THE UNIVERSITY OF MICHIGAN INTERNAL STRUCTURES IN TWO EDRIOASTEROID SPECIES, ISOROPHUS CINCINNATIENSIS (ROEMER) AND CARNEYELLA PILEA (HALL)
|
|
|
- Megan Fleming
- 5 years ago
- Views:
Transcription
1 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN i VOL. XV, No. 14, pp , (9 pls., 4 figs.) NOVEMBER 30, 1960 INTERNAL STRUCTURES IN TWO EDRIOASTEROID SPECIES, ISOROPHUS CINCINNATIENSIS (ROEMER) AND CARNEYELLA PILEA (HALL) BY ROBERT V. KESLING and LEIGH W. MINTZ MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN ANN ARBOR
2 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY Director: LEWIS B. KELLUM The series of contributions from the Museum of Paleontology is a medium for the publication of papers based chiefly upon the collection in the Museum. When the number of pages issued is sufficient to make a volume, a title page and a table of contents will be sent to libraries on the mailing list, and to individuals upon request. A list of the separate papers may also be obtained. Correspondence should be directed to the Museum of Paleontology, The University of Michigan, Ann Arbor, Michigan. VOLS. 11-XIV. Parts of volumes may be obtained if available. VOLUME XV 1. Leioptwia rafinesquii Hall and A New Species of the Genus from the Thedford-Arkona Region of southwestern Ontario, by George M. Ehlers and Edward P. Wright. Pages 1-13, with 2 plates. 2. Quasillitid and Alanellid Ostracods from the Centerfield Limestone of Western New York, by Robert V. Kesling, Emadeddin Kavary, Robert S. Takagi, John R. Tillman, and George R. Wulf. Pages 15-31, with 4 plates. 3. A New Species of Mahonia from the Oligocene Ruby flora of Southwestern Montana, by Herman F. Becker. Pages 33-38, with 1 plate. 4. Some Middle Devonian Stromatoporoids from Michigan and Southwestern Ontario, Including the Types Described by Alexander Winchell and A. W. Grabau, by J. J. Galloway and G. M. Ehlers. Pages , with 13 plates. 5. A Simple Device for Single-lens Stereophotography of Paleontological Specimens, by Herbert W. Wienert. Pages , with 1 plate. 6. Techniques in the Photography of Fossilized Plants, by Herbert W. Wienert. Pages , with 3 plates. 7. The Type Species of the Paleozoic Tabulate Coral Genera Cladopora and Coenites, by Erwin C. Stumm. Pages , with 1 plate. 8. Hydropores in Edrioasteroids, by Robert V. Kesling. Pages , with 13 plates. 9. An Upper Cretaceous Ammonite Bitten by a Mosasaur, by Erle G. Kauffman and Robert V. Kesling. Pages , with 9 plates. 10. A Lepidodendrid Stem from Kansas and its Bearing on the Problem of Cambium and Phloem in Paleozoic Lycopods. Pages , with 4 plates. 11. A New Genus and Four New Species of Fossil Diptera from Montana and Colorado, by Frank M. Hull. Pages , with 4 plates. 12. New Upper Devonian Cypridinacean Ostracod from Southern Indiana, by Robert V. Kesling and Richard A. Ploch. Pages , with 3 plates. 13. Middle Ordovician Black River Ostracods from Michigan, Introduction and Part I, The Nature of Macronotella, by Robert V. Kesling, Frederick S. Crafts, David G. Darby, Kenneth E. Shubak, and Raymond N. Smith. Pages , with 3 plates. 14. Internal Structures in Two Edrioasteroid Species, Zsorophus ciltcinnatiensis (Roemer) and Carnyella pilea (Hall), by Robert V. Kesling and Leigh W. Mintz. Pages , with 9 plates.
3 VOL. XV, NO. 14, pp , (9 pls., 4 figs.) NOVEMBER 30, 1960 INTERNAL STRUCTURES IN TWO EDRIOASTEROID SPECIES, ZSOROPHUS CINCZNNATIENSIS (ROEMER) AND CARNEYELLA PILEA (HALL) BY ROBERT V. KESLING and LEIGH W. MINTZ CONTENTS Introduction 315 Previous work 316 Isorophus cincinnatiensis (Roemer) 320 Aboral surface 320 Polished surfaces 323 Carneyella pilea (Hall) 327 Aboral surface 327 Polished surfaces 330 Conclusions 336 Literature cited 338 Plates.(after) 339 INTRODUCTION T o LEARN MORE about the internal structures in the edrioasteroids Zsorophus and Carneyella, a well-preserved specimen of each of their type species was ground down and studied as a series of polished surfaces. Photographs of these polished surfaces are reproduced in Plates 111-IV and VII-VIII. They show the relationships of structures between the oral and aboral surfaces in these edrioasteroids. Much of our knowledge of internal structures of edrioasteroids has been based on examination of the very few specimens in which the aboral surface is exposed. Previously only one paleontologist attempted grinding down an edrioasteroid to see how aboral and oral features fitted together-mr. W. K. Spencer, who in 1904 did pioneer work on the morphology of Carneyella pilea (Hall). Although study of internal structures from polished surfaces has been sorely needed, no paleontologist has had well-preserved specimens that he was willing to sacrifice for such a project. Edrioasteroids are rare fossils. Only in 1957, when Mr. William Deak collected several hundred from
4 316 KESLING AND MINTZ strata exposed during construction work on Hamilton Avenue, Cincinnati, Ohio, were specimens of Isorophus cincinnatiensis (Roemer) and Carneyella pilea (Hall) available in such numbers. Mr. Deak presented his collection to the Museum of the University of Cincinnati. Recently, Dr. Kenneth E. Caster, who has charge of the Museum, consented to our using one specimen of each species for this study. We are sincerely grateful for this opportunity. Selection of the specimens to be ground down posed a serious consideration. Well-preserved edrioasteroids are very few, but only such specimens can yield reliable information on internal structures. Whereas, to destroy an excellent edrioasteroid is profane to a paleontologist, to acquire data from any other is definitely not sacred to science. In this dilemma we compromised, choosing specimens with good preservation in most of the peristomial region, posterior interambulacrum, and the proximal parts of the ambulacra, but with certain minor defects. We do not believe these defects have influenced the validity of our interpretations. We wish to express our deep gratitude to the generosity of Dr. Kenneth E. Caster and to the perspicacity of Mr. William Deak, without which this study could not have started. Dr. Caster loaned specimens from the University of Cincinnati Museum, Dr. Matthew H. Nitecki from the Walker Museum of Paleontology at the University of Chicago, and Dr. Donald I?. Squires from the American Museum of Natural History. Dr. Chester A. Arnold and Dr. George M. Ehlers critically read the manuscript and offered helpful suggestions. To all who have helped us we are grateful. Repositories are identified by the following abbreviations preceding the catalogue numbers: UCM, University of Cincinnati Museum; UCWM, University of Chicago, Walker Museum; and AMNH, American Museum of Natural History. PREVIOUS WORK In this review and summary of published information on the internal structures of edrioasteroids, we have limited most of our consideration to Isorophus cincinnatiensis, Carneyella pilea, and other species of these genera. In 1873 (pp ) Meek described the aboral surface of a specimen which he assigned to "Agelacrinites (Lepidodiscus) Cincinnatiensis Roemer" but did not illustrate it. His observations on the flooring plates were correct. This description, because of its historical significance, we quote: I have before me a specimen, a little more than an inch in diameter, apparently of this species, belonging to L. B. Case, Esq., of Richmond, Indiana (found by him at that place), that seems to have grown on one of the valves of an Ambonychiu. The
5 EDRIOASTEROID INTERNAL STRUCTURES 317 shell had separated in such a manner as to take with it the under side of the Agelacenites, and leave its upper side in the matrix so situated as to expose its inner surface. The inner side of each arm or ray is here seen to be composed of a single series of quadrangular pieces that are not imbricating... The whereabouts of this specimen are now unknown. Miller and Faber in 1892 described the edrioasteroid catalogued as UCWM No for the first time. Subsequently, Foerste (1914) redescribed and illustrated it, and in this paper we discuss and show it in our Plate V, Figures 1-2. In this specimen of Carneyella pilea, Miller and Faber evidently based their observations of ambulacral plates on ambulacra IV and V, which have the flooring plates missing, for they confused the aboral sides of the covering plates with the flooring plates. They stated (1892, p. 85) that "The under side of the rays, as seen from below, consist of a row of plates on each side of the furrow, which interlock at the bottom of the furrow, and are, therefore, without reference to abutting imbricating plates, pentagonal instead of quadrangular, as Meek described them." Apparently Miller and Faber referred to the outer ends of the covering plates as they continued (p. 85): "They extend beyond the margin of the imbricating plates into the visceral cavity half the depth of the ambulacral furrows, and are separated from each other, in their extensions laterally into the visceral cavity, so as to present a strongly serrated edge, as shown in the illustration (Miller and Faber, P1. 1, Fig. 10)." They probably spoke of the flooring plates, but said: "... the plates are so small and the sutures so indistinct, that they could not be shown, except in a greatly magnified view.'' From their illustration, we assume Miller and Faber regarded the flooring plates, which they called "the covering," as composed of a mosaic of very small plates, a misinterpretation that we cannot understand, because the sutures are clearly defined (see our Plate V, Fig. 2). Miller and Faber also (1892, p. 85) presented a curious analysis of the frame around the peristome. They described a subpentagonal rim in the center of the specimen formed by the union of the "coverings of the rays," which they interpreted to be the structure by which the edrioasteroid attached itself. They correctly noted, however, that (p. 85), "Within this pentagonal rim there is a pit showing the five subovate mouths of the ambulacral canals..." W. K. Spencer (1904, pp , Figs. 8-12) was the first investigator to grind down an edrioasteroid to learn about the internal structures. From photographs of successive polished surfaces of a specimen of Carneyella pilea, he prepared wax models of the plates. His work on the aboral parts of the ambulacral covering plates agrees very well with our conclusions. He also correctly demonstrated the nature of the ambulacral flooring plates
6 318 KESLING AND MINTZ and the frame around the peristome. He did not, however, note the hydropore or passageway for the stone canal, structures which are conspicuous in our polished surfaces. Nor did he mention or illustrate the aboral configuration of the peristomial covering plates. Foerste (1914, pp , , P1. 1, Fig. 5; P1. 2, Fig. 4) redescribed and reillustrated the specimen studied by Miller and Faber. He correctly observed the relationships of the ambulacral covering and flooring plates, described the frame around the peristome (which he called the "substomial chamber"), and called attention to the structure that we interpret to be the passageway for the stone canal. Of the last he stated (1914, p. 429): "This leaves to be accounted for a peculiar margined depression along the proximal part of the right hand margin of the right posterior ray (No. 5)' as viewed from below. This impression involves the two proximal covering plates on the left side of the right posterior ray, while adjoining the right margin of the large posterior peristomial plate, as seen from above. Possibly a duct passed by this path, but its presence could not be verified with confidence." Bather (1915a, p. 55) accepted Spencer's (1904) interpretation of the flooring and ambulacral plates in Carneyella pitea. Later in the same year, he referred to Foerste's (1914) article and repeated many of Dr. Foerste's statements and conclusions. Concerning the "peculiar margined depression" described by Foreste and quoted in the paragraph above, Bather ( 19156,c, p. 266) said: "If, as Dr. Foerste suggests, 'a duct passed by this path,' then the duct in question would most naturally be the hydropore-canal." Williams ( 19 18) discussed edrioasteroid morphology based on much new material. Although he had only a fragment of Carneyella pilea and no Isorophus cincinnatiensis with the aboral side exposed, he had available many specimens of the related Isorophus austini (Foerste), which he called (with variations in punctuation and capitalization) "Agelacrinites austini var. lawshe," many of them with the aboral surface uppermost. He said (p. 70) that "The aboral view of the large peristomial plate P in the posterior interradial space has been obtained in several specimens [of his A. austini var, lawshe].... Foerste's description of it as ridged like the letter W on its surface toward the substomial cavity gives a good idea of that surface." On the following page he stated: "In the best preserved of the Lawshe specimens the substomial chamber is bounded definitely by the five first flooring plates with their projecting lateral processes and the posterior peristomial plate." Williams described the outer ends of ambulacral covering plates as follows (p. 73) : The condition of cover plates in Miller and Faber's specimen of A. pileus as described by Foerste is not common either for the specimens of A. pileus at hand or for
![EDRIOASTEROID INTERNAL STRUCTURES A. austini.... In the Lawshe specimens the aboral [outer] end of cover plates is shown in many cases where the floor plate of the ray has slipped.](/docs-images/94/119258322/images/7-0.jpg "The plate ends rather squarely with two enlargements one the point of articulation with the flooring plate and one more oral in position, which must have been for muscular attachment.")
7 EDRIOASTEROID INTERNAL STRUCTURES A. austini.... In the Lawshe specimens the aboral [outer] end of cover plates is shown in many cases where the floor plate of the ray has slipped. The plate ends rather squarely with two enlargements one the point of articulation with the flooring plate and one more oral in position, which must have been for muscular attachment. It is this enlargment or a point just above it which must be prolonged as the basal extensions of the lateral covering plates in A pileus Miller and Faber and in A. cincinnatiensis.... Might it not be that in aged specimens there was a deposit in the tissues attached to these plates so as to produce these basal extensions? The last sentence quoted is a strange suggestion. It seems more likely that there are specific differences among the outer processes of Carneyella pilea, Isorophus cincinnatiensis, and I. austini. Anderson ( 1939) described the aboral surfaces of several specimens of Anglidiscus fistulosus (Anderson), which he assigned to Lepidodiscus. He reported (p. 74) that the suture between flooring plates were pierced by pairs of pores. He thought that the radial canal lay below the flooring plates, with extensions passing upward through the paired pores. He also described the peristome (1939, p. 79) as "bounded by truncated triangular plates, the five modified proximal flooring plates of the ambulacra, with the posterior peristomial plate." He also stated (p. 80) that "Slightly posterior to the lobate impression and close to the origin of V is the impression of a small canal running from the anal interradius towards the mouth, which may represent the stone-canal as it lies in the position postulated for that organ by Bather." Ehlers and Kesling (1958, pp ) described a structure that they considered might be a genital pore in the posterior interambulacrum of Discocystis laudoni Bassler. In one of the cotypes they found a polygonal depressed area where apparently one or more plates are missing and which contains a narrow, elongate opening bordered by relatively small plates. In the other cotype, this area is overlain by one large polygonal plate. Since they knew of no other edrioasteroid structure save the peristome and ambulacra in which the plates of the oral surface are underlain by additional plates, they suggested the possibility that this structure is a genital pore which (p. 270) "opened below a protective interambulacral plate and connected with some kind of duct leading out between the interambulacral plate and the subthecal plates." They postulated that a groove anterior to the opening may have served as the duct. Kesling (1960, pp. 158, ) identified the raised structures described by Foerste in Isorophus cincinnatiensis and Carneyella pilea as hydropores. He also established the location of hydropores in several other edrioasteroids and (pp ) catalogued hydropores in six groups according to their relative position. Both Carneyella and Ismophus were classed (p. 173) as having the "hydropore along the juncture between a
8 320 KESLING AND MINTZ short proximal ambulacral plate and the right posterior plate of the peristomial region." The hydropore in Carneyella pilea, he reported (p. 167), involves three plates: the proximal covering plate on the left side of ambulacrum V, the posterior peristomial plate, and the unpaired right posterior peristomial plate. The hydropore in Isorophus cincinnatiensis was described similarly (pp ), except that it has an extra peristomial covering plate between the anterior parts of the posterior peristomial plate and the ambulacral covering plate. Isorophus cincinnatiensis (Roemer) Aboral Surface (Pl. I) Insofar as we are aware, the only specimen of this species ever reported in which the aboral surface was exposed is that described by Meek (1873, pp ), quoted in this paper under "Previous Work." It was part of a private collection, and was not subsequently referred to directly. Among the specimens in the collection of the Museum of the University of Cincinnati is UCM No , illustrated in our Plate I. This edrioasteroid lacks the peripheral rim and the distal tips of ambulacra I1 and 111, but otherwise it is remarkably well preserved. In particular, the peristomial region, the proximal parts of all ambulacra, and the posterior interambulacrum are in excellent condition. Our notes on the aboral surface of Isorophus cincinnutiensis are based on this specimen. Frame around peristome.-as seen aborally, this is composed of the proximal parts of the first flooring plates of the ambulacra, disposed in a ring that surrounds the peristome except posteriorly, where there is a gap. The edges of the flooring plates of ambulacra 11-IV form a smooth arch, nearly semicircular but with the ends curved outward. The junctions between these three plates are narrow (Pl. I, Fig. 2). In contrast, the junctions of the first flooring plates of ambulacra I and 11 and of ambulacra IV and V are both wide. Ambulacra I and V are not in contact. Edges of the flooring plates of ambulacra 11-IV are raised (as seen aborally) to form a continuous rim around the anterior border of the peristome. The raised edges of ambulacra I and V are nearly mirror images. The outer part of each is a single ridge, but the inner part bifurcates into narrow proximoanterior and proximoposterior ridges. Although the peristome is now partly filled with matrix, obscuring the opening of the ambulacra, it appears that the ambulacra proper, the tunnels formed between the flooring and covering plates, led directly from the peristome. We suppose that the gut extended distally through the gap in
9 EDRIOASTEROID INTERNAL STRUCTURES 321 the posterior part of the frame. Whether it followed a straight course to the anal pyramid or coiled once around, like that in living sea urchins, is conjectural. Passageway fm stone canal.-as viewed aborally, the edrioasteroid has a subtriangular funnel next to the proximal inner side of ambulacrum V, which is interpreted to be the passageway for the stone canal. The innermost wall of this funnel (Pl. I, Fig. 1) is narrow and steep, projecting posteriorly from the inner end of the proximoposterior ridge of the first flooring plate of ambulacrum V. The posterior wall slopes orally at a much lower angle. The third wall is formed by the inner edges of the first two flooring plates of ambulacrum V. The whole funnel, or passageway, is considered to be part of the peristomial region because (1) it does not include any of the imbricating plates characteristic of the posterior interambulacrum, (2) the plate forming the posterior wall extends orally beyond the level of the posterior interambulacrum, and thus seems clearly to be associated with the other peristomial covering plates, and (3) the innermost wall of the funnel appears to be a crest on the aboral side of one of the large posterior peristomial covering plates. Radial canal into ambulacrum V.-The proximal part of ambulacrum V has flooring plates arranged differently from those in the distal part. This arrangement is not found in other ambulacra. As viewed aborally, the first two flooring plates lie below the level of the others, and the proximomedian* margin of the third flooring plate appears to bear an opening into the ambulacrum. It is more than coincidence, we believe, that this arrangement occurs in the ambulacrum nearest to the passageway for the stone canal. The water-vascular system of edrioasteroids, we assume, was similar to that of living echinoderms, consisting of a hydropore (or madrepore), stone canal, ring canal, and radial canals into the ambulacra. When one considers that the aboral end of the stone canal was outside the frame around the peristome, the ring canal could hardly have been circular. If it were, it would have to cross under the frame twice, one at ambulacrum V and again at ambulacrum I. Any compression of the flexible theca would pinch the ring canal between the frame and the object to which the edrioasteroid was attached. More likely, the posterior part of the ring canal was compressed so that it passed through the gap in the frame between ambulacra V and 1. Such a circuit, however, would place the radial canal to ambulacrum V far from the stone canal, separated from it by the distance from the passageway to the gap in the frame and thence to the ambulacral opening. It is our *In this paper the terms for orienting ambulacral structures are used differently from those for other parts of the edrioasteroid. Prmimd is along the ambulacrum in the direction leading albeit circuitously, toward the peristome; distal is the opposite of proximal, leading away from th; peristome; median, toward the midline of the ambulacrum; and outer, the opposite of median, away from the midline.
