Variable Consequences of Toxic Prey on Generalist Insect Predators
|
|
|
- Chastity Little
- 5 years ago
- Views:
Transcription

1 University of Kentucky UKnowledge Theses and Dissertations--Entomology Entomology 2016 Variable Consequences of Toxic Prey on Generalist Insect Predators Kelly Jackson University of Kentucky, Digital Object Identifier: Click here to let us know how access to this document benefits you. Recommended Citation Jackson, Kelly, "Variable Consequences of Toxic Prey on Generalist Insect Predators" (2016). Theses and Dissertations--Entomology This Master's Thesis is brought to you for free and open access by the Entomology at UKnowledge. It has been accepted for inclusion in Theses and Dissertations--Entomology by an authorized administrator of UKnowledge. For more information, please contact
2 STUDENT AGREEMENT: I represent that my thesis or dissertation and abstract are my original work. Proper attribution has been given to all outside sources. I understand that I am solely responsible for obtaining any needed copyright permissions. I have obtained needed written permission statement(s) from the owner(s) of each thirdparty copyrighted matter to be included in my work, allowing electronic distribution (if such use is not permitted by the fair use doctrine) which will be submitted to UKnowledge as Additional File. I hereby grant to The University of Kentucky and its agents the irrevocable, non-exclusive, and royaltyfree license to archive and make accessible my work in whole or in part in all forms of media, now or hereafter known. I agree that the document mentioned above may be made available immediately for worldwide access unless an embargo applies. I retain all other ownership rights to the copyright of my work. I also retain the right to use in future works (such as articles or books) all or part of my work. I understand that I am free to register the copyright to my work. REVIEW, APPROVAL AND ACCEPTANCE The document mentioned above has been reviewed and accepted by the student s advisor, on behalf of the advisory committee, and by the Director of Graduate Studies (DGS), on behalf of the program; we verify that this is the final, approved version of the student s thesis including all changes required by the advisory committee. The undersigned agree to abide by the statements above. Kelly Jackson, Student Dr. Jennifer White, Major Professor Dr. Charles Fox, Director of Graduate Studies
3 VARIABLE CONSEQUENCES OF TOXIC PREY ON GENERALIST INSECT PREDATORS THESIS A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in Entomology in the College of Agriculture, Food, and Environment at the University of Kentucky By Kelly Ann Jackson Lexington, Kentucky Director: Dr. Jennifer White, Associate Professor of Entomology Lexington, Kentucky 2016 Copyright Kelly Ann Jackson 2016
4 ABSTRACT OF THESIS VARIABLE CONSEQUENCES OF TOXIC PREY ON GENERALIST INSECT PREDATORS Introduced species, like the lady beetle Harmonia axyridis, have been linked to declines of native species through mechanisms including intraguild predation and competitive superiority. However, competitive differentials between species may be mitigated if subdominant species can utilize resources that dominant species cannot. Previous research has shown that some strains of the aphid Aphis craccivora are toxic to H. axyridis. My goal was to investigate use of this resource by both H. axyridis and other lady beetles, to determine whether these aphids might be an exploitable resource for subdominant lady beetle species. I first examined the behavioral responses of adult and larval H. axyridis to toxic strains of A. craccivora. I found that adults invested less time and laid fewer eggs with toxic than nontoxic aphids, and larvae consumed toxic aphids at a slower rate, often refusing them as a food source. I then tested whether six other lady beetle species could use the aphids, monitoring larval development in no-choice environments with different strains of A. craccivora. All species showed increased survival and development rates relative to H. axyridis on toxic aphid strains, suggesting these aphids may allow other coccinellid species to experience competitive release from the otherwise dominant H. axyridis. KEYWORDS: behavior, Coccinellidae, community interactions, competition, defense, predator-prey interactions Kelly Ann Jackson July 27, 2016
5 VARIABLE CONSEQUENCES OF TOXIC PREY ON GENERALIST INSECT PREDATORS By Kelly Ann Jackson Dr. Jennifer A White Director of Thesis Dr. Charles Fox Director of Graduate Studies July 27, 2016
6 DEDICATION For the real Jackson 5
7 ACKNOWLEDGEMENTS I would like to thank my major advisor, Dr. Jen White, for her unwavering support, encouragement, and guidance for the last two years; my development and growth as a scientist and writer would not have happened without you. I would also like to thank my committee members, Dr. John Obrycki and Dr. Ken Haynes, for their assistance and advice in developing my project. I would like to thank all the members of our Endosymbiont Ecology lab for their help with beetle and aphid colony care, statistical help, moral support, and friendship. A very special thanks goes to my parents and sisters, and to my extended Louisville family, for their love, support, and constant encouragement. iii
8 TABLE OF CONTENTS List of Tables...v List of Figures... vi Chapter 1: Introduction...1 Chapter 2: Prey Suitability Determines Behavioral Response in a Generalist Coccinellid Predator...6 Introduction...6 Materials and Methods...9 Results...14 Discussion...16 Tables and Figures...20 Chapter 3: A Window of Opportunity: Subdominant Predators Can Use Suboptimal Prey...26 Introduction...26 Materials and Methods...28 Results...30 Discussion...33 Tables and Figures...36 Chapter 4: Summary and Conclusions...39 References...43 Vita...49 iv
9 LIST OF TABLES Table 3.1: Lady beetle natural history...36 v
10 LIST OF FIGURES Figure 2.1: Mean ± S.E. adult H. axyridis beetle retention over 24h at patches of toxic or nontoxic A. craccivora aphids in an open field experiment...20 Figure 2.2: Mean ± S.E. eggs laid by H. axyridis over 24h at patches of toxic or nontoxic A. craccivora...21 Figure 2.3: Mean ± S.E. A. craccivora consumed by H. axyridis larvae over 24h in toxic or nontoxic aphid treatments...22 Figure 2.4: Mean ± S.E. rejected A. craccivora in toxic or nontoxic aphid treatments, after being preyed upon by H. axyridis for 24h...23 Figure 2.5: Mean time 2 nd instar H. axyridis larva spent per bout of each activity over 60 minutes, following prior exposure to toxic or nontoxic A. craccivora...24 Figure 2.6: Schematic diagram of prey acceptance versus rejection for 2 nd instar H. axyridis larvae encountering nontoxic or toxic A. craccivora...25 Figure 3.1: Survival curves for larvae of five lady beetle species when feeding on either L-strain or A-strain A. craccivora...37 Figure 3.2: Mean ± S.E. adult mass of five lady beetle species after feeding on L- strain or A-strain A. craccivora throughout larval development...38 vi
11 CHAPTER 1: INTRODUCTION Generalist predators are essential to ecosystem function, and play a particularly large role in agricultural systems because of the top-down control they offer (Rosenheim 1998; Snyder et al. 2006). The ecosystem services they provide in controlling a wide variety of pest species have been quantified to be worth more than $4.5 billion annually (Losey & Vaughan 2006). Several beetles in the coccinellid family, or lady beetles, are well studied and easily recognized examples of generalist predators, consuming everything from garden pests to other predator species. While some adult coccinellids primarily consume aphids and scales, larvae typically consume a much broader range of prey items, including other beneficials, conspecifics, and even food sources that might not be very nutritious (Koch et al. 2003; Pell et al. 2008). The suitability of coccinellid prey items can be divided into three categories. First, prey items that allow full larval development and adult reproductive success are termed essential (Hodek & Evans 2012). For example, when Coccinella septempunctata are reared on pea aphid, Acyrthosiphon pisum, they are able to fully develop into adulthood, and as adults, they produce viable eggs (Blackman 1967). Second, "alternative" prey items are those that only maintain survival and do not support larval development or adult fecundity (Hodek & Evans 2012). When feeding on the aphid Aphis sambuci, the lady beetle Adalia bipunctata does not develop, but can survive until it finds a more suitable food source (Hodek 1967). Finally, some food items are not suitable for consumption. For example, Aphis sambuci is toxic to C. septempunctata, with no larvae that consume the aphid surviving to adulthood (Nielsen, Hauge & Toft 2002). 1
12 Likewise, the aphid Aphis nerii has been shown to be toxic to multiple coccinellid species, including C. septempunctata and A. bipunctata (Omkar & Srivastava 2003; Colvin & Yeargan 2014). Toxicity in herbivores is often attributed to secondary chemicals from the host plant that the herbivore consumes (Opitz & Müller 2009). When some herbivorous insects ingest plants, they sequester or store secondary toxic substances in their body tissues or gut, which enhances their own defense and protects them from predation (Opitz & Müller 2009). The monarch butterfly is a standard example, with larvae ingesting cardenolides from their milkweed host plant. Cardenolides are toxic steroids that induce vomiting in predators, and many species of generalist avian predator avoid consuming the monarch because of the presence of these chemicals (Brower et al. 1968; Malcolm & Brower 1989). Another form of sequestration occurs in Neodiprion sertifer. These sawflies consume terpenoids when feeding on their pine host plants, which are then sequestered in their foregut and used in a defensive fluid that is discharged as a resin when they perceive an attack (Eisner et al. 1974). The oleander aphid, Aphis nerii, functions in a similar fashion to the monarch, also sequestering cardenolides that are used as a defense towards coccinellid predators (Malcolm 1990). Other aphids, like Aphis craccivora (the cowpea aphid), also demonstrate toxicity toward coccinellids. In A. craccivora, only aphids associated with some host plants have been shown to be toxic (Hukusima & Kamei 1970; Kamo, Tokuoka & Miyazaki 2010). When the multicolored Asian lady beetle, Harmonia axyridis, consumes A. craccivora from common vetch (Vicia sp.), larval beetles survive to adulthood. Conversely, H. 2
13 axyridis that consume A. craccivora from black locust, Robinia pseudoacacia, experience 100% mortality (Hukusima & Kamei 1970). Based on the assumption that chemicals sequestered from the aphid s host plant were the cause of the differential toxicity, the chemicals present in the foliage of black locust were investigated to determine which ones could be the cause of the toxicity in A. craccivora. Obatake and Suzuki (1985) determined that two compounds, ethanolamine and canavanine, when fed in artificial diets to H. axyridis led to quick mortality in larval beetles. More recently, Kamo et al. (2012) found a third chemical, cyanamide, present in locust that was up to 100 times more effective at inducing larval mortality when fed to H. axyridis than ethanolamine or canavanine. Given that cyanamide was not present in common vetch (Kamo, Tokuoka & Miyazaki 2012) this study furthered the idea that the toxicity of A. craccivora is a byproduct of chemicals present in black locust. However, in each of these studies, the aphids used in the assays were taken directly from their original host plant in the field. These wild-caught aphids from multiple host plants may have also differed from one another in genetic makeup, potentially confounding the cause of the toxicity to H. axyridis. Previous studies in our lab have demonstrated that toxicity in A. craccivora is actually a function of the aphid lineage, rather than the host plant (White et al. 2016). We have taken A. craccivora from both locust and alfalfa (Medicago sativa) and reared them on Vicia faba (fava bean) for over a year, or at least 50 aphid generations, ensuring the removal of any maternal host plant effects. The development of H. axyridis larvae was then monitored while they were being fed these differing aphid lineages (White et al. 2016). The alfalfa-origin aphids were a suitable food source for H. axyridis larvae, with 3
14 larvae surviving to adulthood. The locust-origin aphids, however, were toxic to the larvae regardless of whether they were taken from locust or fava, usually causing lady beetle mortality within 4 days. Even though the fava-reared A. craccivora had not been exposed to locust for over a year, they still retained their toxicity, indicating that the toxicity is intrinsic to the aphid itself instead of being a product of its current host plant. Another facet to consider in the interactions between A. craccivora and lady beetles is the aphid s microbiome. Locust-origin A. craccivora are characteristically infected with the maternally-inherited facultative symbiont Arsenophonus, whereas aphids that are alfalfa-origin are often infected with the facultative symbiont Hamiltonella (Brady & White 2013). Initially, it was hypothesized that the presence or absence of the symbiont Arsenophonus was the cause of the toxicity in the locust-origin aphids; however, this is not the case (White et al. 2016). By curing the aphid with an antibiotic diet, Arsenophonus was removed from locust-origin lines (Wagner et al. 2015). When the locust-origin aphids without Arsenophonus were fed to H. axyridis larvae, they remained toxic (White et al. 2016). While the exact mechanism for the toxicity is still unknown, these results reinforce that the aphid toxicity is intrinsic to the aphid lineage. Harmonia axyridis is a common non-native coccinellid species often associated with biological control (Koch 2003). It has been suggested that this coccinellid is linked to the decline of native coccinellids through intraguild predation and/or competitive superiority (Koch 2003; Harmon, Stephens & Losey 2007; Pell et al. 2008; Smith & Gardiner 2013). In other systems, it has been demonstrated that not all lady beetle species are affected the same way by toxic properties of different aphid species (Hodek & Evans 2012). If other coccinellid species are not affected in the same manner and are able to use 4
