Determining basis for pit size variation in Myrmeleon immaculatus between two lakeshores of Northern Michigan
|
|
|
- Horatio Tyler
- 5 years ago
- Views:
Transcription
1 Determining basis for pit size variation in Myrmeleon immaculatus between two lakeshores of Northern Michigan Maura Niemisto University of Michigan Biological Station EEB 492: Behavioral Ecology 8/14/2013 Prof. Dave Anderson Abstract Environmental factors influencing pit-building behavior of the antlion Myrmeleon immaculatus are fairly well studied. The purpose of this study was to identify and differentiate environmental, genetic, and possible organizational or early learning factors influencing different behavioral phenotypes of two populations at Douglas Lake and Sturgeon Bay in northern Michigan, USA. We collected 60 antlions from each site and measured pit diameters in a common garden design. We distinguished treatments by feeding frequency, site, and day for a total of 22 days under a controlled laboratory setting with a regular disturbance regime. Sturgeon Bay antlions built larger pits that Douglas Lake antlions, animals that were subjected to a higher feeding frequency built smaller pits than a lower feeding frequency, and no apparent trend was gleaned from the day effect due to a presumed temperature effect. As a follow-up experiment we measured the effects of temperature on pit-building using light-bulb warming stations on 64 of our animals. Antlion pits that were given supplemental heat grew at a faster rate than controls. We also compared weight and pit diameters of at least 15 animals from seven sites between Sturgeon Bay and Pine Point. No significant difference was found between any site or lakeshore, though our data suggested that the original Sturgeon Bay study site displayed a larger pit size than any other site. These results indicate that genetic effects on these animals are unlikely, due to the close proximity of all populations. We suggest that organizational or early learning effects are significant determinants of pit-building behavior in this species.
2 2 I grant the Regents of the University of Michigan the non-exclusive right to retain, reproduce, and distribute my paper, titled in electronic formats and at no cost throughout the world. The University of Michigan may make and keep more than one copy of the Paper for purposes of security, backup, preservation and access, and may migrate the Paper to any medium or format for the purpose of preservation and access in the future. Signed, Maura Niemisto
3 3 Introduction The sit-and-wait strategy of predation is found as a foraging technique in several animal phyla. Species adapt this strategy as a way to balance the energetic costs of foraging with the energetic gain and frequency of prey items. Species within the same genus often exhibit either sit and wait or active foraging strategies despite shared environments, depending on differential daily energy requirements (Anderson and Karasov, 1981). Antlion larvae (family: Myrmeleontidae) provide a good model of this tradeoff because their behavior can be categorized into two foraging strategies: sit and wait, and sit and pursue predation (Scharf & Ovadia, 2006). The family Myrmeleontidae consists of over 2000 different species, but few build pit traps to capture their prey in a sit-and-wait fashion (Devetak et al., 2013). Myrmeleon immaculatus is one such species that sits at the bottom of a conical pit in sandy soils during their larval stage and waits until a prey item falls in. Upon encountering a prey item, the larva will fling sand off of its head, causing miniavalanches and impeding escape, thus increasing the amount of time available for capture by the larva s venomous jaws (Deventak, 2005). During its larval stage, M. immaculatus must capture ants and other arthropods to grow and, consequently, increase fecundity in adulthood (Burgess, 2009). The evolution of pit-digging has allowed relatively immobile antlion larvae to reduce the amount of energy spent actively searching for prey items (Ruxton and Hansell, 2009). Creating a large pit allows the animal to maintain a low metabolic cost in foraging, while increasing the area of prey trajectories they can intercept, resulting in a higher intake rate simultaneous to a decreasing amount of time necessary between meals (Ruxton and Hansell, 2009; Arnett and Gotelli, 2001). A study by Griffiths (1986) modeled the cost-benefits of increasing pit size. By increasing the pit volume, an antlion can increase the probability of encountering prey items (increasing pit diameter), or increase the capture rate of prey (increasing slope). Optimal pit size should provide the greatest energy gain by balancing these two parameters. However, in response to varying disturbance and feeding levels, antlions often respond by altering the size of their pit, the amount of energy spent on pit building, or by relocating their pit to a more desirable area (Eltz, 1997). One possible determinant of success in prey capture is the construction of pits large enough to produce a high capture success rate. Animal size has repeatedly been correlated with pit size in many studies, indicating that larger antlions build larger pits (Swenson et al., 2007). However, external constraints, such as disturbance frequency, conspecific density and environmental variability can decrease capture rate and may require a more flexible strategy of antlion pit structure (Klokocovnik et al., 2012). Several studies examined environmental effects on pit construction of M. immaculatus in laboratory and field contexts. In experiments simulating a high disturbance frequency, researchers have observed smaller pit sizes in response to increased disturbance (Barkae et al., 2010; Eltz, 1997). This indicates that differing environments could cause conspecifics to demonstrate varying pit construction outside of a predicted optimum. However, more in depth work on pit size due to disturbance regime is needed to fully understand this relationship. Frequency of feeding has produced varying results when tested experimentally. An early study indicated that decreased feeding frequency leads to decreased pit size (Youthed & Moran, 1969). However, Scharf et al. (2010) found that starving antlions who were exposed to prey items, but not allowed to eat increased their pit size at a constant rate, whereas those who were not exposed to prey items increased their pit size at a decreasing rate. This suggests that exposure to prey items can be a trigger for increased pit digging investment. Similarly, another study found lower feeding frequency to correspond with an increase in pit-building activity (Arnett and Gotelli, 2001). These two results indicate that a risk-taking strategy may be involved in pit-building behavior when feeding frequency is inconsistent. Pit construction has also been linked to soil type. Antlions tend to build pits with larger diameters and depth at low substrate densities resulting in a higher prey capture rate within those environments
4 (Devetak et. al, 2012). Research also indicates that antlions exhibit a preference toward medium-sized sand particles ( mm in diameter), and live at a higher density in this substrate type (Matsura et.al, 2005). Though these external constraints are fairly well studied, few researchers have examined factors of pit-building behavior among conspecific antlion populations outside of environmental condition. Arnett and Gotelli (2001) addressed this question by examining pit-building activity under different feeding and temperature regimes among conspecific populations from two northern and two southernstate populations. Under controlled conditions, southern-state individuals built and maintained pits more often than northern-state individuals, indicating behavioral influences independent of environmental effects. Because of the geographic differences, Arnett and Gotelli attributed these behavioral differences to genetic changes along a latitudinal gradient. However, researchers in Northern Michigan have isolated differences among populations in close geographic areas, indicating effects outside both genetic and environmental effects (Wiers, 2009; Roberts, 2007). Additional factors that may account for conspecific variation in similar geographic regions are organizational effects and early-experience learning. In general, organizational effects are defined as permanent effects on the behavior of an organism when exposed to a specific environment or hormone at a critical moment of development (Elekonich and Robinson, 2000). Unfortunately, this phenomenon is relatively understudied in insects as compared to vertebrates (Creel et al., 2013; Holekamp et al., 2013), though their endocrine systems could facilitate similar mechanisms (Elekonich and Robinson, 2000). The presence or absence of Juvenile Hormone (JH), an important insect chemical for development, has been shown to determine developmental trajectories during one or more critical periods of pre-reproductive growth of insects (Nijhout and Wheeler, 1982). The effect of JH in development and adult behavior has been researched primarily in caste system insects, such as bees and ants, where JH is intentionally upregulated in larvae through diet to determine future behavior and role within the colony (Hartfelder and Engels, 1998; Wheeler and Nijhout, 1984). Some studies have indicated that steroids in conjunction with environmental effects can also act as organizational effects to influence insect behavior in locusts, such as tolerance for conspecific density (Elekonich and Robinson, 2000). This implicates that variation in similar environments, even in close geographic proximity, may have organizational effects on insects that create variation in behavior later in life. Similar, early experience could create variation in behavior between populations of insect species. Though this influence has not been analyzed in M. immaculatus, Hirsch and Tompkins (1994) demonstrated that early experiences in Drosophila melanogaster affected different behavioral strategies later in life. In other words, they showed that the formation of intrinsic responses can be influenced by the animal s environment during development. For example, insects have been shown to change preference of feeding or oviposition sites based on early manipulation and increased exposure to those sites during larval stages (Beach and Jaynes, 1954). These characteristics indicate a development plasticity effecting behavior in later life. For our study we examined two populations of M. immaculate in the upper lower peninsula of Michigan. We sought to replicate and extend a previous study done by the University of Michigan Biological Station s Behavioral Ecology class in 2009 (Wiers, 2009). A difference in M. immaculate pit size has been consistently reported between these two populations by previous surveys. The study in 2009 sought to determine if an intrinsic difference existed separating the behavior of the two populations by testing their reaction to frequent disturbances in a controlled, laboratory environment. If the antlions did not produce significantly different pit sizes, it could be inferred that the observed behavior in the field is due to environmental effects. However, if the antlions retained a clear difference between populations for a large portion of the study, the results would support the presence of an inherent quality, such as genetics, organizational effects or early learning. Because the two populations are inferred to be in range of each other with probable gene flow, the nature of this quality may not be satisfied by a genetic explanation. Explanation for differences amongst populations behavior should accommodate consideration of organizational or early-experiential effects on behavioral variation. 4
