Clap and Fling Mechanism with Interacting Porous Wings in Tiny Insect Flight
|
|
|
- Augustus Hart
- 6 years ago
- Views:
Transcription
1 First posted online on 4 September 2014 as /jeb J Exp Biol Advance Access Online the most Articles. recent version First at posted online on 4 September 2014 as doi: /jeb Access the most recent version at The Journal of Experimental Biology ACCEPTED AUTHOR MANUSCRIPT Clap and Fling Mechanism with Interacting Porous Wings in Tiny Insect Flight Arvind Santhanakrishnan 1, Alice K. Robinson 2, Shannon Jones 4, Audrey Ann Low 3, Sneha Gadi 3, Tyson L. Hedrick 4 and Laura A. Miller 3,4,* 1 School of Mechanical & Aerospace Engineering, Oklahoma State University, 218 Engineering North, Stillwater, OK , USA, 2 University of California, Irvine, Irvine, CA 92697, USA, 3 Department of Mathematics, University of North Carolina, Chapel Hill, NC 27599, USA 4 Department of Biology, University of North Carolina, Chapel Hill, NC 27599, USA * Author for correspondence ( lam9@ .unc.edu) Summary The aerodynamics of flapping flight for the smallest insects such as thrips is often characterized by a 'clap and fling' of the wings at the end of the upstroke and the beginning of the downstroke. These insects fly at Reynolds numbers (Re) on the order of 10 or less where viscous effects are significant. Although this wing motion is known to augment the lift generated during flight, the drag required to fling the wings apart at this scale is an order of magnitude larger than the corresponding force acting on a single wing. Since the opposing forces acting normal to each wing nearly cancel during the fling, these large forces do not have a clear aerodynamic benefit. If flight efficiency is defined as the ratio of lift to drag, the 'clap and fling' motion dramatically reduces efficiency relative to the case of wings that do not aerodynamically interact. In this paper, the effect of a bristled wing characteristic of many of these insects is investigated using computational fluid dynamics. We perform 2D numerical simulations using a porous version of the immersed boundary method. Given the computational complexity involved in modeling flow through exact descriptions of bristled wings, the wing is modeled as a homogenous porous layer as a first approximation. High-speed video recordings of free flying thrips in take-off flight were captured in the laboratory, and an analysis of the wing kinematics was performed. This information was used for the estimation of input parameters for the simulations. As compared to a solid wing (without bristles), the results of the study show that the porous nature of the wings contributes largely to drag reduction across the Re range explored. The aerodynamic efficiency, calculated as the ratio of lift to drag coefficients, was larger for some porosities when compared to solid wings Published by The Company of Biologists Ltd
2 Keywords: tiny insect, aerodynamics, flapping flight, wing kinematics, locomotion, clap and fling, bristled wing, fringed wing, immersed boundary method Introduction Though hard to notice even when they hop, walk, or fly, the smallest flying insects reported in the literature have body lengths less than 1 mm and are of considerable ecological and agricultural importance. For example, over 5500 species of thrips have been described thus far (Morse and Hoddle, 2006). They function in multiple important roles such as: 1) effective transmitters of pollen during feeding (Terry, 2001; Terry, 2002), 2) invasive pests of agriculturally important plants (Crespi et al., 1997; Palmer et al., 1990) and 3) biological vectors of microbial plant pathogens such as Tospoviruses (Ullman et al., 2002; Jones, 2005). Parasitoid wasps consist of a dozen hymenopteran superfamilies that include some of the smallest insects such as Mymar sp. These insects have received much attention in recent years due to their potential as a natural control of agricultural pests (Austin and Dowton, 2008). Understanding the aerodynamics of flapping flight in these tiny insects provides a means of exploring their dispersal strategies and ranges. Understanding the flight of these organisms may also elucidate the principles of active flight in some of the smallest animals (Lewis, 1964; Lewis, 1965). Although many researchers have investigated the complex nature of the aerodynamics of flapping flight in insects ranging from the scale of the hawk moth Manduca sexta (Usherwood and Ellington, 2002; Wang, 2005; Hedrick et al., 2009) to the scale of the fruit fly Drosophila (Vogel, 1962; Dickinson and Götz, 1993; Dickinson et al., 1999), the aerodynamics of flight in the smallest insects remains relatively unexplored (Horridge, 1956; Weis-Fogh, 1973; Ellington, 1980; Miller and Peskin, 2009). The Reynolds number (Re) is commonly used to quantify the ratio of inertial to viscous effects in a fluid and is given as ¹ Ê Ê, where Ê is the density of air, µ is the dynamic viscosity of air, L is the chord length of the wing, and U is the wing tip velocity. Drosophila and Manduca span the Re range from about 100 to 1000, while some of the smallest thrips and parasitoid wasps fly at Re < 10. 2
3 It appears that one of the predominant lift augmenting mechanisms for nearly all insects is flight at high angles of attack ( 45 ). The resulting high lift is possibly due to the presence of a stable leading edge vortex which remains attached to the upper surface of the wing until stroke reversal (Ellington et al, 1996; Van Den Berg and Ellington, 1997a,b; Liu et al., 1998; Usherwood and Ellington, 2002; Birch and Dickinson, 2003; Birch et al., 2004). This vortex creates a negative pressure region above the wing and enhances lift. Lift is also generated when the trailing edge vortex (also known as the starting vortex) is shed from the wing and consequently induces circulation around the wing. Wake capture could also enhance lift as the wing moves back through its wake due in part to the fact that the velocity of the fluid relative to the wing is increased (Wang, 2000). Flight aerodynamics and wing kinematics for the smallest insects differ from those of larger animals such as Drosophila species. Horridge (1956) proposed that tiny insects employ an asymmetric flight stroke via adjusting the angle of attack differentially such that most of the lift is generated during the downstroke and negative lift is minimized during the upstroke. Weis- Fogh (1973) analyzed video recordings capturing the hovering flight of the parasitic tiny wasp Encarsia formosa and discovered the use of a clap and fling mechanism. More specifically, the wings were observed to clap together at the end of each upstroke and fling apart at the beginning of each downstroke (see Figure 1). The fling motion is produced by a rotation of the wings about the common trailing edge, during which time a pair of large leading edge vortices (LEV) are formed (Maxworthy, 1979). Lighthill (1973) analytically showed that the clap and fling mechanism could be used for lift augmentation even in 2D inviscid flows. The clap and fling mechanism has since been observed in other insects such as Thrips physapus (Ellington, 1980, 1984), the parasitoid wasps Muscidifurax raptor and Nasonia vitripennis (Miller and Peskin, 2009), the greenhouse white-fly Trialeurodes vaporariorum (Weis-Fogh, 1975), and some butterflies (Srygley and Thomas, 2002). It is also commonly observed in the tethered flight of larger insects such as Drosophila species (Vogel, 1967). In addition to measurements of tiny insects in free flight, computational fluid dynamics has been used to explore the force generation and resulting flow structures produced during the flight of tiny insects. Here we describe the dimensionless force coefficients as follows: 3
4 Ê, 2 Ê where U is the constant characteristic velocity, describes the lift or vertical component of the force acting against gravity, describes the drag or horizontal component of the force, and is the surface area of the wing. Note that the characteristic velocity, U, does not change in time so that the force coefficients are functions of time. Another way to think about and is to consider them as dimensionless forces. Numerical simulations of flapping wings have shown that lift coefficients are reduced and drag coefficients are increased significantly when the Re is lowered from 100 to 20 or below (Wang, 2000; Miller and Peskin, 2004; Wu and Sun, 2004). Numerical simulations of clap and fling show that lift coefficients can be enhanced by almost 100% for Re in the range of the smallest insects (Sun and Xin, 2003; Miller and Peskin, 2005; Kolomenskiy et al., 2009). The cost of clap and fling for small insects (Re ~10) is that the drag required to fling the wings apart may be an order of magnitude larger than the force required to move a single wing with the same motion (Miller and Peskin, 2009). Since the opposing drag forces on each wing nearly cancel during the fling, these large forces do not have a clear aerodynamic benefit. The previous experimental and computational work has focused on models of solid wings, but the wings of many small insects such as thrips are fringed or bristled rather than continuous (Ellington, 1980) (see Figure 2). The aerodynamic benefits of this type of wing are not clear. Sunada et al. (2002) constructed a dynamically scaled model of a thrips wing and measured lift and drag generated during single wing translation and rotation. Their results showed that lift and drag scaled proportionally with the addition of bristles, and they did not find a clear aerodynamic benefit. Since the forces generated during the fling are up to an order of magnitude greater than the single wing case, it is possible that large differences in bristled wing aerodynamics may be seen when wing-wing interaction is considered. The amount of airflow between the bristles is proportional to the normal force or pressure difference across the wing, and this effect could potentially result in significant airflow through the wings when forces are large. In this study, a porous version of the immersed boundary was used to investigate the aerodynamics of bristled wings during clap and fling. Recordings of thrips in free take-off flight 1 4
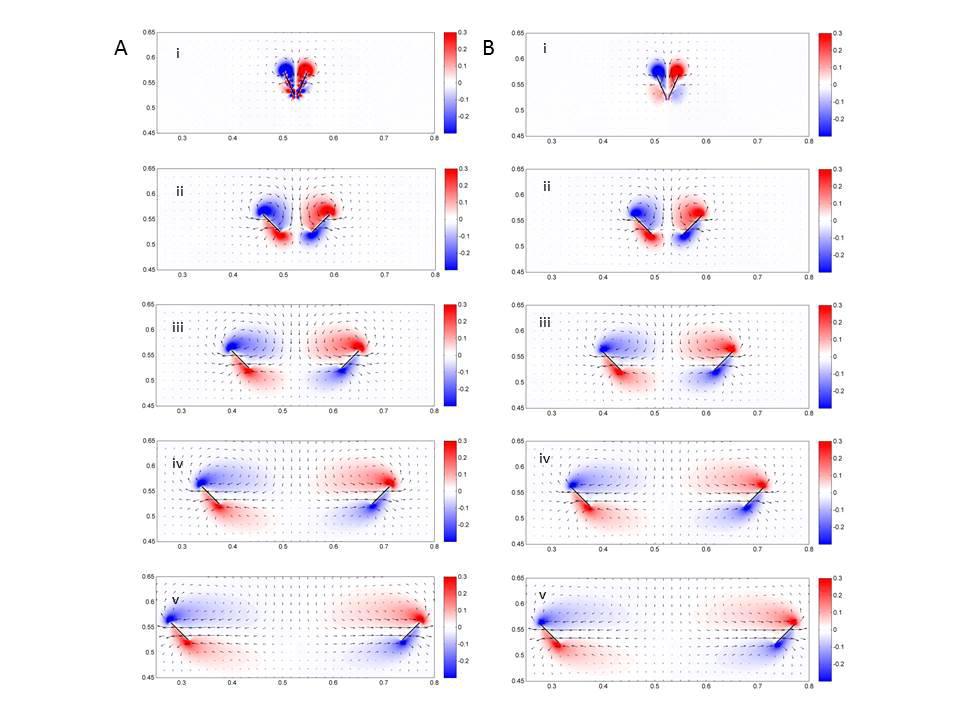
5 were filmed in the laboratory using high-speed videography. Kinematic analyses were performed, and the results were used to select input parameters for the simulations. Given the computational difficulty of resolving the flow through dozens of bristles in three dimensions, the wings were approximated to be homogenous porous layers in two dimensions. Lift and drag coefficients were calculated as functions of time during the flapping cycle. Parametric studies were performed to compare the aerodynamics of solid wings and bristled wings with several levels of porosity for Re 10. Results Kinematic observations Snapshots taken during the clap and fling wing wingbeat of a typical thrips in free takeoff flight are shown in Figure 3. The images were recorded at 4000 frames per second. Note the characteristic bristled wing form. At the end of the upstroke (the clap), the wings are raised normal to the body axis and come within about 1/6 1/10 chord lengths of each other. This clap motion is characterized by a simultaneous rotation of the wing about the base and the trailing edge. At the beginning of the downstroke (the fling), the wings rotate simultaneously about the trailing edge and the wing base, which results in the fling motion. Videos of typical thrips in takeoff flight are shown in Supplementary Material S1-S3. Four of the videos collected show thrips that cease to flap their wings shortly after takeoff. During this time, the insects spread their wings and passively floated downward. This behavior is termed parachuting herein rather than gliding, using the convention given by Alexander (2003). Parachuting refers to the use of drag producing structures in order to reduce the speed of falling. Gliding, on the other hand, relies on lift production. To make an operational distinction between the two, maneuvers using descent angles lower than 45 are considered to be gliding maneuvers and above 45 are considered to parachuting maneuvers. In all cases the descent angles for parachuting thrips were greater than 45 (e.g. the thrips were not gliding). Snapshots taken during a typical parachuting maneuver are shown in Figure 4. In this case, the thrips spreads its four wings above its body about 8 wingbeats after takeoff. The insect then passively parachutes downward until it is out of the field of view of the camera. Supplementary Material S4 shows a video of this maneuver. 5
6 Wing tip speeds as functions of time for two representative thrips are shown in Figure 5. The two forewings were tracked over 3 strokes using DLTdv5 and velocities measured via direct numeric differentiation. Maximum wing tip velocities are on the order of 1.5 m/s. The average flapping frequency measured for 8 individuals (and a total of 45 wing strokes) is 254±32 Hz. A paired two sample t-test shows that the duration of the average upstroke was significantly shorter than the average downstroke (p= ). The duration of the upstroke was 1.87±0.27 ms and the duration of the downstroke was 2.13±0.47 ms. The length of the wings (base to tip) was 0.644±0.193 mm (n=6). If we assume an aspect ratio of about 2 (see Figure 2), this results in a Reynolds number of about 14 based upon the average wing tip velocity and a total sweep of 120 degrees. Looking at the smallest (0.383 mm span) to largest (1.03 mm span) individual gives a range of 4.9 to 35. Mathematical modeling The complex structure of the bristled wing prevalent in tiny insects such as thrips is modeled herein as a porous layer using the method derived by Kim and Peskin (2006) and Stockie (2009). Permeability is incorporated into the immersed boundary method using an interpretation of Darcy s law, which states that the relative velocity of a fluid through a porous medium is proportional to the pressure difference across the boundary: É º 2 Ê where is the volumetric flow rate, κ is the permeability of the layer (with units of m 2 ), [p] is the pressure difference across the layer, is the thickness of the layer, is the surface area of the layer, and Ê is the viscosity of the fluid. Equating the volumetric flow rate to the difference between the local fluid velocity and the velocity of the boundary results in an expression for the slip between the boundary and fluid. This is incorporated into the immersed boundary method by modifying the velocity of the boundary. Rather than moving the boundary at the local fluid velocity, a slip is used that is 6
