This is a provisional PDF comprising this cover note and the manuscript as it was upon acceptance for publication.
|
|
|
- Edgar Luke McCarthy
- 6 years ago
- Views:
Transcription
1 This is a provisional PDF comprising this cover note and the manuscript as it was upon acceptance for publication. A typeset PDF article will be published soon. RNAi-mediated knockdown showing impaired cell survival in Drosophila wing imaginal disc Article type Author(s) Journal name and URL Original Research Makoto Umemori, Okiko Habara, Tatsunori Iwata, Kousuke Maeda, Kana Nishinoue, Atsushi Okabe, Masahiko Takemura, Kuniaki Takahashi, Kaoru Saigo, Ryu Ueda and Takashi Adachi-Yamada Gene Regulation and Systems Biology Accessible at This article is published under the terms of the Creative Commons Attribution License. Copyright in the article, its metadata, and any supplementary data is held by the author or authors. We welcome new submissions to this journal at any time. For further information on this journal and how to submit your research to it please visit the journal home page or send an to the Managing Editor
2 Revised 1 RNAi-mediated knockdown showing impaired cell survival in Drosophila wing imaginal disc Makoto Umemori a, *, Okiko Habara b, *, Tatsunori Iwata c, Kousuke Maeda a, Kana Nishinoue d, Atsushi Okabe c, Masahiko Takemura a, Kuniaki Takahashi e, Kaoru Saigo f, Ryu Ueda e, and Takashi Adachi-Yamada a, b, c, d, ** a b c Department of Biology, Graduate School of Science, Kobe University, Kobe, Japan. Japan Science and Technology Agency, Japan. Department of Sciences for Natural Environment, Graduate School of Cultural Studies and Human Science, Kobe University, Kobe, Japan. d Department of Sciences for Natural Environment, Faculty of Human Development, Kobe University, Kobe, Japan. e f Genetic Strains Research Center, National Institute of Genetics, Mishima, Japan. Department of Biophysics and Biochemistry, Graduate School of Science, University of Tokyo, Tokyo, Japan. * These authors contributed equally to this work. ** Corresponding author. Department of Biology, Graduate School of Science, Kobe University, Kobe , Japan. Tel: Fax: address: yamadach@kobe-u.ac.jp (Takashi Adachi-Yamada)
3 Revised 2 Abstract The genetically amenable organism Drosophila melanogaster has been estimated to have 14,076 protein coding genes in the genome, according to the flybase release note R5.13 ( Recent application of RNA interference (RNAi) to the study of developmental biology in Drosophila has enabled us to carry out a systematic investigation of genes affecting various specific phenotypes. In order to search for genes supporting cell survival, we conducted an immunohistochemical examination in which the RNAi of 2497 genes was independently induced within the dorsal compartment of the wing imaginal disc. Under these conditions, the activities of a stress-activated protein kinase JNK (c-jun N-terminal kinase) and apoptosis-executing factor Caspase-3 were monitored. Approximately half of the genes displayed a strong JNK or Caspase-3 activation when their RNAi was induced. Most of the JNK activation accompanied Caspase-3 activation, while the opposite did not hold true. Interestingly, the area activating Caspase-3 was more broadly seen than that activating JNK, suggesting that JNK is crucial for induction of non-autonomous apoptosis in many cases. Furthermore, the RNAi of essential factors commonly regulating transcription and translation showed a severe and cell-autonomous apoptosis but also elicited another apoptosis at an adjacent area in a non-autonomous way. We also found that the frequency of apoptosis varies depending on the tissues.
4 Revised 3 Introduction In recent years, mechanisms controlling apoptosis have been extensively studied, and various factors are known to be involved in the intrinsic and extrinsic apoptotic pathways (Domingos and Steller 2007; Hay et al 2004; Putcha and Johnson 2004; Steller 2008). Although these pathways play a pivotal role in the execution of most cases of apoptosis, the apoptosis shown in developing animal tissues is also affected by various growth and differentiation signals to promote or repair organ development (Jacobson et al 1997). In general, inhibition of apoptosis accompanies growth induction, whereas reduction of growth conversely leads to apoptosis. However, we can often find exceptions showing an opposite relationship, such as overgrowth-induced apoptosis (Gastman et al 2000; Poggi et al 2005; Zilian et al 1999) and apoptosis-induced overgrowth (Huh et al 2004; Perez-Garijo et al 2004; Ryoo et al 2004), indicating that we do not fully understand these cell survival controls between apoptosis and growth. In order to systematically investigate the apoptosis phenotype caused by reducing each gene function in the developing animal tissues, we employed a genetically amenable fruit fly Drosophila melanogaster, in which each gene can be knocked down by RNAi (Fire et al 1998; Kennerdell and Carthew 1998; Kennerdell and Carthew 2000), to observe the effect on apoptosis induction. RNAi provides an easy and powerful technique for reducing the quantity of mrna derived from endogenous specific genes, and it has recently been applied in many studies to investigate various gene functions (Dietzl et al 2007). In this study, we screened 2497 protein-coding genes of Drosophila to determine whether they were required for prevention of apoptosis in the wing imaginal disc and found that 47 % of them showed an apoptosis induction when their functions were knocked down by RNAi in the developing wing disc. Most of the cases (82 %) with detectable Caspase activation were associated with JNK activation, which was unexpectedly high because JNK has not been observed as essential for all apoptosis. Alternatively, JNK is known to be involved in inducing non-autonomous apoptosis
5 Revised 4 (Adachi-Yamada and O'Connor 2002; Adachi-Yamada and O'Connor 2004), which occurs in cells distant from those cells associated with the primary cause of apoptosis. Interestingly, a major part of the JNK and Caspase-3 activation found in this study occurred in a non-autonomous manner, suggesting that the non-autonomous pathway is a common way to induce apoptosis. Loss of membrane proteins frequently caused JNK activation, which had also been expected because cell-cell communication is presumed to be important for many developmental processes, including apoptosis in multicellular organisms. These results, as well as the database showing the immunofluorescent data, provide an archival source for survey of genes and for fine analysis of each gene in apoptosis regulation using the Drosophila imaginal discs.