5 5 The results of the study were ultimately inconclusive, due to the short 10-day study period. Antlions from the two sites showed a similar pit construction throughout the length of the experiment, indicating that environmental conditions were determinant of pit-building behavior. However, on the last day of the experiment, the two populations appeared to separate and reflect the pit size differences previously observed in the field (Figure 1). Whether or not this trend continues further into a disturbance regime was left unknown. Our study sought to lengthen this previous study to observe whether these two populations of M. immaculatus consistently display behavioral differences beyond 10 days of a disturbance regime. However, the previous study limited the antlion diet to an ant every three days. Because the feeding regime of antlions in natural conditions is unknown, we included an additional feeding regime variable to the original study to identify any effects of food limitation. In a field setting the 2009 study found the two populations exhibit different pit sizes even in the context of the same habitat and sand type (Wiers, 2009). Based on these results, we hypothesized that the differences in behavior consistently observed between the two is due to some other intrinsic factor effecting the populations differently. We predicted that in an extended study where environmental factors were controlled, the animals would display this intrinsic difference in pit-building activity, regardless of feeding frequency. Support for our hypothesis would indicate an important role of genetics, organizational effects or early-experiential factors on development of behavior. If we did not see a difference in pit sizes between the two populations under a controlled laboratory setting, we would find evidence for an alternative hypothesis, indicating that immediate environmental conditions are the main driver in differentiating the two populations pit-building behavior. Either result provides a better understanding of behavior in this insect species, and contributes to previous understandings of antlion behavior. If food availability is the major factor differentiating behavior between the two populations, we would expect pit diameters to be separated by feeding regime, and the site of origin to be irrelevant. We predicted that individuals exposed to a higher feeding-frequency would have the energetic ability to construct larger pit. However, the opposite result would suggest that animals fed at lower frequency adopt a risk-sensitive foraging strategy. One condition that we were able to control in our laboratory set-up was ambient temperature. Because antlions are poikilotherms, we expect them to have a lower metabolic rate at lower temperatures, resulting in slower or less activity (Tribe and Bowler, 1967). We hypothesized that temperature had a significant effect on antlion pit-building behavior. We sought to test this hypothesis in a supplemental study to understand how uncontrolled temperature might affect the results over time. As an additional follow-up to our principle study, we also sought to determine the degree of variation between different sites at the two lakeshores of Douglas Lake and Sturgeon Bay. This would inform on the importance of genetic versus environmental effects on antlions within these two regions, and test our assumption of non-limited gene flow between the two lakes. If we found genetic difference separating the two geographic regions, we would predict Douglas Lake and Sturgeon Bay populations to be more similar in pit-building behavior to populations at their own lake than each other. If the difference separating the two lakes is based mainly on micro-environmental factors, we would predict pit diameter to be characteristic of site, and an equal amount of variation between populations at each lake shore. Methods Time Course Experiment All research animals were collected on July 11 th, We collected 60 antlions from the coastline of Lake Michigan s South Sturgeon Bay (N W ), and 60 from Douglas Lake s Pine Point (N W ), both located in Emmet County, Michigan, USA. Only animals within a 13 to 40 milligram weight range were collected from either site. A total of 120 animals were labeled, weighed, and kept in capped vials for approximately 28 hours before being transferred into a
6 6 controlled laboratory environment. Laboratory temperatures were not controlled and fluctuated based on weather conditions. Deli tubs were filled with the second smallest fraction of sand from a beach on Douglas Lake and labeled according to treatment types. The treatments were arranged in a checkerboard pattern so as to reduce the influence of micro-environmental variation in the lab on the results (Figure 2). In all four treatments, a designated researcher measured pit diameter daily at 5:00 PM using digital calipers. We averaged the longest diameter with its perpendicular measurement to obtain an individual average pit diameter for each animal. Pits that were disrupted by proximity to the edge of the container, we only recorded the longest diameter measurement. We categorized the presence of pits based on whether the antlion had finished creating the pit and was not displaying any activity. We did not measure doodling, or pits under construction. Pits were measured from July 15 th, 2013 to August 5 th, 2013 for a total of 22 days. Following measurements, antlions were fed medium non-formic acid containing ants at a frequency according to their treatment condition (Table 1). All ants were collected from one colony on the campus of the University of Michigan Biological Station and stored in a refrigerator for up to 24 hours. Before feeding the study animal, ants were exposed to a refrigerated environment for a minimum of five minutes in order to facilitate handling. We disturbed all treatment types every three days by filling in the pits and leveling off the sand. The disturbance regime allowed us to continually renew pits to measure, and reflected recent behavior of the animals. We disturbed pits after measurements were taken and the antlions had finished eating their ant meals and had expelled the empty carcass from the pit. Because weights from the two sites were not normally distributed (Shapiro-Wilk test of normality p<0.001), we used a nonparametric Mann-Whitney test to test our assumption that our experimental animal s weights were comparably similar. We ran a factorial ANOVA using each animal s average pit diameter as the dependent variable and day, site, and feeding frequency as our categorical factors. We included Days 1-22, for a total of 22 days, two sites, and two feeding regimes. We evaluated each of the categorical factors as well as the interaction terms between all combinations of factors. We calculated the least squares means of the dependent variable to calculate site, feeding, and day effects. Temperature Variation Experiment In order to understand how the varying temperature affected our results, we followed up the time course experiment by setting up a warming station comprised of four iridescent light bulbs to warm 64 of the time-course-animals from 9:40 AM to 5:00PM daily. This procedure was repeated for three days from August 6 th -8 th, The remaining 56 animals were treated as a control and did not receive supplemental warmth. Feeding, disturbance regimes, and measurements remained consistent as per the methodology of the previous experiment. For analysis, the least squares mean pit diameters of each of the three days of the warming experiment were compared with the corresponding diameters of each of the three days preceding the experiment. We ran a repeated-measures ANOVA, omitting individuals who did not build pits for more than one consecutive day during the six-day sampling. For individuals missing one day, averages were calculated from the preceding and subsequent day diameters. If the 1 st or 6 th pit diameter was missing from any individual, we repeated the adjacent value. Population Variation Study On August 5 th, 2013, we measured four locations on the shoreline of Douglas Lake (DLsite1: N W , DLsite2: N W , DLsite3 N W , DLsite4: N W ) and three along the shoreline of Sturgeon Bay (SBsite1: N W , SBsite2:N W , SBsite3: N W ). Distances between sites at each of the lakeshores were calculated using GPS coordinates.
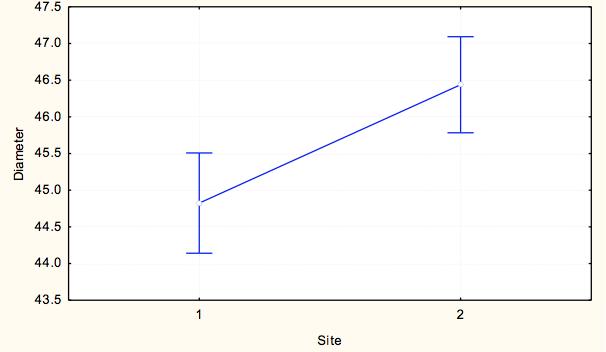
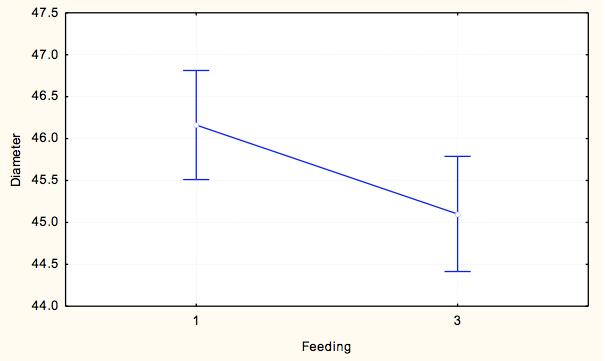
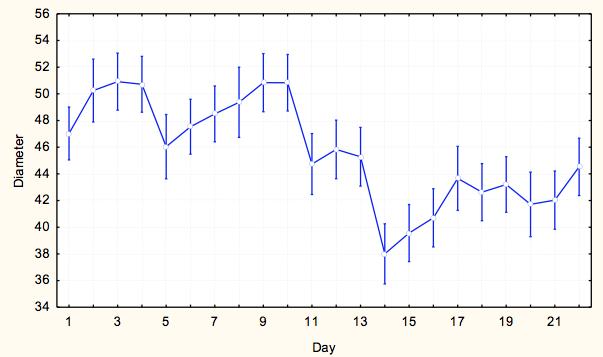
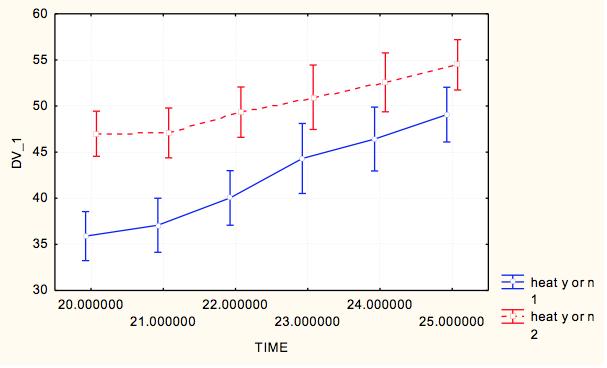
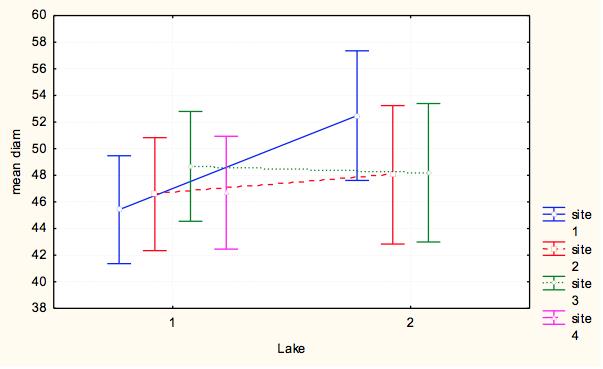
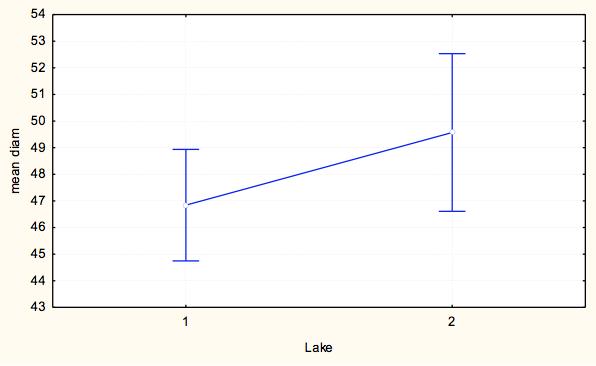
7 7 At each site, we measured pit diameter and animal weight. At each pit, we took two perpendicular measurements of pit diameter, and a measure of pit depth. To take each of these measurements, the same designated measurer from the time course experiment measured the longest diameter and its perpendicular using digital calipers and calculated an average pit diameter for each pit. The animal was then weighed using a digital scale to obtain the weight in milligrams as a measure of relative antlion size. This process was repeated until we collected at least 15 individuals from each site within a milligram weight range. For analysis we grouped the antlions into weight classes of 20-29mg and 30-39mg, which were represented at all sites, to control the weight effect. Exploratory data analysis showed that within 10g weight classes, pit diameter did not depend on weight and average weight within weight class did not differ by lake-site. We ran a two-way ANOVA to test the assumption that no lake*site effect existed (p=0.538) and that we could categorize pit diameters by weight class for further analysis. To test variation within and among lakesites, we ran a nested ANOVA with the dependent variable being an individual s average pit diameter, and the categorical factors being site (nested within lake), lake, and weight class to analyze effects of categorical factors on pit diameters. Results Time Course Experiment For our sample populations, a T-test determined that the two populations were not statistically different (t=-0.287, df=116.61, p=0.775). Based on the calculated least square means, the day, site and feeding effects were all significant, and none of the interaction terms were significant (Table 2). Over the course of the experiment, Sturgeon Bay animals built larger pits than Pine Point animals (F=11.184, df=1,1533, p= ) (Figure 3). Animals that were fed once every three days built larger pits than animals fed daily (F=4.8337, df=1, 1533, p=0.2806) (Figure 4). The day had a large effect on the average pit diameter across all animals, with fluctuating effects over time (F=12.304, df=21, 1533, p<0.0001)(figure 5). Figure 6 displays all main effects. Temperature Variation Micro-experiment Within the 6 day experiment period, when supplemental heat was added to 64 of the animals, average and maximum ambient temperatures of the heated animals increased more than in animals that were not exposed to supplemental heat (Figure 7). In addition, whether the animals were heated or not, the day of measurement, as well as the interaction of day and whether they were heated had a significant effect on average pit diameter (Table 3). Animals that were given supplemental heat increased pit size at a faster rate during the three-day course of heating than did animals that did not receive supplemental heat (F=4.7665, df=5, 375, p=0.0003) (Figure 8). Population Variation Study The results in testing the effectors of pit diameter, lake, site, and weight class all had insignificant effects on pit diameter (Table 4). The sites all had statistically similar average diameters when weight was controlled, although Sturgeon Bay Site 1 gave a suggestion of higher diameter (F= , df=5, 107, p= ) (Figure 9). Any effect indicated from lakeshore site is due to data from Sturgeon Bay site 1 (Figure 10). Distances between sites at each of the lakeshores are shown in Table 5. Discussion Our time course experiment revealed a significant behavior difference between our sample populations of Douglas Lake and Sturgeon Bay. Consistent with observations from previous years, Sturgeon Bay animals generally built larger pits than Pine Point animals, even when subjected to regular disturbance events over a period of 22 days (Figure 3). This finding supports our intrinsic quality
