for nasturtium (%) Cotesia glomerata Cotesia rubecula T TAC TC TA AC Treatments Fig. 10: C. glomerata and C. rubecula: Choice for nasturtium depending on the treatment. T = Training; TC = Training + Cooling; TA = Training + Anisomycin; TAC = Training + Anisomycin + Cooling; AC = Anisomycin + Cooling. For C. rubecula it was not possible to include a naïve control group due to the low availability of the wasps. Results from SMID et al. (submitted) show that naïve C. rubecula are attracted significantly more to cabbage than to nasturtium plants. The preference for cabbage can be compared with the preference observed in naïve C. glomerata, normally % of the wasps choose for cabbage. Trained C. rubecula showed high memory retention after 4 hours; 79 % of the wasps flew to nasturtium (P = ). Trained wasps which were cooled after 3 hours showed no significant difference in preference for nasturtium compared to trained wasps (GLM treatment ² 1 = 0.92, P = ) (Fig. 10). After one conditioning trial there is no significant difference in memory retention between C. glomerata and C. rubecula (GLM species ² 1 = 2.30, P = ). Cooling after three hours led to a significant difference in memory retention between both species (GLM species ² 1 = 10.66, P = ) (Fig. 10). Only results of Training and Training + Cooling of C. rubecula were included into the statistical analysis. Not enough replicates of Training + Anisomycin and Training + Anisomycin + Cooling and could be carried out. Data of these groups are added to figure 10 to show the tendency. Also feeding anisomycin seems not to have an influence on memory retention in C. rubecula.
24 4. Results 19 Table 2: C. glomerata: Choice for nasturtium 4 hours after one learning trial. AC = Anisomycin + Cooling; T = Training; TAC = Training + Anisomycin + Cooling; TC = Training + Cooling; TA = Training + Anisomycin. Treatment Hours after training Nasturtium Cabbage N N responding AC T TAC TC TA Table 3: C. rubecula: Choice for nasturtium 4 hours after one learning trial. T = Training; TC = Training + Cooling; TAC = Training + Anisomycin + Cooling; TA = Training + Anisomycin. Treatment Hours after training Nasturtium Cabbage N N responding T TC TAC TA
25 5. Discussion Discussion 5.1 Cotesia glomerata In this study C. glomerata was trained with one conditioning trial. Disrupting ASM with cooling, and feeding anisomycin before training (to block LTM), resulted in no memory retention after 4 hours; showing that no ARM is present like assumed in the hypothesis. If there is no ARM, ASM is directly consolidated into LTM in this species. This suggests that LTM induction prevents formation of ARM. Data from SMID et al. (submitted) suggested the same results after three spaced trials. Wasps which were fed with anisomycin before training showed no memory retention after 4 hours. 4 hour memory retention after one or three spaced conditioning trials seems to consist exclusively out of protein-synthesis dependent LTM. Unexpectedly, cooling 3 hours after oviposition affected memory retention after 4 hours, showing that ASM is at least present for 3 hours. This result was unexpected and is contradicting to data from WANG (unpublished). WANG (unpublished) also cooled trained wasps after 3 hours but no effect was seen after 4 hours. These results show that in C. glomerata 4 hour memory retention most likely does not consist exclusively out of LTM after one conditioning trial because ASM is still present after 3 hours. Actually planned experiments with cooling after 4 and 5 hours and testing after 5 and 6 hours respectively could not be carried out because of problems with the rearing (see 4.3). These experiments could give more clarity about the duration of ASM. Feeding anisomycin before training also reduced memory retention after 4 hours compared with trained wasps but this reduction was not significant. This result suggests that 4 hour memory retention contains a protein synthesis dependent component, but that this component has not yet been completely consolidated. This suggests that at 4 hours, there is a consolidation process going on from ASM into LTM. The fact that the combination of cooling and anisomycin has an additive effect on 4 h memory retention further suggests that ASM is consolidated into LTM directly. LTM formed after one conditioning trial seems to be different compared to LTM formed after three spaced trials. This is apparent when you look at the duration of LTM after different conditioning trials. In C. glomerata LTM lasts 2-3 days after one trial (GEERVLIET 1997) whereas LTM after three spaced trials lasts at least 5 days (SMID et al. submitted). How these differences can be explained on the neuronal level is unknown so far. It may explain that the effect of anisomycin on 4 hour memory retention in C. glomerata after 3 spaced trials (SMID et al. submitted) is more pronounced than after 1 single trial.
26 5. Discussion 21 In C. glomerata, ASM is consolidated directly into LTM. There is no long lasting memory phase like ARM coexisting with LTM after one conditioning trial. The situation in C. glomerata, where memory phases are mutually exclusive, is similar as described by ISABEL et al. (2004) for Drosophila mutants (ala mutants) in which LTM is deficient but with normal ASM and ARM. In wild-type Drosophila, single and massed conditioning trials induce ASM and ARM. A combined memory trace consisting from ASM, consolidating into ARM and LTM is induced by spaced training (MARGULIES et al. 2005). In ala flies ARM is fully functional and formed after single and massed learning. Training the flies with spaced learning experiences resulted in no memory retention after 5 hours. Normally ARM should be present 5 hours after spaced training, lasting for several days. ISABEL et al. (2004) concluded that ARM and LTM do not coexist in wild-type flies but instead ARM is blocked during LTM-specific training. It is assumed that LTM is formed after spaced conditioning and replaces ARM which is only formed after massed conditioning. LTM is formed in C. glomerata after single, massed and spaced conditioning trials and no ARM seems to be present in the wasp. Therefore also in C. glomerata formation of ARM seems to be prevented by LTM induction. 5.2 Cotesia rubecula C. rubecula was also trained with one conditioning trial but the process of memory consolidation was totally different. Contrary to C. glomerata cooling after 3 hours has no effect on memory retention in C. rubecula after one conditioning trial, showing that ASM is already absent. LTM is not formed in C. rubecula after one trial (SMID submitted) but only after several spaced experiences (Fig. 5a). Therefore ARM has to be present 4 hours after single training. WANG (unpublished) showed that ARM is present at least 8 hours after one trial and wanes between 8 and 24 hours. If C. rubecula receives three spaced learning trials LTM and ARM are formed in parallel and coexist. This was shown by SMID et al. (submitted) by inhibiting LTM with anisomycin; ARM was still present 48 hours after three spaced trials. Coexisting memory phases are described by e.g. TULLY et al. (1994) for Drosophila. After spaced learning trials, ARM and LTM are consolidated out of ASM and appear in parallel. The two memory phases are genetically dissected and dependent on different biochemical pathways. Evidence for this statement came out of the discovery that ARM is not sensitive to protein-synthesis inhibitors like cycloheximide (CXM). In contrast LTM is sensitive to protein-synthesis inhibitors and therefore requires gene expression.
27 5. Discussion 22 In C. glomerata and C rubecula we see clear differences in memory formation and memory retention. My results suggest, according to my hypothesis, that there is no ARM in C. glomerata. If there is no ARM, ASM is directly consolidated into LTM in this species. This suggests that LTM induction prevents formation of ARM in C. glomerata. In C. rubecula LTM is not formed after one conditioning trial and ARM is present after 4 hours. Comparing memory retention of both species we see that in C. glomerata ASM is present at least for 3 hours because cooling reduced memory retention significantly. In C. rubecula ASM lasts shorter; 3 hours after training only ARM is present, cooling had no effect on memory retention. In C. glomerata also feeding trained wasps with anisomycin affected memory retention after 4 hours but this difference was not significant compared to trained wasps. 4 hour memory retention seems to contain a protein synthesis dependent component, but this component has not yet been completely consolidated. C. glomerata can be compared with ala mutants in Drosophila described by ISABEL (2004). In ala mutants ARM is blocked during LTM-specific training. With C. glomerata we found a wild-type species where ARM is also blocked during LTM-specific training (single, massed and spaced). The situation in C. rubecula where ARM is formed after LTM-specific training (spaced) was described by TULLY et al. (1994) for wild-type Drosophila. LTM induction does not prevent formation of ARM in these species. Both models about the pathway of memory formation (Fig. 1, 2) described by TULLY et al. (1994) and ISABEL (2004) seem to have their validity if we look at the results of C. glomerata and C. rubecula. C. glomerata and C rubecula show clear differences in memory formation and memory retention. To find out why these closely related wasps differ so much in preference learning, a closer look has to be taken at memory formation in the context of natural behaviour. During my study unexpected variations in learning were observed in C. glomerata. First I would like to offer some suggestions why these variations might have occurred. 5.3 Variations in learning during the experiments During the first months of my study the rearing of C. glomerata did not work very well and in addition we found that the learning capability of the wasps was very low (see 4.1). Reasons for differences in learning and memory retention are hard to specify. A lot of factors could influence the learning behaviour of the wasps. TAUTZ et al. (2003) showed for honeybees that temperature during pupal development influences behavioural performances in adult bees. Temperature affects their dance communication and also their odour-learning abilities which are important for foraging.
