Evaluating the Involvement of Alternative Sigma Factors SigF and SigG in
|
|
|
- Bryce Robinson
- 6 years ago
- Views:
Transcription
1 IAI Accepts, published online ahead of print on 1 July 0 Infect. Immun. doi:./iai.00- Copyright 0, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. 1 Evaluating the Involvement of Alternative Sigma Factors SigF and SigG in Clostridium perfringens Sporulation and Enterotoxin Synthesis Jihong Li and Bruce A. McClane* University of Pittsburgh School of Medicine, Pittsburgh, PA, USA Running title: Role of SigF and SigG in sporulation and CPE synthesis * To Whom Correspondence Should be Addressed: Bruce A. McClane Department of Microbiology and Molecular Genetics Bridgeside Point II Building 0 Technology Drive University of Pittsburgh School of Medicine Pittsburgh, PA Phone: 1--0 Downloaded from on April, 01 by guest 1 Fax: bamcc@pitt.edu 1
2 Abstract Clostridium perfringens type A food poisoning is the second most commonly-identified bacterial foodborne illness. Sporulation contributes to this disease in two ways, i) most food poisoning strains form exceptionally-resistant spores to facilitate their survival against foodassociated stresses and ii) the enterotoxin (CPE) responsible for the symptoms of this food poisoning is synthesized only during sporulation. In Bacillus subtilis, four alternative sigma factors mediate sporulation. The same four sigma factors are encoded by C. perfringens genomes and two (SigE and SigK) were previously shown (Harry et al., J. Bacteriol. 00, :-) to be necessary for sporulation and CPE production by SM1, a transformable derivative of a C. perfringens food poisoning strain. However, the importance of SigF and SigG for C. perfringens sporulation or CPE production had not yet been assessed. After confirming that sporulating wildtype SM1 cultures produce SigF (from a tricistronic operon) and SigG, the current study prepared isogenic sigf or sigg null mutants. Whereas SM1 formed heat-resistant, phaserefractile spores, the sigf and sigg null mutants were blocked for spore formation. Complementation fully restored sporulation by both mutants. Using these mutants and complementing strains, CPE production was shown to be SigF-dependent, but SigG-independent. This finding apparently involved SigF regulating production of SigE and SigK, which Harry et al showed are necessary for CPE synthesis. Coupling these findings with those previous results, it is now apparent that all four alternative sigma factors are necessary for C. perfringens sporulation, but only SigE, SigF, and SigK are needed for CPE synthesis. Downloaded from on April, 01 by guest
3 Introduction Clostridium perfringens type A isolates producing enterotoxin (CPE) are the second most commonly-identified cause of bacterial foodborne disease in the USA, where an estimated 0,000 cases of this food poisoning occur annually (1, 1). C. perfringens type A food poisoning begins with consumption of foods contaminated with vegetative cells of an enterotoxigenic type A strain. Those ingested bacteria then sporulate in the small intestine and produce CPE, which has been shown to cause the diarrheal and abdominal cramping symptoms that characterize C. perfringens type A food poisoning (1, ). CPE-producing type A strains are also an important cause of human nonfoodborne gastrointestinal diseases (, ). For example, it has been estimated that these bacteria account for -1% of all cases of antibioticassociated diarrhea (). Enterotoxigenic type A isolates only produce CPE during sporulation (1). The sporulation-associated nature of CPE expression is controlled at the transcriptional level, since both Northern blot and reporter construct studies detected enterotoxin gene (cpe) transcription in sporulating cultures, but not vegetative cultures, of CPE-positive C. perfringens strains (, 0). CPE can account for up to 0% of the total protein present in a sporulating C. perfringens cell (). Three strong promoters controlling cpe transcription have been identified upstream of the cpe ORF (0), probably explaining why type A isolates often produce such high levels of CPE during sporulation. Downloaded from on April, 01 by guest
4 Interestingly, the sequence of one cpe promoter (named P1) resembles a B. subtilis SigKdependent promoter, while sequences of the other two cpe promoters (named P and P) show similarity to B. subtilis SigE-dependent promoters (0). SigE and SigK are two of the four sporulation-associated sigma factors produced by Bacillus subtilis, which has traditionally been the prototype for studying sporulation in Gram-positive bacteria (, ). These four sporulationassociated sigma factors are now well-established as major regulators of B. subtilis sporulation, where SigF and SigG regulate gene expression in the forespore, while SigE and SigK control gene expression in the mother cell (, ). In B. subtilis, expression of these sporulationassociated sigma factors occurs in an ordered temporal cascade, with SigF appearing first, followed sequentially by SigE, SigG and SigK (, ). Sporulation has been much less studied in C. perfringens compared to B. subtilis, but homologues of genes encoding SigE, SigF, SigG and SigK have been identified in C. perfringens genomes (, ). In addition, a recent study by Harry et al. () used RT-PCR to detect transcripts of all four sigma factors in sporulating cultures of CPE-positive C. perfringens type A isolate SM1, a transformable derivative of a food poisoning isolate. Furthermore, Western blots performed in that study confirmed SigE and SigK production by SM1 growing in Duncan-Strong (DS) sporulation medium (). Since putative SigK- and SigE-dependent promoters had been previously identified upstream of the cpe gene (0), Harry et al. constructed sigk and sige null mutants of SM1 to evaluate the importance of SigK and SigE for sporulation and CPE production by SM1. Consistent with cpe transcription being dependent Downloaded from on April, 01 by guest
5 upon these two sporulation-associated sigma factors, both mutants failed to drive β- glucuronidase production when transformed with a plasmid construct carrying the cpe promoter region fused to the E. coli reporter gene gusa. In addition, neither the SM1 sigk nor sige null mutants could sporulate The recent Harry et al. study () also suggested differences in some developmental events during sporulation of C. perfringens strain SM1 versus B. subtilis. For example, that recent study suggested the normal accumulation of sigf and sige transcripts in sporulating SM1 cells is dependent upon SigK, whereas SigK is the last sigma factor expressed during B. subtilis sporulation. In addition, sigf transcript accumulation was reportedly delayed in a SM1 sige null mutant, even though sigf is the first transcript produced during B. subtilis sporulation. Harry et al. also suggested that C. perfringens and B. subtilis regulate SigG production differently, since, i) sigg transcripts were detected in SM1 sige and sigk mutants reportedly containing little or no sige or sigf transcript, while ii), sigg transcription by B. subtilis requires both SigE and SigF. Finally, transcripts of all four sigma factor genes were detected much earlier in SM1 than has been reported for B. subtilis. Given those apparent differences between sporulation in C. perfringens versus B. subtilis, the current study constructed isogenic SM1 sigf and sigg null mutants and complementing strains. Those strains were then employed to explore whether these two sigma factors might also be involved in sporulation and CPE production by SM1. Downloaded from on April, 01 by guest
6 Materials and Methods Bacterial strains, media, and reagents. C. perfringens type A isolate SM1, a derivative of food poisoning isolate NCTC (Hobbs serotype ), is transformable by electroporation (0). ATCC is a transformable, CPE-negative type A isolate (). Isolates were stored in cooked-meat medium (Oxoid) at -0 C. FTG (fluid thioglycolate, Difco Laboratories) and TGY broth (% tryptic soy broth [Becton Dickinson and Company], % glucose [Fisher scientific], 1% yeast extract [Difco], and 0.1% sodium thioglycolate [Sigma Chemical]) were used for growing vegetative cultures. Brain heart infusion (BHI) agar (Difco) supplemented with 1 µg/ml of chloramphenicol (Fisher Scientific) was used in screening to identify mutant clones. DS sporulation medium was used to induce sporulation of C. perfringens strains (). E. coli DHα was grown at C in LB broth, with shaking, or on LB agar containing 0µg/ml chloramphenicol. Construction of sigf and sigg null mutants of C. perfringens isolates SM1. The sigf and sigg genes of SM1 were inactivated by insertion of a group II intron using the Clostridiummodified TargeTron (Sigma) insertional mutagenesis system (). Utilizing optimal intron insertion sites identified in the SM1 genome sequence () and the Sigma TargeTron website (www. Sigma-genosys.com/targetron/), an intron was targeted, in the sense orientation, to the sigf ORF / nucleotide site. Primers used for PCR-targeting the intron to the sigf ORF were Downloaded from on April, 01 by guest
7 /s-ibs( -AAAAAAGCTTATAATTATCCTTAGGAAACAAGGAAGTGCGCCCAGAT AGGGTG- ); /s-ebs1d( -CAGATTGTACAAATGTGGTGATAACAGATAAGTCA AGGAAGCTAACTTACCTTTCTTTGT- ) and /s-ebs( - TGAACGCAAGTTTCTAA TTTCGATTTTTCCTCGATAGAGGAAAGTGTCT- ). Another intron was inserted, in the sense orientation, into the sigg ORF 1/ nucleotide site. This intron was prepared by PCR using primers 1/s-IBS( -AAAAAAGCTTATAATTATCCTTAAGAAACAAGAA- GTGCGCCCAGATAGGGTG- ); 1/s-EBS1d ( - CAGATTGTACAAATGTGG- TGATAACAGATAAGTCAAGAAGATTAACTTACCTTTCTTTGT- ) and 1/s-EBS ( - TGAACGCAAGTTTCTAATTTCGATTTTTCTTCGATAGAGGAAAGTGTCT- ). The 0bp PCR products were inserted into pjir0ai to construct sigf- and sigg-specific Targetron plasmids. The resultant plasmids, named (respectively) pjir0sigfi and pjir0siggi, were electroporated into wild-type SM1. The transformation efficiency for SM1 was about transformants/µg plasmid DNA. Transformants were selected on BHI agar plates containing 1 µg/ml of chloramphenicol and cells carrying an intron insertion were screened by PCR using primers sigfkof ( -AACTCTCATAGTCATGGCTAAAG- ) and sigfkor ( - AGTTCTTCCTTATTGTAA-TGAAGC- ) for sigf null mutants (SM1::sigF) or using primers siggkof ( -TGTATAGGACTTATGAAATCTATAG- ) and siggkor ( - ACATATATTGCATCTCCA-CCATC) for sigg null mutants (SM1:: sigg). Downloaded from on April, 01 by guest 0 1 Each reaction mixture was subjected to the following PCR amplification conditions: cycle 1, C for min; cycles through, C for 0 sec, C for 0 sec, and C for 0
8 sec; followed by extension for min at C. An aliquot (0µl) of each PCR sample was electrophoresed on a 1.% agarose gel and then visualized by staining with ethidium bromide A digoxigenin-labeled, intron sequence-specific probe was prepared, as described (), using primers IBS and EBS1d and a DIG-labelling kit (Roche). That probe was then employed for Southern blotting to confirm the presence of a single intron insertion in the SM1::sigF and SM1::sigG mutants. Briefly, DNA from SM1 wild type, the sigf null mutant or the sigg null mutant was isolated using the MasterPure gram-positive DNA purification kit (Epicentre, Wisconsin). A. µg aliquot of each isolated DNA sample was digested overnight with EcoRI according to the manufacturer s (New England Biolabs) instructions. The digested DNA samples were then electrophoresed on a conventional 1% agarose gel. The separated DNA digestion products were transferred onto a nylon member (Roche) for hybridization with the intron probe, as described previously (). Construction of complementing strains for sigf and sigg null mutants. Complementation of the SM1 sigf null mutant was accomplished by cloning the entire sigf operon (see Results), including two upstream ORFs and the sigf ORF, along with 00 bp of upstream sequence and 00 bp of downstream sequence, into the pjir0 E. coli/c. perfringens shuttle vector (1) and then transforming that plasmid into SM1::sigF. Briefly, DNA was isolated from wild-type SM1 as described above. PCR was then performed with that DNA using the Long-Range Taq DNA polymerase from New England Biolabs and primers sigfcomnf ( - Downloaded from on April, 01 by guest
