Lecture 2 and 3: Review of forces (ctd.) and elementary statistical mechanics. Contributions to protein stability
|
|
|
- Vivien Scott
- 6 years ago
- Views:
Transcription
1 Lecture 2 and 3: Review of forces (ctd.) and elementary statistical mechanics. Contributions to protein stability Part I. Review of forces Covalent bonds Non-covalent Interactions: Van der Waals Interactions Electrostatics Hydrogen Bonds Hydrophobic Interactions Part II. Review of key concepts from Stat. Mech. Part III. Contributions to protein stability and binding
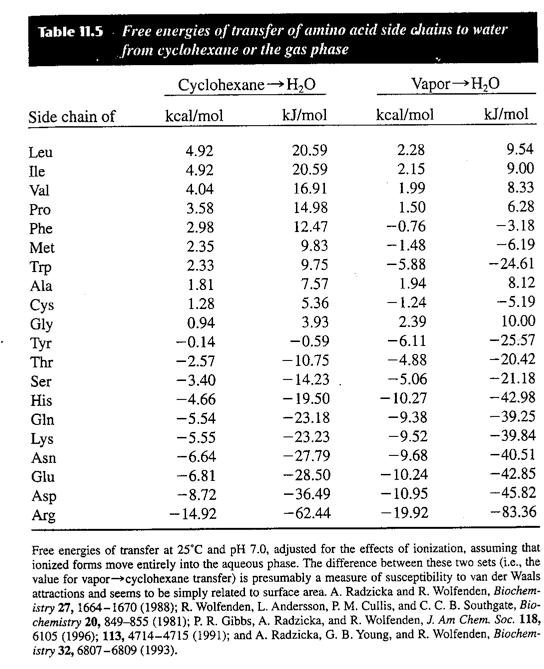
2 Non-Covalent Interactions: Hydrophobic Interactions Solvation/ Hydrophobic interactions The hydrophobic interaction is the pronounced attraction of non-polar solutes in water. Non-polar substances are poorly soluble in water (as is evident in mixing oil and water) and the free energies of transfer of non-polar substances to water are large and positive.
3
4 Origin of the Hydrophobic Effect (1) The origins of the hydrophobic effect are surprisingly still somewhat controversial. It is convenient to divide solvation processes into two steps: 1) the creation of a cavity in the liquid large enough to accommodate the solute, G cavity Bulk H20 (solvent) Solvent + cavity Because water is a strongly cohesive liquid, and because of its small size, the free energy of forming a cavity is higher than in other simple liquids (the probability of finding a reasonably large cavity is quite small). This is probably the main source of the anomalously low solubility of non-polar compounds in water (for polar and charged molecules, this cost is more than offset by the favorable electrostatic and hydrogen bonding interactions that can be formed; see the expression above for transferring an ion to a high dielectric solvent).
5 Origin of the Hydrophobic Effect (2) 2) the placement of the solute into the cavity. + G interactions w/ solvent Solvent + cavity Solute inside cavity The free energy changes associated with 2) are due to interactions between the solvent and the solute that we have already discussed--hydrogen bonding, van der Waals interactions, electrostatics (for non polar compounds, only van der Waals interactions are important). The van der Waals interactions between non-polar solutes and water are of the same order of magnitude as those between water molecules, but to retain hydrogen bonding in the vicinity of the non-polar solutes requires some ordering of water molecules. There is thus also a decrease in entropy associated with exposing non-polar compounds to water and a change in heat capacity which lead to anomalous temperature dependencies that characterize hydrophobic interactions. The first term is unfavorable for all solutes (it depends only in size), while the second is large and favorable for polar and charged solutes, but small for non polar solutes. The sum of these interactions is thus unfavorable only for non-polar solutes-hence the hydrophobic effect.
6 References The topics covered in this lecture are covered in more details in the following textbooks (available in Health Science Library and Biochemistry Library on reserve) Non-covalent interactions relevant to protein structure: Creighton, ch. 4 p139 The same, with an emphasis on Protein-protein interactions: Fersht, ch.11, p324 The three dimensional structure of proteins (α-helix, β-sheet) : Fersht, ch1 p1, Branden & Tooze ch. 1 The Generalized-Born Approximation for Macromolecules: Onufriev et al., Modification of the Generalized Born Model Suitable for Macromolecules, J. Phys. Chem. B. 2000, 104,
7 Some Simple but Key Results from Stat Mech (1) The energy of an isolated system is constant. (2) The entropy is proportional to the logarithm of the number of states of a system: S = k ln Ω (3) The entropy of an isolated system increases in any spontaneous process. (4) For a sub system in thermal equilibrium with a larger system (the outside world), the condition that the entropy of the combined system increases is equivalent to the condition that the free energy of the smaller system, G = E - TS + PV decreases.
8 The Boltzman distribution In most of biochemistry, the PV term is very small, and thus, to determine whether a reaction occurs spontaneously, we must consider the balance between the energy change and the entropy change: ΔG = ΔE - TΔS The probability of observing a particular state of a system with free energy G is: Prob exp [S tot /k] exp [-G/kT] This is a very important result as it relates free energy differences to differences in populations.
9 The Boltzman distribution (ctd.) Consider a protein with two different conformations, conf1 and conf2, that differ in free energy by some amount ΔG. From previous page, Prob(conf1) exp [-G(conf1)/ kt] Prob(conf2) exp [-G(conf2)/ kt] = exp [ -(G(conf1) - ΔG )/kt] The ratio between the populations (concentrations) of the two conformations at equilibrium is called the equilibrium constant (Keq). Keq = Prob(conf1)/Prob(conf2) = exp [-ΔG/kT] Taking the logarithm of both sides gives the familiar expression ΔG = -kt ln K eq which relates the free energy difference in a reaction to the log of the equilibrium constant.
10 The Boltzman distribution (ctd.) For example, if the free energy difference between the two conformations is 1kcal/mol, what is the ratio of the two populations (the equilibrium constant) at 300K? P(conf1)/P(conf2) = exp -(ΔG/kT) = exp -(1/0.6) =.19 It is useful to remember the free energy difference that corresponds to a ten fold difference in the populations of two states in equilibrium: ΔG = -kt ln K eq = -(0.6 ln 10) = kcal/mol
11 Link with Protein Folding/Protein-protein Association Thus, to determine whether a protein will fold or whether two macromolecules will associate, one needs to determine the change in free energy in the process. ΔG = ΔE T ΔS At low temperature, the ΔE term dominates, but as the temperature is increased, the T ΔS term becomes increasingly important. We learned how to compute Δ E for processes involving changes in van der Waals interactions, hydrogen bonds, etc; but how to compute Δ S?
12 Entropic Change in Protein Folding From above, S = k ln (number of states), so we need to determine the change in the number of states during the process. This counting is simplest for amino acid side chains, which adopt a small number of discrete states called rotamers (each torsion angle has three possible values). Example: valine side chains have three possible rotamers (one torsion angle). How much entropy is lost in a change from a conformation in which the valine can adopt all three rotamers to a conformation in which only one rotamer is tolerated? ΔS = k ln 3 k ln 1 = ln 3 0 = kcal/mol K How much free energy does this correspond to? Δfree energy = - T ΔS = (0.0022) = kcal/mol
13 Entropic Change in Protein Folding (cont d) The entropy changes in protein folding are estimated to be ~0.007 kcal/mol K per residue for the main chain, and ~0.003 kcal/mol K per residue for the side chains. For a 100 residue protein the total entropy change in folding is thus ~ 1kcal/mol K; at 300 degrees K (room temperature) this corresponds to ~300 kcal/mol. For a protein to fold, this large unfavorable contribution to the folding free energy must be compensated by the non-covalent interactions discussed last time, which are individually much weaker. (Current Opinion in Structural Biology, 7, )
14 Entropic Change in Macromolecular Association What is the entropic change caused by the association of two macromolecules? There are three components: 1) large decrease in translational entropy 2) decrease in rotational entropy 3) gain in entropy associated with intermolecular motions The net contribution of these three effects is estimated to be ~5-15kcal/mol at 300K.
