A Samoan Hebeloma with phylogenetic ties to the western Pacific
|
|
|
- Whitney Roberts
- 5 years ago
- Views:
Transcription


1 Utah State University Biology Faculty Publications Biology 2014 A Samoan Hebeloma with phylogenetic ties to the western Pacific Bradley R. Kropp Follow this and additional works at: Part of the Biology Commons Recommended Citation Kropp, Bradley R., "A Samoan Hebeloma with phylogenetic ties to the western Pacific" (2014). Biology Faculty Publications. Paper This Article is brought to you for free and open access by the Biology at DigitalCommons@USU. It has been accepted for inclusion in Biology Faculty Publications by an authorized administrator of DigitalCommons@USU. For more information, please contact dylan.burns@usu.edu.
2 In Press at Mycologia, preliminary version published on October 31, 2014 as doi: / Short title: Samoan Hebeloma A Samoan Hebeloma with phylogenetic ties to the western Pacific Bradley R. Kropp 1 Biology Department 5305 Old Main Hall, Utah State University, Logan, Utah Abstract: Hebeloma ifeleletorum is described as a new species from American Samoa. Based on analyses of ITS and combined nlsu-its datasets H. ifeleletorum clusters with but is distinct from described species that have been placed in the genus Anamika by some. The phylogenetic relationship of H. ifeleletorum to the genus Anamika from Asia and to other species from Australia and New Caledonia suggests that H. ifeleletorum has origins in the western Pacific. Key words: Agaricales, Anamika, biogeography, Oceania, South Pacific INTRODUCTION The islands of the South Pacific have received scant attention from mycologists. At first glance, these islands seems too small to harbor much fungal diversity. Whereas that might be true for many of the tiny atolls scattered across the Pacific, some of the larger volcanic islands like those of the Samoan Archipelago hold unique and diverse plant communities. As a consequence they potentially also hold diverse fungal communities (Whistler 1992, 1994; Hawksworth 2001; Schmit and Mueller 2007). Thus far some familiar and widespread agarics such as Chlorophyllum molybdites are known from American Samoa along with one recently described species of Inocybe and another new species of Moniliophthora (Kropp and Albee-Scott 2010, 2012). Other than that, the Samoan mycobiota is poorly known and more undescribed species probably will be be uncovered as work on the material collected in these islands continues. The purposes of this article are to propose a new species of Hebeloma from American Samoa and to assess its phylogenetic and geographic affinities. Copyright 2014 by The Mycological Society of America.
3 MATERIALS AND METHODS Notes were taken from fresh material in the field and a photograph was taken with a gray card as background. Microscopic study of dried material was done with a light microscope after mounting sections in Melzer's reagent, 10% NH 4 OH and cotton blue. All measurements were made with material mounted in 10% NH 4 OH. Color notations are from Munsell Soil Color Charts (Munsell Color 2000) and were taken from digital photographs made in the field. The microscopic measurements were made with oil immersion at 1000 and line drawings were done with the aid of a drawing tube. Basidiospores were photographed with a light microscope at 1000 with DIC illumination. The spore measurements cited are from the holotype and are derived from 20 spores and are reported as an average and a Q value with ranges, whereas measurements for the other cells are given as a size range. The scales proposed by Vesterholt (2005) were used to quantify spore ornamentation (O0 O4), loosening of the perispore (P0 P3), and dextriniod reaction (D0 D4). The holotype was deposited in the Intermountain Herbarium (UTC) at Utah State University. DNA was extracted with a modified CTAB miniprep after crushing a small sample of dried herbarium material in liquid nitrogen (Kropp et al. 1996) and target sequences were amplified with standard PCR protocols (White et al. 1990). The portion of the nuclear large ribosomal subunit (nlsu) in this study was amplified with primers LROR and LR5 (Moncalvo et al. 2000), and primers ITS4 and ITS5 (White et al. 1990) were used to amplify the internal transcribed spacer (ITS) region. Direct sequencing of the amplified PCR products also was done using these primers with an ABI 3730 DNA analyzer. The new sequences generated were deposited in GenBank (ITS = KJ494930, nlsu = KJ494929). Initial BLAST queries indicated that the species from Samoa had affinities to the genus Anamika, thus the aligned dataset of Yang et al. (2005) available at TreeBase (alignment S2122) was used as a base for assessing the phylogenetic position of the Samoan material. To generate the combined nlsu-its phylogram, the Samoan sequences were aligned onto this dataset with Clustal X (Thompson et al. 1997), and the alignments were adjusted by hand where necessary before a phylogenetic analysis was done with MrBayes 3.1 (Ronquist and Huelsenbeck 2003). Taxon sampling for the ITS phylogram also was based on the dataset of Yang et al. (2005) obtained from TreeBASE, however the dataset was pruned to reduce the number of representatives for individual species and some taxa were added from Rees et al. (2013). The dataset was realigned before being analyzed. The realigned nlsu-its and ITS datasets were deposited in TreeBASE (alignments and respectively).
4 Tree searches were performed for both sets of sequences using a time reversible model of evolution (Rodriguez et al. 1990, Maddison 1994) and a gamma distribution with six substitution types and some invariant sites (GTR+G+I). The Bayesian simulations were conducted with eight active MCMC chains and with the temperature set at 0.2. The four MCMC simulations were done with generations and with sampling every one-hundredth tree to approximate posterior probabilities. The potential scale reduction factors for all parameters in both analyses were 0 2, indicating that convergence had occurred. A strict consensus tree was calculated from the last 7500 sampled trees from a tree dataset. Support measures for nodes with less than 70 percent posterior-probability support are not shown on the phylograms. TreeView (Page 1996) was used to visualize the consensus trees obtained from the simulation. The tree generated with the ITS sequences was rooted with species of Naucoria, whereas the tree generated from the combined nlsu-its sequences was rooted with species of Gymnopilus. RESULTS ITS analysis indicates that the Samoan species, Hebeloma ifeleletorum, falls into a wellsupported clade comprising fungi that have been placed in the genus Anamika by Thomas et al. (2002) and Yang et al. (2005). In addition the results of the ITS analysis indicate that Hebeloma ifeleletorum is closely related to Hebeloma (Anamika) indica, H. (Anamika) angustilamellata, H.(Anamika) lactariolens and H. youngii as well as to a number of additional pleurocystidiate Hebeloma species. However it is phylogenetically distinct from each of these species (FIG. 1). The combined nlsu-its analysis produced results similar to those obtained with the ITS sequences alone (FIG. 2) and supports the recognition of a new species. In addition the nlsu- ITS and the ITS analysis also support the conclusion of Rees et al. (2013) that the clade comprising the species of Anamika is derived from within Hebeloma and should not be given generic recognition. DISCUSSION
5 Because Hebeloma is an ectomycorrhizal genus, Hebeloma ifeleletorum is an unexpected element of the Polynesian mycobiota. Except for Inocybe tauensis from American Samoa (Kropp and Albee-Scott 2010) and Tomentella species associated with Pisonia grandis on coral cays in the Great Barrier Reef (Ashford and Allaway 1982, Chambers et al. 2005), few fungi from ectomycorrhizal genera have been reported from the tropical South Pacific. However, given the presence of ectomycorrhizal hosts such as P. grandis and Intsia bijuga, additional ectomycorhizal fungi potentially will be found on these islands (Whistler 1992, 1994). Samoan plant communities are thought to have Australasian connections (van Balgooy et al. (1996), but the geographical origins of the fungal populations found in Samoa are unknown. The meager data available so far indicate that these populations have a complex history. For example, Kropp and Albee Scott (2010) published evidence that Inocybe tauensis, the other putative ectomycorrhizal fungus reported from Samoa, originated in the Paleotropics of the western Pacific. On the other hand, another Samoan species, Moniliophthora aurantiaca, might have a Neotropical rather than a Paleotropical origin (Kropp and Albee-Scott 2012) and some of the fungi that are present such as Chlorophyllum molybdites appear to be Pantropical. In the case of Hebeloma ifeleletorum the phylogenetic and morphological evidence both point to a tight relationship with species that have been placed in the genus Anamika. The fact that all known Anamika species occur in Australia or Asia strongly suggests that H. ifeleletorum has connections to the western Pacific or India (Thomas et al. 2002, Yang et al. 2005, Rees et al. 2013). Whether Hebeloma ifeleletorum is a recent arrival to Samoa or an endemic species is still an open question. Two of the specimens of H. ifeleletorum reported in this paper were collected within a native stand of Intsia bijuga in the National Park of American Samoa and a third
6 specimen was found with saplings of Intsia bijuga that had been planted along a trail within the park. The fact that one collection was found with planted trees initially raised the question of whether the fungus might have been transported with the trees from elsewhere because Intsia bijuga is such a widespread ectomycorrhizal species (Alexander and Högberg 1986). However it is known that the seed used to grow the planted trees was collected within the park (Tavita Togia, National Park of American Samoa, pers comm), eliminating the possibility that the fungus was imported recently with the planted trees. The question of whether this fungus is endemic to Samoa cannot be fully answered without further work on its geographical distribution. Rees et al. (2013) concluded that the clade containing the species placed in the genus Anamika lacks sufficient support to merit generic status. This clade currently comprises Hebeloma (Anamika) indica, H. (Anamika) angustilamellata and H.(Anamika) lactariolens along with H. youngii and H. ifeleletorum (FIG. 1). The phylogeny presented here is consistent not only with the work of Rees et al. (2013) but also of Yang et al. (2005) in showing that the anamika clade is derived from within Hebeloma. As a result accepting Anamika as a genus would render Hebeloma polyphyletic. Furthermore the primary distinguishing morphological character for Anamika is the presence of pleurocystidia, which typically are lacking in Hebeloma species (Smith et al. 1983, Vesterholt 2005). Yet there are several Australasian Hebeloma species that are pleurocystidiate and cluster in a well-supported clade with the species of Anamika (FIG. 1). This indicates that more species could be added to Anamika, however the character is unreliable because two clade members, H. victoriense and H. khogianum, do not possess pleurocystidia (TABLE 1). The other salient morphological character given for Anamika by Thomas et al. (2002) is the presence of cavernae in the basidiospores. Unfortunately, with the exception of Anamika indica, none of the descriptions of the current Anamika species or of the Hebeloma
