Palissya: A global review and reassessment of Eastern Gondwanan material
|
|
|
- Darcy Franklin
- 5 years ago
- Views:
Transcription

1 Accepted Manuscript Palissya: A global review and reassessment of Eastern Gondwanan material G.A. Pattemore, J.F. Rigby, G. Playford PII: S (14) DOI: doi: /j.revpalbo Reference: PALBO 3548 To appear in: Review of Palaeobotany and Palynology Received date: 7 March 2014 Revised date: 29 July 2014 Accepted date: 3 August 2014 Please cite this article as: Pattemore, G.A., Rigby, J.F., Playford, G., Palissya: A global review and reassessment of Eastern Gondwanan material, Review of Palaeobotany and Palynology (2014), doi: /j.revpalbo This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
2 Palissya : a global review and reassessment of Eastern Gondwanan material G.A. Pattemore a,*, J.F. Rigby b, G. Playford a a School of Earth Sciences, The University of Queensland, St. Lucia, Queensland 4072, Australia. b School of Earth, Environmental and Biological Sciences, Queensland University of Technology, GPO Box 2434, Brisbane, Queensland 4001, Australia. Abstract The conifer genus Palissya is reviewed giving special attention to assessing the reliability of Gondwanan material referred to this genus. The collection localities of Australasian specimens are mapped on palaeogeographic reconstructions of Eastern Gondwana at relevant times between 179 Ma 119 Ma. Australasian and other Gondwanan material previously referred to Palissya is considered to differ substantially from the generic diagnosis. Specimens from the Toarcian of Australasia are shown to be allied to Knezourocarpon; this genus is only known from the Toarcian of eastern Australia. Middle Jurassic Lower Cretaceous Australasian specimens previously referred to Palissya are likely allied to the Knezourocarponaceae fam. nov. The higher taxonomic affinity of this family is unknown but broadly, its structure is suggestive of ginkgoalean or pteridospermous affiliation. Southern hemisphere specimens were probably catkin-like and megascopically very different from the northern hemisphere Palissya and the two groups are unlikely to be closely related. Consequently, genuine Palissya cones and foliage appear to be restricted to the Rhaetian Lower Jurassic of the northern hemisphere. * Corresponding author
3 address: (G.A. Pattemore) Keywords: Palissya, Palyssia, Knezourocarpon, Gondwana, Jurassic. 1. Introduction Cones attributed to the Palissyaceae Florin 1958 have been rarely reported from outside of the northern hemisphere. However, in recent decades a considerable amount of material from the Toarcian Aptian of Australasia has been assigned to Palissya Endlicher 1847 emend. Florin Two separate reviews of Palissya differed on major points including the structure and gender of the cone (Parris et al., 1995; Schweitzer and Kirchner, 1996). While each review adopted a global perspective, one focused on the northern hemisphere while the other was largely concerned with Australasian material. These conjectural points have largely been resolved by Wang (2012) prompting a reconsideration of Australasian specimens assigned to this genus. Moreover, the establishment of the genus Knezourocarpon Pattemore 2000 based on specimens from the Toarcian of Queensland, Australia, suggests Australasian specimens previously ascribed to Palissya should be reassessed. The compound structure of the Palissya cone has been conjectural and has been widely used to support conifer affiliation (or otherwise) of the Palissyaceae; however, this contribution does not attempt to further define the higher taxonomic affinity of this family. Rather, this paper seeks to clarify the geographic and temporal distribution of representatives of the Palissyaceae particularly regarding Gondwana. The palaeogeographic position of Eastern Gondwanan representatives of Palissya and Knezourocarpon are mapped based on the modelling of Seton et al. (2012). 2. Palissyaceae Palissyaceae has been regarded historically as a conifer family, but this was doubted by
4 Parris et al. (1995) because they did not regard the cone, Palissya, as compound. This was disputed by Schweitzer and Kirchner (1996). Phylogenetic analysis of conifers by Miller (1999) excluded Palissya due to its uncertain affiliation. Meyen (1984) placed the Palissyaceae within rank Pinales and rank Pinopsida while Taylor et al. (2009) assigned Palissya to the Coniferales Palissya The genus was established monotypically with P. braunii Endlicher 1847 from the Lower Jurassic of Germany. Some authors (eg. Escapa et al., 2008; Bosma et al., 2012) have adopted an alternative orthography; viz. Palyssia. The original spelling is accepted here, being patronymic for Bernard Palissy (Endlicher, 1847). Nathorst (1908) referred Palissya braunii to P. sphenolepis (Braun 1843) Nathorst 1908 emend. Florin 1958 as a correction of priority and described more material from the Lower Jurassic of Sweden. Endlicher s (1847) brief diagnosis specified paired sessile leaves and a cone with loosely overlapping scales. Florin s (1958) emendation of the genus and the type species included a detailed description of ovules, ovule scale complex and foliage cuticle. Reconstructions of Palissya bracts were provided by Hirmer (1936) and Florin (1951); Figure 1 (A B) shows Hirmer s (1936) bract and ovule scale complex reconstruction. Key references for this genus are listed in Table 1. [Figure 1] Over recent decades much new material has been referred to Palissya and the genus has been reviewed by Parris et al. (1995) and Schweitzer and Kirchner (1996). The latter authors considered British specimens previously reported in an unpublished manuscript and added new material from Iran. Both reviews differed on major points, notably the likely structure
5 and gender of the cone. However, each account considered only slightly overlapping subsets of all material referred to Palissya; one focused on the northern hemisphere, the other on Australasian material. Parris et al. (1995) disputed the assertion that the cone was compound but Schweitzer and Kirchner (1996) disagreed. Wang (2012) described Rhaetian specimens from China which are remarkably similar to the European type species, and reported microscopic examination of the ovuliferous scale complex and ovules. He confirmed Florin s (1958) generic diagnosis regarding the structure of the Palissya cone, in particular, stating that the cone is compound and female, thus supporting conifer affiliation. [Table 1] Foliage was originally included in the diagnosis of Palissya as established by Endlicher (1847), and in at least two subsequent emendations (Florin, 1958; Schweitzer and Kirchner, 1996). Following Seward (1919), Parris et al. (1995) discussed the inclusion of foliage without a clearly associated cone and considered a number of such referrals to have caused considerable confusion. Seward s (1919) viewpoint was based on significant differences in foliage cuticle reported by Schenk (1867) and Nathorst (1908), but Florin (1958) considered Seward s (1919) view as going too far. Pott and McLoughlin (2011) also considered Palissya leaf material and referred new foliage specimens to Elatocladus Halle 1913 but within the Palissyaceae. Stachyotaxus Nathorst 1886 comprises Elatocladus-type foliage and male and female cones, and the male cone was considered to be catkin-like (Harris, 1935). Stachyotaxus was regarded as a conifer cone similar to Palissya but with only a single pair of ovules per bract (Nathorst, 1908; Seward, 1919; Harris, 1935; Stewart and Rothwell, 1993; Arndt, 2002; Taylor et al., 2009; Pott and McLoughlin, 2011); Figure 1 (C D) shows Hirmer s (1936) bract reconstruction. Stomata of Stachyotaxus are in two parallel longitudinal zones on the abaxial
6 surface which is also the case in Palissya (Nathorst, 1908). Stachyotaxus has been reported by numerous authors (Table 1). Pott and McLoughlin (2011) regarded Stachyotaxus as restricted to Rhaetian sediments of Greenland and Scania. Jurassic specimens from Antarctica, referred to the genus by Birkenmajer and Ociepa (2008), are insufficiently preserved for confident identification Palissya northern hemisphere Northern hemisphere material ascribed to Palissya has been reviewed by several authors (Nathorst, 1908; Hirmer, 1936; Florin, 1958; Parris et al., 1995; Schweitzer and Kirchner, 1996). The type species, P. sphenolepis, was reported from the Rhaetian Lower Jurassic of Franconia, Germany and Stabbarp, Sweden (Nathorst, 1908; Florin, 1958; Parris et al., 1995); several uppermost Triassic accounts of the genus from Europe and Asia were summarised by Dobruskina (1994). Other poorly preserved and fragmentary material was described from Sofiero, Sweden as Palissya sp. by Nathorst (1908) and Palissya sp. Sofiero by Parris et al. (1995). The reviews by Parris et al. (1995) and Schweitzer and Kirchner (1996) differ considerably in their assessments of Palissya. Schweitzer and Kirchner (1996) regarded Palissya as a compound male cone; by contrast, Parris et al. (1995) considered it as a probable female cone and not compound. Both Parris et al. (1995) and Schweitzer and Kirchner (1996) emended the circumscription of the type species. Whether or not Palissya is a compound cone has proved controversial (Seward, 1919; Florin, 1944, 1958; Parris et al., 1995; Schweitzer and Kirchner, 1996; Pott and McLoughlin, 2011). However, Palissya material from China has confirmed the compound structure of the cone (Wang, 2012). In an unpublished thesis, Hill (1974) identified Palissya in the Middle Jurassic of Yorkshire and described a new species, P. harrisii, comprising foliage and female cones. He recorded cuticle of both bract and foliage, also organic attachment in one specimen
7 between foliage and cone (p.167, specimen B1 and plates 24 25). Although Hill (1974) considered the cones to be female, no ovular attachments were identified. The foliage preserved as part of specimen B1 with the attached cone, is poor and cuticle could not be recovered. An ovuliferous ridge was identified but the extent to which it is separate from the bract is unclear. Bracts are decurrent on the cone axis and bract arrangement is uncertain. Parris et al. (1995) doubted evidence for leaf attachment in the Swedish material. These authors did not emend the diagnosis of Palissya to exclude foliage, but their emended description of the type species did not allude to foliage remains. Schweitzer and Kirchner (1996) emended the diagnosis of the type species to include not only Elatocladus-type leaves and Palissya cones (considered to be male cones by these authors), but also Compsostrobus-type female cones based on the discovery of new Iranian material (discussed below). Metridiostrobus Delevoryas and Hope 1981 and Compsostrobus Delevoryas and Hope 1973 were reported from the Upper Triassic of North Carolina, U.S.A. Schweitzer and Kirchner (1996) considered Palissya as a Metridiostrobus-type cone. According to Delevoryas and Hope (1981), Metridiostrobus is a female cone; however, Schweitzer and Kirchner (1996) re-interpreted the cone as male and as associated with Compsostrobus. Delevoryas and Hope (1973, 1981, 1987) did not refer foliage specimens to Elatocladus, but did identify unattached conifer foliage co-preserved with Compsostrobus. The apparent similarities between the European Palissya type species and North American Metridiostrobus specimens are remarkable but Delevoryas and Hope (1981) discussed some important differences. This stands in stark contrast to the apparent difference between the European Palissya type species and the southern hemisphere Palissya specimens described by Parris et al. (1995), as discussed below. Palissya hunanensis Wang 2012, from the Rhaetian of Hunan Province, China, appears remarkably similar to the type species of Palissya and, as discussed above, supports previous interpretations that the cone is female, compound and conifer-like. The cone structure is
8 shown in Figure 2. Swedish specimens termed Palissya sp. by Nathorst (1908) and Palissya sp. Sofiero by Parris et al. (1995) are too poorly preserved to allow comparative evaluation. Several specimens referred to Palissya from the northern hemisphere were judged by Schweitzer and Kirchner (1996) to be dubious representatives of the genus. [Figure 2] 2.3. Palissya Iran Palissya oleschinskii Schweitzer and Kirchner 1996 was described by Schweitzer and Kirchner (1996) from the Rhaetian Lower Jurassic of Zangerud, Iran. These cones were preserved with Elatocladus-type leaves and isolated cones regarded by these authors as female, and referred to Compsostrobus brevirostratus Schweitzer and Kirchner They reported a likely association between the Elatocladus-type foliage and Compsostrobus-type female cones from another Iranian site, Tazareh, which were found within strata equivalent to those at Zangerud; this forms the basis for their association of all three plant organs. Schweitzer and Kirchner (1996) noted that not all Elatocladus-type leaves are referable to Palissya as other conifer genera are known to bear similar leaves. The bract attachment of Palissya oleschinskii was considered by Schweitzer and Kirchner (1996) to be helically arranged. However, their figured specimen appears more likely to have been bilateral, opposite and sessile. Further, comparing their figured P. oleschinskii specimens and reconstruction, it appears that Schweitzer and Kirchner (1996) considered the end of the cone with the apparently thicker section of axis as the top of the cone. As their reconstruction shows, this places the sporangia on the upper surface of the lamina; thus implying that the cleavage plane must be more-or-less central to the cone axis near the cone s figured apex and be no longer central to the axis near to cone s figured base.
