Response of non-native European terrestrial gastropods to novel climates correlates with biogeographical and biological traits
|
|
|
- Andrea Harvey
- 6 years ago
- Views:
Transcription

1 bs_bs_banner Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2014) 23, RESEARCH PAPER Response of non-native European terrestrial gastropods to novel climates correlates with biogeographical and biological traits César Capinha 1,2 *, Dennis Rödder 2, Henrique Miguel Pereira 1 and Heike Kappes 3,4 1 Centro de Biologia Ambiental, Faculdade de Ciências da Universidade de Lisboa, Lisboa, Portugal, 2 Zoologisches Forschungsmuseum Alexander Koenig, Museumsmeile Bonn, Adenauerallee 160, Bonn, Germany, 3 Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, The Netherlands, 4 Department of Ecology, Cologne Biocenter, University of Cologne, Köln, Germany *Correspondence: César Capinha, Centro de Biologia Ambiental, Faculdade de Ciências da Universidade de Lisboa, Lisboa, Portugal. cesarcapinha@outlook.com ABSTRACT Aim Anticipating the propensity of species to persist outside the climatic conditions in which they are observed is important in assessing the uncertainty in climate-matching when applied to different locations or times. By using data from 27 European terrestrial gastropods (slugs and snails) established in new regions, we measured (1) the degree of climate match between native and non-native ranges and (2) the diversity of novel climatic conditions inhabited. We then tested for species traits as predictors of the patterns found. Location Worldwide. Methods We projected the occurrences of each species onto the climatic space defined by the two main axes of a principal components analysis. On the basis of the convex hull of native occurrences, we measured the proportion of non-native occurrences in novel climates and the diversity of the newly occupied climates. Generalized estimating equations were used to test for associations between climatic responses and species traits, while considering phylogenetic relatedness and introduction effort. Results Approximately half of the species had a large proportion (> 25%) of non-native occurrences in novel climates. Climate mismatch was significantly higher for species with narrow native climatic niches, native ranges elongated in a north south direction or native ranges with the southern limit at lower latitudes. Slugs occupied a higher diversity of novel climatic conditions than snails. Main conclusions Climate matching showed varying levels of accuracy in predicting the non-native ranges of the studied species. Lower accuracy appeared to be mainly related to low levels of distributional equilibrium in native ranges. In addition, coarse-scale predictions for species that are able to take advantage of microclimatic variability as appears to be the case with slugs may be challenging. We conclude that species traits can contribute to understanding uncertainties in climate-matching exercises. Keywords Climate matching, climatic niche, global change, invasive species, niche conservatism, risk assessment, species traits, terrestrial gastropods. INTRODUCTION Climatic requirements play a crucial role in shaping species distributions (Gaston, 2003). Consequently, many researchers have attempted to predict the potential spread of invasive species or the response of biodiversity to climate change by matching the climatic conditions in which species are observed to the conditions in new areas or time periods. Several studies 2014 John Wiley & Sons Ltd DOI: /geb
2 C. Capinha et al. have shown reliable results for this approach (e.g. Thuiller et al., 2005; Hayes & Barry, 2008; Guo et al., 2012), but recent analyses of large-scale biological invasions have shown that several species have become naturalized in novel climatic conditions (e.g. Broennimann et al., 2007; Fitzpatrick et al., 2007; Capinha et al., 2013). These idiosyncratic occurrences reduce the reliability of correlative approaches for predicting invasion risk or biodiversity change (Pearman et al., 2008) and underscore the need for a better understanding of the circumstances and mechanisms that affect the ability of non-native species to persist in novel climates (Pearman et al., 2008; Alexander & Edwards, 2010). Underlying the climate-matching approach is the assumption that, in the absence of source sink dynamics, the environmental conditions in which a species is observed (i.e. the realized niche, sensu Soberón & Nakamura, 2009) provide a reliable representation of the set of abiotic conditions in which it can maintain self-sustaining populations (i.e. the fundamental niche). Because of limited dispersal abilities or negative biotic interactions, however, the realized niche is often a subset of the fundamental niche (Araújo & Pearson, 2005). In addition, a species fundamental niche may include conditions that do not exist in the landscape. Therefore, even if distributional equilibrium occurs (i.e. the full range of existing suitable conditions is occupied), the fundamental niche may still not be fully inhabited (Soberón & Nakamura, 2009; Barve et al., 2011; Jiménez-Valverde et al., 2011). Finally, evolutionary mechanisms such as genetic drift or local adaptation may modify the species ecophysiology, allowing some populations to survive in otherwise unsuitable conditions (Alexander & Edwards, 2010; Hoffmann & Sgrò, 2011). Either independently or in combination, the above factors can cause correlative prediction methods to underestimate the ability of species to cope with novel climates. Assuming that the processes that limit the distribution can be identified, the use of models based on ecophysiology (i.e. mechanistic models) may improve the robustness of predictions, particularly if used in combination with climate-matching (Kearney et al., 2010). Because mechanistic models can be complex and time-consuming, however, their application for large-scale assessments remains impracticable. Alternatively, the identification of indicators of species responses to novel climates can be examined (Pearman et al., 2008). Species traits, either extrinsic or intrinsic, may indicate the tendency of species to conserve or shift their realized or fundamental climatic niches (Lavergne & Molofsky, 2007; Pyšek et al., 2009; Alexander & Edwards, 2010). For instance, species with low competitive or dispersal abilities are often associated with low levels of distributional equilibrium (e.g. Svenning & Skov, 2007), whereas high phenotypic plasticity in traits related to climate adaptation may favour the ability of species to persist in novel climates (Hoffmann & Sgrò, 2011). If consistent associations are observed between variables representing the above or other important mechanisms and patterns of changes in climatic niches, then justifiable levels of confidence can be assigned to agreeing or disagreeing correlative predictions. Despite the potential value of such indicators, the relationship between the ability to survive in novel climates and species traits remains largely untested, particularly for invasive fauna (but see Strubbe et al., 2013, for a recent example with birds). In this study, we evaluate the propensity of 27 European terrestrial gastropods to become established in novel climates in non-native ranges. This group of species is particularly appropriate for our study because they were introduced to their non-native ranges by different routes (e.g. unintentional introduction through horticultural or agricultural import, intentional introduction as biological control agents, or intentional introduction and cultivation for human consumption) and have a long history of introduction into many climatically distinct regions of the world (Cowie, 2011). Moreover, terrestrial gastropods are desiccation-prone ectotherms and are expected to be particularly responsive to climatic conditions (Heller, 2001; Luchtel & Deyrup-Olsen, 2001). We initially determined whether the non-native populations were established in nativelike or novel climates. We also classified the newly colonized climates into four types colder and drier, colder and wetter, warmer and drier, or warmer and wetter than the climate in the species native range. We tested a set of ecological and biological traits, considering phylogenetic relatedness and introduction effort, as predictors of (1) the proportion of non-native populations in novel climatic conditions, (2) the presence or absence of non-native populations in each of the novel climatic types, and (3) the total number of novel climate types inhabited. Our aim was to identify traits that may serve as indicators of species responses to novel climatic conditions that, in turn, may be used to complement predictions of correlative species distribution models applied across space or time. METHODS Species selection and distribution data To select suitable species for our study, we created a list of native European terrestrial gastropods with naturalized ranges (after the year 1500) based on two online databases (White-McLean, 2011; AnimalBase Project Group, 2012). We excluded species for which gene sequence data appropriate for phylogeny reconstruction was not publicly available (see Phylogenetic relatedness section; see also flow-chart in Appendix S1 for the full list of species and a schematic representation of all selection criteria). Frequently misidentified species (cf. White-McLean, 2011) were excluded to reduce the number of taxonomic errors in the distribution data. For the remaining species, we collected native and non-native occurrence data from multiple sources, including monographs, research papers and online databases (Appendix S2). We restricted our searches to observations made after 1950 (the period for which climatic data were obtained; see below). We collected three forms of data geographical coordinates, mapped occurrences and names of locations where specimens were found. We assessed the positional accuracy of the coordinates by projecting them onto a global map in ArcGIS. Records 858 Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd
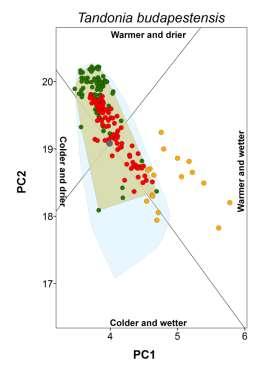
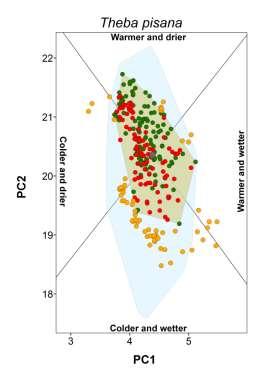
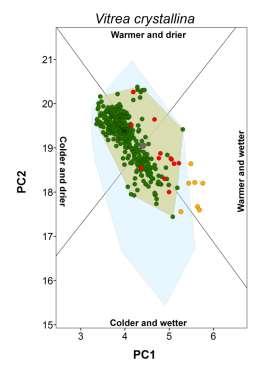
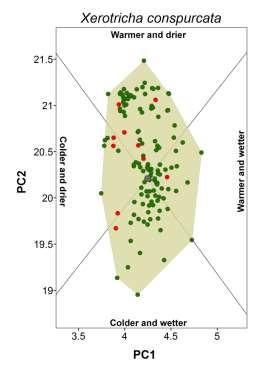
3 Climate-matching and species traits Table 1 Patterns of establishment under novel climatic conditions of 27 species of European terrestrial gastropods. Proportion refers to the percentage of non-native populations falling outside the native climate space. The percentage of non-native populations outside the climate space realized in the native range but still within climatic conditions potentially available in the native region is given in parentheses. Columns from colder and drier to warmer and wetter indicate the presence of non-native populations in that novel climatic setting. The column any change refers to the existence on non-native population in any of the previous conditions. The column diversity corresponds to the sum of novel types of climatic conditions occupied. Species Proportion (%) Colder and drier Colder and wetter Warmer and drier Warmer and wetter Any change Diversity Arianta arbustorum (Linnaeus, 1758) 0.0 no no no no no 0 Arion intermedius (Normand, 1852) 38.9 (9.9) no yes yes yes yes 3 Cepaea hortensis (Müller, 1774) 0.0 no no no no no 0 Cepaea nemoralis (Linnaeus, 1758) 0.0 no no no no no 0 Cernuella virgata (Da Costa, 1778) 3.7 (3.7) no no no no no 0 Chilostoma cingulatum (Studer, 1820) 0.0 no no no no no 0 Cochlicella acuta (Müller, 1774) 40.7 (3.7) no no no yes yes 1 Cornu aspersum (Müller, 1774) 27.6 (9.5) no yes yes yes yes 3 Deroceras reticulatum (Müller, 1774) 34.2 (10.9) yes yes yes yes yes 4 Discus rotundatus (Müller, 1774) 0.0 no no no no no 0 Eobania vermiculata (Müller, 1774) 33.3 (6.6) no no no yes yes 1 Helix lucorum Linnaeus, (3.3) no no no no no 0 Helix pomatia Linnaeus, no no no no no 0 Hygromia cinctella (Draparnaud, 1801) 5.0 (3.8) yes no no no yes 1 Lauria cylindracea (Da Costa, 1778) 3.7 (3.7) no no no no no 0 Limacus flavus Linnaeus, (8.3) no yes yes yes yes 3 Marmorana serpentina (Férussac, 1821) 60.0 (10.0) no no no yes yes 1 Monacha cartusiana (Müller, 1774) 2.9 (2.9) no no no no no 0 Otala lactea (Müller, 1774) 75.0 (12.5) no yes no yes yes 2 Oxychilus alliarius (Miller, 1822) 66.2 (12.9) no no yes yes yes 2 Oxychilus cellarius (Müller, 1774) 33.8 (4.4) no no no yes yes 1 Rumina decollata (Linnaeus, 1758) 42.0 (2.9) no no yes yes yes 2 Tandonia budapestensis (Hazay, 1881) 16.8 (7.9) no yes no yes yes 2 Testacella haliotidea Draparnaud, (6.6) no no no yes yes 1 Theba pisana (Müller, 1774) 40.7 (33.1) yes yes yes yes yes 4 Vitrea crystallina (Müller, 1774) 36.8 (5.3) no no no yes yes 1 Xerotricha conspurcata (Draparnaud, 1801) 0.0 no no no no no 0 showing incorrect placement (e.g. in the sea) were either deleted or corrected (if additional information was available). Maps of occurrences were imported and georeferenced in ArcGIS, and geographical coordinates were assigned to each occurrence point. For locality names, we extracted the coordinates of geographical centre-points using Google Earth (Google, Mountain View, CA, USA; available at: earth/). Localities that could not be unambiguously identified were ignored. In all cases, we evaluated common sources of positional uncertainty (e.g. map scale, extent of localities and arithmetical precision of the given coordinates) and records with a geographical uncertainty of more than 10 km were not considered. Finally, we converted the occurrence datasets of each species to one record per 20 km 20 km grid cell. This level of spatial detail is expected to be compatible with the purposes of our study (e.g. Petitpierre et al., 2012) and allowed most of the collected data to be used. Before the data were treated any further, we confirmed whether the occurrence records we had collected could be used for assessing the climatic conditions occupied by each species in the native and non-native ranges. Using GIS, we drew an approximate representation of the native and non-native range boundaries of each species. Native range boundaries were drawn based mainly on information from Kerney & Cameron (2006), the AnimalBase Project Group (2012) and IUCN (2013). Non-native ranges were based mainly on Herbert (2010) and White-McLean (2011) and references cited therein. In both cases, we also used any other relevant information (see Appendix S2). We superimposed the corresponding set of species occurrences over each of the range maps. Species for which undersampling could be visually detected (e.g. noticeable coverage gaps or the absence of records in a non-native region) were excluded from our study. Our final dataset included 27 species (Table 1) spread across 13 families, with 16,814 occurrences in native ranges (mean 623; max. 3805; min. 13) and 2470 occurrences in naturalized ranges (mean 92; max. 506; min. 7). We have no reason to assume that these species are not representative of the pool of European terrestrial gastropods with non-native ranges. Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd 859
