Multiple steps in the localization of bicoid RNA to the anterior pole of the Drosophila oocyte
|
|
|
- Jonathan Warren Ray
- 5 years ago
- Views:
Transcription

1 Development 1989 Supplement, Printed in Great Britain The Company of Biologists Limited Multiple steps in the localization of bicoid RNA to the anterior pole of the Drosophila oocyte DANIEL ST. JOHNSTON, WOLFGANG DRIEVER, THOMAS BERLETH, SIBYLL RICHSTEIN and CHRISTIANE NOSSLEIN-VOLHARD Max Planck Institut fitr EnlwickJungsbiologie, Abteilung Cenetik, Spemannstrasse 35, 7400 Tubingen, FRG Summary The anterior region of the Drosophila embryonic pattern is determined by a gradient of the bicoid (bed) protein. The correct formation of this gradient requires the localization of bed RNA to the anterior pole of the egg. Here we use a wholemount in situ technique to examine the process of bed RNA localization during oogenesis and embryogenesis. While bed protein becomes distributed in a gradient that extends throughout the anterior two thirds of the early embryo, bed RNA remains restricted to a much smaller region at the anterior pole. The difference between these distributions indicates that the shape of the protein gradient must depend to some extent on the posterior movement of the protein after it has been synthesized. Four distinct phases of bed RNA localization can be distinguished during oogenesis. Between stages 6 and 9 of oogenesis, the RNA accumulates in a ring at the anterior end of the oocyte. During the second phase, in stage 9-10a follicles, the RNA also localizes to the apical regions of the nurse cells, demonstrating that the nurse cells possess an intrinsic polarity. As the nurse cells contract during stages 10b-ll, all of the bed RNA becomes localized to the cortex at the anterior end of the oocyte. During a final phase that must occur between stage 12 of oogenesis and egg deposition, the RNA becomes localized to a spherical region that occupies a slightly dorsal position at the anterior pole. Mutations in the maternal-effect genes, exuperantia (exu) and swallow (sww), lead to an almost uniform distribution of bed RNA in the early embryo, while staufen (stau) mutations produce a gradient of RNA at the anterior pole, exu mutations disrupt the second stage of bed RNA localization during oogenesis, sww mutations disrupt the third, and stau mutations affect the fourth phase. Key words: Drosophila, bicoid, RNA localization, oogenesis, exuperantia, swallow, staufen, gradient formation. Introduction The anterior-posterior pattern of the Drosophila embryo is determined in response to maternal factors that are deposited in the egg during oogenesis. Three groups of maternal genes are required to specify distinct regions of this pattern; the head and thorax (the anterior group), the abdomen (the posterior group) and the acron and telson (the terminal group) (reviewed in Nusslein-Volhard and Roth, 1989; Nusslein-Volhard et al. 1987). The anterior portion of the body plan depends on the product of the bicoid (bed) gene (Frohnhofer and Nusslein-Volhard, 1986). In bicoid~ embryos, the head and thorax are absent, and the abdominal fate map expands anteriorly. In addition, a duplicated telson forms at the anterior end of the embryo. Transplantation experiments have shown that bcd + activity is localized to the anterior pole of the egg, and when transplanted to more posterior positions, can induce the formation of anterior structures at ectopic sites (Frohnh6fer and Nusslein-Volhard, 1986; Frohnhofer et al. 1986). In agreement with the results of the transplantation experiments, bed RNA is localized to the anterior pole of the egg (Frigerio et al. 1986; Berleth et al. 1988). The protein that is translated from this RNA forms a concentration gradient which extends throughout the anterior two thirds of the embryo (Driever and Nusslein-Volhard, 1988fl). This protein gradient appears to determine anterior positional values in a concentrationdependent manner, since changes in the maternal bcd + gene dosage produce complementary shifts in both the protein gradient and the fate map (Driever and Nusslein-Volhard, 19886). Further support for the role of bed protein as the anterior morphogen comes from the results of experiments in which bed RNA synthesized in vitro is injected into early embryos. When the RNA is injected into the middle of a recipient embryo, it can induce the development of ectopic head and thoracic structures (Driever, Siegel and Nusslein-Volhard, in

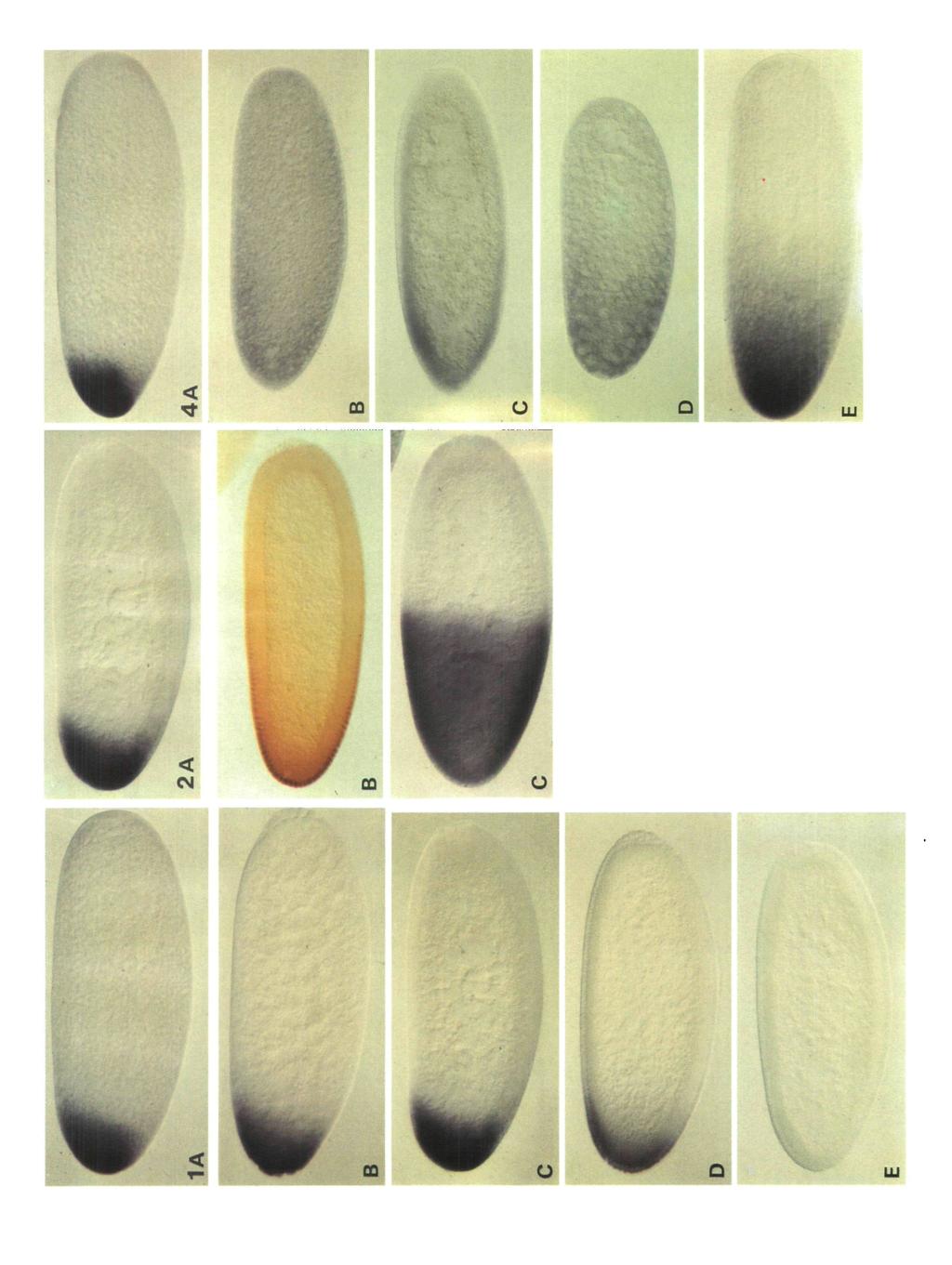
2 14 D. St. Johnston and others preparation). The most anterior pattern elements form closest to the site of injection, with more posterior (thoracic) structures developing on either side. This result indicates not only that the bed protein determines the anterior quality of the structures that develop, but also that the slope of the protein gradient specifies the polarity of the pattern. In addition, the ability of bed RNA to organize anterior pattern in the middle of the embryo shows that no other anteriorly localized molecules are required for this process. The bicoid protein contains a homeodomain, suggesting that bicoid encodes a DNA-binding protein (Frigerio et al. 1986). This makes it attractive to suppose that bed protein specifies anterior positional values by activating or repressing zygotic target genes in a concentration-dependent manner. Genetic and molecular evidence indicates that bed regulates the anterior zygotic expression of the gap gene, hunchback (Lehmann and Nusslein-Volhard, 1987; Schroder et al. 1988; Tautz, 1988). bed protein binds to several sites in the hunchback promoter, and acts as a transcriptional activator of hunchback (Driever and Nusslein-Volhard, 1989). Driever et al. (in press) have made constructs containing synthetic-bed-binding sites fused to the hsp70 promoter and a reporter gene. When transformed into flies, the bed-binding sites drive the anterior expression of the reporter gene in the embryo. The size of this anterior domain of expression is reduced when binding sites with lower affinities for the bed protein are used. The observation that promoters with low-affinity bed-binding sites are only activated at high protein concentrations suggests a model for how the bed protein gradient could activate several zygotic target genes in distinct anterior domains, and thereby determine several levels of