Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA
|
|
|
- Eleanore Fletcher
- 5 years ago
- Views:
Transcription
1 Online Supplemental Results Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA Structure solution and overall architecture of PPDC-3dThDP and PPDC-2HE3dThDP Previously a structure of phenylpyruvate decarboxylase of Azospirillum brasilense (AbPPDC) in complex with the cofactors ThDP and Mg 2+ was solved to a resolution of 1.5 Å (denominated PPDC-ThDP) (1). Binary complexes of AbPPDC with 3deazaThDP (PPDC-3dThDP) and 2-(1- hydroxyethyl)3deazathdp (PPDC-2HE3dThDP) were obtained by incubating AbPPDC, that was purified without addition of ThDP, with an excess of both compounds respectively. After two days of incubation the complete exchange of ThDP with 3dThDP or 2HE3dThDP was confirmed by the total loss of enzyme activity. Crystal structures of PPDC-3dThDP and PPDC-2HE3dThDP were solved to 3.2 and 1.85 Å resolution, using molecular replacement with a subunit of PPDC- ThDP (PDB 2NXW) as a search model. In both structures, clear electron density was present for the cofactor/intermediate analogues in the two active sites of the homodimer in the asymmetric unit (a.u.), and for most of the protein backbone. However, neither the residues of the N-terminal his-tag, nor the last few C-terminal residues were included in the model due to a lack of electron density. Also, in both binary complexes a large peptide region in the vicinity of the active sites, spanning residues 104 to120, could only be modeled partially due to very poorly defined electron density. The asymmetric unit of the crystal structures contains a homodimer, where each subunit adopts the archetypical pyruvate oxidase (POX) fold consisting of three distinct α/β-type domains designated PYR, R and PP domain (1). Each dimer contains two equivalent active sites which are shared between the PYR domain of one subunit and the PP domain of the neighboring subunit. Very little changes in subunit architecture are observed in PPDC-3dThDP and PPDC- 2HE3dThDP compared to PPDC-ThDP (see Supplemental Table 1S online for a list of rms deviations). The biological tetramers (2) of PPDC-3dThDP and PPDC-2HE3dThDP can be obtained from the homodimers in the a.u. through the two-fold crystallographic symmetry axis. However, the nonperpendicular arrangement of non-crystallographic axes relating the monomers in the a.u. and the crystallographic axis results in an asymmetrical tetramer assembly, best described as an asymmetrical dimer of dimers (see Figure 5a and Supplemental Table 2S online for interface surfaces). A same quaternary arrangement was also found for PPDC-ThDP (See Supplemental Table 1S online) (1). Structure solution and overall architecture of PPDC-3dThDP-PPA and PPDC-3dThDP- POVA The catalytically inert binary PPDC-3dThDP complex was subsequently used for cocrystallization with the substrates phenylpyruvic acid (PPA) and 5-phenyl-2-oxo-valeric acid (POVA). Therefore the pre-incubated PPDC-3dThDP complex was incubated for a few hours 1
2 with the substrates before crystallization setups were started. Crystal structures of the PPDC- 3dThDP-PPA and PPDC-3dThDP-POVA ternary complexes were solved to 2.15 and 1.9 Å resolution using the same molecular replacement approach as above. In both structures clear electron density was present for cofactor analogues and substrate molecules in both active sites in the a.u. Although clear electron density was now present for the loop , density was missing for the N-terminal his-tag, the C-terminal residues and a region spanning residues 331 to 340 connecting the R and PP domains. In the two ternary complexes clear electron density corresponding to an additional substrate molecule was also present in a site remote from the active site. This regulatory site is present in every subunit at the juncture of the PYR, R and PP domains, lined by residues Arg60, Arg214, Arg215, Met238, Arg240, Leu395, Met396 and Ala397. In the asymmetric units of the two ternary complexes two subunits, related by a noncrystallographic two-fold axis, form a homodimer. Relatively large differences occur in the overall geometry of the subunits of the ternary complexes (PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA) compared to the binary complexes (PPDC-ThDP, PPDC-3dThDP and PPDC- 2HE3dThDP) as exemplified by large rms- deviations upon superposition (Supplemental Table 1S online). Subunits of PPDC-3dThDP-PPA and PPDC-3dThDP-POVA on the other hand superimpose nearly perfectly. The difference in subunit geometry between the ternary and binary complexes is mainly caused by a difference in domain orientation. When the PYR and PP domains are superimposed, the R domains differ by a rotation of about 12 (Figure 4). This domain rotation is accompanied by the closing of the active sites in the ternary complexes via a large rearrangement of active site loop of the R-domain and the concomitant ordering of the active site loop of the PYR domain of the neighboring subunit. The rearrangement of both active site loops is clearly coupled since they interact with each other in the closed active site conformation. The reorganization of the two active site loops finally leads to a slight tilting of the C-terminal helix which closes further over the active site pocket. Similar to the tetramers of the binary complexes (PPDC-ThDP, PPDC-3dThDP and PPDC- 2HE3dThDP), the tetramers of the tertiary complexes (PPDC-3dThDP-PPA and PPDC-3dThDP- POVA) can be obtained from two dimers related by a two-fold crystallographic symmetry axis. There are however large differences in the assembly of the tetramers (Figure 5). For the ternary complexes, the non-crystallographic symmetry axes relating the two subunits in the dimer intersect with the crystallographic axis at an angle of 90, resulting in a dimer of dimers with pseudo 222 symmetry. In going from the asymmetrical dimer of dimers observed for the binary complexes to the symmetrical dimer of dimers of the ternary complexes, one dimer has to be rotated by about 34, vis-à-vis to the second dimer. This also has implications for the dimer-dimer interfaces. While the AC and BD interfaces are different in the asymmetrical tetramers (see Figure 5, for subunit nomenclature) these interfaces are the same for the symmetrical tetramers. Comparable to the BD interface in the asymmetrical tetramer, the AC and BD dimer-dimer interfaces in the symmetrical tetramers are formed by an extension of the six-stranded β-sheet of the R domains to the adjacent R-domains. Folding and reorganization of the active site loops, where loop of one subunit interacts with loop of the adjacent subunit, cause an increase in the monomer-monomer interface area in the ternary complexes (see Supplemental Table 2S online). On the other hand, the reorganization of the asymmetrical tetramer into a symmetrical tetramer upon substrate binding in the active and regulatory site is not accompanied by a significant change in dimer-dimer interface area. Active sites of the binary PPDC-3dThDP and PPDC-2HE3dThDP complexes 2