9 Figured lamina attachment does not support this view. An alternative view is that the apparently thicker axial section constitutes the base of the cone, while the opposing end has poorer preservation with more sediment obscuration; this would be expected if this end were the more delicate apex of the cone. If the stem thickens as Schweitzer and Kirchner (1996) suggested, then it seems more likely that the cone was oriented in a catkin-like manner. Both these alternatives place the microsporangial surface beneath the lamina, which commonly characterises extant and extinct gymnosperms and pteridophytes by promoting maximum pollen dispersal (Niklas, 1985). The Iranian Palissya oleschinskii, although sessile, appears significantly different to other northern hemisphere species of this genus Palissya Gondwana excluding Australasia Schweitzer and Kirchner (1996) reviewed numerous early descriptions of Palissya from India, Antarctica and South America and rejected all except specimens reported by Frenguelli (1949) as P. conferta (Oldham and Morris 1862) Feistmantel 1877a and P. jabalpurensis Feistmantel 1876b from Chubut, Patagonia. Both these species were originally established from Indian foliage specimens (Feistmantel, 1876a,b, 1877a,b,c, 1879, 1880), while the identifications of them by Frenguelli (1949) comprised foliage and cones. In contrast to Schweitzer and Kirchner s (1996) view, Parris et al. (1995) regarded Frenguelli s (1949) fructifications as unlike the type species. Parris et al. (1995) suggested that Frenguelli s (1949) hand-drawn figures show little resemblance to his figured specimens. These specimens have subsequently been referred elsewhere (Escapa et al., 2008). Additional early descriptions of foliar remains assigned to Palissya from the Jurassic Cretaceous of India were summarised by Lakhanpal et al. (1976); while some of these species are noted as referred elsewhere, the remainder are insufficiently described to confidently be regarded as conforming to the genus. The fructification Palissya antarctica Cantrill 2000, from the Aptian Cerro Negro
10 Formation of Snow Island, Antarctica (Cantrill, 2000), is relatively small compared to other specimens referred to the genus. The adaxial processes were interpreted as likely pear-shaped ovules; however, it is unclear whether these processes represent ovules or sporangia. The laminae appear to be helically arranged and petiolate, and there is no evidence that the fructification is compound (Cantrill, 2000). The circumscription of this species was based on six incomplete specimens and the arguments presented for referral to Palissya were largely informed by the work of Parris et al. (1995) regarding Australasian specimens. It is shown herein that Australasian specimens previously referred to Palissya vary significantly from the generic diagnosis. However, it seems possible that P. antarctica may be related to Victorian Cretaceous specimens hitherto identified as Palissya (see Table 2 and Figure 7). [Figure 3] 2.5. Palissya Australasia Southeast Queensland (Middle Jurassic) Palissya ovalis Parris, Drinnan and Cantrill 1995 was reported from Reynold s Creek and Kalbar in southeast Queensland by Parris et al. (1995), and from Mutdapilly (~22 km north of Kalbar) by McLoughlin and Drinnan (1995); see Figure 3 (point R). These authors considered these sites to be within the Walloon Coal Measures, which were regarded as Bathonian in age (Turner et al., 2009; Cook et al., 2013). No exact locality information was provided for the Reynold s Creek site; however, Queensland Museum specimen records indicate that these were collected from the Parish of Fassifern (3 4 km southwest of Kalbar). Figure 4 (point A) shows these sites juxtaposed with the collection location of similar material from the Callovian of the South Island, New Zealand (discussed below) on a reconstructed map of Eastern Gondwana. Palissya ovalis fructifications are helically arranged with petiolate, ovate laminae; each
11 lamina bears 5 6 pairs of adaxial processes, although specimens reported by McLoughlin and Drinnan (1995) have 7 8 pairs of such processes. These authors postulated that the larger number of sporangial pairs in their specimens reflected better preservation. Parris et al. (1995) regarded their specimens as representing a range of fructifications from open to closed. Their only figured open fructification (Parris et al., 1995, fig. 9A,B) appears to be oppositely branched, whereas the more closed fructifications appear helically arranged; however, both the open and closed fructifications figured by McLoughlin and Drinnan (1995) appear helically branched. If the thicker end of the fructification axis shown by Parris et al. (1995, fig. 9A,C) were oriented upward then the sporangia would be pendulous and the fructification catkin-like. This also seems likely for the P. ovalis fructifications reported by McLoughlin and Drinnan (1995). [Figure 4] South Island, New Zealand (Middle Jurassic) Edwards (1934) and Parris et al. (1995) reported four specimens from the Waikawa locality, South Island, New Zealand, which were assigned to Palissya bartrumii Edwards The collection site is a seaside rock platform in Curio Bay composed of sediments considered latest Callovian in age (Pole, 2001, 2004). Early palaeobotanical studies from this locality were summarised by Arber (1917), who referred a foliage specimen to Palissya that had previously been reported as Taxites manawao Hector The latter, as figured by Hector (1886, fig.30a(2)), is small (albeit with no indication of actual size) and lacks sufficient detail to confirm identification. The Curio Bay site was discussed by Thorn (2001) and fossil wood from there was examined by Thorn (1999). Parris et al. (1995) ascribed several specimens collected from Slope Point (~7 km west of Curio Bay) to Palissya bartrumii. Their material is neither described nor illustrated. The
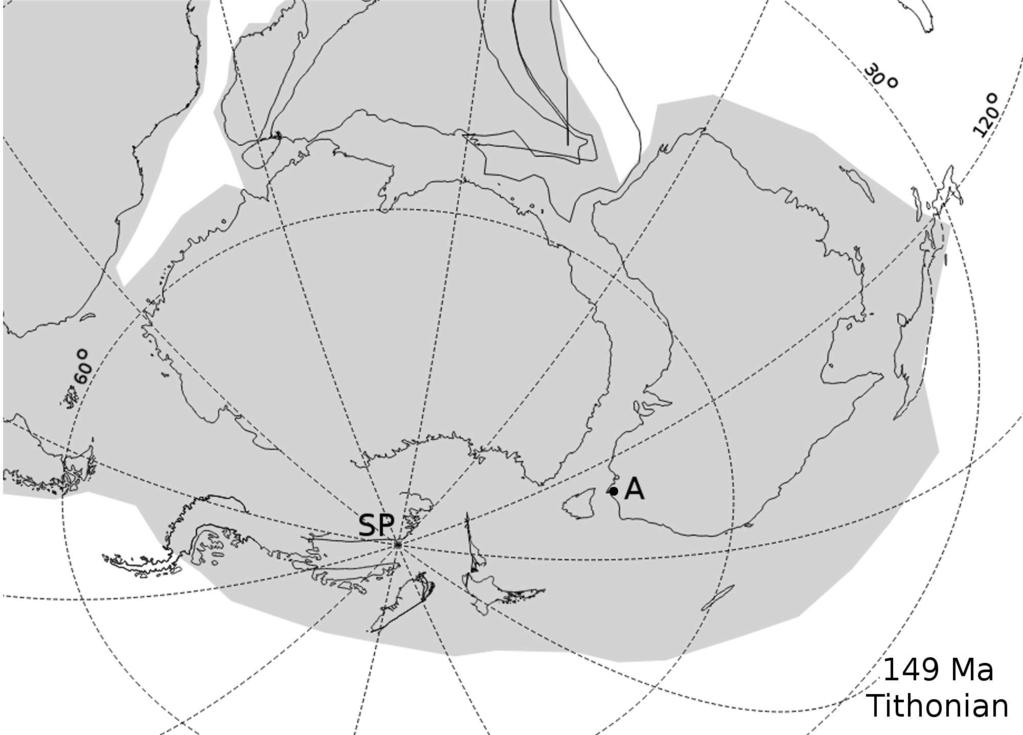
12 host strata were considered upper Callovian but stratigraphically lower than those at Curio Bay (Pole, 2004). Two incomplete specimens from Mokoia, near Gore, were referred to Palissya bartrumii by Edwards (1934); these were originally identified as Stachyotaxus sp. by Arber (1917). Both were substantially larger than other material assigned to P. bartrumii; Edwards (1934) considered them to be the only New Zealand specimens to have reflexed laminae, but this may only be a preservational effect. The specimen figured by Parris et al. (1995, fig. 12) shows remarkably different adaxial processes to other Australasian material assigned to this genus. Their figure suggests the fructification has a uniformly dispersed, probable microsporangial array of investments beneath a sporophyll, rather than paired processes as specified in the generic circumscription of Palissya. The Mokoia site was dated as Callovian by Parris et al. (1995), but they noted that the actual site may have been obliterated by natural processes. All these southern New Zealand sites are within the Murihiku Terrane (Pole, 2009); see Figure 3 (points M and C). Parris et al. (1995) emended the diagnosis of Palissya bartrumii to include helical arrangement of sporophylls. Sporophylls are ovate and petiolate, with 3 4 pairs of adaxial processes per sporophyll, apparently assumed from the dendroidal pattern visible in some specimens. The adaxial processes appear to have likely been pendulous sporangia. The Palissya bartrumii material is poorly preserved and only the holotype can be regarded as approaching completeness. These specimens are very similar to those referred to P. ovalis from the Middle Jurassic of southeast Queensland (discussed above), except that P. bartrumii has fewer reported adaxial processes per sporophyll. Figure 4 (point B) shows the location of these three southern New Zealand sites within a reconstruction of the Eastern Gondwana landmass at approximately the Bathonian Callovian boundary; this is compared with the palaeogeographic position of the Queensland material (point A).
13 North Island, New Zealand (Late Jurassic) A specimen from beds exposed along New Zealand s North Island coastline, at the mouth of the Huriwai Stream, was reported by Bartrum (1921) as?stachyotaxus cf. S. elegans Nathorst 1908, and was later referred to Palissya bartrumii by Edwards (1934). This site is located several kilometres south of Waikato Head (Bartrum, 1921); see Figures 3 (point W), 5 (point A). Parris et al. (1995) considered this specimen to be latest Jurassic Early Cretaceous in age. While the Huriwai Formation at Waikato Head is now considered Tithonian (Parris et al., 1995; Pole, 2009), it is not clear that the sediments at Bartrum s (1921) collection site can be chronocorrelated as implied by Parris et al. (1995); however, as this site appears to be within the Murihiku Terrane, a Late Jurassic age seems probable. Bartrum s (1921) specimen was figured by Edwards (1934); this indifferently preserved specimen shows bilateral symmetry of sporophylls, but the adaxial processes are indistinct. Parris et al. (1995) remarked that the fructification is smaller than other New Zealand material assigned to the genus. This specimen is separated geographically and temporally from other material referred to P. bartrumii (Table 2). [Figure 5] Boola Boola, Victoria (Early Cretaceous) Palissya elegans Parris, Drinnan and Cantrill 1995 has been reported from several sites within the Boola Boola State Forest, Victoria (Parris et al., 1995); see Figures 3 (point B), 6 (point A). Additional specimens from the same area were assigned to this species by McLoughlin et al. (2002). All were dated as Neocomian by Parris et al. (1995); however, all specimens are from the Locmany Member of the Rintoul Creek Formation, Gippsland Basin (McLoughlin et al., 2002), and this member has been dated as Valanginian (Tosolini et al.,
14 1999). Palissya elegans was reported to have helically arranged, petiolate, oval to round laminae with three pairs of adaxial processes per lamina (Parris et al., 1995). Laminae have a ~5 mm long narrow apical projection. The only figured specimen to show this projection is the holotype (Parris et al., 1995, fig. 3), which is also the only complete specimen. No cuticle is preserved. Cone fragments were tested for spores but preparations revealed a diverse, presumably part-allochthonous, assemblage (Parris et al., 1995). The figures by Parris et al. (1995, figs. 3A, 4A-D and 5D) suggest the possibility that laminae are decussate on the axis. The lower laminae in their figure 3A appear decussate; while 4A D suggest bilateral symmetry. The magnified figure 5D shows two adjacent laminae on the same side of the axis. Given the petiole width and the very close proximity of these two laminae, and moreover that the figure shows no apparent stem twist, it would be difficult to physically fit a helical arrangement of attached laminae over such a short axial section. A poor specimen of Palissya elegans figured by McLoughlin et al. (2002, fig. 49A) suggests that this fructification is, in fact, decussate. The figured ovule (Parris et al., 1995, figs. 5D,E) appears to be of a size, shape and orientation that is inconsistent with their reconstruction (Parris et al., 1995, figs. 6B,C) and with the argument that cup-like investments held an ovule. Parris et al. (1995, figs. 4E and 5A,B) showed circular features which were apparently on the lamina and surrounded by adaxial processes somewhat like the type species as described by Nathorst (1908), and this feature was interpreted as a cup. It was suggested that this circular area encompassed an indentation; however, this indentation appears to have been assumed based upon the exposure of sediment within this feature. The indentation is only visible at one point on one figured specimen. The adaxial processes have striations which radiate from the circular feature, thus
15 enhancing the optical effect of the cup. The characteristic dendroidal pattern with a central stalk, often observed in Australasian specimens referred to Palissya, was considered by Parris et al. (1995) to be the broken edges of overlapping processes, with the central stalk inferred to represent adpressed inner margins of the adaxial processes. In considering this explanation, if a specimen cleaved through the adaxial processes, it would be expected instead to produce a series of touching, or nearly touching, circles; ie. in section, similar to that figured by Arndt (2002, pl. 2, fig. 6). The adaxial processes with striations radiating from the circular features are only visible arising from approximately half of each circular feature. It seems likely that these so called cup-shaped processes are illusory (suggested by the striated radiating pattern), and were probably small convex microsporangial leaf-like blades. The apical projections appear more likely to have been a drip-tip (accuminate); if so, the fructification was probably catkin-like. Further, the fine and slender disposition of the fructification strongly suggests that it was catkin-like rather than a rigid upright cone. [Figure 6] Koonwarra Fossil Bed, Victoria (Lower Cretaceous) A few indifferently preserved specimens were referred to Palissya sp. Koonwarra by Parris et al. (1995) from the Koonwarra Fossil Bed, Gippsland Basin, Victoria; see Figures 3 (point K), 7 (point A). The host sediments were considered of Aptian age (McLoughlin et al., 2002) and thus ~17 million years younger than the Boola Boola specimens which were assigned to Palissya elegans (as discussed above). Dettmann (1986) identified a diverse spore-pollen assemblage from this site dated as possibly Aptian. The collection site of Palissya specimens from the Aptian of Antarctica (discussed above) is juxtaposed on Figure 7 (point B). The only complete specimen, described by Drinnan and Chambers (1986, p.70, figs. 37-
16 38A), lacks clearly visible adaxial processes. Additional material provided by Parris et al. (1995) is very fragmentary but appears to show adaxial processes. The specimen reported by Seward (1904, pl. 19, figs. 39, 40) as?ginkgoalean female flower, and partially figured by Parris et al. (1995, fig. 8C), is a very poor, almost unidentifiable specimen. Palissya sp. Koonwarra was interpreted by Parris et al. (1995) as possessing helically arranged petiolate sporophylls, each bearing four pairs of adaxial processes. Sporophyll arrangement on the axis is presumably based on the specimen of Drinnan and Chambers (1986) as that is the only specimen which clearly shows the axis. The detail of sporophylls is presumably determined from the fragmentary new material of Parris et al. (1995). Axis detail in the complete specimen is insufficient for attachment to be regarded as convincingly helical and the bilateral symmetry of preserved sporophylls suggests otherwise. [Figure 7] Discussion Parris et al. (1995) noted that the type species of Palissya appears very different from Australasian species. These authors accounted for this difference on the grounds of preservation and shape and placement of processes on the lamina. They reinterpreted the fructification as neither compound nor conifer-like, but possibly ovuliferous, largely from their examination of a European specimen which has been significantly damaged (Parris et al., 1995) since its study by Florin (1958). Given the recent work of Wang (2012), the reinterpretation of Palissya by Parris et al. (1995) cannot be supported. Key features of the Palissya fructification, as listed below, are based upon the descriptions of Nathorst (1908), Florin (1958), Hill (1974), Schweitzer and Kirchner (1996) and Wang (2012). A woody compound cone structure; bracts helically arranged and lanceolate with an acute or obtuse apex;