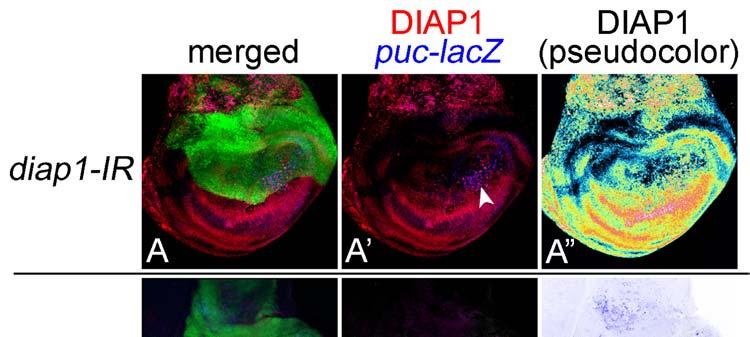
6 Revised 5 Results Rationale for RNAi-mediated screening for genes regulating apoptosis We induced RNAi in the dorsal compartment of the wing disc and monitored the activities of Caspase-3 and JNK. Caspase-3 plays a central role in most apoptosis, while JNK leads to a subgroup of stress-induced apoptosis (Kanda and Miura 2004). In the Drosophila wing disc, JNK activation is usually linked to the activation of Caspase-3 (Adachi-Yamada and O'Connor 2002). Puc is a protein phosphatase specifically inactivating JNK, and its transcription occurs in response to the JNK signal, thereby making a negative-regulatory circuit (Martin-Blanco et al 1998). Thus, the expression of puc reflects the JNK activity and can be used for monitoring it. Before expanding the RNAi analyses to the entire genome, we checked whether the mutant phenotypes caused by previously known apoptosis-regulating genes, such as diap1 (Hay et al 1995) and dark (Kanuka et al 1999; Rodriguez et al 1999; Zhou et al 1999), were reproduced by their RNAi. When diap1 (Drosophila Inhibitor of Apoptosis Protein 1) was knocked down within the dorsal compartment of the imaginal disc, a local but prominent activation of Caspase-3 was detected (Fig. 1B). The position-specificity may be dependent on the difference in sensitivity in the induction of apoptosis, as described later. In contrast, when dark (Drosophila Apaf-1-Related Killer) was knocked down, no apoptosis induction was observed (Fig. 1C). Furthermore, the use of this collection of RNAi strains has already been validated, since they were screened for apoptosis phenotype in the compound eye (Leulier et al 2006). For the non-autonomously induced apoptosis during restoration of morphogenesis, for example, we tested whether the apoptosis shown in several RNAi samples really reflected the non-autonomous apoptosis by conventional gene manipulation in previously studies (Adachi-Yamada and O'Connor 2002). We manipulated signaling factors for a diffusible extracellular ligand Dpp, a homolog of mammalian BMP (Bone Morphogenetic Protein)-2/4. Mad (Mothers
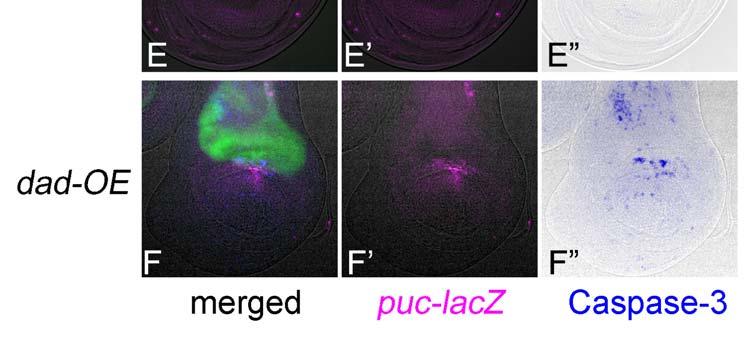
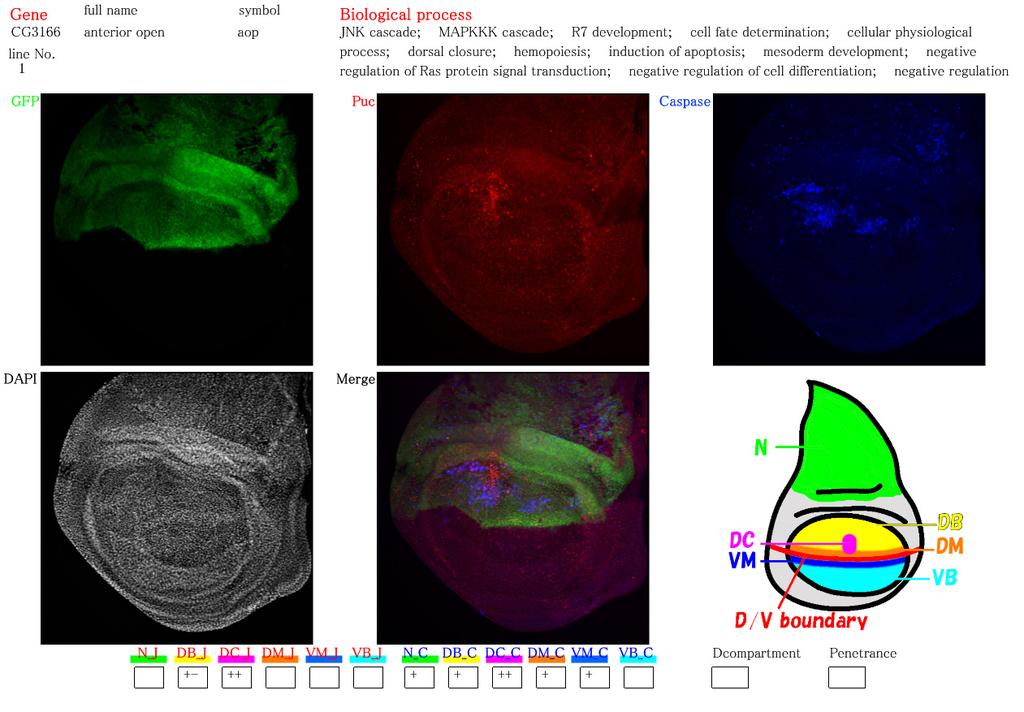
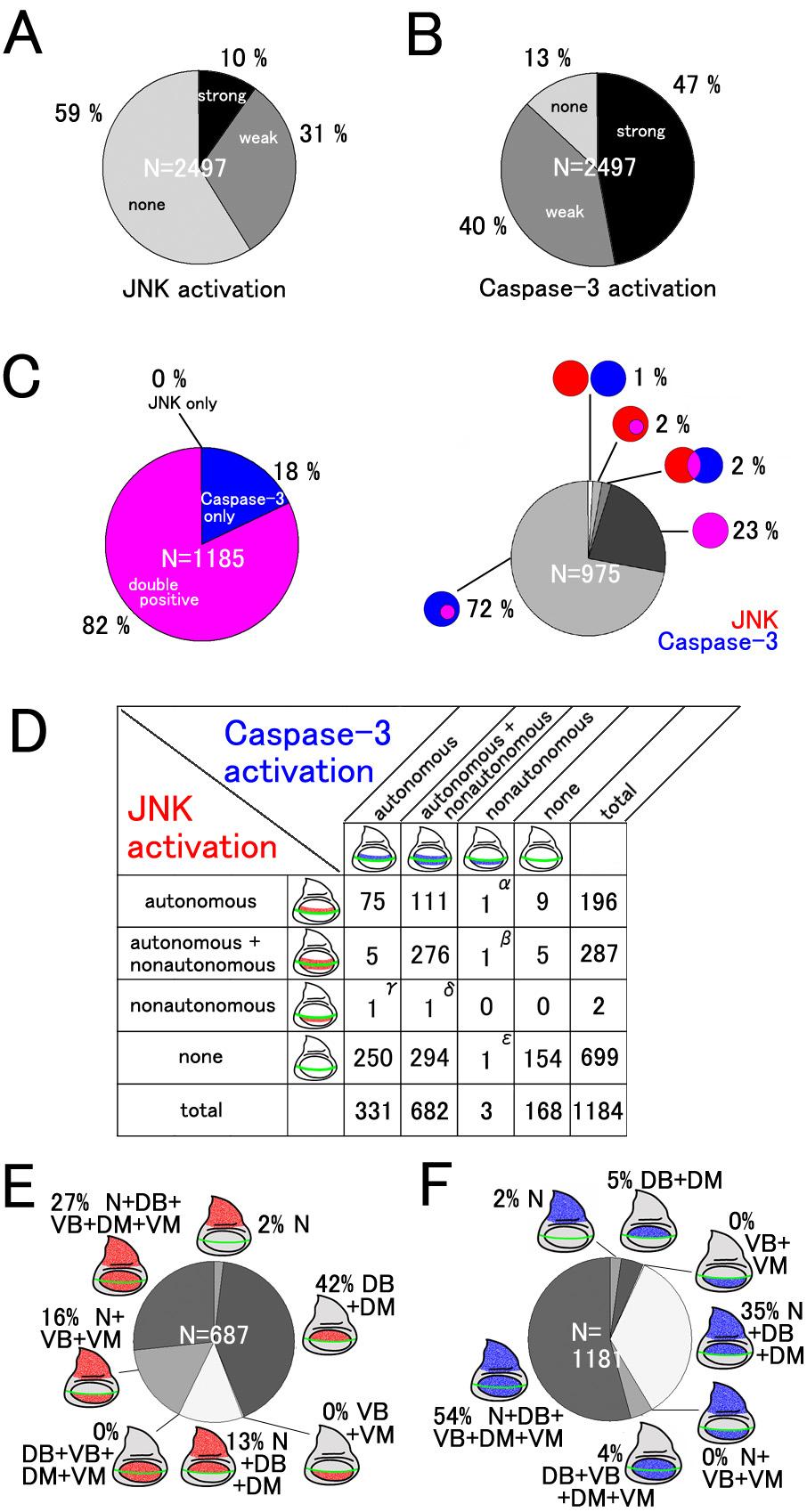
7 Revised 6 against Dpp, (Sekelsky et al 1995), a Drosophila homolog of mammalian r-smad, transmits the intracellular signal caused by Dpp. As shown in Fig. 1E, the RNAi of mad within the dorsal compartment activated JNK and Caspase-3 in both dorsal and ventral compartments of the central wing disc region, which is a typical example of non-cell-autonomous induction of apoptosis. These features are also quite similar to those seen in the case of overexpression of Dad (Daughters against Dpp), a homolog of anti-smad, (Fig. 1F) that can repress Dpp signaling (Tsuneizumi et al 1997). Therefore, RNAi can mimic the conventional gene manipulation in induction of apoptosis, at least in some representative examples. We describe how to choose the genes for RNAi in the section on Experimental Procedures. During observation of our RNAi results, we focused on six areas of the wing disc (Notum [N], Dorsal wing Blade [DB], DorsoCentral spot [DC], Dorsal wing Margin [DM], Ventral wing Margin [VM], and Ventral wing Blade [VB]), and classified the strength of JNK and Caspase-3 activities in each area into three grades (+-, +, and ++). The database was constructed by using FileMaker Pro 7 (FileMaker, Inc.) and contains each immunofluorescence image with the above classification of JNK/Caspase-3 activities in each gene page. First, we noticed that a narrow area in the dorsocentral (DC) wing region showed JNK and Caspase-3 activation too sensitively (e.g. Fig. 2), which was not always correlated with RNAi. Consequently, the results that simply reflected this feature were excluded from all of the analyses. When a survey of all of the RNAi experiments was carried out, both weak and strong activation of JNK and Caspase-3 (shown by + and ++ signs in the database) was observed in 41 % and 87 % of the cases, respectively (Fig. 3A). These proportions seemed much higher than expected because the imaginal disc cells may not actually express such a large number of genes, and they are thought to express several thousand genes (Butler et al 2003; Halder et al 1995; Klebes et al 2002). This large number of strains with weak JNK and/or Caspase-3 activation may reflect false-positive results due to an off-target effect (OTF) or other non-specific effects of dsrna
8 Revised 7 expression. A part of these IR lines was known to cause OTF and the frequency of such possible OTF lines was calculated to be 47% as a maximal estimation. Accordingly, we did not give further consideration to this class of strains and hereafter focused on the results showing high levels of JNK and Caspase-3 activation (shown only by the ++ sign in the database), which amounted to 10 and 47% of cases, respectively (Fig. 3A). The immunofluorescence data can be accessed on the website: (see Experimental Procedures). Screening summary Among these strong cases of JNK and/or Caspase-3 activation, more than 80% showed a coupling of JNK and Caspase-3 activation to various extents (Fig. 3C). Conversely, there were only two cases in which JNK activation did not accompany Caspase-3 activation (calculated as 0.17%). Therefore, these findings are consistent with the previous observation that the JNK activation precedes Caspase-3 activation and is strongly linked to apoptosis in the Drosophila wing (Adachi-Yamada and O'Connor 2004). Furthermore, we carefully assessed the non-cell autonomous effect of RNAi by examining the phenotypes in the vicinity of the DV boundary. Most of the RNAi-induced JNK/Caspase-3 activation showed a striking non-cell autonomy (Fig. 3D), which is similar to the previously known feature in non-cell-autonomous activation of JNK by altered Dpp signaling (Adachi-Yamada and O'Connor 2004). This suggests that the non-cell-autonomous induction of apoptosis is one of the common patterns in cell death induction. It has previously been shown that LRR (Leucine Rich Repeat) family cell adhesion proteins contribute to unique cell affinity. Furthermore, alteration of LRR protein functions causes JNK activation followed by Caspase-3 activation (Adachi-Yamada et al 2005; Milan et al 2002). Therefore, transmembrane and secreted proteins are good candidates for mediators of non-cell-autonomous apoptosis. We thus tested the apoptosis-inducing activity by RNAi of putative secreted proteins selected based on the presence of N-terminal signal sequences. Consistently, the RNAi of such