4 C. Capinha et al. Climatic variables To shape species distributions, climatic factors must affect the organism s physiology (Rödder et al., 2009); for terrestrial gastropods, this involves both temperature and humidity. The variability and interaction between these factors have a direct influence on several key aspects of the biology and phenology of these taxa, including growth rates, egg production and mortality, and the onset of dormancy/activity (Cook, 2001; Heller, 2001). Perhaps most important is the fact that terrestrial gastropods have a permeable integument that makes them particularly vulnerable to dehydration or hyperhydration (Luchtel & Deyrup-Olsen, 2001). Given this strong physiological influence, we collected six spatially explicit climatic variables from WorldClim (Hijmans et al., 2005) representing means, extremes or seasonal variation in temperature and precipitation ( ): annual mean temperature (amt); maximum temperature of the warmest month (maxtwm); minimum temperature of the coldest month (mintcm); annual precipitation (anpre); precipitation of the driest quarter (predrq); and precipitation of the wettest quarter (prewetq). To match the resolution of the species occurrence data, all variables were collected at resolution and projected onto a 20 km 20 km grid in ArcGIS. Identification of establishment under novel climatic conditions The purpose of this analysis was to compare the climatic conditions occupied by the naturalized and native populations of each species. For that purpose, we reduced the dimensionality of the climatic dataset using principal components analysis (PCA). This was performed in ArcGIS after standardizing all the variables to zero mean and unit variance, i.e. PCA on the correlation matrix. We retained the first two axes of the PCA, which accounted for more than 89% of the underlying climatic variation (PC1, 58.6%; PC2, 30.7%). Next, we projected the scores of these two axes onto the geographical space and the value of each was assigned to the overlaying species occurrences. We then used these scores to project the conditions occupied by the native and non-native occurrences of each species onto the twodimensional climatic space. We defined native-like conditions as any climatic combination falling within the convex hull of the cloud of native occurrences. We also captured the climatic space potentially available to each species in the native region. For this, we considered all contiguous climatic conditions in the geographical space within 100 km of each native occurrence to be available and delimited the convex hull of these conditions in the climatic space. We expect this buffer to represent available habitats, because most of the species in our study are synanthropic and are frequently dispersed passively by humans (Kerney & Cameron, 2006; Kappes & Schilthuizen, 2014). We also recognize, however, that this distance may not apply equally well to all species, and consequently, the delimited conditions were used only as general indicators of climatic availability and were not used in responsetrait calculations (see below). Naturalized populations falling outside the native convex hull were further assigned to one of four main types of novel climates: colder and drier; colder and wetter; warmer and drier; or warmer and wetter. We distinguished these conditions based on the intercorrelation of the variables defining the two-dimensional PCA space. On the basis of these relationships, we delimited the relative location of the each type of novel climate by intersecting the two semiplanes perpendicular to the loading vector of annual mean temperature (the central vector of temperature variables) with the two semiplanes perpendicular to the loading vector of annual precipitation (the central vector of precipitation variables). For each species, we then centred the origin of these planes in the centroid of the convex hull of native conditions. Finally, we classified the climatic position of non-native occurrences under the novel conditions based on the planes it overlaid (see Fig. 1 and Appendix S3 for a more detailed explanation, and Appendix S4 for plots for each species). On the basis of the framework described above, we recorded eight distinct responses to novel climates: (1) the proportion of non-native occurrences falling outside the climate space realized in the native range; (2) the proportion of non-native occurrences falling outside the climate space realized in the native range but within the climatic space available in the native region; (3) the presence or absence of non-native populations under any novel climatic conditions; (4 7) the presence or absence of nonnative populations in each of the four types of novel climates; and (8) the diversity of newly colonized climates measured as the number of novel climate types occupied. For the latter six responses, we only classified a species as established outside the native climatic niche if there were three or more occurrence records, to reduce any bias caused by incorrect georeferencing or taxonomic misidentification. Trait data We collected data on egg volume, maximum shell length (snails only), presence or absence of a protective shell, presence or absence of polymorphism in shell colour or banding (snails only) and reproductive strategy (semelparous or iteroparous); data were collected from the literature (e.g. Falkner et al., 2001; Heller, 2001; Kerney & Cameron, 2006; Cowie et al., 2009). These traits were selected because life-history strategies and phenotypic plasticity may be related to the ability of terrestrial gastropods to occupy novel climatic settings (Heath, 1975; Heller, 2001). We also gathered trait data referring to the species ecology and biogeography. These included habitat breadth, humidity preference, breadth of realized native climatic niche, ratio of latitudinal to longitudinal ranges, latitudes of the northern and southern range limits, and latitudinal range between the northern and southern range limits. Habitat breadth corresponded to the sum of first-level macrohabitat categories defined by Falkner et al. (2001). For a few species absent from this database, we verified the use of each of these macrohabitats from complementary literature (Appendix S5). Humidity preference, i.e. the relative occurrence of the species under wet, 860 Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd
.")
5 Climate-matching and species traits Figure 1 Climatic conditions occupied by native and non-native populations of the brown garden snail (Cornu aspersum) along the first two axes of a principal components analysis (PCA). Names of locations are indicated for the most representative non-native ranges that are found outside the climatic conditions occupied in the native range, as defined by the convex hull of native conditions. The inset depicts the correlations between the climatic variables in the PCA and the four sectors that were defined on the basis of the collinearity of the variables in the subspace of the two main axes of the PCA. The climatic variables used were annual mean temperature (amt), maximum temperature of warmest month (maxtwm), minimum temperature of coldest month (mintcm), annual precipitation (anpre), precipitation of the driest quarter (predrq) and precipitation of the wettest quarter (prewetq). moist or dry conditions, was based on Falkner et al. (2001). We used the category with the highest relative occurrence to represent the condition preferred by the species. Information for species not found in the database was retrieved from the literature (Appendix S5). The breadth of the realized native climatic niche corresponded to the area of the convex hull enclosing the climatic conditions occupied by the species in its native range. The relationship between the latitudinal and longitudinal range extent was calculated in GIS and corresponded to the ratio of the Euclidean distance (measured in km) between the 5% most peripheral occurrences in each extreme of the y-axis to its equivalent on the x-axis. Finally, we measured the northernmost and southernmost latitude of the species native ranges from the trimmed range limits. We also aimed to quantify the extent to which the species were dispersed worldwide. Global introduction efforts are expected to bias patterns of climate matching because they can mediate the diversity of climatic conditions made accessible to each species. To represent this factor, we identified the non-native areas of the world (represented by each 20-km grid cell) having climates equivalent to those of the native range (i.e. falling inside the native-range convex hull) for each species. We then measured the proportion of these areas with the species present. By controlling for climatic compatibility, we expected this index to mainly reflect the influence of introduction effort, since these two factors are often considered to be the most important in determining the extent of non-native ranges (e.g. Thuiller et al., 2005; Hayes & Barry, 2008; Pyšek et al., 2009). Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd 861
6 C. Capinha et al. Phylogenetic relatedness Species cannot be treated as independent samples in statistical analyses of comparative data because of phylogenetic autocorrelation. If species are used as independent samples, degrees of freedom may be inflated and statistical significance becomes equivocal (Paradis & Claude, 2002). We therefore built a phylogenetic tree for our 27 species, using nucleotide sequences available from GenBank. The majority of sequences came from Wade et al. (2001, 2006) (3 end of the 5.8S gene, complete ITS-2 region, and the 5 end of the 28S gene), but homologous sequences from other studies were also included (see Appendix S6 for GenBank accession numbers). For two species for which compatible data were unavailable Arion intermedius and Testacella haliotidea we used sequence data from their close relatives Arion hortensis Férussac, 1819 and Testacella scutulum Sowerby, 1821, respectively. The surrogate of A. intermedius was selected based on similarity levels of alternative sequences in blast ( T. scutulum is sometimes considered a variety of T. haliotidea (Kerney, 1999). No congeners of these species occurred in our dataset. We used the opisthobranch Aplysia punctata (Cuvier, 1803) as an outgroup (cf. Wade et al., 2006). Sequences were aligned using muscle (Edgar, 2004) and combined into a single matrix. Finally, we built maximum-likelihood phylogenies in mega (Tamura et al., 2011). We used the general time-reversible model of nucleotide substitution with discrete gamma-distributed rates and a proportion of invariant sites (GTR+γ + I), which was the best fitting according to the Akaike information criterion in jmodeltest 2 (Darriba et al., 2012). Data analyses We analysed the association between the species responses to novel climates and the collected traits using generalized estimating equations (GEEs). GEEs correspond to generalizations of standard generalized linear models that explicitly account for correlation among samples (Paradis & Claude, 2002). We applied GEE using the ape package (Paradis et al., 2004) in R. This implementation translates the species phylogenetic tree into a species-to-species correlation matrix that is used as measure of strength of the dependence among samples (i.e. species) in GEE. Moreover, the degrees of freedom of GEE parameters calculated by ape are based on the procedure suggested by Paradis & Claude (2002), which considers both the number of species analysed and their phylogenetic relatedness. Before running the GEE analyses, we tested for collinearity among trait variables using Kendall s rank correlation. No strong correlations were found (absolute tau > 0.8). Next, we used GEE to test for significant relationships between each response variable and each of the species traits. We included introduction effort as a covariate in all models to account for the confounding effect of this factor. We fitted GEE binomial models for binary response variables (i.e. the presence or absence of non-native distributions outside the native climatic space and presence or absence of non-native distributions in each of the types of novel climates), Gaussian models for the dependent variable representing the proportion of non-native occurrences under novel climates (log-transformed), and Poisson models for the diversity of novel climates colonized (i.e. count data). RESULTS Establishment under novel climatic conditions The proportion of non-native occurrences in novel climates differed markedly among species. Of the 27 species analysed, 14 had more than 25% of non-native occurrences outside their native climatic space and, of these, four exceeded 50% (Table 1). Conversely, 12 species had only marginal (i.e. < 5%) or no establishment in novel conditions. Most of the newly colonized climates were not available in the species native regions (Table 1). In total, 16 species had three or more non-native occurrences in at least one novel climatic type (Table 1). Warmer and wetter climates were the most commonly occupied (15 species), followed by colder and wetter and warmer and drier climates (seven species each); colder and drier climates were the least occupied (three species). The total diversity of novel climates occupied also varied widely among species. Two species were found in all four types of climatic conditions (Table 1), three species were found in three types (see Fig. 1 for one example), four species in two types, and seven species in only one type of novel climate each. Predictors of establishment under novel climatic conditions GEE models showed that the proportion of non-native occurrences in novel climates is unrelated to the introduction effort (P > 0.05) (Appendix S7). However, the models suggested that species displaying wider climatic-niche breadths in their native range have a lower probability of becoming established outside the native climatic niche (d.f. phylogenetic = 8.06, coefficient estimate = , P = 0.040) while the opposite was found for species with native ranges elongated north south (d.f. phylogenetic = 8.06, coefficient estimate = 0.403, P = 0.004). The probability of climate mismatch was also higher for species having southern range limits at lower latitudes (d.f. phylogenetic = 8.06, coefficient estimate = 0.051, P = 0.008). None of the remaining traits was significantly associated with this response variable. For several binomial GEE models, the phylogenetic correlation of the response variable was very high, which caused the effective sample size to be insufficient for model convergence (Appendix S7). When model convergence was achieved, we found no significant association with the tested traits. The GEE models that analysed the diversity of novel climatic conditions occupied identified a significant positive association with introduction effort (d.f. phylogenetic = 8.06, coefficient estimate = 0.664, P = 0.026). In addition, diversity was significantly related to the presence or absence of a protective shell (d.f. phylogenetic = 8.06, coefficient estimate = 0.803, P = 0.034), 862 Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd
7 Climate-matching and species traits which suggests that species without a shell (i.e. slugs) are found in a more diverse range of novel climates than species with a shell (i.e. snails). DISCUSSION Our results show that climate-matching could predict nonnative occurrences of about half of the studied species while, for the remaining half, a considerable proportion of these occurrences (> 25%) was unpredictable. These contrasting levels of accuracy mirror a pattern emerging in the literature that could address the effectiveness of climate-matching at predicting nonnative ranges. In some cases (e.g. Thuiller et al., 2005; Hayes & Barry, 2008; Guo et al., 2012), most of the non-native range was predicted, whereas in others (e.g. Broennimann et al., 2007; Fitzpatrick et al., 2007; Capinha et al., 2013), the predictions failed. Consequently, identification of the circumstances under which species are likely to shift or conserve realized and fundamental niches is crucial for anticipating the reliability of these predictions (Pearman et al., 2008). Although manipulative experiments are necessary for differentiating between shifts in realized or fundamental niches and to identify the processes that foster or hinder these shifts, our results suggest that the analysis of species traits may provide valuable information. We found that species with narrower native climatic niches or with latitudinally elongated native ranges tended to be less predictable. Despite being distinct, these two traits appear to reflect the level of distributional equilibrium of species in their native regions. The breadth of native climatic niches is dictated by several factors, including the species actual physiological tolerances. In the case of our species, however, the effect of geographical barriers to dispersal appears to be one of the most important factors. For example, the two species with narrowest climatic niches were Marmorana serpentina and Otala lactea. Marmorana serpentina is an insular species, whereas O. lactea is a Mediterranean species that occurs only in a small section of the Mediterranean basin (southern Iberian Peninsula, Balearic Islands and Morocco). These two species showed the thirdhighest (60%) and highest (75%) proportion of climate mismatch, respectively. The relationship between vertically elongated native ranges and distributional equilibrium is even more apparent. Variation in the direction of the elongation of species ranges indicates the relative role of climate in shaping range boundaries (Brown et al., 1996). Because of the latitudinal gradient of temperatures worldwide, a north south elongation suggests a reduced role of climate, particularly of temperature, in shaping species distributions. In other words, species with latitudinally elongated range boundaries are expected to be strongly conditioned by non-climatic constraints (e.g. biological enemies or natural barriers) and their distributions are likely to provide a poor indication of the environmental conditions they tolerate (i.e. their fundamental niche). Indeed, previous research has warned about the increased uncertainty of predictions made for species in these circumstances (Araújo & Pearson, 2005). Our results indicate that this uncertainty can, at least to some extent, be assessed a priori by investigating the geographical attributes of the species native ranges. Our results also showed that species native to, but not necessarily restricted to, southern regions (i.e. southern Europe and North Africa) had a higher propensity for climate mismatch. The reason for this relationship is not clear. One possibility is that species restricted to high latitudes inhabit the warm edge of their thermal niche, while some of those occurring further south would not be able to do so because of the Mediterranean Sea (again a manifestation of distributional disequilibrium). Species reaching further south may also be better able to tolerate the climatic conditions at