3 Despite the relatively low resolution of the PPDC-3dThDP structure, the 3dThDP molecules could be unambiguously modeled with full occupancy in the electron density in both active sites of the homodimer in the a.u. (Figure 2a). Upon superposition of the PPDC-3dThDP and PPDC- ThDP structures the cofactors and active site residues superimpose nearly perfectly (see (1) for a detailed description of the active site architecture of PPDC-ThDP). The 3dThDP molecules adopt the typical V conformation, with Φ T = and Φ P = (compare to ThDP in PPDC- ThDP: Φ T = and Φ P = ). The PPDC-2HE3dThDP structure shows clear electron density in the two active sites of the a.u. for both the 3deazaThDP moiety and the 1-hydroyethyl moiety of the intermediate analogue (Figure 2b). The density for the 1-hydroyethyl moiety however can be best interpreted as a mixture of the R and S enantiomers at the Cα atom, consistent with the fact that a racemic mixture was used in the co-crystallization. Consequently both enantiomers were modeled with half occupancy. Slight deviations of the dihedral angles typical for the V conformation of the cofactor are observed relative to ThDP or 3dThDP caused by the covalent addition of the hydroxyethyl moiey (Φ T = and Φ P = for the S enantiomer, and Φ T = and Φ P = for the R enantiomer). The electron density does not a priori allow to distinguish between the methyl and hydroxyl groups attached to the Cα. However, for both enantiomers the hydroxyethyl moieties were modeled with good confidence with their hydroxyl group pointing toward the 4 amino group of the aminopyrimidine ring of 3dThDP based on a distance of 2.5 Å between the N4 and the hydroxyl group. In the R configuration the methyl group is pointing toward the large hydrophobic cavity lined by Met380, Phe385, Met461, Phe465 and Phe532. This configuration thus allows straightforward modeling of the phenyl group of the physiological intermediate 2-(1-hydroxy-2-phenyl-ethyl)-ThDP in the active site (Figure 7a). The S enantiomer does not allow modeling of the phenyl moiety without generating extensive clashes with protein atoms. Therefore we conclude that the R enantiomer is the relevant species, which is used for further interpretation. Except for a small rotation of the side chain of Phe465 and Asp25 (in one active site only) no changes in side chain conformation occur in the active site of PPDC-2HE3dThDP compared to PPDC-ThDP or PPDC-3dThDP. No direct interactions are formed between any amino acid residue and the hydroxyethyl moiety. The C2α-hydroxyl group is located 2.5 Å from the cofactor s N4 amino/imino group, in an ideal orientation to form a strong interaction. Finally we find an enzyme-bound water molecule located at 3.5 Å from the C2α. This water in its turn is hydrogen bonded to the Asp25 carboxylate group and to the side chain hydroxyl of Thr71. Active sites of the ternary complexes PPDC-3dThDP-PPA and PPDC-3dThDP-POVA The PPDC-3dThDP-PPA and PPDC-3dThDP-POVA structures shows the respective substrates, phenylpyruvic acid and 5-phenyl-2-oxo-valeric acid, bound with full occupancy in the two active sites of the homodimer in the asymmetric unit (Figure 2c and d). No significant difference in cofactor analogue geometry is observed compared to PPDC-3dThDP or PPDC-ThDP (Φ T = and Φ P = for PPDC-3dThDP-PPA and Φ T = and Φ P = for PPDC-3dThDP-POVA). Substrate binding leads to a reorganization of the active site loops spanning residues 104 to 120 and 280 to 294 respectively. Folding of the loop over the reaction pocket positions two additional active site residues, His112 and His113. In this process a hydrogen bond is formed between His112 and Asp282 of the loop of the neighboring subunit. The reorganization of the second active site loop also permits some new interactions with the C-terminal helix (e.g. between the side chain of Gln536 and Arg538 and the main chain carbonyls of Asp282 and of Ala 287 and Ser288 respectively) allowing this helix to close further over the active site. 3
4 The keto-acid moieties of PAA and POVA are bound in a similar way in the active site of PPDC (Figure 2). The C2α atoms of the substrates are located about 3.4 Å from the C2 of the 3dThDP, poised for nucleophilic attack by the cofactor. The 2-keto group is located within interaction distance of His113 and the N4 of the cofactor s aminopyrimidine ring. The carboxyl oxygens are hydrogen bonded to the main chain amide of Asp25, the side chain amine of His113 and to a water molecule which is located at an extremely short distance of about 2.3 Å from the carboxyl oxygen. This water molecule is also hydrogen bonded to the side chains of Thr71 and Asp25. Asp25 also forms a hydrogen bond with His112 of the first active site loop, which in turn is hydrogen bonded to Asp282 of the second active site loop. Since Asp282 is exposed to solvent the triad of ionizable residues Asp25-His112-Asp282 could serve as a proton relay shuttling protons into the active site. Leu462 is wedged directly underneath the carboxyl group of the substrate. Interestingly this leucine is not conserved in most other ThDP-dependent decarboxylases such as PDC s and IPDC s, where it is replaced by a glutamate residue (1;2). In the latter enzymes a catalytic role has been proposed for this glutamate (3;4). The methylene groups bridging the keto-acid and aromatic moiety of the substrate are bound in a hydrophobic region lined by residues Met380, Phe385, Ala402, Met461 and Phe465 (Figure 2c and d). The phenyl group of both substrates is stacked on one face in a parallel fashion to the side chain of His112, and on the other face in an anti-parallel fashion to the side chain of Phe532. This latter interaction seems more ideal for POVA than for PPA, which might account for the higher affinity of AbPPDC for POVA (2). The outer part of the active site is mainly lined by residues Asp282 and Thr283 from the loop, and several residues from the C-terminal helix: Phe532, Gln536 and His540. The latter residues move closer over the active site when the smaller substrate PAA is bound, thus suggesting that the C-terminal helix acts as a modular switch which can adapt according to the size of the side group of the substrate. Comparison of the substrate activation mechanism of PPDC to the proposed mechanism for S. cerevisiae PDC The signal transduction pathway observed here for PPDC differs from the most commonly accepted mechanism for substrate activation in S. cerevisiae PDC. In the latter enzyme, a cysteine (Cys221) is considered as the site of covalent binding of the regulatory substrate molecule. This binding information is then proposed to be transmitted via His92, Glu91 and Trp412 to the ThDP cofactor (5;6). Since Cys221 is not conserved in PPDC the same mechanism cannot apply for this enzyme (1). The PPDC regulatory site is situated at a distance of 9 Å from the residue corresponding to Cys221 (Glu212). Although the regulatory binding site in PPDC is different from the proposed site in PDC, both signal transduction routes have in common that the residues involved in this route are located at the interface of the PYR, R and PP domains (see Supplemental Figure1 online). Possibly a signal transduction route has evolved independently (or divergently) in PDC and PPDC. On the other hand, the regulatory substrate binding pocket of PPDC seems to a certain extent conserved in PDC. The two arginine residues which form an interaction with the carboxyl of the substrate in PPDC are conserved in PDC (Arg63 and Arg224). Arg214 which forms a cation-π stacking with the aromatic moiety of the substrate in PPDC is replaced by a serine (Ser223). These residues surround a cavity which might harbor a pyruvate molecule. However, the entrance to this cavity is obstructed by the side chain of Tyr405, but a simple change in side chain rotamer could open up this site for substrate entry. The possible role of this site in PDC remains to be investigated. 4
5 References 1. Versées, W., Spaepen, S., Vanderleyden, J., and Steyaert, J. (2007) FEBS J. 274, Spaepen, S., Versées, W., Gocke, D., Pohl, M., Steyaert, J., and Vanderleyden, J. (2007) J.Bacteriol. In Press 3. Schütz, A., Golbik, R., König, S., Hübner, G., and Tittmann, K. (2005) Biochemistry 44, Sergienko, E. A. and Jordan, F. (2001) Biochemistry 40, Wang, J., Golbik, R., Seliger, B., Spinka, M., Tittmann, K., Hübner, G., and Jordan, F. (2001) Biochemistry 40, Baburina, I., Li, H., Bennion, B., Furey, W., and Jordan, F. (1998) Biochemistry 37,
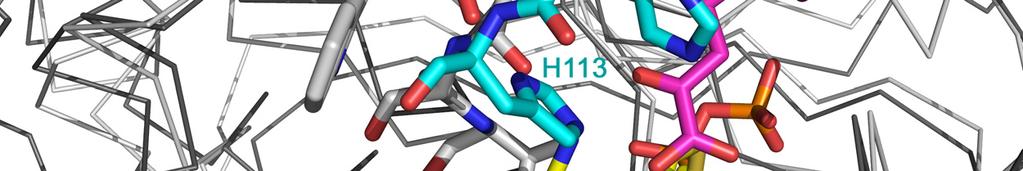
6 Figure legends: Supplemental Figure 1: Comparison of the substrate activation mechanisms of PPDC and S. cerevisiae PDC. A superposition in ribbon representation is shown of PPDC-3dThDP- POVA in dark grey and S. cerevisiae PDC (PDB 1QPB) in light grey. The residues relevant to the activation mechanism in PDC, Cys221, His92, Glu91 and Trp412, are shown as grey sticks. The corresponding residues in PPDC are shown as green and cyan sticks depending on the subunit they belong to. The cofactor analogue is shown in yellow and the substrates in the active and regulatory sites of PPDC in magenta. While both activation mechanisms are clearly different and use a different initial trigger, they both use the interface of the PYR, R and PP domains to propagate the signal from the regulatory site to the active site. 6