7 proportional to the force per unit area acting normal to the boundary (which is equivalent to the pressure jump) and the porosity. The velocity of boundary motion is modified as follows: The Journal of Experimental Biology ACCEPTED AUTHOR MANUSCRIPT ³ º,» ³ º,» Λ ² º,» µ µ 3 where X(q,t) gives the Cartesian coordinates of the boundary point labeled by the Lagrangian parameter q at time t, U is the velocity of the boundary at position q, F(q,t) is the force per unit length applied by the boundary to the fluid as a function of the Lagrangian position q and the time t, n is the unit vector normal to the boundary, and Ç is a proportionality constant termed the porosity by Kim and Peskin (2006) and has units of m 2 /N*s. The physical interpretation of the porosity coefficient, Ç, is that it is equal to the number of pores in an interval multiplied by the conductance of the material per unit arc length. The relationship between the porosity, Ç, and the permeability, κ, is given by Ç = κ/(a p µ). Relating the porosity to the leakiness Another metric that has been used to measure the permeability of a structure is the leakiness, (Cheer and Koehl, 1987). is calculated as the ratio of the actual flux through the structure over the flux in the inviscid (Ê = 0) case, 4 where U is the freestream velocity, Q is the volumetric flow rate. Note that there is no flow through the porous structure when = 0, and the flow is inviscid when = 1. The leakiness of thrips wings and other bristled appendages can be estimated using the analytical results of Cheer and Koehl (1987). These results have also been experimentally validated by Loudon et al. (1994). Thrips have a cylinder diameter to gap ratio of 1:10 and 1:12 at a bristle based Re of 1.0 x 10-2 (Ellington, 1980). This leads to leakiness values ranging from about The porosity, Λ, used by Kim and Peskin can be directly related to the leakiness,, using an appropriate nondimensionalization. The leakiness is calculated at steady state for cylinders arranged normal to the direction of flow. If one divides the porosity by the steady state force per 7
8 unit area at a 90 degree angle of attack,, over the free stream velocity, U, the result is the percent slip between the boundary and the fluid: ¹ Λ 5 The Journal of Experimental Biology ACCEPTED AUTHOR MANUSCRIPT where Le is the dimensionless porosity. When the boundary moves normal to the direction flow at steady state, Le is equivalent the leakiness defined by Cheer and Koehl (1987). This relationship uses the fact that the pressure jump across the porous boundary is equal to the normal force per area acting on boundary, as described in Kim and Peskin (2006). The choice of characteristic force per unit area used in this study was 1 2 Ê, where was set to 6 (the approximate drag coefficient of a Re=10 wing at a 90º angle of attack). Leakiness is then maximized when the wing moves with a 90 angle of attack and would drop to zero at a 0 angle of attack (no flow in the tangential direction). In the simulations that follow, the effective leakiness varies in time as the normal force acting on the boundary changes in time due to wing- wing interactions, acceleration, and changes in angle of attack. Throughout the remainder of the paper we will refer to the dimensionless porosity, Le, as the leakiness. Table 1 shows all parameter values used in the simulations. For easy comparison to thrips and other insects, all simulation parameters are dimensionless using the following relationships:»»»» º º Ê»» where tildas are used to denote the dimensionless parameter. Re was varied from 4 to 10 to span the range of tiny insects from the smallest parasitoid wasps to thrips. Re was adjusted by keeping all length and velocity scales constant and changing the dynamic viscosity appropriately. Le was varied from 0 (solid wing) to Description of prescribed kinematics The simplified flight kinematics of clap and fling used in this paper are similar to those used in a number of experimental, analytical, and computational papers (Lighthill, 1973; Bennett, 1977; Spedding and Maxworthy, 1986; Sun and Xin, 2003; Miller and Peskin, 2005). The translational and angular velocities during each half stroke were constructed using a series of equations to 6 8
9 describe each part of the stroke as described previously (Miller and Peskin, 2009). For the case of clap, the wings translate towards each other at a constant rate at a constant 45º angle of attack. Deceleration and wing rotation then begin simultaneously at the end of the stroke. The wings rotate about the leading edges from 45º to 90º. For the case of fling, the wings rotate apart about the trailing edges from a 90º to a 45º angle of attack. Translation begins halfway during wing rotation. At the end of the translational acceleration phase, the wings continue to translate apart at a constant speed and a constant 45º angle of attack. Plots of translational and angular velocities as functions of time are shown in Figure 6. Either a single clap upstroke or a single fling downstroke was simulated. This simplification was made because the influence of the wake produced by the previous stroke is small for Re < 10. The right wing was the mirror image of the left wing at all times during its motion. At the end of the upstroke and the beginning of the downstroke, the wings were placed 1/10 chord lengths apart unless otherwise noted. Validation of method and choice of kinematics The use of the immersed boundary to study clap and fling with solid wings has been previously validated through a convergence study and comparison to published experimental results (Miller and Peskin, 2005). A convergence study for a porous wing performing a fling at Re and leakiness values relevant to thrips was also carried out. Figure 7 shows lift and drag coefficients as functions of time (fraction of stroke) for a and a grid with a leakiness value of Le = The initial peaks correspond to the forces generated during the fling. Deviations in peak drag between the finer and coarser grids are due to differences in the effective width (and hence the gap) between the wings. A single immersed boundary point interacts with the fluid like a sphere with radius grid cells (Bringley and Peskin, 2008). As a result, the wings have an effective width of about 2.51 grid cells, and the width of the wings are refined as the grid is refined. For the grid the effect gap size is 1.6% of the chord, and for the grid the effective gap size is 5.8% of the chord. Once the wings are sufficiently far apart there is excellent agreement. 9
10 During flight, the distance between the wings at the beginning of the fling varies from about 1/4 to 1/10 of the chord length. For example, Figure 3 shows a gap of about 1/10 of the chord length. To determine the effect of the initial gap, the distances (measured from the middle of each wing) were varied from 1/10 to 1/2 of the chord length. The fact that the wings do not completely touch is fortuitous with regard to the immersed boundary method since the level of mesh refinement required scales inversely with the distance between the wings, and strictly speaking the method does not allow one to resolve the case of wings that are in direct contact. A method that can resolve wing contact for the case of solid rigid wings performing clap and fling has been described by Kolomenskiy et al. (2009, 2011), and their results show larger lift as the distance between the wings approaches zero. The choice of wing kinematics for the translation phase of the simulations is idealized. We were not able to quantitatively analyze angle of attack or stroke plane angle. Given the size of the thrips (~1mm) and the wingbeat frequency ( Hz), lighting and the plane of focus are significant challenges in terms of the amount of quantitative data that can be obtained. Another challenge is that the thrips are constantly changing their orientation and are susceptible to even the smallest breeze in the room. Without a clear measurement of flight direction and orientation, it is not possible to calculate stroke plane angle and angle of attack. The translation portion of the stroke was chosen so as to provide the maximum lift at a 45 degree angle of attack. Since this study is mainly focused on what happens during the initial part of the fling, the kinematics of the translational portion is not significant to the major findings. Single wing studies Single wing studies were performed by placing a porous wing in a moving fluid. Parabolic flow with maximum velocity U max was driven within a computational channel and upstream of the model wing by applying an external force, f ext, to the fluid proportional to the difference between the desired fluid velocity and the actual fluid velocity, as described in Miller et al. (2012). The difference between the actual and desired velocities was controlled with a stiffness parameter, k ext = 10k targ, such that the difference between the two velocities was always less than 0.1%. 10
11 Lift and drag coefficients as functions of time for fixed porous wings at a 45 angle of attack at Re = 10 are shown in Figure 8. The wing was fixed in place, and the background flow was ramped from zero to the characteristic velocity at t = 0.2. Maximum lift coefficients of 1.7 were achieved when the porosity was set zero, which is equivalent to a solid wing. Lift did not significantly drop for leakiness values up to Maximum drag coefficients of about 3.4 were also found when the porosity was set to zero and also did not drop considerably. Clap and fling Drag coefficients as a functions of time (fraction of stroke) for Re =10 are shown in Figure 9 for a single clap and a single fling. Note that Le = 0 corresponds to a solid wing. Four leakiness values ranging from 2.8 x 10-4 (least leaky) to 2.3 x 10-1 (most leaky) were examined. For the clap, the wings accelerate towards each during the first 20% of the stroke. The forces plateau until the end of the stroke as the wings begin to rotated and clap together. For the clap, the peaks in the drag coefficients at the end of the stroke correspond to the large forces required to clap the wings together at low Re (Figure 9A). For the fling, the initial large peak in the drag coefficients correspond to the large forces required to fling the wings apart in a viscous fluid (Figure 9B). The drag coefficients then plateau as the wings translate apart. The peak drag is lowered for the porous case as compared to the solid case, and this drag reducing effect increases rapidly for Le > 2.8 x Figure 10 shows the corresponding lift coefficients as a functions of time (fraction of stroke) for Re =10. The solid wing case and four leakiness values ranging from 2.8 x 10-4 (least leaky) to 2.3 x 10-1 (most leaky) were examined. During the upstroke, the peaks in the lift coefficients at the end of the stroke correspond to the lift augmentation effect generated by the clap (Figure 10A). During the downstroke, the initial large peak in the lift coefficients correspond to the lift augmenting effect of the fling generated during wing rotation (Figure 10B). This lift augmenting effect continues for some time during wing translation. In general, the average and peak lift values are lower for the porous case as compared to the solid case, and this effect increases rapidly for Le > 2.8 x
12 Figure 11 shows the drag coefficients as a functions of time (fraction of stroke) for Re = 4. The dimensionless porosities examined were the same as in Figure 9. During the clap, the wings initially accelerate towards each other, and then the forces plateau until the effect of the other wing is felt. Towards the end of the stroke, the wings begin to rotate and clap together. The large peak in the drag coefficients corresponds to force required to clap the wings together (11A). During the fling, the initial large peaks correspond to the forces required to rapidly fling the wings apart (Figure 11B). The drag coefficients then plateau as the wings translate apart. The peak drag is lowered for the porous cases as compared to the corresponding solid cases. The reduction in peak drag is perceived more for Re = 4 case as compared to Re = 10. Snapshots of the vorticity fields with velocity vectors are shown in Figure 12 for fling at Re = 10 for a solid wing (Le=0) and (B) a porous wing (Le = ). The vorticity and velocity fields are very similar with and without the addition of porosity. During the clap (not shown), a strong downward jet is generated between the wings that is responsible for some lift augmentation. During the fling, two large leading edge vortices are formed, and no trailing edge vortices are formed initially. This vortical asymmetry (strong leading edge vortices and weak trailing edge vortices) increases the circulation about the wings and augments the lift generated. The leading and trailing edge vortices are more diffuse for the porous case, but the basic flow patterns are the same. Effect of initial gap width between wings Figure 13 shows the lift and drag coefficients as a functions of time (fraction of stroke) during a fling for Re =10 and Le = The initial gap width between the centers of the wings was varied from 1/2 to 1/10 chord lengths. The large peaks in the force coefficients correspond to forces required to fling the wings apart. The peak drag forces continue to decrease as the initial gap between the wings is increased. Changes in peak lift force generated were small in comparison to the drag forces. Average and peak forces Peak drag coefficients generated during the fling are shown in Figure 14 as functions of Re. Peak drag coefficient during a fling as a function of Re. While decreasing Re increases the peak drag 12