7 species that cluster with it in the pleurocystidiate clade clearly indicate whether their spores possess cavernae (TABLE 1). Based on the currently available phylogenetic evidence and the inconsistent presence of pleurocystidia and cavernae among the fungi that could be placed in Anamika, it is preferable to follow Rees et al. (2013) by placing all these species into Hebeloma. The Samoan species described here is placed in Hebeloma. However further phylogenetic work and studies on the distribution of cavernae among species in this group should be done before a final decision is made to retain or reject Anamika. The phylogenetic analyses for this study (FIGS. 1, 2) indicate that Hebeloma ifeleletorum is closely related to Hebeloma (Anamika) lactoriolens, H. (Anamika) indica and to H. (Anamika) angustilamellata. However, in addition to being phylogenetically divergent from H. lactariolens, H. ifeleletorum can be differentiated from it by its smooth rather than rugulose pileus surface, by the fact that it lacks a swollen stipe base and by its nondextrinoid spores. Hebeloma indica is morphologically similar to H. ifeleletorum but differs by having larger basidiospores, a solid stipe and versiform pleurocystidia rather than consistently rostrate pleurocystidia such as those found in H. ifeleletorum. Hebeloma angustilamellata is both phylogenetically distinct from H. ifeleletorum and differentiated from it by having much larger basidiomata, slightly rugulose pilei and extremely crowded narrow lamellae. Finally Rees et al. (2013) described an additional species, Hebeloma youngii, from Australia that falls within the anamika clade. Hebeloma youngii and H. ifeleletorum could be considered cryptic species in that they strongly resemble one another morphologically but are phylogenetically distinct. However, in addition to being phylogenetically separated, the two species also have sharply different ecological characteristics. Hebeloma youngii is associated with Eucalyptus and Corymbia in Australia where it was reported from decomposing marsupial remains (Rees et al. 2013), whereas H. ifeleletorum occurs
8 on soil and is associated with Intsia bijuga in American Samoa. There are relatively few reliable morphological differences between the two species. Although the basidiospores of Hebeloma youngii are strongly dextrinoid (D3) whereas the spores of H. ifeleletorum are not dextrinoid (D0). Hebeloma youngii also produces longer cheilocystidia than H.ifeleletorum (45 80 µm versus µm for H. ifeleletorum). TAXONOMY Hebeloma ifeleletorum Kropp, sp. nov. FIGS. 3, 4, 5 MycoBank MB Holotype: USA. AMERICAN SAMOA: National Park of American Samoa, Tutuila Unit, clustered on ground along Mount Alava Trail with Intsia bijuga, 19 May 2009, B.R. Kropp 19-May (HOLOTYPE UTC ). Diagnosis: Hebeloma ifeleletorum differs from H. lactariolens by its smooth pileus and by lacking a swollen stipe base, from Hebeloma indica by having larger basidiospores and rostrate pleurocystidia and from H. angustilamellata by having much smaller basidiomata. It is phylogenetically and ecologically divergent from Hebeloma youngii. Etymology: belonging to Ifelele, the Samoan name for the tree Intsia bijuga. Pileus 3 32 mm diam, at first broadly conic with the margin slightly inrolled, later expanding and becoming convex, smooth, lacking an umbo; reddish brown to buff (5YR 4/6 7.5YR 8/4) at the pileus center, becoming pale buff to cream (10YR 8/3 ) toward the margin; a pale whitish veil is present before expansion. Lamellae mm wide (dry material), narrowly attached, close, with even concolorous edge; pallid or pale pinkish brown when young, dull pinkish brown (5YR 7/4) at maturity, lamellulae 1 2 mm long. Stipe mm, central, terete, equal or slightly enlarged toward the base, hollow, pliant; surface finely pruinose over the upper third of the stipe, otherwise lightly longitudinally fibrillose with faint annular
9 cortinate remains present, some fibrils and the remains of the cortina become brownish mostly from spore deposits; stipe light buff to cream (10YR 8/3 8/2 ), the base becoming reddish brown (5YR 4/4) at maturity. Dried spore print on white paper dull reddish brown (5YR 5/3). Odor faintly fungoid to acidulous. Context pallid, no color changes observed. Basidiospores 9.0(10.6) (6.8)7.5 µm, Q = 1.55, ornamented (O4) with a loosening perispore (P2) but cavernae not obvious under light microscopy, amygdaliform, not cyanophilous, not reacting in Melzer's reagent (D0). Basidia µm, four-spored, clavate. Cheilocystidia µm, numerous, hyaline, thin-walled, variable but mostly clavate to cylindrical or fusiform. Pleurocystidia µm, not abundant, hyaline, thinwalled, mostly rostrate occasionally with an elongated tip. Pileipellis a layer of enlarged cylindrical to subglobose cells, µm wide, with a thin superficial, slightly gelatinized layer; golden brown encrusting material present. Lamellar trama of parallel, smooth hyphae 4 8 µm wide. Stipe surface at apex a loosely woven covering of hyphae with numerous irregular mostly clavate cystidioid tips, µm; at base covered with loosely woven hyphae, 3 6 µm wide, with some golden brown encrusting material. Clamp connections present. Habitat: clustered on the ground in association with Intsia bijuga. Additional specimens examined: USA. AMERICAN SAMOA: National Park of American Samoa, Tutuila Unit, clustered on ground at Siufaga Ridge with Intsia bijuga, 4 Dec 2013, B.R. Kropp 4-Dec (UTC ); National Park of American Samoa, Tutuila Unit, clustered on ground at Siufaga Ridge with Intsia bijuga, 4 December 2013, B.R. Kropp 4-Dec (UTC ). ACKNOWLEDGMENTS The National Geographic Society is gratefully acknowledged for providing the financial support to do field and laboratory work for this project. The National Park of American Samoa and the Department of Marine and Wildlife Resources in American Samoa are thanked for providing the necessary collecting permits and for giving helpful advice about collecting within the national park.
10 LITERATURE CITED Alexander IJ, Högberg P Ectomycorrhizas of tropical angiospermous trees. New Phytol 102: Ashford AE, Allaway WG A sheathing mycorrhiza on Pisonia grandis R. Br. (Nyctaginageae) with development of transfer cells rather than a Hartig net. New Phytol 90: Chambers SM, Hitchcock CJ, Cairney JWG Ectomycorrhizal mycobionts of Pisonia grandis on coral cays in the Capricorn-Bunker group, Great Barrier Reef, Australia. Mycol Res 109: Hawksworth DL The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycol Res 105: Holland AA, Pegler DN Hebeloma victoriense and the genus Metraria. Trans Br Mycol Soc 80: Kropp BR, Albee-Scott S Inocybe tauensis, a new species from the Samoan Archipelago with biogeographic evidence for a Paleotropical origin. Fungal Biol 114: , Moniliophthora aurantiaca sp. nov. a Polynesian species occurring in littoral forests. Mycotaxon 120: , Hansen D, Flint KM, Thomson SV Artificial inoculation and colonization of dyer's woad (Isatis tinctoria) by the systemic fungus Puccinia thlaspeos. Phytopathology 86: Maddison DR Phylogenetic methods for inferring the evolutionary history and processes of change in discretely valued characters. Ann Rev Entomol 39: Moncalvo J-M, Lutzoni FM, Rehner SA, Johnson J, Vilgalys R Phylogenetic relationships of Agaric fungi based on nuclear large subunit ribosomal DNA sequences. Syst Biol 49:
11 Munsell Color Munsell soil color charts. New Windsor, New York: GretagMacbeth Page RDM TreeView: an application to display phylogenetic trees on personal computers. Comput Appl Biosci 12: Rees BJ, Midgley DJ, Marchant A, Perkins A, Orlovich DA Morphological and molecular data for Australian Hebeloma species do not support the generic status of Anamika. Mycologia 105: Rodriguez F, Oliver JL, Marin A, Medina JR The general stochastic model of nucleotide substitution. J Theor Biol 142: Ronquist F, Huelsenbeck JP MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: Schmit PP, Mueller GM An estimate of the lower limit of global fungal diversity. Biodivers Conser 16: Smith AH, Evenson VA, Mitchel DH The veiled species of Hebeloma in the western United States. Ann Arbor: Univ. Mich. Press. p 219. Thomas KA, Peintner U, Moser MM, Manimohan P Anamika, a new mycorrhizal genus of Cortinariaceae from India and its phylogenetic position based on ITS and LSU sequences. MycolRes 106: Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG The Clustal X Windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 24:
12 van Balgooy MMJ, Hovenkamp PH, van Welzen PC Phytogeography of the Pacific floristic and historical distribution patterns in plants. In: Keast A, Miller SE, eds. The origin and evolution of Pacific island biotas, New Guinea to eastern Polynesia: patterns and processes. Amsteredam, the Netherlands: SPB Academic Publishing. p Vesterholt J The genus Hebeloma. Fungi North Eur 3: Whistler WA Botanical inventory of the proposed Ta'u unit of the National Park of American Samoa. National Park Service, Honolulu, Hawaii. Gen Tech. Rep Botanical inventory of the proposed Tutuila and Ofu units of the National Park of American Samoa. National Park Service, Honolulu, Hawaii. Gen. Tech. Rep. 87. White TJ, Bruns T, Lee S, Taylor J Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: PCR protocols: a guide to methods and applications. Innis MA, Gelfand DH, Sninsky JJ, White TJ, eds. New York: Academic Press. p Yang ZL, Matheny PB, Ge ZW, Slot JC, Hibbett DS New Asian species of the genus Anamika (euagarics, hebelomatoid clade) based on morphology and ribosomal DNA sequences. Mycol Res 109: LEGENDS FIG. 1. Bayesian phylogram resulting from an analysis of an ITS dataset showing the position of Hebeloma ifeleletorum relative to the anamika clade and other pleurocystidiate species. Support measures are not shown for nodes with less than 70% posterior probability support. GenBank numbers are shown for the ITS sequences used in the analysis. Following Rees et al. (2013), Anamika indica = Hebeloma indicum, A. lactariolens = H. lactariolens, A. angustilamellata = H. angustilamellatum. FIG. 2. Bayesian phylogram resulting from an analysis of a combined ITS-nLSU dataset showing the position of Hebeloma ifeleletorum within the anamika clade. Support measures are not shown for nodes with less than 70%
13 posterior probability support. Following Rees et al. (2013), Anamika indica = Hebeloma indicum, A. lactariolens = H. lactariolens, A. angustilamellata = H. angustilamellatum. FIG. 3. Basidiomata of the holotype collection for Hebeloma ifeleletorum (UTC253643). Scale bar = 1 mm. FIG. 4. Microscopic characters of Hebeloma ifeleletorum. S = basidiospores, B = basidia, Ch = cheilocystida, Ca = caulocystidia, Pl = pleurocystidia. Scale bar = 10 µm. FIG. 5. Basidiospores of Hebeloma ifeleletorum. P = perispore. Scale bar = 10 µm. FOOTNOTES Submitted 28 Feb 2014; accepted for publication 3 Oct Corresponding author. brad.kropp@usu.edu
14 TABLE I. Salient morphological characters and origins of the species in the pleurocystidiate clade Taxon Pleurocystidia Cavernae b Geographical origin Anamika a angustilamellata Zhu L. Yang & Z.W. Ge + 0 China Anamika indica K.A. Thomas, Peintner, M.M.Moser & Manim. + + India Amanika lactariolens (Clémençon & Hongo) Matheny + Japan Hebeloma aminophilum R.N. Hilton & O.K. Mill. + 0 Australia Hebeloma ifeleletorum + Samoa Hebeloma khogianum Bresinsky 0 New Caledonia Hebeloma subvictoriense B.J. Rees + 0 Australia Hebeloma victoriense A.A. Holland & Pegler + Australia Hebeloma youngii B.J. Rees + 0 Australia a Anamika indica = Hebeloma indicum, A. lactariolens = H. lactariolens, A. angustilamellata = H. angustilamellatum. b Cavernae indicated as + (present), (absent), or 0 (not clearly given or illustrated in the species description). The authors of H. victoriense do not specifically describe cavernae but their basidiospore illustration shows them to be present (Holland and Pegler 1983).