17 bracts strongly decurrent on the axis and sessile (not petiolate laminae); cup-like investments containing ovules, and attached to an axillary structure closely subtended by a bract. [Table 2] Australasian Palissya specimens (summarised in Table 2) diverge considerably from those from the northern hemisphere and probably represent members of more than one genus. In particular: sporophylls are petiolate laminae in contradistinction to the sessile conifer-like bracts of northern hemisphere species; lamina are distinctly branched helically, bilaterally or possibly even decussate from the main axis and show no indication of being decurrent on the axis; they show no indication of possessing cup-like investments mounted on an ovule scale complex; they are long slender fructifications, apparently delicate, not woody, and likely oriented catkin-like with pendulous sporangia mounted beneath the lamina; general form is not suggestive of conifer affiliation, but rather ginkgoalean, or perhaps pteridospermous. Accordingly, the inclusion of the Australasian material in Palissya and the Palissyaceae cannot be substantiated. Further, Palissya specimens from elsewhere in Gondwana are also unlikely to belong to Palissya. Consequently, Palissya can only be confidently regarded as a conifer from the Rhaetian Lower Jurassic of the northern hemisphere. 3. Systematic palaeobotany
18 Incertae ordinus Knezourocarponaceae Pattemore fam. nov. Diagnosis: Catkin-like fructifications with leaf-like lamina borne on a central axis. Lamina constituting an upper surface beneath which are positioned pendulous sporangia or ovules. Knezourocarpon Pattemore 2000 Type species: Knezourocarpon narangbaensis Pattemore 2000; Toarcian, Landsborough Sandstone, Nambour Basin, Queensland, Australia; by monotypy. Knezourocarpon narangbaensis Pattemore 2000 Figure 8A F. Synonymy: 2000 Knezourocarpon narangbaensis Pattemore, p , figs Caytonia cucullata McLoughlin in Jansson, McLoughlin, Vajda and Pole, p.16, figs. 8A F. Diagnosis: see Pattemore (2000). Discussion: Knezourocarpon was based on numerous specimens and comprises female and male fructifications. Circumscription and comparisons with other genera were provided by Pattemore (2000); the collection location is shown on Figures 3 (point N), 9 (point A) herein. Six fragmentary remains of Caytonia cucullata were described by Jansson et al. (2008b) from siltstone of Inverleigh Quarry, in the Clarence-Moreton Basin; see Figures 3 (point I), 9 (point B). The host deposits were considered to be within the Marburg Subgroup and probably upper Gatton Sandstone (Jansson et al., 2008a,b) and are located stratigraphically below an oolitic marker horizon which is present in a number of southern Queensland basins
19 (McKellar, 1993; Cranfield et al., 1994; Cook et al., 2013); the description of the local geology by Jansson et al. (2008b) suggests a likely age range of latest Pliensbachian middle Toarcian (Cook et al., 2013). Specimens referred to Knezourocarpon narangbaensis by Pattemore (2000) from Toarcian sediments of the Narangba Quarry in the Nambour Basin, were reported to be just above this oolitic marker horizon. The Narangba Quarry sediments were considered to be within the uppermost section of the Landsborough Sandstone (Pattemore, 2000) within the Marburg Subgroup (Cook et al., 2013). [Figure 8] Caytonia cucullata was diagnosed as a suboppositely alternately branched, dorsiventral, pinnate megasporophyll with up to 23 ovuliferous lamina. The female fructification, Caytonia Thomas 1925, is known in considerable detail (Thomas, 1925; Harris, 1932, 1933, 1940; Reymanówna, 1973, 1974; Krassilov, 1977), but ovules were not described in the diagnosis or description of C. cucullata by Jansson et al. (2008b) and ovule insertion proved unclear. No likely associated microsporophyll was reported. Re-examination of the Inverleigh Quarry material assigned to Caytonia. cucullata indicates that lamina attachment was incorrectly reported by Jansson et al. (2008b) and should be regarded as opposite to sub-opposite. The adaxial processes appear in most cases to have been overlapping lamina, probably pendulous leaf-like sporangia. One specimen reported by these authors is re-figured herein (Figure 8A). Jansson et al. (2008b) suggested that their specimens of Caytonia were unlike others of Jurassic age in that they lacked the characteristically club shaped cupules and pedicels. They compared their specimens to those which they considered to have kidney-shaped cupules. Caytonia orientale Krassilov 1964, from the Lower Cretaceous of Russia, is a poor and indistinct specimen; comparision with this specimen is most unconvincing. While some
20 aborted cupules of C. indica Bose and Banerji 1984 are reniform, mature cupules are not, and this cannot be regarded as characterising the species. Bose and Banerji s (1984) specimens comprise a very fragmentary collection of cupules; apparently none are attached to an axis and very few were considered to be ovulate. Both C. orientale and C. indica in no way resemble the Inverleigh Quarry specimens. The specimens referred to Caytonia cucullata by Jansson et al. (2008b) clearly conform to Knezourocarpon narangbaensis in every available megascopic detail. Further, as suggested above, the sediments from the Inverleigh and Narangba quarries are approximately coeval; the Inverleigh Quarry specimens are likely latest Pliensbachian to middle Toarcian in age, while the Narangba specimens are middle Toarcian. This new family is proposed with uncertain higher taxonomic position. Although there are some similarities with the Caytoniaceae Thomas 1925, there are some very significant differences as follows. The Caytonia female fructification was formed from a modified leaf lamina which has curled and deformed sufficiently to form a cupule containing ovules; whereas Knezourocarpon has a separate, apparently unprotected, ovule beneath an otherwise unmodified lamina. Caytonia and Caytonanthus Harris 1937 do not appear to have been catkin-like; whereas both male and female Knezourocarpon fructifications were most likely catkin-like. [Figure 9] The catkin-like arrangement of Knezourocarpon suggests possible ginkgoalean affiliation. While poorly preserved specimens reported as Conifer cone sp. A by Pattemore and Rigby (2005) from the Knezourocarpon type locality may be interpreted as short shoots, it seems most unlikely that these were associated with this genus; the abscission marks are too
21 small to suggest attachment to a Knezourocarpon axis. Further, no ginkgoalean foliage was reported from the Inverleigh or Narangba quarry sites by Pattemore (2000), Pattemore and Rigby (2005) or Jansson et al. (2008b). Knezourocarpon sp. cf. K. narangbaensis Pattemore 2000 Synonymy: 1883 Walchia milneana Tenison-Woods, single specimen, identified as male amentum, p , pl. 6 (fig. 8) Cone of Seed-fern of White (1986), p. 163, fig Cone of Pteridosperm of White (1986), p Palissya sp. Talbragar of Parris et al. (1995), p , fig. 10A C. Discussion: Parris et al. (1995) referred a complete specimen and fragments to Palissya sp. Talbragar, which they described as faint and indistinct. This indifferently preserved specimen from the Talbragar River, New South Wales, was originally figured by White (1986); see Figures 3 (point T1), 9 (point C). The Purlawaugh Formation, as exposed along the Talbragar River in the vicinity of the collection locality, was regarded as Toarcian (Parris et al., 1995; Beattie and Avery, 2012). Parris et al. (1995) suggested a stratigraphic correlation with the nearby Talbragar Fish Beds; however, a recent study dates those beds as Late Jurassic (Beattie and Avery, 2012). The fructification s branching arrangement is unclear but the four sporophylls at the apparently thicker end of the fructification axis appear petiolate and bilaterally opposed. The remainder of the fructification is less well preserved. If the branching arrangement is confirmed then this fructification is likely attributable to Knezourocarpon narangbaensis. Tenison-Woods (1883) assigned conifer-like foliage and co-preserved fructifications, identified as male amentum, to his species Walchia milneana from the Ballimore Hill coal
22 mine, east of Dubbo near the Talbragar River (Feistmantel, 1890); see Figures 3 (point T2), 9 (point C). The Ballimore Coal Measures (now Ballimore Formation) are dated as Early Jurassic; its extent is limited to the Ballimore area where it underlies the Purlawaugh Formation (Dulhunty, 1973; Pogson et al., 1999). Tenison-Woods s (1883) diagnosis of W. milneana was based exclusively on foliage. The fructifications were considered associated with the foliage due to their co-preservation; they were described as distichous with scales bearing comb-like projections. His specimen (pl. 6, fig. 8), is very similar to Knezourocarpon narangbaensis in that: (1) laminae are oppositely arranged; (2) laminae bear adaxial processes which were probably pendulous sporangia. The structure of the fructification suggests that it is not coniferous and therefore unlikely to be associated with the co-preserved foliage. However, the figured fructification shows insufficient detail to confidently refer it to K. narangbaensis. 4. Conclusion Australasian specimens previously referred to Palissya are shown to diverge considerably from the type species and from other northern hemisphere material referred to this genus; none are demonstrably allied with the Palissyaceae. Toarcian specimens are considered attributable or akin to Knezourocarpon narangbaensis, while others probably represent new genera and are likely affiliated with the Knezourocarponaceae Pattemore fam. nov. Palissya specimens from elsewhere in Gondwana are likewise considered doubtful attributions to the genus. Consequently, genuine Palissya specimens are likely restricted to the Rhaetian Lower Jurassic of the northern hemisphere. Knezourocarpon is restricted to the Toarcian and possibly the upper Pliensbachian of eastern Australia. Representatives of the Knezourocarponaceae likely range in age from the Toarcian to the Aptian in Eastern Gondwana. This family is unknown elsewhere. It likely has
23 ginkgoalean or pteridospermous affinity. Acknowledgments This work is part of a research project funded by an Australian Postgraduate Award. Prof. G.E. Webb of the University of Queensland has helped with the establishment and progress of this project. Dr. A. Rozefelds of the Queensland Museum assisted with access to the Museum collection. Comments and suggestions by the editor and reviewers were helpful in improving this contribution. The GPlates software performed the palaeogeographic reconstructions. This software and the reconstruction data were provided by the EarthByte Group, School of Geosciences, The University of Sydney. Other geographic data were supplied by Natural Earth and are in the public domain. References Arber, E.A.N., The earlier Mesozoic floras of New Zealand. New Zealand Geological Survey, Palaeontological Bulletin 6, i ii plates I XIV. Arndt, S., Morphologie und Systematik ausgewählter mesozoischer Koniferen. Palaeontographica Abteilung B 262(1 4), plates 1 6. Bartrum, J.A., Note on the Port Waikato Mesozoic flora. New Zealand Journal of Science and Technology 4, 258. Beattie, R.G., Avery, S., Palaeoecology and palaeoenvironment of the Jurassic
24 Talbragar Fossil Fish Bed, Gulgong, New South Wales, Australia. Alcheringa 36(4), Birkenmajer, K., Ociepa, A.M., Plant-bearing Jurassic strata at Hope Bay, Antarctic Peninsula (West Antarctica): geology and fossil-plant description. Studia Geologica Polonica 128, Bose, M.N., Banerji, J., The fossil floras of Kachchh. I Mesozoic megafossils. Palaeobotanist 33, Bosma, H.F., Kunzmann, L., Kvaček, J., van Konijnenburg-van Cittert, J.H.A., Revision of the genus Cunninghamites (fossil conifers), with special reference to nomenclature, taxonomy and geological age. Review of Palaeobotany and Palynology 182, Braun, C.F.W., Beiträge zur Urgeschichte der Pflanzen, in: Münster, G.G. (Ed.), Beiträge zur Petrefacten-Kunde mit zehn doppelten und vier einfachen, nach der Natur gezeichneten Tafeln, unter mitwirking der Herren Professor Göppert, Professor C. Th. von Siebold und Dr. Braun. Bayreuth, Buchner schen Buchhandlung. 6. pp plates Cantrill, D.J., A Cretaceous (Aptian) flora from President Head, Snow Island, Antarctica. Palaeontographica Abteilung B 253(4 6), plates 1 8. Cook, A.G., Byran, S.E., Draper, J.J., Post-orogenic Mesozoic basins and magmatism, in: Jell, P.A. (Ed.), Geology of Queensland. Geological Survey of Queensland, pp
25 Cranfield, L.C., Carmichael, D.C., Wells, A.T., Ferruginous oolite and associated lithofacies from the Clarence-Moreton and related basins in southeast Queensland, in: Wells, A.T., O Brien, P.E. (Eds.), Geology and petroleum potential of the Clarence-Moreton Basin, New South Wales and Queensland. Australian Geological Survey Organisation, Bulletin 241, pp Delevoryas, T., Hope, R.C., Fertile coniferophyte remains from the Late Triassic Deep River Basin, North Carolina. American Journal of Botany 60(8), Delevoryas, T., Hope, R.C., More evidence for conifer diversity in the Upper Triassic of North Carolina. American Journal of Botany 68(7), Delevoryas, T., Hope, R.C., Further observations on the Late Triassic conifers Compsostrobus neotericus and Voltzia andrewsii. Review of Palaeobotany and Palynology 51, Dettmann, M.E., Early Cretaceous palynoflora of subsurface strata correlative with the Koonwarra Fossil Bed, Victoria, in: Jell, P.A., Roberts, J. (Eds.), Plants and Invertebrates from the Lower Cretaceous Koonwarra Fossil Bed, South Gippsland, Victoria. Association of Australasian Palaeontologists, Memoirs 3, pp Dobruskina, I.A., Triassic floras of Eurasia. Österreichische Akademie der Wissenschaften, Schriftenreihe der Erdwissenschaftlichen Kommissionen 10, Drinnan, A.N., Chambers, T.C., Early Cretaceous plants, Koonwarra, in: Jell, P.A., Roberts, J. (Eds.), Plants and Invertebrates from the Lower Cretaceous Koonwarra Fossil Bed,
26 South Gippsland, Victoria. Association of Australasian Palaeontologists, Memoirs 3, pp Dulhunty, J.A., Mesozoic stratigraphy in central western New South Wales. Journal of the Geological Society of Australia 20(3), Edwards, W.N., V. Jurassic plants from New Zealand. Journal of Natural History (10)13, Endlicher, S., Synopsis coniferarum. Sangalli, apud Scheitlin & Zollikofer. Escapa, I.H., Sterli, J., Pol, D., Nicoli, L., Jurassic tetrapods and flora of Cañadón Asfalto Formation in Cerro Cóndor area, Chubut Province. Revista de la Asociación Geológica Argentina 63(4), Feistmantel, O., 1876a. Fossil flora of the Gondwana system. Part 1: Jurassic (Oolitic) flora of the Kach. Memoirs of the Geological Survey of India, Palaeontologia Indica (XI)2, i iv plates I XII. Feistmantel, O., 1876b. Notes on the age of some fossil floras of India. VI, VII and VIII. Records of the Geological Survey of India 9(4), Feistmantel, O., 1877a. 2. Jurassic (Liassic) flora of the Rajmahal group in the Rajmahal Hills. Memoirs of the Geological Survey of India, Palaeontologia Indica (II)1, i iii + i ii (refs.) (also numbered ) + plates XXXVI XLVIII.