9 ttacgaattcgtgtaaatggattgtctgttatag- with an added EcoRI site) and SigFcomR ( - atgcctgcagcatttataatcaatcctcttctag- with an added PstI site), designed according to sequencing results from a previous study (). The PCR reaction was performed in a Techne thermocycler using the following amplification conditions: C for min; cycles of C for 0 sec, C for 0 sec, and C for min; followed by a single extension at C for min. The resultant. kb PCR product was cloned into the Topo.1 vector (Invitrogen). The Topo plasmid was then digested with EcoRI and PstI. The excised insert containing the sigf operon was gel purified and ligated into shuttle plasmid pjir0, which had been digested with EcoRI and PstI, to produce plasmid pjir0sigfcomp. The SM1 sigf null mutant was transformed by electroporation with pjir0sigfcomp; a complementing strain named SM1sigFcomp was then selected from those transformants by growth on a BHI agar plate containing 1 µg/ml of chloramphenicol. Complementation of the SM1 sigg null mutant was accomplished by cloning the sigg ORF and 00bp of upstream sequence into pjir0 and then transforming that new plasmid into SM1::sigG. Briefly, DNA was isolated from wild-type SM1 as described above. PCR was then performed using the Long-Range Taq DNA polymerase from New England Biolabs and primers SigGcomF ( -ttacgaattcttgcaacctatgcatcaagatg- with an added EcoRI site) and SigGcomR ( - atgcctgcagagagctctatacatacttcctc - with an added PstI site) designed according to sequence results from previous study (). The PCR reaction was performed in a Techne thermocycler and used the following amplification conditions: C for min; cycles of C for 0 sec, C for 0 sec, and C for 0 sec; followed by a single Downloaded from on April, 01 by guest
10 extension at C for min. The resultant 1. kb PCR product was cloned into the Topo.1 vector (Invitrogen). The Topo plasmid was then digested with EcoRI and PstI. The excised insert containing sigg was gel-purified and ligated into shuttle plasmid pjir0, which had been digested with EcoRI and PstI, producing the plasmid pjir0siggcomp. The SM1 sigg null mutant was transformed by electroporation with pjir0siggcomp; the complementing strain SM1sigGcomp was then selected on BHI agar plates containing 1 µg/ml of chloramphenicol. RNA extraction and RT-PCR. To conduct a time course study of cpe gene expression during sporulation, a starter culture of wild-type SM1 was first grown overnight at C in FTG. A 0.1 ml aliquot of that culture was then transferred to ml of DS medium, which was incubated at C for 1 to hr, with sample aliquots removed each hour. After centrifugation of each removed culture aliquot, total C. perfringens RNA was extracted from the pelleted cells using saturated phenol (Fisher Scientific), as described in a previous study (). All phenol-extracted samples were then treated with U of DNaseI (Ambion) at C for 0 min. To stop this DNase I activity, a DNase I inhibitor (Ambion) was added to each reaction tube. RNA was quantified by absorbance at 0 nm and stored at -0 C for not more than one month. RT-PCR analysis for cpe gene transcription was then performed with these DNasetreated RNA samples, using the AccessQuick RT-PCR kit from Promega. Briefly, each RNA sample (0 ng) was reverse-transcribed to cdna at C for 1 h and then used as template for PCR with primers targeting cpe sequences or spo0a (as a control house-keeping gene). Control Downloaded from on April, 01 by guest
11 RT-PCR reactions were similarly performed, except for the omission of reverse transcriptase. As an additional control, a PCR reaction amplifying cpe or spo0a sequences was performed using DNA extracted from the MasterPure Gram-Positive DNA Purification Kit as described before. The PCR primers to amplify the cpe gene are F and R (1) and the spo0a primers are Spo0A- F ( -AACAACCAGATTTAGTTGTATTAG- ) and Spo0A-R ( -CTCTATTTGTCCTCT- TCCCCAAGC- ). The RT-PCR conditions used included: C for min; C for 1 h; cycles of C for 0 sec, C for 0 sec, and C for 0 sec; followed by a single extension at C for min. Overlapping RT-PCR analyses of sigf operon expression were performed using 0 ng of RNA extracted from SM1 grown for h in DS medium as described above. The primers used (Table 1) spanned the sigf ORF, three upstream ORFs and one downstream ORF. Each RT-PCR mixture contained 1 µl of template RNA (0 ng), µl of TAQ Complete mix, 0. µl transcriptase, and 1 µl of each primer pair (1 µm final concentration). The RT-PCR conditions used were: C for min; C for 1 h; cycles of C for 0 sec, C for 0 sec, and C for 1 min 0 sec; followed by a single extension at C for min. One control RT-PCR reaction was similarly performed using spo0a primers, except for the omission of reverse transcriptase. Another control reaction used these primers and DNA extracted from the MasterPure Gram-Positive DNA Purification Kit as described earlier. Downloaded from on April, 01 by guest 0
12 Northern blotting analysis of sigf transcription by wild-type SM1, a sigf null mutant and a complementing strain. Total RNA was phenol extracted from SM1, SM1::sigF and SM1sigFcomp using pelleted cells from a h DS culture, as described above. Northern blotting analysis was then performed using the Northern Max kit (a formaldehye-based system for Northern Blots) from Ambion. Briefly, gel preparation, electrophoresis, transfer of RNA onto a nylon membrane (Roche), prehybridization and hybridization were each performed according to the manufacturer s instructions. A digoxigenin-labeled, sigf internal sequencespecific probe was prepared, as described (1, ), using primers sigfkof and sigfkor and the DIG-labelling kit from Roche. Western blotting. For Western blot analysis of CPE production, a 0. ml aliquot of a FTG culture of wild-type SM1, SM1::sigF, SM1sigFcomp, SM1::sigG or SM1sigGcomp was inoculated into ml of DS sporulation medium. After incubation at C for h, each DS culture was visually examined by phase-contrast microscopy to assess the presence of spores. At that timepoint, aliquots of each DS culture were sonicated until more than % of cells had lysed. The sonicated samples were then centrifuged to remove debris and unlysed cells. Supernatant from each centrifuged DS culture lysates was analyzed for the presence of CPE using a previously described CPE Western immunoblot procedure (). Downloaded from on April, 01 by guest For Western immunoblot analysis of alternative sigma factor production, a 1. ml aliquot of a h DS cultures was centrifuged. The pellets were then resuspended in TE buffer and 1µl of 1
13 Ready to Use Lysozyme (Epicentre) was added. After 1 h incubation at C, SDS loading dye was added and the sample was boiled for min. Western blot analysis of these samples to detect the presence of SigE, SigK, SigF and SigG used a 1:00 dilution of rabbit antibodies raised specifically against purified recombinant B. subtilis sigma factors E, F, G or K produced by E. coli. These antibodies were kindly supplied by Drs. Richard Losick and Masaya Fujita Quantitative spore counts. Quantitative spore counts were determined as described previously (1). Briefly, an overnight DS culture of wild-type SM1, SM1::sigF, SM1::sigG, SM1sigFcomp, or SM1sigGcomp was heated at C for 0 min to kill the remaining vegetative cells and to promote germination of mature spores. Each heat-shocked suspension was then serially diluted from - to - with sterile water and plated onto BHI agar plates, which were incubated anaerobically overnight at C prior to colony counting. Quantitative spore counting was performed three times, each, for wild-type SM1, isogenic sigma factor mutants and complementing strains. Photomicroscopy. Spore formation in h DS cultures was documented using a Zeiss phase contrast microscope (00X total magnification). Results Downloaded from on April, 01 by guest 1
14 Construction of SM1 sigf and sigg null mutants. As shown in Fig. 1, inspection of the published C. perfringens strain SM1 genomic sequence () revealed that the sige, sigg and sigk ORFs cluster together, while the sigf ORF is separated from those ORFs by ~00 kb. While, i) a previous study () had demonstrated that sige and sigk are essential for CPE expression and sporulation by SM1 and ii) both SigF and SigG are necessary for B. subtilis sporulation (), the importance (if any) of sigf or sigg for either C. perfringens sporulation or CPE expression had not yet been experimentally evaluated. Since significant differences between B. subtilis and C. perfringens sporulation have been reported (), the current study used sigf and sigg null mutants to directly evaluate whether sigf and sigg are involved in CPE synthesis by, and sporulation of, SM1. The SM1 sigf and sigg null mutants were constructed by a Targetron-mediated insertional mutagenesis approach (). The identity of the putative mutants obtained from this mutagenesis procedure was first evaluated by PCR using primers corresponding to internal sigf ORF sequences or sigg ORF sequences (Fig. A). In wild-type SM1, these internal PCR primers amplified sigf- or sigg-specific products of ~0 bp or ~0 bp, respectively. However, using DNA from the intron-inactivated mutants, the same primers supported PCR amplification of products that are ~00 bp larger than the products amplified using wild-type SM1 template DNA. The larger size of these PCR products amplified from both putative mutants matched the expected size change resulting from an intron insertion into the sigf or sigg ORF. To specifically demonstrate the presence of an intron insertion in the sigf and sigg null mutants, and also to show that these mutants carried only a single intron insertion, Southern blot 1 Downloaded from on April, 01 by guest
15 analysis was performed using an intron-specific probe (Fig. B). As expected, this experiment detected no hybridization of the intron-specific probe to wild-type SM1 DNA. In contrast, these Southern blot results showed the presence of a single intron insertion in the SM1::sigF and SM1::sigG mutants Disruption of sigf or sigg production by the two mutants was then directly assessed using Western blotting (Fig., panels A and B). These analyses first demonstrated that wild-type SM1 produces both SigF and SigG during sporulation. Furthermore, these Western blots also showed the complete loss of SigF or SigG production by sporulating cultures of the sigf or sigg null mutants, respectively. Complementation of the SM1 sigf and sigg null mutants. Complementation of the SM1::sigG mutant was accomplished by PCR-cloning, from wild-type SM1, DNA corresponding to the sigg ORF and 00bp of upstream sequence. This PCR product was cloned into the pjir0 shuttle plasmid and then transformed into the sigg null mutant to create SM1::sigGComp. The presence of a wild-type sigg gene in this complementing strain was confirmed by PCR (data not shown). SigG production by SM1::sigGComp was then directly demonstrated by Western blotting (Fig. B). Complementation of the sigf mutant proved more complicated. Initial experiments indicated that transforming a plasmid carrying a PCR product corresponding to the sigf ORF and ~00bp of upstream sequence failed to restore SigF expression to SM1::sigF (data not shown). Therefore, it was considered that the sigf ORF might be transcribed as part of an operon. 1 Downloaded from on April, 01 by guest
16 To test this hypothesis, overlapping RT-PCR and PCR were first performed. The overlapping PCR primers for these analyses were designed to link sigf with three upstream genes, i.e., the ATP dependent protease gene, sporiiaa, and sporiiab, along with one downstream gene, i.e., sporvac (Fig. A). Using wild-type SM1 DNA as template for this overlap PCR assay, each of the five expected products was amplified (Fig. B), confirming the validity of the primers. However, using SM1 RNA template, the RT-PCR only amplified products from reactions R to R, indicating that the SporIIAA, SporIIAB and SigF ORFs are co-transcribed as a single mrna. Northern blot analysis (Fig. C) supported this finding by indicating that the sigf gene is transcribed as an ~1. kb mrna, matching the expected size of a single mrna encoding SporIIAA, SporIIAB and SigF. In addition, these Northern analyses also failed to detect the presence of any sigf mrna in the sigf null mutant, consistent with the Fig. A Western blot results. Since both the overlap RT-PCR and Northern blot results indicated expression of the sigf gene as part of a tricistronic operon containing two upstream genes, a PCR product containing those three genes and 00bp of upstream and downstream sequence was ligated into shuttle vector pjir0, followed by transformation of the resultant plasmid (pjir0sigfcomp) into the sigf null mutant. Both Western blot and Northern blot analyses demonstrated SigF expression by this complementing strain (Fig. A and C), named SM1::sigFComp. Downloaded from on April, 01 by guest 1