15 toxic A. craccivora as a food source, these aphids could potentially have a mitigating effect on the dominance of H. axyridis by providing a refuge food source for other coccinellids. My goal is to examine the use of toxic A. craccivora by both H. axyridis and other lady beetle species. My first objective, addressed in Chapter 2, is to assess the behavioral responses of adult and larval H. axyridis to toxic strains of A. craccivora, to gain a broader understanding of predator-prey interactions when there is intraspecific variation in toxicity. My second objective, addressed in Chapter 3, is to test how six other lady beetle species are affected by toxic A. craccivora lineages, to determine if these aphids have the potential to function as a refuge food source, providing less dominant coccinellid species with competitor-free space. 5
16 CHAPTER 2: PREY SUITABILITY DETERMINES BEHAVIORAL RESPONSE IN A GENERALIST COCCINELLID PREDATOR Introduction Coccinellids, or lady beetles, are often used in biological control programs because they consume large numbers of aphids and other agricultural and garden pests throughout their entire life cycle (Koch 2003). While some of the species used in these programs are generalists that might be considered indiscriminate consumers, not all aphids are equally acceptable food (Blackman 1967). Some aphid species are unsuitable, or even toxic, and can result in beetle mortality following consumption (Hukusima & Kamei 1970; Hodek & Evans 2012). When coccinellids forage, they typically begin by utilizing broad extensive search behaviors, switching to intensive restricted area searches once they find a food source (Banks 1957; Dixon 1959). After depleting the food source, they resume extensive searching. However, when a larval beetle encounters an unsuitable food source during their extensive search, instead of switching to intensive searching they often respond by ceasing all foraging activities, and remaining immobile (Kalushkov 1999). When adult females encounter toxic prey, they should choose to not lay eggs in association with such a poor food source, so that their less mobile offspring don't encounter unsuitable prey as their initial meal (Fréchette et al. 2006). However, there are also examples in which females lay eggs in association with poor food, perhaps because they are basing their oviposition decisions on other factors, like the presence or absence of conspecific larvae (Hemptinne, Dixon & Coffin 1992). Larvae that hatch in environments with poor food 6
17 choices have been shown under some circumstances to continually consume unsuitable aphids, even if they are toxic (Nedved & Salvucci 2008). On the other hand, they have also been observed to reject unsuitable aphids before full consumption (Blackman 1967). It is possible that predators might not be able to easily distinguish suitable from unsuitable food sources when foraging. Coccinellids often use signals from the aphid, such as contrasting coloration, to determine whether or not to attack (Harmon, Losey & Ives 1998; Mondor & Warren 2000). Some aphid species employ aposematic coloration as a warning signal to predators (Berenbaum & Miliczky 1984). However, there are more subtle defenses that vary even within species that can protect the prey from being consumed. For example, the presence of microbial symbionts in insects has been found to play a role in providing a defense against predators and parasitoids. Fruit flies can be infected with Spiroplasma bacteria that protect them from nematode parasites (Jaenike et al. 2010), and aphids can be infected with Hamiltonella bacteria that protect them from wasp parasitism (Oliver et al. 2003). The cowpea aphid, Aphis craccivora, also shows a wide range of intraspecific variation in toxicity to the lady beetle Harmonia axyridis (Hukusima & Kamei 1970; White et al. 2016). This variation in toxicity is a heritable property that is intrinsic to the individual aphid clones, but is neither attributable to facultative bacterial symbionts nor a function of the aphid's host plant (White et al. 2016). The toxic strains consistently affect both adult and larval H. axyridis (White et al. 2016), but have only been examined in no-choice environments and without observation of individual behavior upon encountering toxic versus nontoxic prey. Determining how both adult and larval H. axyridis behave when encountering an ambiguously toxic food source can illustrate how much this predator might actually 7
18 consume the aphids in the field, which has important implications for both predator and prey. The decisions that adult female beetles make during oviposition ultimately determine the first food source that a larva will encounter (Refsnider & Janzen 2010). If mobile adults avoid ovipositing near unacceptable food, larval offspring may not even come across these unsuitable food sources under natural conditions. Additionally, larval behavior when encountering toxic strains of A. craccivora is unknown, but would determine the rate of aphid consumption, probability of aphid survival, and potential defensive efficacy of the toxic trait. I conducted a number of trials to investigate the behaviors of H. axyridis when faced with either the toxic or nontoxic strain of A. craccivora. First, I examined patch retention time of adult females in an open field choice environment, determining tenure and oviposition to understand future offspring exposure. Second, I quantified larval consumption of unsuitable versus suitable prey items in no-choice environments over time, to assess how much a young larva will consume when encountering their first food source. Third, I directly observed larval rejection behaviors of a toxic food source, for a greater understanding of their foraging decisions. Understanding the behaviors of H. axyridis throughout its lifecycle when interacting with the different strains of A. craccivora can add to the understanding of predator-prey interactions, allowing for the assessment of how well a toxic trait deters a susceptible predator and potentially protects the prey. Predator deterrence can have effects not only on the prey that is escaping predation, but also on other predators that can potentially use that prey item as a resource. 8
19 Materials and Methods Origin and maintenance of insects Adult beetles were collected in Lexington, KY, USA, and kept in incubators at 23 C, 16h:8h light:dark, 65% humidity. They were kept in Petri dishes (100mm 25mm) and provided with pea aphids (Acyrthosiphum pisum) as a food source and a cotton ball saturated with deionized water. I removed the beetle pairs as they mated and placed them into their own Petri dishes, provisioned with Ac. pisum, a DI water cotton ball, and a piece of folded paper for egg deposition. When eggs were laid, I moved the papers to new Petri dishes, and eggs were monitored until they hatched. Aphis craccivora used in these experiments were from laboratory colonies of aphid clones that are either toxic or nontoxic to H. axyridis as described in White et al. (2016). Toxic clones were originally collected from black locust (Robinia pseudoacacia L.) and nontoxic clones from alfalfa (Medicago sativa L.) in central KY, USA, and had been maintained in the lab for at least two years on fava bean (Vicia faba) (Wagner et al. 2015). The toxicity of the aphid clones (or lack thereof) was verified by coccinellid survival and development assays (White et al. 2016). Adult retention time I evaluated the behavior of adult H. axyridis in an open field experiment to determine whether beetles discriminated against toxic A. craccivora. I compared a) the tenure of beetles in patches of toxic versus nontoxic A. craccivora, and b) beetle egg deposition in the two patch types. Aphis craccivora were placed onto young fava bean plants (Vicia faba), with each plant receiving either toxic or nontoxic aphids that were 9
20 allowed to reproduce for two weeks prior to experimental use. Each plant was visually estimated to have more than 3,000 aphids, which was sufficient to ensure that the beetles were not food limited in the 24h timeframe of the beetle retention experiment. I randomly assigned mated adult female H. axyridis to one of the two treatments. I marked each beetle with a dot of paint that corresponded to the treatment to ensure that retention, rather than recruitment, was being recorded. I placed two adult females on each plant and enclosed the plants in mesh bags for transport to Spindletop Research Farm (Lexington, KY; N, W), allowing them at least two hours to forage on the plants. I randomly assigned each plant to a location within an old field plot, within which holding pots had been placed in the ground 5m apart from one another in a 4 4 grid, and removed the mesh bags that were covering the plants. After uncovering the plants, I ensured that all beetles placed on the plants were still present; one plant was excluded due to a missing beetle. The number of beetles present, egg masses, and total number of eggs were counted every 20 minutes for the first hour and every 30 minutes for the next 5 hours. A final count was conducted at 24h. The experiments were conducted on three different days, with 8 plants on day one and 16 plants each on days two and three. I compared number of remaining beetles per plant between treatments for the first six hourly time points using a repeated measures ANOVA. Final 24h beetle counts were initially compared between treatments in a mixed model ANOVA with experimental day as a random effect and aphid toxicity as a fixed effect. I found no significant effect of day, and removed the factor from the model, leaving a simple t-test between toxicity treatments. I also compared total egg deposition per plant between treatments with a t- test; all statistical comparisons were completed in JMP10. 10
21 Larval consumption of aphids over time After observing egg deposition associated with a toxic food source in the field experiment, I sought to characterize the larval feeding patterns of H. axyridis over time, to understand how larvae consume an unsuitable food source in a no-choice environment. In a field setting, neonate larvae do not leave their natal plant even when surrounded by exclusively toxic aphids (Lenhart et al., unpublished data), so a no-choice environment is suitable to study larval consumption of aphids. I removed newly hatched neonate larvae from their egg masses before dispersal, and separated them from each other to prevent sibling cannibalism. After sclerotization, I placed larvae individually into leaf disk arenas containing either nontoxic or toxic A. craccivora. Each enclosed arena consisted of an excised circle of fava bean leaf in a 35mm 10mm Petri dish with 1% agar, infested with a mixture of 1 st, 2 nd, and 3 rd instar aphid nymphs. I conducted two variants of this experiment. In the first round, there were 40 beetle larvae in the toxic treatment and 20 in the nontoxic treatment and each larva received 15 aphids. In the second round there were 50 beetle larvae per treatment and each larva received 20 aphids. I examined each arena at four time points over 24h, counting and categorizing the remaining aphids as uninjured, injured, or partially consumed. Missing aphids were counted as consumed. I conducted a confirmation experiment to ensure that missing aphids were not escaping from the leaf disk arena. I placed 20 aphids into each of 15 leaf disks, and simulated the experiment, opening and closing the arenas over the course of a day at the same intervals as the actual experiment. There was 99.7% (299/300) retention of the aphids in the arenas, indicating that aphid 11
22 escape is rare, and that it is reasonable to assume missing aphids in the actual experimental rounds can be attributed to consumption. In the first round, aphids were counted after 4, 17, 20, and 24 hours, and in the second round aphids were counted after 4, 16, 20, and 24 hours. I compared the number of consumed aphids over time between toxic and nontoxic treatments using repeated measures ANOVA in JMP10. I also compared the final number of rejected (injured and partially consumed) aphids at 24h between treatments using a t-test in JMP10. Given the slight procedural differences between the two rounds of experiments, each round was analyzed separately. Larval behavior observation After observing a substantial number of half-eaten aphid carcasses and injured aphids in the toxic aphid treatment of the previous experiment, I sought to characterize this apparent rejection behavior exhibited by H. axyridis through direct observation. Direct observation of larval foraging behaviors provides more insight into the rejection behaviors by larvae when encountering an unsuitable food item. I placed neonate H. axyridis larvae into groups of four in 60mm 15mm Petri dishes with DI water cotton balls and ad libitum pea aphids until they became pharate, at which time I moved them into individual Petri dishes with just a DI water cotton ball. I used second instars for experimental observation because these larger, more voracious larvae consumed and/or rejected more aphids per unit time than the first instars used in the previous experiment, providing more behavioral events for observation within a reasonable timeframe. After molting to second instar, larvae were given a minimum of 12