27 Feistmantel, O., 1877b. 3. Jurassic (Liassic) flora of the Rajmahal group from Golapili (near Ellore), south Godavari district. Memoirs of the Geological Survey of India, Palaeontologia Indica (II)1, 1 28 (also numbered ) + plates I VIII. Feistmantel, O., 1877c. Fossil flora of the Gondwana system. Part 2: Flora of the Jabulpur group (Upper Gondwanas) in the Son-Narbada region. Memoirs of the Geological Survey of India, Palaeontologia Indica (XI)2, 1 25 (also numbered ) + plates I XIV + index ( ). Feistmantel, O., Outliers on the Madras coast. Memoirs of the Geological Survey of India, Palaeontologia Indica (II)1, i iv (also numbered ) + plates I XVI. Feistmantel, O., Fossil flora of the Gondwana system. Preface. Memoirs of the Geological Survey of India, Palaeontologia Indica (XI)2, i xl. Feistmantel, O., Geological and palaeontological relations of the coal and plant-bearing beds of Palaeozoic and Mesozoic age in eastern Australia and Tasmania. Memoirs of the Geological Survey of New South Wales - Palaeontology No. 3, plates I XXX. Florin, R., Die Koniferen des Oberkarbons und des unteren Perms, siebentes heft. Palaeontographica Abteilung B 85(7), plates CLXXIII CLXXXVI. Florin, R., Evolution in cordaites and conifers. Acta Horti Bergiani 15(11),
28 Florin, R., The female reproductive organs of conifers and taxads. Biological Reviews 29, Florin, R., On Jurassic taxads and conifers from north-western Europe and eastern Greenland. Acta Horti Bergiani 17(10), plates Florin, R., The distribution of conifer and taxad genera in time and space. Acta Horti Bergiani 20(4), Florin, R., The distribution of conifer and taxad genera in time and space: Additions and corrections. Acta Horti Bergiani 20(5), Frenguelli, J., Addenda a la flora del Gondwana Superior en la Argentina. Physis. Revista de la Asociación Argentina de Ciencias Naturales 20(57), Harris, T.M., The fossil flora of Scoresby Sound, east Greenland. Part 3: Caytoniales and Bennettitales. Meddelelser om Grønland 85(5), plates Harris, T.M., A new member of the Caytoniales. New Phytologist 32(2), Harris, T.M., The fossil flora of Scoresby Sound, east Greenland. Part 4. Ekspeditionerne til Scoresbysund distriktet udført i Aarene 1934 og Under ledelse af A. Rosenkrantz. Meddelelser om Grønland 112(1), plates Harris, T.M., The fossil flora of Scoresby Sound, east Greenland. Part 5. Stratigraphic
29 relations of the plant beds. Meddelelser om Grønland 112(2), plate 1 + tables 1 3. Harris, T.M., Caytonia. Annals of Botany 4(16), Hector, J., Indian and colonial exhibition, London, Detailed catalogue and guide to the geological exhibits, in: Handbook of New Zealand. 4 ed.. George Didsbury, Government Printer, Wellington, pp map + appendix. Hill, C.R., Palaeobotanical and sedimentological studies in the lower Bajocian (Middle Jurassic) flora of Yorkshire. PhD thesis. University of Leeds, Leeds, 281 pp. Hirmer, M., Die Blüten der Coniferen. Teil 1: Entwicklungsgeschichte und vergleichende Morphologie des weibliche Blütehzapfens der Coniferen. Bibliotheca Botanica XXVIII(114), plates I VII. Jansson, I.M., McLoughlin, S., Vajda, V., 2008a. Early Jurassic annelid cocoons from eastern Australia. Alcheringa 32(3), Jansson, I.M., McLoughlin, S., Vajda, V., Pole, M., 2008b. An Early Jurassic flora from the Clarence-Moreton Basin, Australia. Review of Palaeobotany and Palynology 150, Krassilov, V.A., New Lower Cretaceous gymnosperms of Primorye. Palaeontological Journal 1964(1), [in Russian]. Krassilov, V.A., Contributions to the knowledge of the Caytoniales. Review of
30 Palaeobotany and Palynology 24, Lakhanpal, R.N., Maheshwari, H.K., Awasthi, N., A catalogue of Indian fossil plants Covering all available records from 1821 to Birbal Sahni Institute of Palaeobotany, Lucknow, India. McKellar, J.L., Stratigraphic relationships in the Nambour Basin, southeastern Queensland. Queensland Geology 5, enclosure 1. McLoughlin, S., Drinnan, A.N., A Middle Jurassic flora from the Walloon Coal Measures, Mutdapilly, Queensland, Australia. Memoirs of the Queensland Museum 38, McLoughlin, S., Tosolini, A.M.P., Nagalingum, N.S., Drinnan, A.N., Early Cretaceous (Neocomian) flora and fauna of the Lower Strzelecki Group, Gippsland Basin, Victoria. Association of Australasian Palaeontologists, Memoir 26, Meyen, S.V., Basic features of gymnosperm systematics and phylogeny as evidenced by the fossil record. The Botanical Review 50(1), Miller, C.N., Implications of fossil conifers for the phylogenetic relationships of living families. The Botanical Review 65(3), Nathorst, A.G., Om floran i Skånes kolförande bildningar. I Florin vid Bjuf ( ). Sveriges Geologiska Undersökning. Series C 85, plates 1 26.
31 Nathorst, A.G., Paläobotanische mitteilungen 7. Über Palissya, Stachyotaxus und Palaeotaxus. Kungliga Svenska Vetenskapsakademiens Handlingar 43(8), plates 1 3. Niklas, K.J., The aerodynamics of wind pollination. The Botanical Review 51(3), Parris, K.M., Drinnan, A.N., Cantrill, D.J., Palissya cones from the Mesozoic of Australia and New Zealand. Alcheringa 19(2), Pattemore, G.A., A new Early Jurassic pteridosperm fructification from Queensland. Journal of African Earth Sciences 31(1), Pattemore, G.A., Rigby, J.F., Fructifications and foliage from the Mesozoic of southeast Queensland. Memoirs of the Queensland Museum 50, Pogson, D.J., Cameron, R.G., Meakin, N.S., Raymond, O.L., Barron, L.M., Henderson, G.A.M., Warren, A.Y.E., Morgan, E.J., Surat Basin, in: Facer, R.A., Stewart, J.R. (Eds.), Explanatory notes. Dubbo 1: geological sheet SI/ ed.. Geological Survey of New South Wales and Australian Geological Survey Organisation, pp Pole, M.S., Repeated flood events and fossil forests at Curio Bay (Middle Jurassic), New Zealand. Sedimentary Geology 144(3 4), Pole, M.S., Early - Middle Jurassic stratigraphy of the Fortrose - Chaslands region,
32 southernmost South Island, New Zealand. New Zealand Journal of Geology and Geophysics 47(1), Pole, M.S., Vegetation and climate of the New Zealand Jurassic. GFF (Journal of the Geological Society of Sweden) 131(1 2), Pott, C., McLoughlin, S., The Rhaetian flora of Rögla, northern Scania, Sweden. Palaeontology 54(5), Reymanówna, M., The Jurassic flora from Grojec near Kraków in Poland. pt.2. Caytoniales and anatomy of Caytonia. Acta Palaeobotanica 14(2), plates I V. Reymanówna, M., On anatomy and morphology of Caytonia, in: Symposium on Morphological and Stratigraphical Palaeobotany. Birbal Sahni Institute of Palaeobotany. Special Publication No. 2, pp Schenk, A., Die fossile Flora der Grenzschichten des Keupers und Lias Frankens. C.W. Kreidel. Schenk, A., Fossile Pflanzen aus der Albourskette, gesammelt von E. Tietze, Chefgeologen der k. k. geologischen Reichsanstalt. Bibliotheca Botanica 6, plates I IX. Schweitzer, H.J., Kirchner, M., Die Rhäto Jurassischen Floren des Iran und Afghanistans: 9. Coniferophyta. Palaeontographica Abteilung B 238(4 6), plates 1 16.
33 Seton, M., Müller, R., Zahirovic, S., Gaina, C., Torsvik, T., Shephard, G., Talsma, A., Gurnis, M., Turner, M., Maus, S., et al., Global continental and ocean basin reconstructions since 200 Ma. Earth-Science Reviews 113(3), Seward, A.C., On a collection of Jurassic plants from Victoria. Records of the Geological Survey of Victoria 1(3), Seward, A.C., Fossil Plants. Vol.4. Ginkgoales, Coniferales, Gnetales. Cambridge University Press, London. Stewart, W.N., Rothwell, G.W., Paleobotany and the evolution of plants. 2nd ed., Cambridge University Press. Taylor, E.L., Taylor, T.N., Krings, M., Paleobotany: the biology and evolution of fossil plants. 2nd ed., Elsevier. Tenison-Woods, J.E., On the fossil flora of the coal deposits of Australia. Proceedings of the Linnean Society of New South Wales (1)8, plates 1 10A. Thomas, H.H., The Caytoniales, a new group of angiospermous plants from the Jurassic rocks of Yorkshire. Philosophical Transactions of the Royal Society of London Series B, 213, Thorn, V., Enigmatic high palaeolatitude forests of Gondwanaland a case study from
34 the New Zealand Jurassic. PhD thesis. School of Geography, Environment and Earth Sciences, Victoria University of Wellington. 2 Vols., 610pp plates + 10 enclosures. Thorn, V., Vegetation communities of a high palaeolatitude Middle Jurassic forest in New Zealand. Palaeogeography, Palaeoclimatology, Palaeoecology 168(3 4), Tosolini, A.M.P., McLoughlin, S., Drinnan, A.N., Stratigraphy and fluvial sedimentary facies of the Neocomian lower Strzelecki Group, Gippsland Basin, Victoria. Australian Journal of Earth Sciences 46(6), Turner, S., Bean, L.B., Dettmann, M., McKellar, J.L., McLoughlin, S., Thulborn, T., Australian Jurassic sedimentary and fossil successions: current work and future prospects for marine and non-marine correlation. GFF (Journal of the Geological Society of Sweden) 131(1 2), Wang, Z., A bizarre Palissya ovulate organ from Upper Triassic strata of the Zixing coal field, Hunan Province, China. Chinese Science Bulletin 57(10), White, M.E., Greening of Gondwana: The 400 million year story of Australia s plants. Reed Australia, Frenchs Forest, New South Wales, Australia.