8 hypothesis. The behavioral differences antlions of similar weights persisted between the two populations despite shared environmental conditions and consistent stress (disturbances). The remaining explanations for consistent differences in behavior are genetic, early learning or organizational effect that conditioned these populations to behave differently throughout their larval stages. A genetic explanation seems unlikely because the two lake shores are within the same geographic area, and no restriction to gene flow is apparent in these researchers. However, genetic differences could not be ruled out without a supplemental study. The remaining explanation available is that some type of early experience, whether learning or organizational effect, separates these two populations and results in diverging behavior patterns later in life. The two sites of study differ. Animals located on a Lake Michigan, with a different soil type and degree of exposure are likely to experience different environmental inputs throughout the year, including but not limited to wind, ground temperature, species composition and others. If antlion larvae are sensitive to micro-environmental conditions early in life, they may be set on a permanently different developmental pathway. Feeding also had a significant effect on pit size within our study. Because pit-building incurs energetic costs, food intake is expected to affect pit size. In fact, some studies have associated starvation with smaller pit size (Youthed & Moran, 1969). However, in contrast to our prediction, antlions who were fed less often tended to build larger pits (Figure 4). This behavior may reflect a risk-sensitive foraging strategy, in which food-stressed individuals increase their pit sizes to raise the probability of prey capture (Abrams, 1991). Animals who are fed regularly, however, should avoid the extra cost of building a larger pit when they are consistently receiving nutrition (Abrams, 1991). High-risk foraging strategies have been found as explanation of spider and bird foraging strategies when subjects are experiencing negative energy budgets, and demonstrate energetically costly behavior to increase probability of a meal (Caraco et al., 1980). This behavior was observed M. immaculatus from both sites, indicating that food availability affects pit-building regardless of site of origin. The day effect was also significant among all antlions, but did not indicate an overall trend (Figure 5). Previous studies found that high disturbance regimes resulted in construction of smaller pits over an extended time period (Barkae et al., 2010; Eltz, 1997). The final pits in the last few days of this studywere smaller than the first few days, but no pattern was made evident. As one of the major uncontrolled conditions in our study, temperature likely had a large effect on metabolic function of the animals. In fact, temperature has been determined as a predictor of pit size (Arnett and Gotelli, 2001). During the course of our experiment, temperature average ranged from F (according to NOAA history for Charlevoix, Michigan). This leads us to believe that the large variation in temperature during our study period majorly confounded the results of our day effect. For this reason, we conducted our temperature variation experiment as a follow up to test this hypothesis. In our temperature variance experiment, we found temperature to have a large effect on antlion pit-digging behavior, supporting our previous lack of confidence in the day effect on antlion pits. Both average and maximum ambient temperature increased for animals given supplemental heat, leading to a faster rate of pit growth over the heated three-day period as compared to the animals that were not given supplemental heat (Figure 8). The non-heated animals had larger pit sizes than the heated animals in the three days preceding the experiment as well as a 0.5 C greater temperature on average. Though they initially built larger pits after the disturbance between day 3 and 4 (the supplemental heat experiment started on day 4), the pits of animals given supplemental heat grew much faster. If the experiment were to continue without disturbance, we suggest that the heated individuals would have been observed to surpass the non-heated individuals in average pit diameter, as predicted by results of previous temperature studies Arnett and Gotelli, 2001). These results indicate that temperature plays an important role in determining pit size, and likely had a large effect on the relationship of day to pit-size during our time course experiment. A better design for a time course experiment would control temperature and humidity changes to more clearly identify a day effect. Our last follow-up experiment measuring variation within and between lakeshores revealed that the two locations of study show fewer differences than we expected based on repeated samplings of our isolated sites of study. When we controlled for weight, neither lake nor site served as predictors for pit 8
9 diameter. From this result we cannot identify a genetic or environmental effect because antlions at all sites are behaved similarly. Our data suggested that animals from Sturgeon Bay site 1, the sample site for the time course experiment, build larger pits than other sites, though the study revealed those differences to be not significant (Figure 9). However, because this site has repeatedly shown larger pit sizes than the Pine Point sample site in the past, a difference likely exists between the animals there and other sample sites. Due to this repeated difference, we suggest that an unidentified factor differentially effects Sturgeon Bay site 1 antlions, which helps us interpret the role of genetic versus environmental factors within our experiment. Distances of Sturgeon Bay site 1 from the other Sturgeon Bay sites are both fewer than 3 km, which is likely too short of a distance to restrict gene flow between populations (Table 5). Though we cannot dismiss genetic factors to be distinguishing site 1 in pit-building behavior completely, the close proximity gives further confidence to our assumption of open gene flow between the two sites, and likely the two lakes. Furthermore, SB site 1 represents a micro-environment, though great variation in environmental conditions from the other SB sites on a day to day basis likely doesn t exist. This indicates another influencing factor of long term larva behavior beyond environmental and genetic differences. This follow-up study increases our confidence that an early experience or organizational effect is acting on the Sturgeon Bay site 1 antlions differently than antlions at all other sites, causing them to act differently in a controlled laboratory setting. In summary, our study finds that antlion behavior is affected by factors other than environmental condition. We suggest that genetics can also be ruled out as an explanation, though further studies are needed to increase confidence in this hypothesis. Slight inconsistency in measuring times and human error may have impacted the results of our study. In addition, we did not differentiate antlions based on age, nor had records of condition during the early life of any of our sample animals. In order to better understand the role that organizational effects and early learning may have on antlion behavior, a longterm experiment could be conducted in antlion rearing, where environmental conditions were manipulated during critical developmental periods of larva. Such an experiment could also be used to weigh the role of genetic effects in later instar behavior. Overall, these findings suggest a role of organizational or early learning effects in development of insect behavior, a fairly understudied topic outside of the order Hymenoptera. 9
10 10 Literature Cited Abrams, P. A Life history and the relationship between food availability and foraging effort. Ecology 72: Anderson, R. A., Karasov, W. H Contrasts in energy intake and expenditure in sit-and-wait and widely foraging lizards. Oecologia 49: Arnett A. M., Gotelli, N. J Pit-building decisions of larval ant lions: effects of larval age, temperature, food and population source. Journal of Insect Behavior 14: Barkae, E. D., Scharf, I., Subach, A., Ovadia, O The involvement of sand disturbance, cannibalism and intra-guild predation in competitive interactions among pit-building antlion larvae. Zoology 113: Beach, F. A., Jaynes, J Effects of early experience on the behavior of animals. Psychological Bulletin 50: Creel, S., Dantzer, B., Goymann, W., Rubenstein, D. R The ecology of stress: effects of the social environment. Functional Ecology 27: Deventak, D Effects of larval antlions Euroleon nostras (Neuroptera, Myrmeleontidae) and their pits on the escape-time of ants. Physiological Entomology 30: Devetak, D., Klokocovnik, V., Lipovsek, S., Bock, E., Leitinger, G Larval morphology of the antlion Myrmecaelurus trigrammus (Pallas, 1771)(Neuroptera, Myrmeleontidae), with notes on larval biology. Zootaxa 3641: Elekonich, M. M., Robinson, G. E Organizational and activational effects of hormones on insect behavior. Journal of Insect Physiology 46: Eltz, T Foraging in the ant-lion Myrmeleon mobilis hagen 1888 (Neuroptera: Myrmeleontidae): Behavioral flexibility of a sit-and-wait predator. Journal of Insect Beavior 10:1-11. Griffiths, D Pit construction by antlion larva: A cost-benefit analysis. Journal of Animal Ecology 55: Hartfelder, K., Engels, W Social insect polymorphism: Hormonal regulation of plasticity in development and reproduction of the honeybee. Current Topics in Developmental Biology 40: Hauber, M.E Variation in pit size of antlion (Myrmeleon carolinus) larvae: the importance of pit construction. Physiological Entomology 24: Hirsch, H. V. B., Tompkins, L The flexible fly-experience-dependent development of complex behaviors in Drosophila melanogaster. Journal of Experimental Biology 195: Holekamp, K. E., Swanson, E. M., Van Meter, P. E Developmental constraints on behavioral flexibility. Philosophical Transactions of the Royal Society Biological Sciences 368: 1-11
11 11 Klokocovnik, V., Devetak, D., Orlacnik, M Behavioral plasticity and variation in pit construction of antlion larvae in substrates with different particle sizes. Ethology 118(11): Lubin, Y., Henschel, J The influence of food supply on foraging behaviour in a desert spider. Oecologia 105: Nijhout, H. F., Wheeler, D Juvinile hormone and the physiological basis of insect polymorphisms. Quarterly Review of Biology 57: Roberts, S Pit building and location strategies of the antlion Myrmeleon immaculatus. Term paper in Behavioral Ecology, University of Michigan Biological Station. < =1> Ruxton, G. D., Hansell, M.H Why are pitfall traps so rare in the natural world? Evolutionary Ecology 23: Scharf, I., Ovadia, O Factors influencing site abandonment and site selection in a sit-and-wait predator: A review of pit-building antlion larvae. Journal of Insect Behavior (19) Scharf, I., Barkae, E. D., Ovadia, O Response of pit-building antlions to repeated unsuccessful encounters with prey. Animal Behaviour 79(1): Tribe, M. A., Bowler, K Temperature dependence of standard metabolic rate in a poikilotherm. Comparitive Biochemistry and Physiology 25: Wheeler, D. E., Nijhout, H. F Soldier determination in the ant Phiedole bicarinata: Inhibition by adult soldiers. Journal of Insect Physiology 30: Wiers, A. E The effects of disturbance and energy budget, intrinsic and environmental factors, and density on antlion pit size. Term paper in Behavioral Ecology, University of Michigan Biological Station. < Youthed, G.J., Moran, V.C Pit construction by Myrmeleontid larvae. Journal of Insect Physiology 15:
12
13 Tables Table 1: Treatment conditions for the time course experiment. All antlions were exposed to a disturbance every 3 days, on the same day that they were fed. Pine Point animals Sturgeon Bay animals Fed every three days (low-feeding frequency) Treatment 1 Treatment 2 Fed daily (high-feeding frequency) Treatment 3 Treatment 4
14 Table 2: Day, site, feeding regime, and interaction term effects on pit diameter for time course experiment. Day, site, and feeding regime all have significant effects on average pit diameter.