anterior development. The formation of the wild-type protein gradient, and thus the determination of a normal anterior pattern, depends upon the localization of bed RNA to the anterior pole. Mutations in the maternal genes exuperantia (exu) and swallow (sww) disrupt this localization during oogenesis, and lead to an almost uniform distribution of bed RNA in the early embryo (Berleth et al. 1988; Stephenson et al. 1988). Both of these mutations result in phenotypes in which anterior structures of the head are deleted, and the thoracic region is expanded (Frohnhofer and Nusslein-Volhard, 1987). Embryos derived from females mutant for the posterior group gene staufen show similar but weaker head defects, in addition to the abdominal deletions characteristic of all mutations in this class (Schiipbach and Wieschaus, 1986; Nusslein-Volhard et al. 1987; Lehmann, 1988). These embryos also show an anterior shift in the fate map, which can be seen in a shift in the position of the cephalic furrow and the first fushi tarazu stripe (Schiipbach and Wieschaus, 1986; Carroll et al. 1986; Lehmann, 1988). The observation that the bed protein gradient is shallower in the mutant embryos suggests that staufen mutations may also alter the distribution of bed RNA (Driever and Nusslein-Volhard, 19886). In this report, we use a non-radioactive, enzymelinked in situ technique developed by Tautz and Pfeifle (1989) to examine the process of bed RNA localization during oogenesis and early embryogenesis. This technique has the advantage that one can perform hybridizations on wholemount preparations. We have used these wholemount stainings to make direct comparisons between the bed RNA distribution and the protein distribution, as revealed by antibody stainings. Results bed RNA localization in wild-type embryos The localization of bed RNA in wild-type embryos has previously been described by Frigerio et al. (1986) and Berleth et al. (1988). We have repeated these investigations in order to gain a clearer understanding of the three-dimensional distribution of bed RNA during the first stages of embryogenesis. In very early (stage 1) embryos, bed RNA staining resembles a flattened ball which is closely apposed to the anterior pole, and which frequently occupies a slightly dorsal position (Fig. 1A). The RNA does not seem to be specifically bound to the cortex of the egg, since much of it is in the interior. As development proceeds through pole cell formation (Fig. IB) to syncytial blastoderm (Fig. 1C), most of the RNA becomes localized to the periphery, in the clear cytoplasm that surrounds each nucleus. This movement to the cortex sometimes results in a slight posteriorwards shift in the RNA distribution. By early nuclear cycle 14, bed RNA begins to disappear (Fig. ID) and midway through cellularization the signal is no longer detectable (Fig. IE). Fig. 1. The distribution of bed RNA in wild-type embryos. The embryos were fixed, and hybridized with a randomprimed probe synthesized from a bicoid cdna clone, following the procedure of Tautz and Pfeifle (1989). In all figures, anterior is to the left and dorsal is uppermost. (A) Early cleavage stage. (B) Late cleavage stage, after pole cell formation. (C) Syncytial blastoderm. (D) Late syncytial blastoderm, after the 13th nuclear division. (E) Early cellularization. These embryos have been overstained in order to clearly show the posterior extent of the bed RNA distribution. This overstating partially obscures the cortical localization of the RNA in syncytial blastoderm embryos (C), which can be clearly seen in understated embryos or in later embryos as the RNA starts to disappear (D). Fig. 2. The distributions of bicoid RNA, bicoid protein and hunchback RNA in syncytial blastoderm embryos. (A) bed RNA (B) bed protein. The embryos were stained with a polyclonal anti-bicoid antibody, as described by Driever and Nusslein-Volhard (1988a). (C) hunchback RNA (Tautz et al. 1987). Fig. 4. The bed RNA distribution in mutant embryos. (A) Wild-type cleavage stage. (B) Cleavage-stage embryo derived from an exu /exu mother, showing a uniform distribution of bed RNA. (C) Syncytial blastoderm embryo derived from an exu PJ /exu PJ mother. By this stage a shallow anterior-posterior gradient of RNA has formed. (D) Cleavage-stage embryo derived from a sww N /sww N mother, showing the early weak RNA gradient. (E) Cleavage-stage embryo derived from a siau D3 /Df(2R) PC4 mother. The RNA is distributed in a steep gradient at the anterior end of the embryo.
3 LXJ HI
 A stage 8 follicle with bed RNA localized to the anterior margins of the oocyte.")
4 6A Fig. 6. The distribution of bed RNA during wild-type oogenesis. (A) Part of an ovariole containing a stage6 follicle with a ring of bed RNA in the presumptive oocyte. The adjacent stage 7 and stage 9 follicles also show bed RNA staining. (B) A stage 8 follicle with bed RNA localized to the anterior margins of the oocyte. A weak signal can also be seen in the nurse cells. (C) A stage 9 follicle showing the apical localization in the 15 nurse cells, in addition to the anterior signal in the oocyte. The stage 8 follicle to the left also shows the anterior ring of RNA. (D) A stage 10a follicle. (E) A stage 10b follicle. The nurse cell localization is disappearing, as the nurse cells contract and the bed RNA accumulates at the anterior pole of the oocyte. (F) A stage 12 follicle. The nurse cells are degenerating, and all bed RNA is now localized to the cortex at the anterior of the oocyte. Fig. 7. The distribution of bed RNA in mutant ovaries. (A) exupj, stage 7-8. The RNA forms a ring at the anterior end of the oocyte, but this ring appears more diffuse. (B) exu, stage 10a. The RNA shows a uniform distribution in the cytoplasm of the nurse cells. (C) sww14', stage 9. The RNA localizes normally in the nurse cells and the oocyte. (D) sww1'4, stage 10b. The cortical bed RNA is no longer localized to the most anterior end of the oocyte but instead extends posteriorly. Much of the RNA entering the oocyte at this stage is not localized to the cortex, and leads to weak staining in the anterior third of the oocyte. (E) staii*', stage 12. (F) staud3, stage 12. The bed RNA localization is completely normal at this stage. We have also examined exuqr and exw ovaries which show the same phenotypes as exufj. C
, and these results are presented graphically in Fig. 3.")
