Determination of the embryonic axes of Drosophila*
|
|
|
- Elfreda Willis
- 5 years ago
- Views:
Transcription

1 Development Supplement 1, 1991, 1-10 Printed in Great Britain The Company of Biologists Limited 199J Determination of the embryonic axes of Drosophila* CHRISTIANE NUSSLEIN-VOLHARD Max-Planck-lnslitul filr Entwicklungsbiologie, Abteilung Genetik, Spemannstrasst 35/111, 7400 Tubingen, Germany * Bateson Memorial Lecture Summary The principles of embryonic pattern formation have been studied extensively in many systems using classical experimental approaches. In Drosophila, a powerful combination of genetics and transplantation experiments, as well as molecular biology, have helped to elucidate the mechanisms that operate during oogenesis and early embryogenesis to establish a set of positional cues required for axis determination in the early embryo. In systematic searches for maternal effect mutations a small number of about 30 genes have been identified that specifically affect the process of determination of the embryonic axes. These 'coordinate' genes define four systems that determine the anteroposterior (AP) axis (three systems) and the dorsoventral (DV) axis (one system) independently. In the anteroposterior axis, the anterior system determines the segmented region of head and thorax, the posterior system determines the segmented abdominal region, and the terminal system is responsible for the formation of the nonsegmented termini at the anterior and posterior egg tips, the acron and telson. In contrast, pattern along the dorsoventral axis is determined by one system only. Although all four systems use different biochemical mechanisms, they share several properties. (1) The product of one gene in each system is localized in a specific region of the freshly laid egg and functions as a spatial signal. (2) In each system, this spatial information finally results in the asymmetrical distribution of one gene product that functions as a transcription factor. (3) This transcription factor is distributed in a concentration gradient that defines the spatial limits of expression of one or more zygotic target genes. The combined action of these three anteroposterior systems as well as the dorsoventral system defines the expression of zygotic target genes in at least seven distinct regions along the anteroposterior and at least three in the dorsoventral axis. These longitudinal and transverse domains provide a coarse spatial prepattern which is then further refined by the action and interaction of zygotic pattern genes. Key words: Drosophila, embryo, embryonic axis, zygotic target genes, zygotic pattern genes. Introduction In the life cycle of higher animals, complex forms alternate with simple ones. An individual begins its life as a zygote (a fertilized egg cell), a morphologically simple structure without recognizable similarity to the body of the adult organism. Development often proceeds through a series of juvenile forms, the structure of which corresponds to the way of life of the respective animals. Thus, in every generation, complex form arises de novo from a much less complex egg cell. How complex is the egg cell really? In what manner and to what extent is the body plan of the living animal contained in the structural organization of the egg cell? How many morphogenetic components are already present in the egg cell, where are they localized, and how does their distribution relate to their function in pattern formation? These fundamental questions have occupied the minds of biologists for a long time. Approaches to an understanding must include experiments in which the informational content of the egg cell is artificially altered. In the ideal experiment, one would wish to remove single morphogenetic components from the system, one by one, without affecting any other parameter of the system. The formation of an aberrant pattern would be the consequence. These kinds of experiments are extremely difficult and, in general, it is not possible experimentally to manipulate a system as complex as the egg cell without inflicting severe, unspecific damage. Extraction, pricking, constriction, irradiation, local destruction - all these methods simultaneously affect all components in the respective region, and the required specificity can be obtained only under extremely favourable circumstances. Nevertheless, experiments of these kinds in insect embryos have shown that determinants localized at the anterior and posterior egg pole are involved in the determination of the anteroposterior axis. Destruction (anterior) or transposition (posterior) of the cytoplasm of these, but not of other regions, had most

2 2 C. Niissleirt-Volhard dramatic consequences on the formation of the embryonic pattern (Kalthoff, 1983; Sander, 1975). These approaches, however, did not lead to the identification and purification of particular morphogenetic components. For the analysis of the pattern-forming processes during embryonic development, we chose the genetical approach. The experimental basis is to eliminate the function of a gene coding for a morphogenetic component by mutation. The resulting phenotype is a specific abnormal pattern formed by the embryo. The kind of deviation from normal development reflects the function of the respective gene. The power of the genetic approach for the analysis of complex metabolic and regulatory pathways in procaryotes as well as eucaryotes has been demonstrated in numerous cases. However, the applicability of genetics is restricted to organisms that can be used in breeding experiments. Among higher organisms, Drosophila is best suited, although initially in chosing it as an object of genetical research, the analysis of embryonic development was not considered at all. For Drosophila, methods have been developed that allow the screening of a very large number of mutational hits, a number that is necessary for the isolation of a representative number of mutations affecting embryonic pattern (Nusslein-Volhard and Wieschaus, 1980; Nusslein-Volhard etal. 1984; Wieschaus and Nusslein-Volhard, 1986). It is possible to identify most, if not all, genes, whose products are specifically involved in a particular process with appropriately large scale mutant screens. The analysis of the phenotypes that result from the lack of function of a single component allows important conclusions to be drawn about the properties of the system and the function of a particular gene. With the modern techniques of molecular biology, it is now possible to clone every gene identified by mutations in order to elucidate the structure of the gene product via the DNA sequence of the gene. In this essay, the analysis of the processes determining the embryonic axes of Drosophila using this genetic approach is described. The identification of coordinate genes The informational content of the egg cell is built during oogenesis. All the substances present in the freshly laid egg are synthesized and deposited in it during oogenesis. Most of these substances serve metabolic functions of the rapidly developing embryo, while only a few components participate in the formation of the embryonic pattern. Several cell types are involved in building the egg. Follicle cells of somatic origin surround a complex of 15 nurse cells and the posteriorly located oocyte. This complex originates from a single germ cell by mitosis and the 16 sister cells are interconnected by cytoplasmic bridges (Fig. 1). The nurse cells are synthetically active during oogenesis and produce RNA species as well as proteins that are transported into the oocyte. A further supply of metabolic substances occurs via uptake from the egg membrane follicle cells pertvitelline space V t eiiine coat I chorion syncytial blastoderm nuclei * pole cells yolk blastoderm cells / Fig. 1. Scheme of oogenesis and early embryogenesis in Drosophila. (A) Follicle at an early stage of oogenesis. The germline-derived nurse cell-oocyte complex is surrounded by a sheet of mesodermally derived follicle cells. (B) Follicle at stage 10. The growing oocyte is located posteriorly to the nurse cells. The follicle cells surrounding the oocyte are thickened. (C) Freshly laid egg. The egg cell is surrounded by two coverings, the vitelline coat and the chorion with its dorsal appendages. Between egg membrane and vitelline coat a space is indicated, the perivitelline space. In the living embryo, this space is only visible at the anterior and posterior tip of the egg, however. (D) Syncytial blastoderm stage embryo (2h after egg deposition). The cleavage nuclei have migrated to the periphery of the egg cell. The pole cells have formed at the posterior pole. In this stage, transcription of the zygotic genome begins. (E) Cellular blastoderm. First cellular stage of the embryo. Monolayer of 6000 uniform cells. hemolymph through the follicle cells. The follicle cells produce the egg coverings, the vitelline coat and chorion. Genetically, all substances in the mature egg are under the control of the maternal genome. Genes that fulfil specific functions during oogenesis and early embryogenesis and that are transcribed during

3 Embryonic axes of Drosophila oogenesis are called, in short, 'maternal genes'. In contrast, those genes that are expressed only after fertilization in the embryo are called 'zygotic genes'. For mutants in maternal genes, the embryonic phenotype is displayed in eggs from genetically mutant females, whereas, in the case of zygotic genes, the genotype of the embryo itself determines its phenotype (Fig. 1). Large-scale mutagenesis experiments for the isolation of maternal effect mutants have been carried out in several laboratories (Gans et al. 1975; Mohler, 1977; Perrimon et al. 1986; Anderson and Nusslein-Volhard, 1986; Schiipbach and Wieschaus, 1986a; Nusslein- Volhard et al. 1987; Schupbach and Wieschaus, 1989). The analysis of phenotypes, genetic complementation tests and mapping experiments resulted in the identification of a small number of genes that have specific functions in embryonic pattern formation, in the establishment of the spatial coordinates of the developing embryo ('coordinate genes': Sander, 1975; Nusslein- Volhard, 1979). Mutations in coordinate genes have certain properties. Homozygous mutant flies are viable if they are derived from heterozygous females. Homozygous mutant females produce eggs of normal egg shell morphology, the embryos developing in these eggs, however, die (independent of their own genotype). The lethal phenotype is not caused by a defect in general functions such as nuclear division, cell formation, cell division and differentiation, but concerns the arrangement and formation of body regions. The spatial organization of the embryo is disturbed. In addition to the strictly maternal coordinate genes, genes with an additional function during the life cycle of the fly are also involved in the establishment of the spatial organisation of the egg. As the lack of function alleles are lethal, their contribution during oogenesis can only be deduced from the existence of viable alleles that only affect the maternal function or the analysis of germ line chimeras (Perrimon et al. 1984; Lehmann and Nusslein-Volhard, 1987). About 30 coordinate genes have been identified so far (Fig. 3). A number of arguments (e.g. frequency of alleles obtained in the various mutagenesis experiments) suggest that these 30 genes represent more than 70% of all coordinate genes. It is not yet possible to estimate the total number of the genes with both maternal and zygotic contribution. The four systems of axis determination Embryos derived from mutants in a coordinate gene generally lack particular body regions while others may be enlarged or duplicated. The number of different phenotypes is smaller than the number of genes, which means that groups of genes show similar or identical phenotypes. This finding suggests that the genes within such a group participate in one pattern-forming system. Each system specifies those regions of the body that are absent in the mutant phenotype of the group. Four pattern-forming systems can be defined by such groups of genes, three of which determine the anteroposterior (AP) axis and one the DV axis. The pattern along the dorsoventral (DV) axis thus is determined independently of that of the AP axis (Nusslein-Volhard et al. 1987; Nusslein-Volhard and Roth, 1989). Within the AP axis, individual regions are determined largely independently. In this manner, the anterior system (A) is responsible for the segmented region of head and thorax, the posterior system (P) determines the segmented abdomen, and a third system, the terminal system (T), determines the nonsegmented acron and telson. In Fig. 2, the phenotypes of the three AP systems are illustrated (Fig. 2B,C,D). The large degree of independence of the systems is indicated by the general additivity of phenotypes; if the function of two of the AP systems is eliminated simultaneously by constructing double mutant embryos, a partial pattern is still formed that reflects the function of the third system (Fig. 2F,G,H). Superposition of the partial patterns of the three possible double mutant combinations results in a fairly complete larval pattern. Only if all three systems are eliminated, does the embryo no longer develop a pattern (Fig. 2E). This means that the three AP systems are necessary and sufficient for the specification of the entire AP axis. There is one exception to the independence that concerns the specification of the terminal nonsegmented regions. The acron depends on both the terminal and anterior systems. If the anterior system is eliminated, the terminal system specifies telson also in the anterior egg region. Whereas each AP system has one principal lack-offunction phenotype (see Fig. 2), the DV system has two: most of the genes display a dorsalisation as the lack-of-function phenotype (Fig. 7B), but null mutations in one gene, in contrast, produce partial ventralisation (Fig. 7C). Before describing the individual systems, their essential common features will be briefly summarized. The components and their relationships are displayed schematically in Fig. 3. Each system starts with the localization of a spatial signal within the egg (Figs 4, 6). In two cases, the signal is represented by an RNA that is localized at the anterior or posterior egg pole respectively. In the two remaining systems, the spatial stimulus emanates from the follicle cells, which produce a spatially restricted signal that is released into the perivitelline space surrounding the egg cell. In each system, the local signal, acting through different mechanisms, finally causes the asymmetrical distribution of a maternal gene product that functions as a transcription factor. This factor is often distributed in the form of a gradient that controls the threshold of expression of one or more zygotic genes along the AP or DV axis (Fig. 6). A superposition of the patterns of expression of the zygotic target genes results in a sequence of at least seven unique domains along the AP axis and at least three along the DV axis (Fig. 4). The anterior system - a morphogenetic gradient The anterior system is the simplest of the four systems;
 Phenotype of the P-System (oskar~ embryo). (D) Phenotype of the T-system (torsolike\" embryo). (E) Phenotype of A\", P~ and T~ (triple mutant bed osk tsl).")