15
16 The Boltzman distribution In most of biochemistry, the PV term is very small, and thus, to determine whether a reaction occurs spontaneously, we must consider the balance between the energy change and the entropy change: ΔG = ΔE - TΔS The probability of observing a particular state of a system with free energy G is: Prob exp [S tot /k] exp [-G/kT] This is a very important result as it relates free energy differences to differences in populations.
17 Last time, considered case where there were two alternative states. What happens if there are many alternative states? We know for each state: Prob exp [S tot /k] exp [-G/kT] The probabilities of all of the possible states must add up to one. This will be true if we write Prob(state) = exp [-G(state)/kT]/ Z where Z is the sum of the exp [G/kT] terms for each of the states Z is called the partition function
18 Concrete example Suppose a protein has three states: Unfolded state different conformations, each with no interactions Intermediate state. 10 different conformations, each with two hydrogen bonds worth 1kcal/M Native state. 1 conformation, with twenty hydrogen bonds worth 1kcal/M What is the fraction of the protein which is folded as a function of temperature?
19 Putting everything together: Contributions to protein stability
20 General Considerations Since proteins are surrounded by water molecules, the energetics of protein folding and binding involve considerable tradeoffs between loss of protein-water interactions and gain of protein-protein interactions. For example, van der Waals interactions are gained between protein atoms and lost between protein and water, and similarly, hydrogen bonds formed between donors and acceptors within a protein chain come at the cost of breaking hydrogen bonds between these atoms and water. Because of these tradeoffs, the contribution of van der Waals interactions and hydrogen bonding to protein stability is relatively small. However, these interactions do have a very important negative influence on protein structures: van der Waals interactions and hydrogen bonds made with water in unfolded or unbound protein chains that are lost during folding or complex formation must be compensated by interactions within the protein or within the complex, or the free energy of folding/ complex formation will have large unfavorable contributions from the lost interactions with water. For this reason, protein structures rarely contain large cavities (which would involve a loss of van der waals interactions) or buried hydrogen bond donors or acceptors that do not make hydrogen bonds.
21 General Considerations (ctd.) In protein folding and binding reactions, the amount of non-polar surface exposed to water may change considerably, and thus the hydrophobic effect plays an important role in these processes. Unfolding, which involves exposure of hydrophobic side chains to water, may be viewed as equivalent to the transfer of these non-polar groups from a non-polar solvent to water. The requirements of retaining hydrogen bonding and attractive van der Waals interactions while minimizing the exposure of nonpolar atoms to water give rise to the hallmarks of globular protein structures: hydrophobic cores with few charged or polar atoms that are well packed to avoid loss of van der waals interactions, largely polar surfaces, and extensive alpha helix and beta sheet secondary structure which allow the polypeptide backbone to retain extensive hydrogen bonding while passing back and forth through the protein.
22 Entropy Loss and the Denatured State As noted in the previous lecture, the hydrophobic effect is probably the main driving force for protein folding: there is a large decrease in the amount of exposed hydrophobic surface area during folding. Opposing this large favorable contribution is an almost equally large unfavorable contribution from chain entropy loss. Recall that the free energy change Δ G = Δ H T Δ S and that the entropy change Δ S is proportional to the logarithm of the change in the number of accessible states. The native state has much lower entropy than the denatured state because the backbone is relatively fixed and most of the side chains adopt single conformations. There are also contributions from the change in entropy of the solvent when the non-polar groups of the protein become buried in the core of the protein. The net Δ S for folding is negative ( a reduction in the number of states), and thus at high temperature proteins unfold (ΔG for folding becomes positive). Denaturants denature proteins by reducing the strength of the hydrophobic effect. (see following table of transfer free energies of the amino acids from water to denaturants)
23
24 Experimental Data How well do these principles account for observed experimental data on protein stability and binding? Unfortunately, because the free energy of protein folding is the difference between two very large contributions: the large chain entropy loss upon folding and the large gain in hydrophobic interactions, it is not currently possible to predict overall protein stability even from high resolution crystal structures. Much more amenable to analysis are the changes in protein stability brought about by single amino acid changes. Studies of the effects of a large number of such sequence changes have led to the general conclusions listed on the next slide. It should be kept in mind, however, that while something may be inferred about effects of the mutations in the native state if a high resolution structure is available, there is considerable uncertainty about the effects of mutations on the denatured state. ΔG = G native - G denatured
25 Given these considerations, what do you expect would be the effect of mutations which 1) Change a non polar residue to a charged residue in the protein core 2) Change a non polar residue to a charged residue on the protein surface 3) Introduce a flexible residue in the protein core 4) Introduce a flexible residue on the protein surface 5) Change a non polar residue to a charged residue at a protein interface 6) Introduce a flexible residue at a protein interface 7) Increase the size of a residue in the protein core 8) Increase the size of a residue on the protein surface
26 Conclusions From Studies of Protein Stability (1) Sequence changes at buried sites almost always have much larger effects on stability than sequence changes at exposed sites. The small change at exposed sites is not surprising given that these residues are likely to have similar environments (ie, largely solvated) in both the denatured and native states. (See figure on λ repressor on next slide)
27
28 Conclusions From Studies of Protein Stability (ctd.) (2) Charged residues and to a lesser extent polar residues are disfavored at buried sites. This is expected given the large energetic cost of burying a charge. (3) Sequence changes which reduce the amount of hydrophobic burial are destabilizing. (see table on next slide)
29 Conclusions From Studies of Protein Stability (ctd.) Substitution G (kcal/mol) average Ile -> Val 1.3+/- 0.4 Ile -> Ala 3.8 +/- 0.7 Leu -> Ala 3.5 +/- 1.1 Val -> Ala 2.5 +/ CH /- 0.9 Met -> Ala 3.0 +/- 0.9 Phe -> Ala 3.8 +/- 0.3
30 Conclusions From Studies of Protein Stability (ctd.) (4) Sequence changes which disrupt side chain packing in the interior or leave large cavities are unfavorable. Surface area of the cavity (5) Salt bridges between oppositely charged residues on the protein surface contribute relatively little to stability, probably because the more favorable electrostatic interactions are offset by the entropic cost of ordering the sidechains. Repulsive interactions between same charged residues on the surface can be quite destabilizing.