17 Regulation of sigma factor production by SigF and SigG. The sigf and sigg null mutants were first used to assess whether SigF or SigG might regulate production of each other. Western blotting (Fig. B) showed that the sigf mutant is impaired for production of SigG, while the SM1::sigFComp complementing strain produces near wild-type levels of SigG. In contrast, the sigg mutant was still able to produce wild-type levels of SigF (Fig. A). The sigf and sigg null mutants were next employed to test whether SigF or SigG might regulate the production of SigE or SigK, which have been previously tied to C. perfringens sporulation and the regulation of CPE production (). Western blots comparing SigE and SigK production by sporulating cultures of wild-type SM1 revealed that, as expected, h DS cultures grown at C produce both SigE and SigK (Fig., panels C and D). The presence of two visible bands for SigE and SigK on Western blot has been observed previously for C. perfingens () and those bands were suggested to correspond to pro-protein or mature, active forms of these two sigma factors. Compared to the Fig. wild-type SM1 results, Western blotting detected sharply reduced or no production of SigE or SigK by the SM1 sigf null mutant grown under the same h incubation conditions at C in DS medium (Fig., panels C and D). Furthermore, complementation of the sigf null mutant fully restored SigE and SigK production after a similar h incubation at C in DS medium (Fig., panels C and D). Downloaded from on April, 01 by guest 1
18 In contrast to these Fig. results indicating that SigF regulates production of SigE and SigK, inactivation of the sigg gene had no affect on production of either SigE or SigK in cultures grown for h in DS medium at C (Fig. ) Evidence that both SigF and SigG are essential for C. perfringens sporulation. To address whether SigF or SigG are required for sporulation by wild-type SM1, the sigf and sigg null mutants (or complementing strains of those mutants) was first examined using phase-contrast microscopy (Fig. ). This analysis showed that, by h incubation at C in DS sporulation medium, wild-type SM1 had already formed refractile spores, with an efficiency of > %. In contrast, the sigg and sigf null mutants failed to form any refractile spores after a similar h incubation in DS medium at C. Furthermore, both mutants remained unable to form refractile spores even when incubated for or h at C in DS medium (not shown). However, complementation of either mutant restored sporulation after an h incubation at C in DS medium, by which time >% of the complementing strain cells had formed refractile spores (Fig. ). The sporulating ability of SM1, the sigg and sigf null mutants, or complementing strains of those mutants was also directly compared by measuring formation of heat-resistant spores after h incubation at C in DS medium. Under these conditions, wild-type SM1 formed ~ heat-resistant spores/ml. However, under the same incubation conditions, no heat-resistant spores were detected for either the SM1 sigg or sigf null mutants. This inability of the sigg and sigf mutants to form heat-resistant spores was not simply due to slower spore production, since no heat-resistant spores were detected even when these mutants 1 Downloaded from on April, 01 by guest
19 were incubated at C in DS sporulation medium for, or days. Complementation of either sigma factor mutant restored the ability to form heat-resistant spores to near wild-type levels following an overnight incubation in DS medium at C. Specifically, SM1sigFcomp formed heat-resistant spores/ml and SM1sigGcomp formed heat-resistant spores/ml. Collectively, these results indicated that SigF and SigG are essential for formation of heat-resistant mature spores by SM1. CPE expression during sporulation is SigF-dependent, but does not require SigG. RT-PCR analyses showed that, for wild-type SM1 growing at C in DS medium, cpe gene transcription starts at ~- h and then increases further over the next several hours (Fig. ). In addition, Western blot analyses demonstrated that, under these same incubation conditions, CPE production by SM1 became detectable within h in DS cultures incubated at C and then increased markedly by - h (Figs. and ). In contrast, no cpe transcription or CPE production was observed in h cultures of SM1 grown at C in TGY medium, a condition that does not trigger C. perfringens sporulation (data not shown). As another control, no immunoreactivity with CPE antibody was observed using h DS cultures of the cpe-negative strain ATCC. Similar Western blot analyses of h DS cultures incubated at C were used to evaluate CPE production by the sigf and sigg null mutants. Results of those CPE Western blot studies detected no CPE production by the SM1::sigF strain (Fig. ). However, under the same incubation conditions, complementation of the sigf mutant to regain SigF expression also restored CPE expression production to near wild-type SM1 levels. Collectively, these results demonstrated that SigF is required for CPE expression. 1 Downloaded from on April, 01 by guest
20 In contrast, similar CPE Western blot experiments revealed that h DS cultures of the sigg null mutant still produce normal levels of CPE, as do similar cultures of the complementing strain for that mutant. Therefore, SigG is necessary for SM1 to produce spores but is not required for CPE production. Discussion The ability to sporulate is an integral contributor to several important C. perfringens diseases. For example, contamination of wounds with spores can lead to gas gangrene (). Spores also contribute to the transmission of C. perfringens type A food poisoning, particularly since survival of food poisoning strains is often facilitated by forming spores that are exceptionally resistant to food environment stresses such as heat, cold and preservatives (1-1, 1,, ). Furthermore, the sporulation process itself is essential for C. perfringens type A food poisoning, i.e., the enterotoxin (CPE) causing the gastrointestinal symptoms of this foodborne illness is produced only by sporulating cells (, 1, 1). Therefore, knowledge of C. perfringens sporulation is important to fully understand the pathogenesis of C. perfringens type A food poisoning and other C. perfringens diseases. While C. perfringens sporulation is still understood only at a rudimentary level, both similarities and differences from B. subtilis sporulation have been reported (). Regarding sporulation similarities, both bacteria were found to utilize the transcription factor Spo0A (, ) 0 Downloaded from on April, 01 by guest
21 and to express four alternative sigma factors that, for B. subtilis, have been shown to regulate sporulation-associated gene expression (, ). Among sporulation differences, C. perfringens lacks the phosphorelay that helps signal the start of B. subtilis sporulation (, ). Also, it was suggested that production of the four alternative sigma factors may be regulated differently in these two bacteria (). Given these apparent differences between B. subtilis vs. C. perfringens sporulation, it has been important to establish whether all four alternative sigma factors are required for C. perfringens sporulation. To begin addressing that question, Harry et al. showed that SigE and SigK are necessary for sporulation of C. perfringens strain SM1 (). The current results have demonstrated that SigF and SigG are also required for C. perfringens sporulation, i.e., mutants unable to produce SigF or SigG were completely blocked for spore formation, with complementation restoring sporulation to near wild-type levels. This complementation of sigf and sigg mutants back to near wild-type sporulation levels notably contrasts with results from the Harry et al. study (), where complementation only restored 1-% of wild-type sporulation ability to sige and sigk null mutants. Harry et al suggested their more limited complementation might be attributable to substantial wild-type sige and sigk transcription from upstream genes, an effect not encoded by their complementing plasmid. This would be similar to our finding that, as in B. subtilis (0), SM1 transcribes sigf as part of a spoiia tricistronic operon containing sigf (spoiiac), spoiiaa and spoiiab genes. This genetic similarity could suggest that, as in B. subtilis (0), SpoIIAA and SpoIIAB regulate SigF activity in SM1. Downloaded from on April, 01 by guest 1
22 The current results add important support for another proposed similarity between the sporulation of SM1 and B. subtilis, where SigE and SigK, but not SigF or SigG, are initially produced as inactive pro-proteins that are then proteolytically-processed to an active, mature form (, ). Specifically, our Western blots showed that antibodies raised against SigF or SigG only reacted with single proteins that correspond to the appropriate sigma factor based upon size and the absence of this immunoreactive band from Western blot lanes containing lysates of sigf or sigg null mutants. In B. subtilis, synthesis of the four alternative sigma factors begins during the initiation of sporulation and follows the temporal pattern: SigF, SigE, SigG and SigK (). In contrast, Harry et al () found that all four sigma factors rapidly accumulate after SM1 is inoculated into sporulation medium and that early accumulation of sigf transcript appeared to be SigKdependent, suggesting that SigK might act prior to SigF in the C. perfringens sporulation regulatory network. Therefore it was interesting that the current study detected little or no or SigK production by the sigf null mutant, although production of that sigma factor was observed after complementation to restore SigF production. As expanded on below, our current results are most consistent with SM1 sporulation involving a cascade of alternative sigma factors reminiscent of B. subtilis, where SigF regulates SigG, SigE and SigK production. Notably, this same classical sporulation-associated sigma factor regulatory cascade has also been described in nonpathogenic Clostridium acetobutylicum (). Harry et al. () also concluded that SigG regulation substantially differs between B. subtilis and C. perfringens. Whereas SigG production in B. subtilis is dependent upon both SigF Downloaded from on April, 01 by guest
23 and SigE, the Harry et al. () study had reported that a SM1 sige null mutant still produces SigG even though producing little or no SigE or SigF. However, our current study found that a SM1 sigf null mutant no longer produces SigG. Consistent with that finding, bioinformatics analysis identified a consensus sigf promoter sequence upstream of sigg. These findings would suggest that, as in B. subtilis, SigG production by C. perfringens requires SigF. As mentioned in the Introduction, the P1 cpe promoter possesses sequence similarity to a SigK-dependent promoter, while the P and P cpe promoter sequences resemble sequences found in SigE-dependent promoters (0). Consistent with those homologies, Harry et al recently used gusa fusion assays to conclude that cpe transcription is blocked in sige and sigk null mutants of SM1 (). Results from our current study now establish that cpe transcription and CPE production are also blocked in a SM1 sigf null mutant. However, we also found that, despite the previously reported () early appearance of sigg transcripts in sporulating SM1 cultures, a sigg null mutant still shows normal levels of cpe transcription and CPE production, thus documenting that not all sporulation-associated sigma factors are required for cpe transcription and CPE production. At least two possibilities could explain how SigF regulates cpe transcription. First, a cpe promoter might be SigF-dependent. This possibility seems less likely since promoter-mapping studies only identified SigE- and SigK-dependent cpe promoter sequences (0). A second possible explanation, shown in the Fig. model, might be that (as for B. subtilis and C. acetylobutylicum) SM1 uses SigF to regulate production of other sporulation-associated sigma factors. This model is supported by the current Western blot results showing little or no Downloaded from on April, 01 by guest
24 production of SigE and SigK by the sigf null mutant. This model also indicates that (like the situation with B. subtilis and C. acetylobutylicum (,, )) SigG production by SM1 apparently requires SigF, consistent with the Western blot results of our current study Because older studies (1) had suggested that CPE production levels may involve posttranscriptional regulatory effects not modeled by transcriptional fusion assays, the current study directly compared the timing of cpe transcription and CPE production in sporulating SM1 cultures. These analyses detected cpe transcription beginning ~ hour post-inoculation into DS medium. The appearance of CPE protein lagged slightly, becoming identifiable by Western blotting at - hours post-inoculation. These results largely agree with the transcriptional fusion assay results of Harry et al (), who noted cpe promoter-driven GusA activity ~- hours postinoculation into a slightly different sporulation medium. However, our results do not necessarily eliminate post-transcriptional contributions to CPE production, a topic worthy of further evaluation. In summary, there is now increasing evidence for both similarities and differences between the sporulation of C. perfringens versus B. subtilis. Sporulation similarities apparently include the involvement of Spo0A and SigF-mediated control of the other sporulation-associated sigma factors, while differences include the absence of a B. subtilis-like phosphorelay from C. perfringens. Future transcriptomic studies should identify transcripts regulated by each alternative sigma factor in C. perfringens. Another largely unstudied topic for C. perfringens sporulation concerns the initial signaling cascade that leads to Spo0A phosphorylation and triggers sporulation. Finally, results of this study indicate a previously unappreciated level of Downloaded from on April, 01 by guest