28 5. Discussion 23 Several studies also focus on parasitic infections and its influence on behavioural traits in vertebrates and invertebrates. Parasitic infections often affect the function of the central nervous system (CNS); directly by destroying neurons or secreting neuroactive compounds or indirectly through substances which are produced by the host s immune system as a response to the infection (ADAMO 1997, 2002; THOMAS et al. 2005). This leads to an impairment of cognitive abilities which might be important for learning, memory and decision-making in animals (GEGEAR et al. 2006). In vertebrates it is well known that there are complex interactions between the immune system and the nervous system. In insects little is known but it is becoming clear that the cost of an immune response affects the survival of the insect itself, and also every day behaviour and memory formation (MALLON et al. 2003). GEGEAR et al. (2006) showed for the social bumble-bee Bombus impatiens that the gut parasite Crithidia bombi influences their ability to utilize floral information impairing foragers to monitor and acquire floral resources. The following food stress increases the mortality rate in infected bees and has therefore detrimental effects on the reproductive success of the colony. Although the parasite is found exclusively in the gut it can influence indirectly the CNS by the immune response of the bee. This study is one example how parasitic infections can disrupt ecologically important aspects of cognition in animals (e.g. KAVALIERS et al. 1995). MALLON et al. (2003) infected honeybees with a non-pathogenic immune elicitor (lipopolysaccharide) to simulate an immune response. The honeybees performed, after conditioning experiments, a reduced ability to associate an odour with a sugar reward, showing that the immune system seems to interfere with learning and/or memory formation. If both processes (immune response and memory formation) rely on the same intercellular communication system, using the same substances or messengers, there could be a trade-off in substrates. Specific substances like octopamine, which is important in immune response (WIESNER et al. 1996), might be used for the immune response instead for associative learning and/or memory formation (MALLON et al. 2003). Studies in vertebrates showed a way of signalling between the nervous system and the immune system (ADER et al. 1991, PUGH et al. 2001). Substances released by the immune response affect the hippocampus, and reduce memory consolidation (PUGH et al. 2001). In insects a similar signalling pathway could be possible, substances like eicosanoids are known to be involved in immune responses but also in learning processes (PIOMELLI 1994). Generally it is thought that immune responses must have substantial energetic costs and are therefore traded off against other costly functions (SHELDON & VERHULST 1996). For
29 5. Discussion 24 C. glomerata no parasitic infections are known so far but the high mortality rates and the low memory performances support the assumption that the individuals have been infected by a parasite or a pathogen. The differences in memory formation and consolidation between the wasps used by WANG (unpublished) and my population leads to the conclusion that the condition of the individuals has a huge impact on memory formation and consolidation. Learning is a costly process affecting animal fitness (DUKAS 1998), so maybe C. glomerata could not afford to spend energy on learning and memory formation. If the formation of LTM, which requires gene expression, is too costly, only ASM can be formed in C. glomerata because ARM seems to be absent. ASM and ARM do not require gene expression and are therefore also called a form of low cost memory (MERY & KAWECKI 2005). In C. glomerata the infection might have a higher impact on the wasp compared to other species because ARM, as a form of long lasting memory phase, is absent and only ASM can be formed. It would be interesting to know how long ASM lasts in individuals with a bad state of health. In these experiments ASM is still present after 3 hours whereas in the population of WANG (unpublished) LTM was exclusively present after 3 hours. Future studies should try to find out if infections play a role in variations of learning behaviour in Cotesia species. As a first step the mortality rate of the individuals should be correlated to the learning performance of the wasps. 5.4 Costs and benefits of learning To find out why C. glomerata and C. rubecula, two closely related species, differ in their learning behaviour you have to look at several factors. Learning and storing memory is a costly process which affects animal fitness. Energetic costs are involved to form and maintain memory and there are also ecological costs to learning; learning takes time and is vulnerable to mistakes (DUKAS 1998). Therefore innate behaviour might be favourable in some cases. Why should animals learn at all if it affects their fitness and why did learning evolve? It was thought that learning has developed because animals live in unpredictable environments (e.g. THORPE 1963, MACKINTOSH 1983). If the environmental situation would be completely stable, learning would be useless and innate behaviour favourable. STEPHENS (1993) rephrased a compromise between these two extreme situations; relatively little change is required for learning to evolve. For STEPHENS (1993) learning makes sense if the environment changes unpredictably between generations but is predictable within generations. For example if there are several generations of an insect in one year, which live in different seasons and exploit different food sources. In this case the environment is predictable within
30 5. Discussion 25 the generation and learning is favoured above innate behaviour. If the environment is very variable within a generation innate behaviour is beneficial. Animals are then confronted with so many different decisions that they would spend too much energy and time for changing behaviour. During their lifetime animals encounter a lot of new stimuli which they have to assess for its importance. Information is only valuable if it has the potential to change behaviour considerably (STEPHENS 1993). How do animals know which information is useful and reliable? When should they adapt to changes in their environment by changing their innate behaviour? One way of assessing the value of information takes place when information is repeated several times. Learning and the subsequent storage of memory in short- or long-term memory is always dependent on the frequency of the stimuli but most of all on the time between the repetitions. Single experiences lead to the storage of memory in STM and ARM whereas only spaced experiences lead to LTM. Still this is not true for every species as we can see e.g. in C. glomerata. C. glomerata does not need confirmative experiences (spaced learning) to form LTM showing the value and the reliability of the information. Learning and memory formation is a process, which proceeds differently in various species. In some species a considerable amount of time is needed for consolidation of memory phases but this time is not required for the underlying neuronal processes; these can react and change very quickly (FREY & MORRIS 1997). Instead, memory formation is adapted to the specific demands and constraints of every single species in its natural environment. A more ecological view of learning contains that memory formation has to be seen in the context of natural behaviour and the specific requirements to which animals are exposed (MENZEL 1999). Learning always means to find a balance between costs and benefits. E.g. learning fast can be a disadvantage by storing wrong information but it might also be advantageous if the information is reliable. Referring to memory formation this means that new memories have to be evaluated on the basis of their relevance. Forming new memory is a costly process indicating that new memories should only be consolidated when they are reliable and of importance. 5.5 Cotesia glomerata and Cotesia rubecula why are they so different in learning? Looking at the natural behaviour, differences in preference learning might be an adaptation to the specific differences in host-finding behaviour between the two wasp species. If we look at the generalist C. glomerata we see a gregarious species which lays in average 20 eggs mainly
31 5. Discussion 26 in the caterpillars of P. brassicae. P. brassicae is also gregarious and a generalist, it uses different host plants (cruciferous species) to lay its eggs on. For the wasp this causes a problem in its searching behaviour. On all cruciferous plants hosts could be present and therefore the host plants have a low predictive value for the wasps. If there are a lot of potential plants which might offer hosts, a lot of time and energy is wasted for host searching. GEERVLIET et al. (1994) showed that C. glomerata is attracted innately by cabbage plants with caterpillars feeding on it but still the wasp encounters hosts on plants different than cabbage. If this happens associative learning takes place. The wasp associates the odour of the new encountered plant with the presence of suitable hosts and this process causes temporal specialization (DUKAS 1998). Specialization is thought to be a way out of inefficient searching behaviour. If C. glomerata encounters a host on a specific plant species this is reliable information for the wasp. It can be sure to find other hosts on the surrounding plants of the same species due to the egg laying performance of its host P. brassicae. P. brassicae uses different host plants but if it lays its egg clusters it chooses a stand of plants of the same species to ensure the food supply for its offspring. The caterpillars are conditioned on the first species they feed on and need to find more plants of the same species. Therefore the wasp will find enough hosts for its own offspring if it orientates itself towards the odour of that plant species. For this reason C. glomerata not only changes its behaviour after one experience but it also stores this memory directly in LTM. For the generalist C. glomerata, parasitizing a generalist host, learning is more valuable than retaining to its innate behaviour. Through learning it benefits from the advantages of specialization (SMID 2006). For the solitary specialist C. rubecula, finding a host on a certain plant species is not reliable information. Its host P. rapae lays only one egg per plant and spreads its eggs widely in its habitat on different plant species (ROOT & KAREIVA 1984). For C. rubecula the presence of one caterpillar does not indicate the presence of other suitable hosts on other plants of the same species. It would be maladaptive for the wasp to change its innate behaviour and start searching for this plant species. C. rubecula is able to associate the odour of a plant with a suitable host but this memory is stored in ASM and ARM and wanes within one day. The wasp does not change its innate behaviour (BLEEKER 2006, SMID submitted). In C. rubecula, memory formation works as a filter keeping out unreliable information. Only after several verifying experiences information is stored in LTM. The wasp is called a slow learner which might sound negative but in the case of C. rubecula learning slowly is the way to successful oviposition.
32 5. Discussion 27 Comparing the memory formation in C. glomerata and C. rubecula we see a quick and a slow learner but both ways of learning are successful in their natural environment. My results show that it seems not to be important for C. glomerata to have an additional long lasting memory phase like ARM. The information the wasp gets is so reliable that LTM is directly consolidated out of ASM. LTM requires new gene expression but although the formation is connected to high energetic costs, these costs are worth to spend. The benefits from forming LTM outweigh the disadvantage of spending energy for the formation. Maybe the wasp can also save some energy by leaving out ARM. In C. rubecula the situation is different, the predictability of its environment is very low and innate behaviour is favoured. Still associations are formed after one learning experience but these are stored in ASM and ARM. ASM and ARM are a form of low cost memories, demanding less energetic costs than the formation of LTM (MERY & KAWECKI 2005). Like this C. rubecula found a successful way to benefit on the one hand of innate behaviour but also from learning without wasting too much energy. Closely related parasitic wasps which differ in their learning behaviour are useful model organisms to study more about learning and memory formation. Results of my study offer some interesting findings concerning differences in memory consolidation in C. glomerata and C. rubecula. Still a lot of questions remain and future studies could focus on these. Together with a more ecological approach on learning with behavioural tests in the wind tunnel, biomolecular techniques can help to find out more about underlying mechanisms of memory formation. Working six months intensively with parasitic wasps was a fascinating experience for me. Although I read a lot about learning in insects it is still different to experience it by yourself. Wasps which were put for some seconds on a nasturtium leaf with suitable hosts on it remembered this odour 4 hours later! Watching them in the wind tunnel, seeing them move their antennae in agitation, and flying directly to the new learned plant odour fascinated me every time again.
33 6. References References ADAMO, S.A.: How parasites alter the behaviour of their insect hosts. In: N. Beckage (ed.): Parasites and pathogens. Effects on host hormone and behaviour. Chapman & Hall, , ADAMO, S.A.: Modulating the modulators: parasites, neuromodulators and host behavioural change. Brain Behaviour and Evolution 60, , ADER, R., D.L. FELTEN, N. COHEN: Psychoneuroimmunologie. San Diego, CA: Academic, Bleeker, M.: Associative learning in two closely related parasitoid wasps: a neuroecological approach. Thesis Wageningen University, CHEW, F.S.: Food plant preferences of Pieris caterpillars. Oecologia 46, , COLLATZ, J., C. MÜLLER, J.L.M. STEIDLE: Protein synthesis-dependent long-term memory induced by one single associative training trial in the parasitic wasp Lariophagus distinguendus. Learning & Memory 13, , DUKAS, R.: Evolutionary ecology of learning. In : R. Dukas (ed.): Cognitive ecology: The evolutionary ecology of information processing and decision making. University of Chicago press, Chicago, London, , FELTWELL, J.: Large white butterfly. The biology, biochemistry and physiology of Pieris brassicae (Linnaeus). Dr. W. Junk Publishers, The Hague, Boston, London, FREY, U., R.G.M. MORRIS: Synaptic tagging and long-term potentation. Nature 385, FULTON, D., I. KEMENES, R.J. ANDREW, P.R. BENJAMIN: A single time-window for protein synthesis dependent long-term memory formation after one-trial appetitive conditioning. European Journal of Neurosccience 21, , GEERVLIET, J.B.F., L.E.M. VET, M. DICKE: Volatiles from damaged plants as major cues in long-range host-searching by the specialist parasitoid Cotesia rubecula. Entomologia Experimentalis et Applicata 73, , GEERVLIET, J.B.F., L.E.M. VET, M. DICKE: Innate responses of the parasitoids Cotesia glomerata and C. rubecula (Hymenpotera: Braconidae) to volatiles from different plantherbivore complexes. Journal of Insect Behaviour 9, , GEERVLIET, J.B.F.: Infochemical use by insect parasitoids in a tritrophic context: comparison of a generalist and a specialist. Thesis. Laboratory of Entomology, Wageningen University, 1997.