10 322 KESLING AND MINTZ theory that, as the frame, the ambulacra, and the water-vascular system evolved, the radial canal to ambulacrum V came to bypass the first two flooring plates in Zsorophus cincinnutiensis. The structure of arnbulacrum V indicates that the radial canal joined the ring canal very close to the junction of the latter with the stone canal and entered into ambulacrum V through the opening at the proximomedian margin of the third flooring plate. In the other four ambulacra, the radial canals seem to have entered from the peristome. Outer ends of ambulacral covering plates.-this specimen also shows the tips of the ambulacral covering plates extending beyond the flooring plates (PI. I, Fig. 1). As Foerste suggested (1914, p. 423) from his study of oral surfaces in which the adjacent interambulacral plates were missing: "Immediately beyond the point where the covering plate passes beneath the adjacent interambulacral plate, the covering plate begins to narrow rapidly, and then the sides become approximately parallel, forming the basal extension, which only slightly exceeds one-fourth the total length of the plate." The outer ends of the covering plates are not nearly as long nor as broad as those in Carneyella pilea. Ambulacral flooring plates.-these plates taper throughout the length of the ambulacra. Except in curved parts of the ambulacra, where they are trapezoidal, all plates except the proximal are subquadrate. The modification of the proximal two flooring plates in abulacrum V has already been mentioned. The plates fit closely together, and no evidence of pores through their junctions could be found. Interambulacral plates.-these imbricate on the aboral surface in the opposite manner to that on the oral surface, that is, overlapping with their distal edges rather than proximal. Each plate has a faint crest along its distal margin (Pl. I, Fig. 1). Anal pyramid.-each plate in the anal pyramid bears an aboral ridge tapering from its outer border to the center of the pyramid. Possibly, these ridges served as places of attachment for closing muscles. Although the plates of the pyramid are clearly differentiated from those of the surrounding posterior interambulacrum, we could not distinguish the way in which the two kind of plates articulated. Genital duct?--near the left anterior margin (as seen aborally) of the anal pyramid, the posterior interambulacrum appears to consist of two layers of plates (PI. I, Fig. 2). This disposition of plates is very similar to that described in the corresponding area in Discocystis laudoni by Ehlers and Kesling (1958, p. 270): In No. S3SS6b there is a structure in the posterior interambulacrum that has the general form of a pore, and its position suggests that it may have been the genital pore.
11 EDRIOASTEROID INTERNAL STRUCTURES 323 This feature lies to the right of the anal pyramid. The posterior interambulacrum has a polygonal depressed area, apparently where one or more large interambulacral plates are missing from the specimen, which contains an elongate, narrow opening bordered by plates. The bordering plates are much smaller than those in the adjacent area. In No. S3886a, the corresponding area is covered by one large polygonal plate. We can discern no opening between the two layers of plates, but if one were present it would probably be filled with matrix and obscured. Polished Surfaces (Figs. 1-2 ; Pls. 11-IV) The specimen selected for polished surfaces, shown before grinding in Plate 11, has a few defects in preservation. The right anterior peristomial covering plate was found to be shoved inward and twisted, so that the oral part was out of position (Pl. 111, Figs. 2-3) and the aboral termination was no longer properly inserted between the expanded proximal edges of the first flooring plates of ambulacra I11 and IV (Pl. 111, Fig. 7). The proximal part of ambulacrum I had been elevated, so that it was the first part of the specimen to be ground off (PI. 111, Figs. 1-6). Some of the plates in interambulacrum I were tipped up, out of their normal relationship to other plates. Apart from these defects, the specimen had very good preservation in the ambulacra, the peristomial region, the interambulacra, and the peripheral rim. The anal pyramid was complete and undistorted. An interesting feature of the specimen was a small round hole drilled through the plates, partly in interambulacrum 3 and partly in the left side of ambulacrum IV (Pl. 11, Figs. 1-2 ; P1. 111, Figs. 1-8). We assume that this hole was made by some predator; possibly, much of the edrioasteroid's soft parts were thus eaten, or the physiological damage resulted in death. We point out that the most elevated ambulacrum, as fossilized, is the one diametrically opposite the hole. Peristomial covering plates.-the left anterior plate, which was preserved in place, has its proximal edges sloping aborally away from the outer surface (Figs. 2a-b; P1. 111, Figs. 1-3). A longitudinally narrow but transversely wide aboral extension of this plate is inserted in a notch between the expanded proximal edges of the flooring plates of ambulacra I1 and I11 (Figs. 2c-d; P1. 111, Fig. 7). The large posterior plate bordering the hydropore has a thin bladelike process on its aboral side, which forms the left side of the passageway for the stone canal (Fig. 26; PI. 111, Figs. 3-5). Frame around peristome.-as correctly interpreted by Spencer (1904), Foerste (1914), and Williams (1918), the frame around the peristome on
12 324 KESLING AND MINTZ the aboral side in Carneyella and Zsmophus is composed of the expanded proximal edges of the first flooring plates of the ambulacra (Fig. 2e). Just above the aboral surface of the peristome, however, the inserted edges of the anterior peristomial covering plates form integral parts of the frame in FIG. 1. ZSOYO~~US ciminnatiensis (Roemer). Restoration of oral view of specimen ground down and studied as a series of polished surfaces. Photographs of this specimen are shown on Plate 11. interambulacral positions, as described in the paragraph above (Figs. 2c-d; P1.111, Fig. 7). Passageway for stone canal.-from the hydropore, a short triangular funnel expands aborally (Fig. 2a; P1. 111, Figs. 1-4). Near the hydropore, this opening is bordered only by the same three plates which form the hydropore (Pl. 111, Figs. 3-4), but below this it is also bordered by other plates of ambulacrum V (Fig. 26; P1. 111, Fig. 5). The bladelike process on the large posterior peristomial plate, as noted above, forms the left wall of the passageway.
.")
13 EDRIOASTEROID INTERNAL STRUCTURES 325 Radial canal into ambulacrum V.-The third flooring plate of ambulacrum V shows a narrow opening passing through it radially (Fig. 2c; P1. 111, Figs. 8-9). As we analyze the series of polished surfaces, this opening led proximally on a diagonal from the ambulacral groove to the aboral sur- FIG. 2. Isorophus cincinnatiensis (Roemer). Restorations of oral views of selected polished surfaces of the specimen shown in Fig. 1 and PI. 11. a, based on Pl. III, Figs. 1-3 ; b, based on PI. 111, Fig. 5; c, based on P1. 111, Figs. 6-9; d, based on PI. III, Figs. 7-8 and P1. IV, Fig. 1; and e, based on P1. 111, Fig. 8, and P1. JY, Figs face of the ambulacrum. Other ambulacra and other plates of ambulacrum V have no such opening. Because the distal end of this tubelike opening lies in the groove of the ambulacrum and the proximal end lies at the location of the opening observed on the aboral side of the ambulacrum in UCM No (see P1. I, Fig. 1-2), we believe this opening accommodated the radial canal into ambulacrum V, bypassing the first two flooring plates.
14 326 KESLING AND MINTZ Ambulacral covering plates.-aborally the small median covering plates expand so that they are only slightly narrower than the outer covering plates and their ends extend as far as the outer border of the flooring plates (PI. 111, Fig. 9). This agrees generally with the statement by Williams (1918, p. 68) that "the outer covering plates [in I. cincinnatiensis] are grooved on their sides and the cylindrical median plate fills the space left by the grooving of two adjacent lateral plates. Its free end only is visible, alternating with the opposite median plate and both entirely covered from the side by the two rows of lateral covering plates." We did not, however, observe any aboral increase in width of the outer covering plates, as would be the case if they were grooved on their sides. Instead, they appear to narrow gradually to their aboral sides. As we interpret the junctions of the two kinds of covering plates, the edges of the larger overlap orally the beveled edges of the intercalated smaller. Only the larger covering plates have outer processes extending beyond the outer border of the flooring plates, and these are noticeably shorter than those in Carneyella pilea. Williams ( 1918, p. 73) found no outer processes on any covering plates in Isorophus austini; hence it is possible that, despite their lack of projections beyond the border of the flooring plates, the median plates in Isorophus cincinnatiensis were opened directly by muscles, rather than indirectly by the action of the outer covering plates. Ambulacral flooring plates.-the flooring plates are seen in oblique section on many polished surfaces (PI. 111, Figs. 6-9; P1. IV, Figs. 1-6). All suture lines appear as straight lines on the surfaces, indicating that the junctions between flooring plates are essentially vertical planes. Although the junctions are not closely fitted, we have seen no evidence of pores along them. The ambulacra have well-defined median grooves (Figs. 2c-d; P1.111, Figs. 6-8). Genital duct?-as we ground through the region of the anal pyramid, we looked for a double layer of plates, like that observed in UCM No in aboral view. Our results were inconclusive. The plates near the right anterior edge of the anal pyramid seem to be thicker than other interambulacral plates (Pl. IV, Figs. 4-S), but it is possible that there were two layers at this place and the junction was passed through in grinding without our being aware of it. Another specimen will have to be sacrificed at some future time to decide whether there is a structure in this region that could qualify as a genital pore. Other features.-the larger plates in the anal pyramid have beveled edges that fit over the sides of the smaller plates. On the aboral side, the plates are nearly equal in width. This arrangement would permit the plates to remain in contact even when the pyramid was at least partly open,
15 EDRIOASTEROID INTERNAL STRUCTURES 327 The sides of the interambulacral plates taper rather abruptly to sharp edges, which fit closely between the similarly tapered edges of the laterally adjacent plates immediately distal and proximal. Such overlapping of the margins would still retain a "shingled" covering of the interambulacra when the theca was considerably inflated. The large plates in the proximal part of the peripheral ring fit together in the same manner as those of the interambulacra. Because they stand at a rather high angle on the inner margin, each polished surface intersects the peripheral ring through a thickness of about four plates (Pl. IV, Fig. 6). Carneyella pilea (Hall) Aboral Surface (PIS. v, 1x1 Our description of the aboral surface in Caryneyella pilea is based on two specimens, both of which were studied, described, and figured by Foerste ( 1914, pp , P1. 1, Fig. 5 ; P1. 2, Figs. 3-4), UCWM NO (here shown in P1. V) and AMNH No x (here shown in PI. IX). In the first of these specimens (Pl. V, Figs. 1-2)) the posterior interambulacrum, including the anal pyramid, is poorly preserved, ambulacra I and I1 are distorted, the distal end of ambulacrum IV is crushed, all the flooring plates of ambulacra IV and V are missing, and only the proximal flooring plates of ambulacra I1 and I11 are present. Despite these deficiencies, this specimen provides valuable information on the frame around the peristome, the passageway for the stone canal, and the relationships of the ambulacral covering and flooring plates. In the second specimen (Pl. IX, Figs. 1-2), the left posterior region is missing from the middle of the posterior interambulacrum to ambulacrum 111. The anal pyramid is present, but not clearly exposed. The frame around the peristome is distorted on the left side of the specimen (the right side as seen aborally) and incomplete posteriorly. In contrast, ambulacra 111-V are complete and interambulacra 2-4 are well exposed. The entire aboral surface of the specimen is somewhat weathered. Frame around peristome.-although the frame is more nearly complete in AMNH No x (PI. IX, Figs. 1-2)) it is more distorted than that in UCWM No (Pl. V, Figs. 1-2). From a study of both, however, nearly all of the characteristics of the frame can be learned. It is noteworthy that the frame in Carneyella pilea is not proportionally much larger than that in Isorophus cincinnatiensis (compare P1. IX, Fig. 1, with P1. I, Fig. I), although the peristomial region as seen orally is much larger in C. pilea than in I. cincinnatiensis (compare Fig. 3 with Fig. 1). One may
16 328 KESLING AND MINTZ conclude that the peristome in Carneyella does not expand as much aborally as that in Isorophus. Insofar as we may judge from AMNH No x, the lateral tips of the frame do not project beyond the junction of ambulacra I and I1 and ambulacra IV and V, as they do in Isorophus cincinnatiensis. The part of the frame formed by the edges of the flooring plates of ambulacra 11-IV is semicircular. In AMNH No x (Pl. IX, Fig. 2) these edges are much narrower than those in UCWM No (Pl. V, Fig. 2), leading us to believe that the aboral surface of frame in the latter has been planed off by abrasion. In fact, the configuration of the suture lines between the anterior peristomial covering plates and the preserved parts of the proximal flooring plates in ambulacra 11-IV strongly suggest that the aboral surface has been abraded nearly to the level at which the peristomial plates are inserted in the frame (PI. V, Fig. 2). The radially directed aboral ridges on the anterior peristomial covering plates can be seen in both specimens, but they are especially prominent in UCWM No The aboral surface of the large posterior peristomial covering plate, which lies in the posterior gap in the frame, is better preserved in UCWM No (Pl. V, Fig. 2), although we strongly suspect, by comparison with polished surfaces (Pl. VII, Figs. 9-10; PI. VIII, Figs. 1-9), that the median aboral ridge in this specimen has had its distal part broken off. Passageway for stone canal.-the aboral opening of the passageway is clearly visible in both specimens, along the right side (as viewed aborally) of ambulacrum V, between the flooring plates of the ambulacrum and the large posterior peristomial covering plate. AMNH No x, in which the flooring plates of ambulacrum V are preserved, does not show clearly the deep indentation shared by the first two flooring plates as seen on polished surfaces (PI. VIII, Figs. 5-10). We assume that this is partly filled with matrix. Ambulacral covekng plates.-as pointed out by Foerste (1914, p. 422), in UCWM No. 8825, "the basal extensions are wider on the convex side of the rays and narrower on the concave side, where there is less room." Foerste also described in this specimen (p. 422): "Along that part of the base of each covering plate which serves as its fulcrum, there are two striations, which extend in a direction parallel with the ray, and a moderate distance apart. These striations seem to fit against the inner margin of the narrow lateral groove which is seen on the upper side of the flooring plates in certain species." In our opinion, the "striations" are simply the two borders of a low ridge, which crosses the aboral "base" of each covering plate in a proximo-distal direction (PI. V, Fig. 1). Without a coating of sublimated ammonium chloride (as in P1. V, Fig. 2) these ridges are scarcely

17 EDRIOASTEROID INTERNAL STRUCTURES 329 discernible; for this reason, they may have been overlooked by Foerste. These ridges, as Foerste suggested, probably served as fulcra for the plates, fitting into grooves on the oral margins of the flooring plates. Aborally, each covering plate is seen to have a median ridge projecting from the proximomedian side of its base (PI. V, Fig. 2). These structures are not clearly indicated on any of the polished surfaces, possibly because they are very thin. Considered in conjunction with the "braided" pattern of projections on the aboral sides of the covering plates (Pl. VII, Figs. 4-10), however, they indicate that the covering plates of one side fit with those of the opposite side much in the manner of the teeth in a zipper, so that Foerste's suggestion (1914, p. 412) that each tooth "interlocks with the opposing" may be correct. This arrangement is further discussed below under "Polished Surfaces." Flooring plates.-adjacent flooring plates have rather close-fitting junctions. We can find no evidence of any pores between plates. The first two flooring plates of ambulacrum V (Pl. IX, Fig. 2) have an indentation between their right parts (as viewed aborally), but they are not depressed below the level of the other flooring plates of the ambulacrum as are the corresponding plates in Isorophus cincinnatiensis (Pl. I, Fig. 2). Interambulacral plates.-these plates differ in appearance in UCWM No (PI. V, Fig. 1) and AMNH No x (Pl. IX, Fig. 1). In the former the surfaces appear to overlap successively outward, whereas in the latter they appear to overlap successively inward. In other words, the direction of imbrication of plates in one seems to be opposite to that in the other. The plates in the American Museum specimen (PI. IX, Fig. 1) seem to retain ornamentation on their aboral surfaces and are much thicker than those in the Walker Museum specimen (Pl. V, Fig. I), which are now quite thin, possibly the result of corrosion. The difference leads us to believe that the American Museum specimen retains interambulacral plates more nearly in their original form. If this is true, then each ambulacral plate must taper to a thin edge distally, fitting into a groove on the proximal edge of the distally adjacent plate. Thus, the plates on both oral and aboral sides would appear to imbricate or overlap in the same manner. With such an arrangement the theca of the edrioasteroid would have an aboral limit to which it could be compressed without distortion or disruption of the plates. Other features.-the anal pyramid is crushed in the Walker Museum specimen (Pl. V, Fig. 2) and partly filled with matrix in the American Museum specimen (PI. IX, Fig. 2). We can learn very little about the anal pyramid and the surrounding plates. There is no apparent double layer of plates near the pyramid, as was found in the specimen of Ismophus cincinnutiensis.