31 Conclusions From Studies of Protein Stability (ctd.) (6) Interactions between negatively charged residues at the N termini of alpha helices with the helix dipole formed by the lining up of all of the dipoles in the individual peptide bonds are stabilizing (this was first noted by one of the more senior lecturers in this course!) A method for predicting the effect of mutations on stability of proteins/protein interfaces is available on the web: (Interface Alanine Scanning)
32 References Most of what has been covered in the lecture on Protein Stability can be found mainly in Creighton, ch4 especially sections 4.1, 4.3 and 4.4. (available on HSB and BIOCHEM libraries)
Lecture 2-3: Review of forces (ctd.) and elementary statistical mechanics. Contributions to protein stability
 and elementary statistical mechanics. Contributions to protein stability") Lecture 2-3: Review of forces (ctd.) and elementary statistical mechanics. Contributions to protein stability Part I. Review of forces Covalent bonds Non-covalent Interactions Van der Waals Interactions
Lecture 2-3: Review of forces (ctd.) and elementary statistical mechanics. Contributions to protein stability Part I. Review of forces Covalent bonds Non-covalent Interactions Van der Waals Interactions
Biochemistry,530:,, Introduc5on,to,Structural,Biology, Autumn,Quarter,2015,
 Biochemistry,530:,, Introduc5on,to,Structural,Biology, Autumn,Quarter,2015, Course,Informa5on, BIOC%530% GraduateAlevel,discussion,of,the,structure,,func5on,,and,chemistry,of,proteins,and, nucleic,acids,,control,of,enzyma5c,reac5ons.,please,see,the,course,syllabus,and,
Biochemistry,530:,, Introduc5on,to,Structural,Biology, Autumn,Quarter,2015, Course,Informa5on, BIOC%530% GraduateAlevel,discussion,of,the,structure,,func5on,,and,chemistry,of,proteins,and, nucleic,acids,,control,of,enzyma5c,reac5ons.,please,see,the,course,syllabus,and,
Biochemistry 530: Introduction to Structural Biology. Autumn Quarter 2014 BIOC 530
 Biochemistry 530: Introduction to Structural Biology Autumn Quarter 2014 Course Information Course Description Graduate-level discussion of the structure, function, and chemistry of proteins and nucleic
Biochemistry 530: Introduction to Structural Biology Autumn Quarter 2014 Course Information Course Description Graduate-level discussion of the structure, function, and chemistry of proteins and nucleic
BIOC : Homework 1 Due 10/10
 Contact information: Name: Student # BIOC530 2012: Homework 1 Due 10/10 Department Email address The following problems are based on David Baker s lectures of forces and protein folding. When numerical
Contact information: Name: Student # BIOC530 2012: Homework 1 Due 10/10 Department Email address The following problems are based on David Baker s lectures of forces and protein folding. When numerical
BIOC 530 Fall, 2011 BIOC 530
 Fall, 2011 Course Information Course Description Graduate-level discussion of the structure, function, and chemistry of proteins and nucleic acids, control of enzymatic reactions. Please see the course
Fall, 2011 Course Information Course Description Graduate-level discussion of the structure, function, and chemistry of proteins and nucleic acids, control of enzymatic reactions. Please see the course
= (-22) = +2kJ /mol
 = +2kJ /mol") Lecture 8: Thermodynamics & Protein Stability Assigned reading in Campbell: Chapter 4.4-4.6 Key Terms: DG = -RT lnk eq = DH - TDS Transition Curve, Melting Curve, Tm DH calculation DS calculation van der
Lecture 8: Thermodynamics & Protein Stability Assigned reading in Campbell: Chapter 4.4-4.6 Key Terms: DG = -RT lnk eq = DH - TDS Transition Curve, Melting Curve, Tm DH calculation DS calculation van der
Why Proteins Fold. How Proteins Fold? e - ΔG/kT. Protein Folding, Nonbonding Forces, and Free Energy
 Why Proteins Fold Proteins are the action superheroes of the body. As enzymes, they make reactions go a million times faster. As versatile transport vehicles, they carry oxygen and antibodies to fight
Why Proteins Fold Proteins are the action superheroes of the body. As enzymes, they make reactions go a million times faster. As versatile transport vehicles, they carry oxygen and antibodies to fight
Section Week 3. Junaid Malek, M.D.
 Section Week 3 Junaid Malek, M.D. Biological Polymers DA 4 monomers (building blocks), limited structure (double-helix) RA 4 monomers, greater flexibility, multiple structures Proteins 20 Amino Acids,
Section Week 3 Junaid Malek, M.D. Biological Polymers DA 4 monomers (building blocks), limited structure (double-helix) RA 4 monomers, greater flexibility, multiple structures Proteins 20 Amino Acids,
Solutions and Non-Covalent Binding Forces
 Chapter 3 Solutions and Non-Covalent Binding Forces 3.1 Solvent and solution properties Molecules stick together using the following forces: dipole-dipole, dipole-induced dipole, hydrogen bond, van der
Chapter 3 Solutions and Non-Covalent Binding Forces 3.1 Solvent and solution properties Molecules stick together using the following forces: dipole-dipole, dipole-induced dipole, hydrogen bond, van der
Biochemistry Prof. S. DasGupta Department of Chemistry Indian Institute of Technology Kharagpur. Lecture - 06 Protein Structure IV
 Biochemistry Prof. S. DasGupta Department of Chemistry Indian Institute of Technology Kharagpur Lecture - 06 Protein Structure IV We complete our discussion on Protein Structures today. And just to recap
Biochemistry Prof. S. DasGupta Department of Chemistry Indian Institute of Technology Kharagpur Lecture - 06 Protein Structure IV We complete our discussion on Protein Structures today. And just to recap
The protein folding problem consists of two parts:
 Energetics and kinetics of protein folding The protein folding problem consists of two parts: 1)Creating a stable, well-defined structure that is significantly more stable than all other possible structures.
Energetics and kinetics of protein folding The protein folding problem consists of two parts: 1)Creating a stable, well-defined structure that is significantly more stable than all other possible structures.
Protein Structure. W. M. Grogan, Ph.D. OBJECTIVES
 Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
From Amino Acids to Proteins - in 4 Easy Steps
 From Amino Acids to Proteins - in 4 Easy Steps Although protein structure appears to be overwhelmingly complex, you can provide your students with a basic understanding of how proteins fold by focusing
From Amino Acids to Proteins - in 4 Easy Steps Although protein structure appears to be overwhelmingly complex, you can provide your students with a basic understanding of how proteins fold by focusing
Free energy, electrostatics, and the hydrophobic effect
 Protein Physics 2016 Lecture 3, January 26 Free energy, electrostatics, and the hydrophobic effect Magnus Andersson magnus.andersson@scilifelab.se Theoretical & Computational Biophysics Recap Protein structure
Protein Physics 2016 Lecture 3, January 26 Free energy, electrostatics, and the hydrophobic effect Magnus Andersson magnus.andersson@scilifelab.se Theoretical & Computational Biophysics Recap Protein structure
Lecture 34 Protein Unfolding Thermodynamics
 Physical Principles in Biology Biology 3550 Fall 2018 Lecture 34 Protein Unfolding Thermodynamics Wednesday, 21 November c David P. Goldenberg University of Utah goldenberg@biology.utah.edu Clicker Question
Physical Principles in Biology Biology 3550 Fall 2018 Lecture 34 Protein Unfolding Thermodynamics Wednesday, 21 November c David P. Goldenberg University of Utah goldenberg@biology.utah.edu Clicker Question
Protein Folding & Stability. Lecture 11: Margaret A. Daugherty. Fall How do we go from an unfolded polypeptide chain to a
 Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2004 How do we go from an unfolded polypeptide chain to a compact folded protein? (Folding of thioredoxin, F. Richards) Structure - Function
Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2004 How do we go from an unfolded polypeptide chain to a compact folded protein? (Folding of thioredoxin, F. Richards) Structure - Function
Other Cells. Hormones. Viruses. Toxins. Cell. Bacteria
 Other Cells Hormones Viruses Toxins Cell Bacteria ΔH < 0 reaction is exothermic, tells us nothing about the spontaneity of the reaction Δ H > 0 reaction is endothermic, tells us nothing about the spontaneity
Other Cells Hormones Viruses Toxins Cell Bacteria ΔH < 0 reaction is exothermic, tells us nothing about the spontaneity of the reaction Δ H > 0 reaction is endothermic, tells us nothing about the spontaneity
THE TANGO ALGORITHM: SECONDARY STRUCTURE PROPENSITIES, STATISTICAL MECHANICS APPROXIMATION
 THE TANGO ALGORITHM: SECONDARY STRUCTURE PROPENSITIES, STATISTICAL MECHANICS APPROXIMATION AND CALIBRATION Calculation of turn and beta intrinsic propensities. A statistical analysis of a protein structure
THE TANGO ALGORITHM: SECONDARY STRUCTURE PROPENSITIES, STATISTICAL MECHANICS APPROXIMATION AND CALIBRATION Calculation of turn and beta intrinsic propensities. A statistical analysis of a protein structure
Biophysics II. Hydrophobic Bio-molecules. Key points to be covered. Molecular Interactions in Bio-molecular Structures - van der Waals Interaction
 Biophysics II Key points to be covered By A/Prof. Xiang Yang Liu Biophysics & Micro/nanostructures Lab Department of Physics, NUS 1. van der Waals Interaction 2. Hydrogen bond 3. Hydrophilic vs hydrophobic
Biophysics II Key points to be covered By A/Prof. Xiang Yang Liu Biophysics & Micro/nanostructures Lab Department of Physics, NUS 1. van der Waals Interaction 2. Hydrogen bond 3. Hydrophilic vs hydrophobic
Chapter 1. Topic: Overview of basic principles
 Chapter 1 Topic: Overview of basic principles Four major themes of biochemistry I. What are living organism made from? II. How do organism acquire and use energy? III. How does an organism maintain its
Chapter 1 Topic: Overview of basic principles Four major themes of biochemistry I. What are living organism made from? II. How do organism acquire and use energy? III. How does an organism maintain its
Biology Chemistry & Physics of Biomolecules. Examination #1. Proteins Module. September 29, Answer Key
 Biology 5357 Chemistry & Physics of Biomolecules Examination #1 Proteins Module September 29, 2017 Answer Key Question 1 (A) (5 points) Structure (b) is more common, as it contains the shorter connection
Biology 5357 Chemistry & Physics of Biomolecules Examination #1 Proteins Module September 29, 2017 Answer Key Question 1 (A) (5 points) Structure (b) is more common, as it contains the shorter connection
Problem Set 1
 2006 7.012 Problem Set 1 Due before 5 PM on FRIDAY, September 15, 2006. Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. For each of the following parts, pick
2006 7.012 Problem Set 1 Due before 5 PM on FRIDAY, September 15, 2006. Turn answers in to the box outside of 68-120. PLEASE WRITE YOUR ANSWERS ON THIS PRINTOUT. 1. For each of the following parts, pick
BIBC 100. Structural Biochemistry
 BIBC 100 Structural Biochemistry http://classes.biology.ucsd.edu/bibc100.wi14 Papers- Dialogue with Scientists Questions: Why? How? What? So What? Dialogue Structure to explain function Knowledge Food
BIBC 100 Structural Biochemistry http://classes.biology.ucsd.edu/bibc100.wi14 Papers- Dialogue with Scientists Questions: Why? How? What? So What? Dialogue Structure to explain function Knowledge Food
schematic diagram; EGF binding, dimerization, phosphorylation, Grb2 binding, etc.