13 coefficient, the inclusion of wing leakiness decreases this value for a particular Re. For leakiness values relevant to thrips (Le>0.15), the reduction in peak drag is on the order of 50% or more. The scaled porosity was set to 0 (for the case of the solid wing), 2 x 10-5, 2 x 10-3, 8 x 10-3, and 16 x Note that the peak drag coefficients for the solid wing cases are an order of magnitude larger than the forces required to translate a single wing. Decreasing Re increases the peak drag coefficient from about 90 at Re = 10 to about 160 at Re = 4. The inclusion of wing porosity decreases the peak drag coefficient for each Re. The reduction in drag is significant for λ > The average lift to drag ratios calculated during the fling downstroke are shown in Figure 15 as functions of the leakiness, Le. The Re was set to 10, 8, 6, and 4. Figure 15A shows that average lift over the entire downstroke is maximized for the case of a solid wing (Le = 0) for all Reynolds numbers. The average lift slowly decreases until ¹ and then lift begins to quickly decrease for ¹ Average drag over the entire downstroke is also highest for the case of the solid wing (Le=0) and increases with decreasing Re. The average drag also decreases with increasing leakiness. Lift over drag is maximized when Le is set to about 7.5 x 10-2 for all Re (Figure 15C). In these cases, the large peaks in drag generated as the wings are brought together are substantially reduced, but the lift coefficients are not significantly decreased. Discussion As a result of many experimental, computational, and theoretical studies, much is known about the aerodynamic mechanisms that generate lift in larger insects (Dickinson et al., 1999; Ellington, 1999; Liu et al., 1998; Ramamurtri and Sandberg, 2002; Birch and Dickinson, 2003; Sun and Wu, 2003, Usherwood, 2003). Insect aerodynamics are fundamentally different, however, at the lowest Re observed for insects due to the significant viscous effects that characterize these flows. Computational work shows that at lower Re, the leading and trailing edge vortices remain attached to single wings during translation, resulting in reduced lift coefficients and increased drag coefficients (Miller and Peskin, 2004). An aerodynamic feature observed in the flight of small insects that might compensate for this loss of lift is the predominant use of clap and fling (Ellington, 1999; Dudley, 2000). Numerical simulations 13
14 suggest that the clap and fling mechanism works best in terms of lift production at lower Re, so it is perhaps not surprising that most tiny insects use it (Miller and Peskin, 2005, 2009). What has been largely ignored in most analytical and numerical studies, however, are the very large drag forces that are generated during the clap and fling at lower Re. Results from a computational study by Miller and Peskin (2009) suggest that flexibility reduces the large drag forces generated during clap and fling while maintaining lift for a certain range of wing flexibilities. However, the drag forces generated for during clap and fling are still 3-5 times greater than during single wing translation for biologically realistic flexibilities. In this paper, the role of bristled wing structures are explored as another mechanism to reduce the drag required to clap together and fling apart wings at lower Re. Although it has been suggested that this wing design enhances flight performance at low Re, no experimental or computational study has supported this claim. In fact, Sunada et al. (2002) found no aerodynamic benefit when they compared lift and drag coefficients generated in physical models of fringed and solid wings at a Reynolds number of 10. When wing-wing interactions are considered, however, the highest lift over drag ratios are found for leakiness values of about ¹ This is primarily due to the reduced cost of rapidly flapping together and flinging apart porous wings in a viscous fluid. It has been shown previously in experiments with physical models that the movement of bristles near a boundary increases the flow between the bristles (Loudon et al., 1994). Similarly, the motion of two wings in close proximity can also increase the flow between the bristles and reduce the drag forces acting on each structure. The relevant range of dimensionless porosities or leakiness for thrips wings are in the range of These values are beyond the range where lift over drag is optimized, and these results suggest that the bristles are reducing the total force required to clap the wings together and fling them apart rather than to augment lift or lift over drag. Since the majority of the force acting on each wing during the end of the clap and the beginning of the fling are in opposite directions and cancel, it seems reasonable to reduce this force so as to decrease both the cost and peak force requirements for flight. Since both the lift and the drag forces are reduced proportionally, the changes in lift over drag are modest for biologically relevant ranges of porosity. 14
15 Possible rarefied effects At the scale of flow between each individual bristle, the continuum assumption for the fluid may introduce errors (Liu & Aono, 2009). The Knudsen number, Kn, gives a measure of the mean free path of a molecule over the characteristic length scale and is given by the equation The Journal of Experimental Biology ACCEPTED AUTHOR MANUSCRIPT º º É 7 2ÊÊ º where º is the Boltzmann constant, T is the thermodynamic temperature, p is the total pressure, Ê is the particle hard shell diameter, É is the mean free path, and is the characteristic length scale. In this case, is the length between bristles. Assuming that thrips bristles are 2 microns in diameter with a 20 micron gap (Ellington, 1980) and standard temperature and pressure, the Knudsen number is about Fluids are well described by continuum laws for Kn < For < Kn < 1, the no slip condition is violated, and these effects maybe be account for by increasing the amount of slip between the boundary and the fluid (Dyson et al., 2008). These effects may be accounted for by increasing the porosity or leakiness in the simulations. Intermittent parachuting as a dispersal mechanism This paper also presents the first analysis of the wing motion of a tiny insect with bristled wings. The clap and fling motion was observed for all individuals, although the wings did not come close enough to touch during the clap. We also obtained some of the first footage of a passive parachuting mechanism being employed by the smallest flying insects. Although we were not able to track the thrips for long periods of flight, the footage does suggest that thrips may alternate between passive and active flight modes as they disperse over distances of miles. The advantage here is that periods of passive flight could conserve significant amounts of energy during long distance dispersal. Longer distance tracking studies are needed, however, to verify this claim. Materials and Methods Video recordings of free-flying tiny insects 15
16 Thrips were collected in Chapel Hill, NC during late July to early August, 2009 from daylilies and gardenia flowers. The insects and flowers were brought to the lab and filmed within 8 hours of collection. A pipette tip was placed over the thrips to allow them to crawl onto the inner surface of the tip. The pipette was then suspended upside down in the field of view of the cameras so that the thrips could crawl down the tube and takeoff from the tip. Two high-speed cameras (Phantom v7.1, Vision Research, Wayne NJ) were each equipped with a 55mm micro-nikkor lens and 27.5mm extension tube and arranged perpendicularly for stereo filming (see Figure 16). Magnifying lenses were used to aim the beams from halogen bulbs directly into the camera apertures. The thrips were filmed at 4000 frames per second with a 15 to 30 microsecond exposure time. The cameras were calibrated by waving a wand of known dimension in the field of view. The calibration and analysis of the wing motion was performed using DLTdv5 software (Hedrick, 2008) with additional custom routines to perform the structure-from-motion wand-wave calibration. DLTdv5 is a digitizing environment written in MATLAB designed to acquire 3D coordinates from multiple video sources calibrated via a set of Direct Linear Transformation (DLT) coefficients. Recording of the thrips began when they were observed to prepare for flight by raising their wings. Computational method The immersed boundary method (Peskin, 2002) was used to solve the fully coupled fluidstructure interaction problem of two porous wings performing a clap and a fling. The full Navier- Stokes equations are solved on a fixed Cartesian grid, and the elastic boundary equations are discretized on a moving Lagrangian grid. The immersed boundary method provides an efficient way of and handling the interaction between these two grids. A description of the two-dimensional, non-porous immersed boundary method is given below. The equations of fluid motion are the incompressible Navier-Stokes equations: Ê,»,»,» º,» Ê,»,» 8 476,» 0, 9 16
17 where u(x,t) is the fluid velocity at Cartesian coordinate x and time t, p(x,t) is the pressure, and f(x,t) is the force per unit area acting on the fluid. The interaction equations between the fluid and the boundary are given by the following equations:,» ² º,» É ³ º,» ¹º 10 ³ º,» ³ ³ º,»,» É ³ º,» ¹, 11 The Journal of Experimental Biology ACCEPTED AUTHOR MANUSCRIPT where É is a two-dimensional delta function. Equation 10 applies force from the boundary to the fluid grid, and Equation 11 evaluates the local fluid velocity at the boundary. The boundary is then moved at the local fluid velocity, and this enforces the no-slip condition. The exact discretization used in these simulations has been described elsewhere (Peskin and McQueen, 1996; Miller and Peskin, 2009). The force f(q,t) is specific to the problem. Boundaries have been modeled in the immersed boundary framework that include active elastic forces due to muscle contraction (Peskin and McQueen, 1996), cohesive forces between boundaries or cells (Fogelson and Guy, 2008), and the action of dynein molecular motors (Dillon et al., 2007). In a simple case where boundary points are tethered to target points, the equation describing the force applied to the fluid by the boundary is ² º,» º ³ º,» ³ º,», 12 where k targ is a stiffness coefficient and Y(q,t) is the prescribed position of the target boundary. Acknowledgements This research was supported by Burroughs Wellcome Fund (CASI award ID # ) and NSF (DMS FRG ) to L.A.M. and NSF IOS to T.L.H. The authors would like to thank Charles Peskin for advice on the porous immersed boundary method and William Kier for insightful discussions on working with invertebrates. 17
18 References Alexander, M. R. (2003). Principles of Animal Locomotion. Princeton: Princeton University Press. Austin, A. and Dowton, M. (2000). Hymenoptera: Evolution, biodiversity and biological control. Csiro Publishing, Australia. Bennett, L. (1977). Clap and fling aerodynamics an experimental evaluation. J. Exp. Biol. 69, Birch, J. M. and Dickinson, M. H. (2003). The influence of wing-wake interactions on the production of aerodynamic forces in flapping flight. J. Exp. Biol. 206, Birch, J. M., Dickson, W. B. and Dickinson, M. H. (2004). Force production and flow structure of the leading edge vortex on flapping wings at high and low Reynolds numbers. J. Exp. Biol. 207, Bringley, T. T. and Peskin, C. S. (2008). Validation of a simple method for representing spheres and slender bodies in an immersed boundary method for Stokes flow on an unbounded domain. J. Comp. Phys. 227, Cheer, A. Y. L. and M. A. R. Koehl (1987) Paddles and rakes: Fluid flow through bristled appendages of small organisms. J. Theor. Biol. 129: Crespi, B. J., Carmean, D. A. and Chapman, T. W. (1997). Ecology and evolution of galling thrips and their allies. Annu. Rev. Entomol. 42, Dickinson, M. H. and Götz, K. G. (1993). Unsteady aerodynamic performance of model wings at low Reynolds numbers. J. Exp. Biol. 174,
19 Dickinson, M. H., Lehmann, F. O. and Sane, S. (1999). Wing rotation and the aerodynamic basis of insect flight. Science 284, Dillon, R. H., Fauci, L. J., Omoto, C., and Yang, X. (2007). Fluid dynamic models of flagellar and ciliary beating. Annals of the New York Academy of Sciences, 1101, Dudley, R. (2000). The biomechanics of insect flight: form, function, evolution. Princeton: Princeton University Press. Dyson, P. Ransing, R., Williams, P. J., Williams, R. (2008). Fluid properties at Nano/Meso Scale: A Numerical Treatment. John Wiley & Sons, Ltd. Ellington, C. P. (1980). Wing mechanics and take-off preparation of thrips (Thysanoptera). J. Exp. Biol. 85, Ellington, C. P. (1984). The aerodynamics of hovering insect flight. III. Kinematics. Phil. Trans. R. Soc. Lond. B 305, Ellington, C. P., Van Den Berg, C., Willmont, A. P. and Thomas, A. L. R. (1996). Leading edge vortices in insect flight. Nature 348, Ellington, C. P. (1999). The novel aerodynamics of insect flight: applications to micro-air vehicles. J. Exp. Biol. 202, Fogelson, A. L. and Guy, R. D. (2008). Immersed-Boundary-Motivated Models of Intravascular Platelet Aggregation, Comput. Methods Appl. Mech. Eng., 197, Hedrick, T. (2008). Software techniques for two- and three-dimensional kinematic measurements of biological and biomimetic systems. Bioinspir. Biomim. 3,
20 Hedrick, T. L., Cheng, B. and Deng, X. (2009). Wingbeat Time and the Scaling of Passive Rotational Damping in Flapping Flight. Science 324(5924), Horridge, G. A. (1956). The flight of very small insects. Nature 178, Jones, D. R. (2005). Plant viruses transmitted by thrips. European Journal of Plant Pathology 113, Kim, Y. and Peskin, C. S. (2006). 2-D parachute simulation by the immersed boundary method. SIAM J. Sci. Comput. 28, Kolomenskiy, D., Moffatt, H. K., Farge, M., and Schneider, K. (2009). Vorticity generation during the clap-fling-sweep of some hovering insects. Theoretical and Computational Fluid Dynamics 24, Kolomenskiy, D., Moffatt, H. K., Farge, M., and Schneider, K. (2011). Two- and threedimensional numerical simulations of the clap fling sweep of hovering insects. Journal of Fluids and Structures 27, Lewis, T. (1964). The weather and mass flights of Thysanoptera. Ann. Appl. Biol. 53, Lewis, T. (1965). The species, aerial density and sexual maturity of Thysanoptera caught in mass flights. Ann. Appl. Biol. 55, Lighthill, M. J. (1973). On the Weis-Fogh mechanism of lift generation. J. Fluid Mech. 60, Liu, H., Ellington, C. P., Kawachi, C., Van den Berg, C., and Willmott, A. P. (1998). A computational fluid dynamic study of hawkmoth hovering. J. Exp. Biol. 201,
21 Liu, H. and Aono, H. (2009). Size effects on insect hovering aerodynamics: an integrated computation study. Bioinsp. Biomm. 4, Loudon, C., Best, B., and Koehl, M. A. R. (1994). When does motion relative to neighboring surfaces alter the flow through arrays of hairs? J. Exp. Bio. 193, Maxworthy, T. (1979). Experiments on the Weis-Fogh mechanism of lift generation by insects in hovering flight. Part I. Dynamics of the fling. J. Fluid Mech. 93, Miller, L. A., Santhanakrishnan A., Jones S., Hamlet C., Mertens K. and Zhu, L. (2012). Reconfiguration and the reduction of vortex-induced vibrations in broad leaves. J. Exp. Biol. 215, Miller, L. A. and Peskin, C. S. (2004). When vortices stick: an aerodynamic transition in tiny insect flight. J. Exp. Biol. 207, Miller, L. A. and Peskin, C. S. (2005). A computational fluid dynamics of clap and fling in the smallest flying insects. J. Exp. Biol. 208, Miller, L. A. and Peskin, C. S. (2009). Flexible clap and fling in tiny insect flight. J. Exp. Biol. 212, Morse, J. G. and Hoddle, M. S. (2006). Invasion biology of thrips. Annu. Rev. Entomol. 51, Palmer, J. M., Reddy, D. V. R., Wightman, J. A. and Ranga Rao, G. V. (1990). New information on the thrips vectors of tomato spotted wilt virus in groundnut crops in India. International Arachis Newsletter 7, Peskin, C. S. (2002). The immersed boundary method. Acta Numerica, 11,