18 330 KESLING AND MINTZ Polished Surfaces (Figs. 3-4 ; Pls. VI-VIII) The specimen ground down is shown in its entirety in Plate VI. It is far from perfect. The left posterior part of the peripheral ring, the outer halves of interambulacra 1 and 3, the middle one-third of ambulacrum 11, FIG. 3. Carneyella pilea (Hall). Restoration of oral view of specimen ground down and studied as a series of polished surfaces. Photographs of this specimen are shown on Plate VI. and the distal two-thirds of ambulacrum IV are missing. In addition, the middle of ambulacrum I11 is bent and broken and plates are missing in interambulacrum 4. The specimen is adequate for our study, however, because the central part, posterior interambulacrum, and ambulacra I and V are complete and well preserved. Internally, we found, the aboral side of the frame around the peristome is cracked and slightly distorted and the flooring plates of ambulacrum IV are somewhat askew. Inasmuch as the corresponding structures are in good condition in other parts of the edrio-
. Restorations of oral views of selected polished surfaces of the specimen shown in Fig. 3 and P1. VI. a, based on PI. VII, Fig. 3 ; b, based on PI. VII, Fig. 6; c, based on P1.")
19 EDRZOASTEROZD INTERNAL STRUCTURES 33 1 asteroid, we do not consider these defects as serious obstacles in our investigation. Peristomial covering plates.-these plates can readily be followed FIG. 4. Carneyella pilea (Roemer). Restorations of oral views of selected polished surfaces of the specimen shown in Fig. 3 and P1. VI. a, based on PI. VII, Fig. 3 ; b, based on PI. VII, Fig. 6; c, based on P1. VII, Fig. 10, and PI. VIII, Figs. 1-2; d, based on P1. VIII, Figs. 6-9 ; and e, based on P1. VIII, Figs through the sequence of polished surfaces, since there are only four and each is large and distinctive. The two anterior peristomial covering plates are nearly mirror images and lie in interambulacral positions (Fig. 4a; P1. VI, Figs. 1-2; P1. VII, Figs. 1-3). Near the oral surface, the arrangement is like that described and illustrated by Spencer (1904, p. 40, Fig. 9). Proximally, each plate has thin edges and a thick central part. A short distance below the surface, how-
20 332 KESLING AND MINT2 ever, when the thin edges have been ground away, only the thick central part projects toward the center of the peristome (Fig. 4b; P1. VII, Figs. 5-6). The thick central parts of these two plates are the two radially disposed ridges which are so prominently displayed on the aboral surface of UCWM No (Pl. V, Fig. 2). Still deeper, the proximal tips of these ridges are ground off, and each plate becomes subrhomboidal (Pl. VII, Figs. 7-8). The distal parts of the two anterior peristomial covering plates are much thicker. In its aboral part, each of the plates has a cross section in the shape of the head of a battle-ax (Fig. 4c; P1. VIII, Fig. 1-4). The aboral terminations of the plates are wedge-shaped, inserted in notches between the expanded proximal edges of the flooring plates as units in the frame around the peristome (Pl. VIII, Figs. 3-4). The large posterior peristomial covering plate is complex. Its anterior edge (at the junction with the two anterior covering plates just described) is nearly a straight line on the oral surface (Pl. VI, Fig. I), but a short distance below this surface it becomes a broad V (Pl. VII, Figs. 2-3). At the oral surface this plate is bounded on the left by one covering plate of ambulacrum I (Pl. VII, Figs. 1-2), but below it is bounded by two of these covering plates (Fig. 4b-c; P1. VII, Figs. 5-7). At the oral surface the anterior margin of the plate is nearly straight (PI. VI, Fig. 2) ; deeper, it is V-shaped (Pl. VII, Figs. 3-4) ; still deeper, it has a central anterior projection (Fig. 4b; P1. VII, Figs. 6-7) ; and, aborally, it is W-shaped (Fig. 4c; PI. VII, Figs. 8-10). The central anterior projection is long and deep (Pl. VIII, Figs. 1-9). We suspect that part of it is broken off in UCWM No (Pl. V, Fig. 2). Aborally, the proximal tips of the plate are notched (Pl. VII, Figs. 7-10), just as in UCWM No (Pl. V, Fig. 2). This plate forms the left wall of the passageway for the stone canal, but does not extend nearly to the aboral end of the passageway (Pl. VII, Figs. 3-10). The small posterior peristomial covering plate forms the rear boundary of the hydropore (Pl. VII, Fig. 1) and the rear wall of the passageway for the stone canal (Pl. VII, Figs. 3-10). It is a triangular plate that slopes down posteriorly. Proximally, it is nearly flat (Pl. VII, Figs, 3-5), but distally it is slightly arched, so that it is crescentic in cross section (Pl. VII, Figs. 9-10; P1. VIII, Fig. 1). The distal end of this plate appears to be underlain directly by a lateral projection from the second flooring plate of ambulacrum V (Pl. VIII, Figs. 4-5) ; however, the exact limits of the plate could not be determined. Frame around peristome.-the frame is composed of the expanded proximal edges of the first flooring plates of the ambulacra and the distal parts of the two anterior peristomial covering plates. The former are the
21 EDRIOASTEROID INTERNAL STRUCTURES 333 only plates in the aboral part of the frame (Pl. V, Fig. 2), and the latter are inserted like tenons a little higher in the theca. Near the aboral limit of the peristome, the polished surfaces intersect only the liplike edges of the flooring plates (Fig. 4e-f; P1. VIII, Figs ). Slightly higher, they intersect the expanded proximal margins of the flooring plates Fig. 4d; P1. VIII, Figs. 7-9), and, still higher, they also cut through the peristomial covering plates (Pl. VIII, Figs. 3-4). The covering plates, as pointed out above, have thick distal parts which are wedge-shaped aborally. The left anterior covering plate is inserted between the flooring plates of ambulacra I1 and 111, whereas the right anterior covering plate is inserted between ambulacra I11 and IV. Passageway for stone canal.-the passageway for the stone canal is much longer in Carneyella pilea than in Isorophus cincinnatiensis. It extends from the hydropore (Pl. VII, Figs. 1-2) to the aboral surface of the first flooring plate of ambulacrum V, involving different plates in the oral and aboral parts. The oral part is a subtriangular funnel expanding downward, formed of the same three plates that bound the hydropore: the two posterior peristomial covering plates and the first covering plate of ambulacrum V (Figs. 4a-b; PI. VII, Figs. 3-6). A little deeper, it is bounded on the right by the first two covering plates of ambulacrum V (Pl. VII, Figs. 7-9). The aboral part is a deep troughlike indentation in the first two flooring plates of ambulacrum V (Fig. 4d; PI. VIII, Figs. 4-9). Each of the first two flooring plates has a long, thin lip projecting toward the left, curved around the sides of the subcircular indentation like the horns of a crescent (PI. VIII, Figs. 4-8). The aboral end of the passageway seems to be an indentation in only the first flooring plate (Fig. 4e; P1. VIII, Figs ). The stone canal, if it extended along the full length of this passageway, projected aborally beyond the adit of the ambulacrum. Radial canal into ambulacrum V.-There is no conclusive evidence on the exact course of the radial canal leading to ambulacrum V. The radial canal evidently entered the ambulacrum where the passageway for the stone canal is confluent with the ambulacrum (Pl. VIII, Fig. 3). Possibly, if the radial canal branched from the ring canal below the peristome, it shared the lower part of the passageway wiiii tlie siolle canal. It is conceivabie, however, that the radial canal branched directly from the stone canal rather than from the ring canal and entered ambulacrum V at the same level (Pl. VII, Fig. 3). Such a circuit is unknown in living echinoderms, but in edrioasteroids the water-vascular system was strongly affected by the frame around the peristome and the oral location of the hydropore.
22 334 KESLING AND MZNTZ Ambu1acra.-Ambulacra I and I1 are confluent at the left side of the peristome, and ambulacra IV and V at the right side (Figs. 4b-c; PI. VII, Figs. 6-10). Ambulacrum I11 joins the peristome directly. Within the peristome, therefore, the passages are broadly trimerous. Ambulacral covering plates.-the shape of the ambulacral covering plates in Carneyella pilea is far more complex than has been described. Spencer (1904, pp ) stated: "Viewed externally the ambulacra of Lepidodiscus pileus are seen to be roofed over by an alternating series of plates which are shaped like a bent finger, fig. 9. These do not completely enclose the groove. There is no trace of any separate side plates. We may therefore call this upper series of plates 'side-covering plates.' " His Fig. 8, a diagram of a cross section of an ambulacrum, shows the oral side of the flooring plate to be a flat surface. This is not so. As shown in our P1. VIII, Figs. 7-10, the flooring plates are definitely channeled, forming the lower half of the ambulacral cavity. Furthermore, Spencer's conclusion that the covering plates do not completely enclose the groove is in error, apparently based on the polished surface shown in his Fig. 9, which lies at some distance below the oral sides of the plates. If the covering plates of his specimen were shaped like a "bent finger" when "viewed externally," we can only conclude that the distal part had been eroded away before he started grinding on it; in well-preserved edrioasteroids, as in the one we ground, the oral aspect of the covering plates is not at all like a "bent finger" (see P1. VI, Fig. 1) and the plates fit closely together. From his observation of isolated plates, Foerste (1914, p. 412) described their median parts as follows: In by far the greater number of specimens the proximal side of each cover plate is elongated into an acute spinous tooth which projects across the median line of the ray and interlocks with the opposing cover plate. The latter also has a spinous prolongation on the proximal side, and both cover plates are curved concavely on the distal side of this spine, along the median line of the ray, so as to admit the interlocking. As shown in a complete specimen (Pl. VI, Fig. 1) these descriptions apply to the aboral parts of the plates but not to the oral surfaces. Bather (1915a, p. 55), accepting the interpretation of Spencer, said that the cover plates of Agelacrinus (in which he included Carneyella pilea), Hemicystis (error for Hemicystites), Cystaster, and Streptaster were "boot-shaped, with the sole of the boot adoral (= proximal) and the toe of the boot admedian, and in consequence of this shape they cannot close in the groove so completely as do the symmetrical cover-plates of the Edrioasteridae." This is an endorsement of error. Successive polished surfaces show that the previous descriptions failed to relate the oral and aboral parts of the covering plates, which form very
 and in the distal part of ambulacrum I (PI. VI, Fig. 1, and P1. VIII, Fig. 13).")