5 Localization of bicoid RNA 15 Fig. 2 compares the distributions of bed RNA and protein at the syncytial blastoderm stage. The levels of RNA and protein were quantified using the procedure described by Driever and Nusslein-Volhard (1988a), and these results are presented graphically in Fig. 3. These measurements clearly demonstrate that the RNA is more tightly localized to the anterior end of the embryo, than is the protein, bed RNA forms a steep gradient at the anterior end of the embryo, in which 90 % of the RNA is restricted to the region anterior to 82% egg length (0% EL is the posterior pole). In contrast, bed protein is distributed in a much shallower gradient, in which only 57% of the protein falls within this anterior region. The protein must therefore move posteriorly after it has been synthesized. One of the functions of bed protein is to activate the anterior zygotic expression of hunchback (SchrSder et al. 1988; Tautz, 1988; Driever and Nusslein-Volhard, 1989). This anterior hunchback domain is shown in Fig. 2C, and illustrates that the bed protein gradient must extend to at least 55 % egg length, a position at which bed RNA is not detectable above background " 400" bed protein bed RNA position (% egg length) Fig. 3. The distribution of bed mrna (solid line) and bed protein (dotted line) in the embryo. Whole wild-type embryos were stained for bed protein, using an immunohistochemical method (Driever and Nusslein- Volhard, 1988a), or for bed mrna, using the enzyme linked in situ detection technique. Video images of wholemount embryos were taken and a background image subtracted. The distribution of the stain intensities along the anterior posterior midline was recorded for five embryos (nuclear cycle 13) and the average values at 30 equidistant positions calculated. Signal linearity of the in situ detection method was tested by performing the enzymatic colour reaction for 3 and a half or 16 minutes, respectively. Values in the anteriormost 15% of the embryos appear to be nonlinear in the 16min reaction and were corrected accordingly. The areas under the RNA and protein curves have been made equal in order to allow a comparison of the two distribution profiles. Anterior is to the left. bed RNA localization in mutant embryos In early (stage 1) embryos derived from exu homozygous mothers, bed RNA is uniformly distributed throughout the egg cytoplasm (Fig. 4B). However, by syncytial blastoderm a shallow anterior-posterior gradient has formed (Fig. 4C). Berleth et al. (1987) have found that bed RNA remains uniformly distributed in eggs laid by mothers which are mutant for both exu and the posterior group gene vasa. This suggests that the late bed RNA gradient is created by the degradation of the RNA in the posterior region of the embryo, due to the activity of the posterior organizing centre (Nusslein-Volhard et al. 1987). Embryos laid by sww homozygotes have a more variable distribution of bed RNA. While some embryos show no localization, most contain a weak anterior-toposterior gradient (Fig. 4D). As is the case for exu mutant embryos, the gradient becomes more pronounced as development proceeds, due to the posterior degradation of the RNA. The variability of the bed RNA distribution in sww mutant embryos is reflected in the variability in the final cuticular phenotype (Frohnhdfer and Nusslein-Volhard, 1986). In embryos derived from staufen mutant mothers, bed RNA forms a gradient in the anterior region of the embryo (Fig. 4E). This phenotype is clearly distinct from that produced by exu or sww mutations, and cannot depend upon the posterior degradation of the RNA, since there is no localized posterior activity in stau mutant embryos (Nusslein-Volhard et al. 1987; Lehmann and Nusslein-Volhard, unpublished results). This partial mislocalization of bed RNA leads to a shallower protein gradient (Driever and Nusslein-Volhard, 1988b). The bed RNA distributions in exu, sww, and stau mutant embryos are compared to the wild-type distribution in Fig. 5. Since these comparisons were performed using syncytial blastoderm embryos, exu and sww both show a shallow anterior-to-posterior RNA gradient. The process of bed RNA localization during wild-type oogenesis bed RNA can first be detected in late previtellogenic follicles (stages 5-6 of King (1970)) accumulating in a single, posteriorly located cell of the germ cell cluster (Fig. 6A). As oogenesis proceeds and the oocyte grows larger than the fifteen nurse cells, it becomes clear that this cell is the oocyte. At these stages, very little RNA can be seen in the nurse cells, and the RNA forms a ring around the anterior margin of the developing oocyte (Fig. 6B). This ring increases in size as the follicle grows. During stages 9-10a, large amounts of bed RNA accumulate in the nurse cells (Fig. 6C,D). Unlike other maternal RNAs, bed RNA is not uniformly distributed within the nurse cell cytoplasm, but is concentrated in a peripheral region adjacent to each nurse cell nucleus. During stages 10b and 11, the localization within the nurse cells gradually disappears, as the nurse cells contract and transfer their cytoplasm to the oocyte (Fig. 6E). Upon entering the oocyte, the bed RNA becomes localized to the cortex of the anterior pole. At

6 16 D. St. Johnston and others the final few hours of oogenesis, stages 13 and 14, the follicle cells that surround the oocyte secrete the chorion (King, 1970). We have been unable to analyze bed RNA localization during these stages because the vitelline membrane and chorion prevent the entry of the probe into the oocyte. However, the distribution of the RNA in stage 12 oocytes is different from that observed in very young eggs. In the oocytes, the RNA is localized to the cortex and is more concentrated ventrally, whereas in the early embryo, the RNA is found in a spherical region that extends into the interior of the egg and which is often located slightly dorsally. These differences suggest that there is a redistribution of bed RNA, either during stages 13 and 14 of oogenesis, or immediately after fertilization position (% egg length) Fig. 5. Distribution of bed mrna in wild-type embryos and in embryos from females mutant for exu, sww and stau. The distribution of bed mrna in early nuclear cycle 13 embryos was visualized and measured as described in the legend to Fig. 3. The RNA distribution profiles of embryos derived from mutant females (solid lines; (A) exu PJ /exu PJ \ (B) SWW N /SWH-"\ (C) stau D3 /Df(2R) PC4) were plotted together with those of wild-type embryos (dotted lines; (A and B) wild-type controls stained in parallel to the mutant embryos; (C) wild-type embryos stained in the same batch as the stau mutant embryos, which were identified by the lack of polecells). this stage, the RNA is no longer restricted to a ring around the anterior pole, and instead covers most of the anterior end of the oocyte. By stage 12, when the nurse cells are degenerating, bed RNA is localized in a cap at the anterior end of the egg, with more of the RNA being found ventrally than dorsally (Fig. 6F). During bed RNA localization in mutant ovaries In order to understand how the mutations that alter the bed RNA distribution in the embryo affect the four phases of RNA localization during oogenesis, we have performed in situ hybridizations on exu, sww and stau mutant ovaries. The earliest differences from wild-type bed RNA localization are observed in exu mutant ovaries. The initial accumulation of bed RNA during stages 5-7 occurs normally. As the oocyte increases in size, the RNA still forms a ring at the anterior end, but this ring often appears more diffuse (Fig. 7A). The first major deviation from normal oogenesis becomes apparent during stage 10a, when bed RNA is not localized to the apical regions of the nurse cells, and instead shows a uniform distribution in the nurse cell cytoplasm (Fig. 7B). When this RNA enters the oocyte, it is not retained at the anterior pole, and from stage 10 onwards no localization within the oocyte is visible. In the ovaries of sww homozygous females, the process of bed RNA localization appears entirely normal up to stage 10a. The ring of RNA forms at the anterior end of the oocyte, and RNA accumulates in the apical regions of the nurse cells (Fig. 7C). During stages 10b and 11, the anterior ring of RNA seems to slip posteriorly and become more diffuse (Fig. 7D). In addition, much of the RNA that enters the oocyte during this time is not localized to the cortex. By stage 12, all bed RNA appears to have been released from the cortex and forms a shallow anterior-to-posterior gradient. In homozygous stau ovaries, the process of bed RNA localization appears completely normal up until stage 12 (Fig. 7E,F), the latest stage that we have examined. Since the RNA is distributed in a gradient in early embryos, it must be released from the anterior pole after stage 12 of oogenesis. Discussion The experiments presented in this report use a wholemount in situ technique to provide a detailed picture of the process of bed RNA localization during oogenesis and embryogenesis. These results confirm and extend the previous analyses of bed RNA localization, which