15 Table 3: Presence of heat, site, and interaction term effects on pit diameter for temperature variation experiment. Presence of heat, day(time), and presence of heat*day(time) interaction terms are statistically significant.
16 Table 4: Lake, site nested within lake, and interaction term effects on pit diameter for population variation study. No predictor has any significant effect on average pit diameter.
17 Table 5: Distances between sites in the population variation experiment. Calculated by GPS coordinates DL1 DL2 DL3 DL4 SB1 SB2 SB3 DL1 DL2 DL3 DL4 SB1 SB2 SB km 0.71 km 1.52 km 0.68 km 2.73 km
18 Figure Legends Figure 1: Average pit diameter results from previous class study (Wiers, 2010). Average pit diameters were significantly different between sites on day 0 (Z=4.932, p<0.001) and day 10 (Z=2.428, p=0.015). Days 1-9 were insignificant. Figure 2: Checkerboard layout of animals in time course experiment. This pattern was repeated twice for a total of 120 animals. Figure 3: Average pit diameter (mm) separated by site (1=Pine Point, 2=Sturgeon Bay) for time course experiment Figure 4: Average pit diameters (mm) separated by feeding regimes (1=one ant every three days (low feeding frequency), 3=one ant daily (high feeding frequency)) for time course experiment Figure 5: Average pit diameters (mm) separated by day of study for time course experiment Figure 6: Average pit diameter (mm) separated by day, feeding regimes, and site. Vertical lines represent disturbance events every three days. Figure 7: Maximum and average ambient temperature of antlions given supplemental heat and not given supplemental heat, three days prior (day 1-3), and three days during (day 4-6) the temperature variance experiment. Figure 8: Time and presence of heat interaction effect on average pit diameter (mm) for temperature variation experiment. Figure 9: Site effect at each lakeshore (1=Pine Point, 2=Sturgeon Bay) on average pit diameter (mm) for population variation study. Figure 10: Lakeshore effect (1=Pine Point, 2=Sturgeon Bay) on average pit diameter (mm) for population variation study.
19 Figures Figure 1

20 Figure 2
21 Figure 3
22 Figure 4
23 Figure 5
24 Pit Diameter Across Disturbance Regime 65 Mean Pit Diameter (mm) PP fed every 3 days PP fed daily SSB fed every 3 days SSB fed daily Day Figure 6
25 28 Temperature (C) Max no heat Max w/ heat Avg. no heat Avg. w/ heat Day Figure 7
26 Figure 8
27 Figure 9
28 Figure 10
Effects of rain and foot disturbances on pit size and location preference of antlions (Myrmeleon immaculatus)
") Effects of rain and foot disturbances on pit size and location preference of antlions (Myrmeleon immaculatus) Angela Feng University of Michigan Biological Station EEB 381 Ecology August 17, 2010 Professor
Effects of rain and foot disturbances on pit size and location preference of antlions (Myrmeleon immaculatus) Angela Feng University of Michigan Biological Station EEB 381 Ecology August 17, 2010 Professor
Taylor Glick. Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor, MI August 2013
 1 A Common Garden Experiment Comparing Antlion Pit Sizes at Douglas Lake and Sturgeon Bay: Genotype by Environment Interactions Taylor Glick Department of Ecology and Evolutionary Biology, University of
1 A Common Garden Experiment Comparing Antlion Pit Sizes at Douglas Lake and Sturgeon Bay: Genotype by Environment Interactions Taylor Glick Department of Ecology and Evolutionary Biology, University of
Pit building and Location Strategies of the Antlion Myrmeleon immaculatus.
 Roberts 1 of 17 Pit building and Location Strategies of the Antlion Myrmeleon immaculatus. Sophia R. Roberts Department of Biology, University of Michigan Biological Station, Pellston, Michigan Abstract
Roberts 1 of 17 Pit building and Location Strategies of the Antlion Myrmeleon immaculatus. Sophia R. Roberts Department of Biology, University of Michigan Biological Station, Pellston, Michigan Abstract
Variations in the Pit Size of Cueta sauteri (Neuroptera: Myrmeleontidae) Larvae in Response to Past Pit-Building Experience and Food Limitation
 Larvae in Response to Past Pit-Building Experience and Food Limitation") Zoological Studies 49(1): 102-107 (2010) Variations in the Pit Size of Cueta sauteri (Neuroptera: Myrmeleontidae) Larvae in Response to Past Pit-Building Experience and Food Limitation Shih-Hsiung Liang
Zoological Studies 49(1): 102-107 (2010) Variations in the Pit Size of Cueta sauteri (Neuroptera: Myrmeleontidae) Larvae in Response to Past Pit-Building Experience and Food Limitation Shih-Hsiung Liang
The effect of sand depth, feeding regime, density, and body mass on the foraging behaviour of a pit-building antlion
 Ecological Entomology (2009), 34, 26 33 DOI: 10.1111/j.1365-2311.2008.01038.x The effect of sand depth, feeding regime, density, and body mass on the foraging behaviour of a pit-building antlion INON SCHARF,
Ecological Entomology (2009), 34, 26 33 DOI: 10.1111/j.1365-2311.2008.01038.x The effect of sand depth, feeding regime, density, and body mass on the foraging behaviour of a pit-building antlion INON SCHARF,
The Flow of Aquatic Nitrogen from Ants to Antlions on the Sturgeon Bay Dunes
 Cara Fuentes Ecology-Scholtens Group 3 8/13/2008 The Flow of Aquatic Nitrogen from Ants to Antlions on the Sturgeon Bay Dunes Abstract In order to determine the effect of distance from the shoreline on
Cara Fuentes Ecology-Scholtens Group 3 8/13/2008 The Flow of Aquatic Nitrogen from Ants to Antlions on the Sturgeon Bay Dunes Abstract In order to determine the effect of distance from the shoreline on
Factors Influencing Site Abandonment and Site Selection in a Sit-and-Wait Predator: A Review of Pit-Building Antlion Larvae
 Journal of Insect Behavior, Vol. 19, No. 2, March 2006 ( C 2006) DOI: 10.1007/s10905-006-9017-4 Factors Influencing Site Abandonment and Site Selection in a Sit-and-Wait Predator: A Review of Pit-Building
Journal of Insect Behavior, Vol. 19, No. 2, March 2006 ( C 2006) DOI: 10.1007/s10905-006-9017-4 Factors Influencing Site Abandonment and Site Selection in a Sit-and-Wait Predator: A Review of Pit-Building
Niche The sum of all interactions a species has with biotic/abiotic components of the environment N-dimensional hypervolume
 Niche The sum of all interactions a species has with biotic/abiotic components of the environment N-dimensional hypervolume Each dimension is a biotic or abiotic resource Ecomorphology Ecology (niche)
Niche The sum of all interactions a species has with biotic/abiotic components of the environment N-dimensional hypervolume Each dimension is a biotic or abiotic resource Ecomorphology Ecology (niche)
Polyphenic Insects. genotype X environment = phenotype POLYPHENISM. genetic polymorphism vs polyphenism. the peppered moth.
 What makes for differences between individuals? Polyphenic Insects genes environment genotype X environment = phenotype POLYPHENISM poly many (more than one anyway) phen - form genetic polymorphism vs
What makes for differences between individuals? Polyphenic Insects genes environment genotype X environment = phenotype POLYPHENISM poly many (more than one anyway) phen - form genetic polymorphism vs
Chronic malnutrition favours smaller critical size for metamorphosis initiation in Drosophila melanogaster
 Vijendravarma et al: Experimental evolution of critical size 1 Chronic malnutrition favours smaller critical size for metamorphosis initiation in Drosophila melanogaster Roshan K. Vijendravarma, Sunitha
Vijendravarma et al: Experimental evolution of critical size 1 Chronic malnutrition favours smaller critical size for metamorphosis initiation in Drosophila melanogaster Roshan K. Vijendravarma, Sunitha
Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603)
") NCEA Level 3 Biology (91603) 2013 page 1 of 6 Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Assessment Criteria
NCEA Level 3 Biology (91603) 2013 page 1 of 6 Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Assessment Criteria
Foraging. This week in Animal Behaviour. How do animals choose? Do animals choose their food?
