Autophagic clearance of proteasomes in yeast requires the conserved sorting nexin Snx4
|
|
|
- Phoebe Stanley
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on November 6, 2017 as Manuscript M The latest version is at Autophagic clearance of proteasomes in yeast requires the conserved sorting nexin Snx4 Antonia A. Nemec *#, Lauren A. Howell *#, Anna K. Peterson *, Matthew A. Murray *, and Robert J. Tomko Jr * * Department of Biomedical Sciences, College of Medicine, Florida State University, Tallahassee, FL Running Title: Proteasome autophagy requires Snx4 # These authors contributed equally to this work. To whom correspondence should be addressed: Robert J. Tomko Jr., 1115 W. Call St., Tallahassee, FL USA. Tel (850) , robert.tomko@med.fsu.edu Key words: Autophagy, proteasome, sorting nexin (SNX), protein degradation, proteolysis ABSTRACT Turnover of the 26S proteasome by autophagy is an evolutionarily conserved process that governs cellular proteolytic capacity and eliminates inactive particles. In most organisms, proteasomes are located in both the nucleus and cytoplasm. However, the specific autophagy routes for nuclear and cytoplasmic proteasomes are unclear. Here, we investigate the spatial control of autophagic proteasome turnover in budding yeast (Saccharomyces cerevisiae). We found that nitrogen starvation-induced proteasome autophagy is independent of known nucleophagy pathways, but is compromised when nuclear protein export is blocked. Further, via pharmacological tethering of proteasomes to chromatin or the plasma membrane, we provide evidence that nuclear proteasomes at least partially disassemble before autophagic turnover whereas cytoplasmic proteasomes remain largely intact. A targeted screen of autophagy genes identified a requirement for the conserved sorting nexin Snx4 in the autophagic turnover of proteasomes and several other large multisubunit complexes. We demonstrate that Snx4 cooperates with sorting nexins Snx41 and Snx42 to mediate proteasome turnover, and is required for the formation of cytoplasmic proteasome puncta that accumulate when autophagosome formation is blocked. Together, our results support distinct mechanistic paths in the turnover of nuclear versus cytoplasmic proteasomes, and point to a critical role for Snx4 in cytoplasmic agglomeration of proteasomes en route to autophagic destruction. In eukaryotes, two main pathways exist for the regulated destruction of proteins: macroautophagy (hereafter autophagy) and the ubiquitin-proteasome system (UPS) (1). The UPS catalyzes the ubiquitin-dependent degradation of protein substrates by the proteasome, a highly conserved, 2.5 MDa multisubunit ATP-dependent protease complex (2,3). The proteasome consists of a barrel-shaped proteolytic core particle (CP) that houses the peptidase active sites in its interior. The CP can be capped on one or both ends by the 19S regulatory particle (RP). The RP can be further divided into lid and base subcomplexes. The lid contains an enzymatic activity that removes the polyubiquitin targeting signal from the substrate. The base contains a ring of AAA+ family ATPases that use mechanical force derived from ATP to unfold the substrate and translocate it into the interior of the CP for destruction. In contrast to the UPS, autophagy comprises the engulfment of cellular contents in a double-membraned vesicle called an autophagosome for delivery to and destruction in the lysosome (vacuole in yeasts) (4). In the canonical, nonselective Downloaded from at Florida State Univ Med Li on November 9, Copyright 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
2 form of autophagy, a portion of the cytoplasm is delivered in bulk to the vacuole for destruction. This process is directed by the covalent attachment of the small ubiquitin-like protein, Atg8, onto growing autophagosomal membranes to promote membrane expansion and subsequent engulfment of cellular components. Fusion of the autophagosome with the vacuolar membrane delivers the contents into the vacuolar lumen, where the contents are destroyed by various hydrolases. Whereas the UPS can recycle only proteins, autophagy can recycle proteins, lipids, and nucleic acids (5,6). Further, because substrates are typically encapsulated en masse in the cytosol, autophagy is better suited than the UPS for the destruction of exceptionally large or complicated cargoes, such as organelles (7), pathogenic organisms(8), or multisubunit complexes (9-11). In addition to the canonical nonselective (bulk) autophagy, individual cargoes can be earmarked for turnover by autophagy in a process termed selective autophagy. Selective autophagy has recently emerged as a critical homeostatic paradigm and is deregulated in numerous human diseases (12). In selective autophagy, a particular cargo is typically recruited to the assembling autophagosomal membrane, called the phagophore assembly site (PAS), via a cargo-selective autophagy receptor. This receptor binds both the substrate and Atg8-coated PAS membrane to promote cargo engulfment (13). The selective autophagic turnover of the proteasome has recently been demonstrated in mammals (14), plants (15), and in the budding yeast S. cerevisiae (16,17), and occurs in response to nitrogen starvation or treatment of cells with proteasome inhibitors. This conserved pathway serves to control both the abundance and the quality of proteasomes to preserve organismal health. In response to the proteasome inhibitor MG132, proteasomes undergo ubiquitination-dependent delivery to the autophagosome by the autophagy receptor Cue5. The selective recognition of proteasomes for autophagy in response to nitrogen starvation is thus far poorly understood, but appears to involve distinct paths for the RP and CP subcomplexes, based in part on differential requirements for the ubiquitin protease Ubp3 for delivery of RP and CP subunits to the vacuole (17). For both proteasome inhibitortriggered and nitrogen starvation-induced proteasome autophagy, formation of cytoplasmic puncta that are apparently distinct from the PAS precedes delivery to the vacuole. In response to the chemical proteasome inhibitor MG132, proteasomes destined for autophagic turnover colocalize with markers of insoluble protein deposits (16), whereas in nitrogen-starved cells unable to form the PAS due to deletion of ATG17, proteasomes form cytoplasmic puncta of unknown origin or composition (17). Together, these observations suggest that proteasomes may be concentrated or encapsulated prior to their delivery to the PAS, but the mechanism(s) and mediator(s) of this cytoplasmic agglomeration are unknown. One potential barrier to the recruitment of proteasomes to the PAS is that a substantial proportion of proteasomes are located in the nucleus in yeasts (18,19) and in many mammalian cells (20-22), whereas encapsulation of cargo by the growing autophagosome occurs solely in the cytosol. Very little is known about autophagy of nuclear components. In mammals, autophagy of the nuclear lamina has been reported (23). In budding yeast, a pathway called piecemeal autophagy of the nucleus (PMN) has been shown to dispose of nuclear components via direct engulfment of nuclear blebs by the vacuole. This process requires the formation of nucleus-vacuole junctions via Nvj1 (24) and Lam6 (and potentially its homolog, Lam5) (25). More recently, selective autophagy of nuclear components was shown to be mediated by the receptor Atg39 (26). This Atg39-dependent nucleophagy appears to be distinct from PMN. The mechanism by which nuclear proteasomes may be turned over by autophagy, as well as the relative contributions of these known forms of nucleophagy, remain unclear. 2
3 The proteasome, the core autophagy machinery, and autophagic proteasome turnover are each highly conserved from yeasts to humans. Budding yeast therefore serves as an excellent model both for probing the pathway(s) and mediators of proteasome autophagy, as well as for unveiling conserved mechanistic underpinnings of selective autophagy. In this study, we utilize this highly tractable model eukaryote to investigate how proteasomes are sourced and packaged for destruction in the vacuole via selective autophagy. Our data supports a model of distinct mechanistic paths for nuclear vs. cytoplasmic proteasomes, in which nuclear proteasomes are disassembled in the nucleus prior to autophagy. Further, our data suggest that proteasomes transiently accumulate in cytoplasmic puncta in a manner dependent on the conserved sorting nexins Snx4/41/42, before Atg8-dependent delivery to the vacuole for destruction. Lastly, we show that Snx4 dependence for autophagy is shared with at least two other soluble multisubunit complexes. In summary, our study clarifies selective autophagy of proteasomes and unveils a requirement for sorting nexins in autophagy of diverse multisubunit complexes. 3
4 RESULTS Proteasome autophagy requires nuclear export and is independent of known forms of nuclear autophagy. To determine if known forms of nucleophagy mediate the destruction of proteasomes, we first created a yeast strain expressing green fluorescent protein (GFP)-tagged α4 core particle subunit (Figure 1A), encoded by the PRE6 gene, from its chromosomal locus. In yeast, delivery of GFP-fused proteins to the vacuole via autophagy releases a 27 kda GFP fragment that is resistant to proteolysis and can be readily detected via GFP immunoblotting (27). The α4-gfp protein was found in fully assembled CP and in CP capped with regulatory particles (RP 2CP and RP 1CP), as gauged by in-gel GFP fluorescence imaging of proteasomes in whole cell extracts via native polyacrylamide electrophoresis (Figure 1B). These proteasomes were proteolytically active, as inferred from the cleavage of a wellcharacterized fluorogenic peptidase substrate, suc-llvy-amc (Figure 1B). This α4-gfp protein fusion was efficiently delivered to the vacuole upon nitrogen starvation as gauged by accumulation of a GFP fragment on immunoblots, and this process was compromised in cells lacking the core autophagy gene ATG8 or the main vacuolar protease PEP4 (detailed below in Figures 3, 4, and 6), confirming the GFP fragment results from autophagy. We next introduced the chromosomal PRE6-GFP allele into a panel of yeast strains lacking genes known to be required for PMN (NVJ1, LAM5, LAM6) or Atg39-dependent nucleophagy (24-26). To our surprise, none of the tested strains were appreciably compromised for α4-gfp autophagy (Figure 1C), indicating that proteasomes were turned over independent of PMN or Atg39- dependent nucleophagy. The nuclear pore complex mediates the bulk of nucleocytoplasmic traffic, and most proteins are exported from the nucleus via the exportin Crm1. To test a role for nuclear export in proteasome autophagy, we exploited a well-characterized temperaturesensitive CRM1 allele, xpo1-1, that is nearcompletely defective for Crm1-dependent nuclear export at 37 o C (28). Because the core autophagy machinery is located in the cytosol (4), we anticipated that blockade of nuclear export would not globally impair autophagy over our experimental time course. To test this, we monitored nitrogen starvationinduced cleavage of GFP-Atg8, a widely accepted marker of autophagic flux (27), at the permissive (25 o C) or nonpermissive (37 o C) temperature. Although blockade of nuclear export was maintained for at least 12 hours (Figure 1D), no appreciable difference in GFP accumulation from GFP-Atg8 was observed, even after prolonged incubation of xpo1-1 cells at the nonpermissive temperature (Figure 1E). Thus, as anticipated, the cytosolic autophagy machinery remained intact upon disruption of nuclear export, at least over our experimental time course. We next tested the effect of blocking nuclear export on autophagic turnover of GFP-tagged proteasome subunits. As we did for α4, we generated strains expressing GFPtagged alleles of the lid subunit Rpn5 and the base subunit Rpn2 from their chromosomal loci as reporters for these subcomplexes (Figure 1A). As for the α4-gfp fusion, both Rpn5-GFP and Rpn2-GFP were near-fully incorporated into proteolytically active 26S proteasomes (RP 2CP and RP 1CP) based on in-gel GFP fluorescence imaging of proteasomes in whole cell extracts separated by nondenaturing PAGE and peptidase assay (Figure 1B). We then introduced these three alleles individually into the xpo1-1 mutant background. At the permissive temperature, we observed a time-dependent increase in free GFP cleaved from α4-gfp (Figure 1F) in xpo1-1 cells upon nitrogen starvation. In contrast, GFP accumulation was significantly attenuated at the nonpermissive temperature, although some small amount of GFP accumulated in these cells. This GFP signal may be derived from proteasomes that were already cytoplasmic at the time of the temperature shift, as nuclear export blockade remained largely intact over our time course (Figure 1D). We observed similar results 4
5 when a subunit of the base (Rpn2-GFP, Figure 1G) or lid (Rpn5-GFP, Figure 1H) were tested. Rpn11 has previously been reported to interact physically with Crm1 (29). However, despite extensive efforts, we were unable to copurify proteasome subunits of the lid, base, or CP with Crm1 (not shown). Although we cannot exclude the possibility that interaction with Crm1 was too transient or labile to detect under our experimental conditions, we suggest that failed autophagy of proteasome subunits in the xpo1-1 mutant may potentially reflect the failed export of some other essential factor(s) rather than of proteasome subunits themselves. Regardless, because bulk autophagy was not impaired by blockade of nuclear export (Figure 1E), and because autophagy of proteasomes was compromised (Figures 1F- H), we conclude that Crm1-dependent nuclear export of an as-yet unknown factor(s) is required for efficient autophagy of proteasomes. Subcellular tethering of proteasomal subcomplexes provides evidence for disassembly in the nucleus prior to autophagy. We next sought to ask whether nuclear proteasomes exit the nucleus intact, or if they instead can be disassembled prior to nuclear egress. Conflicting reports exist on whether proteasomes disassemble prior to autophagy (16,17). Both of these studies investigated proteasome assembly state in whole cell extracts that presumably contained mixtures of nuclear and cytoplasmic proteasomes, confounding analysis of a particular subcellular proteasome population. To circumvent this limitation, we utilized the anchor-away approach (30) to enrich or deplete proteasomes from the nucleus. The anchor-away approach affords control over the subcellular localization of a protein of interest by inducibly tethering it to an "anchor" protein that is invariantly located either in the cytoplasm or the nucleus. We reasoned that if proteasomes must exit the nucleus intact, then trapping one subcomplex of the proteasome, such as the lid, in the nucleus would prevent autophagy of base or CP subunits. Alternatively, if the base and/or CP is able to dissociate from the lid prior to nuclear egress, then such trapping would have no effect (Figure 2A). We utilized two anchors: the histone Htb2, which localizes exclusively to the nucleus as part of chromatin, and the plasma membrane protein Pma1, which localizes specifically to the plasma membrane. Importantly, we confirmed that Htb2 is not turned over by autophagy in response to nitrogen starvation (Figure 2B), and others have demonstrated the same for Pma1 (31). We introduced chromosomal alleles encoding α4-gfp or Rpn2-GFP into cells expressing lid subunit Rpn11 as a fusion to mcherry and the rapamycin-binding FRB domain (Rpn11-FRB-mCherry). The mcherry tag allowed for visual confirmation of rapamycin-dependent subcellular tethering. These strains also expressed either a nuclear (Htb2-FKBP12) or plasma membrane (PM) (Pma1-2xFKBP12) anchor protein fusion (Figure 2A). In this way, addition of rapamycin tethers Rpn11-FRBmCherry either to chromatin or to the cytosolic face of the PM. Although rapamycin can induce autophagy via inhibition of Tor1 in WT cells, the rapamycin-insensitive TOR1-1 mutation in the anchor-away cells rendered them insensitive to rapamycin-induced autophagy (Figure 2C), and thus rapamycin treatment did not interfere with our approach. As expected, addition of DMSO alone or addition of rapamycin to cells lacking an anchor protein had no effect on the localization of Rpn11-FRB-mCherry or α4- GFP (Figure 2D, compare panels i-vi). Rapamycin addition to the nuclear anchor strain enriched Rpn11-FRB-mCherry and α4- GFP in the nucleus (Figure 2D, panels viixii), whereas rapamycin addition to the PM anchor strain depleted these proteins from the nucleus (Figure 2D, panels xiii-xviii). This relocalization occurred rapidly and was maintained for at least 24 hours in nitrogenstarved cells (Figure 2D). Thus, our anchor system effectively relocalized proteasomes to or from the nucleus under nitrogen starvation conditions. Further, the relocalization of 5