7 exu sww Localization of bicoid RNA stau 17 stage 13, 14 fertilization egg deposition Stage 8 Stage 9 Stage 10b Stage 12 embryo Fig. 8. A drawing showing the four phases of bed RNA localization during oogenesis and the points at which exu, sww, and stau are required. The drawings are based on King (1972). were performed using radioactive probes on sectioned material (Frigerio et al. 1986; Berleth et al. 1987; Stephenson et al. 1988). The wholemount procedure allows direct comparisons between the bed RNA and protein distributions. As shown in Figs 2 and 3, the two distributions are quite different. Given that no RNA can be detected posterior to 60% egg length even in overstained embryos, whereas the protein gradient extends at least to 30 % EL, the shape of the protein gradient must depend to some extent on the movement of protein molecules towards the posterior after they have been translated. Simple diffusion of the protein can account for this distribution (Driever and Nusslein- Volhard, 1988a). The in situ hybridizations to embryos derived from staufen mutant females reveal that, like exuperantia and swallow, staufen is required for the correct localization of bed RNA to the anterior pole. However, staufen mutations only cause a partial mislocalization of bed RNA. The anterior RNA gradient produced by these mutations results in a shallower bed protein distribution, in which the anterior levels of bed protein are strongly reduced compared to wild-type, and more posterior levels are slightly increased (Driever and Nilsslein-Volhard, 1988/?). The anterior reduction in bed protein concentration accounts for the loss of anterior head structures observed in staufen embryos (Schiipbach and Wieschaus, 1986). In addition to the head defects, staufen mutations produce a typical posterior group phenotype in which the abdomen is deleted and pole cells do not form. The phenotypes produced in double mutant combinations between staufen and other maternal effect mutations strongly suggest that the posterior effects of staufen are due to a failure to transport pole plasm constituents to the posterior pole (R. Lehmann, personal communication). Thus the staufen gene product is implicated in the localization of maternal factors to both the anterior and posterior poles of the egg. Based on the geometry of the follicle, Frohnhofer and Niisslein-Volhard (1987) have proposed a simple model for bed RNA localization, in which RNA entering at the anterior end of the oocyte is trapped and attached to the cytoskeleton by factors that are uniformly distributed in the egg. The present data suggest that the process of localization is likely to be more complex. The in situ hybridizations to wild-type ovaries reveal at least four phases of bed RNA localization. In the first phase, which extends from stage 6 to early stage 9 of oogenesis, bed RNA is found in a ring at the anterior end of the oocyte. During stages 9-10a, RNA is still found in this ring but bed RNA also accumulates to high levels in the apical regions of the nurse cells. Stephenson et al. (1988) have also noticed the nonuniform distribution of bed RNA in the nurse cells of sectioned ovaries. The apical localization of bed RNA within the nurse cells is unusual since all other maternal RNAs that have been examined show a uniform distribution in the nurse cell cytoplasm, and no specialized cytological structures have been observed in these regions (for example Kobayashi et al. 1988; Sprenger et al. 1989). In the third phase, during stages 10b-12, the nurse cell localization disappears and all bed RNA becomes localized to the cortex at the anterior pole of the egg. A final phase of RNA redistribution must occur after stage 12, to produce the spherical pattern of bed staining seen in early embryos. The bed RNA distributions during these four phases are presented schematically in Fig. 8. The second and fourth phases of bed RNA localization cannot be explained by the simple model, since, during the second phase, the RNA is localized in the nurse cells before it enters the egg, and, in the fourth phase, the RNA is redistributed within the egg. Since exu mutations lead to a uniform distribution of bed RNA in the nurse cell cytoplasm as well as in the oocyte, it seems likely that in wild-type ovaries both localizations occur by similar mechanisms. The apical region of a nurse cell can be considered to be the anterior end of the cell, since it lies on the opposite side

8 18 D. St. Johnston and others of the cell from the ring canals which connect to the other nurse cells and the oocyte. Thus bed RNA may be transiently localized to the anterior ends of the nurse cells in a similar fashion to its localization within the oocyte, suggesting that the nurse cells also possess an intrinsic anterior-posterior polarity. The molecules that are required for bed RNA localization in the oocyte are most probably synthesized in the nurse cells. These molecules could therefore mediate the transient RNA localization within the nurse cells, before they themselves are transported into the oocyte. Since bed RNA is synthesized within the nurse cells, this localization cannot depend upon the polar entry of the RNA into one side of the cell. Thus the localization mechanism can function, at least within the nurse cells, in the absence of an asymmetric source of the RNA. If the process of localization within the egg is similar to that in nurse cells, the anterior accumulation of bed RNA within the oocyte may involve a more active mechanism than the simple trapping of the RNA as it enters at the anterior pole. None of the mutations examined in this study completely disrupts all phases of bed RNA localization. The earliest phenotypes are seen in exu mutant ovaries, in which the RNA does not become restricted to the apical regions of the nurse cells during stages 9-10a. exu mutations also seem to have a weak effect on the first phase of bed RNA localization, since the anterior ring often appears more diffuse. Although we have used three different strong exu alleles, which all produce the same phenotype, it is possible that none of these is an amorphic mutation. In an exu null genotype, even the earliest phase of bed RNA localization might be abolished. Berleth et al. (1988) have proposed that the exu gene product binds directly to bed RNA, and mediates its attachment to the cytoskeleton. Since exu mutations abolish the localization in nurse cells, if exu does bind the RNA, it must first do so within the nurse cells. This raises the possibility that bed RNA enters the oocyte as part of a ribonucleoprotein complex that also contains exu protein. If this is the case, one might expect exu protein to colocalize with bed RNA at the anterior pole of the embryo. In sww mutant ovaries, the third phase of bed RNA localization is disrupted. As Berleth et al. (1988) and Stephenson et al. (1988) have previously noted, the RNA is gradually released from the cortex of the oocyte during stages 10b-ll of oogenesis. Since sww mutations also cause defects in nuclear migration and cellularization during embryogenesis, Frohnhofer and Niisslein- Volhard (1987) and Stephenson et al. (1988) have suggested that the swallow protein is a component of the cytoskeleton. In sww mutants, the lack of this protein would cause a destabilization of the cytoskeletal elements that anchor bed RNA to the cortex of the oocyte and lead to a gradual release of the RNA. Our observation that sww mutations have no effect on the apical localization of the RNA in nurse cells provide support for the idea that swallow encodes an oocytespecific component of the cytoskeleton. In staufen homozygotes, the process of bed RNA localization appears completely normal up until stage 12 of oogenesis, yet the RNA is not correctly localized in early embryos. The difference between these distributions indicates that the RNA must be released from the anterior pole sometime between stage 12 and egg deposition. This suggests that the staufen gene product may be required for the movement of bed RNA from the anterior/ventral cortex to the more dorsally located spherical region observed in early embryos, and provides further evidence for a fourth phase of bed RNA localization. The anterior gradient of RNA observed in staufen mutant embryos most probably results from the diffusion of the RNA during a short period of time between its release from the anterior pole and the start of embryogenesis. In general, there is a correlation between the stage at which a mutation affects bed RNA during oogenesis and the distribution of the RNA in embryos, exu mutations show the earliest phenotype (stage 9) and result in a uniform distribution in the embryo, sww mutations disrupt localization during stages and lead to a very shallow embryonic RNA gradient, while staufen alleles seem to cause a release of the RNA after stage 12, producing a much steeper gradient at the anterior pole. This study has identified four phases of bed RNA localization during oogenesis, and has demonstrated that exu, sww, and stau mutations each affect a different stage of this process. At present, we still lack any information on how this localization is achieved at a biochemical or cell biological level. Macdonald and Struhl (1988) have identified a 3' untranslated region of of the bed RNA that is sufficient for localization to the anterior half of the embryo. As the molecular analysis of the trans-acting factors involved in this process advances, we may discover how this region of the bed RNA is recognized, and what components of the cytoarchitecture of the oocyte participate in each phase of the localization of bed RNA to the anterior pole. We thank Maria Leptin, Leslie Stevens, Frank Sprenger and Dominique Ferrandon for their constructive criticisms of the manuscript, and Roswitha Gr6mke-Lutz and Doris Eder for help with the Figures. This work was supported by an EMBO fellowship to D. St. J, and by the Leibniz program of the Deutsche Forschungsgemeinschaft. References BERLETH, T., BURRI, M., THOMA, G., BOPP, D., RICHSTEIN, S.. FRIGERIO, G., NOLL, M. AND NOSSLEIN-VOLHARD, C. (1988). The role of bicoid RNA in organizing the anterior pattern of the Drosophila embryo. EMBO J 7, CARROLL, S. B., WINSLOW, G. M., SCHCPBACH, T. AND SCOTT. M P. (1986). Maternal control of Drosophila segmentation gene expression. Nature. Lond. 323, DRIEVER, W. AND NOSSLEIN-VOLHARD. C. (1988a). A gradient of bicoid protein in Drosophila embryos. Cell 54, DRIEVER, W. AND NOSSLEIN-VOLHARD, C. (1988b). The bicoid protein determines position in the Drosophila embryo in a concentration-dependent manner. Cell 54, DRIEVER, W. AND NOSSLEIN-VOLHARD, C. (1989). The bicoid protein is a positive regulator of hunchback transcription in the early Drosophila embryo. Nature. Lond. 337, FRIGERIO, G., BURRI, M.. BOPP, D., BAUMGARTNER, S. AND NOLL,
. Manipulating the anteroposterior pattern of the Drosophila embryo. J. Embryol. exp. Morph. 97 Supplement 169-179. FROHNHOFER, H.-G. & NOSSLEIN-VOLHARD, C. (1986). Organization of anterior pattern in the Drosophila embryo by the maternal gene bicoid.")