 This week in Animal Behaviour Lecture 22: Cooperation Lecture 23: Foraging Lecture 24: TBA Text chapters 10-11 LAB: Project 2 proposal seminars Midterm 2 on Tuesday Nov. 8 Foraging 1. Models of foraging
This week in Animal Behaviour Lecture 22: Cooperation Lecture 23: Foraging Lecture 24: TBA Text chapters 10-11 LAB: Project 2 proposal seminars Midterm 2 on Tuesday Nov. 8 Foraging 1. Models of foraging
Selection for late pupariation affects diapause incidence and duration in the flesh fly, Sarcophaga bullata
 Selection for late pupariation affects diapause incidence and duration in the flesh fly, Sarcophaga bullata By: Vincent C. Henrich and David L. Denlinger Henrich, V.C., and D.L. Denlinger (1982) Selection
Selection for late pupariation affects diapause incidence and duration in the flesh fly, Sarcophaga bullata By: Vincent C. Henrich and David L. Denlinger Henrich, V.C., and D.L. Denlinger (1982) Selection
What Shapes an Ecosystem Section 4-2
 What Shapes an Ecosystem Section 4-2 Biotic and Abiotic Factors Ecosystems are influenced by a combination of biological and physical factors. Biotic factors are the biological influences on an organism.
What Shapes an Ecosystem Section 4-2 Biotic and Abiotic Factors Ecosystems are influenced by a combination of biological and physical factors. Biotic factors are the biological influences on an organism.
Adaptation. Biotic and Abiotic Environments. Eric R. Pianka
 Adaptation Eric R. Pianka To survive and reproduce, all living organisms must adjust to conditions imposed on them by their environments. An organism's environment includes everything impinging upon it,
Adaptation Eric R. Pianka To survive and reproduce, all living organisms must adjust to conditions imposed on them by their environments. An organism's environment includes everything impinging upon it,
The relationship between current speed and shell morphology in the freshwater snail, Elimia livescens, in two Northern Michigan streams
 The relationship between current speed and shell morphology in the freshwater snail, Elimia livescens, in two Northern Michigan streams Katherine L. Anderson & Abigail R. DeBofsky University of Michigan
The relationship between current speed and shell morphology in the freshwater snail, Elimia livescens, in two Northern Michigan streams Katherine L. Anderson & Abigail R. DeBofsky University of Michigan
A A A A B B1
 LEARNING OBJECTIVES FOR EACH BIG IDEA WITH ASSOCIATED SCIENCE PRACTICES AND ESSENTIAL KNOWLEDGE Learning Objectives will be the target for AP Biology exam questions Learning Objectives Sci Prac Es Knowl
LEARNING OBJECTIVES FOR EACH BIG IDEA WITH ASSOCIATED SCIENCE PRACTICES AND ESSENTIAL KNOWLEDGE Learning Objectives will be the target for AP Biology exam questions Learning Objectives Sci Prac Es Knowl
SC741 W12: Division of Labor Part I: Fixed- and Variable- Threshold Algorithms
 SC741 W12: Division of Labor Part I: Fixed- and Variable- Threshold Algorithms Outline Division of labor in natural systems Ants Bees, wasps Models and mechanisms Fixed-threshold mechanisms Variable-threshold
SC741 W12: Division of Labor Part I: Fixed- and Variable- Threshold Algorithms Outline Division of labor in natural systems Ants Bees, wasps Models and mechanisms Fixed-threshold mechanisms Variable-threshold
Assessment of microhabitat differences between Drosera rotundifolia and Drosera intermedia in a northern Michigan bog
 Assessment of microhabitat differences between Drosera rotundifolia and Drosera intermedia in a northern Michigan bog Andrew David Abstract University of Michigan Biological Station EEB 381, General Ecology
Assessment of microhabitat differences between Drosera rotundifolia and Drosera intermedia in a northern Michigan bog Andrew David Abstract University of Michigan Biological Station EEB 381, General Ecology
Biology Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod. Introduction
 1 Biology 317 - Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod Introduction The determination of how natural selection acts in contemporary populations constitutes
1 Biology 317 - Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod Introduction The determination of how natural selection acts in contemporary populations constitutes
In recent years bat populations on the eastern seaboard and throughout the. Midwest United States have greatly decreased due to the fungal infection
 Burrell 1 Determining proper bat activity survey methods for the monitoring of the effect of Pseudogymnoascus destructans in Michigan bat populations Galen E. Burrell Abstract Pseudogymnoascus destructans,
Burrell 1 Determining proper bat activity survey methods for the monitoring of the effect of Pseudogymnoascus destructans in Michigan bat populations Galen E. Burrell Abstract Pseudogymnoascus destructans,
The effects of larval predation on the morphology of juvenile wood frogs (Rana sylvatica)
") The effects of larval predation on the morphology of juvenile wood frogs (Rana sylvatica) Maria Correa BIOS 35502: Practicum in Field Biology Advisor: Dr. Matthew Michel 2011 2 Abstract Organisms that
The effects of larval predation on the morphology of juvenile wood frogs (Rana sylvatica) Maria Correa BIOS 35502: Practicum in Field Biology Advisor: Dr. Matthew Michel 2011 2 Abstract Organisms that
Chapter 44. Table of Contents. Section 1 Development of Behavior. Section 2 Types of Animal Behavior. Animal Behavior
 Animal Behavior Table of Contents Section 1 Development of Behavior Section 2 Types of Animal Behavior Section 1 Development of Behavior Objectives Identify four questions asked by biologists who study
Animal Behavior Table of Contents Section 1 Development of Behavior Section 2 Types of Animal Behavior Section 1 Development of Behavior Objectives Identify four questions asked by biologists who study
SCOTCAT Credits: 20 SCQF Level 7 Semester 1 Academic year: 2018/ am, Practical classes one per week pm Mon, Tue, or Wed
 Biology (BL) modules BL1101 Biology 1 SCOTCAT Credits: 20 SCQF Level 7 Semester 1 10.00 am; Practical classes one per week 2.00-5.00 pm Mon, Tue, or Wed This module is an introduction to molecular and
Biology (BL) modules BL1101 Biology 1 SCOTCAT Credits: 20 SCQF Level 7 Semester 1 10.00 am; Practical classes one per week 2.00-5.00 pm Mon, Tue, or Wed This module is an introduction to molecular and
Georgia Performance Standards for Urban Watch Restoration Field Trips
 Georgia Performance Standards for Field Trips 6 th grade S6E3. Students will recognize the significant role of water in earth processes. a. Explain that a large portion of the Earth s surface is water,
Georgia Performance Standards for Field Trips 6 th grade S6E3. Students will recognize the significant role of water in earth processes. a. Explain that a large portion of the Earth s surface is water,
Interactions among Land, Water, and Vegetation in Shoreline Arthropod Communities
 AMERICAN JOURNAL OF UNDERGRADUATE RESEARCH VOL., NO.. () Interactions among Land, Water, and Vegetation in Shoreline Arthropod Communities Randall D. Willoughby and Wendy B. Anderson Department of Biology
AMERICAN JOURNAL OF UNDERGRADUATE RESEARCH VOL., NO.. () Interactions among Land, Water, and Vegetation in Shoreline Arthropod Communities Randall D. Willoughby and Wendy B. Anderson Department of Biology
Foraging behaviour and habitat selection in pit-building antlion larvae in constant light or dark conditions
 ANIMAL BEHAVIOUR, 28, 76, 249e7 doi:1.116/j.anbehav.28.8.23 Available online at www.sciencedirect.com Foraging behaviour and habitat selection in pit-building antlion larvae in constant light or dark conditions
ANIMAL BEHAVIOUR, 28, 76, 249e7 doi:1.116/j.anbehav.28.8.23 Available online at www.sciencedirect.com Foraging behaviour and habitat selection in pit-building antlion larvae in constant light or dark conditions
AP Curriculum Framework with Learning Objectives
 Big Ideas Big Idea 1: The process of evolution drives the diversity and unity of life. AP Curriculum Framework with Learning Objectives Understanding 1.A: Change in the genetic makeup of a population over
Big Ideas Big Idea 1: The process of evolution drives the diversity and unity of life. AP Curriculum Framework with Learning Objectives Understanding 1.A: Change in the genetic makeup of a population over
Adaptive Traits. Natural selection results in evolution of adaptations. Adaptation: trait that enhances an organism's survival and reproduction
 Adaptive Traits Adaptive Traits Natural selection results in evolution of adaptations Adaptation: trait that enhances an organism's survival and reproduction Nothing in biology makes sense except in the
Adaptive Traits Adaptive Traits Natural selection results in evolution of adaptations Adaptation: trait that enhances an organism's survival and reproduction Nothing in biology makes sense except in the
A trade-off between growth and starvation endurance in a pit-building antlion
 Oecologia (29) 16:453 46 DOI 1.17/s442-9-1316-y PHYSIOLOGICAL ECOLOGY - ORIGINAL PAPER A trade-off between growth and starvation endurance in a pit-building antlion Inon Scharf Æ Ido Filin Æ Ofer Ovadia
Oecologia (29) 16:453 46 DOI 1.17/s442-9-1316-y PHYSIOLOGICAL ECOLOGY - ORIGINAL PAPER A trade-off between growth and starvation endurance in a pit-building antlion Inon Scharf Æ Ido Filin Æ Ofer Ovadia
Group selection, individual selection. Group selection (GS) concept. Can behaviors ever arise that benefit the group and not the individual?
 concept. Can behaviors ever arise that benefit the group and not the individual?") Group selection, individual selection Can behaviors ever arise that benefit the group and not the individual? Group selection (GS) concept GS scenarios can be constructed, but they are not helpful in real
Group selection, individual selection Can behaviors ever arise that benefit the group and not the individual? Group selection (GS) concept GS scenarios can be constructed, but they are not helpful in real
Tatia Bauer. University of Michigan Biological Station. EEB 381 General Ecology. August 19, Cathy Bach
 The densities of goldenrod galls (Eurosta solidaginis) and their goldenrod host plants (Solidago canadensis) while directly related to each other, are not impacted by soil nitrogen or soil moisture Tatia
The densities of goldenrod galls (Eurosta solidaginis) and their goldenrod host plants (Solidago canadensis) while directly related to each other, are not impacted by soil nitrogen or soil moisture Tatia
Degree-days and Phenology Models
 Page 1 of 14 Degree-days and Phenology Models Using the UC IPM Degree-day Calculator Specify source of temperature data Data from UC IPM weather database Data from your file Data you enter online Run models
Page 1 of 14 Degree-days and Phenology Models Using the UC IPM Degree-day Calculator Specify source of temperature data Data from UC IPM weather database Data from your file Data you enter online Run models
OPTIMAL FORAGING MODELS
 26 OPTIMAL FORAGING MODELS In collaboration with David N. Bonter Objectives Develop a spreadsheet model of foraging choices among two prey types, prey 1 and prey 2. Determine the conditions in which individuals
26 OPTIMAL FORAGING MODELS In collaboration with David N. Bonter Objectives Develop a spreadsheet model of foraging choices among two prey types, prey 1 and prey 2. Determine the conditions in which individuals
Enduring understanding 1.A: Change in the genetic makeup of a population over time is evolution.