34 6. References 29 GEERVLIET, J.B.F., A.I. VREUGDENHIL, M. DICKE, L.E.M. VET: Learning to discriminate between infochemicals form different plant-host complexes by the parasitoids Cotesia glomerata and C. rubecula. Entomologia Experimentalis et Applicata 86, , GEERVLIET, J.B.F., M.S.W. VEDEL, H. SNELLEN, J. SCHAUB. M. DICKE, L.E.M. VET: Coexistence and niche sgragation by field populations of the parasitoids Cotesia glomerata and C. rubecula in The Netherlands: predicting field performance from laboratory data. Oecologia 124, 55-63, 2000a. GEERVLIET, J.B.F., S. ARIENS, A.I. VREUGDENHIL, M. DICKE: Volatiles form damaged plants act as major cues in long-range host-searching by the parasitoid Cotesia rubecula. Entomologia Experimentalis et Applicata 73, , 2000b. GEGEAR, R.J., M.C. OTTERSTATTER, J.D. THOMSON: Bumble-bee foragers infected by a gut parasite have an impaired ability to utilize floral information. Proceedings of the Royal Society B 273, , IGAZ, L.M., M.R.M. VIANNA, J.H. MEDINA, I. IZQUIERDO: Two Time Periods of Hippocampal mrna Synthesis Are Required for Memory Consolidation of Fear-Motivated Learning. The Journal of Neuroscience 22, , ISABEL, G., A. PASCUAL, T. PREAT: Exclusive Consolidated Memory Phases in Drosophila. Science 304, , JAMES, W.: The Principles of Psychology. Holt, New York, KAVALIERS, M.,D.D. COLWELL, L.A.M.GALEA: Parasitic infecion impairs spatial learning in mice. Animal Behaviour 50, , LE MASURIER, A.D.: Costs and benefits of egg clustering in Pieris brassicae. Journal of Animal Ecology 63, LUTZ, J.: Learning and Memory. Waveland Press, Long Grove, MACKINTOSH, N.J.: General principles of learning. In: T. Halliday, P.J.B. Later (eds.): Animal Behaviour. Vol. 3: Genes, Development and Learning. W.H. Freeman, New York, , MALLON, E.B., A. BROCKMANN, P. SCHMID-HEMPEL: Immune response inhibits associative learning in insects. Proceedings of the Royal Society B 270, , MARGULIES, C., T. TULLY, J. DUBNAU: Deconstructing Memory in Drosophila. Current Biology 15, R700-R713, MENZEL, R.: Behavioural access to short-term memory in bees. Nature 281, , MENZEL, R.: Memory dynamics in the honeybee. Journal of Comparative Physiology 185, , 1999.
35 6. References 30 MERY, F., T.J. KAWECKI: A Cost of Long-Term Memory in Drosophila. Science 308, 1148, PAVLOV, I.P.: Conditioned reflexes (F.V. Anrep. Trans.). Oxford University Press, Oxford, PIOMELLI, D.: Eicosanoids in synaptic transmission. Critical Reviews in Neurobiology 8, 65-83, PUGH, C.R., M. FLESHNER, L.R. WATKINS, S.F. MAIER, J.W. RUDY: The immune system and memory consolidation: a role for the cytokine IL-1. Neuroscience and Biobehavioral Reviews 25, 29-41, ROOT, R.B., P.M. KAREIVA: The search for resources by cabbage butterflies (Pieris rapae) Ecological consequences and adaptive significance of markovian movements in a patchy environment. Ecology 65, , ROSENZWEIG, M.R., E.L. BENNET, P.J. COLOMBO, D.W. LEE, P.A. SERRANO: Short-term, intermediate-term, and long-term memories. Behavioural Brain Research 57, , ROTHSCHILD, M.: Speculations concerning the Large White Butterfly (Pieris brassicae L.): do females assess the number of suitable host plants present? In: Chapman, R.F., E.A. Bernays, J.G.J. Stoffolano (eds.): Perspectives in chemoreception and behavior. Springer Verlag, New York, , SHELDON, B.C., S. VERHULST: Ecological immunology: costly parasite defences and tradeoffs in evolutionary ecology. TREE 11, , SKINNER, B.F.: The behaviour of organisms. Appleton-Century, New York, SMID, H.: Variation in learning of herbivory-induced plant odours by parasitic wasps: From brain to behaviour. In: Dicke, M.; W. Takken (eds.): Proceedings of the Frontis Workshop on Chemical Ecology: From Gene to Ecosystem. Springer, Doordrecht, , Smid, H., G. Wang, T. Bukovinszky, J.L.M. Steidle, M.A.K. Bleeker, J.J.A. van Loon, L.E.M. Vet: Species-specific acquisition and consolidation of long-term memory in parasitic wasps. (Submitted). STEPHENS, D.W.: Learning and Behavioral Ecology: Incomplete Information and Environmental Predictability. In: Papaj, D.R., A.C. Lewis (eds): Insect Learning: Ecological and Evolutionary Perspectives. Chapman & Hall, New York, London, , 1993.
36 6. References 31 TAUTZ, J., S. MAIER, C. GROH, W. RÖSSLER, A. BROCKMANN: Behavioral performance in adult honeybees is influenced by the temperature experienced during their pupal development. Proceedings of the National Academy of Scienes 100, , THOMAS, F., S. ADAMO, J. MOORE: Parasitic manipulation: where are we and where should we go? Behavioural Processes 68, , THORNDICKE, E.L.: Animal intelligence: An experimental study of the associative processes in animals. Psychological Review Monograph Supplement 2, 1-109, THORPE, W.H.: Learning and Instinct in Animals. Methuen, London, TULLY, T., T. PREAT, S.C. BOYNTON, M. DEL VECCHIO: Genetic Dissection of Consolidation Memory in Drosophila. Cell 79, 35-47, TUMLINSON, J.H., W.J. LEWIS, L.E.M. VET: Parasitic wasps, chemically guided intelligent foragers. Scientific American 268, , VET, L.E.M., M. DICKE: Ecology of infochemical use by natural enemies in a tritrophic context. Annual Review of Entomology 37, , VOS, M., L.E.M. VET: Geographic variation in host acceptance by an insect parasitoid: genotype versus experience. Evolutionary Ecology Research 6, , WIESNER, A., D. WITTWER, P. GOTZ: A small phagocytosis stimulating factor is released by and acts on phagocytosing Galleria mellonella haemocytes in vivo. Journal of Insect Physiology 24, , YIN, J.C., J.S. WALLACH, M. DEL VECCHIO, E.L. WILDER, H. ZHOU, W.G. QUINN, T. TULLY: Induction of a dominant negative CREB transgene specifically blocks long-term memory in Drosophila. Cell 79, 49-59, 1994.
37 1. Introduction 32 Appendix: The role of larval mobility in the transition from solitary to gregarious development in Cotesia glomerata and Cotesia rubecula 1. Introduction 1.1 Solitary and gregarious development In parasitic wasps two different ways of developing are known, solitary and gregarious. In solitary species only one egg can develop in one host. If more than one egg is laid into the host only one of the larvae survives (e.g. SALT 1961) because of physical attack, physiological suppression or resource competition (GODFRAY 1994). Despite the fact that adult females are able to destroy larvae or eggs of other females (ovicide) with their ovipositor (GODFRAY 1994) or by injecting venoms (HÅGVAR 1988, MACKAUER 1990), mostly the behaviour of the immature parasites determines their own survival. First instar solitary larvae have strong mandibles and they search actively for conspecific competitors and individuals of other species. The death of rivals is not necessarily caused by fighting itself but wounding makes sure that invaders are recognized by the host s immune system and die (GODFRAY 1994). Older larvae are able to eliminate younger larvae by physiological suppression. They are thought to excrete chemicals that inhibit or kill competitors (TIMBERLAKE 1910, 1912; MACKAUER 1986). Also the competition for limited resources is a possibility to fight against opponents. The host can be already consumed before a competitor hatches (FISKE & THOMPSON 1909) or e.g. the oxygen content in the host s haemolymph can be reduced (asphyxation) and younger more sensible larvae or eggs perish (FISHER 1961, 1963). For the young larvae which have fighting capabilities mobility plays the key role for finding and killing competitors to exploit the whole resources of the host. Solitary development seems to be the ancestral condition in parasitoid Hymenoptera. Still gregariousness has evolved at least 43 times in 26 different families of Hymenoptera (ROSENHEIM 1993, MAYHEW 1998) and is stable in these cases. In gregarious species several eggs are laid in one host and the larvae develop successfully together (ODE & ROSENHEIM
38 1. Introduction ). Gregarious development has the advantage that the adult insect can exploit its hosts better, the larger the host the more eggs are laid into it. In evolutionary biology the transition of solitary to gregarious development has been of interest for a long time (MOCK & PARKER 1997). Most research has been done on birds (GODFRAY & HARPER 1990) and insect parasitoids (GODFRAY 1987, MAYHEW 1998). The question is how the transition from solitary and siblicidal to gregarious and nonsiblicidal individuals took place. If the individuals are siblicidal there should be a selection towards solitary individuals, laying only one egg in one host. How can nonsiblicidal behaviour enter under these conditions in siblicidal populations? HARVEY and PARTRIDGE (1987) called this situation an evolutionary black hole because this evolution seems to be irreversible. Normally nonsiblicidal brood mates would always be killed by their siblicidal mates (MAYHEW et al. 1998). In addition why should solitary species start to lay more than one egg in a host? Several hypotheses are proposed until now. Beside the suggestion, that this could happen by accident (GODFRAY 1987), it is proposed that parasitoids may lay additional eggs to ensure the successful development of their offspring (PARKER & COURTNEY 1984, GODFRAY 1987). If the first egg is not viable, the second egg has a chance to develop and survive (GODFRAY 1994). Additional eggs are also an advantage when the risk of multiparasitism (interspecific superparasitism) is high. Two or more eggs of the same species enhance the chance to be the winner in competitive interactions between the developing larvae of other species (PARKER & MOCK 1987, VAN ALPHEN & VISSER 1990). Also the discouragement of conspecifics is seen as a possibility. If conspecific adults are able to determine the number of eggs being present in a host they might be prevented from parasitizing if there are enough hosts in the patch. For the offspring the chance of surviving decreases with the number of eggs already present in the host. Therefore additional eggs are a good option to ensure the survival of the own offspring (VAN ALPHEN & VISSER 1990). Eggs of the parasitoid wasps also have to deal with the immune system of its hosts, haemocytes of the host can form multicellular capsules which surround the eggs and the larvae are not able to hatch any more (BAUER 1999, VAN ALPHEN & VISSER 1990). In this case superparasitism or self-superparasitism can be advantageous because the encapsulation of the first egg in the host might have exhausted the haemocyte supply of the host (VAN ALPHEN & VISSER 1990). Normally, brood reduction by fighting larvae seems to be advantageous because there is a higher amount of host resources left over for the winner. Still the laying of additional eggs can
39 1. Introduction 34 be of advantage when the host can support the development of more than one individual and there is a fitness advantage for several individuals developing together. Improvement of host quality through host shift or change in host size might be a reason why gregarious development has been evolved in some cases (GODFRAY 1994). Still the question remains how the transition from solitary to gregarious development took place. 1.2 Transition from solitary to gregarious development One assumption of the successfully transition to gregarious development has been that the brood mates tolerate each other through a loss of aggression while the larvae retain mobile (MAYHEW & VANALPHEN 1999). Still, the invasion of an aggressive individual would disturb the transition to tolerant behaviour. In Argochrysis armilla the spread of tolerant behaviour seems to be supported by single sex broods. If a tolerant gene enters a population it has a bigger chance to be spread if the brood is closely related to each other (PEXTON & MAYHEW 2001). BOIVIN and BAAREN (2000) assume that instead of a loss of aggression (and retaining mobility), the larvae retain aggressive behaviour but have a reduced mobility. They compared the mobility of Anaphes listronoti, a gregarious egg parasitoid of Listronotus oregonensis, with the solitary A. victus (Fig. 1). Larvae of A. victus were more mobile (twisting and folding) than larvae of A. listronoti. Both first instar larvae have highly developed mandibles and are able to defend themselves against siblicidal brood mates. The assumption is that the brood mates of gregarious species don t search actively for conspecific larvae and they develop beyond the aggressive first larval instar, without encountering each other. If they encounter a siblicideous larvae invading the population they have equal chances to survive because of their developed mandibles. To investigate if this phenomenon also occurs in other species the mobility of larvae of the gregarious wasp Cotesia glomerata (L.) and the solitary Cotesia rubecula (Marshall) (Hymenoptera: Braconidae) will be compared in Pieris caterpillars. 1.3 Cotesia glomerata and Cotesia rubecula The parasitic wasps C. glomerata and C. rubecula lay their eggs in young larvae of Pieris butterflies. C. rubecula is classified as solitary and prefers the larvae of the solitary small cabbage white Piries rapae (L.) (Lepidoptera: Piridae) as a host. Only one egg is laid in a single caterpillar (GEERVLIET et al. 2000, SMID 2006). If more than one egg is laid into a host only one larva will survive (HARVEY, unpublished).