9 Localization of bicoid RNA 19 M. (1986). Structure of the segmentation gene paired and the Drosophila PRD gene set as part of a gene network. Cell 47, FKOHNHOFER, H.-G., LEHMANN, R. AND NOSSLEIN-VOLHARD, C. (1986). Manipulating the anteroposterior pattern of the Drosophila embryo. J. Embryol. exp. Morph. 97 Supplement FROHNHOFER, H.-G. & NOSSLEIN-VOLHARD, C. (1986). Organization of anterior pattern in the Drosophila embryo by the maternal gene bicoid. Nature, Lond. 324, FROHNHOFER, H.-G. AND NOSSLEIN-VOLHARD, C. (1987). Maternal genes required for the anterior localization of bicoid activity in the embryo of Drosophila. Genes and Development 1, KING, R. C. (1970). Ovarian Development in Drosophila melanogaster. New York: Academic Press. KOBAYASHI, S., MIZUNO, H. AND OKADA, M. (1988). Accumulation and spatial distribution of poly(a) + RNA in oocytes and early embryos of Drosophila melanogaster. Develop. Growth & Differ. 30, LEHMANN, R. (1988). Maternal and zygotic control of Drosophila segmentation. Development 104 Supplement, LEHMANN, R. AND NOSSLEIN-VOLHARD, C. (1987). hunchback, a gene required for segmentation of an anterior and posterior region of the Drosophila embryo. Devi Biol. 119, MACDONALD, P. M. AND STRUHL, G. (1988). Cis-acting sequences responsible for anterior localization of bicoid mrna in Drosophila embryos. Nature, Lond. 336, NOSSLEIN-VOLHARD, C, FROHNHOFER, H.-G. AND LEHMANN, R. (1987). Determination of anteroposterior polarity in Drosophila. Science 238, NOSSLEIN-VOLHARD, C. AND ROTH, S. (1989). Axis determination in insect embryos. In Cellular Basis of Morphogenesis (Ciba Foundation Symposium 144), pp Chichester: Wiley. SCHRODER, C, TAUTZ, D., SEIFERT, E. AND JACKLE, H. (1988). Differential regulation of the two transcripts from the Drosophila gap segmentation gene hunchback. EMBO J. 7, SCHOPBACH, T. AND WIESCHAUS, E. (1986). Maternal-effect mutations altering the anterior-posterior pattern of the Drosophila embryo. Roux's Arch devl Biol. 195, SPRENGER, F., STEVENS, L. M. AND NOSSLEIN-VOLHARD, C. (1989). The Drosophila gene torso encodes a putative receptor tyrosine kinase. Nature, Lond. 338, STEPHENSON, E. C, CHAO, Y. AND FACKENTHAL, J. D. (1988). Molecular analysis of the swallow gene of Drosophila melanogaster. Genes and Development 2, TAUTZ, D. (1988). Regulation of the Drosophila segmentation gene hunchback by two maternal morphogenetic centres. Nature, Lond. 332, TAUTZ, D., LEHMANN, R., SCHNORCH, H., SCHUH, R., SEIFERT, E., KIENLIN, A., JONES, K. AND JACKLE, H. (1987). Finger protein of novel structure encoded by hunchback, a second member of the gap class of Drosophila segmentation genes. Nature, Lond. 327, TAUTZ, D. AND PFEIFLE, C. (1989). A non-radioactive in situ hybridization method for the localization of specific RNAs in Drosophila embryos reveals translational control of the segmentation gene hunchback. Chromosoma 98,
10
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION
 MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Autonomous concentration-dependent activation and repression of Krüppel by hunchback in the Drosophila embryo
 Development 120, 3043-3049 (1994) Printed in Great Britain The Company of Biologists Limited 1994 3043 Autonomous concentration-dependent activation and repression of Krüppel by hunchback in the Drosophila
Development 120, 3043-3049 (1994) Printed in Great Britain The Company of Biologists Limited 1994 3043 Autonomous concentration-dependent activation and repression of Krüppel by hunchback in the Drosophila
ULRIKE GAUL 1 * and HERBERT JACKLE 2
 Development 107, 651-662 (1989) Printed in Great Britain The Company of Biologists Limited 1989 651 Analysis of maternal effect mutant combinations elucidates regulation and function of the overlap of
Development 107, 651-662 (1989) Printed in Great Britain The Company of Biologists Limited 1989 651 Analysis of maternal effect mutant combinations elucidates regulation and function of the overlap of
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
MCB 141 Midterm I Feb. 19, 2009
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Determination of the embryonic axes of Drosophila*
 Development Supplement 1, 1991, 1-10 Printed in Great Britain The Company of Biologists Limited 199J Determination of the embryonic axes of Drosophila* CHRISTIANE NUSSLEIN-VOLHARD Max-Planck-lnslitul filr
Development Supplement 1, 1991, 1-10 Printed in Great Britain The Company of Biologists Limited 199J Determination of the embryonic axes of Drosophila* CHRISTIANE NUSSLEIN-VOLHARD Max-Planck-lnslitul filr
Cellular automata for exploring gene regulation in Drosophila segmentation
 Cellular automata for exploring gene regulation in Drosophila segmentation Matthew J. Berryman a, Andrew Allison a, and Derek Abbott a a Centre for Biomedical Engineering and School of Electrical and Electronic
Cellular automata for exploring gene regulation in Drosophila segmentation Matthew J. Berryman a, Andrew Allison a, and Derek Abbott a a Centre for Biomedical Engineering and School of Electrical and Electronic
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Gratuitous mrna localization in the Drosophila oocyte
 Development 121, 3013-3021(1995) Printed in Great Britain The Company of Biologists Limited 1995 3013 Gratuitous mrna localization in the Drosophila oocyte Thomas L. Serano 1,2, and Robert S. Cohen 1,
Development 121, 3013-3021(1995) Printed in Great Britain The Company of Biologists Limited 1995 3013 Gratuitous mrna localization in the Drosophila oocyte Thomas L. Serano 1,2, and Robert S. Cohen 1,
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Gradients and insect segmentation
 Development 104 Supplement, 3-1,6 (1988) Printed in Great Britain @ The Company of Biologists Limited 1988 Gradients and insect segmentation VERNON FRENCH Department of Zoology, University of Edinburgh,
Development 104 Supplement, 3-1,6 (1988) Printed in Great Britain @ The Company of Biologists Limited 1988 Gradients and insect segmentation VERNON FRENCH Department of Zoology, University of Edinburgh,
BIS &003 Answers to Assigned Problems May 23, Week /18.6 How would you distinguish between an enhancer and a promoter?
 Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
MCB 141 Midterm I Feb. 14, 2012
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 14, 2012 Question #1 Question #2 Question #3 Question #4 BONUS / 28 pts / 27 pts / 25 pts / 20 pts / 1 pt TOTAL / 100 pts
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 14, 2012 Question #1 Question #2 Question #3 Question #4 BONUS / 28 pts / 27 pts / 25 pts / 20 pts / 1 pt TOTAL / 100 pts
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Biology 4361 Developmental Biology The Genetics of Axis Specification in Drosophila November 2, 2006
 Biology 4361 Developmental Biology The Genetics of Axis Specification in Drosophila November 2, 2006 EARLY DROSOPHILA DEVELOPMENT Fertilization 1) Drosophila egg activation occurs at ovulation - eggs are
Biology 4361 Developmental Biology The Genetics of Axis Specification in Drosophila November 2, 2006 EARLY DROSOPHILA DEVELOPMENT Fertilization 1) Drosophila egg activation occurs at ovulation - eggs are
Development Team. Developmental Biology Axis Specification in Drosophila. Head, Department of Zoology, University of Delhi
 Paper No. : 11 Module : 6 Development Team Principal Investigator: Prof. Neeta Sehgal Head, Department of Zoology, University of Delhi Paper Coordinator: Prof. Namita Agrawal Department of Zoology, University
Paper No. : 11 Module : 6 Development Team Principal Investigator: Prof. Neeta Sehgal Head, Department of Zoology, University of Delhi Paper Coordinator: Prof. Namita Agrawal Department of Zoology, University
Extranuclear Inheritance
 Extranuclear Inheritance Extranuclear Inheritance The past couple of lectures, we ve been exploring exceptions to Mendel s principles of transmission inheritance. Scientists have observed inheritance patterns
Extranuclear Inheritance Extranuclear Inheritance The past couple of lectures, we ve been exploring exceptions to Mendel s principles of transmission inheritance. Scientists have observed inheritance patterns
Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila
 Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila Jordi Casanova and Gary Struhl Howard Hughes Medical Institute, Center for Neurobiology and
Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila Jordi Casanova and Gary Struhl Howard Hughes Medical Institute, Center for Neurobiology and
Chapter 18 Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Axis determination in flies. Sem 9.3.B.5 Animal Science
 Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Homeotic genes in flies. Sem 9.3.B.6 Animal Science
 Homeotic genes in flies Sem 9.3.B.6 Animal Science So far We have seen that identities of each segment is determined by various regulators of segment polarity genes In arthopods, and in flies, each segment
Homeotic genes in flies Sem 9.3.B.6 Animal Science So far We have seen that identities of each segment is determined by various regulators of segment polarity genes In arthopods, and in flies, each segment
Drosophila germline stem cells
 Development 125, 679-690 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV3776 679 Nanos and Pumilio have critical roles in the development and function of Drosophila germline
Development 125, 679-690 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV3776 679 Nanos and Pumilio have critical roles in the development and function of Drosophila germline
RNA sorting in Drosophila oocytes and embryos
 RNA sorting in Drosophila oocytes and embryos PAUL LASKO 1 Departments of Biology and Anatomy and Cell Biology, McGill University, Montréal, Québec, Canada H3A 1B1 ABSTRACT Many RNAs involved in determination
RNA sorting in Drosophila oocytes and embryos PAUL LASKO 1 Departments of Biology and Anatomy and Cell Biology, McGill University, Montréal, Québec, Canada H3A 1B1 ABSTRACT Many RNAs involved in determination
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology June 16, 2008 Overview What forces affect embryonic development? The embryonic environment: external and internal How do forces
Biology 4361 Developmental Biology Principles of Experimental Embryology June 16, 2008 Overview What forces affect embryonic development? The embryonic environment: external and internal How do forces
Gaining New Insights into Primitive Strategies for Embryonic Axis Specification Using the Wasp Nasonia
 Functional Development and Embryology 2007 Global Science Books Gaining New Insights into Primitive Strategies for Embryonic Axis Specification Using the Wasp Nasonia Eugenia C. Olesnicky Claude Desplan
Functional Development and Embryology 2007 Global Science Books Gaining New Insights into Primitive Strategies for Embryonic Axis Specification Using the Wasp Nasonia Eugenia C. Olesnicky Claude Desplan
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
High Bicoid levels render the terminal system dispensable for Drosophila head development
 Development 127, 3993-3999 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2563 3993 High Bicoid levels render the terminal system dispensable for Drosophila head development
Development 127, 3993-3999 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2563 3993 High Bicoid levels render the terminal system dispensable for Drosophila head development
Role for mrna localization in translational activation but not spatial restriction of nanos RNA
 Development 126, 659-669 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV7673 659 Role for mrna localization in translational activation but not spatial restriction of nanos RNA
Development 126, 659-669 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV7673 659 Role for mrna localization in translational activation but not spatial restriction of nanos RNA
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
Developmental Biology
 Developmental Biology 376 (2013) 99 112 Contents lists available at SciVerse ScienceDirect Developmental Biology journal homepage: www.elsevier.com/locate/developmentalbiology Genomes and Developmental
Developmental Biology 376 (2013) 99 112 Contents lists available at SciVerse ScienceDirect Developmental Biology journal homepage: www.elsevier.com/locate/developmentalbiology Genomes and Developmental
Formation of the bicoid morphogen gradient: an mrna gradient dictates the protein gradient
 RESEARCH ARTICLE 605 Development 136, 605-614 (2009) doi:10.1242/dev.031195 Formation of the bicoid morphogen gradient: an mrna gradient dictates the protein gradient Alexander Spirov 1, Khalid Fahmy 2,
RESEARCH ARTICLE 605 Development 136, 605-614 (2009) doi:10.1242/dev.031195 Formation of the bicoid morphogen gradient: an mrna gradient dictates the protein gradient Alexander Spirov 1, Khalid Fahmy 2,
Temporal regulation of gene expression in the blastoderm Drosophila embryo
 Temporal regulation of gene expression in the blastoderm Drosophila embryo Glenn K. Yasuda, 1,3 Jayne Baker, 2 and Gerold Schubiger 2 1Department of Genetics, SK-50, and 2Department of Zoology, NJ-15,
Temporal regulation of gene expression in the blastoderm Drosophila embryo Glenn K. Yasuda, 1,3 Jayne Baker, 2 and Gerold Schubiger 2 1Department of Genetics, SK-50, and 2Department of Zoology, NJ-15,
Model for the robust establishment of precise proportions in the early Drosophila embryo
 Journal of Theoretical Biology 234 (2005) 13 19 www.elsevier.com/locate/yjtbi Model for the robust establishment of precise proportions in the early Drosophila embryo Tinri Aegerter-Wilmsen a, Christof
Journal of Theoretical Biology 234 (2005) 13 19 www.elsevier.com/locate/yjtbi Model for the robust establishment of precise proportions in the early Drosophila embryo Tinri Aegerter-Wilmsen a, Christof
Thoracic Patterning by the Drosophila Gap Gene hunchback
 Developmental Biology 237, 79 92 (2001) doi:10.1006/dbio.2001.0355, available online at http://www.idealibrary.com on Thoracic Patterning by the Drosophila Gap Gene hunchback Xuelin Wu,* Vikram Vasisht,*
Developmental Biology 237, 79 92 (2001) doi:10.1006/dbio.2001.0355, available online at http://www.idealibrary.com on Thoracic Patterning by the Drosophila Gap Gene hunchback Xuelin Wu,* Vikram Vasisht,*
Seeing Is Believing: The Bicoid Morphogen Gradient Matures
 Cell, Vol. 116, 143 152, January 23, 2004, Copyright 2004 by Cell Press Seeing Is Believing: The Bicoid Morphogen Gradient Matures Review Anne Ephrussi 1, * and Daniel St Johnston 2, * 1 European Molecular
Cell, Vol. 116, 143 152, January 23, 2004, Copyright 2004 by Cell Press Seeing Is Believing: The Bicoid Morphogen Gradient Matures Review Anne Ephrussi 1, * and Daniel St Johnston 2, * 1 European Molecular
Morphogens in biological development: Drosophila example
 LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
Drosophila Somatic Anterior-Posterior Axis (A-P Axis) Formation
 Formation") Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics
Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics
178 Part 3.2 SUMMARY INTRODUCTION
 178 Part 3.2 Chapter # DYNAMIC FILTRATION OF VARIABILITY WITHIN EXPRESSION PATTERNS OF ZYGOTIC SEGMENTATION GENES IN DROSOPHILA Surkova S.Yu. *, Samsonova M.G. St. Petersburg State Polytechnical University,
178 Part 3.2 Chapter # DYNAMIC FILTRATION OF VARIABILITY WITHIN EXPRESSION PATTERNS OF ZYGOTIC SEGMENTATION GENES IN DROSOPHILA Surkova S.Yu. *, Samsonova M.G. St. Petersburg State Polytechnical University,
AP Biology Gene Regulation and Development Review
 AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
Two distinct mechanisms for differential positioning of gene expression borders involving the Drosophila gap protein giant
 Development 125, 3765-3774 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5218 3765 Two distinct mechanisms for differential positioning of gene expression borders involving
Development 125, 3765-3774 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5218 3765 Two distinct mechanisms for differential positioning of gene expression borders involving
SPATIAL PROGRAMMING OF GENE EXPRESSION IN EARLY DROSOPHILA EMBRYOGENESIS
 Ann. Rev. Cell Biol. 1986.2 : 49-80 Copyright 0 1986 by Annual Reviews Inc. All rights reserved SPATIAL PROGRAMMING OF GENE EXPRESSION IN EARLY DROSOPHILA EMBRYOGENESIS Matthew P. Scott Department of Molecular,
Ann. Rev. Cell Biol. 1986.2 : 49-80 Copyright 0 1986 by Annual Reviews Inc. All rights reserved SPATIAL PROGRAMMING OF GENE EXPRESSION IN EARLY DROSOPHILA EMBRYOGENESIS Matthew P. Scott Department of Molecular,
DIFFERENTIATION MORPHOGENESIS GROWTH HOW CAN AN IDENTICAL SET OF GENETIC INSTRUCTIONS PRODUCE DIFFERENT TYPES OF CELLS?