 The AP Biology course is designed to enable you to develop advanced inquiry and reasoning skills, such as designing a plan for collecting data, analyzing data, applying mathematical routines, and connecting
The AP Biology course is designed to enable you to develop advanced inquiry and reasoning skills, such as designing a plan for collecting data, analyzing data, applying mathematical routines, and connecting
Regents Review Assignment #8-A08 Living Environment: Comet Part A Questions
 Part A Questions 1. A student notices that fruit flies with the curlywing trait develop straight wings if kept at a temperature of 16 C, but develop curly wings if kept at 25 C. The best explanation for
Part A Questions 1. A student notices that fruit flies with the curlywing trait develop straight wings if kept at a temperature of 16 C, but develop curly wings if kept at 25 C. The best explanation for
Efficiency of antlion trap construction
 351 The Journal of Experimental Biology 29, 351-3515 Published by The Company of Biologists 26 doi:1.1242/jeb.241 Efficiency of antlion trap construction Arnold Fertin* and Jérôme Casas Université de Tours,
351 The Journal of Experimental Biology 29, 351-3515 Published by The Company of Biologists 26 doi:1.1242/jeb.241 Efficiency of antlion trap construction Arnold Fertin* and Jérôme Casas Université de Tours,
Current controversies in Marine Ecology with an emphasis on Coral reef systems. Niche Diversification Hypothesis Assumptions:
 Current controversies in Marine Ecology with an emphasis on Coral reef systems Open vs closed populations (already Discussed) The extent and importance of larval dispersal Maintenance of Diversity Equilibrial
Current controversies in Marine Ecology with an emphasis on Coral reef systems Open vs closed populations (already Discussed) The extent and importance of larval dispersal Maintenance of Diversity Equilibrial
Population Ecology. Study of populations in relation to the environment. Increase population size= endangered species
 Population Basics Population Ecology Study of populations in relation to the environment Purpose: Increase population size= endangered species Decrease population size = pests, invasive species Maintain
Population Basics Population Ecology Study of populations in relation to the environment Purpose: Increase population size= endangered species Decrease population size = pests, invasive species Maintain
BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences
 BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences D. POPULATION & COMMUNITY DYNAMICS Week 13. Herbivory, predation & parasitism: Lecture summary: Predation:
BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences D. POPULATION & COMMUNITY DYNAMICS Week 13. Herbivory, predation & parasitism: Lecture summary: Predation:
A Game Theory Based Predation Behavior Model
 A Game Theory Based Predation Behavior Model Shi Chen,Sheng Bao 2008-01-09 1 Abstract In this paper we first use a simplified non-cooperative zero sum Game Theory model model to investigate how predators
A Game Theory Based Predation Behavior Model Shi Chen,Sheng Bao 2008-01-09 1 Abstract In this paper we first use a simplified non-cooperative zero sum Game Theory model model to investigate how predators
NOTES: CH 4 Ecosystems & Communities
 NOTES: CH 4 Ecosystems & Communities 4.1 - Weather & Climate: WEATHER = day-to-day conditions of Earth s atmosphere CLIMATE= refers to average conditions over long periods; defined by year-afteryear patterns
NOTES: CH 4 Ecosystems & Communities 4.1 - Weather & Climate: WEATHER = day-to-day conditions of Earth s atmosphere CLIMATE= refers to average conditions over long periods; defined by year-afteryear patterns
Big Idea 1: The process of evolution drives the diversity and unity of life.
 Big Idea 1: The process of evolution drives the diversity and unity of life. understanding 1.A: Change in the genetic makeup of a population over time is evolution. 1.A.1: Natural selection is a major
Big Idea 1: The process of evolution drives the diversity and unity of life. understanding 1.A: Change in the genetic makeup of a population over time is evolution. 1.A.1: Natural selection is a major
Effects of Predator Chemical Cues On Snail Behavior
 Effects of Predator Chemical Cues On Snail Behavior BIOS 35502: Practicum in Field Biology Bryan J. Armajo; Mentor Shayna Sura July 23, 2012 1 Effects of Predator Chemical Cues On Snail Behavior Bryan
Effects of Predator Chemical Cues On Snail Behavior BIOS 35502: Practicum in Field Biology Bryan J. Armajo; Mentor Shayna Sura July 23, 2012 1 Effects of Predator Chemical Cues On Snail Behavior Bryan
because more individuals are heterozygous than homozygous recessive.
 1. A pesticide that was rarely used in 1932 was used with increasing frequency until it was banned altogether by 1972. Fruit flies (Drosophila melanogaster) that are resistant to this pesticide carry the
1. A pesticide that was rarely used in 1932 was used with increasing frequency until it was banned altogether by 1972. Fruit flies (Drosophila melanogaster) that are resistant to this pesticide carry the
Lesson Overview. Niches and Community Interactions. Lesson Overview. 4.2 Niches and Community Interactions
 Lesson Overview 4.2 Niches and Community Interactions The Niche What is a niche? A niche is the range of physical and biological conditions in which a species lives and the way the species obtains what
Lesson Overview 4.2 Niches and Community Interactions The Niche What is a niche? A niche is the range of physical and biological conditions in which a species lives and the way the species obtains what
Assessment Schedule 2016 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603)
") NCEA Level 3 Biology (91603) 2016 page 1 of 6 Assessment Schedule 2016 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Evidence Statement
NCEA Level 3 Biology (91603) 2016 page 1 of 6 Assessment Schedule 2016 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Evidence Statement
FACTORS FOR INSECTS ABUNDANCE. 1. More number of species: In the animal kingdom more than 85 per cent of the species
 FACTORS FOR INSECTS ABUNDANCE Measures of dominance 1. More number of species: In the animal kingdom more than 85 per cent of the species belongs to insect group. Total number of insects described so far
FACTORS FOR INSECTS ABUNDANCE Measures of dominance 1. More number of species: In the animal kingdom more than 85 per cent of the species belongs to insect group. Total number of insects described so far
5. Which graph represents a population that grew and is maintained at the carrying capacity of its ecosystem?
 Date: Pd. Regents Review Assignment #5 Living Environment 2 Part A Questions 1. Which structures carry out life functions within cells? (1) tissues (3) organelles (2) organ systems (4) organs 2. The sorting
Date: Pd. Regents Review Assignment #5 Living Environment 2 Part A Questions 1. Which structures carry out life functions within cells? (1) tissues (3) organelles (2) organ systems (4) organs 2. The sorting
Testing for Grazer Adaptation to Toxic Algae
 Testing for Grazer Adaptation to Toxic Algae by Michael B. Finiguerra, Hans G. Dam, and David E. Avery Part I Introduction and Background Phytoplankton, microscopic single-celled algae, are natural components
Testing for Grazer Adaptation to Toxic Algae by Michael B. Finiguerra, Hans G. Dam, and David E. Avery Part I Introduction and Background Phytoplankton, microscopic single-celled algae, are natural components
SYMPOSIUM Student Journal of Science & Math. Volume 2 Issue 1
 SYMPOSIUM Student Journal of Science & Math Volume 2 Issue 1 biology 117 B82.731 OBSERVATIONAL LEARNING IN EUSOCIAL INSECTS Background A RESEARCH PROPOSAL by Avity Norman Ants (order Hymenoptera, family
SYMPOSIUM Student Journal of Science & Math Volume 2 Issue 1 biology 117 B82.731 OBSERVATIONAL LEARNING IN EUSOCIAL INSECTS Background A RESEARCH PROPOSAL by Avity Norman Ants (order Hymenoptera, family
NICHE BREADTH AND RESOURCE PARTIONING
 22 NICHE BREADTH AND RESOURCE PARTIONING Objectives Compute niche breadth for two organisms coexisting in a community. Compute niche overlap for the two coexisting organisms. Use the Solver function to
22 NICHE BREADTH AND RESOURCE PARTIONING Objectives Compute niche breadth for two organisms coexisting in a community. Compute niche overlap for the two coexisting organisms. Use the Solver function to
Current controversies in Marine Ecology with an emphasis on Coral reef systems
 Current controversies in Marine Ecology with an emphasis on Coral reef systems Open vs closed populations (already discussed) The extent and importance of larval dispersal Maintenance of Diversity Equilibrial
Current controversies in Marine Ecology with an emphasis on Coral reef systems Open vs closed populations (already discussed) The extent and importance of larval dispersal Maintenance of Diversity Equilibrial
Rocky Intertidal Ecology -- part II The development of experimental ecology. Connell and the experimental revolution
 Rocky Intertidal Ecology -- part II The development of experimental ecology I. Intertidal Zonation, part II 1. Follow ups on Connell 2. Predation 3. Exceptions II. Horizontal Distribution 1. Variation
Rocky Intertidal Ecology -- part II The development of experimental ecology I. Intertidal Zonation, part II 1. Follow ups on Connell 2. Predation 3. Exceptions II. Horizontal Distribution 1. Variation
Chapter Chemical Uniqueness 1/23/2009. The Uses of Principles. Zoology: the Study of Animal Life. Fig. 1.1
 Fig. 1.1 Chapter 1 Life: Biological Principles and the Science of Zoology BIO 2402 General Zoology Copyright The McGraw Hill Companies, Inc. Permission required for reproduction or display. The Uses of
Fig. 1.1 Chapter 1 Life: Biological Principles and the Science of Zoology BIO 2402 General Zoology Copyright The McGraw Hill Companies, Inc. Permission required for reproduction or display. The Uses of
Biology. Slide 1 of 39. End Show. Copyright Pearson Prentice Hall
 Biology 1 of 39 4-2 What Shapes an Ecosystem? 2 of 39 Biotic and Abiotic Factors Biotic and Abiotic Factors Ecosystems are influenced by a combination of biological and physical factors. Biotic biological
Biology 1 of 39 4-2 What Shapes an Ecosystem? 2 of 39 Biotic and Abiotic Factors Biotic and Abiotic Factors Ecosystems are influenced by a combination of biological and physical factors. Biotic biological
4. Identify one bird that would most likely compete for food with the large tree finch. Support your answer. [1]
![4. Identify one bird that would most likely compete for food with the large tree finch. Support your answer. [1]](/thumbs/84/91138454.jpg "4. Identify one bird that would most likely compete for food with the large tree finch. Support your answer. [1]") Name: Topic 5B 1. A hawk has a genetic trait that gives it much better eyesight than other hawks of the same species in the same area. Explain how this could lead to evolutionary change within this species
Name: Topic 5B 1. A hawk has a genetic trait that gives it much better eyesight than other hawks of the same species in the same area. Explain how this could lead to evolutionary change within this species
Lecture 2: Individual-based Modelling
 Lecture 2: Individual-based Modelling Part I Steve Railsback Humboldt State University Department of Mathematics & Lang, Railsback & Associates Arcata, California USA www.langrailsback.com 1 Outline 1.