6 much of the α4-gfp signal along with Rpn11-FRB-mCherry upon rapamycin treatment indicates that the majority of proteasomes are fully assembled in vivo, consistent with previous reports (32). We then pretreated cells harboring the nuclear or PM anchor with rapamycin to induce tethering, and then induced autophagy via nitrogen starvation. We then monitored the release of GFP fragment from α4-gfp or Rpn2-GFP as a measure of delivery to the vacuole. Importantly, no compromise of α4- GFP (Figure 2E, left panel) or Rpn2-GFP (Figure 2F, left panel) cleavage were observed upon tethering of Rpn11 to the chromatin, even though Rpn11 was nearcompletely nuclear based on mcherry fluorescence (Figure 2D, panels vii-xii). This strongly suggests that in nuclear proteasomes targeted for autophagy, the CP and the base can dissociate from the lid prior to nuclear egress. In contrast, tethering of Rpn11 to the plasma membrane compromised autophagy of both α4 (Figure 2E, right panel) and Rpn2 (Figure 2F, right panel). This result further confirms that rapamycin-induced tethering was effective under our experimental conditions, and strengthens our conclusion that proteasomes can be disassembled prior to nuclear export for autophagy. Importantly, our finding that autophagy of proteasome subunits Rpn2 and α4 were compromised when Rpn11 was tethered to the cell periphery suggests that disassembly precedes destruction for nuclear, but not cytoplasmic, proteasomes. A targeted genetic screen identifies a requirement for the conserved sorting nexin Snx4 in proteasome autophagy. Because known forms of nucleophagy were dispensable for α4-gfp autophagy, we next performed a targeted screen to identify the full complement of ATG gene products required for turnover of proteasomes upon nitrogen starvation. We picked each of the known ATG gene knockout strains from the yeast knockout library (33) to create a targeted autophagy system knockout library. We then introduced the chromosomal PRE6- GFP allele into this library via standard yeast genetic array methodology (34), allowing us to search for strains that failed to accrue the diagnostic GFP cleavage fragment from α4- GFP upon nitrogen starvation. As anticipated, deletion of ATG genes encoding the core autophagy machinery (ATG1-10; ATG12-16; ATG18) or ATG genes that are specifically required for nitrogen starvationinduced autophagy (ATG17; ATG29; ATG31) completely abolished cleavage of α4-gfp (Figure 3). In contrast, α4-gfp autophagy was retained in nearly all strains harboring deletions of other ATG genes, with the sole exception being deletion of ATG24 (standard name and hereafter SNX4), which completely compromised cleavage of α4- GFP (Figure 3). We thus focused our further efforts on SNX4. SNX4 encodes a member of the sorting nexin containing a Bin/amphiphisin/Rvs (BAR) domain (SNX- BAR) protein family and has obvious orthologs in most eukaryotes, including yeasts and humans (35). SNX-BAR proteins typically contain an N-terminal Phox homology (PX) domain that binds phosphatidylinositol-3-phosphate (PI3P), and a C-terminal BAR domain that promotes association with membranes of defined curvature (36). PI3P is enriched on endosomal membranes, and this association with curved PI3P-containing membranes drives formation of highly tubulated endosomal microdomains for cargo enrichment that then bud off for subsequent cargo transport. In addition to its roles in endosomal sorting (37-41), Snx4 has previously been implicated in the autophagy of mitochondria, peroxisomes, and ER (10,42-45), and in the budding yeast-specific cytoplasm-to-vacuole targeting (CVT) pathway (44), which delivers a select number of resident enzymes to the vacuole in a manner mechanistically similar to selective autophagy. However, the functional mechanism(s) of Snx4 in these processes, as well as its contributions to other forms of selective autophagy, are poorly defined. To better understand how Snx4 regulates autophagy of proteasomes, we first tested whether deletion of SNX4 was simply 6
7 slowing α4-gfp turnover or instead was compromising it completely. We measured the time-dependent accumulation of cleaved GFP in nitrogen-starved WT and snx4δ cells, and in atg8δ cells as a control for general compromise of autophagy. As was the case in atg8δ cells, no detectable accumulation of GFP was observed in snx4δ cells, even after prolonged nitrogen starvation (Figure 4A). Deletion of SNX4 similarly compromised turnover of the base subunit Rpn2-GFP and the lid subunit Rpn5-GFP (Figure 4B), suggesting that Snx4 is required for turnover of all three proteasomal subcomplexes: lid, base, and CP. Proteasome autophagy has been demonstrated recently in response to treatment of cells with the proteasome inhibitor MG132 (15,16). We therefore asked whether Snx4 function was required for turnover of proteasomes in response to MG132. We used cells harboring a deletion in the ATP-binding cassette transporter PDR5 to enhance intracellular accumulation of MG132 by reducing drug efflux (46). As was previously reported (16), MG132 induced autophagy of proteasomes, as evidenced by accumulation of the diagnostic GFP proteolytic fragment (Figure 4C). In contrast to reports by others (15,16), neither MG132 nor the FDA-approved proteasome inhibitor bortezomib (not shown) robustly induced autophagy of proteasomes in our hands; this may result in part from our use of a PDR5 deletion to enhance uptake of these small molecules rather than the previously used and pleiotropic ERG6 mutation (16), which disrupts membrane composition and permeability. Nevertheless, formation of this GFP fragment was ablated upon deletion of ATG8 or the vacuolar protease PEP4, confirming this fragment was produced via autophagy. Importantly, deletion of SNX4 also completely compromised accumulation of GFP, indicating that Snx4 is required for proteasome autophagy in response to both nitrogen starvation and treatment with proteasome inhibitors. We next sought to test the selectivity of Snx4 for proteasome autophagy vs. other soluble protein cargoes. We utilized a synthetic reporter consisting of a GFP-RFP fusion, called Rosella (47), as well as a GFP fusion to an endogenous protein, Pgk1 (48), for reporters of bulk (nonselective) autophagy. In WT cells, both Rosella (Figure 4D) and Pgk1-GFP (Figure 4E) were efficiently cleaved to GFP in response to nitrogen starvation. Importantly, these substrates were both processed to GFP fragments as effectively in cells lacking SNX4 as in WT cells, in agreement with previous reports that SNX4 is dispensable for bulk autophagy (44). Snx4 is required for autophagic turnover of other, unrelated multisubunit complexes. Recently, a requirement for SNX4 in the turnover of the fatty acid synthase (FAS) complex by autophagy was reported, although the mechanism was not investigated (10). As FAS is also a large, soluble, multisubunit complex, we considered the possibility that SNX4 represents a shared requirement for the autophagic turnover of large multisubunit complexes. To test this, we evaluated the impact of SNX4 deletion on nitrogen starvation-induced autophagy of the FAS complex, which has a similar size to the proteasome but is localized exclusively to the cytosol. We also investigated the requirement for SNX4 in the turnover of ribosomal large and small subunits, which are also primarily cytosolic but consist both of protein and nucleic acids. We generated strains expressing from their chromosomal loci fusions of GFP to FAS complex subunit Fas2, or the ribosomal large and small subunits Rpl25 and Rps2, respectively. All three of these genes are essential, so the viability of these strains indicates that the resultant fusion proteins assemble properly into their respective multisubunit complexes and are functional. As reported previously (10), we observed robust turnover of Fas2- GFP by autophagy after 16 hours of nitrogen starvation, as evidenced by the accumulation of a GFP cleavage product. As anticipated, deletion of SNX4 near-completely compromised autophagic turnover of the FAS subunit Fas2-GFP in response to nitrogen starvation as gauged by loss of the 7
8 diagnostic GFP fragment (Figure 5A). We next tested autophagy of Rpl25-GFP (Figure 5B) and Rps2-GFP (Figure 5C) in response to nitrogen starvation. In WT cells, both reporters were readily cleaved to a free GFP fragment, indicating successful delivery to the vacuole. This GFP fragment was absent both for Rpl25-GFP (Figure 5B) and Rps2- GFP (Figure 5C) when SNX4 was deleted, indicating a shared requirement for SNX4 for turnover of the proteasome, FAS, and the ribosome during starvation-induced autophagy. Importantly, these three multisubunit complexes have distinct subcellular distributions (49), are composed of different macromolecules (e.g., proteins vs. proteins and nucleic acids), and apparently share only their large size and multisubunit nature in common, suggesting SNX4 may be uniformly required for selective autophagy of macromolecular complexes. In agreement with an important role in the regulated turnover of several essential multisubunit complexes, cells lacking SNX4 displayed a severe growth defect compared to WT cells in the presence of a low concentration of the starvation mimetic rapamycin (Figure 5D). Snx4 functions prior to vacuolar delivery of α4-gfp and promotes the accumulation of cytoplasmic proteasome puncta when autophagosome formation is disrupted. We next investigated the function of Snx4 in α4- GFP autophagy via microscopy. We first asked whether α4-gfp arrived in the vacuole in nitrogen starved snx4δ cells. For this, we stabilized the protein contents of the vacuole by utilizing cells in which the gene encoding the major vacuolar peptidase, PEP4, had been deleted. A pattern of bright nuclear and diffuse cytosolic fluorescence typical of proteasomes (18,19) was observed in unstarved cells (not shown), and was generally retained upon nitrogen starvation (Figure 6A). Importantly, GFP fluorescence accumulated in the vacuole (circumscribed by the vacuolar membrane protein Vph1- mcherry) upon nitrogen starvation (Figure 6A). In contrast, the GFP fluorescence intensity was much lower in the vacuole of nearly all snx4δ cells. Quantitative vacuolar fluorescence analysis confirmed this observation (Figure 6B), suggesting the defect in snx4δ cells precedes vacuolar delivery. Further, we observed a slight but significant increase in the nuclear fluorescence intensity in nitrogen-starved snx4δ cells, consistent with a compromise of proteasome autophagy (Figure 6B). We next examined the localization of α4-gfp upon nitrogen starvation in atg8δ cells, which are compromised for autophagosome formation (50). Interestingly, we observed the presence of cytoplasmic GFP puncta in atg8δ cells (Figure 6C), similar to those previously observed by others in atg17δ cells (17). Although they were rare, we occasionally observed such puncta in ATG8 WT cells (not shown and Figure 6D). The accumulation of these species in atg8δ cells suggests that they are normally transient, and were accumulating when autophagosome formation was compromised via ATG8 deletion. In contrast to the PAS, which is invariantly perivacuolar in yeast, these puncta were often juxtanuclear and clearly separated from the vacuole in atg8δ cells (Figure 6C, arrowhead). However, they did not colocalize with the PAS or with known perinuclear structures such as the juxtanuclear quality control compartment (51) or the spindle pole body (not shown). Importantly, these puncta were nearcompletely absent in snx4δ cells (Figure 6C,D), indicating that their accumulation requires Snx4. This dependence on Snx4 did not reflect a nonspecific consequence of disrupting endosomal sorting because deletion of the VPS35 gene, which encodes the cargo-binding subunit of the retromer complex essential for retromer-dependent endosomal sorting (52), did not affect accumulation of these puncta (Figure 6C, D). Thus, Snx4 mediates the formation of cytoplasmic puncta that accumulate when autophagy is impaired. Further, the presence of these puncta at low frequency in WT cells, coupled with the impaired delivery of α4- GFP to the vacuole in snx4δ cells (Figure 6A, B), suggests that Snx4 may promote the 8
9 formation of these puncta for subsequent Atg8-dependent delivery to the vacuole. Snx4 cooperates with both Snx41 and Snx42 to mediate proteasome autophagy. SNX4- family nexins typically function as dimers, and in budding yeast, Snx4 can heterodimerize with two other PX-BAR family nexins, Snx41 and Snx42, to form Snx4-Snx41 and Snx4-Snx42 complexes with distinct and overlapping functions (37,40,41,43). In Saccharomyces cerevisiae, Snx4 cooperates with Snx41 to promote retrograde sorting of the autophagy integral membrane protein Atg27, whereas Snx42 is dispensable (37). In contrast, Snx4 and Snx42 cooperate to promote autophagy of peroxisomes and mitochondria (45), and is also required for the CVT pathway (44). Deletion of the Schizosaccharomyces pombe homologs of Snx4, Snx41, or Snx42 alone have no impact on nitrogen starvationinduced autophagy of ER or mitochondria, but deletion of select combinations of these factors compromises autophagy of these organelles (43). To investigate whether Snx4 functions with Snx41 and/or Snx42 in proteasome autophagy, we created strains harboring single deletions of SNX41 and SNX42, as well as a double deletion strain lacking both genes. We then introduced the PRE6-GFP allele into these deletion strains, and assessed the impact on α4-gfp autophagy in response to nitrogen starvation. Deletion of SNX41 alone had no measurable effect on α4-gfp autophagy as ascertained by accumulation of the GFP fragment upon nitrogen starvation, whereas deletion of SNX42 alone caused a small but reproducible defect when autophagy was induced in cells first grown in rich media (Figure 7A). This effect was consistently less pronounced in cells grown in minimal media (Figure 7B and not shown). Importantly, codeletion of SNX41 and SNX42 caused a complete loss of proteasome autophagy upon nitrogen starvation (Figure 7A), indicating that these genes function redundantly with one another. Overexpression of Snx4 in snx41δ snx42δ cells failed to restore α4-gfp autophagy (Figure 7B), supporting a model where Snx4 functioned in the same pathway as Snx41 and Snx42 as Snx4-Snx41 and Snx4-Snx42 heterodimers. Such a redundant function for Snx4-Snx41 and Snx4-Snx42 also rationalizes the complete compromise of proteasome autophagy in snx4δ cells or when both SNX41 and SNX42 were deleted, but not in single snx41δ or snx42δ mutants (Figure 7A). To better understand the relationship between Snx4, Snx41, Snx42, and proteasome autophagy, we investigated the effect of SNX41 and SNX42 deletion on Snx4 subcellular localization. We expressed Snx4- GFP from the chromosomal locus, and verified that this allele was functional (Figure 7C). In nitrogen-starved WT cells expressing Snx4-GFP, we observed primarily cytoplasmic puncta of Snx4 (Figure 7D), as is typical of endosomal proteins and as has recently been reported by others (37). We were unable to detect colocalization of mcherry-tagged proteasome subunits with these cytoplasmic Snx4-GFP puncta in atg8δ cells (not shown), indicating they are either distinct structures from the α4-gfp puncta (Figure 6C) or possibly that they colocalize transiently or only rarely. Nonetheless, deletion of SNX41 alone had no apparent effect on the appearance of Snx4-GFP puncta (Figure 7D), and no statistically significant effect on their frequency (Figure 7E); this was in agreement with the robust α4-gfp autophagy observed in snx41δ cells (Figure 7A). In contrast, deletion of SNX42 alone caused a small but reproducible decrease in the frequency of Snx4-GFP puncta (Figure 7E), and the concomitant appearance of diffuse cytoplasmic fluorescence in most cells (Figure 7D), again consistent with the partial loss of α4-gfp autophagy in this mutant (Figure 7A). Importantly, codeletion of SNX41 and SNX42 resulted in a nearcomplete loss of Snx4-GFP puncta and a further increase in diffuse cytoplasmic fluorescence (Figure 7D, E). This strongly suggests that proper localization of Snx4 to such puncta is important for proteasome autophagy. 9
10 Snx4 localizes to membranes via its PI3P-binding PX domain, and mutation of a critical tyrosine residue in this domain to alanine (Y79A) has previously been shown to disrupt PI3P-binding and cause partial mislocalization of Snx4 (44). We thus tested the impact of this mutation on α4-gfp autophagy. We introduced plasmids expressing either WT SNX4 or the snx4(y79a) PX domain mutant into snx4δ cells. As expected, provision of WT SNX4 to snx4δ cells restored turnover of α4-gfp upon nitrogen starvation. In contrast, provision of the snx4(y79a) mutant restored α4-gfp autophagy to only approx. 50% of that observed with the WT SNX4 allele (Figure 7F). Taken together, these results indicate that Snx41 and Snx42 promote proper localization of Snx4 to perivacuolar puncta, and that disruption of this localization, either via reducing PI3P-binding by Snx4 or by disrupting formation of Snx4- containing heterodimers, compromises proteasome autophagy. Consistent with this model, codeletion of SNX41 and SNX42 severely compromised formation of Snx4- dependent cytoplasmic α4-gfp puncta in atg8δ cells upon nitrogen starvation (Figure 7G). We note that the relative loss of α4-gfp puncta in snx41δ, snx42δ, or snx41δ snx42δ cells closely mirrored the impact both on α4- GFP autophagy (Figure 7A) and delocalization of Snx4-GFP (Figure 7E). Thus, we conclude that Snx4 cooperates with Snx41 and Snx42 to drive the formation of cytoplasmic proteasome puncta, likely en route to the vacuole for destruction, in a manner dependent on proper localization of Snx4 to PI3P-containing membranes. 10