25 complexity for the regulation of cpe transcription (i.e., the involvement of SigF). However, in combination with the earlier results of Harry et al, the now demonstrated involvement of three sporulation-associated sigma factors in regulating cpe transcription offers an explanation for the ~0 year-old observation reporting a strong linkage between CPE production and sporulation Acknowledgements This research was generously supported by grant R AI1- from the National Institute of Allergy and Infectious Diseases. The authors thank Drs. Richard Losick and Masaya Fujita for supplying the sigma factor antibodies used in this study. Figure Legends Fig. 1. Arrangement of the sigma factor ORFs in C. perfringens SM1 based upon previous genome sequencing results (). A) Arrangement of the sigf locus in SM1. Results from the current study (see text) indicate that SigF, SporIIAB and SporIIAA are transcribed in one operon (dotted arrows). B) Arrangement of the sigg, sige and sigk locus in SM1. As indicated by the dotted black arrows, SigK and CPR_1 are also transcribed in one operon, according to (). A 1 kb scale marker is included below panel B for size comparisons. Fig.. Intron-based insertional mutagenesis to create SM1 sigf and sigg null mutants. A) Internal sigf and sigg-specific PCR results for wild-type SM1; SM1::sigF, and SM1::sigG. Migration of 0 bp DNA markers is shown at left and migration of 1 kb DNA markers is shown at right. B) Southern blot analysis of wild-type, SM1::sigF and Downloaded from on April, 01 by guest
26 SM1::sigG null mutant strains with a DIG-labeled, intron-specific probe. DNA size markers are shown at right Fig.. Western blot analyses of alternative sigma factor production by SM1. Shown are Western blot results for production of SigF (Panel A), SigG (Panel B), SigK (Panel C) and SigE (Panel D) by, as indicated, DS cultures of wild-type, SM1::sigF, SM1sigFcomp, SM1::sigG, and SM1sigGcomp grown for h at C. See Results for discussion of the two SigE or SigK bands present in some lanes. Fig.. The sigf ORF is transcribed as part of a tricistronic operon. A) Arrangement of the sigf locus in SM1, based upon sequencing results (). Location of primers used in panel B for overlap PCR is indicated by R1-R. B) Overlap PCR analysis of the sigf locus region in five reactions (R1 to R). The upper figure shows results for a DNA PCR and the lower figure shows results for a RNA RT-PCR using reaction R1 to R primers (Table 1). Size markers are shown at right. C) Northern blot analysis using a DIG-labeled sigf probe. Results shown are for wild-type, SM1::sigF and SM1sigFcomp strains. Size markers are shown at left. Fig.. Formation of phase-refractile spores by wild-type SM1, the sigf and sigg null mutants, and complementing strains. Shown are DS cultures of each strain after h growth at C in DS. Photographs were taken using a phase-contrast microscope (00 ). Fig.. Kinetics of cpe gene expression and CPE production by DS cultures of SM1. A) RT- PCR analysis of cpe gene transcription in cultures growing from 1- hr in DS medium at C. Downloaded from on April, 01 by guest
27 The Spo0A gene was used as a housekeeping gene and a sample lacking RT was run to show the absence of DNA contamination in these samples. The size of products is shown at right of each gel based upon comparison against DNA size markers (not shown). B) Western blot analyses of CPE production after 1- hr growth in DS medium at C. To show that CPE production is dependent on sporulation, this figure shows the absence of CPE production by an h culture of SM1 grown at C in TGY, a medium that does not induce sporulation. Size of the immunoreactive protein is indicated at right, based upon migration of prestained markers (not shown). Fig.. Comparison of CPE production by wild-type SM1, sigf and sigg null mutants and complementing strains. Western blot results for CPE production are shown for sporulating hr DS cultures incubated at C. As controls, similar analyses are shown for an h DS culture of CPE-negative strain ATCC grown at C and a lane run with purified CPE. The size of the immuno-reactive proteins is indicated at right, based upon migration of prestained markers (not shown). Fig.. Model describing sigma factor regulation of C. perfringens CPE production and sporulation. The model incorporates findings from the current work and research by Harry et al (). Not shown is the proven involvement of Spo0A in C. perfringens sporulation () since it is not yet clear how that protein leads to sporulation-associated sigma factor production by C. perfringens. Downloaded from on April, 01 by guest
28 Reference: Bannam, T. L., and J. I. Rood. 1. Clostridium perfringens-escherichia coli shuttle vectors that carry single antibiotic resistance determinants. Plasmid. :-.. Carman, R. J. 1. Clostridium perfringens in spontaneous and antibiotic-associated diarrhoea of man and other animals. Rev. Med. Microbiol., supplement 1:S-S.. Chen, Y., B. A. McClane, D. J. Fisher, J. I. Rood, and P. Gupta. 00. Construction of an alpha toxin gene knockout mutant of Clostridium perfringens type A by use of a mobile group II intron. Appl. Environ. Microbiol. 1:-.. Czeczulin, J. R., R. E. Collie, and B. A. McClane. 1. Regulated expression of Clostridium perfringens enterotoxin in naturally cpe-negative type A, B, and C isolates of C. perfringens. Infect. Immun. : Czeczulin, J. R., P. C. Hanna, and B. A. McClane. 1. Cloning, nucleotide sequencing, and expression of the Clostridium perfringens enterotoxin gene in Escherichia coli. Infect. Immun. 1:-.. Haldenwang, W. G. 1. The sigma factors of Bacillus subtilis. Microbiol. Rev :1-0.. Harry, K. H., R. Zhou, L. Kroos, and S. B. Melville. 00. Sporulation and Enterotoxin (CPE) synthesis are controlled by the sporulation-specific factors SigE and SigK in Clostridium perfringens. J. Bacteriol. :-.. Huang, I. H., M. Waters, R. R. Grau, and M. R. Sarker. 00. Disruption of the gene (spo0a) encoding sporulation transcription factor blocks endospore formation and Downloaded from on April, 01 by guest
29 enterotoxin productioin in enterotoxigenic Clostridium perfringens type A. FEMS Microbiol. Lett. :-0.. Jones, S. W., C. J. Paredes, B. Tracy, N. Cheng, R. Sillers, R. S. Senger, and E. T. Papoutsakis. 00. The transcriptional program underlying the physiology of clostridial sporulation. Genome Biology. :R.. Kokai-Kun, J. F., J. G. Songer, J. R. Czeczulin, F. Chen, and B. A. McClane. 1. Comparison of Western immunoblots and gene detection assays for identification of potentially enterotoxigenic isolates of Clostridium perfringens. J. Clin. Microbiol. :-.. Kroos, L. 00. The Bacillus and Myxococcus developmental netwoorks and their transcriptional regulators.. Annu. Rev. Genet. 1: Labbe, R. G., and C. L. Duncan. 1. Evidence for stable messenger ribonucleic acid during sporulation and enterotoxin synthesis by Clostridium perfringens type A. J. Bacteriol. 1:-. 1. Li, J., and B. A. McClane. 00. A novel small acid soluble protein variant is important for spore resistance of most Clostridium perfringens food poisoning isolates. PLoS Pathogens :e Li, J., and B. A. McClane. 00. Comparative effects of osmotic, sodium nitrite-induced, and ph-induced stress on growth and survival of Clostridium perfringens type A isolates carrying chromosomal or plasmid-borne enterotoxin genes. Appl. Environ. Microbiol. :0-. Downloaded from on April, 01 by guest
30 Li, J., and B. A. McClane. 00. Further comparison of temperature effects on growth and survival of Clostridium perfringens type A isolates carrying a chromosomal or plasmid-borne enterotoxin gene. Appl. Environ. Microbiol. : Li, J., K. Miyamoto, and B. A. McClane. 00. Comparison of virulence plasmids among Clostridium perfringens type E isolates. Infect. Immun. : Li, J., D. Paredes-Sabja, M. R. Sarker, and B. A. McClane. 00. Further characterization of Clostridium perfringens small acid soluble protein- (Ssp) properties and expression. PLoS One :e. 1. McClane, B. A. 00. Clostridium perfringens, p. -. In M. P. Doyle and L. R. Beuchat (ed.), Food Microbiology, rd ed. ASM press, Washington D.C. 1. McClane, B. A., F. A. Uzal, M. F. Miyakawa, D. Lyerly, and T. Wilkins. 00. The Enterotoxic Clostridia, p. -. In M. Dworkin, S. Falkow, E. Rosenburg, H. Schleifer, and E. Stackebrandt (ed.), The Prokaryotes, rd ed. Springer NY, New York. 0. Min, K., C. M. Hilditch, B. Diedetich, J. Errington, and M. D. Yudkin. 1. σf, the first compartment-specific transcription factor of Bacillus subtilis, is regulated by an antiσ factor that is also a protein kinase. Cell. :-. 1. Miyamoto, K., Q. Wen, and B. A. McClane. 00. Multiplex PCR genotyping assay that distinguishes between isolates of Clostridium perfringens type A carrying a chromosomal enterotoxin gene (cpe) locus, a plasmid cpe locus with an IS10-like sequence or a plasmid cpe locus with an IS1 sequence. J. Clin. Microbiol. 1:1-1.. Myers, G. S., D. A. Rasko, J. K. Cheung, J. Ravel, R. Seshadri, R. T. DeBoy, Q. Ren, J. Varga, M. M. Awad, L. M. Brinkac, S. C. Daugherty, D. H. Haft, R. J. Dodson, R. 0 Downloaded from on April, 01 by guest
31 Madupu, W. C. Nelson, M. J. Rosovitz, S. A. Sullivan, K. H., G. I. Dimitrov, K. L. Watkins, S. Mulligan, J. Benton, D. Radune, D. J. Fisher, H. S. Atkins, T. Hiscox, B. H. Jost, S. J. Billington, J. G. Songer, B. A. McClane, R. W. Titball, J. I. Rood, S. B. Melville, and I. T. Paulsen. 00. Skewed genomic variability in strains of the toxigenic bacterial pathogen, Clostridium perfringens. Genome Res. 1:1-0.. Orsburn, B., S. B. Melville, and D. Popham. 00. Factors contributing to heat resistance of Clostridium perfringens endospores. Appl. Environ. Microbiol. :-.. Sarker, M. R., R. J. Carman, and B. A. McClane. 1. Inactivation of the gene (cpe) encoding Clostridium perfringens enterotoxin eliminates the ability of two cpe-positive C. perfringens type A human gastrointestinal disease isolates to affect rabbit ileal loops. Molec. Microbiol. :-.. Sarker, M. R., R. P. Shivers, S. G. Sparks, V. K. Juneja, and B. A. McClane Comparative experiments to examine the effects of heating on vegetative cells and spores of Clostridium perfringens isolates carrying plasmid versus chromosomal enterotoxin genes. Appl. Environ. Microbiol. :-0.. Sayeed, S., J. Li, and B. A. McClane. 00. Virulence plasmid diversity in Clostridium perfringens type D isolates. Infect. Immun. :1-.. Shimizu, T., K. Ohtani, H. Hirakawa, K. Ohshima, A. Yamashita, T. Shiba, N. Ogasawara, M. Hattori, S. Kuhara, and H. Hayashi. 00. Complete genome sequence of Clostridium perfringens, an anaerobic flesh-eater. Proc. Natl. Acad. Sci. USA. :-01. Downloaded from on April, 01 by guest 1
32 . Stevens, D. L., and J. I. Rood. 00. Histotoxic Clostridia. p. 1-. In V. A. Fischetti, R. P. Novick, J. J. Ferretti, D. A. Portnoy, and J. I. Rood (ed.), Gram-positive pathogens, nd ed. ASM press, Washington, DC.. Vidal, J. E., K. Ohtani, T. Shimizu, and B. A. McClane. 00. Contact with enterocyte-like Caco cells induces rapid upregulation of toxin production by Clostridium perfringens type C isolates. Cell Microbiol. : Zhao, Y., and S. B. Melville. 1. Identification and characterization of sporulationdependent promoters upstream of the enterotoxin gene (cpe) of Clostridium perfringens. J. Bacteriol. :1-1. Downloaded from on April, 01 by guest
33 Table 1. Primers using sigf operon overlap PCR Primers Sequence Product size (bp) sigfcomnf -GTGTAAATGGATTGTCTGTTATAG- bp sigfnr -TTCTGCACTATGATGATCTAATTC- sigfcompnf -AATTAGATCATCATAGTGCAGAAG- 00bp sigfr1 sigfcompf sigfkor sigff sigfr sigffn sigfr -CTTCAATAGTTGGATCTAACTGAG- -CTCAGTTAGATCCAACTATTGAAG- -AGTTCTTCCTTATTGTAATGAAGC- -GCTTCATTACAATAAGGAAGAACT- -CTCTATTCTAGAAACTTGAACTTG- -CAAGTTCAAGTTTCTAGAATAGAG- -CTATTTTATCATATAATCCAAGTCC- bp bp 0bp Downloaded from on April, 01 by guest
34 Fig. 1 (A) sporvae sporvac sporiiab sporvad sigf sporiiaa Targetron KO-/ sense (B) CPR_ spoiiga CPR_1 sigg sige ftsz ftsa pilt sigk Targetron KO-1/ sense 1kb Downloaded from on April, 01 by guest
 SigG SM1::SigG Marker kb")

35 Fig. 00bp 00bp 00bp Marker SigF SM1 SM1::SigF SM1 (A) SigG SM1::SigG Marker kb 1kb SM1::sigF SM1::sigG WT (B) kb kb Downloaded from on April, 01 by guest

36 Fig. SigF SigG SM1 SM1::sigF SM1SigFComp SM1::sigG SM1SigGComp (A) (B) Downloaded from SigK SigE (C) (D) on April, 01 by guest

 0bp SporVAC SporVADSporVAE 1.kb 1.")