Lecture 2: Individual-based Modelling Part I Steve Railsback Humboldt State University Department of Mathematics & Lang, Railsback & Associates Arcata, California USA www.langrailsback.com 1 Outline 1.
Bees and Flowers. Unit 1: Qualitative and Graphical Approaches
 Bees and Flowers Often scientists use rate of change equations in their stu of population growth for one or more species. In this problem we stu systems of rate of change equations designed to inform us
Bees and Flowers Often scientists use rate of change equations in their stu of population growth for one or more species. In this problem we stu systems of rate of change equations designed to inform us
Chapter 8. Biogeographic Processes. Upon completion of this chapter the student will be able to:
 Chapter 8 Biogeographic Processes Chapter Objectives Upon completion of this chapter the student will be able to: 1. Define the terms ecosystem, habitat, ecological niche, and community. 2. Outline how
Chapter 8 Biogeographic Processes Chapter Objectives Upon completion of this chapter the student will be able to: 1. Define the terms ecosystem, habitat, ecological niche, and community. 2. Outline how
Biodiversity Classwork Classwork #1
 Biodiversity Classwork Classwork #1 1. What is biodiversity? 2. In the boxes below, create two ecosystems: one with low biodiversity and one with high biodiversity. Explain the difference. Biodiversity
Biodiversity Classwork Classwork #1 1. What is biodiversity? 2. In the boxes below, create two ecosystems: one with low biodiversity and one with high biodiversity. Explain the difference. Biodiversity
chatper 17 Multiple Choice Identify the choice that best completes the statement or answers the question.
 chatper 17 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. If a mutation introduces a new skin color in a lizard population, which factor might determine
chatper 17 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. If a mutation introduces a new skin color in a lizard population, which factor might determine
Goldenrod Galls and the Scientific Method
 Goldenrod Galls and the Scientific Method Overview Groups of students are given several goldenrod stems with galls. They are asked to make observations, come up with questions and make hypotheses. They
Goldenrod Galls and the Scientific Method Overview Groups of students are given several goldenrod stems with galls. They are asked to make observations, come up with questions and make hypotheses. They
Yakın Doğu Üniversitesi Mimarlık Fakültesi Peyzaj Mimarlığı Bölümü. PM 317 Human and Environment Assoc. Prof. Dr. Salih GÜCEL
 Yakın Doğu Üniversitesi Mimarlık Fakültesi Peyzaj Mimarlığı Bölümü PM 317 Human and Environment Assoc. Prof. Dr. Salih GÜCEL Ecology & Ecosystems Principles of Ecology Ecology is the study of the interactions
Yakın Doğu Üniversitesi Mimarlık Fakültesi Peyzaj Mimarlığı Bölümü PM 317 Human and Environment Assoc. Prof. Dr. Salih GÜCEL Ecology & Ecosystems Principles of Ecology Ecology is the study of the interactions
Adaptation and genetics. Block course Zoology & Evolution 2013, Daniel Berner
 Adaptation and genetics Block course Zoology & Evolution 2013, Daniel Berner 2 Conceptual framework Evolutionary biology tries to understand the mechanisms that lead from environmental variation to biological
Adaptation and genetics Block course Zoology & Evolution 2013, Daniel Berner 2 Conceptual framework Evolutionary biology tries to understand the mechanisms that lead from environmental variation to biological
4-2 What Shapes an Ecosystem?
 4-2 What Shapes an Ecosystem? Biotic and Abiotic Factors Ecosystems are influenced by a combination of biological and physical factors. Biotic biological factors predation competition resources Biotic
4-2 What Shapes an Ecosystem? Biotic and Abiotic Factors Ecosystems are influenced by a combination of biological and physical factors. Biotic biological factors predation competition resources Biotic
Testing adaptive hypotheses What is (an) adaptation? Testing adaptive hypotheses What is (an) adaptation?
 adaptation? Testing adaptive hypotheses What is (an) adaptation?") What is (an) adaptation? 1 A trait, or integrated set of traits, that increases the fitness of an organism. The process of improving the fit of phenotype to environment through natural selection What is
What is (an) adaptation? 1 A trait, or integrated set of traits, that increases the fitness of an organism. The process of improving the fit of phenotype to environment through natural selection What is
Map of AP-Aligned Bio-Rad Kits with Learning Objectives
 Map of AP-Aligned Bio-Rad Kits with Learning Objectives Cover more than one AP Biology Big Idea with these AP-aligned Bio-Rad kits. Big Idea 1 Big Idea 2 Big Idea 3 Big Idea 4 ThINQ! pglo Transformation
Map of AP-Aligned Bio-Rad Kits with Learning Objectives Cover more than one AP Biology Big Idea with these AP-aligned Bio-Rad kits. Big Idea 1 Big Idea 2 Big Idea 3 Big Idea 4 ThINQ! pglo Transformation
School of Biology. Biology (BL) modules. Biology & 2000 Level /8 - August BL1101 Biology 1
 modules. Biology & 2000 Level /8 - August BL1101 Biology 1") School of Biology Biology (BL) modules BL1101 Biology 1 SCOTCAT Credits: 20 SCQF Level 7 Semester: 1 10.00 am; Practical classes one per week 2.00-5.00 pm Mon, Tue, or Wed This module is an introduction
School of Biology Biology (BL) modules BL1101 Biology 1 SCOTCAT Credits: 20 SCQF Level 7 Semester: 1 10.00 am; Practical classes one per week 2.00-5.00 pm Mon, Tue, or Wed This module is an introduction
Microscale patterns of habitat fragmentation and disturbance events as a result of chemical applications:
 Microscale patterns of habitat fragmentation and disturbance events as a result of chemical applications: effects on Folsomia candida (Collembola) populations Mattia Meli Annemette Palmqvist, Valery E.
Microscale patterns of habitat fragmentation and disturbance events as a result of chemical applications: effects on Folsomia candida (Collembola) populations Mattia Meli Annemette Palmqvist, Valery E.
RECOMMENDATIONS/PREREQUISITES: No pre-requisites beyond the admissions standards required to gain a place on the course.
 1 MODULE: MODULE NUMBER: JACS CODE: Ecology BIO00001C C180 STAGE / YEAR: 1 CREDITS: 20 ORGANISER: PROGRAMME COMMITTEE: Thorunn Helgason BIO VERSION: August 2013 TERMS TAUGHT: Au/Sp/Su 2013/14 RECOMMENDATIONS/PREREQUISITES:
1 MODULE: MODULE NUMBER: JACS CODE: Ecology BIO00001C C180 STAGE / YEAR: 1 CREDITS: 20 ORGANISER: PROGRAMME COMMITTEE: Thorunn Helgason BIO VERSION: August 2013 TERMS TAUGHT: Au/Sp/Su 2013/14 RECOMMENDATIONS/PREREQUISITES:
Foraging decisions and behavioural flexibility in trap-building predators: a review
 Biol. Rev. (2011), 86, pp. 626 639. 626 doi: 10.1111/j.1469-185X.2010.00163.x Foraging decisions and behavioural flexibility in trap-building predators: a review Inon Scharf 1, Yael Lubin 2 and Ofer Ovadia
Biol. Rev. (2011), 86, pp. 626 639. 626 doi: 10.1111/j.1469-185X.2010.00163.x Foraging decisions and behavioural flexibility in trap-building predators: a review Inon Scharf 1, Yael Lubin 2 and Ofer Ovadia
TEMPERATURE, PREDATION RISK AND GRASSHOPPER BEHAVIOR. BIOS 569 Field Practicum in Environmental Biology. Molly Chambers
 Chambers TEMPERATURE, PREDATION RISK AND GRASSHOPPER BEHAVIOR BIOS 9 Field Practicum in Environmental Biology Molly Chambers Wheaton College, Wheaton, IL 87 Angela Laws UNDERC Chambers ABSTRACT A field
Chambers TEMPERATURE, PREDATION RISK AND GRASSHOPPER BEHAVIOR BIOS 9 Field Practicum in Environmental Biology Molly Chambers Wheaton College, Wheaton, IL 87 Angela Laws UNDERC Chambers ABSTRACT A field
VI) Population and Community Stability
 Population and Community Stability") Proportional abundance VI) Population and Community Stability I. Background / questions - refer back to succession A) Do marine communities trend toward climax states? B) Is there a single climax state?
Proportional abundance VI) Population and Community Stability I. Background / questions - refer back to succession A) Do marine communities trend toward climax states? B) Is there a single climax state?
Environmental Influences on Adaptation
 Have you ever noticed how the way you feel sometimes mirrors the emotions of the people with whom you spend a lot of time? For example, when you re around happy people, do you tend to become happy? Since
Have you ever noticed how the way you feel sometimes mirrors the emotions of the people with whom you spend a lot of time? For example, when you re around happy people, do you tend to become happy? Since
Behavioural Processes
 Behavioural Processes 80 (2009) 224 232 Contents lists available at ScienceDirect Behavioural Processes journal homepage: www.elsevier.com/locate/behavproc Learning in a sedentary insect predator: Antlions
Behavioural Processes 80 (2009) 224 232 Contents lists available at ScienceDirect Behavioural Processes journal homepage: www.elsevier.com/locate/behavproc Learning in a sedentary insect predator: Antlions
Weather is the day-to-day condition of Earth s atmosphere.
 4.1 Climate Weather and Climate Weather is the day-to-day condition of Earth s atmosphere. Climate refers to average conditions over long periods and is defined by year-after-year patterns of temperature
4.1 Climate Weather and Climate Weather is the day-to-day condition of Earth s atmosphere. Climate refers to average conditions over long periods and is defined by year-after-year patterns of temperature
FLOWERS AND POLLINATION. This activity introduces the relationship between flower structures and pollination.