35 Figure 1
36 Figure 2
37 Figure 3

38 Figure 4
39 Figure 5
40 Figure 6

41 Figure 7

42 Figure 8 AC CE P TE D MA NU SC RI PT

43 Figure 9
Emendation of Sorosaccus gracilis Harris 1935, a gymnospermous pollen cone 1, 2 LIU Xiu-Qun 3 Francis M. HUEBER 1 LI Cheng-Sen * 1 WANG Yu-Fei
 43 2 : 182 190 2005 Acta Phytotaxonomica Sinica Emendation of Sorosaccus gracilis Harris 1935, a gymnospermous pollen cone 1, 2 LIU Xiu-Qun 3 Francis M. HUEBER 1 LI Cheng-Sen * 1 WANG Yu-Fei 1 (Laboratory
43 2 : 182 190 2005 Acta Phytotaxonomica Sinica Emendation of Sorosaccus gracilis Harris 1935, a gymnospermous pollen cone 1, 2 LIU Xiu-Qun 3 Francis M. HUEBER 1 LI Cheng-Sen * 1 WANG Yu-Fei 1 (Laboratory
Laboratory 8: Ginkgo, Cycads, and Gnetophytes
 IB 168 Plant Systematics Laboratory 8: Ginkgo, Cycads, and Gnetophytes This is the third and final lab concerning the gymnosperms. Today we are looking at Ginkgo, the Cycads, and the Gnetophytes, the so-called
IB 168 Plant Systematics Laboratory 8: Ginkgo, Cycads, and Gnetophytes This is the third and final lab concerning the gymnosperms. Today we are looking at Ginkgo, the Cycads, and the Gnetophytes, the so-called
[92] THE NEW EVIDENCE OF ISOSPORY IN PALAEOZOIC SEED PLANTS
![[92] THE NEW EVIDENCE OF ISOSPORY IN PALAEOZOIC SEED PLANTS](/thumbs/92/110747357.jpg "[92] THE NEW EVIDENCE OF ISOSPORY IN PALAEOZOIC SEED PLANTS") [92] THE NEW EVIDENCE OF ISOSPORY IN PALAEOZOIC SEED PLANTS BY M. BENSON (With 4 figures in the text) AN intensive study of some sections of the Palaeozoic fructifica- J\ tion Schnetzia Bennieana Kidston,
[92] THE NEW EVIDENCE OF ISOSPORY IN PALAEOZOIC SEED PLANTS BY M. BENSON (With 4 figures in the text) AN intensive study of some sections of the Palaeozoic fructifica- J\ tion Schnetzia Bennieana Kidston,
Glossopterids: Suzerains of Geologically Past Indian Forests
 Glossopterids: Suzerains of Geologically Past Indian Forests Dipanjan Ghosh Dipanjan Ghosh is doing his PhD in Microbiology at the Department of Botany, University of Burdwan. His research interests include:
Glossopterids: Suzerains of Geologically Past Indian Forests Dipanjan Ghosh Dipanjan Ghosh is doing his PhD in Microbiology at the Department of Botany, University of Burdwan. His research interests include:
Adjustments to Life in Air
 1 Adjustments to Life in Air Plants and Animals no buoyancy, need skeletal support desiccation physical-chemical environmental extremes (temperature, humidity, salinity) no nutrients in air reproductive
1 Adjustments to Life in Air Plants and Animals no buoyancy, need skeletal support desiccation physical-chemical environmental extremes (temperature, humidity, salinity) no nutrients in air reproductive
Jurassic liverworts from Mount Flora, Hope Bay, Antarctic Peninsula
 vol. 28, no. 1, pp. 31 36, 2007 Jurassic liverworts from Mount Flora, Hope Bay, Antarctic Peninsula Anna Maria OCIEPA Instytut Botaniki, Uniwersytet Jagielloński, Kopernika 27, 31 501 Kraków, Poland
vol. 28, no. 1, pp. 31 36, 2007 Jurassic liverworts from Mount Flora, Hope Bay, Antarctic Peninsula Anna Maria OCIEPA Instytut Botaniki, Uniwersytet Jagielloński, Kopernika 27, 31 501 Kraków, Poland
Plan of Development Mountain Valley Pipeline Project. APPENDIX P Plan for Unanticipated Discovery of Paleontological Resources
 APPENDIX P Plan for Unanticipated Discovery of Paleontological Resources Appendix P Plan for Unanticipated Discovery of Paleontological Resources Prepared by: TABLE OF CONTENTS 1.0 INTRODUCTION... 1 1.1
APPENDIX P Plan for Unanticipated Discovery of Paleontological Resources Appendix P Plan for Unanticipated Discovery of Paleontological Resources Prepared by: TABLE OF CONTENTS 1.0 INTRODUCTION... 1 1.1
EPIDERMAL STRUCTURE AND DEVELOPMENT OF STOMATA IN EPHEDRA FOLIATA BOISS.
 EPIDERMAL STRUCTURE AND DEVELOPMENT OF STOMATA IN EPHEDRA FOLIATA BOISS. BY D. D. PANT AND BHARATI MEHRA Department of Botany, The University, Allahabad, India {Received z August 1963) SUMMARY The epidermal
EPIDERMAL STRUCTURE AND DEVELOPMENT OF STOMATA IN EPHEDRA FOLIATA BOISS. BY D. D. PANT AND BHARATI MEHRA Department of Botany, The University, Allahabad, India {Received z August 1963) SUMMARY The epidermal
3. The diagram below shows how scientists think some of Earth's continents were joined together in the geologic past.
 1. The map below shows the present-day locations of South America and Africa. Remains of Mesosaurus, an extinct freshwater reptile, have been found in similarly aged bedrock formed from lake sediments
1. The map below shows the present-day locations of South America and Africa. Remains of Mesosaurus, an extinct freshwater reptile, have been found in similarly aged bedrock formed from lake sediments
Geology of the Batemans Bay region. Geological evolution. The Lachlan Orogen
 Australian Journal of Earth Sciences 1 The word orogen is derived from the ancient Greek language word for mountain building. The Lachlan Orogen The rocks exposed in the Batemans Bay are part of the geological
Australian Journal of Earth Sciences 1 The word orogen is derived from the ancient Greek language word for mountain building. The Lachlan Orogen The rocks exposed in the Batemans Bay are part of the geological
MUHAMMAD S TAMANNAI, DOUGLAS WINSTONE, IAN DEIGHTON & PETER CONN, TGS Nopec Geological Products and Services, London, United Kingdom
 Geological and Geophysical Evaluation of Offshore Morondava Frontier Basin based on Satellite Gravity, Well and regional 2D Seismic Data Interpretation MUHAMMAD S TAMANNAI, DOUGLAS WINSTONE, IAN DEIGHTON
Geological and Geophysical Evaluation of Offshore Morondava Frontier Basin based on Satellite Gravity, Well and regional 2D Seismic Data Interpretation MUHAMMAD S TAMANNAI, DOUGLAS WINSTONE, IAN DEIGHTON
PLATE TECTONICS THEORY
 PLATE TECTONICS THEORY Continental drift Sea floor spreading CONTINENTAL DRIFT CONTINENTAL DRIFT 1. The fitness of continents and Continental Reconstruction Earth ~200 million years ago 1.1 Geometrical
PLATE TECTONICS THEORY Continental drift Sea floor spreading CONTINENTAL DRIFT CONTINENTAL DRIFT 1. The fitness of continents and Continental Reconstruction Earth ~200 million years ago 1.1 Geometrical
ESTIMATION OF CONSERVATISM OF CHARACTERS BY CONSTANCY WITHIN BIOLOGICAL POPULATIONS
 ESTIMATION OF CONSERVATISM OF CHARACTERS BY CONSTANCY WITHIN BIOLOGICAL POPULATIONS JAMES S. FARRIS Museum of Zoology, The University of Michigan, Ann Arbor Accepted March 30, 1966 The concept of conservatism
ESTIMATION OF CONSERVATISM OF CHARACTERS BY CONSTANCY WITHIN BIOLOGICAL POPULATIONS JAMES S. FARRIS Museum of Zoology, The University of Michigan, Ann Arbor Accepted March 30, 1966 The concept of conservatism
ASPECTS OF THE GEOLOGY OF UPPERMOST JURASSIC STRATA AT PORT WAIKATO. by J.A. Kenny, T.C. Leonard, G.P. O'Leary and S.S. Palmer INTRODUCTION
 TANE 23, 1977 ASPECTS OF THE GEOLOGY OF UPPERMOST JURASSIC STRATA AT PORT WAIKATO by J.A. Kenny, T.C. Leonard, G.P. O'Leary and S.S. Palmer Department of Geology, University of Auckland, Private Bag, Auckland
TANE 23, 1977 ASPECTS OF THE GEOLOGY OF UPPERMOST JURASSIC STRATA AT PORT WAIKATO by J.A. Kenny, T.C. Leonard, G.P. O'Leary and S.S. Palmer Department of Geology, University of Auckland, Private Bag, Auckland
Ediacaria and Hiemalora - Disc-like Attachments. of Benthic Organisms from the White Sea and. Arctic Siberia (Russia)
") Ediacaria and Hiemalora - Disc-like Attachments of Benthic Organisms from the White Sea and Arctic Siberia (Russia) E. A. Serezhnikova Paleontological Institute, Russian Academy of Sciences, 123 Profsoyuznaya,
Ediacaria and Hiemalora - Disc-like Attachments of Benthic Organisms from the White Sea and Arctic Siberia (Russia) E. A. Serezhnikova Paleontological Institute, Russian Academy of Sciences, 123 Profsoyuznaya,
Robert S. Sawin Stratigraphic Research Section Kansas Geological Survey. Open-File Report
 Copies of Archived Correspondence Pertinent to the Decision to Classify the Exposure at Point of Rocks, Morton County, Kansas, as Jurassic by the Kansas Geological Survey in 1967 Robert S. Sawin Stratigraphic
Copies of Archived Correspondence Pertinent to the Decision to Classify the Exposure at Point of Rocks, Morton County, Kansas, as Jurassic by the Kansas Geological Survey in 1967 Robert S. Sawin Stratigraphic
A new species of Wiffiumsoniu from the Rajmahal Hills, India
 J.Linn.80~. (Bot.),61,384,pp. 121-127 With 2 plates and 3 led-figures Printed in Great Britain July, 1968 A new species of Wiffiumsoniu from the Rajmahal Hills, India BY M. N. BOSE Birbal Sahni Institute
J.Linn.80~. (Bot.),61,384,pp. 121-127 With 2 plates and 3 led-figures Printed in Great Britain July, 1968 A new species of Wiffiumsoniu from the Rajmahal Hills, India BY M. N. BOSE Birbal Sahni Institute
The structure of umkomasiacean fructifications from the Triassic of Queensland
 Acta Palaeobotanica 56(1): 17 40, 2016 DOI: 10.1515/acpa-2016-0001 The structure of umkomasiacean fructifications from the Triassic of Queensland GARY A. PATTEMORE School of Earth Sciences, The University
Acta Palaeobotanica 56(1): 17 40, 2016 DOI: 10.1515/acpa-2016-0001 The structure of umkomasiacean fructifications from the Triassic of Queensland GARY A. PATTEMORE School of Earth Sciences, The University
Structural Geology of the Mountains
 Structural Geology of the Mountains Clinton R. Tippett Shell Canada Limited, Calgary, Alberta clinton.tippett@shell.ca INTRODUCTION The Southern Rocky Mountains of Canada (Figure 1) are made up of several
Structural Geology of the Mountains Clinton R. Tippett Shell Canada Limited, Calgary, Alberta clinton.tippett@shell.ca INTRODUCTION The Southern Rocky Mountains of Canada (Figure 1) are made up of several
[279] A NOTE ON THE ORIGIN OF LATERAL ROOTS AND THE STRUCTURE OF THE ROOT-APEX OF LYGINOPTERIS OLDHAMIA
![[279] A NOTE ON THE ORIGIN OF LATERAL ROOTS AND THE STRUCTURE OF THE ROOT-APEX OF LYGINOPTERIS OLDHAMIA](/thumbs/83/87080534.jpg "[279] A NOTE ON THE ORIGIN OF LATERAL ROOTS AND THE STRUCTURE OF THE ROOT-APEX OF LYGINOPTERIS OLDHAMIA") [279] A NOTE ON THE ORIGIN OF LATERAL ROOTS AND THE STRUCTURE OF THE ROOT-APEX OF LYGINOPTERIS OLDHAMIA BY A. C. HALKET (With Plate XI and i figure in the text) E 'GlNOPTERis oi.dh.imi.i, a plant of the
[279] A NOTE ON THE ORIGIN OF LATERAL ROOTS AND THE STRUCTURE OF THE ROOT-APEX OF LYGINOPTERIS OLDHAMIA BY A. C. HALKET (With Plate XI and i figure in the text) E 'GlNOPTERis oi.dh.imi.i, a plant of the
PALMS IN THE UPPER PALEOCENE OF THE RANCHERÍA RIVER
 PALMS IN THE UPPER PALEOCENE OF THE RANCHERÍA RIVER VALLEY (GUAJIRA DEPARTMENT, COLOMBIA) CAROLINA GÓMEZ NAVARRO Instituto de Biología, Universidad de Antioquia (Medellín, Colombia) email: karogomez@yahoo.com
PALMS IN THE UPPER PALEOCENE OF THE RANCHERÍA RIVER VALLEY (GUAJIRA DEPARTMENT, COLOMBIA) CAROLINA GÓMEZ NAVARRO Instituto de Biología, Universidad de Antioquia (Medellín, Colombia) email: karogomez@yahoo.com
NOTES ON GINKGO BILOBA'
 NOTES ON GINKGO BILOBA' WALTER WV. TUPPER (WITH PLATE xx) Among the gymnosperms, one of the groups most interesting from a morphological standpoint is the Ginkgoales, the only living representative of
NOTES ON GINKGO BILOBA' WALTER WV. TUPPER (WITH PLATE xx) Among the gymnosperms, one of the groups most interesting from a morphological standpoint is the Ginkgoales, the only living representative of
GeoConvention 2012: Vision 1
 Cretaceous possibilities: Sedimentology and reservoir potential of the Early Cretaceous Isachsen Formation of the Sverdrup Basin, Ellef Ringnes Island, Arctic Canada Dylan Tullius Department of Geoscience
Cretaceous possibilities: Sedimentology and reservoir potential of the Early Cretaceous Isachsen Formation of the Sverdrup Basin, Ellef Ringnes Island, Arctic Canada Dylan Tullius Department of Geoscience
EROMANGA BASIN SYMPOSIUM JOINTLY SPONSORED BY THE GEOLOGICAL SOCIETY OF AUSTRALIA AND THE PETROLEUM EXPLORATION SOCIETY OF AUSTRALIA (S.A.