22 Peskin, C. S. and McQueen, D. M. (1996). Fluid dynamics of the heart and its valves. In H. G. Othmer, F. R. Adler, M. A. Lewis, and J. C. Dallon, editors, Case Studies in Mathematical Modeling: Ecology, Physiology, and Cell Biology. Prentice-Hall, New Jersey, 2nd edition. Ramamutri, R. and Sandberg, W. C. (2002). A three-dimensional computational study of the aerodynamic mechanisms of insect flight. J. Exp. Biol. 205, Spedding, G. R. and Maxworthy, T. (1986). The generation of circulation and lift in a rigid two-dimensional fling. J. Fluid Mech. 165, Srygley, R. B. and Thomas, A. L. R. (2002). Unconventional lift-generating mechanisms in free-flying butterflies. Nature 420, Stockie, J. M. (2009). Modelling and simulation of porous immersed boundaries. Comput. Struct. 87, Sun, M. and Wu, H. J. (2003). Aerodynamic force generation and power requirements in forward flight in a fruit fly with modeled wing motion. J. Exp. Biol. 206, Sun, M. and Xin, Y. (2003). Flow around two airfoils performing fling and subsequent translation and translation and subsequent flap. Acta Mech. Sin.19, Sunada, S., Takashima, H., Hattori, T., Yasuda, K. and Kawachi, K. (2002). Fluid dynamic characteristics of a bristled wing. J. Exp. Biol. 205, Terry, I. (2001). Thrips and weevils as dual, specialist pollinators of the Australian Cycad Macrozamia communis (Zamiaceae). Int. J. Plant Sci. 162, Terry, I. (2002). Thrips- the primeval pollinators? Seventh International conference on Thrips biology In Mound, L. and Marullo, R. (eds). Proceedings of Thrips, Plants, Tospoviruses: The Millenial Review. Reggio de Calabria. 22
23 Ullman, D.E., Meideros, R., Campbell, L.R., Whitfield, A.E., Sherwood, J. L. and German, T.L. (2002). Thrips as vectors of tospoviruses. Adv. Bot. Res. 36, Usherwood, J. (2003). More than a flitting tour of flapping flight: The biomechanics of insect flight. J. Exp. Biol. 206, Usherwood, J. R. and Ellington, C. P. (2002). The aerodynamics of revolving wings I. Model hawkmoth wings. J. Exp. Biol. 205, Van den Berg, C. and Ellington, C. P. (1997a). The three-dimensional leading-edge vortex of a hovering model hawkmoth. Phil. Trans. R. Soc. Lond. B 352, Van den Berg, C. and Ellington, C. P. (1997b). The vortex wake of a hovering model hawkmoth. Phil. Trans. R. Soc. Lond. B 352, Vogel, S. (1962). A possible role of the boundary layer in insect flight. Nature 193, Vogel, S. (1967). Flight in Drosophila. II. Variations in stroke parameters and wing contour. J. Exp. Biol. 46, Wang, Z. J. (2000). Two-dimensional mechanism for insect hovering. Phys. Rev. Lett. 85, Wang, Z. J. (2005). Dissecting insect flight. Annu. Rev. Fluid Mech. 37, Weis-Fogh, T. (1973). Quick estimates of flight fitness in hovering animals, including novel mechanisms for lift production. J. Exp. Biol. 59, Weis-Fogh, T. (1975). Unusual mechanisms for the generation of lift in flying animals. Scient. Am. 233,
24 Wu, J. H. and Sun, M. (2004). Unsteady aerodynamic forces of flapping flight. J. Exp. Biol. 207, The Journal of Experimental Biology ACCEPTED AUTHOR MANUSCRIPT 24
25 693 Table 1 Dimensionless parameters Symbol Value Chord length L 1 Domain length D 20 Reynolds number Re 4-10 Dimensionless porosity (Leakiness) Spatial step size dx The Journal of Experimental Biology ACCEPTED AUTHOR MANUSCRIPT 694 Time step size dt Maximum translational velocity U max 1 Maximum rotational velocity Ω max 9.0 Target stiffness k targ
26 Fig. 1. Diagram showing a two-dimensional simplified clap and fling from Miller and Peskin (2005). Top: The three-dimensional motion of the insect. Bottom: The corresponding twodimensional approximation. A) At the beginning of the upstroke the wings rotate together about the leading edges to perform the clap. B) At the beginning of the downstroke, the wings rotate apart about the trailing edges to perform the fling. Fig. 2. Images showing some of the variation in the number and spacing of bristles present in tiny insects wings. A) The bristled wing of an adult female parasitoid wasp Mymar sp. from New Zealand (courtesy S.E. Thorpe). B) The bristled wings of an adult thrips collected in Chapel Hill, NC. Fig. 3. Successive snapshots taken during clap and fling of a thrips in free takeoff flight. The images were recorded at 4000 frames per second. Note the characteristic bristled wing form. Top: At the end of the upstroke, the wings are raised vertically with respect to the body and come within about 1/6 1/10 chord lengths of each other. Bottom: At the beginning of the downstroke, the wings rotate about the trailing edge and the root performing the fling motion. Fig. 4. A passive parachuting maneuver from a 4000 fps video recording. Snapshots shown are taken every 1.25 ms about eight wingbeats after takeoff. (i) shows the wings clapped together at the beginning of the downstroke. Rather than completing the stroke, the thrips spreads its wings (ii-v) and passively parachutes downward until it leaves the field of view (vi-viii). Fig. 5. Wing tip speed of the left and right wing tips as a function of time for two representative thrips. Maximum wing tip velocities are on the order of 1.5 m/s. Flapping frequencies are on the order of 250 Hz, and the upstroke was significantly shorter than the downstroke. Fig. 6. Dimensionless translational and rotational velocities of the computational wings as functions of dimensionless time. (A) Kinematics for the clap with 100% overlap between translation and rotation at the end of the stroke. (B) Kinematics for the fling with 50% overlap between the beginning of rotation and the subsequent beginning of wing translation. 26
27 Fig. 7. Lift and drag coefficients as functions of time (fraction of stroke) for a and a grid with a leakiness value of Le = Deviations in peak drag are due to differences in the effective width (and hence the gap) between the wings. For the grid the effect gap size is 1.6 % of the chord, and for the grid the effective gap size is 5.8% of the chord. Once the wings are sufficient far apart there is excellent agreement. The Journal of Experimental Biology ACCEPTED AUTHOR MANUSCRIPT 27
28 Fig. 8. Force coefficients for a single wing in background flow at a 45 angle of attack at Re = 10 for varying values of the porosity. The wing is fixed in place, and the background flow is ramped from zero to the characteristic velocity at t = 0.2. A) Lift coefficients as functions of dimensionless time. Maximum lift is achieved with the porosity/leakiness is zero which is equivalent to a solid wing. B) Drag coefficients as functions of time. Fig. 9. Drag coefficients as a functions of time (fraction of stroke) for Re =10. A) Drag coefficients generated during the clap. Note that Le = 0 corresponds to a solid wing. The peaks in the drag coefficients at the end of the stroke correspond to the large forces required to clap the wings together at low Re. (b) Drag coefficients generated during fling. The initial large peak in the drag coefficients correspond to the large forces required to fling the wings apart in a viscous fluid. The peak drag force is reduced for the porous case as compared to the solid case. Fig. 10. Lift coefficients as a functions of time (fraction of stroke) for Re = 10. A) Lift coefficients generated during the clap. The large peaks in the lift coefficients correspond to the augmented lift generated when the wings are clapped together. B) Lift coefficients generated during the fling. The initial large peaks correspond to the augmented lift generated during the fling. Peak lift is reduced for the porous case, but not as much as the peak drag. Fig. 11. Drag coefficients as a functions of time (fraction of stroke) for Re = 4. A) Drag coefficients generated during the clap. The large peaks in the drag coefficients correspond to force required to clap the wings together. B) Drag coefficients generated during the fling. The initial large peaks correspond to the forces required to rapidly fling the wings apart. The peak drag forces are lowered for the porous cases as compared to the corresponding solid cases. Fig. 12. Snapshots of the vorticity fields with velocity vectors for the fling at Re = 10 for (A) a solid wing (Le=0) and (B) a porous wing (Le = ). During the fling, two large leading edge vortices are formed that enhance the lift acting on each wing. The larger leading edge vortices remain for some time as the wings translate apart. The vorticity and velocity fields for 28
29 the porous and solid wings are not substantially different, but the magnitude of vorticity is slightly decreased for the porous case. Fig. 13. Lift (a) and drag (b) coefficients as a functions of time (fraction of stroke) during a fling for Re =10, Le = , and varying initial gap widths between the wings as a fraction of the chord length, L. The large peaks in the force coefficients correspond to forces required to fling the wings apart. The peak drag forces continue to decrease as the initial gap between the wings is increased. Fig. 14. Peak drag coefficient during a fling as a function of Re. While decreasing Re increases the peak drag coefficient, the inclusion of wing leakiness decreases this value for a particular Re. For leakiness values relevant to thrips (Le>0.15), the reduction in peak drag is on the order of 50% or more. Fig. 15. Average lift and drag ratios during the entire downstroke as a function of the leakiness Le for Re = 10, 8, 6, and 4. (a) Average lift over the entire downstroke is maximized for the case of a solid wing (Le = 0). Average lift slowly decreases until ¹ and then lift begins to quickly decrease. (b) Average drag over the entire downstroke is highest for the case of the solid wing (Le=0) and decreases with increasing leakiness. (c) Lift over drag is maximized when Le is set to about 7.5 x For this case, the large peak in drag generated as the wings are brought together is substantially reduced, but the lift coefficients are not significantly decreased. Fig 16. Diagram of the experimental setup, not to scale. Thrips were filmed in stereo using two high speed cameras (Phantom v.7.1, Vision Research, Wayne NJ) operating at 4000 frames per second with a shutter duration of 15 to 30 µs. Lighting was provided by 300 W halogen bulbs positioned such that the beam of light was focused directly on the camera aperature using a magnifying lens. The cameras were focused on a pipette tip which held the thrips; recordings were collected as the thrips emerged from the pipette tip and flew away. 29
30

31

32 ournal of Experimental Biology ACCEPTED AUTHOR MANUSCR

33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54

55
Clap and fling mechanism with interacting porous wings in tiny insect flight
 14. Published by The Company of iologists Ltd (14) 17, 3898-399 doi:1.14/jeb.84897 RESERCH RTICLE Clap and fling mechanism with interacting porous wings in tiny insect flight rvind Santhanakrishnan 1,
14. Published by The Company of iologists Ltd (14) 17, 3898-399 doi:1.14/jeb.84897 RESERCH RTICLE Clap and fling mechanism with interacting porous wings in tiny insect flight rvind Santhanakrishnan 1,
Many of the smallest flying insects clap their wings together at the end of each upstroke
 DRAFT Miller, L. A. and Peskin, C. S. Title: Flexible clap and fling in tiny insect flight. Abstract Many of the smallest flying insects clap their wings together at the end of each upstroke and fling
DRAFT Miller, L. A. and Peskin, C. S. Title: Flexible clap and fling in tiny insect flight. Abstract Many of the smallest flying insects clap their wings together at the end of each upstroke and fling
Flexible clap and fling in tiny insect flight
 376 The Journal of Experimental Biology, 376-39 Published by The Company of Biologists 9 doi:./jeb.866 Flexible clap and fling in tiny insect flight Laura A. Miller* and Charles S. Peskin Department of
376 The Journal of Experimental Biology, 376-39 Published by The Company of Biologists 9 doi:./jeb.866 Flexible clap and fling in tiny insect flight Laura A. Miller* and Charles S. Peskin Department of
A COMPUTATIONAL FLUID DYNAMICS STUDY OF CLAP AND FLING IN THE SMALLEST INSECTS. Laura A. Miller* and Charles S. Peskin**
 A COMPUTATIONAL FLUID DYNAMICS STUDY OF CLAP AND FLING IN THE SMALLEST INSECTS Laura A. Miller* and Charles S. Peskin** *Department of Mathematics, University of Utah, 155 South 1400 East, Salt Lake City,
A COMPUTATIONAL FLUID DYNAMICS STUDY OF CLAP AND FLING IN THE SMALLEST INSECTS Laura A. Miller* and Charles S. Peskin** *Department of Mathematics, University of Utah, 155 South 1400 East, Salt Lake City,
A computational fluid dynamics of clap and fling in the smallest insects
 The Journal of Experimental Biology 8, 95- Published by The Company of Biologists 5 doi:.4/jeb.376 95 A computational fluid dynamics of clap and fling in the smallest insects Laura A. Miller, * and Charles
The Journal of Experimental Biology 8, 95- Published by The Company of Biologists 5 doi:.4/jeb.376 95 A computational fluid dynamics of clap and fling in the smallest insects Laura A. Miller, * and Charles
Unsteady aerodynamic forces of a flapping wing
 The Journal of Experimental Biology 7, 37-5 Published by The Company of Biologists 4 doi:.4/jeb.868 37 Unsteady aerodynamic forces of a flapping wing Jiang Hao Wu and Mao Sun* Institute of Fluid Mechanics,
The Journal of Experimental Biology 7, 37-5 Published by The Company of Biologists 4 doi:.4/jeb.868 37 Unsteady aerodynamic forces of a flapping wing Jiang Hao Wu and Mao Sun* Institute of Fluid Mechanics,
ABSTRACT. travels through the wing reducing the drag forces generated. When the wings are in
 ABSTRACT Tiny insects use a process called clap and fling to augment the lift forces generated during flight. The one disadvantage to using this method is the drag forces created when the wings fling apart
ABSTRACT Tiny insects use a process called clap and fling to augment the lift forces generated during flight. The one disadvantage to using this method is the drag forces created when the wings fling apart
When vortices stick: an aerodynamic transition in tiny insect flight
 The Journal of Experimental Biology 7, 7-88 Published by The Company of Biologists 4 doi:.4/jeb.8 7 When vortices stick: an aerodynamic transition in tiny insect flight Laura A. Miller* and Charles S.