4 C. Niisslein-Volhard Fig. 2. Phenotype of mutant embryos of the 3 AP systems. Cuticle preparations of differentiated embryos. (A) Normal larva. (B) Phenotype of the A-system (bicoid~ embryo). (C) Phenotype of the P-System (oskar~ embryo). (D) Phenotype of the T-system (torsolike" embryo). (E) Phenotype of A", P~ and T~ (triple mutant bed osk tsl). This embryo is still in the vitelline coat. (F) Phenotype of P~ T~ (double mutant osk tsf). (G) Phenotype of A~ T" (double mutant bed is!). (H) Phenotype of A" P~ (double mutant bed osk).
, is indispensable for the determination of all anterior structures, while mutants of the other genes of the system have only partial effects on anterior pattern")
5 Embryonic axes of Drosophila further, it is for the time being the best understood. Only one gene, bicoid (bed), is indispensable for the determination of all anterior structures, while mutants of the other genes of the system have only partial effects on anterior pattern (Frohnhofer and Niisslein-Volhard, 1986, 1987; Schupbach and Wieschaus, 1986a). The products of the bed gene provide both the localized signal and the transcription factor. The bed mrna is synthesized during oogenesis in the nurse cells and deposited at the anterior egg pole (Frigerio et al. 1986; Berleth et al. 1988; St. Johnston et al. 1989) where it functions as the source of a bed protein gradient (Fig. 5). The gradient probably is formed by diffusion away from the local source and dispersed decay (Driever and Nusslein-Volhard, 1988a). The bed protein contains a homeobox (Frigerio et al. 1986; Berleth et al. 1988) and functions as an activator of transcription of zygotic target genes (Driever and Nusslein-Volhard, 1989; Driever et al ; Struhl et al. 1989). One of these target genes is the gap gene hunchback (hb). hb is transcribed at uniform levels above a particular bed protein concentration (Struhl et al. 1989). The threshold concentration that is required for hb transcription is reached at about 50 % egg length (Fig. 5). It is determined by the affinity of the promoter for the bed protein. In experiments involving artificial promoter constructs, it has been demonstrated that lowering the affinity of the promoter leads to an anterior shift of the boundary of the expression domain. Assuming the existence of other target genes whose promoters differ in their affinity for the bed protein, the smooth bed gradient thus leads to a subdivision of the egg into several clearly defined domains (Driever et al. 1989a) (Fig. 6). Candidates for such target genes are btd, ems and otd (Finkelstein and Perrimon, 1990; Dalton et al. 1989; Cohen and Jiirgens, 1990). The localization of the bed RNA at the anterior egg pole during oogenesis is dependent on the activity of at least three coordinate genes of the anterior system. The elimination of the function of exuperantla (exu), swallow (swa) or staufen (stau) results in a spread of the mrna towards more posterior regions and a corresponding change in the embryonic fate map (Frohnhofer and Nusslein-Volhard, 1987; Berleth et al. 1988; Driever and Nusslein-Volhard, 1988 >; St. Johnston et al. 1989). The analysis of the distribution of bed mrna during oogenesis and early stages of embryogenesis by in situ hybridization has revealed several steps in localization. The RNA is already localized in the nurse cells to apical regions. Upon entering the oocyte, it is bound to the cortex at the anterior of the oocyte. In the freshly laid egg, the RNA occupies a more central position at the anterior dorsal tip of the egg (Fig. 5). The exu product is already required at an early stage for the localization in the nurse cells, while the sww product appears to be involved in attaching the bed mrna (perhaps bound to the exu protein) to the cortex of the oocyte. In stau embryos, the bed mrna is distributed in a shallow anterior gradient in the egg, while all the early processes occur normally (St Johnston et al. 1989). The 3' nontranslated end of the bed mrna contains nucleotide sequences that are required for the anterior localization (MacDonald and Struhl, 1988). These sequences may include sites for specific interaction with the proteins involved in RNA localization, bed RNA, when transplanted into the egg cell, is not transported to the anterior pole; rather it remains localized, as other RNA molecules, at the site of transplantation and may function as the source of the protein gradient at the respective location. In such experiments, a dramatic reorganization of the entire spatial pattern of the embryo can be induced. The bed RNA thus has properties of an organizer determining polarity and pattern with long range influence (Frohnh6fer and Nusslein-Volhard, 1986; Driever et al. 1990). The posterior system - double negative control In several ways, the posterior system is similar to the anterior system. Like the anterior cytoplasm, the posterior pole plasm, when transplanted, displays long range effects on pattern formation, and the posterior group of genes is responsible for the formation and activity of this localized source (Lehmann and Niisslein- Volhard, 1986, 1987; Sander and Lehmann, 1988; Lehmann and Frohnhofer, 1989; Lehmann and Nusslein-Volhard, 1991). A closer analysis of the components of the posterior system with embryological, genetic and molecular approaches has revealed, however, that it functions in a strikingly different way from the anterior system (Hiilskamp et al. 1989; Irish et al. 1989; Struhl, 1989; Lehmann and Frohnhofer, 1989). A central component is the product of the gene nanos, (nos) that was identified as a localized activity that can induce abdomen formation in mutant embryos of the posterior system (Lehmann and Nusslein-Volhard, 1991). Recent results indicate that it is the nos mrna that is localized at the posterior pole (Wang and Lehmann, 1991). There is evidence that the nos dependent activity spreads anteriorly to about the middle of the egg. For this spread, the function of the gene pumilio is required (Lehmann and Niisslein- Volhard, 1987). Cytoplasmic transplantation experiments in which posterior pole plasm was injected into embryos of various mutant genotypes at various positions indicated that, in the case of nos, the activity does not determine polarity and pattern in an autonomous and concentration-dependent manner, as was observed with bed; while the nos activity determines abdominal structures, their segmental quality within the abdomen, as well as their polarity, seems to depend on the influence of zygotic target genes expressed in the adjacent regions (Lehmann and Frohnhofer, 1989). In other words, nos is required for abdomen formation, but patterning within the abdomen is determined by the interaction of the products of zygotic target genes of the gap class that depend on all three maternal systems. The nos function is required for the transcriptional activation of the zygotic gap gene knirps. Surprisingly, nos is not a transcription factor. Its function is indirect, and it acts by elimination of a transcriptional repressor of knirps. This repressor is normally present throughout the embryo and is removed from the posterior egg half