23 EDRZOASTEROZD INTERNAL STRUCTURES different patterns by their junctions. For two examples, compare the oral and aboral parts of these plates in the distal part of ambulacrurn 111 (PI. VI, Fig. 1, and P1. VII, Fig. 9) and in the distal part of ambulacrum I (PI. VI, Fig. 1, and P1. VIII, Fig. 13). On the oral surface, each covering plate has parallel sides and a pointed median end that fits tightly between the ends of the two opposite covering plates. Below this surface, however, each plate has a proximomedian extension that lies between similar extensions of the two opposite plates (Spencer's "bent finger," Foerste's "spinous tooth,'' and Bather's "toe of the boot"). Here, the extensions appear to be "braided" but this is an illusion, since they terminate flush and are not intertwined. Although the junctions between plates on one side and those on the other form a serrate line on both the oral and aboral surfaces, the serrations on the aboral surface are offset proximally from those on the oral surface. At and near the oral surface, the median end of each plate is bluntly acuminate (Fig. 3; P1. VII, Fig. 1) ; slightly below, it is indented in the middle (Fig. 4a; P1. VII, Figs. 2-3); and farther below, it projects proximally and is truncated by the extension of plate opposite and proximal to it (Fig. 4b; P1. VII, Fig. 4). From these relationships we can reconstruct the shape of each plate and explain how it fits against the others. Each plate has a proximomedian extension from the aboral half of its proximal edge. As pointed out correctly by Foerste (1914, p. 412), these extensions project under and beyond the oral position of the median line of the ambulacrum. Each extension fits into a rabbet or recess in the aboral half of the distomedian part of the covering plate lying opposite and proximal to it. Furthermore, it is prevented from moving outward by a small distomedian tip on the aboral half of the opposite and proximal plate. In similar fashion, the plate itself is rabbeted under its distomedian part for reception of the proximomedian extension from the covering plate opposite and distal. As pointed out above in the discussion of the "Aboral Surface," there are median ridges projecting from the proximomedian sides of the bases of the covering plates (Pl. V, Fig. 2), which do not show up clearly on the polished surfaces and which we presume were very thin. In this unusually complicated arrangement, the covering plates seem to have interlocked much like the teeth in a zipper. A plate could move from the dosed positior. only if the plztes sn the epposite side of :he amhlaci-urn also moved, inasmuch as its extension was overlain by a thin oral shelf of the opposing plate and underlain by the thin median end of the ridge from the proximomedian side of the base of the opposing plate. Thus, covering plates on both sides of the ambulacrum had to swing open simultaneously. We think that the median ends of the covering plates were mor-
24 336 KESLING AND MINTZ tised, rather than dovetailed, so that our comparison with a zipper may tend to overemphasize the permanence of their junction in the closed position. It is particularly appropriate, however, as regards the form and relationships of the plates. The significance and use of this complex closure is unknown. It gave great rigidity and strength to the ambulacral cover, particularly in the closed position. It would also have increased the filtering action of the plates in the open position. The outer processes of the covering plates project beyond the flooring plates, as also observed in the aboral view of UCWM No (Pl. V, Fig. 2). As in the latter specimen, also, the processes on the convex side of the ambulacrum are much broader than those on the concave side (PI. VIII, Figs. 7-10, 13). The covering plates are more numerous than the flooring plates, but do not appear to have been spaced on them in any particular arrangement. Ambulacral flooring plates.-except for the first flooring plates of the ambulacra, which form part of the frame around the peristome, the plates taper rather evenly toward the distal end of the ambulacrum. Suture lines between flooring plates are nearly straight on polished surfaces, so that there do not seem to be any configurations on the contact surfaces (PI. VIII, fig. 13). Other features.-the interambulacral plates, like those in Isorophus cincinnatiensis, fit closely together, with the tapered edges of one plate inserted between similar edges of the bordering plates. No evidence on the nature of the genital duct was found in this specimen. CONCLUSIONS From our observations of oral, aboral, and polished surfaces, we conclude that Isorophus cincinnatiensis and Carneyella pdea both have: (1) three plates, two in the right posterior part of the peristomial region and one in the adjacent left side of ambulacrum V, which form a low pyramid and which are underlain directly by a passageway to the interior sf the edrioasteroid. We believe that the three plates formed the hydropore and the passageway below accommodated the stone canal; (2) the aboral part of the frame around the peristome composed of the expanded proximal parts of the first flooring plates of the ambulacra, as previously reported in literature; (3) the aboral ends of the anterior peristomial plates inserted wedgelike between the first flooring plates of the three anterior ambulacra, forming an integral part of the frame around the peristome; (4) a gap in the posterior part of the frame around the peristome, which is overlain by the
25 EDRZOASTEROZD INTERNAL STRUCTURES covering plates of the posterior part of the peristome; (5) ambulacral covering plates with basal extensions, which presumably were for attachment of muscles; (6) flooring plates of the ambulacra which fitted closely together and had no pores along their junctions; and (7) interambulacral plates beveled at their lateral edges to fit between similarly beveled plates lying immediately proximal and distal at the sides. We note as differences between the two species that: (1) the arrangement of ambulacral covering plates is much more complicated in Caryneyella pilea, although it lacks the median series of plates which is found in Isorophus cincinnatiensis; (2) although the peristomial covering plates in Isorophus cincinnatiensis covers a smaller area than that in Carneyella pilea, its peristome expands aborally so that the frame around the peristome is proportionally about the same size in both species; (3) the flooring plates in the proximal part of ambulacrum V have different configurations in the two species; and (4) the basal extensions on the ambulacral covering plates are longer in Carneyella pilea than in Isorophus cincinnatiensis. In Carneyella pilea we found that the bases of the ambulacral covering plates have ridges on their aboral surfaces, presumably for articulation with grooves in the oral sides of the flooring plates, and that the covering plates of one side of an ambulacrum fit with those on the other side much like the teeth of a zipper, preventing the plates of one side from opening or closing independently of those opposite. In Isorophus cincinnatiensis the small ambulacral covering plates increase aborally, so that on the aboral side they are nearly the same size as the larger plates. In like manner, the smaller plates of the anal pyramid also expand aborally. Each plate of the anal pyramid has an aboral radial ridge, thought to be for attachment of muscles. The following four of our interpretations need additional confirmation: f. In I. cincinnatiensis, the aboral view shows a double layer of plates adjacent to the anal pyramid, comparable in position to the structure in Discocystis laudoni described by Ehlers and Kesling (1955) and suggested to be a genital pore. We observed no pore leading between the two layers of plates. 2. In I. cincinnatiensis, the first two flooring plates in ambulacrum V do not extend aborally as far as the others, and the third flooring plate has a tube leading into the ambulacrum. Whether this tube served as the passageway for the radial canal, bypassing the first two flooring plates, as we suggested, is speculative. 3. In C. pilea, there is a deep notch between the first and second flooring plates, on the left side of ambulacrum V. We do not know whether this served as the adit for the radial canal or had some other function.
26 338 KESLZNG AND MZNTZ 4. In C. pilea, comparison of oral and aboral surfaces leads us to believe that each interambulacral plate is distally beveled to fit into a groove of the plate beyond it. This could not be confirmed in polished surfaces. All in all, their type species are constructed so similarly that we seriously question that Zsorophus and CarneyeUa belong to different families of edrioasteroids. LITERATURE CITED ANDERSON, F. W Lepidodiscus fistulosus sp. nov. from Lower Carboniferous Rocks, Northumberland. Bull. Geol. Surv. Great Britain, Vol. 1, pp , 5 pls., 10 figs. BATHER, F. A. 1915a. VI. Pyrgocystis n.g. Geol. Mag., Decade 6, Vol. 2, pp , 3 pls b. VII. Morphology and bionomics of the Edrioasteridae. Geol. Mag., Decade 6, Vol. 2, pp , c. Studies in Edrioasteroidea. (Bather's articles Edrioasteroidea published in the Geological Magazine between 1898 and 1915 were reissued, including plates and retaining original pagination, with an added preface and index, as a book published by the author at "Fabo," Marryat Road, Wimbledon, England.) EHLERS, G. M., and KESLING, R. V Cyclic Pattern of Ambulacral Covering Plates in Discocystis laudoni and Its Taxonomic Implication. Contrib. Mus. Paleontol. Univ. Mich., Vol. 14, No. 15, pp , 3 pls. FOERSTE, A. F Notes on Agelacrinidae and Lepadocystinae, with Descriptions of Thresherodiscus and Brockocystis. Bull. Sci. Lab. Denison Univ., Vol. 17, pp ,6 pls., 8 figs. KESLING, R. V Hydropores in Edrioasteroids. Contrib. Mus. Paleontol. Univ. Mich., Vol. 15, No. 8, pp , 13 pls., 14 figs. MEEE, F. B Descriptions of Invertebrate Fossils of the Silurian and Devonian Systems. Rept. Geol. Surv. Ohio, Vol. 1, Geology and Palaeontology, Pt. 2, Palaeontology, Sec. 1, pp , Pls M~LER, S. A., and FABER, CHAS Some New Species and New Structural Parts of Fossils. Journ. Cincinnati Soc. Nat. Hist., Vol. 15, No. 2, pp , 1 pl. SPENCER, W. K On the Structure and Affinities of Palaeodiscus and Agelacrinus. Proc. Roy. Soc. London, Vol. 74, pp , 1 pl. 12 figs. WILLIAMS, S. R Concerning the Structure of Agelacrinites and Streptaster, Edrioasteroidea of the Richmond and Maysville Divisions of the Ordovician. Ohio Journ. Sci., Vol. 19, No. 1, pp , Pls Submitted for publication July 25,1960
27 PLATES
28 KESLING AND MINTZ EXPLANATION OF PLATE I PAGE Isorophus cincinnatiensis (Roemer) FIGS Aboral views of UCM No ; from the Upper Ordovician Maysville group, McMillan formation, Corryville member; Stonelick Creek, 100 yards below bridge, 1 mile west of Newtonville, Ohio; collected by John K. Pope in Figure 1 is a stereogram of the specimen lightly coated with sublimated ammonium chloride (X 4). Figure 2 is a view of specimen submersed in xylol X 7%). Ambulacrum V, at the lower left in each figure, shows the offset of the proximal two flooring plates and what is interpreted as an opening at their distal edge. Near the left anterior margin (as seen aborally) of the anal pyramid, the posterior interambulacrum appears to consist of two layers of plates; possibly, the genital duct passed between them.

29 PLATE I
30
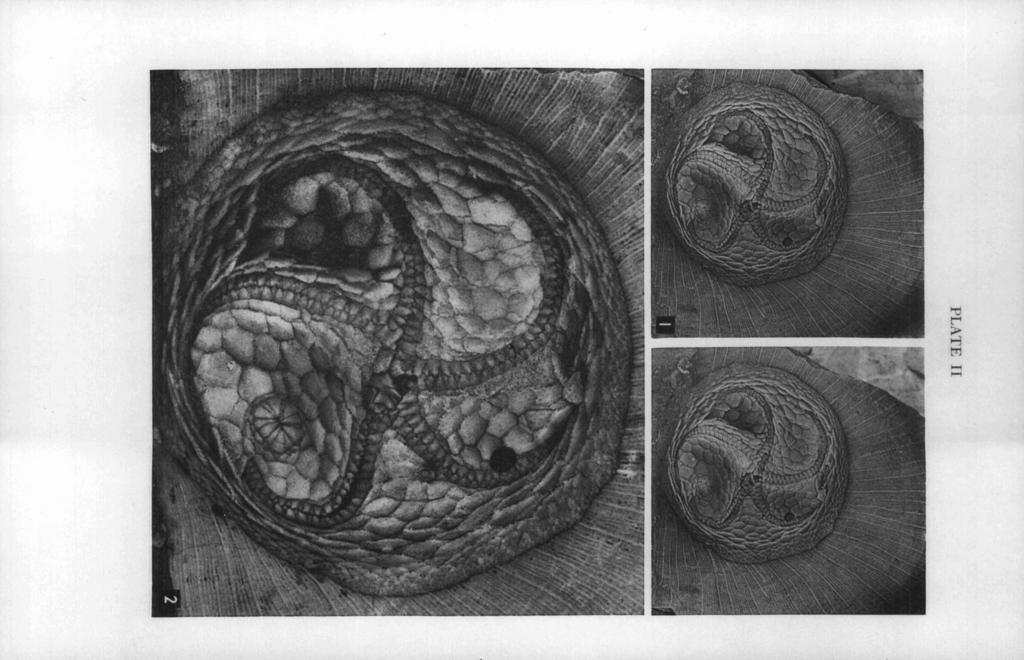
31 EDRIOASTEROID INTERNAL STRUCTURES 34 1 EXPLANATION OF PLATE I1 Isorofihus cincinnatiensis (Roemer) PAGE FIGS Oral views of specimen which was subsequently ground down in the study of its internal structures; from the Upper Ordovician Maysville group, McMillan formation, Bellevue member; Hamilton Avenue, opposite Frisch's Big Boy Restaurant, Cincinnati, Ohio; collected by Mr. William Deak in Figure 1 is a stereogram of specimen lightly coated with sublimated ammonium chloride (X 2). Figure 2 is a view of specimen submersed in xylol (X 5). Polished surfaces of this specimen are shown in Plates 111 and IV.
32 KESLZNG AND MZNTZ EXPLANATION OF PLATE I11 PAGE Isorophus cincinnatiensis (Roemer) FIGS Oral views of successive polished surfaces of specimen shown in Plate 11, each submersed in xylol (X 2%). This series shows the passageway for the stone canal, the relationships of some of the ambulacral covering plates to the underlying flooring plates, the frame around the peristome (formed by the proximal flooring plates of the ambulacra), and what seems to be a duct through the third flooring plate of ambulacrum V. Additional polished surfaces of this specimen are shown in Plate IV. FIGS. 4, 9. Enlargements of parts of Figures 3 and 8 (X 7%). Figure 4 shows the passageway for the stone canal immediately below the hydropore. Figure 9 shows part of the frame around the peristome, the proximal flooring plates of ambulacrum IV, and what seems to be a duct through the third flooring plate of ambulacrum V.

33
34
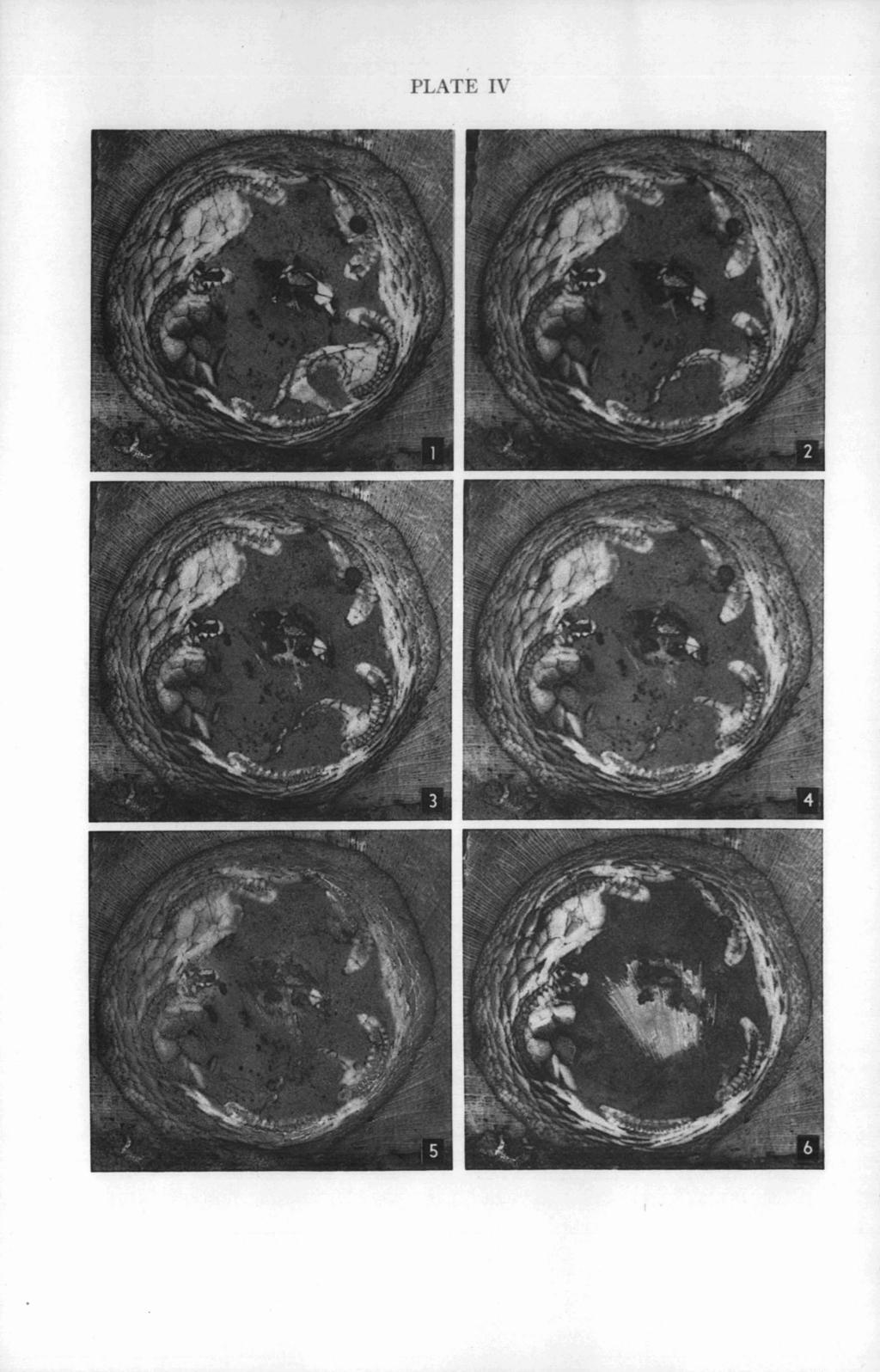
35 EDRIOASTEROID INTERNAL STRUCTURES EXPLANATION OF PLATE IV (All figures x 2%) PAGE Isorophus cincinnatiensis (Roemer) FIGS Oral views of successive polished surfaces of specimen shown in Plate 11, each submersed in xylol. These surfaces follow those shown in Plate 111. This series shows the aboral part of the posterior interambulacrum around the anal pyramid and oblique surfaces through the ambulacra.