The Journal of Experimental Biology 7, 7-88 Published by The Company of Biologists 4 doi:.4/jeb.8 7 When vortices stick: an aerodynamic transition in tiny insect flight Laura A. Miller* and Charles S.
The wings and the body shape of Manduca sexta and Agrius convolvuli are compared in
 1 Wing and body shape of Manduca sexta and Agrius convolvuli The wings and the body shape of Manduca sexta and Agrius convolvuli are compared in terms of the aspect ratio of forewing AR fw (wing length
1 Wing and body shape of Manduca sexta and Agrius convolvuli The wings and the body shape of Manduca sexta and Agrius convolvuli are compared in terms of the aspect ratio of forewing AR fw (wing length
Computational Analysis of Hovering Hummingbird Flight
 Computational Analysis of Hovering Hummingbird Flight Zongxian Liang 1 and Haibo Dong 2 Department of Mechanical & Materials Engineering, Wright State University, Dayton, OH 45435 Mingjun Wei 3 Department
Computational Analysis of Hovering Hummingbird Flight Zongxian Liang 1 and Haibo Dong 2 Department of Mechanical & Materials Engineering, Wright State University, Dayton, OH 45435 Mingjun Wei 3 Department
Computational Analysis of Hovering Hummingbird Flight
 48th AIAA Aerospace Sciences Meeting Including the New Horizons Forum and Aerospace Exposition 4-7 January 2010, Orlando, Florida AIAA 2010-555 Computational Analysis of Hovering Hummingbird Flight Zongxian
48th AIAA Aerospace Sciences Meeting Including the New Horizons Forum and Aerospace Exposition 4-7 January 2010, Orlando, Florida AIAA 2010-555 Computational Analysis of Hovering Hummingbird Flight Zongxian
Two-Dimensional Aerodynamic Models of Insect Flight for Robotic Flapping Wing Mechanisms of Maximum Efficiency
 Journal of Bionic Engineering 5 (2008) 1 11 Two-Dimensional Aerodynamic Models of Insect Flight for Robotic Flapping Wing Mechanisms of Maximum Efficiency Thien-Tong Nguyen 1, Doyoung Byun 2 1. Department
Journal of Bionic Engineering 5 (2008) 1 11 Two-Dimensional Aerodynamic Models of Insect Flight for Robotic Flapping Wing Mechanisms of Maximum Efficiency Thien-Tong Nguyen 1, Doyoung Byun 2 1. Department
A flow control mechanism in wing flapping with stroke asymmetry during insect forward flight
 Acta Mech Sinica (2005) 21, 218 227 DOI 10.1007/s10409-005-0032-z RESEARCH PAPER Yongliang Yu Binggang Tong A flow control mechanism in wing flapping with stroke asymmetry during insect forward flight
Acta Mech Sinica (2005) 21, 218 227 DOI 10.1007/s10409-005-0032-z RESEARCH PAPER Yongliang Yu Binggang Tong A flow control mechanism in wing flapping with stroke asymmetry during insect forward flight
Wing Kinematics in a Hovering Dronefly Minimize Power Expenditure
 Wing Kinematics in a Hovering Dronefly Minimize Power Expenditure Authors: J. H. Wu, and M. Sun Source: Journal of Insect Science, 14(159) : 1-8 Published By: Entomological Society of America URL: https://doi.org/10.1093/jisesa/ieu021
Wing Kinematics in a Hovering Dronefly Minimize Power Expenditure Authors: J. H. Wu, and M. Sun Source: Journal of Insect Science, 14(159) : 1-8 Published By: Entomological Society of America URL: https://doi.org/10.1093/jisesa/ieu021
NUMERICAL SIMULATION OF SELF-PROPELLED FLYING OF A THREE-DIMENSIONAL BIRD WITH FLAPPING WINGS
 NUMERICAL SIMULATION OF SELF-PROPELLED FLYING OF A THREE-DIMENSIONAL BIRD WITH FLAPPING WINGS WU Chui-Jie, ZHU Lin-Lin State Key Laboratory of Structural Analysis for Industrial Equipment, School of Aeronautics
NUMERICAL SIMULATION OF SELF-PROPELLED FLYING OF A THREE-DIMENSIONAL BIRD WITH FLAPPING WINGS WU Chui-Jie, ZHU Lin-Lin State Key Laboratory of Structural Analysis for Industrial Equipment, School of Aeronautics
RESEARCH ARTICLE Aerodynamic effects of corrugation in flapping insect wings in hovering flight
 3 The Journal of Experimental iology, 3-. Published by The Company of iologists Ltd doi:./jeb.6375 RESERCH RTIE erodynamic effects of corrugation in flapping insect wings in hovering flight Xue Guang Meng*,
3 The Journal of Experimental iology, 3-. Published by The Company of iologists Ltd doi:./jeb.6375 RESERCH RTIE erodynamic effects of corrugation in flapping insect wings in hovering flight Xue Guang Meng*,
SENSITIVITY ANALYSIS OF THE FACTORS AFFECTING FORCE GENERATION BY WING FLAPPING MOTION
 Proceedings of the ASME 2013 International Mechanical Engineering Congress and Exposition IMECE2013 November 15-21, 2013, San Diego, California, USA IMECE2013-65472 SENSITIVITY ANALYSIS OF THE FACTORS
Proceedings of the ASME 2013 International Mechanical Engineering Congress and Exposition IMECE2013 November 15-21, 2013, San Diego, California, USA IMECE2013-65472 SENSITIVITY ANALYSIS OF THE FACTORS
FUNDAMENTAL ANALYSIS OF THREE-DIMENSIONAL NEAR FLING
 J. exp. Biol. 83, 27 248 (993) Printed in Great Britain The Company of Biologists Limited 993 27 FUNDAMENTAL ANALYSIS OF THREE-DIMENSIONAL NEAR FLING SHIGERU SUNADA Exploratory Research for Advanced Technology,
J. exp. Biol. 83, 27 248 (993) Printed in Great Britain The Company of Biologists Limited 993 27 FUNDAMENTAL ANALYSIS OF THREE-DIMENSIONAL NEAR FLING SHIGERU SUNADA Exploratory Research for Advanced Technology,
Near-Hover Dynamics and Attitude Stabilization of an Insect Model
 21 American Control Conference Marriott Waterfront, Baltimore, MD, USA June 3-July 2, 21 WeA1.4 Near-Hover Dynamics and Attitude Stabilization of an Insect Model B. Cheng and X. Deng Abstract In this paper,
21 American Control Conference Marriott Waterfront, Baltimore, MD, USA June 3-July 2, 21 WeA1.4 Near-Hover Dynamics and Attitude Stabilization of an Insect Model B. Cheng and X. Deng Abstract In this paper,
Smart wing rotation and ingenious leading edge vortex control modulate the unconventional forces during insects flights
 Smart wing rotation and ingenious leading edge vortex control modulate the unconventional forces during insects flights Yang Xiang, Haotian Hang, and Hong Liu J.C.Wu Center for Aerodynamics, School of
Smart wing rotation and ingenious leading edge vortex control modulate the unconventional forces during insects flights Yang Xiang, Haotian Hang, and Hong Liu J.C.Wu Center for Aerodynamics, School of
Lift and power requirements of hovering flight in Drosophila virilis
 The Journal of Experimental Biology 5, 37 () Printed in Great Britain The ompany of Biologists Limited JEB6 3 Lift and power requirements of hovering flight in Drosophila virilis Mao Sun* and Jian Tang
The Journal of Experimental Biology 5, 37 () Printed in Great Britain The ompany of Biologists Limited JEB6 3 Lift and power requirements of hovering flight in Drosophila virilis Mao Sun* and Jian Tang
Numerical Analysis of Unsteady Viscous Flow through a Weis-Fogh-Type Water Turbine
 International Conference on Emerging Trends in Computer and Image Processing (ICETCIP'014) Dec. 15-16, 014 Pattaya (Thailand) Numerical Analysis of Unsteady Viscous Flow through a Weis-Fogh-Type Water
International Conference on Emerging Trends in Computer and Image Processing (ICETCIP'014) Dec. 15-16, 014 Pattaya (Thailand) Numerical Analysis of Unsteady Viscous Flow through a Weis-Fogh-Type Water
Lift Enhancement by Dynamically Changing Wingspan. in Forward Flapping Flight (09/10/2013)
") Lift Enhancement by Dynamically Changing Wingspan in Forward Flapping Flight Shizhao Wang 1, Xing Zhang 1, Guowei He 1a), ianshu Liu 2,1 (09/10/2013) 1 he State Key Laboratory of Nonlinear Mechanics, Institute
Lift Enhancement by Dynamically Changing Wingspan in Forward Flapping Flight Shizhao Wang 1, Xing Zhang 1, Guowei He 1a), ianshu Liu 2,1 (09/10/2013) 1 he State Key Laboratory of Nonlinear Mechanics, Institute
THE CONTROL OF FLIGHT FORCE BY A FLAPPING WING: LIFT AND DRAG PRODUCTION
 The Journal of Experimental Biology 24, 2672626 (2) Printed in Great Britain The Company of Biologists Limited 2 JEB34 267 THE CONTROL OF FLIGHT FORCE BY A FLAPPING WING: LIFT AND DRAG PRODUCTION SANJAY
The Journal of Experimental Biology 24, 2672626 (2) Printed in Great Britain The Company of Biologists Limited 2 JEB34 267 THE CONTROL OF FLIGHT FORCE BY A FLAPPING WING: LIFT AND DRAG PRODUCTION SANJAY
Vortex Dynamics in Near Wake of a Hovering Hawkmoth
 46th AIAA Aerospace Sciences Meeting and Exhibit 7-1 January 28, Reno, Nevada AIAA 28-583 Vortex Dynamics in Near Wake of a Hovering Hawkmoth Hikaru Aono 1 Graduate School of Science and Technology, Chiba
46th AIAA Aerospace Sciences Meeting and Exhibit 7-1 January 28, Reno, Nevada AIAA 28-583 Vortex Dynamics in Near Wake of a Hovering Hawkmoth Hikaru Aono 1 Graduate School of Science and Technology, Chiba
Analysis of a Hinge-Connected Flapping Plate with an Implemented Torsional Spring Model
 Analysis of a Hinge-Connected Flapping Plate with an Implemented Torsional Spring Model Zach Gaston 1, Hui Wan 2 and Haibo Dong 3 Department of Mechanical & Materials Engineering, Wright State University,
Analysis of a Hinge-Connected Flapping Plate with an Implemented Torsional Spring Model Zach Gaston 1, Hui Wan 2 and Haibo Dong 3 Department of Mechanical & Materials Engineering, Wright State University,
I nsect flight has long been considered a paradox, defeating the expectations of steady state aerodynamics. We
 OPEN SUBJECT AREAS: BIOMECHANICS FLUID DYNAMICS Received 15 May 2013 Accepted 5 November 2013 Published 20 November 2013 Correspondence and requests for materials should be addressed to L.C.J. (Christoffer.
OPEN SUBJECT AREAS: BIOMECHANICS FLUID DYNAMICS Received 15 May 2013 Accepted 5 November 2013 Published 20 November 2013 Correspondence and requests for materials should be addressed to L.C.J. (Christoffer.