40 2. Materials and methods 35 Fig. 1: Mean numbers (± SEM) of twisting (lateral torsion) and folding (vertical bending) observed for 5 min in mymariform larvae of the solitary A. victus and the gregarious A. listronoti (BOIVIN & BAAREN 2000). In contrast, C. glomerata is classified as gregarious, i.e. the wasp lays up to 20 eggs in a single host. Although C. glomerata is regarded as a generalist, the wasp prefers to lay its eggs in the larvae of the gregarious large cabbage white Pieris brassicae (L.) (Lepidoptera: Piridae) (VOS & VET 2004). The larvae develop successfully together without killing each other, i.e. they are regarded as nonsiblicidal (PEXTON et al. 2003). To see if the loss of mobility plays also an important role in these two species for the transition from solitary to gregarious development a similar approach as in the experiments of BOIVIN and BAAREN (2000) will be done. 2. Materials and methods 2.1 Experimental setup Insects and plants were reared as explained before. Freshly hatched larvae of P. brassicae and P. rapae were transferred to cabbage plants and were allowed to feed for 24 hours. After 24 hours the larvae were parasitized by C. glomerata and C. rubecula respectively. The wasp larvae hatched after 5 days and the Pieris caterpillars were dissected after 5, 6, 7 and 8 days. Larvae of the wasps were observed with a microscope and filmed in physiological saline for a period of 5 min. 15 larvae of each species were analysed at the specific days.
41 3. Results 36 To compare the morphology of the mouthparts, pictures of the head of both species were taken with a JEOL JSM 6300F field emission scanning electron microscope (SEM). Pieris caterpillars were parasitized as described above and the wasp larvae were dissected out of the caterpillars after 5, 6, 7 and 8 days. The larvae were transferred to 100 % ethanol, critical point dried and sputtered with 10 nm platinum for observation in the microscope. Unfortunately larvae of C. glomerata at day 5 and 6 were so small that they were washed through the gauze during the drying process. 2.2 Statistical analysis Generalized linear modelling (GLM) procedures using procedure GENMOD in SAS version 8.02 (Proc GENMOD, SAS Inc, Chicago, IL, U.S.A.) were used for data with a binominal distribution of error variance and a logit-link function. When main effects or their interactions were found significantly different, further separation of the treatment levels was carried out by acquiring specific contrasts for particular comparisons, which are given in the text. Data collected on different experimental days were considered as replicates. We tested whether time (4 levels: 5, 6, 7 or 8 days) and species (2 levels: C. glomerata and C. rubecula) had an effect on twisting in Cotesia larvae (GLM treatment ² 6 = 41.42, P < ). 3. Results Observing the larvae of both species, the lateral bending were counted. Five days after oviposition early first instar larvae of C. rubecula twisted significantly more than larvae of C. glomerata (GLM, P = ) (Fig. 2). The larvae bended with quick and dynamic movements, enabling them to move through the body of the host. Movements of C. glomerata were more slowly and consistent with breaks in between. From day 6 on there was no difference in movements any more between both species. In C. rubecula a rapid decline of movements from 5 to 8 days after oviposition is shown (GLM, P < ) whereas in C. glomerata the twisting remain at the same level (GLM, P = ).


42 3. Results 37 twistings in 5 minutes Cotesia glomerata Cotesia rubecula days after oviposition Fig. 2: Mean numbers of lateral bending of the solitary C. rubecula and the gregarious C. glomerata within 5 minutes, 5 to 8 days after oviposition. Table 1: Mean numbers of bending in C. glomerata and C. rubecula, 5, 6, 7 and 8 days after oviposition. Species 5 days 6 days 7 days 8 days C. glomerata C. rubecula Fig. 3: Larvae of C. glomerata and C. rubecula 6 days after oviposition. Bar = 300 m.












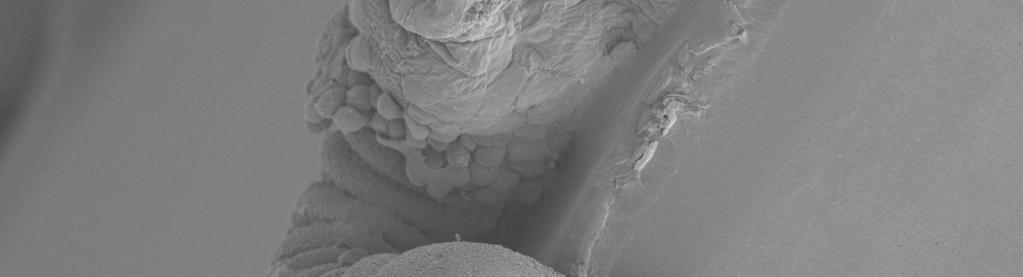




43 3. Results A 38 B mp C D mp md E lp F Fig. 4: Larvae of C. glomerata (A, B) and C. rubecula (C F). A, B: Mouthparts and ventral view of C. glomerata on day 7 and 8. C: Lateral view of C. rubecula on day 5. D: Ventral-frontal view of C. rubecula and mouthparts on day 5. E, F: Dorsal and ventral view of the head of C. rubecula on day 8. Note the absence of mandibles and labial palpes in C. glomerata. mp = maxillary palpus; md = mandible; lp = labial palpus.
WAGENINGEN UNIVERSITY LABORATORY OF ENTOMOLOGY
 WAGENINGEN UNIVERSITY LABORATORY OF ENTOMOLOGY Host discrimination by Cotesia glomerata (L.) (Hymenoptera: Braconidae), a parasitoid of Pieris brassicae (L.) (Lepidoptera: Pieridae), as affected by experience
WAGENINGEN UNIVERSITY LABORATORY OF ENTOMOLOGY Host discrimination by Cotesia glomerata (L.) (Hymenoptera: Braconidae), a parasitoid of Pieris brassicae (L.) (Lepidoptera: Pieridae), as affected by experience
Mr.checker Wikipedia Commons. Louise Docker from sydney, Australia, Wikimedia Commons. Angelique Paulk
 Mr.checker Wikipedia Commons Louise Docker from sydney, Australia, Wikimedia Commons Angelique Paulk Objectives How does learning and memory operate at the behavioral level? How can you train an insect?
Mr.checker Wikipedia Commons Louise Docker from sydney, Australia, Wikimedia Commons Angelique Paulk Objectives How does learning and memory operate at the behavioral level? How can you train an insect?