9 Revised 8 genes caused a higher frequency of strong JNK activation (74 out of 296 cases, 25%) when compared with the frequency of strong JNK activation in all of the RNAi cases (248 out of 2497 cases, 10%). Moreover, the RNAi of various transcription factors frequently showed non-autonomous apoptosis, suggesting that they are highly involved in regulation of morphogenesis and that their aberration likely induces non-autonomous apoptosis. These results suggest that a relatively large number of secreted proteins and transcription factors are involved in the prevention of non-cell-autonomous apoptosis. Alternatively, functional redundancy may have prevented identification of such molecules. We also noted that there was a significant tendency for various examples of RNAi-mediated apoptosis to be preferentially found in the wing blade region but not outside of this region. For example, the RNAi of taf6 (TBP-Associated Factor 6) leads to an autonomous activation of Caspase-3 at high levels in the wing blade region but at lower levels in the wing hinge region (Fig. 4). Similar traits have also been reported in the case of the apoptosis induced by a reduced-dpp signal (Adachi-Yamada and O'Connor 2004). Furthermore, when we surveyed all of the results, the activation of JNK/Caspase-3 in the wing blade was found to be much more frequent than those in the notum (Figs. 3E and F). Consequently, localization of apoptosis in the wing blade region may not be a feature specific to the alteration of a particular cell signal (such as Dpp) but instead a general feature found in all cases of apoptosis in the wing disc. This suggests that the number of stimuli to activate JNK or apoptosis varies depending on the tissue, or that the sensitivities to alterations of gene expression in induction of apoptosis are quite different between tissues. Non-autonomously induced apoptosis As we reported previously, aberrations of some morphogenetic signaling induce JNK activation followed by Caspase-3 activation at the boundary between cell populations with different levels of signaling intensities. This non-autonomous apoptosis is thought to be important for
10 Revised 9 restoration of abnormally developing tissues (Adachi-Yamada and O'Connor 2004). Various examples of apoptosis are probably induced in a similar non-autonomous way. To determine to what extent this applies to non-autonomous apoptosis, we surveyed the relationship in cell autonomy between JNK and Caspase-3 activation by focusing on the DV boundary at which the two cell populations come in contact (Fig. 3D). Around this position, there is an apparent tendency for JNK and Caspase-3 activation to occur simultaneously, which was observed in 471 cases, as shown in the 9 upper-left boxes in Fig. 3D s grid. As stated above, the most frequent pattern is that both JNK and Caspase-3 are both autonomously and non-autonomously activated. However, 111 out of 196 cases with autonomous JNK activation (57%) displayed a non-autonomous Caspase-3 activation (upper-most row in Fig. 3D grid). In contrast, except for one case, autonomous Caspase-3 activation (81 cases) did not show non-autonomous JNK activation (left-most column in Fig. 3D grid). Accordingly, these data strongly suggest that JNK activation is also crucial for priming non-autonomous apoptosis, whereas Caspase-3 is not. Furthermore, when observed throughout the wing disc, the activation patterns of JNK and Caspase-3 are different (Figs. 3E and F). JNK activation seems to be found unevenly in the dorsal region ( =57%), whereas the Caspase-3 activation only within the dorsal region is less (2+5+35=42%). On the other hand, JNK with non-autonomous activation is minor ( =43%), whereas non-autonomous Caspase-3 is major ( =58%). We were interested in the fact that some RNAi examples resulted in a non-autonomous apoptosis similar to that seen previously (Adachi-Yamada and O'Connor 2004). The RNAi of basal transcription factor Taf6 showed an autonomous activation of Caspase-3 in the blade region and a non-autonomous activation of JNK in the hinge region (Fig. 4A, B), the latter of which was unexpected because Taf6 is known to be necessary for the function of TBP (TATA-Binding Protein), suggesting its ubiquitous requirement for most of the transcription by RNA polymerase II. Accordingly, the RNAi of taf6 is expected to induce a severe autonomous apoptosis in all of the
11 Revised 10 tissues, as is the case for the RNAi of ribosomal protein genes (e.g. RpS14, Fig. 4C). Thus, the non-autonomy in JNK activation in the hinge region in taf6 RNAi suggests a morphogenetic function rather than its common transcriptional function and/or a difference in the sensitivity of decreased transcription leading to apoptosis between the tissues. The difference in responses between the blade and hinge regions was previously described in the apoptosis associated with homeotic transformation by overexpression of spineless (Adachi-Yamada et al 2005). Through these RNAi experiments, we discovered numerous cases of non-autonomous apoptosis cases. Among these, there are particular cases in which an autonomous apoptosis must be induced as a primary response (e.g. RpL17, shotgun) while an additional non-autonomous apoptosis may be further induced as a secondary response, which is probably caused by a juxtaposition of a normal area and a wide apoptotic area, as proposed previously (Milan et al 1996). Therefore, we tested a model case in which the proapoptotic gene reaper (rpr) is temporarily induced within the dorsal compartment by combination with a temperature sensitive-gal80 (McGuire et al 2003) (Fig. 5). As a result, at around 24 hours after rpr expression, non-autonomous apoptosis could be observed in the ventral compartment (Fig. 5A), although no similar non-autonomy was observed earlier or later (Figs. 5B and C). Thus, a wide area of autonomous apoptosis induction can cause a secondary non-autonomous apoptosis. This phenomenon seems to be a repair mechanism for fitting the adjacent tissue size (Milan 1996).