The Interaction of Wings in Different Flight Modes of a Dragonfly
 The Interaction of Wings in Different Flight Modes of a Dragonfly Csaba Hefler 1, Huihe Qiu 1,*, Wei Shyy 1 1: Department of Mechanical and Aerospace Engineering, The Hong Kong University of Science and
The Interaction of Wings in Different Flight Modes of a Dragonfly Csaba Hefler 1, Huihe Qiu 1,*, Wei Shyy 1 1: Department of Mechanical and Aerospace Engineering, The Hong Kong University of Science and
Effects of unsteady deformation of flapping wing on its aerodynamic forces
 Appl. Math. Mech. -Engl. Ed., 2008, 29(6):731 743 DOI 10.1007/s10483-008-0605-9 c Editorial Committee of Appl. Math. Mech. and Springer-Verlag 2008 Applied Mathematics and Mechanics (English Edition) Effects
Appl. Math. Mech. -Engl. Ed., 2008, 29(6):731 743 DOI 10.1007/s10483-008-0605-9 c Editorial Committee of Appl. Math. Mech. and Springer-Verlag 2008 Applied Mathematics and Mechanics (English Edition) Effects
The aerodynamic effects of wing rotation and a revised quasi-steady model of flapping flight
 The Journal of Experimental Biology 5, 87 96 () Printed in Great Britain The Company of Biologists Limited JEB396 87 The aerodynamic effects of wing rotation and a revised quasi-steady model of flapping
The Journal of Experimental Biology 5, 87 96 () Printed in Great Britain The Company of Biologists Limited JEB396 87 The aerodynamic effects of wing rotation and a revised quasi-steady model of flapping
We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists. International authors and editors
 We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists 3,800 116,000 120M Open access books available International authors and editors Downloads Our
We are IntechOpen, the world s leading publisher of Open Access books Built by scientists, for scientists 3,800 116,000 120M Open access books available International authors and editors Downloads Our
ABSTRACT PARAMETRIC INVESTIGATIONS INTO FLUID-STRUCTURE INTERACTIONS IN HOVERING FLAPPING FLIGHT. Jesse Maxwell, 2013
 ABSTRACT Title of Thesis: PARAMETRIC INVESTIGATIONS INTO FLUID-STRUCTURE INTERACTIONS IN HOVERING FLAPPING FLIGHT Jesse Maxwell, 2013 Thesis Directed By: Professor Balakumar Balachandran Department of
ABSTRACT Title of Thesis: PARAMETRIC INVESTIGATIONS INTO FLUID-STRUCTURE INTERACTIONS IN HOVERING FLAPPING FLIGHT Jesse Maxwell, 2013 Thesis Directed By: Professor Balakumar Balachandran Department of
Dynamic flight stability of a hovering model insect: lateral motion
 Acta Mech Sin (010) 6:175 190 DOI 10.1007/s10409-009-00-1 RESEARCH PAPER Dynamic flight stability of a hovering model insect: lateral motion Yanlai Zhang Mao Sun Received: 18 May 009 / Revised: 5 August
Acta Mech Sin (010) 6:175 190 DOI 10.1007/s10409-009-00-1 RESEARCH PAPER Dynamic flight stability of a hovering model insect: lateral motion Yanlai Zhang Mao Sun Received: 18 May 009 / Revised: 5 August
THE MECHANICS OF FLIGHT IN THE HAWKMOTH MANDUCA SEXTA
 The Journal of Experimental Biology 00, 73 745 (1997) Printed in Great Britain The Company of Biologists Limited 1997 JEB0994 73 THE MECHANICS OF FLIGHT IN THE HAWKMOTH MANDUCA SEXTA II. AERODYNAMIC CONSEQUENCES
The Journal of Experimental Biology 00, 73 745 (1997) Printed in Great Britain The Company of Biologists Limited 1997 JEB0994 73 THE MECHANICS OF FLIGHT IN THE HAWKMOTH MANDUCA SEXTA II. AERODYNAMIC CONSEQUENCES
DISSECTING INSECT FLIGHT
 Annu. Rev. Fluid Mech. 2005. 37:183 210 doi: 10.1146/annurev.fluid.36.050802.121940 Copyright c 2005 by Annual Reviews. All rights reserved DISSECTING INSECT FLIGHT Z. Jane Wang Theoretical and Applied
Annu. Rev. Fluid Mech. 2005. 37:183 210 doi: 10.1146/annurev.fluid.36.050802.121940 Copyright c 2005 by Annual Reviews. All rights reserved DISSECTING INSECT FLIGHT Z. Jane Wang Theoretical and Applied
Modeling of Instantaneous Passive Pitch of Flexible Flapping Wings
 Fluid Dynamics and Co-located Conferences June 24-27, 2013, San Diego, CA 43rd Fluid Dynamics Conference AIAA 2013-2469 Modeling of Instantaneous Passive Pitch of Flexible Flapping Wings Chang-kwon Kang
Fluid Dynamics and Co-located Conferences June 24-27, 2013, San Diego, CA 43rd Fluid Dynamics Conference AIAA 2013-2469 Modeling of Instantaneous Passive Pitch of Flexible Flapping Wings Chang-kwon Kang
SIMULATION OF GAS FLOW OVER MICRO-SCALE AIRFOILS USING A HYBRID CONTINUUM-PARTICLE APPROACH
 33rd AIAA Fluid Dynamics Conference and Exhibit 3-6 June 3, Orlando, Florida AIAA 3-44 33 rd AIAA Fluid Dynamics Conference and Exhibit / Orlando, Florida / 3-6 Jun 3 SIMULATION OF GAS FLOW OVER MICRO-SCALE
33rd AIAA Fluid Dynamics Conference and Exhibit 3-6 June 3, Orlando, Florida AIAA 3-44 33 rd AIAA Fluid Dynamics Conference and Exhibit / Orlando, Florida / 3-6 Jun 3 SIMULATION OF GAS FLOW OVER MICRO-SCALE
Numerical and experimental investigation of the role of flexibility in flapping wing flight
 36th AIAA Fluid Dynamics Conference and Exhibit 5-8 June 26, San Francisco, California AIAA 26-3211 Numerical and experimental investigation ohe role of flexibility in flapping wing flight Jonathan Toomey
36th AIAA Fluid Dynamics Conference and Exhibit 5-8 June 26, San Francisco, California AIAA 26-3211 Numerical and experimental investigation ohe role of flexibility in flapping wing flight Jonathan Toomey
A. Bottaro, D. Venkataraman & F. Negrello Università di Genova, Italy
 A. Bottaro, D. Venkataraman & F. Negrello Università di Genova, Italy A. Bottaro, D. Venkataraman & F. Negrello & G. Tadmor Università di Genova, Italy Focus on passive actuators, what works, why it does,
A. Bottaro, D. Venkataraman & F. Negrello Università di Genova, Italy A. Bottaro, D. Venkataraman & F. Negrello & G. Tadmor Università di Genova, Italy Focus on passive actuators, what works, why it does,
PIV and force measurements on the flapping-wing MAV DelFly II
 Master of Science Thesis PIV and force measurements on the flapping-wing MAV DelFly II An aerodynamic and aeroelastic investigation into vortex development M.A. Groen 2 December 21 Ad Faculty of Aerospace
Master of Science Thesis PIV and force measurements on the flapping-wing MAV DelFly II An aerodynamic and aeroelastic investigation into vortex development M.A. Groen 2 December 21 Ad Faculty of Aerospace
Dynamic pitching of an elastic rectangular wing in hovering motion
 Under consideration for publication in J. Fluid Mech. Dynamic pitching of an elastic rectangular wing in hovering motion Hu Dai, Haoxiang Luo, and James F. Doyle 2 Department of Mechanical Engineering,
Under consideration for publication in J. Fluid Mech. Dynamic pitching of an elastic rectangular wing in hovering motion Hu Dai, Haoxiang Luo, and James F. Doyle 2 Department of Mechanical Engineering,
Dynamic flight stability of a hovering bumblebee
 The Journal of Experimental iology 28, 447-459 Published by The Company of iologists 25 doi:1.1242/jeb.147 447 Dynamic flight stability of a hovering bumblebee Mao Sun* and Yan Xiong Institute of Fluid
The Journal of Experimental iology 28, 447-459 Published by The Company of iologists 25 doi:1.1242/jeb.147 447 Dynamic flight stability of a hovering bumblebee Mao Sun* and Yan Xiong Institute of Fluid
THE INERTIAL CAUSE OF WING ROTATION IN DIPTERA
 J. exp. Biol. 140, 161-169 (1988) 161 Printed in Great Britain The Company of Biologists Limited 1988 THE INERTIAL CAUSE OF WING ROTATION IN DIPTERA BY A. ROLAND ENNOS* Department of Biological Sciences,
J. exp. Biol. 140, 161-169 (1988) 161 Printed in Great Britain The Company of Biologists Limited 1988 THE INERTIAL CAUSE OF WING ROTATION IN DIPTERA BY A. ROLAND ENNOS* Department of Biological Sciences,
Force production and flow structure of the leading edge vortex on flapping wings at high and low Reynolds numbers
 The Journal of Experimental iology 27, 163-172 Published by The Company of iologists 24 doi:1.1242/jeb.848 163 Force production and flow structure of the leading edge vortex on flapping wings at high and
The Journal of Experimental iology 27, 163-172 Published by The Company of iologists 24 doi:1.1242/jeb.848 163 Force production and flow structure of the leading edge vortex on flapping wings at high and
International Journal of Micro Air Vehicles
 Reliable Force Predictions for a Flapping-wing Micro Air Vehicle: A Vortex-lift Approach W. Thielicke, A.B. Kesel and E.J. Stamhuis Reprinted from International Journal of Micro Air Vehicles Volume 3 Number
Reliable Force Predictions for a Flapping-wing Micro Air Vehicle: A Vortex-lift Approach W. Thielicke, A.B. Kesel and E.J. Stamhuis Reprinted from International Journal of Micro Air Vehicles Volume 3 Number
APS Flapping flight from flexible wings : tuning of wing stiffness for flight? Tom Daniel, Stacey Combes,, & Sanjay Sane
 APS 2004 Flapping flight from flexible wings : tuning of wing stiffness for flight? Tom Daniel, Stacey Combes,, & Sanjay Sane CNS SENSORY INPUT MOTOR OUTPUT FLIGHT MUSCLES FORCE, STRAIN WING HINGE How
APS 2004 Flapping flight from flexible wings : tuning of wing stiffness for flight? Tom Daniel, Stacey Combes,, & Sanjay Sane CNS SENSORY INPUT MOTOR OUTPUT FLIGHT MUSCLES FORCE, STRAIN WING HINGE How
Coupling of the wings and the body dynamics enhances damselfly maneuverability
 Coupling of the wings and the body dynamics enhances damselfly maneuverability Samane Zeyghami, Haibo Dong Mechanical and Aerospace Engineering, University of Virginia, Charlottesville, VA, 22904 In flapping
Coupling of the wings and the body dynamics enhances damselfly maneuverability Samane Zeyghami, Haibo Dong Mechanical and Aerospace Engineering, University of Virginia, Charlottesville, VA, 22904 In flapping
Vortex wake and energy transitions of an oscillating cylinder at low Reynolds number
 ANZIAM J. 46 (E) ppc181 C195, 2005 C181 Vortex wake and energy transitions of an oscillating cylinder at low Reynolds number B. Stewart J. Leontini K. Hourigan M. C. Thompson (Received 25 October 2004,
ANZIAM J. 46 (E) ppc181 C195, 2005 C181 Vortex wake and energy transitions of an oscillating cylinder at low Reynolds number B. Stewart J. Leontini K. Hourigan M. C. Thompson (Received 25 October 2004,
Fig. 1. Bending-Torsion Foil Flutter
 27 TH INTERNATIONAL CONGRESS OF THE AERONAUTICAL SCIENCES EXTRACTING POWER IN JET STREAMS: PUSHING THE PERFORMANCE OF FLAPPING WING TECHNOLOGY M.F. Platzer*, M.A. Ashraf**, J. Young**, and J.C.S. Lai**
27 TH INTERNATIONAL CONGRESS OF THE AERONAUTICAL SCIENCES EXTRACTING POWER IN JET STREAMS: PUSHING THE PERFORMANCE OF FLAPPING WING TECHNOLOGY M.F. Platzer*, M.A. Ashraf**, J. Young**, and J.C.S. Lai**
Rotational lift: something different or more of the same?
 University of Southern Maine USM Digital Commons iological Sciences College of Science, Technology, and Health 12-22 Rotational lift: something different or more of the same? Jeffrey. Walker University
University of Southern Maine USM Digital Commons iological Sciences College of Science, Technology, and Health 12-22 Rotational lift: something different or more of the same? Jeffrey. Walker University
Enclosure enhancement of flight performance
 THEORETICAL & APPLIED MECHANICS LETTERS, 23 (21) Enclosure enhancement of flight performance Mehdi Ghommem, 1, a) Daniel Garcia, 2 Victor M. Calo 3 1) Center for Numerical Porous Media (NumPor), King Abdullah
THEORETICAL & APPLIED MECHANICS LETTERS, 23 (21) Enclosure enhancement of flight performance Mehdi Ghommem, 1, a) Daniel Garcia, 2 Victor M. Calo 3 1) Center for Numerical Porous Media (NumPor), King Abdullah
Introduction to immersed boundary method
 Introduction to immersed boundary method Ming-Chih Lai mclai@math.nctu.edu.tw Department of Applied Mathematics Center of Mathematical Modeling and Scientific Computing National Chiao Tung University 1001,
Introduction to immersed boundary method Ming-Chih Lai mclai@math.nctu.edu.tw Department of Applied Mathematics Center of Mathematical Modeling and Scientific Computing National Chiao Tung University 1001,
(This is a sample cover image for this issue. The actual cover is not yet available at this time.)