11 DISCUSSION Here, we provide some of the first insights into the subcellular trafficking of proteasomes to the vacuole for autophagy. We demonstrate that nuclear proteasomes can be disassembled prior to autophagy, and that proteasome autophagy is independent of known nucleophagy genes and is impaired when Crm1-dependent nuclear export is compromised. Turnover of proteasomes is completely compromised by loss of Snx4, which mediates the agglomeration of proteasomes into cytoplasmic puncta prior to delivery to the vacuole (Figure 8). These data support a model in which autophagic turnover of nuclear and cytoplasmic proteasomes converge at the stage of Snx4- dependent agglomeration prior to delivery to the vacuole for destruction. Importantly, Snx4 mediates the turnover of diverse multisubunit complexes (Figure 5), including the ribosome and FAS, indicating it is broadly involved in selective autophagy of soluble multisubunit complexes. In yeasts and many mammalian cells, a substantial portion of mature proteasomes are located in the nucleus, an organelle that was generally considered to be spared from autophagic destruction until only recently. Although autophagy of proteasomes has been reported in yeast, plants, and mammals, whether nuclear and cytoplasmic proteasomes were recognized and processed via the same mechanism was not known. Our data suggest that in yeast, nuclear proteasomes are disassembled prior to autophagy (Figure 2, Figure 8). These findings are consistent with recent reports that at least some fraction of proteasomes are disassembled prior to nitrogen starvationinduced autophagy (17). Why some nuclear autophagy substrates are turned over via nucleus-specific selective autophagy pathways such as PMN or Atg39-dependent nucleophagy whereas others may be independent is thus far unknown, but it may allow greater control over turnover of particular sets of nuclear autophagic substrates. Alternatively, some feature(s) of the substrate, such as membrane embedment or association, may necessitate a particular route of delivery to the vacuole from the nucleus. Our findings that autophagic turnover of both the RP base and the CP are unaffected when the lid is tethered to chromatin suggests that, at minimum, the lid dissociates from the base and CP during autophagy of nuclear proteasomes. We suspect that all three subcomplexes dissociate from one another. In support of this, we observed different amounts of GFP liberated from lid, base, and CP subunits upon nitrogen starvation (Figure 1F-H, permissive temperature), and different requirements for autophagy of the base subunit Rpn1 and the CP subunit β5 were reported by others (17). The reason for disassembly of nuclear but not cytoplasmic proteasomes prior to autophagy is unknown. Although export through the nuclear pore has not to our knowledge been demonstrated for the proteasome, fully assembled proteasomes have been reported to pass intact through the nuclear pore into the nucleus (32,53), arguing against the possibility that proteasomes cannot otherwise fit. Instead, we suggest that this disassembly may be necessary to limit a sudden influx of proteasome activity into the cytosol prior to autophagic engulfment that could otherwise disrupt cellular processes. Indeed, the activities of individual proteasomal subcomplexes are much lower than in the holocomplex (54,55). Thus, such dissociation into subcomplexes may serve to inactivate or attenuate these proteasomes until they can be encapsulated by the autophagosome and shielded from cytosolic proteasome substrates. An additional question arising is the nature of the signal for disassembly of nuclear proteasomes upon nitrogen starvation-induced autophagy. Although the interaction between the RP base and the CP has been demonstrated to be sensitive to ATP levels in vitro (56), nitrogen starvation does not impact the intracellular ATP concentration, at least in yeast (57,58). Also, we are not aware of any evidence that lid and base subcomplexes dissociate in a nucleotide-dependent manner. Further, such a nucleotide-dependent disassembly would 11
12 be anticipated to similarly affect cytosolic proteasomes. Instead, we speculate that a nucleus-specific signal promotes disassembly of these subcomplexes. Phosphorylation may be a likely candidate. We note that nitrogen starvation regulates the activity as well as the subcellular localization of several kinases (reviewed in (59)). Further, phosphorylation has recently been shown to recruit the proteasome-binding protein Ecm29 (60), which disassembles proteasomes in response to other stimuli (61). Based on the loss of α4-gfp puncta in snx4δ cells, we propose that Snx4 is involved either in the clustering of soluble proteasomes into phase-separated granules similar to those observed in response to carbon deprivation (62), or is instead involved in the enrichment of proteasomes in membrane-encased structures that are subsequently undergo Atg8-dependent engulfment and delivery to the vacuole. Considering that Snx4 is a PI3P-binding protein involved in membrane protein sorting (37,40,41) and that its membrane binding function is necessary for proteasome autophagy (Figure 7), we favor the latter possibility. Indeed, while this manuscript was being prepared, a computational clustering study proposed a role for Snx4 in the membrane engulfment of selective autophagy cargos (63), although the relationship to Atg8-dependent autophagosomal membrane expansion is not yet known. Although bulk and selective autophagy pathways share the core autophagy machinery, differences have emerged in the requirements for auxiliary factors for individual cargoes. For example, deletion of SNX4 alone has no effect on starvation-induced autophagy of the vacuolar enzyme Ape1 (64), on organelle autophagy (43), or on bulk autophagy (Figs. 4D,E), whereas codeletion of SNX42 with SNX4 compromises turnover both of Ape1 and organelles. In contrast, deletion of SNX4 alone completely compromises autophagy of proteasomes and two other large, macromolecular complexes the FAS complex and the ribosome (Figure 5). This suggests that these cargoes may be differentially dependent on Snx4-containing complexes. The mechanistic basis for this remains to be determined, but individual roles for Snx4-Snx41 and Snx4-Snx42 complexes are emerging and are likely to provide additional insight. The underlying function of Snx4 in proteasome autophagy, as well as in other forms of selective autophagy, remains unclear. Numerous studies have documented a contribution of the endosomal sorting machinery to selective autophagy (reviewed in (65)), and early endosomal markers have been observed in autophagic vesicles (66). Specifically, roles in membrane delivery to the autophagosome from the Golgi (64) and trafficking of autophagy proteins (37) have been proposed for Snx4. However, each of these roles required deletion of multiple genes to manifest a defect in selective autophagy of other cargoes. Our finding that deletion of Snx4 completely compromised proteasome autophagy suggests a more central role for Snx4 in this process. We have anecdotally observed that the α4-gfp puncta that accumulate in atg8δ cells are often juxtaposed to the early endosome marker mcherry-tlg1 (unpublished results), raising the intriguing possibility that Snx4 may mediate engulfment of proteasomes at the endosome, or instead may promote extrusion of proteasomes destined for the vacuole from the endosomal compartment. The direct delivery of cargo to the autophagosome from early endosomes via vesicle fusion is not entirely unprecedented (66,67). Our observation that any perturbations that disrupt Snx4 localization compromise proteasome autophagy (Figure 7) is consistent with this hypothesis. Live-cell microscopy studies will be necessary to determine whether proteasomes pass through the endosome en route to the vacuole. Further, identification of additional genes necessary for the accumulation of α4-gfp puncta in atg8δ cells will provide valuable insight into the origin of these species, as well as into the mechanism of Snx4-dependent cargo processing for autophagy. 12
13 EXPERIMENTAL PROCEDURES Yeast strains and media. Yeast manipulations were carried out according to standard protocols (68). Strains used in this study are listed in Supplementary Table S1. For nitrogen starvation, yeast were grown in YPD or synthetic dropout medium to midlog, centrifuged at 4,000 x g for two minutes, washed once with 15 ml of dh 2O, centrifuged again, and then resuspended to an OD 600 of 1.0 in SD-N media (0.67% yeast nitrogen base without amino acids or ammonium sulfate, 2% glucose). For temperature shift experiments using the xpo1-1 allele, cells were grown at 25 o C and subjected to nitrogen starvation as above. After three hours, cultures were split in two; one half continued to be incubated at 25 o C whereas the other half was subsequently incubated at 37 o C to inactivate the xpo1-1 mutant allele. Where indicated, mid-log phase yeast grown in YPD were treated with 50 µm MG132 in DMSO as described in the figure legends. For anchor-away experiments, yeast were grown to mid-log in YPD, and then treated for three hours with 1 µg/ml rapamycin (Enzo Life Sciences) prior to nitrogen starvation as above in SD-N supplemented with 1 µg/ml rapamycin. For growth assays, the indicated strains were spotted as six-fold serial dilutions in water onto the indicated media. Plasmids. All plasmids were constructed using standard molecular cloning techniques using TOP10 F (Invitrogen) as a host strain. Plasmids used in this study are listed in Supplementary Table S2. Complete sequences and construction details are available upon request. Genetic Array Analysis. ATG deletion strains were hand-picked from the YKO library (Open Biosystems) and arrayed in 96- well format. A query strain derived from Y7092 (34) was produced by sequentially integrating PRE6-yEGFP:caURA3MX4 and pdr5δ::natmx4 cassettes into the respective chromosomal loci. After confirmation of integration, this resultant query strain (RTY1207) was mated to the ATG deletion array to generate haploid triple mutant strains. The presence of the pdr5δ allele did not affect nitrogen starvation-induced autophagy of α4-gfp in our hands. SDS-PAGE and immunoblotting. Cell extracts were prepared from equal numbers of cells via the alkaline lysis method (69), cleared via centrifugation, and the supernatants were loaded directly onto 12% SDS-PAGE gels. After electrophoresis, proteins were transferred to PVDF membranes (EMD Millipore) and probed with antibodies against GFP (Roche, Cat# , 1:2000 dilution). Blots were imaged on an Azure Biosystems c300 or a Bio-Rad ChemiDoc MP using HRPconjugated secondary antibodies (GE Lifesciences, Cat# , 1:5000) and ECL reagent. The percentage of free GFP liberated from a given protein was calculated for each sample by dividing the band intensity from the free GFP product by the sum of the intensities of free GFP plus the intact GFP fusion protein, and thus is internally normalized across sample lanes. Band intensities were confirmed to be in the linear signal range using ImageLab software (Bio-Rad), and were quantified from raw image files. GFP cleavage measurements reported in the figures are of the blot shown, but are representative of multiple experiments. Non-denaturing polyacrylamide electrophoresis of cell extracts and fluorescence analyses. Cell extracts (100 μg total protein) were separated by nondenaturing polyacrylamide electrophoresis exactly as described previously (70). To visualize in-gel GFP fluorescence, gels were washed once in deionized water after electrophoresis, and were imaged with a Typhoon Imager (GE Life Sciences) using excitation at 488 nm, and measurement of emission through a 520 nm bandpass filter. For measurement of suc-llvy-amc hydrolysis, nondenaturing gels in which 100 μg of cell extract had been separated were incubated in Overlay buffer (50 mm Tris-Cl, ph 7.5, 5 mm MgCl 2, 10% glycerol, 1 mm 13
14 ATP) containing 50 μm suc-llvy-amc for 30 minutes at 30 o C with occasional gentle agitation. Liberated AMC was detected in a Bio-Rad Gel-doc XR imaging system with the pre-programmed excitation and emission settings for ethidium bromide. For anti-20s immunoblotting, 50 μg of cell extract was separated exactly as above before transfer to PVDF membranes and immunoblotting with anti-20s antibodies (Enzo, Cat# BML- PW9355, 1:2000 dilution). Microscopy. All micrographs were collected on an EVOS FL Cell Imaging System (Thermo Fisher Scientific) equipped with GFP and RFP filter sets. Identical exposure times and light intensities were used for each image in a given experiment. Briefly, cells were collected via centrifugation at 10,000 x g for 30 seconds, followed by resuspension in the appropriate medium at 1/10 th of the original culture volume. To minimize cell movement during imaging, pre-cleaned microscope slides were overlaid with a thin pad of 3% agarose containing the appropriate culture medium onto which the cells were applied. In Figure 6B, vacuolar or nuclear fluorescence intensity was measured using ImageJ on raw image files, and individual cell measurements are plotted. Statistical analysis and reproducibility. All experiments were performed three times or more to ensure reproducibility. Statistical analysis was carried out using Graph Pad Prism 7.0 software, via two-tailed Mann- Whitney test (Figure 6B), one-way ANOVA with Tukey s test for multiple comparisons (Figure 6D), two-way ANOVA with Sidak s test for multiple comparisons (Figure 7E), or one-way ANOVA with Dunnett test for multiple comparisons (Figure 7G). Unless otherwise indicated, at least 100 cells per condition were analyzed for statistical analysis of micrographs. Statistical significance was considered p <
15 ACKNOWLEDGMENTS This work was supported by start-up funds from the Florida State University College of Medicine and NIH/NIGMS grant 1R01GM to R.J.T.Jr. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. The authors thank Rodney Devenish (Monash University), Akash Gunjan (FSU-COM), and Daniel Klionsky (University of Michigan) for reagents, and the FSU yeast community for helpful comments and feedback. 15
16 CONFLICT OF INTEREST The authors declare no competing financial interests. 16
17 AUTHOR CONTRIBUTIONS A.A.N and R. J. T. Jr. conceived and designed the study. Most experiments were replicated by more than one author. L.A.H performed or contributed reagents to Figures 1D-H, Figure 2, Figure 6, and Figure 7. A.A.N. performed experiments in Figures 1C, Figures 2-4, Figure 6 C and D. A.K.P. contributed reagents to all figures and helped conduct the screen in Figure 3. M.A.M performed experiments in Figure 5. R.J.T.Jr. contributed reagents and performed experiments in Figures 1B, 6A, and 6B. R.J.T. Jr. wrote the manuscript with input from all authors. 17
18 REFERENCES 1. Cohen-Kaplan, V., Livneh, I., Avni, N., Cohen-Rosenzweig, C., and Ciechanover, A. (2016) The ubiquitin-proteasome system and autophagy: Coordinated and independent activities. Int J Biochem Cell Biol 79, Finley, D. (2009) Recognition and processing of ubiquitin-protein conjugates by the proteasome. Annual review of biochemistry 78, Tomko, R. J., Jr., and Hochstrasser, M. (2013) Molecular architecture and assembly of the eukaryotic proteasome. Annual review of biochemistry 82, Feng, Y., He, D., Yao, Z., and Klionsky, D. J. (2014) The machinery of macroautophagy. Cell Res 24, Welter, E., and Elazar, Z. (2015) Autophagy mediates nonselective RNA degradation in starving yeast. EMBO J 34, Jaishy, B., and Abel, E. D. (2016) Lipids, lysosomes, and autophagy. Journal of lipid research 57, Anding, A. L., and Baehrecke, E. H. (2017) Cleaning House: Selective Autophagy of Organelles. Dev Cell 41, Wileman, T. (2013) Autophagy as a defence against intracellular pathogens. Essays Biochem 55, Kraft, C., Deplazes, A., Sohrmann, M., and Peter, M. (2008) Mature ribosomes are selectively degraded upon starvation by an autophagy pathway requiring the Ubp3p/Bre5p ubiquitin protease. Nat Cell Biol 10, Shpilka, T., Welter, E., Borovsky, N., Amar, N., Shimron, F., Peleg, Y., and Elazar, Z. (2015) Fatty acid synthase is preferentially degraded by autophagy upon nitrogen starvation in yeast. Proc Natl Acad Sci U S A 112, Le Fourn, V., Park, S., Jang, I., Gaplovska-Kysela, K., Guhl, B., Lee, Y., Cho, J. W., Zuber, C., and Roth, J. (2013) Large protein complexes retained in the ER are dislocated by non-copii vesicles and degraded by selective autophagy. Cellular and molecular life sciences : CMLS 70, Mizushima, N., Levine, B., Cuervo, A. M., and Klionsky, D. J. (2008) Autophagy fights disease through cellular self-digestion. Nature 451, Stolz, A., Ernst, A., and Dikic, I. (2014) Cargo recognition and trafficking in selective autophagy. Nat Cell Biol 16, Cohen-Kaplan, V., Livneh, I., Avni, N., Fabre, B., Ziv, T., Kwon, Y. T., and Ciechanover, A. (2016) p62- and ubiquitin-dependent stress-induced autophagy of the mammalian 26S proteasome. Proc Natl Acad Sci U S A 113, E7490-E Marshall, R. S., Li, F., Gemperline, D. C., Book, A. J., and Vierstra, R. D. (2015) Autophagic Degradation of the 26S Proteasome Is Mediated by the Dual ATG8/Ubiquitin Receptor RPN10 in Arabidopsis. Mol Cell 58, Marshall, R. S., McLoughlin, F., and Vierstra, R. D. (2016) Autophagic Turnover of Inactive 26S Proteasomes in Yeast Is Directed by the Ubiquitin Receptor Cue5 and the Hsp42 Chaperone. Cell reports 16, Waite, K. A., De-La Mota-Peynado, A., Vontz, G., and Roelofs, J. (2016) Starvation Induces Proteasome Autophagy with Different Pathways for Core and Regulatory Particles. J Biol Chem 291, Enenkel, C., Lehmann, A., and Kloetzel, P. M. (1999) GFP-labelling of 26S proteasomes in living yeast: insight into proteasomal functions at the nuclear envelope/rough ER. Molecular biology reports 26, Enenkel, C., Lehmann, A., and Kloetzel, P. M. (1998) Subcellular distribution of proteasomes implicates a major location of protein degradation in the nuclear envelope- ER network in yeast. EMBO J 17,