6 C. Niisslein-Volhard by the nos function (Fig. 6). If this repressor is absent from the beginning, nos function is no longer required for the specification of the abdomen. The repressor of knirps has been identified as the maternal product of the gap gene hunchback which is homogeneously distributed in the freshly laid egg (Hiilskamp et al, 1989; Irish et al. 1989; Struhl, 1989). As already mentioned, the hunchback gene has a later function as a zygotic target gene of bicoid. The rationale for this dual function and for the rather complex double negative control is not obvious. Perhaps it can be understood only in the context of the origin of the pattern-forming systems during evolution. Most of the posterior group genes (Fig. 3) are required for the localization of the nos product in the posterior pole plasm of the egg cell. In mutants of five of the seven posterior group genes, the pole plasm is not formed and thus the localization of nos mrna and in addition the formation of the pole cells, the germ line precursors, is blocked (Schupbach and Wieschaus, I986a,b; Lehmann and Niisslein-Volhard, 1991). For one of these genes, vasa, it is shown that the protein product but not the mrna is localized in the pole plasm (Lasko and Ashburner, 1988; Hay et al. 1988). Preliminary evidence suggests that also the products of other posterior group genes, in the form of mrna or protein, are localized in the pole plasm (Ephrussi et al. 1991; St Johnston et al. 1991). How these products become localized to the posterior pole after being synthesized in the anterior nurse cells, is still obscure. The terminal system - local activation of a receptor While, in the anterior and posterior system, the origins of polarity reside within the germ line, in both the terminal and the dorsoventral system inductive influences coming from the follicle cells play a leading role. In the case of the terminal system, five maternal coordinate genes have been identified (Fig. 3) that share a common phenotype: deletion of the anteriormost and posterior-most regions of the embryo, acron and telson (Schupbach and Wieschaus, 1986a; Niisslein- Volhard et al. 1987; Klingler et al. 1988). In germ line chimeras and, more recently, in clones induced by mitotic recombination in the follicle cells, it has been shown that one of these genes, torsolike, is active in a subpopulation of follicle cells located at the anterior and posterior tips of the oocyte (Stevens et al. 1990). These experiments suggest that these cells produce a signal (perhaps the torsolike product) that can activate the egg cell at the anterior and posterior egg pole (Fig. 6). The receptor of this local signal is the protein product of the gene torso (Sprenger et al. 1989). Among the genes of the terminal system, torso is exceptional as dominant alleles exist, which produce a phenotype that is complementary to the lack-offunction phenotype. In embryos from such mutant females, the segmented middle region is defective while the termini are enlarged (Klingler et al. 1988). Genetic experiments indicated that, in these dominant mutants, active torso product is also present in the middle region of the egg, which is normally segmented, torso activity inhibits segmentation in this middle region by activating the target genes of the terminal system in the wrong position (Klingler et al. 1988; Strecker et al. 1989). The molecular analysis of the torso gene has helped to understand this striking behavior. It encodes a membrane-bound receptor tyrosine kinase (Sprenger et al. 1989), which is incorporated in the egg cell membrane (oolemma) of the early embryo (Casanova and Struhl, 1990). The present working hypothesis is that torso, in order to fulfill its function, has to be activated by a ligand. This activation normally only occurs at the ends where the ligand is present. Dominant alleles code for a ligand-independent, constitutively active product (Sprenger et al. in preparation). The ligand itself may be the product of the gene torsolike (Stevens et al. 1990). Activation of the torso gene product at the egg poles results in a signal transduction chain that leads to a positive control of transcription of the zygotic target genes huckebein and tailless (Klingler et al. 1988; Weigel et al. 1990). The gene encoding the transcription factor for the terminal system (gene Y) has not yet been identified. We predict that the product of gene Y determines the domains of expression of huckebein and tailless in a concentration-dependent manner, defining two thresholds in a similar fashion to bicoid (Fig. 6). The dorsoventral system - control of the nuclear morphogen concentration Of all the four systems of axis determination, the DV system is the most complex. Polarity in this system, as in the terminal system, results from local induction by the follicle cells. Position along the DV axis, like in the anterior system, is determined by a concentration gradient of a morphogen that functions as a transcription factor. However, the formation of this gradient involves an entirely new mechanism: the spatially controlled, graded uptake of the morphogen into the nuclei of the syncytial blastoderm stage embryo. Eleven of the twelve genes of the DV system, the dorsal group, display a complete dorsalization as the lack-of-function phenotype: only pattern elements that normally derive from the dorsal-most egg region are formed, with ventral and lateral elements lacking (Nusslein-Volhard, 1979) (Fig. 7B). In weak alleles, partially dorsalized embryos are formed which lack only the ventral-most elements. For some of the genes (easter, Toll), alleles have been isolated that have a lateralized or partially ventralized phenotype (Anderson and Nusslein-Volhard, 1984; Anderson et al. 1985a,b- Anderson and Nusslein-Volhard, 1986). Finally, the gene cactus shows partial ventralization as a lack-of-function phenotype; pattern elements normally derived from the dorsal and dorsolateral region are absent in mutant embryos, while ventral and ventrolateral elements are formed along the entire DV axis (Schupbach and Wieschaus, 1989; Roth et al. 1989) (Fig. 7C). Most double mutants of cactus and loss of function alleles of genes of the dorsal group display an apolar, lateralized pattern (Fig. 7D); only the double

7 DORSO-VENTRAL POSTERIOR tmout, exuperantia ttaufen fwallow otkar Btaufen tudor valoit vata trunk MVpotehoU fxvnomrath nudel pipe windbeutel tnake tp&tzle torso UVpoUhole stw pelu tube../-v- Fig. 3. The genes of the 4 systems of axis determination in Drosphila. Above the dotted line: maternal genes, below the dotted line: zygotic target genes of the maternal systems. Yellow: genes encoding the localized signal. In the case of the terminal and dorsoventral system, these have not yet been identified, candidates are the somadependent genes tsl, ndl, pipe and wind. Red: genes encoding a membrane-bound receptor. Blue: genes encoding the maternal transcription factor that is asymmetrically distributed. In the anterior system, the transcription factor is the product of the signal, thus both are encoded by the same gene, bicoid. Green: putative zygotic target genes of the maternal systems that encode transcription factors. Maternal signals Zygotic prepattern Fig. 4. Model for the development of complexity of the pattern during early embryogenesis. The maternally provided prepattern consists of 4 localized components and specifies a zygotic prepattern of at least 7 unique domains along the AP and 4 along the DV axis. Fig. 5. The distribution of signal, transcription factor and response RNA in the case of the anterior system. (A) bed RNA in the syncytial blastoderm stage embryo. In situ hybridization. (B) bed protein distribution in a syncytial blastoderm stage embryo, bicoid protein antibody staining. (C) The expression pattern of hunchback in the early embryo. In situ hybridization.
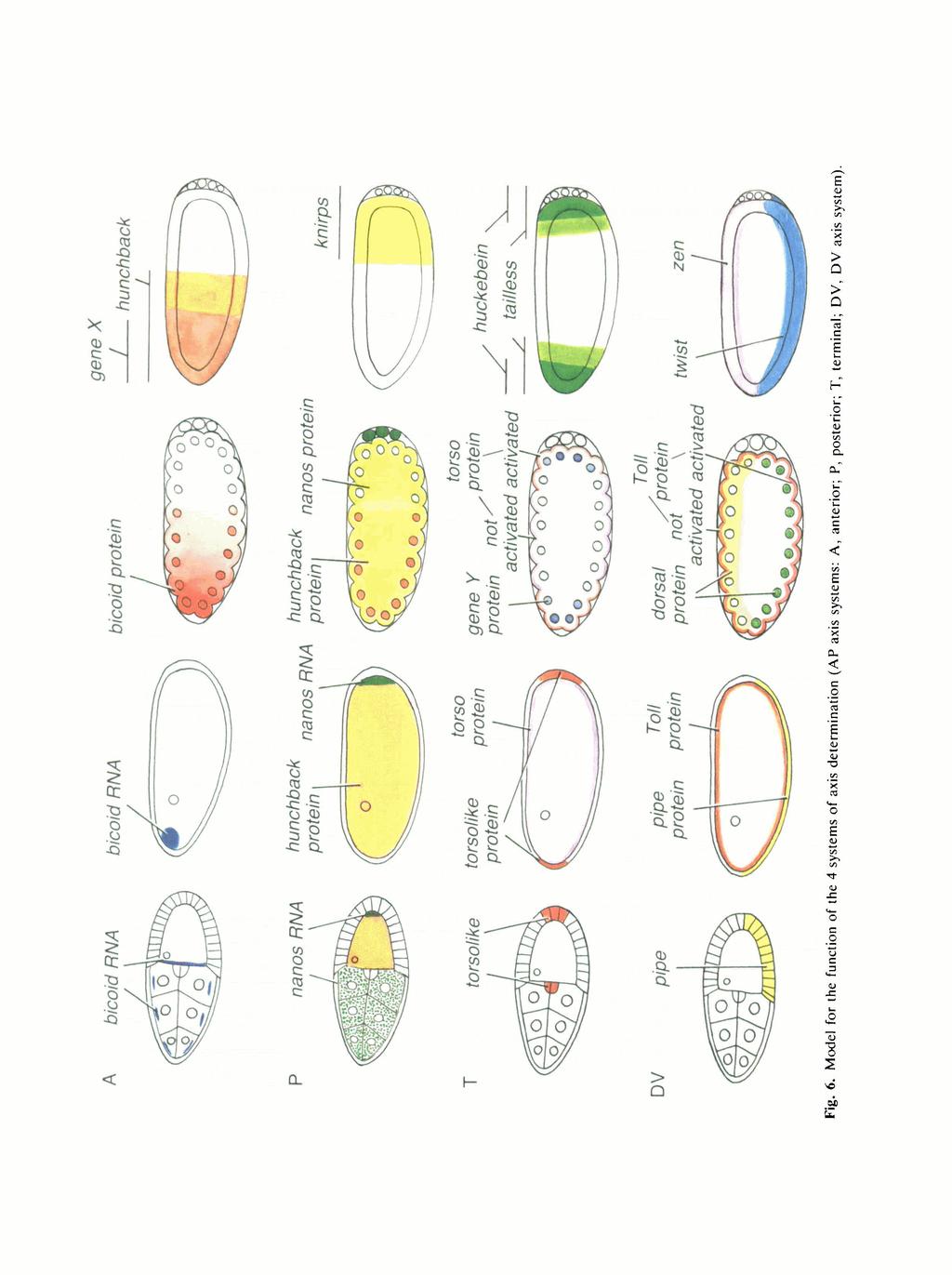
8
 Dorsalized embryo, only dorsal cuticle is differentiated (dorsal~ embryo). (C) Partially ventralized embryo.")