12 Revised 11 Discussion We created a database to show the immunofluorescent images for JNK and Caspase-3 activities in each RNAi experiment. In addition to our purpose of surveying genes affecting cell survival, the database may also be useful for searching for genes regulating tissue growth and patterning. For example, the RNAi of mad displays an apparent shrinkage of the compartment size without showing severe apoptosis except at the DV boundary (Fig. 1E). This phenotype strongly suggests the involvement of this gene in tissue growth and/or patterning. In contrast, the wing disc in which the dorsal compartment cells overexpress rpr showed a wide and severe apoptosis so that most of the dorsal cells disappeared (Fig. 5C). This case represents a typical phenotype, showing that the gene plays a role exclusively for apoptosis. These findings provide insight into the roles of genes for regulating various developmental processes. As is the case for the above-mentioned possibility of weak activation of JNK and/or Caspase-3, the OTF should also be considered for all phenotypes (Ma et al 2006). As an effective initial examination, the RNAi phenotype must be ameliorated by addition of the wild type transgene that is targeted by RNAi. Furthermore, two ways to distinguish the real RNAi effect from OTF have been proposed (Echeverri et al 2006). One is a test of a dsrna corresponding to the other part of the same mrna for displaying the same phenotype. The other is a test of an artificially altered transgene that is not targeted by the dsrna but that encodes the same amino acid sequence for complete rescue of the phenotype. Of course, classical analysis using the loss-of-function mutant may be another reliable way to judge the involvement of the gene in each phenotype. In either case, further analyses are required to demonstrate each phenotype as a real loss-of-function phenotype of the gene under focus. The database will be updated when we check the phenotype by the above examinations or accumulate the data from other RNAi constructs.
13 Revised 12 Experimental Procedures Materials Various fly strains harboring a transcribable inverted repeat sequence (IR) driven by UAS were prepared in the National Institute of Genetics (NIG) in Japan, as previously described (Leulier et al 2002). Briefly, a cdna fragment with nucleotide position of the coding sequence was obtained by PCR and was inserted as an IR in a head-to-head manner into a modified Bluescript vector, psc1. Then IR-fragments were excised by NotI and were subcloned into puast, a germline transformation vector containing UAS (upstream activation sequence) (Brand and Perrimon 1993). In the earlier stages of this research, we did not select the IR strains but randomly employed them according to the order in which NIG collected them. In the later stages, we preferentially focused on 302 genes that were predicted to encode secretory proteins (Sakurai et al 2007). Each UAS-IR fly strain was crossed with another strain carrying ap-gal4, UAS-GFP and puc-lacz. The offspring larvae possessing these four transgenes were reared at 25 C on a standard diet and then dissected at the late third instar larval stage for immunological staining. Immunological staining The dissected carcasses with the imaginal discs were fixed in 4% formaldehyde for 20 min at room temperature and washed with PBS (phosphate buffered saline) containing Triton X-100 (0.02 %). The puc-lacz expression was detected by indirect immunofluorescence using the murine anti-β-galactosidase antibody (Promega, #Z378B, 1/200 dilution). Active Caspase-3 was detected by the rabbit anti-cleaved Caspase-3 antibody (Cell Signaling Technology, #9661, 1/200 dilution), which is known to bind to the cleaved (activated) forms of mammalian Caspase-3 and its Drosophila homolog Drice. DIAP1 was detected by the rabbit anti-diap1 antibody (1/500 dilution) (Kuranaga et al 2006). Specific binding of these primary antibodies was visualized by fluorescent secondary
14 Revised 13 antibodies, such as the anti-mouse Ig-Cy3 (# , Jackson Immunoresearch) and anti-rabbit Ig-Cy5 (# , Jackson Immunoresearch) antibodies. Incubation with these primary or secondary antibodies was performed at 37C for 1 hour or at 4C overnight. Microscopic observation was performed without antifade reagents by the Leica deconvolution system Q550FW. Prediction of secreted proteins in the Drosophila genome Putative secreted proteins were searched based on the presence of hydrophobic residues in the N-terminal amino acids. When the average hydrophobicity index in the 25 amino acids between positions 6 and 30 exceeded 0.953, the protein was assumed to be secreted. The predicted protein data set from BDGP release 4.2 was searched using the program Ahiru (Sakurai et al 2007), which was written based on the algorithm described in (Bannai et al 2002) ( this yielded a list of 2184 candidate genes encoding secreted proteins. Among them, 296 genes available in the RNAi strains in NIG were used for the screen. How to access the immunofluorescence images To access the fluorescent images, go to the middle of the right column of the web page ( and click the line of "Browse All RNAi Stocks". In the newly appeared page, you can see all of the IR strains. When you click each Stock ID name which has the "wing disc" icon on the right column, you can see a set of immunofluorescence images at the bottom of the further next page.
15 Revised 14 Acknowledgements We are grateful to Yoshihiro H. Inoue, Guillermo Marques, Enrique Martin-Blanco, Alfonso Martinez-Arias, Tetsuya Tabata, Daisuke Yamamoto, and the Bloomington Stock Center (University of Indiana) for fly stock, and to Masayuki Miura and Erina Kuranaga for anti-diap1 antibody. We also thank Shigeo Hayashi and Kagayaki Kato for providing us the information on putative secretory proteins before its publication, and Shigeo Hayashi for his critical reading of the manuscript. This work was supported by grants from the Japan Science and Technology Agency and the Kato Memorial Bioscience Foundation.
16 Revised 15 References Adachi-Yamada, T., et al Wing-to-Leg homeosis by spineless causes apoptosis regulated by Fish-lips, a novel leucine-rich repeat transmembrane protein. Mol Cell Biol, 25: Adachi-Yamada, T., O'Connor, M. B Morphogenetic apoptosis: a mechanism for correcting discontinuities in morphogen gradients. Dev Biol, 251: Adachi-Yamada, T., O'Connor, M. B Mechanisms for removal of developmentally abnormal cells: cell competition and morphogenetic apoptosis. J Biochem (Tokyo), 136: Bannai, H., et al Extensive feature detection of N-terminal protein sorting signals. Bioinformatics, 18: Brand, A. H., Perrimon, N Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development, 118: Butler, M. J., et al Discovery of genes with highly restricted expression patterns in the Drosophila wing disc using DNA oligonucleotide microarrays. Development, 130: Dietzl, G., et al A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature, 448: Domingos, P. M., Steller, H Pathways regulating apoptosis during patterning and development. Curr Opin Genet Dev, 17: Echeverri, C. J., et al Minimizing the risk of reporting false positives in large-scale RNAi screens. Nat Methods, 3: Fire, A., et al Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature, 391: Gastman, B. R., et al Tumor-induced apoptosis of T cells: amplification by a mitochondrial cascade. Cancer Res, 60: Halder, G., Callaerts, P., Gehring, W. J Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science, 267: Hay, B. A., Huh, J. R., Guo, M The genetics of cell death: approaches, insights and opportunities in Drosophila. Nat Rev Genet, 5: Hay, B. A., Wassarman, D. A., Rubin, G. M Drosophila homologs of baculovirus inhibitor of apoptosis proteins function to block cell death. Cell, 83: Huh, J. R., Guo, M., Hay, B. A Compensatory proliferation induced by cell death in the Drosophila wing disc requires activity of the apical cell death caspase Dronc in a nonapoptotic role. Curr Biol, 14:
17 Revised 16 Jacobson, M. D., Weil, M., Raff, M. C Programmed cell death in animal development. Cell, 88: Kanda, H., Miura, M Regulatory roles of JNK in programmed cell death. J Biochem (Tokyo), 136: 1-6. Kanuka, H., et al Control of the cell death pathway by Dapaf-1, a Drosophila Apaf-1/CED-4-related caspase activator. Mol Cell, 4: Kennerdell, J. R., Carthew, R. W Use of dsrna-mediated genetic interference to demonstrate that frizzled and frizzled 2 act in the wingless pathway. Cell, 95: Kennerdell, J. R., Carthew, R. W Heritable gene silencing in Drosophila using double-stranded RNA. Nat Biotechnol, 18: Klebes, A., et al Expression profiling of Drosophila imaginal discs. Genome Biol, 3: RESEARCH0038. Kuranaga, E., et al Drosophila IKK-related kinase regulates nonapoptotic function of caspases via degradation of IAPs. Cell, 126: Leulier, F., et al Systematic in vivo RNAi analysis of putative components of the Drosophila cell death machinery. Cell Death Differ, 13: Leulier, F., et al Inducible expression of double-stranded RNA reveals a role for dfadd in the regulation of the antibacterial response in Drosophila adults. Curr Biol, 12: Ma, Y., et al Prevalence of off-target effects in Drosophila RNA interference screens. Nature, 443: Martin-Blanco, E., et al puckered encodes a phosphatase that mediates a feedback loop regulating JNK activity during dorsal closure in Drosophila. Genes Dev, 12: McGuire, S. E., et al Spatiotemporal rescue of memory dysfunction in Drosophila. Science, 302: Milan, M., Campuzano, S., Garcia-Bellido, A Cell cycling and patterned cell proliferation in the wing primordium of Drosophila. Proc Natl Acad Sci U S A, 93: Milan, M., Perez, L., Cohen, S. M Short-range cell interactions and cell survival in the Drosophila wing. Dev Cell, 2: Perez-Garijo, A., Martin, F. A., Morata, G Caspase inhibition during apoptosis causes abnormal signalling and developmental aberrations in Drosophila. Development, 131: Poggi, A., et al Tumor-induced apoptosis of human IL-2-activated NK cells: role of natural cytotoxicity receptors. J Immunol, 174: Putcha, G. V., Johnson, E. M., Jr Men are but worms: neuronal cell death in C elegans and
18 Revised 17 vertebrates. Cell Death Differ, 11: Rodriguez, A., et al Dark is a Drosophila homologue of Apaf-1/CED-4 and functions in an evolutionarily conserved death pathway. Nat Cell Biol, 1: Ryoo, H. D., Gorenc, T., Steller, H Apoptotic cells can induce compensatory cell proliferation through the JNK and the Wingless signaling pathways. Dev Cell, 7: Sakurai, K. T., et al Differential control of cell affinity required for progression and refinement of cell boundary during Drosophila leg segmentation. Dev Biol, 309: Sekelsky, J. J., et al Genetic characterization and cloning of mothers against dpp, a gene required for decapentaplegic function in Drosophila melanogaster. Genetics, 139: Steller, H Regulation of apoptosis in Drosophila. Cell Death Differ, 15: Tsuneizumi, K., et al Daughters against dpp modulates dpp organizing activity in Drosophila wing development. Nature, 389: Zhou, L., et al HAC-1, a Drosophila homolog of APAF-1 and CED-4 functions in developmental and radiation-induced apoptosis. Mol Cell, 4: Zilian, O., et al double-time is identical to discs overgrown, which is required for cell survival, proliferation and growth arrest in Drosophila imaginal discs. Development, 126:
19 Revised 18 LEGENDS TO FIGURES Figure 1. Similarity of RNAi and conventional gene manipulation in their apoptotic phenotypes. (A-A ) RNAi of diap1 by expressing its inverted repeat (IR) sequence in the dorsal compartment displayed a reduction of DIAP1 protein levels specifically in the dorsal compartment. Expression of a dorsal compartment marker apterous (green), DIAP1 (red), and JNK indicator puc-lacz (blue) are shown. A shows a pseudocolor image representing DIAP1 protein levels. Arrowhead in A indicates a cluster of high level staining of DIAP1 and puc-lacz, which is caused by the presence of massive apoptotic bodies. (B-F) Expression of apterous (green), puc-lacz (magenta), and activated Caspase-3 (blue) are shown. (B-B ) RNAi of diap1 in the dorsal compartment led to an activation of Caspase-3 at the specific position in the dorsal compartment. When the contrast of B image is elevated, puc-lacz expression can be observed at around the apoptotic bodies as in A (not shown). (C-C ) RNAi of dark in the dorsal compartment as a negative control experiment. No activation of JNK and faint activation of Caspase-3 were observed. (D-D ) Wild type. JNK and Caspase-3 are not activated in wild type. (E-E ) RNAi of mad in the dorsal compartment led to a non-autonomous activation of JNK and Caspase-3 in the medial region of the DV boundary. (F-F ) Overexpression (OE) of dad in the dorsal compartment, which results in a reduction of Dpp signaling, induces a shrinkage of the dorsal compartment as well as a non-autonomous activation of JNK and Caspase-3 in the medial region of the DV boundary. Figure 2. Web page showing the database for immunofluorescent images of the imaginal discs. The page for anterior open (aop) is shown here as an example. The gene s full name, CG number, gene symbol, strain line number, and biological process are described at the top according to the Flybase ( flybase.bio.indiana.edu/). The top three photographs are images for each color channel (green [apterous], red [JNK], and blue [Caspase-3]). The center-bottom photograph shows an image
20 Revised 19 merged with the three colors. In the left-bottom photographs, the DAPI staining is displayed to recognize the position and shape of the imaginal discs in the other photos. In some photographs, three kinds of imaginal discs (leg, haltere and wing) are shown in a single frame. In the right-bottom corner of the figure, abbreviations for the position in the wing imaginal discs are indicated using a cartoon. Small boxes at the bottom box row indicate the levels of JNK and Caspase-3 activation by symbols as follows: +-, +, ++. In this example, strong activation of JNK and Caspase-3 can be seen only around the DC spot area in the wing blade region. The abbreviations are explained in the fourth paragraph of the Results section. Figure 3. Categorization of RNAi-induced JNK and Caspase-3 activation. (A) Frequency of genes in which RNAi induces JNK activation. Cases with activation only at the dorsocentral spot (e.g. aop shown in Fig. 2) are excluded. (B) Frequency of genes in which RNAi induces Caspase-3 activation. Again, cases with activation only at the dorsocentral spot are excluded. (C) Relationship between JNK and Caspase-3. Cases showing strong activation of at least either JNK or Caspase-3 are analyzed. Left: Overlap of JNK and Caspase-3 activation. In most cases, both are simultaneously activated. Caspase-3 are sometimes activated solely (e.g. Fig. 1B), whereas JNK activation without Caspase-3 activation is rare. Right: Pattern of relative position of JNK-activating area (red) and Caspase-3-activating area (blue). Overlapping areas are shown in magenta. (D) Relation of cell autonomy in JNK activation and that in Caspase-3 activation around the DV boundary in the wing blade. Examples showing rare patterns in the table are RpL17 (α), shotgun (β), CG14122 (γ), CG14072 (δ), and E(spl)m5 (ε). (E) Frequencies of each JNK activation pattern with regard to wing disc subdomains. (F) Frequencies of each Caspase-3 activation pattern with regard to wing disc subdomains. Figure 4. Examples showing non-autonomous activation of JNK and Caspase-3. (A-A ) An example
21 Revised 20 of Taf6 RNAi. A non-autonomously induced JNK activation around the DV boundary in the hinge region is indicated by arrows. The approximate position of the boundary between the wing blade and hinge regions is indicated by the circle with the broken line. (B-B ) Another example of Taf6 RNAi, which shows a more severe autonomous apoptosis in the dorsal cells in the wing blade region. (C-C ) RNAi of RpS14. Similar to B-B, a severe autonomous apoptosis can be observed. puc-lacz expression (magenta). Caspase-3 activation (blue). Figure 5. Induction of autonomous apoptosis in the wide area of the wing disc leads to a non-autonomous apoptosis in an adjacent position. (A-A ) Just after induction of rpr expression. Active Caspase-3 is observed in the dorsal compartment. UAS-GFP expression in the dorsal compartment is not visualized yet at this point. Broken lines indicate the position of the DV boundary. (B-B ) 24 hrs after induction of rpr expression. A non-autonomously induced Caspase-3 activation is indicated by the arrows. (C-C ) 48 hrs after induction of rpr expression. Non-autonomous Caspase-3 activation is no longer induced. (D-D ) Induction of rpr expression without GAL80 ts. Most of the area in the dorsal compartment has already disappeared. puc-lacz expression (magenta). Caspase-3 activation (blue).
22 Figure 1
23 Figure 2
24 Figure 3
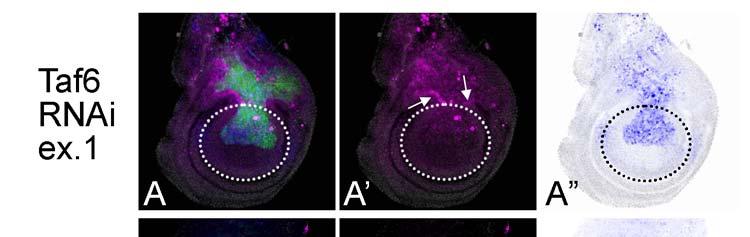
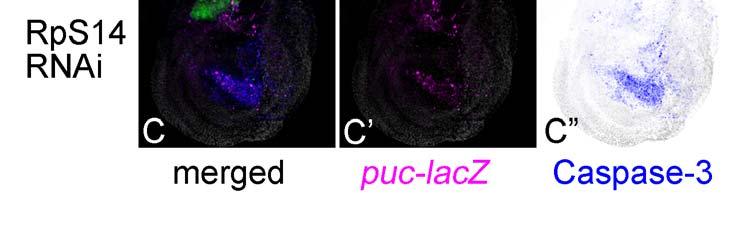
25 Figure 4
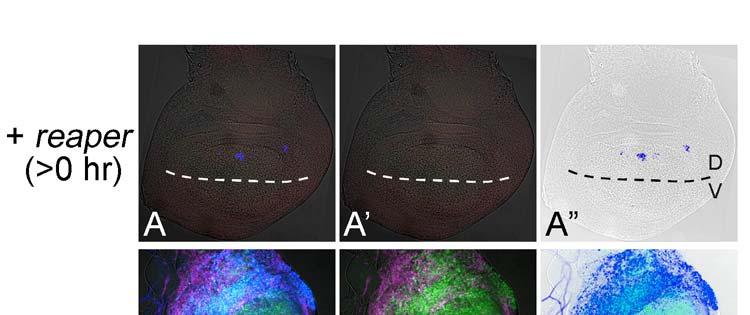
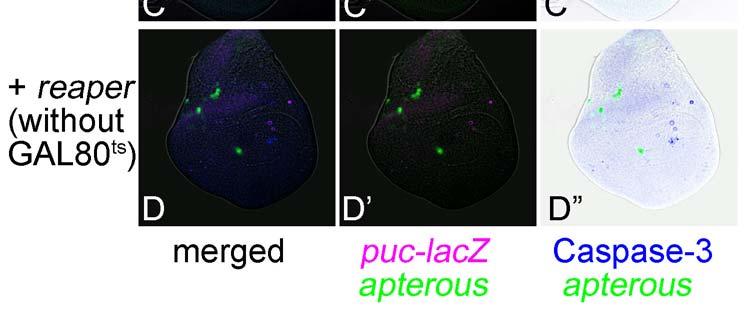
26 Figure 5
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Wing-to-Leg Homeosis by Spineless Causes Apoptosis Regulated by Fish-lips, a Novel Leucine-Rich Repeat Transmembrane Protein
 MOLECULAR AND CELLULAR BIOLOGY, Apr. 2005, p. 3140 3150 Vol. 25, No. 8 0270-7306/05/$08.00 0 doi:10.1128/mcb.25.8.3140 3150.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved.