 Lecture 1: Noncovalent Biomolecular Interactions Bioengineering and Modeling of biological processes -e.g. tissue engineering, cancer, autoimmune disease Example: RTK signaling, e.g. EGFR Growth responses
Lecture 1: Noncovalent Biomolecular Interactions Bioengineering and Modeling of biological processes -e.g. tissue engineering, cancer, autoimmune disease Example: RTK signaling, e.g. EGFR Growth responses
Lecture 11: Protein Folding & Stability
 Structure - Function Protein Folding: What we know Lecture 11: Protein Folding & Stability 1). Amino acid sequence dictates structure. 2). The native structure represents the lowest energy state for a
Structure - Function Protein Folding: What we know Lecture 11: Protein Folding & Stability 1). Amino acid sequence dictates structure. 2). The native structure represents the lowest energy state for a
Protein Folding & Stability. Lecture 11: Margaret A. Daugherty. Fall Protein Folding: What we know. Protein Folding
 Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2003 Structure - Function Protein Folding: What we know 1). Amino acid sequence dictates structure. 2). The native structure represents
Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2003 Structure - Function Protein Folding: What we know 1). Amino acid sequence dictates structure. 2). The native structure represents
3. Solutions W = N!/(N A!N B!) (3.1) Using Stirling s approximation ln(n!) = NlnN N: ΔS mix = k (N A lnn + N B lnn N A lnn A N B lnn B ) (3.
 (3.1) Using Stirling s approximation ln(n!) = NlnN N: ΔS mix = k (N A lnn + N B lnn N A lnn A N B lnn B ) (3.") 3. Solutions Many biological processes occur between molecules in aqueous solution. In addition, many protein and nucleic acid molecules adopt three-dimensional structure ( fold ) in aqueous solution.
3. Solutions Many biological processes occur between molecules in aqueous solution. In addition, many protein and nucleic acid molecules adopt three-dimensional structure ( fold ) in aqueous solution.
MCB100A/Chem130 MidTerm Exam 2 April 4, 2013
 MCBA/Chem Miderm Exam 2 April 4, 2 Name Student ID rue/false (2 points each).. he Boltzmann constant, k b sets the energy scale for observing energy microstates 2. Atoms with favorable electronic configurations
MCBA/Chem Miderm Exam 2 April 4, 2 Name Student ID rue/false (2 points each).. he Boltzmann constant, k b sets the energy scale for observing energy microstates 2. Atoms with favorable electronic configurations
Aqueous solutions. Solubility of different compounds in water
 Aqueous solutions Solubility of different compounds in water The dissolution of molecules into water (in any solvent actually) causes a volume change of the solution; the size of this volume change is
Aqueous solutions Solubility of different compounds in water The dissolution of molecules into water (in any solvent actually) causes a volume change of the solution; the size of this volume change is
Dihedral Angles. Homayoun Valafar. Department of Computer Science and Engineering, USC 02/03/10 CSCE 769
 Dihedral Angles Homayoun Valafar Department of Computer Science and Engineering, USC The precise definition of a dihedral or torsion angle can be found in spatial geometry Angle between to planes Dihedral
Dihedral Angles Homayoun Valafar Department of Computer Science and Engineering, USC The precise definition of a dihedral or torsion angle can be found in spatial geometry Angle between to planes Dihedral
Biotechnology of Proteins. The Source of Stability in Proteins (III) Fall 2015
 Fall 2015") Biotechnology of Proteins The Source of Stability in Proteins (III) Fall 2015 Conformational Entropy of Unfolding It is The factor that makes the greatest contribution to stabilization of the unfolded
Biotechnology of Proteins The Source of Stability in Proteins (III) Fall 2015 Conformational Entropy of Unfolding It is The factor that makes the greatest contribution to stabilization of the unfolded
Protein Folding experiments and theory
 Protein Folding experiments and theory 1, 2,and 3 Protein Structure Fig. 3-16 from Lehninger Biochemistry, 4 th ed. The 3D structure is not encoded at the single aa level Hydrogen Bonding Shared H atom
Protein Folding experiments and theory 1, 2,and 3 Protein Structure Fig. 3-16 from Lehninger Biochemistry, 4 th ed. The 3D structure is not encoded at the single aa level Hydrogen Bonding Shared H atom
Dana Alsulaibi. Jaleel G.Sweis. Mamoon Ahram
 15 Dana Alsulaibi Jaleel G.Sweis Mamoon Ahram Revision of last lectures: Proteins have four levels of structures. Primary,secondary, tertiary and quaternary. Primary structure is the order of amino acids
15 Dana Alsulaibi Jaleel G.Sweis Mamoon Ahram Revision of last lectures: Proteins have four levels of structures. Primary,secondary, tertiary and quaternary. Primary structure is the order of amino acids
Flexibility of Protein Structure
 Flexibility of Protein Structure Proteins show varying degree of conformational flexibility Due to movements of atoms in molecules vibration in bond length and angles Reflects the existence of populations
Flexibility of Protein Structure Proteins show varying degree of conformational flexibility Due to movements of atoms in molecules vibration in bond length and angles Reflects the existence of populations
LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor
 LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor Note: Adequate space is given for each answer. Questions that require a brief explanation should
LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor Note: Adequate space is given for each answer. Questions that require a brief explanation should
Chem. 27 Section 1 Conformational Analysis Week of Feb. 6, TF: Walter E. Kowtoniuk Mallinckrodt 303 Liu Laboratory
 Chem. 27 Section 1 Conformational Analysis TF: Walter E. Kowtoniuk wekowton@fas.harvard.edu Mallinckrodt 303 Liu Laboratory ffice hours are: Monday and Wednesday 3:00-4:00pm in Mallinckrodt 303 Course
Chem. 27 Section 1 Conformational Analysis TF: Walter E. Kowtoniuk wekowton@fas.harvard.edu Mallinckrodt 303 Liu Laboratory ffice hours are: Monday and Wednesday 3:00-4:00pm in Mallinckrodt 303 Course
Charged amino acids (side-chains)
") Proteins are composed of monomers called amino acids There are 20 different amino acids Amine Group Central ydrocarbon N C C R Group Carboxyl Group ALL amino acids have the exact same structure except
Proteins are composed of monomers called amino acids There are 20 different amino acids Amine Group Central ydrocarbon N C C R Group Carboxyl Group ALL amino acids have the exact same structure except
Packing of Secondary Structures
 7.88 Lecture Notes - 4 7.24/7.88J/5.48J The Protein Folding and Human Disease Professor Gossard Retrieving, Viewing Protein Structures from the Protein Data Base Helix helix packing Packing of Secondary
7.88 Lecture Notes - 4 7.24/7.88J/5.48J The Protein Folding and Human Disease Professor Gossard Retrieving, Viewing Protein Structures from the Protein Data Base Helix helix packing Packing of Secondary
Supplementary Information
 1 Supplementary Information Figure S1 The V=0.5 Harker section of an anomalous difference Patterson map calculated using diffraction data from the NNQQNY crystal at 1.3 Å resolution. The position of the
1 Supplementary Information Figure S1 The V=0.5 Harker section of an anomalous difference Patterson map calculated using diffraction data from the NNQQNY crystal at 1.3 Å resolution. The position of the
What binds to Hb in addition to O 2?