9 Embryonic axes of Drosophila \ ' f ini I * Fig. 7. The phenotypes of the genes of the DV system. (A) Normal larva, anterior left, ventral bottom. (B) Dorsalized embryo, only dorsal cuticle is differentiated (dorsal~ embryo). (C) Partially ventralized embryo. Ventral denticle bands surround the entire embryo while dorsal cuticle is lacking (cactus embryo). (D) Lateralized embryo. Ventrolateral denticle bands surround the embryo while ventral and dorsal cuticle is lacking. During gastrulation, the embryos in B and D are without polarity while the partially ventralized embryo displays a polar gastrulation pattern with mesodermal invagination at the ventral side only. mutant cactus dorsal has a dorsalized phenotype (Roth et al. 1989; Roth et at. 1991). The continuous spectrum of common phenotypes that is observed in mutants of the DV system is best described with a gradient model in which the local concentration of a morphogen determines position along the DV axis (Niisslein-Volhard, 1979). For several reasons, the gene dorsal is the best candidate for the gene coding for this morphogen (Santamaria and Nusslein-Volhard, 1983; Anderson etal. 1985b). Recent molecular findings support this hypothesis (Roth et al. 1989). The mrna of dorsal (Steward et al. 1985) as well as its protein product are homogeneously distributed in the freshly laid egg (Roth et al. 1989; Rushlow et al. 1989; Steward, 1989). As soon as the nuclei of the syncytial embryo reach the periphery of the egg, however, a gradient of nuclear concentration of dorsal protein is observed. In the nuclei of the ventral side of the embryo, the dorsal protein is enriched, while it remains in the cytoplasm at the dorsal side (Roth et al. 1989; Rushlow et al. 1989; Steward, 1989) (Fig. 8A). This gradient therefore is not based, as is the case for bicoid, on a net asymmetry of distribution of the protein, but in contrast, the total dorsal protein concentration is uniform along the DV axis. This fact is most apparent during the mitoses of the cleaving embryo, during which the dorsal protein is released from the nuclei (Roth et al. 1989). The dorsal gene codes for a protein with homology to the oncogene rel (Steward, 1987), and the transcription factor NF*rB (Kieran et al. 1990; Ghosh et al. 1990). It presumably functions as a transcription factor. The domains of expression of the zygotic genes twist and zen, which are normally located in longitudinal stripes along the ventral (twist, Thisse et al. 1988) and dorsal (zen, Rushlow et al. 1987) midline, are dependent on the local nuclear concentration of the dorsal protein (Roth et al. 1989; Rushlow et al. 1989; Steward, 1989). It appears that dorsal can influence the transcription of target genes in both a positive (twist) and a negative (zen, Doyle et al. 1989) fashion. The nuclear uptake of the dorsal protein is controlled by the genes of the dorsal group and by cactus. In mutant embryos of all these genes, the dorsal protein is present in normal amounts (Roth et al. 1989). In dorsalized mutants of the dorsal group, it is not taken up by the nuclei but remains in the cytoplasm even at ventral positions, while in cactus it is taken up by the nuclei also at the dorsal side (Fig. 8B,C). In lateralized embryos, it is equally distributed between cytoplasm and nuclei (Roth et al. 1989). Therefore, cactus functions as an inhibitor of the nuclear uptake of the dorsal protein, and the genes of the dorsal group, in contrast, provide a positive stimulus for the nuclear uptake on the ventral side of the egg. In providing the ventral stimulus, among the dorsal group genes the Toll gene plays a central role (Anderson et al. 1985a,b). Toll, like torso, has gain-offunction alleles with a phenotype complementary to the dorsalized lack-of-function phenotype. Toll encodes a membrane protein that is evenly distributed in the egg membrane (Hashimoto etal. 1988), and therefore may, like torso, function as a receptor. Recent experiments (Stein et al. 1991) have shown that the expression of three of the dorsal group genes (nudel, pipe, windbeutel) is required in the follicle cells. By analogy to torsolike, they may be involved in providing a ligand for the Toll receptor. Two other dorsal group genes, snake and easter, have been shown to encode secreted
 Normal embryo. Gradient of nuclear dorsal protein concentration with maximum in ventral midline10h(bottom). (B) Dorsalized embryo (pelle~). dorsal protein is exclusively in the cytoplasm.")
10 C. Niisslein-Volhard Fig. 8. Distribution of the dorsal protein in normal and mutant embryos in the blastoderm stage. Staining with a polyclonal dorsal protein antibody. loj/m araldite sections. (A) Normal embryo. Gradient of nuclear dorsal protein concentration with maximum in ventral midline10h(bottom). (B) Dorsalized embryo (pelle~). dorsal protein is exclusively in the cytoplasm. (C) Ventralized embryo (77 embryo), dorsal protein is almost exclusively in the nuclei all around the embryo. serine proteases (DeLotto and Spierer, 1986; Chasan and Anderson, 1989). Analogies to the terminal system (Sprenger et al. 1989, and in preparation; Stevens et al. 1989) as well as recent transplantation experiments (Stein et al. 1991) suggest the following model (Fig. 8). A ligand for the Toll receptor is produced by specialized follicle cells facing the ventral side of the oocyte. The ligand is localized in the perivitelline space, perhaps attached to the vitelline coat. During early stages of embryogenesis, it is released by the products of easier and snake and bound by the Toll receptor, thereby activating it. The activation of Toll results, in a manner not yet understood, in the nuclear uptake of the dorsal protein (Fig. 6). Synopsis The origin of the embryonic prepattern Starting from the four localized signals, the four maternal systems of axis determination culminate in the asymmetric distribution of four components, the gene products of the genes bicoid, nanos, gene Y and dorsal. With the exception of nanos, these are morphogens in the classical meaning of the word: they are distributed in gradients and determine positions along the axes in a concentration-dependent manner. The maternal morphogens presumably act as transcription factors that control the spatial domains of transcription of zygotic pattern genes (Fig. 6). The target genes of the morphogens of the AP systems are the gap genes (Niisslein-Volhard and Wieschaus, 1980). Those of the DV systems are a number of zygotic genes that are expressed in longitudinal domains along the DV axis. Target genes of different affinity to the morphogen respond with different threshold concentrations, such that the graded distribution of a morphogen can define a sequence of several regions (Driever et al. 1989a,b). Considering the function of each individual system separately, the bicoid gradient determines at least two zones, nanos one (as mentioned earlier, in this case not in a direct manner but through a double negative control involving the transcription factor hunchback), and the morphogen of the terminal system specifying two symmetrically duplicated regions (Fig. 6). The interaction of the three systems finally results in the formation of a pattern of at least seven domains along the AP axis. In the case of the anterior terminal region, these interactions are combinatorial, here gene Y, together with a high bed concentration, defines acron, while in the absence of bed it defines telson. Further, interactions between the products of the target genes play an important role. The description of these interactions is beyond the scope of this article. They are important for the exact definition of the boundaries that initially are defined in a rather course manner by the maternal transcription factors. In the case of the DV axis, the dorsal gradient alone appears to define at least three, most likely four domains along the DV axis. The dorsal protein functions in this process both as an activator (target: twi) and a repressor (zen). The molecular prepattern of the embryo in the blastoderm stage, that is determined by the four systems of axis determination, is thus already much more complex than the distribution of the four signals present in the freshly laid egg (Fig. 4). The origin of polarity The spatial arrangement of the signals in the egg has its basis in the architecture of the follicle as it arises in the ovarium. In two of the systems, the signal is represented by localized mrna. Synthesis as well as anchoring of this RNA occurs within the germ line-derived oocytenurse cell complex. In contrast, the two other systems, the terminal and the dorsoventral, show striking similarities with inductive systems. For the origin of polarity, a close contact between two different cell types, follicle cells and oocyte, is imperative. The signal is presumably created in a spatially restricted region

11 Embryonic axes of Drosophila within the follicle cell sheet and is received by the activation of receptor molecules in the egg membrane. A special feature of the maternal systems, in this context, is the temporal delay between the production and the function of the signal. The signal is active in the early embryo, at a time when the follicle cells have long disappeared and have left only inviable egg coverings, the chorion and the vitelline coat, behind. Despite this peculiarity, there are several parallels in other systems of cell biology that help to explain the molecular details of these signal transduction mechanisms. The intrinsic polarizing processes that are reflected in the localization of the signals originate during the development of the follicle and its polarity. These processes precede, both temporally and functionally, the processes in which the maternal coordinate genes are involved. The polarization of the nurse cell-oocyte complex, in particular the singling out of one of the 16 daughter cells as the oocyte, determines the orientation of the gradients of the AP axis. It is to be expected that an elaborate process, comparable to the patternforming process of axis determination, is responsible for pattern formation within the follicle cell epithelium. Genes whose phenotypes affect the pattern of the chorion (derived from the follicle cells) as well as that of the embryo are involved in this earlier process (Wieschaus el al. 1978; Schiipbach, 1987; Manseau and Schiipbach, 1989). For a complete understanding of axis determination, the elucidation of these early processes is of prime importance. I would like to thank all my previous and present collaborators, who with their contributions helped to develop the concepts described in this essay. In the isolation of mutants and their phenotypic and genetical analysis, Gerd Jiirgens, Kathryn Anderson, Ruth Lehmann, Hans-Georg FrohnhSfer, Martin Klingler and Siegfried Roth took major parts. The more recent results on the molecular biology in my laboratory were obtained by Wolfgang Driever, Frank Sprenger, Leslie Stevens, Daniel St Johnston, Dave Stein and Thomas Berleth. I would like to thank Daniel St Johnston, Maria Leptin and Phil Ingham for critical comments on the manuscript. References ANDERSON, K. V., BOKLA, L. AND NOSSLEIN-VOLHARD, C. (1985b). Establishment of dorsal-ventral polarity in the Drosophila embryo: the induction of polarity by the Toll gene product. Cell 42, 79J-798. ANDERSON, K. V., JORGENS, G. AND NOSSLEIN-VOLHARD, C. (1985a). Establishment of dorsal-ventral polarity in the Drosophila embryo: genetic studies on the role of the Toll gene product. Cell 42, ANDERSON, K. V. AND NOSSLEIN-VOLHARD, C. (1984). Information for the dorso-ventral pattern of the Drosophila embryo is stored as maternal mrna. Nature 311, ANDERSON, K. V. AND NOSSLEIN-VOLHARD, C. (1986). Dorsalgroup genes in Drosophila. In Gametogenesis and the Early Embryo (ed. J. Gall), Symp. Soc. devl Biol. 43, pp New York: Alan Liss. Inc. BERLETH, T., BURRI, M., THOMA, G., BOPP, D., RICHSTEIN, S., FRIGERIO, G., NOLL, M. AND NOSSLEIN-VOLHARD, C. (1988). The role of localisation of bicoid RNA in organizing the anterior pattern of the Drosophila embryo. EMBO J. 7, CASANOVA, J. AND STRUHL, G. (1990). Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body patterns in Drosophila. Genes Dev. 3, CHASAN, R. AND ANDERSON, K. V. (1989). The role of easier, an apparent serine protease, in organizing the dorsal-ventral pattern of the Drosophila embryo. Cell 56, COHEN, S. M. AND JORGENS, G. (1990). Mediation of Drosophila head development by gap like segmentation genes. Nature 346, DALTON, D., CHADWICK, R. AND MCGINNIS, W. (1989). Expression and embryonic function of empty spiracles: a Drosophila homoeobox gene with two patterning functions on the anterior posterior axis of the embryo. Genes Dev. 3, DELOTTO, R. AND SPIERER, P. (1986). A gene required for the specification of dorsal-ventral pattern in Drosophila appears to encode a serine protease. Nature 323, DOYLE, H., KRAUT, R. AND LEVINE, M. (1989). Spatial regulation of zerkniillt: a dorsal-ventral patterning gene in Drosphda. Genes Dev. 3, DRIEVER, W., MA, J., NOSSLEIN-VOLHARD, C. AND PTASHNE, M. (1989ft). Rescue of bicoid mutant Drosophila embryos by bicoid fusion proteins containing heterologous activating sequences. Nature 342, DRIEVER, W. AND NOSSLEIN-VOLHARD, C. (198&a). A gradient of bicoid protein in the Drosophila embryo. Cell 54, DRIEVER, W. AND NOSSLEIN-VOLHARD, C. (1988ft). The bicoid protein gradient determines position in the Drosophila embryo in a concentration dependent manner. Cell 54, DRIEVER, W. AND NOSSLEIN-VOLHARD, C. (1989). The bicoid protein is a positive regulator of hunchback transcription in the early Drosophila embryo. Nature 337, DRIEVER, W., SIEGEL, V. AND NOSSLEIN-VOLHARD, C. (1990). Autonomous determination of anterior structures in the early Drosophila embryo by the maternal bicoid morphogen. Development 109, DRIEVER, W., THOMA, G. AND NOSSLEIN-VOLHARD, C. (1989a). Determination of spatial domains of zygotic gene expression in the Drosophila embryo by the affinity of binding sites for the bicoid morphogen. Nature 340, EPHRUSSI, A., DICKINSON, L. K. AND LEHMANN, R. (1991). Oskar organizes the germ plasm and directs localization of the posterior determinant nanos. Submitted. FINKELSTEIN, R. AND PERRIMON, N. (1990). The orthodenticle gene is regulated by bicoid and torso and specifies Drosophila head development. Nature 346, FRIGERIO, G., BURRI, M., BOPP, D., BAUMGARTNER, S. AND NOLL, M. (1987). Structure of the segmentation gene paired and the Drosophila PRD gene set as a part of a gene network. Cell 47, FROHNHOFER, H. G. AND NOSSLEIN-VOLHARD, C. (1986). Organization of anterior pattern in the Drosophila embryo by the maternal gene bicoid. Nature 324, FROHNHOFER, H. G. AND NOSSLEIN-VOLHARD, C. (1987). Maternal genes required for the anterior localization of bicoid activity in the embryo of Drosophila. Genes Dev. 1, GANS, M., AUDIT, C. AND MASSON, M. (1975). Isolation and characterization of sex-linked female sterile mutants in Drosophila melanogaster. Genetics 81, GHOSH, S., GIFFORD, A. M., RIVIERE, L. R., TEMPST, P., NOLAN, G. P. AND BALTIMORE, D. (1990). Cloning of the p50 DNA binding subunit of NFicB: homology to rel and dorsal. Cell 62, HASHIMOTO, C, HUDSON, K. L. AND ANDERSON, K. V. (1988). The Toll gene of Drosophila, required for dorsal-ventral embryonic polarity, appears to encode a transmembrane protein. Celt 52, HAY, B., JAN, L. Y. AND JAN, Y. N. (1988). A protein component of Drosophila polar granules is encoded by vasa and has extensive sequence similarity to ATP-dependent helicases. Cell 55, HOLSKAMP, M., SCHRODER, C, PFEIFLE, C, JACKLE, H. AND TAUTZ, D. (1989). Posterior segmentation of the Drosophila embryo in the absence of a maternal posterior organizer gene. Nature 338,

12 10 C. Niisslein-Volhard IRISH, V.. LEHMANN. R. AND AKAM, M. (1989). The Drosophila posterior-group gene nanos functions by repressing hunchback activity. Nature 338, KALTHOFF, K. (1983). Cytoplasmic determinants in dipteran eggs. In Time, Space, and Pattern in Embryonic Development, pp New York: Alan R. Liss Inc. KIERAN, M., BLANK, V., LOCEAT, F., VANDEKERCKHOVE. J.. LOTTSPEICH, F.. LEBAIL. O., URBAN. M. B.. KOURILSKY. P.. BAEUERLE. P. A. AND ISRAEL. A. (1990). The DNA binding subunit of NFicB is identical to factor KBF1 and homologous to the ret oncogene product. Cell KLINGLER. M., ERDELYI, M.. SZABAD, J. AND NOSSLEIN-VOLHARD. C. (1988). Function of torso in determining the terminal anlagen of the Drosophila embryo. Nature LASKO, P. F AND ASHBURNER, M. (1988). The product of the Drosophila gene vasa is very similar to eukaryotic initiation factor-4a. Nature 335, LEHMANN, R AND FROHNHOFER. H. G. (1989). Segment polarity and identity in the abdomen of Drosophila is controlled by the relative positioning of gap gene expression. Development, 107 Supplement LEHMANN. R. AND NOSSLEIN-VOLHARD, C. (1986). Abdominal segmentation, pole cell formation, and embryonic polarity require the localized activity of oskar, a maternal gene in Drosophila. Cell LEHMANN, R. AND NOSSLEIN-VOLHARD. C. (1987). Involvement of the pumilio gene in the transport of an abdominal signal in the Drosophila embryo. Nature 329, LEHMANN. R. AND NOSSLEIN-VOLHARD, C. (1991). The maternal gene nanos has a central role in posterior pattern formation in the Drosophila embryo. Development (in press). MACDONALD, P. M. AND STRUHL. G (1988). Cis-acting sequences responsible for the anterior localization of bicoid mrna in Drosophila embryos. Nature 336, MANSEAU, L AND SCHOPBACH, T. (1989). cappuccino and spire' two unique maternal-effect loci required for both the anteroposterior and dorsovental patterns of the Drosophila embryo. Genes Dev. 3, MOHLER, J. D. (1977) Developmental genetics of the Drosophila egg. Genetics 85, NOSSLEIN-VOLHARD. C. (1979). Maternal effect mutations that alter the spatial coordinates of the embryo of Drosophila melanogaster. In Determinants of Spatial Organisation (ed. I. Konigsberg and S. Subtelney). pp New York and London: Academic Press. NOSSLEIN-VOLHARD, C FROHNHOFER. H. G. AND LEHMANN. R. (1987) Determination of antero-postenor polarity in Drosophila. Science NOSSLEIN-VOLHARD. C. AND ROTH, S. (1989). Axis determination in insect embryos. In Cellular Basis of Morphogenesis. CIBA Foundation Symposium 144, pp New York: John Wiley & Sons. NOSSLEIN-VOLHARD, C. AND WIESCHAUS. E (1980) Mutations affecting segment number and polarity. Nature NOSSLEIN-VOLHARD. C. WIESCHAUS. E. AND KLUDING. H. (1984) Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster I: Zygotic loci on the second chromosome. Wiihelm Rotix's Arch devl Biol. 193, PERRIMON, N., ENCSTROM. L. AND MAHOWALD, A. P. (1984). The effects of zygotic lethal mutations on female germ-line functions in Drosophila. Devi Biol PERRIMON, N., MOHLER, D., ENGSTROM, L. AND MAHOWALD. A. P. (1986). X-linked female sterile loci in Drosophila melanogaster. Genetics 113, ROTH. S, HIROMI. Y.. GODT. D. AND NOSSLEIN-VOLHARD, C. (1991). Cactus, a maternal gene required for proper formation of the dorsoventral morphogen gradient in Drosophila embryos. Submitted. ROTH, S., STEIN, D. AND NOSSLEIN-VOLHARD, C. (1989) A gradient of nuclear localization of the dorsal protein determines dorsoventral pattern in the Drosophila embryo. Cell 59, RUSHLOW. C. A.. FRASCH. M.. DOYLE, H. AND LEVINE, M. (1987). Maternal regulations of zerknidlf. a homoeobox gene controlling differentiation of dorsal tissues in Drosophila. Nature 330, RUSHLOW. C. A.. HAN. K.. MANLEY. J. L. AND LEVINE, M. (1989). The graded distribution of the dorsal morphogen is initiated by selective nuclear transport in Drosophila. Cell SANDER. K. (1975). Pattern specification in the insect embryo. In Cell Patterning. Ciba Foundation Symposium 29. pp Elsevier, Amsterdam. SANDER, K. AND LEHMANN. R. (1988). Drosophila nurse cells produce a posterior signal required for embryonic segmentation and polarity. Nature SANTAMARIA. P. AND NOSSLEIN-VOLHARD. C. (1983). Partial rescue of dorsal, a maternal effect mutation affecting the dorso-ventral pattern of the Drosophila embryo, by the injection of wild-type cytoplasm. EMBO J. 2, SCHOPBACH. T. (1987). Germ line and soma cooperate during oogenesis to establish the dorsoventral pattern of egg shell and embryo in Drosophila melanogaster. Cell SCHOPBACH. T. AND WIESCHAUS, E. (1986O). Maternal-effect mutations altering the anterior-posterior pattern of the Drosophila embryo. ROILX'S Arch, devl Biol. 195, SCHOPBACH. T. AND WIESCHAUS. E. (1986ft). Germline autonomy of maternal-effect mutations altering the embryonic body pattern of Drosophila. Devl Biol. 113, SCHOPBACH, T. AND WIESCHAUS. E. (1989) Female sterile mutations on the second chromosome of Drosophila melanogaster. I. Maternal effect mutations. Genetics SPRENGER, F., STEVENS. L. M. AND NOSSLEIN-VOLHARD. C. (1989) The Drosophila gene torso encodes a putative receptor tyrosine kinase. Nature 338, 478^*83. STEIN. D., ROTH, S.. VOGELSANG. E. AND NOSSLEIN-VOLHARD, C. (1991). The polarity of the dorsoventral axis in the Drosophila embryo is defined by an external signal Cell (in press). STEVENS. L. M.. FROHNHOFER. H. G.. KLINGLER. M. AND NOSSLEIN-VOLHARD, C. (1990) Localized requirement for torsohke expression in follicle cells for the development of terminal anlagen of the Drosophila embryo. Nature STEWARD. R. (1987). Dorsal, an embryonic polarity gene in Drosophila, is homologous to the vertebrate proto-oncogene, c-rel. Science STEWARD. R. (1989). Relocalization of the dorsal protein from the cytoplasm to the nucleus correlates with its function. Cell STEWARD. R., AMBROSION, L. AND SCHEDL. P (1985). Expression of the dorsal gene. Cold Spring Harb. Svmp. quant. Biol ST JOHNSTON. D, BEUCHLE. D. AND NOSSLEIN-VOLHARD. C (1991). Staufen. a gene required to localize maternal RNAs in the Drosophila egg. Submitted. ST JOHNSTON. D.. DRIEVER, W.. BERLETH. T.. RICHSTEIN. S. AND NOSSLEIN-VOLHARD. C. (1989). Multiple steps in the localization of bicoid RNA to the anterior pole of the Drosophila oocyte Development 107 Supplement ST JOHNSTON. D. AND GELBART. W. M. (1987). Decapentaplegic transcripts are localized along the dorsal-ventral axis of the Drosophila embryo. EMBO J STRECKER. T. R.. HALSELL. S R. FISHER. W. W. AND LIPSHITZ. (1989). Reciprocal effects of hyper- and hypoactivity mutations in the Drosophila pattern gene torso. Science STRUHL. G. (1989). Differing strategies for organizing anterior and posterior body pattern in Drosophila embryos. Nature STRUHL. G., STRUHL. K. AND MACDONALD, P M. (1989). The gradient morphogen bicoid is a concentration-dependent transcnptional activator. Cell ST THISSE. B.. STOETZEL. C GOROSTIZA-THISSE. C. AND PERRIN- SCHMITT, F. (1988). Sequence of the rwist gene and nuclear localization of its protein in endomesodermal cells of early Drosophila embryos EMBO J WANG. C. AND LEHMANN. R. (1991). Nanos acts as the posterior determinant in Drosophila. Submitted. WEIGEL. D.. JORGENS. G.. KLINGLER. M. AND JACKLE. H. (1990). Two gap genes mediate maternal terminal pattern information in Drosophila. Science 248, S. WIESCHAUS. E.. MARSH. J. L. AND GEHRING. W (1978). fs(l)k10. a germline-dependent female sterile mutation causing abnormal chorion morphology in Drosophila melanogaster. Wiihelm Roux's Arch, devl Biol WIESCHAUS. E. AND NOSSLEIN-VOLHARD. C. (1986). Looking at Embryos. In Drosophila, a Practical Approach (ed. D. B. Roberts), pp Oxford' IRL Press.