 FLOWERS AND POLLINATION This activity introduces the relationship between flower structures and pollination. Objectives for Exam #1 1. Identify flower structures and match those structures to specific
FLOWERS AND POLLINATION This activity introduces the relationship between flower structures and pollination. Objectives for Exam #1 1. Identify flower structures and match those structures to specific
Computational Ecology Introduction to Ecological Science. Sonny Bleicher Ph.D.
 Computational Ecology Introduction to Ecological Science Sonny Bleicher Ph.D. Ecos Logos Defining Ecology Interactions: Organisms: Plants Animals: Bacteria Fungi Invertebrates Vertebrates The physical
Computational Ecology Introduction to Ecological Science Sonny Bleicher Ph.D. Ecos Logos Defining Ecology Interactions: Organisms: Plants Animals: Bacteria Fungi Invertebrates Vertebrates The physical
Biology 211 (2) Week 1 KEY!
 Week 1 KEY!") Biology 211 (2) Week 1 KEY Chapter 1 KEY FIGURES: 1.2, 1.3, 1.4, 1.5, 1.6, 1.7 VOCABULARY: Adaptation: a trait that increases the fitness Cells: a developed, system bound with a thin outer layer made of
Biology 211 (2) Week 1 KEY Chapter 1 KEY FIGURES: 1.2, 1.3, 1.4, 1.5, 1.6, 1.7 VOCABULARY: Adaptation: a trait that increases the fitness Cells: a developed, system bound with a thin outer layer made of
The Royal Entomological Society Journals
 Read the latest Virtual Special Issues from The Royal Entomological Society Journals Click on the buttons below to view the Virtual Special Issues Agricultural and Forest Pests Introduction This virtual
Read the latest Virtual Special Issues from The Royal Entomological Society Journals Click on the buttons below to view the Virtual Special Issues Agricultural and Forest Pests Introduction This virtual
Chapter 04 Lecture Outline
 Chapter 04 Lecture Outline William P. Cunningham University of Minnesota Mary Ann Cunningham Vassar College Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. 1
Chapter 04 Lecture Outline William P. Cunningham University of Minnesota Mary Ann Cunningham Vassar College Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. 1
BIOS 6150: Ecology Dr. Stephen Malcolm, Department of Biological Sciences
 BIOS 6150: Ecology Dr. Stephen Malcolm, Department of Biological Sciences Week 6: Predation and predatory behavior: Lecture summary: Nature of predation. Diet breadth & choice. Optimal foraging. Functional
BIOS 6150: Ecology Dr. Stephen Malcolm, Department of Biological Sciences Week 6: Predation and predatory behavior: Lecture summary: Nature of predation. Diet breadth & choice. Optimal foraging. Functional
Eye size in Drosophila melanogaster and how it affects peripheral motion vision. Abstract
 Varkey 1 Justin Varkey Anthony McGoron 4 December 2015 Eye size in Drosophila melanogaster and how it affects peripheral motion vision Abstract Drosophila melanogaster, the fruit fly, is a holometabolous
Varkey 1 Justin Varkey Anthony McGoron 4 December 2015 Eye size in Drosophila melanogaster and how it affects peripheral motion vision Abstract Drosophila melanogaster, the fruit fly, is a holometabolous
Class Announcements. Extended Example: Bird Song Learning. White-crowned sparrows and Social Learning. White-crowned sparrows and Social Learning
 Class Announcements Section reading Paper for next week is up online. The paper is Sexual Selection, Temperature, and the Lion's Mane. Section reading must be done before coming to section. Web recording
Class Announcements Section reading Paper for next week is up online. The paper is Sexual Selection, Temperature, and the Lion's Mane. Section reading must be done before coming to section. Web recording
LACK OF TASK DIFFERENTIATION DURING PREY CAPTURE IN THE GROUP LIVING SPIDER STEGODYPHUS MIMOSARUM (ARANEAE, ERESIDAE)
") 2002. The Journal of Arachnology 30:39 46 LACK OF TASK DIFFERENTIATION DURING PREY CAPTURE IN THE GROUP LIVING SPIDER STEGODYPHUS MIMOSARUM (ARANEAE, ERESIDAE) Cheron Ainsworth and Rob Slotow: School of
2002. The Journal of Arachnology 30:39 46 LACK OF TASK DIFFERENTIATION DURING PREY CAPTURE IN THE GROUP LIVING SPIDER STEGODYPHUS MIMOSARUM (ARANEAE, ERESIDAE) Cheron Ainsworth and Rob Slotow: School of
Principles of Ecology BL / ENVS 402 Exam II Name:
 Principles of Ecology BL / ENVS 402 Exam II 10-26-2011 Name: There are three parts to this exam. Use your time wisely as you only have 50 minutes. Part One: Circle the BEST answer. Each question is worth
Principles of Ecology BL / ENVS 402 Exam II 10-26-2011 Name: There are three parts to this exam. Use your time wisely as you only have 50 minutes. Part One: Circle the BEST answer. Each question is worth
Adaptation. Analysis of form and function
 Adaptation Analysis of form and function What is adaptation Trait, or set of related traits, that increases the fitness of the organism that has it Adaptations created by natural selection Requires mutation
Adaptation Analysis of form and function What is adaptation Trait, or set of related traits, that increases the fitness of the organism that has it Adaptations created by natural selection Requires mutation
Evolution Common Assessment 1
 Evolution Common Assessment 1 1. The field of biology that includes the study of the origin of new species through time is known as 5. A. biochemistry B. evolution C. ecology D. embryology 2. Evidence
Evolution Common Assessment 1 1. The field of biology that includes the study of the origin of new species through time is known as 5. A. biochemistry B. evolution C. ecology D. embryology 2. Evidence
Natural Selection in Action
 3 What You Will Learn Genetic variation and environmental factors affect evolution by natural selection. Separation, adaptation, and reproductive isolation can produce new species. Extinction occurs when
3 What You Will Learn Genetic variation and environmental factors affect evolution by natural selection. Separation, adaptation, and reproductive isolation can produce new species. Extinction occurs when
Joseph Priestly ECOSYSTEMS. Part
 ECOSYSTEMS Part 2 Joseph Priestly Joseph Priestly was an English clergyman and scientist. He noticed that if he put a burning candle in a jar, the candle went out after a few minutes and when he put a
ECOSYSTEMS Part 2 Joseph Priestly Joseph Priestly was an English clergyman and scientist. He noticed that if he put a burning candle in a jar, the candle went out after a few minutes and when he put a
Eusocial species. Eusociality. Phylogeny showing only eusociality Eusocial insects. Eusociality: Cooperation to the extreme
 Eusociality: Cooperation to the extreme Groups form colonies with reproductive and worker castes. Eusociality has evolved most often in insects: Ants Eusocial species Honeybees Termites Wasps Phylogeny
Eusociality: Cooperation to the extreme Groups form colonies with reproductive and worker castes. Eusociality has evolved most often in insects: Ants Eusocial species Honeybees Termites Wasps Phylogeny
CHAPTER - V. POPULATION STUDIES OF THE TERMITE O.wallonensis
 CHAPTER - V POPULATION STUDIES OF THE TERMITE O.wallonensis 5.1. Introduction Andrews (1911) and Grasses (1939) studied the total population and relative percentage of various castes in different species
CHAPTER - V POPULATION STUDIES OF THE TERMITE O.wallonensis 5.1. Introduction Andrews (1911) and Grasses (1939) studied the total population and relative percentage of various castes in different species
Animal Behavior (Ch. 51)
") Animal Behavior (Ch. 51) Behavioral Ecology Two types of questions: Proximate questions: Focus on environmental stimuli that trigger behavior and physiology behind response How? Ultimate questions: Focus
Animal Behavior (Ch. 51) Behavioral Ecology Two types of questions: Proximate questions: Focus on environmental stimuli that trigger behavior and physiology behind response How? Ultimate questions: Focus
ENVE203 Environmental Engineering Ecology (Nov 05, 2012)
") ENVE203 Environmental Engineering Ecology (Nov 05, 2012) Elif Soyer Ecosystems and Living Organisms Population Density How Do Populations Change in Size? Maximum Population Growth Environmental Resistance
ENVE203 Environmental Engineering Ecology (Nov 05, 2012) Elif Soyer Ecosystems and Living Organisms Population Density How Do Populations Change in Size? Maximum Population Growth Environmental Resistance
Bee Colony Activities Throughout The Year
 Bee Colony Activities Throughout The Year Written by Khalil Hamdan Apeldoorn The Netherlands A honeybee gathering nectar from a flower. Photo source: forestwander.com Bee collecting pollen. Photo source:
Bee Colony Activities Throughout The Year Written by Khalil Hamdan Apeldoorn The Netherlands A honeybee gathering nectar from a flower. Photo source: forestwander.com Bee collecting pollen. Photo source:
Existing modelling studies on shellfish
 Existing modelling studies on shellfish Laboratoire Ressources Halieutiques IFREMER Port-en-Bessin, France Worldwide production of cultured shellfish GENIMPACT February 2007 Main species and producers
Existing modelling studies on shellfish Laboratoire Ressources Halieutiques IFREMER Port-en-Bessin, France Worldwide production of cultured shellfish GENIMPACT February 2007 Main species and producers
8/23/2014. Introduction to Animal Diversity
 Introduction to Animal Diversity Chapter 32 Objectives List the characteristics that combine to define animals Summarize key events of the Paleozoic, Mesozoic, and Cenozoic eras Distinguish between the
Introduction to Animal Diversity Chapter 32 Objectives List the characteristics that combine to define animals Summarize key events of the Paleozoic, Mesozoic, and Cenozoic eras Distinguish between the
Aggregations on larger scales. Metapopulation. Definition: A group of interconnected subpopulations Sources and Sinks
 Aggregations on larger scales. Metapopulation Definition: A group of interconnected subpopulations Sources and Sinks Metapopulation - interconnected group of subpopulations sink source McKillup and McKillup
Aggregations on larger scales. Metapopulation Definition: A group of interconnected subpopulations Sources and Sinks Metapopulation - interconnected group of subpopulations sink source McKillup and McKillup
Chapter 6 Lecture. Life History Strategies. Spring 2013
 Chapter 6 Lecture Life History Strategies Spring 2013 6.1 Introduction: Diversity of Life History Strategies Variation in breeding strategies, fecundity, and probability of survival at different stages
Chapter 6 Lecture Life History Strategies Spring 2013 6.1 Introduction: Diversity of Life History Strategies Variation in breeding strategies, fecundity, and probability of survival at different stages