lower latitudes (e.g. in subtropical or tropical areas). For instance, most species native to the Mediterranean climate can tolerate high temperatures and drought and may therefore be pre-adapted to cooler and more humid conditions (i.e. the novel climates are within their fundamental niche; Alexander & Edwards, 2010). Our results show that most species native to southern Europe or North Africa have nonnative populations in climates that are wetter and warmer (Table 1). However, because of data limitations (see Results), we could not test the statistical significance of the association. We found that slugs colonized a wider diversity of novel climates than snails. There are many examples of range expansion of European slugs into novel climates (White-McLean, 2011) and our findings further support this pattern. Perhaps the bestknown example is the European invasion of the slug Arion lusitanicus auct. non Mabille, 1868, a species that was originally confined to a limited area of Western Europe. Although one could expect snails to inhabit a wider diversity of climatic conditions, because having a shell provides protection from heat and dehydration, our findings suggest that the opposite is true. The reason for this may be the greater ability of slugs to take advantage of microclimates. Because slugs are shell-less, their bodies are more plastic, which may allow slugs to better search for and exploit suitable microenvironments (Luchtel & Deyrup- Olsen, 2001). Slugs also appear to be very selective in choosing these microenvironments and regularly exhibit homing behaviour (Cook, 2001), a strategy that ensures refuge from adverse macroclimates. Recently, Suggitt et al. (2011) showed that local thermal variability might compensate to some extent for thermal changes in extreme climate change. It seems likely that the ability to exploit microclimatic refuges in new areas may also enhance resistance to adverse macroclimates. Our results also show clear differences between the types of novel climatic conditions inhabited by species in their nonnative ranges, and show that warmer and wetter regions were the most commonly colonized. We did not, however, find any indication of these patterns being related to any of the analysed traits. We also found no significant association between any of the patterns of climatic niche change and traits related to the species life history (i.e. egg volume, semelparity versus iteroparity, or shell size). Some studies have suggested that species that produce large numbers of offspring that grow and reproduce quickly but are small-sized and have a short life-span (i.e. r-strategists) will be better able to become successfully established outside their native regions (see examples in Sakai Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd 863
8 C. Capinha et al. et al., 2001). Our results suggest that, if these attributes do facilitate establishment, it is because they aid the species in overcoming challenges other than the change in climatic conditions (e.g. demographic stochasticity). The same conclusion appears to apply to habitat preferences or habitat generalism, because no relationship was found between the traits and any of the climatic responses measured. The albedo of shells determines the amount of solar radiation absorbed by snails and, consequently, their body temperature (Heath, 1975); plasticity in this trait may therefore favour survival in stressful climates through selection. We also found no distinct climatic responses between snails having variability in shell colour or banding and those that do not, however. Our results agree with other studies (e.g. Heller, 1981) in which this relationship was not observed. There are several other methods for evaluating climatematching predictions. Most distinctive from our approach are those that restrict predictions to climatic conditions shared between native and non-native regions (e.g. Petitpierre et al., 2012) in an attempt to exclude climate mismatch due to preadaptation. As shown by our results, however, many of the climatic combinations occupied by species in non-native areas appear not to be available in native regions. Thus, disregarding species responses to novel climatic conditions is likely to lead to an underestimation of their potential ranges (Webber et al., 2012). Of course, physiologically based predictions are preferable in this context, but, because they may not be obtainable, recent studies have also investigated how the use of species distribution data can be optimized for new environments (e.g. Elith et al., 2010; Owens et al., 2013). For instance, Owens et al. (2013) introduced the mobility-orientated parity metric, a modification and extension of the multivariate environmental similarity surface (Elith et al., 2010), which allows the locations in which extrapolation takes place to be mapped, and the degree of novelty of the environmental conditions to be measured. Combining techniques such as this with an a priori knowledge of species attributes that consistently relate to niche conservatism or shift may be of great help in evaluating the uncertainty of climate-matching predictions made for new spatial or temporal domains. CONCLUSIONS We have identified variable levels of accuracy of native-rangebased climate matching in predicting the worldwide non-native occurrences of European terrestrial gastropods. This variability was related to the level of climatic equilibrium of the species native ranges: species with lower levels of distributional equilibrium had a greater propensity to becoming established in novel climates. We also found that slugs ability to take advantage of microenvironments might increase their likelihood of survival in a wide diversity of macroclimates. Such aptitude may severely reduce the accuracy of coarse-scale climate-matching predictions. Although there is widespread agreement that ecophysiological experiments and genetic approaches are necessary to more precisely predict where a species could become established (Kearney et al., 2010; Hoffmann & Sgrò, 2011), our findings suggest that, in the absence of such approaches, analyses of species traits may provide information about the uncertainty of correlative predictions. We therefore strongly encourage further studies that test the generality of relationships between species traits and accuracy of climate matching. ACKNOWLEDGEMENTS C.C. is supported by a post-doctoral grant from the Portuguese FCT (SFRH/BPD/84422/2012). H.K. received support from the Netherlands Organisation for Scientific Research (NWO ALW ). We thank David Robinson and Kenneth Hayes for providing unpublished species occurrences and Rolanda Matos for sharing information on species distribution ranges. We also acknowledge two anonymous referees for constructive comments on earlier versions of the manuscript. REFERENCES Alexander, J.M. & Edwards, P.J. (2010) Limits to the niche and range margins of alien species. Oikos, 119, AnimalBase Project Group (2012) AnimalBase v University of Göttingen. Available at: (accessed 29 March 2013). Araújo, M.B. & Pearson, R.G. (2005) Equilibrium of species distributions with climate. Ecography, 28, Barve, N., Barve, V., Jiménez-Valverde, A., Lira-Noriega, A., Maher, S.P., Peterson, A.T., Soberón, J. & Villalobos, F. (2011) The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecological Modelling, 222, Broennimann, O., Treier, U.A., Müller-Schärer, H., Thuiller, W., Peterson, A.T. & Guisan, A. (2007) Evidence of climatic niche shift during biological invasion. Ecology Letters, 10, Brown, J.H., Stevens, G.C. & Kaufman, D.M. (1996) The geographic range: size, shape, boundaries, and internal structure. Annual Review of Ecology and Systematics, 27, Capinha, C., Brotons, L. & Anastácio, P. (2013) Geographical variability in propagule pressure and climatic suitability explain the European distribution of two highly invasive crayfish. Journal of Biogeography, 40, Cook, A. (2001) Behavioural ecology: on doing the right thing, in the right place at the right time. The biology of terrestrial molluscs (ed. by G.M. Barker), pp CABI, Wallingford, UK. Cowie, R.H. (2011) Snails and slugs. Encyclopedia of biological invasions. (ed. by D. Simberloff and M. Rejmánek), pp University of California Press, Berkeley, CA. Cowie, R.H., Dillon, R.T., Jr, Robinson, D.G. & Smith, J.W. (2009) Alien non-marine snails and slugs of priority quarantine importance in the United States: a preliminary risk assessment. American Malacological Bulletin, 27, Darriba, D., Taboada, G.L., Doallo, R. & Posada, D. (2012) jmodeltest 2: more models, new heuristics and parallel computing. Nature Methods, 9, Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd
9 Climate-matching and species traits Edgar, R.C. (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research, 32, Elith, J., Kearney, M. & Phillips, S. (2010) The art of modelling range-shifting species. Methods in Ecology and Evolution, 1, Falkner, G., Obrdlík, P., Castella, E. & Speight, M.C.D. (2001) Shelled Gastropoda of Western Europe. Verlag der Friedrich- Held-Gesellschaft, Munich. Fitzpatrick, M.C., Weltzin, J.F., Sanders, N.J. & Dunn, R.R. (2007) The biogeography of prediction error: why does the introduced range of the fire ant over-predict its native range? Global Ecology and Biogeography, 16, Gaston, K.J. (2003) The structure and dynamics of geographic ranges. Oxford University Press, Oxford. Guo, Q., Sax, D.F., Qian, H. & Early, R. (2012) Latitudinal shifts of introduced species: possible causes and implications. Biological Invasions, 14, Hayes, K.R. & Barry, S.C. (2008) Are there any consistent predictors of invasion success? Biological Invasions, 10, Heath, D.J. (1975) Colour, sunlight and internal temperatures in the land-snail Cepaea nemoralis (L.). Oecologia, 19, Heller, J. (1981) Visual versus climatic selection of shell banding in the landsnail Theba pisana in Israel. Journal of Zoology, 194, Heller, J. (2001) Life history strategies. The biology of terrestrial molluscs. (ed. by G.M. Barker), pp CABI, Wallingford, UK. Herbert, D.G. (2010) The introduced terrestrial Mollusca of South Africa. SANBI Biodiversity Series 15. South African National Biodiversity Institute, Pretoria. Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G. & Jarvis, A. (2005) Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, Hoffmann, A.A. & Sgrò, C.M. (2011) Climate change and evolutionary adaptation. Nature, 470, IUCN (2013) IUCN Red List of Threatened Species. Version International Union for Conservation of Nature and Natural Resources, Gland, Switzerland. Available at: Jiménez-Valverde, A., Peterson, A.T., Soberón, J., Overton, J.M., Aragón, P. & Lobo, J. (2011) Use of niche models in invasive species risk assessments. Biological Invasions, 13, Kappes, H. & Schilthuizen, M. (2014) Habitat effects on slug assemblages and introduced species. Journal of Molluscan Studies, 80, Kearney, M.R., Wintle, B.A. & Porter, W.P. (2010) Correlative and mechanistic models of species distribution provide congruent forecasts under climate change. Conservation Letters, 3, Kerney, M.P. (1999) Atlas of the land and freshwater molluscs of Britain and Ireland. Harley Books, Colchester. Kerney, M.P. & Cameron, R.A.D. (2006) Guide des escargots et limaces d Europe. Delachaux et Niestlé, Paris. Lavergne, S. & Molofsky, J. (2007) Increased genetic variation and evolutionary potential drive the success of an invasive grass. Proceedings of the National Academy of Sciences of the United States of America, 104, Luchtel, D.L. & Deyrup-Olsen, I. (2001) Body wall: form and function. The biology of terrestrial molluscs (ed. by G.M. Barker), pp CABI, Wallingford, UK. Owens, H.L., Campbell, L.P., Dornak, L.L., Saupe, E.E., Barve, N., Soberón, J., Ingenloff, K., Lira-Noriega, A., Hensz, C.M., Myers, C.E. & Peterson, A.T. (2013) Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecological Modelling, 263, Paradis, E. & Claude, J. (2002) Analysis of comparative data using generalized estimating equations. Journal of Theoretical Biology, 218, Paradis, E., Claude, J. & Strimmer, K. (2004) APE: analyses of phylogenetics and evolution in R language. Bioinformatics, 20, Pearman, P.B., Guisan, A., Broennimann, O. & Randin, C.F. (2008) Niche dynamics in space and time. Trends in Ecology and Evolution, 23, Petitpierre, B., Kueffer, C., Broennimann, O., Randin, C., Daehler, C. & Guisan, A. (2012) Climatic niche shifts are rare among terrestrial plant invaders. Science, 335, Pyšek, P., Jarošík, V., Pergl, J., Randall, R., Chytrý, M., Kühn, I., Tichý, L., Danihelka, J., Chrtek, J., Jr & Sádlo, J. (2009) The global invasion success of Central European plants is related to distribution characteristics in their native range and species traits. Diversity and Distributions, 15, Rödder, D., Schmidtlein, S., Veith, M. & Lötters, S. (2009) Alien invasive slider turtle in unpredicted habitat: a matter of niche shift or of predictors studied? PLoS ONE, 4, e7843. Sakai, A.K., Allendorf, F.W., Holt, J.S., Lodge, D.M., Molofsky, J., With, K.A., Baughman, S., Cabin, R.J., Cohen, J.E., Ellstrand, N.C., McCauley, D.E., O Neil, P., Parker, I.M., Thompson, J.N. & Weller, S.G. (2001) The population biology of invasive species. Annual Review of Ecology and Systematics, 32, Soberón, J. & Nakamura, M. (2009) Niches and distributional areas: concepts, methods, and assumptions. Proceedings of the National Academy of Sciences of the United States of America, 106, Strubbe, D., Broennimann, O., Chiron, F. & Matthysen, E. (2013) Niche conservatism in non-native birds in Europe: niche unfilling rather than niche expansion. Global Ecology and Biogeography, 22, Suggitt, A.J., Gillingham, P.K., Hill, J.K., Huntley, B., Kunin, W.E., Roy, D.B. & Thomas, C.D. (2011) Habitat microclimates drive fine-scale variation in extreme temperatures. Oikos, 120, 1 8. Svenning, J.-C. & Skov, F. (2007) Could the tree diversity pattern in Europe be generated by postglacial dispersal limitation? Ecology Letters, 10, Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. & Kumar, S. (2011) MEGA5: molecular evolutionary genetics Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd 865
10 C. Capinha et al. analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 28, Thuiller, W., Richardson, D.M., Pyšek, P., Midgley, G.F., Hughes, G.O. & Rouget, M. (2005) Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Global Change Biology, 11, Wade, C.M., Mordan, P.B. & Clarke, B. (2001) A phylogeny of the land snails (Gastropoda: Pulmonata). Proceedings of the Royal Society B: Biological Sciences, 268, Wade, C.M., Mordan, P.B. & Naggs, F. (2006) Evolutionary relationships among the pulmonate land snails and slugs (Pulmonata, Stylommatophora). Biological Journal of the Linnean Society, 87, Webber, B.L., Le Maitre, D.C. & Kriticos, D.J. (2012) Comment on Climatic niche shifts are rare among terrestrial plant invaders. Science, 338, 193. White-McLean, J.A. (2011) Terrestrial mollusc tool. USDA/ APHIS/PPQ Center for Plant Health Science and Technology and the University of Florida. Available at: id/mollusc. SUPPORTING INFORMATION Additional supporting information may be found in the online version of this article at the publisher s web-site. Figure S1 Schematics of the procedure used to classify the types of novel climatic conditions occupied by species in non-native ranges. Appendix S1 Schematic representation of the criteria used for species selection. Appendix S2 Sources of species occurrence data. Appendix S3 Classification of novel climatic conditions colonized. Appendix S4 Climatic conditions occupied by each species in native and non-native ranges. Appendix S5 Alternative sources of data for macrohabitat usage and humidity preferences. Appendix S6 GenBank accession numbers. Appendix S7 Results of the generalized estimating equations. BIOSKETCH César Capinha is a postdoctoral researcher with research interests in invasion ecology and biogeography. He has focused mainly on assessing the relative role of the distinct factors that determine the distribution of non-native species, particularly climate. Author contributions: C.C. led the project and the writing with contributions from the remaining authors. C.C and H.K. performed the data collection. All authors contributed in the interpretation of results. Editor: Joshua Lawler 866 Global Ecology and Biogeography, 23, , 2014 John Wiley & Sons Ltd
11 Global Ecology and Biogeography SUPPORTING INFORMATION Response of non-native European terrestrial gastropods to novel climates correlates with biogeographical and biological traits César Capinha, Dennis Rödder, Henrique Miguel Pereira and Heike Kappes Appendix S1 Schematic representation of the criteria used for species selection. Appendix S2 Sources of species occurrence data. Appendix S3 Classification of novel climatic conditions colonized. Appendix S4 Climatic conditions occupied by species in native and non-native ranges. Appendix S5 Alternative sources of data for macrohabitat usage and humidity preferences. Appendix S6 GenBank accession numbers. Appendix S7 Results of the generalized estimating equations.