36 KESLING AND MINTZ EXPLANATION OF PLATE V Carneyella pilea (Hall) PAGE FIGS Aboral views of UCWM No. 8825; from the Upper Ordovician Maysville group, McMillan formation, Corryville shale member; Cincinnati, Ohio. This specimen was figured by Miller and Faber (1892, P1. I, Fig. 10) and later by Foerste (1914, PI. 11, Fig. 4). It shows the lateral extensions of the ambulacral covering plates particularly well. Figure 1 is a stereogram of the specimen lightly coated with sublimated ammonium chloride (X 3%). Figure 2 is a view of specimen submersed in xylol (X 7).

37 PLATE V

38 PLATE VI
39 EDRIOASTEROID INTERNAL STRUCTURES 345 EXPLANATION OF PLATE VI PAGE Carneyella pilea (Hall) FIGS Oral views of specimen which was subsequently ground down in the study of its internal structures; from the Upper Ordovician Maysville group, McMillan formation, Bellevue member; Hamilton Avenue, opposite Frisch's Big Boy Restaurant, Cincinnati, Ohio; collected by Mr. William Deak in Figure 1 is a stereogram of specimen lightly coated with sublimated ammonium chloride (X 4). Figure 2 is a view of specimen submersed in xylol (X 7%). Polished surfaces of this specimen are shown in Plates VII and VIII.
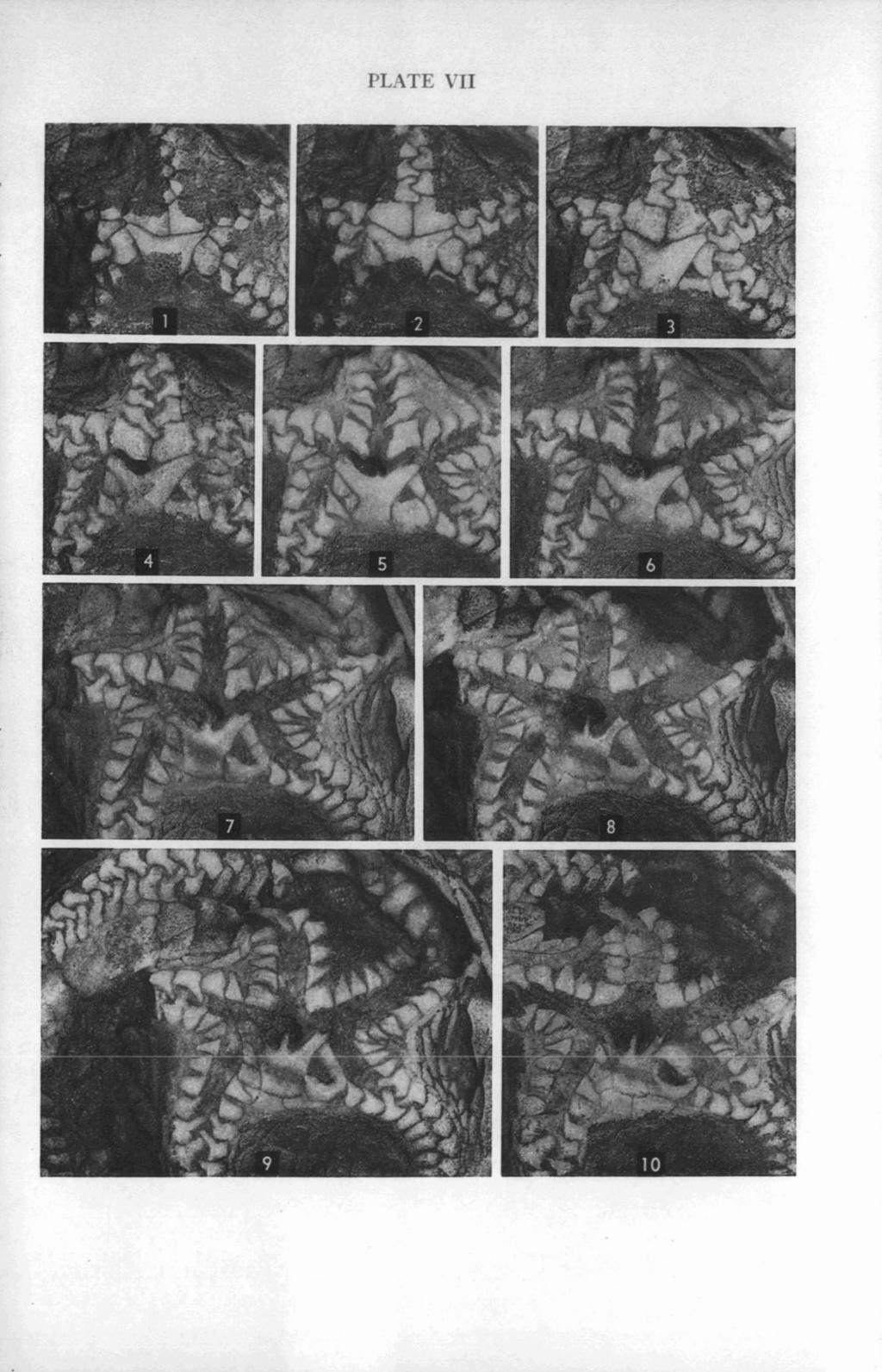
40 KESLING AND MINTZ EXPLANATION OF PLATE VII (All figures X 7%) PAGE Carneyella pilea (Hall) FIGS Oral views of successive polished surfaces of specimen shown in Plate VI, each submersed in xylol. This series shows the passageway for the stone canal leading down from the hydropore, the peristomial covering plates (particularly the W-shaped anterior part of the large posterior plate), and the junctions and lateral extensions of the ambulacral covering plates. Additional polished surfaces of this specimen are shown in Plate VIII.
41
42
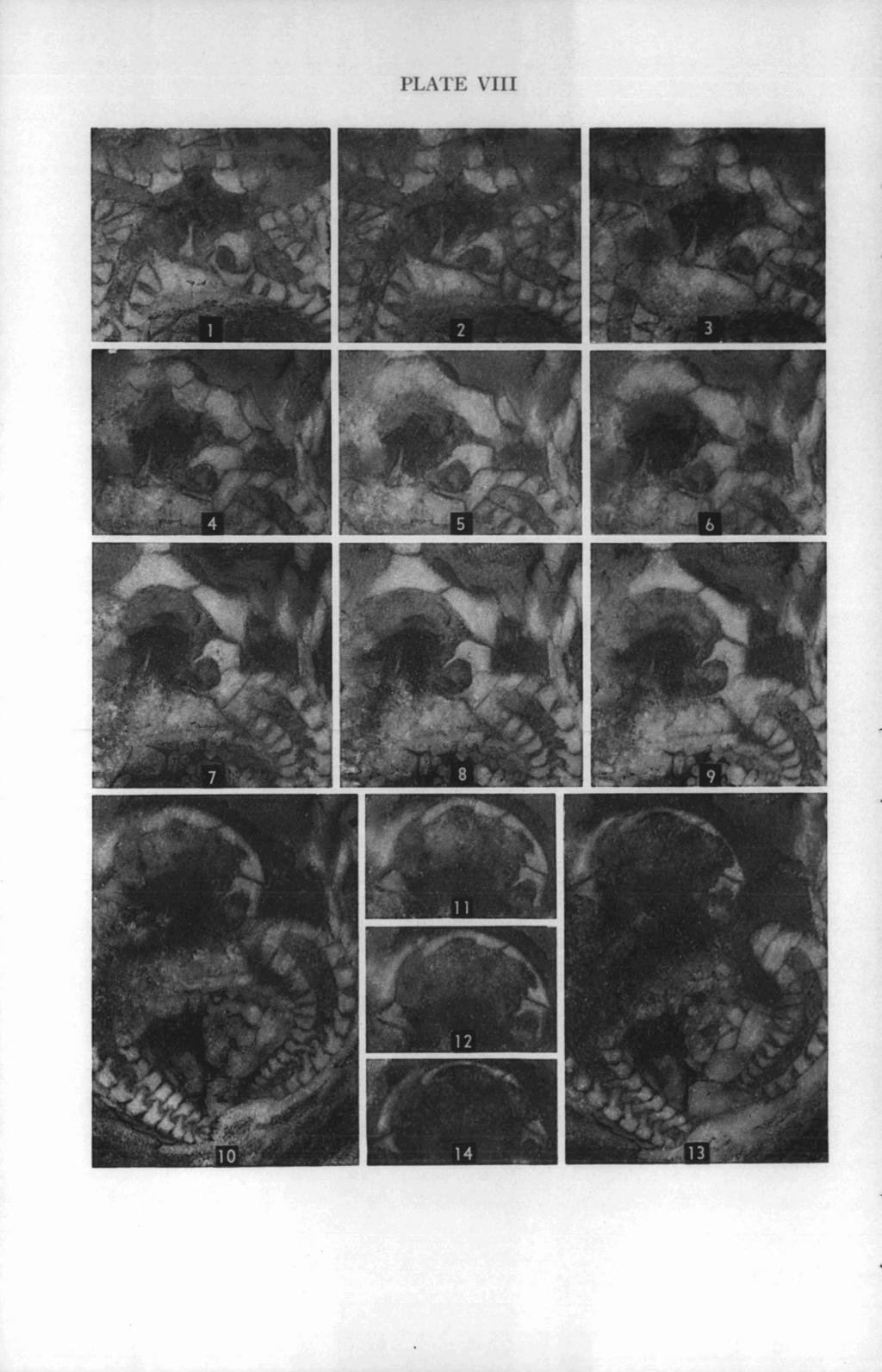
43 EDRIOASTEROID INTERNAL STRUCTURES 347 EXPLANATION OF PLATE VIII (All figures x 7%) PACE Carneyella pilea (Hall) FIGS Oral views of successive polished surfaces of specimen shown in Plate VI, each submersed in xylol. These surfaces follow those shown in Plate VII. This series shows the aboral extension of the passageway for the stone canal, the frame around the peristome (formed by the proximal flooring plates of the ambulacra), and the relationships of some of the ambulacral covering plates to the underlying flooring plates.
44 KESLZNG AND MZNTZ EXPLANATION OF PLATE IX PAGE Carneyella pilea (Hall) FIGS Aboral views of AMNH No x; from the Upper Ordovician Maysville group, McMillan formation, Corryville shale member; Cincinnati, Ohio. This specimen was described and figured by Foerste (1914, pp. 418, 428, PI. 11, Fig. 3). Figure 1 is a stereogram of the specimen rather heavily coated with sublimated ammonium chloride (X 5). Figure 2 is a view of specimen very lightly coated with sublimated ammonium chloride (approximately x 8).

45 PLATE IX
THE UNIVERSITY OF MICHIGAN
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY f THE UNIVERSITY OF MICHIGAN VOL. XV, NO. 8, pp. 139-192 (13 pls., 14 figs.) JANUARY 22, 1960 HYDROPORES IN EDRIOASTEROIDS BY ROBERT V. KESLING MUSEUM OF PALEONTOLOGY
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY f THE UNIVERSITY OF MICHIGAN VOL. XV, NO. 8, pp. 139-192 (13 pls., 14 figs.) JANUARY 22, 1960 HYDROPORES IN EDRIOASTEROIDS BY ROBERT V. KESLING MUSEUM OF PALEONTOLOGY
UNVERSITY OF MICHIGAN THREE NEW SPECIES OF THE CYSTID GENUS LIPSANOCYSTIS FROM THE MIDDLE DEVONIAN TRAVERSE GROUP OF MICHIGAN
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNVERSITY OF MICHIGAN VOL. XII, No. 6, pp. 97-103 (2 pls.) APRIL 8, 1955 THREE NEW SPECIES OF THE CYSTID GENUS LIPSANOCYSTIS FROM THE MIDDLE DEVONIAN TRAVERSE
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNVERSITY OF MICHIGAN VOL. XII, No. 6, pp. 97-103 (2 pls.) APRIL 8, 1955 THREE NEW SPECIES OF THE CYSTID GENUS LIPSANOCYSTIS FROM THE MIDDLE DEVONIAN TRAVERSE
CONCERNING THE STRUCTURE OF AGELACRINITES AND STREPTASTER, EDRIOASTEROIDEA OF THE RICHMOND AND MAYSVILLE DIVISIONS OF THE ORDOVICIAN.
 CONCERNING THE STRUCTURE OF AGELACRINITES AND STREPTASTER, EDRIOASTEROIDEA OF THE RICHMOND AND MAYSVILLE DIVISIONS OF THE ORDOVICIAN. STEPHEN R. WILLIAMS. CONTENTS. Introduction and Sources of Material
CONCERNING THE STRUCTURE OF AGELACRINITES AND STREPTASTER, EDRIOASTEROIDEA OF THE RICHMOND AND MAYSVILLE DIVISIONS OF THE ORDOVICIAN. STEPHEN R. WILLIAMS. CONTENTS. Introduction and Sources of Material
THE UNIVERSITY OF MICHIGAN PROCTOTHYLACOCRIN US ESSERI, A NEW CRINOID FROM THE MIDDLE DEVONIAN SILICA FORMATION OF NORTHWESTERN OHIO
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XX, NO. 4, pp. 75-87 (5 pls., 1 fig.) OCTOBER 15, 1965 PROCTOTHYLACOCRIN US ESSERI, A NEW CRINOID FROM THE MIDDLE DEVONIAN
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XX, NO. 4, pp. 75-87 (5 pls., 1 fig.) OCTOBER 15, 1965 PROCTOTHYLACOCRIN US ESSERI, A NEW CRINOID FROM THE MIDDLE DEVONIAN
NEW AMERICAN PALEOZOIC OSTRACODA.
 New American Paleozoic Ostracoda. 179 NEW AMERICAN PALEOZOIC OSTRACODA. BY E. O. ULRICH. No. 1. CTKNOBOLBINA AND KIRKBYA. Since the publication of my paper on " New and Ivittle Known American Paleozoic
New American Paleozoic Ostracoda. 179 NEW AMERICAN PALEOZOIC OSTRACODA. BY E. O. ULRICH. No. 1. CTKNOBOLBINA AND KIRKBYA. Since the publication of my paper on " New and Ivittle Known American Paleozoic
OF PLANALVEOLITES POUGHTI
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XXI, No. 2, pp. 67-72 (1 pl.) MAP 10, 1967 PLANALVEOLITELLA, A NEW GENUS OF DEVONIAN TABULATE CORALS, WITH A REDESCRIPTION
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XXI, No. 2, pp. 67-72 (1 pl.) MAP 10, 1967 PLANALVEOLITELLA, A NEW GENUS OF DEVONIAN TABULATE CORALS, WITH A REDESCRIPTION
THE UNIVERSITY OF MICHIGAN UNUSUAL PRESERVATION FROM THE MIDDLE DEVONIAN SILICA FORMATION OF OHIO
 COXTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 24, NO. 2, p. 9-15 (2 pls., 3 text-figs.) APRE 11, 1972 STRATASTER DEVONZCUS, A NEW BRITTLE-STAR WITH UNUSUAL PRESERVATION
COXTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 24, NO. 2, p. 9-15 (2 pls., 3 text-figs.) APRE 11, 1972 STRATASTER DEVONZCUS, A NEW BRITTLE-STAR WITH UNUSUAL PRESERVATION
UNIVERSITY OF MICHIGAN. A NEW AND UNUSUAL SPECIES OF THE OSTRACOD GENUS HERRMAlVNIlVA FROM THE MIDDLE SILURIAN HENDRICKS DOLOMITE OF MICHIGAN
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL. SIV, Xo. 9, pp. 143-148 (1 pl.) FEBRUARY 28, 1958 A NEW AND UNUSUAL SPECIES OF THE OSTRACOD GENUS HERRMAlVNIlVA FROM THE MIDDLE
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL. SIV, Xo. 9, pp. 143-148 (1 pl.) FEBRUARY 28, 1958 A NEW AND UNUSUAL SPECIES OF THE OSTRACOD GENUS HERRMAlVNIlVA FROM THE MIDDLE
ACINETASTER KONIECKII, A NEW BRITTLE-STAR FROM THE MIDDLE DEVONIAN ARKONA SHALE
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 26, NO. 5, p. 71-81 (2 plates) December 2 1, 1982 ACINETASTER KONIECKII, A NEW BRITTLE-STAR FROM THE MIDDLE DEVONIAN ARKONA
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 26, NO. 5, p. 71-81 (2 plates) December 2 1, 1982 ACINETASTER KONIECKII, A NEW BRITTLE-STAR FROM THE MIDDLE DEVONIAN ARKONA
DOLATOCRINUS AND STEREOCRINUS, ITS JUNIOR SYNONYM
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN T - VOL. XVIII, No. 14, pp. 229-237 (2 pls.) OCTOBER 22, 1963 DOLATOCRINUS AND STEREOCRINUS, ITS JUNIOR SYNONYM BY ROBERT V. KESLING
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN T - VOL. XVIII, No. 14, pp. 229-237 (2 pls.) OCTOBER 22, 1963 DOLATOCRINUS AND STEREOCRINUS, ITS JUNIOR SYNONYM BY ROBERT V. KESLING
Lecture 19. A Sieve Plate with large Sieve Pores. Secondary Phloem. Secondary phloem (cont d)
") Lecture 19 Secondary phloem (cont d) Secondary Phloem in Tilia americana (American Basswood) Secondary Phloem of Tilia Stained with Toluidine Blue & viewed with Crossed Polarizers. Secondary Phloem A Sieve
Lecture 19 Secondary phloem (cont d) Secondary Phloem in Tilia americana (American Basswood) Secondary Phloem of Tilia Stained with Toluidine Blue & viewed with Crossed Polarizers. Secondary Phloem A Sieve
Exercise 10 Fossil Lab Part 5: Crinoids, Blastoids, Fusulinids, Plants
 Exercise 10 Fossil Lab Part 5: Crinoids, Blastoids, Fusulinids, Plants ECHINODERMS (CRINOIDS AND BLASTOIDS): Echinoderms are an extremely diverse group of advanced invertebrates including such familiar
Exercise 10 Fossil Lab Part 5: Crinoids, Blastoids, Fusulinids, Plants ECHINODERMS (CRINOIDS AND BLASTOIDS): Echinoderms are an extremely diverse group of advanced invertebrates including such familiar
GEOLOGICAL SURVEY OF OHIO.