23 six hours to fully sclerotize before being placed individually into enclosed leaf disk arenas with either nontoxic or toxic A. craccivora. A total of 30 mixed age aphid nymphs (second, third, and fourth instar) were provided per arena. I allowed the larvae to forage undisturbed for three hours, and then I observed them for one hour. This timeframe was chosen because the larvae had sufficient time to experience the toxic or nontoxic aphids as a food source prior to monitoring and recording their behavioral responses. I conducted this experiment in a room with only nondirectional overhead lighting, augmented by a circular Daylight Company magnifying lamp that I placed directly over the Petri dishes while they were being observed. I placed the closed Petri dishes on one sheet of white paper. The Petri dishes remained closed for the first three hours, and were uncovered only during the observation period. Tape between the dishes prevented potential cross-contamination of aphids. I conducted observations on either 4 or 6 beetles at a time, for a total of 20 beetles per treatment. One beetle was subsequently excluded from the toxic treatment after escaping the arena. I recorded observed behaviors with a voice recorder and transcribed the data, calculating time spent in 5 activities: Walk, Stand, Drink, Groom, and Aphid Encounter. The latter category was subdivided into either initial acceptance, where the larvae attacked and accepted the aphid, or initial rejection, where the larvae physically touched the aphid but did not attack. For encounters in which the aphid was accepted, I recorded the duration of the feeding bout, and further categorized whether the larvae completely consumed the aphid, or rejected the aphid after some consumption, leaving an injured aphid or a partially consumed carcass. If the feeding bout was still in progress at the end of the observation period, it was categorized as a complete consumption. Some of these events might ultimately have 13
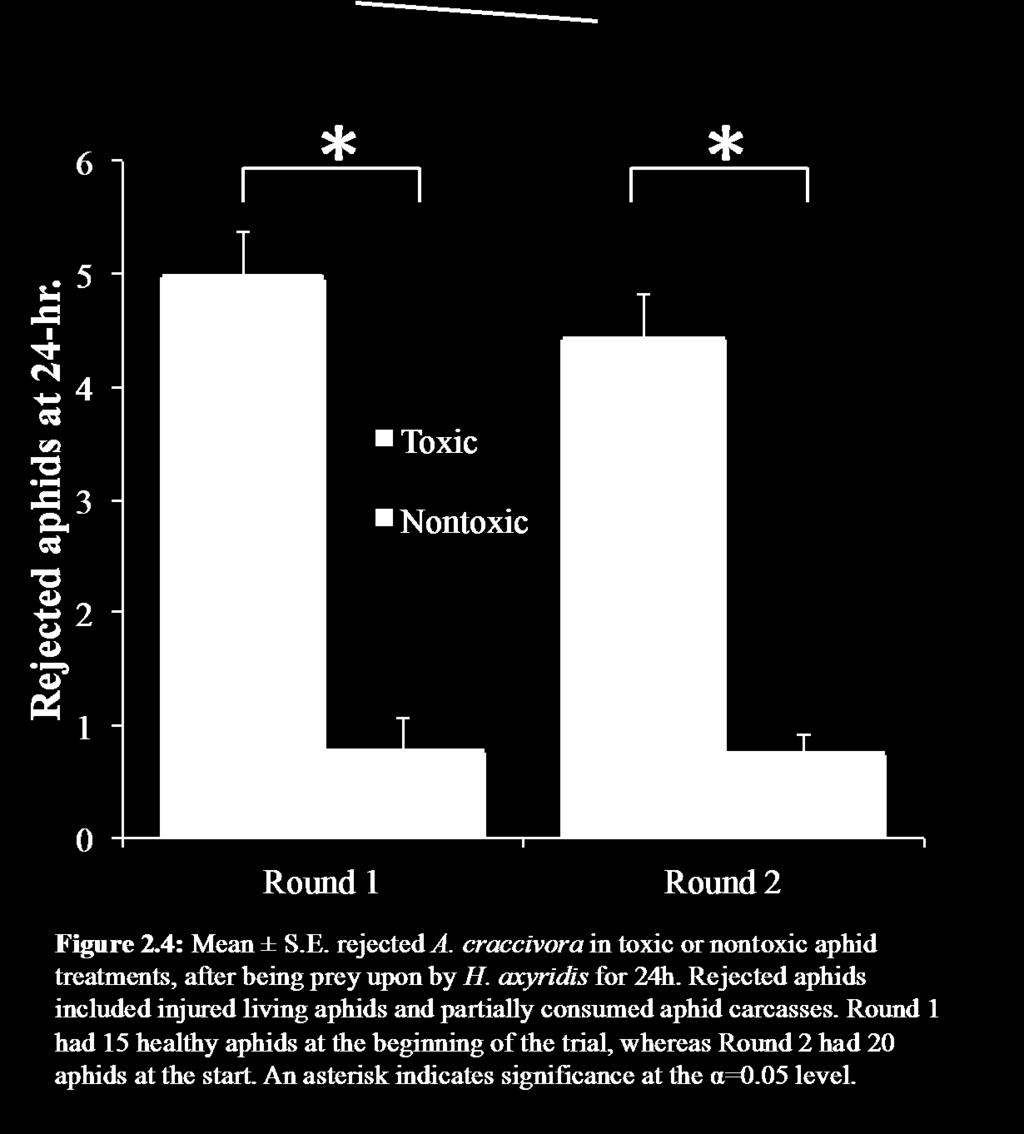
24 ended in a rejection, thus my estimates of rejection frequency are potentially conservative underestimates. The number of aphids that were accepted or rejected, and at what point the rejection occurred were recorded. I compared total time spent eating between treatments using a t-test (JMP 10). I then compared a) the likelihood of a larva to initially reject or accept an aphid, and b) subsequently reject or completely consume the aphid using two separate logistic regressions, with Williams' corrections to account for moderate overdispersion of the data (Arc v1.04) (Williams 1982). Results Adult retention time When comparing adult beetle retention on plants with either toxic or nontoxic aphids, beetle numbers declined over the first six hours (Time: F3,144=19.43, p<0.001; Fig. 2.1), but there was no significant difference between the two aphid treatments (Trt: F1,37=1.09, p=0.068; Time*Trt: F3,144=1.94, p=0.108). However, by the 24h time check, a majority of the beetles placed on the plants with the toxic aphids had left. Most plants with toxic aphids had zero beetles left, and only a few had one beetle (Mean ±S.E.= 0.21 ±0.10 beetles per plant). This was significantly less than the plants with nontoxic aphids, which averaged more than one beetle per plant (1.15 ±0.21 beetles per plant; t26=4.09, p<0.001; Fig. 2.1). Despite reduced beetle retention in the toxic aphid treatment, beetles laid an average of 8.5 ±2.3 eggs in association with toxic aphids. However, this was less than half the number of eggs laid with nontoxic aphids (Mean=20.0 ±4.5 eggs; t28=2.26, p=0.032; Fig. 2.2). 14
25 Larval consumption of aphids over time First instar H. axyridis larvae that were given toxic aphids consumed fewer aphids over 24h than larvae that were given nontoxic aphids. In both rounds of the experiment, there were significant time, toxicity, and time toxicity effects over the 24h, with slower aphid consumption in the toxic treatment than in the nontoxic treatment (Round 1: Trt: F1,57=77.34, p<0.001, Time: F2.3,131.9=284.84, p<0.001, Time*Trt: F2.3,131.9=81.68, p<0.001; Round 2: Trt: F1,46=56.63, p<0.001, Time: F1.7,76.2=289.88, p<0.001, Time*Trt: F1.7,76.2=70.58, p<0.001; Fig. 2.3). At the final 24h time check, the larvae with the nontoxic aphids had consumed 4 the number of aphids as larvae with the toxic aphids in Round 1, and 3 more in Round 2. Additionally, there were more aphids in the injured and partially consumed categories in the toxic treatment than in the nontoxic at the end of the 24h. Both of these categories consisted of aphids that had been wounded but not completely consumed, and thus could be construed to be rejected by the larvae. In both rounds of the experiment, at least 5 more aphids were rejected in the toxic treatment than in the nontoxic treatments (Fig. 2.4; Round 1: t56=9.12, p<0.001; Round 2: t62=9.45, p<0.001). Larval behavioral observation As depicted in Fig. 2.5, larvae in both aphid treatments distributed their time similarly across the five categories during the hour-long observational window. The average eating bout lasted 10.2 ± 1.3 min for larvae in the nontoxic treatment, and 11.5 ± 2.2 min for larvae in the toxic treatment (t80=0.49, p=0.629). 15
26 Overall, the larvae in the toxic treatment encountered a total of 94 aphids, where the larvae in the nontoxic treatment encountered 60. When the larvae in the toxic treatment encountered an aphid, 62% were immediately rejected as a food source, unlike the nontoxic aphids in which only 28% were immediately rejected; the odds of the larvae immediately rejecting an aphid were 3 higher for larva in the toxic than the nontoxic treatment (Fig. 2.6; Wald=2.32, p=0.02). For aphids that were initially accepted as a food source, the odds of subsequent rejection were 20 higher for larvae with toxic than nontoxic aphids (Fig. 2.6; F29=3.00, p=0.016), leaving behind more injured aphids and partially consumed carcasses. Over half of the toxic aphids were ultimately rejected after being initially accepted by the larvae, whereas only 2% of the nontoxic aphids that were initially accepted by larvae were later rejected. Discussion When given the choice, adult female H. axyridis ultimately left patches of unsuitable toxic aphids. Foraging theory predicts that a predator will leave a patch of food once the cost of staying outweighs the benefits, and coccinellids have been used as examples of invertebrates that utilize optimal foraging theory (MacArthur & Pianka 1966; Carter & Dixon 1982). The consumption of toxic aphids is less beneficial to the beetles than consuming the nontoxic aphids, and H. axyridis followed the predictions by leaving more quickly from the patches of toxic aphids than the nontoxic aphids. Food preference of the mother can sometimes be motivated by oviposition preferences (Jensen et al. 2012), which was also seen in H. axyridis, with females laying fewer eggs on the plants with the toxic than nontoxic aphids. Though the time spent in a patch of toxic 16
27 aphids and the number of eggs laid was lower, some eggs were still laid in association with the toxic aphids. Maternal decisions determine the principal food source that first instars will encounter (Refsnider & Janzen 2010). While it might seem illogical for H. axyridis to lay any eggs with an unsuitable food source, there are a number of possible explanations for why this might occur. Other factors, such as the presence or absence of conspecific larvae, can affect the insect's decision-making, resulting in an ultimately poor placement of an egg mass for larval growth and development (Valladares & Lawton 1991; Hemptinne, Dixon & Coffin 1992). Additionally, for H. axyridis, the adult females might still lay eggs near a poor food source due to a temporal limitation, like a limited lifespan or seasonal variation (Hemptinne, Dixon & Coffin 1992; Heimpel & Rosenheim 1998; Díaz-Fleischer & Aluja 2003). Though a location might not be idea for egg oviposition and larval development might be stunted, it is still more beneficial for an adult female to lay eggs where larvae have a chance of survival instead of refraining from laying any eggs at all. Based on these observed egg-laying behaviors of adults, at least some first instar H. axyridis will likely encounter a toxic food source as their first meal. In separate field trials, we have observed very little emigration of first instar H. axyridis, even when on a plant with toxic aphids as the sole food source indicating that the neonate larvae are at the mercy of the mother s oviposition choices (Lenhart et al., unpublished data). Though my field experiments were conducted in a less than natural setting, neonate and first instar larvae have restricted mobility, and the clonal nature of aphids likely limits the variability in toxicity available to newly hatched larvae within a given 17
28 resource patch. Consequently, natural conditions experienced by these larvae may be reasonably analogous to the no-choice experiments I used to examine larval behavior. In these behavioral assays, I found that H. axyridis larvae with toxic aphids ate fewer aphids than the larvae that were feeding on the nontoxic aphids, but continued to try to consume the toxic prey throughout the 24h assay. The larvae in the toxic aphid treatment consumed aphids at a slower rate than those that were in the nontoxic treatment, with an accumulation of aphid carcasses occurring instead as the larvae left behind the half-eaten aphids. This continual attempted consumption reduced the potential defensive benefit of the toxic trait for individual aphids; even if not fully consumed, the toxic aphids likely perished from the attack. Even over the short 24h assay, however, more toxic aphids survived unharmed than nontoxic aphids, and eventually, once the larvae cease consumption or die, unharmed clonal aphid sisters will benefit from the toxic trait. Additionally, older, more mobile H. axyridis larvae may behave similarly to adults, leaving toxic colonies altogether and further promoting the defensive value of the toxic trait. Field trials conducted with older H. axyridis larvae have shown that they quickly leave patches of A. craccivora (Lenhart et al., unpublished data). Discrimination against the toxic aphids is not innate in the beetles, with naïve beetles readily attacking toxic aphids. However, after 3 hours of experience, larvae in the toxic treatment were 3 times more likely to reject an aphid immediately upon encounter, and 20 times more likely to reject an aphid after tasting it than larvae in the nontoxic treatment. The continued attempts suggest that the larvae were motivated, potentially by hunger, and also suggest that the larvae might not readily switch between intensive and extensive foraging strategies. The rejections demonstrate that the larvae are not being 18
29 deterred by visual or volatile signals that can be discerned from a distance; from a human perspective, the two strains of aphids are indistinguishable. The rejection of the first aphids tried occur after the larva bites into the aphids, likely after the beetles have tasted the hemolymph. The ability of the larva to identify the aphids as bad through a contact signal after trying multiple aphids also suggests some level of associative learning. Aphid rejection and learning behaviors have been seen in other coccinellids (Boivin et al. 2010), though the literature on these rejection behaviors is still relatively limited. Future experiments should include examining larval behavior when switching between toxic and nontoxic aphid strains, to determine whether the larvae can distinguish between the two lines. Understanding the behaviors of both the adult and larval life stages of H. axyridis when encountering a toxic prey item can not only be applied to H. axyridis, but to other predators that face prey that might have intraspecific variations in defense, yet are only subtly distinguishable, if at all (Oliver et al. 2003; Vorburger, Gehrer & Rodriguez 2009; Oliver et al. 2012). It is probable that other generalist predators encounter food sources that are not ideal for their development, reproduction, or survival, but might be difficult to distinguish from suitable prey items. Additionally, the rejection of a potential food source opens up the potential for other community effects. Toxic A. craccivora are not being consumed by a dominant coccinellid species; if other, subdominant, species can utilize the toxic aphids as a food source, A. craccivora is a potential refuge food item, allowing coccinellid species to coexist despite niche overlap. This topic is addressed more fully in the next chapter. 19

30 20


31 21

32 22

33 23


34 24

35 25