 EROMANGA BASIN SYMPOSIUM JOINTLY SPONSORED BY THE GEOLOGICAL SOCIETY OF AUSTRALIA AND THE PETROLEUM EXPLORATION SOCIETY OF AUSTRALIA (S.A. BRANCHES) ABSTRACT VOLUME EDITED BY P.S. MOORE & T.J. MOUNT GEOLOGICAL
EROMANGA BASIN SYMPOSIUM JOINTLY SPONSORED BY THE GEOLOGICAL SOCIETY OF AUSTRALIA AND THE PETROLEUM EXPLORATION SOCIETY OF AUSTRALIA (S.A. BRANCHES) ABSTRACT VOLUME EDITED BY P.S. MOORE & T.J. MOUNT GEOLOGICAL
NATHALIE S. NAGALINGUM 1, ANDREW N. DRINNAN 1 & STEPHEN MCLOUGHLIN 2
 RSV 117-2 proceedings FINAL 3/5/06 3:07 PM Page 295 A NEW FOSSIL CONIFER, BELLARINEA RICHARDSII, FROM THE EARLY CRETACEOUS STRZELECKI GROUP, SOUTHEASTERN VICTORIA NATHALIE S. NAGALINGUM 1, ANDREW N. DRINNAN
RSV 117-2 proceedings FINAL 3/5/06 3:07 PM Page 295 A NEW FOSSIL CONIFER, BELLARINEA RICHARDSII, FROM THE EARLY CRETACEOUS STRZELECKI GROUP, SOUTHEASTERN VICTORIA NATHALIE S. NAGALINGUM 1, ANDREW N. DRINNAN
Petroleum geology framework, West Coast offshore region
 Petroleum geology framework, West Coast offshore region James W. Haggart* Geological Survey of Canada, Vancouver, BC jhaggart@nrcan.gc.ca James R. Dietrich Geological Survey of Canada, Calgary, AB and
Petroleum geology framework, West Coast offshore region James W. Haggart* Geological Survey of Canada, Vancouver, BC jhaggart@nrcan.gc.ca James R. Dietrich Geological Survey of Canada, Calgary, AB and
Meandering Miocene Deep Sea Channel Systems Offshore Congo, West Africa
 Meandering Miocene Deep Sea Channel Systems Offshore Congo, West Africa S. Baer* (PGS), J. E. Comstock (PGS), K. Vrålstad (PGS), R. Borsato (PGS), M. Martin (PGS), J.P. Saba (SNPC), B. Débi-Obambé (SNPC)
Meandering Miocene Deep Sea Channel Systems Offshore Congo, West Africa S. Baer* (PGS), J. E. Comstock (PGS), K. Vrålstad (PGS), R. Borsato (PGS), M. Martin (PGS), J.P. Saba (SNPC), B. Débi-Obambé (SNPC)
The puzzle presented by the famous stumps of Gilboa, New York, finds a solution in the
 PALAEOBOTANY A tree without leaves Brigitte Meyer-Berthaud and Anne-Laure Decombeix The puzzle presented by the famous stumps of Gilboa, New York, finds a solution in the discovery of two fossil specimens
PALAEOBOTANY A tree without leaves Brigitte Meyer-Berthaud and Anne-Laure Decombeix The puzzle presented by the famous stumps of Gilboa, New York, finds a solution in the discovery of two fossil specimens
EARLY CRETACEOUS FLORAL DIVERSITY AND ECOLOGY IN THE PRANHITA-GODAVARI BASIN, EAST COAST OF INDIA
 FLORAL DIVERSITY AND ECOLOGY DURING EARLY CRETACEOUS IN THE PRANHITA-GODAVARI BASIN 189 Journal of the Palaeontological Society of India ISSN 0522-9630 Volume 61(2), December 31, 2016: 189-214 EARLY CRETACEOUS
FLORAL DIVERSITY AND ECOLOGY DURING EARLY CRETACEOUS IN THE PRANHITA-GODAVARI BASIN 189 Journal of the Palaeontological Society of India ISSN 0522-9630 Volume 61(2), December 31, 2016: 189-214 EARLY CRETACEOUS
Also examine a pollen cone from the genus Emporia (= Lebachia). How does this compare to that of Cordaianthus?
. How does this compare to that of Cordaianthus?") PBIO 3080/5080 STRUCTURAL BOTANY FALL 2014 LAB 10: CORDAITES & CONIFERS ORDER CORDAITALES Cordaites are an important group of Carboniferous plants that are closely related to conifers. Rudolf Florin (1951,
PBIO 3080/5080 STRUCTURAL BOTANY FALL 2014 LAB 10: CORDAITES & CONIFERS ORDER CORDAITALES Cordaites are an important group of Carboniferous plants that are closely related to conifers. Rudolf Florin (1951,
PALEONTOLOGICAL CONTRIBUTIONS
 THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS August 18, 1978 Paper 94 SPHENOPTERIS SELLARDSII, A PROBLEMATICAL PTERIDOSPERM FROM THE PERMIAN OF KANSAS' ROBERT W. BAXTER University of Kansas,
THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS August 18, 1978 Paper 94 SPHENOPTERIS SELLARDSII, A PROBLEMATICAL PTERIDOSPERM FROM THE PERMIAN OF KANSAS' ROBERT W. BAXTER University of Kansas,
Leaf Identification Kit
 Introduction Leaf Identification Kit Catalog No. FB0490 Publication No. 10673 Leaves can be found in a wide variety of sizes, shapes and colors. Each species of tree produces its own variation of leaf.
Introduction Leaf Identification Kit Catalog No. FB0490 Publication No. 10673 Leaves can be found in a wide variety of sizes, shapes and colors. Each species of tree produces its own variation of leaf.
GCE A level 1214/01 GEOLOGY GL4 Interpreting the Geological Record
 Surname Centre Number Candidate Number Other Names 2 GCE A level 1214/01 GEOLOGY GL4 Interpreting the Geological Record P.M. MONDAY, 6 June 2016 2 hours S16-1214-01 For s use Question Maximum Mark Mark
Surname Centre Number Candidate Number Other Names 2 GCE A level 1214/01 GEOLOGY GL4 Interpreting the Geological Record P.M. MONDAY, 6 June 2016 2 hours S16-1214-01 For s use Question Maximum Mark Mark
Structural Styles and Geotectonic Elements in Northwestern Mississippi: Interpreted from Gravity, Magnetic, and Proprietary 2D Seismic Data
 Structural Styles and Geotectonic Elements in Northwestern Mississippi: Interpreted from Gravity, Magnetic, and Proprietary 2D Seismic Data Nick Loundagin 1 and Gary L. Kinsland 2 1 6573 W. Euclid Pl.,
Structural Styles and Geotectonic Elements in Northwestern Mississippi: Interpreted from Gravity, Magnetic, and Proprietary 2D Seismic Data Nick Loundagin 1 and Gary L. Kinsland 2 1 6573 W. Euclid Pl.,
Relinquishment Report
 Relinquishment Report Licence P1403 Block 13/22d Chevron North Sea Limited Korean National Oil Company Chevron North Sea Limited December 2009 1 SYNOPSIS... 3 2 INTRODUCTION. 3 2.1 Licence Terms.. 3 2.2
Relinquishment Report Licence P1403 Block 13/22d Chevron North Sea Limited Korean National Oil Company Chevron North Sea Limited December 2009 1 SYNOPSIS... 3 2 INTRODUCTION. 3 2.1 Licence Terms.. 3 2.2
2. PHYSICAL FEATURES OF INDIA
 2. PHYSICAL FEATURES OF INDIA Question 1: Choose the right answer from the four alternatives given below. (i) A landmass bound by sea on three sides is referred to as (a) Coast (c) Peninsula (b) Island
2. PHYSICAL FEATURES OF INDIA Question 1: Choose the right answer from the four alternatives given below. (i) A landmass bound by sea on three sides is referred to as (a) Coast (c) Peninsula (b) Island
Phanerozoic (last 0.54 by) Tectonics Climate Life
 Tectonics Climate Life") Phanerozoic (last 0.54 by) Tectonics Climate Life Tools for Locating Past Continent Positions Fossils depending on climate Alignment of geological features Geometrical fit of continental margins Similarity
Phanerozoic (last 0.54 by) Tectonics Climate Life Tools for Locating Past Continent Positions Fossils depending on climate Alignment of geological features Geometrical fit of continental margins Similarity
SPECIALIST GEOLOGY SERVICES
 SPECIALIST GEOLOGY SERVICES rpsgroup.com/energy RPS ENERGY an independent global energy consultancy RPS Energy is part of RPS Group which has been listed on the main London Stock Exchange since 1995. It
SPECIALIST GEOLOGY SERVICES rpsgroup.com/energy RPS ENERGY an independent global energy consultancy RPS Energy is part of RPS Group which has been listed on the main London Stock Exchange since 1995. It
OBSERVATIONS ON FOSSIL PLANTS FROM THE DEVONIAN OF EASTERN NORTH AMERICA
 CONTRIBUTIONS F'ROM THE MUSEUM OF PAI;EONTOLOGY UNIVERSITY OF MICHIGAN VOL VI, No. 3, pp. 63-57 (1 PI.) OCIOBEB 1,1941 OBSERVATIONS ON FOSSIL PLANTS FROM THE DEVONIAN OF EASTERN NORTH AMERICA V. RYENIA
CONTRIBUTIONS F'ROM THE MUSEUM OF PAI;EONTOLOGY UNIVERSITY OF MICHIGAN VOL VI, No. 3, pp. 63-57 (1 PI.) OCIOBEB 1,1941 OBSERVATIONS ON FOSSIL PLANTS FROM THE DEVONIAN OF EASTERN NORTH AMERICA V. RYENIA
Before Plate Tectonics: Theory of Continental Drift
 Before Plate Tectonics: Theory of Continental Drift Predecessor to modern plate tectonics Shape and fit of the continents was the initial evidence Snider-Pelligrini (1858) Taylor (1908) Wegner (1915) Fig.
Before Plate Tectonics: Theory of Continental Drift Predecessor to modern plate tectonics Shape and fit of the continents was the initial evidence Snider-Pelligrini (1858) Taylor (1908) Wegner (1915) Fig.
Name: Lab section? 1:30 or 3:30 (circle one) FOR 320 Spring 2012 First Hourly Exam Feb 10, 2012
 FOR 320 Spring 2012 First Hourly Exam Feb 10, 2012") Name: Lab section? 1:30 or 3:30 (circle one) FOR 320 Spring 2012 First Hourly Exam Feb 10, 2012 1) (27 pts) On the left are all of the species that we covered in this section of the course. Match the letter
Name: Lab section? 1:30 or 3:30 (circle one) FOR 320 Spring 2012 First Hourly Exam Feb 10, 2012 1) (27 pts) On the left are all of the species that we covered in this section of the course. Match the letter
WEIGHING THE EVIDENCE FOR A MASS EXTINCTION PART 2: ON LAND
 WEIGHING THE EVIDENCE FOR A MASS EXTINCTION PART 2: ON LAND LESSON INTRODUCTION TEACHER MATERIALS The iridium-rich clay layer at the boundary between the Cretaceous (K) and Tertiary (T) periods is not
WEIGHING THE EVIDENCE FOR A MASS EXTINCTION PART 2: ON LAND LESSON INTRODUCTION TEACHER MATERIALS The iridium-rich clay layer at the boundary between the Cretaceous (K) and Tertiary (T) periods is not
A FOSSIL CRAB FROM THE LAKES ENTRANCE OIL SHAFT, GIPPSLAND, VICTORIA
 gtopt JtoAtty 0f WitUxm. -..., ' -: / '.. v - - ^.. ' ' * A FOSSIL CRAB FROM THE LAKES ENTRANCE OIL SHAFT, GIPPSLAND, VICTORIA BY.'. " ; IRENE CRESPIN, R. A. Read 6** June, 1946 ' Reprinted from Proo.
gtopt JtoAtty 0f WitUxm. -..., ' -: / '.. v - - ^.. ' ' * A FOSSIL CRAB FROM THE LAKES ENTRANCE OIL SHAFT, GIPPSLAND, VICTORIA BY.'. " ; IRENE CRESPIN, R. A. Read 6** June, 1946 ' Reprinted from Proo.
Asian Bush Honeysuckles. Lonicera morrowii, L. tatarica, L. x bella, L. maackii
 Asian Bush Honeysuckles Lonicera morrowii, L. tatarica, L. x bella, L. maackii Table comparing nonnative shrubby Lonicera spp. Table adapted from: A guide to Nonnative Invasive Plants Inventoried in the
Asian Bush Honeysuckles Lonicera morrowii, L. tatarica, L. x bella, L. maackii Table comparing nonnative shrubby Lonicera spp. Table adapted from: A guide to Nonnative Invasive Plants Inventoried in the
Lake Levels and Climate Change in Maine and Eastern North America during the last 12,000 years
 Maine Geologic Facts and Localities December, 2000 Lake Levels and Climate Change in Maine and Eastern North America during the last 12,000 years Text by Robert A. Johnston, Department of Agriculture,
Maine Geologic Facts and Localities December, 2000 Lake Levels and Climate Change in Maine and Eastern North America during the last 12,000 years Text by Robert A. Johnston, Department of Agriculture,
This book focuses mostly on Proteas, but also considers some of the other Proteaceae genera that are more widely cultivated.