 DIFFERENTIATION HOW CAN AN IDENTICAL SET OF GENETIC INSTRUCTIONS PRODUCE DIFFERENT TYPES OF CELLS? MORPHOGENESIS HOW CAN CELLS FORM ORDERED STRUCTURES? GROWTH HOW DO OUR CELLS KNOW WHEN TO STOP DIVIDING
DIFFERENTIATION HOW CAN AN IDENTICAL SET OF GENETIC INSTRUCTIONS PRODUCE DIFFERENT TYPES OF CELLS? MORPHOGENESIS HOW CAN CELLS FORM ORDERED STRUCTURES? GROWTH HOW DO OUR CELLS KNOW WHEN TO STOP DIVIDING
Chapter 10 Development and Differentiation
 Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Developmental Biology Lecture Outlines
 Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
1(1)pole hole is required maternally for pattern formation in the terminal regions of the embryo
pole hole is required maternally for pattern formation in the terminal regions of the embryo") Development 106, 145-158 (1989) Printed in Great Britain The Company of Biologists Limited 1989 145 1(1)pole hole is required maternally for pattern formation in the terminal regions of the embryo LINDA
Development 106, 145-158 (1989) Printed in Great Britain The Company of Biologists Limited 1989 145 1(1)pole hole is required maternally for pattern formation in the terminal regions of the embryo LINDA
Development of Developmental System System(Mathematical Topics in Biolo. Citation 数理解析研究所講究録 (1993), 827:
, 827:") Title Development of Developmental System System(Mathematical Topics in Biolo Author(s) Takeda, Yasuhiko Citation 数理解析研究所講究録 (1993), 827: 57-74 Issue Date 1993-03 URL http://hdl.handle.net/2433/83294 Right
Title Development of Developmental System System(Mathematical Topics in Biolo Author(s) Takeda, Yasuhiko Citation 数理解析研究所講究録 (1993), 827: 57-74 Issue Date 1993-03 URL http://hdl.handle.net/2433/83294 Right
Manipulating the anteroposterior pattern of the Drosophila embryo
 /. Embryol. exp. Morph. 97 Supplement, 169-179 (1986) 169 Printed in Great Britain The Company of Biologists Limited 1986 Manipulating the anteroposterior pattern of the Drosophila embryo HANS GEORG FROHNHOFER,
/. Embryol. exp. Morph. 97 Supplement, 169-179 (1986) 169 Printed in Great Britain The Company of Biologists Limited 1986 Manipulating the anteroposterior pattern of the Drosophila embryo HANS GEORG FROHNHOFER,
TRANSLATIONAL REGULATION AND RNA LOCALIZATION IN DROSOPHILA OOCYTES AND EMBRYOS
 Annu. Rev. Genet. 2001. 35:365 406 Copyright c 2001 by Annual Reviews. All rights reserved TRANSLATIONAL REGULATION AND RNA LOCALIZATION IN DROSOPHILA OOCYTES AND EMBRYOS Oona Johnstone and Paul Lasko
Annu. Rev. Genet. 2001. 35:365 406 Copyright c 2001 by Annual Reviews. All rights reserved TRANSLATIONAL REGULATION AND RNA LOCALIZATION IN DROSOPHILA OOCYTES AND EMBRYOS Oona Johnstone and Paul Lasko
REVIEW mrna Localisation during Development
 DEVELOPMENTAL BIOLOGY 172, 377 395 (1995) REVIEW mrna Localisation during Development David R. Micklem Wellcome/CRC Institute and Department of Genetics, University of Cambridge, Tennis Court Road, Cambridge
DEVELOPMENTAL BIOLOGY 172, 377 395 (1995) REVIEW mrna Localisation during Development David R. Micklem Wellcome/CRC Institute and Department of Genetics, University of Cambridge, Tennis Court Road, Cambridge
Caenorhabditis elegans
 Caenorhabditis elegans Why C. elegans? Sea urchins have told us much about embryogenesis. They are suited well for study in the lab; however, they do not tell us much about the genetics involved in embryogenesis.
Caenorhabditis elegans Why C. elegans? Sea urchins have told us much about embryogenesis. They are suited well for study in the lab; however, they do not tell us much about the genetics involved in embryogenesis.
BILD7: Problem Set. 2. What did Chargaff discover and why was this important?
 BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
Interactions of the Drosophila gap gene giant with maternal and zygotic pattern-forming genes
 Development 111, 367-378 (1991) Printed in Great Britain The Company of Biologists Limited 1991 367 Interactions of the Drosophila gap gene giant with maternal and zygotic pattern-forming genes ELIZABETH
Development 111, 367-378 (1991) Printed in Great Britain The Company of Biologists Limited 1991 367 Interactions of the Drosophila gap gene giant with maternal and zygotic pattern-forming genes ELIZABETH
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018
 Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
The pattern of cell death in fushi tarazu, a segmentation gene of Drosophila
 Development 104, 447-451 (1988) Printed in Great Britain The Company of Biologists Limited 1988 447 The pattern of cell death in fushi tarazu, a segmentation gene of Drosophila LORENZO MAGRASSI and PETER
Development 104, 447-451 (1988) Printed in Great Britain The Company of Biologists Limited 1988 447 The pattern of cell death in fushi tarazu, a segmentation gene of Drosophila LORENZO MAGRASSI and PETER
Mechanisms of dorsal-ventral axis determination in Drosophila embryos revealed by cytoplasmic transplantations
 Development 117, 1385-1396 (1993) Printed in Great Britain The Company of Biologists Limited 1993 1385 Mechanisms of dorsal-ventral axis determination in Drosophila embryos revealed by cytoplasmic transplantations
Development 117, 1385-1396 (1993) Printed in Great Britain The Company of Biologists Limited 1993 1385 Mechanisms of dorsal-ventral axis determination in Drosophila embryos revealed by cytoplasmic transplantations
Novel modes of localization and function of nanos in the wasp Nasonia
 RESEARCH ARTICLE 3813 Development 137, 3813-3821 (2010) doi:10.1242/dev.054213 2010. Published by The Company of Biologists Ltd Novel modes of localization and function of nanos in the wasp Nasonia Jeremy
RESEARCH ARTICLE 3813 Development 137, 3813-3821 (2010) doi:10.1242/dev.054213 2010. Published by The Company of Biologists Ltd Novel modes of localization and function of nanos in the wasp Nasonia Jeremy
Effects of ectopic expression of caudal during Drosophila development
 Development 109, 271-277 (1990) Printed in Great Britain The Company of Biologists Limited 1990 271 Effects of ectopic expression of caudal during Drosophila development MAREK MLODZIK*, GREG GIBSONt and
Development 109, 271-277 (1990) Printed in Great Britain The Company of Biologists Limited 1990 271 Effects of ectopic expression of caudal during Drosophila development MAREK MLODZIK*, GREG GIBSONt and
Roles of wingless in patterning the larval epidermis of Drosophila
 Development 113, 471-485 (1991) Printed in Great Britain (E) The Company of Biologists Limited 1991 471 Roles of wingless in patterning the larval epidermis of Drosophila AMY BEJSOVEC* and ALFONSO MARTINEZ
Development 113, 471-485 (1991) Printed in Great Britain (E) The Company of Biologists Limited 1991 471 Roles of wingless in patterning the larval epidermis of Drosophila AMY BEJSOVEC* and ALFONSO MARTINEZ
Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos by the polyhomeotic gene
 Development 103, 733-741 (1988) Printed in Great Britain The Company of Biologists Limited 1988 733 Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos
Development 103, 733-741 (1988) Printed in Great Britain The Company of Biologists Limited 1988 733 Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos
AP3162D: Lecture 4 - Basic modelling frameworks for developmental biology and cell-fate decisions
 AP162D: Lecture 4 - Basic modelling frameworks for developmental biology and cell-fate decisions Hyun Youk Delft University of Technology (Dated: March 15, 2018) In this lecture, we will derive the Berg-Purcell
AP162D: Lecture 4 - Basic modelling frameworks for developmental biology and cell-fate decisions Hyun Youk Delft University of Technology (Dated: March 15, 2018) In this lecture, we will derive the Berg-Purcell
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Trans-regulatory functions in the Abdominal-B gene of the bithorax complex
 Development 101, 117-122 (1987) Printed in Great Britain The Company of Biologists Limited 1987 117 Trans-regulatory functions in the Abdominal-B gene of the bithorax complex JORDI CASANOVA 1 and ROBERT
Development 101, 117-122 (1987) Printed in Great Britain The Company of Biologists Limited 1987 117 Trans-regulatory functions in the Abdominal-B gene of the bithorax complex JORDI CASANOVA 1 and ROBERT
Exam 2 ID#: November 9, 2006
 Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Rui Dilão NonLinear Dynamics Group, IST
 1st Conference on Computational Interdisciplinary Sciences (CCIS 2010) 23-27 August 2010, INPE, São José dos Campos, Brasil Modeling, Simulating and Calibrating Genetic Regulatory Networks: An Application
1st Conference on Computational Interdisciplinary Sciences (CCIS 2010) 23-27 August 2010, INPE, São José dos Campos, Brasil Modeling, Simulating and Calibrating Genetic Regulatory Networks: An Application
Modelling the Bicoid gradient
 REVIEW 2253 Development 137, 2253-2264 (2010) doi:10.1242/dev.032409 2010. Published by The Company of Biologists Ltd Modelling the Bicoid gradient Oliver Grimm 1, *,, Mathieu Coppey 2, *, and Eric Wieschaus
REVIEW 2253 Development 137, 2253-2264 (2010) doi:10.1242/dev.032409 2010. Published by The Company of Biologists Ltd Modelling the Bicoid gradient Oliver Grimm 1, *,, Mathieu Coppey 2, *, and Eric Wieschaus
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis
 18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
SUPPLEMENTARY INFORMATION
 med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
An anterior function for the Drosophila posterior determinant Pumilio
 Development 129, 2699-2710 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV5016 2699 An anterior function for the Drosophila posterior determinant Pumilio Chiara Gamberi, David
Development 129, 2699-2710 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV5016 2699 An anterior function for the Drosophila posterior determinant Pumilio Chiara Gamberi, David
NIH Public Access Author Manuscript Curr Opin Genet Dev. Author manuscript; available in PMC 2009 August 1.