15 Naucoria escharoides AF Hebeloma crustuliniforme AF Hebeloma crustuliniforme AF Hebeloma pusillum AF Hebeloma helodes AF Hebeloma crustuliniforme AF Hebeloma pusillum AF Hebeloma cavipes AF Hebeloma hiemale AF anamika clade 0.91 Hebeloma helodes AF Hebeloma helodes AF Hebeloma lacteocoffeatum GU Hebeloma lacteocoffeatum GU Anamika angustilamellata AY Anamika angustilamellata AY Anamika lindica AF Hebeloma ifeleletorum KJ Hebeloma youngii GU Anamika lactariolens AY Hebeloma aminophilum KC Hebeloma victoriense GU Hebeloma victoriense GU Hebeloma khogianum GU Hebeloma subvictoriense GU Hebeloma incarnatulum AF Hebeloma velutipes AF Hebeloma velutipes AY Hebeloma velutipes AF Hebeloma bulbiferum AF Hebeloma radicosum AF Hebeloma sarcophyllum AF Hebeloma sacchariolens AF Hebeloma tomentosum AF Hebeloma collariatum AF Hebeloma mesophaeum AF Hebeloma sinapizans AF Hebeloma truncatum AF Hebeloma danicum AF Hebeloma edurum AF Hebeloma birrus AF Hebeloma cylindrosporum AF Hebeloma circinans AF Hymenogaster sp AF Hymenogaster subalpinus AF Hymenogaster populetorum AF Hymenogaster bulliardii AF Naucoria amarescens AY Naucoria geraniolens AY Naucoria tantilla AY pleurocystidiate clade
16 Gymnopilus spectabilis Gymnopilus sapineus Anamika indica Anamika angustilamellata Anamika angustilamellata Hebeloma ifeleletorum Anamika lactariolens Hebeloma fastibile Hebeloma crustuliniforme Hebeloma velutipes Naucoria bohemica Naucoria escharoides Agrocybe praecox Agrocybe praecox Hypholoma sublateritium Stropharia ambigua 0.1

17
18 Ca S B Pl Ch

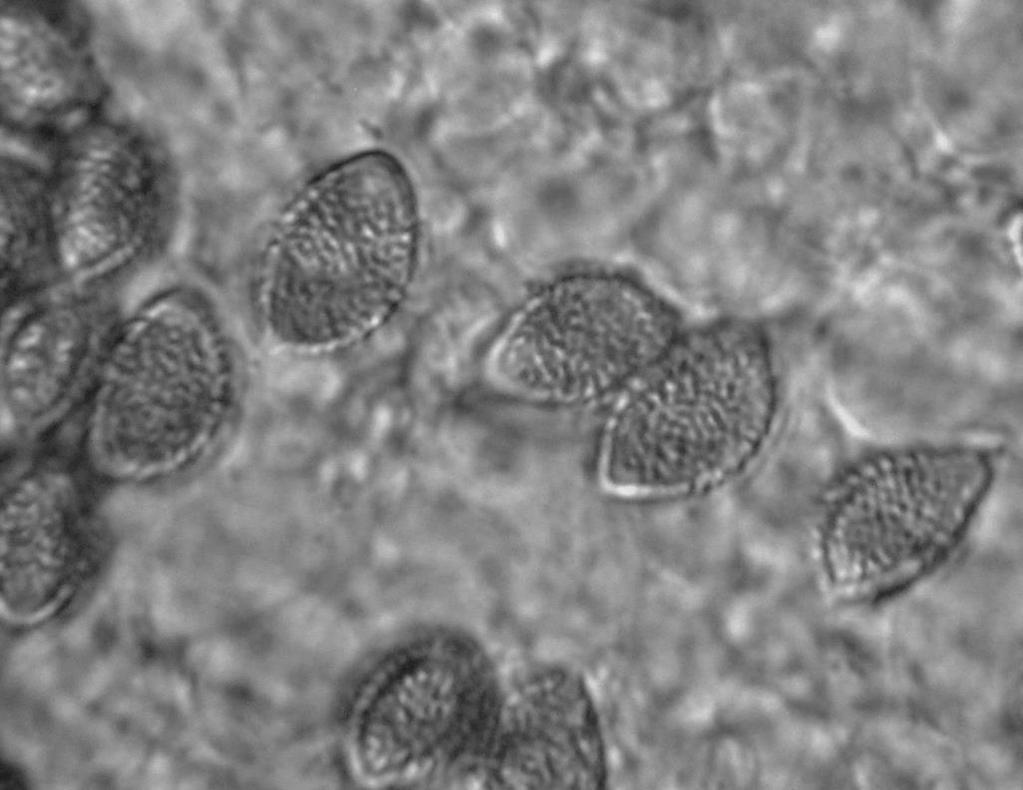
19 P
Clitocella (Entolomataceae) - a new genus record for India
 - a new genus record for India") Studies in Fungi 1 (1): 130 134(2016) ISSN 2465-4973 Article Doi 10.5943/sif/1/1/13 Copyright Mushroom Research Foundation 2016 Clitocella (Entolomataceae) - a new genus record for India Kour H 1, Kumar
Studies in Fungi 1 (1): 130 134(2016) ISSN 2465-4973 Article Doi 10.5943/sif/1/1/13 Copyright Mushroom Research Foundation 2016 Clitocella (Entolomataceae) - a new genus record for India Kour H 1, Kumar
Caloplaca citrina and C. lactea are incorrectly understood in the British Isles
 Caloplaca citrina and C. lactea are incorrectly understood in the British Isles Introduction In the British Isles the genus Caloplaca is rich in species and new taxa are still being described from there
Caloplaca citrina and C. lactea are incorrectly understood in the British Isles Introduction In the British Isles the genus Caloplaca is rich in species and new taxa are still being described from there
Ectomycorrhizal fungi above and below ground in a small, isolated aspen stand: A simple system reveals fungal fruiting strategies an an edge effect
 Ectomycorrhizal fungi above and below ground in a small, isolated aspen stand: A simple system reveals fungal fruiting strategies an an edge effect Cripps, C. L. (Department of Plant Sciences and Plant
Ectomycorrhizal fungi above and below ground in a small, isolated aspen stand: A simple system reveals fungal fruiting strategies an an edge effect Cripps, C. L. (Department of Plant Sciences and Plant
MYCOTAXON. Volume 100, pp April June Zhu L. Yang
 MYCOTAXON Volume 100, pp. 279 287 April June 2007 Type studies on Chamaeota species described from China Zhu L. Yang fungi@mail.kib.ac.cn, fungiamanita@gmail.com Key Laboratory of Biodiversity and Biogeography,
MYCOTAXON Volume 100, pp. 279 287 April June 2007 Type studies on Chamaeota species described from China Zhu L. Yang fungi@mail.kib.ac.cn, fungiamanita@gmail.com Key Laboratory of Biodiversity and Biogeography,
Short guide to some common mycological terms
 Short guide to some common mycological terms Thomas Læssøe & Jens H. Petersen Macro-morphology English (latinised English) Bulb-like (= bulbous) used for swollen stem bases, can be rimmed (= marginate).
Short guide to some common mycological terms Thomas Læssøe & Jens H. Petersen Macro-morphology English (latinised English) Bulb-like (= bulbous) used for swollen stem bases, can be rimmed (= marginate).
Lactocollybia subvariicystis, a new species of little known genus Lactocollybia from subtropical south China
 Mycosphere 7 (6): 794 800 (2016) ISSN 2077 7019 www.mycosphere.org Article Mycosphere Copyright 2016 Online Edition Doi 10.5943/mycosphere/7/6/10 Lactocollybia subvariicystis, a new species of little known
Mycosphere 7 (6): 794 800 (2016) ISSN 2077 7019 www.mycosphere.org Article Mycosphere Copyright 2016 Online Edition Doi 10.5943/mycosphere/7/6/10 Lactocollybia subvariicystis, a new species of little known
SPORE-FORMS IN SPOROPHORES OF GANODERMA LUCIDUM (LEYSS.) KARST.