Multiple steps in the localization of bicoid RNA to the anterior pole of the Drosophila oocyte
 Development 1989 Supplement, 13-19 Printed in Great Britain The Company of Biologists Limited 1989 13 Multiple steps in the localization of bicoid RNA to the anterior pole of the Drosophila oocyte DANIEL
Development 1989 Supplement, 13-19 Printed in Great Britain The Company of Biologists Limited 1989 13 Multiple steps in the localization of bicoid RNA to the anterior pole of the Drosophila oocyte DANIEL
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION
 MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
MOLECULAR CONTROL OF EMBRYONIC PATTERN FORMATION Drosophila is the best understood of all developmental systems, especially at the genetic level, and although it is an invertebrate it has had an enormous
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Mechanisms of dorsal-ventral axis determination in Drosophila embryos revealed by cytoplasmic transplantations
 Development 117, 1385-1396 (1993) Printed in Great Britain The Company of Biologists Limited 1993 1385 Mechanisms of dorsal-ventral axis determination in Drosophila embryos revealed by cytoplasmic transplantations
Development 117, 1385-1396 (1993) Printed in Great Britain The Company of Biologists Limited 1993 1385 Mechanisms of dorsal-ventral axis determination in Drosophila embryos revealed by cytoplasmic transplantations
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
MCB 141 Midterm I Feb. 19, 2009
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 19, 2009 Circle the name of your TA Jessica Lyons Alberto Stolfi Question #1 Question #2 Question #3 Question #4 TOTAL
Autonomous concentration-dependent activation and repression of Krüppel by hunchback in the Drosophila embryo
 Development 120, 3043-3049 (1994) Printed in Great Britain The Company of Biologists Limited 1994 3043 Autonomous concentration-dependent activation and repression of Krüppel by hunchback in the Drosophila
Development 120, 3043-3049 (1994) Printed in Great Britain The Company of Biologists Limited 1994 3043 Autonomous concentration-dependent activation and repression of Krüppel by hunchback in the Drosophila
Development Team. Developmental Biology Axis Specification in Drosophila. Head, Department of Zoology, University of Delhi
 Paper No. : 11 Module : 6 Development Team Principal Investigator: Prof. Neeta Sehgal Head, Department of Zoology, University of Delhi Paper Coordinator: Prof. Namita Agrawal Department of Zoology, University
Paper No. : 11 Module : 6 Development Team Principal Investigator: Prof. Neeta Sehgal Head, Department of Zoology, University of Delhi Paper Coordinator: Prof. Namita Agrawal Department of Zoology, University
ULRIKE GAUL 1 * and HERBERT JACKLE 2
 Development 107, 651-662 (1989) Printed in Great Britain The Company of Biologists Limited 1989 651 Analysis of maternal effect mutant combinations elucidates regulation and function of the overlap of
Development 107, 651-662 (1989) Printed in Great Britain The Company of Biologists Limited 1989 651 Analysis of maternal effect mutant combinations elucidates regulation and function of the overlap of
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Lecture 7. Development of the Fruit Fly Drosophila
 BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
BIOLOGY 205/SECTION 7 DEVELOPMENT- LILJEGREN Lecture 7 Development of the Fruit Fly Drosophila 1. The fruit fly- a highly successful, specialized organism a. Quick life cycle includes three larval stages
Cellular automata for exploring gene regulation in Drosophila segmentation
 Cellular automata for exploring gene regulation in Drosophila segmentation Matthew J. Berryman a, Andrew Allison a, and Derek Abbott a a Centre for Biomedical Engineering and School of Electrical and Electronic
Cellular automata for exploring gene regulation in Drosophila segmentation Matthew J. Berryman a, Andrew Allison a, and Derek Abbott a a Centre for Biomedical Engineering and School of Electrical and Electronic
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Biology 4361 Developmental Biology The Genetics of Axis Specification in Drosophila November 2, 2006
 Biology 4361 Developmental Biology The Genetics of Axis Specification in Drosophila November 2, 2006 EARLY DROSOPHILA DEVELOPMENT Fertilization 1) Drosophila egg activation occurs at ovulation - eggs are
Biology 4361 Developmental Biology The Genetics of Axis Specification in Drosophila November 2, 2006 EARLY DROSOPHILA DEVELOPMENT Fertilization 1) Drosophila egg activation occurs at ovulation - eggs are
Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila
 Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila Jordi Casanova and Gary Struhl Howard Hughes Medical Institute, Center for Neurobiology and
Localized surface activity of torso, a receptor tyrosine kinase, specifies terminal body pattern m Drosophila Jordi Casanova and Gary Struhl Howard Hughes Medical Institute, Center for Neurobiology and
Morphogens in biological development: Drosophila example
 LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
High Bicoid levels render the terminal system dispensable for Drosophila head development
 Development 127, 3993-3999 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2563 3993 High Bicoid levels render the terminal system dispensable for Drosophila head development
Development 127, 3993-3999 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV2563 3993 High Bicoid levels render the terminal system dispensable for Drosophila head development
Gradients and insect segmentation
 Development 104 Supplement, 3-1,6 (1988) Printed in Great Britain @ The Company of Biologists Limited 1988 Gradients and insect segmentation VERNON FRENCH Department of Zoology, University of Edinburgh,
Development 104 Supplement, 3-1,6 (1988) Printed in Great Britain @ The Company of Biologists Limited 1988 Gradients and insect segmentation VERNON FRENCH Department of Zoology, University of Edinburgh,
MCB 141 Midterm I Feb. 14, 2012
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 14, 2012 Question #1 Question #2 Question #3 Question #4 BONUS / 28 pts / 27 pts / 25 pts / 20 pts / 1 pt TOTAL / 100 pts
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 14, 2012 Question #1 Question #2 Question #3 Question #4 BONUS / 28 pts / 27 pts / 25 pts / 20 pts / 1 pt TOTAL / 100 pts
Gratuitous mrna localization in the Drosophila oocyte
 Development 121, 3013-3021(1995) Printed in Great Britain The Company of Biologists Limited 1995 3013 Gratuitous mrna localization in the Drosophila oocyte Thomas L. Serano 1,2, and Robert S. Cohen 1,
Development 121, 3013-3021(1995) Printed in Great Britain The Company of Biologists Limited 1995 3013 Gratuitous mrna localization in the Drosophila oocyte Thomas L. Serano 1,2, and Robert S. Cohen 1,
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
Chapter 18 Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Chapter 18 Regulation of Gene Expression Differential gene expression Every somatic cell in an individual organism contains the same genetic information and replicated from the same original fertilized
Developmental Biology Lecture Outlines
 Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Localized Requirements for windbeutel and pipe Reveal a Dorsoventral Prepattern within the Follicular Epithelium of the Drosophila Ovary
 Cell, Vol. 93, 253 262, April 17, 1998, Copyright 1998 by Cell Press Localized Requirements for windbeutel and pipe Reveal a Dorsoventral Prepattern within the Follicular Epithelium of the Drosophila Ovary
Cell, Vol. 93, 253 262, April 17, 1998, Copyright 1998 by Cell Press Localized Requirements for windbeutel and pipe Reveal a Dorsoventral Prepattern within the Follicular Epithelium of the Drosophila Ovary
BIS &003 Answers to Assigned Problems May 23, Week /18.6 How would you distinguish between an enhancer and a promoter?
 Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Week 9 Study Questions from the textbook: 6 th Edition: Chapter 19-19.6, 19.7, 19.15, 19.17 OR 7 th Edition: Chapter 18-18.6 18.7, 18.15, 18.17 19.6/18.6 How would you distinguish between an enhancer and
Homeotic genes in flies. Sem 9.3.B.6 Animal Science
 Homeotic genes in flies Sem 9.3.B.6 Animal Science So far We have seen that identities of each segment is determined by various regulators of segment polarity genes In arthopods, and in flies, each segment
Homeotic genes in flies Sem 9.3.B.6 Animal Science So far We have seen that identities of each segment is determined by various regulators of segment polarity genes In arthopods, and in flies, each segment
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Interactions of the Drosophila gap gene giant with maternal and zygotic pattern-forming genes
 Development 111, 367-378 (1991) Printed in Great Britain The Company of Biologists Limited 1991 367 Interactions of the Drosophila gap gene giant with maternal and zygotic pattern-forming genes ELIZABETH
Development 111, 367-378 (1991) Printed in Great Britain The Company of Biologists Limited 1991 367 Interactions of the Drosophila gap gene giant with maternal and zygotic pattern-forming genes ELIZABETH
Axis determination in flies. Sem 9.3.B.5 Animal Science
 Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Drosophila Somatic Anterior-Posterior Axis (A-P Axis) Formation
 Formation") Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics
Home Biol 4241 Luria-Delbruck 1943 Hershey-Chase 1952 Meselson-Stahl 1958 Garapin et al. 1978 McClintock 1953 King-Wilson 1975 Sanger et al. 1977 Rothberg et al. 2011 Jeffreys et al. 1985 Bacterial Genetics
cactus, a maternal gene required for proper formation of the dorsoventral morphogen gradient in Drosophila embryos
 Development 11, 371388 (1991) Printed in Great Britain The Company of Biologists Limited 1991 371 cactus, a maternal gene required for proper formation of the dorsoventral morphogen gradient in Drosophila
Development 11, 371388 (1991) Printed in Great Britain The Company of Biologists Limited 1991 371 cactus, a maternal gene required for proper formation of the dorsoventral morphogen gradient in Drosophila
Gaining New Insights into Primitive Strategies for Embryonic Axis Specification Using the Wasp Nasonia
 Functional Development and Embryology 2007 Global Science Books Gaining New Insights into Primitive Strategies for Embryonic Axis Specification Using the Wasp Nasonia Eugenia C. Olesnicky Claude Desplan
Functional Development and Embryology 2007 Global Science Books Gaining New Insights into Primitive Strategies for Embryonic Axis Specification Using the Wasp Nasonia Eugenia C. Olesnicky Claude Desplan
Development of Developmental System System(Mathematical Topics in Biolo. Citation 数理解析研究所講究録 (1993), 827:
, 827:") Title Development of Developmental System System(Mathematical Topics in Biolo Author(s) Takeda, Yasuhiko Citation 数理解析研究所講究録 (1993), 827: 57-74 Issue Date 1993-03 URL http://hdl.handle.net/2433/83294 Right
Title Development of Developmental System System(Mathematical Topics in Biolo Author(s) Takeda, Yasuhiko Citation 数理解析研究所講究録 (1993), 827: 57-74 Issue Date 1993-03 URL http://hdl.handle.net/2433/83294 Right
Exam 2 ID#: November 9, 2006
 Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
NIH Public Access Author Manuscript Curr Opin Genet Dev. Author manuscript; available in PMC 2009 August 1.