12 Appendix S1 Schematic representation of the criteria used for species selection.
13 Appendix S2 Sources of species occurrence data. (a) Online species distribution databases* Analyse Educatie en Marien Oecologisch Onderzoek (ANEMOON): Atlas of Living Australia: Electronic Atlas of the Wildlife of British Columbia (e-fauna BC): biodiversity/efauna/index.shtml Caucasian Land Snails: Global Biodiversity Information Facility (GBIF): Inter-American Biodiversity Information Network (IABIN): Inventaire National du Patrimoine Naturel (INPN): Montana Natural Heritage MapViewer: NatureServe Explorer: Sys Tax: (b) Scientific literature Abbes, I., Liberto, F., Castillejo, J. & Nouira, S. (2010) A review of slugs and semi-slugs of Tunisia (Testacellidae, Milacidae and Limacidae). Journal of Conchology, 40, Al-Khayat, J.A. (2010) First record of five terrestrial snails in the State of Qatar. Turkish Journal of Zoology, 34, Agudo-Padrón, A.I. (2011) Exotic molluscs (Mollusca, Gastropoda et Bivalvia) in Santa Catarina State, Southern Brazil region: check list and regional spatial distribution. Biodiversity Journal, 2, Anderson, F.E. & Coppolino, M.L. (2009) Land snail abundance and diversity with associated ecological variables in six southern Illinois counties. MS Thesis, Southern Illinois University, Carbondale, IL. Arnason, E. & Grant, P.R. (1976) Climatic selection in Cepaea hortensis at the northern limit of its range in Iceland. Evolution, 30, Baker, G.H. (2008) The population dynamics of the mediterranean snails Cernuella virgata, Cochlicella acuta (Hygromiidae) and Theba pisana (Helicidae) in pasture cereal rotations in South Australia: a 20-year study. Australian Journal of Experimental Agriculture, 48, Baker, G.H. (2012) The population dynamics of the mediterranean snail, Cernuella virgata (da Costa, 1778) (Hygromiidae), in continuous-cropping rotations in South Australia. Journal of Molluscan Studies, 78, Barker, G.M. (1999) Naturalized terrestrial Stylommatophora (Mollusca: Gastropoda). Fauna of New Zealand, vol. 38. Manaaki Whenua Press, Lincoln, New Zealand. Baur, B., Schileyko, A.A. & Baur, A. (2000) Ecological observations on Arianta aethiops aethiops (Helicidae), a land snail endemic to the South Carpathian mountains, Romania. Journal of Molluscan Studies, 66, Bößneck, U. (2011) New records of freshwater and land molluscs from Lebanon (Mollusca: Gastropoda & Bivalvia). Zoology in the Middle East, 54, Branson, B.A. (1980) The Recent Gastropoda of Oklahoma, part VIII. The slug families Limacidae, Arionidae, Veronicellidae, and Philomycidae. Proceedings of the Oklahoma Academy of Science, 60,
14 Bronson, C. & Gaskalla, R. (2003) Milk snail survey, Otala lactea, in Nassau County, Florida March Florida Cooperative Agricultural Pest Survey Program, Gainesville, FL. Cádiz, F.J. & Gallardo, C.S. (2007) Arion intermedius (Gastropoda: Stylommatophora); first record of this introduced slug in Chile, with notes on its anatomy and natural history. Revista Chilena de Historia Natural, 80, Campos, J. & Calvo, A. (2006) Moluscos introducidos en Uruguay. Comunicaciones de la Sociedad Malacológica del Uruguay, 9, Castillejo, J. (1986) Caracoles terrestres de Galicia. Familia Helicidae (Gastropoda: Pulmonata). Universidad de Santiago de Compostela, Santiago de Compostela, Spain. Chambers, B. (2008) Port of Tacoma Cernuella virgata (C. virgata) eradication program in Pierce County, Washington. Environmental Assessment June United States Department of Agriculture, Washington, DC. Cianfanelli, S. & Lori, E. (2007) La malacofauna del comprensorio delle Tre Limentre Reno. Museo di Storia Naturale dell Università degli Studi di Firenze, Florence, Italy. Cowie, R.H. (1990) Climatic selection on body colour in the land snail Theba pisana (Pulmonata: Helicidae). Heredity, 65, Däumer, C., Greve, C., Hutterer, R., Misof, B. & Haase, M. (2012) Phylogeography of an invasive land snail: natural range expansion versus anthropogenic dispersal in Theba pisana pisana. Biological Invasions, 14, De Francesco, C.G. & Lagiglia, H. (2007) A predatory land snail invades central-western Argentina. Biological Invasions, 9, Desouky, M.M.A. & Busais, S. (2012) Phylogenetic relationships of the land snail; Eobania vermiculata (Müller, 1774) from Egypt and Saudi Arabia. A combined morphological and molecular analysis. Journal of Basic and Applied Zoology, 65, Dvořák, L., Čejka, T. & Horsák, M. (2003) Present knowledge of distribution of Tandonia budapestensis (Hazay, 1881) in the Czech and Slovak Republics (Gastropoda: Milacidae). Malakológiai Tájékoztató, 21, Dvořáková, J., Ložek, V., Horsák, M. & Pechanec, V. (2011) Atlas rozšíření suchozemských plžů v CHKO Bílé Karpaty. Acta Carpathica Occidentalis Supplementum 1, Vsetín, Czech Republic. Fechter, R. & Falkner, G. (1989) Weichtiere. Die farbigen Naturführer. Mosaik-Verlag, Munich. Ferreri, D., Bodon, M. & Manganelli, G. (2005) Molluschi terrestri della provincia di Lecce. Thalassia Salentina, 28, Fiorentino, V., Salomone, N., Manganelli, G. & Giusti, F. (2008) Phylogeography and morphological variability in land snails: the Sicilian Marmorana (Pulmonata, Helicidae). Biological Journal of the Linnean Society, 94, Frank, C. (2000) Mollusca (Gastropoda et Bivalvia): eine historische Sammlung aus Hollabrunn (Niederösterreich). Wissenschaftliche Mitteilungen aus dem Niederösterreichischen Landesmuseum, 13, Gargominy, O. (2011) Marmorana serpentina. IUCN Red List of Threatened Species, Version International Union for the Conservation of Nature and Natural Resources, Gland, Switzerland. Available at: Gill, R. (1996) Xerotricha conspurcata. California Plant Pest and Disease Report, 15, Girod, A. (2002) La malacofauna del Bronzo antico al Riparo del Santuario di Lasino (Trento) - Scavi Annali del Museo Civico di Rovereto, 18, Górka, M. (2005) The invasion continues a new locality of Monacha cartusiana (O. F. Müller, 1774) (Gastropoda: Pulmonata: Helicidae) in the Świętokrzyskie Mts (Central Poland). Folia Malacologica, 13,
15 Guiller, A. & Madec, L. (2010) Historical biogeography of the land snail Cornu aspersum: a new scenario inferred from haplotype distribution in the Western Mediterranean basin. BMC Evolutionary Biology, 10, 18. Guiller, A., Martin, M.-C., Hiraux, C. & Madec, L. (2012) Tracing the invasion of the mediterranean land snail Cornu aspersum aspersum becoming an agricultural and garden pest in areas recently introduced. PLoS ONE, 7, e Haase, M. & Bisenberger, A. (2003) Allozymic differentiation in the land snail Arianta arbustorum (Stylommatophora, Helicidae): historical inferences. Journal of Zoological Systematics and Evolutionary Research, 41, Haase, M. & Misof, B. (2009) Dynamic gastropods: stable shell polymorphism despite gene flow in the land snail Arianta arbustorum. Journal of Zoological Systematics and Evolutionary Research, 47, Hanna, D.G. (1966) Introduced mollusks of western North America. Occasional Papers of the California Academia of Sciences, 48, Hayes, K.A., Yeung, N.W., Kim, J.R. & Cowie, R.H. (2012) New records of alien Gastropoda in the Hawaiian Islands: Bishop Museum Occasional Papers, 112, Henk, K.M. & Rittner, O. (2010) On the presence of Helix lucorum Linnaeus, 1758 (Mollusca, Gastropoda, Helicidae) in Le Vesinet, a western suburb of Paris. MalaCo, 6, Herbert, D.G. (2010) The introduced terrestrial Mollusca of South Africa. SANBI Biodiversity Series 15. South African National Biodiversity Institute, Pretoria. Holland, K.D., McDonnell, M.J. & Williams, N.S.G. (2007) Abundance, species richness and feeding preferences of introduced molluscs in native grasslands of Victoria, Australia. Austral Ecology, 32, Hotopp, K. & Pearce, T.A. (2007) Land snails in New York: statewide distribution and talus site faunas. Final Report for contract #NYHER submitted to New York State Biodiversity Research Institute. New York State Museum, New York. Kantor, I.Y., Schileyko, A.A., Vinarski, M.V. & Sysoev, A.V. (2009) Catalogue of the continental mollusks of Russia and adjacent countries. Moscow State University, Russia. Available at: Kebapçı, Ü., Yıldırım, M.Z., Gülle, İ., Öztop, M. & Çağlan, D.C. (2012) The land snail fauna of Mut District (Mersin Province, Turkey). Turkish Journal of Zoology, 36, Kerney, M.P. (1999) Atlas of the land and freshwater molluscs of Britain and Ireland. Harley Books, Colchester. Kierdorf-Traut, G. (2012) Notizen zur Verbreitung von Chilostoma cingulatum peregrini Falkner, 1998 in Nordtirol (Gastropoda: Helicidae). Gredleriana, 12, Kiss, Y. (2008) Die Weichtierfauna (Mollusca: Gastropoda, Bivalvia) des Schlerngebietes (Südtirol, Italien). Gredleriana, 8, Lesicki, A. & Koralewska-Batura, E. (2007) Monacha cartusiana (O. F. Müller, 1774) (Gastropoda: Pulmonata: Hygromiidae) becomes more frequent in Poland. Folia Malacologica, 15, Madec, L., Bellido, A. & Guiller, A. (2003) Shell shape of the land snail Cornu aspersum in North Africa: unexpected evidence of a phylogeographical splitting. Heredity, 91, Martín, S.M., César, I.I. & Liberto, R. (2009) Distribution of Deroceras reticulatum (Müller, 1774) (Pulmonata Stylommatophora) in Argentina with first record of the Reserva de Usos Múltiples Isla Martin Garcia, Río de la Plata superior. Brazilian Journal of Biology, 69,
16 McAlpine, D.F., Schueler, F.W., Maunder, J.E., Noseworthy, R.G. & Sollows, M.C. (2009) Establishment and persistence of the copse snail, Arianta arbustorum (Linnaeus, 1758) (Gastropoda: Helicidae) in Canada. The Nautilus, 123, Mc Donnell, R.J., Paine T.D. & Gormally, M.J. (2009) Slugs: a guide to the invasive and native fauna of California. University of California Press, Oakland, CA. Mc Donnell, R.J., Paine, T.D., Stouthamer, R., Gormally, M.J. & Harwood, J.D. (2008) Molecular and morphological evidence for the occurrence of two new species of invasive slugs in Kentucky, Arion intermedius Normand and Arion hortensis Férussac (Arionidae: Stylommatophora). Journal of the Kentucky Academy of Science, 69, Michaelidis, B., Pallidou, A. & Vakouftsi, P. (1999) Effects of anoxia on the extra- and intracellular acid base status in the land snail Helix lucorum (L.): lack of evidence for a relationship between pyruvate kinase down-regulation and acid base status. Journal of Experimental Biology, 202, Michalak, P.S. (2010) New pest response guidelines: temperate terrestrial gastropods. United States Department of Agriculture, Washington, DC. Míkovcová, A., Juřičková, L. & Petrusek, A. (2007) Ekologie invazního druhu Monacha cartusiana. Charles University, Prague. Available at: Noma, T., Colunga-Garcia, M., Brewer, M., Landis, J., Gooch, A. & Philip, M. (2010) Carthusian snail Monacha cartusiana. Michigan State University s invasive species factsheets. Michigan State University, Lansing, MI. Novák, J. & Novák, M. (2009) Dvě nové lokality tmavoretky bělavé Monacha cartusiana (O.F. Müller, 1774) na Moravě. Malacologica Bohemoslovaca, 8, Örstan, A. (2003) The first record of Discus rotundatus from Turkey. Triton, 7, 27. Örstan, A. (2010) Gastropoda, Pulmonata, Helicidae, Cepaea nemoralis (Linnaeus, 1758): new records for Montreal, Canada. Check List, 6, Örstan, A., Sparks, J.L., Jr & Pearce, T.A. (2011) Wayne Grimm s legacy: a 40-year experiment on the dispersal of Cepaea nemoralis in Frederick County, Maryland. American Malacological Bulletin, 29, Ożgo, M. (2012) Shell polymorphism in the land-snail Cepaea nemoralis (L.) along a West East transect in continental Europe. Folia Malacologica, 20, Pech, P. & Pechová, H. (2009) Monacha cartusiana (Gastropoda: Hygromiidae) in South Bohemia. Malacologica Bohemoslovaca, 8, 28. Pelbárt, J. (2000) Data to the mollusc fauna of the Carpathian Biosphere Reserve, Ukraine (Mollusca). Miscellanea Zoologica Hungarica, 13, Peltanová, A., Dvořák, L. & Juřičková, L. (2012) The spread of non-native Cepaea nemoralis and Monacha cartusiana (Gastropoda: Pulmonata) in the Czech Republic with comments on other land snail immigrants. Biologia, 67, Peltanová, A., Petrusek, A., Kment, P. & Juřičková, L. (2012) A fast snail s pace: colonization of Central Europe by Mediterranean gastropods. Biological Invasions, 14, Quiñonero Salgado, S., López Alabau, A. & García Meseguer, A.J. (2010) Nuevas localidades de Helix lucorum (Linnaeus, 1758) para la península Ibérica. Spira, 3, Reise, H., Hutchinson, J.M.C. & Robinson, D.G. (2006) Two introduced pest slugs: Tandonia budapestensis new to the Americas, and Deroceras panormitanum new to the Eastern USA. The Veliger, 48, Richards, A.V. & Murray, J. (1975) The relation of phenotype to habitat in an introduced colony of Cepaea nemoralis. Heredity, 34,
17 Říhová, D. & Juřičková, L. (2011) The Girdled Snail Hygromia cinctella (Draparnaud, 1801) new to the Czech Republic. Malacologica Bohemoslovaca, 10, Rosenbauer, A. (2011) Vorkommen südeuropäischer Schneckenarten in Steinmetzbetrieben. Mitteilungen der Deutschen Malakozoologischen Gesellschaft, 85, Selander, R.K. & Foltz, D.W. (1981) Gametic disequilibrium between esterase loci in populations of Cepaea nemoralis in western New York. Evolution, 35, Steinke, D., Albrecht, C. & Pfenninger, M. (2004) Molecular phylogeny and character evolution in the Western Palaearctic Helicidae s.l. (Gastropoda: Stylommatophora). Molecular Phylogenetics and Evolution, 32, Ueshima, R., Okamoto, M. & Saito, Y. (2004) Eobania vermiculata, a land snail newly introduced into Japan. Chiribotan, 35, Vardinoyannis, K., Demetropoulos, S., Mylonas, M., Triantis, K.A., Makris, C., Georgiou, G., Wiktor, A. & Demetropoulos, A. (2012) Terrestrial slugs (Gastropoda, Pulmonata) in the NATURA 2000 areas of Cyprus island. ZooKeys, 174, Whitson, M. (2005) Cepaea nemoralis (Gastropoda, Helicidae): the invited invader. Journal of the Kentucky Academy of Science, 66, Yıldırım, M.Z. & Kebapçı, Ü. (2004) Slugs (Gastropoda: Pulmonata) of the Lakes Region (Göller Bölgesi) in Turkey. Turkish Journal of Zoology, 28, (c) Non-scientific internet websites* 1. htm 2. (d) Unpublished data C.C., unpubl. data Iberian Peninsula H.K., unpubl. data continental Europe David Robinson (USDA/APHIS/PPQ, Philadelphia, PA, USA), unpubl. data continental USA Kenneth A. Hayes (Howard University, Washington, DC, USA), unpubl. data Hawaii *All online data sources were consulted between 8 October 2012 and 29 March 2013.