37 Fig (B) (A) bp ATP dependent protease R1 R EcoRI Sig F complement bp 0bp SporIIAASporIIAB bp SigF R1 R R R R R R spo0a DNA-PCR RNA-RT-PCR 00bp 00bp 0bp 00bp 00bp 0bp (C) 0bp SporVAC SporVADSporVAE 1.kb 1.0kb R PstI SM1 SM1::sigF Northern blot SM1sigFcomp Downloaded from on April, 01 by guest




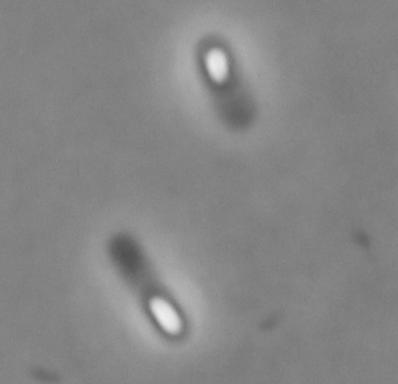
38 Fig. SM1 SM1::sigF SM1sigFcomp SM1::sigG SM1sigGcomp Downloaded from on April, 01 by guest
39 Fig. (A) 1hr hr hr hr hr hr hr DNA spo0a-rt 00bp spo0a+rt 00bp cpe+rt 00bp Downloaded from (B) TGYhr DShr DS1hr DShr DShr DShr DShr DShr DShr KD on April, 01 by guest

40 Fig. SM1::sigF SM1sigFcomp SM1::sigG SM1sigGcomp SM1 ATCC CPE Downloaded from KD on April, 01 by guest
AND BRUCE A. MCCLANE 1 *
 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 2000, p. 3234 3240 Vol. 66, No. 8 0099-2240/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Comparative Experiments To Examine
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 2000, p. 3234 3240 Vol. 66, No. 8 0099-2240/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Comparative Experiments To Examine
AND BRUCE A. MCCLANE 1 *
 JOURNAL OF CLINICAL MICROBIOLOGY, Mar. 2001, p. 883 888 Vol. 39, No. 3 0095-1137/01/$04.00 0 DOI: 10.1128/JCM.39.3.883 888.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Genotyping
JOURNAL OF CLINICAL MICROBIOLOGY, Mar. 2001, p. 883 888 Vol. 39, No. 3 0095-1137/01/$04.00 0 DOI: 10.1128/JCM.39.3.883 888.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Genotyping
and Cellular Biology of Rosario, Sala 9, Rosario 2000, Argentina
 FEMS Microbiology Letters 233 (2004) 233 240 www.fems-microbiology.org Disruption of the gene (spo0a) encoding sporulation transcription factor blocks endospore formation and enterotoxin production in
FEMS Microbiology Letters 233 (2004) 233 240 www.fems-microbiology.org Disruption of the gene (spo0a) encoding sporulation transcription factor blocks endospore formation and enterotoxin production in
Further Characterization of Clostridium perfringens Small Acid Soluble Protein-4 (Ssp4) Properties and Expression
 Properties and Expression") Further Characterization of Clostridium perfringens Small Acid Soluble Protein-4 (Ssp4) Properties and Expression Jihong Li 1, Daniel Paredes-Sabja 2, Mahfuzur R. Sarker 2,3, Bruce A. McClane 1 * 1 Department
Further Characterization of Clostridium perfringens Small Acid Soluble Protein-4 (Ssp4) Properties and Expression Jihong Li 1, Daniel Paredes-Sabja 2, Mahfuzur R. Sarker 2,3, Bruce A. McClane 1 * 1 Department
Characterization of germinants and their receptors for spores of non-food-borne Clostridium perfringens strain F4969
 Microbiology (2016), 162, 1972 1983 DOI 10.1099/mic.0.000378 Characterization of germinants and their receptors for spores of non-food-borne Clostridium perfringens strain F4969 Saeed Banawas, 1,2,3 Daniel
Microbiology (2016), 162, 1972 1983 DOI 10.1099/mic.0.000378 Characterization of germinants and their receptors for spores of non-food-borne Clostridium perfringens strain F4969 Saeed Banawas, 1,2,3 Daniel
Association of beta2 toxin production with Clostridium perfringens type A human gastrointestinal disease isolates carrying a plasmid enterotoxin gene
 Blackwell Science, LtdOxford, UKMMIMolecular Microbiology1365-2958Blackwell Publishing Ltd, 2005? 2005563747762Original rticlecpb2/cpe-positive C. perfringens disease isolatesd. J. Fisher et al. Molecular
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology1365-2958Blackwell Publishing Ltd, 2005? 2005563747762Original rticlecpb2/cpe-positive C. perfringens disease isolatesd. J. Fisher et al. Molecular
Clostridium perfringens Spore Germination: Characterization of Germinants and Their Receptors
 JOURNAL OF BACTERIOLOGY, Feb. 2008, p. 1190 1201 Vol. 190, No. 4 0021-9193/08/$08.00 0 doi:10.1128/jb.01748-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Clostridium perfringens
JOURNAL OF BACTERIOLOGY, Feb. 2008, p. 1190 1201 Vol. 190, No. 4 0021-9193/08/$08.00 0 doi:10.1128/jb.01748-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Clostridium perfringens
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Comparison of Virulence Plasmids among Clostridium perfringens Type E Isolates
 INFECTION AND IMMUNITY, Apr. 2007, p. 1811 1819 Vol. 75, No. 4 0019-9567/07/$08.00 0 doi:10.1128/iai.01981-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Comparison of Virulence
INFECTION AND IMMUNITY, Apr. 2007, p. 1811 1819 Vol. 75, No. 4 0019-9567/07/$08.00 0 doi:10.1128/iai.01981-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Comparison of Virulence
Characterization of Clostridium perfringens Spores That Lack SpoVA Proteins and Dipicolinic Acid
 JOURNAL OF BACTERIOLOGY, July 2008, p. 4648 4659 Vol. 190, No. 13 0021-9193/08/$08.00 0 doi:10.1128/jb.00325-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. Characterization
JOURNAL OF BACTERIOLOGY, July 2008, p. 4648 4659 Vol. 190, No. 13 0021-9193/08/$08.00 0 doi:10.1128/jb.00325-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. Characterization
Data Sheet. Azide Cy5 RNA T7 Transcription Kit
 Cat. No. Size 1. Description PP-501-Cy5 10 reactions à 40 µl For in vitro use only Quality guaranteed for 12 months Store all components at -20 C. Avoid freeze and thaw cycles. DBCO-Sulfo-Cy5 must be stored
Cat. No. Size 1. Description PP-501-Cy5 10 reactions à 40 µl For in vitro use only Quality guaranteed for 12 months Store all components at -20 C. Avoid freeze and thaw cycles. DBCO-Sulfo-Cy5 must be stored
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha
 Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha Plasmids 1. Extrachromosomal DNA, usually circular-parasite 2. Usually encode ancillary
Gene expression in prokaryotic and eukaryotic cells, Plasmids: types, maintenance and functions. Mitesh Shrestha Plasmids 1. Extrachromosomal DNA, usually circular-parasite 2. Usually encode ancillary
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
A Gene (sleb) Encoding a Spore Cortex-Lytic Enzyme from Bacillus subtilis and Response of the Enzyme to
 Encoding a Spore Cortex-Lytic Enzyme from Bacillus subtilis and Response of the Enzyme to") JOURNAL OF BACTERIOLOGY, Oct. 1996, p. 6059 6063 Vol. 178, No. 20 0021-9193/96/$04.00 0 Copyright 1996, American Society for Microbiology A Gene (sleb) Encoding a Spore Cortex-Lytic Enzyme from Bacillus
JOURNAL OF BACTERIOLOGY, Oct. 1996, p. 6059 6063 Vol. 178, No. 20 0021-9193/96/$04.00 0 Copyright 1996, American Society for Microbiology A Gene (sleb) Encoding a Spore Cortex-Lytic Enzyme from Bacillus
of the work reported here was to define the point in the developmental process at which the curing salts act to prevent outgrowth.
 APPLIED MICROBIOLOGY, Feb. 1968, p. 406-411 Copyright 1968 American Society for Microbiology Vol. 16, No. 2 Printed in U.S.A. Effect of Sodium Nitrite, Sodium Chloride, and Sodium Nitrate on Germination
APPLIED MICROBIOLOGY, Feb. 1968, p. 406-411 Copyright 1968 American Society for Microbiology Vol. 16, No. 2 Printed in U.S.A. Effect of Sodium Nitrite, Sodium Chloride, and Sodium Nitrate on Germination
Component Product # Product # Cell Lysis Reagent 100 ml 500 ml Product Insert 1 1
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Cell Lysis Reagent Product # 18800 (100 ml) Product # 18801 (500
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Cell Lysis Reagent Product # 18800 (100 ml) Product # 18801 (500
mrna Isolation Kit for Blood/Bone Marrow For isolation mrna from blood or bone marrow lysates Cat. No
 For isolation mrna from blood or bone marrow lysates Cat. No. 1 934 333 Principle Starting material Application Time required Results Key advantages The purification of mrna requires two steps: 1. Cells
For isolation mrna from blood or bone marrow lysates Cat. No. 1 934 333 Principle Starting material Application Time required Results Key advantages The purification of mrna requires two steps: 1. Cells
Killing of Bacillus Spores by High-Intensity Ultraviolet Light
 Killing of Bacillus Spores by High-Intensity Ultraviolet Light STUDY ON EFFECTS OF PULSED LIGHT Abraham L. Sonenshein, PhD Professor and Deputy Chair Department of Molecular Biology and Microbiology Tufts
Killing of Bacillus Spores by High-Intensity Ultraviolet Light STUDY ON EFFECTS OF PULSED LIGHT Abraham L. Sonenshein, PhD Professor and Deputy Chair Department of Molecular Biology and Microbiology Tufts
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
Growth from Spores of Clostridium perfringens
 APpuE MicRoBioLOGY, Feb. 1970, p. 353-359 Copyright 1970 American Society for Microbiology Vol. 19, No. 2 Printed in U.S.A. Growth from Spores of Clostridium perfringens in the Presence of Sodium Nitrite'
APpuE MicRoBioLOGY, Feb. 1970, p. 353-359 Copyright 1970 American Society for Microbiology Vol. 19, No. 2 Printed in U.S.A. Growth from Spores of Clostridium perfringens in the Presence of Sodium Nitrite'
Biology 112 Practice Midterm Questions
 Biology 112 Practice Midterm Questions 1. Identify which statement is true or false I. Bacterial cell walls prevent osmotic lysis II. All bacterial cell walls contain an LPS layer III. In a Gram stain,
Biology 112 Practice Midterm Questions 1. Identify which statement is true or false I. Bacterial cell walls prevent osmotic lysis II. All bacterial cell walls contain an LPS layer III. In a Gram stain,
AN ABSTRACT OF THE THESIS OF. Colton B. Bond for the degree of Master of Science in Microbiology presented on June 3, 2011.