36 CHAPTER THREE: A WINDOW OF OPPORTUNITY: SUBDOMINANT PREDATORS CAN USE SUBOPTIMAL PREY Introduction Competition often plays a large role in shaping community structure (Menge 1976; Price & Kirkpatrick 2009). Species that are the most successful in securing food and habitat resources can establish themselves as dominant species, potentially excluding other species that are less adept (Fretwell 1969; Goldberg 1987). For these subdominant species, survival in a community then becomes contingent on exploiting alternative resources that cannot or will not be utilized by the dominant species (Hill & Lodge 1994; Messing & Wang 2009). When invasive species that enter novel ecosystems are competitively dominant, they are able to disrupt established community interactions (Blossey & Notzold 1995). Globally, there are many examples of community compositional shifts as a result of introduced species (Mooney & Cleland 2001). The multicolored Asian lady beetle, Harmonia axyridis (Pallas), is a notably dominant invasive predator species. This beetle originated in Asia and has spread to at least 38 new countries since 1988 (Brown et al. 2011b). The ability of H. axyridis to rapidly expand its range and establish itself in novel communities is largely attributed to its success as an intraguild predator and a superior competitor (Lucas, Gagne & Coderre 2002; Ware & Majerus 2008). Additionally, high dispersal capability, multivoltinism, an ability to resist fungal pathogens, and the capability to survive in a wide variety of habitats have been hypothesized to contribute to the dominance of H. axyridis in novel ecosystems (Roy & Brown 2015). Overall, native lady beetle populations and species diversity have been in decline (Harmon, Stephens & 26
37 Losey 2007), and it is thought that H. axyridis has played a role in the decreasing biodiversity in some coccinellid communities (Brown et al. 2011a; Bahlai et al. 2015). Harmonia axyridis is an aphidophagous generalist, yet there are some aphids that cause delayed growth and mortality when consumed by the beetles, e.g. Megoura viciae Buckton and Aulacorthum magnolia Essig et Kuwana (Tsaganou et al. 2004; Fukunaga & Akimoto 2007). Interestingly, the cowpea aphid, Aphis craccivora Koch, varies in suitability as food for H. axyridis, with some strains causing toxicity while other strains support lady beetle development and reproduction (Hukusima & Kamei 1970; Kamo, Tokuoka & Miyazaki 2010). Strains found on black locust (Robinia pseudoacacia L.), have been documented to be toxic, inducing 100% mortality in H. axyridis larvae (Hukusima & Kamei 1970; White et al. 2016). In contrast, A. craccivora strains that originate from alfalfa (Medicago sativa L.) are not toxic to H. axyridis larvae (White et al. 2016). This difference between the strains is a result of intrinsic intraspecific variation, and not a function of host plant chemistry. The aphid strains that originated on locust are consistently and heritably toxic to H. axyridis, even after many generations of rearing on alternate host plant species (White et al. 2016). Despite the strong negative effects that the locust-origin aphids have on H. axyridis, the toxicity might not be ubiquitous across coccinellid predator species. Previous studies on the suitability of A. craccivora as a food source have been conducted with other coccinellid species, often with the results suggesting that they are an acceptable prey item that yields adult coccinellids (Omkar & Srivastava 2003; Omkar & Mishra 2005; Ferrer, Dixon & Hemptinne 2008). However, it is not clear which strains of aphids were evaluated in these trials. They may have used intrinsically suitable A. 27
38 craccivora that even H. axyridis would do well on, or they may have used strains that would be toxic to H. axyridis, thus indicating that these other lady beetles are less susceptible to the toxic strains. If other coccinellid species can use these aphids as a food source, the presence of selectively toxic aphids in an environment could mitigate competitive differentials between H. axyridis and other subdominant lady beetle species. Here, I examined whether locust-origin A. craccivora is suitable food for several other coccinellid species. By determining if these aphids could be exploited by other lady beetle species, I ultimately hope to better predict how selectively-toxic aphids might structure coccinellid community composition. Such differentially suitable food sources can facilitate niche partitioning, predator coexistence, and diversity, which in turn can provide more stable ecosystem services (Letourneau et al. 2011). Materials and Methods We evaluated the development and survival of six lady beetle species: Anatis labiculata (Say), Coccinella septempunctata L., Coleomegilla maculata DeGeer, Cycloneda munda (Say), Hippodamia convergens Guerin, and Hippodamia parenthesis (Say) (Table 3.1). Each species naturally occurs with both A. craccivora and H. axyridis in the field. Wild caught beetles were collected from Lexington, KY, USA in 2014 and 2015 and brought into the lab and kept in colony. The beetles were grouped by species and life stage in Petri dishes (100mm 25mm). Both juvenile and adult beetles were fed pea aphids (Acyrthosiphum pisum Harris). When mating occurred between adult beetles, the paired male and female were removed from the colony and placed in their own Petri dish with folded paper for egg deposition. Egg papers were regularly removed from the 28
39 parents Petri dish, to prevent cannibalism. One incubator (25 C, 16h:8h light:dark, 65% humidity) was used both to house the lady beetle colonies and to conduct rearing experiments. All aphids (A. craccivora and Ac. pisum) also originated from clones originally collected in Lexington, Kentucky, USA and were maintained in colonies on fava bean (Vicia faba) in the laboratory at ambient room temperature. Aphis craccivora clones were initially collected from either black locust or alfalfa as described in Wagner et al. (2015). To date, all clones collected from black locust have been toxic to H. axyridis, and all clones collected from alfalfa are nontoxic to H. axyridis (White et al. 2016). The toxicity status of the two aphid strains is unknown for non-harmonia coccinellid species, as such, hereafter, the locust-origin A. craccivora will be referred to as L-strain and the alfalfaorigin A. craccivora will be referred to as A-strain. For each lady beetle species, we compared beetle development time and survival on the two strains of A. craccivora in no-choice environments. Neonate larvae were removed from their egg mass before sibling cannibalism could occur and were placed individually in Petri dishes (35mm 10mm) that had an excised circle of fava bean leaf embedded in 1% agar. Each larva was randomly assigned to an aphid treatment. Three lady beetle species (C. septempunctata, Co. maculata, and Cy. munda) had one of two treatments: either L-strain or A-strain A. craccivora. The remaining three species (A. labiculata, H. convergens, and H. parenthesis) also included a pea aphid control treatment to control for overall poor survival. The larvae were fed their assigned aphids ad libitum for the duration of development, and were monitored daily for mortality and developmental stage. Once the beetles reached the third instar, they were moved to larger 29
40 Petri dishes (60mm 15mm) and provided with a cotton ball soaked in DI water, along with their aphid treatment. For the beetles that survived to adulthood, teneral adults were allowed to sclerotize for one day before weighing to the nearest milligram. Sex of each adult beetle was determined through mating observations of the adults. Sex did not statistically affect the differences between treatments, and was removed from the analysis. Sample size varied among species. Hippodamia parenthesis was included in trials, but excluded from statistical analysis due to low sample size. For the other species, survival to adulthood among treatments was statistically compared using either chisquared contingency table analysis, or Fisher's exact test. Time until pupation and adult weight, being normally distributed and exhibiting equal variance, were compared between treatments with t-tests for the beetle species that had sufficient sample size in JMP10. Results In contrast with H. axyridis, which experiences 100% mortality when exposed to L-strain A. craccivora, the tested lady beetles showed a wide range of tolerance. Two species, Cy. munda and Co. maculata, were only slightly affected by the L-strain of A. craccivora. For Cy. munda, all of the beetles reared on both L-strain (n=13/13) and A- strain (n=14/14) survived to adulthood (Figure 3.1A). There was a slight delay in development, with larvae reared on the L-strain taking about 10% longer to pupate than those on the A-strain (L-strain mean ± S.E. = 11.7 ± 0.2 d, A-strain = 10.8 ± 0.2 d; t19=2.97, p=0.008). However, there was no significant difference between the adult weights of Cy. munda reared on the two treatments (Figure 3.2A; t18=0.59, p=0.562). 30
41 Similarly, Co. maculata experienced greater than 93% survival on both strains of the aphid (Figure 3.1B; χ 2 =0.06, d.f.=1, p=0.813). However, C. maculata experienced slower development on L-strain than A-strain A. craccivora, with L-strain taking 12% longer to develop (L-strain = 18.5 ± 2.3 d, A-strain = 16.6 ± 2.1 d ; t46=2.90, p=0.006). Though the development was slower, there was no difference in the adult weight between treatments (Figure 3.2B; t45=1.04, p=0.304). Coccinella septempunctata showed a moderate response to L-strain A. craccivora. There was a trend towards reduced survival on the L-strain, with 76.9% (n=10/13) surviving, relative to 100% (n=14/14) survival on A-strain (Figure 3.1C; Fisher's exact test p=0.098). The beetles feeding on the L-strain aphids took about one day longer (7.5%) to pupate than the beetles feeding on the A-strain (L-strain = 15.9 ± 0.46 d, A- strain = 14.7 ± 0.61 d; t11=2.40, p=0.035), and showed a 24% reduction in adult weight (Figure 3.2C; t21=4.32, p<0.001). The remaining three species, A. labiculata, H. convergens, and H. parenthesis, had generally poor survival, and additionally included a pea aphid control treatment. For A. labiculata, there was 0% (n=0/13) survival on the L-strain, but only 15% (n=2/13) survival on the A-strain and 50% (n=6/12) on the pea aphid control (Figure 3.1D; Fisher s exact test for L-strain vs. A-strain A. craccivora p=0.48, Fisher s exact test for A. craccivora vs. Ac. pisum p=0.007). This level of mortality on L-strain aphids was equivalent to that seen previously in H. axyridis (White et al. 2016), but for A. labiculata, larvae showed some development on L-strain A. craccivora before mortality occurred. When feeding on the L-strain, 54% of beetles survived into the second instar, and 23% survived to the third, with an average survival time of 10.8±1.1 days. This contrasts with 31
42 previously reported H. axyridis results, in which larvae rarely survived to the second instar (White et al. 2016). Due to the low survival across all treatments, there was too small of a sample size for a statistical comparison of development time or adult weight, though they tended to be slower to pupate and weigh less on the A-strain A. craccivora when compared to the Ac. pisum control (Time to pupation: A-strain = 16.5 ± 0.50 d, Control = 15.2 ± 0.47 d). The fifth species, H. convergens, also performed poorly in all treatments. On the L-strain, 6.67% (n=2/30) survived, and on the A-strain 3.8% (n=1/26) survived to adulthood (Figure 3.1E; Fisher s exact test p=1.00). Even in the control pea aphid treatment, only 34.6% (n=9/26) survived, though the beetles in the control aphid treatments had increased survival over those on both A. craccivora strains when the strains were pooled (Fisher s exact test p=0.001). Additionally, the development times and adult weights of those on the A. craccivora treatments tended to be slower and lower than the survivors on the control treatment, though there were too few survivors for statistical analysis (L-strain = 19.0 ± 1 d, A-strain = 20 d (n=1), Control = 16.0 ± 0.42 d; Figure 3.2E). In the last species, H. parenthesis, I was only able to evaluate 12 beetles. Similar to H. convergens, survival was low across all treatments: 0% (n=0/2) survived on the control treatment, 40% (n=2/5) survived on the L-strain, and 20% (n=1/5) survived on the A-strain. Low sample size precluded this species from both development and adult weight comparisons. The low survival across all treatments suggests that A. craccivora, no matter the strain, is a poor food choice for A. labiculata, H. convergens and H. parenthesis. 32
43 Discussion The coccinellid species I examined varied in their ability to use the L-strain of A. craccivora, but all performed better than H. axyridis. Three species, Cy. munda, Co. maculata and C. septempunctata, showed slight effects; all three species had slightly slower development on the L-strain, though most beetles survived to adulthood on both strains of A. craccivora. Only C. septempunctata demonstrated a lower adult weight when feeding on the L-strain. Lower adult weight, especially in females, can imply lower adult fitness (Honěk 1993). The remaining three beetle species, H. convergens, H. parenthesis, and A. labiculata, performed poorly on L-strain A. craccivora, but for all three species this seemed more a reflection of generally poor adaptation to the rearing environment. Each of these three species had a pea aphid control group, which is frequently used for rearing coccinellids in a laboratory setting due to its status as an essential food source for many coccinellid species (Obrycki & Orr 1990; Hodek & Evans 2012); even on pea aphids, survival was low for all species (0-50%). The overall lack of survival to adulthood makes it difficult to compare among the treatments. While A. labiculata seemed to show a particularly negative response to L-strain A. craccivora versus the A-strain, it is still worth noting that there was some development past the first instar when feeding on the L- strain. Aphis craccivora might therefore be considered an "alternative" food source for A. labiculata, one that facilitates some survival but not full development (Hodek & Evans 2012). For both H. convergens and H. parenthesis, there is no indication that either species reacts more poorly to L-strain than A-strain. Despite the overall low survival for both H. convergens and H. parenthesis, some larvae survived to adulthood on the L- 33