 KARST.") SPORE-FORMS IN SPOROPHORES OF GANODERMA LUCIDUM (LEYSS.) KARST. BY SACHINDRANATH BANERJEE AND ANJALI SARKAR (Department of Botany, University o[ Calcutta) Received September 12, 1958 (Communicated by Dr.
SPORE-FORMS IN SPOROPHORES OF GANODERMA LUCIDUM (LEYSS.) KARST. BY SACHINDRANATH BANERJEE AND ANJALI SARKAR (Department of Botany, University o[ Calcutta) Received September 12, 1958 (Communicated by Dr.
Post-doc fellowships to non-eu researchers FINAL REPORT. Home Institute: Centro de Investigaciones Marinas, Universidad de La Habana, CUBA
 Recipient: Maickel Armenteros Almanza. Post-doc fellowships to non-eu researchers FINAL REPORT Home Institute: Centro de Investigaciones Marinas, Universidad de La Habana, CUBA Promoter: Prof. Dr. Wilfrida
Recipient: Maickel Armenteros Almanza. Post-doc fellowships to non-eu researchers FINAL REPORT Home Institute: Centro de Investigaciones Marinas, Universidad de La Habana, CUBA Promoter: Prof. Dr. Wilfrida
Micropsalliota brunneosquamata, a New Species from Thailand
 Chiang Mai J. Sci. 2016; 43(4) 689 Chiang Mai J. Sci. 2016; 43(4) : 689-694 http://epg.science.cmu.ac.th/ejournal/ Contributed Paper Micropsalliota brunneosquamata, a New Species from Thailand Jie Chen
Chiang Mai J. Sci. 2016; 43(4) 689 Chiang Mai J. Sci. 2016; 43(4) : 689-694 http://epg.science.cmu.ac.th/ejournal/ Contributed Paper Micropsalliota brunneosquamata, a New Species from Thailand Jie Chen
A new species of Psathyrella (Psathyrellaceae, Agaricales) collected on dung from Punjab, India
 collected on dung from Punjab, India") Journal on New Biological Reports 2(3): 275-280 (2013) ISSN 2319 1104 (Online) A new species of Psathyrella (Psathyrellaceae, Agaricales) collected on dung from Punjab, India Amandeep Kaur 1*, NS Atri
Journal on New Biological Reports 2(3): 275-280 (2013) ISSN 2319 1104 (Online) A new species of Psathyrella (Psathyrellaceae, Agaricales) collected on dung from Punjab, India Amandeep Kaur 1*, NS Atri
The utility of morphological, ITS molecular and combined datasets. in estimating the phylogeny of the. cortinarioid sequestrate fungi.
 The utility of morphological, ITS molecular and combined datasets in estimating the phylogeny of the cortinarioid sequestrate fungi by Anthony Francis This thesis is presented for the degree of Doctor
The utility of morphological, ITS molecular and combined datasets in estimating the phylogeny of the cortinarioid sequestrate fungi by Anthony Francis This thesis is presented for the degree of Doctor
Two new species of Mycena Section Longisetae from Mexico
 Current Research in Environmental & Applied Mycology 9(1): 36 43 (2019) ISSN 2229-2225 www.creamjournal.org Article Doi 10.5943/cream/9/1/4 Two new species of Mycena Section Longisetae from Mexico Cortés-Pérez
Current Research in Environmental & Applied Mycology 9(1): 36 43 (2019) ISSN 2229-2225 www.creamjournal.org Article Doi 10.5943/cream/9/1/4 Two new species of Mycena Section Longisetae from Mexico Cortés-Pérez
Vesicular-arbuscular mycorrhizal fungal sporocarps associated with Pennisetum pedicillatum
 Proc. lndian Acad. Sci. (Plant Sci.), Vol. 96, No. 2, June 1986, pp. 153--158. 9 Printed in India. Vesicular-arbuscular mycorrhizal fungal sporocarps associated with Pennisetum pedicillatum K AMMANI, K
Proc. lndian Acad. Sci. (Plant Sci.), Vol. 96, No. 2, June 1986, pp. 153--158. 9 Printed in India. Vesicular-arbuscular mycorrhizal fungal sporocarps associated with Pennisetum pedicillatum K AMMANI, K
Peter Gault Kennedy CURRICULUM VITAE. 321 Koshland Hall phone: University of California, Berkeley fax: Berkeley, CA 94720
 Peter Gault Kennedy CURRICULUM VITAE Department of Plant and Microbial Biology pkennedy@berkeley.edu 321 Koshland Hall phone: 510-643-5483 University of California, fax: 510-642-4995, CA 94720 Professional
Peter Gault Kennedy CURRICULUM VITAE Department of Plant and Microbial Biology pkennedy@berkeley.edu 321 Koshland Hall phone: 510-643-5483 University of California, fax: 510-642-4995, CA 94720 Professional
Managing Mycological Mysteries
 Managing Mycological Mysteries (Systematics and the Identification of Fungi) NPDN meeting March 2016 Megan Romberg USDA APHIS PPQ PHP NIS APHIS NIS Beltsville APHIS CPHST Beltsville APHIS NIS (Mycology)
Managing Mycological Mysteries (Systematics and the Identification of Fungi) NPDN meeting March 2016 Megan Romberg USDA APHIS PPQ PHP NIS APHIS NIS Beltsville APHIS CPHST Beltsville APHIS NIS (Mycology)
Minor Research Project
 Executive Summary Minor Research Project DNA BARCODING OF MURDANNIA (COMMELINACEAE) IN WESTERN GHATS MRP (S)-1409/11-12/KLMG002/UGC-SWRO By Rogimon P. Thomas Assistant Professor Department of Botany CMS
Executive Summary Minor Research Project DNA BARCODING OF MURDANNIA (COMMELINACEAE) IN WESTERN GHATS MRP (S)-1409/11-12/KLMG002/UGC-SWRO By Rogimon P. Thomas Assistant Professor Department of Botany CMS
A Study of the Moss Parasite Eocronartium muscicola By: Alicia Knudson Advisor: Dr. Elizabeth Frieders
 A Study of the Moss Parasite Eocronartium muscicola By: Alicia Knudson Advisor: Dr. Elizabeth Frieders Abstract The genus Eocronartium contains a single described species of parasitic fungus on moss plants
A Study of the Moss Parasite Eocronartium muscicola By: Alicia Knudson Advisor: Dr. Elizabeth Frieders Abstract The genus Eocronartium contains a single described species of parasitic fungus on moss plants
Effects of Gap Open and Gap Extension Penalties
 Brigham Young University BYU ScholarsArchive All Faculty Publications 200-10-01 Effects of Gap Open and Gap Extension Penalties Hyrum Carroll hyrumcarroll@gmail.com Mark J. Clement clement@cs.byu.edu See
Brigham Young University BYU ScholarsArchive All Faculty Publications 200-10-01 Effects of Gap Open and Gap Extension Penalties Hyrum Carroll hyrumcarroll@gmail.com Mark J. Clement clement@cs.byu.edu See
Quantum Dots: A New Technique to Assess Mycorrhizal Contributions to Plant Nitrogen Across a Fire-Altered Landscape
 2006-2011 Mission Kearney Foundation of Soil Science: Understanding and Managing Soil-Ecosystem Functions Across Spatial and Temporal Scales Progress Report: 2006007, 1/1/2007-12/31/2007 Quantum Dots:
2006-2011 Mission Kearney Foundation of Soil Science: Understanding and Managing Soil-Ecosystem Functions Across Spatial and Temporal Scales Progress Report: 2006007, 1/1/2007-12/31/2007 Quantum Dots:
Mycological Notes 35. New Zealand Marasmiaceae. Jerry Cooper, 14 th Dec. 2016
 Mycological Notes 35 New Zealand Marasmiaceae Jerry Cooper, 14 th Dec. 2016 The following genera are placed within the family Marasmiaceae: Campanella, Cellypha, Crinipellis, Chaetocalathus, Lactocollybia,
Mycological Notes 35 New Zealand Marasmiaceae Jerry Cooper, 14 th Dec. 2016 The following genera are placed within the family Marasmiaceae: Campanella, Cellypha, Crinipellis, Chaetocalathus, Lactocollybia,
Basidiospore homoplasy and variation in the Inocybe chelanensis group in North America
 Mycologia, 96(2), 2004, pp. 295 309. 2004 by The Mycological Society of America, Lawrence, KS 66044-8897 Basidiospore homoplasy and variation in the Inocybe chelanensis group in North America Bradley R.
Mycologia, 96(2), 2004, pp. 295 309. 2004 by The Mycological Society of America, Lawrence, KS 66044-8897 Basidiospore homoplasy and variation in the Inocybe chelanensis group in North America Bradley R.