 AN ABSTRACT OF THE THESIS OF Colton B. Bond for the degree of Master of Science in Microbiology presented on June 3, 2011. Title: Role of Plc in the Germination of Clostridium perfringens Food Poisoning
AN ABSTRACT OF THE THESIS OF Colton B. Bond for the degree of Master of Science in Microbiology presented on June 3, 2011. Title: Role of Plc in the Germination of Clostridium perfringens Food Poisoning
Redacted for Privacy
 AN ABSTRACT OF THE THESIS OF Ben Harrison for the degree of Master of Science in Microbiology presented on June 19, 2003. Title: Genotypic and Phenotypic Characterization of Enterotoxigenic Clostridium
AN ABSTRACT OF THE THESIS OF Ben Harrison for the degree of Master of Science in Microbiology presented on June 19, 2003. Title: Genotypic and Phenotypic Characterization of Enterotoxigenic Clostridium
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
Detection of Enterotoxic Bacillus cereus Producing Hemolytic and Non Hemolytic Enterotoxins by PCR Test
 Polish Journal of Microbiology 2006, Vol. 55, No 2, 113 118 Detection of Enterotoxic Bacillus cereus Producing Hemolytic and Non Hemolytic Enterotoxins by PCR Test EL BIETA O TUSZAK-WALCZAK *, PIOTR WALCZAK
Polish Journal of Microbiology 2006, Vol. 55, No 2, 113 118 Detection of Enterotoxic Bacillus cereus Producing Hemolytic and Non Hemolytic Enterotoxins by PCR Test EL BIETA O TUSZAK-WALCZAK *, PIOTR WALCZAK
Microbial DNA qpcr Multi-Assay Kit Clostridium perfringens Pathogenicity
 Microbial DNA qpcr Multi-Assay Kit Clostridium perfringens Pathogenicity Cat. no. 330043 BBID-1507ZR-3 For real-time PCR-based, application-specific microbial identification or profiling The Clostridium
Microbial DNA qpcr Multi-Assay Kit Clostridium perfringens Pathogenicity Cat. no. 330043 BBID-1507ZR-3 For real-time PCR-based, application-specific microbial identification or profiling The Clostridium
Epidemiological studies on Clostridium perfringens food poisoning in retail foods
 Rev. Sci. Tech. Off. Int. Epiz., 2017, 36 (3),... -... Epidemiological studies on Clostridium perfringens food poisoning in retail foods This paper (No. 25092017-00109-EN) has been peer-reviewed, accepted,
Rev. Sci. Tech. Off. Int. Epiz., 2017, 36 (3),... -... Epidemiological studies on Clostridium perfringens food poisoning in retail foods This paper (No. 25092017-00109-EN) has been peer-reviewed, accepted,
CYTOLOGICAL CHANGES IN AGING BACTERIAL CULTURES
 CYTOLOGICAL CHANGES IN AGING BACTERIAL CULTURES B. R. CHATTERJEE AND ROBERT P. WILLIAMS Department of Microbiology, Baylor University College of Medicine, Houston, Texas Received for publication March
CYTOLOGICAL CHANGES IN AGING BACTERIAL CULTURES B. R. CHATTERJEE AND ROBERT P. WILLIAMS Department of Microbiology, Baylor University College of Medicine, Houston, Texas Received for publication March
U.S. Patent No. 9,051,563 and other pending patents. Ver
 INSTRUCTION MANUAL Direct-zol 96 RNA Catalog Nos. R2054, R2055, R2056 & R2057 Highlights Quick, 96-well purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and all other
INSTRUCTION MANUAL Direct-zol 96 RNA Catalog Nos. R2054, R2055, R2056 & R2057 Highlights Quick, 96-well purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and all other
About OMICS Group Conferences
 About OMICS Group OMICS Group International is an amalgamation of Open Access publications and worldwide international science conferences and events. Established in the year 2007 with the sole aim of
About OMICS Group OMICS Group International is an amalgamation of Open Access publications and worldwide international science conferences and events. Established in the year 2007 with the sole aim of
Epsilon-Toxin Plasmids of Clostridium perfringens Type D Are Conjugative
 JOURNAL OF BACTERIOLOGY, Nov. 2007, p. 7531 7538 Vol. 189, No. 21 0021-9193/07/$08.00 0 doi:10.1128/jb.00767-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Epsilon-Toxin Plasmids
JOURNAL OF BACTERIOLOGY, Nov. 2007, p. 7531 7538 Vol. 189, No. 21 0021-9193/07/$08.00 0 doi:10.1128/jb.00767-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Epsilon-Toxin Plasmids
Three types of RNA polymerase in eukaryotic nuclei
 Three types of RNA polymerase in eukaryotic nuclei Type Location RNA synthesized Effect of α-amanitin I Nucleolus Pre-rRNA for 18,.8 and 8S rrnas Insensitive II Nucleoplasm Pre-mRNA, some snrnas Sensitive
Three types of RNA polymerase in eukaryotic nuclei Type Location RNA synthesized Effect of α-amanitin I Nucleolus Pre-rRNA for 18,.8 and 8S rrnas Insensitive II Nucleoplasm Pre-mRNA, some snrnas Sensitive
Supplementary Information
 Supplementary Information Supplementary figures % Occupancy 90 80 70 60 50 40 30 20 Wt tol-1(nr2033) Figure S1. Avoidance behavior to S. enterica was not observed in wild-type or tol-1(nr2033) mutant nematodes.
Supplementary Information Supplementary figures % Occupancy 90 80 70 60 50 40 30 20 Wt tol-1(nr2033) Figure S1. Avoidance behavior to S. enterica was not observed in wild-type or tol-1(nr2033) mutant nematodes.
Substrate Requirements for Regulated Intramembrane Proteolysis of Bacillus subtilis Pro- K
 JOURNAL OF BACTERIOLOGY, Feb. 2005, p. 961 971 Vol. 187, No. 3 0021-9193/05/$08.00 0 doi:10.1128/jb.187.3.961 971.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Substrate
JOURNAL OF BACTERIOLOGY, Feb. 2005, p. 961 971 Vol. 187, No. 3 0021-9193/05/$08.00 0 doi:10.1128/jb.187.3.961 971.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Substrate
CHAPTER : Prokaryotic Genetics
 CHAPTER 13.3 13.5: Prokaryotic Genetics 1. Most bacteria are not pathogenic. Identify several important roles they play in the ecosystem and human culture. 2. How do variations arise in bacteria considering
CHAPTER 13.3 13.5: Prokaryotic Genetics 1. Most bacteria are not pathogenic. Identify several important roles they play in the ecosystem and human culture. 2. How do variations arise in bacteria considering
AN ABSTRACT OF THE THESIS OF
 AN ABSTRACT OF THE THESIS OF Saeed S. Banawas for the degree of Master of Science in Veterinary Science presented on November 11, 2010. Title: Characterization of Germinant Receptors in Clostridium perfringens
AN ABSTRACT OF THE THESIS OF Saeed S. Banawas for the degree of Master of Science in Veterinary Science presented on November 11, 2010. Title: Characterization of Germinant Receptors in Clostridium perfringens
Integration and amplification of the Bacillus sp cellulase gene in the Bacillus subtilis 168 chromosome
 J. Gen. Appl. Microbiol., 44, 107 111 (1998) Short Communication Integration and amplification of the Bacillus sp. 79-23 cellulase gene in the Bacillus subtilis 168 chromosome Kyung Hwa Jung, Dae-Hee Lee,
J. Gen. Appl. Microbiol., 44, 107 111 (1998) Short Communication Integration and amplification of the Bacillus sp. 79-23 cellulase gene in the Bacillus subtilis 168 chromosome Kyung Hwa Jung, Dae-Hee Lee,
2. Yeast two-hybrid system
 2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
2. Yeast two-hybrid system I. Process workflow a. Mating of haploid two-hybrid strains on YPD plates b. Replica-plating of diploids on selective plates c. Two-hydrid experiment plating on selective plates
This is a repository copy of Evidence for antibiotic induced Clostridium perfringens diarrhoea.
 This is a repository copy of Evidence for antibiotic induced Clostridium perfringens diarrhoea. White Rose Research Online URL for this paper: http://eprints.whiterose.ac.uk/256/ Article: Modi, N. and
This is a repository copy of Evidence for antibiotic induced Clostridium perfringens diarrhoea. White Rose Research Online URL for this paper: http://eprints.whiterose.ac.uk/256/ Article: Modi, N. and
ydci GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC
 Table S1. DNA primers used in this study. Name ydci P1ydcIkd3 Sequence GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC Kd3ydcIp2 lacz fusion YdcIendP1 YdcItrgP2 GAC AGC
Table S1. DNA primers used in this study. Name ydci P1ydcIkd3 Sequence GTC TGT TTG AAC GCG GGC GAC TGG GCG CGC AAT TAA CGG TGT GTA GGC TGG AGC TGC TTC Kd3ydcIp2 lacz fusion YdcIendP1 YdcItrgP2 GAC AGC
Use of the 3M Molecular Detection System for Salmonella and Listeria spp.
 Use of the 3M Molecular Detection System for Salmonella and Listeria spp. March 11, 213 Prof Steve Forsythe Pathogen Research Centre, School of Science and Technology Nottingham Trent University Clifton
Use of the 3M Molecular Detection System for Salmonella and Listeria spp. March 11, 213 Prof Steve Forsythe Pathogen Research Centre, School of Science and Technology Nottingham Trent University Clifton
Helical Macrofiber Formation in Bacillus subtilis: Inhibition by Penicillin G
 JOURNAL OF BACTERIOLOGY, June 1984, p. 1182-1187 0021-9193/84/061182-06$02.00/0 Copyright C 1984, American Society for Microbiology Vol. 158, No. 3 Helical Macrofiber Formation in Bacillus subtilis: Inhibition
JOURNAL OF BACTERIOLOGY, June 1984, p. 1182-1187 0021-9193/84/061182-06$02.00/0 Copyright C 1984, American Society for Microbiology Vol. 158, No. 3 Helical Macrofiber Formation in Bacillus subtilis: Inhibition
Evidence for cyclic-di-gmp-mediated signaling pathway in Bacillus subtilis by Chen Y. et al.
 Supplemental materials for Evidence for cyclic-di-gmp-mediated signaling pathway in Bacillus subtilis by Chen Y. et al. 1. Table S1. Strains used in this study 2. Table S2. Plasmids used in this study
Supplemental materials for Evidence for cyclic-di-gmp-mediated signaling pathway in Bacillus subtilis by Chen Y. et al. 1. Table S1. Strains used in this study 2. Table S2. Plasmids used in this study
Mini-Tn7 Derivative Construction and Characterization. Mini-Tn7 derivatives for
 Supplemental Methods Mini-Tn7 Derivative Construction and Characterization. Mini-Tn7 derivatives for constitutive expression of fluorescent proteins in S. oneidensis were constructed as follows. The EcoRI-XbaI
Supplemental Methods Mini-Tn7 Derivative Construction and Characterization. Mini-Tn7 derivatives for constitutive expression of fluorescent proteins in S. oneidensis were constructed as follows. The EcoRI-XbaI
World Journal of Pharmaceutical Research SJIF Impact Factor 8.074
 SJIF Impact Factor 8.074 Volume 7, Issue 5, 966-973. Research Article ISSN 2277 7105 MOLECULAR DETECTION OF ENTEROTOXIGENIC ISOLATES OF SALMONELLA TYPHIMURIUM, SHIGELLA FLEXNERI AND STAPHYLOCOCCUS AUREUS
SJIF Impact Factor 8.074 Volume 7, Issue 5, 966-973. Research Article ISSN 2277 7105 MOLECULAR DETECTION OF ENTEROTOXIGENIC ISOLATES OF SALMONELLA TYPHIMURIUM, SHIGELLA FLEXNERI AND STAPHYLOCOCCUS AUREUS
Bacterial strains, plasmids, and growth conditions. Bacterial strains and
 I Text I Materials and Methods acterial strains, plasmids, and growth conditions. acterial strains and plasmids used in this study are listed in I Table. almonella enterica serovar Typhimurium strains
I Text I Materials and Methods acterial strains, plasmids, and growth conditions. acterial strains and plasmids used in this study are listed in I Table. almonella enterica serovar Typhimurium strains
Salt-sensitivity of SigH and Spo0A prevents Bacillus subtilis sporulation at high osmolarity avoiding death during cellular differentiation
 Salt-sensitivity of SigH and Spo0A prevents Bacillus subtilis sporulation at high osmolarity avoiding death during cellular differentiation Nils Widderich 1,, Christopher D.A. Rodrigues 2,, Fabian M. Commichau
Salt-sensitivity of SigH and Spo0A prevents Bacillus subtilis sporulation at high osmolarity avoiding death during cellular differentiation Nils Widderich 1,, Christopher D.A. Rodrigues 2,, Fabian M. Commichau
Supplemental Data. Chen and Thelen (2010). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Data. Chen and Thelen (2010). Plant Cell 10.1105/tpc.109.071837 1 C Total 5 kg 20 kg 100 kg Transmission Image 100 kg soluble pdtpi-gfp Plastid (PDH-alpha) Mito (PDH-alpha) GFP Image vector
Supplemental Data. Chen and Thelen (2010). Plant Cell 10.1105/tpc.109.071837 1 C Total 5 kg 20 kg 100 kg Transmission Image 100 kg soluble pdtpi-gfp Plastid (PDH-alpha) Mito (PDH-alpha) GFP Image vector
Genetically Engineering Yeast to Understand Molecular Modes of Speciation
 Genetically Engineering Yeast to Understand Molecular Modes of Speciation Mark Umbarger Biophysics 242 May 6, 2004 Abstract: An understanding of the molecular mechanisms of speciation (reproductive isolation)
Genetically Engineering Yeast to Understand Molecular Modes of Speciation Mark Umbarger Biophysics 242 May 6, 2004 Abstract: An understanding of the molecular mechanisms of speciation (reproductive isolation)
Biology 105/Summer Bacterial Genetics 8/12/ Bacterial Genomes p Gene Transfer Mechanisms in Bacteria p.