 Reading: Ch5; 158-169, 162-166, 169-174 Problems: Ch5 (text); 3,7,8,10 Ch5 (study guide-facts); 1,2,3,4,5,8 Ch5 (study guide-apply); 2,3 Remember Today at 5:30 in CAS-522 is the second chance for the MB
Reading: Ch5; 158-169, 162-166, 169-174 Problems: Ch5 (text); 3,7,8,10 Ch5 (study guide-facts); 1,2,3,4,5,8 Ch5 (study guide-apply); 2,3 Remember Today at 5:30 in CAS-522 is the second chance for the MB
Lec.1 Chemistry Of Water
 Lec.1 Chemistry Of Water Biochemistry & Medicine Biochemistry can be defined as the science concerned with the chemical basis of life. Biochemistry can be described as the science concerned with the chemical
Lec.1 Chemistry Of Water Biochemistry & Medicine Biochemistry can be defined as the science concerned with the chemical basis of life. Biochemistry can be described as the science concerned with the chemical
Biomolecules: lecture 10
 Biomolecules: lecture 10 - understanding in detail how protein 3D structures form - realize that protein molecules are not static wire models but instead dynamic, where in principle every atom moves (yet
Biomolecules: lecture 10 - understanding in detail how protein 3D structures form - realize that protein molecules are not static wire models but instead dynamic, where in principle every atom moves (yet
Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability
 Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability Dr. Andrew Lee UNC School of Pharmacy (Div. Chemical Biology and Medicinal Chemistry) UNC Med
Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability Dr. Andrew Lee UNC School of Pharmacy (Div. Chemical Biology and Medicinal Chemistry) UNC Med
PROTEIN STRUCTURE AMINO ACIDS H R. Zwitterion (dipolar ion) CO 2 H. PEPTIDES Formal reactions showing formation of peptide bond by dehydration:
 CO 2 H. PEPTIDES Formal reactions showing formation of peptide bond by dehydration:") PTEI STUTUE ydrolysis of proteins with aqueous acid or base yields a mixture of free amino acids. Each type of protein yields a characteristic mixture of the ~ 20 amino acids. AMI AIDS Zwitterion (dipolar
PTEI STUTUE ydrolysis of proteins with aqueous acid or base yields a mixture of free amino acids. Each type of protein yields a characteristic mixture of the ~ 20 amino acids. AMI AIDS Zwitterion (dipolar
MCB100A/Chem130 MidTerm Exam 2 April 4, 2013
 MCB1A/Chem13 MidTerm Exam 2 April 4, 213 Name Student ID True/False (2 points each). 1. The Boltzmann constant, k b T sets the energy scale for observing energy microstates 2. Atoms with favorable electronic
MCB1A/Chem13 MidTerm Exam 2 April 4, 213 Name Student ID True/False (2 points each). 1. The Boltzmann constant, k b T sets the energy scale for observing energy microstates 2. Atoms with favorable electronic
Introduction to Comparative Protein Modeling. Chapter 4 Part I
 Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Short Announcements. 1 st Quiz today: 15 minutes. Homework 3: Due next Wednesday.
 Short Announcements 1 st Quiz today: 15 minutes Homework 3: Due next Wednesday. Next Lecture, on Visualizing Molecular Dynamics (VMD) by Klaus Schulten Today s Lecture: Protein Folding, Misfolding, Aggregation
Short Announcements 1 st Quiz today: 15 minutes Homework 3: Due next Wednesday. Next Lecture, on Visualizing Molecular Dynamics (VMD) by Klaus Schulten Today s Lecture: Protein Folding, Misfolding, Aggregation
Protein folding. Today s Outline
 Protein folding Today s Outline Review of previous sessions Thermodynamics of folding and unfolding Determinants of folding Techniques for measuring folding The folding process The folding problem: Prediction
Protein folding Today s Outline Review of previous sessions Thermodynamics of folding and unfolding Determinants of folding Techniques for measuring folding The folding process The folding problem: Prediction
Protein Folding. I. Characteristics of proteins. C α
 I. Characteristics of proteins Protein Folding 1. Proteins are one of the most important molecules of life. They perform numerous functions, from storing oxygen in tissues or transporting it in a blood
I. Characteristics of proteins Protein Folding 1. Proteins are one of the most important molecules of life. They perform numerous functions, from storing oxygen in tissues or transporting it in a blood
Thermodynamics. Entropy and its Applications. Lecture 11. NC State University
 Thermodynamics Entropy and its Applications Lecture 11 NC State University System and surroundings Up to this point we have considered the system, but we have not concerned ourselves with the relationship
Thermodynamics Entropy and its Applications Lecture 11 NC State University System and surroundings Up to this point we have considered the system, but we have not concerned ourselves with the relationship
BCH 4053 Exam I Review Spring 2017
 BCH 4053 SI - Spring 2017 Reed BCH 4053 Exam I Review Spring 2017 Chapter 1 1. Calculate G for the reaction A + A P + Q. Assume the following equilibrium concentrations: [A] = 20mM, [Q] = [P] = 40fM. Assume
BCH 4053 SI - Spring 2017 Reed BCH 4053 Exam I Review Spring 2017 Chapter 1 1. Calculate G for the reaction A + A P + Q. Assume the following equilibrium concentrations: [A] = 20mM, [Q] = [P] = 40fM. Assume
Proton Acidity. (b) For the following reaction, draw the arrowhead properly to indicate the position of the equilibrium: HA + K + B -
 For the following reaction, draw the arrowhead properly to indicate the position of the equilibrium: HA + K + B -") Proton Acidity A01 Given that acid A has a pk a of 15 and acid B has a pk a of 10, then: (a) Which of the two acids is stronger? (b) For the following reaction, draw the arrowhead properly to indicate
Proton Acidity A01 Given that acid A has a pk a of 15 and acid B has a pk a of 10, then: (a) Which of the two acids is stronger? (b) For the following reaction, draw the arrowhead properly to indicate
Dental Biochemistry Exam The total number of unique tripeptides that can be produced using all of the common 20 amino acids is
 Exam Questions for Dental Biochemistry Monday August 27, 2007 E.J. Miller 1. The compound shown below is CH 3 -CH 2 OH A. acetoacetate B. acetic acid C. acetaldehyde D. produced by reduction of acetaldehyde
Exam Questions for Dental Biochemistry Monday August 27, 2007 E.J. Miller 1. The compound shown below is CH 3 -CH 2 OH A. acetoacetate B. acetic acid C. acetaldehyde D. produced by reduction of acetaldehyde
Central Dogma. modifications genome transcriptome proteome
 entral Dogma DA ma protein post-translational modifications genome transcriptome proteome 83 ierarchy of Protein Structure 20 Amino Acids There are 20 n possible sequences for a protein of n residues!
entral Dogma DA ma protein post-translational modifications genome transcriptome proteome 83 ierarchy of Protein Structure 20 Amino Acids There are 20 n possible sequences for a protein of n residues!