MOLECULAR AND CELLULAR BIOLOGY, Apr. 2005, p. 3140 3150 Vol. 25, No. 8 0270-7306/05/$08.00 0 doi:10.1128/mcb.25.8.3140 3150.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved.
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Cell Death & Trophic Factors II. Steven McLoon Department of Neuroscience University of Minnesota
 Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Cell Death & Trophic Factors II Steven McLoon Department of Neuroscience University of Minnesota 1 Remember? Neurotrophins are cell survival factors that neurons get from their target cells! There is a
Supplementary Information
 Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Drosophila Apoptosis and the Regulation of the Caspase Cascade
 Drosophila Apoptosis and the Regulation of the Caspase Cascade Kate Stafford March 18, 2005 Abstract The caspase cascade in Drosophila is controlled primarily by DIAP1 (Drosophila inhibitor of apoptosis),
Drosophila Apoptosis and the Regulation of the Caspase Cascade Kate Stafford March 18, 2005 Abstract The caspase cascade in Drosophila is controlled primarily by DIAP1 (Drosophila inhibitor of apoptosis),
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Honors Biology Reading Guide Chapter 11
 Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Supplementary Materials for
 www.sciencemag.org/cgi/content/full/science.1244624/dc1 Supplementary Materials for Cytoneme-Mediated Contact-Dependent Transport of the Drosophila Decapentaplegic Signaling Protein Sougata Roy, Hai Huang,
www.sciencemag.org/cgi/content/full/science.1244624/dc1 Supplementary Materials for Cytoneme-Mediated Contact-Dependent Transport of the Drosophila Decapentaplegic Signaling Protein Sougata Roy, Hai Huang,
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
Lesson Overview. Gene Regulation and Expression. Lesson Overview Gene Regulation and Expression
 13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
Drosophila IAP1-mediated ubiquitylation controls activation of the initiator caspase DRONC independent of protein degradation
 University of Massachusetts Medical School escholarship@umms Molecular, Cell and Cancer Biology Publications Molecular, Cell and Cancer Biology 9-1-2011 Drosophila IAP1-mediated ubiquitylation controls
University of Massachusetts Medical School escholarship@umms Molecular, Cell and Cancer Biology Publications Molecular, Cell and Cancer Biology 9-1-2011 Drosophila IAP1-mediated ubiquitylation controls
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Apoptosis in Mammalian Cells
 Apoptosis in Mammalian Cells 7.16 2-10-05 Apoptosis is an important factor in many human diseases Cancer malignant cells evade death by suppressing apoptosis (too little apoptosis) Stroke damaged neurons
Apoptosis in Mammalian Cells 7.16 2-10-05 Apoptosis is an important factor in many human diseases Cancer malignant cells evade death by suppressing apoptosis (too little apoptosis) Stroke damaged neurons
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Chapter 4: A Deficiency Screen to Isolate Novel Regulators of DIAP1
 70 Chapter 4: A Deficiency Screen to Isolate Novel Regulators of DIAP1 4.1 Abstract DIAP1, a ubiquitin E3 ligase, is the key Drosophila regulator of the family of cysteine proteases known as caspases.
70 Chapter 4: A Deficiency Screen to Isolate Novel Regulators of DIAP1 4.1 Abstract DIAP1, a ubiquitin E3 ligase, is the key Drosophila regulator of the family of cysteine proteases known as caspases.
Caspase inhibition during apoptosis causes abnormal signalling and developmental aberrations in Drosophila
 Research article 5591 Caspase inhibition during apoptosis causes abnormal signalling and developmental aberrations in Drosophila Ainhoa Pérez-Garijo, Francisco A. Martín and Ginés Morata* Centro de Biología
Research article 5591 Caspase inhibition during apoptosis causes abnormal signalling and developmental aberrations in Drosophila Ainhoa Pérez-Garijo, Francisco A. Martín and Ginés Morata* Centro de Biología
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Developmental Biology
 Developmental Biology 334 (2009) 161 173 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology Wingless signaling and the control of cell
Developmental Biology 334 (2009) 161 173 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology Wingless signaling and the control of cell
DEVELOPMENT. Ainhoa Pérez-Garijo, Evgeny Shlevkov and Ginés Morata*
 RESEARCH ARTICLE 1169 Development 136, 1169-1177 (2009) doi:10.1242/dev.034017 The role of Dpp and Wg in compensatory proliferation and in the formation of hyperplastic overgrowths caused by apoptotic
RESEARCH ARTICLE 1169 Development 136, 1169-1177 (2009) doi:10.1242/dev.034017 The role of Dpp and Wg in compensatory proliferation and in the formation of hyperplastic overgrowths caused by apoptotic
Genes, Development, and Evolution
 14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
Morphogens in biological development: Drosophila example
 LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
S Phase Coupled E2f1 Destruction Ensures Homeostasis in Proliferating Tissues
 S Phase Coupled E2f1 Destruction Ensures Homeostasis in Proliferating Tissues Jean M. Davidson 1, Robert J. Duronio 1,2,3 * 1 Department of Biology, The University of North Carolina at Chapel Hill, Chapel
S Phase Coupled E2f1 Destruction Ensures Homeostasis in Proliferating Tissues Jean M. Davidson 1, Robert J. Duronio 1,2,3 * 1 Department of Biology, The University of North Carolina at Chapel Hill, Chapel
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Tuesday, December 27, 16
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
13.4 Gene Regulation and Expression
 13.4 Gene Regulation and Expression Lesson Objectives Describe gene regulation in prokaryotes. Explain how most eukaryotic genes are regulated. Relate gene regulation to development in multicellular organisms.
13.4 Gene Regulation and Expression Lesson Objectives Describe gene regulation in prokaryotes. Explain how most eukaryotic genes are regulated. Relate gene regulation to development in multicellular organisms.