Grade 7 Lesson Instructions Friend or Foe? Preparation: Background information: Activity:
 Instructions Friend or Foe? You can use monarchs to teach about many things! Stone Mountain Memorial Association (SMMA) uses the monarch butterfly to help students apply their knowledge in other contexts
Instructions Friend or Foe? You can use monarchs to teach about many things! Stone Mountain Memorial Association (SMMA) uses the monarch butterfly to help students apply their knowledge in other contexts
Bee Colony Activities Throughout The Year
 Bee Colony Activities Throughout The Year Written by Khalil Hamdan Apeldoorn The Netherlands A honeybee gathering nectar from a flower. Photo source: forestwander.com Bee collecting pollen. Photo source:
Bee Colony Activities Throughout The Year Written by Khalil Hamdan Apeldoorn The Netherlands A honeybee gathering nectar from a flower. Photo source: forestwander.com Bee collecting pollen. Photo source:
28 3 Insects Slide 1 of 44
 1 of 44 Class Insecta contains more species than any other group of animals. 2 of 44 What Is an Insect? What Is an Insect? Insects have a body divided into three parts head, thorax, and abdomen. Three
1 of 44 Class Insecta contains more species than any other group of animals. 2 of 44 What Is an Insect? What Is an Insect? Insects have a body divided into three parts head, thorax, and abdomen. Three
EUROPEAN JOURNAL OF ENTOMOLOGY
 EUROPEAN JOURNAL OF ENTOMOLOGY ISSN (online): 1802-8829 http://www.eje.cz Eur. J. Entomol. 115: 25 29, 2018 ORIGINAL ARTICLE Host-plant leaf-surface preferences of young caterpillars of three species of
EUROPEAN JOURNAL OF ENTOMOLOGY ISSN (online): 1802-8829 http://www.eje.cz Eur. J. Entomol. 115: 25 29, 2018 ORIGINAL ARTICLE Host-plant leaf-surface preferences of young caterpillars of three species of
Natural variation in learning rate and memory dynamics in parasitoid wasps: opportunities for converging ecology and neuroscience
 Natural variation in learning rate and memory dynamics in parasitoid wasps: opportunities for converging ecology and neuroscience Katja M. Hoedjes, H. Marjolein Kruidhof, Martinus E. Huigens, Marcel Dicke,
Natural variation in learning rate and memory dynamics in parasitoid wasps: opportunities for converging ecology and neuroscience Katja M. Hoedjes, H. Marjolein Kruidhof, Martinus E. Huigens, Marcel Dicke,
Community Involvement in Research Monitoring Pollinator Populations using Public Participation in Scientific Research
 Overview Community Involvement in Research Monitoring Pollinator Populations using Public Participation in Scientific Research Public Participation in Scientific Research (PPSR) is a concept adopted by
Overview Community Involvement in Research Monitoring Pollinator Populations using Public Participation in Scientific Research Public Participation in Scientific Research (PPSR) is a concept adopted by
Biology Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod. Introduction
 1 Biology 317 - Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod Introduction The determination of how natural selection acts in contemporary populations constitutes
1 Biology 317 - Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod Introduction The determination of how natural selection acts in contemporary populations constitutes
Biology 322 Fall 2009 Wasp Genetics: Genetic Heterogeneity and Complementation Revisted
 Biology 322 Fall 2009 Wasp Genetics: Genetic Heterogeneity and Complementation Revisted Required Reading: Deaf by Design Nature 431: 894-896 October 21, 2004 http://fire.biol.wwu.edu/trent/trent/naturedeafdesign.pdf
Biology 322 Fall 2009 Wasp Genetics: Genetic Heterogeneity and Complementation Revisted Required Reading: Deaf by Design Nature 431: 894-896 October 21, 2004 http://fire.biol.wwu.edu/trent/trent/naturedeafdesign.pdf
Population Ecology. Study of populations in relation to the environment. Increase population size= endangered species
 Population Basics Population Ecology Study of populations in relation to the environment Purpose: Increase population size= endangered species Decrease population size = pests, invasive species Maintain
Population Basics Population Ecology Study of populations in relation to the environment Purpose: Increase population size= endangered species Decrease population size = pests, invasive species Maintain
Peter Dutton. 28 August Halesworth& District
 Peter Dutton 28 August 2014 Halesworth& District Bees Hymenoptera - 150,000+ species Sawflies, Wasps, Bees, Ants, Bees - 20,000+ species Most solitary, a few social Honey Bee - The only one which produces
Peter Dutton 28 August 2014 Halesworth& District Bees Hymenoptera - 150,000+ species Sawflies, Wasps, Bees, Ants, Bees - 20,000+ species Most solitary, a few social Honey Bee - The only one which produces
Whitney Cranshaw Colorado State University
 Natural and Biological Controls of Shade Tree Insect Pests Whitney Cranshaw Colorado State University Natural Controls Natural Enemies Abiotic (Weather) Controls Topographic Limitations Temperature Extremes
Natural and Biological Controls of Shade Tree Insect Pests Whitney Cranshaw Colorado State University Natural Controls Natural Enemies Abiotic (Weather) Controls Topographic Limitations Temperature Extremes
Plant Stimuli pp Topic 3: Plant Behaviour Ch. 39. Plant Behavioural Responses. Plant Hormones. Plant Hormones pp
 Topic 3: Plant Behaviour Ch. 39 Plants exist in environments that are constantly changing. Like animals, plants must be able to detect and react to stimuli in the environment. Unlike animals, plants can
Topic 3: Plant Behaviour Ch. 39 Plants exist in environments that are constantly changing. Like animals, plants must be able to detect and react to stimuli in the environment. Unlike animals, plants can
What is behavior? What questions can we ask? Why study behavior? Evolutionary perspective. Innate behaviors 4/8/2016.
 What is behavior? Animal Behavior Behavior everything an animal does & how it does it response to stimuli in its environment Innate (instinct) inherited automatic & consistent learned ability to learn
What is behavior? Animal Behavior Behavior everything an animal does & how it does it response to stimuli in its environment Innate (instinct) inherited automatic & consistent learned ability to learn
Brown Hairstreak (Early Stages)
") 01 February 2014 Vince Massimo Citation: Massimo, V. (2014). Brown Hairstreak (Early Stages) [Online]. Available from http://www.dispar.org/reference.php?id=18 [Accessed February 1, 2014]. Brown Hairstreak
01 February 2014 Vince Massimo Citation: Massimo, V. (2014). Brown Hairstreak (Early Stages) [Online]. Available from http://www.dispar.org/reference.php?id=18 [Accessed February 1, 2014]. Brown Hairstreak
Eusocial species. Eusociality. Phylogeny showing only eusociality Eusocial insects. Eusociality: Cooperation to the extreme
 Eusociality: Cooperation to the extreme Groups form colonies with reproductive and worker castes. Eusociality has evolved most often in insects: Ants Eusocial species Honeybees Termites Wasps Phylogeny
Eusociality: Cooperation to the extreme Groups form colonies with reproductive and worker castes. Eusociality has evolved most often in insects: Ants Eusocial species Honeybees Termites Wasps Phylogeny
Pollinator Activity #1: How to Raise a Butterfly
 How to Raise a Butterfly How to Raise a Butterfly A Conversation Where do you most often see butterflies? What are they doing when you see them? Have you ever seen a butterfly in another form? They have
How to Raise a Butterfly How to Raise a Butterfly A Conversation Where do you most often see butterflies? What are they doing when you see them? Have you ever seen a butterfly in another form? They have
Approximate Pacing for First Grade Insects and Plants Unit
 Approximate Pacing for First Grade Insects and Plants Unit p.1 = Part 1 p.2 = Part 2 p.3 = Part 3 The schedule for this unit is almost COMPLETELY dependent on what the living organisms are doing and where
Approximate Pacing for First Grade Insects and Plants Unit p.1 = Part 1 p.2 = Part 2 p.3 = Part 3 The schedule for this unit is almost COMPLETELY dependent on what the living organisms are doing and where
Experimental Design, Data, and Data Summary
 Chapter Six Experimental Design, Data, and Data Summary Tests of Hypotheses Because science advances by tests of hypotheses, scientists spend much of their time devising ways to test hypotheses. There
Chapter Six Experimental Design, Data, and Data Summary Tests of Hypotheses Because science advances by tests of hypotheses, scientists spend much of their time devising ways to test hypotheses. There
MEXICAN BROMELIAD WEEVIL REPORT 12 JANUARY 2013
 1 MEXICAN BROMELIAD WEEVIL REPORT 12 JANUARY 2013 Ronald D. Cave 1, Teresa M. Cooper 1, and J. Howard Frank 2 1 Indian River Research & Education Center, UF, Ft. Pierce, FL 2 Entomology & Nematology Department,
1 MEXICAN BROMELIAD WEEVIL REPORT 12 JANUARY 2013 Ronald D. Cave 1, Teresa M. Cooper 1, and J. Howard Frank 2 1 Indian River Research & Education Center, UF, Ft. Pierce, FL 2 Entomology & Nematology Department,
Bio 1M: The evolution of apes. 1 Example. 2 Patterns of evolution. Similarities and differences. History
 Bio 1M: The evolution of apes 1 Example Humans are an example of a biological species that has evolved Possibly of interest, since many of your friends are probably humans Humans seem unique: How do they
Bio 1M: The evolution of apes 1 Example Humans are an example of a biological species that has evolved Possibly of interest, since many of your friends are probably humans Humans seem unique: How do they
Report No Yehua Li Msc Plant Science Thesis ENT June 2010-January 2011 Supervosors: Roxina Soler Ana Pineda Examiner: Marcel Dicke
 Plant-mediated inter-guild interactions: influence of the phloem feeder Brevicoryne brassicae on the host preference of the leaf chewer Pieris brassicae, and its parasitoid Cotesia glomerata Report No.
Plant-mediated inter-guild interactions: influence of the phloem feeder Brevicoryne brassicae on the host preference of the leaf chewer Pieris brassicae, and its parasitoid Cotesia glomerata Report No.
Labdien! こんにちは Kon nichi wa. I am Naota Ohsaki, PhD of National Yamagata University, Japan.
 Labdien! こんにちは Kon nichi wa I am Naota Ohsaki, PhD of National Yamagata University, Japan. I am a professor of Yamagata University in charge of International Exchange. My mission is to stay at several
Labdien! こんにちは Kon nichi wa I am Naota Ohsaki, PhD of National Yamagata University, Japan. I am a professor of Yamagata University in charge of International Exchange. My mission is to stay at several
Ecology: The science concerned with the relationships among living things and their environment.
 ECOSYSTEM VOCABULARY IN PACKET Ecology: The science concerned with the relationships among living things and their environment. Ecosystem: A community that includes all the living and nonliving things
ECOSYSTEM VOCABULARY IN PACKET Ecology: The science concerned with the relationships among living things and their environment. Ecosystem: A community that includes all the living and nonliving things
Insect Success. Insects are one of the most successful groups of living organisms on earth
 Insect Success Insects are one of the most successful groups of living organisms on earth Why Insects are so successful Insects comprise about 95% of all known animal species. Actually it is insects instead
Insect Success Insects are one of the most successful groups of living organisms on earth Why Insects are so successful Insects comprise about 95% of all known animal species. Actually it is insects instead
Biology Unit Overview and Pacing Guide
 This document provides teachers with an overview of each unit in the Biology curriculum. The Curriculum Engine provides additional information including knowledge and performance learning targets, key
This document provides teachers with an overview of each unit in the Biology curriculum. The Curriculum Engine provides additional information including knowledge and performance learning targets, key
o f the Dark Phase on Diapause Determination in Papilio xuthus L.