 NIH Public Access Author Manuscript Published in final edited form as: Curr Opin Genet Dev. 2008 August ; 18(4): 342 347. doi:10.1016/j.gde.2008.06.002. Dynamics of maternal morphogen gradients in the
NIH Public Access Author Manuscript Published in final edited form as: Curr Opin Genet Dev. 2008 August ; 18(4): 342 347. doi:10.1016/j.gde.2008.06.002. Dynamics of maternal morphogen gradients in the
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology June 16, 2008 Overview What forces affect embryonic development? The embryonic environment: external and internal How do forces
Biology 4361 Developmental Biology Principles of Experimental Embryology June 16, 2008 Overview What forces affect embryonic development? The embryonic environment: external and internal How do forces
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Extranuclear Inheritance
 Extranuclear Inheritance Extranuclear Inheritance The past couple of lectures, we ve been exploring exceptions to Mendel s principles of transmission inheritance. Scientists have observed inheritance patterns
Extranuclear Inheritance Extranuclear Inheritance The past couple of lectures, we ve been exploring exceptions to Mendel s principles of transmission inheritance. Scientists have observed inheritance patterns
1(1)pole hole is required maternally for pattern formation in the terminal regions of the embryo
pole hole is required maternally for pattern formation in the terminal regions of the embryo") Development 106, 145-158 (1989) Printed in Great Britain The Company of Biologists Limited 1989 145 1(1)pole hole is required maternally for pattern formation in the terminal regions of the embryo LINDA
Development 106, 145-158 (1989) Printed in Great Britain The Company of Biologists Limited 1989 145 1(1)pole hole is required maternally for pattern formation in the terminal regions of the embryo LINDA
Chapter 10 Development and Differentiation
 Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
BILD7: Problem Set. 2. What did Chargaff discover and why was this important?
 BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
AP Biology Gene Regulation and Development Review
 AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
Rui Dilão NonLinear Dynamics Group, IST
 1st Conference on Computational Interdisciplinary Sciences (CCIS 2010) 23-27 August 2010, INPE, São José dos Campos, Brasil Modeling, Simulating and Calibrating Genetic Regulatory Networks: An Application
1st Conference on Computational Interdisciplinary Sciences (CCIS 2010) 23-27 August 2010, INPE, São José dos Campos, Brasil Modeling, Simulating and Calibrating Genetic Regulatory Networks: An Application
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Drosophila germline stem cells
 Development 125, 679-690 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV3776 679 Nanos and Pumilio have critical roles in the development and function of Drosophila germline
Development 125, 679-690 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV3776 679 Nanos and Pumilio have critical roles in the development and function of Drosophila germline
Developmental processes Differential gene expression Introduction to determination The model organisms used to study developmental processes
 Date Title Topic(s) Learning Outcomes: Sept 28 Oct 3 1. What is developmental biology and why should we care? 2. What is so special about stem cells and gametes? Developmental processes Differential gene
Date Title Topic(s) Learning Outcomes: Sept 28 Oct 3 1. What is developmental biology and why should we care? 2. What is so special about stem cells and gametes? Developmental processes Differential gene
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
Manipulating the anteroposterior pattern of the Drosophila embryo
 /. Embryol. exp. Morph. 97 Supplement, 169-179 (1986) 169 Printed in Great Britain The Company of Biologists Limited 1986 Manipulating the anteroposterior pattern of the Drosophila embryo HANS GEORG FROHNHOFER,
/. Embryol. exp. Morph. 97 Supplement, 169-179 (1986) 169 Printed in Great Britain The Company of Biologists Limited 1986 Manipulating the anteroposterior pattern of the Drosophila embryo HANS GEORG FROHNHOFER,
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
Thoracic Patterning by the Drosophila Gap Gene hunchback
 Developmental Biology 237, 79 92 (2001) doi:10.1006/dbio.2001.0355, available online at http://www.idealibrary.com on Thoracic Patterning by the Drosophila Gap Gene hunchback Xuelin Wu,* Vikram Vasisht,*
Developmental Biology 237, 79 92 (2001) doi:10.1006/dbio.2001.0355, available online at http://www.idealibrary.com on Thoracic Patterning by the Drosophila Gap Gene hunchback Xuelin Wu,* Vikram Vasisht,*
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis
 18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
Temporal regulation of gene expression in the blastoderm Drosophila embryo
 Temporal regulation of gene expression in the blastoderm Drosophila embryo Glenn K. Yasuda, 1,3 Jayne Baker, 2 and Gerold Schubiger 2 1Department of Genetics, SK-50, and 2Department of Zoology, NJ-15,
Temporal regulation of gene expression in the blastoderm Drosophila embryo Glenn K. Yasuda, 1,3 Jayne Baker, 2 and Gerold Schubiger 2 1Department of Genetics, SK-50, and 2Department of Zoology, NJ-15,
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018
 Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
!!!!!!!! DB3230 Midterm 2 12/13/2013 Name:
 1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
Chromosome Chr Duplica Duplic t a ion Pixley
 Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos by the polyhomeotic gene
 Development 103, 733-741 (1988) Printed in Great Britain The Company of Biologists Limited 1988 733 Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos
Development 103, 733-741 (1988) Printed in Great Britain The Company of Biologists Limited 1988 733 Tissue- and stage-specific control of homeotic and segmentation gene expression in Drosophila embryos
178 Part 3.2 SUMMARY INTRODUCTION
 178 Part 3.2 Chapter # DYNAMIC FILTRATION OF VARIABILITY WITHIN EXPRESSION PATTERNS OF ZYGOTIC SEGMENTATION GENES IN DROSOPHILA Surkova S.Yu. *, Samsonova M.G. St. Petersburg State Polytechnical University,
178 Part 3.2 Chapter # DYNAMIC FILTRATION OF VARIABILITY WITHIN EXPRESSION PATTERNS OF ZYGOTIC SEGMENTATION GENES IN DROSOPHILA Surkova S.Yu. *, Samsonova M.G. St. Petersburg State Polytechnical University,
CARL HASHIMOTO*, SUZANNE GERTTULA and KATHRYN V. ANDERSON
 Development 111, 1021-1028 (1991) Printed in Great Britain The Company of Biologists Limited 1991 1021 Plasma membrane localization of the Toll protein in the syncytial Drosophila embryo: importance of
Development 111, 1021-1028 (1991) Printed in Great Britain The Company of Biologists Limited 1991 1021 Plasma membrane localization of the Toll protein in the syncytial Drosophila embryo: importance of
STUDY UNIT 1 MITOSIS AND MEIOSIS. Klug, Cummings & Spencer Chapter 2. Morphology of eukaryotic metaphase chromosomes. Chromatids
 STUDY UNIT 1 MITOSIS AND MEIOSIS Klug, Cummings & Spencer Chapter 2 Life depends on cell division and reproduction of organisms. Process involves transfer of genetic material. New somatic (body) cells
STUDY UNIT 1 MITOSIS AND MEIOSIS Klug, Cummings & Spencer Chapter 2 Life depends on cell division and reproduction of organisms. Process involves transfer of genetic material. New somatic (body) cells
Full file at CHAPTER 2 Genetics
 CHAPTER 2 Genetics MULTIPLE CHOICE 1. Chromosomes are a. small linear bodies. b. contained in cells. c. replicated during cell division. 2. A cross between true-breeding plants bearing yellow seeds produces
CHAPTER 2 Genetics MULTIPLE CHOICE 1. Chromosomes are a. small linear bodies. b. contained in cells. c. replicated during cell division. 2. A cross between true-breeding plants bearing yellow seeds produces
Genes, Development, and Evolution
 14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
14 Genes, Development, and Evolution Chapter 14 Genes, Development, and Evolution Key Concepts 14.1 Development Involves Distinct but Overlapping Processes 14.2 Changes in Gene Expression Underlie Cell
Novel modes of localization and function of nanos in the wasp Nasonia
 RESEARCH ARTICLE 3813 Development 137, 3813-3821 (2010) doi:10.1242/dev.054213 2010. Published by The Company of Biologists Ltd Novel modes of localization and function of nanos in the wasp Nasonia Jeremy
RESEARCH ARTICLE 3813 Development 137, 3813-3821 (2010) doi:10.1242/dev.054213 2010. Published by The Company of Biologists Ltd Novel modes of localization and function of nanos in the wasp Nasonia Jeremy
SPATIAL PROGRAMMING OF GENE EXPRESSION IN EARLY DROSOPHILA EMBRYOGENESIS
 Ann. Rev. Cell Biol. 1986.2 : 49-80 Copyright 0 1986 by Annual Reviews Inc. All rights reserved SPATIAL PROGRAMMING OF GENE EXPRESSION IN EARLY DROSOPHILA EMBRYOGENESIS Matthew P. Scott Department of Molecular,
Ann. Rev. Cell Biol. 1986.2 : 49-80 Copyright 0 1986 by Annual Reviews Inc. All rights reserved SPATIAL PROGRAMMING OF GENE EXPRESSION IN EARLY DROSOPHILA EMBRYOGENESIS Matthew P. Scott Department of Molecular,
Reading: Chapter 5, pp ; Reference chapter D, pp Problem set F
 Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Mosaic analyses reveal the function of Drosophila Ras in embryonic dorsoventral patterning and dorsal follicle cell morphogenesis
 Development 129, 2209-2222 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV3618 2209 Mosaic analyses reveal the function of Drosophila Ras in embryonic dorsoventral patterning
Development 129, 2209-2222 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV3618 2209 Mosaic analyses reveal the function of Drosophila Ras in embryonic dorsoventral patterning
Two distinct mechanisms for differential positioning of gene expression borders involving the Drosophila gap protein giant
 Development 125, 3765-3774 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5218 3765 Two distinct mechanisms for differential positioning of gene expression borders involving