7 Supplemental Table 1S. Comparison of PPDC structures: Rms-deviation (in Å) upon superposition (510 Cα atoms) PPDC- 3dThDP PPDC- 2HE3dThDP PPDC- 3dThDP- PPA PPDC-ThDP Monomer-monomer Dimer-dimer Tetramer-tetramer PPDC-3dThDP Monomer-monomer Dimer-dimer Tetramer-tetramer PPDC-2HE3dThDP Monomer-monomer Dimer-dimer Tetramer-tetramer PPDC-3dThDP-PPA Monomer-monomer 0.25 Dimer-dimer 0.29 Tetramer-tetramer 0.32 PPDC- 3dThDP- POVA 7
8 Supplemental Table 2S. Tetramer architecture: accessible surface area buried in the subunit interfaces (in Å 2 ) Monomer-monomer interface Dimer-dimer interface PPDC-ThDP a PPDC-3dThDP PPDC-2HE3dThDP PPDC-3dThDP-PPA PPDC-3dThDP- POVA a Data from 1 8





9 Supplemental Figure 1 9
Properties of amino acids in proteins
 Properties of amino acids in proteins one of the primary roles of DNA (but not the only one!) is to code for proteins A typical bacterium builds thousands types of proteins, all from ~20 amino acids repeated
Properties of amino acids in proteins one of the primary roles of DNA (but not the only one!) is to code for proteins A typical bacterium builds thousands types of proteins, all from ~20 amino acids repeated
Introduction to Comparative Protein Modeling. Chapter 4 Part I
 Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Peptides And Proteins
 Kevin Burgess, May 3, 2017 1 Peptides And Proteins from chapter(s) in the recommended text A. Introduction B. omenclature And Conventions by amide bonds. on the left, right. 2 -terminal C-terminal triglycine
Kevin Burgess, May 3, 2017 1 Peptides And Proteins from chapter(s) in the recommended text A. Introduction B. omenclature And Conventions by amide bonds. on the left, right. 2 -terminal C-terminal triglycine
Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase Cwc27
 Acta Cryst. (2014). D70, doi:10.1107/s1399004714021695 Supporting information Volume 70 (2014) Supporting information for article: Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase
Acta Cryst. (2014). D70, doi:10.1107/s1399004714021695 Supporting information Volume 70 (2014) Supporting information for article: Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase
Conformational Analysis
 Conformational Analysis C01 3 C C 3 is the most stable by 0.9 kcal/mole C02 K eq = K 1-1 * K 2 = 0.45-1 * 0.048 = 0.11 C04 The intermediate in the reaction of 2 has an unfavorable syn-pentane interaction,
Conformational Analysis C01 3 C C 3 is the most stable by 0.9 kcal/mole C02 K eq = K 1-1 * K 2 = 0.45-1 * 0.048 = 0.11 C04 The intermediate in the reaction of 2 has an unfavorable syn-pentane interaction,
Advanced Certificate in Principles in Protein Structure. You will be given a start time with your exam instructions
 BIRKBECK COLLEGE (University of London) Advanced Certificate in Principles in Protein Structure MSc Structural Molecular Biology Date: Thursday, 1st September 2011 Time: 3 hours You will be given a start
BIRKBECK COLLEGE (University of London) Advanced Certificate in Principles in Protein Structure MSc Structural Molecular Biology Date: Thursday, 1st September 2011 Time: 3 hours You will be given a start
1. Amino Acids and Peptides Structures and Properties
 1. Amino Acids and Peptides Structures and Properties Chemical nature of amino acids The!-amino acids in peptides and proteins (excluding proline) consist of a carboxylic acid ( COOH) and an amino ( NH
1. Amino Acids and Peptides Structures and Properties Chemical nature of amino acids The!-amino acids in peptides and proteins (excluding proline) consist of a carboxylic acid ( COOH) and an amino ( NH
Solutions In each case, the chirality center has the R configuration
 CAPTER 25 669 Solutions 25.1. In each case, the chirality center has the R configuration. C C 2 2 C 3 C(C 3 ) 2 D-Alanine D-Valine 25.2. 2 2 S 2 d) 2 25.3. Pro,, Trp, Tyr, and is, Trp, Tyr, and is Arg,
CAPTER 25 669 Solutions 25.1. In each case, the chirality center has the R configuration. C C 2 2 C 3 C(C 3 ) 2 D-Alanine D-Valine 25.2. 2 2 S 2 d) 2 25.3. Pro,, Trp, Tyr, and is, Trp, Tyr, and is Arg,
Exam I Answer Key: Summer 2006, Semester C
 1. Which of the following tripeptides would migrate most rapidly towards the negative electrode if electrophoresis is carried out at ph 3.0? a. gly-gly-gly b. glu-glu-asp c. lys-glu-lys d. val-asn-lys
1. Which of the following tripeptides would migrate most rapidly towards the negative electrode if electrophoresis is carried out at ph 3.0? a. gly-gly-gly b. glu-glu-asp c. lys-glu-lys d. val-asn-lys
Packing of Secondary Structures
 7.88 Lecture Notes - 4 7.24/7.88J/5.48J The Protein Folding and Human Disease Professor Gossard Retrieving, Viewing Protein Structures from the Protein Data Base Helix helix packing Packing of Secondary
7.88 Lecture Notes - 4 7.24/7.88J/5.48J The Protein Folding and Human Disease Professor Gossard Retrieving, Viewing Protein Structures from the Protein Data Base Helix helix packing Packing of Secondary
Final Chem 4511/6501 Spring 2011 May 5, 2011 b Name
 Key 1) [10 points] In RNA, G commonly forms a wobble pair with U. a) Draw a G-U wobble base pair, include riboses and 5 phosphates. b) Label the major groove and the minor groove. c) Label the atoms of
Key 1) [10 points] In RNA, G commonly forms a wobble pair with U. a) Draw a G-U wobble base pair, include riboses and 5 phosphates. b) Label the major groove and the minor groove. c) Label the atoms of
LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor
 LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor Note: Adequate space is given for each answer. Questions that require a brief explanation should
LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor Note: Adequate space is given for each answer. Questions that require a brief explanation should
Physiochemical Properties of Residues
 Physiochemical Properties of Residues Various Sources C N Cα R Slide 1 Conformational Propensities Conformational Propensity is the frequency in which a residue adopts a given conformation (in a polypeptide)
Physiochemical Properties of Residues Various Sources C N Cα R Slide 1 Conformational Propensities Conformational Propensity is the frequency in which a residue adopts a given conformation (in a polypeptide)
CHAPTER 29 HW: AMINO ACIDS + PROTEINS
 CAPTER 29 W: AMI ACIDS + PRTEIS For all problems, consult the table of 20 Amino Acids provided in lecture if an amino acid structure is needed; these will be given on exams. Use natural amino acids (L)
CAPTER 29 W: AMI ACIDS + PRTEIS For all problems, consult the table of 20 Amino Acids provided in lecture if an amino acid structure is needed; these will be given on exams. Use natural amino acids (L)
PROTEIN STRUCTURE AMINO ACIDS H R. Zwitterion (dipolar ion) CO 2 H. PEPTIDES Formal reactions showing formation of peptide bond by dehydration:
 CO 2 H. PEPTIDES Formal reactions showing formation of peptide bond by dehydration:") PTEI STUTUE ydrolysis of proteins with aqueous acid or base yields a mixture of free amino acids. Each type of protein yields a characteristic mixture of the ~ 20 amino acids. AMI AIDS Zwitterion (dipolar
PTEI STUTUE ydrolysis of proteins with aqueous acid or base yields a mixture of free amino acids. Each type of protein yields a characteristic mixture of the ~ 20 amino acids. AMI AIDS Zwitterion (dipolar
The Structure and Functions of Proteins
 Wright State University CORE Scholar Computer Science and Engineering Faculty Publications Computer Science and Engineering 2003 The Structure and Functions of Proteins Dan E. Krane Wright State University
Wright State University CORE Scholar Computer Science and Engineering Faculty Publications Computer Science and Engineering 2003 The Structure and Functions of Proteins Dan E. Krane Wright State University
Table 1. Crystallographic data collection, phasing and refinement statistics. Native Hg soaked Mn soaked 1 Mn soaked 2
 Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
NH 2. Biochemistry I, Fall Term Sept 9, Lecture 5: Amino Acids & Peptides Assigned reading in Campbell: Chapter
 Biochemistry I, Fall Term Sept 9, 2005 Lecture 5: Amino Acids & Peptides Assigned reading in Campbell: Chapter 3.1-3.4. Key Terms: ptical Activity, Chirality Peptide bond Condensation reaction ydrolysis
Biochemistry I, Fall Term Sept 9, 2005 Lecture 5: Amino Acids & Peptides Assigned reading in Campbell: Chapter 3.1-3.4. Key Terms: ptical Activity, Chirality Peptide bond Condensation reaction ydrolysis
Translation. A ribosome, mrna, and trna.