18 Appendix S3 Classification of novel climatic conditions colonized. We distinguished four types of novel climates based on the correlation of the climatic variables in the plane defined by the two main axes of a PCA (see main manuscript). For that purpose we initially plotted the loadings of all variables in ARCGIS using a Cartesian coordinate plane (Fig. S1a, below). Because the two axes accounted for about 89% of total variance, the loading vectors in this plane approximate well the correlation among variables. We performed the division of climatic conditions based on the loading vectors of annual mean temperature (amt) and annual precipitation (anpre). This is because the orientation of their vectors is central to that of their counterpart variables to which they are closely associated and both are also highly discriminating in the twodimensional plane (i.e. have long vectors). Any condition in the climatic space is positively related with a variable in the two-dimensional plane if the vector connecting it to the origin and the vector representing the variable form an angle less than 90, whereas angles greater than 90 indicate a negative relationship. Based on this, we traced the semi-planes perpendicular to the orientation of the vectors of annual mean temperature and annual precipitation (Fig. S1bc). This was made in ARCGIS using the COGO extension. At the intersection of these semi-planes we obtained four sectors, each representing a distinct climatic context: cold and dry; cold and wet; warm and dry and warm and wet (Fig. S1d). For each species we then used the Spatial Adjust tool to centre the origin of the sectors on the centroid of the native-based convex hull (see main manuscript; Fig. S1e). Finally, we classified the type of novel climate encompassed by the species (i.e. non-native occurrences falling outside the native-based convex hull) based on the climatic sectors it overlaid. Figure S1e illustrates the convex hull of native conditions (in grey) and three non-native species occurrences (triangles) established under warmer and drier conditions.
19 Figure S1 Schematics of the procedure used to classify the types of novel climatic conditions occupied by species in non-native ranges (see details above).


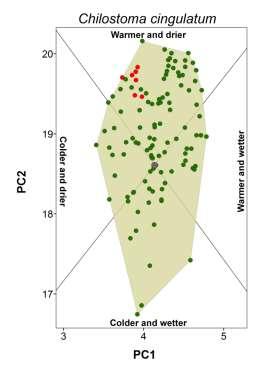
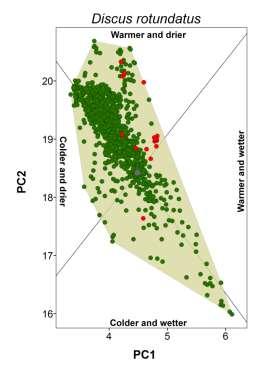
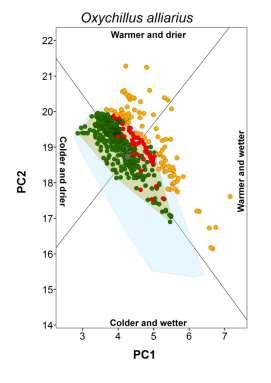
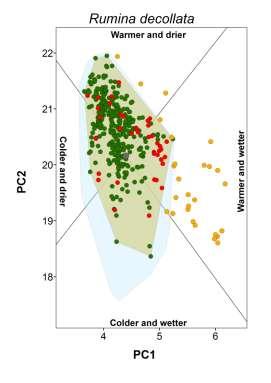
20 Appendix S4 Climatic conditions occupied by each species in native and non-native ranges. The plot for the brown garden snail (Cornu aspersum) is shown in the main document.
21
22
23
24
25
26
Spatial Effects on Current and Future Climate of Ipomopsis aggregata Populations in Colorado Patterns of Precipitation and Maximum Temperature
 A. Kenney GIS Project Spring 2010 Amanda Kenney GEO 386 Spring 2010 Spatial Effects on Current and Future Climate of Ipomopsis aggregata Populations in Colorado Patterns of Precipitation and Maximum Temperature
A. Kenney GIS Project Spring 2010 Amanda Kenney GEO 386 Spring 2010 Spatial Effects on Current and Future Climate of Ipomopsis aggregata Populations in Colorado Patterns of Precipitation and Maximum Temperature
Welcome! Text: Community Ecology by Peter J. Morin, Blackwell Science ISBN (required) Topics covered: Date Topic Reading
 Topics covered: Date Topic Reading") Welcome! Text: Community Ecology by Peter J. Morin, Blackwell Science ISBN 0-86542-350-4 (required) Topics covered: Date Topic Reading 1 Sept Syllabus, project, Ch1, Ch2 Communities 8 Sept Competition
Welcome! Text: Community Ecology by Peter J. Morin, Blackwell Science ISBN 0-86542-350-4 (required) Topics covered: Date Topic Reading 1 Sept Syllabus, project, Ch1, Ch2 Communities 8 Sept Competition
Species Distribution Models
 Species Distribution Models Whitney Preisser ESSM 689 Quantitative Methods in Ecology, Evolution, and Biogeography Overview What are SDMs? What are they used for? Assumptions and Limitations Data Types
Species Distribution Models Whitney Preisser ESSM 689 Quantitative Methods in Ecology, Evolution, and Biogeography Overview What are SDMs? What are they used for? Assumptions and Limitations Data Types
Georgia Performance Standards for Urban Watch Restoration Field Trips
 Georgia Performance Standards for Field Trips 6 th grade S6E3. Students will recognize the significant role of water in earth processes. a. Explain that a large portion of the Earth s surface is water,
Georgia Performance Standards for Field Trips 6 th grade S6E3. Students will recognize the significant role of water in earth processes. a. Explain that a large portion of the Earth s surface is water,
"PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200 Spring 2014 University of California, Berkeley
 "PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200 Spring 2014 University of California, Berkeley D.D. Ackerly April 16, 2014. Community Ecology and Phylogenetics Readings: Cavender-Bares,
"PRINCIPLES OF PHYLOGENETICS: ECOLOGY AND EVOLUTION" Integrative Biology 200 Spring 2014 University of California, Berkeley D.D. Ackerly April 16, 2014. Community Ecology and Phylogenetics Readings: Cavender-Bares,
What is the range of a taxon? A scaling problem at three levels: Spa9al scale Phylogene9c depth Time
 What is the range of a taxon? A scaling problem at three levels: Spa9al scale Phylogene9c depth Time 1 5 0.25 0.15 5 0.05 0.05 0.10 2 0.10 0.10 0.20 4 Reminder of what a range-weighted tree is Actual Tree
What is the range of a taxon? A scaling problem at three levels: Spa9al scale Phylogene9c depth Time 1 5 0.25 0.15 5 0.05 0.05 0.10 2 0.10 0.10 0.20 4 Reminder of what a range-weighted tree is Actual Tree
SIF_7.1_v2. Indicator. Measurement. What should the measurement tell us?