") (This is a sample cover image for this issue. The actual cover is not yet available at this time.) This is an open access article which appeared in a ournal published by Elsevier. This article is free
(This is a sample cover image for this issue. The actual cover is not yet available at this time.) This is an open access article which appeared in a ournal published by Elsevier. This article is free
Numerical Investigation of Thermal Performance in Cross Flow Around Square Array of Circular Cylinders
 Numerical Investigation of Thermal Performance in Cross Flow Around Square Array of Circular Cylinders A. Jugal M. Panchal, B. A M Lakdawala 2 A. M. Tech student, Mechanical Engineering Department, Institute
Numerical Investigation of Thermal Performance in Cross Flow Around Square Array of Circular Cylinders A. Jugal M. Panchal, B. A M Lakdawala 2 A. M. Tech student, Mechanical Engineering Department, Institute
Lab 6: Lift and Bernoulli
 Lab 6: Lift and Bernoulli Bio427 Biomechanics In this lab, we explore the flows and fluid dynamic forces on wings and other structures. We deploy force measurement techniques, wind meters, and a variety
Lab 6: Lift and Bernoulli Bio427 Biomechanics In this lab, we explore the flows and fluid dynamic forces on wings and other structures. We deploy force measurement techniques, wind meters, and a variety
THE ADVANTAGES OF AN UNSTEADY PANEL METHOD IN MODELLING THE AERODYNAMIC FORCES ON RIGID FLAPPING WINGS
 The Journal of Experimental Biology 199, 1073 1083 (1996) Printed in Great Britain The Company of Biologists Limited 1996 JEB9635 1073 THE ADVANTAGES OF AN UNSTEADY PANEL METHOD IN MODELLING THE AERODYNAMIC
The Journal of Experimental Biology 199, 1073 1083 (1996) Printed in Great Britain The Company of Biologists Limited 1996 JEB9635 1073 THE ADVANTAGES OF AN UNSTEADY PANEL METHOD IN MODELLING THE AERODYNAMIC
On the Aerodynamic Performance of Dragonfly Wing Section in Gliding Mode
 Advances in Aerospace Science and Applications. ISSN 2277-3223 Volume 3, Number 3 (2013), pp. 227-234 Research India Publications http://www.ripublication.com/aasa.htm On the Aerodynamic Performance of
Advances in Aerospace Science and Applications. ISSN 2277-3223 Volume 3, Number 3 (2013), pp. 227-234 Research India Publications http://www.ripublication.com/aasa.htm On the Aerodynamic Performance of
Detailed Outline, M E 521: Foundations of Fluid Mechanics I
 Detailed Outline, M E 521: Foundations of Fluid Mechanics I I. Introduction and Review A. Notation 1. Vectors 2. Second-order tensors 3. Volume vs. velocity 4. Del operator B. Chapter 1: Review of Basic
Detailed Outline, M E 521: Foundations of Fluid Mechanics I I. Introduction and Review A. Notation 1. Vectors 2. Second-order tensors 3. Volume vs. velocity 4. Del operator B. Chapter 1: Review of Basic
Title: Aerodynamics characteristics of butterfly flight through measurement of threedimensional unsteady velocity field using TR-PIV system
 Title: Aerodynamics characteristics of butterfly flight through measurement of threedimensional unsteady velocity field using TR-PIV system REF: AOARD-09-4102 Contract No. FA23860914102 PI: Debopam Das
Title: Aerodynamics characteristics of butterfly flight through measurement of threedimensional unsteady velocity field using TR-PIV system REF: AOARD-09-4102 Contract No. FA23860914102 PI: Debopam Das
Jun 22-25, 2009/San Antonio, TX
 19th AIAA Computational Fluid Dynamics 22-25 June 2009, San Antonio, Texas AIAA 2009-4273 AIAA 2009 4273 An Assessment of Dual-Time Stepping, Time Spectral and Artificial Compressibility based Numerical
19th AIAA Computational Fluid Dynamics 22-25 June 2009, San Antonio, Texas AIAA 2009-4273 AIAA 2009 4273 An Assessment of Dual-Time Stepping, Time Spectral and Artificial Compressibility based Numerical
Spontaneous Symmetry Breaking of Hinged Flapping Filament Generates Lift
 Spontaneous Symmetry Breaking of Hinged Flapping Filament Generates Lift Shervin Bagheri (KTH, Stockholm) Andrea Mazzino & Alessandro Bottaro (Genova University, Italy) Fluid & Elasticity 2012 La Jolla,
Spontaneous Symmetry Breaking of Hinged Flapping Filament Generates Lift Shervin Bagheri (KTH, Stockholm) Andrea Mazzino & Alessandro Bottaro (Genova University, Italy) Fluid & Elasticity 2012 La Jolla,
A wing characterization method for flapping-wing robotic insects
 213 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS) November 3-7, 213. Tokyo, Japan A wing characterization method for flapping-wing robotic insects Alexis Lussier Desbiens*,
213 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS) November 3-7, 213. Tokyo, Japan A wing characterization method for flapping-wing robotic insects Alexis Lussier Desbiens*,
Active Control of Separated Cascade Flow
 Chapter 5 Active Control of Separated Cascade Flow In this chapter, the possibility of active control using a synthetic jet applied to an unconventional axial stator-rotor arrangement is investigated.
Chapter 5 Active Control of Separated Cascade Flow In this chapter, the possibility of active control using a synthetic jet applied to an unconventional axial stator-rotor arrangement is investigated.
Fluid Dynamics: Theory, Computation, and Numerical Simulation Second Edition
 Fluid Dynamics: Theory, Computation, and Numerical Simulation Second Edition C. Pozrikidis m Springer Contents Preface v 1 Introduction to Kinematics 1 1.1 Fluids and solids 1 1.2 Fluid parcels and flow
Fluid Dynamics: Theory, Computation, and Numerical Simulation Second Edition C. Pozrikidis m Springer Contents Preface v 1 Introduction to Kinematics 1 1.1 Fluids and solids 1 1.2 Fluid parcels and flow
Energy-minimizing kinematics in hovering insect flight
 J. Fluid Mech. (7), vol. 58, pp. 153 168. c 7 Cambridge University Press doi:1.117/s11769 Printed in the United Kingdom 153 Energy-minimizing kinematics in hovering insect flight GORDON J. BERMAN 1 AND
J. Fluid Mech. (7), vol. 58, pp. 153 168. c 7 Cambridge University Press doi:1.117/s11769 Printed in the United Kingdom 153 Energy-minimizing kinematics in hovering insect flight GORDON J. BERMAN 1 AND
CFD DESIGN OF A GENERIC CONTROLLER FOR VORTEX-INDUCED RESONANCE
 Seventh International Conference on CFD in the Minerals and Process Industries CSIRO, Melbourne, Australia 9-11 December 2009 CFD DESIGN OF A GENERIC CONTROLLER FOR VORTEX-INDUCED RESONANCE Andrew A. ANTIOHOS,
Seventh International Conference on CFD in the Minerals and Process Industries CSIRO, Melbourne, Australia 9-11 December 2009 CFD DESIGN OF A GENERIC CONTROLLER FOR VORTEX-INDUCED RESONANCE Andrew A. ANTIOHOS,
Contents. I Introduction 1. Preface. xiii
 Contents Preface xiii I Introduction 1 1 Continuous matter 3 1.1 Molecules................................ 4 1.2 The continuum approximation.................... 6 1.3 Newtonian mechanics.........................
Contents Preface xiii I Introduction 1 1 Continuous matter 3 1.1 Molecules................................ 4 1.2 The continuum approximation.................... 6 1.3 Newtonian mechanics.........................
Spontaneous Symmetry Breaking of a Hinged Flapping Filament Generates Lift
 Spontaneous Symmetry Breaking of a Hinged Flapping Filament Generates Lift A. Bottaro (DICCA, University of Genova) S. Bagheri (KTH, Stockholm) A. Mazzino (DICCA, Genova) J. Favier (AMU, Marseille) A.
Spontaneous Symmetry Breaking of a Hinged Flapping Filament Generates Lift A. Bottaro (DICCA, University of Genova) S. Bagheri (KTH, Stockholm) A. Mazzino (DICCA, Genova) J. Favier (AMU, Marseille) A.
COMPUTATIONAL STUDY ON THE INFLUENCE OF DYNAMIC STALL ON THE UNSTEADY AERODYNAMICS OF FLAPPING WING ORNITHOPTER
 COMPUTATIONAL STUDY ON THE INFLUENCE OF DYNAMIC STALL ON THE UNSTEADY AERODYNAMICS OF FLAPPING WING ORNITHOPTER Alif Syamim Syazwan Ramli and Harijono Djojodihardjo Department of Aerospace Engineering,
COMPUTATIONAL STUDY ON THE INFLUENCE OF DYNAMIC STALL ON THE UNSTEADY AERODYNAMICS OF FLAPPING WING ORNITHOPTER Alif Syamim Syazwan Ramli and Harijono Djojodihardjo Department of Aerospace Engineering,
Physics problems for life scientists Pushing on the Environment (aka collisions for biologists)
") Physics problems for life scientists Pushing on the Environment (aka collisions for biologists) Overview of problem set: Animals move around by pushing on the environment (e.g., the ground, the air or
Physics problems for life scientists Pushing on the Environment (aka collisions for biologists) Overview of problem set: Animals move around by pushing on the environment (e.g., the ground, the air or
Given the water behaves as shown above, which direction will the cylinder rotate?