19 20. Tanaka, K., Kumatori, A., Ii, K., and Ichihara, A. (1989) Direct evidence for nuclear and cytoplasmic colocalization of proteasomes (multiprotease complexes) in liver. Journal of cellular physiology 139, Palmer, A., Rivett, A. J., Thomson, S., Hendil, K. B., Butcher, G. W., Fuertes, G., and Knecht, E. (1996) Subpopulations of proteasomes in rat liver nuclei, microsomes and cytosol. Biochem J 316 ( Pt 2), Brooks, P., Fuertes, G., Murray, R. Z., Bose, S., Knecht, E., Rechsteiner, M. C., Hendil, K. B., Tanaka, K., Dyson, J., and Rivett, J. (2000) Subcellular localization of proteasomes and their regulatory complexes in mammalian cells. Biochem J 346 Pt 1, Dou, Z., Xu, C., Donahue, G., Shimi, T., Pan, J. A., Zhu, J., Ivanov, A., Capell, B. C., Drake, A. M., Shah, P. P., Catanzaro, J. M., Ricketts, M. D., Lamark, T., Adam, S. A., Marmorstein, R., Zong, W. X., Johansen, T., Goldman, R. D., Adams, P. D., and Berger, S. L. (2015) Autophagy mediates degradation of nuclear lamina. Nature 527, Mijaljica, D., Prescott, M., and Devenish, R. J. (2012) A late form of nucleophagy in Saccharomyces cerevisiae. PloS one 7, e Elbaz-Alon, Y., Eisenberg-Bord, M., Shinder, V., Stiller, S. B., Shimoni, E., Wiedemann, N., Geiger, T., and Schuldiner, M. (2015) Lam6 Regulates the Extent of Contacts between Organelles. Cell reports 12, Mochida, K., Oikawa, Y., Kimura, Y., Kirisako, H., Hirano, H., Ohsumi, Y., and Nakatogawa, H. (2015) Receptor-mediated selective autophagy degrades the endoplasmic reticulum and the nucleus. Nature 522, Klionsky, D. J., Abdelmohsen, K., Abe, A., Abedin, M. J., Abeliovich, H., Acevedo Arozena, A., Adachi, H., Adams, C. M., Adams, P. D., Adeli, K., Adhihetty, P. J., Adler, S. G., Agam, G., Agarwal, R., Aghi, M. K., Agnello, M., Agostinis, P., Aguilar, P. V., Aguirre-Ghiso, J., Airoldi, E. M., Ait-Si-Ali, S., Akematsu, T., Akporiaye, E. T., Al- Rubeai, M., Albaiceta, G. M., Albanese, C., Albani, D., Albert, M. L., Aldudo, J., Algul, H., Alirezaei, M., Alloza, I., Almasan, A., Almonte-Beceril, M., Alnemri, E. S., Alonso, C., Altan-Bonnet, N., Altieri, D. C., Alvarez, S., Alvarez-Erviti, L., Alves, S., Amadoro, G., Amano, A., Amantini, C., Ambrosio, S., Amelio, I., Amer, A. O., Amessou, M., Amon, A., An, Z., Anania, F. A., Andersen, S. U., Andley, U. P., Andreadi, C. K., Andrieu-Abadie, N., Anel, A., Ann, D. K., Anoopkumar-Dukie, S., Antonioli, M., Aoki, H., Apostolova, N., Aquila, S., Aquilano, K., Araki, K., Arama, E., Aranda, A., Araya, J., Arcaro, A., Arias, E., Arimoto, H., Ariosa, A. R., Armstrong, J. L., Arnould, T., Arsov, I., Asanuma, K., Askanas, V., Asselin, E., Atarashi, R., Atherton, S. S., Atkin, J. D., Attardi, L. D., Auberger, P., Auburger, G., Aurelian, L., Autelli, R., Avagliano, L., Avantaggiati, M. L., Avrahami, L., Awale, S., Azad, N., Bachetti, T., Backer, J. M., Bae, D. H., Bae, J. S., Bae, O. N., Bae, S. H., Baehrecke, E. H., Baek, S. H., Baghdiguian, S., Bagniewska- Zadworna, A., Bai, H., Bai, J., Bai, X. Y., Bailly, Y., Balaji, K. N., Balduini, W., Ballabio, A., Balzan, R., Banerjee, R., Banhegyi, G., Bao, H., Barbeau, B., Barrachina, M. D., Barreiro, E., Bartel, B., Bartolome, A., Bassham, D. C., Bassi, M. T., Bast, R. C., Jr., Basu, A., Batista, M. T., Batoko, H., Battino, M., Bauckman, K., Baumgarner, B. L., Bayer, K. U., Beale, R., Beaulieu, J. F., Beck, G. R., Jr., Becker, C., Beckham, J. D., Bedard, P. A., Bednarski, P. J., Begley, T. J., Behl, C., Behrends, C., Behrens, G. M., Behrns, K. E., Bejarano, E., Belaid, A., Belleudi, F., Benard, G., Berchem, G., Bergamaschi, D., Bergami, M., Berkhout, B., Berliocchi, L., Bernard, A., Bernard, M., Bernassola, F., Bertolotti, A., Bess, A. S., Besteiro, S., Bettuzzi, S., Bhalla, S., Bhattacharyya, S., Bhutia, S. K., Biagosch, C., Bianchi, M. W., Biard-Piechaczyk, M., Billes, V., Bincoletto, C., Bingol, B., Bird, S. W., Bitoun, M., Bjedov, I., Blackstone, C., Blanc, L., Blanco, G. A., Blomhoff, H. K., Boada-Romero, E., Bockler, S., Boes, M., Boesze-Battaglia, K., Boise, L. H., Bolino, A., Boman, A., Bonaldo, P., Bordi, M., 19
20 Bosch, J., Botana, L. M., Botti, J., Bou, G., Bouche, M., Bouchecareilh, M., Boucher, M. J., Boulton, M. E., Bouret, S. G., Boya, P., Boyer-Guittaut, M., Bozhkov, P. V., Brady, N., Braga, V. M., Brancolini, C., Braus, G. H., Bravo-San Pedro, J. M., Brennan, L. A., Bresnick, E. H., Brest, P., Bridges, D., Bringer, M. A., Brini, M., Brito, G. C., Brodin, B., Brookes, P. S., Brown, E. J., Brown, K., Broxmeyer, H. E., Bruhat, A., Brum, P. C., Brumell, J. H., Brunetti-Pierri, N., Bryson-Richardson, R. J., Buch, S., Buchan, A. M., Budak, H., Bulavin, D. V., Bultman, S. J., Bultynck, G., Bumbasirevic, V., Burelle, Y., Burke, R. E., Burmeister, M., Butikofer, P., Caberlotto, L., Cadwell, K., Cahova, M., Cai, D., Cai, J., Cai, Q., Calatayud, S., Camougrand, N., Campanella, M., Campbell, G. R., Campbell, M., Campello, S., Candau, R., Caniggia, I., Cantoni, L., Cao, L., Caplan, A. B., Caraglia, M., Cardinali, C., Cardoso, S. M., Carew, J. S., Carleton, L. A., Carlin, C. R., Carloni, S., Carlsson, S. R., Carmona-Gutierrez, D., Carneiro, L. A., Carnevali, O., Carra, S., Carrier, A., Carroll, B., Casas, C., Casas, J., Cassinelli, G., Castets, P., Castro- Obregon, S., Cavallini, G., Ceccherini, I., Cecconi, F., Cederbaum, A. I., Cena, V., Cenci, S., Cerella, C., Cervia, D., Cetrullo, S., Chaachouay, H., Chae, H. J., Chagin, A. S., Chai, C. Y., Chakrabarti, G., Chamilos, G., Chan, E. Y., Chan, M. T., Chandra, D., Chandra, P., Chang, C. P., Chang, R. C., Chang, T. Y., Chatham, J. C., Chatterjee, S., Chauhan, S., Che, Y., Cheetham, M. E., Cheluvappa, R., Chen, C. J., Chen, G., Chen, G. C., Chen, G., Chen, H., Chen, J. W., Chen, J. K., Chen, M., Chen, M., Chen, P., Chen, Q., Chen, Q., Chen, S. D., Chen, S., Chen, S. S., Chen, W., Chen, W. J., Chen, W. Q., Chen, W., Chen, X., Chen, Y. H., Chen, Y. G., Chen, Y., Chen, Y., Chen, Y., Chen, Y. J., Chen, Y. Q., Chen, Y., Chen, Z., Chen, Z., Cheng, A., Cheng, C. H., Cheng, H., Cheong, H., Cherry, S., Chesney, J., Cheung, C. H., Chevet, E., Chi, H. C., Chi, S. G., Chiacchiera, F., Chiang, H. L., Chiarelli, R., Chiariello, M., Chieppa, M., Chin, L. S., Chiong, M., Chiu, G. N., Cho, D. H., Cho, S. G., Cho, W. C., Cho, Y. Y., Cho, Y. S., Choi, A. M., Choi, E. J., Choi, E. K., Choi, J., Choi, M. E., Choi, S. I., Chou, T. F., Chouaib, S., Choubey, D., Choubey, V., Chow, K. C., Chowdhury, K., Chu, C. T., Chuang, T. H., Chun, T., Chung, H., Chung, T., Chung, Y. L., Chwae, Y. J., Cianfanelli, V., Ciarcia, R., Ciechomska, I. A., Ciriolo, M. R., Cirone, M., Claerhout, S., Clague, M. J., Claria, J., Clarke, P. G., Clarke, R., Clementi, E., Cleyrat, C., Cnop, M., Coccia, E. M., Cocco, T., Codogno, P., Coers, J., Cohen, E. E., Colecchia, D., Coletto, L., Coll, N. S., Colucci-Guyon, E., Comincini, S., Condello, M., Cook, K. L., Coombs, G. H., Cooper, C. D., Cooper, J. M., Coppens, I., Corasaniti, M. T., Corazzari, M., Corbalan, R., Corcelle-Termeau, E., Cordero, M. D., Corral-Ramos, C., Corti, O., Cossarizza, A., Costelli, P., Costes, S., Cotman, S. L., Coto-Montes, A., Cottet, S., Couve, E., Covey, L. R., Cowart, L. A., Cox, J. S., Coxon, F. P., Coyne, C. B., Cragg, M. S., Craven, R. J., Crepaldi, T., Crespo, J. L., Criollo, A., Crippa, V., Cruz, M. T., Cuervo, A. M., Cuezva, J. M., Cui, T., Cutillas, P. R., Czaja, M. J., Czyzyk-Krzeska, M. F., Dagda, R. K., Dahmen, U., Dai, C., Dai, W., Dai, Y., Dalby, K. N., Dalla Valle, L., Dalmasso, G., D'Amelio, M., Damme, M., Darfeuille-Michaud, A., Dargemont, C., Darley-Usmar, V. M., Dasarathy, S., Dasgupta, B., Dash, S., Dass, C. R., Davey, H. M., Davids, L. M., Davila, D., Davis, R. J., Dawson, T. M., Dawson, V. L., Daza, P., de Belleroche, J., de Figueiredo, P., de Figueiredo, R. C., de la Fuente, J., De Martino, L., De Matteis, A., De Meyer, G. R., De Milito, A., De Santi, M., de Souza, W., De Tata, V., De Zio, D., Debnath, J., Dechant, R., Decuypere, J. P., Deegan, S., Dehay, B., Del Bello, B., Del Re, D. P., Delage-Mourroux, R., Delbridge, L. M., Deldicque, L., Delorme-Axford, E., Deng, Y., Dengjel, J., Denizot, M., Dent, P., Der, C. J., Deretic, V., Derrien, B., Deutsch, E., Devarenne, T. P., Devenish, R. J., Di Bartolomeo, S., Di Daniele, N., Di Domenico, F., Di Nardo, A., Di Paola, S., Di Pietro, A., Di Renzo, L., DiAntonio, A., Diaz-Araya, G., Diaz-Laviada, I., Diaz-Meco, M. T., Diaz-Nido, J., Dickey, C. A., Dickson, R. C., Diederich, M., Digard, P., Dikic, I., Dinesh- Kumar, S. P., Ding, C., Ding, W. X., Ding, Z., Dini, L., Distler, J. H., Diwan, A., 20
21 Djavaheri-Mergny, M., Dmytruk, K., Dobson, R. C., Doetsch, V., Dokladny, K., Dokudovskaya, S., Donadelli, M., Dong, X. C., Dong, X., Dong, Z., Donohue, T. M., Jr., Doran, K. S., D'Orazi, G., Dorn, G. W., 2nd, Dosenko, V., Dridi, S., Drucker, L., Du, J., Du, L. L., Du, L., du Toit, A., Dua, P., Duan, L., Duann, P., Dubey, V. K., Duchen, M. R., Duchosal, M. A., Duez, H., Dugail, I., Dumit, V. I., Duncan, M. C., Dunlop, E. A., Dunn, W. A., Jr., Dupont, N., Dupuis, L., Duran, R. V., Durcan, T. M., Duvezin-Caubet, S., Duvvuri, U., Eapen, V., Ebrahimi-Fakhari, D., Echard, A., Eckhart, L., Edelstein, C. L., Edinger, A. L., Eichinger, L., Eisenberg, T., Eisenberg-Lerner, A., Eissa, N. T., El- Deiry, W. S., El-Khoury, V., Elazar, Z., Eldar-Finkelman, H., Elliott, C. J., Emanuele, E., Emmenegger, U., Engedal, N., Engelbrecht, A. M., Engelender, S., Enserink, J. M., Erdmann, R., Erenpreisa, J., Eri, R., Eriksen, J. L., Erman, A., Escalante, R., Eskelinen, E. L., Espert, L., Esteban-Martinez, L., Evans, T. J., Fabri, M., Fabrias, G., Fabrizi, C., Facchiano, A., Faergeman, N. J., Faggioni, A., Fairlie, W. D., Fan, C., Fan, D., Fan, J., Fang, S., Fanto, M., Fanzani, A., Farkas, T., Faure, M., Favier, F. B., Fearnhead, H., Federici, M., Fei, E., Felizardo, T. C., Feng, H., Feng, Y., Feng, Y., Ferguson, T. A., Fernandez, A. F., Fernandez-Barrena, M. G., Fernandez-Checa, J. C., Fernandez-Lopez, A., Fernandez-Zapico, M. E., Feron, O., Ferraro, E., Ferreira-Halder, C. V., Fesus, L., Feuer, R., Fiesel, F. C., Filippi-Chiela, E. C., Filomeni, G., Fimia, G. M., Fingert, J. H., Finkbeiner, S., Finkel, T., Fiorito, F., Fisher, P. B., Flajolet, M., Flamigni, F., Florey, O., Florio, S., Floto, R. A., Folini, M., Follo, C., Fon, E. A., Fornai, F., Fortunato, F., Fraldi, A., Franco, R., Francois, A., Francois, A., Frankel, L. B., Fraser, I. D., Frey, N., Freyssenet, D. G., Frezza, C., Friedman, S. L., Frigo, D. E., Fu, D., Fuentes, J. M., Fueyo, J., Fujitani, Y., Fujiwara, Y., Fujiya, M., Fukuda, M., Fulda, S., Fusco, C., Gabryel, B., Gaestel, M., Gailly, P., Gajewska, M., Galadari, S., Galili, G., Galindo, I., Galindo, M. F., Galliciotti, G., Galluzzi, L., Galluzzi, L., Galy, V., Gammoh, N., Gandy, S., Ganesan, A. K., Ganesan, S., Ganley, I. G., Gannage, M., Gao, F. B., Gao, F., Gao, J. X., Garcia Nannig, L., Garcia Vescovi, E., Garcia-Macia, M., Garcia-Ruiz, C., Garg, A. D., Garg, P. K., Gargini, R., Gassen, N. C., Gatica, D., Gatti, E., Gavard, J., Gavathiotis, E., Ge, L., Ge, P., Ge, S., Gean, P. W., Gelmetti, V., Genazzani, A. A., Geng, J., Genschik, P., Gerner, L., Gestwicki, J. E., Gewirtz, D. A., Ghavami, S., Ghigo, E., Ghosh, D., Giammarioli, A. M., Giampieri, F., Giampietri, C., Giatromanolaki, A., Gibbings, D. J., Gibellini, L., Gibson, S. B., Ginet, V., Giordano, A., Giorgini, F., Giovannetti, E., Girardin, S. E., Gispert, S., Giuliano, S., Gladson, C. L., Glavic, A., Gleave, M., Godefroy, N., Gogal, R. M., Jr., Gokulan, K., Goldman, G. H., Goletti, D., Goligorsky, M. S., Gomes, A. V., Gomes, L. C., Gomez, H., Gomez-Manzano, C., Gomez-Sanchez, R., Goncalves, D. A., Goncu, E., Gong, Q., Gongora, C., Gonzalez, C. B., Gonzalez- Alegre, P., Gonzalez-Cabo, P., Gonzalez-Polo, R. A., Goping, I. S., Gorbea, C., Gorbunov, N. V., Goring, D. R., Gorman, A. M., Gorski, S. M., Goruppi, S., Goto- Yamada, S., Gotor, C., Gottlieb, R. A., Gozes, I., Gozuacik, D., Graba, Y., Graef, M., Granato, G. E., Grant, G. D., Grant, S., Gravina, G. L., Green, D. R., Greenhough, A., Greenwood, M. T., Grimaldi, B., Gros, F., Grose, C., Groulx, J. F., Gruber, F., Grumati, P., Grune, T., Guan, J. L., Guan, K. L., Guerra, B., Guillen, C., Gulshan, K., Gunst, J., Guo, C., Guo, L., Guo, M., Guo, W., Guo, X. G., Gust, A. A., Gustafsson, A. B., Gutierrez, E., Gutierrez, M. G., Gwak, H. S., Haas, A., Haber, J. E., Hadano, S., Hagedorn, M., Hahn, D. R., Halayko, A. J., Hamacher-Brady, A., Hamada, K., Hamai, A., Hamann, A., Hamasaki, M., Hamer, I., Hamid, Q., Hammond, E. M., Han, F., Han, W., Handa, J. T., Hanover, J. A., Hansen, M., Harada, M., Harhaji-Trajkovic, L., Harper, J. W., Harrath, A. H., Harris, A. L., Harris, J., Hasler, U., Hasselblatt, P., Hasui, K., Hawley, R. G., Hawley, T. S., He, C., He, C. Y., He, F., He, G., He, R. R., He, X. H., He, Y. W., He, Y. Y., Heath, J. K., Hebert, M. J., Heinzen, R. A., Helgason, G. V., Hensel, M., Henske, E. P., Her, C., Herman, P. K., Hernandez, A., Hernandez, C., Hernandez- 21
22 Tiedra, S., Hetz, C., Hiesinger, P. R., Higaki, K., Hilfiker, S., Hill, B. G., Hill, J. A., Hill, W. D., Hino, K., Hofius, D., Hofman, P., Hoglinger, G. U., Hohfeld, J., Holz, M. K., Hong, Y., Hood, D. A., Hoozemans, J. J., Hoppe, T., Hsu, C., Hsu, C. Y., Hsu, L. C., Hu, D., Hu, G., Hu, H. M., Hu, H., Hu, M. C., Hu, Y. C., Hu, Z. W., Hua, F., Hua, Y., Huang, C., Huang, H. L., Huang, K. H., Huang, K. Y., Huang, S., Huang, S., Huang, W. P., Huang, Y. R., Huang, Y., Huang, Y., Huber, T. B., Huebbe, P., Huh, W. K., Hulmi, J. J., Hur, G. M., Hurley, J. H., Husak, Z., Hussain, S. N., Hussain, S., Hwang, J. J., Hwang, S., Hwang, T. I., Ichihara, A., Imai, Y., Imbriano, C., Inomata, M., Into, T., Iovane, V., Iovanna, J. L., Iozzo, R. V., Ip, N. Y., Irazoqui, J. E., Iribarren, P., Isaka, Y., Isakovic, A. J., Ischiropoulos, H., Isenberg, J. S., Ishaq, M., Ishida, H., Ishii, I., Ishmael, J. E., Isidoro, C., Isobe, K., Isono, E., Issazadeh-Navikas, S., Itahana, K., Itakura, E., Ivanov, A. I., Iyer, A. K., Izquierdo, J. M., Izumi, Y., Izzo, V., Jaattela, M., Jaber, N., Jackson, D. J., Jackson, W. T., Jacob, T. G., Jacques, T. S., Jagannath, C., Jain, A., Jana, N. R., Jang, B. K., Jani, A., Janji, B., Jannig, P. R., Jansson, P. J., Jean, S., Jendrach, M., Jeon, J. H., Jessen, N., Jeung, E. B., Jia, K., Jia, L., Jiang, H., Jiang, H., Jiang, L., Jiang, T., Jiang, X., Jiang, X., Jiang, X., Jiang, Y., Jiang, Y., Jimenez, A., Jin, C., Jin, H., Jin, L., Jin, M., Jin, S., Jinwal, U. K., Jo, E. K., Johansen, T., Johnson, D. E., Johnson, G. V., Johnson, J. D., Jonasch, E., Jones, C., Joosten, L. A., Jordan, J., Joseph, A. M., Joseph, B., Joubert, A. M., Ju, D., Ju, J., Juan, H. F., Juenemann, K., Juhasz, G., Jung, H. S., Jung, J. U., Jung, Y. K., Jungbluth, H., Justice, M. J., Jutten, B., Kaakoush, N. O., Kaarniranta, K., Kaasik, A., Kabuta, T., Kaeffer, B., Kagedal, K., Kahana, A., Kajimura, S., Kakhlon, O., Kalia, M., Kalvakolanu, D. V., Kamada, Y., Kambas, K., Kaminskyy, V. O., Kampinga, H. H., Kandouz, M., Kang, C., Kang, R., Kang, T. C., Kanki, T., Kanneganti, T. D., Kanno, H., Kanthasamy, A. G., Kantorow, M., Kaparakis-Liaskos, M., Kapuy, O., Karantza, V., Karim, M. R., Karmakar, P., Kaser, A., Kaushik, S., Kawula, T., Kaynar, A. M., Ke, P. Y., Ke, Z. J., Kehrl, J. H., Keller, K. E., Kemper, J. K., Kenworthy, A. K., Kepp, O., Kern, A., Kesari, S., Kessel, D., Ketteler, R., Kettelhut Ido, C., Khambu, B., Khan, M. M., Khandelwal, V. K., Khare, S., Kiang, J. G., Kiger, A. A., Kihara, A., Kim, A. L., Kim, C. H., Kim, D. R., Kim, D. H., Kim, E. K., Kim, H. Y., Kim, H. R., Kim, J. S., Kim, J. H., Kim, J. C., Kim, J. H., Kim, K. W., Kim, M. D., Kim, M. M., Kim, P. K., Kim, S. W., Kim, S. Y., Kim, Y. S., Kim, Y., Kimchi, A., Kimmelman, A. C., Kimura, T., King, J. S., Kirkegaard, K., Kirkin, V., Kirshenbaum, L. A., Kishi, S., Kitajima, Y., Kitamoto, K., Kitaoka, Y., Kitazato, K., Kley, R. A., Klimecki, W. T., Klinkenberg, M., Klucken, J., Knaevelsrud, H., Knecht, E., Knuppertz, L., Ko, J. L., Kobayashi, S., Koch, J. C., Koechlin-Ramonatxo, C., Koenig, U., Koh, Y. H., Kohler, K., Kohlwein, S. D., Koike, M., Komatsu, M., Kominami, E., Kong, D., Kong, H. J., Konstantakou, E. G., Kopp, B. T., Korcsmaros, T., Korhonen, L., Korolchuk, V. I., Koshkina, N. V., Kou, Y., Koukourakis, M. I., Koumenis, C., Kovacs, A. L., Kovacs, T., Kovacs, W. J., Koya, D., Kraft, C., Krainc, D., Kramer, H., Kravic-Stevovic, T., Krek, W., Kretz-Remy, C., Krick, R., Krishnamurthy, M., Kriston-Vizi, J., Kroemer, G., Kruer, M. C., Kruger, R., Ktistakis, N. T., Kuchitsu, K., Kuhn, C., Kumar, A. P., Kumar, A., Kumar, A., Kumar, D., Kumar, D., Kumar, R., Kumar, S., Kundu, M., Kung, H. J., Kuno, A., Kuo, S. H., Kuret, J., Kurz, T., Kwok, T., Kwon, T. K., Kwon, Y. T., Kyrmizi, I., La Spada, A. R., Lafont, F., Lahm, T., Lakkaraju, A., Lam, T., Lamark, T., Lancel, S., Landowski, T. H., Lane, D. J., Lane, J. D., Lanzi, C., Lapaquette, P., Lapierre, L. R., Laporte, J., Laukkarinen, J., Laurie, G. W., Lavandero, S., Lavie, L., LaVoie, M. J., Law, B. Y., Law, H. K., Law, K. B., Layfield, R., Lazo, P. A., Le Cam, L., Le Roch, K. G., Le Stunff, H., Leardkamolkarn, V., Lecuit, M., Lee, B. H., Lee, C. H., Lee, E. F., Lee, G. M., Lee, H. J., Lee, H., Lee, J. K., Lee, J., Lee, J. H., Lee, J. H., Lee, M., Lee, M. S., Lee, P. J., Lee, S. W., Lee, S. J., Lee, S. J., Lee, S. Y., Lee, S. H., Lee, S. S., Lee, S. J., Lee, S., Lee, Y. R., Lee, Y. J., Lee, Y. H., Leeuwenburgh, C., Lefort, S., Legouis, R., Lei, J., Lei, Q. Y., Leib, 22
23 D. A., Leibowitz, G., Lekli, I., Lemaire, S. D., Lemasters, J. J., Lemberg, M. K., Lemoine, A., Leng, S., Lenz, G., Lenzi, P., Lerman, L. O., Lettieri Barbato, D., Leu, J. I., Leung, H. Y., Levine, B., Lewis, P. A., Lezoualc'h, F., Li, C., Li, F., Li, F. J., Li, J., Li, K., Li, L., Li, M., Li, M., Li, Q., Li, R., Li, S., Li, W., Li, W., Li, X., Li, Y., Lian, J., Liang, C., Liang, Q., Liao, Y., Liberal, J., Liberski, P. P., Lie, P., Lieberman, A. P., Lim, H. J., Lim, K. L., Lim, K., Lima, R. T., Lin, C. S., Lin, C. F., Lin, F., Lin, F., Lin, F. C., Lin, K., Lin, K. H., Lin, P. H., Lin, T., Lin, W. W., Lin, Y. S., Lin, Y., Linden, R., Lindholm, D., Lindqvist, L. M., Lingor, P., Linkermann, A., Liotta, L. A., Lipinski, M. M., Lira, V. A., Lisanti, M. P., Liton, P. B., Liu, B., Liu, C., Liu, C. F., Liu, F., Liu, H. J., Liu, J., Liu, J. J., Liu, J. L., Liu, K., Liu, L., Liu, L., Liu, Q., Liu, R. Y., Liu, S., Liu, S., Liu, W., Liu, X. D., Liu, X., Liu, X. H., Liu, X., Liu, X., Liu, X., Liu, Y., Liu, Y., Liu, Z., Liu, Z., Liuzzi, J. P., Lizard, G., Ljujic, M., Lodhi, I. J., Logue, S. E., Lokeshwar, B. L., Long, Y. C., Lonial, S., Loos, B., Lopez-Otin, C., Lopez-Vicario, C., Lorente, M., Lorenzi, P. L., Lorincz, P., Los, M., Lotze, M. T., Lovat, P. E., Lu, B., Lu, B., Lu, J., Lu, Q., Lu, S. M., Lu, S., Lu, Y., Luciano, F., Luckhart, S., Lucocq, J. M., Ludovico, P., Lugea, A., Lukacs, N. W., Lum, J. J., Lund, A. H., Luo, H., Luo, J., Luo, S., Luparello, C., Lyons, T., Ma, J., Ma, Y., Ma, Y., Ma, Z., Machado, J., Machado-Santelli, G. M., Macian, F., MacIntosh, G. C., MacKeigan, J. P., Macleod, K. F., MacMicking, J. D., MacMillan-Crow, L. A., Madeo, F., Madesh, M., Madrigal-Matute, J., Maeda, A., Maeda, T., Maegawa, G., Maellaro, E., Maes, H., Magarinos, M., Maiese, K., Maiti, T. K., Maiuri, L., Maiuri, M. C., Maki, C. G., Malli, R., Malorni, W., Maloyan, A., Mami- Chouaib, F., Man, N., Mancias, J. D., Mandelkow, E. M., Mandell, M. A., Manfredi, A. A., Manie, S. N., Manzoni, C., Mao, K., Mao, Z., Mao, Z. W., Marambaud, P., Marconi, A. M., Marelja, Z., Marfe, G., Margeta, M., Margittai, E., Mari, M., Mariani, F. V., Marin, C., Marinelli, S., Marino, G., Markovic, I., Marquez, R., Martelli, A. M., Martens, S., Martin, K. R., Martin, S. J., Martin, S., Martin-Acebes, M. A., Martin-Sanz, P., Martinand-Mari, C., Martinet, W., Martinez, J., Martinez-Lopez, N., Martinez- Outschoorn, U., Martinez-Velazquez, M., Martinez-Vicente, M., Martins, W. K., Mashima, H., Mastrianni, J. A., Matarese, G., Matarrese, P., Mateo, R., Matoba, S., Matsumoto, N., Matsushita, T., Matsuura, A., Matsuzawa, T., Mattson, M. P., Matus, S., Maugeri, N., Mauvezin, C., Mayer, A., Maysinger, D., Mazzolini, G. D., McBrayer, M. K., McCall, K., McCormick, C., McInerney, G. M., McIver, S. C., McKenna, S., McMahon, J. J., McNeish, I. A., Mechta-Grigoriou, F., Medema, J. P., Medina, D. L., Megyeri, K., Mehrpour, M., Mehta, J. L., Mei, Y., Meier, U. C., Meijer, A. J., Melendez, A., Melino, G., Melino, S., de Melo, E. J., Mena, M. A., Meneghini, M. D., Menendez, J. A., Menezes, R., Meng, L., Meng, L. H., Meng, S., Menghini, R., Menko, A. S., Menna- Barreto, R. F., Menon, M. B., Meraz-Rios, M. A., Merla, G., Merlini, L., Merlot, A. M., Meryk, A., Meschini, S., Meyer, J. N., Mi, M. T., Miao, C. Y., Micale, L., Michaeli, S., Michiels, C., Migliaccio, A. R., Mihailidou, A. S., Mijaljica, D., Mikoshiba, K., Milan, E., Miller-Fleming, L., Mills, G. B., Mills, I. G., Minakaki, G., Minassian, B. A., Ming, X. F., Minibayeva, F., Minina, E. A., Mintern, J. D., Minucci, S., Miranda-Vizuete, A., Mitchell, C. H., Miyamoto, S., Miyazawa, K., Mizushima, N., Mnich, K., Mograbi, B., Mohseni, S., Moita, L. F., Molinari, M., Molinari, M., Moller, A. B., Mollereau, B., Mollinedo, F., Mongillo, M., Monick, M. M., Montagnaro, S., Montell, C., Moore, D. J., Moore, M. N., Mora-Rodriguez, R., Moreira, P. I., Morel, E., Morelli, M. B., Moreno, S., Morgan, M. J., Moris, A., Moriyasu, Y., Morrison, J. L., Morrison, L. A., Morselli, E., Moscat, J., Moseley, P. L., Mostowy, S., Motori, E., Mottet, D., Mottram, J. C., Moussa, C. E., Mpakou, V. E., Mukhtar, H., Mulcahy Levy, J. M., Muller, S., Munoz-Moreno, R., Munoz-Pinedo, C., Munz, C., Murphy, M. E., Murray, J. T., Murthy, A., Mysorekar, I. U., Nabi, I. R., Nabissi, M., Nader, G. A., Nagahara, Y., Nagai, Y., Nagata, K., Nagelkerke, A., Nagy, P., Naidu, S. R., Nair, S., Nakano, H., Nakatogawa, H., 23