 READING: 14.2 Bacterial Genomes p. 481 14.3 Gene Transfer Mechanisms in Bacteria p. 486 Suggested Problems: 1, 7, 13, 14, 15, 20, 22 BACTERIAL GENETICS AND GENOMICS We still consider the E. coli genome
READING: 14.2 Bacterial Genomes p. 481 14.3 Gene Transfer Mechanisms in Bacteria p. 486 Suggested Problems: 1, 7, 13, 14, 15, 20, 22 BACTERIAL GENETICS AND GENOMICS We still consider the E. coli genome
AP Bio Module 16: Bacterial Genetics and Operons, Student Learning Guide
 Name: Period: Date: AP Bio Module 6: Bacterial Genetics and Operons, Student Learning Guide Getting started. Work in pairs (share a computer). Make sure that you log in for the first quiz so that you get
Name: Period: Date: AP Bio Module 6: Bacterial Genetics and Operons, Student Learning Guide Getting started. Work in pairs (share a computer). Make sure that you log in for the first quiz so that you get
Vital Statistics Derived from Complete Genome Sequencing (for E. coli MG1655)
") We still consider the E. coli genome as a fairly typical bacterial genome, and given the extensive information available about this organism and it's lifestyle, the E. coli genome is a useful point of
We still consider the E. coli genome as a fairly typical bacterial genome, and given the extensive information available about this organism and it's lifestyle, the E. coli genome is a useful point of
Quantification of Protein Half-Lives in the Budding Yeast Proteome
 Supporting Methods Quantification of Protein Half-Lives in the Budding Yeast Proteome 1 Cell Growth and Cycloheximide Treatment Three parallel cultures (17 ml) of each TAP-tagged strain were grown in separate
Supporting Methods Quantification of Protein Half-Lives in the Budding Yeast Proteome 1 Cell Growth and Cycloheximide Treatment Three parallel cultures (17 ml) of each TAP-tagged strain were grown in separate
Introduction to Microbiology BIOL 220 Summer Session I, 1996 Exam # 1
 Name I. Multiple Choice (1 point each) Introduction to Microbiology BIOL 220 Summer Session I, 1996 Exam # 1 B 1. Which is possessed by eukaryotes but not by prokaryotes? A. Cell wall B. Distinct nucleus
Name I. Multiple Choice (1 point each) Introduction to Microbiology BIOL 220 Summer Session I, 1996 Exam # 1 B 1. Which is possessed by eukaryotes but not by prokaryotes? A. Cell wall B. Distinct nucleus
Genetic Organization and Distribution of Tetracycline Resistance Determinants in Clostridium perfringens
 ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Nov. 1996, p. 2500 2504 Vol. 40, No. 11 0066-4804/96/$04.00 0 Copyright 1996, American Society for Microbiology Genetic Organization and Distribution of Tetracycline
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Nov. 1996, p. 2500 2504 Vol. 40, No. 11 0066-4804/96/$04.00 0 Copyright 1996, American Society for Microbiology Genetic Organization and Distribution of Tetracycline
Sporulation Phenotype of a Bacillus subtilis Mutant Expressing an Unprocessable but Active E Transcription Factor
 JOURNAL OF BACTERIOLOGY, Apr. 2004, p. 1999 2005 Vol. 186, No. 7 0021-9193/04/$08.00 0 DOI: 10.1128/JB.186.7.1999 2005.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Sporulation
JOURNAL OF BACTERIOLOGY, Apr. 2004, p. 1999 2005 Vol. 186, No. 7 0021-9193/04/$08.00 0 DOI: 10.1128/JB.186.7.1999 2005.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Sporulation
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11419 Supplementary Figure 1 Schematic representation of innate immune signaling pathways induced by intracellular Salmonella in cultured macrophages. a, During the infection Salmonella
doi:10.1038/nature11419 Supplementary Figure 1 Schematic representation of innate immune signaling pathways induced by intracellular Salmonella in cultured macrophages. a, During the infection Salmonella
Analysis of Escherichia coli amino acid transporters
 Ph.D thesis Analysis of Escherichia coli amino acid transporters Presented by Attila Szvetnik Supervisor: Dr. Miklós Kálmán Biology Ph.D School University of Szeged Bay Zoltán Foundation for Applied Research
Ph.D thesis Analysis of Escherichia coli amino acid transporters Presented by Attila Szvetnik Supervisor: Dr. Miklós Kálmán Biology Ph.D School University of Szeged Bay Zoltán Foundation for Applied Research
CURRICULUM VITAE, ABBREVIATED. Professor, Department of Food Science, University of Massachusetts, Amherst
 CURRICULUM VITAE, ABBREVIATED Professor, Department of Food Science, University of Massachusetts, Amherst Education: Department of Bacteriology and Food Research Institute, University of Wisconsin, Madison;
CURRICULUM VITAE, ABBREVIATED Professor, Department of Food Science, University of Massachusetts, Amherst Education: Department of Bacteriology and Food Research Institute, University of Wisconsin, Madison;
Microbial Pathogenesis
 Microbial Pathogenesis 85 (2015) 1e10 Contents lists available at ScienceDirect Microbial Pathogenesis journal homepage: www.elsevier.com/locate/micpath In vitro cytotoxicity induced by Clostridium perfringens
Microbial Pathogenesis 85 (2015) 1e10 Contents lists available at ScienceDirect Microbial Pathogenesis journal homepage: www.elsevier.com/locate/micpath In vitro cytotoxicity induced by Clostridium perfringens
U.S. Patent No. 9,051,563 and other pending patents. Ver
 INSTRUCTION MANUAL Direct-zol RNA MiniPrep Catalog Nos. R050, R05, R05, & R053 Highlights Quick, spin column purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and other
INSTRUCTION MANUAL Direct-zol RNA MiniPrep Catalog Nos. R050, R05, R05, & R053 Highlights Quick, spin column purification of high-quality (DNA-free) total RNA directly from TRIzol, TRI Reagent and other
What Organelle Makes Proteins According To The Instructions Given By Dna
 What Organelle Makes Proteins According To The Instructions Given By Dna This is because it contains the information needed to make proteins. assemble enzymes and other proteins according to the directions
What Organelle Makes Proteins According To The Instructions Given By Dna This is because it contains the information needed to make proteins. assemble enzymes and other proteins according to the directions
Cloning, Nucleotide Sequencing, and Expression of the Clostridium perfringens Enterotoxin Gene in Eschenichia coli
 INFECTION AND IMMUNIrY, Aug. 1993, p. 3429-3439 0019-9567/93/083429-11$02.00/0 Copyright 1993, American Society for Microbiology Vol. 61, No. 8 Cloning, Nucleotide Sequencing, and Expression of the Clostridium
INFECTION AND IMMUNIrY, Aug. 1993, p. 3429-3439 0019-9567/93/083429-11$02.00/0 Copyright 1993, American Society for Microbiology Vol. 61, No. 8 Cloning, Nucleotide Sequencing, and Expression of the Clostridium
CRISPR-SeroSeq: A Developing Technique for Salmonella Subtyping
 Department of Biological Sciences Seminar Blog Seminar Date: 3/23/18 Speaker: Dr. Nikki Shariat, Gettysburg College Title: Probing Salmonella population diversity using CRISPRs CRISPR-SeroSeq: A Developing
Department of Biological Sciences Seminar Blog Seminar Date: 3/23/18 Speaker: Dr. Nikki Shariat, Gettysburg College Title: Probing Salmonella population diversity using CRISPRs CRISPR-SeroSeq: A Developing
surface of each plate and spread evenly with a sterile glass rod. Inoculated media were incubated The stock cultures of the C. perfringens strains
 STUDIES OF THE L-FORMS OF CLOSTRIDIUM PERFRINGENS I. RELATIONSHIP OF COLONY MORPHOLOGY AND REVERSIBILITY TOSHIO KAWATOMARI Department of Bacteriology, 406th Medical General Laboratory, APO 343, San Francisco,
STUDIES OF THE L-FORMS OF CLOSTRIDIUM PERFRINGENS I. RELATIONSHIP OF COLONY MORPHOLOGY AND REVERSIBILITY TOSHIO KAWATOMARI Department of Bacteriology, 406th Medical General Laboratory, APO 343, San Francisco,
In vitro cytotoxicity induced by Clostridium perfringens isolate carrying a chromosomal
 In vitro cytotoxicity induced by Clostridium perfringens isolate carrying a chromosomal cpe gene is exclusively dependent on sporulation and enterotoxin production Mayo Yasugi 1*, Yuki Sugahara 1, Hidenobu
In vitro cytotoxicity induced by Clostridium perfringens isolate carrying a chromosomal cpe gene is exclusively dependent on sporulation and enterotoxin production Mayo Yasugi 1*, Yuki Sugahara 1, Hidenobu
Controlling Gene Expression
 Controlling Gene Expression Control Mechanisms Gene regulation involves turning on or off specific genes as required by the cell Determine when to make more proteins and when to stop making more Housekeeping
Controlling Gene Expression Control Mechanisms Gene regulation involves turning on or off specific genes as required by the cell Determine when to make more proteins and when to stop making more Housekeeping
Bio 119 Bacterial Genomics 6/26/10
 BACTERIAL GENOMICS Reading in BOM-12: Sec. 11.1 Genetic Map of the E. coli Chromosome p. 279 Sec. 13.2 Prokaryotic Genomes: Sizes and ORF Contents p. 344 Sec. 13.3 Prokaryotic Genomes: Bioinformatic Analysis
BACTERIAL GENOMICS Reading in BOM-12: Sec. 11.1 Genetic Map of the E. coli Chromosome p. 279 Sec. 13.2 Prokaryotic Genomes: Sizes and ORF Contents p. 344 Sec. 13.3 Prokaryotic Genomes: Bioinformatic Analysis
RNA-Solv Reagent. R preps R preps R preps
 RNA-Solv Reagent R6830-00 5 preps R6830-01 100 preps R6830-02 200 preps March 2013 RNA-Solv Reagent Table of Contents Introduction...2 Kit Contents/Storage and Stability...3 Before Beginning...4 Analysis
RNA-Solv Reagent R6830-00 5 preps R6830-01 100 preps R6830-02 200 preps March 2013 RNA-Solv Reagent Table of Contents Introduction...2 Kit Contents/Storage and Stability...3 Before Beginning...4 Analysis
RNA-Solv Reagent. R preps R preps R preps
 RNA-Solv Reagent R6830-00 5 preps R6830-01 100 preps R6830-02 200 preps March 2013 RNA-Solv Reagent Table of Contents Introduction...2 Kit Contents/Storage and Stability...3 Before Beginning...4 Analysis
RNA-Solv Reagent R6830-00 5 preps R6830-01 100 preps R6830-02 200 preps March 2013 RNA-Solv Reagent Table of Contents Introduction...2 Kit Contents/Storage and Stability...3 Before Beginning...4 Analysis
ANALYSIS OF MICROBIAL COMPETITION
 ANALYSIS OF MICROBIAL COMPETITION Eric Pomper Microbiology 9 Pittsburgh Central Catholic High School Grade 9 Introduction Escherichia coli (E. coli) and Saccharomyces cerevisiae (Yeast) were grown together
ANALYSIS OF MICROBIAL COMPETITION Eric Pomper Microbiology 9 Pittsburgh Central Catholic High School Grade 9 Introduction Escherichia coli (E. coli) and Saccharomyces cerevisiae (Yeast) were grown together
RNA Transport. R preps R preps
 RNA Transport R0527-00 5 preps R0527-01 50 preps July 2014 RNA Transport Table of Contents Introduction...2 Kit Contents/Storage and Stability...3 Protocol...4 Storage Procedure...4 Recovery Procedure...5
RNA Transport R0527-00 5 preps R0527-01 50 preps July 2014 RNA Transport Table of Contents Introduction...2 Kit Contents/Storage and Stability...3 Protocol...4 Storage Procedure...4 Recovery Procedure...5
Experiences with the Coulter Counter in Bacteriology1
 Experiences with the Coulter Counter in Bacteriology1 ELLEN M. SWANTON, WILLIAM A. CTJRBY, AND HOWARD E. LIND Sias Laboratories, Brooks Hospital, Brookline, Massachusetts Received for publication May 24,
Experiences with the Coulter Counter in Bacteriology1 ELLEN M. SWANTON, WILLIAM A. CTJRBY, AND HOWARD E. LIND Sias Laboratories, Brooks Hospital, Brookline, Massachusetts Received for publication May 24,
A sensitive whole-cell biosensor for the simultaneous detection of a broad-spectrum of toxic heavy metal ions
 Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2015 SUPPORTING INFORMATION A sensitive whole-cell biosensor for the simultaneous detection of a broad-spectrum
Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2015 SUPPORTING INFORMATION A sensitive whole-cell biosensor for the simultaneous detection of a broad-spectrum
Bacillus subtilis Food Grade Expression System
 MoBiTec GmbH 2017 Page 1 Bacillus subtilis Food Grade Expression System MoBiTec GmbH 2017 Page 2 Contents Introduction... 3 The Food Grade Selection System... 3 Storage and Handling Instructions... 5 Protocols...