 CHAPTER 1: INTRODUCING THE PROTEA FAMILY There are around 1700 species and 79 genera of plants in the Proteaceae (Protea) family, and most are indigenous to the southern hemisphere. Around half of these
CHAPTER 1: INTRODUCING THE PROTEA FAMILY There are around 1700 species and 79 genera of plants in the Proteaceae (Protea) family, and most are indigenous to the southern hemisphere. Around half of these
Geology, Coal and Gas Resources of the Clarence Moreton Basin. Mark Dawson 05/04/2017
 Geology, Coal and Gas Resources of the Clarence Moreton Basin Mark Dawson 05/04/2017 2 Talk Outline Who am I? Regional Geology Stratigraphy Previous Mining Future Resource Areas 3 Who am I? Graduated from
Geology, Coal and Gas Resources of the Clarence Moreton Basin Mark Dawson 05/04/2017 2 Talk Outline Who am I? Regional Geology Stratigraphy Previous Mining Future Resource Areas 3 Who am I? Graduated from
Fig Available seismic reflection, refraction, and magnetic profiles from 107 the Offshore Indus Basin close to the representative profile GCDH,
 List of Figures Page No. Fig. 1.1 Generalized physiography of the Indian Ocean along with 2 selected (200 m, 1000 m, 2000 m, and 3000 m) bathymetric contours. Fig. 1.2 Lithospheric plates in the Indian
List of Figures Page No. Fig. 1.1 Generalized physiography of the Indian Ocean along with 2 selected (200 m, 1000 m, 2000 m, and 3000 m) bathymetric contours. Fig. 1.2 Lithospheric plates in the Indian
Major Domain of the Earth
 Major Domain of the Earth The surface of the earth is a complex zone in which three main components of the environment meet, overlap and interact. The solid portion of the earth on which we live is called
Major Domain of the Earth The surface of the earth is a complex zone in which three main components of the environment meet, overlap and interact. The solid portion of the earth on which we live is called
Reproductive Morphology
 Week 3; Wednesday Announcements: 1 st lab quiz TODAY Reproductive Morphology Reproductive morphology - any portion of a plant that is involved with or a direct product of sexual reproduction Example: cones,
Week 3; Wednesday Announcements: 1 st lab quiz TODAY Reproductive Morphology Reproductive morphology - any portion of a plant that is involved with or a direct product of sexual reproduction Example: cones,
Chapter 2: Plate Tectonics: A Unifying Theory
 Chapter 2: Plate Tectonics: A Unifying Theory Chapter Outline 2.1 Introduction 2.2 Early Ideas About Continental Drift 2.3 What Is the Evidence for Continental Drift? 2.4 Features of the Seafloor 2.5 Earth
Chapter 2: Plate Tectonics: A Unifying Theory Chapter Outline 2.1 Introduction 2.2 Early Ideas About Continental Drift 2.3 What Is the Evidence for Continental Drift? 2.4 Features of the Seafloor 2.5 Earth
Pteridophytes: Pteridophytes are vascular cryptogams. They are the earliest know vascular plants which originated in the Silurian period of
 Pteridophytes: Pteridophytes are vascular cryptogams. They are the earliest know vascular plants which originated in the Silurian period of Palaeozoic era. They are the successful colonizers on land habit.
Pteridophytes: Pteridophytes are vascular cryptogams. They are the earliest know vascular plants which originated in the Silurian period of Palaeozoic era. They are the successful colonizers on land habit.
On the occurrence of fossil conifers with affinities to Geinitzia in the late Cretaceous
 On the occurrence of fossil conifers with affinities to Geinitzia in the late Cretaceous (Campanian) Mesaverde Group, Williams Fork Formation of northeastern Utah, U.S.A. 1 1 1 1 1 1 1 1 0 1 Benjamin J.
On the occurrence of fossil conifers with affinities to Geinitzia in the late Cretaceous (Campanian) Mesaverde Group, Williams Fork Formation of northeastern Utah, U.S.A. 1 1 1 1 1 1 1 1 0 1 Benjamin J.
High-resolution Sequence Stratigraphy of the Glauconitic Sandstone, Upper Mannville C Pool, Cessford Field: a Record of Evolving Accommodation
 Page No. 069-1 High-resolution Sequence Stratigraphy of the Glauconitic Sandstone, Upper Mannville C Pool, Cessford Field: a Record of Evolving Accommodation Thérèse Lynch* and John Hopkins, Department
Page No. 069-1 High-resolution Sequence Stratigraphy of the Glauconitic Sandstone, Upper Mannville C Pool, Cessford Field: a Record of Evolving Accommodation Thérèse Lynch* and John Hopkins, Department
PLATE TECTONICS. SECTION 17.1 Drifting Continents
 Date Period Name PLATE TECTONICS SECTION.1 Drifting Continents In your textbook, read about continental drift. Circle the letter of the choice that best completes each statement. 1. Early mapmakers thought
Date Period Name PLATE TECTONICS SECTION.1 Drifting Continents In your textbook, read about continental drift. Circle the letter of the choice that best completes each statement. 1. Early mapmakers thought
First Record of the Rudist Bivalve Mitrocaprina tschoppi (Palmer) from the Maastrichtian of Jamaica
 from the Maastrichtian of Jamaica") 392 Caribbean Journal of Science, Vol. 40, No. 3, 392-396, 2004 Copyright 2004 College of Arts and Sciences University of Puerto Rico, Mayagüez First Record of the Rudist Bivalve Mitrocaprina tschoppi
392 Caribbean Journal of Science, Vol. 40, No. 3, 392-396, 2004 Copyright 2004 College of Arts and Sciences University of Puerto Rico, Mayagüez First Record of the Rudist Bivalve Mitrocaprina tschoppi
GCE A level 1215/03 GEOLOGY - GL5 THEMATIC UNIT 3 GEOLOGICAL EVOLUTION OF BRITAIN
 Surname Other Names Centre Number 2 Candidate Number GCE A level 1215/03 GEOLOGY - GL5 THEMATIC UNIT 3 GEOLOGICAL EVOLUTION OF BRITAIN P.M. FRIDAY, 10 June 2011 ONE of TWO units to be completed in 2 hours
Surname Other Names Centre Number 2 Candidate Number GCE A level 1215/03 GEOLOGY - GL5 THEMATIC UNIT 3 GEOLOGICAL EVOLUTION OF BRITAIN P.M. FRIDAY, 10 June 2011 ONE of TWO units to be completed in 2 hours
LEARN * DREAM * AWAKEN* DISCOVER * ENLIGHTEN * INVESTIGATE * QUESTION * EXPLORE
 S C I N C P L A N T S PLANTS Ancient Plants in Focus This nrichment4you e-guide provides a brief overview of ancient leaves. In this e-guide you will: *Learn basic Information About Fossilized Leaves *Learn
S C I N C P L A N T S PLANTS Ancient Plants in Focus This nrichment4you e-guide provides a brief overview of ancient leaves. In this e-guide you will: *Learn basic Information About Fossilized Leaves *Learn
GEOPHYSICS GRAVITY DATA COVERAGE
 GEOPHYSICS DATA COVERAGE The Mudgee-Gulgong district lies within the Dubbo 1:250,000 sheet area. This area is now covered by high res_olution gravity, magnetic and radiometric data. The aeromagnetic and
GEOPHYSICS DATA COVERAGE The Mudgee-Gulgong district lies within the Dubbo 1:250,000 sheet area. This area is now covered by high res_olution gravity, magnetic and radiometric data. The aeromagnetic and
Structural Deformation and Karst in the Devonian Waterways Formation: Examples from Outcrops along the Athabasca River*
 Structural Deformation and Karst in the Devonian Waterways Formation: Examples from Outcrops along the Athabasca River* Chris L. Schneider 1 and Matthias Grobe 2 Search and Discovery Article #51073 (2015)
Structural Deformation and Karst in the Devonian Waterways Formation: Examples from Outcrops along the Athabasca River* Chris L. Schneider 1 and Matthias Grobe 2 Search and Discovery Article #51073 (2015)
Figure 1. Examples of vector displacement diagrams for two and three-plate systems.
 Figure 1. Examples of vector displacement diagrams for two and three-plate systems. Figure 2. Relationships between pole of rotation, great circles, ridge segments, small circles, transforms and fracture
Figure 1. Examples of vector displacement diagrams for two and three-plate systems. Figure 2. Relationships between pole of rotation, great circles, ridge segments, small circles, transforms and fracture
Chapter 02 The Sea Floor
 Chapter 02 The Sea Floor Multiple Choice Questions 1. One of the following is not one of the world's major ocean basins: A. Atlantic Ocean B. Arctic Ocean C. Indian Ocean D. Antarctic Ocean E. Pacific
Chapter 02 The Sea Floor Multiple Choice Questions 1. One of the following is not one of the world's major ocean basins: A. Atlantic Ocean B. Arctic Ocean C. Indian Ocean D. Antarctic Ocean E. Pacific
Orphan Basin, Offshore Newfoundland: New seismic data and hydrocarbon plays for a dormant Frontier Basin
 Orphan Basin, Offshore Newfoundland: New seismic data and hydrocarbon plays for a dormant Frontier Basin Jerry Smee* G&G Exploration Consulting, 301 400-3rd Avenue SW, Calgary, AB, T2P 4H2 Sam Nader, Paul
Orphan Basin, Offshore Newfoundland: New seismic data and hydrocarbon plays for a dormant Frontier Basin Jerry Smee* G&G Exploration Consulting, 301 400-3rd Avenue SW, Calgary, AB, T2P 4H2 Sam Nader, Paul
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS White, Mary E., 1981. Cylomeia undulata (Burges) gen.et comb.nov., a lycopod of the Early Triassic strata of New South Wales. Records of the Australian Museum
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS White, Mary E., 1981. Cylomeia undulata (Burges) gen.et comb.nov., a lycopod of the Early Triassic strata of New South Wales. Records of the Australian Museum
Appendix Z. Groundwater Assessment Supplementary Report. Document Name i Insert Month/Year
 Appendix Z Groundwater Assessment Supplementary Report Document Name i Insert Month/Year Australasian Groundwater and Environmental Consultants Pty Ltd Level 2 / 15 Mallon Street Bowen Hills, QLD 4006
Appendix Z Groundwater Assessment Supplementary Report Document Name i Insert Month/Year Australasian Groundwater and Environmental Consultants Pty Ltd Level 2 / 15 Mallon Street Bowen Hills, QLD 4006
Yukon Presentation. January 2015
 Yukon Presentation January 2015 Disclaimer This presentation contains forward looking statements. All statements, other than statements of historical fact, included herein, including without limitation,
Yukon Presentation January 2015 Disclaimer This presentation contains forward looking statements. All statements, other than statements of historical fact, included herein, including without limitation,
A New Locality of Fossombronia mylioides (Fossombroniaceae, Marchantiophyta)
") Bull. Natl. Mus. Nat. Sci., Ser. B, 42(1), pp. 19 23, February 22, 2016 A New Locality of Fossombronia mylioides (Fossombroniaceae, Marchantiophyta) Masanobu Higuchi Department of Botany, National Museum
Bull. Natl. Mus. Nat. Sci., Ser. B, 42(1), pp. 19 23, February 22, 2016 A New Locality of Fossombronia mylioides (Fossombroniaceae, Marchantiophyta) Masanobu Higuchi Department of Botany, National Museum
Laboratory #7: Plate Tectonics
 Materials Needed: 1. Pencil 2. Colored Pencils 3. Metric/Standard Ruler 4. Calculator 5. Tracing Paper Laboratory #7: Plate Tectonics Plate Tectonics The Earth is composed of layers. At the center is a
Materials Needed: 1. Pencil 2. Colored Pencils 3. Metric/Standard Ruler 4. Calculator 5. Tracing Paper Laboratory #7: Plate Tectonics Plate Tectonics The Earth is composed of layers. At the center is a
REPORT OF GEOLOGICAL MAPPING ON THE MICHEL COAL LICENSE #5177. LOCATION: KOOTENAY LAND DISTRICT NTS 82-G-7 4Y028 Lat f Long.
 i *. REPORT OF GEOLOGICAL MAPPING ON THE MICHEL COAL LICENSE #5177 LOCATION: KOOTENAY LAND DISTRICT NTS 82-G-7 4Y028 Lat. 114 40f Long. ESSO RESOURCES CANADA LIMITED WARD E. KILBY WORK: OCTOBER 17 - OCTOBER
i *. REPORT OF GEOLOGICAL MAPPING ON THE MICHEL COAL LICENSE #5177 LOCATION: KOOTENAY LAND DISTRICT NTS 82-G-7 4Y028 Lat. 114 40f Long. ESSO RESOURCES CANADA LIMITED WARD E. KILBY WORK: OCTOBER 17 - OCTOBER
Topic 22. Introduction to Vascular Plants: The Lycophytes
 Topic 22. Introduction to Vascular Plants: The Lycophytes Introduction to Vascular Plants Other than liverworts, hornworts, and mosses, all plants have vascular tissues. As discussed earlier, the mosses
Topic 22. Introduction to Vascular Plants: The Lycophytes Introduction to Vascular Plants Other than liverworts, hornworts, and mosses, all plants have vascular tissues. As discussed earlier, the mosses
A MEGASECOPTERON FROM UPPER CARBONIFEROUS BY F. M. CARPENTER. In I962 Professor F. Stockmans, of the Institut Royal des Sciences STRATA IN SPAIN
 A MEGASECOPTERON FROM UPPER CARBONIFEROUS STRATA IN SPAIN BY F. M. CARPENTER Harvard University In I962 Professor F. Stockmans, of the Institut Royal des Sciences Naturelles de. Belgique, kindly sent me
A MEGASECOPTERON FROM UPPER CARBONIFEROUS STRATA IN SPAIN BY F. M. CARPENTER Harvard University In I962 Professor F. Stockmans, of the Institut Royal des Sciences Naturelles de. Belgique, kindly sent me
162. Protosequoia (n, g.) in Taxodiaceae from Pinus tri f olia Beds in Central Honshu, Japan
 in Taxodiaceae from Pinus tri f olia Beds in Central Honshu, Japan") No. 8] Proc. Japan Acad., 45 (1969) 727 162. Protosequoia (n, g.) in Taxodiaceae from Pinus tri f olia Beds in Central Honshu, Japan By Shigeru MIKI Mukogawa Women's Univ., Nishinomiya City, Hyogo (Comm.
No. 8] Proc. Japan Acad., 45 (1969) 727 162. Protosequoia (n, g.) in Taxodiaceae from Pinus tri f olia Beds in Central Honshu, Japan By Shigeru MIKI Mukogawa Women's Univ., Nishinomiya City, Hyogo (Comm.