24 Nanjundan, M., Napolitano, G., Naqvi, N. I., Nardacci, R., Narendra, D. P., Narita, M., Nascimbeni, A. C., Natarajan, R., Navegantes, L. C., Nawrocki, S. T., Nazarko, T. Y., Nazarko, V. Y., Neill, T., Neri, L. M., Netea, M. G., Netea-Maier, R. T., Neves, B. M., Ney, P. A., Nezis, I. P., Nguyen, H. T., Nguyen, H. P., Nicot, A. S., Nilsen, H., Nilsson, P., Nishimura, M., Nishino, I., Niso-Santano, M., Niu, H., Nixon, R. A., Njar, V. C., Noda, T., Noegel, A. A., Nolte, E. M., Norberg, E., Norga, K. K., Noureini, S. K., Notomi, S., Notterpek, L., Nowikovsky, K., Nukina, N., Nurnberger, T., O'Donnell, V. B., O'Donovan, T., O'Dwyer, P. J., Oehme, I., Oeste, C. L., Ogawa, M., Ogretmen, B., Ogura, Y., Oh, Y. J., Ohmuraya, M., Ohshima, T., Ojha, R., Okamoto, K., Okazaki, T., Oliver, F. J., Ollinger, K., Olsson, S., Orban, D. P., Ordonez, P., Orhon, I., Orosz, L., O'Rourke, E. J., Orozco, H., Ortega, A. L., Ortona, E., Osellame, L. D., Oshima, J., Oshima, S., Osiewacz, H. D., Otomo, T., Otsu, K., Ou, J. H., Outeiro, T. F., Ouyang, D. Y., Ouyang, H., Overholtzer, M., Ozbun, M. A., Ozdinler, P. H., Ozpolat, B., Pacelli, C., Paganetti, P., Page, G., Pages, G., Pagnini, U., Pajak, B., Pak, S. C., Pakos-Zebrucka, K., Pakpour, N., Palkova, Z., Palladino, F., Pallauf, K., Pallet, N., Palmieri, M., Paludan, S. R., Palumbo, C., Palumbo, S., Pampliega, O., Pan, H., Pan, W., Panaretakis, T., Pandey, A., Pantazopoulou, A., Papackova, Z., Papademetrio, D. L., Papassideri, I., Papini, A., Parajuli, N., Pardo, J., Parekh, V. V., Parenti, G., Park, J. I., Park, J., Park, O. K., Parker, R., Parlato, R., Parys, J. B., Parzych, K. R., Pasquet, J. M., Pasquier, B., Pasumarthi, K. B., Patschan, D., Patterson, C., Pattingre, S., Pattison, S., Pause, A., Pavenstadt, H., Pavone, F., Pedrozo, Z., Pena, F. J., Penalva, M. A., Pende, M., Peng, J., Penna, F., Penninger, J. M., Pensalfini, A., Pepe, S., Pereira, G. J., Pereira, P. C., Perez-de la Cruz, V., Perez-Perez, M. E., Perez-Rodriguez, D., Perez-Sala, D., Perier, C., Perl, A., Perlmutter, D. H., Perrotta, I., Pervaiz, S., Pesonen, M., Pessin, J. E., Peters, G. J., Petersen, M., Petrache, I., Petrof, B. J., Petrovski, G., Phang, J. M., Piacentini, M., Pierdominici, M., Pierre, P., Pierrefite-Carle, V., Pietrocola, F., Pimentel-Muinos, F. X., Pinar, M., Pineda, B., Pinkas-Kramarski, R., Pinti, M., Pinton, P., Piperdi, B., Piret, J. M., Platanias, L. C., Platta, H. W., Plowey, E. D., Poggeler, S., Poirot, M., Polcic, P., Poletti, A., Poon, A. H., Popelka, H., Popova, B., Poprawa, I., Poulose, S. M., Poulton, J., Powers, S. K., Powers, T., Pozuelo-Rubio, M., Prak, K., Prange, R., Prescott, M., Priault, M., Prince, S., Proia, R. L., Proikas-Cezanne, T., Prokisch, H., Promponas, V. J., Przyklenk, K., Puertollano, R., Pugazhenthi, S., Puglielli, L., Pujol, A., Puyal, J., Pyeon, D., Qi, X., Qian, W. B., Qin, Z. H., Qiu, Y., Qu, Z., Quadrilatero, J., Quinn, F., Raben, N., Rabinowich, H., Radogna, F., Ragusa, M. J., Rahmani, M., Raina, K., Ramanadham, S., Ramesh, R., Rami, A., Randall-Demllo, S., Randow, F., Rao, H., Rao, V. A., Rasmussen, B. B., Rasse, T. M., Ratovitski, E. A., Rautou, P. E., Ray, S. K., Razani, B., Reed, B. H., Reggiori, F., Rehm, M., Reichert, A. S., Rein, T., Reiner, D. J., Reits, E., Ren, J., Ren, X., Renna, M., Reusch, J. E., Revuelta, J. L., Reyes, L., Rezaie, A. R., Richards, R. I., Richardson, D. R., Richetta, C., Riehle, M. A., Rihn, B. H., Rikihisa, Y., Riley, B. E., Rimbach, G., Rippo, M. R., Ritis, K., Rizzi, F., Rizzo, E., Roach, P. J., Robbins, J., Roberge, M., Roca, G., Roccheri, M. C., Rocha, S., Rodrigues, C. M., Rodriguez, C. I., de Cordoba, S. R., Rodriguez-Muela, N., Roelofs, J., Rogov, V. V., Rohn, T. T., Rohrer, B., Romanelli, D., Romani, L., Romano, P. S., Roncero, M. I., Rosa, J. L., Rosello, A., Rosen, K. V., Rosenstiel, P., Rost-Roszkowska, M., Roth, K. A., Roue, G., Rouis, M., Rouschop, K. M., Ruan, D. T., Ruano, D., Rubinsztein, D. C., Rucker, E. B., 3rd, Rudich, A., Rudolf, E., Rudolf, R., Ruegg, M. A., Ruiz-Roldan, C., Ruparelia, A. A., Rusmini, P., Russ, D. W., Russo, G. L., Russo, G., Russo, R., Rusten, T. E., Ryabovol, V., Ryan, K. M., Ryter, S. W., Sabatini, D. M., Sacher, M., Sachse, C., Sack, M. N., Sadoshima, J., Saftig, P., Sagi-Eisenberg, R., Sahni, S., Saikumar, P., Saito, T., Saitoh, T., Sakakura, K., Sakoh-Nakatogawa, M., Sakuraba, Y., Salazar-Roa, M., Salomoni, P., Saluja, A. K., Salvaterra, P. M., Salvioli, R., Samali, A., Sanchez, A. M., 24
25 Sanchez-Alcazar, J. A., Sanchez-Prieto, R., Sandri, M., Sanjuan, M. A., Santaguida, S., Santambrogio, L., Santoni, G., Dos Santos, C. N., Saran, S., Sardiello, M., Sargent, G., Sarkar, P., Sarkar, S., Sarrias, M. R., Sarwal, M. M., Sasakawa, C., Sasaki, M., Sass, M., Sato, K., Sato, M., Satriano, J., Savaraj, N., Saveljeva, S., Schaefer, L., Schaible, U. E., Scharl, M., Schatzl, H. M., Schekman, R., Scheper, W., Schiavi, A., Schipper, H. M., Schmeisser, H., Schmidt, J., Schmitz, I., Schneider, B. E., Schneider, E. M., Schneider, J. L., Schon, E. A., Schonenberger, M. J., Schonthal, A. H., Schorderet, D. F., Schroder, B., Schuck, S., Schulze, R. J., Schwarten, M., Schwarz, T. L., Sciarretta, S., Scotto, K., Scovassi, A. I., Screaton, R. A., Screen, M., Seca, H., Sedej, S., Segatori, L., Segev, N., Seglen, P. O., Segui-Simarro, J. M., Segura-Aguilar, J., Seki, E., Sell, C., Seiliez, I., Semenkovich, C. F., Semenza, G. L., Sen, U., Serra, A. L., Serrano-Puebla, A., Sesaki, H., Setoguchi, T., Settembre, C., Shacka, J. J., Shajahan-Haq, A. N., Shapiro, I. M., Sharma, S., She, H., Shen, C. K., Shen, C. C., Shen, H. M., Shen, S., Shen, W., Sheng, R., Sheng, X., Sheng, Z. H., Shepherd, T. G., Shi, J., Shi, Q., Shi, Q., Shi, Y., Shibutani, S., Shibuya, K., Shidoji, Y., Shieh, J. J., Shih, C. M., Shimada, Y., Shimizu, S., Shin, D. W., Shinohara, M. L., Shintani, M., Shintani, T., Shioi, T., Shirabe, K., Shiri-Sverdlov, R., Shirihai, O., Shore, G. C., Shu, C. W., Shukla, D., Sibirny, A. A., Sica, V., Sigurdson, C. J., Sigurdsson, E. M., Sijwali, P. S., Sikorska, B., Silveira, W. A., Silvente-Poirot, S., Silverman, G. A., Simak, J., Simmet, T., Simon, A. K., Simon, H. U., Simone, C., Simons, M., Simonsen, A., Singh, R., Singh, S. V., Singh, S. K., Sinha, D., Sinha, S., Sinicrope, F. A., Sirko, A., Sirohi, K., Sishi, B. J., Sittler, A., Siu, P. M., Sivridis, E., Skwarska, A., Slack, R., Slaninova, I., Slavov, N., Smaili, S. S., Smalley, K. S., Smith, D. R., Soenen, S. J., Soleimanpour, S. A., Solhaug, A., Somasundaram, K., Son, J. H., Sonawane, A., Song, C., Song, F., Song, H. K., Song, J. X., Song, W., Soo, K. Y., Sood, A. K., Soong, T. W., Soontornniyomkij, V., Sorice, M., Sotgia, F., Soto-Pantoja, D. R., Sotthibundhu, A., Sousa, M. J., Spaink, H. P., Span, P. N., Spang, A., Sparks, J. D., Speck, P. G., Spector, S. A., Spies, C. D., Springer, W., Clair, D. S., Stacchiotti, A., Staels, B., Stang, M. T., Starczynowski, D. T., Starokadomskyy, P., Steegborn, C., Steele, J. W., Stefanis, L., Steffan, J., Stellrecht, C. M., Stenmark, H., Stepkowski, T. M., Stern, S. T., Stevens, C., Stockwell, B. R., Stoka, V., Storchova, Z., Stork, B., Stratoulias, V., Stravopodis, D. J., Strnad, P., Strohecker, A. M., Strom, A. L., Stromhaug, P., Stulik, J., Su, Y. X., Su, Z., Subauste, C. S., Subramaniam, S., Sue, C. M., Suh, S. W., Sui, X., Sukseree, S., Sulzer, D., Sun, F. L., Sun, J., Sun, J., Sun, S. Y., Sun, Y., Sun, Y., Sun, Y., Sundaramoorthy, V., Sung, J., Suzuki, H., Suzuki, K., Suzuki, N., Suzuki, T., Suzuki, Y. J., Swanson, M. S., Swanton, C., Sward, K., Swarup, G., Sweeney, S. T., Sylvester, P. W., Szatmari, Z., Szegezdi, E., Szlosarek, P. W., Taegtmeyer, H., Tafani, M., Taillebourg, E., Tait, S. W., Takacs-Vellai, K., Takahashi, Y., Takats, S., Takemura, G., Takigawa, N., Talbot, N. J., Tamagno, E., Tamburini, J., Tan, C. P., Tan, L., Tan, M. L., Tan, M., Tan, Y. J., Tanaka, K., Tanaka, M., Tang, D., Tang, D., Tang, G., Tanida, I., Tanji, K., Tannous, B. A., Tapia, J. A., Tasset-Cuevas, I., Tatar, M., Tavassoly, I., Tavernarakis, N., Taylor, A., Taylor, G. S., Taylor, G. A., Taylor, J. P., Taylor, M. J., Tchetina, E. V., Tee, A. R., Teixeira-Clerc, F., Telang, S., Tencomnao, T., Teng, B. B., Teng, R. J., Terro, F., Tettamanti, G., Theiss, A. L., Theron, A. E., Thomas, K. J., Thome, M. P., Thomes, P. G., Thorburn, A., Thorner, J., Thum, T., Thumm, M., Thurston, T. L., Tian, L., Till, A., Ting, J. P., Titorenko, V. I., Toker, L., Toldo, S., Tooze, S. A., Topisirovic, I., Torgersen, M. L., Torosantucci, L., Torriglia, A., Torrisi, M. R., Tournier, C., Towns, R., Trajkovic, V., Travassos, L. H., Triola, G., Tripathi, D. N., Trisciuoglio, D., Troncoso, R., Trougakos, I. P., Truttmann, A. C., Tsai, K. J., Tschan, M. P., Tseng, Y. H., Tsukuba, T., Tsung, A., Tsvetkov, A. S., Tu, S., Tuan, H. Y., Tucci, M., Tumbarello, D. A., Turk, B., Turk, V., Turner, R. F., Tveita, A. A., Tyagi, S. C., Ubukata, M., Uchiyama, Y., Udelnow, A., Ueno, T., Umekawa, M., Umemiya-Shirafuji, 25
26 R., Underwood, B. R., Ungermann, C., Ureshino, R. P., Ushioda, R., Uversky, V. N., Uzcategui, N. L., Vaccari, T., Vaccaro, M. I., Vachova, L., Vakifahmetoglu-Norberg, H., Valdor, R., Valente, E. M., Vallette, F., Valverde, A. M., Van den Berghe, G., Van Den Bosch, L., van den Brink, G. R., van der Goot, F. G., van der Klei, I. J., van der Laan, L. J., van Doorn, W. G., van Egmond, M., van Golen, K. L., Van Kaer, L., van Lookeren Campagne, M., Vandenabeele, P., Vandenberghe, W., Vanhorebeek, I., Varela-Nieto, I., Vasconcelos, M. H., Vasko, R., Vavvas, D. G., Vega-Naredo, I., Velasco, G., Velentzas, A. D., Velentzas, P. D., Vellai, T., Vellenga, E., Vendelbo, M. H., Venkatachalam, K., Ventura, N., Ventura, S., Veras, P. S., Verdier, M., Vertessy, B. G., Viale, A., Vidal, M., Vieira, H. L., Vierstra, R. D., Vigneswaran, N., Vij, N., Vila, M., Villar, M., Villar, V. H., Villarroya, J., Vindis, C., Viola, G., Viscomi, M. T., Vitale, G., Vogl, D. T., Voitsekhovskaja, O. V., von Haefen, C., von Schwarzenberg, K., Voth, D. E., Vouret- Craviari, V., Vuori, K., Vyas, J. M., Waeber, C., Walker, C. L., Walker, M. J., Walter, J., Wan, L., Wan, X., Wang, B., Wang, C., Wang, C. Y., Wang, C., Wang, C., Wang, C., Wang, D., Wang, F., Wang, F., Wang, G., Wang, H. J., Wang, H., Wang, H. G., Wang, H., Wang, H. D., Wang, J., Wang, J., Wang, M., Wang, M. Q., Wang, P. Y., Wang, P., Wang, R. C., Wang, S., Wang, T. F., Wang, X., Wang, X. J., Wang, X. W., Wang, X., Wang, X., Wang, Y., Wang, Y., Wang, Y., Wang, Y. J., Wang, Y., Wang, Y., Wang, Y. T., Wang, Y., Wang, Z. N., Wappner, P., Ward, C., Ward, D. M., Warnes, G., Watada, H., Watanabe, Y., Watase, K., Weaver, T. E., Weekes, C. D., Wei, J., Weide, T., Weihl, C. C., Weindl, G., Weis, S. N., Wen, L., Wen, X., Wen, Y., Westermann, B., Weyand, C. M., White, A. R., White, E., Whitton, J. L., Whitworth, A. J., Wiels, J., Wild, F., Wildenberg, M. E., Wileman, T., Wilkinson, D. S., Wilkinson, S., Willbold, D., Williams, C., Williams, K., Williamson, P. R., Winklhofer, K. F., Witkin, S. S., Wohlgemuth, S. E., Wollert, T., Wolvetang, E. J., Wong, E., Wong, G. W., Wong, R. W., Wong, V. K., Woodcock, E. A., Wright, K. L., Wu, C., Wu, D., Wu, G. S., Wu, J., Wu, J., Wu, M., Wu, M., Wu, S., Wu, W. K., Wu, Y., Wu, Z., Xavier, C. P., Xavier, R. J., Xia, G. X., Xia, T., Xia, W., Xia, Y., Xiao, H., Xiao, J., Xiao, S., Xiao, W., Xie, C. M., Xie, Z., Xie, Z., Xilouri, M., Xiong, Y., Xu, C., Xu, C., Xu, F., Xu, H., Xu, H., Xu, J., Xu, J., Xu, J., Xu, L., Xu, X., Xu, Y., Xu, Y., Xu, Z. X., Xu, Z., Xue, Y., Yamada, T., Yamamoto, A., Yamanaka, K., Yamashina, S., Yamashiro, S., Yan, B., Yan, B., Yan, X., Yan, Z., Yanagi, Y., Yang, D. S., Yang, J. M., Yang, L., Yang, M., Yang, P. M., Yang, P., Yang, Q., Yang, W., Yang, W. Y., Yang, X., Yang, Y., Yang, Y., Yang, Z., Yang, Z., Yao, M. C., Yao, P. J., Yao, X., Yao, Z., Yao, Z., Yasui, L. S., Ye, M., Yedvobnick, B., Yeganeh, B., Yeh, E. S., Yeyati, P. L., Yi, F., Yi, L., Yin, X. M., Yip, C. K., Yoo, Y. M., Yoo, Y. H., Yoon, S. Y., Yoshida, K., Yoshimori, T., Young, K. H., Yu, H., Yu, J. J., Yu, J. T., Yu, J., Yu, L., Yu, W. H., Yu, X. F., Yu, Z., Yuan, J., Yuan, Z. M., Yue, B. Y., Yue, J., Yue, Z., Zacks, D. N., Zacksenhaus, E., Zaffaroni, N., Zaglia, T., Zakeri, Z., Zecchini, V., Zeng, J., Zeng, M., Zeng, Q., Zervos, A. S., Zhang, D. D., Zhang, F., Zhang, G., Zhang, G. C., Zhang, H., Zhang, H., Zhang, H., Zhang, H., Zhang, J., Zhang, J., Zhang, J., Zhang, J., Zhang, J. P., Zhang, L., Zhang, L., Zhang, L., Zhang, L., Zhang, M. Y., Zhang, X., Zhang, X. D., Zhang, Y., Zhang, Y., Zhang, Y., Zhang, Y., Zhang, Y., Zhao, M., Zhao, W. L., Zhao, X., Zhao, Y. G., Zhao, Y., Zhao, Y., Zhao, Y. X., Zhao, Z., Zhao, Z. J., Zheng, D., Zheng, X. L., Zheng, X., Zhivotovsky, B., Zhong, Q., Zhou, G. Z., Zhou, G., Zhou, H., Zhou, S. F., Zhou, X. J., Zhu, H., Zhu, H., Zhu, W. G., Zhu, W., Zhu, X. F., Zhu, Y., Zhuang, S. M., Zhuang, X., Ziparo, E., Zois, C. E., Zoladek, T., Zong, W. X., Zorzano, A., and Zughaier, S. M. (2016) Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 12, Stade, K., Ford, C. S., Guthrie, C., and Weis, K. (1997) Exportin 1 (Crm1p) is an essential nuclear export factor. Cell 90,
27 29. Kaake, R. M., Milenkovic, T., Przulj, N., Kaiser, P., and Huang, L. (2010) Characterization of cell cycle specific protein interaction networks of the yeast 26S proteasome complex by the QTAX strategy. Journal of proteome research 9, Haruki, H., Nishikawa, J., and Laemmli, U. K. (2008) The anchor-away technique: rapid, conditional establishment of yeast mutant phenotypes. Mol Cell 31, Muller, M., Schmidt, O., Angelova, M., Faserl, K., Weys, S., Kremser, L., Pfaffenwimmer, T., Dalik, T., Kraft, C., Trajanoski, Z., Lindner, H., and Teis, D. (2015) The coordinated action of the MVB pathway and autophagy ensures cell survival during starvation. Elife 4, e Pack, C. G., Yukii, H., Toh-e, A., Kudo, T., Tsuchiya, H., Kaiho, A., Sakata, E., Murata, S., Yokosawa, H., Sako, Y., Baumeister, W., Tanaka, K., and Saeki, Y. (2014) Quantitative live-cell imaging reveals spatio-temporal dynamics and cytoplasmic assembly of the 26S proteasome. Nature communications 5, Giaever, G., Chu, A. M., Ni, L., Connelly, C., Riles, L., Veronneau, S., Dow, S., Lucau- Danila, A., Anderson, K., Andre, B., Arkin, A. P., Astromoff, A., El-Bakkoury, M., Bangham, R., Benito, R., Brachat, S., Campanaro, S., Curtiss, M., Davis, K., Deutschbauer, A., Entian, K. D., Flaherty, P., Foury, F., Garfinkel, D. J., Gerstein, M., Gotte, D., Guldener, U., Hegemann, J. H., Hempel, S., Herman, Z., Jaramillo, D. F., Kelly, D. E., Kelly, S. L., Kotter, P., LaBonte, D., Lamb, D. C., Lan, N., Liang, H., Liao, H., Liu, L., Luo, C., Lussier, M., Mao, R., Menard, P., Ooi, S. L., Revuelta, J. L., Roberts, C. J., Rose, M., Ross-Macdonald, P., Scherens, B., Schimmack, G., Shafer, B., Shoemaker, D. D., Sookhai-Mahadeo, S., Storms, R. K., Strathern, J. N., Valle, G., Voet, M., Volckaert, G., Wang, C. Y., Ward, T. R., Wilhelmy, J., Winzeler, E. A., Yang, Y., Yen, G., Youngman, E., Yu, K., Bussey, H., Boeke, J. D., Snyder, M., Philippsen, P., Davis, R. W., and Johnston, M. (2002) Functional profiling of the Saccharomyces cerevisiae genome. Nature 418, Tong, A. H., and Boone, C. (2006) Synthetic genetic array analysis in Saccharomyces cerevisiae. Methods Mol Biol 313, Haft, C. R., de la Luz Sierra, M., Barr, V. A., Haft, D. H., and Taylor, S. I. (1998) Identification of a family of sorting nexin molecules and characterization of their association with receptors. Mol Cell Biol 18, Gallon, M., and Cullen, P. J. (2015) Retromer and sorting nexins in endosomal sorting. Biochemical Society transactions 43, Ma, M., Burd, C. G., and Chi, R. J. (2017) Distinct complexes of yeast Snx4 family SNX-BARs mediate retrograde trafficking of Snc1 and Atg27. Traffic 18, Solis, G. P., Hulsbusch, N., Radon, Y., Katanaev, V. L., Plattner, H., and Stuermer, C. A. (2013) Reggies/flotillins interact with Rab11a and SNX4 at the tubulovesicular recycling compartment and function in transferrin receptor and E-cadherin trafficking. Mol Biol Cell 24, Traer, C. J., Rutherford, A. C., Palmer, K. J., Wassmer, T., Oakley, J., Attar, N., Carlton, J. G., Kremerskothen, J., Stephens, D. J., and Cullen, P. J. (2007) SNX4 coordinates endosomal sorting of TfnR with dynein-mediated transport into the endocytic recycling compartment. Nat Cell Biol 9, Hettema, E. H., Lewis, M. J., Black, M. W., and Pelham, H. R. (2003) Retromer and the sorting nexins Snx4/41/42 mediate distinct retrieval pathways from yeast endosomes. EMBO J 22, Bean, B. D., Davey, M., and Conibear, E. (2017) Cargo selectivity of yeast sorting nexins. Traffic 18, Maruzs, T., Lorincz, P., Szatmari, Z., Szeplaki, S., Sandor, Z., Lakatos, Z., Puska, G., Juhasz, G., and Sass, M. (2015) Retromer Ensures the Degradation of Autophagic Cargo by Maintaining Lysosome Function in Drosophila. Traffic 16,