MoBiTec GmbH 2017 Page 1 Bacillus subtilis Food Grade Expression System MoBiTec GmbH 2017 Page 2 Contents Introduction... 3 The Food Grade Selection System... 3 Storage and Handling Instructions... 5 Protocols...
Increased Numbers of Heat-resistant Spores Produced by Two Strains of Clostridium perfringens Bearing Temperate Phage s9
 Journal of General Microbiology (I977), 103, 45-50. Printed in Great Britain 45 Increased Numbers of Heat-resistant Spores Produced by Two Strains of Clostridium perfringens Bearing Temperate Phage s9
Journal of General Microbiology (I977), 103, 45-50. Printed in Great Britain 45 Increased Numbers of Heat-resistant Spores Produced by Two Strains of Clostridium perfringens Bearing Temperate Phage s9
Supramolecular stabilization of the acid tolerant L-arabinose isomerase from the food-grade Lactobacillus sakei
 Supporting Information Supramolecular stabilization of the acid tolerant L-arabinose isomerase from the food-grade Lactobacillus sakei Said Jebors a, Yannick Tauran a, Nushin Aghajari b, Samira Boudebbouze
Supporting Information Supramolecular stabilization of the acid tolerant L-arabinose isomerase from the food-grade Lactobacillus sakei Said Jebors a, Yannick Tauran a, Nushin Aghajari b, Samira Boudebbouze
Genetic biomarkers for high heat resistance of Bacillus spores: relevance for optimal design of heat treatment
 Genetic biomarkers for high heat resistance of Bacillus spores: relevance for optimal design of heat treatment IAFP s 12th European Symposium on Food Safety marjon.wells-bennik@nizo.com 11-13 May 2016
Genetic biomarkers for high heat resistance of Bacillus spores: relevance for optimal design of heat treatment IAFP s 12th European Symposium on Food Safety marjon.wells-bennik@nizo.com 11-13 May 2016
Germination of Individual Bacillus subtilis Spores with Alterations in the GerD and SpoVA Proteins, Which Are Important in Spore Germination
 JOURNAL OF BACTERIOLOGY, May 2011, p. 2301 2311 Vol. 193, No. 9 0021-9193/11/$12.00 doi:10.1128/jb.00122-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. Germination of Individual
JOURNAL OF BACTERIOLOGY, May 2011, p. 2301 2311 Vol. 193, No. 9 0021-9193/11/$12.00 doi:10.1128/jb.00122-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. Germination of Individual
Role of GerD in Germination of Bacillus subtilis Spores
 JOURNAL OF BACTERIOLOGY, Feb. 2007, p. 1090 1098 Vol. 189, No. 3 0021-9193/07/$08.00 0 doi:10.1128/jb.01606-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Role of GerD in Germination
JOURNAL OF BACTERIOLOGY, Feb. 2007, p. 1090 1098 Vol. 189, No. 3 0021-9193/07/$08.00 0 doi:10.1128/jb.01606-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Role of GerD in Germination
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Supplemental material
 Supplemental material Table 1- Segregation analysis of sgt1a sgt1b double mutant plants Parental genotype No. of lines Normal seeds Phenotype Abnormal seeds Ratio of abnormal seeds (%) WT 3 171 2 1.16
Supplemental material Table 1- Segregation analysis of sgt1a sgt1b double mutant plants Parental genotype No. of lines Normal seeds Phenotype Abnormal seeds Ratio of abnormal seeds (%) WT 3 171 2 1.16
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Chapter 15 Active Reading Guide Regulation of Gene Expression
 Name: AP Biology Mr. Croft Chapter 15 Active Reading Guide Regulation of Gene Expression The overview for Chapter 15 introduces the idea that while all cells of an organism have all genes in the genome,
Name: AP Biology Mr. Croft Chapter 15 Active Reading Guide Regulation of Gene Expression The overview for Chapter 15 introduces the idea that while all cells of an organism have all genes in the genome,
Chapter 5. Partial purification of granule bound Pi-fA synthase
 Chapter 5 Partial purification of granule bound Pi-fA synthase 5.1 INTRODUCTION The enzyme PHA synthase occurs inside the bacterial cells both, as soluble and granule bound form (Haywood et al., 1989).
Chapter 5 Partial purification of granule bound Pi-fA synthase 5.1 INTRODUCTION The enzyme PHA synthase occurs inside the bacterial cells both, as soluble and granule bound form (Haywood et al., 1989).
Supporting Information
 Supporting Information López et al. 10.1073/pnas.0810940106 1. Ivey DM, et al. (1993) Cloning and characterization of a putative Ca2 /H antiporter gene from Escherichia coli upon functional complementation
Supporting Information López et al. 10.1073/pnas.0810940106 1. Ivey DM, et al. (1993) Cloning and characterization of a putative Ca2 /H antiporter gene from Escherichia coli upon functional complementation
camp Direct Immunoassay Kit
 camp Direct Immunoassay Kit Catalog Number KA0886 100 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
camp Direct Immunoassay Kit Catalog Number KA0886 100 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
PETER J. LEWIS, LING JUAN WU, AND JEFFERY ERRINGTON* Sir William Dunn School of Pathology, University of Oxford, Oxford OX1 3RE, United Kingdom
 JOURNAL OF BACTERIOLOGY, July 1998, p. 3276 3284 Vol. 180, No. 13 0021-9193/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Establishment of Prespore-Specific Gene Expression
JOURNAL OF BACTERIOLOGY, July 1998, p. 3276 3284 Vol. 180, No. 13 0021-9193/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Establishment of Prespore-Specific Gene Expression
NukEx Nucleic Acid Release Reagent
 Instruction for Use NukEx Nucleic Acid Release Reagent For general laboratory use. For in vitro use only. Reagent for the enzymatic release of nucleic acid from tissue samples, ticks, insects and swabs.
Instruction for Use NukEx Nucleic Acid Release Reagent For general laboratory use. For in vitro use only. Reagent for the enzymatic release of nucleic acid from tissue samples, ticks, insects and swabs.
Initiation of DNA Replication Lecture 3! Linda Bloom! Office: ARB R3-165! phone: !
 Initiation of DNA Replication Lecture 3! Linda Bloom! Office: ARB R3-165! email: lbloom@ufl.edu! phone: 392-8708! 1! Lecture 3 Outline! Students will learn! Basic techniques for visualizing replicating
Initiation of DNA Replication Lecture 3! Linda Bloom! Office: ARB R3-165! email: lbloom@ufl.edu! phone: 392-8708! 1! Lecture 3 Outline! Students will learn! Basic techniques for visualizing replicating
Bacterial Genetics & Operons
 Bacterial Genetics & Operons The Bacterial Genome Because bacteria have simple genomes, they are used most often in molecular genetics studies Most of what we know about bacterial genetics comes from the
Bacterial Genetics & Operons The Bacterial Genome Because bacteria have simple genomes, they are used most often in molecular genetics studies Most of what we know about bacterial genetics comes from the
Topic 4 - #14 The Lactose Operon
 Topic 4 - #14 The Lactose Operon The Lactose Operon The lactose operon is an operon which is responsible for the transport and metabolism of the sugar lactose in E. coli. - Lactose is one of many organic
Topic 4 - #14 The Lactose Operon The Lactose Operon The lactose operon is an operon which is responsible for the transport and metabolism of the sugar lactose in E. coli. - Lactose is one of many organic
Three different fusions led to three basic ideas: 1) If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes
 If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes") Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
PREVALENCE OF CLOSTRIDIUM PERFRINGENS TYPE A ISOLATES IN DIFFERENT TISSUES OF BROILER CHICKENS
 Bulgarian Journal of Veterinary Medicine, 2017, 20, No 1, 80 86 ISSN 1311-1477; DOI: 10.15547/bjvm.919 Short communication PREVALENCE OF CLOSTRIDIUM PERFRINGENS TYPE A ISOLATES IN DIFFERENT TISSUES OF
Bulgarian Journal of Veterinary Medicine, 2017, 20, No 1, 80 86 ISSN 1311-1477; DOI: 10.15547/bjvm.919 Short communication PREVALENCE OF CLOSTRIDIUM PERFRINGENS TYPE A ISOLATES IN DIFFERENT TISSUES OF
Nitric Oxide Synthase Ultrasensitive Colorimetric Assay
 Package Insert Nitric Oxide Synthase Ultrasensitive Colorimetric Assay 96 Wells For Research Use Only v. 2.0 09.20.17 Eagle Biosciences, Inc. 20A NW Blvd., Suite 112, Nashua, NH 03063 Phone: 866-419-2019
Package Insert Nitric Oxide Synthase Ultrasensitive Colorimetric Assay 96 Wells For Research Use Only v. 2.0 09.20.17 Eagle Biosciences, Inc. 20A NW Blvd., Suite 112, Nashua, NH 03063 Phone: 866-419-2019
Enterotoxigenecity and Typing of Clostridium perfringens Isolated from Raw Meat, Meat Products and Water Samples in Kashmir, India
 International Journal of Current Microbiology and Applied Sciences ISSN: 39-7706 Volume 7 Number 08 (08) Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/0.0546/ijcmas.08.708.053
International Journal of Current Microbiology and Applied Sciences ISSN: 39-7706 Volume 7 Number 08 (08) Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/0.0546/ijcmas.08.708.053
Risk Assessment of Staphylococcus aureus and Clostridium perfringens in ready to eat Egg Products
 Risk Assessment of Staphylococcus aureus and Clostridium perfringens in ready to eat Egg Products Introduction Egg products refer to products made by adding other types of food or food additives to eggs
Risk Assessment of Staphylococcus aureus and Clostridium perfringens in ready to eat Egg Products Introduction Egg products refer to products made by adding other types of food or food additives to eggs
Introduction to Molecular and Cell Biology
 Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the molecular basis of disease? What
Introduction to Molecular and Cell Biology Molecular biology seeks to understand the physical and chemical basis of life. and helps us answer the following? What is the molecular basis of disease? What