 GEOLOGICAL SURVEY OF OHIO. VOL. 1. PART II. PALÆONTOLOGY. SECTION I. DESCRIPTIONS OF INVERTEBRATE FOSSILS OF THE SILURIAN AND DEVONIAN SYSTEMS. BY F. B. MEEK. Digital version copyrighted 2012 by Don Chesnut.
GEOLOGICAL SURVEY OF OHIO. VOL. 1. PART II. PALÆONTOLOGY. SECTION I. DESCRIPTIONS OF INVERTEBRATE FOSSILS OF THE SILURIAN AND DEVONIAN SYSTEMS. BY F. B. MEEK. Digital version copyrighted 2012 by Don Chesnut.
STRUCTURAL FEATURES OF COCKROACH EGG CAPSULES. II. THE OOTHECA OF CARIBLATTA LUTEA LUTEA (ORTHOPTERA: BLATTIDAE) 1
 1") STRUCTURAL FEATURES OF COCKROACH EGG CAPSULES. II. THE OOTHECA OF CARIBLATTA LUTEA LUTEA (ORTHOPTERA: BLATTIDAE) 1 FRED A. LAWSON Department of Zoology and Entomology, The University of Tennessee, Knoxville
STRUCTURAL FEATURES OF COCKROACH EGG CAPSULES. II. THE OOTHECA OF CARIBLATTA LUTEA LUTEA (ORTHOPTERA: BLATTIDAE) 1 FRED A. LAWSON Department of Zoology and Entomology, The University of Tennessee, Knoxville
JOEL CRACRAFT. N 1913 Shufeldt described a new fossil bird, Palaeophasianus meleagroides,
 REALLOCATION PALAEOPHASIANUS OF THE EOCENE MELEAGROIDES FOSSIL SHUFELDTl I JOEL CRACRAFT N 1913 Shufeldt described a new fossil bird, Palaeophasianus meleagroides, from the early Eocene of Wyoming. Shufeldt
REALLOCATION PALAEOPHASIANUS OF THE EOCENE MELEAGROIDES FOSSIL SHUFELDTl I JOEL CRACRAFT N 1913 Shufeldt described a new fossil bird, Palaeophasianus meleagroides, from the early Eocene of Wyoming. Shufeldt
Be able to understand the processes which occurred during the last ice age.
 Glaciation Learning Intentions Be able to understand the processes which occurred during the last ice age. Be able to describe and explain the formation of features formed during glacial periods. 1 Water
Glaciation Learning Intentions Be able to understand the processes which occurred during the last ice age. Be able to describe and explain the formation of features formed during glacial periods. 1 Water
Acrobotrys tritubus Riedel
 151 Acrobotrys tritubus Riedel Acrobotrys tritubus Riedel, 1957, p.80, pl.1, fig.5 DESCRIPTION Cephalis trilobate, with large subglobular [antecephalic] lobe, smaller globular [cephalic] lobe, and inflated-conical
151 Acrobotrys tritubus Riedel Acrobotrys tritubus Riedel, 1957, p.80, pl.1, fig.5 DESCRIPTION Cephalis trilobate, with large subglobular [antecephalic] lobe, smaller globular [cephalic] lobe, and inflated-conical
AN OCCURRENCE OF PERFORATED TRACHEIDS IN THUJA OCCIDENTALIS L.
 AN OCCURRENCE OF PERFORATED TRACHEIDS IN THUJA OCCIDENTALIS L. BY M. W. B ANN AN Department of Botany, University of Toronto {Received 28 February 1957) (With Plate and i figure in the text) In a recent
AN OCCURRENCE OF PERFORATED TRACHEIDS IN THUJA OCCIDENTALIS L. BY M. W. B ANN AN Department of Botany, University of Toronto {Received 28 February 1957) (With Plate and i figure in the text) In a recent
OF THE LEMNA FROND MORPHOLOGY
 MORPHOLOGY OF THE LEMNA FROND FREDERICK H. BLODGETT (WITH PLATE XIV AND ONE FIGURE) In the case of structure simplified by reduction, it is sometimes necessary to trace the development of the parts through
MORPHOLOGY OF THE LEMNA FROND FREDERICK H. BLODGETT (WITH PLATE XIV AND ONE FIGURE) In the case of structure simplified by reduction, it is sometimes necessary to trace the development of the parts through
THE UNIVERSITY OF MICHIGAN PROCTOTHYLACOCRZNUS BERRYORUM, A NEW CRINOID FROM THE MIDDLE DEVONIAN ARKONA SHALE OF ONTARIO
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 23, NO. 21, p. 343-347 (2 pls.) SEPTEMBER 17, 1971 PROCTOTHYLACOCRZNUS BERRYORUM, A NEW CRINOID FROM THE MIDDLE DEVONIAN ARKONA
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 23, NO. 21, p. 343-347 (2 pls.) SEPTEMBER 17, 1971 PROCTOTHYLACOCRZNUS BERRYORUM, A NEW CRINOID FROM THE MIDDLE DEVONIAN ARKONA
Section 7. Reading the Geologic History of Your Community. What Do You See? Think About It. Investigate. Learning Outcomes
 Chapter 3 Minerals, Rocks, and Structures Section 7 Reading the Geologic History of Your Community What Do You See? Learning Outcomes In this section, you will Goals Text Learning Outcomes In this section,
Chapter 3 Minerals, Rocks, and Structures Section 7 Reading the Geologic History of Your Community What Do You See? Learning Outcomes In this section, you will Goals Text Learning Outcomes In this section,
THE OHIO JOURNAL OF SCIENCE
 THE OHIO JOURNAL OF SCIENCE VOL. XL MARCH, 1940 No. 2 CRINOIDS FROM THE SILICA SHALE, DEVONIAN, OF OHIO GRACE A. STEWART, Department of Geology, Ohio State University Recently the writer has had the privilege
THE OHIO JOURNAL OF SCIENCE VOL. XL MARCH, 1940 No. 2 CRINOIDS FROM THE SILICA SHALE, DEVONIAN, OF OHIO GRACE A. STEWART, Department of Geology, Ohio State University Recently the writer has had the privilege
What is weathering and how does it change Earth s surface? Answer the question using
 7 th Grade Lesson What is weathering and how does it change Earth s surface? Answer the question using the sentence frame. You have 4 minutes. Weathering is. This changes the Earth s surface because. 1
7 th Grade Lesson What is weathering and how does it change Earth s surface? Answer the question using the sentence frame. You have 4 minutes. Weathering is. This changes the Earth s surface because. 1
ON THE OCCURRENCE KINNEGRAPTUS SKOGLUND IN NORWAY
 NORSK GEOLOGISK TIDSSKRIFT 42 ON THE OCCURRENCE OF KINNEGRAPTUS SKOGLUND IN NORWAY By O. M. B. BuLMAN and CYNTHIA M. Cowm (Sedgwick Museum, Cambridge) Abs t r a c t. Kinnegraptus kinnekullensis SKoGLUND,
NORSK GEOLOGISK TIDSSKRIFT 42 ON THE OCCURRENCE OF KINNEGRAPTUS SKOGLUND IN NORWAY By O. M. B. BuLMAN and CYNTHIA M. Cowm (Sedgwick Museum, Cambridge) Abs t r a c t. Kinnegraptus kinnekullensis SKoGLUND,
Galileoscope Assembly Instructions
 Galileoscope Assembly Instructions To begin, open the exterior box flaps to reveal the interior box. Under one flap, but not the other, you ll find two holes, as shown at right. Place a finger and thumb
Galileoscope Assembly Instructions To begin, open the exterior box flaps to reveal the interior box. Under one flap, but not the other, you ll find two holes, as shown at right. Place a finger and thumb
Monday 3 June 2013 Morning
 Monday 3 June 2013 Morning A2 GCE GEOLOGY F795/01 Evolution of Life, Earth and Climate *F713200613* Candidates answer on the Question Paper. OCR supplied materials: None Other materials required: Electronic
Monday 3 June 2013 Morning A2 GCE GEOLOGY F795/01 Evolution of Life, Earth and Climate *F713200613* Candidates answer on the Question Paper. OCR supplied materials: None Other materials required: Electronic
AND A NEW SPECIES OF THIS BRACHIOPOD
 L i CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XVIII, NO. 12, pp. 197-204 (1 pl.) OCTOBER 4, 1963 CYRTINA HAMILTONENSIS (HALL) AND A NEW SPECIES OF THIS BRACHIOPOD GENUS
L i CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XVIII, NO. 12, pp. 197-204 (1 pl.) OCTOBER 4, 1963 CYRTINA HAMILTONENSIS (HALL) AND A NEW SPECIES OF THIS BRACHIOPOD GENUS
By Sudha Vashisht P.G.G.C.G. -11 CHD
 By Sudha Vashisht P.G.G.C.G. -11 CHD Pila is the common Apple snail which is found in ponds ditches, rivers and in watery fileds. The soft body is covered by a coiled shell. The first chamber of the shell
By Sudha Vashisht P.G.G.C.G. -11 CHD Pila is the common Apple snail which is found in ponds ditches, rivers and in watery fileds. The soft body is covered by a coiled shell. The first chamber of the shell
Of the University of Colorado, Boulder.
 SOME EOCENE INSECTS OF THE FAMILY FULGORIDA~ By T. D. A. COCKERELL and GRACE SANDTIOUSE, Of the University of Colorado, Boulder. The remarkable abundance and variety of the Homopterous family Fulgoridae
SOME EOCENE INSECTS OF THE FAMILY FULGORIDA~ By T. D. A. COCKERELL and GRACE SANDTIOUSE, Of the University of Colorado, Boulder. The remarkable abundance and variety of the Homopterous family Fulgoridae
Pratice Surface Processes Test
 1. The cross section below shows the movement of wind-driven sand particles that strike a partly exposed basalt cobble located at the surface of a windy desert. Which cross section best represents the
1. The cross section below shows the movement of wind-driven sand particles that strike a partly exposed basalt cobble located at the surface of a windy desert. Which cross section best represents the
The Upper Course of a River
 The Upper Course of a River I. Introduction A river is split along the long profile into three different sections; the upper, middle and lower course. As GCSE students you are expected to know the characteristics
The Upper Course of a River I. Introduction A river is split along the long profile into three different sections; the upper, middle and lower course. As GCSE students you are expected to know the characteristics
THE method of operating upon stem apices and leaf primordia which we have
 THE DETERMINATION OF AXILLARY BUDS BY MARY SNOW AND R. SNOW (With 10 figures in the text) THE method of operating upon stem apices and leaf primordia which we have practised for other purposes (1931, 1935)
THE DETERMINATION OF AXILLARY BUDS BY MARY SNOW AND R. SNOW (With 10 figures in the text) THE method of operating upon stem apices and leaf primordia which we have practised for other purposes (1931, 1935)
The new physiology of vision-chapter The colours of interference. SIR C V RAMAN Received October 14, 1965
 Proc. Indian Acad. Sci. A62 243-248 (1965) The new physiology of vision-chapter The colours of interference XXVII. SIR C V RAMAN Received October 14, 1965 The characteristic features and properties of
Proc. Indian Acad. Sci. A62 243-248 (1965) The new physiology of vision-chapter The colours of interference XXVII. SIR C V RAMAN Received October 14, 1965 The characteristic features and properties of
THE UNIVERSITY OF MICHIGAN ORDOVICIAN STREPTELASMID RUGOSE CORALS FROM MICHIGAN
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XVIII, No. 2, pp. 23-31 (2 pls.) MARCH 20, 1963 ORDOVICIAN STREPTELASMID RUGOSE CORALS FROM MICHIGAN BY ERWIN C. STUMM FROM
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XVIII, No. 2, pp. 23-31 (2 pls.) MARCH 20, 1963 ORDOVICIAN STREPTELASMID RUGOSE CORALS FROM MICHIGAN BY ERWIN C. STUMM FROM
Lithosphere Features Of Glacial Erosion. Corrie;
 Corrie; Lithosphere Features Of Glacial Erosion These are bowl like hollows in the sides of mountains formed by glacial erosion A small hollow on the side of a mountain will start to fill with snow all
Corrie; Lithosphere Features Of Glacial Erosion These are bowl like hollows in the sides of mountains formed by glacial erosion A small hollow on the side of a mountain will start to fill with snow all
Lab 4 Identifying metazoan phyla and plant groups
 Geol G308 Paleontology and Geology of Indiana Name: Lab 4 Identifying metazoan phyla and plant groups The objective of this lab is to classify all of the fossils from your site to phylum (or to plant group)
Geol G308 Paleontology and Geology of Indiana Name: Lab 4 Identifying metazoan phyla and plant groups The objective of this lab is to classify all of the fossils from your site to phylum (or to plant group)
FRANCIS DE CASTELNAU'S ESSAI SUR LE SYSTEME SILURIEN DE L'AMBRIQUE SEPTENTRIONALE AND THE STATUS OF HIS SPIRIFER HURONIENSIS
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL. Xm, No. 7, pp. 175-180 (2 pls.) NOYEMBEB 20, 1957 FRANCIS DE CASTELNAU'S ESSAI SUR LE SYSTEME SILURIEN DE L'AMBRIQUE SEPTENTRIONALE
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL. Xm, No. 7, pp. 175-180 (2 pls.) NOYEMBEB 20, 1957 FRANCIS DE CASTELNAU'S ESSAI SUR LE SYSTEME SILURIEN DE L'AMBRIQUE SEPTENTRIONALE
THE TOPOGRAPHY AND GEOLOGY OF THE GRAND PORTAGE^
 THE TOPOGRAPHY AND GEOLOGY OF THE GRAND PORTAGE^ Few areas within the boundaries of Minnesota are of greater historical interest than the region surrounding the Grand Portage, which long before the first
THE TOPOGRAPHY AND GEOLOGY OF THE GRAND PORTAGE^ Few areas within the boundaries of Minnesota are of greater historical interest than the region surrounding the Grand Portage, which long before the first
GEOMETRIC THEORY OF FRESNEL DIFFRACTION PATTERNS
 GEOMETRIC THEORY OF FRESNEL DIFFRACTION PATTERNS Part V. Elliptic Obstacles and Apertures BY Y. V. KATHAVATE (From the Department of Physics, Indian Institute of Science, Bangalore) Received April 2, 1945
GEOMETRIC THEORY OF FRESNEL DIFFRACTION PATTERNS Part V. Elliptic Obstacles and Apertures BY Y. V. KATHAVATE (From the Department of Physics, Indian Institute of Science, Bangalore) Received April 2, 1945
CALLISPHENUS GRACILIS, N. GEN., N. SP. A FOSSIL ALGA FROM THE WENLOCK OF THE OSLO REGION
 CALLISPHENUS GRACILIS, N. GEN., N. SP. A FOSSIL ALGA FROM THE WENLOCK OF THE OSLO REGION BY OVE HØEG WITH 2 PLATES The specimen described below was found in August, 191 O, on the small island of Kommersøy
CALLISPHENUS GRACILIS, N. GEN., N. SP. A FOSSIL ALGA FROM THE WENLOCK OF THE OSLO REGION BY OVE HØEG WITH 2 PLATES The specimen described below was found in August, 191 O, on the small island of Kommersøy
THE UNIVERSITY OF MICHIGAN
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XIX, NO. 8, pp. 105-114 (1 pl., 1 fig.) NOVEMBER 25, 1964 A NEW SPIRACULATE BLASTOID, PYRAMIBLASTUS, FROM THE MISSISSIPPIAN
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XIX, NO. 8, pp. 105-114 (1 pl., 1 fig.) NOVEMBER 25, 1964 A NEW SPIRACULATE BLASTOID, PYRAMIBLASTUS, FROM THE MISSISSIPPIAN
Characteristics of Echinoderms
 Characteristics of Echinoderms Adult echinoderms have a body plan with five parts organized symmetrically around a center Does not have an anterior nor posterior end or a brain Most echinoderms are two
Characteristics of Echinoderms Adult echinoderms have a body plan with five parts organized symmetrically around a center Does not have an anterior nor posterior end or a brain Most echinoderms are two
A NEW OCCURRENCE OF THE DEVONIAN ARTHRODIRE HOLONEMA'
 A NEW OCCURRENCE OF THE DEVONIAN ARTHRODIRE HOLONEMA' STEVEN W. MITCHELL Department of Geology, Wayne State University, Detroit, Michigan 4-8202 ABSTRACT A single, incomplete, paranuchal plate of the cranial
A NEW OCCURRENCE OF THE DEVONIAN ARTHRODIRE HOLONEMA' STEVEN W. MITCHELL Department of Geology, Wayne State University, Detroit, Michigan 4-8202 ABSTRACT A single, incomplete, paranuchal plate of the cranial
\ SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 153, NUMBER 2
 (91' \ SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 153, NUMBER 2 Smithsonian Publication 4738 Cfjatless B. anb iularp "^aux Malcott ^Res^earcf) jfunb Echinoids from the Middle Eocene Lake City Formation
(91' \ SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 153, NUMBER 2 Smithsonian Publication 4738 Cfjatless B. anb iularp "^aux Malcott ^Res^earcf) jfunb Echinoids from the Middle Eocene Lake City Formation
Glaciers Earth 9th Edition Chapter 18 Glaciers: summary in haiku form Key Concepts Glaciers Glaciers Glaciers Glaciers
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Earth 9 th Edition Chapter 18 : summary in haiku form Ten thousand years thence big glaciers began to melt - called "global warming." Key Concepts and types of glaciers.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 Earth 9 th Edition Chapter 18 : summary in haiku form Ten thousand years thence big glaciers began to melt - called "global warming." Key Concepts and types of glaciers.