28 43. Zhao, D., Liu, X. M., Yu, Z. Q., Sun, L. L., Xiong, X., Dong, M. Q., and Du, L. L. (2016) Atg20- and Atg24-family proteins promote organelle autophagy in fission yeast. J Cell Sci 129, Nice, D. C., Sato, T. K., Stromhaug, P. E., Emr, S. D., and Klionsky, D. J. (2002) Cooperative binding of the cytoplasm to vacuole targeting pathway proteins, Cvt13 and Cvt20, to phosphatidylinositol 3-phosphate at the pre-autophagosomal structure is required for selective autophagy. J Biol Chem 277, Kanki, T., Wang, K., Baba, M., Bartholomew, C. R., Lynch-Day, M. A., Du, Z., Geng, J., Mao, K., Yang, Z., Yen, W. L., and Klionsky, D. J. (2009) A genomic screen for yeast mutants defective in selective mitochondria autophagy. Mol Biol Cell 20, Collins, G. A., Gomez, T. A., Deshaies, R. J., and Tansey, W. P. (2010) Combined chemical and genetic approach to inhibit proteolysis by the proteasome. Yeast 27, Rosado, C. J., Mijaljica, D., Hatzinisiriou, I., Prescott, M., and Devenish, R. J. (2008) Rosella: a fluorescent ph-biosensor for reporting vacuolar turnover of cytosol and organelles in yeast. Autophagy 4, Welter, E., Thumm, M., and Krick, R. (2010) Quantification of nonselective bulk autophagy in S. cerevisiae using Pgk1-GFP. Autophagy 6, Huh, W. K., Falvo, J. V., Gerke, L. C., Carroll, A. S., Howson, R. W., Weissman, J. S., and O'Shea, E. K. (2003) Global analysis of protein localization in budding yeast. Nature. 425, Kirisako, T., Baba, M., Ishihara, N., Miyazawa, K., Ohsumi, M., Yoshimori, T., Noda, T., and Ohsumi, Y. (1999) Formation process of autophagosome is traced with Apg8/Aut7p in yeast. The Journal of cell biology 147, Kaganovich, D., Kopito, R., and Frydman, J. (2008) Misfolded proteins partition between two distinct quality control compartments. Nature 454, Nothwehr, S. F., Ha, S. A., and Bruinsma, P. (2000) Sorting of yeast membrane proteins into an endosome-to-golgi pathway involves direct interaction of their cytosolic domains with Vps35p. The Journal of cell biology 151, Savulescu, A. F., Shorer, H., Kleifeld, O., Cohen, I., Gruber, R., Glickman, M. H., and Harel, A. (2011) Nuclear import of an intact preassembled proteasome particle. Mol Biol Cell 22, Dambacher, C. M., Worden, E. J., Herzik, M. A., Martin, A., and Lander, G. C. (2016) Atomic structure of the 26S proteasome lid reveals the mechanism of deubiquitinase inhibition. Elife 5, e Groll, M., Bajorek, M., Kohler, A., Moroder, L., Rubin, D. M., Huber, R., Glickman, M. H., and Finley, D. (2000) A gated channel into the proteasome core particle. Nature structural biology 7, Kleijnen, M. F., Roelofs, J., Park, S., Hathaway, N. A., Glickman, M., King, R. W., and Finley, D. (2007) Stability of the proteasome can be regulated allosterically through engagement of its proteolytic active sites. Nat Struct Mol Biol 14, Thomsson, E., Larsson, C., Albers, E., Nilsson, A., Franzen, C. J., and Gustafsson, L. (2003) Carbon starvation can induce energy deprivation and loss of fermentative capacity in Saccharomyces cerevisiae. Applied and environmental microbiology 69, Nilsson, A., Pahlman, I. L., Jovall, P. A., Blomberg, A., Larsson, C., and Gustafsson, L. (2001) The catabolic capacity of Saccharomyces cerevisiae is preserved to a higher extent during carbon compared to nitrogen starvation. Yeast 18, Rodkaer, S. V., and Faergeman, N. J. (2014) Glucose- and nitrogen sensing and regulatory mechanisms in Saccharomyces cerevisiae. FEMS Yeast Res 14,
29 60. Wani, P. S., Suppahia, A., Capalla, X., Ondracek, A., and Roelofs, J. (2016) Phosphorylation of the C-terminal tail of proteasome subunit alpha7 is required for binding of the proteasome quality control factor Ecm29. Scientific reports 6, Wang, X., Yen, J., Kaiser, P., and Huang, L. (2010) Regulation of the 26S proteasome complex during oxidative stress. Science signaling 3, ra Laporte, D., Salin, B., Daignan-Fornier, B., and Sagot, I. (2008) Reversible cytoplasmic localization of the proteasome in quiescent yeast cells. The Journal of cell biology 181, Kramer, M. H., Farre, J. C., Mitra, K., Yu, M. K., Ono, K., Demchak, B., Licon, K., Flagg, M., Balakrishnan, R., Cherry, J. M., Subramani, S., and Ideker, T. (2017) Active Interaction Mapping Reveals the Hierarchical Organization of Autophagy. Mol Cell 65, e Ohashi, Y., and Munro, S. (2010) Membrane delivery to the yeast autophagosome from the Golgi-endosomal system. Mol Biol Cell 21, Tooze, S. A., Abada, A., and Elazar, Z. (2014) Endocytosis and autophagy: exploitation or cooperation? Cold Spring Harb Perspect Biol 6, a Berg, T. O., Fengsrud, M., Stromhaug, P. E., Berg, T., and Seglen, P. O. (1998) Isolation and characterization of rat liver amphisomes. Evidence for fusion of autophagosomes with both early and late endosomes. J Biol Chem 273, Razi, M., Chan, E. Y., and Tooze, S. A. (2009) Early endosomes and endosomal coatomer are required for autophagy. The Journal of cell biology 185, Guthrie, C., and Fink, G. R. (1991) Guide to Yeast Genetics and Molecular Biology, Academic Press, San Diego 69. Kushnirov, V. V. (2000) Rapid and reliable protein extraction from yeast. Yeast 16, Tomko, R. J., Jr., and Hochstrasser, M. (2014) The intrinsically disordered Sem1 protein functions as a molecular tether during proteasome lid biogenesis. Mol Cell 53,
30 FOOTNOTES The abbreviations used are: BAR, Bin/amphiphysin/Rvs; CP, core particle; CVT, cytoplasm-tovacuole targeting; FAS, fatty acid synthase; GFP, green fluorescent protein; PAS, phagophore assembly site; PI3P, phosphatidylinositol 3-phosphate; PM, plasma membrane; PMN, piecemeal autophagy of the nucleus; PX, phox homology; RP, regulatory particle; UPS, ubiquitin-proteasome system 30
31 FIGURE LEGENDS Figure 1. Proteasome autophagy is independent of known nucleophagy genes and is impaired by blockade of nuclear export. A, Proteasome subunit-gfp fusion autophagy reporters used in Figure 1. Rpn5 and Rpn11 are subunits of the lid subcomplex, Rpn2 of the base, and α4 of the core particle subcomplex. B, whole cell extracts of the indicated strains expressing proteasome subunits as GFP fusions from their chromosomal loci were prepared under non-denaturing conditions and separated by non-denaturing polyacrylamide electrophoresis before in-gel imaging of GFP fluorescence (left), peptidase activity (middle), or immunoblotting with antibodies against the core particle (right). Uncapped CP displays comparatively weak peptidase activity because of restricted access of the substrate to the peptidase sites in the absence of the RP. C, the indicated strains were subjected to nitrogen starvation for the indicated times, followed by immunoblotting of cell extracts with anti-gfp antibodies. D, cells containing the xpo1-1 nuclear export mutation and expressing a nucleocytoplasmic shuttling reporter, NLS-NES-GFP, were incubated in log phase at 25 o C before shift to 37 o C for 15 minutes or 12 hours as shown. E, cells harboring the xpo1-1 conditional nuclear export mutant allele and expressing GFP-Atg8 (E), α4-gfp (F), Rpn2-GFP (G), or Rpn5-GFP (H) were grown at permissive temperature in rich media and then transferred to SD-N medium. After three hours, the cells were either shifted to 37 o C (black circles) or left at 25 o C before harvest at the indicated times for immunoblotting of cell extracts with antibodies against GFP. The percentage of GFP fragment is shown as % cleaved ± standard deviation (n 3). Figure 2. Nuclear proteasomes can disassemble prior to autophagy. A, Experimental scheme to test whether proteasomal subcomplexes can disassemble prior to autophagy. Rpn2-GFP or α4- GFP was expressed from the chromosomal locus in tether-anchor strains harboring chromosomal RPN11-mCherry-FRB (tether) and either no anchor, HTB2-FKBP12 or PMA1-2xFKBP12 alleles. The strains were then treated with DMSO vehicle or with 1 µg/ml rapamycin, followed by nitrogen starvation for different time periods before examination of GFP cleavage by immunoblotting. B, histone Htb2-GFP is not appreciably turned over by autophagy in response to nitrogen starvation. Yeast expressing Htb2-GFP from the chromosomal locus were nitrogen-starved for the indicated time periods before harvesting. Cell extracts were separated via SDS-PAGE and subjected to immunoblotting with anti-gfp antibodies. An asterisk indicates post-translationally modified Htb2-GFP. The blot was intentionally overexposed to emphasize that essentially no free GFP is released upon nitrogen starvation. C, cells harboring WT TOR1 or the rapamycin-insensitive TOR1-1 allele were treated with DMSO vehicle or 1 μg/ml rapamycin for the indicated times. Cell extracts from treated cells were then subjected to immunoblotting with anti-gfp antibodies. D, confirmation of subcellular tethering of proteasome subunits. Cells expressing no anchor (top panels), the Htb2-FKBP12 nuclear anchor (middle panels), or the cytosol-facing plasma membrane anchor Pma1-2xFKBP12 (bottom panels) were pre-treated for three hours with 1 μg/ml rapamycin, at which point they were washed and transferred to SD-N media containing 1 μg/ml rapamycin, and incubated for 24 hours at 30 o C. Representative micrographs of α4-gfp and Rpn11-mCherry- FRB fluorescence, and merged fluorescence images are shown. Scale bar = 10 μm. E, cells expressing Rpn11-mCherry-FRB, and α4-gfp as well as either the nuclear anchor Htb2-FKBP12 (left panel) or the cytosol-facing plasma membrane anchor Pma1-2xFKBP12 (right panel) were treated with DMSO or rapamycin as indicated and described in (D) before nitrogen starvation for the indicated times. Cell extracts were prepared and subjected to immunoblotting with anti-gfp antibodies. F, as in (E), but with cells expressing Rpn2-GFP instead of α4-gfp. Asterisk, autophagy-independent proteolytic fragment of Rpn2-GFP. This fragment was present coincident with a decrease in full-length Rpn2-GFP upon nitrogen starvation even in atg8δ cells (not shown), indicating that Rpn2-GFP may undergo some nonspecific autophagy-independent proteolysis in vivo. 31
32 Figure 3. A targeted genetic screen to define ATG genes required for proteasome autophagy. The indicated strains were nitrogen-starved for 16 hours before harvesting for immunoblotting of whole cell extracts with anti-gfp antibodies. An arrowhead highlights the complete loss of autophagy in the atg24δ (snx4δ) strain. Inset table, description of the autophagy-specific functions of genes that when deleted, demonstrated a complete loss of GFP formation. Genes encoding components of the core autophagy machinery are listed in purple both in the immunoblots and in the table. Genes specific to nitrogen starvation are shown in blue, and ATG24/SNX4 is shown in red. Figure 4. Snx4 is required for selective autophagy of the proteasome. A, The indicated yeast strains were nitrogen starved for the times shown before harvesting and analysis of whole cell extracts by anti-gfp immunoblotting as in Figure 3. B, The indicated strains were nitrogen starved for the times shown and then subjected to anti-gfp immunoblotting. Asterisk, nonspecific Rpn2- GFP cleavage product unrelated to autophagy. C, The indicated yeast strains, each of which harbored a deletion of PDR5, were treated for 24 hours with 50 μm MG132 before analysis as in (A). Autophagy of α4-gfp in response to cellular treatment with proteasome inhibitors is also compromised in snx4δ yeast. Cells were treated for the indicated times with 50 µm MG132 (black circles) or DMSO (no circle), followed by harvesting for immunoblotting with antibodies against GFP. D and E, deletion of SNX4 does not compromise nonselective autophagy. Turnover of the bulk autophagy substrates Rosella (D) or Pgk1-GFP (E) upon nitrogen starvation in the indicated strains was assessed as in (A). In (D), an asterisk indicates a nonspecific proteolytic product unrelated to autophagy. Figure 5. Snx4 is required for autophagic turnover of two other multisubunit complexes. A, WT or snx4δ cells expressing Fas2-GFP from the chromosomal locus were nitrogen-starved for the indicated times before harvesting for SDS-PAGE and immunoblotting with anti-gfp antibodies. An asterisk indicates a nonspecific proteolytic fragment unrelated to autophagy. B, As in (A), but with cells expressing the large ribosomal subunit Rpl25 as a GFP fusion instead of Fas2- GFP. C, as in (A), but with cells expressing the small ribosomal subunit Rps2 as a GFP fusion. D, Equal numbers of WT or snx4δ cells were spotted in six-fold serial dilutions on YPD plates containing DMSO vehicle or 50 ng/μl rapamycin before incubation as shown and imaging. Figure 6. Snx4 is required for the formation of cytoplasmic α4-gfp puncta. A, WT or snx4δ cells expressing α4-gfp and Vph1-mCherry (as a vacuolar membrane marker) were nitrogen starved for 16 hours before imaging. All strains lacked the vacuolar peptidase PEP4 to stabilize vacuolar contents. Representative micrographs are shown. Scale bar = 5 µm. B, statistical dot plot with means (black bars) of the intensity of GFP signal from the vacuole (left) or nucleus for WT (n = 23) and snx4δ (n = 21) cells (two-tailed Mann-Whitney test). C, the indicated strains expressing α4-gfp were nitrogen-starved for 16 hours before imaging. Representative micrographs of atg8δ, snx4δ atg8δ, or vps35δ atg8δ cells expressing α4-gfp are shown. White arrowheads indicate GFP puncta. Scale bar = 5 μm. D, quantification of cytoplasmic α4-gfp puncta in the indicated strains after nitrogen starvation for 16 hours (n > 100 cells; one-way ANOVA with Tukey's test for multiple comparisons; error bars = SEM). Figure 7. Snx4 cooperates with Snx41 and Snx42 to promote autophagy of proteasomes. A, the indicated strains were nitrogen-starved, followed by analysis of whole-cell extracts by SDS- PAGE and immunoblotting with anti-gfp antibodies. The percentage of GFP fragment is shown as % cleaved. B, overexpression of SNX4 fails to rescue the autophagy defect in snx41δ snx42δ cells. The indicated cells were transformed with either empty plasmid (no circles) or a plasmid that overproduces SNX4. Cells were then nitrogen-starved or not as indicated, followed by analysis of whole-cell extracts as in (A). C, SNX4-GFP is functional. Equal numbers of the indicated cells 32
33 were spotted in six-fold serial dilutions on YPD plates containing DMSO vehicle or 50 ng/μl rapamycin before incubation as shown and imaging. D, micrographs of Snx4-GFP fluorescence in the indicated strains after nitrogen starvation for 16 hours. Scale bar = 5 μm. E, Analysis of punctate or diffuse Snx4-GFP localization (Two-way ANOVA; error bars = SEM). ns, not significantly different from WT; ***, p = vs. WT; ****, p < vs. WT; ^^^^, p < vs. WT. F, snx4δ cells were transformed with plasmids expressing either SNX4 or snx4(y79a) from the endogenous SNX4 promoter. Transformants were then subjected to nitrogen starvation and analysis of whole cell extracts by anti-gfp immunoblotting as in (A). The percentage of GFP fragment is shown as % cleaved. G, quantification of cytoplasmic α4-gfp puncta in the indicated strains after nitrogen starvation for 16 hours (n > 100 cells; one-way ANOVA with Tukey's test for multiple comparisons; error bars = SEM). Figure 8. Model for delivery of proteasomes to the vacuole for destruction. The data support a model in which nuclear proteasomes are disassembled prior to autophagy and within the nucleus, whereas cytoplasmic proteasomes remain intact. Snx4 promotes the agglomeration of proteasomes into cytoplasmic puncta that accumulate in a step prior to Atg8-dependent encapsulation in the autophagosomal membrane (orange circle). Although the model depicts these Snx4-dependent puncta as membrane-enclosed (black circle), it is not yet clear if this is the case. Atg8 then mediates traditional encapsulation in the autophagosomal membrane for delivery to and destruction in the vacuole. 33

")


 GFP-Atg8 GFP % cleaved ± 0 ± 2 ±17 ±")
 37 C shift @ 3 h N-starve (h) α4-gfp")
 D G kda 150 100 75 50 37 25 H")




34 A B Base Core Particle Reporter Rpn5- or Rpn11-GFP Rpn2-GFP α4-gfp RP 2 CP RP 1 CP CP GFP fluorescence Peptidase activity (suc-llvy-amc fluorescence) Anti-20S (CP blot) C E F kda kda kda N-starve (h) α4-gfp GFP 37 C 3 h N-starve (h) GFP-Atg8 GFP % cleaved ± 0 ± 2 ±17 ± 3 ± 0 ± 8 ± 5 Std. Dev. (%) 37 C 3 h N-starve (h) α4-gfp GFP % cleaved ± 5 ±7 ± 6 ± 4 ± 7 ± 7 Std. Dev. (%) D G kda H kda C 37 C, 15 min 37 C, 12 h NLS-NES-GFP NLS-NES-GFP NLS-NES-GFP 37 C 3 h N-starve (h) Rpn2-GFP GFP % cleaved ± 4 ±2 ±3 ± 6 ± 7 ± 5 Std. Dev. (%) 37 C 3 h N-starve (h) Rpn5-GFP GFP % cleaved ± 8 ±9 ± 5 ± 2 ± 3 ± 2 Std. Dev. (%) Nemec et al., Figure 1







")


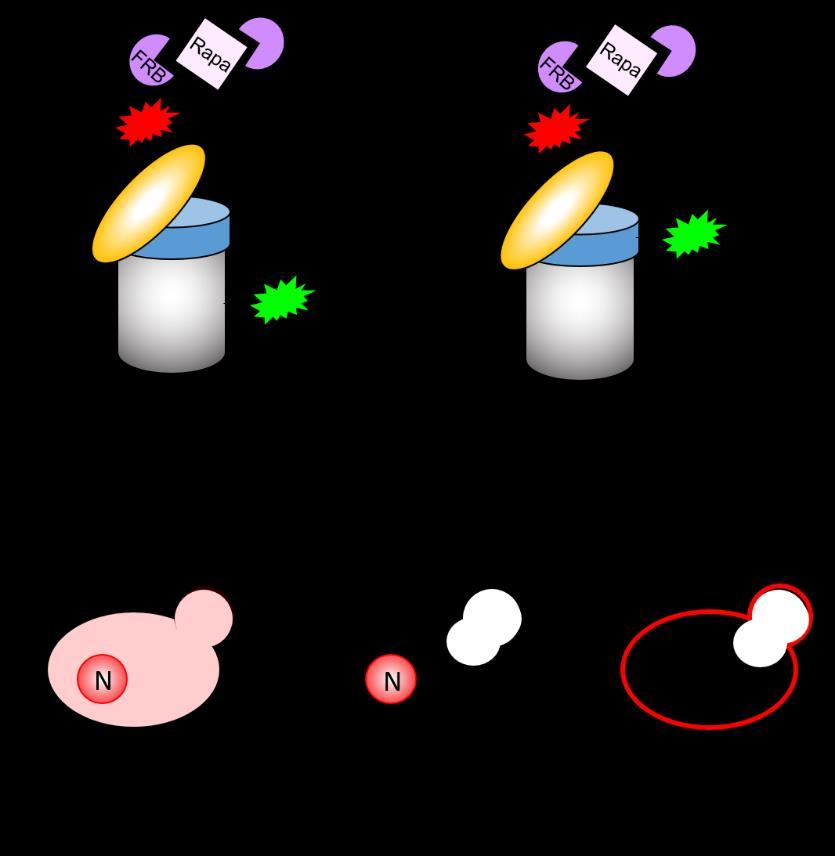
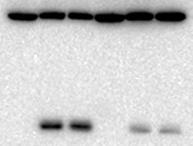
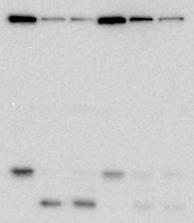
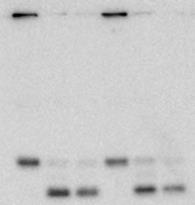
35 A B kda N-starve (h) * Htb2-GFP 25 GFP D No anchor Nuclear anchor PM anchor C kda DMSO Rapamycin DMSO Rapamycin DMSO Rapamycin i ii iii iv v vi vii viii ix x xi xii xiii xiv xv xvi xvii xviii Rapamycin (h) α4-gfp GFP α4-gfp Rpn11-mCherry-FRB Merged E F kda Nuclear anchor PM anchor Rapa N-starve (h) α4-gfp GFP kda Rapa N-starve (h) Rpn2-GFP Nuclear anchor PM anchor GFP Nemec et al., Figure 2 *



36 kda α4-gfp GFP kda α4-gfp GFP Role in autophagy Core Machinery Nitrogen Starvation Other ATG mutants with a complete loss of α4-gfp autophagy ATG1,2,3,4,5,6,7,8,9,10,12,13,14, 15,16,18 ATG17,29,31 ATG24 (SNX4) Nemec et al., Figure 3
 α4-gfp GFP kda 75 50 37 0 24 24 24 N-starve (h) Rosella")
 PDR5")
 Pgk1-GFP GFP Nemec et al.")