Lobe and Serrate are required for cell survival during early eye development in Drosophila
 RESEARCH ARTICLE 4771 Development 133, 4771-4781 (2006) doi:10.1242/dev.02686 Lobe and Serrate are required for cell survival during early eye development in Drosophila Amit Singh 1, *, Xiao Shi 1 and
RESEARCH ARTICLE 4771 Development 133, 4771-4781 (2006) doi:10.1242/dev.02686 Lobe and Serrate are required for cell survival during early eye development in Drosophila Amit Singh 1, *, Xiao Shi 1 and
Lobe and Serrate are Required for Cell Survival during Early Eye Development in Drosophila
 University of Dayton ecommons Biology Faculty Publications Department of Biology 11-2006 Lobe and Serrate are Required for Cell Survival during Early Eye Development in Drosophila Amit Singh University
University of Dayton ecommons Biology Faculty Publications Department of Biology 11-2006 Lobe and Serrate are Required for Cell Survival during Early Eye Development in Drosophila Amit Singh University
Biol403 - Receptor Serine/Threonine Kinases
 Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
On the Mechanism of Wing Size Determination in Fly Development
 On the Mechanism of Wing Size Determination in Fly Development PNAS Paper Authors: Lars Hufnagel, Aurelio A. Teleman, Herve Rouault, Stephen M. Cohen, and Boris I. Shraiman Group Members: Rebekah Starks,
On the Mechanism of Wing Size Determination in Fly Development PNAS Paper Authors: Lars Hufnagel, Aurelio A. Teleman, Herve Rouault, Stephen M. Cohen, and Boris I. Shraiman Group Members: Rebekah Starks,
Initiation of translation in eukaryotic cells:connecting the head and tail
 Initiation of translation in eukaryotic cells:connecting the head and tail GCCRCCAUGG 1: Multiple initiation factors with distinct biochemical roles (linking, tethering, recruiting, and scanning) 2: 5
Initiation of translation in eukaryotic cells:connecting the head and tail GCCRCCAUGG 1: Multiple initiation factors with distinct biochemical roles (linking, tethering, recruiting, and scanning) 2: 5
Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Written Exam 15 December Course name: Introduction to Systems Biology Course no
 Technical University of Denmark Written Exam 15 December 2008 Course name: Introduction to Systems Biology Course no. 27041 Aids allowed: Open book exam Provide your answers and calculations on separate
Technical University of Denmark Written Exam 15 December 2008 Course name: Introduction to Systems Biology Course no. 27041 Aids allowed: Open book exam Provide your answers and calculations on separate
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
Zool 3200: Cell Biology Exam 5 4/27/15
 Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Nature Neuroscience: doi: /nn.2662
 Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Drosophila wing. Temporal regulation of Apterous activity during development of the. Marco Milán and Stephen M. Cohen* SUMMARY
 Development 127, 3069-3078 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2547 3069 Temporal regulation of Apterous activity during development of the Drosophila wing Marco Milán
Development 127, 3069-3078 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2547 3069 Temporal regulation of Apterous activity during development of the Drosophila wing Marco Milán
Prokaryotic Regulation
 Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Prokaryotic Regulation Control of transcription initiation can be: Positive control increases transcription when activators bind DNA Negative control reduces transcription when repressors bind to DNA regulatory
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Nature Biotechnology: doi: /nbt Supplementary Figure 1. Overexpression of YFP::GPR-1 in the germline.
 Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Role of Mitochondrial Remodeling in Programmed Cell Death in
 Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290
 PLoS Biol 4(9): e290") Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Shavenbaby Couples Patterning to Epidermal Cell Shape Control. Chanut-Delalande H, Fernandes I, Roch F, Payre F, Plaza S (2006) PLoS Biol 4(9): e290 Question (from Introduction): How does svb control the
Upstream Elements Regulating mir-241 and mir-48 Abstract Introduction
 Upstream Elements Regulating mir-241 and mir-48 Hanna Vollbrecht, Tamar Resnick, and Ann Rougvie University of Minnesota: Twin Cities Undergraduate Research Scholarship 2012-2013 Abstract Caenorhabditis
Upstream Elements Regulating mir-241 and mir-48 Hanna Vollbrecht, Tamar Resnick, and Ann Rougvie University of Minnesota: Twin Cities Undergraduate Research Scholarship 2012-2013 Abstract Caenorhabditis
Systematic in vivo RNAi analysis of putative components of the Drosophila cell death machinery Introduction UNCORRECTED PROOF
 (2006) 00, 1 12 & 2006 Nature Publishing Group All rights reserved 1350-9047/06 $30.00 www.nature.com/cdd Systematic in vivo RNAi analysis of putative components of the Drosophila cell death machinery
(2006) 00, 1 12 & 2006 Nature Publishing Group All rights reserved 1350-9047/06 $30.00 www.nature.com/cdd Systematic in vivo RNAi analysis of putative components of the Drosophila cell death machinery
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION
 MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Characterization of head involution defective (hid) as a pro-apoptotic gene in Megasalia scalaris
 as a pro-apoptotic gene in Megasalia scalaris") University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange University of Tennessee Honors Thesis Projects University of Tennessee Honors Program 5-2015 Characterization of head
University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange University of Tennessee Honors Thesis Projects University of Tennessee Honors Program 5-2015 Characterization of head
Robustness of Tissue Patterns*
 MCBU Project II - 2014 Robustness of Tissue Patterns* June, 2014 Frederic Y.M. Wan Mathematics University of California, Irvine Supported by: NIH Grants R01-GM67247 P50-GM66051 Biological Patterning The
MCBU Project II - 2014 Robustness of Tissue Patterns* June, 2014 Frederic Y.M. Wan Mathematics University of California, Irvine Supported by: NIH Grants R01-GM67247 P50-GM66051 Biological Patterning The
Drosophila Somatic Anterior-Posterior Axis (A-P Axis) Formation
 Formation") Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics
Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics
1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms.
 Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
PRACTICE EXAM. 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos.
 PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
Autophagy-independent function of Atg1 for apoptosis-induced compensatory proliferation
 University of Massachusetts Medical School escholarship@umms Molecular, Cell and Cancer Biology Publications Molecular, Cell and Cancer Biology 8-19-2016 Autophagy-independent function of Atg1 for apoptosis-induced
University of Massachusetts Medical School escholarship@umms Molecular, Cell and Cancer Biology Publications Molecular, Cell and Cancer Biology 8-19-2016 Autophagy-independent function of Atg1 for apoptosis-induced
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila
 Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
9/4/2015 INDUCTION CHAPTER 1. Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology. Fig 1.
 INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
Computational Structural Bioinformatics
 Computational Structural Bioinformatics ECS129 Instructor: Patrice Koehl http://koehllab.genomecenter.ucdavis.edu/teaching/ecs129 koehl@cs.ucdavis.edu Learning curve Math / CS Biology/ Chemistry Pre-requisite
Computational Structural Bioinformatics ECS129 Instructor: Patrice Koehl http://koehllab.genomecenter.ucdavis.edu/teaching/ecs129 koehl@cs.ucdavis.edu Learning curve Math / CS Biology/ Chemistry Pre-requisite
Chapter 18 Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
SUPPLEMENTARY INFORMATION
 Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Genetic transcription and regulation
 Genetic transcription and regulation Central dogma of biology DNA codes for DNA DNA codes for RNA RNA codes for proteins not surprisingly, many points for regulation of the process DNA codes for DNA replication
Genetic transcription and regulation Central dogma of biology DNA codes for DNA DNA codes for RNA RNA codes for proteins not surprisingly, many points for regulation of the process DNA codes for DNA replication
Chapter 3: Novel Regulators of the C. elegans Caspase CED-3
 53 Chapter 3: Novel Regulators of the C. elegans Caspase CED-3 3.1 Abstract The hallmark of apoptosis is the presence of activated caspases, a conserved family of cysteine proteases. Proper regulation
53 Chapter 3: Novel Regulators of the C. elegans Caspase CED-3 3.1 Abstract The hallmark of apoptosis is the presence of activated caspases, a conserved family of cysteine proteases. Proper regulation
Proteome-wide High Throughput Cell Based Assay for Apoptotic Genes
 John Kenten, Doug Woods, Pankaj Oberoi, Laura Schaefer, Jonathan Reeves, Hans A. Biebuyck and Jacob N. Wohlstadter 9238 Gaither Road, Gaithersburg, MD 2877. Phone: 24.631.2522 Fax: 24.632.2219. Website:
John Kenten, Doug Woods, Pankaj Oberoi, Laura Schaefer, Jonathan Reeves, Hans A. Biebuyck and Jacob N. Wohlstadter 9238 Gaither Road, Gaithersburg, MD 2877. Phone: 24.631.2522 Fax: 24.632.2219. Website:
Welcome to Class 21!
 Welcome to Class 21! Introductory Biochemistry! Lecture 21: Outline and Objectives l Regulation of Gene Expression in Prokaryotes! l transcriptional regulation! l principles! l lac operon! l trp attenuation!
Welcome to Class 21! Introductory Biochemistry! Lecture 21: Outline and Objectives l Regulation of Gene Expression in Prokaryotes! l transcriptional regulation! l principles! l lac operon! l trp attenuation!