 Translation The basic processes of translation are conserved among prokaryotes and eukaryotes. Prokaryotic Translation A ribosome, mrna, and trna. In the initiation of translation in prokaryotes, the Shine-Dalgarno
Translation The basic processes of translation are conserved among prokaryotes and eukaryotes. Prokaryotic Translation A ribosome, mrna, and trna. In the initiation of translation in prokaryotes, the Shine-Dalgarno
Read more about Pauling and more scientists at: Profiles in Science, The National Library of Medicine, profiles.nlm.nih.gov
 2018 Biochemistry 110 California Institute of Technology Lecture 2: Principles of Protein Structure Linus Pauling (1901-1994) began his studies at Caltech in 1922 and was directed by Arthur Amos oyes to
2018 Biochemistry 110 California Institute of Technology Lecture 2: Principles of Protein Structure Linus Pauling (1901-1994) began his studies at Caltech in 1922 and was directed by Arthur Amos oyes to
Chapter 4: Amino Acids
 Chapter 4: Amino Acids All peptides and polypeptides are polymers of alpha-amino acids. lipid polysaccharide enzyme 1940s 1980s. Lipids membrane 1960s. Polysaccharide Are energy metabolites and many of
Chapter 4: Amino Acids All peptides and polypeptides are polymers of alpha-amino acids. lipid polysaccharide enzyme 1940s 1980s. Lipids membrane 1960s. Polysaccharide Are energy metabolites and many of
The Structure of Enzymes!
 The Structure of Enzymes Levels of Protein Structure 0 order amino acid composition Primary Secondary Motifs Tertiary Domains Quaternary ther sequence repeating structural patterns defined by torsion angles
The Structure of Enzymes Levels of Protein Structure 0 order amino acid composition Primary Secondary Motifs Tertiary Domains Quaternary ther sequence repeating structural patterns defined by torsion angles
The Structure of Enzymes!
 The Structure of Enzymes Levels of Protein Structure 0 order amino acid composition Primary Secondary Motifs Tertiary Domains Quaternary ther sequence repeating structural patterns defined by torsion angles
The Structure of Enzymes Levels of Protein Structure 0 order amino acid composition Primary Secondary Motifs Tertiary Domains Quaternary ther sequence repeating structural patterns defined by torsion angles
3. Results Results Crystal structure of the N-terminal domain of human SHBG in complex with DHT
 3. Results 33 3. Results 3.1. Crystal structure of the N-terminal domain of human SHBG in complex with DHT 3.1.1. Crystallization For crystallization experiments the amino-terminal laminin G-like domain
3. Results 33 3. Results 3.1. Crystal structure of the N-terminal domain of human SHBG in complex with DHT 3.1.1. Crystallization For crystallization experiments the amino-terminal laminin G-like domain
Section Week 3. Junaid Malek, M.D.
 Section Week 3 Junaid Malek, M.D. Biological Polymers DA 4 monomers (building blocks), limited structure (double-helix) RA 4 monomers, greater flexibility, multiple structures Proteins 20 Amino Acids,
Section Week 3 Junaid Malek, M.D. Biological Polymers DA 4 monomers (building blocks), limited structure (double-helix) RA 4 monomers, greater flexibility, multiple structures Proteins 20 Amino Acids,
Central Dogma. modifications genome transcriptome proteome
 entral Dogma DA ma protein post-translational modifications genome transcriptome proteome 83 ierarchy of Protein Structure 20 Amino Acids There are 20 n possible sequences for a protein of n residues!
entral Dogma DA ma protein post-translational modifications genome transcriptome proteome 83 ierarchy of Protein Structure 20 Amino Acids There are 20 n possible sequences for a protein of n residues!
Major Types of Association of Proteins with Cell Membranes. From Alberts et al
 Major Types of Association of Proteins with Cell Membranes From Alberts et al Proteins Are Polymers of Amino Acids Peptide Bond Formation Amino Acid central carbon atom to which are attached amino group
Major Types of Association of Proteins with Cell Membranes From Alberts et al Proteins Are Polymers of Amino Acids Peptide Bond Formation Amino Acid central carbon atom to which are attached amino group
Protein Structure Bioinformatics Introduction
 1 Swiss Institute of Bioinformatics Protein Structure Bioinformatics Introduction Basel, 27. September 2004 Torsten Schwede Biozentrum - Universität Basel Swiss Institute of Bioinformatics Klingelbergstr
1 Swiss Institute of Bioinformatics Protein Structure Bioinformatics Introduction Basel, 27. September 2004 Torsten Schwede Biozentrum - Universität Basel Swiss Institute of Bioinformatics Klingelbergstr
Using Higher Calculus to Study Biologically Important Molecules Julie C. Mitchell
 Using Higher Calculus to Study Biologically Important Molecules Julie C. Mitchell Mathematics and Biochemistry University of Wisconsin - Madison 0 There Are Many Kinds Of Proteins The word protein comes
Using Higher Calculus to Study Biologically Important Molecules Julie C. Mitchell Mathematics and Biochemistry University of Wisconsin - Madison 0 There Are Many Kinds Of Proteins The word protein comes
Supplementary figure 1. Comparison of unbound ogm-csf and ogm-csf as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine
 Supplementary figure 1. Comparison of unbound and as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine GM-CSF (slate) with bound GM-CSF in the GIF:GM-CSF complex (GIF: green,
Supplementary figure 1. Comparison of unbound and as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine GM-CSF (slate) with bound GM-CSF in the GIF:GM-CSF complex (GIF: green,
Amino Acids and Peptides
 Amino Acids Amino Acids and Peptides Amino acid a compound that contains both an amino group and a carboxyl group α-amino acid an amino acid in which the amino group is on the carbon adjacent to the carboxyl
Amino Acids Amino Acids and Peptides Amino acid a compound that contains both an amino group and a carboxyl group α-amino acid an amino acid in which the amino group is on the carbon adjacent to the carboxyl
Bacterial protease uses distinct thermodynamic signatures for substrate recognition
 Bacterial protease uses distinct thermodynamic signatures for substrate recognition Gustavo Arruda Bezerra, Yuko Ohara-Nemoto, Irina Cornaciu, Sofiya Fedosyuk, Guillaume Hoffmann, Adam Round, José A. Márquez,
Bacterial protease uses distinct thermodynamic signatures for substrate recognition Gustavo Arruda Bezerra, Yuko Ohara-Nemoto, Irina Cornaciu, Sofiya Fedosyuk, Guillaume Hoffmann, Adam Round, José A. Márquez,