Basics of Thermodynamics: Easy learning by Dr. Anjana Sen
 Basics of Thermodynamics: Easy learning by Dr. Anjana Sen Part 1: Theory and concept Part 2: Definitions and equations Part 3: Laws of Thermodynamics Part 1: theory and concept Thermodynamics means conversion
Basics of Thermodynamics: Easy learning by Dr. Anjana Sen Part 1: Theory and concept Part 2: Definitions and equations Part 3: Laws of Thermodynamics Part 1: theory and concept Thermodynamics means conversion
Secondary Structure. Bioch/BIMS 503 Lecture 2. Structure and Function of Proteins. Further Reading. Φ, Ψ angles alone determine protein structure
 Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
16 years ago TODAY (9/11) at 8:46, the first tower was hit at 9:03, the second tower was hit. Lecture 2 (9/11/17)
 at 8:46, the first tower was hit at 9:03, the second tower was hit. Lecture 2 (9/11/17)") 16 years ago TODAY (9/11) at 8:46, the first tower was hit at 9:03, the second tower was hit By Anthony Quintano - https://www.flickr.com/photos/quintanomedia/15071865580, CC BY 2.0, https://commons.wikimedia.org/w/index.php?curid=38538291
16 years ago TODAY (9/11) at 8:46, the first tower was hit at 9:03, the second tower was hit By Anthony Quintano - https://www.flickr.com/photos/quintanomedia/15071865580, CC BY 2.0, https://commons.wikimedia.org/w/index.php?curid=38538291
IV. SOLVATION - CLASSICAL THERMODYNAMICS - STABILITY
 1 IV. SOLVATION - CLASSICAL THERMODYNAMICS - STABILITY 1. Driving forces in protein folding a. Hydrophobicity is the dominant force driving protein folding. Protein folding was initially thought to be
1 IV. SOLVATION - CLASSICAL THERMODYNAMICS - STABILITY 1. Driving forces in protein folding a. Hydrophobicity is the dominant force driving protein folding. Protein folding was initially thought to be
Biological Thermodynamics
 Biological Thermodynamics Classical thermodynamics is the only physical theory of universal content concerning which I am convinced that, within the framework of applicability of its basic contents, will
Biological Thermodynamics Classical thermodynamics is the only physical theory of universal content concerning which I am convinced that, within the framework of applicability of its basic contents, will
Denaturation and renaturation of proteins
 Denaturation and renaturation of proteins Higher levels of protein structure are formed without covalent bonds. Therefore, they are not as stable as peptide covalent bonds which make protein primary structure
Denaturation and renaturation of proteins Higher levels of protein structure are formed without covalent bonds. Therefore, they are not as stable as peptide covalent bonds which make protein primary structure
CHEM 3653 Exam # 1 (03/07/13)
") 1. Using phylogeny all living organisms can be divided into the following domains: A. Bacteria, Eukarya, and Vertebrate B. Archaea and Eukarya C. Bacteria, Eukarya, and Archaea D. Eukarya and Bacteria
1. Using phylogeny all living organisms can be divided into the following domains: A. Bacteria, Eukarya, and Vertebrate B. Archaea and Eukarya C. Bacteria, Eukarya, and Archaea D. Eukarya and Bacteria
Overview. The peptide bond. Page 1
 Overview Secondary structure: the conformation of the peptide backbone The peptide bond, steric implications Steric hindrance and sterically allowed conformations. Ramachandran diagrams Side chain conformations
Overview Secondary structure: the conformation of the peptide backbone The peptide bond, steric implications Steric hindrance and sterically allowed conformations. Ramachandran diagrams Side chain conformations
A) at equilibrium B) endergonic C) endothermic D) exergonic E) exothermic.
 at equilibrium B) endergonic C) endothermic D) exergonic E) exothermic.") CHEM 2770: Elements of Biochemistry Mid Term EXAMINATION VERSION A Date: October 29, 2014 Instructor: H. Perreault Location: 172 Schultz Time: 4 or 6 pm. Duration: 1 hour Instructions Please mark the Answer
CHEM 2770: Elements of Biochemistry Mid Term EXAMINATION VERSION A Date: October 29, 2014 Instructor: H. Perreault Location: 172 Schultz Time: 4 or 6 pm. Duration: 1 hour Instructions Please mark the Answer
Introduction into Biochemistry. Dr. Mamoun Ahram Lecture 1
 Introduction into Biochemistry Dr. Mamoun Ahram Lecture 1 Course information Recommended textbooks Biochemistry; Mary K. Campbell and Shawn O. Farrell, Brooks Cole; 7 th edition Instructors Dr. Mamoun
Introduction into Biochemistry Dr. Mamoun Ahram Lecture 1 Course information Recommended textbooks Biochemistry; Mary K. Campbell and Shawn O. Farrell, Brooks Cole; 7 th edition Instructors Dr. Mamoun
Full file at https://fratstock.eu
 Chapter 03 1. a. DG=DH-TDS Δ G = 80 kj ( 98 K) 0.790 kj = 44.6 kj K b. ΔG = 0 @ T m. Unfolding will be favorable at temperatures above the T m. Δ G =Δ H TΔ S 0 kj kj 80 ( xk) 0.790 K 0 Δ G = = 354.4 K
Chapter 03 1. a. DG=DH-TDS Δ G = 80 kj ( 98 K) 0.790 kj = 44.6 kj K b. ΔG = 0 @ T m. Unfolding will be favorable at temperatures above the T m. Δ G =Δ H TΔ S 0 kj kj 80 ( xk) 0.790 K 0 Δ G = = 354.4 K
Physiochemical Properties of Residues
 Physiochemical Properties of Residues Various Sources C N Cα R Slide 1 Conformational Propensities Conformational Propensity is the frequency in which a residue adopts a given conformation (in a polypeptide)
Physiochemical Properties of Residues Various Sources C N Cα R Slide 1 Conformational Propensities Conformational Propensity is the frequency in which a residue adopts a given conformation (in a polypeptide)
Water. 2.1 Weak Interactions in Aqueous Sy stems Ionization of Water, Weak Acids, and Weak Bases 58
 Home http://www.macmillanhighered.com/launchpad/lehninger6e... 1 of 1 1/6/2016 3:07 PM 2 Printed Page 47 Water 2.1 Weak Interactions in Aqueous Sy stems 47 2.2 Ionization of Water, Weak Acids, and Weak
Home http://www.macmillanhighered.com/launchpad/lehninger6e... 1 of 1 1/6/2016 3:07 PM 2 Printed Page 47 Water 2.1 Weak Interactions in Aqueous Sy stems 47 2.2 Ionization of Water, Weak Acids, and Weak
Lattice protein models
 Lattice protein models Marc R. Roussel epartment of Chemistry and Biochemistry University of Lethbridge March 5, 2009 1 Model and assumptions The ideas developed in the last few lectures can be applied
Lattice protein models Marc R. Roussel epartment of Chemistry and Biochemistry University of Lethbridge March 5, 2009 1 Model and assumptions The ideas developed in the last few lectures can be applied
Saba Al Fayoumi. Tamer Barakat. Dr. Mamoun Ahram + Dr. Diala Abu-Hassan
 1 Saba Al Fayoumi Tamer Barakat Dr. Mamoun Ahram + Dr. Diala Abu-Hassan What is BIOCHEMISTRY??? Biochemistry = understanding life Chemical reactions are what makes an organism (An organism is simply atoms
1 Saba Al Fayoumi Tamer Barakat Dr. Mamoun Ahram + Dr. Diala Abu-Hassan What is BIOCHEMISTRY??? Biochemistry = understanding life Chemical reactions are what makes an organism (An organism is simply atoms
Dental Biochemistry EXAM I
 Dental Biochemistry EXAM I August 29, 2005 In the reaction below: CH 3 -CH 2 OH -~ ethanol CH 3 -CHO acetaldehyde A. acetoacetate is being produced B. ethanol is being oxidized to acetaldehyde C. acetaldehyde
Dental Biochemistry EXAM I August 29, 2005 In the reaction below: CH 3 -CH 2 OH -~ ethanol CH 3 -CHO acetaldehyde A. acetoacetate is being produced B. ethanol is being oxidized to acetaldehyde C. acetaldehyde
AN INVESTIGATION INTO THE DEPENDENCE OF EGG PROTEIN DENATURATION ON TEMPERATURE.