44 strain. It is not unusual for H. convergens to exhibit low survival on pea aphids in the laboratory, suggesting that the species might be sensitive to an unknown component of lab-rearing conditions that might have contributed to poor survival (Costopoulos et al. 2014). Given how few of the tested beetle species truly responded to the toxicity of L- strain A. craccivora, it is debatable whether this strain of aphid should be considered to possess an evolved defensive trait. Typically, indigestible or toxic prey items present some sort of warning to the predator species, e.g. aposematic coloration, and are broadly protected against any number of generalist predators (Berenbaum & Miliczky 1984). Any defensive cues or visual differences that might be present between the strains of A. craccivora have yet to be discovered. After determining that the toxic effects are relatively restricted to H. axyridis, it might be more appropriate to consider the effect as a deficiency on the part of H. axyridis. Regardless of mechanism, the ultimate fate of H. axyridis consuming L-strain A. craccivora is mortality. Our results demonstrate that L- strain A. craccivora has differential effects on different members of the coccinellid community, and the presence or absence of selectively toxic food sources, such as L- strain A. craccivora, has the potential to determine the composition of the lady beetle community. Harmonia axyridis is one of the most dominant species within coccinellid communities (Hesler, Kieckhefer & Catangui 2004), and a food resource that they are unable to use becomes one that other less dominant species might be able to exploit. My research demonstrates that despite belonging to similar size classes, generalist predators don t necessarily have overlapping food sources. While niche partitioning is well studied 34
45 in herbivores, it is not a broadly applied concept to generalist predator species; a suboptimal resource that serves as a refuge resource can facilitate broader niche partitioning among the dominant and subdominant predator species (Holt & Lawton 1993; Messing & Wang 2009). Niche partitioning improves the potential for coexistence within a community and aids in maintaining greater predator biodiversity, which in turn is associated with greater ecosystem services and improved biological control (Letourneau et al. 2011). 35
46 Table 3.1: Lady beetle natural history (Honek & Hodek 1996) Lady beetle Species Status in North America Habitat Voltinism Harmonia axyridis Anatis labiculata Non-native Native Semi-arboreal, Habitat generalist Arboreal, Habitat specialist Multivoltine Univoltine Coccinella septempunctata Non-native Field crops, Habitat generalist Multivoltine Coleomegilla maculata Native Field crops, Habitat generalist Multivoltine Cycloneda munda Native Field crops, Habitat generalist Multivoltine Hippodamia convergens Native Field crops, Habitat generalist Multivoltine Hippodamia parenthesis Native Field crops, Habitat generalist Multivoltine 36

47 37


48 38
MAGNETIC PROPERTIES OF Nb/Ni SUPERCONDUCTING / FERROMAGNETIC MULTILAYERS
 University of Kentucky UKnowledge Theses and Dissertations--Physics and Astronomy Physics and Astronomy 2012 MAGNETIC PROPERTIES OF Nb/Ni SUPERCONDUCTING / FERROMAGNETIC MULTILAYERS Sergiy A. Kryukov University
University of Kentucky UKnowledge Theses and Dissertations--Physics and Astronomy Physics and Astronomy 2012 MAGNETIC PROPERTIES OF Nb/Ni SUPERCONDUCTING / FERROMAGNETIC MULTILAYERS Sergiy A. Kryukov University
General comments about aphid biological control
 General comments about aphid biological control Aphid advantages: Rapid colonization (winged forms) followed by rapid reproduction (wingless forms) The aphid s best defense is its reproductive rate Results
General comments about aphid biological control Aphid advantages: Rapid colonization (winged forms) followed by rapid reproduction (wingless forms) The aphid s best defense is its reproductive rate Results
Comparison of the wing polyphenic response of pea aphids (Acyrthosiphon pisum) to crowding and predator cues
 to crowding and predator cues") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Brigitte Tenhumberg Papers Papers in the Biological Sciences 2014 Comparison of the wing polyphenic response of pea aphids
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Brigitte Tenhumberg Papers Papers in the Biological Sciences 2014 Comparison of the wing polyphenic response of pea aphids
Grade 7 Lesson Instructions Friend or Foe? Preparation: Background information: Activity:
 Instructions Friend or Foe? You can use monarchs to teach about many things! Stone Mountain Memorial Association (SMMA) uses the monarch butterfly to help students apply their knowledge in other contexts
Instructions Friend or Foe? You can use monarchs to teach about many things! Stone Mountain Memorial Association (SMMA) uses the monarch butterfly to help students apply their knowledge in other contexts
Biology Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod. Introduction
 1 Biology 317 - Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod Introduction The determination of how natural selection acts in contemporary populations constitutes
1 Biology 317 - Principles of Ecology Oct. 20 and 27, 2011 Natural Selection on Gall Flies of Goldenrod Introduction The determination of how natural selection acts in contemporary populations constitutes
Unit 6 Populations Dynamics
 Unit 6 Populations Dynamics Define these 26 terms: Commensalism Habitat Herbivory Mutualism Niche Parasitism Predator Prey Resource Partitioning Symbiosis Age structure Population density Population distribution
Unit 6 Populations Dynamics Define these 26 terms: Commensalism Habitat Herbivory Mutualism Niche Parasitism Predator Prey Resource Partitioning Symbiosis Age structure Population density Population distribution
BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences
 BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences D. POPULATION & COMMUNITY DYNAMICS Week 13. Herbivory, predation & parasitism: Lecture summary: Predation:
BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences D. POPULATION & COMMUNITY DYNAMICS Week 13. Herbivory, predation & parasitism: Lecture summary: Predation:
Social Insects. Social Insects. Subsocial. Social Insects 4/9/15. Insect Ecology
 Social Insects Social Insects Insect Ecology Sociality evolved multiple times in insects Much of Earth s fauna consists of social insects They play major roles in entire ecosystems Proliferation of ants
Social Insects Social Insects Insect Ecology Sociality evolved multiple times in insects Much of Earth s fauna consists of social insects They play major roles in entire ecosystems Proliferation of ants
Social Insects. Insect Ecology
 Social Insects Insect Ecology Social Insects Sociality evolved multiple times in insects Much of Earth s fauna consists of social insects They play major roles in entire ecosystems Proliferation of ants
Social Insects Insect Ecology Social Insects Sociality evolved multiple times in insects Much of Earth s fauna consists of social insects They play major roles in entire ecosystems Proliferation of ants
ACCURACY OF MODELS FOR PREDICTING PHENOLOGY OF BLACKHEADED FIREWORM AND IMPLICATIONS FOR IMPROVED PEST MANAGEMENT
 ACCURACY OF MODELS FOR PREDICTING PHENOLOGY OF BLACKHEADED FIREWORM AND IMPLICATIONS FOR IMPROVED PEST MANAGEMENT Stephen D. Cockfield and Daniel L. Mahr Department of Entomology University of Wisconsin-Madison
ACCURACY OF MODELS FOR PREDICTING PHENOLOGY OF BLACKHEADED FIREWORM AND IMPLICATIONS FOR IMPROVED PEST MANAGEMENT Stephen D. Cockfield and Daniel L. Mahr Department of Entomology University of Wisconsin-Madison
Ecology Symbiotic Relationships
 Ecology Symbiotic Relationships Overview of the Co-evolution and Relationships Exhibited Among Community Members What does Symbiosis mean? How do we define Symbiosis? Symbiosis in the broadest sense is
Ecology Symbiotic Relationships Overview of the Co-evolution and Relationships Exhibited Among Community Members What does Symbiosis mean? How do we define Symbiosis? Symbiosis in the broadest sense is
FIELD TEST OF THE EFFECTIVENESS OF LADYBIRDS IN CONTROLLING APHIDS
 FIELD TEST OF THE EFFECTIVENESS OF LADYBIRDS IN CONTROLLING APHIDS Pavel KINDLMANN 1, Hironori YASUDA 2, Yukie KAJITA 2, and Anthony F.G. DIXON 3 1 Inst. Landscape Ecol. AS CR University of South Bohemia
FIELD TEST OF THE EFFECTIVENESS OF LADYBIRDS IN CONTROLLING APHIDS Pavel KINDLMANN 1, Hironori YASUDA 2, Yukie KAJITA 2, and Anthony F.G. DIXON 3 1 Inst. Landscape Ecol. AS CR University of South Bohemia
Chapter 6 Reading Questions
 Chapter 6 Reading Questions 1. Fill in 5 key events in the re-establishment of the New England forest in the Opening Story: 1. Farmers begin leaving 2. 3. 4. 5. 6. 7. Broadleaf forest reestablished 2.
Chapter 6 Reading Questions 1. Fill in 5 key events in the re-establishment of the New England forest in the Opening Story: 1. Farmers begin leaving 2. 3. 4. 5. 6. 7. Broadleaf forest reestablished 2.
Approximate Pacing for First Grade Insects and Plants Unit
 Approximate Pacing for First Grade Insects and Plants Unit p.1 = Part 1 p.2 = Part 2 p.3 = Part 3 The schedule for this unit is almost COMPLETELY dependent on what the living organisms are doing and where
Approximate Pacing for First Grade Insects and Plants Unit p.1 = Part 1 p.2 = Part 2 p.3 = Part 3 The schedule for this unit is almost COMPLETELY dependent on what the living organisms are doing and where
Lecture 8 Insect ecology and balance of life
 Lecture 8 Insect ecology and balance of life Ecology: The term ecology is derived from the Greek term oikos meaning house combined with logy meaning the science of or the study of. Thus literally ecology
Lecture 8 Insect ecology and balance of life Ecology: The term ecology is derived from the Greek term oikos meaning house combined with logy meaning the science of or the study of. Thus literally ecology
Community Structure. Community An assemblage of all the populations interacting in an area
 Community Structure Community An assemblage of all the populations interacting in an area Community Ecology The ecological community is the set of plant and animal species that occupy an area Questions
Community Structure Community An assemblage of all the populations interacting in an area Community Ecology The ecological community is the set of plant and animal species that occupy an area Questions
Chapter 6 Population and Community Ecology
 Chapter 6 Population and Community Ecology Friedland and Relyea Environmental Science for AP, second edition 2015 W.H. Freeman and Company/BFW AP is a trademark registered and/or owned by the College Board,
Chapter 6 Population and Community Ecology Friedland and Relyea Environmental Science for AP, second edition 2015 W.H. Freeman and Company/BFW AP is a trademark registered and/or owned by the College Board,
What is insect forecasting, and why do it
 Insect Forecasting Programs: Objectives, and How to Properly Interpret the Data John Gavloski, Extension Entomologist, Manitoba Agriculture, Food and Rural Initiatives Carman, MB R0G 0J0 Email: jgavloski@gov.mb.ca
Insect Forecasting Programs: Objectives, and How to Properly Interpret the Data John Gavloski, Extension Entomologist, Manitoba Agriculture, Food and Rural Initiatives Carman, MB R0G 0J0 Email: jgavloski@gov.mb.ca
IMPACT OF INTRASPECIFIC AND INTRAGUILD PREDATION ON PREDATOR INVASION AND COEXISTENCE: CAN EXOTIC LADYBEETLES DISPLACE NATIVE SPECIES?
 Van Rijn et al. IMPACT OF INTRASPECIFIC AND INTRAGUILD PREDATION ON PREDATOR INVASION AND COEXISTENCE: CAN EXOTIC LADYBEETLES DISPLACE NATIVE SPECIES? Paul C. J. VAN RIJN 1, Giovanni BURGIO 2, and Matt
Van Rijn et al. IMPACT OF INTRASPECIFIC AND INTRAGUILD PREDATION ON PREDATOR INVASION AND COEXISTENCE: CAN EXOTIC LADYBEETLES DISPLACE NATIVE SPECIES? Paul C. J. VAN RIJN 1, Giovanni BURGIO 2, and Matt
Testing for Grazer Adaptation to Toxic Algae
 Testing for Grazer Adaptation to Toxic Algae by Michael B. Finiguerra, Hans G. Dam, and David E. Avery Part I Introduction and Background Phytoplankton, microscopic single-celled algae, are natural components
Testing for Grazer Adaptation to Toxic Algae by Michael B. Finiguerra, Hans G. Dam, and David E. Avery Part I Introduction and Background Phytoplankton, microscopic single-celled algae, are natural components
Population Ecology. Study of populations in relation to the environment. Increase population size= endangered species
 Population Basics Population Ecology Study of populations in relation to the environment Purpose: Increase population size= endangered species Decrease population size = pests, invasive species Maintain
Population Basics Population Ecology Study of populations in relation to the environment Purpose: Increase population size= endangered species Decrease population size = pests, invasive species Maintain
Survival of the Sweetest
 Biology Survival of the Sweetest A Tasty Tale of Natural Selection and Community Dynamics MATERIALS AND RESOURCES EACH GROUP teacher-provided materials 2 dice TEACHER bags, brown paper, small 3 bags Skittles,
Biology Survival of the Sweetest A Tasty Tale of Natural Selection and Community Dynamics MATERIALS AND RESOURCES EACH GROUP teacher-provided materials 2 dice TEACHER bags, brown paper, small 3 bags Skittles,
BIOS 6150: Ecology Dr. Stephen Malcolm, Department of Biological Sciences
 BIOS 6150: Ecology Dr. Stephen Malcolm, Department of Biological Sciences Week 7: Dynamics of Predation. Lecture summary: Categories of predation. Linked prey-predator cycles. Lotka-Volterra model. Density-dependence.
BIOS 6150: Ecology Dr. Stephen Malcolm, Department of Biological Sciences Week 7: Dynamics of Predation. Lecture summary: Categories of predation. Linked prey-predator cycles. Lotka-Volterra model. Density-dependence.
Pollinator Activity #1: How to Raise a Butterfly
 How to Raise a Butterfly How to Raise a Butterfly A Conversation Where do you most often see butterflies? What are they doing when you see them? Have you ever seen a butterfly in another form? They have
How to Raise a Butterfly How to Raise a Butterfly A Conversation Where do you most often see butterflies? What are they doing when you see them? Have you ever seen a butterfly in another form? They have
Community and Population Ecology Populations & Communities Species Diversity Sustainability and Environmental Change Richness and Sustainability
 1 2 3 4 Community and Population Ecology Chapter 6 Populations & Communities Biosphere> ecosystems> communities> populations> individuals A population is all of the individuals of the same species in a
1 2 3 4 Community and Population Ecology Chapter 6 Populations & Communities Biosphere> ecosystems> communities> populations> individuals A population is all of the individuals of the same species in a
Biology 11 Unit 1: Fundamentals. Lesson 1: Ecology
 Biology 11 Unit 1: Fundamentals Lesson 1: Ecology Objectives In this section you will be learning about: ecosystem structure energy flow through an ecosystem photosynthesis and cellular respiration factors
Biology 11 Unit 1: Fundamentals Lesson 1: Ecology Objectives In this section you will be learning about: ecosystem structure energy flow through an ecosystem photosynthesis and cellular respiration factors
Chapter 6 Population and Community Ecology. Thursday, October 19, 17
 Chapter 6 Population and Community Ecology Module 18 The Abundance and Distribution of After reading this module you should be able to explain how nature exists at several levels of complexity. discuss
Chapter 6 Population and Community Ecology Module 18 The Abundance and Distribution of After reading this module you should be able to explain how nature exists at several levels of complexity. discuss
Ch.5 Evolution and Community Ecology How do organisms become so well suited to their environment? Evolution and Natural Selection
 Ch.5 Evolution and Community Ecology How do organisms become so well suited to their environment? Evolution and Natural Selection Gene: A sequence of DNA that codes for a particular trait Gene pool: All
Ch.5 Evolution and Community Ecology How do organisms become so well suited to their environment? Evolution and Natural Selection Gene: A sequence of DNA that codes for a particular trait Gene pool: All
Investigating Use of Biocontrol Agents to Control Spotted Knapweed
 Investigating Use of Biocontrol Agents to Control Spotted Knapweed Target Grade Level: 5 th Created and Adapted by: Rachel Loehman UNIVERSITY OF MONTANA GK-12 PROGRAM 1 Investigating Use of Biocontrol
Investigating Use of Biocontrol Agents to Control Spotted Knapweed Target Grade Level: 5 th Created and Adapted by: Rachel Loehman UNIVERSITY OF MONTANA GK-12 PROGRAM 1 Investigating Use of Biocontrol
Chapter 44. Table of Contents. Section 1 Development of Behavior. Section 2 Types of Animal Behavior. Animal Behavior
 Animal Behavior Table of Contents Section 1 Development of Behavior Section 2 Types of Animal Behavior Section 1 Development of Behavior Objectives Identify four questions asked by biologists who study
Animal Behavior Table of Contents Section 1 Development of Behavior Section 2 Types of Animal Behavior Section 1 Development of Behavior Objectives Identify four questions asked by biologists who study
6 2 Insects and plants
 6 2 Insects and plants Insect DIY 1. Find plant habitat 2. Find plant 3. Accept plant 4. Eat survive, reproduce Plant characteristics Shape structure Mechanical defenses trichomes Chemical defenses sap,
6 2 Insects and plants Insect DIY 1. Find plant habitat 2. Find plant 3. Accept plant 4. Eat survive, reproduce Plant characteristics Shape structure Mechanical defenses trichomes Chemical defenses sap,
4. Identify one bird that would most likely compete for food with the large tree finch. Support your answer. [1]
![4. Identify one bird that would most likely compete for food with the large tree finch. Support your answer. [1]](/thumbs/84/91138454.jpg "4. Identify one bird that would most likely compete for food with the large tree finch. Support your answer. [1]") Name: Topic 5B 1. A hawk has a genetic trait that gives it much better eyesight than other hawks of the same species in the same area. Explain how this could lead to evolutionary change within this species
Name: Topic 5B 1. A hawk has a genetic trait that gives it much better eyesight than other hawks of the same species in the same area. Explain how this could lead to evolutionary change within this species
University of Groningen. Seasonal timing in a warming world Salis, Lucia
 University of Groningen Seasonal timing in a warming world Salis, Lucia IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the
University of Groningen Seasonal timing in a warming world Salis, Lucia IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the
PREDATION POTENTIAL OF A BIOCONTROL AGENT, HIPPODAMIA VARIEGATA AGAINST THE APHID, APHIS GOSSYPII ABSTRACT
 PREDATION POTENTIAL OF A BIOCONTROL AGENT, HIPPODAMIA VARIEGATA AGAINST THE APHID, APHIS GOSSYPII Ahmad Pervez 1*, Pooja 1 & Hakan Bozdoğan 2 1 Biocontrol Laboratory, Department of Zoology, Radhey Hari
PREDATION POTENTIAL OF A BIOCONTROL AGENT, HIPPODAMIA VARIEGATA AGAINST THE APHID, APHIS GOSSYPII Ahmad Pervez 1*, Pooja 1 & Hakan Bozdoğan 2 1 Biocontrol Laboratory, Department of Zoology, Radhey Hari
Bio112 Home Work Community Structure
 Bio112 Home Work Community Structure Multiple Choice Identify the choice that best completes the statement or answers the question. 1. All of the populations of different species that occupy and are adapted
Bio112 Home Work Community Structure Multiple Choice Identify the choice that best completes the statement or answers the question. 1. All of the populations of different species that occupy and are adapted
Yakın Doğu Üniversitesi Mimarlık Fakültesi Peyzaj Mimarlığı Bölümü. PM 317 Human and Environment Assoc. Prof. Dr. Salih GÜCEL
 Yakın Doğu Üniversitesi Mimarlık Fakültesi Peyzaj Mimarlığı Bölümü PM 317 Human and Environment Assoc. Prof. Dr. Salih GÜCEL Ecology & Ecosystems Principles of Ecology Ecology is the study of the interactions
Yakın Doğu Üniversitesi Mimarlık Fakültesi Peyzaj Mimarlığı Bölümü PM 317 Human and Environment Assoc. Prof. Dr. Salih GÜCEL Ecology & Ecosystems Principles of Ecology Ecology is the study of the interactions
Invasive Species Test. 30 Stations 90 seconds each -or- 15 stations (2/seat) 3 minutes each
 3 minutes each") Invasive Species Test 30 Stations 90 seconds each -or- 15 stations (2/seat) 3 minutes each Station 1 A. The insect transmits Huanglongbing killing the plant upon which it feeds. How was this species introduced
Invasive Species Test 30 Stations 90 seconds each -or- 15 stations (2/seat) 3 minutes each Station 1 A. The insect transmits Huanglongbing killing the plant upon which it feeds. How was this species introduced
Effect of Prey Limitation on Competitive Interactions Between a Native Lady Beetle, Coccinella novemnotata
 Effect of Prey Limitation on Competitive Interactions Between a Native Lady Beetle, Coccinella novemnotata, and an Invasive Lady Beetle, Coccinella septempunctata (Coleoptera: Coccinellidae) Author(s):
Effect of Prey Limitation on Competitive Interactions Between a Native Lady Beetle, Coccinella novemnotata, and an Invasive Lady Beetle, Coccinella septempunctata (Coleoptera: Coccinellidae) Author(s):
ENVE203 Environmental Engineering Ecology (Nov 05, 2012)
") ENVE203 Environmental Engineering Ecology (Nov 05, 2012) Elif Soyer Ecosystems and Living Organisms Population Density How Do Populations Change in Size? Maximum Population Growth Environmental Resistance
ENVE203 Environmental Engineering Ecology (Nov 05, 2012) Elif Soyer Ecosystems and Living Organisms Population Density How Do Populations Change in Size? Maximum Population Growth Environmental Resistance
Ecology Regulation, Fluctuations and Metapopulations
 Ecology Regulation, Fluctuations and Metapopulations The Influence of Density on Population Growth and Consideration of Geographic Structure in Populations Predictions of Logistic Growth The reality of
Ecology Regulation, Fluctuations and Metapopulations The Influence of Density on Population Growth and Consideration of Geographic Structure in Populations Predictions of Logistic Growth The reality of
BIOL EVOLUTION OF QUANTITATIVE CHARACTERS
 1 BIOL2007 - EVOLUTION OF QUANTITATIVE CHARACTERS How do evolutionary biologists measure variation in a typical quantitative character? Let s use beak size in birds as a typical example. Phenotypic variation
1 BIOL2007 - EVOLUTION OF QUANTITATIVE CHARACTERS How do evolutionary biologists measure variation in a typical quantitative character? Let s use beak size in birds as a typical example. Phenotypic variation
Arthropod Containment in Plant Research. Jian J Duan & Jay Bancroft USDA ARS Beneficial Insects Research Unit Newark, Delaware
 Arthropod Containment in Plant Research Jian J Duan & Jay Bancroft USDA ARS Beneficial Insects Research Unit Newark, Delaware What we do at USDA ARS BIIRU - To develop biological control programs against
Arthropod Containment in Plant Research Jian J Duan & Jay Bancroft USDA ARS Beneficial Insects Research Unit Newark, Delaware What we do at USDA ARS BIIRU - To develop biological control programs against
Charles Darwin published The Origin of Species in In his book,
 Survival of the Sweetest Biology Survival of the Sweetest Biology A Tasty Tale of Natural Selection and Community Dynamics MATERIALS teacher-provided materials Charles Darwin published The Origin of Species
Survival of the Sweetest Biology Survival of the Sweetest Biology A Tasty Tale of Natural Selection and Community Dynamics MATERIALS teacher-provided materials Charles Darwin published The Origin of Species
Gibbs: The Investigation of Competition
 ESSAI Volume 5 Article 21 1-1-2007 The Investigation of Competition Between Eurosta Solidaginis (Fitch) and Rhopalomyia Solidaginis (Loew), Two Gall makers of Solidago Altissima (Asteraceae) Jessica Gibbs
ESSAI Volume 5 Article 21 1-1-2007 The Investigation of Competition Between Eurosta Solidaginis (Fitch) and Rhopalomyia Solidaginis (Loew), Two Gall makers of Solidago Altissima (Asteraceae) Jessica Gibbs
Understanding Populations Section 1. Chapter 8 Understanding Populations Section1, How Populations Change in Size DAY ONE
 Chapter 8 Understanding Populations Section1, How Populations Change in Size DAY ONE What Is a Population? A population is a group of organisms of the same species that live in a specific geographical
Chapter 8 Understanding Populations Section1, How Populations Change in Size DAY ONE What Is a Population? A population is a group of organisms of the same species that live in a specific geographical
BIOS 3010: Ecology Lecture 8: Predator foraging & prey defense. 2. Predation: 3. Predator diet breadth and preference:
 BIOS 3010: Ecology Lecture 8: Predator foraging & prey defense 1. Lecture Summary: What is predation? Predator diet breadth. Preference & switching. Optimal foraging. Marginal value theorem. Functional
BIOS 3010: Ecology Lecture 8: Predator foraging & prey defense 1. Lecture Summary: What is predation? Predator diet breadth. Preference & switching. Optimal foraging. Marginal value theorem. Functional
AP Environmental Science I. Unit 1-2: Biodiversity & Evolution
 NOTE/STUDY GUIDE: Unit 1-2, Biodiversity & Evolution AP Environmental Science I, Mr. Doc Miller, M.Ed. North Central High School Name: ID#: NORTH CENTRAL HIGH SCHOOL NOTE & STUDY GUIDE AP Environmental
NOTE/STUDY GUIDE: Unit 1-2, Biodiversity & Evolution AP Environmental Science I, Mr. Doc Miller, M.Ed. North Central High School Name: ID#: NORTH CENTRAL HIGH SCHOOL NOTE & STUDY GUIDE AP Environmental
Oenopia conglobata L.