Development 125, 3765-3774 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV5218 3765 Two distinct mechanisms for differential positioning of gene expression borders involving
Honors Biology Reading Guide Chapter 11
 Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Honors Biology Reading Guide Chapter 11 v Promoter a specific nucleotide sequence in DNA located near the start of a gene that is the binding site for RNA polymerase and the place where transcription begins
Model for the robust establishment of precise proportions in the early Drosophila embryo
 Journal of Theoretical Biology 234 (2005) 13 19 www.elsevier.com/locate/yjtbi Model for the robust establishment of precise proportions in the early Drosophila embryo Tinri Aegerter-Wilmsen a, Christof
Journal of Theoretical Biology 234 (2005) 13 19 www.elsevier.com/locate/yjtbi Model for the robust establishment of precise proportions in the early Drosophila embryo Tinri Aegerter-Wilmsen a, Christof
EDWIN L. FERGUSON and KATHRYN V. ANDERSON
 Development 11, 58-597 (199) Printed in Great Britain The Company of Biologists Limited 199 58 Localized enhancement and repression of the activity of the TGF-fi family member, decapentaplegic, is necessary
Development 11, 58-597 (199) Printed in Great Britain The Company of Biologists Limited 199 58 Localized enhancement and repression of the activity of the TGF-fi family member, decapentaplegic, is necessary
Developmental Biology
 Developmental Biology 376 (2013) 99 112 Contents lists available at SciVerse ScienceDirect Developmental Biology journal homepage: www.elsevier.com/locate/developmentalbiology Genomes and Developmental
Developmental Biology 376 (2013) 99 112 Contents lists available at SciVerse ScienceDirect Developmental Biology journal homepage: www.elsevier.com/locate/developmentalbiology Genomes and Developmental
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Tuesday, December 27, 16
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. Enduring understanding 3.B: Expression of genetic information involves cellular and molecular
Seeing Is Believing: The Bicoid Morphogen Gradient Matures
 Cell, Vol. 116, 143 152, January 23, 2004, Copyright 2004 by Cell Press Seeing Is Believing: The Bicoid Morphogen Gradient Matures Review Anne Ephrussi 1, * and Daniel St Johnston 2, * 1 European Molecular
Cell, Vol. 116, 143 152, January 23, 2004, Copyright 2004 by Cell Press Seeing Is Believing: The Bicoid Morphogen Gradient Matures Review Anne Ephrussi 1, * and Daniel St Johnston 2, * 1 European Molecular
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
10/03/2014. Eukaryotic Development. + Differentiation vs. Development. Differentiation. Development
 Differentiation vs. Development What comes to mind when you think of differentiation? Eukaryotic Development What about development? Presented by: Sean, Daria, Emily, and Maggie Example: Human Development
Differentiation vs. Development What comes to mind when you think of differentiation? Eukaryotic Development What about development? Presented by: Sean, Daria, Emily, and Maggie Example: Human Development
Role for mrna localization in translational activation but not spatial restriction of nanos RNA
 Development 126, 659-669 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV7673 659 Role for mrna localization in translational activation but not spatial restriction of nanos RNA
Development 126, 659-669 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV7673 659 Role for mrna localization in translational activation but not spatial restriction of nanos RNA
DIFFERENTIATION MORPHOGENESIS GROWTH HOW CAN AN IDENTICAL SET OF GENETIC INSTRUCTIONS PRODUCE DIFFERENT TYPES OF CELLS?
 DIFFERENTIATION HOW CAN AN IDENTICAL SET OF GENETIC INSTRUCTIONS PRODUCE DIFFERENT TYPES OF CELLS? MORPHOGENESIS HOW CAN CELLS FORM ORDERED STRUCTURES? GROWTH HOW DO OUR CELLS KNOW WHEN TO STOP DIVIDING
DIFFERENTIATION HOW CAN AN IDENTICAL SET OF GENETIC INSTRUCTIONS PRODUCE DIFFERENT TYPES OF CELLS? MORPHOGENESIS HOW CAN CELLS FORM ORDERED STRUCTURES? GROWTH HOW DO OUR CELLS KNOW WHEN TO STOP DIVIDING
Review. Dorsoventral Axis Formation in the Drosophila Embryo Shaping and Transducing a Morphogen Gradient. Bernard Moussian 1 and Siegfried Roth 2
 Current Biology, Vol. 15, R887 R899, November 8, 2005, 2005 Elsevier Ltd. All rights reserved. DOI 10.1016/j.cub.2005.10.026 Dorsoventral Axis Formation in the Drosophila Embryo Shaping and Transducing
Current Biology, Vol. 15, R887 R899, November 8, 2005, 2005 Elsevier Ltd. All rights reserved. DOI 10.1016/j.cub.2005.10.026 Dorsoventral Axis Formation in the Drosophila Embryo Shaping and Transducing
Early Development in Invertebrates
 Developmental Biology Biology 4361 Early Development in Invertebrates October 25, 2006 Early Development Overview Cleavage rapid cell divisions divisions of fertilized egg into many cells Gastrulation
Developmental Biology Biology 4361 Early Development in Invertebrates October 25, 2006 Early Development Overview Cleavage rapid cell divisions divisions of fertilized egg into many cells Gastrulation
The Genetic Basis of Development
 Chapter 21 The Genetic Basis of Development Lecture Outline Overview The application of genetic analysis and DNA technology to the study of development has brought about a revolution in our understanding
Chapter 21 The Genetic Basis of Development Lecture Outline Overview The application of genetic analysis and DNA technology to the study of development has brought about a revolution in our understanding
The mago nashi locus encodes an essential product required for germ plasm assembly in Drosophila
 Development 120, 1303-1313 (1994) Printed in Great Britain The Company of Biologists Limited 1994 1303 The mago nashi locus encodes an essential product required for germ plasm assembly in Drosophila Phillip
Development 120, 1303-1313 (1994) Printed in Great Britain The Company of Biologists Limited 1994 1303 The mago nashi locus encodes an essential product required for germ plasm assembly in Drosophila Phillip
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
DNA Structure and Function
 DNA Structure and Function Nucleotide Structure 1. 5-C sugar RNA ribose DNA deoxyribose 2. Nitrogenous Base N attaches to 1 C of sugar Double or single ring Four Bases Adenine, Guanine, Thymine, Cytosine
DNA Structure and Function Nucleotide Structure 1. 5-C sugar RNA ribose DNA deoxyribose 2. Nitrogenous Base N attaches to 1 C of sugar Double or single ring Four Bases Adenine, Guanine, Thymine, Cytosine
Chapter 10, 11, 14: Gene Expression, Regulation, and Development Exam
 Chapter 10, 11, 14: Gene Expression, Regulation, and Development Exam Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Why did the original one-gene, one-enzyme
Chapter 10, 11, 14: Gene Expression, Regulation, and Development Exam Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Why did the original one-gene, one-enzyme
Chromosome duplication and distribution during cell division
 CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms.
 Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
Practicing Biology BIG IDEA 3.A 1. Draw, label and describe the structure of DNA and RNA including bonding mechanisms. 2. Using at least 2 well-known experiments, describe which features of DNA and RNA
Cellular Division. copyright cmassengale
 Cellular Division 1 Cell Division All cells are derived from pre- existing cells New cells are produced for growth and to replace damaged or old cells Differs in prokaryotes (bacteria) and eukaryotes (protists,
Cellular Division 1 Cell Division All cells are derived from pre- existing cells New cells are produced for growth and to replace damaged or old cells Differs in prokaryotes (bacteria) and eukaryotes (protists,
7.32/7.81J/8.591J. Rm Rm (under construction) Alexander van Oudenaarden Jialing Li. Bernardo Pando. Rm.
 Alexander van Oudenaarden Jialing Li. Bernardo Pando. Rm.") Introducing... 7.32/7.81J/8.591J Systems Biology modeling biological networks Lectures: Recitations: ti TR 1:00-2:30 PM W 4:00-5:00 PM Rm. 6-120 Rm. 26-204 (under construction) Alexander van Oudenaarden
Introducing... 7.32/7.81J/8.591J Systems Biology modeling biological networks Lectures: Recitations: ti TR 1:00-2:30 PM W 4:00-5:00 PM Rm. 6-120 Rm. 26-204 (under construction) Alexander van Oudenaarden
Control of Gene Expression
 Control of Gene Expression Mechanisms of Gene Control Gene Control in Eukaryotes Master Genes Gene Control In Prokaryotes Epigenetics Gene Expression The overall process by which information flows from
Control of Gene Expression Mechanisms of Gene Control Gene Control in Eukaryotes Master Genes Gene Control In Prokaryotes Epigenetics Gene Expression The overall process by which information flows from
Recruitment of Tube and Pelle to signaling sites at the surface of the
 Development 125, 2443-2450 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV3817 2443 Recruitment of Tube and Pelle to signaling sites at the surface of the Drosophila embryo Par
Development 125, 2443-2450 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV3817 2443 Recruitment of Tube and Pelle to signaling sites at the surface of the Drosophila embryo Par
The maternal NF-κB/Dorsal gradient of Tribolium castaneum: dynamics of early dorsoventral patterning in a short-germ beetle
 Development 127, 5145-5156 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV7840 5145 The maternal NF-κB/Dorsal gradient of Tribolium castaneum: dynamics of early dorsoventral
Development 127, 5145-5156 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV7840 5145 The maternal NF-κB/Dorsal gradient of Tribolium castaneum: dynamics of early dorsoventral
Mitosis, development, regeneration and cell differentiation
 Mitosis, development, regeneration and cell differentiation Mitosis is a type of cell division by binary fission (splitting in two) which occurs in certain eukaryotic cells. Mitosis generates new body
Mitosis, development, regeneration and cell differentiation Mitosis is a type of cell division by binary fission (splitting in two) which occurs in certain eukaryotic cells. Mitosis generates new body