Topic 2: Plant Structure & Growth Ch. 35 Angiosperms are the most complex plants. They are composed of cells, tissues, organs and organ systems.
 Topic 2: Plant Structure & Growth Ch. 35 Angiosperms are the most complex plants. They are composed of cells, tissues, organs and organ systems. Fig. 35.8 Plant Cells pp.798-802 Types of plant cells Include:
Topic 2: Plant Structure & Growth Ch. 35 Angiosperms are the most complex plants. They are composed of cells, tissues, organs and organ systems. Fig. 35.8 Plant Cells pp.798-802 Types of plant cells Include:
THE UNIVERSITY OF MICHIGAN AN INTERPRETATION OF RHOMBIFERA BOHEMICA BARRANDE, 1867, AN UNUSUAL HYDROPHORIDEAN CYSTOID
 i i CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XVII, No. 13, pp. 277-289 (2 pls., 2 figs.) OCTOBER 11, 1962 AN INTERPRETATION OF RHOMBIFERA BOHEMICA BARRANDE, 1867, AN
i i CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XVII, No. 13, pp. 277-289 (2 pls., 2 figs.) OCTOBER 11, 1962 AN INTERPRETATION OF RHOMBIFERA BOHEMICA BARRANDE, 1867, AN
LIBRARY OF THE UNIVERSITY OF ILLINOIS AT URBANA-CHAMPAIGN v. 16. t 5 LIBRARY GEOLOGY
 . LIBRARY OF THE UNIVERSITY OF ILLINOIS AT URBANA-CHAMPAIGN 550.5 FI v. 16 t 5 LIBRARY AT URBANA-CHAMPAIGN GEOLOGY St wavwisd on or before the Latest Date stamped belou. mutilation, and underlining of
. LIBRARY OF THE UNIVERSITY OF ILLINOIS AT URBANA-CHAMPAIGN 550.5 FI v. 16 t 5 LIBRARY AT URBANA-CHAMPAIGN GEOLOGY St wavwisd on or before the Latest Date stamped belou. mutilation, and underlining of
UNIVERSITY OF MICHIGAN
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL. XIII, No. 6, pp. 161-173, (4 plates) DECEMBER 21, 1956 A NEW CALAMITE FROM COLORADO BY CHESTER A. ARNOLD From the Ermine Cowles
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL. XIII, No. 6, pp. 161-173, (4 plates) DECEMBER 21, 1956 A NEW CALAMITE FROM COLORADO BY CHESTER A. ARNOLD From the Ermine Cowles
ZOOPHYTOLOGY. On some MADEIRAN POLYZOA. Collected by J. YATES JOHNSON, Esq. (Continued from No. XXIV, p. 263.)
") On some MADEIRAN POLYZOA. Collected by J. YATES JOHNSON, Esq. (Continued from No. XXIV, p. 263.) WE continue the account of zoophytes, brought by Mr. J. Y. Johnson from Madeira, and to which he has made
On some MADEIRAN POLYZOA. Collected by J. YATES JOHNSON, Esq. (Continued from No. XXIV, p. 263.) WE continue the account of zoophytes, brought by Mr. J. Y. Johnson from Madeira, and to which he has made
Glaciers. (Shaping Earth s Surface, Part 6) Science 330 Summer 2005
 Science 330 Summer 2005") Glaciers (Shaping Earth s Surface, Part 6) Science 330 Summer 2005 1 Glaciers Glaciers are parts of two basic cycles Hydrologic cycle Rock cycle Glacier a thick mass of ice that originates on land from
Glaciers (Shaping Earth s Surface, Part 6) Science 330 Summer 2005 1 Glaciers Glaciers are parts of two basic cycles Hydrologic cycle Rock cycle Glacier a thick mass of ice that originates on land from
Objectives: Define Relative Age, Absolute Age
 S6E5. Students will investigate the scientific view of how the earth s surface is formed. c. Classify rocks by their process of formation. g. Describe how fossils show evidence of the changing surface
S6E5. Students will investigate the scientific view of how the earth s surface is formed. c. Classify rocks by their process of formation. g. Describe how fossils show evidence of the changing surface
A PORCUPINE FROM THE PLEISTOCENE OF
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XVIII, No. 16, pp. 245-250 (1 pl., 2 figs.) NOVEMBER 22, 1963 A PORCUPINE FROM THE PLEISTOCENE OF AGUASCALIENTES, MEXICO BY
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. XVIII, No. 16, pp. 245-250 (1 pl., 2 figs.) NOVEMBER 22, 1963 A PORCUPINE FROM THE PLEISTOCENE OF AGUASCALIENTES, MEXICO BY
Tribology Prof. Dr. Harish Hirani Department of Mechanical Engineering Indian Institute of Technology, Delhi
 Tribology Prof. Dr. Harish Hirani Department of Mechanical Engineering Indian Institute of Technology, Delhi Lecture No. # 29 Rolling Element Bearings (Contd.) Welcome to 29 th lecture of video course
Tribology Prof. Dr. Harish Hirani Department of Mechanical Engineering Indian Institute of Technology, Delhi Lecture No. # 29 Rolling Element Bearings (Contd.) Welcome to 29 th lecture of video course
A MEGASECOPTERON FROM UPPER CARBONIFEROUS BY F. M. CARPENTER. In I962 Professor F. Stockmans, of the Institut Royal des Sciences STRATA IN SPAIN
 A MEGASECOPTERON FROM UPPER CARBONIFEROUS STRATA IN SPAIN BY F. M. CARPENTER Harvard University In I962 Professor F. Stockmans, of the Institut Royal des Sciences Naturelles de. Belgique, kindly sent me
A MEGASECOPTERON FROM UPPER CARBONIFEROUS STRATA IN SPAIN BY F. M. CARPENTER Harvard University In I962 Professor F. Stockmans, of the Institut Royal des Sciences Naturelles de. Belgique, kindly sent me
Galileo Telescope Solar Viewer Joseph Hora, Elizabeth Hora 2017/09/18
 Galileo Telescope Solar Viewer Joseph Hora, Elizabeth Hora 2017/09/18 17 7.75 5 2 1.5 3 2 1.5 Materials: (all dimensions in inches) 3x plywood sheet 17 x 7.75 x ½ 3x wood block cut from 2x4: 5 x 2 x 1.5
Galileo Telescope Solar Viewer Joseph Hora, Elizabeth Hora 2017/09/18 17 7.75 5 2 1.5 3 2 1.5 Materials: (all dimensions in inches) 3x plywood sheet 17 x 7.75 x ½ 3x wood block cut from 2x4: 5 x 2 x 1.5
NEW STANDARD SEXTANT OF THE U. S. NAVY
 NEW STANDARD SEXTANT OF THE U. S. NAVY Director of the United States of America Hydrographic Office has sent to the International Hydrographic Bureau the following data relating to the new Standard Sextant
NEW STANDARD SEXTANT OF THE U. S. NAVY Director of the United States of America Hydrographic Office has sent to the International Hydrographic Bureau the following data relating to the new Standard Sextant
Prismatic cleavage and steep rhombohedral form in s-quartz.
 259 Prismatic cleavage and steep rhombohedral form in s-quartz. By JULIEN DRUGM*~, M.Sc., PhD. [Read November 3, 1938.] (l) AN EXAMPLE OF ~-QUARTZ SHOWING GOOD :PRISMATIC CLEAVAGES. S is well known, quartz
259 Prismatic cleavage and steep rhombohedral form in s-quartz. By JULIEN DRUGM*~, M.Sc., PhD. [Read November 3, 1938.] (l) AN EXAMPLE OF ~-QUARTZ SHOWING GOOD :PRISMATIC CLEAVAGES. S is well known, quartz
1 Conducting Units: Tracheids and Vessels 1.1 Evolutionary Specialization
 1 Conducting Units: Tracheids and Vessels 1.1 Evolutionary Specialization The development of upright land plants depended on the development of a waterconducting system. Many of the earliest land plants,
1 Conducting Units: Tracheids and Vessels 1.1 Evolutionary Specialization The development of upright land plants depended on the development of a waterconducting system. Many of the earliest land plants,
Chapter. Transport in. Structure of. 1- Epidermis: 2- Cortex: All plants 2- a specialized. In higher moving by. hydra and. with cuticles) 1-2-
 1-2-") Chapter 2 Transport in living organisms The concept of transport and the need for it: All plants need CO 2, water and mineral salts to perform photosynthesis In primitive plants such as algae these materials
Chapter 2 Transport in living organisms The concept of transport and the need for it: All plants need CO 2, water and mineral salts to perform photosynthesis In primitive plants such as algae these materials
Multiple origins of respiratory structures of sea urchins OR I Didn t Know They Could Breathe!
 Multiple origins of respiratory structures of sea urchins OR I Didn t Know They Could Breathe! Cara Marie Breslin Dr. Rich Mooi Department of Invertebrate Zoology and Geology Slide 1: Title and introduction.
Multiple origins of respiratory structures of sea urchins OR I Didn t Know They Could Breathe! Cara Marie Breslin Dr. Rich Mooi Department of Invertebrate Zoology and Geology Slide 1: Title and introduction.
First Record of the Rudist Bivalve Mitrocaprina tschoppi (Palmer) from the Maastrichtian of Jamaica
 from the Maastrichtian of Jamaica") 392 Caribbean Journal of Science, Vol. 40, No. 3, 392-396, 2004 Copyright 2004 College of Arts and Sciences University of Puerto Rico, Mayagüez First Record of the Rudist Bivalve Mitrocaprina tschoppi
392 Caribbean Journal of Science, Vol. 40, No. 3, 392-396, 2004 Copyright 2004 College of Arts and Sciences University of Puerto Rico, Mayagüez First Record of the Rudist Bivalve Mitrocaprina tschoppi
Specific Characters.-Smaller than Amynodon advenus, A. antiquus and. Introduction.-Since the description' of a skull of Amynodontopsis bodei
 270 PALEONTOLOG Y: C. STOCK PROC. N. A. S. EOCENE AMYNODONTS FROM SOUTHERN CALIFORNIA BY CHESTER STOCK BALCH GRADUATE SCHOOL OF THE GEOLOGICAL SCIENCES, CALIFORNIA INSTITUTE OF TECHNOLOGY Communicated
270 PALEONTOLOG Y: C. STOCK PROC. N. A. S. EOCENE AMYNODONTS FROM SOUTHERN CALIFORNIA BY CHESTER STOCK BALCH GRADUATE SCHOOL OF THE GEOLOGICAL SCIENCES, CALIFORNIA INSTITUTE OF TECHNOLOGY Communicated
FIELD IDENTIFICATION CARDS FOR INVASIVE NON-NATIVE PLANT SPECIES KNOWN TO THREATEN ARIZONA WILDLANDS.
 FIELD IDENTIFICATION CARDS FOR INVASIVE NON-NATIVE PLANT SPECIES KNOWN TO THREATEN ARIZONA WILDLANDS. PACKET CONTAINS INFORMATION ON 74 INVASIVE NON-NATIVE PLANTS INCLUDING: morphologic descriptions photos
FIELD IDENTIFICATION CARDS FOR INVASIVE NON-NATIVE PLANT SPECIES KNOWN TO THREATEN ARIZONA WILDLANDS. PACKET CONTAINS INFORMATION ON 74 INVASIVE NON-NATIVE PLANTS INCLUDING: morphologic descriptions photos
Assembly Instructions for the 1-Wire Weather Station V2.0/V3.0
 Assembly Instructions for the 1-Wire Weather Station V2.0/V3.0 Tools and Supplies Required (not included): Phillips screwdriver 3/8" wrench Drill with 5/32" bit "U" type mounting bracket (i.e., part number
Assembly Instructions for the 1-Wire Weather Station V2.0/V3.0 Tools and Supplies Required (not included): Phillips screwdriver 3/8" wrench Drill with 5/32" bit "U" type mounting bracket (i.e., part number
Cornu s method for the determination of elastic constants of a Perspex beam Surjeet Singh
 Cornu s method for the determination of elastic constants of a Perspex beam Surjeet Singh Indian Institute of Science Education and Research Pune surjeet.singh@iiserpune.ac.in Objective: Using Cornu s
Cornu s method for the determination of elastic constants of a Perspex beam Surjeet Singh Indian Institute of Science Education and Research Pune surjeet.singh@iiserpune.ac.in Objective: Using Cornu s
THE ROOTS OF WILD RICE. ZIZANIA AQUATICA L.