MYCOTAXON. Annulate Pluteus species, a study of the genus Chamaeota in the United States
 MYCOTAXON Volume 96, pp. 31 39 April June 2006 Annulate Pluteus species, a study of the genus Chamaeota in the United States Andrew M. Minnis 1*, Walter J. Sundberg 1, Andrew S. Methven 2, Sedonia D. Sipes
MYCOTAXON Volume 96, pp. 31 39 April June 2006 Annulate Pluteus species, a study of the genus Chamaeota in the United States Andrew M. Minnis 1*, Walter J. Sundberg 1, Andrew S. Methven 2, Sedonia D. Sipes
Supporting Information Table S1 and Methods S1
 Supporting Information Table S1 and Methods S1 Methods S1 Power analysis for multivariate data. Recent developments in computational biology have made it possible to conduct power analyses on multivariate
Supporting Information Table S1 and Methods S1 Methods S1 Power analysis for multivariate data. Recent developments in computational biology have made it possible to conduct power analyses on multivariate
ISSN (print) Mycotaxon, Ltd. ISSN (online) MYCOTAXON
 Mycotaxon, Ltd. ISSN (online) MYCOTAXON") ISSN (print) 0093-4666 2013. Mycotaxon, Ltd. ISSN (online) 2154-8889 MYCOTAXON http://dx.doi.org/10.5248/123.169 Volume 123, pp. 169 181 January March 2013 Two striking Inocybe species from Yunnan Province,
ISSN (print) 0093-4666 2013. Mycotaxon, Ltd. ISSN (online) 2154-8889 MYCOTAXON http://dx.doi.org/10.5248/123.169 Volume 123, pp. 169 181 January March 2013 Two striking Inocybe species from Yunnan Province,
Comparison of two main mycorrhizal types
 Comparison of two main mycorrhizal types VAM (Endos) Ectos Plant hosts Most vascular plants, including herbs, shrubs, trees. examples of tree you know: Maples, Ash, giant Sequoia, Sequoia, Incense Cedar
Comparison of two main mycorrhizal types VAM (Endos) Ectos Plant hosts Most vascular plants, including herbs, shrubs, trees. examples of tree you know: Maples, Ash, giant Sequoia, Sequoia, Incense Cedar
Working with Mycorrhizas in Forestry and Agriculture
 Working with Mycorrhizas in Forestry and Agriculture SUB Gdttingen 206 384661 Mark Brundrett, Neale Bougher, Bernie Dell, Tim Grove and Nick Malajczuk CONTENTS Chapter I. INTRODUCTION 1.1. MYCORRHIZAL
Working with Mycorrhizas in Forestry and Agriculture SUB Gdttingen 206 384661 Mark Brundrett, Neale Bougher, Bernie Dell, Tim Grove and Nick Malajczuk CONTENTS Chapter I. INTRODUCTION 1.1. MYCORRHIZAL
Overview. Revised through 30 June Initial Groups ("naked-eye" characters)
") Overview Revised through 30 June 2010 Initial Groups ("naked-eye" characters) Plants essentially leafless, consisting of strongly inclined, highly asymmetric capsules on a stout papillose seta; the "bug-on-a-stick"
Overview Revised through 30 June 2010 Initial Groups ("naked-eye" characters) Plants essentially leafless, consisting of strongly inclined, highly asymmetric capsules on a stout papillose seta; the "bug-on-a-stick"
This thesis is an outcome of efforts made on studies on the diversity, ecology and
 Chapter V SUMMARY This thesis is an outcome of efforts made on studies on the diversity, ecology and activities of microfungi associated with some medicinal plants of the forests of Western Ghats in Goa,
Chapter V SUMMARY This thesis is an outcome of efforts made on studies on the diversity, ecology and activities of microfungi associated with some medicinal plants of the forests of Western Ghats in Goa,
PHYLOGENY AND SYSTEMATICS
 AP BIOLOGY EVOLUTION/HEREDITY UNIT Unit 1 Part 11 Chapter 26 Activity #15 NAME DATE PERIOD PHYLOGENY AND SYSTEMATICS PHYLOGENY Evolutionary history of species or group of related species SYSTEMATICS Study
AP BIOLOGY EVOLUTION/HEREDITY UNIT Unit 1 Part 11 Chapter 26 Activity #15 NAME DATE PERIOD PHYLOGENY AND SYSTEMATICS PHYLOGENY Evolutionary history of species or group of related species SYSTEMATICS Study
(Pl. VI Fig. 36) Ramaria maculatipes sp. nov.
 Ramaria maculatipes sp. nov.") 103 Ramaria maculatipes sp. nov. (Pl. VI Fig. 36) Basidiocarpia terrestria 10 cm alta 6 cm crassa, stipite simplici 2.0-4.0 x 1.5-2.0 cm, e basi usque ad septuplo sursum ramificantia, apicibus polynodulosis,
103 Ramaria maculatipes sp. nov. (Pl. VI Fig. 36) Basidiocarpia terrestria 10 cm alta 6 cm crassa, stipite simplici 2.0-4.0 x 1.5-2.0 cm, e basi usque ad septuplo sursum ramificantia, apicibus polynodulosis,
PHYTOTAXA ISSN (online edition)
") Phytotaxa 175 (3): 133 140 www.mapress.com/phytotaxa/ ISSN 1179-3155 (print edition) Copyright 2014 Magnolia Press Article PHYTOTAXA ISSN 1179-3163 (online edition) http://dx.doi.org/10.11646/phytotaxa.175.3.2
Phytotaxa 175 (3): 133 140 www.mapress.com/phytotaxa/ ISSN 1179-3155 (print edition) Copyright 2014 Magnolia Press Article PHYTOTAXA ISSN 1179-3163 (online edition) http://dx.doi.org/10.11646/phytotaxa.175.3.2
Literature. Morphology. Morphology of the mycorrhizal system. Morphology of the unramified ends
 Literature references Müller WR, Rauscher T, Agerer R, Chevalier G (1996) Tuber aestivum Vitt. + Corylus avellana L.Descr Ectomyc 1: 167-172. Rauscher T, Müller WR, Chevalier G, Agerer R (1996) Tuber aestivum.
Literature references Müller WR, Rauscher T, Agerer R, Chevalier G (1996) Tuber aestivum Vitt. + Corylus avellana L.Descr Ectomyc 1: 167-172. Rauscher T, Müller WR, Chevalier G, Agerer R (1996) Tuber aestivum.
Clitopilus chalybescens, a new species from Thailand
 Fungal Diversity Clitopilus chalybescens, a new species from Thailand Timothy J. Baroni t, Dennis E. Desjardin 2 and Nigel Hywel-Jones 3 'Department of Biological Sciences, State University of New York
Fungal Diversity Clitopilus chalybescens, a new species from Thailand Timothy J. Baroni t, Dennis E. Desjardin 2 and Nigel Hywel-Jones 3 'Department of Biological Sciences, State University of New York
Phylogenetic diversity and conservation
 Phylogenetic diversity and conservation Dan Faith The Australian Museum Applied ecology and human dimensions in biological conservation Biota Program/ FAPESP Nov. 9-10, 2009 BioGENESIS Providing an evolutionary
Phylogenetic diversity and conservation Dan Faith The Australian Museum Applied ecology and human dimensions in biological conservation Biota Program/ FAPESP Nov. 9-10, 2009 BioGENESIS Providing an evolutionary
Digital ESF. SUNY College of Environmental Science and Forestry. Max Hermanson. Silus Weckel. Alex Kozisky.
 SUNY College of Environmental Science and Forestry Digital Commons @ ESF Cranberry Lake Biological Station Environmental and Forest Biology 2017 Session D, 2017 First Place: Under the Sphagnum: An Observational
SUNY College of Environmental Science and Forestry Digital Commons @ ESF Cranberry Lake Biological Station Environmental and Forest Biology 2017 Session D, 2017 First Place: Under the Sphagnum: An Observational
Biogeography expands:
 Biogeography expands: Phylogeography Ecobiogeography Due to advances in DNA sequencing and fingerprinting methods, historical biogeography has recently begun to integrate relationships of populations within
Biogeography expands: Phylogeography Ecobiogeography Due to advances in DNA sequencing and fingerprinting methods, historical biogeography has recently begun to integrate relationships of populations within
Psathyloma, a new genus in Hymenogastraceae described from New Zealand
 In Press at Mycologia, preliminary version published on January 7, 2016 as doi:10.3852/15-143 Psathyloma sp. nov. Psathyloma, a new genus in Hymenogastraceae described from New Zealand Karl Soop 1 Swedish
In Press at Mycologia, preliminary version published on January 7, 2016 as doi:10.3852/15-143 Psathyloma sp. nov. Psathyloma, a new genus in Hymenogastraceae described from New Zealand Karl Soop 1 Swedish
Mycological Notes 7: Resupinatus species in New Zealand
 Mycological Notes 7: Resupinatus species in New Zealand Jerry Cooper, June 2012 Resupinatus is a genus of small pleurotoid or cupulate dark coloured fungi on wood. Stevenson had a fairly broad concept
Mycological Notes 7: Resupinatus species in New Zealand Jerry Cooper, June 2012 Resupinatus is a genus of small pleurotoid or cupulate dark coloured fungi on wood. Stevenson had a fairly broad concept
Two remarkable xylariaceous ascomycetes associated with elephant dung
 Two remarkable xylariaceous ascomycetes associated with elephant dung Deepna Latha KP and Manimohan P* Department of Botany, University of Calicut, Kerala, 673 635, India Deepna Latha KP, Manimohan P 2012
Two remarkable xylariaceous ascomycetes associated with elephant dung Deepna Latha KP and Manimohan P* Department of Botany, University of Calicut, Kerala, 673 635, India Deepna Latha KP, Manimohan P 2012
UoN, CAS, DBSC BIOL102 lecture notes by: Dr. Mustafa A. Mansi. The Phylogenetic Systematics (Phylogeny and Systematics)
") - Phylogeny? - Systematics? The Phylogenetic Systematics (Phylogeny and Systematics) - Phylogenetic systematics? Connection between phylogeny and classification. - Phylogenetic systematics informs the
- Phylogeny? - Systematics? The Phylogenetic Systematics (Phylogeny and Systematics) - Phylogenetic systematics? Connection between phylogeny and classification. - Phylogenetic systematics informs the
Basidiomycota. Botany 201 Laboratory Spring 2007
 Botany 201 Laboratory Spring 2007 Basidiomycota As was the case of the Ascomycota, this phylum represents a very variable group of fungi. This only characteristic that is common to all species in this
Botany 201 Laboratory Spring 2007 Basidiomycota As was the case of the Ascomycota, this phylum represents a very variable group of fungi. This only characteristic that is common to all species in this
1 Towards Ecological Relevance Progress and Pitfalls in the Path Towards an Understanding of Mycorrhizal Functions in Nature... 3 D.J.
 Contents Section A: Introduction 1 Towards Ecological Relevance Progress and Pitfalls in the Path Towards an Understanding of Mycorrhizal Functions in Nature... 3 D.J. Read 1.1 Summary.............................