1. What is an ångstrom unit, and why is it used to describe molecular structures?
 1. What is an ångstrom unit, and why is it used to describe molecular structures? The ångstrom unit is a unit of distance suitable for measuring atomic scale objects. 1 ångstrom (Å) = 1 10-10 m. The diameter
1. What is an ångstrom unit, and why is it used to describe molecular structures? The ångstrom unit is a unit of distance suitable for measuring atomic scale objects. 1 ångstrom (Å) = 1 10-10 m. The diameter
From Amino Acids to Proteins - in 4 Easy Steps
 From Amino Acids to Proteins - in 4 Easy Steps Although protein structure appears to be overwhelmingly complex, you can provide your students with a basic understanding of how proteins fold by focusing
From Amino Acids to Proteins - in 4 Easy Steps Although protein structure appears to be overwhelmingly complex, you can provide your students with a basic understanding of how proteins fold by focusing
Biological Macromolecules
 Introduction for Chem 493 Chemistry of Biological Macromolecules Dr. L. Luyt January 2008 Dr. L. Luyt Chem 493-2008 1 Biological macromolecules are the molecules of life allow for organization serve a
Introduction for Chem 493 Chemistry of Biological Macromolecules Dr. L. Luyt January 2008 Dr. L. Luyt Chem 493-2008 1 Biological macromolecules are the molecules of life allow for organization serve a
26.7 Laboratory Synthesis of Peptides
 S Hornback_Ch26_1123-1161 12/15/04 8:18 PM Page 1148 1148 CHAPTER 26 AMI ACIDS, PEPTIDES, AD PRTEIS A chain B chain Gly Ile Val Glu Intramolecular disulfide bridge Gln Cys S S Cys Thr Ser Ile Cys Ser Leu
S Hornback_Ch26_1123-1161 12/15/04 8:18 PM Page 1148 1148 CHAPTER 26 AMI ACIDS, PEPTIDES, AD PRTEIS A chain B chain Gly Ile Val Glu Intramolecular disulfide bridge Gln Cys S S Cys Thr Ser Ile Cys Ser Leu
A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility
 proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility") (P&S Ch 5; Fer Ch 2, 9; Palm Ch 10,11; Zub Ch 9) A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility B.
(P&S Ch 5; Fer Ch 2, 9; Palm Ch 10,11; Zub Ch 9) A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility B.
Chemical Properties of Amino Acids
 hemical Properties of Amino Acids Protein Function Make up about 15% of the cell and have many functions in the cell 1. atalysis: enzymes 2. Structure: muscle proteins 3. Movement: myosin, actin 4. Defense:
hemical Properties of Amino Acids Protein Function Make up about 15% of the cell and have many functions in the cell 1. atalysis: enzymes 2. Structure: muscle proteins 3. Movement: myosin, actin 4. Defense:
CHMI 2227 EL. Biochemistry I. Test January Prof : Eric R. Gauthier, Ph.D.
 CHMI 2227 EL Biochemistry I Test 1 26 January 2007 Prof : Eric R. Gauthier, Ph.D. Guidelines: 1) Duration: 55 min 2) 14 questions, on 7 pages. For 70 marks (5 marks per question). Worth 15 % of the final
CHMI 2227 EL Biochemistry I Test 1 26 January 2007 Prof : Eric R. Gauthier, Ph.D. Guidelines: 1) Duration: 55 min 2) 14 questions, on 7 pages. For 70 marks (5 marks per question). Worth 15 % of the final
Model Mélange. Physical Models of Peptides and Proteins
 Model Mélange Physical Models of Peptides and Proteins In the Model Mélange activity, you will visit four different stations each featuring a variety of different physical models of peptides or proteins.
Model Mélange Physical Models of Peptides and Proteins In the Model Mélange activity, you will visit four different stations each featuring a variety of different physical models of peptides or proteins.
Dental Biochemistry Exam The total number of unique tripeptides that can be produced using all of the common 20 amino acids is
 Exam Questions for Dental Biochemistry Monday August 27, 2007 E.J. Miller 1. The compound shown below is CH 3 -CH 2 OH A. acetoacetate B. acetic acid C. acetaldehyde D. produced by reduction of acetaldehyde
Exam Questions for Dental Biochemistry Monday August 27, 2007 E.J. Miller 1. The compound shown below is CH 3 -CH 2 OH A. acetoacetate B. acetic acid C. acetaldehyde D. produced by reduction of acetaldehyde
Biomolecules: lecture 10
 Biomolecules: lecture 10 - understanding in detail how protein 3D structures form - realize that protein molecules are not static wire models but instead dynamic, where in principle every atom moves (yet
Biomolecules: lecture 10 - understanding in detail how protein 3D structures form - realize that protein molecules are not static wire models but instead dynamic, where in principle every atom moves (yet
Principles of Biochemistry
 Principles of Biochemistry Fourth Edition Donald Voet Judith G. Voet Charlotte W. Pratt Chapter 4 Amino Acids: The Building Blocks of proteins (Page 76-90) Chapter Contents 1- Amino acids Structure: 2-
Principles of Biochemistry Fourth Edition Donald Voet Judith G. Voet Charlotte W. Pratt Chapter 4 Amino Acids: The Building Blocks of proteins (Page 76-90) Chapter Contents 1- Amino acids Structure: 2-
4 Examples of enzymes
 Catalysis 1 4 Examples of enzymes Adding water to a substrate: Serine proteases. Carbonic anhydrase. Restrictions Endonuclease. Transfer of a Phosphoryl group from ATP to a nucleotide. Nucleoside monophosphate
Catalysis 1 4 Examples of enzymes Adding water to a substrate: Serine proteases. Carbonic anhydrase. Restrictions Endonuclease. Transfer of a Phosphoryl group from ATP to a nucleotide. Nucleoside monophosphate
Lecture 15: Enzymes & Kinetics. Mechanisms ROLE OF THE TRANSITION STATE. H-O-H + Cl - H-O δ- H Cl δ- HO - + H-Cl. Margaret A. Daugherty.
 Lecture 15: Enzymes & Kinetics Mechanisms Margaret A. Daugherty Fall 2004 ROLE OF THE TRANSITION STATE Consider the reaction: H-O-H + Cl - H-O δ- H Cl δ- HO - + H-Cl Reactants Transition state Products
Lecture 15: Enzymes & Kinetics Mechanisms Margaret A. Daugherty Fall 2004 ROLE OF THE TRANSITION STATE Consider the reaction: H-O-H + Cl - H-O δ- H Cl δ- HO - + H-Cl Reactants Transition state Products
What makes a good graphene-binding peptide? Adsorption of amino acids and peptides at aqueous graphene interfaces: Electronic Supplementary
 Electronic Supplementary Material (ESI) for Journal of Materials Chemistry B. This journal is The Royal Society of Chemistry 21 What makes a good graphene-binding peptide? Adsorption of amino acids and
Electronic Supplementary Material (ESI) for Journal of Materials Chemistry B. This journal is The Royal Society of Chemistry 21 What makes a good graphene-binding peptide? Adsorption of amino acids and
Viewing and Analyzing Proteins, Ligands and their Complexes 2
 2 Viewing and Analyzing Proteins, Ligands and their Complexes 2 Overview Viewing the accessible surface Analyzing the properties of proteins containing thousands of atoms is best accomplished by representing
2 Viewing and Analyzing Proteins, Ligands and their Complexes 2 Overview Viewing the accessible surface Analyzing the properties of proteins containing thousands of atoms is best accomplished by representing
Basic structures of proteins
 Basic structures of proteins Structural Hierarchy of Protein Primary structure Functional elements : α-helix, strands, β-sheet, loops.. - Structure, affinity, activity, specificity, stability etc. Secondary
Basic structures of proteins Structural Hierarchy of Protein Primary structure Functional elements : α-helix, strands, β-sheet, loops.. - Structure, affinity, activity, specificity, stability etc. Secondary
Biochemistry Quiz Review 1I. 1. Of the 20 standard amino acids, only is not optically active. The reason is that its side chain.