 No. 6] Proc. Japan Acad., 46 (1970) 541 127. Effect o f Non 24 Hour Photo period and Light Interruption o f the Dark Phase on Diapause Determination in Papilio xuthus L. By Toshitaka HIDAKA and Yoshio
No. 6] Proc. Japan Acad., 46 (1970) 541 127. Effect o f Non 24 Hour Photo period and Light Interruption o f the Dark Phase on Diapause Determination in Papilio xuthus L. By Toshitaka HIDAKA and Yoshio
Chapter 24-Flowering Plant and Animal Coevolution
 Chapter 24-Flowering Plant and Animal Coevolution coevolutionary plant-animal associations alliances that have influenced the evoluton of both partners. These examples show that plants have acquired traits
Chapter 24-Flowering Plant and Animal Coevolution coevolutionary plant-animal associations alliances that have influenced the evoluton of both partners. These examples show that plants have acquired traits
Parasites and Animal Behaviour. Karen Loxton Department of Zoology
 Parasites and Animal Behaviour Karen Loxton Department of Zoology loxtonk@tcd.ie Lecture Outline 1. Life Cycles of parasites 2. Behaviours that enhance transmission 3. Behaviours that reduce infection
Parasites and Animal Behaviour Karen Loxton Department of Zoology loxtonk@tcd.ie Lecture Outline 1. Life Cycles of parasites 2. Behaviours that enhance transmission 3. Behaviours that reduce infection
GRADE 6 SCIENCE REVISED 2014
 QUARTER 1 Developing and Using Models Develop and use a model to describe phenomena. (MS-LS1-2) Develop a model to describe unobservable mechanisms. (MS-LS1-7) Planning and Carrying Out Investigations
QUARTER 1 Developing and Using Models Develop and use a model to describe phenomena. (MS-LS1-2) Develop a model to describe unobservable mechanisms. (MS-LS1-7) Planning and Carrying Out Investigations
Desert Patterns. Plants Growth and reproduction Water loss prevention Defenses. Animals Growth and reproduction Water loss prevention Defenses
 Desert Patterns Plants Growth and reproduction Water loss prevention Defenses Animals Growth and reproduction Water loss prevention Defenses Abiotic Features Introduction A major emphasis in ecology is
Desert Patterns Plants Growth and reproduction Water loss prevention Defenses Animals Growth and reproduction Water loss prevention Defenses Abiotic Features Introduction A major emphasis in ecology is
3 Types of Interactions
 CHAPTER 18 3 Types of Interactions SECTION Interactions of Living Things BEFORE YOU READ After you read this section, you should be able to answer these questions: What determines an area s carrying capacity?
CHAPTER 18 3 Types of Interactions SECTION Interactions of Living Things BEFORE YOU READ After you read this section, you should be able to answer these questions: What determines an area s carrying capacity?
Adaptations. Organisms have specific adaptations in their environments In order to accomplish tasks they must have successful traits
 Adaptations Organisms have specific adaptations in their environments In order to accomplish tasks they must have successful traits Adaptation: beneficial trait that allows an organism to be best suited
Adaptations Organisms have specific adaptations in their environments In order to accomplish tasks they must have successful traits Adaptation: beneficial trait that allows an organism to be best suited
4 Questions relating to Behavior
 Chapter 51: Animal Behavior 1. Stimulus & Response 2. Learned Behavior 3. Connecting Behavior to Survival & Reproduction 4 Questions relating to Behavior The Dutch behavioral scientist Niko Tinbergen proposed
Chapter 51: Animal Behavior 1. Stimulus & Response 2. Learned Behavior 3. Connecting Behavior to Survival & Reproduction 4 Questions relating to Behavior The Dutch behavioral scientist Niko Tinbergen proposed
What is insect forecasting, and why do it
 Insect Forecasting Programs: Objectives, and How to Properly Interpret the Data John Gavloski, Extension Entomologist, Manitoba Agriculture, Food and Rural Initiatives Carman, MB R0G 0J0 Email: jgavloski@gov.mb.ca
Insect Forecasting Programs: Objectives, and How to Properly Interpret the Data John Gavloski, Extension Entomologist, Manitoba Agriculture, Food and Rural Initiatives Carman, MB R0G 0J0 Email: jgavloski@gov.mb.ca
Goldenrod Galls and the Scientific Method
 Goldenrod Galls and the Scientific Method Overview Groups of students are given several goldenrod stems with galls. They are asked to make observations, come up with questions and make hypotheses. They
Goldenrod Galls and the Scientific Method Overview Groups of students are given several goldenrod stems with galls. They are asked to make observations, come up with questions and make hypotheses. They
Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603)
") NCEA Level 3 Biology (91603) 2013 page 1 of 6 Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Assessment Criteria
NCEA Level 3 Biology (91603) 2013 page 1 of 6 Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Assessment Criteria
Generation Date: 12/07/2015 Generated By: Tristan Wiley Title: Bio I Winter Packet
 Generation Date: 12/07/2015 Generated By: Tristan Wiley Title: Bio I Winter Packet 1. Many natural ecosystems have been destroyed by human activity. To better manage our remaining natural ecosystems, we
Generation Date: 12/07/2015 Generated By: Tristan Wiley Title: Bio I Winter Packet 1. Many natural ecosystems have been destroyed by human activity. To better manage our remaining natural ecosystems, we
Holt McDougal ScienceFusion Student Edition 2012 Grades 6 8. correlated to the. Minnesota Academic Standards Science Grade 7
 Holt McDougal ScienceFusion Student Edition 2012 Grades 6 8 correlated to the Minnesota Academic Science Grade 7 7.1. The Nature of Science and Engineering 7.1.1. The Practice of Science 7.1.1.1. Science
Holt McDougal ScienceFusion Student Edition 2012 Grades 6 8 correlated to the Minnesota Academic Science Grade 7 7.1. The Nature of Science and Engineering 7.1.1. The Practice of Science 7.1.1.1. Science
The Cell The building blocks of life
 The Cell The building blocks of life Cells are not the smallest structure that make up living organisms. They are merely the smallest functional units. Cells themselves contain smaller units called o Tiny
The Cell The building blocks of life Cells are not the smallest structure that make up living organisms. They are merely the smallest functional units. Cells themselves contain smaller units called o Tiny
Mun. Ent. Zool. Vol. 6, No. 1, January 2011
 176 PARASITISM OF PIERIS BRASSICAE (L.) (LEP.: PIERIDAE) ON CABBAGE FARMS IN COMPARISON WITH WILD HOSTS AND STUDY ON USE OF PTEROMALUS PUPARUM (L.) (HYM. PTEROMALIDAE), AS A BIOLOGICAL CONTROL AGENT VERSUS
176 PARASITISM OF PIERIS BRASSICAE (L.) (LEP.: PIERIDAE) ON CABBAGE FARMS IN COMPARISON WITH WILD HOSTS AND STUDY ON USE OF PTEROMALUS PUPARUM (L.) (HYM. PTEROMALIDAE), AS A BIOLOGICAL CONTROL AGENT VERSUS
Abstract. Introduction
 NEW METHOD FOR RAISING QUEENS Hossein Yeganehrad Caspian Apiaries P.O. Box 16058 617, New Westminster, British Columbia, Canada, V3M 6W6 radbees@hotmail.com Paper 138, Oral Presentation XXXVIIIth Apimondia
NEW METHOD FOR RAISING QUEENS Hossein Yeganehrad Caspian Apiaries P.O. Box 16058 617, New Westminster, British Columbia, Canada, V3M 6W6 radbees@hotmail.com Paper 138, Oral Presentation XXXVIIIth Apimondia
What is wrong with deer on Haida Gwaii?
 What is wrong with deer on Haida Gwaii? A school curriculum by the Research Group on Introduced Species 2007 Forests of Haida Gwaii Haida Gwaii is an archipelago. It consists of a great number of islands,
What is wrong with deer on Haida Gwaii? A school curriculum by the Research Group on Introduced Species 2007 Forests of Haida Gwaii Haida Gwaii is an archipelago. It consists of a great number of islands,
(Write your name on every page. One point will be deducted for every page without your name!)
") POPULATION GENETICS AND MICROEVOLUTIONARY THEORY FINAL EXAMINATION (Write your name on every page. One point will be deducted for every page without your name!) 1. Briefly define (5 points each): a) Average
POPULATION GENETICS AND MICROEVOLUTIONARY THEORY FINAL EXAMINATION (Write your name on every page. One point will be deducted for every page without your name!) 1. Briefly define (5 points each): a) Average
Characteristics of Animals
 Animal Adaptations Notes Characteristics of Animals Animals are in Kingdom a. b. eukaryotic c. cells lack. do not have a backbone. Ex. do have a backbone. Ex. Animals survive by doing the following essential
Animal Adaptations Notes Characteristics of Animals Animals are in Kingdom a. b. eukaryotic c. cells lack. do not have a backbone. Ex. do have a backbone. Ex. Animals survive by doing the following essential
Interspecific competition between Diadegma semiclausum and Oomyzus sokolowskii, parasitoids of diamondback moth, Plutella xylostella
 Interspecific competition between Diadegma semiclausum and Oomyzus sokolowskii, parasitoids of diamondback moth, Plutella xylostella Zu-hua Shi, Qin-bao Li, Xin Li and Shu-sheng Liu Institute of Applied
Interspecific competition between Diadegma semiclausum and Oomyzus sokolowskii, parasitoids of diamondback moth, Plutella xylostella Zu-hua Shi, Qin-bao Li, Xin Li and Shu-sheng Liu Institute of Applied
Chapter 44. Table of Contents. Section 1 Development of Behavior. Section 2 Types of Animal Behavior. Animal Behavior
 Animal Behavior Table of Contents Section 1 Development of Behavior Section 2 Types of Animal Behavior Section 1 Development of Behavior Objectives Identify four questions asked by biologists who study
Animal Behavior Table of Contents Section 1 Development of Behavior Section 2 Types of Animal Behavior Section 1 Development of Behavior Objectives Identify four questions asked by biologists who study
University of Kentucky Department of Entomology Insects in the Classroom: Lesson Plan No. 105
 University of Kentucky Department of Entomology Insects in the Classroom: Lesson Plan No. 105 BENEFICIAL BUG SCAVENGER HUNT Prepared by Blake Newton, Extension Specialist Developed from an activity designed
University of Kentucky Department of Entomology Insects in the Classroom: Lesson Plan No. 105 BENEFICIAL BUG SCAVENGER HUNT Prepared by Blake Newton, Extension Specialist Developed from an activity designed
Ecology Notes CHANGING POPULATIONS
 Ecology Notes TEK 8.11 (B) Investigate how organisms and populations in an ecosystem depend on and may compete for biotic and abiotic factors such as quantity of light, water, range of temperatures, or
Ecology Notes TEK 8.11 (B) Investigate how organisms and populations in an ecosystem depend on and may compete for biotic and abiotic factors such as quantity of light, water, range of temperatures, or
FACTORS FOR INSECTS ABUNDANCE. 1. More number of species: In the animal kingdom more than 85 per cent of the species
 FACTORS FOR INSECTS ABUNDANCE Measures of dominance 1. More number of species: In the animal kingdom more than 85 per cent of the species belongs to insect group. Total number of insects described so far
FACTORS FOR INSECTS ABUNDANCE Measures of dominance 1. More number of species: In the animal kingdom more than 85 per cent of the species belongs to insect group. Total number of insects described so far
Pollination A Sticky Situation! A lesson from the New Jersey Agricultural Society s Learning Through Gardening program
 Pollination A Sticky Situation! A lesson from the New Jersey Agricultural Society s Learning Through Gardening program OVERVIEW: Pollination is a sticky situation. In this active lesson, students learn
Pollination A Sticky Situation! A lesson from the New Jersey Agricultural Society s Learning Through Gardening program OVERVIEW: Pollination is a sticky situation. In this active lesson, students learn
Adaptation. Biotic and Abiotic Environments. Eric R. Pianka
 Adaptation Eric R. Pianka To survive and reproduce, all living organisms must adjust to conditions imposed on them by their environments. An organism's environment includes everything impinging upon it,
Adaptation Eric R. Pianka To survive and reproduce, all living organisms must adjust to conditions imposed on them by their environments. An organism's environment includes everything impinging upon it,
Lecture 8 Insect ecology and balance of life
 Lecture 8 Insect ecology and balance of life Ecology: The term ecology is derived from the Greek term oikos meaning house combined with logy meaning the science of or the study of. Thus literally ecology
Lecture 8 Insect ecology and balance of life Ecology: The term ecology is derived from the Greek term oikos meaning house combined with logy meaning the science of or the study of. Thus literally ecology
How to use this book. How the book is organised. Answering questions. Learning and using the terminology. Developing skills
 How to use this book Welcome to the beginning of your Human and Social Biology course! We hope that you really enjoy your course, and that this book will help you to understand your work, and to do well
How to use this book Welcome to the beginning of your Human and Social Biology course! We hope that you really enjoy your course, and that this book will help you to understand your work, and to do well
Social Insects. Social Insects. Subsocial. Social Insects 4/9/15. Insect Ecology
 Social Insects Social Insects Insect Ecology Sociality evolved multiple times in insects Much of Earth s fauna consists of social insects They play major roles in entire ecosystems Proliferation of ants
Social Insects Social Insects Insect Ecology Sociality evolved multiple times in insects Much of Earth s fauna consists of social insects They play major roles in entire ecosystems Proliferation of ants
Social Insects. Insect Ecology
 Social Insects Insect Ecology Social Insects Sociality evolved multiple times in insects Much of Earth s fauna consists of social insects They play major roles in entire ecosystems Proliferation of ants
Social Insects Insect Ecology Social Insects Sociality evolved multiple times in insects Much of Earth s fauna consists of social insects They play major roles in entire ecosystems Proliferation of ants
Polyphenic Insects. genotype X environment = phenotype POLYPHENISM. genetic polymorphism vs polyphenism. the peppered moth.