Contents Section A: Introduction 1 Towards Ecological Relevance Progress and Pitfalls in the Path Towards an Understanding of Mycorrhizal Functions in Nature... 3 D.J. Read 1.1 Summary.............................
Lecture 11 Friday, October 21, 2011
 Lecture 11 Friday, October 21, 2011 Phylogenetic tree (phylogeny) Darwin and classification: In the Origin, Darwin said that descent from a common ancestral species could explain why the Linnaean system
Lecture 11 Friday, October 21, 2011 Phylogenetic tree (phylogeny) Darwin and classification: In the Origin, Darwin said that descent from a common ancestral species could explain why the Linnaean system
Horizontal gene transfer from trees to ectomycorrhizal fungi: Lessons from laboratory and host plant liberation experiments
 Horizontal gene transfer from trees to ectomycorrhizal fungi: Lessons from laboratory and host plant liberation experiments Dr. Uwe Nehls 1,2, Dr. Chi Zhang 1, Dr. Mika Tarkka 1, Andrea Bock 1 1: University
Horizontal gene transfer from trees to ectomycorrhizal fungi: Lessons from laboratory and host plant liberation experiments Dr. Uwe Nehls 1,2, Dr. Chi Zhang 1, Dr. Mika Tarkka 1, Andrea Bock 1 1: University
HISTOCHEMICAL STUDIES ON THE DEVELOPMENT OF CARPOPHORE OF POLYPORELLUS BRUMALIS (PERS. EX FR.) KARST.
 KARST.") J. Gen. Appl. Microbiol., 21, 211--216 (1975) HISTOCHEMICAL STUDIES ON THE DEVELOPMENT OF CARPOPHORE OF POLYPORELLUS BRUMALIS (PERS. EX FR.) KARST. MASAHIKO OKUNISHI AND KAZUO KOMAGATAI Central Research
J. Gen. Appl. Microbiol., 21, 211--216 (1975) HISTOCHEMICAL STUDIES ON THE DEVELOPMENT OF CARPOPHORE OF POLYPORELLUS BRUMALIS (PERS. EX FR.) KARST. MASAHIKO OKUNISHI AND KAZUO KOMAGATAI Central Research
Phylogenomics. Jeffrey P. Townsend Department of Ecology and Evolutionary Biology Yale University. Tuesday, January 29, 13
 Phylogenomics Jeffrey P. Townsend Department of Ecology and Evolutionary Biology Yale University How may we improve our inferences? How may we improve our inferences? Inferences Data How may we improve
Phylogenomics Jeffrey P. Townsend Department of Ecology and Evolutionary Biology Yale University How may we improve our inferences? How may we improve our inferences? Inferences Data How may we improve
following citations: 7832 Correction numbered or omitted in the paper. The citation referred to by be replaced with the following citations:
 7832 Correction Proc. Natl. Acad. Sci. USA 91 (1994) Evolution. In the article "Ancient and recent patterns of geographic speciation in the oyster mushroom Pleurotus revealed by phylogenetic analysis of
7832 Correction Proc. Natl. Acad. Sci. USA 91 (1994) Evolution. In the article "Ancient and recent patterns of geographic speciation in the oyster mushroom Pleurotus revealed by phylogenetic analysis of
Rapid evolution of the cerebellum in humans and other great apes
 Rapid evolution of the cerebellum in humans and other great apes Article Accepted Version Barton, R. A. and Venditti, C. (2014) Rapid evolution of the cerebellum in humans and other great apes. Current
Rapid evolution of the cerebellum in humans and other great apes Article Accepted Version Barton, R. A. and Venditti, C. (2014) Rapid evolution of the cerebellum in humans and other great apes. Current
Harvesting and harnessing data for biogeographical research
 How do we know what grows where? Harvesting and harnessing data for biogeographical research A. Geography Tree B. Species Tree inventories and surveys natural areas, preserves, state forests, private properties
How do we know what grows where? Harvesting and harnessing data for biogeographical research A. Geography Tree B. Species Tree inventories and surveys natural areas, preserves, state forests, private properties
InDel 3-5. InDel 8-9. InDel 3-5. InDel 8-9. InDel InDel 8-9
 Lecture 5 Alignment I. Introduction. For sequence data, the process of generating an alignment establishes positional homologies; that is, alignment provides the identification of homologous phylogenetic
Lecture 5 Alignment I. Introduction. For sequence data, the process of generating an alignment establishes positional homologies; that is, alignment provides the identification of homologous phylogenetic
Using phylogenetics to estimate species divergence times... Basics and basic issues for Bayesian inference of divergence times (plus some digression)
") Using phylogenetics to estimate species divergence times... More accurately... Basics and basic issues for Bayesian inference of divergence times (plus some digression) "A comparison of the structures
Using phylogenetics to estimate species divergence times... More accurately... Basics and basic issues for Bayesian inference of divergence times (plus some digression) "A comparison of the structures
Impact of increased inorganic nitrogen deposition on the mycorrhizal community
 Eastern CANUSA Forest Science Conference Impact of increased inorganic nitrogen deposition on the mycorrhizal community Adam Bordeleau, Hubert Morin, Sergio Rossi et Daniel Houle 1 Ectomycorrhiza Symbiotic
Eastern CANUSA Forest Science Conference Impact of increased inorganic nitrogen deposition on the mycorrhizal community Adam Bordeleau, Hubert Morin, Sergio Rossi et Daniel Houle 1 Ectomycorrhiza Symbiotic
Trichaptum (Basidiomycota, Polyporaceae) in China
 in China") Mycosystema 菌物学报 15 July 2008, 27(4): 510-514 jwxt@im.ac.cn ISSN1672-6472 CN11-5180Q 2008 Institute of Microbiology, CAS, all rights reserved. Trichaptum (Basidiomycota, Polyporaceae) in China DAI Yu-Cheng
Mycosystema 菌物学报 15 July 2008, 27(4): 510-514 jwxt@im.ac.cn ISSN1672-6472 CN11-5180Q 2008 Institute of Microbiology, CAS, all rights reserved. Trichaptum (Basidiomycota, Polyporaceae) in China DAI Yu-Cheng
MiGA: The Microbial Genome Atlas
 December 12 th 2017 MiGA: The Microbial Genome Atlas Jim Cole Center for Microbial Ecology Dept. of Plant, Soil & Microbial Sciences Michigan State University East Lansing, Michigan U.S.A. Where I m From
December 12 th 2017 MiGA: The Microbial Genome Atlas Jim Cole Center for Microbial Ecology Dept. of Plant, Soil & Microbial Sciences Michigan State University East Lansing, Michigan U.S.A. Where I m From
Taxonomy of Pluteus eugraptus and morphologically similar taxa
 Mycologia, 103(3), 2011, pp. 646 655. DOI: 10.3852/10-280 # 2011 by The Mycological Society of America, Lawrence, KS 66044-8897 Taxonomy of Pluteus eugraptus and morphologically similar taxa Alfredo Justo
Mycologia, 103(3), 2011, pp. 646 655. DOI: 10.3852/10-280 # 2011 by The Mycological Society of America, Lawrence, KS 66044-8897 Taxonomy of Pluteus eugraptus and morphologically similar taxa Alfredo Justo
Supplemental Data. Perea-Resa et al. Plant Cell. (2012) /tpc
 /tpc") Supplemental Data. Perea-Resa et al. Plant Cell. (22)..5/tpc.2.3697 Sm Sm2 Supplemental Figure. Sequence alignment of Arabidopsis LSM proteins. Alignment of the eleven Arabidopsis LSM proteins. Sm and
Supplemental Data. Perea-Resa et al. Plant Cell. (22)..5/tpc.2.3697 Sm Sm2 Supplemental Figure. Sequence alignment of Arabidopsis LSM proteins. Alignment of the eleven Arabidopsis LSM proteins. Sm and
A new species of Leucopaxillus BOURS. (Agaricales) from Western Australia
 from Western Australia") A new species of Leucopaxillus BOURS. (Agaricales) from Western Australia N. L. BOUGHER Botany Department, University of Western Australia, Nedlands, W. A. 6009. Abstract. - Leucopaxillus lilacinus is
A new species of Leucopaxillus BOURS. (Agaricales) from Western Australia N. L. BOUGHER Botany Department, University of Western Australia, Nedlands, W. A. 6009. Abstract. - Leucopaxillus lilacinus is
Title ghost-tree: creating hybrid-gene phylogenetic trees for diversity analyses
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 Title ghost-tree: creating hybrid-gene phylogenetic trees for diversity analyses
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 Title ghost-tree: creating hybrid-gene phylogenetic trees for diversity analyses
Hirticlavula elegans, a new clavarioid fungus from Scandinavia
 Karstenia 54: 1 8, 2014 Hirticlavula elegans, a new clavarioid fungus from Scandinavia JENS H. PETERSEN, MARIE L. DAVEY and THOMAS LÆSSØE PETERSEN, J.H., DAVEY, M.L. & LÆSSØE, T. 2014: Hirticlavula elegans,
Karstenia 54: 1 8, 2014 Hirticlavula elegans, a new clavarioid fungus from Scandinavia JENS H. PETERSEN, MARIE L. DAVEY and THOMAS LÆSSØE PETERSEN, J.H., DAVEY, M.L. & LÆSSØE, T. 2014: Hirticlavula elegans,
Lack of mutualisms as barrier for Pinaceae invasion* Biological invasions are a problem
 Lack of mutualisms as barrier for Pinaceae invasion* Martin A. Nuñez 1, Thomas R. Horton 2, & Daniel Simberloff 1 1 Dept. of Ecology and Evolutionary Biology, The University of Tennessee 2 Department of
Lack of mutualisms as barrier for Pinaceae invasion* Martin A. Nuñez 1, Thomas R. Horton 2, & Daniel Simberloff 1 1 Dept. of Ecology and Evolutionary Biology, The University of Tennessee 2 Department of
Phylogeny of Pluteus section Celluloderma including eight new species from Brazil
 Mycologia, 107(6), 2015, pp. 1205 1220. DOI: 10.3852/14-312 # 2015 by The Mycological Society of America, Lawrence, KS 66044-8897 Phylogeny of Pluteus section Celluloderma including eight new species from
Mycologia, 107(6), 2015, pp. 1205 1220. DOI: 10.3852/14-312 # 2015 by The Mycological Society of America, Lawrence, KS 66044-8897 Phylogeny of Pluteus section Celluloderma including eight new species from
Mycorrhiza Fungus + Plant Host (Root)
") Mycorrhiza Fungus + Plant Host (Root) Root Anatomy Mycorrhizal fungi Cryptomycota http://www.mykoweb.com/articles/index.html#apm1_4 Summary Mycorrhizal symbioses are mutualistic Fungal benefits carbohydrates