 Biochemistry Quiz Review 1I A general note: Short answer questions are just that, short. Writing a paragraph filled with every term you can remember from class won t improve your answer just answer clearly,
Biochemistry Quiz Review 1I A general note: Short answer questions are just that, short. Writing a paragraph filled with every term you can remember from class won t improve your answer just answer clearly,
Supplementary Figure 3 a. Structural comparison between the two determined structures for the IL 23:MA12 complex. The overall RMSD between the two
 Supplementary Figure 1. Biopanningg and clone enrichment of Alphabody binders against human IL 23. Positive clones in i phage ELISA with optical density (OD) 3 times higher than background are shown for
Supplementary Figure 1. Biopanningg and clone enrichment of Alphabody binders against human IL 23. Positive clones in i phage ELISA with optical density (OD) 3 times higher than background are shown for
Rotamers in the CHARMM19 Force Field
 Appendix A Rotamers in the CHARMM19 Force Field The people may be made to follow a path of action, but they may not be made to understand it. Confucius (551 BC - 479 BC) ( ) V r 1 (j),r 2 (j),r 3 (j),...,r
Appendix A Rotamers in the CHARMM19 Force Field The people may be made to follow a path of action, but they may not be made to understand it. Confucius (551 BC - 479 BC) ( ) V r 1 (j),r 2 (j),r 3 (j),...,r
Part II => PROTEINS and ENZYMES. 2.1 AMINO ACIDS 2.1a Nomenclature 2.1b Stereochemistry 2.1c Derivatives
 Part II => PROTEINS and ENZYMES 2.1 AMINO ACIDS 2.1a Nomenclature 2.1b Stereochemistry 2.1c Derivatives Section 2.1a: Nomenclature Synopsis 2.1a - Proteins (or polypeptides) are polymers made up of building
Part II => PROTEINS and ENZYMES 2.1 AMINO ACIDS 2.1a Nomenclature 2.1b Stereochemistry 2.1c Derivatives Section 2.1a: Nomenclature Synopsis 2.1a - Proteins (or polypeptides) are polymers made up of building
Chemistry Chapter 22
 hemistry 2100 hapter 22 Proteins Proteins serve many functions, including the following. 1. Structure: ollagen and keratin are the chief constituents of skin, bone, hair, and nails. 2. atalysts: Virtually
hemistry 2100 hapter 22 Proteins Proteins serve many functions, including the following. 1. Structure: ollagen and keratin are the chief constituents of skin, bone, hair, and nails. 2. atalysts: Virtually
B O C 4 H 2 O O. NOTE: The reaction proceeds with a carbonium ion stabilized on the C 1 of sugar A.
 hbcse 33 rd International Page 101 hemistry lympiad Preparatory 05/02/01 Problems d. In the hydrolysis of the glycosidic bond, the glycosidic bridge oxygen goes with 4 of the sugar B. n cleavage, 18 from
hbcse 33 rd International Page 101 hemistry lympiad Preparatory 05/02/01 Problems d. In the hydrolysis of the glycosidic bond, the glycosidic bridge oxygen goes with 4 of the sugar B. n cleavage, 18 from
Membrane Proteins: 1. Integral proteins: 2. Peripheral proteins: 3. Amphitropic proteins:
 Membrane Proteins: 1. Integral proteins: proteins that insert into/span the membrane bilayer; or covalently linked to membrane lipids. (Interact with the hydrophobic part of the membrane) 2. Peripheral
Membrane Proteins: 1. Integral proteins: proteins that insert into/span the membrane bilayer; or covalently linked to membrane lipids. (Interact with the hydrophobic part of the membrane) 2. Peripheral
What binds to Hb in addition to O 2?
 Reading: Ch5; 158-169, 162-166, 169-174 Problems: Ch5 (text); 3,7,8,10 Ch5 (study guide-facts); 1,2,3,4,5,8 Ch5 (study guide-apply); 2,3 Remember Today at 5:30 in CAS-522 is the second chance for the MB
Reading: Ch5; 158-169, 162-166, 169-174 Problems: Ch5 (text); 3,7,8,10 Ch5 (study guide-facts); 1,2,3,4,5,8 Ch5 (study guide-apply); 2,3 Remember Today at 5:30 in CAS-522 is the second chance for the MB
Protein Struktur (optional, flexible)
") Protein Struktur (optional, flexible) 22/10/2009 [ 1 ] Andrew Torda, Wintersemester 2009 / 2010, AST nur für Informatiker, Mathematiker,.. 26 kt, 3 ov 2009 Proteins - who cares? 22/10/2009 [ 2 ] Most important
Protein Struktur (optional, flexible) 22/10/2009 [ 1 ] Andrew Torda, Wintersemester 2009 / 2010, AST nur für Informatiker, Mathematiker,.. 26 kt, 3 ov 2009 Proteins - who cares? 22/10/2009 [ 2 ] Most important
NMR, X-ray Diffraction, Protein Structure, and RasMol
 NMR, X-ray Diffraction, Protein Structure, and RasMol Introduction So far we have been mostly concerned with the proteins themselves. The techniques (NMR or X-ray diffraction) used to determine a structure
NMR, X-ray Diffraction, Protein Structure, and RasMol Introduction So far we have been mostly concerned with the proteins themselves. The techniques (NMR or X-ray diffraction) used to determine a structure
Catalytic Mechanism of the Glycyl Radical Enzyme 4-Hydroxyphenylacetate Decarboxylase from Continuum Electrostatic and QC/MM Calculations
 Catalytic Mechanism of the Glycyl Radical Enzyme 4-Hydroxyphenylacetate Decarboxylase from Continuum Electrostatic and QC/MM Calculations Supplementary Materials Mikolaj Feliks, 1 Berta M. Martins, 2 G.
Catalytic Mechanism of the Glycyl Radical Enzyme 4-Hydroxyphenylacetate Decarboxylase from Continuum Electrostatic and QC/MM Calculations Supplementary Materials Mikolaj Feliks, 1 Berta M. Martins, 2 G.
Problem Set 1
 2006 7.012 Problem Set 1 Due before 5 PM on FRIDAY, September 15, 2006. Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. For each of the following parts, pick
2006 7.012 Problem Set 1 Due before 5 PM on FRIDAY, September 15, 2006. Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. For each of the following parts, pick
NMR study of complexes between low molecular mass inhibitors and the West Nile virus NS2B-NS3 protease
 University of Wollongong Research Online Faculty of Science - Papers (Archive) Faculty of Science, Medicine and Health 2009 NMR study of complexes between low molecular mass inhibitors and the West Nile
University of Wollongong Research Online Faculty of Science - Papers (Archive) Faculty of Science, Medicine and Health 2009 NMR study of complexes between low molecular mass inhibitors and the West Nile
Protein structure. Protein structure. Amino acid residue. Cell communication channel. Bioinformatics Methods
 Cell communication channel Bioinformatics Methods Iosif Vaisman Email: ivaisman@gmu.edu SEQUENCE STRUCTURE DNA Sequence Protein Sequence Protein Structure Protein structure ATGAAATTTGGAAACTTCCTTCTCACTTATCAGCCACCT...
Cell communication channel Bioinformatics Methods Iosif Vaisman Email: ivaisman@gmu.edu SEQUENCE STRUCTURE DNA Sequence Protein Sequence Protein Structure Protein structure ATGAAATTTGGAAACTTCCTTCTCACTTATCAGCCACCT...