 What makes for differences between individuals? Polyphenic Insects genes environment genotype X environment = phenotype POLYPHENISM poly many (more than one anyway) phen - form genetic polymorphism vs
What makes for differences between individuals? Polyphenic Insects genes environment genotype X environment = phenotype POLYPHENISM poly many (more than one anyway) phen - form genetic polymorphism vs
Abiotic Stress in Crop Plants
 1 Abiotic Stress in Crop Plants Mirza Hasanuzzaman, PhD Professor Department of Agronomy Sher-e-Bangla Agricultural University E-mail: mhzsauag@yahoo.com Stress Stress is usually defined as an external
1 Abiotic Stress in Crop Plants Mirza Hasanuzzaman, PhD Professor Department of Agronomy Sher-e-Bangla Agricultural University E-mail: mhzsauag@yahoo.com Stress Stress is usually defined as an external
6 2 Insects and plants
 6 2 Insects and plants Insect DIY 1. Find plant habitat 2. Find plant 3. Accept plant 4. Eat survive, reproduce Plant characteristics Shape structure Mechanical defenses trichomes Chemical defenses sap,
6 2 Insects and plants Insect DIY 1. Find plant habitat 2. Find plant 3. Accept plant 4. Eat survive, reproduce Plant characteristics Shape structure Mechanical defenses trichomes Chemical defenses sap,
Biology 11 Unit 1: Fundamentals. Lesson 1: Ecology
 Biology 11 Unit 1: Fundamentals Lesson 1: Ecology Objectives In this section you will be learning about: ecosystem structure energy flow through an ecosystem photosynthesis and cellular respiration factors
Biology 11 Unit 1: Fundamentals Lesson 1: Ecology Objectives In this section you will be learning about: ecosystem structure energy flow through an ecosystem photosynthesis and cellular respiration factors
BUTTERFLY SCIENCE. 9 Science Activities for PreK, K & EarthsBirthday.org
 BUTTERFLY SCIENCE 9 Science Activities for PreK, K & 1-3 1 800 698 4438 EarthsBirthday.org CONTENTS Butterfly Life Cycle Song 4 Changing Butterfly Dance 5 What Is a Caterpillar? 6 Caterpillar & Pupa Timelines
BUTTERFLY SCIENCE 9 Science Activities for PreK, K & 1-3 1 800 698 4438 EarthsBirthday.org CONTENTS Butterfly Life Cycle Song 4 Changing Butterfly Dance 5 What Is a Caterpillar? 6 Caterpillar & Pupa Timelines
Flower Power!! Background knowledge material and dissection directions.
 Flower Power!! Background knowledge material and dissection directions. 96 Plant Dissection 3.2 Plants Essential Question: Why do plants have flowers? Questions: As you read the lab background, complete
Flower Power!! Background knowledge material and dissection directions. 96 Plant Dissection 3.2 Plants Essential Question: Why do plants have flowers? Questions: As you read the lab background, complete
INSTRUCTIONS TO TEACHERS. Bee Life Grade 2 Science and Technology Unit
 INSTRUCTIONS TO TEACHERS Bee Life Grade 2 Science and Technology Unit ONTARIO TEACHER-FOCUSED TOOLKIT INSTRUCTIONS TO TEACHERS Bee Life Instructions to Teachers This unit is a Science and Technology lesson
INSTRUCTIONS TO TEACHERS Bee Life Grade 2 Science and Technology Unit ONTARIO TEACHER-FOCUSED TOOLKIT INSTRUCTIONS TO TEACHERS Bee Life Instructions to Teachers This unit is a Science and Technology lesson
 Honeybees There are more than 22,000 types of bees in the world. Do you know which bees make honey? Without honeybees, you wouldn t have any honey to spread on your toast in the morning. You may have seen
Honeybees There are more than 22,000 types of bees in the world. Do you know which bees make honey? Without honeybees, you wouldn t have any honey to spread on your toast in the morning. You may have seen
2/3/2017. Behavioral Ecology. Genetic and Environmental Components of Behavior
 Behavioral Ecology Genetic and Environmental Components of Behavior 1 Behavioral Ecology Behavior lets organisms respond rapidly and adaptively to their environment. Usually in a beneficial way. Examples?
Behavioral Ecology Genetic and Environmental Components of Behavior 1 Behavioral Ecology Behavior lets organisms respond rapidly and adaptively to their environment. Usually in a beneficial way. Examples?
Exploring Matthaei s Ecosystems
 Name: Exploring Matthaei s Ecosystems As you walk on the trails, look for evidence of each of the following components of an ecosystem. Draw and describe what you observed and where you found it. Component
Name: Exploring Matthaei s Ecosystems As you walk on the trails, look for evidence of each of the following components of an ecosystem. Draw and describe what you observed and where you found it. Component
Science Review Notes for Parents and Students
 Science Review Notes for Parents and Students Grade 3 4th Nine Weeks 2017-2018 Page 1 Science Review Notes for Parents and Students Grade 3 Science: Fourth Nine Weeks 2017-2018 April, 2015 This resource
Science Review Notes for Parents and Students Grade 3 4th Nine Weeks 2017-2018 Page 1 Science Review Notes for Parents and Students Grade 3 Science: Fourth Nine Weeks 2017-2018 April, 2015 This resource
Red Admiral (Early Stages)
") 01 February 2014 Vince Massimo Citation: Massimo, V. (2014). Red Admiral (Early Stages) [Online]. Available from http://www.dispar.org/reference.php?id=25 [Accessed February 1, 2014]. Red Admiral (Early
01 February 2014 Vince Massimo Citation: Massimo, V. (2014). Red Admiral (Early Stages) [Online]. Available from http://www.dispar.org/reference.php?id=25 [Accessed February 1, 2014]. Red Admiral (Early
Biology (Biology_Hilliard)
") Name: Date: 1. There are two types of modern whales: toothed whales and baleen whales. Baleen whales filter plankton from the water using baleen, plates made of fibrous proteins that grow from the roof
Name: Date: 1. There are two types of modern whales: toothed whales and baleen whales. Baleen whales filter plankton from the water using baleen, plates made of fibrous proteins that grow from the roof
Bee Behavior. Summary of an article by. Stephen Taber III from Beekeeping in the United States
 Bee Behavior Summary of an article by Stephen Taber III from Beekeeping in the United States Bees Sense Organs: Vision Each compound eye is spherical in shape and comprised of some 6,300 cone-shaped facets
Bee Behavior Summary of an article by Stephen Taber III from Beekeeping in the United States Bees Sense Organs: Vision Each compound eye is spherical in shape and comprised of some 6,300 cone-shaped facets
12. Social insects. Is it better to be social? Is it better to be social? What is social? Some costs of being social
 Is it better to be social? 12. Social insects Cost and benefit viewpoint Social behavior is not always adaptive (costs exceed benefits) What are some costs of being social? What are some benefits of being
Is it better to be social? 12. Social insects Cost and benefit viewpoint Social behavior is not always adaptive (costs exceed benefits) What are some costs of being social? What are some benefits of being
Disciplinary Core List of Standards (NGSS) for 6-8 Life Science Progression
 for 6-8 Life Science Progression") LS1.A: Structure and Function - All living things are made up of cells, which is the smallest unit that can be said to be alive. An organism may consist of one single cell (unicellular) or many different
LS1.A: Structure and Function - All living things are made up of cells, which is the smallest unit that can be said to be alive. An organism may consist of one single cell (unicellular) or many different
Where in the world does your food come from?