Class XI Chapter 6 Anatomy of Flowering Plants Biology
 Class XI Chapter 6 Anatomy of Flowering Plants Biology Question 1: State the location and function of different types of meristem. Meristems are specialised regions of plant growth. The meristems mark
Class XI Chapter 6 Anatomy of Flowering Plants Biology Question 1: State the location and function of different types of meristem. Meristems are specialised regions of plant growth. The meristems mark
The Continental Drift Hypothesis
 Lesson 1 The Continental Drift Hypothesis Scan Lesson 1. Then write three questions that you have about continental drift in your Science Journal. Try to answer your questions as you read. Pangaea Define
Lesson 1 The Continental Drift Hypothesis Scan Lesson 1. Then write three questions that you have about continental drift in your Science Journal. Try to answer your questions as you read. Pangaea Define
GNS Science, Lower Hutt, New Zealand NZSEE Conference
 A Ground Shaking Amplification Map for New Zealand U. Destegul, G. Dellow & D. Heron GNS Science, Lower Hutt, New Zealand. 2008 NZSEE Conference ABSTRACT: A ground shaking amplification map of New Zealand
A Ground Shaking Amplification Map for New Zealand U. Destegul, G. Dellow & D. Heron GNS Science, Lower Hutt, New Zealand. 2008 NZSEE Conference ABSTRACT: A ground shaking amplification map of New Zealand
=%REPORT RECONNAISSANCE OF CHISHOLM LAKE PROSPECT. October 25, 1977
 =%REPORT ON FIELD RECONNAISSANCE OF CHISHOLM LAKE PROSPECT October 25, 1977 Bruce D. Vincent Imperial Oil Limited, Minerals - Coal, CALGARY, ALBERTA CHISHOLM LAKE PROSPECT Introduction The Chisholm Lake
=%REPORT ON FIELD RECONNAISSANCE OF CHISHOLM LAKE PROSPECT October 25, 1977 Bruce D. Vincent Imperial Oil Limited, Minerals - Coal, CALGARY, ALBERTA CHISHOLM LAKE PROSPECT Introduction The Chisholm Lake
A new observation of Ovummuridae, from the mid-silurian (Wenlock) strata of Wenlock Edge, Shropshire, UK: A preliminary report
 strata of Wenlock Edge, Shropshire, UK: A preliminary report") A NEW OBSERVATION FROM WENLOCK EDGE A new observation of Ovummuridae, from the mid-silurian (Wenlock) strata of Wenlock Edge, Shropshire, UK: A preliminary report Steven Rogers 1, Joel Blackburn 1,2 and
A NEW OBSERVATION FROM WENLOCK EDGE A new observation of Ovummuridae, from the mid-silurian (Wenlock) strata of Wenlock Edge, Shropshire, UK: A preliminary report Steven Rogers 1, Joel Blackburn 1,2 and
DEEP TIME it's discovery and interpretation
 DEEP TIME it's discovery and interpretation from ancient times to the 1600s, where time was thought of at all, it was supposed either continuous & 'circular' or of rather short duration either way, the
DEEP TIME it's discovery and interpretation from ancient times to the 1600s, where time was thought of at all, it was supposed either continuous & 'circular' or of rather short duration either way, the
Quarterly Report Quarter ended 30 June 2010
 28 July 2010 The Company Announcement Officer Australian Securities Exchange Ltd via electronic lodgement Quarterly Report Quarter ended 30 June 2010 Highlights PhosEnergy - Uranium Extraction Technology
28 July 2010 The Company Announcement Officer Australian Securities Exchange Ltd via electronic lodgement Quarterly Report Quarter ended 30 June 2010 Highlights PhosEnergy - Uranium Extraction Technology
Case Study of the Structural and Depositional-Evolution Interpretation from Seismic Data*
 Case Study of the Structural and Depositional-Evolution Interpretation from Seismic Data* Yun Ling 1, Xiangyu Guo 1, Jixiang Lin 1, and Desheng Sun 1 Search and Discovery Article #20143 (2012) Posted April
Case Study of the Structural and Depositional-Evolution Interpretation from Seismic Data* Yun Ling 1, Xiangyu Guo 1, Jixiang Lin 1, and Desheng Sun 1 Search and Discovery Article #20143 (2012) Posted April
Chapter 1-Plants in Our World
 Chapter 1-Plants in Our World Formation of earth-4.5-4.6 billion years ago Evidence of life from organic material-3.8 billion years ago Many cyanobacteria are photosynthetic, but these microscopic organisms
Chapter 1-Plants in Our World Formation of earth-4.5-4.6 billion years ago Evidence of life from organic material-3.8 billion years ago Many cyanobacteria are photosynthetic, but these microscopic organisms
Patterns of evolution
 To branch or not to branch Patterns of evolution Chapter 3 Cladogenesis lineages branch into two or more lines Anagenesis evolutionary change in a lineage without branching Anagenesis and Cladogenesis
To branch or not to branch Patterns of evolution Chapter 3 Cladogenesis lineages branch into two or more lines Anagenesis evolutionary change in a lineage without branching Anagenesis and Cladogenesis
Mesozoic Earth History
 Mesozoic Earth History The Mesozoic Era 251-66 MYA Breakup of Pangea Changes in air and oceanic currents Evolution of new terrestrial and marine life Opening of the Atlantic Ocean Basin Rocky Mountains
Mesozoic Earth History The Mesozoic Era 251-66 MYA Breakup of Pangea Changes in air and oceanic currents Evolution of new terrestrial and marine life Opening of the Atlantic Ocean Basin Rocky Mountains
THE DISCOVERY OF A NEW CONTINENT ON PLANET EARTH: Zealandia
 THE DISCOVERY OF A NEW CONTINENT ON PLANET EARTH: Zealandia By Vashaanth Gowri-Kriszyk, Science Editor February 2017 Recently, in February 2017, scientists have discovered a new continent called Zealandia,
THE DISCOVERY OF A NEW CONTINENT ON PLANET EARTH: Zealandia By Vashaanth Gowri-Kriszyk, Science Editor February 2017 Recently, in February 2017, scientists have discovered a new continent called Zealandia,
Hildebrand Analysis: Topic 4
 Steven Plescia 12/9/16 Hildebrand Analysis: Topic 4 Hildebrand interprets much of the classic late pc-paloezoic Cordilleran miogeocline as exotic to North America Is there a break in the detrital zircons
Steven Plescia 12/9/16 Hildebrand Analysis: Topic 4 Hildebrand interprets much of the classic late pc-paloezoic Cordilleran miogeocline as exotic to North America Is there a break in the detrital zircons
Comparative Taphonomy of the Vendian Genera. Beltanelloides and Nemiana as a Key to their True. Nature
 Comparative Taphonomy of the Vendian Genera Beltanelloides and Nemiana as a Key to their True Nature M. V. Leonov Paleontological Institute, Russian Academy of Science, 123 Profsoyuznaya, Moscow, 117868,
Comparative Taphonomy of the Vendian Genera Beltanelloides and Nemiana as a Key to their True Nature M. V. Leonov Paleontological Institute, Russian Academy of Science, 123 Profsoyuznaya, Moscow, 117868,
EXISTING GEOLOGICAL INFORMATION
 CHAPER 3 EXISTING GEOLOGICAL INFORMATION 3-1 General Geology of the Surrounding Area (1) General geology and ore deposits in Mongolia Geographically, Mongolia is a country located between Russia to the
CHAPER 3 EXISTING GEOLOGICAL INFORMATION 3-1 General Geology of the Surrounding Area (1) General geology and ore deposits in Mongolia Geographically, Mongolia is a country located between Russia to the
IVANHOE COMMENCES RESOURCE DELINEATION DRILLING AT NARIIN SUKHAIT COAL PROJECT IN MONGOLIA S SOUTH GOBI
 January 12, 2005 IVANHOE COMMENCES RESOURCE DELINEATION DRILLING AT NARIIN SUKHAIT COAL PROJECT IN MONGOLIA S SOUTH GOBI ULAANBAATAR, MONGOLIA Ivanhoe Mines Chairman Robert Friedland and President John
January 12, 2005 IVANHOE COMMENCES RESOURCE DELINEATION DRILLING AT NARIIN SUKHAIT COAL PROJECT IN MONGOLIA S SOUTH GOBI ULAANBAATAR, MONGOLIA Ivanhoe Mines Chairman Robert Friedland and President John
60% water. Big Bang: 14,000 millions years ago The Earth originated about 4,500 millions years ago its orbit allows water to exist in a liquid state!
 Ch2. The Sea Floor #1 Why geology of the oceans? Marine habitats are directly shaped by geological processes The form of the coastlines The depth of the water Type of bottom (muddy, sandy, rocky) #2 Geological
Ch2. The Sea Floor #1 Why geology of the oceans? Marine habitats are directly shaped by geological processes The form of the coastlines The depth of the water Type of bottom (muddy, sandy, rocky) #2 Geological
Geography of Evolution
 Geography of Evolution Biogeography - the study of the geographic distribution of organisms. The current distribution of organisms can be explained by historical events and current climatic patterns. Darwin
Geography of Evolution Biogeography - the study of the geographic distribution of organisms. The current distribution of organisms can be explained by historical events and current climatic patterns. Darwin
Evolutionary Morphology of Land Plants
 MHRD Scheme on Global Initiative on Academic Network (GIAN) Evolutionary Morphology of Land Plants Overview This century will be dominated by Life Sciences for well reasons of understanding evolution,
MHRD Scheme on Global Initiative on Academic Network (GIAN) Evolutionary Morphology of Land Plants Overview This century will be dominated by Life Sciences for well reasons of understanding evolution,
FLOWER MORPHOLOGY AND PLANT TYPES WITHIN JUNCUS ROEMERlANUS
 FLOWER MORPHOLOGY AND PLANT TYPES WITHIN JUNCUS ROEMERlANUS LIONEL N. ELEUTERIUS Gulf Coast Research Laboratory Ocean Springs, Mississippi 39564 ABSTRACT Two plant forms were found to occur within funcus
FLOWER MORPHOLOGY AND PLANT TYPES WITHIN JUNCUS ROEMERlANUS LIONEL N. ELEUTERIUS Gulf Coast Research Laboratory Ocean Springs, Mississippi 39564 ABSTRACT Two plant forms were found to occur within funcus
Early theories: Joseph Hooker (1853) vs. Charles Darwin (1859)
 vs. Charles Darwin (1859)") Gondwanan Plants of the Sydney Region Presentation Dr Peter Weston 25/11/2017 Honorary Research Associate, Science and Conservation Branch, Royal Botanic Gardens and Domain Trust Summary: Dr Marilyn Cross,
Gondwanan Plants of the Sydney Region Presentation Dr Peter Weston 25/11/2017 Honorary Research Associate, Science and Conservation Branch, Royal Botanic Gardens and Domain Trust Summary: Dr Marilyn Cross,
Lecture #13 notes, Geology 3950 Spring 2006: CR Stern Magnetic reversals (text pages th edition and in the 5 th edition)
") Lecture #13 notes, Geology 3950 Spring 2006: CR Stern Magnetic reversals (text pages 35-37 4 th edition and 53-55 in the 5 th edition) The earth has a magnetic field generated by circulation of charged
Lecture #13 notes, Geology 3950 Spring 2006: CR Stern Magnetic reversals (text pages 35-37 4 th edition and 53-55 in the 5 th edition) The earth has a magnetic field generated by circulation of charged
IRAQ. Target Exploration. Geodynamic Evolutions of The Sedimentary Basins of. This study is a major reference for Petroleum
 barr Target Exploration Target Exploration Geodynamic Evolutions of The Sedimentary Basins of IRAQ This study is a major reference for Petroleum Explorationists on the tectonics, stratigraphy, sedimentary
barr Target Exploration Target Exploration Geodynamic Evolutions of The Sedimentary Basins of IRAQ This study is a major reference for Petroleum Explorationists on the tectonics, stratigraphy, sedimentary
2013 GEOLOGICAL ASSESSMENT REPORT SHERIDAN HILL PROPERTY
 2013 GEOLOGICAL ASSESSMENT REPORT ON THE SHERIDAN HILL PROPERTY NEW WESTMINSTER MINING DIVISION BRITISH COLUMBIA NTS 092G07 49 16 31 NORTH LATITUDE, 122 39 48 WEST LONGITUDE PREPARED FOR Sheridan Hill
2013 GEOLOGICAL ASSESSMENT REPORT ON THE SHERIDAN HILL PROPERTY NEW WESTMINSTER MINING DIVISION BRITISH COLUMBIA NTS 092G07 49 16 31 NORTH LATITUDE, 122 39 48 WEST LONGITUDE PREPARED FOR Sheridan Hill
Tuesday 10 June 2014 Afternoon
 Tuesday 10 June 2014 Afternoon A2 GCE GEOLOGY F795/01 Evolution of Life, Earth and Climate *1242977619* Candidates answer on the Question Paper. OCR supplied materials: None Other materials required: Electronic
Tuesday 10 June 2014 Afternoon A2 GCE GEOLOGY F795/01 Evolution of Life, Earth and Climate *1242977619* Candidates answer on the Question Paper. OCR supplied materials: None Other materials required: Electronic
Controls on clastic systems in the Angoche basin, Mozambique: tectonics, contourites and petroleum systems
 P2-2-13 Controls on clastic systems in the Angoche basin, Mozambique: tectonics, contourites and petroleum systems Eva Hollebeek, Olivia Osicki, Duplo Kornpihl Schlumberger, London, UK Introduction Offshore
P2-2-13 Controls on clastic systems in the Angoche basin, Mozambique: tectonics, contourites and petroleum systems Eva Hollebeek, Olivia Osicki, Duplo Kornpihl Schlumberger, London, UK Introduction Offshore
Swaledale and Arkengarthdale Archaeology Group
 Swaledale and Arkengarthdale Archaeology Group Bellerby Moor GPS Earthwork Survey Report prepared by: Tim Laurie and Stephen Eastmead. Swaledale and Arkengarthdale Archaeology Group. January 27/01/2012.
Swaledale and Arkengarthdale Archaeology Group Bellerby Moor GPS Earthwork Survey Report prepared by: Tim Laurie and Stephen Eastmead. Swaledale and Arkengarthdale Archaeology Group. January 27/01/2012.
Geologic History of Texas: The Making of Texas Over 1.5 Billion Years
 # 4 Geologic History of Texas: The Making of Texas Over 1.5 Billion Years Dr. Richard Kyle March 24, 2000 Produced by and for Hot Science - Cool Talks by the Environmental Science Institute. We request
# 4 Geologic History of Texas: The Making of Texas Over 1.5 Billion Years Dr. Richard Kyle March 24, 2000 Produced by and for Hot Science - Cool Talks by the Environmental Science Institute. We request