CHEM 3653 Exam # 1 (03/07/13)
") 1. Using phylogeny all living organisms can be divided into the following domains: A. Bacteria, Eukarya, and Vertebrate B. Archaea and Eukarya C. Bacteria, Eukarya, and Archaea D. Eukarya and Bacteria
1. Using phylogeny all living organisms can be divided into the following domains: A. Bacteria, Eukarya, and Vertebrate B. Archaea and Eukarya C. Bacteria, Eukarya, and Archaea D. Eukarya and Bacteria
BCH 4053 Exam I Review Spring 2017
 BCH 4053 SI - Spring 2017 Reed BCH 4053 Exam I Review Spring 2017 Chapter 1 1. Calculate G for the reaction A + A P + Q. Assume the following equilibrium concentrations: [A] = 20mM, [Q] = [P] = 40fM. Assume
BCH 4053 SI - Spring 2017 Reed BCH 4053 Exam I Review Spring 2017 Chapter 1 1. Calculate G for the reaction A + A P + Q. Assume the following equilibrium concentrations: [A] = 20mM, [Q] = [P] = 40fM. Assume
Examples of Protein Modeling. Protein Modeling. Primary Structure. Protein Structure Description. Protein Sequence Sources. Importing Sequences to MOE
 Examples of Protein Modeling Protein Modeling Visualization Examination of an experimental structure to gain insight about a research question Dynamics To examine the dynamics of protein structures To
Examples of Protein Modeling Protein Modeling Visualization Examination of an experimental structure to gain insight about a research question Dynamics To examine the dynamics of protein structures To
Proteins: Characteristics and Properties of Amino Acids
 SBI4U:Biochemistry Macromolecules Eachaminoacidhasatleastoneamineandoneacidfunctionalgroupasthe nameimplies.thedifferentpropertiesresultfromvariationsinthestructuresof differentrgroups.thergroupisoftenreferredtoastheaminoacidsidechain.
SBI4U:Biochemistry Macromolecules Eachaminoacidhasatleastoneamineandoneacidfunctionalgroupasthe nameimplies.thedifferentpropertiesresultfromvariationsinthestructuresof differentrgroups.thergroupisoftenreferredtoastheaminoacidsidechain.
NAME IV. /22. I. MULTIPLE CHOICE. (48 points; 2 pts each) Choose the BEST answer to the question by circling the appropriate letter.
 Choose the BEST answer to the question by circling the appropriate letter.") NAME Exam I I. /48 September 25, 2017 Biochemistry I II. / 4 BI/CH 421/621 III. /26 IV. /22 TOTAL /100 I. MULTIPLE CHOICE. (48 points; 2 pts each) Choose the BEST answer to the question by circling the
NAME Exam I I. /48 September 25, 2017 Biochemistry I II. / 4 BI/CH 421/621 III. /26 IV. /22 TOTAL /100 I. MULTIPLE CHOICE. (48 points; 2 pts each) Choose the BEST answer to the question by circling the
It s the amino acids!
 Catalytic Mechanisms HOW do enzymes do their job? Reducing activation energy sure, but HOW does an enzyme catalysis reduce the energy barrier ΔG? Remember: The rate of a chemical reaction of substrate
Catalytic Mechanisms HOW do enzymes do their job? Reducing activation energy sure, but HOW does an enzyme catalysis reduce the energy barrier ΔG? Remember: The rate of a chemical reaction of substrate
Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets
 Supporting information Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets Wan-Na Chen, Christoph Nitsche, Kala Bharath Pilla, Bim Graham, Thomas
Supporting information Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets Wan-Na Chen, Christoph Nitsche, Kala Bharath Pilla, Bim Graham, Thomas
Key Concepts.
 Lectures 13-14: Enzyme Catalytic Mechanisms [PDF] Reading: Berg, Tymoczko & Stryer, Chapter 9, pp. 241-254 Updated on: 2/7/07 at 9:15 pm movie of chemical mechanism of serine proteases (from Voet & Voet,
Lectures 13-14: Enzyme Catalytic Mechanisms [PDF] Reading: Berg, Tymoczko & Stryer, Chapter 9, pp. 241-254 Updated on: 2/7/07 at 9:15 pm movie of chemical mechanism of serine proteases (from Voet & Voet,
CHEM J-9 June 2014
 CEM1611 2014-J-9 June 2014 Alanine (ala) and lysine (lys) are two amino acids with the structures given below as Fischer projections. The pk a values of the conjugate acid forms of the different functional
CEM1611 2014-J-9 June 2014 Alanine (ala) and lysine (lys) are two amino acids with the structures given below as Fischer projections. The pk a values of the conjugate acid forms of the different functional
Exam III. Please read through each question carefully, and make sure you provide all of the requested information.
 09-107 onors Chemistry ame Exam III Please read through each question carefully, and make sure you provide all of the requested information. 1. A series of octahedral metal compounds are made from 1 mol
09-107 onors Chemistry ame Exam III Please read through each question carefully, and make sure you provide all of the requested information. 1. A series of octahedral metal compounds are made from 1 mol
A. Two of the common amino acids are analyzed. Amino acid X and amino acid Y both have an isoionic point in the range of
 Questions with Answers- Amino Acids & Peptides A. Two of the common amino acids are analyzed. Amino acid X and amino acid Y both have an isoionic point in the range of 5.0-6.5 (Questions 1-4) 1. Which
Questions with Answers- Amino Acids & Peptides A. Two of the common amino acids are analyzed. Amino acid X and amino acid Y both have an isoionic point in the range of 5.0-6.5 (Questions 1-4) 1. Which
Review of General & Organic Chemistry
 Review of General & Organic Chemistry Diameter of a nucleus is only about 10-15 m. Diameter of an atom is only about 10-10 m. Fig 3.1 The structure of an atom Periodic Table, shown below, is a representation
Review of General & Organic Chemistry Diameter of a nucleus is only about 10-15 m. Diameter of an atom is only about 10-10 m. Fig 3.1 The structure of an atom Periodic Table, shown below, is a representation
DOCKING TUTORIAL. A. The docking Workflow
 2 nd Strasbourg Summer School on Chemoinformatics VVF Obernai, France, 20-24 June 2010 E. Kellenberger DOCKING TUTORIAL A. The docking Workflow 1. Ligand preparation It consists in the standardization
2 nd Strasbourg Summer School on Chemoinformatics VVF Obernai, France, 20-24 June 2010 E. Kellenberger DOCKING TUTORIAL A. The docking Workflow 1. Ligand preparation It consists in the standardization
Biomolecules: lecture 9
 Biomolecules: lecture 9 - understanding further why amino acids are the building block for proteins - understanding the chemical properties amino acids bring to proteins - realizing that many proteins
Biomolecules: lecture 9 - understanding further why amino acids are the building block for proteins - understanding the chemical properties amino acids bring to proteins - realizing that many proteins
Department of Structural Two refined structures, differing in alkali metal ion content, of the bifunctional,
 J. Mol. Biol. (1995) 245, 151 179 Structural and Mechanistic Analysis of Two Refined Crystal Structures of the Pyridoxal Phosphate-dependent Enzyme Dialkylglycine Decarboxylase Michael D. Toney 1, Erhard
J. Mol. Biol. (1995) 245, 151 179 Structural and Mechanistic Analysis of Two Refined Crystal Structures of the Pyridoxal Phosphate-dependent Enzyme Dialkylglycine Decarboxylase Michael D. Toney 1, Erhard
BSc and MSc Degree Examinations
 Examination Candidate Number: Desk Number: BSc and MSc Degree Examinations 2018-9 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes Marking
Examination Candidate Number: Desk Number: BSc and MSc Degree Examinations 2018-9 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes Marking
Supplementary Information
 1 Supplementary Information Figure S1 The V=0.5 Harker section of an anomalous difference Patterson map calculated using diffraction data from the NNQQNY crystal at 1.3 Å resolution. The position of the
1 Supplementary Information Figure S1 The V=0.5 Harker section of an anomalous difference Patterson map calculated using diffraction data from the NNQQNY crystal at 1.3 Å resolution. The position of the
12/6/12. Dr. Sanjeeva Srivastava IIT Bombay. Primary Structure. Secondary Structure. Tertiary Structure. Quaternary Structure.