 Pollinators come in all species, sizes, shapes and shades Where in the world does your food come from? Do you eat fruits? vegetables? nuts? seeds? grains? Where do you get them? Usually Mom or Dad go to
Pollinators come in all species, sizes, shapes and shades Where in the world does your food come from? Do you eat fruits? vegetables? nuts? seeds? grains? Where do you get them? Usually Mom or Dad go to
Predation. Vine snake eating a young iguana, Panama. Vertebrate predators: lions and jaguars
 Predation Vine snake eating a young iguana, Panama Vertebrate predators: lions and jaguars 1 Most predators are insects Parasitoids lay eggs in their hosts, and the larvae consume the host from the inside,
Predation Vine snake eating a young iguana, Panama Vertebrate predators: lions and jaguars 1 Most predators are insects Parasitoids lay eggs in their hosts, and the larvae consume the host from the inside,
L. fabarum L. fabarum.
 (201-210) 1391 1 43 Aphis fabae Lysiphlebus fabarum 3 3 2 *1 1 2 (90/12/2 : 90/6/5 : ).. - - Lysiphlebus fabarum (Marshall) -.. ( 15 ). -. -... - -... L. fabarum L. fabarum. : E-mail:arasekh@ut.ac.ir 09126603166
(201-210) 1391 1 43 Aphis fabae Lysiphlebus fabarum 3 3 2 *1 1 2 (90/12/2 : 90/6/5 : ).. - - Lysiphlebus fabarum (Marshall) -.. ( 15 ). -. -... - -... L. fabarum L. fabarum. : E-mail:arasekh@ut.ac.ir 09126603166
KNES Primary School Year 5 Science Course Outline:
 KNES Primary School Year 5 Science Course Outline: 2017-2018 Year Overview. The Year 5 program of study for Science will take students on a scientific adventure through various activities which will be
KNES Primary School Year 5 Science Course Outline: 2017-2018 Year Overview. The Year 5 program of study for Science will take students on a scientific adventure through various activities which will be
ECOSYSTEMS AND THEIR LIVING COMMUNITIES
 ECOSYSTEMS AND THEIR LIVING COMMUNITIES COMMUNITY Each community is made up of populations of various organisms living in the same location at the same time. community 1 = popln 1 + popln 2 + popln 3 Each
ECOSYSTEMS AND THEIR LIVING COMMUNITIES COMMUNITY Each community is made up of populations of various organisms living in the same location at the same time. community 1 = popln 1 + popln 2 + popln 3 Each
14.1 Habitat And Niche
 14.1 Habitat And Niche A habitat differs from a niche. Habitat physical area in which an organism lives Niche each species plays a specific role in an ecosystem niche includes the species habitat, feeding
14.1 Habitat And Niche A habitat differs from a niche. Habitat physical area in which an organism lives Niche each species plays a specific role in an ecosystem niche includes the species habitat, feeding
Campbell Essential Biology, 4/e (Simon/Reece/Dickey)
") Campbell Essential Biology, 4/e (Simon/Reece/Dickey) Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) Using the branching tree of life for
Campbell Essential Biology, 4/e (Simon/Reece/Dickey) Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) Using the branching tree of life for
o Can you find any nectar? Brood? Honey? o Can you find any drones and drone cells? o Can you find the queen bee?
 o Can you find any nectar? Brood? Honey? o Can you find any drones and drone cells? o Can you find the queen bee? *NOTE: The queen in Observation Hive #1 has a yellow mark on her. Put the wooden panels
o Can you find any nectar? Brood? Honey? o Can you find any drones and drone cells? o Can you find the queen bee? *NOTE: The queen in Observation Hive #1 has a yellow mark on her. Put the wooden panels
Garlic Mustard Biocontrol An Update. Jeanie Katovich, Esther Gerber, Hariet Hinz, Luke Skinner, David Ragsdale and Roger Becker
 Garlic Mustard Biocontrol An Update Jeanie Katovich, Esther Gerber, Hariet Hinz, Luke Skinner, David Ragsdale and Roger Becker Ceutorhynchus scrobicollis McCornack C. scrobicollis Life Cycle Summer Adult
Garlic Mustard Biocontrol An Update Jeanie Katovich, Esther Gerber, Hariet Hinz, Luke Skinner, David Ragsdale and Roger Becker Ceutorhynchus scrobicollis McCornack C. scrobicollis Life Cycle Summer Adult
PERFORMANCE OF NATURAL ENEMIES REARED ON ARTIFICIAL DIETS J.E. Carpenter 1 and S. Bloem 2 1
 Performance of natural enemies reared on artificial diets 143 PERFORMANCE OF NATURAL ENEMIES REARED ON ARTIFICIAL DIETS J.E. Carpenter 1 and S. Bloem 2 1 U.S. Department of Agriculture, Agricultural Research
Performance of natural enemies reared on artificial diets 143 PERFORMANCE OF NATURAL ENEMIES REARED ON ARTIFICIAL DIETS J.E. Carpenter 1 and S. Bloem 2 1 U.S. Department of Agriculture, Agricultural Research
Regents Review Assignment #8-A08 Living Environment: Comet Part A Questions
 Part A Questions 1. A student notices that fruit flies with the curlywing trait develop straight wings if kept at a temperature of 16 C, but develop curly wings if kept at 25 C. The best explanation for
Part A Questions 1. A student notices that fruit flies with the curlywing trait develop straight wings if kept at a temperature of 16 C, but develop curly wings if kept at 25 C. The best explanation for
BENEFICIAL INSECTS GOING BUGGY
 BENEFICIAL INSECTS GOING BUGGY GOALS FOR THIS STATION Understand the importance of beneficial insects Understand the importance of native bees Understand the importance of conserving insect habitats ROLES
BENEFICIAL INSECTS GOING BUGGY GOALS FOR THIS STATION Understand the importance of beneficial insects Understand the importance of native bees Understand the importance of conserving insect habitats ROLES
Community and Population Ecology Populations & Communities Species Diversity Sustainability and Environmental Change Richness and Sustainability
 1 2 3 4 Community and Population Ecology Chapter 6 Populations & Communities Biosphere> ecosystems> communities> populations> individuals A population is all of the individuals of the same species in a
1 2 3 4 Community and Population Ecology Chapter 6 Populations & Communities Biosphere> ecosystems> communities> populations> individuals A population is all of the individuals of the same species in a
The effect of light on the Vanessa cardui. By Michael Muransky
 The effect of light on the Vanessa cardui By Michael Muransky The subject of this experiment is painted ladies caterpillars. Their eggs are pale green and are placed on the upper- side of the leaves. They
The effect of light on the Vanessa cardui By Michael Muransky The subject of this experiment is painted ladies caterpillars. Their eggs are pale green and are placed on the upper- side of the leaves. They
Understand how living things function and how they interact with one another and their environment.
 District of Columbia Public Schools Essential Knowledge and Skills for Science (Grade 7) SCIENCE, Grade 7: Life Science Content Standard 2: Understand how living things function and how they interact with
District of Columbia Public Schools Essential Knowledge and Skills for Science (Grade 7) SCIENCE, Grade 7: Life Science Content Standard 2: Understand how living things function and how they interact with
CHEMICALS IN HOST PARASITOID AND PREY PREDATOR RELATIONS
 CHEMICALS IN HOST PARASITOID AND PREY PREDATOR RELATIONS Lozano C. Estacion Experimental del Zaidin, Consejo Superior de Investigaciones Científicas, Granada, Spain Keywords: semiochemicals, pesticides,
CHEMICALS IN HOST PARASITOID AND PREY PREDATOR RELATIONS Lozano C. Estacion Experimental del Zaidin, Consejo Superior de Investigaciones Científicas, Granada, Spain Keywords: semiochemicals, pesticides,
Characteristics of Life
 Name Date Characteristics of Life Most people feel confident that they could identify a living thing from a nonliving thing, but sometimes it s not so easy. Scientists have argued for centuries over the
Name Date Characteristics of Life Most people feel confident that they could identify a living thing from a nonliving thing, but sometimes it s not so easy. Scientists have argued for centuries over the
FLOWERS AND POLLINATION. This activity introduces the relationship between flower structures and pollination.
 FLOWERS AND POLLINATION This activity introduces the relationship between flower structures and pollination. Objectives for Exam #1 1. Identify flower structures and match those structures to specific
FLOWERS AND POLLINATION This activity introduces the relationship between flower structures and pollination. Objectives for Exam #1 1. Identify flower structures and match those structures to specific
Pollination Lab Bio 220 Ecology and Evolution Fall, 2016
 Pollination Lab Bio 220 Ecology and Evolution Fall, 2016 Journal reading: Comparison of pollen transfer dynamics by multiple floral visitors: experiments with pollen and fluorescent dye Introduction: Flowers
Pollination Lab Bio 220 Ecology and Evolution Fall, 2016 Journal reading: Comparison of pollen transfer dynamics by multiple floral visitors: experiments with pollen and fluorescent dye Introduction: Flowers
Evolution. 1. The figure below shows the classification of several types of prairie dogs.
 Name: Date: 1. The figure below shows the classification of several types of prairie dogs. 3. Which statement describes the best evidence that two species share a recent common ancestor? A. The species
Name: Date: 1. The figure below shows the classification of several types of prairie dogs. 3. Which statement describes the best evidence that two species share a recent common ancestor? A. The species
Characteristics of Life
 Characteristics of Life All living things share some basic characteristics: 1. Organization 2. Movement 3. Made up of cells 4. Reproduce 5. Grow and / or develop 6. Obtain and use energy 7. Respond to
Characteristics of Life All living things share some basic characteristics: 1. Organization 2. Movement 3. Made up of cells 4. Reproduce 5. Grow and / or develop 6. Obtain and use energy 7. Respond to
Name Date Period Handout A: Characteristics of Life
 Name Date Period Handout A: Characteristics of Life Directions: 1. Read and highlight the main ideas in each passage. 2. Circle Key vocabulary 3. Answer the questions below each passage. Most people feel
Name Date Period Handout A: Characteristics of Life Directions: 1. Read and highlight the main ideas in each passage. 2. Circle Key vocabulary 3. Answer the questions below each passage. Most people feel
A population is a group of individuals of the same species, living in a shared space at a specific point in time.
 A population is a group of individuals of the same species, living in a shared space at a specific point in time. A population size refers to the number of individuals in a population. Increase Decrease
A population is a group of individuals of the same species, living in a shared space at a specific point in time. A population size refers to the number of individuals in a population. Increase Decrease
Lesson Overview. Gene Regulation and Expression. Lesson Overview Gene Regulation and Expression
 13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
Project. Aim: How does energy flow in Arctic and Antarctic ecosystems? Explore. The four food webs are:
 Name: Date: Aim: How does energy flow in Arctic and Antarctic ecosystems? Explore The four food webs are: o Antarctic Marine Food Web o Arctic Marine Food Web o Tundra Land Food Web o Tundra Freshwater
Name: Date: Aim: How does energy flow in Arctic and Antarctic ecosystems? Explore The four food webs are: o Antarctic Marine Food Web o Arctic Marine Food Web o Tundra Land Food Web o Tundra Freshwater
Science 9 - Unit A Review Sheet
 Science 9 - Unit A Review Sheet Learning Outcomes Can you? describe the relative abundance of species on Earth and in different environments describe examples of variation among species and within species
Science 9 - Unit A Review Sheet Learning Outcomes Can you? describe the relative abundance of species on Earth and in different environments describe examples of variation among species and within species
Lesson 4: Insect Life Cycles
 Life Cycles -> 4: Insect Life Cycles Lesson 4: Insect Life Cycles Getting Started? Big Ideas P P P How are the life cycles of plants and animals similar and different? Where can cycles be found in nature?
Life Cycles -> 4: Insect Life Cycles Lesson 4: Insect Life Cycles Getting Started? Big Ideas P P P How are the life cycles of plants and animals similar and different? Where can cycles be found in nature?