 AN INVESTIGATION INTO THE DEPENDENCE OF EGG PROTEIN DENATURATION ON TEMPERATURE. Aim The aim of this investigation was to investigate how the rate of denaturation of egg white proteins is dependent on
AN INVESTIGATION INTO THE DEPENDENCE OF EGG PROTEIN DENATURATION ON TEMPERATURE. Aim The aim of this investigation was to investigate how the rate of denaturation of egg white proteins is dependent on
Biomolecules: lecture 9
 Biomolecules: lecture 9 - understanding further why amino acids are the building block for proteins - understanding the chemical properties amino acids bring to proteins - realizing that many proteins
Biomolecules: lecture 9 - understanding further why amino acids are the building block for proteins - understanding the chemical properties amino acids bring to proteins - realizing that many proteins
High Specificity and Reversibility
 Lecture #8 The Cell as a Machine High Specificity and Reversibility In considering the problem of transcription factor binding in the nucleus and the great specificity that is called for to transcribe
Lecture #8 The Cell as a Machine High Specificity and Reversibility In considering the problem of transcription factor binding in the nucleus and the great specificity that is called for to transcribe
Some properties of water
 Some properties of water Hydrogen bond network Solvation under the microscope 1 Water solutions Oil and water does not mix at equilibrium essentially due to entropy Substances that does not mix with water
Some properties of water Hydrogen bond network Solvation under the microscope 1 Water solutions Oil and water does not mix at equilibrium essentially due to entropy Substances that does not mix with water
Life Sciences 1a Lecture Slides Set 10 Fall Prof. David R. Liu. Lecture Readings. Required: Lecture Notes McMurray p , O NH
 Life ciences 1a Lecture lides et 10 Fall 2006-2007 Prof. David R. Liu Lectures 17-18: The molecular basis of drug-protein binding: IV protease inhibitors 1. Drug development and its impact on IV-infected
Life ciences 1a Lecture lides et 10 Fall 2006-2007 Prof. David R. Liu Lectures 17-18: The molecular basis of drug-protein binding: IV protease inhibitors 1. Drug development and its impact on IV-infected
6 Hydrophobic interactions
 The Physics and Chemistry of Water 6 Hydrophobic interactions A non-polar molecule in water disrupts the H- bond structure by forcing some water molecules to give up their hydrogen bonds. As a result,
The Physics and Chemistry of Water 6 Hydrophobic interactions A non-polar molecule in water disrupts the H- bond structure by forcing some water molecules to give up their hydrogen bonds. As a result,
Water. Dr. Diala Abu-Hassan, DDS, PhD Lecture 2 MD summer Dr. Diala Abu-Hassan
 Water, DDS, PhD Dr.abuhassand@gmail.com Lecture 2 MD summer 2014 1 Lecture Content Importance of water in biological systems Noncovalent interactions Water structure Water properties Water as a solvent
Water, DDS, PhD Dr.abuhassand@gmail.com Lecture 2 MD summer 2014 1 Lecture Content Importance of water in biological systems Noncovalent interactions Water structure Water properties Water as a solvent
BIOCHEMISTRY Unit 2 Part 4 ACTIVITY #6 (Chapter 5) PROTEINS
 PROTEINS") BIOLOGY BIOCHEMISTRY Unit 2 Part 4 ACTIVITY #6 (Chapter 5) NAME NAME PERIOD PROTEINS GENERAL CHARACTERISTICS AND IMPORTANCES: Polymers of amino acids Each has unique 3-D shape Vary in sequence of amino
BIOLOGY BIOCHEMISTRY Unit 2 Part 4 ACTIVITY #6 (Chapter 5) NAME NAME PERIOD PROTEINS GENERAL CHARACTERISTICS AND IMPORTANCES: Polymers of amino acids Each has unique 3-D shape Vary in sequence of amino
Essential Forces in Protein Folding
 Essential Forces in Protein Folding Dr. Mohammad Alsenaidy Department of Pharmaceutics College of Pharmacy King Saud University Office: AA 101 msenaidy@ksu.edu.sa Previously on PHT 426!! Amino Acid sequence
Essential Forces in Protein Folding Dr. Mohammad Alsenaidy Department of Pharmaceutics College of Pharmacy King Saud University Office: AA 101 msenaidy@ksu.edu.sa Previously on PHT 426!! Amino Acid sequence
Water and Aqueous Solutions. 2. Solvation and Hydrophobicity. Solvation
 Water and Aqueous Solutions. Solvation and Hydrophobicity Solvation Solvation describes the intermolecular interactions of a molecule or ion in solution with the surrounding solvent, which for our purposes
Water and Aqueous Solutions. Solvation and Hydrophobicity Solvation Solvation describes the intermolecular interactions of a molecule or ion in solution with the surrounding solvent, which for our purposes
Protein structure forces, and folding
 Harvard-MIT Division of Health Sciences and Technology HST.508: Quantitative Genomics, Fall 2005 Instructors: Leonid Mirny, Robert Berwick, Alvin Kho, Isaac Kohane Protein structure forces, and folding
Harvard-MIT Division of Health Sciences and Technology HST.508: Quantitative Genomics, Fall 2005 Instructors: Leonid Mirny, Robert Berwick, Alvin Kho, Isaac Kohane Protein structure forces, and folding
Introduction to" Protein Structure
 Introduction to" Protein Structure Function, evolution & experimental methods Thomas Blicher, Center for Biological Sequence Analysis Learning Objectives Outline the basic levels of protein structure.
Introduction to" Protein Structure Function, evolution & experimental methods Thomas Blicher, Center for Biological Sequence Analysis Learning Objectives Outline the basic levels of protein structure.