 Dr. anjeeva rivastava Primary tructure econdary tructure Tertiary tructure Quaternary tructure Amino acid residues α Helix Polypeptide chain Assembled subunits 2 1 Amino acid sequence determines 3-D structure
Dr. anjeeva rivastava Primary tructure econdary tructure Tertiary tructure Quaternary tructure Amino acid residues α Helix Polypeptide chain Assembled subunits 2 1 Amino acid sequence determines 3-D structure
Rex-Family Repressor/NADH Complex
 Kasey Royer Michelle Lukosi Rex-Family Repressor/NADH Complex Part A The biological sensing protein that we selected is the Rex-family repressor/nadh complex. We chose this sensor because it is a calcium
Kasey Royer Michelle Lukosi Rex-Family Repressor/NADH Complex Part A The biological sensing protein that we selected is the Rex-family repressor/nadh complex. We chose this sensor because it is a calcium
Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate
 and IDH2 (bottom) with isocitrate") SUPPLEMENTARY FIGURE LEGENDS Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate dehydrogenase from Escherichia coli [ICD, pdb 1PB1, Mesecar, A. D., and Koshland,
SUPPLEMENTARY FIGURE LEGENDS Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate dehydrogenase from Escherichia coli [ICD, pdb 1PB1, Mesecar, A. D., and Koshland,
Supporting Information
 Supporting Information Micelle-Triggered b-hairpin to a-helix Transition in a 14-Residue Peptide from a Choline-Binding Repeat of the Pneumococcal Autolysin LytA HØctor Zamora-Carreras, [a] Beatriz Maestro,
Supporting Information Micelle-Triggered b-hairpin to a-helix Transition in a 14-Residue Peptide from a Choline-Binding Repeat of the Pneumococcal Autolysin LytA HØctor Zamora-Carreras, [a] Beatriz Maestro,
Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits. AhpC and AhpF from Escherichia coli
 Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits AhpC and AhpF from Escherichia coli Phat Vinh Dip 1,#, Neelagandan Kamariah 2,#, Malathy Sony Subramanian Manimekalai
Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits AhpC and AhpF from Escherichia coli Phat Vinh Dip 1,#, Neelagandan Kamariah 2,#, Malathy Sony Subramanian Manimekalai
SUPPLEMENTARY INFORMATION
 Table of Contents Page Supplementary Table 1. Diffraction data collection statistics 2 Supplementary Table 2. Crystallographic refinement statistics 3 Supplementary Fig. 1. casic1mfc packing in the R3
Table of Contents Page Supplementary Table 1. Diffraction data collection statistics 2 Supplementary Table 2. Crystallographic refinement statistics 3 Supplementary Fig. 1. casic1mfc packing in the R3
Supporting Information
 Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
2015 AP Biology Unit 2 PRETEST- Introduction to the Cell and Biochemistry
 Name: Class: _ Date: _ 2015 AP Biology Unit 2 PRETEST- Introduction to the Cell and Biochemistry Multiple Choice Identify the choice that best completes the statement or answers the question. 1) In what
Name: Class: _ Date: _ 2015 AP Biology Unit 2 PRETEST- Introduction to the Cell and Biochemistry Multiple Choice Identify the choice that best completes the statement or answers the question. 1) In what
SUPPLEMENTARY INFORMATION
 Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Protein Structure. Role of (bio)informatics in drug discovery. Bioinformatics
informatics in drug discovery. Bioinformatics") Bioinformatics Protein Structure Principles & Architecture Marjolein Thunnissen Dep. of Biochemistry & Structural Biology Lund University September 2011 Homology, pattern and 3D structure searches need
Bioinformatics Protein Structure Principles & Architecture Marjolein Thunnissen Dep. of Biochemistry & Structural Biology Lund University September 2011 Homology, pattern and 3D structure searches need
UNIT TWELVE. a, I _,o "' I I I. I I.P. l'o. H-c-c. I ~o I ~ I / H HI oh H...- I II I II 'oh. HO\HO~ I "-oh
 UNT TWELVE PROTENS : PEPTDE BONDNG AND POLYPEPTDES 12 CONCEPTS Many proteins are important in biological structure-for example, the keratin of hair, collagen of skin and leather, and fibroin of silk. Other
UNT TWELVE PROTENS : PEPTDE BONDNG AND POLYPEPTDES 12 CONCEPTS Many proteins are important in biological structure-for example, the keratin of hair, collagen of skin and leather, and fibroin of silk. Other
Secondary Structure. Bioch/BIMS 503 Lecture 2. Structure and Function of Proteins. Further Reading. Φ, Ψ angles alone determine protein structure
 Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
4. The Michaelis-Menten combined rate constant Km, is defined for the following kinetic mechanism as k 1 k 2 E + S ES E + P k -1
 Fall 2000 CH 595C Exam 1 Answer Key Multiple Choice 1. One of the reasons that enzymes are such efficient catalysts is that a) the energy level of the enzyme-transition state complex is much higher than
Fall 2000 CH 595C Exam 1 Answer Key Multiple Choice 1. One of the reasons that enzymes are such efficient catalysts is that a) the energy level of the enzyme-transition state complex is much higher than
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
7.012 Problem Set 1. i) What are two main differences between prokaryotic cells and eukaryotic cells?
 What are two main differences between prokaryotic cells and eukaryotic cells?") ame 7.01 Problem Set 1 Section Question 1 a) What are the four major types of biological molecules discussed in lecture? Give one important function of each type of biological molecule in the cell? b)
ame 7.01 Problem Set 1 Section Question 1 a) What are the four major types of biological molecules discussed in lecture? Give one important function of each type of biological molecule in the cell? b)
Biophysics 490M Project
 Biophysics 490M Project Dan Han Department of Biochemistry Structure Exploration of aa 3 -type Cytochrome c Oxidase from Rhodobacter sphaeroides I. Introduction: All organisms need energy to live. They
Biophysics 490M Project Dan Han Department of Biochemistry Structure Exploration of aa 3 -type Cytochrome c Oxidase from Rhodobacter sphaeroides I. Introduction: All organisms need energy to live. They
Chemistry Problem Set #9 Due on Thursday 11/15/18 in class.
 Chemistry 391 - Problem Set #9 Due on Thursday 11/15/18 in class. Name 1. There is a real enzyme called cocaine esterase that is produced in bacteria that live at the base of the coca plant. The enzyme
Chemistry 391 - Problem Set #9 Due on Thursday 11/15/18 in class. Name 1. There is a real enzyme called cocaine esterase that is produced in bacteria that live at the base of the coca plant. The enzyme
Supplemental Information for: Characterizing the Membrane-Bound State of Cytochrome P450 3A4: Structure, Depth of Insertion and Orientation
 Supplemental Information for: Characterizing the Membrane-Bound State of Cytochrome P450 3A4: Structure, Depth of Insertion and Orientation Javier L. Baylon, Ivan L. Lenov, Stephen G. Sligar and Emad Tajkhorshid
Supplemental Information for: Characterizing the Membrane-Bound State of Cytochrome P450 3A4: Structure, Depth of Insertion and Orientation Javier L. Baylon, Ivan L. Lenov, Stephen G. Sligar and Emad Tajkhorshid
Protein Structure. W. M. Grogan, Ph.D. OBJECTIVES
 Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Nature Structural & Molecular Biology doi: /nsmb Supplementary Figure 1. CRBN binding assay with thalidomide enantiomers.
 Supplementary Figure 1 CRBN binding assay with thalidomide enantiomers. (a) Competitive elution assay using thalidomide-immobilized beads coupled with racemic thalidomide. Beads were washed three times
Supplementary Figure 1 CRBN binding assay with thalidomide enantiomers. (a) Competitive elution assay using thalidomide-immobilized beads coupled with racemic thalidomide. Beads were washed three times
BA, BSc, and MSc Degree Examinations
 Examination Candidate Number: Desk Number: BA, BSc, and MSc Degree Examinations 2017-8 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes
Examination Candidate Number: Desk Number: BA, BSc, and MSc Degree Examinations 2017-8 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes