Nature Structural and Molecular Biology: doi: /nsmb.2938
|
|
|
- Malcolm Elliott
- 5 years ago
- Views:
Transcription


1
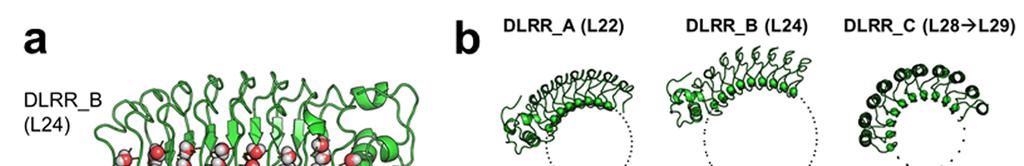
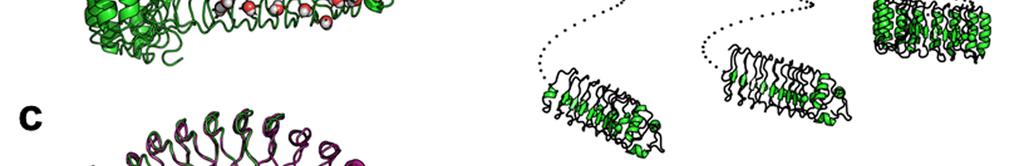
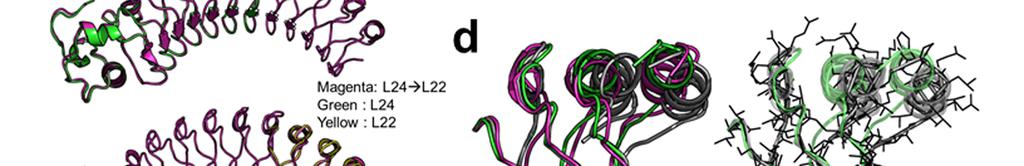
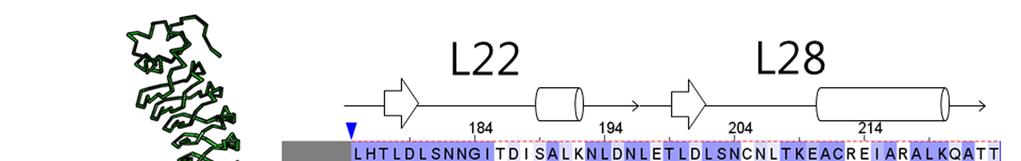
2 Supplementary Figure 1 Characterization of designed leucine-rich-repeat proteins. (a) Water-mediate hydrogen-bond network is frequently visible in the convex region of LRR crystal structures. Examples are shown for the idealized L24 (DLRR_B) and L24 L28 fusion structure (DLRR_G3). Water molecules participating in the hydrogen bond (yellow dots) network are represented by spheres. (b) Super-helical shapes of the three idealized building block repeats. For clear visualization, dots tracing the global super-helix defined by the fitted parameters are overlaid with the LRR structures (rotation angle < 720 ). The highly conserved leucine residues used for the parameter fitting are represented by spheres. See Supplementary Table 1 for the helical parameter estimation. (c) Structural alignments of the partial Ncap-L24 5 structure in DLRR_B (top) and L22 5 structure in DLRR_A (bottom) into the crystal structure of DLRR_E. Cα r.m.s. deviations for the alignments of DLRR_B and DLRR_A are 0.4 Å and 0.3 Å, respectively. (d) Structural defects in the initial fusion model of DLRR_G3. The crystal structure (magenta) of the junction module in DLRR_G3 is aligned with the initial model structure before design (gray) and the final model structure after design (green). The initial model contains large cavity and side chain clashes in the junction module, which are improved in the subsequent design procedure as shown in the final model structure (green). (e) SEC-MALS experiments for DLRR_D, DLRR_E, DLRR_I, DLRR_J, DLRR_K, and DLRR_L. Most of designs are monomeric even though some soluble aggregates/oligomers are observed in DLRR_I and DLRR_K.

3
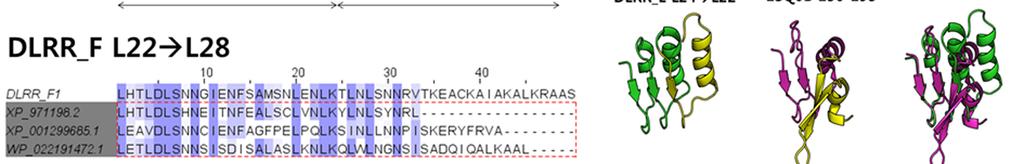
4 Supplementary Figure 2 Experimental characterization of six L22 L28 designs (DLRR_F). In the top row, structure alignment (left) and sequence alignment (right) of the six junction module designs are represented. The building block sequences (L22 + L28) are shown in the first row of the sequence alignment for comparison. Far-UV CD spectra, thermal denaturation at 218 nm, and SEC-MALS are shown from left to right for each design.
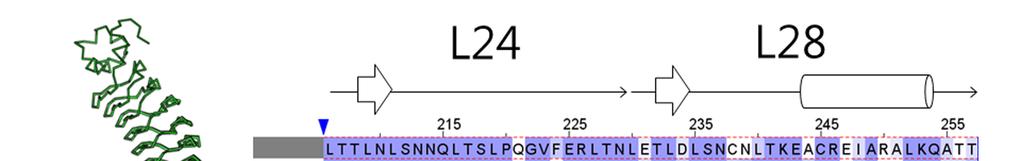

5
6 Supplementary Figure 3 Experimental characterization of six L24 L28 designs (DLRR_G). In the top row, structure alignment (left) and sequence alignment (right) of the six junction module designs are represented. The building block sequences (L24 + L28) are shown in the first row of the sequence alignment for comparison. Far-UV CD spectra, thermal denaturation at 218 nm, and SEC-MALS are shown from left to right for each design. DLRR_G6 has one less {L28 L29} module than the others. The crystal structure of DLRR_G3 is shown in Figure 3d.

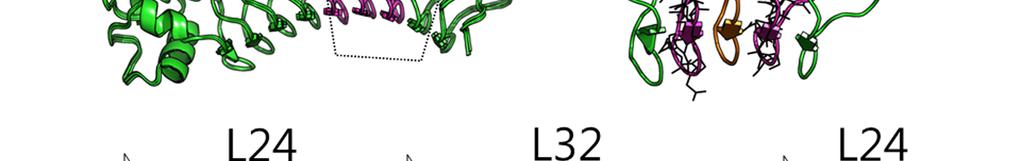
7
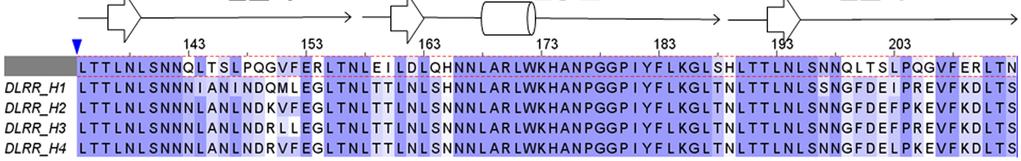
8 Supplementary Figure 4 Experimental characterization of four L24 L32 L24 designs (DLRR_H). In the top row, structural alignment of the four wedge module designs is represented with the structure. Sequence alignment of the four wedge module designs is shown with the building block and the native L32 module sequence (L24 + L32 + L24) in the first row of the alignment for comparison. Far-UV CD spectra, thermal denaturation at 218 nm, and SEC-MALS are shown from left to right for each design. Design DLRR_I has two identical L32 modules derived from DLRR_H1 (Supplementary Table 2). In SEC-MALS experiments, some soluble aggregates/oligomers are observed in addition to the monomeric status. The crystal structure of DLRR_H2 is shown in Figure 3e.


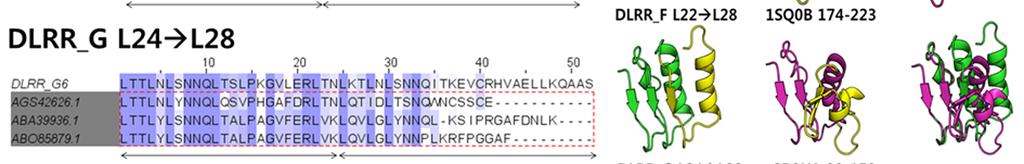

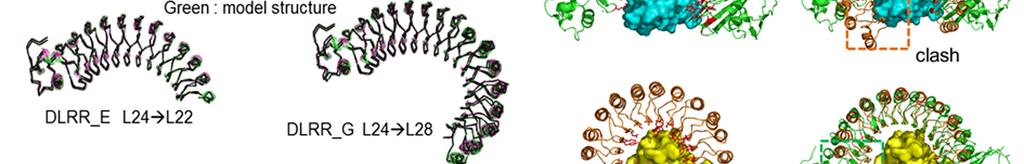
9 Supplementary Figure 5
10 Characterization of designed junction modules. (a) Sequence alignments between the designed junction modules and the top 3 naturally occurring sequences (square block) found in BLAST 1 search for the non-redundant (NR) database. There are numerous sequence differences between the designed modules and the closest sequence in NR. Indeed, BLAST fails to find full length alignments for most of the junction sequences. (b) Comparison of structures of designed and naturally occurring junctions between LRR modules. Left: designed junction modules, Middle: the closest structural matches found in the PDB using TMalign 2, Right: structural alignment. The TMalign searches were carried out with the twounit junction module structures (green) and one or two module structures next to the junction module are shown for both designed and natural structures (yellow) to make the ideality (lack of ideality) of the different structures clearer. Most junctions between different length LRR modules in the native structures occur near the caps where the structure becomes much less regular. This irregularity, evident in the right side of the images from native structures, makes it not possible to generate novel LRR s with controlled curvature by combining multiple different types of modules simply using junctions already existing in the PDB. (c) Structural comparison between crystal structures and model structures generated by the iterative module assembly protocol described in Method. All model structures show high consistency to the crystal structures (r.m.s. deviation g in Table 2). (d) Native LRR proteins, internalin A (InlA, PDB ID: 1O6S, top left) and ribonuclease inhibitor (RI, PDB ID: 1A4Y, bottom left), achieve high affinity and specificity by having shapes closely conforming to the surfaces of the target proteins (human E-cadherin and ribonuclease A, respectively). Each protein has a curvature optimized to its target, resulting in well-packed complementary protein-protein interfaces with hot-spot clusters (shown by red sticks) at both the N and C termini. In contract, swapping the respective target for each of the LRR proteins (i.e. RI:E-cadherin, orange-cyan complex in the top right and InlA:ribonuclease, green-yellow complex in the bottom right) makes the clashes and large gaps in the binding interface.
11 Supplementary Table 1 Super-helical parameters of building block modules LRR type Rise (Å) Radius (Å) Rotation angle (radian) Number of repeat units used for fitting Fitted RMSD (Å) L L {L28 L29} The L22, L24 and {L28 L29} repeats form unique solenoid shapes which can be described by three super-helical parameters (radius: distance to the helical axis, rise: projected distance along the helical axis between adjacent units, and rotation angle: rotation angle about the helical axis between units). The global helical shapes and parameters are estimated by fitting the three parameters to the repeat protein structures. For the parameter fitting, one of the highly conserved positions, the second Leu in LxxLxLxxN/C motif, is used as a representative for each repeat module. The Cα coordinates of the representative positions are obtained from the crystal structures of DLRR_A (L22) and DLRR_B (L24), and from the model structure of DLRR_C ({L28 L29}). Eight to ten Cα coordinates are used to fit the same number of coordinates arbitrary generated from the three helical parameters. RMSD between the two coordinate sets is minimized by using non-linear optimization algorithm (constroptim.nl) in alabama R package 3,4. Initial helical parameters, the input of the optimization procedure, are inferred from the transformation matrix between the first two modules of the building block structures. After performing the optimization procedure, the parameter of the lowest RMSD is used to represent the global helical shape of the idealized building block structures (Supplementary Fig. 1b).
12 Supplementary Table 2 Module organization and module origins of the multiple fusion designs in Figure 4c. Design name Module organization Individual modules Original design Ncap L24 2 DLRR_B DLRR_I Ncap L24 2 JN L24 L32 L24 JN L24 L32 L24 L24 2 DLRR_J Ncap L22 4 L24 2 JN L24 L28 L29 [L28 L29] 2 JN L24 L32 L24 JN L24 L32 L24 L24 2 Ncap L22 4 L24 2 JN L24 L28 L29 [L28 L29] 2 Ncap L24 2 JN L24 L32 L24 DLRR_H1 DLRR_H1 DLRR_B DLRR_A DLRR_B DLRR_G3 DLRR_G3 DLRR_G3 DLRR_B DLRR_H2 DLRR_K Ncap L24 2 JN L24 L32 L24 L24 3 JN L24 L28 L29 [L28 L29] 2 L24 3 JN L24 L28 DLRR_B DLRR_G6 L29 DLRR_G6 [L28 L29] 2 DLRR_G6 Ncap L22 3 DLRR_A L24 3 DLRR_B DLRR_L Ncap L22 3 L24 3 JN L24 L32 L24 L24 3 JN L24 L28 L29 [L28 L29] 2 JN L24 L32 L24 L24 3 DLRR_H2 DLRR_B JN L24 L28 DLRR_G6 L29 DLRR_G6 [L28 L29] 2 DLRR_G6
13 Supplementary Table 3 Number of possible fusion LRR structures with respect to the number of repeat units. Number of repeat units Number of possible LRR structures Fold change (i) (i+1) , , , , , , , , ,076, ,417, ,433, LRR structures are generated by recursively following the edges of the network in Figure 4a. The general module assembly starts from Ncap-L22 or Ncap-L24 in the network except {L28 L29} n and each assembly (transition in the network) adds one repeat unit to the structure. The number of repeat units in the table only considers the internal repeat units excluding N-terminal capping domain.
14 Supplementary Table 4 Crystallization conditions Design names DLRR_A Crystallization conditions 22% PEG3350 w/v, 0.1 M MES ph 6.0, 0.2 M NaCl DLRR_E 20% PEG 1000 v/v, 0.1 M Na/K phosphate ph 6.2 DLRR_G3 2 M ammonium sulfate, 0.1 M Bis-Tris Ph 5.5 DLRR_H2 DLRR_I DLRR_K 22% PEG 3350 w/v, 300 mm Ammonium sulfate, unbuffered 24% PEG 3350 w/v, 0.2 M ammonium sulfate, 0.1 M HEPES ph 7.5, 0.1 M proline 20% PEG-3000, 0.1 M Tris ph 7.0, 0.2 M Ca(OAc)2
15 Supplementary Table 5 Designed sequences > DLRR_A ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLHTLDLSNNGITDISALKNLDNLETLDLRNNGITDKSALKNLNNLKgslehhhhhh >DLRR_B KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNQLTSLPQGVFERLT NLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLTSLPQGVFERLTNLKTL NLSNNQLQSLPTGVDEKLTQLTgshhhhhh >DLRR_C LDLSNQNKTKEDCREIARELKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARAL KQAASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEG AAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDL SNCNLTKEACREIARALKQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_D ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLP QGVFERLTSLTTLNLSNNQLTSLPQGVFERLTNLKTLNLSNNQLQSLPTGVDEKLTQLTgshhhhhh >DLRR_E KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNQLTSLPQGVFERLT NLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLHTLDLSNNGITDISALKNLDNLHTLDL SNNGITDISALKNLDNLHTLDLSNNGITDISALKNLTSLHTLDLSNNGITDISALKNLDNLETLDLRNNGITDKSALKN LNNLKgslehhhhhh >DLRR_F1
16 ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLHTLDLSNNGIENFSAMSNLENLKTLNLSNNRVTKEACKAIAKALKRATSLHELHLSNNNIGE EGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETL DLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALK QATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_F2 ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLHTLDLSNNGIENFNALRNLENLKTLNLSNNRVTKDACEAIAEALKRATSLHELHLSNNNIGE EGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETL DLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALK QATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_F3 ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLHTLDLSNNGIENFEAMRNLENLKTLNLSNNRLTKEACKAVAEALKRATSLHELHLSNNNIG EEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLET LDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARAL KQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_F4 ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLHTLDLSNNGITNVSALKNLENLKTLNLSNNNITKEACKAIAEALKRATSLHELHLSNNNIGEE GAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLD LSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQ ATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_F5 ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL
17 KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLHTLDLSNNGIRNLEAMRNLENLKTLNLSNNNVTKEACSALAEALKRATSLHELHLSNNNIG EEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLET LDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARAL KQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_F6 ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLHTLDLSNNGIRNFEAMRNLENLKTLNLSNNNFTKEACSALAEALKRATSLHELHLSNNNIG EEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLET LDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARAL KQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_G1 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNQLTSLPQGVFERLT NLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLTSLPDGVLERLTNLKTL NLSNNQITKEVCRHVAKILKQAASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALK QATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGA AELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshh hhhh >DLRR_G2 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNQLTSLPQGVFERLT NLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLTSLPDGVFERLTNLKTL NLSNNQLTKEACRIVAKMLKQLASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALK QATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGA AELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshh hhhh >DLRR_G3
18 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNQLTSLPQGVFERLT NLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLTSLPDGVFERLTNLKTL NLSNNQLTKEACRAVANALKQAASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARAL KQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEG AAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgsh hhhhh >DLRR_G4 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNQLTSLPQGVFERLT NLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLTSLPDGVLERLTNLKTL NLSNNQITKEVCRLVAKFLKQLASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALK QATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEGA AELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshh hhhh >DLRR_G5 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNQLTSLPQGVFERLT NLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLTSLPDGVFERLTNLKTL NLSNNQITKEVCRMVAKVLKQAASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARAL KQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLHELHLSNNNIGEEG AAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATSLHELHLSNNNIGEEGKAWLEEARRHPGSTLETgsh hhhhh >DLRR_G6 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNQLTSLPQGVFERLT NLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLTSLPKGVLERLTNLKTL NLSNNQITKEVCRHVAELLKQAASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALK QATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATSLHELHLSNNNIGEEGK AWLEEARRHPGSTLETgshhhhhh
19 >DLRR_H1 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNNIANINDQMLEGLT NLTTLNLSHNNLARLWKHANPGGPIYFLKGLTNLTTLNLSSNGFDEIPREVFKDLTSLTTLNLSNNQLTSLPQGVFE RLTNLKTLNLSNNQLQSLPTGVDEKLTQLTgshhhhhh >DLRR_H2 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNNLANLNDKVFEGLT NLTTLNLSNNNLARLWKHANPGGPIYFLKGLTNLTTLNLSNNGFDEFPKEVFKDLTSLTTLNLSNNQLTSLPQGVF ERLTNLKTLNLSNNQLQSLPTGVDEKLTQLTgshhhhhh >DLRR_H3 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNNLANLNDRLLEGLT NLTTLNLSNNNLARLWKHANPGGPIYFLKGLTNLTTLNLSNNGFDEFPREVFKDLTSLTTLNLSNNQLTSLPQGVF ERLTNLKTLNLSNNQLQSLPTGVDEKLTQLTgshhhhhh >DLRR_H4 KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNNLANLNDRVFEGLT NLTTLNLSNNNLARLWKHANPGGPIYFLKGLTNLTTLNLSNNGFDELPKEVFKDLTSLTTLNLSNNQLTSLPQGVF ERLTNLKTLNLSNNQLQSLPTGVDEKLTQLTgshhhhhh >DLRR_I KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNNIANINDQMLEGLT NLTTLNLSHNNLARLWKHANPGGPIYFLKGLTNLTTLNLSSNGFDEIPREVFKDLTSLTTLNLSNNNIANINDQMLE GLTNLTTLNLSHNNLARLWKHANPGGPIYFLKGLTNLTTLNLSSNGFDEIPREVFKDLTSLTTLNLSNNQLTSLPQG VFERLTNLKTLNLSNNQLQSLPTGVDEKLTQLTgshhhhhh >DLRR_J ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL
20 KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLDNLHTLDLS NNGITDISALKNLTSLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLP QGVFERLTNLKTLNLSNNQLTKEACRAVANALKQAASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCN LTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATTLH ELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_K KELTNLGWLNLSNNQLETLPQGVFEKLTNLTTLNLSNNQLTSLPQGVFERLASLTTLNLSNNNLANLNDRVFEGLT NLTTLNLSNNNLARLWKHANPGGPIYFLKGLTNLTTLNLSNNGFDELPKEVFKDLTSLTTLNLSNNQLTSLPQGVF ERLTNLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLTSLPKGVLERLTN LKTLNLSNNQITKEVCRHVAELLKQAASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIA RALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATSLHELHLSNNNIG EEGKAWLEEARRHPGSTLETgshhhhhh >DLRR_L ETITVSTPIKQIFPDDAFAETIKANLKKKSVTDAVTQNELNSIDQIIANNSDIKSVQGIQYLPNLKTLKLSNNKITDISAL KQLNNLGWLDLSNNGITDISALKNLASLHTLDLSNNGITDISALKNLDNLHTLDLSNNGITDISALKNLTSLTTLNLSN NQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNNLA NLNDRVFEGLTNLTTLNLSNNNLARLWKHANPGGPIYFLKGLTNLTTLNLSNNGFDELPKEVFKDLTSLTTLNLSN NQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTNLTTLNLSNNQLTSLPQGVFERLTSLTTLNLSNNQLT SLPKGVLERLTNLKTLNLSNNQITKEVCRHVAELLKQAASLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSN CNLTKEACREIARALKQATTLHELHLSNNNIGEEGAAELVEALLHPGSTLETLDLSNCNLTKEACREIARALKQATS LHELHLSNNNIGEEGKAWLEEARRHPGSTLETgshhhhhh *C-terminal linkers and 6x His tags are shown in lower case. *AS or TS in the regular repeat sequences are for inserting the restriction sites (NheI and SpeI).
21 Supplementary References 1. Altschul, S.F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25, (1997). 2. Y. Zhang, J. Skolnick, TM-align: A protein structure alignment algorithm based on TM-score, Nucleic Acids Research 33, (2005) 3. R Core Team R: A language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria. ISBN , URL (2012) 4. Ravi Varadhan, alabama: Constrained nonlinear optimization. R package version (2012)
Presenter: She Zhang
 Presenter: She Zhang Introduction Dr. David Baker Introduction Why design proteins de novo? It is not clear how non-covalent interactions favor one specific native structure over many other non-native
Presenter: She Zhang Introduction Dr. David Baker Introduction Why design proteins de novo? It is not clear how non-covalent interactions favor one specific native structure over many other non-native
Nitrogenase MoFe protein from Clostridium pasteurianum at 1.08 Å resolution: comparison with the Azotobacter vinelandii MoFe protein
 Acta Cryst. (2015). D71, 274-282, doi:10.1107/s1399004714025243 Supporting information Volume 71 (2015) Supporting information for article: Nitrogenase MoFe protein from Clostridium pasteurianum at 1.08
Acta Cryst. (2015). D71, 274-282, doi:10.1107/s1399004714025243 Supporting information Volume 71 (2015) Supporting information for article: Nitrogenase MoFe protein from Clostridium pasteurianum at 1.08
Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R)
") Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R) Shown in cyan and green are two adjacent tetramers from the crystallographic lattice of COP, forming the only unique inter-tetramer
Supplementary Figure 1 Crystal contacts in COP apo structure (PDB code 3S0R) Shown in cyan and green are two adjacent tetramers from the crystallographic lattice of COP, forming the only unique inter-tetramer
Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate
 and IDH2 (bottom) with isocitrate") SUPPLEMENTARY FIGURE LEGENDS Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate dehydrogenase from Escherichia coli [ICD, pdb 1PB1, Mesecar, A. D., and Koshland,
SUPPLEMENTARY FIGURE LEGENDS Supplementary Figure 1. Aligned sequences of yeast IDH1 (top) and IDH2 (bottom) with isocitrate dehydrogenase from Escherichia coli [ICD, pdb 1PB1, Mesecar, A. D., and Koshland,
SUPPLEMENTARY INFORMATION
 Table of Contents Page Supplementary Table 1. Diffraction data collection statistics 2 Supplementary Table 2. Crystallographic refinement statistics 3 Supplementary Fig. 1. casic1mfc packing in the R3
Table of Contents Page Supplementary Table 1. Diffraction data collection statistics 2 Supplementary Table 2. Crystallographic refinement statistics 3 Supplementary Fig. 1. casic1mfc packing in the R3
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/309/5742/1868/dc1 Supporting Online Material for Toward High-Resolution de Novo Structure Prediction for Small Proteins Philip Bradley, Kira M. S. Misura, David Baker*
www.sciencemag.org/cgi/content/full/309/5742/1868/dc1 Supporting Online Material for Toward High-Resolution de Novo Structure Prediction for Small Proteins Philip Bradley, Kira M. S. Misura, David Baker*
SUPPLEMENTARY INFORMATION. doi: /nature07461
 Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
SUPPLEMENTARY INFORMATION
 Dph2 SeMet (iron-free) # Dph2 (iron-free) Dph2-[4Fe-4S] Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 58.26, 82.08, 160.42 58.74, 81.87, 160.01 55.70, 80.53,
Dph2 SeMet (iron-free) # Dph2 (iron-free) Dph2-[4Fe-4S] Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 58.26, 82.08, 160.42 58.74, 81.87, 160.01 55.70, 80.53,
SUPPLEMENTARY FIGURES
 SUPPLEMENTARY FIGURES Supplementary Figure 1 Protein sequence alignment of Vibrionaceae with either a 40-residue insertion or a 44-residue insertion. Identical residues are indicated by red background.
SUPPLEMENTARY FIGURES Supplementary Figure 1 Protein sequence alignment of Vibrionaceae with either a 40-residue insertion or a 44-residue insertion. Identical residues are indicated by red background.
Supplementary Figures
 1 Supplementary Figures Supplementary Figure 1 Type I FGFR1 inhibitors (a) Chemical structures of a pyrazolylaminopyrimidine inhibitor (henceforth referred to as PAPI; PDB-code of the FGFR1-PAPI complex:
1 Supplementary Figures Supplementary Figure 1 Type I FGFR1 inhibitors (a) Chemical structures of a pyrazolylaminopyrimidine inhibitor (henceforth referred to as PAPI; PDB-code of the FGFR1-PAPI complex:
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
SUPPLEMENTARY INFORMATION
 www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
SUPPLEMENTARY INFORMATION
 Supplementary Table 1: Amplitudes of three current levels. Level 0 (pa) Level 1 (pa) Level 2 (pa) TrkA- TrkH WT 200 K 0.01 ± 0.01 9.5 ± 0.01 18.7 ± 0.03 200 Na * 0.001 ± 0.01 3.9 ± 0.01 12.5 ± 0.03 200
Supplementary Table 1: Amplitudes of three current levels. Level 0 (pa) Level 1 (pa) Level 2 (pa) TrkA- TrkH WT 200 K 0.01 ± 0.01 9.5 ± 0.01 18.7 ± 0.03 200 Na * 0.001 ± 0.01 3.9 ± 0.01 12.5 ± 0.03 200
SUPPLEMENTARY INFORMATION
 Supplementary materials Figure S1 Fusion protein of Sulfolobus solfataricus SRP54 and a signal peptide. a, Expression vector for the fusion protein. The signal peptide of yeast dipeptidyl aminopeptidase
Supplementary materials Figure S1 Fusion protein of Sulfolobus solfataricus SRP54 and a signal peptide. a, Expression vector for the fusion protein. The signal peptide of yeast dipeptidyl aminopeptidase
THE CRYSTAL STRUCTURE OF THE SGT1-SKP1 COMPLEX: THE LINK BETWEEN
 THE CRYSTAL STRUCTURE OF THE SGT1-SKP1 COMPLEX: THE LINK BETWEEN HSP90 AND BOTH SCF E3 UBIQUITIN LIGASES AND KINETOCHORES Oliver Willhoft, Richard Kerr, Dipali Patel, Wenjuan Zhang, Caezar Al-Jassar, Tina
THE CRYSTAL STRUCTURE OF THE SGT1-SKP1 COMPLEX: THE LINK BETWEEN HSP90 AND BOTH SCF E3 UBIQUITIN LIGASES AND KINETOCHORES Oliver Willhoft, Richard Kerr, Dipali Patel, Wenjuan Zhang, Caezar Al-Jassar, Tina
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11085 Supplementary Tables: Supplementary Table 1. Summary of crystallographic and structure refinement data Structure BRIL-NOP receptor Data collection Number of crystals 23 Space group
doi:10.1038/nature11085 Supplementary Tables: Supplementary Table 1. Summary of crystallographic and structure refinement data Structure BRIL-NOP receptor Data collection Number of crystals 23 Space group
Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1.
 Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1. PDZK1 constru cts Amino acids MW [kda] KD [μm] PEPT2-CT- FITC KD [μm] NHE3-CT- FITC KD [μm] PDZK1-CT-
Table S1. Overview of used PDZK1 constructs and their binding affinities to peptides. Related to figure 1. PDZK1 constru cts Amino acids MW [kda] KD [μm] PEPT2-CT- FITC KD [μm] NHE3-CT- FITC KD [μm] PDZK1-CT-
Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase Cwc27
 Acta Cryst. (2014). D70, doi:10.1107/s1399004714021695 Supporting information Volume 70 (2014) Supporting information for article: Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase
Acta Cryst. (2014). D70, doi:10.1107/s1399004714021695 Supporting information Volume 70 (2014) Supporting information for article: Structure and evolution of the spliceosomal peptidyl-prolyl cistrans isomerase
SUPPLEMENTARY INFORMATION
 Figure S1. Secondary structure of CAP (in the camp 2 -bound state) 10. α-helices are shown as cylinders and β- strands as arrows. Labeling of secondary structure is indicated. CDB, DBD and the hinge are
Figure S1. Secondary structure of CAP (in the camp 2 -bound state) 10. α-helices are shown as cylinders and β- strands as arrows. Labeling of secondary structure is indicated. CDB, DBD and the hinge are
Diphthamide biosynthesis requires a radical iron-sulfur enzyme. Pennsylvania State University, University Park, Pennsylvania 16802, USA
 Diphthamide biosynthesis requires a radical iron-sulfur enzyme Yang Zhang, 1,4 Xuling Zhu, 1,4 Andrew T. Torelli, 1 Michael Lee, 2 Boris Dzikovski, 1 Rachel Koralewski, 1 Eileen Wang, 1 Jack Freed, 1 Carsten
Diphthamide biosynthesis requires a radical iron-sulfur enzyme Yang Zhang, 1,4 Xuling Zhu, 1,4 Andrew T. Torelli, 1 Michael Lee, 2 Boris Dzikovski, 1 Rachel Koralewski, 1 Eileen Wang, 1 Jack Freed, 1 Carsten
SUPPLEMENTARY INFORMATION
 Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Prediction and refinement of NMR structures from sparse experimental data
 Prediction and refinement of NMR structures from sparse experimental data Jeff Skolnick Director Center for the Study of Systems Biology School of Biology Georgia Institute of Technology Overview of talk
Prediction and refinement of NMR structures from sparse experimental data Jeff Skolnick Director Center for the Study of Systems Biology School of Biology Georgia Institute of Technology Overview of talk
Orientational degeneracy in the presence of one alignment tensor.
 Orientational degeneracy in the presence of one alignment tensor. Rotation about the x, y and z axes can be performed in the aligned mode of the program to examine the four degenerate orientations of two
Orientational degeneracy in the presence of one alignment tensor. Rotation about the x, y and z axes can be performed in the aligned mode of the program to examine the four degenerate orientations of two
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
Computational engineering of cellulase Cel9A-68 functional motions through mutations in its linker region. WT 1TF4 (crystal) -90 ERRAT PROVE VERIFY3D
 -90 ERRAT PROVE VERIFY3D") Electronic Supplementary Material (ESI) for Physical Chemistry Chemical Physics. This journal is the Owner Societies 218 Supplementary Material: Computational engineering of cellulase Cel9-68 functional
Electronic Supplementary Material (ESI) for Physical Chemistry Chemical Physics. This journal is the Owner Societies 218 Supplementary Material: Computational engineering of cellulase Cel9-68 functional
Supporting Information
 Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis Sergio de Cima, Luis M. Polo, Carmen Díez-Fernández, Ana I. Martínez, Javier
SUPPLEMENTARY INFORMATION Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis Sergio de Cima, Luis M. Polo, Carmen Díez-Fernández, Ana I. Martínez, Javier
Supplemental Data SUPPLEMENTAL FIGURES
 Supplemental Data CRYSTAL STRUCTURE OF THE MG.ADP-INHIBITED STATE OF THE YEAST F 1 C 10 ATP SYNTHASE Alain Dautant*, Jean Velours and Marie-France Giraud* From Université Bordeaux 2, CNRS; Institut de
Supplemental Data CRYSTAL STRUCTURE OF THE MG.ADP-INHIBITED STATE OF THE YEAST F 1 C 10 ATP SYNTHASE Alain Dautant*, Jean Velours and Marie-France Giraud* From Université Bordeaux 2, CNRS; Institut de
Supplementary Figure 1 Crystal packing of ClR and electron density maps. Crystal packing of type A crystal (a) and type B crystal (b).
 and type B crystal (b).") Supplementary Figure 1 Crystal packing of ClR and electron density maps. Crystal packing of type A crystal (a) and type B crystal (b). Crystal contacts at B-C loop are magnified and stereo view of A-weighted
Supplementary Figure 1 Crystal packing of ClR and electron density maps. Crystal packing of type A crystal (a) and type B crystal (b). Crystal contacts at B-C loop are magnified and stereo view of A-weighted
CMPS 3110: Bioinformatics. Tertiary Structure Prediction
 CMPS 3110: Bioinformatics Tertiary Structure Prediction Tertiary Structure Prediction Why Should Tertiary Structure Prediction Be Possible? Molecules obey the laws of physics! Conformation space is finite
CMPS 3110: Bioinformatics Tertiary Structure Prediction Tertiary Structure Prediction Why Should Tertiary Structure Prediction Be Possible? Molecules obey the laws of physics! Conformation space is finite
CMPS 6630: Introduction to Computational Biology and Bioinformatics. Tertiary Structure Prediction
 CMPS 6630: Introduction to Computational Biology and Bioinformatics Tertiary Structure Prediction Tertiary Structure Prediction Why Should Tertiary Structure Prediction Be Possible? Molecules obey the
CMPS 6630: Introduction to Computational Biology and Bioinformatics Tertiary Structure Prediction Tertiary Structure Prediction Why Should Tertiary Structure Prediction Be Possible? Molecules obey the
SUPPLEMENTARY INFORMATION
 Supplementary Table 1: Data collection, phasing and refinement statistics ChbC/Ta 6 Br 12 Native ChbC Data collection Space group P4 3 2 1 2 P4 3 2 1 2 Cell dimensions a, c (Å) 132.75, 453.57 132.81, 452.95
Supplementary Table 1: Data collection, phasing and refinement statistics ChbC/Ta 6 Br 12 Native ChbC Data collection Space group P4 3 2 1 2 P4 3 2 1 2 Cell dimensions a, c (Å) 132.75, 453.57 132.81, 452.95
SUPPLEMENTARY INFORMATION
 doi:1.138/nature1737 Supplementary Table 1 variant Description FSEC - 2B12 a FSEC - 6A1 a K d (leucine) c Leucine uptake e K (wild-type like) K (Y18F) K (TS) K (TSY) K288A mutant, lipid facing side chain
doi:1.138/nature1737 Supplementary Table 1 variant Description FSEC - 2B12 a FSEC - 6A1 a K d (leucine) c Leucine uptake e K (wild-type like) K (Y18F) K (TS) K (TSY) K288A mutant, lipid facing side chain
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature17991 Supplementary Discussion Structural comparison with E. coli EmrE The DMT superfamily includes a wide variety of transporters with 4-10 TM segments 1. Since the subfamilies of the
doi:10.1038/nature17991 Supplementary Discussion Structural comparison with E. coli EmrE The DMT superfamily includes a wide variety of transporters with 4-10 TM segments 1. Since the subfamilies of the
Introduction to Comparative Protein Modeling. Chapter 4 Part I
 Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Basics of protein structure
 Today: 1. Projects a. Requirements: i. Critical review of one paper ii. At least one computational result b. Noon, Dec. 3 rd written report and oral presentation are due; submit via email to bphys101@fas.harvard.edu
Today: 1. Projects a. Requirements: i. Critical review of one paper ii. At least one computational result b. Noon, Dec. 3 rd written report and oral presentation are due; submit via email to bphys101@fas.harvard.edu
Protein Structure Prediction, Engineering & Design CHEM 430
 Protein Structure Prediction, Engineering & Design CHEM 430 Eero Saarinen The free energy surface of a protein Protein Structure Prediction & Design Full Protein Structure from Sequence - High Alignment
Protein Structure Prediction, Engineering & Design CHEM 430 Eero Saarinen The free energy surface of a protein Protein Structure Prediction & Design Full Protein Structure from Sequence - High Alignment
Performing a Pharmacophore Search using CSD-CrossMiner
 Table of Contents Introduction... 2 CSD-CrossMiner Terminology... 2 Overview of CSD-CrossMiner... 3 Searching with a Pharmacophore... 4 Performing a Pharmacophore Search using CSD-CrossMiner Version 2.0
Table of Contents Introduction... 2 CSD-CrossMiner Terminology... 2 Overview of CSD-CrossMiner... 3 Searching with a Pharmacophore... 4 Performing a Pharmacophore Search using CSD-CrossMiner Version 2.0
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10955 Supplementary Figures Supplementary Figure 1. Electron-density maps and crystallographic dimer structures of the motor domain. (a f) Stereo views of the final electron-density maps
doi:10.1038/nature10955 Supplementary Figures Supplementary Figure 1. Electron-density maps and crystallographic dimer structures of the motor domain. (a f) Stereo views of the final electron-density maps
Identifying Interaction Hot Spots with SuperStar
 Identifying Interaction Hot Spots with SuperStar Version 1.0 November 2017 Table of Contents Identifying Interaction Hot Spots with SuperStar... 2 Case Study... 3 Introduction... 3 Generate SuperStar Maps
Identifying Interaction Hot Spots with SuperStar Version 1.0 November 2017 Table of Contents Identifying Interaction Hot Spots with SuperStar... 2 Case Study... 3 Introduction... 3 Generate SuperStar Maps
Table 1. Crystallographic data collection, phasing and refinement statistics. Native Hg soaked Mn soaked 1 Mn soaked 2
 Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
of the Guanine Nucleotide Exchange Factor FARP2
 Structure, Volume 21 Supplemental Information Structural Basis for Autoinhibition of the Guanine Nucleotide Exchange Factor FARP2 Xiaojing He, Yi-Chun Kuo, Tyler J. Rosche, and Xuewu Zhang Inventory of
Structure, Volume 21 Supplemental Information Structural Basis for Autoinhibition of the Guanine Nucleotide Exchange Factor FARP2 Xiaojing He, Yi-Chun Kuo, Tyler J. Rosche, and Xuewu Zhang Inventory of
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11744 Supplementary Table 1. Crystallographic data collection and refinement statistics. Wild-type Se-Met-BcsA-B SmCl 3 -soaked EMTS-soaked Data collection Space
SUPPLEMENTARY INFORMATION doi:10.1038/nature11744 Supplementary Table 1. Crystallographic data collection and refinement statistics. Wild-type Se-Met-BcsA-B SmCl 3 -soaked EMTS-soaked Data collection Space
Nature Structural and Molecular Biology: doi: /nsmb Supplementary Figure 1. Definition and assessment of ciap1 constructs.
 Supplementary Figure 1 Definition and assessment of ciap1 constructs. (a) ciap1 constructs used in this study are shown as primary structure schematics with domains colored as in the main text. Mutations
Supplementary Figure 1 Definition and assessment of ciap1 constructs. (a) ciap1 constructs used in this study are shown as primary structure schematics with domains colored as in the main text. Mutations
SUPPLEMENTARY FIGURES. Figure S1
 SUPPLEMENTARY FIGURES Figure S1 The substrate for DH domain (2R,3R,4R,6R,7S,8S,9R)-3,7,9-trihydroxy-5-oxo-2,4,6,8 tetramethylundecanoate) was docked as two separate fragments shown in magenta and blue
SUPPLEMENTARY FIGURES Figure S1 The substrate for DH domain (2R,3R,4R,6R,7S,8S,9R)-3,7,9-trihydroxy-5-oxo-2,4,6,8 tetramethylundecanoate) was docked as two separate fragments shown in magenta and blue
Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits. AhpC and AhpF from Escherichia coli
 Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits AhpC and AhpF from Escherichia coli Phat Vinh Dip 1,#, Neelagandan Kamariah 2,#, Malathy Sony Subramanian Manimekalai
Structure, mechanism and ensemble formation of the Alkylhydroperoxide Reductase subunits AhpC and AhpF from Escherichia coli Phat Vinh Dip 1,#, Neelagandan Kamariah 2,#, Malathy Sony Subramanian Manimekalai
Acta Crystallographica Section D
 Supporting information Acta Crystallographica Section D Volume 70 (2014) Supporting information for article: Structural characterization of the virulence factor Nuclease A from Streptococcus agalactiae
Supporting information Acta Crystallographica Section D Volume 70 (2014) Supporting information for article: Structural characterization of the virulence factor Nuclease A from Streptococcus agalactiae
Introduction to" Protein Structure
 Introduction to" Protein Structure Function, evolution & experimental methods Thomas Blicher, Center for Biological Sequence Analysis Learning Objectives Outline the basic levels of protein structure.
Introduction to" Protein Structure Function, evolution & experimental methods Thomas Blicher, Center for Biological Sequence Analysis Learning Objectives Outline the basic levels of protein structure.
Bioengineering & Bioinformatics Summer Institute, Dept. Computational Biology, University of Pittsburgh, PGH, PA
 Pharmacophore Model Development for the Identification of Novel Acetylcholinesterase Inhibitors Edwin Kamau Dept Chem & Biochem Kennesa State Uni ersit Kennesa GA 30144 Dept. Chem. & Biochem. Kennesaw
Pharmacophore Model Development for the Identification of Novel Acetylcholinesterase Inhibitors Edwin Kamau Dept Chem & Biochem Kennesa State Uni ersit Kennesa GA 30144 Dept. Chem. & Biochem. Kennesaw
Supporting Information
 Supporting Information Oxaliplatin binding to human copper chaperone Atox1 and protein dimerization Benny D. Belviso, 1 Angela Galliani, 2 Alessia Lasorsa, 2 Valentina Mirabelli, 1,3 Rocco Caliandro, 1
Supporting Information Oxaliplatin binding to human copper chaperone Atox1 and protein dimerization Benny D. Belviso, 1 Angela Galliani, 2 Alessia Lasorsa, 2 Valentina Mirabelli, 1,3 Rocco Caliandro, 1
SUPPLEMENTARY INFORMATION
 Fig. 1 Influences of crystal lattice contacts on Pol η structures. a. The dominant lattice contact between two hpol η molecules (silver and gold) in the type 1 crystals. b. A close-up view of the hydrophobic
Fig. 1 Influences of crystal lattice contacts on Pol η structures. a. The dominant lattice contact between two hpol η molecules (silver and gold) in the type 1 crystals. b. A close-up view of the hydrophobic
Supporting Information How does Darunavir prevent HIV-1 protease dimerization?
 Supporting Information How does Darunavir prevent HIV- protease dimerization? Danzhi Huang and Amedeo Caflisch a Department of Biochemistry University of Zürich, Winterthurerstrasse 9 CH-7 Zürich, Switzerland
Supporting Information How does Darunavir prevent HIV- protease dimerization? Danzhi Huang and Amedeo Caflisch a Department of Biochemistry University of Zürich, Winterthurerstrasse 9 CH-7 Zürich, Switzerland
Lecture 11: Protein Folding & Stability
 Structure - Function Protein Folding: What we know Lecture 11: Protein Folding & Stability 1). Amino acid sequence dictates structure. 2). The native structure represents the lowest energy state for a
Structure - Function Protein Folding: What we know Lecture 11: Protein Folding & Stability 1). Amino acid sequence dictates structure. 2). The native structure represents the lowest energy state for a
Protein Folding & Stability. Lecture 11: Margaret A. Daugherty. Fall Protein Folding: What we know. Protein Folding
 Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2003 Structure - Function Protein Folding: What we know 1). Amino acid sequence dictates structure. 2). The native structure represents
Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2003 Structure - Function Protein Folding: What we know 1). Amino acid sequence dictates structure. 2). The native structure represents
Protein Structure. W. M. Grogan, Ph.D. OBJECTIVES
 Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Supporting Information
 Electronic Supplementary Material (ESI) for Physical Chemistry Chemical Physics. This journal is the Owner Societies 2016 Supporting Information Lipid molecules can induce an opening of membrane-facing
Electronic Supplementary Material (ESI) for Physical Chemistry Chemical Physics. This journal is the Owner Societies 2016 Supporting Information Lipid molecules can induce an opening of membrane-facing
Comparing crystal structure of M.HhaI with and without DNA1, 2 (PDBID:1hmy and PDBID:2hmy),
,") Supporting Information 1. Constructing the starting structure Comparing crystal structure of M.HhaI with and without DNA1, 2 (PDBID:1hmy and PDBID:2hmy), we find that: the RMSD of overall structure and
Supporting Information 1. Constructing the starting structure Comparing crystal structure of M.HhaI with and without DNA1, 2 (PDBID:1hmy and PDBID:2hmy), we find that: the RMSD of overall structure and
Protein Structure. Hierarchy of Protein Structure. Tertiary structure. independently stable structural unit. includes disulfide bonds
 Protein Structure Hierarchy of Protein Structure 2 3 Structural element Primary structure Secondary structure Super-secondary structure Domain Tertiary structure Quaternary structure Description amino
Protein Structure Hierarchy of Protein Structure 2 3 Structural element Primary structure Secondary structure Super-secondary structure Domain Tertiary structure Quaternary structure Description amino
Cryo-EM data collection, refinement and validation statistics
 1 Table S1 Cryo-EM data collection, refinement and validation statistics Data collection and processing CPSF-160 WDR33 (EMDB-7114) (PDB 6BM0) CPSF-160 WDR33 (EMDB-7113) (PDB 6BLY) CPSF-160 WDR33 CPSF-30
1 Table S1 Cryo-EM data collection, refinement and validation statistics Data collection and processing CPSF-160 WDR33 (EMDB-7114) (PDB 6BM0) CPSF-160 WDR33 (EMDB-7113) (PDB 6BLY) CPSF-160 WDR33 CPSF-30
Alpha-helical Topology and Tertiary Structure Prediction of Globular Proteins Scott R. McAllister Christodoulos A. Floudas Princeton University
 Alpha-helical Topology and Tertiary Structure Prediction of Globular Proteins Scott R. McAllister Christodoulos A. Floudas Princeton University Department of Chemical Engineering Program of Applied and
Alpha-helical Topology and Tertiary Structure Prediction of Globular Proteins Scott R. McAllister Christodoulos A. Floudas Princeton University Department of Chemical Engineering Program of Applied and
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
Supporting Information
 Supporting Information Critical role of inter-domain interactions on the conformational change and catalytic mechanism of Endoplasmic Reticulum Aminopeptidase 1 Athanasios Stamogiannos 1, Zachary Maben
Supporting Information Critical role of inter-domain interactions on the conformational change and catalytic mechanism of Endoplasmic Reticulum Aminopeptidase 1 Athanasios Stamogiannos 1, Zachary Maben
Structural characterization of NiV N 0 P in solution and in crystal.
 Supplementary Figure 1 Structural characterization of NiV N 0 P in solution and in crystal. (a) SAXS analysis of the N 32-383 0 -P 50 complex. The Guinier plot for complex concentrations of 0.55, 1.1,
Supplementary Figure 1 Structural characterization of NiV N 0 P in solution and in crystal. (a) SAXS analysis of the N 32-383 0 -P 50 complex. The Guinier plot for complex concentrations of 0.55, 1.1,
4 Proteins: Structure, Function, Folding W. H. Freeman and Company
 4 Proteins: Structure, Function, Folding 2013 W. H. Freeman and Company CHAPTER 4 Proteins: Structure, Function, Folding Learning goals: Structure and properties of the peptide bond Structural hierarchy
4 Proteins: Structure, Function, Folding 2013 W. H. Freeman and Company CHAPTER 4 Proteins: Structure, Function, Folding Learning goals: Structure and properties of the peptide bond Structural hierarchy
Viewing and Analyzing Proteins, Ligands and their Complexes 2
 2 Viewing and Analyzing Proteins, Ligands and their Complexes 2 Overview Viewing the accessible surface Analyzing the properties of proteins containing thousands of atoms is best accomplished by representing
2 Viewing and Analyzing Proteins, Ligands and their Complexes 2 Overview Viewing the accessible surface Analyzing the properties of proteins containing thousands of atoms is best accomplished by representing
Design of a Novel Globular Protein Fold with Atomic-Level Accuracy
 Design of a Novel Globular Protein Fold with Atomic-Level Accuracy Brian Kuhlman, Gautam Dantas, Gregory C. Ireton, Gabriele Varani, Barry L. Stoddard, David Baker Presented by Kate Stafford 4 May 05 Protein
Design of a Novel Globular Protein Fold with Atomic-Level Accuracy Brian Kuhlman, Gautam Dantas, Gregory C. Ireton, Gabriele Varani, Barry L. Stoddard, David Baker Presented by Kate Stafford 4 May 05 Protein
Tiffany Samaroo MB&B 452a December 8, Take Home Final. Topic 1
 Tiffany Samaroo MB&B 452a December 8, 2003 Take Home Final Topic 1 Prior to 1970, protein and DNA sequence alignment was limited to visual comparison. This was a very tedious process; even proteins with
Tiffany Samaroo MB&B 452a December 8, 2003 Take Home Final Topic 1 Prior to 1970, protein and DNA sequence alignment was limited to visual comparison. This was a very tedious process; even proteins with
Modeling for 3D structure prediction
 Modeling for 3D structure prediction What is a predicted structure? A structure that is constructed using as the sole source of information data obtained from computer based data-mining. However, mixing
Modeling for 3D structure prediction What is a predicted structure? A structure that is constructed using as the sole source of information data obtained from computer based data-mining. However, mixing
Nature Structural and Molecular Biology: doi: /nsmb Supplementary Figure 1. Experimental approach for enhancement of unbiased Fo Fc maps.
 Supplementary Figure 1 Experimental approach for enhancement of unbiased Fo Fc maps. a, c, Unbiased Fo-Fc maps of the Tth 70S post-catalysis complex at 2.55 Å resolution with (a) or without (c) bulk solvent
Supplementary Figure 1 Experimental approach for enhancement of unbiased Fo Fc maps. a, c, Unbiased Fo-Fc maps of the Tth 70S post-catalysis complex at 2.55 Å resolution with (a) or without (c) bulk solvent
Bio nformatics. Lecture 23. Saad Mneimneh
 Bio nformatics Lecture 23 Protein folding The goal is to determine the three-dimensional structure of a protein based on its amino acid sequence Assumption: amino acid sequence completely and uniquely
Bio nformatics Lecture 23 Protein folding The goal is to determine the three-dimensional structure of a protein based on its amino acid sequence Assumption: amino acid sequence completely and uniquely
Lipid Regulated Intramolecular Conformational Dynamics of SNARE-Protein Ykt6
 Supplementary Information for: Lipid Regulated Intramolecular Conformational Dynamics of SNARE-Protein Ykt6 Yawei Dai 1, 2, Markus Seeger 3, Jingwei Weng 4, Song Song 1, 2, Wenning Wang 4, Yan-Wen 1, 2,
Supplementary Information for: Lipid Regulated Intramolecular Conformational Dynamics of SNARE-Protein Ykt6 Yawei Dai 1, 2, Markus Seeger 3, Jingwei Weng 4, Song Song 1, 2, Wenning Wang 4, Yan-Wen 1, 2,
Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA
 Online Supplemental Results Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA Structure solution and overall architecture
Online Supplemental Results Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA Structure solution and overall architecture
Supplementary Information
 Supplementary Information Resveratrol Serves as a Protein-Substrate Interaction Stabilizer in Human SIRT1 Activation Xuben Hou,, David Rooklin, Hao Fang *,,, Yingkai Zhang Department of Medicinal Chemistry
Supplementary Information Resveratrol Serves as a Protein-Substrate Interaction Stabilizer in Human SIRT1 Activation Xuben Hou,, David Rooklin, Hao Fang *,,, Yingkai Zhang Department of Medicinal Chemistry
Protein folding. α-helix. Lecture 21. An α-helix is a simple helix having on average 10 residues (3 turns of the helix)
") Computat onal Biology Lecture 21 Protein folding The goal is to determine the three-dimensional structure of a protein based on its amino acid sequence Assumption: amino acid sequence completely and uniquely
Computat onal Biology Lecture 21 Protein folding The goal is to determine the three-dimensional structure of a protein based on its amino acid sequence Assumption: amino acid sequence completely and uniquely
ml. ph 7.5 ph 6.5 ph 5.5 ph 4.5. β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS
 a UV28 absorption (mau) 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS β 2 AR-Gs complex dissociated complex excess nucleotides b 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex
a UV28 absorption (mau) 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS β 2 AR-Gs complex dissociated complex excess nucleotides b 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Identification of the ScDcp2 minimal region interacting with both ScDcp1 and the ScEdc3 LSm domain. Pull-down experiment of untagged ScEdc3 LSm with various ScDcp1-Dcp2-His 6 fragments.
Supplementary Figure 1 Identification of the ScDcp2 minimal region interacting with both ScDcp1 and the ScEdc3 LSm domain. Pull-down experiment of untagged ScEdc3 LSm with various ScDcp1-Dcp2-His 6 fragments.
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12045 Supplementary Table 1 Data collection and refinement statistics. Native Pt-SAD X-ray source SSRF BL17U SPring-8 BL41XU Wavelength (Å) 0.97947 1.07171 Space group P2 1 2 1 2 1 P2
doi:10.1038/nature12045 Supplementary Table 1 Data collection and refinement statistics. Native Pt-SAD X-ray source SSRF BL17U SPring-8 BL41XU Wavelength (Å) 0.97947 1.07171 Space group P2 1 2 1 2 1 P2
Structural basis for catalytically restrictive dynamics of a high-energy enzyme state
 Supplementary Material Structural basis for catalytically restrictive dynamics of a high-energy enzyme state Michael Kovermann, Jörgen Ådén, Christin Grundström, A. Elisabeth Sauer-Eriksson, Uwe H. Sauer
Supplementary Material Structural basis for catalytically restrictive dynamics of a high-energy enzyme state Michael Kovermann, Jörgen Ådén, Christin Grundström, A. Elisabeth Sauer-Eriksson, Uwe H. Sauer
Supplementary Figure 3 a. Structural comparison between the two determined structures for the IL 23:MA12 complex. The overall RMSD between the two
 Supplementary Figure 1. Biopanningg and clone enrichment of Alphabody binders against human IL 23. Positive clones in i phage ELISA with optical density (OD) 3 times higher than background are shown for
Supplementary Figure 1. Biopanningg and clone enrichment of Alphabody binders against human IL 23. Positive clones in i phage ELISA with optical density (OD) 3 times higher than background are shown for
Examples of Protein Modeling. Protein Modeling. Primary Structure. Protein Structure Description. Protein Sequence Sources. Importing Sequences to MOE
 Examples of Protein Modeling Protein Modeling Visualization Examination of an experimental structure to gain insight about a research question Dynamics To examine the dynamics of protein structures To
Examples of Protein Modeling Protein Modeling Visualization Examination of an experimental structure to gain insight about a research question Dynamics To examine the dynamics of protein structures To
Plasmid Relevant features Source. W18N_D20N and TrXE-W18N_D20N-anti
 Table S1. E. coli plasmids Plasmid Relevant features Source pdg680 T. reesei XynII AA 2-190 with C-terminal His 6 tag optimized for E. coli expression in pjexpress401 Wan et al. (in press) psbn44d psbn44h
Table S1. E. coli plasmids Plasmid Relevant features Source pdg680 T. reesei XynII AA 2-190 with C-terminal His 6 tag optimized for E. coli expression in pjexpress401 Wan et al. (in press) psbn44d psbn44h
Esser et al. Crystal Structures of R. sphaeroides bc 1
 Esser et al. Crystal Structures of R. sphaeroides bc Supplementary Information Trivariate Gaussian Probability Analysis The superposition of six structures results in sextets of 3D coordinates for every
Esser et al. Crystal Structures of R. sphaeroides bc Supplementary Information Trivariate Gaussian Probability Analysis The superposition of six structures results in sextets of 3D coordinates for every
Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions. Implicated in Dimerization and Autoinhibition
 Reveals Regions. Implicated in Dimerization and Autoinhibition") JBC Papers in Press. Published on November 1, 2000 as Manuscript M006502200 Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions Implicated in Dimerization and Autoinhibition 1 Copyright
JBC Papers in Press. Published on November 1, 2000 as Manuscript M006502200 Crystal Structure of Fibroblast Growth Factor 9 (FGF9) Reveals Regions Implicated in Dimerization and Autoinhibition 1 Copyright
Supplementary figure 1. Comparison of unbound ogm-csf and ogm-csf as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine
 Supplementary figure 1. Comparison of unbound and as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine GM-CSF (slate) with bound GM-CSF in the GIF:GM-CSF complex (GIF: green,
Supplementary figure 1. Comparison of unbound and as captured in the GIF:GM-CSF complex. Alignment of two copies of unbound ovine GM-CSF (slate) with bound GM-CSF in the GIF:GM-CSF complex (GIF: green,
Sunhats for plants. How plants detect dangerous ultraviolet rays
 Sunhats for plants How plants detect dangerous ultraviolet rays Anyone who has ever suffered sunburn will know about the effects of too much ultraviolet (UV) radiation, in particular UV-B (from 280-315
Sunhats for plants How plants detect dangerous ultraviolet rays Anyone who has ever suffered sunburn will know about the effects of too much ultraviolet (UV) radiation, in particular UV-B (from 280-315
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Cryo-EM structure and model of the C. thermophilum 90S preribosome. a, Gold standard FSC curve showing the average resolution of the 90S preribosome masked and unmasked (left). FSC
Supplementary Figure 1 Cryo-EM structure and model of the C. thermophilum 90S preribosome. a, Gold standard FSC curve showing the average resolution of the 90S preribosome masked and unmasked (left). FSC
Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets
 Supporting information Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets Wan-Na Chen, Christoph Nitsche, Kala Bharath Pilla, Bim Graham, Thomas
Supporting information Sensitive NMR Approach for Determining the Binding Mode of Tightly Binding Ligand Molecules to Protein Targets Wan-Na Chen, Christoph Nitsche, Kala Bharath Pilla, Bim Graham, Thomas
Homologous proteins have similar structures and structural superposition means to rotate and translate the structures so that corresponding atoms are
 1 Homologous proteins have similar structures and structural superposition means to rotate and translate the structures so that corresponding atoms are as close to each other as possible. Structural similarity
1 Homologous proteins have similar structures and structural superposition means to rotate and translate the structures so that corresponding atoms are as close to each other as possible. Structural similarity
Procheck output. Bond angles (Procheck) Structure verification and validation Bond lengths (Procheck) Introduction to Bioinformatics.
 Structure verification and validation Bond lengths (Procheck) Introduction to Bioinformatics.") Structure verification and validation Bond lengths (Procheck) Introduction to Bioinformatics Iosif Vaisman Email: ivaisman@gmu.edu ----------------------------------------------------------------- Bond
Structure verification and validation Bond lengths (Procheck) Introduction to Bioinformatics Iosif Vaisman Email: ivaisman@gmu.edu ----------------------------------------------------------------- Bond
Secondary and sidechain structures
 Lecture 2 Secondary and sidechain structures James Chou BCMP201 Spring 2008 Images from Petsko & Ringe, Protein Structure and Function. Branden & Tooze, Introduction to Protein Structure. Richardson, J.
Lecture 2 Secondary and sidechain structures James Chou BCMP201 Spring 2008 Images from Petsko & Ringe, Protein Structure and Function. Branden & Tooze, Introduction to Protein Structure. Richardson, J.
Week 10: Homology Modelling (II) - HHpred
 - HHpred") Week 10: Homology Modelling (II) - HHpred Course: Tools for Structural Biology Fabian Glaser BKU - Technion 1 2 Identify and align related structures by sequence methods is not an easy task All comparative
Week 10: Homology Modelling (II) - HHpred Course: Tools for Structural Biology Fabian Glaser BKU - Technion 1 2 Identify and align related structures by sequence methods is not an easy task All comparative
Supplementary Figure S1. Urea-mediated buffering mechanism of H. pylori. Gastric urea is funneled to a cytoplasmic urease that is presumably attached
 Supplementary Figure S1. Urea-mediated buffering mechanism of H. pylori. Gastric urea is funneled to a cytoplasmic urease that is presumably attached to HpUreI. Urea hydrolysis products 2NH 3 and 1CO 2
Supplementary Figure S1. Urea-mediated buffering mechanism of H. pylori. Gastric urea is funneled to a cytoplasmic urease that is presumably attached to HpUreI. Urea hydrolysis products 2NH 3 and 1CO 2
Protein Folding & Stability. Lecture 11: Margaret A. Daugherty. Fall How do we go from an unfolded polypeptide chain to a
 Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2004 How do we go from an unfolded polypeptide chain to a compact folded protein? (Folding of thioredoxin, F. Richards) Structure - Function
Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2004 How do we go from an unfolded polypeptide chain to a compact folded protein? (Folding of thioredoxin, F. Richards) Structure - Function
Packing of Secondary Structures
 7.88 Lecture Notes - 4 7.24/7.88J/5.48J The Protein Folding and Human Disease Professor Gossard Retrieving, Viewing Protein Structures from the Protein Data Base Helix helix packing Packing of Secondary
7.88 Lecture Notes - 4 7.24/7.88J/5.48J The Protein Folding and Human Disease Professor Gossard Retrieving, Viewing Protein Structures from the Protein Data Base Helix helix packing Packing of Secondary
Cks1 CDK1 CDK1 CDK1 CKS1. are ice- lobe. conserved. conserved
 Cks1 d CKS1 Supplementary Figure 1 The -Cks1 crystal lattice. (a) Schematic of the - Cks1 crystal lattice. -Cks1 crystallizes in a lattice that contains c 4 copies of the t - Cks1 dimer in the crystallographic
Cks1 d CKS1 Supplementary Figure 1 The -Cks1 crystal lattice. (a) Schematic of the - Cks1 crystal lattice. -Cks1 crystallizes in a lattice that contains c 4 copies of the t - Cks1 dimer in the crystallographic
Secondary Structure. Bioch/BIMS 503 Lecture 2. Structure and Function of Proteins. Further Reading. Φ, Ψ angles alone determine protein structure
 Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
Homology modeling of Ferredoxin-nitrite reductase from Arabidopsis thaliana
 www.bioinformation.net Hypothesis Volume 6(3) Homology modeling of Ferredoxin-nitrite reductase from Arabidopsis thaliana Karim Kherraz*, Khaled Kherraz, Abdelkrim Kameli Biology department, Ecole Normale
www.bioinformation.net Hypothesis Volume 6(3) Homology modeling of Ferredoxin-nitrite reductase from Arabidopsis thaliana Karim Kherraz*, Khaled Kherraz, Abdelkrim Kameli Biology department, Ecole Normale
SI Text S1 Solution Scattering Data Collection and Analysis. SI references
 SI Text S1 Solution Scattering Data Collection and Analysis. The X-ray photon energy was set to 8 kev. The PILATUS hybrid pixel array detector (RIGAKU) was positioned at a distance of 606 mm from the sample.
SI Text S1 Solution Scattering Data Collection and Analysis. The X-ray photon energy was set to 8 kev. The PILATUS hybrid pixel array detector (RIGAKU) was positioned at a distance of 606 mm from the sample.
Supplementary Figure 1
 A R R RA-selective pocket Cl Adenine pocket and hinge-binding moiety Cl ulfonamide series PLX7 PLX Br BR BR TV PLX RI TQ D RI9 C B PLX7 M ulfonamide concentration Monomer Dimer RA-elective Pocket Unoccupied
A R R RA-selective pocket Cl Adenine pocket and hinge-binding moiety Cl ulfonamide series PLX7 PLX Br BR BR TV PLX RI TQ D RI9 C B PLX7 M ulfonamide concentration Monomer Dimer RA-elective Pocket Unoccupied
Biology Chemistry & Physics of Biomolecules. Examination #1. Proteins Module. September 29, Answer Key
 Biology 5357 Chemistry & Physics of Biomolecules Examination #1 Proteins Module September 29, 2017 Answer Key Question 1 (A) (5 points) Structure (b) is more common, as it contains the shorter connection
Biology 5357 Chemistry & Physics of Biomolecules Examination #1 Proteins Module September 29, 2017 Answer Key Question 1 (A) (5 points) Structure (b) is more common, as it contains the shorter connection