Mycorrhiza Fungus + Plant Host (Root) Root Anatomy Mycorrhizal fungi Cryptomycota http://www.mykoweb.com/articles/index.html#apm1_4 Summary Mycorrhizal symbioses are mutualistic Fungal benefits carbohydrates
Chapters 25 and 26. Searching for Homology. Phylogeny
 Chapters 25 and 26 The Origin of Life as we know it. Phylogeny traces evolutionary history of taxa Systematics- analyzes relationships (modern and past) of organisms Figure 25.1 A gallery of fossils The
Chapters 25 and 26 The Origin of Life as we know it. Phylogeny traces evolutionary history of taxa Systematics- analyzes relationships (modern and past) of organisms Figure 25.1 A gallery of fossils The
Yinglong Chen BSc. (Biology), MSc. (Microbiology)
, MSc. (Microbiology)") Optimization of Scleroderma spore inoculum for Eucalyptus nurseries in south China By Yinglong Chen BSc. (Biology), MSc. (Microbiology) This thesis is submitted in fulfilment of the requirements for the
Optimization of Scleroderma spore inoculum for Eucalyptus nurseries in south China By Yinglong Chen BSc. (Biology), MSc. (Microbiology) This thesis is submitted in fulfilment of the requirements for the
Bayesian Inference using Markov Chain Monte Carlo in Phylogenetic Studies
 Bayesian Inference using Markov Chain Monte Carlo in Phylogenetic Studies 1 What is phylogeny? Essay written for the course in Markov Chains 2004 Torbjörn Karfunkel Phylogeny is the evolutionary development
Bayesian Inference using Markov Chain Monte Carlo in Phylogenetic Studies 1 What is phylogeny? Essay written for the course in Markov Chains 2004 Torbjörn Karfunkel Phylogeny is the evolutionary development
Some Agarics new to India
 Some Agarics new to India P. MANIMOHAN & K.M. LEELAVATHY Department of Botany, University of Calicut, Kerala 673 635, India MANIMOHAN, P. & K.M. LEELAVATHY (1989). Some Agarics new to India. - SYD- OWIA
Some Agarics new to India P. MANIMOHAN & K.M. LEELAVATHY Department of Botany, University of Calicut, Kerala 673 635, India MANIMOHAN, P. & K.M. LEELAVATHY (1989). Some Agarics new to India. - SYD- OWIA
Community Interactions. Community An assemblage of all the populations interacting in an area
 Community Interactions Community An assemblage of all the populations interacting in an area Populations are affected by: Available living space habitat Resource Availability niche Species interactions
Community Interactions Community An assemblage of all the populations interacting in an area Populations are affected by: Available living space habitat Resource Availability niche Species interactions
Phylogenies Scores for Exhaustive Maximum Likelihood and Parsimony Scores Searches
 Int. J. Bioinformatics Research and Applications, Vol. x, No. x, xxxx Phylogenies Scores for Exhaustive Maximum Likelihood and s Searches Hyrum D. Carroll, Perry G. Ridge, Mark J. Clement, Quinn O. Snell
Int. J. Bioinformatics Research and Applications, Vol. x, No. x, xxxx Phylogenies Scores for Exhaustive Maximum Likelihood and s Searches Hyrum D. Carroll, Perry G. Ridge, Mark J. Clement, Quinn O. Snell
Bioinformatics tools for phylogeny and visualization. Yanbin Yin
 Bioinformatics tools for phylogeny and visualization Yanbin Yin 1 Homework assignment 5 1. Take the MAFFT alignment http://cys.bios.niu.edu/yyin/teach/pbb/purdue.cellwall.list.lignin.f a.aln as input and
Bioinformatics tools for phylogeny and visualization Yanbin Yin 1 Homework assignment 5 1. Take the MAFFT alignment http://cys.bios.niu.edu/yyin/teach/pbb/purdue.cellwall.list.lignin.f a.aln as input and
MYCOTAXON. Volume 96, pp April June 2006
 MYCOTAXON Volume 96, pp. 133 140 April June 2006 The sequestrate genus Rhodactina (Basidiomycota, Boletales) in northern Thailand Zhu L. Yang 1, James M. Trappe 2, Manfred Binder 3, Rarunee Sanmee 4, Pipob
MYCOTAXON Volume 96, pp. 133 140 April June 2006 The sequestrate genus Rhodactina (Basidiomycota, Boletales) in northern Thailand Zhu L. Yang 1, James M. Trappe 2, Manfred Binder 3, Rarunee Sanmee 4, Pipob
Reconstructing the history of lineages
 Reconstructing the history of lineages Class outline Systematics Phylogenetic systematics Phylogenetic trees and maps Class outline Definitions Systematics Phylogenetic systematics/cladistics Systematics
Reconstructing the history of lineages Class outline Systematics Phylogenetic systematics Phylogenetic trees and maps Class outline Definitions Systematics Phylogenetic systematics/cladistics Systematics
A new species of Bertiella (Melanommataceae) from Brazil and a key to accepted species
 from Brazil and a key to accepted species") Mycosphere 8 (4): 392 396 (2017) www.mycosphere.org ISSN 2077 7019 Article Doi 10.5943/mycosphere/8/4/1 Copyright Guizhou Academy of Agricultural Sciences A new species of Bertiella (Melanommataceae) from
Mycosphere 8 (4): 392 396 (2017) www.mycosphere.org ISSN 2077 7019 Article Doi 10.5943/mycosphere/8/4/1 Copyright Guizhou Academy of Agricultural Sciences A new species of Bertiella (Melanommataceae) from
Need for systematics. Applications of systematics. Linnaeus plus Darwin. Approaches in systematics. Principles of cladistics
 Topics Need for systematics Applications of systematics Linnaeus plus Darwin Approaches in systematics Principles of cladistics Systematics pp. 474-475. Systematics - Study of diversity and evolutionary
Topics Need for systematics Applications of systematics Linnaeus plus Darwin Approaches in systematics Principles of cladistics Systematics pp. 474-475. Systematics - Study of diversity and evolutionary
Geography of Evolution
 Geography of Evolution Biogeography - the study of the geographic distribution of organisms. The current distribution of organisms can be explained by historical events and current climatic patterns. Darwin
Geography of Evolution Biogeography - the study of the geographic distribution of organisms. The current distribution of organisms can be explained by historical events and current climatic patterns. Darwin
Integrative Biology 200 "PRINCIPLES OF PHYLOGENETICS" Spring 2018 University of California, Berkeley
 Integrative Biology 200 "PRINCIPLES OF PHYLOGENETICS" Spring 2018 University of California, Berkeley B.D. Mishler Feb. 14, 2018. Phylogenetic trees VI: Dating in the 21st century: clocks, & calibrations;
Integrative Biology 200 "PRINCIPLES OF PHYLOGENETICS" Spring 2018 University of California, Berkeley B.D. Mishler Feb. 14, 2018. Phylogenetic trees VI: Dating in the 21st century: clocks, & calibrations;
Amira A. AL-Hosary PhD of infectious diseases Department of Animal Medicine (Infectious Diseases) Faculty of Veterinary Medicine Assiut
 Faculty of Veterinary Medicine Assiut") Amira A. AL-Hosary PhD of infectious diseases Department of Animal Medicine (Infectious Diseases) Faculty of Veterinary Medicine Assiut University-Egypt Phylogenetic analysis Phylogenetic Basics: Biological
Amira A. AL-Hosary PhD of infectious diseases Department of Animal Medicine (Infectious Diseases) Faculty of Veterinary Medicine Assiut University-Egypt Phylogenetic analysis Phylogenetic Basics: Biological
Creating an e-flora for South Africa
 SANBI POLICY DOCUMENT DIVISION: Biosystematics Research and Biodiversity Collections EFFECTIVE DATE: 1 April 2014 Compiler: Marianne le Roux & Janine Victor POLICY NUMBER: LAST AMENDED: Creating an e-flora
SANBI POLICY DOCUMENT DIVISION: Biosystematics Research and Biodiversity Collections EFFECTIVE DATE: 1 April 2014 Compiler: Marianne le Roux & Janine Victor POLICY NUMBER: LAST AMENDED: Creating an e-flora
Last Time. Biology of FUNgi. Lecture 24 Community ecology - what rules fungal communities. ! We wrapped up the good, the bad and the ugly.
 Biology of FUNgi Lecture 24 Community ecology - what rules fungal communities Last Time! We wrapped up the good, the bad and the ugly.! Systemic fungal infections: blastomycosis, coccidiomycosis, cryptococcosis,
Biology of FUNgi Lecture 24 Community ecology - what rules fungal communities Last Time! We wrapped up the good, the bad and the ugly.! Systemic fungal infections: blastomycosis, coccidiomycosis, cryptococcosis,
a-dB. Code assigned:
 This form should be used for all taxonomic proposals. Please complete all those modules that are applicable (and then delete the unwanted sections). For guidance, see the notes written in blue and the
This form should be used for all taxonomic proposals. Please complete all those modules that are applicable (and then delete the unwanted sections). For guidance, see the notes written in blue and the
Taxonomy. Content. How to determine & classify a species. Phylogeny and evolution
 Taxonomy Content Why Taxonomy? How to determine & classify a species Domains versus Kingdoms Phylogeny and evolution Why Taxonomy? Classification Arrangement in groups or taxa (taxon = group) Nomenclature
Taxonomy Content Why Taxonomy? How to determine & classify a species Domains versus Kingdoms Phylogeny and evolution Why Taxonomy? Classification Arrangement in groups or taxa (taxon = group) Nomenclature
"PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200 Spring 2014 University of California, Berkeley
 "PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200 Spring 2014 University of California, Berkeley D.D. Ackerly April 16, 2014. Community Ecology and Phylogenetics Readings: Cavender-Bares,
"PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200 Spring 2014 University of California, Berkeley D.D. Ackerly April 16, 2014. Community Ecology and Phylogenetics Readings: Cavender-Bares,
Department of Botany, University of Dhaka, Dhaka 1000, Bangladesh. Key words: Seaweeds, Marine algae, Kallymenia spp., St. Martin's Is.