MATHEMATICAL MODELS - Vol. III - Mathematical Modeling and the Human Genome - Hilary S. Booth MATHEMATICAL MODELING AND THE HUMAN GENOME
 MATHEMATICAL MODELING AND THE HUMAN GENOME Hilary S. Booth Australian National University, Australia Keywords: Human genome, DNA, bioinformatics, sequence analysis, evolution. Contents 1. Introduction:
MATHEMATICAL MODELING AND THE HUMAN GENOME Hilary S. Booth Australian National University, Australia Keywords: Human genome, DNA, bioinformatics, sequence analysis, evolution. Contents 1. Introduction:
Evolution of the Complex Eye and Pax6 Gene
 Evolution of the Complex Eye and Pax6 Gene Rachel Thomsen Sarah Kim Jenia Ostrovskaya Key Points Definition of an Eye o Types of Eyes Origin of Species: Difficulties on Theory Pax family Pax6 gene why
Evolution of the Complex Eye and Pax6 Gene Rachel Thomsen Sarah Kim Jenia Ostrovskaya Key Points Definition of an Eye o Types of Eyes Origin of Species: Difficulties on Theory Pax family Pax6 gene why
Cell biology: Death drags down the neighbourhood
 Cell biology: Death drags down the neighbourhood The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published Publisher Vasquez,
Cell biology: Death drags down the neighbourhood The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published Publisher Vasquez,
REVIEW SESSION. Wednesday, September 15 5:30 PM SHANTZ 242 E
 REVIEW SESSION Wednesday, September 15 5:30 PM SHANTZ 242 E Gene Regulation Gene Regulation Gene expression can be turned on, turned off, turned up or turned down! For example, as test time approaches,
REVIEW SESSION Wednesday, September 15 5:30 PM SHANTZ 242 E Gene Regulation Gene Regulation Gene expression can be turned on, turned off, turned up or turned down! For example, as test time approaches,
Warm-Up. Explain how a secondary messenger is activated, and how this affects gene expression. (LO 3.22)
") Warm-Up Explain how a secondary messenger is activated, and how this affects gene expression. (LO 3.22) Yesterday s Picture The first cell on Earth (approx. 3.5 billion years ago) was simple and prokaryotic,
Warm-Up Explain how a secondary messenger is activated, and how this affects gene expression. (LO 3.22) Yesterday s Picture The first cell on Earth (approx. 3.5 billion years ago) was simple and prokaryotic,
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Molecular Developmental Physiology and Signal Transduction
 Prof. Dr. J. Vanden Broeck (Animal Physiology and Neurobiology - Dept. of Biology - KU Leuven) Molecular Developmental Physiology and Signal Transduction My Research Team Insect species under study +
Prof. Dr. J. Vanden Broeck (Animal Physiology and Neurobiology - Dept. of Biology - KU Leuven) Molecular Developmental Physiology and Signal Transduction My Research Team Insect species under study +
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Gene regulation I Biochemistry 302. Bob Kelm February 25, 2005
 Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Regulation of gene expression. Premedical - Biology
 Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Chapter 15 Active Reading Guide Regulation of Gene Expression
 Name: AP Biology Mr. Croft Chapter 15 Active Reading Guide Regulation of Gene Expression The overview for Chapter 15 introduces the idea that while all cells of an organism have all genes in the genome,
Name: AP Biology Mr. Croft Chapter 15 Active Reading Guide Regulation of Gene Expression The overview for Chapter 15 introduces the idea that while all cells of an organism have all genes in the genome,
Control of Gene Expression
 Control of Gene Expression Mechanisms of Gene Control Gene Control in Eukaryotes Master Genes Gene Control In Prokaryotes Epigenetics Gene Expression The overall process by which information flows from
Control of Gene Expression Mechanisms of Gene Control Gene Control in Eukaryotes Master Genes Gene Control In Prokaryotes Epigenetics Gene Expression The overall process by which information flows from
Measuring TF-DNA interactions
 Measuring TF-DNA interactions How is Biological Complexity Achieved? Mediated by Transcription Factors (TFs) 2 Regulation of Gene Expression by Transcription Factors TF trans-acting factors TF TF TF TF
Measuring TF-DNA interactions How is Biological Complexity Achieved? Mediated by Transcription Factors (TFs) 2 Regulation of Gene Expression by Transcription Factors TF trans-acting factors TF TF TF TF
NOPO modulates Egr-induced JNK-independent cell death in Drosophila
 ORIGINAL ARTICLE Cell Research (2012) 22:425-431. 2012 IBCB, SIBS, CAS All rights reserved 1001-0602/12 $ 32.00 www.nature.com/cr npg NOPO modulates Egr-induced JNK-independent cell death in Drosophila
ORIGINAL ARTICLE Cell Research (2012) 22:425-431. 2012 IBCB, SIBS, CAS All rights reserved 1001-0602/12 $ 32.00 www.nature.com/cr npg NOPO modulates Egr-induced JNK-independent cell death in Drosophila
Bio 3411, Fall 2006, Lecture 19-Cell Death.
 Types of Cell Death Questions : Apoptosis (Programmed Cell Death) : Cell-Autonomous Stereotypic Rapid Clean (dead cells eaten) Necrosis : Not Self-Initiated Not Stereotypic Can Be Slow Messy (injury can
Types of Cell Death Questions : Apoptosis (Programmed Cell Death) : Cell-Autonomous Stereotypic Rapid Clean (dead cells eaten) Necrosis : Not Self-Initiated Not Stereotypic Can Be Slow Messy (injury can
15.2 Prokaryotic Transcription *
 OpenStax-CNX module: m52697 1 15.2 Prokaryotic Transcription * Shannon McDermott Based on Prokaryotic Transcription by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons
OpenStax-CNX module: m52697 1 15.2 Prokaryotic Transcription * Shannon McDermott Based on Prokaryotic Transcription by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons
Characterization of the apoptotic functions of the HID hmolog isolated from Megaselia scalaris
 University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange University of Tennessee Honors Thesis Projects University of Tennessee Honors Program 5-2013 Characterization of the apoptotic
University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange University of Tennessee Honors Thesis Projects University of Tennessee Honors Program 5-2013 Characterization of the apoptotic
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Proteomics. Yeast two hybrid. Proteomics - PAGE techniques. Data obtained. What is it?
 Proteomics What is it? Reveal protein interactions Protein profiling in a sample Yeast two hybrid screening High throughput 2D PAGE Automatic analysis of 2D Page Yeast two hybrid Use two mating strains
Proteomics What is it? Reveal protein interactions Protein profiling in a sample Yeast two hybrid screening High throughput 2D PAGE Automatic analysis of 2D Page Yeast two hybrid Use two mating strains
Transcription Regulation And Gene Expression in Eukaryotes UPSTREAM TRANSCRIPTION FACTORS
 Transcription Regulation And Gene Expression in Eukaryotes UPSTREAM TRANSCRIPTION FACTORS RG. Clerc March 26. 2008 UPSTREAM TRANSCRIPTION FACTORS Experimental approaches DNA binding domains (DBD) Transcription
Transcription Regulation And Gene Expression in Eukaryotes UPSTREAM TRANSCRIPTION FACTORS RG. Clerc March 26. 2008 UPSTREAM TRANSCRIPTION FACTORS Experimental approaches DNA binding domains (DBD) Transcription
RANK. Alternative names. Discovery. Structure. William J. Boyle* SUMMARY BACKGROUND
 RANK William J. Boyle* Department of Cell Biology, Amgen, Inc., One Amgen Center Drive, Thousand Oaks, CA 91320-1799, USA * corresponding author tel: 805-447-4304, fax: 805-447-1982, e-mail: bboyle@amgen.com
RANK William J. Boyle* Department of Cell Biology, Amgen, Inc., One Amgen Center Drive, Thousand Oaks, CA 91320-1799, USA * corresponding author tel: 805-447-4304, fax: 805-447-1982, e-mail: bboyle@amgen.com
MCB 141 Midterm I Feb. 14, 2012
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 14, 2012 Question #1 Question #2 Question #3 Question #4 BONUS / 28 pts / 27 pts / 25 pts / 20 pts / 1 pt TOTAL / 100 pts
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 14, 2012 Question #1 Question #2 Question #3 Question #4 BONUS / 28 pts / 27 pts / 25 pts / 20 pts / 1 pt TOTAL / 100 pts
COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION
 COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION I. Aksan 1, M. Sen 2, M. K. Araz 3, and M. L. Kurnaz 3 1 School of Biological Sciences, University of Manchester,
COMPUTER SIMULATION OF DIFFERENTIAL KINETICS OF MAPK ACTIVATION UPON EGF RECEPTOR OVEREXPRESSION I. Aksan 1, M. Sen 2, M. K. Araz 3, and M. L. Kurnaz 3 1 School of Biological Sciences, University of Manchester,
UNIVERSITY OF YORK BIOLOGY. Developmental Biology
 Examination Candidate Number: UNIVERSITY OF YORK BSc Stage 2 Degree Examinations 2017-18 Department: BIOLOGY Title of Exam: Developmental Biology Desk Number: Time allowed: 1 hour and 30 minutes Total
Examination Candidate Number: UNIVERSITY OF YORK BSc Stage 2 Degree Examinations 2017-18 Department: BIOLOGY Title of Exam: Developmental Biology Desk Number: Time allowed: 1 hour and 30 minutes Total
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its
 The Wnt/ -catenin signalling pathway and its") Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday
 Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday 1. What is the Central Dogma? 2. How does prokaryotic DNA compare to eukaryotic DNA? 3. How is DNA
Complete all warm up questions Focus on operon functioning we will be creating operon models on Monday 1. What is the Central Dogma? 2. How does prokaryotic DNA compare to eukaryotic DNA? 3. How is DNA