 Indicator 7 Area of natural and semi-natural habitat Measurement 7.1 Area of natural and semi-natural habitat What should the measurement tell us? Natural habitats are considered the land and water areas
Indicator 7 Area of natural and semi-natural habitat Measurement 7.1 Area of natural and semi-natural habitat What should the measurement tell us? Natural habitats are considered the land and water areas
Rainfall variability and uncertainty in water resource assessments in South Africa
 New Approaches to Hydrological Prediction in Data-sparse Regions (Proc. of Symposium HS.2 at the Joint IAHS & IAH Convention, Hyderabad, India, September 2009). IAHS Publ. 333, 2009. 287 Rainfall variability
New Approaches to Hydrological Prediction in Data-sparse Regions (Proc. of Symposium HS.2 at the Joint IAHS & IAH Convention, Hyderabad, India, September 2009). IAHS Publ. 333, 2009. 287 Rainfall variability
Chapter 8. Biogeographic Processes. Upon completion of this chapter the student will be able to:
 Chapter 8 Biogeographic Processes Chapter Objectives Upon completion of this chapter the student will be able to: 1. Define the terms ecosystem, habitat, ecological niche, and community. 2. Outline how
Chapter 8 Biogeographic Processes Chapter Objectives Upon completion of this chapter the student will be able to: 1. Define the terms ecosystem, habitat, ecological niche, and community. 2. Outline how
Changing Planet: Changing Mosquito Genes
 Changing Planet: Changing Mosquito Genes Name Background As the climate changes around the globe, organisms will need to adapt in order to survive. But what does it mean to adapt? When you put on a sweater
Changing Planet: Changing Mosquito Genes Name Background As the climate changes around the globe, organisms will need to adapt in order to survive. But what does it mean to adapt? When you put on a sweater
Eichhornia crassipes (water hyacinth) Tristylous, clonal
 Tristylous, clonal") Plant of the Day Eichhornia crassipes (water hyacinth) Native to South America Tristylous, clonal Invasive in Asia, Africa, North America, Australia Clogs waterways, blocks sunlight and reduces oxygen
Plant of the Day Eichhornia crassipes (water hyacinth) Native to South America Tristylous, clonal Invasive in Asia, Africa, North America, Australia Clogs waterways, blocks sunlight and reduces oxygen
AP Environmental Science I. Unit 1-2: Biodiversity & Evolution
 NOTE/STUDY GUIDE: Unit 1-2, Biodiversity & Evolution AP Environmental Science I, Mr. Doc Miller, M.Ed. North Central High School Name: ID#: NORTH CENTRAL HIGH SCHOOL NOTE & STUDY GUIDE AP Environmental
NOTE/STUDY GUIDE: Unit 1-2, Biodiversity & Evolution AP Environmental Science I, Mr. Doc Miller, M.Ed. North Central High School Name: ID#: NORTH CENTRAL HIGH SCHOOL NOTE & STUDY GUIDE AP Environmental
Energy Systems, Structures and Processes Essential Standard: Analyze patterns of global climate change over time Learning Objective: Differentiate
 Energy Systems, Structures and Processes Essential Standard: Analyze patterns of global climate change over time Learning Objective: Differentiate between weather and climate Global Climate Focus Question
Energy Systems, Structures and Processes Essential Standard: Analyze patterns of global climate change over time Learning Objective: Differentiate between weather and climate Global Climate Focus Question
Metacommunities Spatial Ecology of Communities
 Spatial Ecology of Communities Four perspectives for multiple species Patch dynamics principles of metapopulation models (patchy pops, Levins) Mass effects principles of source-sink and rescue effects
Spatial Ecology of Communities Four perspectives for multiple species Patch dynamics principles of metapopulation models (patchy pops, Levins) Mass effects principles of source-sink and rescue effects
Ryan P. Shadbolt * Central Michigan University, Mt. Pleasant, Michigan
 14A.1 RECENT CLIMATE CHANGE IN THE HIGH ELEVATIONS OF THE SOUTHERN APPALACHIANS Ryan P. Shadbolt * Central Michigan University, Mt. Pleasant, Michigan 1. INTRODUCTION Island species are often vulnerable
14A.1 RECENT CLIMATE CHANGE IN THE HIGH ELEVATIONS OF THE SOUTHERN APPALACHIANS Ryan P. Shadbolt * Central Michigan University, Mt. Pleasant, Michigan 1. INTRODUCTION Island species are often vulnerable
Chapter 6 Lecture. Life History Strategies. Spring 2013
 Chapter 6 Lecture Life History Strategies Spring 2013 6.1 Introduction: Diversity of Life History Strategies Variation in breeding strategies, fecundity, and probability of survival at different stages
Chapter 6 Lecture Life History Strategies Spring 2013 6.1 Introduction: Diversity of Life History Strategies Variation in breeding strategies, fecundity, and probability of survival at different stages
Marine Resources Development Foundation/MarineLab Grades: 9, 10, 11, 12 States: AP Biology Course Description Subjects: Science
 Marine Resources Development Foundation/MarineLab Grades: 9, 10, 11, 12 States: AP Biology Course Description Subjects: Science Highlighted components are included in Tallahassee Museum s 2016 program
Marine Resources Development Foundation/MarineLab Grades: 9, 10, 11, 12 States: AP Biology Course Description Subjects: Science Highlighted components are included in Tallahassee Museum s 2016 program
Introduction to Biosystematics. Course Website: Lecture 1: Introduction to Biological Systematics Outline: The role and value of Systematics
 Introduction to Biosystematics Course Website: http://homepages.ucalgary.ca/~dsikes/courses.htm Check weekly for lecture updates, readings, etc. D. S. Sikes University of Calgary There have been many authorities
Introduction to Biosystematics Course Website: http://homepages.ucalgary.ca/~dsikes/courses.htm Check weekly for lecture updates, readings, etc. D. S. Sikes University of Calgary There have been many authorities
American Harris mud crab Rhithropanopeus harrisii
 American Harris mud crab Rhithropanopeus harrisii (Gould, 1841) in the Gulf of Gdańsk (southern Baltic Sea): distribution, population structure and basic physiological processes Joanna Hegele-Drywa Alien
American Harris mud crab Rhithropanopeus harrisii (Gould, 1841) in the Gulf of Gdańsk (southern Baltic Sea): distribution, population structure and basic physiological processes Joanna Hegele-Drywa Alien
CHAPTER 1: INTRODUCTION
 CHAPTER 1: INTRODUCTION There is now unequivocal evidence from direct observations of a warming of the climate system (IPCC, 2007). Despite remaining uncertainties, it is now clear that the upward trend
CHAPTER 1: INTRODUCTION There is now unequivocal evidence from direct observations of a warming of the climate system (IPCC, 2007). Despite remaining uncertainties, it is now clear that the upward trend
Kristina Enciso. Brian Leung. McGill University Quebec, Canada
 Embracing uncertainty to incorporate biotic interactions into species distribution modeling: creating community assemblages using interactive community distribution models Kristina Enciso Brian Leung McGill
Embracing uncertainty to incorporate biotic interactions into species distribution modeling: creating community assemblages using interactive community distribution models Kristina Enciso Brian Leung McGill
Climate Dynamics (PCC 587): Hydrologic Cycle and Global Warming
: Hydrologic Cycle and Global Warming") Climate Dynamics (PCC 587): Hydrologic Cycle and Global Warming D A R G A N M. W. F R I E R S O N U N I V E R S I T Y O F W A S H I N G T O N, D E P A R T M E N T O F A T M O S P H E R I C S C I E N C
Climate Dynamics (PCC 587): Hydrologic Cycle and Global Warming D A R G A N M. W. F R I E R S O N U N I V E R S I T Y O F W A S H I N G T O N, D E P A R T M E N T O F A T M O S P H E R I C S C I E N C
Chapter 5 Evolution of Biodiversity. Sunday, October 1, 17
 Chapter 5 Evolution of Biodiversity CHAPTER INTRO: The Dung of the Devil Read and Answer Questions Provided Module 14 The Biodiversity of Earth After reading this module you should be able to understand
Chapter 5 Evolution of Biodiversity CHAPTER INTRO: The Dung of the Devil Read and Answer Questions Provided Module 14 The Biodiversity of Earth After reading this module you should be able to understand
How Can Evolution Explain the Frequency of White-Striped Clovers? *Adapted from National Center for Case Study Teaching in Science
 How Can Evolution Explain the Frequency of White-Striped Clovers? *Adapted from National Center for Case Study Teaching in Science Learning Objectives Understand the process of natural selection Acquire
How Can Evolution Explain the Frequency of White-Striped Clovers? *Adapted from National Center for Case Study Teaching in Science Learning Objectives Understand the process of natural selection Acquire
The Environmental Classification of Europe, a new tool for European landscape ecologists
 The Environmental Classification of Europe, a new tool for European landscape ecologists De Environmental Classification of Europe, een nieuw gereedschap voor Europese landschapsecologen Marc Metzger Together
The Environmental Classification of Europe, a new tool for European landscape ecologists De Environmental Classification of Europe, een nieuw gereedschap voor Europese landschapsecologen Marc Metzger Together
Jean-Marc Dufour-dror
 Israel Nature and Parks Authority Science & Conservation Division The Jerusalem Institute for Israel Studies The Center for Environmental Policy Invasive plant species in Israel s natural areas: Distribution,
Israel Nature and Parks Authority Science & Conservation Division The Jerusalem Institute for Israel Studies The Center for Environmental Policy Invasive plant species in Israel s natural areas: Distribution,
Part I History and ecological basis of species distribution modeling
 Part I History and ecological basis of species distribution modeling Recent decades have seen an explosion of interest in species distribution modeling. This has resulted from a confluence of the growing
Part I History and ecological basis of species distribution modeling Recent decades have seen an explosion of interest in species distribution modeling. This has resulted from a confluence of the growing
Data Dictionary for Network of Conservation Areas Transcription Reports from the Colorado Natural Heritage Program
 Data Dictionary for Network of Conservation Areas Transcription Reports from the Colorado Natural Heritage Program This Data Dictionary defines terms used in Network of Conservation Areas (NCA) Reports
Data Dictionary for Network of Conservation Areas Transcription Reports from the Colorado Natural Heritage Program This Data Dictionary defines terms used in Network of Conservation Areas (NCA) Reports
ENMTools: a toolbox for comparative studies of environmental niche models
 Ecography 33: 607611, 2010 doi: 10.1111/j.1600-0587.2009.06142.x # 2010 The Authors. Journal compilation # 2010 Ecography Subject Editor: Thiago Rangel. Accepted 4 September 2009 ENMTools: a toolbox for
Ecography 33: 607611, 2010 doi: 10.1111/j.1600-0587.2009.06142.x # 2010 The Authors. Journal compilation # 2010 Ecography Subject Editor: Thiago Rangel. Accepted 4 September 2009 ENMTools: a toolbox for
Statistical Forecast of the 2001 Western Wildfire Season Using Principal Components Regression. Experimental Long-Lead Forecast Bulletin
 Statistical Forecast of the 2001 Western Wildfire Season Using Principal Components Regression contributed by Anthony L. Westerling 1, Daniel R. Cayan 1,2, Alexander Gershunov 1, Michael D. Dettinger 2
Statistical Forecast of the 2001 Western Wildfire Season Using Principal Components Regression contributed by Anthony L. Westerling 1, Daniel R. Cayan 1,2, Alexander Gershunov 1, Michael D. Dettinger 2
Chapter 22: Descent with Modification 1. BRIEFLY summarize the main points that Darwin made in The Origin of Species.
 AP Biology Chapter Packet 7- Evolution Name Chapter 22: Descent with Modification 1. BRIEFLY summarize the main points that Darwin made in The Origin of Species. 2. Define the following terms: a. Natural
AP Biology Chapter Packet 7- Evolution Name Chapter 22: Descent with Modification 1. BRIEFLY summarize the main points that Darwin made in The Origin of Species. 2. Define the following terms: a. Natural
Open projects for BSc & MSc
 Next Generation Sequencing New sequencing technologies enable biologists to obtain complete genome and New sequencing technologies enable biologists to obtain complete transcriptome data of non-model organisms.
Next Generation Sequencing New sequencing technologies enable biologists to obtain complete genome and New sequencing technologies enable biologists to obtain complete transcriptome data of non-model organisms.
Monthly Long Range Weather Commentary Issued: APRIL 18, 2017 Steven A. Root, CCM, Chief Analytics Officer, Sr. VP,
 Monthly Long Range Weather Commentary Issued: APRIL 18, 2017 Steven A. Root, CCM, Chief Analytics Officer, Sr. VP, sroot@weatherbank.com MARCH 2017 Climate Highlights The Month in Review The average contiguous
Monthly Long Range Weather Commentary Issued: APRIL 18, 2017 Steven A. Root, CCM, Chief Analytics Officer, Sr. VP, sroot@weatherbank.com MARCH 2017 Climate Highlights The Month in Review The average contiguous
PREDICTING DROUGHT VULNERABILITY IN THE MEDITERRANEAN
 J.7 PREDICTING DROUGHT VULNERABILITY IN THE MEDITERRANEAN J. P. Palutikof and T. Holt Climatic Research Unit, University of East Anglia, Norwich, UK. INTRODUCTION Mediterranean water resources are under
J.7 PREDICTING DROUGHT VULNERABILITY IN THE MEDITERRANEAN J. P. Palutikof and T. Holt Climatic Research Unit, University of East Anglia, Norwich, UK. INTRODUCTION Mediterranean water resources are under
Phylogenetic diversity and conservation
 Phylogenetic diversity and conservation Dan Faith The Australian Museum Applied ecology and human dimensions in biological conservation Biota Program/ FAPESP Nov. 9-10, 2009 BioGENESIS Providing an evolutionary
Phylogenetic diversity and conservation Dan Faith The Australian Museum Applied ecology and human dimensions in biological conservation Biota Program/ FAPESP Nov. 9-10, 2009 BioGENESIS Providing an evolutionary
What is the IPCC? Intergovernmental Panel on Climate Change
 IPCC WG1 FAQ What is the IPCC? Intergovernmental Panel on Climate Change The IPCC is a scientific intergovernmental body set up by the World Meteorological Organization (WMO) and by the United Nations
IPCC WG1 FAQ What is the IPCC? Intergovernmental Panel on Climate Change The IPCC is a scientific intergovernmental body set up by the World Meteorological Organization (WMO) and by the United Nations
Chapter 5. Evolution of Biodiversity
 Chapter 5. Evolution of Biodiversity I. Earth s tremendous diversity A. life comes in many forms B. Recall 1. we can think of biodiversity in three ways a) genetic diversity b) species diversity c) ecosystem
Chapter 5. Evolution of Biodiversity I. Earth s tremendous diversity A. life comes in many forms B. Recall 1. we can think of biodiversity in three ways a) genetic diversity b) species diversity c) ecosystem
DEPARTMENT OF ANIMAL AND PLANT SCIENCES Autumn Semester ANIMAL POPULATION & COMMUNITY ECOLOGY
 APS208 DEPARTMENT OF ANIMAL AND PLANT SCIENCES Autumn Semester 2006-2007 ANIMAL POPULATION & COMMUNITY ECOLOGY Your answers should include named examples, and diagrams where appropriate. Answer TWO questions.
APS208 DEPARTMENT OF ANIMAL AND PLANT SCIENCES Autumn Semester 2006-2007 ANIMAL POPULATION & COMMUNITY ECOLOGY Your answers should include named examples, and diagrams where appropriate. Answer TWO questions.
Using Predictive Modelling to Aid Planning in the Mineral Exploration and Mining Sector A Case Study using the Powelliphanta Land Snail
 Using Predictive Modelling to Aid Planning in the Mineral Exploration and Mining Sector A Case Study using the Powelliphanta Land Snail C. Wildman 1 and K. Peters 2 1 Kenex Limited, PO Box 41136, Eastbourne,
Using Predictive Modelling to Aid Planning in the Mineral Exploration and Mining Sector A Case Study using the Powelliphanta Land Snail C. Wildman 1 and K. Peters 2 1 Kenex Limited, PO Box 41136, Eastbourne,
thesis_kokshoorn:thesis :47 Pagina 65 5 the sierra del Cadí
 5 the sierra del Cadí thesis_kokshoorn:thesis 31-10-2008 9:48 Pagina 67 the sierra del cadí in the chapters 6, 7 and 8 new data on the land snail species Abida secale (draparnaud, 1801) are presented.
5 the sierra del Cadí thesis_kokshoorn:thesis 31-10-2008 9:48 Pagina 67 the sierra del cadí in the chapters 6, 7 and 8 new data on the land snail species Abida secale (draparnaud, 1801) are presented.
Eric R. Larson 1 *, Rachael V. Gallagher 2, Linda J. Beaumont 2 and Julian D. Olden 3
 Diversity and Distributions, (Diversity Distrib.) (2014) 20, 1296 1306 A Journal of Conservation Biogeography Diversity and Distributions BIODIVERSITY RESEARCH 1 Department of Ecology and Evolutionary
Diversity and Distributions, (Diversity Distrib.) (2014) 20, 1296 1306 A Journal of Conservation Biogeography Diversity and Distributions BIODIVERSITY RESEARCH 1 Department of Ecology and Evolutionary
Taxonomy: Classification of slugs and snails
 Hawaii Island Rat Lungworm Working Group Daniel K. Inouye College of Pharmacy University of Hawaii, Hilo Rat Lungworm IPM RLWL-4 Taxonomy: Classification of slugs and snails Standards addressed: Next Generation
Hawaii Island Rat Lungworm Working Group Daniel K. Inouye College of Pharmacy University of Hawaii, Hilo Rat Lungworm IPM RLWL-4 Taxonomy: Classification of slugs and snails Standards addressed: Next Generation
Some preliminary results from Climex and Maxent distribution modelling of Drosophila suzukii. Version 2.