37 A WT snx4δ atg8δ D kda N-starve (h) α4-gfp GFP kda N-starve (h) Rosella * B Rpn2-GFP Rpn5-GFP 25 GFP kda C kda WT WT Δ Δ Δ Δ Δ Δ WT WT Δ SNX N-starve (h) PDR5 MG-132 α4-gfp GFP Rpn2-GFP Rpn5-GFP * GFP E kda WT snx4δ N-starve (h) Pgk1-GFP GFP Nemec et al., Figure 4



38 A kda N-starve (h) Fas2-GFP * B kda WT snx4δ N-starve (h) Rpl25-GFP 25 GFP 25 GFP C D kda N-starve (h) Rps2-GFP GFP WT snx4δ DMSO 3d, 30 o C 50 ng/μl Rapa, 6d, 30 o C Nemec et al., Figure 5

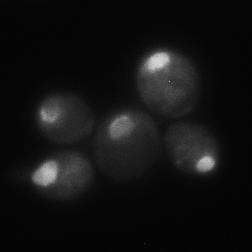
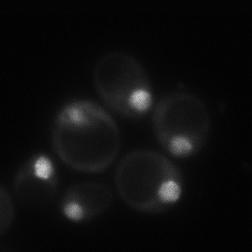
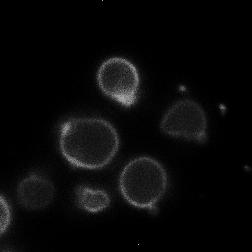
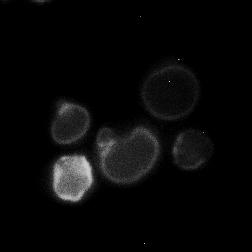
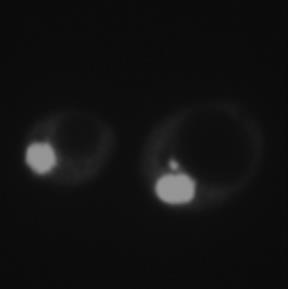
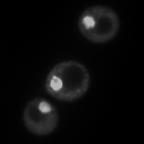
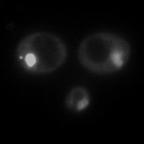
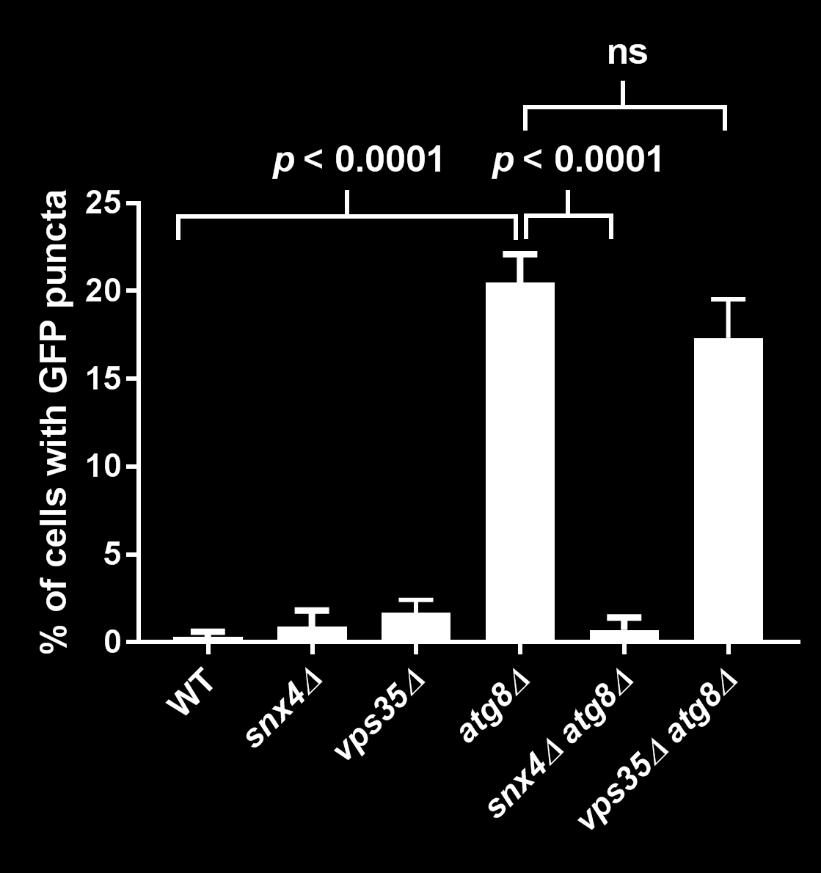
39 A B WT α4-gfp Vph1-mCherry Merge snx4δ C atg8δ snx4δ atg8δ vps35δ atg8δ D α4-gfp α4-gfp α4-gfp Nemec et al., Figure 6



 α4-gfp 37 37 25")




 α4-gfp 37 25 29")

40 A B snx4δ snx41δ snx42δ snx41δ snx42δ O/E SNX4 kda N-starve (h) α4-gfp kda N-starve (h) α4-gfp GFP 25 GFP % cleaved C snx4δ D WT snx41δ Wild-type SNX4-GFP E DMSO 3d, 30 o C 50 ng/μl Rapa, 6d, 30 o C Snx4- GFP G snx42δ snx41δ snx42δ F kda snx4δ SNX4 snx4-y79a N-starve (h) α4-gfp GFP % cleaved Nemec et al., Figure 7
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins
 13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
Lecture 6 - Intracellular compartments and transport I
 01.26.11 Lecture 6 - Intracellular compartments and transport I Intracellular transport and compartments 1. Protein sorting: How proteins get to their appropriate destinations within the cell 2. Vesicular
01.26.11 Lecture 6 - Intracellular compartments and transport I Intracellular transport and compartments 1. Protein sorting: How proteins get to their appropriate destinations within the cell 2. Vesicular
CELB40060 Membrane Trafficking in Animal Cells. Prof. Jeremy C. Simpson. Lecture 2 COPII and export from the ER
 CELB40060 Membrane Trafficking in Animal Cells Prof. Jeremy C. Simpson Lecture 2 COPII and export from the ER Today s lecture... The COPII coat - localisation and subunits Formation of the COPII coat at
CELB40060 Membrane Trafficking in Animal Cells Prof. Jeremy C. Simpson Lecture 2 COPII and export from the ER Today s lecture... The COPII coat - localisation and subunits Formation of the COPII coat at
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
Protein Sorting, Intracellular Trafficking, and Vesicular Transport
 Protein Sorting, Intracellular Trafficking, and Vesicular Transport Noemi Polgar, Ph.D. Department of Anatomy, Biochemistry and Physiology Email: polgar@hawaii.edu Phone: 692-1422 Outline Part 1- Trafficking
Protein Sorting, Intracellular Trafficking, and Vesicular Transport Noemi Polgar, Ph.D. Department of Anatomy, Biochemistry and Physiology Email: polgar@hawaii.edu Phone: 692-1422 Outline Part 1- Trafficking
7.06 Problem Set
 7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces cerevisiae.
 Genetics: Published Articles Ahead of Print, published on July 30, 2012 as 10.1534/genetics.112.143818 A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces
Genetics: Published Articles Ahead of Print, published on July 30, 2012 as 10.1534/genetics.112.143818 A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Chapter 12: Intracellular sorting
 Chapter 12: Intracellular sorting Principles of intracellular sorting Principles of intracellular sorting Cells have many distinct compartments (What are they? What do they do?) Specific mechanisms are
Chapter 12: Intracellular sorting Principles of intracellular sorting Principles of intracellular sorting Cells have many distinct compartments (What are they? What do they do?) Specific mechanisms are
Cell Biology Review. The key components of cells that concern us are as follows: 1. Nucleus
 Cell Biology Review Development involves the collective behavior and activities of cells, working together in a coordinated manner to construct an organism. As such, the regulation of development is intimately
Cell Biology Review Development involves the collective behavior and activities of cells, working together in a coordinated manner to construct an organism. As such, the regulation of development is intimately
7.06 Cell Biology EXAM #3 KEY
 7.06 Cell Biology EXAM #3 KEY May 2, 2006 This is an OPEN BOOK exam, and you are allowed access to books, a calculator, and notes BUT NOT computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 KEY May 2, 2006 This is an OPEN BOOK exam, and you are allowed access to books, a calculator, and notes BUT NOT computers or any other types of electronic devices. Please write
Transport between cytosol and nucleus
 of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
of 60 3 Gated trans Lectures 9-15 MBLG 2071 The n GATED TRANSPORT transport between cytoplasm and nucleus (bidirectional) controlled by the nuclear pore complex active transport for macro molecules e.g.
!"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%%
*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%%") !"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%% !"#$%&'(")*++*%,*'-&'./%/,*#01#%-2)#3&)/% 4'(")*++*% % %5"0)%-2)#3&) %%% %67'2#72'*%%%%%%%%%%%%%%%%%%%%%%%4'(")0/./% % 8$+&'&,+"/7 % %,$&7&/9)7$*/0/%%%%%%%%%%
!"#$%&'%()*%+*,,%-&,./*%01%02%/*/3452*%3&.26%&4752*,,*1%% !"#$%&'(")*++*%,*'-&'./%/,*#01#%-2)#3&)/% 4'(")*++*% % %5"0)%-2)#3&) %%% %67'2#72'*%%%%%%%%%%%%%%%%%%%%%%%4'(")0/./% % 8$+&'&,+"/7 % %,$&7&/9)7$*/0/%%%%%%%%%%
Importance of Protein sorting. A clue from plastid development
 Importance of Protein sorting Cell organization depend on sorting proteins to their right destination. Cell functions depend on sorting proteins to their right destination. Examples: A. Energy production
Importance of Protein sorting Cell organization depend on sorting proteins to their right destination. Cell functions depend on sorting proteins to their right destination. Examples: A. Energy production
Chapter 6: A Tour of the Cell
 Chapter 6: A Tour of the Cell 1. The study of cells has been limited by their small size, and so they were not seen and described until 1665, when Robert Hooke first looked at dead cells from an oak tree.
Chapter 6: A Tour of the Cell 1. The study of cells has been limited by their small size, and so they were not seen and described until 1665, when Robert Hooke first looked at dead cells from an oak tree.
Chapter 4 Active Reading Guide A Tour of the Cell
 Name: AP Biology Mr. Croft Chapter 4 Active Reading Guide A Tour of the Cell Section 1 1. The study of cells has been limited by their small size, and so they were not seen and described until 1665, when
Name: AP Biology Mr. Croft Chapter 4 Active Reading Guide A Tour of the Cell Section 1 1. The study of cells has been limited by their small size, and so they were not seen and described until 1665, when
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 2: Phagocytosis Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research Salmonella Gram negative bacteria
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 2: Phagocytosis Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research Salmonella Gram negative bacteria
Richik N. Ghosh, Linnette Grove, and Oleg Lapets ASSAY and Drug Development Technologies 2004, 2:
 1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
Biological Process Term Enrichment
 Biological Process Term Enrichment cellular protein localization cellular macromolecule localization intracellular protein transport intracellular transport generation of precursor metabolites and energy
Biological Process Term Enrichment cellular protein localization cellular macromolecule localization intracellular protein transport intracellular transport generation of precursor metabolites and energy
Chapter 6: A Tour of the Cell
 AP Biology Reading Guide Fred and Theresa Holtzclaw Chapter 6: A Tour of the Cell Name Period Chapter 6: A Tour of the Cell Concept 6.1 To study cells, biologists use microscopes and the tools of biochemistry
AP Biology Reading Guide Fred and Theresa Holtzclaw Chapter 6: A Tour of the Cell Name Period Chapter 6: A Tour of the Cell Concept 6.1 To study cells, biologists use microscopes and the tools of biochemistry
Division Ave. High School AP Biology
 Tour of the Cell 1 Types of cells Prokaryote bacteria cells - no organelles - organelles Eukaryote animal cells Eukaryote plant cells Why organelles? Specialized structures u specialized functions cilia
Tour of the Cell 1 Types of cells Prokaryote bacteria cells - no organelles - organelles Eukaryote animal cells Eukaryote plant cells Why organelles? Specialized structures u specialized functions cilia
MOLECULAR CELL BIOLOGY
 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
Heather Currinn, Benjamin Guscott, Zita Balklava, Alice Rothnie and Thomas Wassmer*
 Online Resources APP controls the formation of PI(3,5)P 2 vesicles through its binding of the PIKfyve complex. Cellular and Molecular Life Sciences Heather Currinn, Benjamin Guscott, Zita Balklava, Alice
Online Resources APP controls the formation of PI(3,5)P 2 vesicles through its binding of the PIKfyve complex. Cellular and Molecular Life Sciences Heather Currinn, Benjamin Guscott, Zita Balklava, Alice
Chapter 6: A Tour of the Cell
 Chapter 6: A Tour of the Cell Concept 6.2 Eukaryotic cells have internal membranes that compartmentalize their functions 1. Which two domains consist of prokaryotic cells? 2. A major difference between
Chapter 6: A Tour of the Cell Concept 6.2 Eukaryotic cells have internal membranes that compartmentalize their functions 1. Which two domains consist of prokaryotic cells? 2. A major difference between
Cell (Learning Objectives)
") Cell (Learning Objectives) 1. Understand & describe the basic components necessary for a functional cell. 2. Review the order of appearance of cells on earth and explain the endosymbiotic theory. 3. Compare
Cell (Learning Objectives) 1. Understand & describe the basic components necessary for a functional cell. 2. Review the order of appearance of cells on earth and explain the endosymbiotic theory. 3. Compare
Regulation of gene expression. Premedical - Biology
 Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Regulation of gene expression Premedical - Biology Regulation of gene expression in prokaryotic cell Operon units system of negative feedback positive and negative regulation in eukaryotic cell - at any
Concept 6.1 To study cells, biologists use microscopes and the tools of biochemistry
 Name Period Chapter 6: A Tour of the Cell Concept 6.1 To study cells, biologists use microscopes and the tools of biochemistry 1. The study of cells has been limited by their small size, and so they were
Name Period Chapter 6: A Tour of the Cell Concept 6.1 To study cells, biologists use microscopes and the tools of biochemistry 1. The study of cells has been limited by their small size, and so they were
The Cell. C h a p t e r. PowerPoint Lecture Slides prepared by Jason LaPres North Harris College Houston, Texas
 C h a p t e r 2 The Cell PowerPoint Lecture Slides prepared by Jason LaPres North Harris College Houston, Texas Copyright 2009 Pearson Education, Inc., publishing as Pearson Benjamin Cummings Introduction
C h a p t e r 2 The Cell PowerPoint Lecture Slides prepared by Jason LaPres North Harris College Houston, Texas Copyright 2009 Pearson Education, Inc., publishing as Pearson Benjamin Cummings Introduction
2011 The Simple Homeschool Simple Days Unit Studies Cells
 1 We have a full line of high school biology units and courses at CurrClick and as online courses! Subscribe to our interactive unit study classroom and make science fun and exciting! 2 A cell is a small
1 We have a full line of high school biology units and courses at CurrClick and as online courses! Subscribe to our interactive unit study classroom and make science fun and exciting! 2 A cell is a small
Name: TF: Section Time: LS1a ICE 5. Practice ICE Version B
 Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
CELL PART Expanded Definition Cell Structure Illustration Function Summary Location ALL CELLS DNA Common in Animals Uncommon in Plants Lysosome
 CELL PART Expanded Definition Cell Structure Illustration Function Summary Location is the material that contains the Carry genetic ALL CELLS information that determines material inherited characteristics.
CELL PART Expanded Definition Cell Structure Illustration Function Summary Location is the material that contains the Carry genetic ALL CELLS information that determines material inherited characteristics.
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
targets. clustering show that different complex pathway
 Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Introduction. Gene expression is the combined process of :
 1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
1 To know and explain: Regulation of Bacterial Gene Expression Constitutive ( house keeping) vs. Controllable genes OPERON structure and its role in gene regulation Regulation of Eukaryotic Gene Expression
DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS. Scientific Background on the Nobel Prize in Medicine 2013
 DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS Scientific Background on the Nobel Prize in Medicine 2013 Daniela Scalet 6/12/2013 The Nobel Prize in Medicine
DISCOVERIES OF MACHINERY REGULATING VESICLE TRAFFIC, A MAJOR TRANSPORT SYSTEM IN OUR CELLS Scientific Background on the Nobel Prize in Medicine 2013 Daniela Scalet 6/12/2013 The Nobel Prize in Medicine
Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes
 9 The Nucleus Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes Explain general structures of Nuclear Envelope, Nuclear Lamina, Nuclear Pore Complex Explain movement of proteins
9 The Nucleus Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes Explain general structures of Nuclear Envelope, Nuclear Lamina, Nuclear Pore Complex Explain movement of proteins
Components of a functional cell. Boundary-membrane Cytoplasm: Cytosol (soluble components) & particulates DNA-information Ribosomes-protein synthesis
 & particulates DNA-information Ribosomes-protein synthesis") Cell (Outline) - Components of a functional cell - Major Events in the History of Earth: abiotic and biotic phases; anaerobic and aerobic atmosphere - Prokaryotic cells impact on the biosphere - Origin
Cell (Outline) - Components of a functional cell - Major Events in the History of Earth: abiotic and biotic phases; anaerobic and aerobic atmosphere - Prokaryotic cells impact on the biosphere - Origin
Chapter 4: Cells: The Working Units of Life
 Name Period Chapter 4: Cells: The Working Units of Life 1. What are the three critical components of the cell theory? 2. What are the two important conceptual implications of the cell theory? 3. Which
Name Period Chapter 4: Cells: The Working Units of Life 1. What are the three critical components of the cell theory? 2. What are the two important conceptual implications of the cell theory? 3. Which
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Breker et al., http://www.jcb.org/cgi/content/full/jcb.201301120/dc1 Figure S1. Single-cell proteomics of stress responses. (a) Using
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Breker et al., http://www.jcb.org/cgi/content/full/jcb.201301120/dc1 Figure S1. Single-cell proteomics of stress responses. (a) Using
Reconstructing Mitochondrial Evolution?? Morphological Diversity. Mitochondrial Diversity??? What is your definition of a mitochondrion??