 2011 (5) (33) _ Tishreen University Journal for Research and Scientific Studies - Biological Sciences Series Vol. (33) No. (5) 2011 Oenopia conglobata L. * ** Myzus persicae (Sulzer) (2011 / 9 / 25.2011
2011 (5) (33) _ Tishreen University Journal for Research and Scientific Studies - Biological Sciences Series Vol. (33) No. (5) 2011 Oenopia conglobata L. * ** Myzus persicae (Sulzer) (2011 / 9 / 25.2011
Niche The sum of all interactions a species has with biotic/abiotic components of the environment N-dimensional hypervolume
 Niche The sum of all interactions a species has with biotic/abiotic components of the environment N-dimensional hypervolume Each dimension is a biotic or abiotic resource Ecomorphology Ecology (niche)
Niche The sum of all interactions a species has with biotic/abiotic components of the environment N-dimensional hypervolume Each dimension is a biotic or abiotic resource Ecomorphology Ecology (niche)
Ch 5. Evolution, Biodiversity, and Population Ecology. Part 1: Foundations of Environmental Science
 Ch 5 Evolution, Biodiversity, and Population Ecology Part 1: Foundations of Environmental Science PowerPoint Slides prepared by Jay Withgott and Heidi Marcum Copyright 2006 Pearson Education, Inc., publishing
Ch 5 Evolution, Biodiversity, and Population Ecology Part 1: Foundations of Environmental Science PowerPoint Slides prepared by Jay Withgott and Heidi Marcum Copyright 2006 Pearson Education, Inc., publishing
Ch. 14 Interactions in Ecosystems
 Ch. 14 Interactions in Ecosystems 1 14.1 Habitat vs. Niche Habitat all biotic and abiotic factors where an organism lives WHERE a species lives 2 Ecological Niche All physical, chemical, and biological
Ch. 14 Interactions in Ecosystems 1 14.1 Habitat vs. Niche Habitat all biotic and abiotic factors where an organism lives WHERE a species lives 2 Ecological Niche All physical, chemical, and biological
A population is a group of individuals of the same species occupying a particular area at the same time
 A population is a group of individuals of the same species occupying a particular area at the same time Population Growth As long as the birth rate exceeds the death rate a population will grow Immigration
A population is a group of individuals of the same species occupying a particular area at the same time Population Growth As long as the birth rate exceeds the death rate a population will grow Immigration
Principles of Ecology BL / ENVS 402 Exam II Name:
 Principles of Ecology BL / ENVS 402 Exam II 10-26-2011 Name: There are three parts to this exam. Use your time wisely as you only have 50 minutes. Part One: Circle the BEST answer. Each question is worth
Principles of Ecology BL / ENVS 402 Exam II 10-26-2011 Name: There are three parts to this exam. Use your time wisely as you only have 50 minutes. Part One: Circle the BEST answer. Each question is worth
2. Overproduction: More species are produced than can possibly survive
 Name: Date: What to study? Class notes Graphic organizers with group notes Review sheets What to expect on the TEST? Multiple choice Short answers Graph Reading comprehension STRATEGIES Circle key words
Name: Date: What to study? Class notes Graphic organizers with group notes Review sheets What to expect on the TEST? Multiple choice Short answers Graph Reading comprehension STRATEGIES Circle key words
(Write your name on every page. One point will be deducted for every page without your name!)
") POPULATION GENETICS AND MICROEVOLUTIONARY THEORY FINAL EXAMINATION (Write your name on every page. One point will be deducted for every page without your name!) 1. Briefly define (5 points each): a) Average
POPULATION GENETICS AND MICROEVOLUTIONARY THEORY FINAL EXAMINATION (Write your name on every page. One point will be deducted for every page without your name!) 1. Briefly define (5 points each): a) Average
Polyphenic Insects. genotype X environment = phenotype POLYPHENISM. genetic polymorphism vs polyphenism. the peppered moth.
 What makes for differences between individuals? Polyphenic Insects genes environment genotype X environment = phenotype POLYPHENISM poly many (more than one anyway) phen - form genetic polymorphism vs
What makes for differences between individuals? Polyphenic Insects genes environment genotype X environment = phenotype POLYPHENISM poly many (more than one anyway) phen - form genetic polymorphism vs
let s examine pupation rates. With the conclusion of that data collection, we will go on to explore the rate at which new adults appear, a process
 Population Dynamics and Initial Population Size (Module website: http://web.as.uky.edu/biology/faculty/cooper/population%20dynamics%20examples%20 with%20fruit%20flies/theamericanbiologyteacher-populationdynamicswebpage.html
Population Dynamics and Initial Population Size (Module website: http://web.as.uky.edu/biology/faculty/cooper/population%20dynamics%20examples%20 with%20fruit%20flies/theamericanbiologyteacher-populationdynamicswebpage.html
REVIEW OF PHYLOGENY AND EUKARYOTIC ORIGINS (QUIZ MON)
") REVIEW OF PHYLOGENY AND EUKARYOTIC ORIGINS (QUIZ MON) Why is biodiversity important? What are phylogeny and systematics? What does it mean for a phylogenetic tree to be rooted? What is a dichotomous key?
REVIEW OF PHYLOGENY AND EUKARYOTIC ORIGINS (QUIZ MON) Why is biodiversity important? What are phylogeny and systematics? What does it mean for a phylogenetic tree to be rooted? What is a dichotomous key?
Science 9 - Unit A Review Sheet
 Science 9 - Unit A Review Sheet Learning Outcomes Can you? describe the relative abundance of species on Earth and in different environments describe examples of variation among species and within species
Science 9 - Unit A Review Sheet Learning Outcomes Can you? describe the relative abundance of species on Earth and in different environments describe examples of variation among species and within species
Exploring Matthaei s Ecosystems
 Name: Exploring Matthaei s Ecosystems As you walk on the trails, look for evidence of each of the following components of an ecosystem. Draw and describe what you observed and where you found it. Component
Name: Exploring Matthaei s Ecosystems As you walk on the trails, look for evidence of each of the following components of an ecosystem. Draw and describe what you observed and where you found it. Component
BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences
 BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences Week 5. Plant defense theory 2: Development: Lecture summary: Resource Availability Theory: Coley, Bryant
BIOS 5970: Plant-Herbivore Interactions Dr. Stephen Malcolm, Department of Biological Sciences Week 5. Plant defense theory 2: Development: Lecture summary: Resource Availability Theory: Coley, Bryant
Minute Pirate Bug: A Beneficial Generalist Insect Predator
 Minute Pirate Bug: A Beneficial Generalist Insect Predator Veronica Johnson* and Cerruti R 2 Hooks $ University of Maryland Dept. of Entomology * Graduate student and $ Associate professor and Extension
Minute Pirate Bug: A Beneficial Generalist Insect Predator Veronica Johnson* and Cerruti R 2 Hooks $ University of Maryland Dept. of Entomology * Graduate student and $ Associate professor and Extension
Qualitative and Quantitative Prey Requirements of two Aphidophagous Coccinellids, Adalia tetraspilota and Hippodamia variegata
 Qualitative and Quantitative Prey Requirements of two Aphidophagous Coccinellids, Adalia tetraspilota and Hippodamia variegata Authors: Mohd Abas Shah, and Akhtar Ali Khan Source: Journal of Insect Science,
Qualitative and Quantitative Prey Requirements of two Aphidophagous Coccinellids, Adalia tetraspilota and Hippodamia variegata Authors: Mohd Abas Shah, and Akhtar Ali Khan Source: Journal of Insect Science,
Page # Herbivory. I. Introduction A. Functional types of heterotrophs. Predators. Parasites. Herbivores. How do they differ?
 Herbivory I. Introduction A. Functional types of heterotrophs Predators Parasites Herbivores How do they differ? Functional types of heterotrophs Predators - kill and eat several animals (prey) over lifetime
Herbivory I. Introduction A. Functional types of heterotrophs Predators Parasites Herbivores How do they differ? Functional types of heterotrophs Predators - kill and eat several animals (prey) over lifetime
Beadle Plasticus Evolution Teacher Information
 STO-125 Beadle Plasticus Evolution Teacher Information Summary Students model the effects of two different environments on the frequencies of characteristics in a simulated Beadle population. Core Concepts
STO-125 Beadle Plasticus Evolution Teacher Information Summary Students model the effects of two different environments on the frequencies of characteristics in a simulated Beadle population. Core Concepts
a) Understand how light, temperature and water impact living things and how these living things adapt themselves to their environment.
 Understand how light, temperature and water impact living things and how these living things adapt themselves to their environment.") This area deals with organisms living in a particular environment and ecosystems that contain components that affect the lives of organisms. In this area, students come to understand the complex array
This area deals with organisms living in a particular environment and ecosystems that contain components that affect the lives of organisms. In this area, students come to understand the complex array
ISSN Pak. J. Agri., Agril. Engg., Vet. Sci., 2012, 28 (2):
:") ISSN 1023-1072 Pak. J. Agri., Agril. Engg., Vet. Sci., 2012, 28 (2): 150-159 BIOLOGY AND FEEDING POTENTIAL OF PREDATOR, HIPPODAMIA CONVERGENS GUIR, (COLEOPTERA: COCCINELLIDAE) ON MUSTARD APHID, LIPAPHIS
ISSN 1023-1072 Pak. J. Agri., Agril. Engg., Vet. Sci., 2012, 28 (2): 150-159 BIOLOGY AND FEEDING POTENTIAL OF PREDATOR, HIPPODAMIA CONVERGENS GUIR, (COLEOPTERA: COCCINELLIDAE) ON MUSTARD APHID, LIPAPHIS
May 11, Aims: Agenda
 May 11, 2017 Aims: SWBAT explain how survival of the fittest and natural selection have contributed to the continuation, extinction, and adaptation of species. Agenda 1. Do Now 2. Class Notes 3. Guided
May 11, 2017 Aims: SWBAT explain how survival of the fittest and natural selection have contributed to the continuation, extinction, and adaptation of species. Agenda 1. Do Now 2. Class Notes 3. Guided
Goldenrod Galls and the Scientific Method
 Goldenrod Galls and the Scientific Method Overview Groups of students are given several goldenrod stems with galls. They are asked to make observations, come up with questions and make hypotheses. They
Goldenrod Galls and the Scientific Method Overview Groups of students are given several goldenrod stems with galls. They are asked to make observations, come up with questions and make hypotheses. They
BIO S380T Page 1 Summer 2005: Exam 2
 BIO S380T Page 1 Part I: Definitions. [5 points for each term] For each term, provide a brief definition that also indicates why the term is important in ecology or evolutionary biology. Where I ve provided
BIO S380T Page 1 Part I: Definitions. [5 points for each term] For each term, provide a brief definition that also indicates why the term is important in ecology or evolutionary biology. Where I ve provided
Structure Learning of a Behavior Network for Context Dependent Adaptability
 Georgia State University ScholarWorks @ Georgia State University Computer Science Theses Department of Computer Science 12-7-26 Structure Learning of a Behavior Network for Context Dependent Adaptability
Georgia State University ScholarWorks @ Georgia State University Computer Science Theses Department of Computer Science 12-7-26 Structure Learning of a Behavior Network for Context Dependent Adaptability
Outline. Ecology: Succession and Life Strategies. Interactions within communities of organisms. Key Concepts:
 Ecology: Succession and Life Strategies Interactions within communities of organisms u 1. Key concepts Outline u 2. Ecosystems and communities u 3. Competition, Predation, Commensalism, Mutualism, Parasitism
Ecology: Succession and Life Strategies Interactions within communities of organisms u 1. Key concepts Outline u 2. Ecosystems and communities u 3. Competition, Predation, Commensalism, Mutualism, Parasitism
14.1 Habitat And Niche
 14.1 Habitat And Niche A habitat differs from a niche. Habitat physical area in which an organism lives Niche each species plays a specific role in an ecosystem niche includes the species habitat, feeding
14.1 Habitat And Niche A habitat differs from a niche. Habitat physical area in which an organism lives Niche each species plays a specific role in an ecosystem niche includes the species habitat, feeding
Interspecific ant competition over novel aphid resources and changes in plant chemistry. due to ant-aphid mutualisms on milkweed plants
 Liesl Oeller 7/27/14 Ecology Summer 2014 Interspecific ant competition over novel aphid resources and changes in plant chemistry due to ant-aphid mutualisms on milkweed plants Abstract Ants and aphids
Liesl Oeller 7/27/14 Ecology Summer 2014 Interspecific ant competition over novel aphid resources and changes in plant chemistry due to ant-aphid mutualisms on milkweed plants Abstract Ants and aphids
Unit G: Pest Management. Lesson 2: Managing Crop Diseases
 Unit G: Pest Management Lesson 2: Managing Crop Diseases 1 Terms Abiotic disease Bacteria Biotic disease Cultural disease control Disease avoidance Disease resistance Disease tolerance Fungi Infectious
Unit G: Pest Management Lesson 2: Managing Crop Diseases 1 Terms Abiotic disease Bacteria Biotic disease Cultural disease control Disease avoidance Disease resistance Disease tolerance Fungi Infectious
APES Chapter 9 Study Guide. 1. Which of the following statements about sea otters is false?