 THE ROOTS OF WILD RICE. ZIZANIA AQUATICA L. E. L. STOVER, Eastern Illinois State Teachers College. This grass grows from Maine to Minnesota in aquatic habitats (2 and 5). It is common in marsh lands all
THE ROOTS OF WILD RICE. ZIZANIA AQUATICA L. E. L. STOVER, Eastern Illinois State Teachers College. This grass grows from Maine to Minnesota in aquatic habitats (2 and 5). It is common in marsh lands all
Prentice Hall EARTH SCIENCE
 Prentice Hall EARTH SCIENCE Tarbuck Lutgens Running Water and Groundwater Running Water The Water Cycle Water constantly moves among the oceans, the atmosphere, the solid Earth, and the biosphere. This
Prentice Hall EARTH SCIENCE Tarbuck Lutgens Running Water and Groundwater Running Water The Water Cycle Water constantly moves among the oceans, the atmosphere, the solid Earth, and the biosphere. This
Paleocurrents. Why measure paleocurrent directions? Features that give paleocurrent directions. Correction to paleocurrent measurements
 Why measure paleocurrent directions? Paleocurrent measurements can provide valuable information on ancient flow conditions, which can often shed light on paleogeography. For example, paleocurrent data
Why measure paleocurrent directions? Paleocurrent measurements can provide valuable information on ancient flow conditions, which can often shed light on paleogeography. For example, paleocurrent data
2013 Antiquarian Horological Society. Reproduction prohibited without permission.
 2013 Antiquarian Horological Society. Reproduction prohibited without permission. ANTIQUARIAN HOROLOGY Picture Gallery 30-hour iron wall clock dated 1748 The painted dial with the minute hand shorter than
2013 Antiquarian Horological Society. Reproduction prohibited without permission. ANTIQUARIAN HOROLOGY Picture Gallery 30-hour iron wall clock dated 1748 The painted dial with the minute hand shorter than
1/6/ th Grade Earth s Surface. Chapter 3: Erosion and Deposition. Lesson 1 (Mass Movement)
") Lesson 1 (Mass Movement) 7 th Grade Earth s Surface Chapter 3: Erosion and Deposition Weathering the chemical and physical processes that break down rock at Earth s surface Mechanical weathering when rock
Lesson 1 (Mass Movement) 7 th Grade Earth s Surface Chapter 3: Erosion and Deposition Weathering the chemical and physical processes that break down rock at Earth s surface Mechanical weathering when rock
THE UNIVERSITY OF MICHIGAN CRINOID FROM THE MIDDLE DEVONIAN SILICA FORMATION IN NORTHWESTERN OHIO
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 22, NO. 11, p. 155-162 (3 pls., 2 text-figs.) NOVEMBER 22, 1968 AMELIACRINUS BENDERI, A NEW DICYCLIC CAMERATE CRINOID FROM
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 22, NO. 11, p. 155-162 (3 pls., 2 text-figs.) NOVEMBER 22, 1968 AMELIACRINUS BENDERI, A NEW DICYCLIC CAMERATE CRINOID FROM
2 - Machining Fundamentals Measurement. Manufacturing Processes - 2, IE-352 Ahmed M El-Sherbeeny, PhD Spring-2015
 2 - Machining Fundamentals Measurement Manufacturing Processes - 2, IE-352 Ahmed M El-Sherbeeny, PhD Spring-2015 Learning Objectives Measure to 1/64 (.5 mm) with a steel rule Reading an Inch-based Vernier
2 - Machining Fundamentals Measurement Manufacturing Processes - 2, IE-352 Ahmed M El-Sherbeeny, PhD Spring-2015 Learning Objectives Measure to 1/64 (.5 mm) with a steel rule Reading an Inch-based Vernier
Bulletin Zoölogisch Museum
 Bulletin Zoölogisch Museum UNIVERSITEIT VAN AMSTERDAM Vol.11 No. 15 1988 Redescription of johanna Monod, 1926 Virgin Isls (Isopoda) from St. John, Hans Georg Müller Summary Based on the type material,
Bulletin Zoölogisch Museum UNIVERSITEIT VAN AMSTERDAM Vol.11 No. 15 1988 Redescription of johanna Monod, 1926 Virgin Isls (Isopoda) from St. John, Hans Georg Müller Summary Based on the type material,
What type of land feature is located at Point A? A Cliff B Delta C Mountain D Valley
 1 What type of land feature is located at Point A? A Cliff B Delta C Mountain D Valley Alfred Wegener s theory of continental drift was 2 not accepted by scientists when the theory was first proposed.
1 What type of land feature is located at Point A? A Cliff B Delta C Mountain D Valley Alfred Wegener s theory of continental drift was 2 not accepted by scientists when the theory was first proposed.
The luminescence of diamond-i
 Curr. Sci. 19 357-363 (1950) The luminescence of diamond-i SIR C V RAMAN 1. Introduction' No less than seventy-five distinct papers which concerned themselves with the structure and properties of diamond
Curr. Sci. 19 357-363 (1950) The luminescence of diamond-i SIR C V RAMAN 1. Introduction' No less than seventy-five distinct papers which concerned themselves with the structure and properties of diamond
Tuesday, February 11, Chapter 3. Load and Stress Analysis. Dr. Mohammad Suliman Abuhaiba, PE
 1 Chapter 3 Load and Stress Analysis 2 Chapter Outline Equilibrium & Free-Body Diagrams Shear Force and Bending Moments in Beams Singularity Functions Stress Cartesian Stress Components Mohr s Circle for
1 Chapter 3 Load and Stress Analysis 2 Chapter Outline Equilibrium & Free-Body Diagrams Shear Force and Bending Moments in Beams Singularity Functions Stress Cartesian Stress Components Mohr s Circle for
Wikipedia and Centrifugal Force
 Wikipedia and Centrifugal Force Frederick David Tombe, sirius184@hotmail.com 27 th July 2015 Abstract. Wikipedia is the on-line encyclopaedia that anybody can edit. The content changes on a continual basis.
Wikipedia and Centrifugal Force Frederick David Tombe, sirius184@hotmail.com 27 th July 2015 Abstract. Wikipedia is the on-line encyclopaedia that anybody can edit. The content changes on a continual basis.
F795. GEOLOGY Evolution of Life, Earth and Climate ADVANCED GCE. Tuesday 1 February 2011 Afternoon
 ADVANCED GCE GEOLOGY Evolution of Life, Earth and Climate F795 *OCE/30912* Candidates answer on the question paper. OCR supplied materials: None Other materials required: Electronic calculator Ruler (cm/mm)
ADVANCED GCE GEOLOGY Evolution of Life, Earth and Climate F795 *OCE/30912* Candidates answer on the question paper. OCR supplied materials: None Other materials required: Electronic calculator Ruler (cm/mm)
ESTIMATION OF CONSERVATISM OF CHARACTERS BY CONSTANCY WITHIN BIOLOGICAL POPULATIONS
 ESTIMATION OF CONSERVATISM OF CHARACTERS BY CONSTANCY WITHIN BIOLOGICAL POPULATIONS JAMES S. FARRIS Museum of Zoology, The University of Michigan, Ann Arbor Accepted March 30, 1966 The concept of conservatism
ESTIMATION OF CONSERVATISM OF CHARACTERS BY CONSTANCY WITHIN BIOLOGICAL POPULATIONS JAMES S. FARRIS Museum of Zoology, The University of Michigan, Ann Arbor Accepted March 30, 1966 The concept of conservatism
VARIATION IN THE SIZE OF RAY PITS OF CONIFERS.*
 VARIATION IN THE SIZE OF RAY PITS OF CONIFERS.* FOREST B. H. BROWN. Since Haeckel proposed the word Ecology in 88, there has been an ever growing interest in the influence which environmental factors may
VARIATION IN THE SIZE OF RAY PITS OF CONIFERS.* FOREST B. H. BROWN. Since Haeckel proposed the word Ecology in 88, there has been an ever growing interest in the influence which environmental factors may
6.1 Water. The Water Cycle
 6.1 Water The Water Cycle Water constantly moves among the oceans, the atmosphere, the solid Earth, and the biosphere. This unending circulation of Earth s water supply is the water cycle. The Water Cycle
6.1 Water The Water Cycle Water constantly moves among the oceans, the atmosphere, the solid Earth, and the biosphere. This unending circulation of Earth s water supply is the water cycle. The Water Cycle
THE UNIVERSITY OF MICHIGAN
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 25, NO. 10, p. 221-234 (5 text-figs.; 1 plate) December 3 1, 198 1 THE CRINOID TRIBOLOPORUS CRYPTOPLICATUS AND THE EVOLUTION
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 25, NO. 10, p. 221-234 (5 text-figs.; 1 plate) December 3 1, 198 1 THE CRINOID TRIBOLOPORUS CRYPTOPLICATUS AND THE EVOLUTION
PALEONTOLOGICAL CONTRIBUTIONS
 THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS November 30, 1967 Paper 28 NANNOFOSSIL SPECIES RELATED TO CYCLOCOCCOLITHUS LEPTOPORUS (MURRAY & BLACKMAN) STEFAN GARTNER, JR. [Esso Production Research
THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS November 30, 1967 Paper 28 NANNOFOSSIL SPECIES RELATED TO CYCLOCOCCOLITHUS LEPTOPORUS (MURRAY & BLACKMAN) STEFAN GARTNER, JR. [Esso Production Research
FoSSil Puzzler (1 Hour)
") FoSSil Puzzler (1 Hour) Addresses NGSS Level of Difficulty: 2 Grade Range: 3-5 OVERVIEW Students will examine and sketch various trace fossils from an unknown creature and then construct an image of the
FoSSil Puzzler (1 Hour) Addresses NGSS Level of Difficulty: 2 Grade Range: 3-5 OVERVIEW Students will examine and sketch various trace fossils from an unknown creature and then construct an image of the
5. Crater Rim Uplift and Crater Wall Collapse
 5. Crater Rim Uplift and Crater Wall Collapse The basic processes involved in crater rim uplift are understood, but there is a lot of evidence at the crater that has not been fully explored and may eventually
5. Crater Rim Uplift and Crater Wall Collapse The basic processes involved in crater rim uplift are understood, but there is a lot of evidence at the crater that has not been fully explored and may eventually
The Shoot System: Primary Stem Structure - 1
 The Shoot System: Primary Stem Structure - 1 Shoot System The shoot system comprises the leaves and stems of plants. Leaves are located at nodes on the stem; the distance along the stem between nodes is
The Shoot System: Primary Stem Structure - 1 Shoot System The shoot system comprises the leaves and stems of plants. Leaves are located at nodes on the stem; the distance along the stem between nodes is
LOOKING AT PLANT STEMS
 Activity 4.17 Student Sheet LOOKING AT PLANT STEMS Purpose To look at the structure of xylem vessels, phloem sieve tubes and sclerenchyma fibres. To locate the position of these tissues within the stem.
Activity 4.17 Student Sheet LOOKING AT PLANT STEMS Purpose To look at the structure of xylem vessels, phloem sieve tubes and sclerenchyma fibres. To locate the position of these tissues within the stem.
+Galileo Teacher s Network
 +Galileo Teacher s Network The Moon (and more) Presenters: Erin Wood Matthew Benjamin Alex Thom Think-Pair-Share Rules: Think and write (1 minute) Discuss with a partner (3 minutes) Share with the group
+Galileo Teacher s Network The Moon (and more) Presenters: Erin Wood Matthew Benjamin Alex Thom Think-Pair-Share Rules: Think and write (1 minute) Discuss with a partner (3 minutes) Share with the group
Paramecium. Sub-Order Peniculina. Genus Paramecium
 Paramecium Kingdom Animalia Phylum Protozoa Sub-Phylum Ciliophora Class Ciliata or Infusoria Sub-Class Holotricha Order Hymenostomatida Sub-Order Peniculina Family Paramecidae Genus Paramecium Introduction:
Paramecium Kingdom Animalia Phylum Protozoa Sub-Phylum Ciliophora Class Ciliata or Infusoria Sub-Class Holotricha Order Hymenostomatida Sub-Order Peniculina Family Paramecidae Genus Paramecium Introduction:
EROSION AND DEPOSITION
 CHAPTER 8 EROSION AND DEPOSITION SECTION 8 1 Changing Earth s Surface (pages 252-255) This section explains how sediment is carried away and deposited elsewhere to wear down and build up Earth s surface.
CHAPTER 8 EROSION AND DEPOSITION SECTION 8 1 Changing Earth s Surface (pages 252-255) This section explains how sediment is carried away and deposited elsewhere to wear down and build up Earth s surface.
THE UNIVERSITY OF MICHIGAN DOLATOCRINUS KIITASII, A NEW CRINOID FROM THE MIDDLE DEVONIAN BELL SHALE OF MICHIGAN
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 23, NO. 12, p. 201-211 (5 pls., 1 text-fig.) FEBRUARY 22, 1971 DOLATOCRINUS KIITASII, A NEW CRINOID FROM THE MIDDLE DEVONIAN
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 23, NO. 12, p. 201-211 (5 pls., 1 text-fig.) FEBRUARY 22, 1971 DOLATOCRINUS KIITASII, A NEW CRINOID FROM THE MIDDLE DEVONIAN
SPECTROSCOPIC STUDY OF LUMINESCENCE PATTERNS IN DIAMOND BY ANNA MANI. Received August 21, 1944 (Communicated by Sir C. V. Raman, Kt., F.R.S., N.L.
 SPECTROSCOPIC STUDY OF LUMINESCENCE PATTERNS IN DIAMOND BY ANNA MANI (From the Department of Physics, Indian Institute of Science, Bangalore) Received August 21, 1944 (Communicated by Sir C. V. Raman,
SPECTROSCOPIC STUDY OF LUMINESCENCE PATTERNS IN DIAMOND BY ANNA MANI (From the Department of Physics, Indian Institute of Science, Bangalore) Received August 21, 1944 (Communicated by Sir C. V. Raman,
Directed Reading. Section: Volcanoes and Plate Tectonics
 Skills Worksheet Directed Reading Section: Volcanoes and Plate Tectonics 1. Some volcanic eruptions can be more powerful than a(n) a. hand grenade. b. earthquake. c. geyser. d. atomic bomb. 2. The cause
Skills Worksheet Directed Reading Section: Volcanoes and Plate Tectonics 1. Some volcanic eruptions can be more powerful than a(n) a. hand grenade. b. earthquake. c. geyser. d. atomic bomb. 2. The cause
 Question 1: State the location and function of different types of meristem. Meristems are specialised regions of plant growth. The meristems mark the regions where active cell division and rapid division
Question 1: State the location and function of different types of meristem. Meristems are specialised regions of plant growth. The meristems mark the regions where active cell division and rapid division
Section 3 Deforming Earth s Crust
 Section 3 Deforming Earth s Crust Key Concept Tectonic plate motions deform Earth s crust. Deformation causes rock layers to bend and break and causes mountains to form. What You Will Learn Stress is placed
Section 3 Deforming Earth s Crust Key Concept Tectonic plate motions deform Earth s crust. Deformation causes rock layers to bend and break and causes mountains to form. What You Will Learn Stress is placed
Granite School District Parent Guides Utah Core State Standards for Mathematics Grades K-6
 Granite School District Parent Guides Grades K-6 GSD Parents Guide for Kindergarten The addresses Standards for Mathematical Practice and Standards for Mathematical Content. The standards stress not only
Granite School District Parent Guides Grades K-6 GSD Parents Guide for Kindergarten The addresses Standards for Mathematical Practice and Standards for Mathematical Content. The standards stress not only
OSCILLATIONS OF A SPRING-MASS SYSTEM AND A TORSIONAL PENDULUM
 EXPERIMENT Spring-Mass System and a Torsional Pendulum OSCILLATIONS OF A SPRING-MASS SYSTEM AND A TORSIONAL PENDULUM Structure.1 Introduction Objectives. Determination of Spring Constant Static Method
EXPERIMENT Spring-Mass System and a Torsional Pendulum OSCILLATIONS OF A SPRING-MASS SYSTEM AND A TORSIONAL PENDULUM Structure.1 Introduction Objectives. Determination of Spring Constant Static Method
4. What type of glacier forms in a sloping valley between rock walls? a. firn glacier b. ice sheet c. cirque d. alpine glacier
 Multiple Choice Questions 1. The term means the loss of snow and ice by evaporation and melting. a. sublimation b. ablation c. erosion d. abrasion 2. What condition must be met for a glacier to begin flowing
Multiple Choice Questions 1. The term means the loss of snow and ice by evaporation and melting. a. sublimation b. ablation c. erosion d. abrasion 2. What condition must be met for a glacier to begin flowing
Lecture 4 Root Put line under your answer! There is only one correct answer in the multiple choice questions
 Lecture 4 Root Put line under your answer! There is only one correct answer in the multiple choice questions 1. The perception of gravity by a root is thought to take place in a) root hairs b) the region
Lecture 4 Root Put line under your answer! There is only one correct answer in the multiple choice questions 1. The perception of gravity by a root is thought to take place in a) root hairs b) the region