 Reconstructing Mitochondrial Evolution?? What is your definition of a mitochondrion?? Morphological Diversity Mitochondria as we all know them: Suprarenal gland Liver cell Plasma cell Adrenal cortex Mitochondrial
Reconstructing Mitochondrial Evolution?? What is your definition of a mitochondrion?? Morphological Diversity Mitochondria as we all know them: Suprarenal gland Liver cell Plasma cell Adrenal cortex Mitochondrial
Chapter 4 A Tour of the Cell*
 Chapter 4 A Tour of the Cell* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Fundamental Units of Life Cells
Chapter 4 A Tour of the Cell* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Fundamental Units of Life Cells
Overview of Cells. Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory
 Overview of Cells Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory Prokaryotic Cells Archaea Bacteria Come in many different shapes and sizes.5 µm 2 µm, up to 60 µm long Have large
Overview of Cells Prokaryotes vs Eukaryotes The Cell Organelles The Endosymbiotic Theory Prokaryotic Cells Archaea Bacteria Come in many different shapes and sizes.5 µm 2 µm, up to 60 µm long Have large
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2647 Figure S1 Other Rab GTPases do not co-localize with the ER. a, Cos-7 cells cotransfected with an ER luminal marker (either KDEL-venus or mch-kdel) and mch-tagged human Rab5 (mch-rab5,
DOI: 10.1038/ncb2647 Figure S1 Other Rab GTPases do not co-localize with the ER. a, Cos-7 cells cotransfected with an ER luminal marker (either KDEL-venus or mch-kdel) and mch-tagged human Rab5 (mch-rab5,
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Quiz answers. Allele. BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA)
 Lecture 17: The Quiz (and back to Eukaryotic DNA)") BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA) http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Quiz answers Kinase: An enzyme
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 17: The Quiz (and back to Eukaryotic DNA) http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Quiz answers Kinase: An enzyme
7.06 Cell Biology EXAM #3 April 21, 2005
 7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
Yeast Genome-wide Screens to Ascertain the Genetic Landscape for Barth Syndrome. Christopher R. McMaster, PhD Dalhousie University
 Yeast Genome-wide Screens to Ascertain the Genetic Landscape for Barth Syndrome Christopher R. McMaster, PhD Dalhousie University Using Systematic Genetics to Identify Modifies Genes that Affect Fitness
Yeast Genome-wide Screens to Ascertain the Genetic Landscape for Barth Syndrome Christopher R. McMaster, PhD Dalhousie University Using Systematic Genetics to Identify Modifies Genes that Affect Fitness
A. The Cell: The Basic Unit of Life. B. Prokaryotic Cells. D. Organelles that Process Information. E. Organelles that Process Energy
 The Organization of Cells A. The Cell: The Basic Unit of Life Lecture Series 4 The Organization of Cells B. Prokaryotic Cells C. Eukaryotic Cells D. Organelles that Process Information E. Organelles that
The Organization of Cells A. The Cell: The Basic Unit of Life Lecture Series 4 The Organization of Cells B. Prokaryotic Cells C. Eukaryotic Cells D. Organelles that Process Information E. Organelles that
Chapter 1. DNA is made from the building blocks adenine, guanine, cytosine, and. Answer: d
 Chapter 1 1. Matching Questions DNA is made from the building blocks adenine, guanine, cytosine, and. Answer: d 2. Matching Questions : Unbranched polymer that, when folded into its three-dimensional shape,
Chapter 1 1. Matching Questions DNA is made from the building blocks adenine, guanine, cytosine, and. Answer: d 2. Matching Questions : Unbranched polymer that, when folded into its three-dimensional shape,
Supplemental Information. Expanded Coverage of the 26S Proteasome. Conformational Landscape Reveals. Mechanisms of Peptidase Gating
 Cell Reports, Volume 24 Supplemental Information Expanded Coverage of the 26S Proteasome Conformational Landscape Reveals Mechanisms of Peptidase Gating Markus R. Eisele, Randi G. Reed, Till Rudack, Andreas
Cell Reports, Volume 24 Supplemental Information Expanded Coverage of the 26S Proteasome Conformational Landscape Reveals Mechanisms of Peptidase Gating Markus R. Eisele, Randi G. Reed, Till Rudack, Andreas
The neuron as a secretory cell
 The neuron as a secretory cell EXOCYTOSIS ENDOCYTOSIS The secretory pathway. Transport and sorting of proteins in the secretory pathway occur as they pass through the Golgi complex before reaching the
The neuron as a secretory cell EXOCYTOSIS ENDOCYTOSIS The secretory pathway. Transport and sorting of proteins in the secretory pathway occur as they pass through the Golgi complex before reaching the
S1 Gene ontology (GO) analysis of the network alignment results
 analysis of the network alignment results") 1 Supplementary Material for Effective comparative analysis of protein-protein interaction networks by measuring the steady-state network flow using a Markov model Hyundoo Jeong 1, Xiaoning Qian 1 and
1 Supplementary Material for Effective comparative analysis of protein-protein interaction networks by measuring the steady-state network flow using a Markov model Hyundoo Jeong 1, Xiaoning Qian 1 and
Transmembrane Domains (TMDs) of ABC transporters
 of ABC transporters") Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Molecular Biology (9)
") Molecular Biology (9) Translation Mamoun Ahram, PhD Second semester, 2017-2018 1 Resources This lecture Cooper, Ch. 8 (297-319) 2 General information Protein synthesis involves interactions between three
Molecular Biology (9) Translation Mamoun Ahram, PhD Second semester, 2017-2018 1 Resources This lecture Cooper, Ch. 8 (297-319) 2 General information Protein synthesis involves interactions between three
Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus:
 m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
m Eukaryotic mrna processing Newly made RNA is called primary transcript and is modified in three ways before leaving the nucleus: Cap structure a modified guanine base is added to the 5 end. Poly-A tail
O.k., Now Starts the Good Stuff (Part II) Eukaryotic Cell Structure and Function
 Eukaryotic Cell Structure and Function") O.k., Now Starts the Good Stuff (Part II) Eukaryotic Cell Structure and Function Eukaryotic Cells These cells have membrane-bound structures called organelles. Cell processes occur in these organelles.
O.k., Now Starts the Good Stuff (Part II) Eukaryotic Cell Structure and Function Eukaryotic Cells These cells have membrane-bound structures called organelles. Cell processes occur in these organelles.
Biology of Fungi. Fungal Structure and Function. Lecture: Structure/Function, Part A BIOL 4848/ Fall Overview of the Hypha
 Biology of Fungi Fungal Structure and Function Overview of the Hypha The hypha is a rigid tube containing cytoplasm Growth occurs at the tips of hyphae Behind the tip, the cell is aging Diagram of hyphal
Biology of Fungi Fungal Structure and Function Overview of the Hypha The hypha is a rigid tube containing cytoplasm Growth occurs at the tips of hyphae Behind the tip, the cell is aging Diagram of hyphal
Supplementary Information
 Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Supplementary Information MAP2/Hoechst Hyp.-AP ph 6.5 Hyp.-SD ph 7.2 Norm.-SD ph 7.2 Supplementary Figure 1. Mitochondrial elongation in cortical neurons by acidosis. Representative images of neuronal
Host-Pathogen interaction-ii. Pl Path 604 PN Sharma Department of Plant Pathology CSK HPKV, Palampur
 Host-Pathogen interaction-ii Pl Path 604 PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 It was originally believed that gene-for-gene resistance was conferred by a direct interaction
Host-Pathogen interaction-ii Pl Path 604 PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 It was originally believed that gene-for-gene resistance was conferred by a direct interaction
Three different fusions led to three basic ideas: 1) If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes
 If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes") Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Supplementary information Supplementary figures
 Supplementary information Supplementary figures Supplementary Figure 1. Quantitation by FCS of GFP-tagged proteasomes in supernatant of cell extract (a) In vitro assay for evaluation of the hydrodynamic
Supplementary information Supplementary figures Supplementary Figure 1. Quantitation by FCS of GFP-tagged proteasomes in supernatant of cell extract (a) In vitro assay for evaluation of the hydrodynamic
9/25/2011. Outline. Overview: The Energy of Life. I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V.
 Chapter 8 Introduction to Metabolism Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes Overview: The Energy of Life Figure 8.1 The living cell is a miniature
Chapter 8 Introduction to Metabolism Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes Overview: The Energy of Life Figure 8.1 The living cell is a miniature
CHAPTER 3. Cell Structure and Genetic Control. Chapter 3 Outline
 CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
CHAPTER 3 Cell Structure and Genetic Control Chapter 3 Outline Plasma Membrane Cytoplasm and Its Organelles Cell Nucleus and Gene Expression Protein Synthesis and Secretion DNA Synthesis and Cell Division
4) Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.
 Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.") Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Biochemistry: A Review and Introduction
 Biochemistry: A Review and Introduction CHAPTER 1 Chem 40/ Chem 35/ Fundamentals of 1 Outline: I. Essence of Biochemistry II. Essential Elements for Living Systems III. Classes of Organic Compounds IV.
Biochemistry: A Review and Introduction CHAPTER 1 Chem 40/ Chem 35/ Fundamentals of 1 Outline: I. Essence of Biochemistry II. Essential Elements for Living Systems III. Classes of Organic Compounds IV.
Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter
 Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter 9/10/2008 1 Learning Objectives Explain why a cell cycle was selected for during evolution
Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter 9/10/2008 1 Learning Objectives Explain why a cell cycle was selected for during evolution
Chapter 6 A Tour of the Cell
 Chapter 6 A Tour of the Cell The cell is the basic unit of life Although cells differ substantially from one another, they all share certain characteristics that reflect a common ancestry and remind us
Chapter 6 A Tour of the Cell The cell is the basic unit of life Although cells differ substantially from one another, they all share certain characteristics that reflect a common ancestry and remind us
Chapter 6- An Introduction to Metabolism*
 Chapter 6- An Introduction to Metabolism* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Energy of Life
Chapter 6- An Introduction to Metabolism* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Energy of Life
A. The Cell: The Basic Unit of Life. B. Prokaryotic Cells. C. Eukaryotic Cells. D. Organelles that Process Information
 The Organization of Cells A. The Cell: The Basic Unit of Life Lecture Series 4 The Organization of Cells B. Prokaryotic Cells C. Eukaryotic Cells D. Organelles that Process Information E. Organelles that
The Organization of Cells A. The Cell: The Basic Unit of Life Lecture Series 4 The Organization of Cells B. Prokaryotic Cells C. Eukaryotic Cells D. Organelles that Process Information E. Organelles that
7.L.1.2 Plant and Animal Cells. Plant and Animal Cells
 7.L.1.2 Plant and Animal Cells Plant and Animal Cells Clarifying Objective: 7.L.1.2 Compare the structures and functions of plant and animal cells; include major organelles (cell membrane, cell wall, nucleus,
7.L.1.2 Plant and Animal Cells Plant and Animal Cells Clarifying Objective: 7.L.1.2 Compare the structures and functions of plant and animal cells; include major organelles (cell membrane, cell wall, nucleus,
Course Outcomes Guide (COG)
") Course Outcomes Guide (COG) Directions: Please complete this form to document your progress toward improving student learning. For each item, indicate your progress and your anticipated next steps. Thank
Course Outcomes Guide (COG) Directions: Please complete this form to document your progress toward improving student learning. For each item, indicate your progress and your anticipated next steps. Thank
CHARACTERISTICS OF LIFE ORGANIZATION OF LIFE CELL THEORY TIMELINE
 CHARACTERISTICS OF LIFE 1. composed of cells either uni/multi 2. reproduce sexual and/or asexual 3. contain DNA in cells 4. grow and develop 5. use material/energy in metabolic reactions 6. respond to
CHARACTERISTICS OF LIFE 1. composed of cells either uni/multi 2. reproduce sexual and/or asexual 3. contain DNA in cells 4. grow and develop 5. use material/energy in metabolic reactions 6. respond to
Biology: Life on Earth
 Teresa Audesirk Gerald Audesirk Bruce E. Byers Biology: Life on Earth Eighth Edition Lecture for Chapter 4 Cell Structure and Function Copyright 2008 Pearson Prentice Hall, Inc. Chapter 4 Outline 4.1 What
Teresa Audesirk Gerald Audesirk Bruce E. Byers Biology: Life on Earth Eighth Edition Lecture for Chapter 4 Cell Structure and Function Copyright 2008 Pearson Prentice Hall, Inc. Chapter 4 Outline 4.1 What
Vacuole Biogenesis in Saccharomyces cerevisiae: Protein Transport Pathways to the Yeast Vacuole
 MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, Mar. 1998, p. 230 247 Vol. 62, No. 1 1092-2172/98/$04.00 0 Copyright 1998, American Society for Microbiology Vacuole Biogenesis in Saccharomyces cerevisiae:
MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, Mar. 1998, p. 230 247 Vol. 62, No. 1 1092-2172/98/$04.00 0 Copyright 1998, American Society for Microbiology Vacuole Biogenesis in Saccharomyces cerevisiae:
Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question.
 Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) The receptors for a group of signaling molecules known as growth factors are
Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) The receptors for a group of signaling molecules known as growth factors are
Cell Is the basic structural, functional, and biological unit of all known living organisms. Cells are the smallest unit of life and are often called
 The Cell Cell Is the basic structural, functional, and biological unit of all known living organisms. Cells are the smallest unit of life and are often called the "building blocks of life". The study of
The Cell Cell Is the basic structural, functional, and biological unit of all known living organisms. Cells are the smallest unit of life and are often called the "building blocks of life". The study of
7.06 Spring 2004 PS 6 KEY 1 of 14
 7.06 Spring 2004 PS 6 KEY 1 of 14 Problem Set 6. Question 1. You are working in a lab that studies hormones and hormone receptors. You are tasked with the job of characterizing a potentially new hormone
7.06 Spring 2004 PS 6 KEY 1 of 14 Problem Set 6. Question 1. You are working in a lab that studies hormones and hormone receptors. You are tasked with the job of characterizing a potentially new hormone
Tuesday 9/6/2018 Mike Mueckler
 Tuesday 9/6/2018 Mike Mueckler mmueckler@wustl.edu Intracellular Targeting of Nascent Polypeptides Mitochondria are the Sites of Oxidative ATP Production Sugars Triglycerides Figure 14-10 Molecular Biology
Tuesday 9/6/2018 Mike Mueckler mmueckler@wustl.edu Intracellular Targeting of Nascent Polypeptides Mitochondria are the Sites of Oxidative ATP Production Sugars Triglycerides Figure 14-10 Molecular Biology
Takehome group assignment #1 - Individual worksheet NAME
 Takehome group assignment #1 - Individual worksheet NAME Answer the questions below, on your own (pp. 1-5). Then get together with your group, discuss your answers and complete the final group worksheet
Takehome group assignment #1 - Individual worksheet NAME Answer the questions below, on your own (pp. 1-5). Then get together with your group, discuss your answers and complete the final group worksheet
Lecture 4. Protein Translocation & Nucleocytoplasmic Transport
 Lecture 4 Protein Translocation & Nucleocytoplasmic Transport Chapter 12 MBoC (5th Edition) Alberts et al. Reference paper: Tran and Wente, Cell 125, 1041-1053, 2006 2/8/2012 1 Page 713 Molecular Biology
Lecture 4 Protein Translocation & Nucleocytoplasmic Transport Chapter 12 MBoC (5th Edition) Alberts et al. Reference paper: Tran and Wente, Cell 125, 1041-1053, 2006 2/8/2012 1 Page 713 Molecular Biology
Name # Class Date Regents Review: Cells & Cell Transport
 Name # Class Date Regents Review: Cells & Cell Transport 1. All of the following are true regarding cells except? A) All cells have genetic material B) All cells have cell walls C) All cells have plasma
Name # Class Date Regents Review: Cells & Cell Transport 1. All of the following are true regarding cells except? A) All cells have genetic material B) All cells have cell walls C) All cells have plasma
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Reviewers' comments: Reviewer #1 (Remarks to the Author):
:") Reviewers' comments: Reviewer #1 (Remarks to the Author): The manuscript by Malli and coworkers reports the successful development and characterization of the first K+ specific FRET-based sensor protein.
Reviewers' comments: Reviewer #1 (Remarks to the Author): The manuscript by Malli and coworkers reports the successful development and characterization of the first K+ specific FRET-based sensor protein.
Big Idea 2: Biological systems utilize free energy and molecular building blocks to grow, to reproduce and to maintain dynamic homeostasis.
 Big Idea 2: Biological systems utilize free energy and molecular building blocks to grow, to reproduce and to maintain dynamic homeostasis. Enduring understanding 2.B: Growth, reproduction and dynamic
Big Idea 2: Biological systems utilize free energy and molecular building blocks to grow, to reproduce and to maintain dynamic homeostasis. Enduring understanding 2.B: Growth, reproduction and dynamic
Class Work 31. Describe the function of the Golgi apparatus? 32. How do proteins travel from the E.R. to the Golgi apparatus? 33. After proteins are m
 Eukaryotes Class Work 1. What does the word eukaryote mean? 2. What is the one major difference between eukaryotes and prokaryotes? 3. List the different kingdoms of the eukaryote domain in the order in
Eukaryotes Class Work 1. What does the word eukaryote mean? 2. What is the one major difference between eukaryotes and prokaryotes? 3. List the different kingdoms of the eukaryote domain in the order in
T HE C ELL C H A P T E R 1 P G. 4-23
 T HE C ELL C H A P T E R 1 P G. 4-23 A CELL IS THE SMALLEST LIVING UNIT KNOWN. IT IS OFTEN CALLED THE BUILDING BLOCK OF THE BODY, AND IS THE BASIC STRUCTURAL AND FUNCTIONAL UNIT OF AN ORGANISM. CELL THEORY
T HE C ELL C H A P T E R 1 P G. 4-23 A CELL IS THE SMALLEST LIVING UNIT KNOWN. IT IS OFTEN CALLED THE BUILDING BLOCK OF THE BODY, AND IS THE BASIC STRUCTURAL AND FUNCTIONAL UNIT OF AN ORGANISM. CELL THEORY
Cells. A. The iodine diffused into the bag. B. The starch was changed to sugar.
 Name: Date: 1. A student filled a bag of dialysis tubing with a milky-white starch solution and placed the bag in a beaker of iodine-water as shown in the diagram. An hour later, the student observed that
Name: Date: 1. A student filled a bag of dialysis tubing with a milky-white starch solution and placed the bag in a beaker of iodine-water as shown in the diagram. An hour later, the student observed that
Reading Assignments. A. Genes and the Synthesis of Polypeptides. Lecture Series 7 From DNA to Protein: Genotype to Phenotype
 Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
Lecture Series 7 From DNA to Protein: Genotype to Phenotype Reading Assignments Read Chapter 7 From DNA to Protein A. Genes and the Synthesis of Polypeptides Genes are made up of DNA and are expressed
What is the central dogma of biology?
 Bellringer What is the central dogma of biology? A. RNA DNA Protein B. DNA Protein Gene C. DNA Gene RNA D. DNA RNA Protein Review of DNA processes Replication (7.1) Transcription(7.2) Translation(7.3)
Bellringer What is the central dogma of biology? A. RNA DNA Protein B. DNA Protein Gene C. DNA Gene RNA D. DNA RNA Protein Review of DNA processes Replication (7.1) Transcription(7.2) Translation(7.3)
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
DOI: 10.1038/ncb2362 Figure S1 CYLD and CASPASE 8 genes are co-regulated. Analysis of gene expression across 79 tissues was carried out as described previously [Ref: PMID: 18636086]. Briefly, microarray
REVIEW 2: CELLS & CELL COMMUNICATION. A. Top 10 If you learned anything from this unit, you should have learned:
 Name AP Biology REVIEW 2: CELLS & CELL COMMUNICATION A. Top 10 If you learned anything from this unit, you should have learned: 1. Prokaryotes vs. eukaryotes No internal membranes vs. membrane-bound organelles
Name AP Biology REVIEW 2: CELLS & CELL COMMUNICATION A. Top 10 If you learned anything from this unit, you should have learned: 1. Prokaryotes vs. eukaryotes No internal membranes vs. membrane-bound organelles
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016
 Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 2 November 8, 2016 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Cellular Neuroanatomy I The Prototypical Neuron: Soma. Reading: BCP Chapter 2
 Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Cellular Neuroanatomy I The Prototypical Neuron: Soma Reading: BCP Chapter 2 Functional Unit of the Nervous System The functional unit of the nervous system is the neuron. Neurons are cells specialized
Molecular Cell Biology 5068 In Class Exam 1 September 30, Please print your name:
 Molecular Cell Biology 5068 In Class Exam 1 September 30, 2014 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Molecular Cell Biology 5068 In Class Exam 1 September 30, 2014 Exam Number: Please print your name: Instructions: Please write only on these pages, in the spaces allotted and not on the back. Write your
Honors Biology summer assignment. Review the notes and study them. There will be a test on this information the 1 st week of class
 Honors Biology summer assignment Review the notes and study them. There will be a test on this information the 1 st week of class Biomolecules Molecules that make up living things. There are 4 molecules
Honors Biology summer assignment Review the notes and study them. There will be a test on this information the 1 st week of class Biomolecules Molecules that make up living things. There are 4 molecules
Protein-Protein Interactions of the Cytoplasmic Loops of the Yeast Multidrug Resistance Protein,PDR 5: A Two -Hybrid Based Analysis.
 Researcher : Rao Subba G (2001) SUMMARY Guide :Gupta,C.M.(Dr.) Protein-Protein Interactions of the Cytoplasmic Loops of the Yeast Multidrug Resistance Protein,PDR 5: A Two -Hybrid Based Analysis. Mukidrug
Researcher : Rao Subba G (2001) SUMMARY Guide :Gupta,C.M.(Dr.) Protein-Protein Interactions of the Cytoplasmic Loops of the Yeast Multidrug Resistance Protein,PDR 5: A Two -Hybrid Based Analysis. Mukidrug
9/8/2010. Chapter 4. Structures Internal to the Cell Wall. The Plasma Membrane. Functional Anatomy of Prokaryotic and Eukaryotic Cells
 Chapter 4 Functional Anatomy of Prokaryotic and Eukaryotic Cells Johana Meléndez Part II slides 39-87 Lectures prepared by Christine L. Case Structures Internal to the Cell Wall Learning Objectives 4-8
Chapter 4 Functional Anatomy of Prokaryotic and Eukaryotic Cells Johana Meléndez Part II slides 39-87 Lectures prepared by Christine L. Case Structures Internal to the Cell Wall Learning Objectives 4-8
Class IX: Biology Chapter 5: The fundamental unit of life. Chapter Notes. 1) In 1665, Robert Hooke first discovered and named the cells.
 In 1665, Robert Hooke first discovered and named the cells.") Class IX: Biology Chapter 5: The fundamental unit of life. Key learnings: Chapter Notes 1) In 1665, Robert Hooke first discovered and named the cells. 2) Cell is the structural and functional unit of all
Class IX: Biology Chapter 5: The fundamental unit of life. Key learnings: Chapter Notes 1) In 1665, Robert Hooke first discovered and named the cells. 2) Cell is the structural and functional unit of all