 APES Chapter 9 Study Guide 1. Which of the following statements about sea otters is false? They use tools, They have the thickest fur of any mammal. They can eat 25% of their weight per day in sea urchins
APES Chapter 9 Study Guide 1. Which of the following statements about sea otters is false? They use tools, They have the thickest fur of any mammal. They can eat 25% of their weight per day in sea urchins
CHAPTER. Population Ecology
 CHAPTER 4 Population Ecology Chapter 4 TOPIC POPULATION ECOLOGY Indicator Species Serve as Biological Smoke Alarms Indicator species Provide early warning of damage to a community Can monitor environmental
CHAPTER 4 Population Ecology Chapter 4 TOPIC POPULATION ECOLOGY Indicator Species Serve as Biological Smoke Alarms Indicator species Provide early warning of damage to a community Can monitor environmental
module, with the exception that the vials are larger and you only use one initial population size.
 Population Dynamics and Space Availability (http://web.as.uky.edu/biology/faculty/cooper/population%20dynamics%20examples%2 0with%20fruit%20flies/TheAmericanBiologyTeacher- PopulationDynamicsWebpage.html
Population Dynamics and Space Availability (http://web.as.uky.edu/biology/faculty/cooper/population%20dynamics%20examples%2 0with%20fruit%20flies/TheAmericanBiologyTeacher- PopulationDynamicsWebpage.html
Campbell Essential Biology, 5e (Simon/Yeh) Chapter 1 Introduction: Biology Today. Multiple-Choice Questions
 Chapter 1 Introduction: Biology Today. Multiple-Choice Questions") Campbell Essential Biology, 5e (Simon/Yeh) Chapter 1 Introduction: Biology Today Multiple-Choice Questions 1) In what way(s) is the science of biology influencing and changing our culture? A) by helping
Campbell Essential Biology, 5e (Simon/Yeh) Chapter 1 Introduction: Biology Today Multiple-Choice Questions 1) In what way(s) is the science of biology influencing and changing our culture? A) by helping
Assessment Schedule 2016 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603)
") NCEA Level 3 Biology (91603) 2016 page 1 of 6 Assessment Schedule 2016 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Evidence Statement
NCEA Level 3 Biology (91603) 2016 page 1 of 6 Assessment Schedule 2016 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Evidence Statement
Jeopardy. Final Jeopardy. Topic 1 Topic 2 Topic 3 Topic 4 Topic 5 $100 $100 $100 $100 $100 $200 $200 $200 $200 $200 $300 $300 $300 $300 $400 $400
 Jeopardy Topic 1 Topic 2 Topic 3 Topic 4 Topic 5 $100 $100 $100 $100 $100 $200 $200 $200 $200 $200 $300 $300 $300 $300 $300 $400 $400 $400 $400 $400 $500 $500 $500 $500 $500 Final Jeopardy 1 - $100 n Although
Jeopardy Topic 1 Topic 2 Topic 3 Topic 4 Topic 5 $100 $100 $100 $100 $100 $200 $200 $200 $200 $200 $300 $300 $300 $300 $300 $400 $400 $400 $400 $400 $500 $500 $500 $500 $500 Final Jeopardy 1 - $100 n Although
FACTORS FOR INSECTS ABUNDANCE. 1. More number of species: In the animal kingdom more than 85 per cent of the species
 FACTORS FOR INSECTS ABUNDANCE Measures of dominance 1. More number of species: In the animal kingdom more than 85 per cent of the species belongs to insect group. Total number of insects described so far
FACTORS FOR INSECTS ABUNDANCE Measures of dominance 1. More number of species: In the animal kingdom more than 85 per cent of the species belongs to insect group. Total number of insects described so far
Campbell Essential Biology, 4/e (Simon/Reece/Dickey)
") Campbell Essential Biology, 4/e (Simon/Reece/Dickey) Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) Using the branching tree of life for
Campbell Essential Biology, 4/e (Simon/Reece/Dickey) Name MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) Using the branching tree of life for
Regents Review Assignment #8-A08 Living Environment: Comet Part A Questions
 Part A Questions 1. A student notices that fruit flies with the curlywing trait develop straight wings if kept at a temperature of 16 C, but develop curly wings if kept at 25 C. The best explanation for
Part A Questions 1. A student notices that fruit flies with the curlywing trait develop straight wings if kept at a temperature of 16 C, but develop curly wings if kept at 25 C. The best explanation for
MIGRATORY PATTERNS AND DIAPAUSE OF HIPPODAMIA CONVERGENS (CONVERGENT LADYBUG)
") S. E. Grossett Migratory Patterns and Diapause of Hippodamia convergens 1 ABSTRACT MIGRATORY PATTERNS AND DIAPAUSE OF HIPPODAMIA CONVERGENS (CONVERGENT LADYBUG) SARA ELIZABETH GROSSETT SARA.GROSSETT@GMAIL.COM
S. E. Grossett Migratory Patterns and Diapause of Hippodamia convergens 1 ABSTRACT MIGRATORY PATTERNS AND DIAPAUSE OF HIPPODAMIA CONVERGENS (CONVERGENT LADYBUG) SARA ELIZABETH GROSSETT SARA.GROSSETT@GMAIL.COM
14.1. KEY CONCEPT Every organism has a habitat and a niche. 38 Reinforcement Unit 5 Resource Book
 14.1 HABITAT AND NICHE KEY CONCEPT Every organism has a habitat and a niche. A habitat is all of the living and nonliving factors in the area where an organism lives. For example, the habitat of a frog
14.1 HABITAT AND NICHE KEY CONCEPT Every organism has a habitat and a niche. A habitat is all of the living and nonliving factors in the area where an organism lives. For example, the habitat of a frog
Groups of organisms living close enough together for interactions to occur.
 Community ecology: First, let's define a community: Groups of organisms living close enough together for interactions to occur. First we probably want to describe the community a bit, so we look at: Biodiversity
Community ecology: First, let's define a community: Groups of organisms living close enough together for interactions to occur. First we probably want to describe the community a bit, so we look at: Biodiversity
Copyright 2014 Edmentum - All rights reserved.
 Copyright 2014 Edmentum - All rights reserved. AP Biology Unity and Diversity Blizzard Bag 2014-20151. The sawfish, also known as the carpenter shark, lives in estuaries off the coast of Australia. A scientist
Copyright 2014 Edmentum - All rights reserved. AP Biology Unity and Diversity Blizzard Bag 2014-20151. The sawfish, also known as the carpenter shark, lives in estuaries off the coast of Australia. A scientist
Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603)
") NCEA Level 3 Biology (91603) 2013 page 1 of 6 Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Assessment Criteria
NCEA Level 3 Biology (91603) 2013 page 1 of 6 Assessment Schedule 2013 Biology: Demonstrate understanding of the responses of plants and animals to their external environment (91603) Assessment Criteria
Current controversies in Marine Ecology with an emphasis on Coral reef systems
 Current controversies in Marine Ecology with an emphasis on Coral reef systems Open vs closed populations (already discussed) The extent and importance of larval dispersal Maintenance of Diversity Equilibrial
Current controversies in Marine Ecology with an emphasis on Coral reef systems Open vs closed populations (already discussed) The extent and importance of larval dispersal Maintenance of Diversity Equilibrial
PERFORMANCE OF NATURAL ENEMIES REARED ON ARTIFICIAL DIETS J.E. Carpenter 1 and S. Bloem 2 1
 Performance of natural enemies reared on artificial diets 143 PERFORMANCE OF NATURAL ENEMIES REARED ON ARTIFICIAL DIETS J.E. Carpenter 1 and S. Bloem 2 1 U.S. Department of Agriculture, Agricultural Research
Performance of natural enemies reared on artificial diets 143 PERFORMANCE OF NATURAL ENEMIES REARED ON ARTIFICIAL DIETS J.E. Carpenter 1 and S. Bloem 2 1 U.S. Department of Agriculture, Agricultural Research
BIOLOGICAL PARAMETERS AND PREY CONSUMPTION BY ZIGZAG BEETLE
 Sarhad J. Agric. Vol. 23, No. 4, 2007 BIOLOGICAL PARAMETERS AND PREY CONSUMPTION BY ZIGZAG BEETLE Menochilus sexamaculatus fab. AGAINST Rhopalosiphum maidis fitch, Aphis gossypii glov. AND Therioaphis
Sarhad J. Agric. Vol. 23, No. 4, 2007 BIOLOGICAL PARAMETERS AND PREY CONSUMPTION BY ZIGZAG BEETLE Menochilus sexamaculatus fab. AGAINST Rhopalosiphum maidis fitch, Aphis gossypii glov. AND Therioaphis
Pulse Knowledge. Pea Aphid. Identification and Life Cycle. Host Crops and Crop Injury. Scouting and Economic Thresholds. Jennifer Bogdan, P.Ag.
 Pulse Knowledge Pea Aphid Jennifer Bogdan, P.Ag., CCA The pea aphid (Acyrthosiphon pisum (Harris)) is a common insect found wherever pulses are grown in Saskatchewan. Pea aphids cause damage to their host
Pulse Knowledge Pea Aphid Jennifer Bogdan, P.Ag., CCA The pea aphid (Acyrthosiphon pisum (Harris)) is a common insect found wherever pulses are grown in Saskatchewan. Pea aphids cause damage to their host
28 3 Insects Slide 1 of 44
 1 of 44 Class Insecta contains more species than any other group of animals. 2 of 44 What Is an Insect? What Is an Insect? Insects have a body divided into three parts head, thorax, and abdomen. Three
1 of 44 Class Insecta contains more species than any other group of animals. 2 of 44 What Is an Insect? What Is an Insect? Insects have a body divided into three parts head, thorax, and abdomen. Three
Maintenance of species diversity
 1. Ecological succession A) Definition: the sequential, predictable change in species composition over time foling a disturbance - Primary succession succession starts from a completely empty community
1. Ecological succession A) Definition: the sequential, predictable change in species composition over time foling a disturbance - Primary succession succession starts from a completely empty community
Current controversies in Marine Ecology with an emphasis on Coral reef systems. Niche Diversification Hypothesis Assumptions:
 Current controversies in Marine Ecology with an emphasis on Coral reef systems Open vs closed populations (already Discussed) The extent and importance of larval dispersal Maintenance of Diversity Equilibrial
Current controversies in Marine Ecology with an emphasis on Coral reef systems Open vs closed populations (already Discussed) The extent and importance of larval dispersal Maintenance of Diversity Equilibrial
Unit D: Controlling Pests and Diseases in the Orchard. Lesson 5: Identify and Control Diseases in the Orchard
 Unit D: Controlling Pests and Diseases in the Orchard Lesson 5: Identify and Control Diseases in the Orchard 1 Terms Abiotic disease Bacteria Biotic diseases Cultural disease control Disease avoidance
Unit D: Controlling Pests and Diseases in the Orchard Lesson 5: Identify and Control Diseases in the Orchard 1 Terms Abiotic disease Bacteria Biotic diseases Cultural disease control Disease avoidance
Impact of Pollinators in Rangelands. months. Insects are vital to humans, and without them, humans would disappear, too. Insects
 Alger 1 April Alger Impact of Pollinators in Rangelands E.O. Wilson, an american biologist, said If we were to wipe out insects alone on this planet, the rest of life and humanity with it would mostly
Alger 1 April Alger Impact of Pollinators in Rangelands E.O. Wilson, an american biologist, said If we were to wipe out insects alone on this planet, the rest of life and humanity with it would mostly
What do plants compete for? What do animals compete for? What is a gamete and what do they carry? What is a gene?
 How are these animals adapted to their surroundings: - a) Polar bear b) Camel c) Cactus What do plants compete for? What do animals compete for? What is a gamete and what do they carry? What is a gene?
How are these animals adapted to their surroundings: - a) Polar bear b) Camel c) Cactus What do plants compete for? What do animals compete for? What is a gamete and what do they carry? What is a gene?
Multiple choice 2 pts each): x 2 = 18) Essay (pre-prepared) / 15 points. 19) Short Answer: / 2 points. 20) Short Answer / 5 points
: x 2 = 18) Essay (pre-prepared) / 15 points. 19) Short Answer: / 2 points. 20) Short Answer / 5 points") P 1 Biology 217: Ecology Second Exam Fall 2004 There should be 7 ps in this exam - take a moment and count them now. Put your name on the first p of the exam, and on each of the ps with short answer questions.
P 1 Biology 217: Ecology Second Exam Fall 2004 There should be 7 ps in this exam - take a moment and count them now. Put your name on the first p of the exam, and on each of the ps with short answer questions.