Chapter 1 1) Biological Molecules a) Only a small subset of the known elements are found in living systems i) Most abundant- C, N, O, and H ii) Less
 Biological Molecules a) Only a small subset of the known elements are found in living systems i) Most abundant- C, N, O, and H ii) Less") Chapter 1 1) Biological Molecules a) Only a small subset of the known elements are found in living systems i) Most abundant- C, N, O, and H ii) Less abundant- Ca, P, K, S, Cl, Na, and Mg b) Cells contain
Chapter 1 1) Biological Molecules a) Only a small subset of the known elements are found in living systems i) Most abundant- C, N, O, and H ii) Less abundant- Ca, P, K, S, Cl, Na, and Mg b) Cells contain
Lecture 1. Conformational Analysis in Acyclic Systems
 Lecture 1 Conformational Analysis in Acyclic Systems Learning Outcomes: by the end of this lecture and after answering the associated problems, you will be able to: 1. use Newman and saw-horse projections
Lecture 1 Conformational Analysis in Acyclic Systems Learning Outcomes: by the end of this lecture and after answering the associated problems, you will be able to: 1. use Newman and saw-horse projections
Macromolecule Stability Curves
 Chem728 page 1 Spring 2012 Macromolecule Stability Curves Macromolecule Transitions - We have discussed in class the factors that determine the spontaneity of processes using conformational transitions
Chem728 page 1 Spring 2012 Macromolecule Stability Curves Macromolecule Transitions - We have discussed in class the factors that determine the spontaneity of processes using conformational transitions
Biological Macromolecules
 Introduction for Chem 493 Chemistry of Biological Macromolecules Dr. L. Luyt January 2008 Dr. L. Luyt Chem 493-2008 1 Biological macromolecules are the molecules of life allow for organization serve a
Introduction for Chem 493 Chemistry of Biological Macromolecules Dr. L. Luyt January 2008 Dr. L. Luyt Chem 493-2008 1 Biological macromolecules are the molecules of life allow for organization serve a
CHAPTER 29 HW: AMINO ACIDS + PROTEINS
 CAPTER 29 W: AMI ACIDS + PRTEIS For all problems, consult the table of 20 Amino Acids provided in lecture if an amino acid structure is needed; these will be given on exams. Use natural amino acids (L)
CAPTER 29 W: AMI ACIDS + PRTEIS For all problems, consult the table of 20 Amino Acids provided in lecture if an amino acid structure is needed; these will be given on exams. Use natural amino acids (L)
EXAM 1 Fall 2009 BCHS3304, SECTION # 21734, GENERAL BIOCHEMISTRY I Dr. Glen B Legge
 EXAM 1 Fall 2009 BCHS3304, SECTION # 21734, GENERAL BIOCHEMISTRY I 2009 Dr. Glen B Legge This is a Scantron exam. All answers should be transferred to the Scantron sheet using a #2 pencil. Write and bubble
EXAM 1 Fall 2009 BCHS3304, SECTION # 21734, GENERAL BIOCHEMISTRY I 2009 Dr. Glen B Legge This is a Scantron exam. All answers should be transferred to the Scantron sheet using a #2 pencil. Write and bubble
Many proteins spontaneously refold into native form in vitro with high fidelity and high speed.
 Macromolecular Processes 20. Protein Folding Composed of 50 500 amino acids linked in 1D sequence by the polypeptide backbone The amino acid physical and chemical properties of the 20 amino acids dictate
Macromolecular Processes 20. Protein Folding Composed of 50 500 amino acids linked in 1D sequence by the polypeptide backbone The amino acid physical and chemical properties of the 20 amino acids dictate
Molecular Modelling. part of Bioinformatik von RNA- und Proteinstrukturen. Sonja Prohaska. Leipzig, SS Computational EvoDevo University Leipzig
 part of Bioinformatik von RNA- und Proteinstrukturen Computational EvoDevo University Leipzig Leipzig, SS 2011 Protein Structure levels or organization Primary structure: sequence of amino acids (from
part of Bioinformatik von RNA- und Proteinstrukturen Computational EvoDevo University Leipzig Leipzig, SS 2011 Protein Structure levels or organization Primary structure: sequence of amino acids (from
Carbohydrate- Protein interac;ons are Cri;cal in Life and Death. Other Cells. Hormones. Viruses. Toxins. Cell. Bacteria
 ther Cells Carbohydrate- Protein interac;ons are Cri;cal in Life and Death ormones Viruses Toxins Cell Bacteria ow to Model Protein- ligand interac;ons? Protein Protein Protein DNA/RNA Protein Carbohydrate
ther Cells Carbohydrate- Protein interac;ons are Cri;cal in Life and Death ormones Viruses Toxins Cell Bacteria ow to Model Protein- ligand interac;ons? Protein Protein Protein DNA/RNA Protein Carbohydrate
Phys 102 Lecture 2 Coulomb s Law & Electric Dipoles
 Phys 102 Lecture 2 Coulomb s Law & Electric Dipoles 1 Today we will... Get practice using Coulomb s law & vector addition Learn about electric dipoles Apply these concepts! Molecular interactions Polar
Phys 102 Lecture 2 Coulomb s Law & Electric Dipoles 1 Today we will... Get practice using Coulomb s law & vector addition Learn about electric dipoles Apply these concepts! Molecular interactions Polar
Student Questions and Answers October 8, 2002
 Student Questions and Answers October 8, 2002 Q l. Is the Cα of Proline also chiral? Answer: FK: Yes, there are 4 different residues bound to this C. Only in a strictly planar molecule this would not hold,
Student Questions and Answers October 8, 2002 Q l. Is the Cα of Proline also chiral? Answer: FK: Yes, there are 4 different residues bound to this C. Only in a strictly planar molecule this would not hold,
Protein Structure Bioinformatics Introduction
 1 Swiss Institute of Bioinformatics Protein Structure Bioinformatics Introduction Basel, 27. September 2004 Torsten Schwede Biozentrum - Universität Basel Swiss Institute of Bioinformatics Klingelbergstr
1 Swiss Institute of Bioinformatics Protein Structure Bioinformatics Introduction Basel, 27. September 2004 Torsten Schwede Biozentrum - Universität Basel Swiss Institute of Bioinformatics Klingelbergstr
Protein Structure Basics
 Protein Structure Basics Presented by Alison Fraser, Christine Lee, Pradhuman Jhala, Corban Rivera Importance of Proteins Muscle structure depends on protein-protein interactions Transport across membranes
Protein Structure Basics Presented by Alison Fraser, Christine Lee, Pradhuman Jhala, Corban Rivera Importance of Proteins Muscle structure depends on protein-protein interactions Transport across membranes
Energetics of Protein Folding
 doi:10.1016/j.jmb.2007.05.078 J. Mol. Biol. (2007) 371, 283 301 REVIEW Energetics of Protein Folding Robert L. Baldwin Department of Biochemistry, Beckman Center, Stanford University Medical Center, Stanford,
doi:10.1016/j.jmb.2007.05.078 J. Mol. Biol. (2007) 371, 283 301 REVIEW Energetics of Protein Folding Robert L. Baldwin Department of Biochemistry, Beckman Center, Stanford University Medical Center, Stanford,
CHEM 4170 Problem Set #1
 CHEM 4170 Problem Set #1 0. Work problems 1-7 at the end of Chapter ne and problems 1, 3, 4, 5, 8, 10, 12, 17, 18, 19, 22, 24, and 25 at the end of Chapter Two and problem 1 at the end of Chapter Three
CHEM 4170 Problem Set #1 0. Work problems 1-7 at the end of Chapter ne and problems 1, 3, 4, 5, 8, 10, 12, 17, 18, 19, 22, 24, and 25 at the end of Chapter Two and problem 1 at the end of Chapter Three
Molecular Modeling Lecture 11 side chain modeling rotamers rotamer explorer buried cavities.
 Molecular Modeling 218 Lecture 11 side chain modeling rotamers rotamer explorer buried cavities. Sidechain Rotamers Discrete approximation of the continuous space of backbone angles. Sidechain conformations
Molecular Modeling 218 Lecture 11 side chain modeling rotamers rotamer explorer buried cavities. Sidechain Rotamers Discrete approximation of the continuous space of backbone angles. Sidechain conformations
Using Higher Calculus to Study Biologically Important Molecules Julie C. Mitchell
 Using Higher Calculus to Study Biologically Important Molecules Julie C. Mitchell Mathematics and Biochemistry University of Wisconsin - Madison 0 There Are Many Kinds Of Proteins The word protein comes
Using Higher Calculus to Study Biologically Important Molecules Julie C. Mitchell Mathematics and Biochemistry University of Wisconsin - Madison 0 There Are Many Kinds Of Proteins The word protein comes
1) Here we review the various types of interactions that can take place between and among molecules.
 Here we review the various types of interactions that can take place between and among molecules.") Chem 431A-L02-W'05 page 1 of 6 Chem 431A-L02-W'05 Summary of lecture topics discussed in lecture 2-3: 1) Here we review the various types of interactions that can take place between and among molecules.
Chem 431A-L02-W'05 page 1 of 6 Chem 431A-L02-W'05 Summary of lecture topics discussed in lecture 2-3: 1) Here we review the various types of interactions that can take place between and among molecules.