 Some preliminary results from Climex and Maxent distribution modelling of Drosophila suzukii. Version 2. Changes from Version 1: Oregon locations where D. suzukii is found now georeferenced. Maxent model
Some preliminary results from Climex and Maxent distribution modelling of Drosophila suzukii. Version 2. Changes from Version 1: Oregon locations where D. suzukii is found now georeferenced. Maxent model
СПИСАНИЯ с Импакт фактор за 2016 година в областта на екологията
 Rank СПИСАНИЯ с Импакт фактор за 2016 година в областта на екологията Full Journal Title Total Cites Journal Impact Factor Eigenfactor Score 1 Trends in ecology & evolution 32,332 15.268 0.039290 2 Annual
Rank СПИСАНИЯ с Импакт фактор за 2016 година в областта на екологията Full Journal Title Total Cites Journal Impact Factor Eigenfactor Score 1 Trends in ecology & evolution 32,332 15.268 0.039290 2 Annual
Chapter 52 An Introduction to Ecology and the Biosphere
 Chapter 52 An Introduction to Ecology and the Biosphere Ecology The study of the interactions between organisms and their environment. Ecology Integrates all areas of biological research and informs environmental
Chapter 52 An Introduction to Ecology and the Biosphere Ecology The study of the interactions between organisms and their environment. Ecology Integrates all areas of biological research and informs environmental
Functional Diversity. By Morgan Davies and Emily Smith
 Functional Diversity By Morgan Davies and Emily Smith Outline Introduction to biodiversity and functional diversity How do we measure functional diversity Why do we care about functional diversity Applications
Functional Diversity By Morgan Davies and Emily Smith Outline Introduction to biodiversity and functional diversity How do we measure functional diversity Why do we care about functional diversity Applications
OCR (A) Biology A-level
 Biology A-level") OCR (A) Biology A-level Topic 4.2: Biodiversity Notes Biodiversity is the variety of living organisms, over time the variety of life on Earth has become more extensive but now it is being threatened by
OCR (A) Biology A-level Topic 4.2: Biodiversity Notes Biodiversity is the variety of living organisms, over time the variety of life on Earth has become more extensive but now it is being threatened by
Overview. How many species are there? Major patterns of diversity Causes of these patterns Conserving biodiversity
 Overview How many species are there? Major patterns of diversity Causes of these patterns Conserving biodiversity Biodiversity The variability among living organisms from all sources, including, inter
Overview How many species are there? Major patterns of diversity Causes of these patterns Conserving biodiversity Biodiversity The variability among living organisms from all sources, including, inter
Appendix 1: UK climate projections
 Appendix 1: UK climate projections The UK Climate Projections 2009 provide the most up-to-date estimates of how the climate may change over the next 100 years. They are an invaluable source of information
Appendix 1: UK climate projections The UK Climate Projections 2009 provide the most up-to-date estimates of how the climate may change over the next 100 years. They are an invaluable source of information
Dominant climate influences on North American bird distributionsgeb_
 Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2011) 20, 114 118 RESEARCH PAPER Dominant climate influences on North American bird distributionsgeb_574 114..118 Alberto Jiménez-Valverde*, Narayani
Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2011) 20, 114 118 RESEARCH PAPER Dominant climate influences on North American bird distributionsgeb_574 114..118 Alberto Jiménez-Valverde*, Narayani
Pelecanus erythrorhynchos
 Published on Climate Change Sensitivity Database (http://climatechangesensitivity.org) Pelecanus erythrorhynchos This species is complete. March 17, 2010 by Jorge Tomasevic Author(s) Expertise: Print species
Published on Climate Change Sensitivity Database (http://climatechangesensitivity.org) Pelecanus erythrorhynchos This species is complete. March 17, 2010 by Jorge Tomasevic Author(s) Expertise: Print species
Journal of Pharmacognosy and Phytochemistry 2017; 6(4): Sujitha E and Shanmugasundaram K
: Sujitha E and Shanmugasundaram K") 2017; 6(4): 452-457 E-ISSN: 2278-4136 P-ISSN: 2349-8234 JPP 2017; 6(4): 452-457 Received: 01-05-2017 Accepted: 02-06-2017 Sujitha E Research Scholar, Department of Soil and Water Conservation Engineering,
2017; 6(4): 452-457 E-ISSN: 2278-4136 P-ISSN: 2349-8234 JPP 2017; 6(4): 452-457 Received: 01-05-2017 Accepted: 02-06-2017 Sujitha E Research Scholar, Department of Soil and Water Conservation Engineering,
Microhabitat choice and polymorphism
 Heredity 65 (1990) 449 454 The Genetical Society of Great Britain Received 19 April 1990 Microhabitat choice and polymorphism in the land snail Theba pisana (Muller) W. N. Hazel* and M. S. Johnsont * Department
Heredity 65 (1990) 449 454 The Genetical Society of Great Britain Received 19 April 1990 Microhabitat choice and polymorphism in the land snail Theba pisana (Muller) W. N. Hazel* and M. S. Johnsont * Department
LATITUDINAL VARIATION IN SPECIATION MECHANISMS IN FROGS
 ORIGINAL ARTICLE doi:10.1111/j.1558-5646.2009.00836.x LATITUDINAL VARIATION IN SPECIATION MECHANISMS IN FROGS Xia Hua 1,2 and John J. Wiens 1,3 1 Department of Ecology and Evolution, Stony Brook University,
ORIGINAL ARTICLE doi:10.1111/j.1558-5646.2009.00836.x LATITUDINAL VARIATION IN SPECIATION MECHANISMS IN FROGS Xia Hua 1,2 and John J. Wiens 1,3 1 Department of Ecology and Evolution, Stony Brook University,
The Origin of Species
 Chapter 24 The Origin of Species PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Chapter 24 The Origin of Species PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Rank-abundance. Geometric series: found in very communities such as the
 Rank-abundance Geometric series: found in very communities such as the Log series: group of species that occur _ time are the most frequent. Useful for calculating a diversity metric (Fisher s alpha) Most
Rank-abundance Geometric series: found in very communities such as the Log series: group of species that occur _ time are the most frequent. Useful for calculating a diversity metric (Fisher s alpha) Most
Resilience potential of the Ethiopian coffee sector under climate change
 In the format provided by the authors and unedited. SUPPLEMENTARY INFORMATION VOLUME: 3 ARTICLE NUMBER: 17081 Resilience potential of the Ethiopian coffee sector under climate change Justin Moat 1,2 *,
In the format provided by the authors and unedited. SUPPLEMENTARY INFORMATION VOLUME: 3 ARTICLE NUMBER: 17081 Resilience potential of the Ethiopian coffee sector under climate change Justin Moat 1,2 *,
Conquering the Cold: Climate suitability predictions for the Asian clam in cold temperate North America
 Conquering the Cold: Climate suitability predictions for the Asian clam in cold temperate North America Morden AL, Ricciardi A Redpath Museum McGill University International Conference on Aquatic Invasive
Conquering the Cold: Climate suitability predictions for the Asian clam in cold temperate North America Morden AL, Ricciardi A Redpath Museum McGill University International Conference on Aquatic Invasive
Chapter 19: Taxonomy, Systematics, and Phylogeny
 Chapter 19: Taxonomy, Systematics, and Phylogeny AP Curriculum Alignment Chapter 19 expands on the topics of phylogenies and cladograms, which are important to Big Idea 1. In order for students to understand
Chapter 19: Taxonomy, Systematics, and Phylogeny AP Curriculum Alignment Chapter 19 expands on the topics of phylogenies and cladograms, which are important to Big Idea 1. In order for students to understand
Observed Climate Variability and Change: Evidence and Issues Related to Uncertainty
 Observed Climate Variability and Change: Evidence and Issues Related to Uncertainty David R. Easterling National Climatic Data Center Asheville, North Carolina Overview Some examples of observed climate
Observed Climate Variability and Change: Evidence and Issues Related to Uncertainty David R. Easterling National Climatic Data Center Asheville, North Carolina Overview Some examples of observed climate
PATTERNS OF PLANT SPECIES RICHNESS IN THE CONTIGUOUS UNITED STATES INTRODUCTION
 Middle States Geographer, 2012, 44:57-64 PATTERNS OF PLANT SPECIES RICHNESS IN THE CONTIGUOUS UNITED STATES Erika Y. Chin Department of Geography State University of New York at Binghamton Binghamton,
Middle States Geographer, 2012, 44:57-64 PATTERNS OF PLANT SPECIES RICHNESS IN THE CONTIGUOUS UNITED STATES Erika Y. Chin Department of Geography State University of New York at Binghamton Binghamton,
FIG S1: Plot of rarefaction curves for Borrelia garinii Clp A allelic richness for questing Ixodes
 FIG S1: Plot of rarefaction curves for Borrelia garinii Clp A allelic richness for questing Ixodes ricinus sampled in continental Europe, England, Scotland and grey squirrels (Sciurus carolinensis) from
FIG S1: Plot of rarefaction curves for Borrelia garinii Clp A allelic richness for questing Ixodes ricinus sampled in continental Europe, England, Scotland and grey squirrels (Sciurus carolinensis) from
The Climatology of Clouds using surface observations. S.G. Warren and C.J. Hahn Encyclopedia of Atmospheric Sciences.
 The Climatology of Clouds using surface observations S.G. Warren and C.J. Hahn Encyclopedia of Atmospheric Sciences Gill-Ran Jeong Cloud Climatology The time-averaged geographical distribution of cloud
The Climatology of Clouds using surface observations S.G. Warren and C.J. Hahn Encyclopedia of Atmospheric Sciences Gill-Ran Jeong Cloud Climatology The time-averaged geographical distribution of cloud
Climate Change Vulnerability Assessment for Species
 Climate Change Vulnerability Assessment for Species SPECIES: Specify whether you are assessing the entire species or particular populations: This tool assesses the vulnerability or resilience of species
Climate Change Vulnerability Assessment for Species SPECIES: Specify whether you are assessing the entire species or particular populations: This tool assesses the vulnerability or resilience of species
Supplemental Information The geography of climate change: implications for conservation biogeography
 Supplemental Information The geography of climate change: implications for conservation biogeography D.D. Ackerly, S.R. Loarie, W.K. Cornwell, S.B. Weiss, H. Hamilton, R. Branciforte and N.J.B. Kraft Diversity
Supplemental Information The geography of climate change: implications for conservation biogeography D.D. Ackerly, S.R. Loarie, W.K. Cornwell, S.B. Weiss, H. Hamilton, R. Branciforte and N.J.B. Kraft Diversity
THE ROLE OF BBSRC IN BIODIVERSITY RESEARCH
 THE ROLE OF BBSRC IN BIODIVERSITY RESEARCH INTRODUCTION The aim of this document is to explain the role of biodiversity research in the delivery of the BBSRC mission, and thereby to provide guidance to
THE ROLE OF BBSRC IN BIODIVERSITY RESEARCH INTRODUCTION The aim of this document is to explain the role of biodiversity research in the delivery of the BBSRC mission, and thereby to provide guidance to
A Study of the Uncertainty in Future Caribbean Climate Using the PRECIS Regional Climate Model
 A Study of the Uncertainty in Future Caribbean Climate Using the PRECIS Regional Climate Model by Abel Centella and Arnoldo Bezanilla Institute of Meteorology, Cuba & Kenrick R. Leslie Caribbean Community
A Study of the Uncertainty in Future Caribbean Climate Using the PRECIS Regional Climate Model by Abel Centella and Arnoldo Bezanilla Institute of Meteorology, Cuba & Kenrick R. Leslie Caribbean Community
Figure 1. Time series of Western Sahel precipitation index and Accumulated Cyclone Energy (ACE).