 Bangladesh J. Bot. 37(2): 173-178, 2008 (December) MARINE ALGAE OF THE ST. MARTIN S ISLAND, BANGLADESH. VI. NEW RECORDS OF SPECIES OF THE GENUS KALLYMENIA J. AG. (RHODOPHYTA) ABDUL AZIZ, A.K.M. NURUL ISLAM
Bangladesh J. Bot. 37(2): 173-178, 2008 (December) MARINE ALGAE OF THE ST. MARTIN S ISLAND, BANGLADESH. VI. NEW RECORDS OF SPECIES OF THE GENUS KALLYMENIA J. AG. (RHODOPHYTA) ABDUL AZIZ, A.K.M. NURUL ISLAM
NEW CHARACTERISTICS FOR MORPHOTAXONOMY OF GIGASPORA SPECIES BELONGING TO ARBUSCULAR MYCORRHIZAL FUNGI
 SHARDA W. KHADE J. Plant Develop. 18(2011): 71-80 NEW CHARACTERISTICS FOR MORPHOTAXONOMY OF GIGASPORA SPECIES BELONGING TO ARBUSCULAR MYCORRHIZAL FUNGI SHARDA W. KHADE 1 Abstract: New characteristics for
SHARDA W. KHADE J. Plant Develop. 18(2011): 71-80 NEW CHARACTERISTICS FOR MORPHOTAXONOMY OF GIGASPORA SPECIES BELONGING TO ARBUSCULAR MYCORRHIZAL FUNGI SHARDA W. KHADE 1 Abstract: New characteristics for
The process by which the genetic structure of populations changes over time.
 Evolution The process by which the genetic structure of populations changes over time. Divergent evolution is the accumulation of differences between groups which can lead to the formation of new species.
Evolution The process by which the genetic structure of populations changes over time. Divergent evolution is the accumulation of differences between groups which can lead to the formation of new species.
Cleistocybe, a new genus of Agaricales
 Mycoscience (2007) 48:282 289 The Mycological Society of Japan and Springer 2007 DOI 10.1007/s10267-007-0365-5 FULL PAPER Joseph F. Ammirati Andrew D. Parker P. Brandon Matheny Cleistocybe, a new genus
Mycoscience (2007) 48:282 289 The Mycological Society of Japan and Springer 2007 DOI 10.1007/s10267-007-0365-5 FULL PAPER Joseph F. Ammirati Andrew D. Parker P. Brandon Matheny Cleistocybe, a new genus
8/23/2014. Phylogeny and the Tree of Life
 Phylogeny and the Tree of Life Chapter 26 Objectives Explain the following characteristics of the Linnaean system of classification: a. binomial nomenclature b. hierarchical classification List the major
Phylogeny and the Tree of Life Chapter 26 Objectives Explain the following characteristics of the Linnaean system of classification: a. binomial nomenclature b. hierarchical classification List the major
Phaeocalicium populneum
 Phaeocalicium populneum markpowell222@btinternet.com After conducting a survey of the RHS garden at Wisley on 18 th August 2018, Fay Newbery kindly showed me the colony of P. populneum at Esher Common.
Phaeocalicium populneum markpowell222@btinternet.com After conducting a survey of the RHS garden at Wisley on 18 th August 2018, Fay Newbery kindly showed me the colony of P. populneum at Esher Common.
Adaptive Radiations. Future of Molecular Systematics. Phylogenetic Ecology. Phylogenetic Ecology. ... Systematics meets Ecology...
 Future of Molecular Systematics... Systematics meets Ecology... 1. Biogeography 2. Ecology 3. Genomics Ecology and phylogenetics intertwined in a number of new fields of study called Phylogenetic Ecology:
Future of Molecular Systematics... Systematics meets Ecology... 1. Biogeography 2. Ecology 3. Genomics Ecology and phylogenetics intertwined in a number of new fields of study called Phylogenetic Ecology:
MYCOTAXON. Vol. XI, No. 2, pp July-September 1980 THE FRUITING AND DEVELOPMENT OF RHODOTUS PALMATUS IN CULTURE ORSON K. MILLER, JR.
 MYCOTAXON Vol. XI, No. 2, pp. 409-419 July-September 1980 THE FRUITING AND DEVELOPMENT OF RHODOTUS PALMATUS IN CULTURE ORSON K. MILLER, JR. Department of Biology Virginia Polytechnic Institute & state
MYCOTAXON Vol. XI, No. 2, pp. 409-419 July-September 1980 THE FRUITING AND DEVELOPMENT OF RHODOTUS PALMATUS IN CULTURE ORSON K. MILLER, JR. Department of Biology Virginia Polytechnic Institute & state
a,bD (modules 1 and 10 are required)
") This form should be used for all taxonomic proposals. Please complete all those modules that are applicable (and then delete the unwanted sections). For guidance, see the notes written in blue and the
This form should be used for all taxonomic proposals. Please complete all those modules that are applicable (and then delete the unwanted sections). For guidance, see the notes written in blue and the
Notes on the Algae -- III
 Butler University Botanical Studies Volume 8 Article 10 Notes on the Algae -- III William A. Daily Follow this and additional works at: http://digitalcommons.butler.edu/botanical The Butler University
Butler University Botanical Studies Volume 8 Article 10 Notes on the Algae -- III William A. Daily Follow this and additional works at: http://digitalcommons.butler.edu/botanical The Butler University
OMICS Journals are welcoming Submissions
 OMICS Journals are welcoming Submissions OMICS International welcomes submissions that are original and technically so as to serve both the developing world and developed countries in the best possible
OMICS Journals are welcoming Submissions OMICS International welcomes submissions that are original and technically so as to serve both the developing world and developed countries in the best possible
Key to West Coast Spartina Species Based on Vegetative Characters
 Glossary Accuminate - tapering to a sharp tip and having concave sides just before the tip. Caespitose - growing in tufts. Glabrous - having a smooth, shiny surface. Internode - the region of a stem between
Glossary Accuminate - tapering to a sharp tip and having concave sides just before the tip. Caespitose - growing in tufts. Glabrous - having a smooth, shiny surface. Internode - the region of a stem between
The diversity of plant communities mediates mycorrhizal fungal diversity
 The diversity of plant communities mediates mycorrhizal fungal diversity Or, How graduate school is going to be way harder than I thought Marlene Tyner, University of Michigan R. Michael Miller, Argonne
The diversity of plant communities mediates mycorrhizal fungal diversity Or, How graduate school is going to be way harder than I thought Marlene Tyner, University of Michigan R. Michael Miller, Argonne
Postgraduate teaching for the next generation of taxonomists
 Postgraduate teaching for the next generation of taxonomists Alfried P. Vogler Professor of Molecular Systematics Imperial College London and Natural History Museum MSc in Taxonomy and Biodiversity MRes
Postgraduate teaching for the next generation of taxonomists Alfried P. Vogler Professor of Molecular Systematics Imperial College London and Natural History Museum MSc in Taxonomy and Biodiversity MRes
INTRODUCTION budding, binary fission hyphae mycelium Figure 1.
 INTRODUCTION Although most of our work in this lab is done on bacteria, fungi are nonetheless an important aspect in microbiology. Besides being important food providers, fungi play central roles in recycling
INTRODUCTION Although most of our work in this lab is done on bacteria, fungi are nonetheless an important aspect in microbiology. Besides being important food providers, fungi play central roles in recycling
Fungi are absorptive heterotrophs that secrete digestive enzymes and are major decomposers of dead organic material
 Fungi 1 2002 Prentice Hall, Inc The scarlet hood (Hygrocybe coccinea) Fungi are absorptive heterotrophs that secrete digestive enzymes and are major decomposers of dead organic material 2 Animals 3 Myxozoa
Fungi 1 2002 Prentice Hall, Inc The scarlet hood (Hygrocybe coccinea) Fungi are absorptive heterotrophs that secrete digestive enzymes and are major decomposers of dead organic material 2 Animals 3 Myxozoa
Community phylogenetics review/quiz
 Community phylogenetics review/quiz A. This pattern represents and is a consequent of. Most likely to observe this at phylogenetic scales. B. This pattern represents and is a consequent of. Most likely
Community phylogenetics review/quiz A. This pattern represents and is a consequent of. Most likely to observe this at phylogenetic scales. B. This pattern represents and is a consequent of. Most likely
Plant Systematics. What is Systematics? or Why Study Systematics? Botany 400. What is Systematics or Why Study Systematics?
 Plant Systematics Botany 400 http://botany.wisc.edu/courses/botany_400/ What is Systematics? or Why Kenneth J. Sytsma Melody Sain Kelsey Huisman Botany Department University of Wisconsin Pick up course
Plant Systematics Botany 400 http://botany.wisc.edu/courses/botany_400/ What is Systematics? or Why Kenneth J. Sytsma Melody Sain Kelsey Huisman Botany Department University of Wisconsin Pick up course
PLANT BIOLOGY (PBIO) Plant Biology (PBIO) 1
 Plant Biology (PBIO) 1") Plant Biology (PBIO) 1 PLANT BIOLOGY (PBIO) PBIO 1052 How Plants Shaped Our World (LN) Description: This course is an eclectic dive into the world of plants and their influence on human society. Students
Plant Biology (PBIO) 1 PLANT BIOLOGY (PBIO) PBIO 1052 How Plants Shaped Our World (LN) Description: This course is an eclectic dive into the world of plants and their influence on human society. Students