 water stream fixed but free to rotate Given the water behaves as shown above, which direction will the cylinder rotate? ) Clockwise 2) Counter-clockwise 3) Not enough information F y U 0 U F x V=0 V=0
water stream fixed but free to rotate Given the water behaves as shown above, which direction will the cylinder rotate? ) Clockwise 2) Counter-clockwise 3) Not enough information F y U 0 U F x V=0 V=0
Vortex structures in the wake of a buoyant tethered cylinder at moderate to high reduced velocities
 European Journal of Mechanics B/Fluids 23 (2004) 127 135 Vortex structures in the wake of a buoyant tethered cylinder at moderate to high reduced velocities K. Ryan, M.C. Thompson, K. Hourigan Fluids Laboratory
European Journal of Mechanics B/Fluids 23 (2004) 127 135 Vortex structures in the wake of a buoyant tethered cylinder at moderate to high reduced velocities K. Ryan, M.C. Thompson, K. Hourigan Fluids Laboratory
ν δ - 1 -
 ν δ - 1 - δ ν ν δ ν ν - 2 - ρ δ ρ θ θ θ δ τ ρ θ δ δ θ δ δ δ δ τ μ δ μ δ ν δ δ δ - 3 - τ ρ δ ρ δ ρ δ δ δ δ δ δ δ δ δ δ δ - 4 - ρ μ ρ μ ρ ρ μ μ ρ - 5 - ρ τ μ τ μ ρ δ δ δ - 6 - τ ρ μ τ ρ μ ρ δ θ θ δ θ - 7
ν δ - 1 - δ ν ν δ ν ν - 2 - ρ δ ρ θ θ θ δ τ ρ θ δ δ θ δ δ δ δ τ μ δ μ δ ν δ δ δ - 3 - τ ρ δ ρ δ ρ δ δ δ δ δ δ δ δ δ δ δ - 4 - ρ μ ρ μ ρ ρ μ μ ρ - 5 - ρ τ μ τ μ ρ δ δ δ - 6 - τ ρ μ τ ρ μ ρ δ θ θ δ θ - 7
Fluid Mechanics Prof. T.I. Eldho Department of Civil Engineering Indian Institute of Technology, Bombay. Lecture - 17 Laminar and Turbulent flows
 Fluid Mechanics Prof. T.I. Eldho Department of Civil Engineering Indian Institute of Technology, Bombay Lecture - 17 Laminar and Turbulent flows Welcome back to the video course on fluid mechanics. In
Fluid Mechanics Prof. T.I. Eldho Department of Civil Engineering Indian Institute of Technology, Bombay Lecture - 17 Laminar and Turbulent flows Welcome back to the video course on fluid mechanics. In
SHAPE, FLAPPING AND FLEXION: WING AND FIN DESIGN FOR FORWARD FLIGHT
 The Journal of Experimental Biology 4, 73 85 () Printed in Great Britain The Company of Biologists Limited JEB37 73 SHAPE, FLAPPING AND FLEXION: WING AND FIN DESIGN FOR FORWARD FLIGHT S. A. COMBES* AND
The Journal of Experimental Biology 4, 73 85 () Printed in Great Britain The Company of Biologists Limited JEB37 73 SHAPE, FLAPPING AND FLEXION: WING AND FIN DESIGN FOR FORWARD FLIGHT S. A. COMBES* AND
arxiv: v1 [physics.flu-dyn] 27 Mar 2014
![arxiv: v1 [physics.flu-dyn] 27 Mar 2014](/thumbs/86/93690098.jpg "arxiv: v1 [physics.flu-dyn] 27 Mar 2014") A study on aerodynamics and mechanisms of elementary morphing models for flapping wing in bat forward flight arxiv:143.684v1 [physics.flu-dyn] 7 Mar 14 GUAN Zi-Wu 1,, YU Yong-Liang 1 (1. The Laboratory
A study on aerodynamics and mechanisms of elementary morphing models for flapping wing in bat forward flight arxiv:143.684v1 [physics.flu-dyn] 7 Mar 14 GUAN Zi-Wu 1,, YU Yong-Liang 1 (1. The Laboratory
Computational Aerodynamics of Low Reynolds Number Plunging, Pitching and Flexible Wings for MAV Applications
 46th AIAA Aerospace Sciences Meeting and Exhibit 7-10 January 2008, Reno, Nevada AIAA 2008-523 Computational Aerodynamics of Low Reynolds Number Plunging, Pitching and Flexible Wings for MAV Applications
46th AIAA Aerospace Sciences Meeting and Exhibit 7-10 January 2008, Reno, Nevada AIAA 2008-523 Computational Aerodynamics of Low Reynolds Number Plunging, Pitching and Flexible Wings for MAV Applications
arxiv: v1 [physics.flu-dyn] 30 May 2012
![arxiv: v1 [physics.flu-dyn] 30 May 2012](/thumbs/93/114279992.jpg "arxiv: v1 [physics.flu-dyn] 30 May 2012") Under consideration for publication in J. Fluid Mech. Lift Enhancement of a Vortex-Sink Attached to a Flat Plate arxiv:.68v [physics.flu-dyn] May X. X I A, A N D J. M U L L E N, A N D K. M O H S E N I,,
Under consideration for publication in J. Fluid Mech. Lift Enhancement of a Vortex-Sink Attached to a Flat Plate arxiv:.68v [physics.flu-dyn] May X. X I A, A N D J. M U L L E N, A N D K. M O H S E N I,,
Computational Fluid-Structure Interaction of a Deformable Flapping Wing for Micro Air Vehicle Applications
 46th AIAA Aerospace Sciences Meeting and Exhibit 7-10 January 008, Reno, Nevada AIAA 008-615 Computational Fluid-Structure Interaction of a Deformable Flapping Wing for Micro Air Vehicle Applications Jian
46th AIAA Aerospace Sciences Meeting and Exhibit 7-10 January 008, Reno, Nevada AIAA 008-615 Computational Fluid-Structure Interaction of a Deformable Flapping Wing for Micro Air Vehicle Applications Jian
K. M. Isaac, P. Shivaram + University of Missouri-Rolla Rolla, MO T. DalBello NASA Glenn Research Center
 Low Re, High α Aerodynamics with Controlled Wing Kinematics K. M. Isaac, P. Shivaram + University of Missouri-Rolla Rolla, MO 65409 isaac@umr.edu T. DalBello NASA Glenn Research Center ABSTRACT Numerical
Low Re, High α Aerodynamics with Controlled Wing Kinematics K. M. Isaac, P. Shivaram + University of Missouri-Rolla Rolla, MO 65409 isaac@umr.edu T. DalBello NASA Glenn Research Center ABSTRACT Numerical
Effect of Blockage on Spanwise Correlation in a Circular Cylinder Wake
 Effect of Blockage on Spanwise Correlation in a Circular Cylinder Wake H. M. Blackburn Department of Mechanical Engineering, Monash University May 15, 2003 Summary A short series of experiments was conducted
Effect of Blockage on Spanwise Correlation in a Circular Cylinder Wake H. M. Blackburn Department of Mechanical Engineering, Monash University May 15, 2003 Summary A short series of experiments was conducted
Department of Energy Sciences, LTH
 Department of Energy Sciences, LTH MMV11 Fluid Mechanics LABORATION 1 Flow Around Bodies OBJECTIVES (1) To understand how body shape and surface finish influence the flow-related forces () To understand
Department of Energy Sciences, LTH MMV11 Fluid Mechanics LABORATION 1 Flow Around Bodies OBJECTIVES (1) To understand how body shape and surface finish influence the flow-related forces () To understand
CLAP AND FLING AERODYNAMICS - AN EXPERIMENTAL EVALUATION
 J. exp. Biol. (i977). 69, 261-273 26l With 8 figures 'tinted in Great Britain CLAP AND FLING AERODYNAMICS - AN EXPERIMENTAL EVALUATION BY LEON BENNETT Department of Applied Science, New York University,
J. exp. Biol. (i977). 69, 261-273 26l With 8 figures 'tinted in Great Britain CLAP AND FLING AERODYNAMICS - AN EXPERIMENTAL EVALUATION BY LEON BENNETT Department of Applied Science, New York University,
High Speed Aerodynamics. Copyright 2009 Narayanan Komerath
 Welcome to High Speed Aerodynamics 1 Lift, drag and pitching moment? Linearized Potential Flow Transformations Compressible Boundary Layer WHAT IS HIGH SPEED AERODYNAMICS? Airfoil section? Thin airfoil
Welcome to High Speed Aerodynamics 1 Lift, drag and pitching moment? Linearized Potential Flow Transformations Compressible Boundary Layer WHAT IS HIGH SPEED AERODYNAMICS? Airfoil section? Thin airfoil
Effect of Pivot Point on Aerodynamic Force and Vortical Structure of Pitching Flat Plate Wings
 5st AIAA Aerospace Sciences Meeting including the New Horizons Forum and Aerospace Exposition 7 - January 23, Grapevine (Dallas/Ft. Worth Region), Texas AIAA 23-792 Effect of Pivot Point on Aerodynamic
5st AIAA Aerospace Sciences Meeting including the New Horizons Forum and Aerospace Exposition 7 - January 23, Grapevine (Dallas/Ft. Worth Region), Texas AIAA 23-792 Effect of Pivot Point on Aerodynamic
NUMERICAL SIMULATION OF THE FLOW AROUND A SQUARE CYLINDER USING THE VORTEX METHOD
 NUMERICAL SIMULATION OF THE FLOW AROUND A SQUARE CYLINDER USING THE VORTEX METHOD V. G. Guedes a, G. C. R. Bodstein b, and M. H. Hirata c a Centro de Pesquisas de Energia Elétrica Departamento de Tecnologias
NUMERICAL SIMULATION OF THE FLOW AROUND A SQUARE CYLINDER USING THE VORTEX METHOD V. G. Guedes a, G. C. R. Bodstein b, and M. H. Hirata c a Centro de Pesquisas de Energia Elétrica Departamento de Tecnologias
CFD Time Evolution of Heat Transfer Around A Bundle of Tubes In Staggered Configuration. G.S.T.A. Bangga 1*, W.A. Widodo 2
 CFD Time Evolution of Heat Transfer Around A Bundle of Tubes In Staggered Configuration G.S.T.A. Bangga 1*, W.A. Widodo 2 1,2 Department of mechanical engineering Field of study energy conversion Institut
CFD Time Evolution of Heat Transfer Around A Bundle of Tubes In Staggered Configuration G.S.T.A. Bangga 1*, W.A. Widodo 2 1,2 Department of mechanical engineering Field of study energy conversion Institut
ScienceDirect. Experimental Validation on Lift Increment of a Flapping Rotary Wing with Boring-hole Design
 Available online at www.sciencedirect.com ScienceDirect Procedia Engineering 99 (2015 ) 1543 1547 APISAT2014, 2014 Asia-Pacific International Symposium on Aerospace Technology, APISAT2014 Experimental
Available online at www.sciencedirect.com ScienceDirect Procedia Engineering 99 (2015 ) 1543 1547 APISAT2014, 2014 Asia-Pacific International Symposium on Aerospace Technology, APISAT2014 Experimental
Airfoils and Wings. Eugene M. Cliff
 Airfoils and Wings Eugene M. Cliff 1 Introduction The primary purpose of these notes is to supplement the text material related to aerodynamic forces. We are mainly interested in the forces on wings and
Airfoils and Wings Eugene M. Cliff 1 Introduction The primary purpose of these notes is to supplement the text material related to aerodynamic forces. We are mainly interested in the forces on wings and
Implementing a Partitioned Algorithm for Fluid-Structure Interaction of Flexible Flapping Wings within Overture
 10 th Symposimum on Overset Composite Grids and Solution Technology, NASA Ames Research Center Moffett Field, California, USA 1 Implementing a Partitioned Algorithm for Fluid-Structure Interaction of Flexible
10 th Symposimum on Overset Composite Grids and Solution Technology, NASA Ames Research Center Moffett Field, California, USA 1 Implementing a Partitioned Algorithm for Fluid-Structure Interaction of Flexible
Detailed Outline, M E 320 Fluid Flow, Spring Semester 2015
 Detailed Outline, M E 320 Fluid Flow, Spring Semester 2015 I. Introduction (Chapters 1 and 2) A. What is Fluid Mechanics? 1. What is a fluid? 2. What is mechanics? B. Classification of Fluid Flows 1. Viscous
Detailed Outline, M E 320 Fluid Flow, Spring Semester 2015 I. Introduction (Chapters 1 and 2) A. What is Fluid Mechanics? 1. What is a fluid? 2. What is mechanics? B. Classification of Fluid Flows 1. Viscous
A Momentum Exchange-based Immersed Boundary-Lattice. Boltzmann Method for Fluid Structure Interaction
 APCOM & ISCM -4 th December, 03, Singapore A Momentum Exchange-based Immersed Boundary-Lattice Boltzmann Method for Fluid Structure Interaction Jianfei Yang,,3, Zhengdao Wang,,3, and *Yuehong Qian,,3,4
APCOM & ISCM -4 th December, 03, Singapore A Momentum Exchange-based Immersed Boundary-Lattice Boltzmann Method for Fluid Structure Interaction Jianfei Yang,,3, Zhengdao Wang,,3, and *Yuehong Qian,,3,4
Scaling Law I. BioSensing & BioMEMS 530/ Jeff Wang Johns Hopkins University 1
 Scaling Law I Jeff Wang Johns Hopkins University 1 Why scaling is important at the micro and nano scale Micro and nano devices are orders of magnitude smaller than macro counterparts As the sizes shrink,
Scaling Law I Jeff Wang Johns Hopkins University 1 Why scaling is important at the micro and nano scale Micro and nano devices are orders of magnitude smaller than macro counterparts As the sizes shrink,
Unsteady aerodynamics of fluttering and tumbling plates
 J. Fluid Mech. (25), vol. 541, pp. 65 9. c 25 Cambridge University Press doi:1.117/s221125594x Printed in the United Kingdom 65 Unsteady aerodynamics of fluttering and tumbling plates By A. ANDERSEN 1,
J. Fluid Mech. (25), vol. 541, pp. 65 9. c 25 Cambridge University Press doi:1.117/s221125594x Printed in the United Kingdom 65 Unsteady aerodynamics of fluttering and tumbling plates By A. ANDERSEN 1,
Study of design parameters of flapping-wings
 Study of design parameters of flapping-wings Q. Wang, J.F.L. Goosen, and F. van Keulen Structural Optimization & Mechanics Group Faculty of Mechanical, Maritime and Materials Engineering Delft University
Study of design parameters of flapping-wings Q. Wang, J.F.L. Goosen, and F. van Keulen Structural Optimization & Mechanics Group Faculty of Mechanical, Maritime and Materials Engineering Delft University
Analytical model for instantaneous lift and shape deformation of an insect-scale flapping wing in hover
 Downloaded from http://rsif.royalsocietypublishing.org/ on November 19, 18 rsif.royalsocietypublishing.org Research Cite this article: Kang C-k, Shyy W. 14 Analytical model for instantaneous lift and shape
Downloaded from http://rsif.royalsocietypublishing.org/ on November 19, 18 rsif.royalsocietypublishing.org Research Cite this article: Kang C-k, Shyy W. 14 Analytical model for instantaneous lift and shape
A Biologically Inspired Computational Study of Flow Past Tandem Flapping Foils
 A Biologically Inspired Computational Study of Flow Past andem Flapping Foils I. Akhtar * and R. Mittal Department of Mechanical & Aerospace Engineering he George Washington University, Washington DC 20052
A Biologically Inspired Computational Study of Flow Past andem Flapping Foils I. Akhtar * and R. Mittal Department of Mechanical & Aerospace Engineering he George Washington University, Washington DC 20052
Lecture-4. Flow Past Immersed Bodies
 Lecture-4 Flow Past Immersed Bodies Learning objectives After completing this lecture, you should be able to: Identify and discuss the features of external flow Explain the fundamental characteristics
Lecture-4 Flow Past Immersed Bodies Learning objectives After completing this lecture, you should be able to: Identify and discuss the features of external flow Explain the fundamental characteristics
Visualization of flow pattern over or around immersed objects in open channel flow.
 EXPERIMENT SEVEN: FLOW VISUALIZATION AND ANALYSIS I OBJECTIVE OF THE EXPERIMENT: Visualization of flow pattern over or around immersed objects in open channel flow. II THEORY AND EQUATION: Open channel:
EXPERIMENT SEVEN: FLOW VISUALIZATION AND ANALYSIS I OBJECTIVE OF THE EXPERIMENT: Visualization of flow pattern over or around immersed objects in open channel flow. II THEORY AND EQUATION: Open channel:
Fundamentals of Fluid Dynamics: Ideal Flow Theory & Basic Aerodynamics
 Fundamentals of Fluid Dynamics: Ideal Flow Theory & Basic Aerodynamics Introductory Course on Multiphysics Modelling TOMASZ G. ZIELIŃSKI (after: D.J. ACHESON s Elementary Fluid Dynamics ) bluebox.ippt.pan.pl/
Fundamentals of Fluid Dynamics: Ideal Flow Theory & Basic Aerodynamics Introductory Course on Multiphysics Modelling TOMASZ G. ZIELIŃSKI (after: D.J. ACHESON s Elementary Fluid Dynamics ) bluebox.ippt.pan.pl/