.") 2B.6 THE NON-STATIONARY CORRELATION BETWEEN SAHEL PRECIPITATION INDICES AND ATLANTIC HURRICANE ACTIVITY Andreas H. Fink 1 and Jon M. Schrage 2 1 Institute for Geophysics 2 Department of and Meteorology
2B.6 THE NON-STATIONARY CORRELATION BETWEEN SAHEL PRECIPITATION INDICES AND ATLANTIC HURRICANE ACTIVITY Andreas H. Fink 1 and Jon M. Schrage 2 1 Institute for Geophysics 2 Department of and Meteorology
UON, CAS, DBSC, General Biology II (BIOL102) Dr. Mustafa. A. Mansi. The Origin of Species
 Dr. Mustafa. A. Mansi. The Origin of Species") The Origin of Species Galápagos Islands, landforms newly emerged from the sea, despite their geologic youth, are filled with plants and animals known no-where else in the world, Speciation: The origin
The Origin of Species Galápagos Islands, landforms newly emerged from the sea, despite their geologic youth, are filled with plants and animals known no-where else in the world, Speciation: The origin
ENSO-DRIVEN PREDICTABILITY OF TROPICAL DRY AUTUMNS USING THE SEASONAL ENSEMBLES MULTIMODEL
 1 ENSO-DRIVEN PREDICTABILITY OF TROPICAL DRY AUTUMNS USING THE SEASONAL ENSEMBLES MULTIMODEL Based on the manuscript ENSO-Driven Skill for precipitation from the ENSEMBLES Seasonal Multimodel Forecasts,
1 ENSO-DRIVEN PREDICTABILITY OF TROPICAL DRY AUTUMNS USING THE SEASONAL ENSEMBLES MULTIMODEL Based on the manuscript ENSO-Driven Skill for precipitation from the ENSEMBLES Seasonal Multimodel Forecasts,
Lecture Outlines PowerPoint. Chapter 20 Earth Science 11e Tarbuck/Lutgens
 Lecture Outlines PowerPoint Chapter 20 Earth Science 11e Tarbuck/Lutgens 2006 Pearson Prentice Hall This work is protected by United States copyright laws and is provided solely for the use of instructors
Lecture Outlines PowerPoint Chapter 20 Earth Science 11e Tarbuck/Lutgens 2006 Pearson Prentice Hall This work is protected by United States copyright laws and is provided solely for the use of instructors
Conceptually, we define species as evolutionary units :
 Bio 1M: Speciation 1 How are species defined? S24.1 (2ndEd S26.1) Conceptually, we define species as evolutionary units : Individuals within a species are evolving together Individuals of different species
Bio 1M: Speciation 1 How are species defined? S24.1 (2ndEd S26.1) Conceptually, we define species as evolutionary units : Individuals within a species are evolving together Individuals of different species
Priority areas for grizzly bear conservation in western North America: an analysis of habitat and population viability INTRODUCTION METHODS
 Priority areas for grizzly bear conservation in western North America: an analysis of habitat and population viability. Carroll, C. 2005. Klamath Center for Conservation Research, Orleans, CA. Revised
Priority areas for grizzly bear conservation in western North America: an analysis of habitat and population viability. Carroll, C. 2005. Klamath Center for Conservation Research, Orleans, CA. Revised
Biogeography. An ecological and evolutionary approach SEVENTH EDITION. C. Barry Cox MA, PhD, DSc and Peter D. Moore PhD
 Biogeography An ecological and evolutionary approach C. Barry Cox MA, PhD, DSc and Peter D. Moore PhD Division of Life Sciences, King's College London, Fmnklin-Wilkins Building, Stamford Street, London
Biogeography An ecological and evolutionary approach C. Barry Cox MA, PhD, DSc and Peter D. Moore PhD Division of Life Sciences, King's College London, Fmnklin-Wilkins Building, Stamford Street, London
AN INTERNATIONAL SOLAR IRRADIANCE DATA INGEST SYSTEM FOR FORECASTING SOLAR POWER AND AGRICULTURAL CROP YIELDS
 AN INTERNATIONAL SOLAR IRRADIANCE DATA INGEST SYSTEM FOR FORECASTING SOLAR POWER AND AGRICULTURAL CROP YIELDS James Hall JHTech PO Box 877 Divide, CO 80814 Email: jameshall@jhtech.com Jeffrey Hall JHTech
AN INTERNATIONAL SOLAR IRRADIANCE DATA INGEST SYSTEM FOR FORECASTING SOLAR POWER AND AGRICULTURAL CROP YIELDS James Hall JHTech PO Box 877 Divide, CO 80814 Email: jameshall@jhtech.com Jeffrey Hall JHTech
Climate Change and Invasive Plants in the Pacific Northwest
 Climate Change and Invasive Plants in the Pacific Northwest David W Peterson Becky K Kerns Ecosystem Dynamics and Environmental Change Team Threat Characterization and Management Program Pacific Northwest
Climate Change and Invasive Plants in the Pacific Northwest David W Peterson Becky K Kerns Ecosystem Dynamics and Environmental Change Team Threat Characterization and Management Program Pacific Northwest
July Forecast Update for Atlantic Hurricane Activity in 2017
 July Forecast Update for Atlantic Hurricane Activity in 2017 Issued: 4 th July 2017 by Professor Mark Saunders and Dr Adam Lea Dept. of Space and Climate Physics, UCL (University College London), UK Forecast
July Forecast Update for Atlantic Hurricane Activity in 2017 Issued: 4 th July 2017 by Professor Mark Saunders and Dr Adam Lea Dept. of Space and Climate Physics, UCL (University College London), UK Forecast
EXAM PRACTICE. 12 questions * 4 categories: Statistics Background Multivariate Statistics Interpret True / False
 EXAM PRACTICE 12 questions * 4 categories: Statistics Background Multivariate Statistics Interpret True / False Stats 1: What is a Hypothesis? A testable assertion about how the world works Hypothesis
EXAM PRACTICE 12 questions * 4 categories: Statistics Background Multivariate Statistics Interpret True / False Stats 1: What is a Hypothesis? A testable assertion about how the world works Hypothesis
4. is the rate at which a population of a given species will increase when no limits are placed on its rate of growth.
 Population Ecology 1. Populations of mammals that live in colder climates tend to have shorter ears and limbs than populations of the same species in warm climates (coyotes are a good example of this).
Population Ecology 1. Populations of mammals that live in colder climates tend to have shorter ears and limbs than populations of the same species in warm climates (coyotes are a good example of this).
The Environment Habitat and Niche
 The Environment Habitat and Niche Ecology By KAPMAN LIFE SCIENCE ACADEMY Call us at +91-97296-37935 1 Ecology Scientific study of the relationships between organisms and their environment. Environment
The Environment Habitat and Niche Ecology By KAPMAN LIFE SCIENCE ACADEMY Call us at +91-97296-37935 1 Ecology Scientific study of the relationships between organisms and their environment. Environment
3.3 Threats to Biodiversity
 Name: Date: Class: IB Environmental Systems and Societies 3.3 Threats to Biodiversity Significant ideas: While global biodiversity is difficult to quantify, it is decreasing rapidly due to human activity.
Name: Date: Class: IB Environmental Systems and Societies 3.3 Threats to Biodiversity Significant ideas: While global biodiversity is difficult to quantify, it is decreasing rapidly due to human activity.
Observed changes in climate and their effects
 1 1.1 Observations of climate change Since the TAR, progress in understanding how climate is changing in space and time has been gained through improvements and extensions of numerous datasets and data
1 1.1 Observations of climate change Since the TAR, progress in understanding how climate is changing in space and time has been gained through improvements and extensions of numerous datasets and data
Cuba. General Climate. Recent Climate Trends. UNDP Climate Change Country Profiles. Temperature. C. McSweeney 1, M. New 1,2 and G.
 UNDP Climate Change Country Profiles Cuba C. McSweeney 1, M. New 1,2 and G. Lizcano 1 1. School of Geography and Environment, University of Oxford. 2. Tyndall Centre for Climate Change Research http://country-profiles.geog.ox.ac.uk
UNDP Climate Change Country Profiles Cuba C. McSweeney 1, M. New 1,2 and G. Lizcano 1 1. School of Geography and Environment, University of Oxford. 2. Tyndall Centre for Climate Change Research http://country-profiles.geog.ox.ac.uk
Our climate system is based on the location of hot and cold air mass regions and the atmospheric circulation created by trade winds and westerlies.
 CLIMATE REGIONS Have you ever wondered why one area of the world is a desert, another a grassland, and another a rainforest? Or have you wondered why are there different types of forests and deserts with
CLIMATE REGIONS Have you ever wondered why one area of the world is a desert, another a grassland, and another a rainforest? Or have you wondered why are there different types of forests and deserts with
Community phylogenetics review/quiz
 Community phylogenetics review/quiz A. This pattern represents and is a consequent of. Most likely to observe this at phylogenetic scales. B. This pattern represents and is a consequent of. Most likely
Community phylogenetics review/quiz A. This pattern represents and is a consequent of. Most likely to observe this at phylogenetic scales. B. This pattern represents and is a consequent of. Most likely
Gynura aurantiaca (purple velvet plant)
") Australia/New Zealand Weed Risk Assessment adapted for Florida. Data used for analysis published in: Gordon, D.R., D.A. Onderdonk, A.M. Fox, R.K. Stocker, and C. Gantz. 28. Predicting Invasive Plants in
Australia/New Zealand Weed Risk Assessment adapted for Florida. Data used for analysis published in: Gordon, D.R., D.A. Onderdonk, A.M. Fox, R.K. Stocker, and C. Gantz. 28. Predicting Invasive Plants in
Advanced Placement Biology Union City High School Summer Assignment 2011 Ecology Short Answer Questions
 Summer Assignment 2011 Ecology Short Answer Questions 1. Each of the terrestrial biomes have very different characteristics that determine the niches of the organisms that live within that biome. (a) Select
Summer Assignment 2011 Ecology Short Answer Questions 1. Each of the terrestrial biomes have very different characteristics that determine the niches of the organisms that live within that biome. (a) Select
Historical contingency, niche conservatism and the tendency for some taxa to be more diverse towards the poles
 Electronic Supplementary Material Historical contingency, niche conservatism and the tendency for some taxa to be more diverse towards the poles Ignacio Morales-Castilla 1,2 *, Jonathan T. Davies 3 and
Electronic Supplementary Material Historical contingency, niche conservatism and the tendency for some taxa to be more diverse towards the poles Ignacio Morales-Castilla 1,2 *, Jonathan T. Davies 3 and
A Simulation of the Process of Evolution Modified from Biology Labs On-Line (Pearson)
") A Simulation of the Process of Evolution Modified from Biology Labs On-Line (Pearson) Biology Labs On-line EvolutionLab is a simulation which allows you to study the principles and processes behind the
A Simulation of the Process of Evolution Modified from Biology Labs On-Line (Pearson) Biology Labs On-line EvolutionLab is a simulation which allows you to study the principles and processes behind the
From basic taxonomic data towards a synthesis in plant invasion ecology. Petr Pyšek
 From basic taxonomic data towards a synthesis in plant invasion ecology Petr Pyšek Alien flora of the Czech Republic ca 2700 native + 1378 alien taxa Aliens form 33% of the Czech flora Pyšek P., Sádlo
From basic taxonomic data towards a synthesis in plant invasion ecology Petr Pyšek Alien flora of the Czech Republic ca 2700 native + 1378 alien taxa Aliens form 33% of the Czech flora Pyšek P., Sádlo
Prediction of Snow Water Equivalent in the Snake River Basin
 Hobbs et al. Seasonal Forecasting 1 Jon Hobbs Steve Guimond Nate Snook Meteorology 455 Seasonal Forecasting Prediction of Snow Water Equivalent in the Snake River Basin Abstract Mountainous regions of
Hobbs et al. Seasonal Forecasting 1 Jon Hobbs Steve Guimond Nate Snook Meteorology 455 Seasonal Forecasting Prediction of Snow Water Equivalent in the Snake River Basin Abstract Mountainous regions of
VCS MODULE VMD0018 METHODS TO DETERMINE STRATIFICATION
 VMD0018: Version 1.0 VCS MODULE VMD0018 METHODS TO DETERMINE STRATIFICATION Version 1.0 16 November 2012 Document Prepared by: The Earth Partners LLC. Table of Contents 1 SOURCES... 2 2 SUMMARY DESCRIPTION
VMD0018: Version 1.0 VCS MODULE VMD0018 METHODS TO DETERMINE STRATIFICATION Version 1.0 16 November 2012 Document Prepared by: The Earth Partners LLC. Table of Contents 1 SOURCES... 2 2 SUMMARY DESCRIPTION
The importance of sampling multidecadal variability when assessing impacts of extreme precipitation
 The importance of sampling multidecadal variability when assessing impacts of extreme precipitation Richard Jones Research funded by Overview Context Quantifying local changes in extreme precipitation
The importance of sampling multidecadal variability when assessing impacts of extreme precipitation Richard Jones Research funded by Overview Context Quantifying local changes in extreme precipitation
Climate and Biomes. Adapted by T.Brunetto from: Developed by Steven Taylor Wichmanowski based in part on Pearson Environmental Science by Jay Withgott
 Climate and Biomes Adapted by T.Brunetto from: Developed by Steven Taylor Wichmanowski based in part on Pearson Environmental Science by Jay Withgott Remember that an ecosystem consists of all the biotic
Climate and Biomes Adapted by T.Brunetto from: Developed by Steven Taylor Wichmanowski based in part on Pearson Environmental Science by Jay Withgott Remember that an ecosystem consists of all the biotic
Assessing state-wide biodiversity in the Florida Gap analysis project
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Nebraska Cooperative Fish & Wildlife Research Unit -- Staff Publications Nebraska Cooperative Fish & Wildlife Research Unit
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Nebraska Cooperative Fish & Wildlife Research Unit -- Staff Publications Nebraska Cooperative Fish & Wildlife Research Unit
Evolution Common Assessment 1
 Evolution Common Assessment 1 1. The field of biology that includes the study of the origin of new species through time is known as 5. A. biochemistry B. evolution C. ecology D. embryology 2. Evidence
Evolution Common Assessment 1 1. The field of biology that includes the study of the origin of new species through time is known as 5. A. biochemistry B. evolution C. ecology D. embryology 2. Evidence
Grenada. General Climate. Recent Climate Trends. UNDP Climate Change Country Profiles. Temperature. Precipitation
 UNDP Climate Change Country Profiles C. McSweeney 1, M. New 1,2 and G. Lizcano 1 1. School of Geography and Environment, University of Oxford. 2. Tyndall Centre for Climate Change Research http://country-profiles.geog.ox.ac.uk
UNDP Climate Change Country Profiles C. McSweeney 1, M. New 1,2 and G. Lizcano 1 1. School of Geography and Environment, University of Oxford. 2. Tyndall Centre for Climate Change Research http://country-profiles.geog.ox.ac.uk
What is Climate? Understanding and predicting climatic changes are the basic goals of climatology.
 What is Climate? Understanding and predicting climatic changes are the basic goals of climatology. Climatology is the study of Earth s climate and the factors that affect past, present, and future climatic
What is Climate? Understanding and predicting climatic changes are the basic goals of climatology. Climatology is the study of Earth s climate and the factors that affect past, present, and future climatic
Climate Chapter 19. Earth Science, 10e. Stan Hatfield and Ken Pinzke Southwestern Illinois College
 Climate Chapter 19 Earth Science, 10e Stan Hatfield and Ken Pinzke Southwestern Illinois College The climate system A. Climate is an aggregate of weather B. Involves the exchanges of energy and moisture
Climate Chapter 19 Earth Science, 10e Stan Hatfield and Ken Pinzke Southwestern Illinois College The climate system A. Climate is an aggregate of weather B. Involves the exchanges of energy and moisture
Stochastic Hydrology. a) Data Mining for Evolution of Association Rules for Droughts and Floods in India using Climate Inputs
 Data Mining for Evolution of Association Rules for Droughts and Floods in India using Climate Inputs") Stochastic Hydrology a) Data Mining for Evolution of Association Rules for Droughts and Floods in India using Climate Inputs An accurate prediction of extreme rainfall events can significantly aid in policy
Stochastic Hydrology a) Data Mining for Evolution of Association Rules for Droughts and Floods in India using Climate Inputs An accurate prediction of extreme rainfall events can significantly aid in policy