 NIH Public Access Author Manuscript Published in final edited form as: Curr Opin Genet Dev. 2008 August ; 18(4): 342 347. doi:10.1016/j.gde.2008.06.002. Dynamics of maternal morphogen gradients in the
NIH Public Access Author Manuscript Published in final edited form as: Curr Opin Genet Dev. 2008 August ; 18(4): 342 347. doi:10.1016/j.gde.2008.06.002. Dynamics of maternal morphogen gradients in the
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Three different fusions led to three basic ideas: 1) If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes
 If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes") Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Meiosis and Sexual Reproduction
 Meiosis and Sexual Reproduction Asexual Reproduction Single parent produces offspring All offspring are genetically identical to one another and to parent Produces identical somatic (body) cells Sexual
Meiosis and Sexual Reproduction Asexual Reproduction Single parent produces offspring All offspring are genetically identical to one another and to parent Produces identical somatic (body) cells Sexual
Genes, Development, and Evolution
 14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
The effect of lethal mutations and deletions within the bithorax complex upon the identity of caudal metameres in the Drosophila embryo
 /. Embryol. exp. Morph. 93, 153-166 (1986) 153 Printed in Great Britain The Company of Biologists Limited 1986 The effect of lethal mutations and deletions within the bithorax complex upon the identity
/. Embryol. exp. Morph. 93, 153-166 (1986) 153 Printed in Great Britain The Company of Biologists Limited 1986 The effect of lethal mutations and deletions within the bithorax complex upon the identity
Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p
 Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p.110-114 Arrangement of information in DNA----- requirements for RNA Common arrangement of protein-coding genes in prokaryotes=
Organization of Genes Differs in Prokaryotic and Eukaryotic DNA Chapter 10 p.110-114 Arrangement of information in DNA----- requirements for RNA Common arrangement of protein-coding genes in prokaryotes=
Development RESEARCH ARTICLE. David Cheung 1, Cecelia Miles 2,3, Martin Kreitman 2 and Jun Ma 1,4, *
 2014. Published by The Company of Biologists Ltd (2014) 141, 124-135 doi:10.1242/dev.098640 RESEARCH ARTICLE Adaptation of the length scale and amplitude of the Bicoid gradient profile to achieve robust
2014. Published by The Company of Biologists Ltd (2014) 141, 124-135 doi:10.1242/dev.098640 RESEARCH ARTICLE Adaptation of the length scale and amplitude of the Bicoid gradient profile to achieve robust
The mago nashi locus encodes an essential product required for germ plasm assembly in Drosophila
 Development 120, 1303-1313 (1994) Printed in Great Britain The Company of Biologists Limited 1994 1303 The mago nashi locus encodes an essential product required for germ plasm assembly in Drosophila Phillip
Development 120, 1303-1313 (1994) Printed in Great Britain The Company of Biologists Limited 1994 1303 The mago nashi locus encodes an essential product required for germ plasm assembly in Drosophila Phillip
PROTEIN SYNTHESIS INTRO
 MR. POMERANTZ Page 1 of 6 Protein synthesis Intro. Use the text book to help properly answer the following questions 1. RNA differs from DNA in that RNA a. is single-stranded. c. contains the nitrogen
MR. POMERANTZ Page 1 of 6 Protein synthesis Intro. Use the text book to help properly answer the following questions 1. RNA differs from DNA in that RNA a. is single-stranded. c. contains the nitrogen
10/03/2014. Eukaryotic Development. + Differentiation vs. Development. Differentiation. Development
 Differentiation vs. Development What comes to mind when you think of differentiation? Eukaryotic Development What about development? Presented by: Sean, Daria, Emily, and Maggie Example: Human Development
Differentiation vs. Development What comes to mind when you think of differentiation? Eukaryotic Development What about development? Presented by: Sean, Daria, Emily, and Maggie Example: Human Development
Evolutionary Developmental Biology
 Evolutionary Developmental Biology a.k.a. EVO-DEVO Paedomorphosis is common among salamanders. Note how this hellbender (top) and mudpuppy (bottom) both have gills, paddle tails, and weaker limbs... Top:
Evolutionary Developmental Biology a.k.a. EVO-DEVO Paedomorphosis is common among salamanders. Note how this hellbender (top) and mudpuppy (bottom) both have gills, paddle tails, and weaker limbs... Top:
!!!!!!!! DB3230 Midterm 2 12/13/2013 Name:
 1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
Chapter 2 Cells and Cell Division
 Chapter 2 Cells and Cell Division MULTIPLE CHOICE 1. The process of meiosis results in: A. the production of four identical cells B. no change in chromosome number from parental cells C. a doubling of
Chapter 2 Cells and Cell Division MULTIPLE CHOICE 1. The process of meiosis results in: A. the production of four identical cells B. no change in chromosome number from parental cells C. a doubling of
Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes
 9 The Nucleus Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes Explain general structures of Nuclear Envelope, Nuclear Lamina, Nuclear Pore Complex Explain movement of proteins
9 The Nucleus Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes Explain general structures of Nuclear Envelope, Nuclear Lamina, Nuclear Pore Complex Explain movement of proteins
Reading Assignments. A. Genes and the Synthesis of Polypeptides. Lecture Series 7 From DNA to Protein: Genotype to Phenotype
 Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
SUPPLEMENTARY INFORMATION
 Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Supplementary Discussion Rationale for using maternal ythdf2 -/- mutants as study subject To study the genetic basis of the embryonic developmental delay that we observed, we crossed fish with different
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
cactus, a maternal gene required for proper formation of the dorsoventral morphogen gradient in Drosophila embryos
 Development 11, 371388 (1991) Printed in Great Britain The Company of Biologists Limited 1991 371 cactus, a maternal gene required for proper formation of the dorsoventral morphogen gradient in Drosophila
Development 11, 371388 (1991) Printed in Great Britain The Company of Biologists Limited 1991 371 cactus, a maternal gene required for proper formation of the dorsoventral morphogen gradient in Drosophila